Embed Size (px)
Citation preview

2 MANUAL DE DERMATOLOGÍA, 2.ª EDICIÓN - VOLUMEN I© 2018. Grupo Aula Médica
ANATOMÍA Y FISIOLOGÍA DE LA PIEL
Agustín Buendía Eisman1, José Mazuecos Blanca2 y Francisco M. Camacho Martínez3
1 Profesor Titular de Dermatología. Facultad de Medicina. Universidad de Granada.2 Catedrático de Escuela Universitaria. Área de Dermatología. Facultad de Medicina. Universidad de Sevilla. 3 Presidente de Honor de la Academia Española de Dermatología. Catedrático de Dermatología. Universidad de Sevilla.
1
Características generales
◗ La piel consta de tres capas bien diferenciadas: epidermis, dermis e hipodermis.
◗ La epidermis está constituida por queratinocitos, melanocitos, células de Largenhans y células de Merkel.
◗ La capa más externa de la epidermis o capa córnea se forma por la apoptosis de los queratinocitos.
◗ Las uniones entre queratinocitos son mediante desmosomas; y entre la capa basal y la dermis, mediante hemidesmosomas.
◗ La dermis es una capa conjuntiva que alberga los plexos vasculonerviosos y sirve de sostén a la epidermis y a sus anejos.
Está formada por fibras, como las de colágeno y las elásticas, y por células, como los fibrocitos, mastocitos e histiocitos. Tiene dos áreas bien distinguibles: superior, o dermis papilar, e inferior o dermis reticular.
◗ La hipodermis es la tercera capa, encargada de almacenar lípidos para aportar energía al organismo y aislante térmico.
◗ Las funciones de la piel son: protección, termorregulación, sensorial, secretora y excretora, inmunológica y producción de vitamina D.
Contenido
• Estructura general de la piel
• Funciones de la piel
• Embriología de la piel
• Estructura de la epidermis
• Sustancia fundamental
• Anejos cutáneos
• Manto cutáneo ácido lipídico
• Hipodermis
• Vasos y nervios cutáneos
• Tendencias en la investigación del desarrollo y estructura de la piel
• Bibliografía
Manual de Dermatología, 2.ª edición. Editores: J. Conejo-Mir, J. C. Moreno, F. M. Camacho, pp. 2-27. ISBN Volumen I: 978-84-7885-628-2.ISBN Obra completa: 978-84-7885-627-5. ISBN Volumen II: 978-84-7885-629-9.
MANUAL DE DERMATOLOGÍA, 2.ª EDICIÓN - VOLUMEN I 3
Imágenes clave
Los desmosomas forman una unión simétrica, tienenun diámetro de 0,1-0,5 nm y están formados por unoscomponentes transmembrana y otros de la propia placaelectrón-densa, con un espacio intercelular central (Figs.7, 8 y 11). Los componentes transmembrana son unasglicoproteínas, familias de las cadherinas, formando aso-ciaciones heterófilas de desmogleínas y de desmocoli-nas; hay tres desmogleínas (desmogleínas 1-3) y otrastres desmocolinas (desmocolinas 1-3), y los de la placason otras tres proteínas (desmoplaquina, placoglobinay placofilina), que son las que fijan las cadherinas a lostonofilamentos de queratina, que se disponen perpen-dicularmente a ella (Fig. 12).
Las uniones adherentes son estructuras transmembrana elec-trón-densas compuestas por cadherina E, la cual forma interac-ciones calcio-dependientes con la cadherina E de la célula adya-cente. Estas interacciones se conexionan con la membranaplasmática por medio de una red de proteínas adhesivas queincluyen α-catenina, β-catenina y p120ctn.
Las uniones tipo hendidura (gap) son un grupo de canalesintercelulares conocidos como conexones, que forman cone-xiones entre los citoplasmas de los queratinocitos adyacen-tes. Los conexones se originan en el aparato de Golgimediante el ensamblaje de seis subunidades de conexinas,sintetizadas en el retículo endoplásmica, siendo despuéstransportados a la membrana plasmática, donde se agregancon otros conexones y, en combinación con los agregados delqueratinocito adyacente, forman la unión gap. La función deestas uniones tipo hendidura es la transferencia de moléculasde bajo peso molecular (< 1000 Da) y de intercambio de ionesentre células vecinas.
Las uniones ajustadas están compuestas de moléculastransmembrana e intracelulares, como la ocludina y las clau-dinas. Tienen como función la de controlar la permeabilidadepidérmica y la de mantener la polaridad celular.
EpidermopoyesisEn la cinética celular de la epidermis, el concepto más impor-tante es el tiempo de regeneración celular (turnover time),que define el tiempo medio de una población celular parareproducirse a sí misma; es decir, el intervalo entre dos mito-sis sucesivas de las células germinativas epidérmicas.
Aunque turnover time también se emplea para definir elpaso de una célula desde el estrato germinativo a la capa cór-nea epitelial, nos parece más lógico emplear el términotiempo de tránsito para denominar este periodo. Ya hemosindicado que es de 52 a 75 días; sin embargo, un epitelio pso-riásico sólo necesita de 8 a 10 días. El hecho de que el tamañode la epidermis permanezca constante, hace pensar que latasa de producción celular en el estrato germinativo está equi-librada con la pérdida celular en el estrato córneo.
La capa basal tiene una serie de controles estimuladoresy/o inhibidores que permiten, en determinadas circunstan-cias, producir más o menos células.
Factores estimuladores� Familia del Factor de crecimiento epidérmico (EGF,
«Epidermal Growth Factor»). El EGF humano es un polipép-tido que estimula la proliferación y diferenciación celularen un amplio rango de tejidos. Se encuentra en las glándu-las salivales, plaquetas y algunas glándulas del duodeno.El EGF incrementa la epidermopoyesis uniéndose a unos
Tem
as g
ener
ales
10
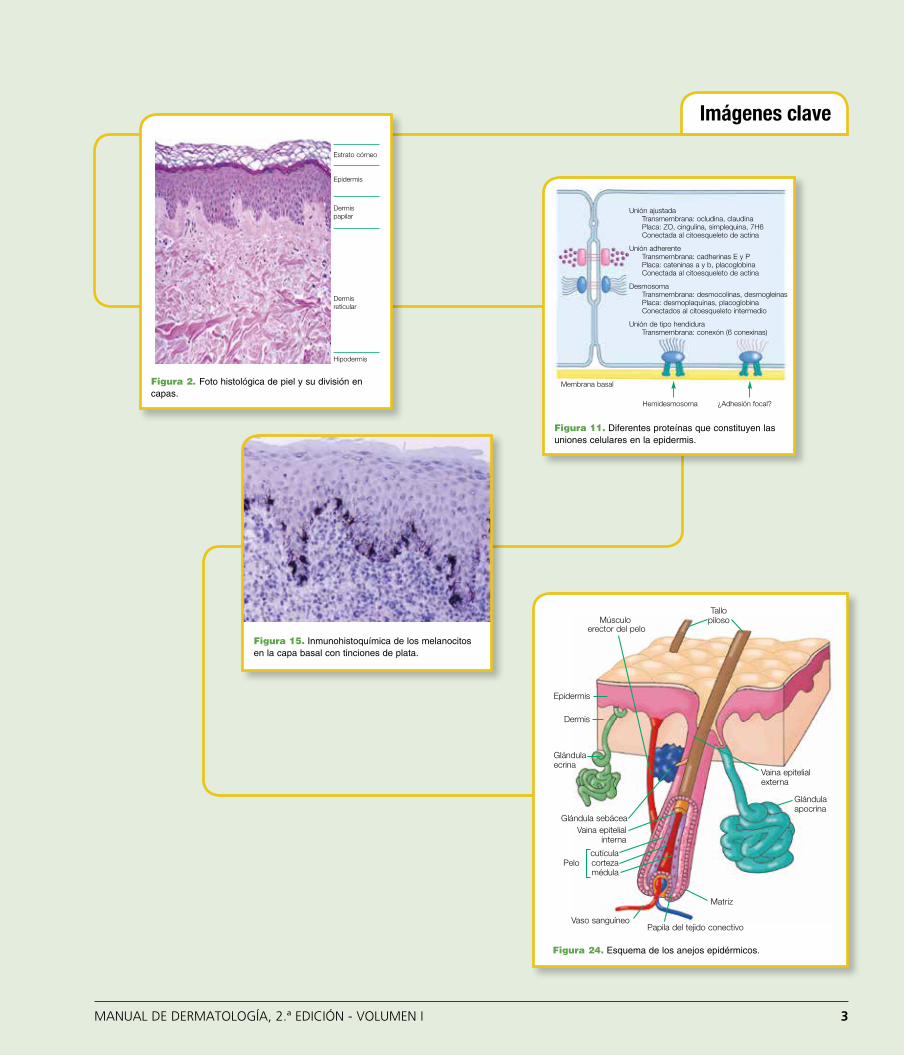
Figura 11. Diferentes proteínas que constituyen las uniones celula-res en la epidermis.
Unión ajustadaTransmembrana: ocludina, claudinaPlaca: ZO, cingulina, simplequina, 7H6Conectada al citoesqueleto de actina
Unión adherenteTransmembrana: cadherinas E y PPlaca: cateninas a y b, placoglobinaConectada al citoesqueleto de actina
DesmosomaTransmembrana: desmocolinas, desmogleinasPlaca: desmoplaquinas, placoglobinaConectados al citoesqueleto intermedio
Unión de tipo hendiduraTransmembrana: conexón (6 conexinas)
Membrana basal
Hemidesmosoma ¿Adhesión focal?
Figura 12. Componentes de los desmosomas.
Desmocolina
Desmogleína
Placoglobina
Placofilina
Desmoplaquina
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 10
Figura 11. Diferentes proteínas que constituyen las uniones celulares en la epidermis.
Figura 15. Inmunohistoquímica de los melanocitos en la capa basal con tinciones de plata.
Figura 2. Foto histológica de piel y su división en capas.
Tem
as g
ener
ales
4
Figura 1. Dibujo esquemático de la piel.
Figura 2. Foto histológica de piel y su división en capas.
Estrato córneo
Epidermis
Dermispapilar
Dermisreticular
Hipodermis
vasos y nervios, y se ve perforado por los anejos, unos glan-dulares (glándulas sebáceas y sudoríparas ecrinas y apocri-nas) y otros queratinizados (pelos y uñas).
Inmediatamente por debajo, y separada por la unióndermo-epidérmica, se encuentra la dermis, estratoconjuntivo 20 a 30 veces mayor que la capa anterior,que alberga en su interior los plexos vasculonervio-sos y sirve de sostén a la epidermis y a sus anejos.Está formada por un componente fibroso, queincluye fibras de colágeno (principal estructura de ladermis) y fibras elásticas. Sus células constitutivasson los fibroblastos, como las células más importantes,los mastocitos y los histiocitos. Estos dos componen-tes se encuentran dentro de una sustancia funda-mental, en la que predominan los mucopolisacári-dos hidratados, con gran capacidad para reteneragua.Por debajo de la dermis se encuentra la hipodermis,panículo adiposo o tejido celular subcutáneo, que,aparte de contener algunos elementos vasculonervio-sos, es un perfecto aislante térmico y sirve de pro tec-ción frente a los traumatismos a los órganos internos.Ya debajo existe una fascia fibrosa profunda, que seconsidera el límite cutáneo.
Figura 3. Esquema de las capas de la epidermis.
Capa córnea
Estrato lúcido
Capa granulosa
Capa espinosa
Capa basal
Dermis
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 4
Figura 24. Esquema de los anejos epidérmicos.
y paralelas a la superficie cutánea en la dermis profunda,siendo más gruesas. La síntesis de las fibras elásticas porlos fibrocitos consta de dos etapas. En la primera se secre-tan al espacio extracelular las microfibrillas agrupadas deforma lineal produciéndose una polimerización posterior-mente. En la segunda se produce la secreción de la tropoe-lastina dentro de los cilindros preformados de microfibri-llas; la elastina se une a ellas por medio de unionescovalentes conocidas como desmosinas e isodesmosinas.Estas fibras son muy numerosas al nacer, observándosecomo bandas de material amorfo rodeadas de un grannúmero de microfibrillas de elastina. Pocos meses despuésdel nacimiento, aumenta la cantidad de elastina amorfa yya se comprueba la organización típica de las fibras elásti-cas maduras. Durante la vida adulta, las fibras elásticasmuestran pocas variaciones a excepción de una discretareducción de las microfibrillas y la presencia de algunasinclusiones electrón-densas y de márgenes irregulares, loque se hace más evidente sobre los cincuenta años.
Morfométricamente, las fibras elásticas representan el3-4% del total de la dermis en el recién nacido, nocambiando significativamente hasta la quinta décadade la vida, a partir de la cual aumentan al 5% y en losancianos llegan al 7-8%. Este aumento se debe almayor diámetro de las fibras y al mayor número porunidad de área. Las mujeres ancianas muestran másfibras elásticas que los varones.
La degradación de la elastina por medio de las elastasasda lugar a la formación de unos fragmentos peptídicos deno-minados elastocinas, para remarcar sus propiedades de cito-cinas, ya que se ha visto que representan una importanteseñal de reparación tisular.
SUSTANCIA FUNDAMENTAL
Es un material extracelular amorfo que se encuentra entre lasfibras propias de la dermis, siendo también producida por losfibrocitos. Está constituida por agua, electrolitos, proteínasplasmáticas y proteoglicanos. Los proteoglicanos se tratan decadenas de polisacáridos aminados (glicosaminoglicanos) uni-das a proteínas centrales.
En la piel, los glicosaminoglicanos más abundantes sondermatan-sulfato y ácido hialurónico junto con pequeñas can-tidades de condroitin-6-sulfato, heparan-sulfato y heparina.Los glicosaminoglicanos son muy higroscópicos reteniendogran cantidad de agua.
En la matriz extracelular también se encuentran diversasglicoproteínas de adhesión como fibronectina, vitronectina,trombospondinas y tenascinas. Estas glicoproteínas se unen alas superficies celulares, a los glicosaminoglicanos y a lasfibras de colágeno provocando unas interacciones estables.
Las fibronectinas que se unen a las fibras de colágeno tipoIII, anteriormente denominadas «fibras reticulares», son lasresponsables de las reacciones argirófilas de esas fibras.
ANEJOS CUTÁNEOS
Como señalamos antes, los hay queratinizados y glandulares;unos claramente separados y otros en íntima relación, confor-mando el denominado folículo pilosebáceo.
Anejos queratinizadosSon dos: el pelo, que forma parte del folículo piloso, y la uña.
Folículo pilosoEl folículo piloso tiene una porción distal conocida como«bulbo», compuesto por la matriz pilosa y la papila y uncuerpo cilíndrico en el que, de abajo arriba, se observa lainserción del músculo erector del pelo por medio del tendónelástico de Nagel, constituido por fibras elásticas elaunínicasy oxitalánicas, la glándula sebácea y la glándula sudoríparaapocrina (Figs. 24, 25 y 26).
Las células matriciales, al multiplicarse y diferenciarse, for-man el pelo y la vaina epitelial interna. Entre las matricialesindiferenciadas y las ya diferenciadas, queda un límite que se
1.A
nato
mía
y f
isio
logí
a de
la p
iel
19
Figura 24. Esquema de los anejos epidérmicos.
Epidermis
Dermis
Glándulaecrina
Músculoerector del pelo
Glándula sebáceaVaina epitelial
interna
Pelocutículacortezamédula
Vaso sanguíneoPapila del tejido conectivo
Matriz
Glándulaapocrina
Vaina epitelialexterna
Tallopiloso
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 19
4
Manual de derMatología. Sección I: Temas generales
ESTRUCTURA GENERAL DE LA PIEL
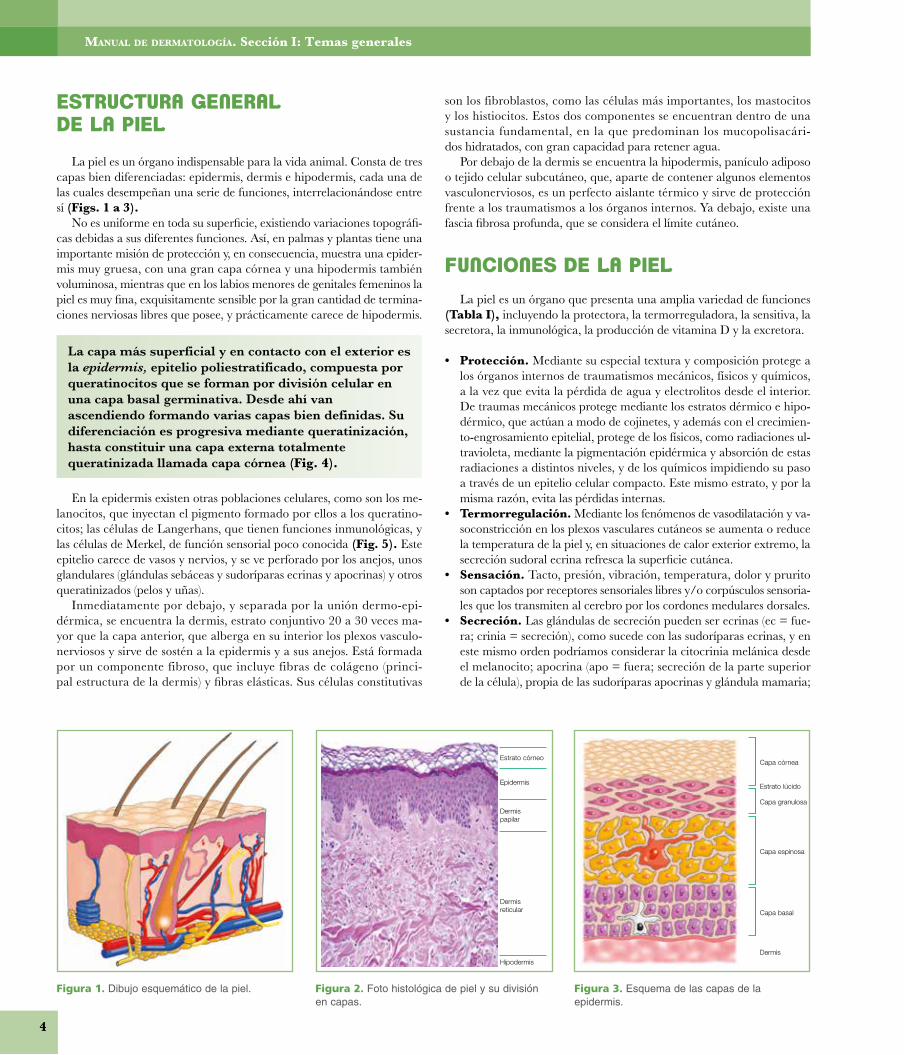
La piel es un órgano indispensable para la vida animal. Consta de tres capas bien diferenciadas: epidermis, dermis e hipodermis, cada una de las cuales desempeñan una serie de funciones, interrelacionándose entre sí (Figs. 1 a 3).
No es uniforme en toda su superficie, existiendo variaciones topográfi-cas debidas a sus diferentes funciones. Así, en palmas y plantas tiene una importante misión de protección y, en consecuencia, muestra una epider-mis muy gruesa, con una gran capa córnea y una hipodermis también voluminosa, mientras que en los labios menores de genitales femeninos la piel es muy fina, exquisitamente sensible por la gran cantidad de termina-ciones nerviosas libres que posee, y prácticamente carece de hipodermis.
La capa más superficial y en contacto con el exterior es la epidermis, epitelio poliestratificado, compuesta por queratinocitos que se forman por división celular en una capa basal germinativa. Desde ahí van ascendiendo formando varias capas bien definidas. Su diferenciación es progresiva mediante queratinización, hasta constituir una capa externa totalmente queratinizada llamada capa córnea (Fig. 4).
En la epidermis existen otras poblaciones celulares, como son los me-lanocitos, que inyectan el pigmento formado por ellos a los queratino-citos; las células de Langerhans, que tienen funciones inmunológicas, y las células de Merkel, de función sensorial poco conocida (Fig. 5). Este epitelio carece de vasos y nervios, y se ve perforado por los anejos, unos glandulares (glándulas sebáceas y sudoríparas ecrinas y apocrinas) y otros queratinizados (pelos y uñas).
Inmediatamente por debajo, y separada por la unión dermo-epi-dérmica, se encuentra la dermis, estrato conjuntivo 20 a 30 veces ma-yor que la capa anterior, que alberga en su interior los plexos vasculo-nerviosos y sirve de sostén a la epidermis y a sus anejos. Está formada por un componente fibroso, que incluye fibras de colágeno (princi-pal estructura de la dermis) y fibras elásticas. Sus células constitutivas
son los fibroblastos, como las células más importantes, los mastocitos y los histiocitos. Estos dos componentes se encuentran dentro de una sustancia fundamental, en la que predominan los mucopolisacári- dos hidratados, con gran capacidad para retener agua.
Por debajo de la dermis se encuentra la hipodermis, panículo adiposo o tejido celular subcutáneo, que, aparte de contener algunos elementos vasculonerviosos, es un perfecto aislante térmico y sirve de pro tección frente a los traumatismos a los órganos internos. Ya debajo, existe una fascia fibrosa profunda, que se considera el límite cutáneo.
FUNCIONES DE LA PIEL
La piel es un órgano que presenta una amplia variedad de funciones (Tabla I), incluyendo la protectora, la termorreguladora, la sensitiva, la secretora, la inmunológica, la producción de vitamina D y la excretora.
• Protección. Mediante su especial textura y composición protege a los órganos internos de traumatismos mecánicos, físicos y químicos, a la vez que evita la pérdida de agua y electrolitos desde el interior. De traumas mecánicos protege mediante los estratos dérmico e hipo-dérmico, que actúan a modo de cojinetes, y además con el crecimien-to-engrosamiento epitelial, protege de los físicos, como radiaciones ul-travioleta, mediante la pigmentación epidérmica y absorción de estas radiaciones a distintos niveles, y de los químicos impidiendo su paso a través de un epitelio celular compacto. Este mismo estrato, y por la misma razón, evita las pérdidas internas.
• Termorregulación. Mediante los fenómenos de vasodilatación y va-soconstricción en los plexos vasculares cutáneos se aumenta o reduce la temperatura de la piel y, en situaciones de calor exterior extremo, la secreción sudoral ecrina refresca la superficie cutánea.
• Sensación. Tacto, presión, vibración, temperatura, dolor y prurito son captados por receptores sensoriales libres y/o corpúsculos sensoria-les que los transmiten al cerebro por los cordones medulares dorsales.
• Secreción. Las glándulas de secreción pueden ser ecrinas (ec = fue-ra; crinia = secreción), como sucede con las sudoríparas ecrinas, y en este mismo orden podríamos considerar la citocrinia melánica desde el melanocito; apocrina (apo = fuera; secreción de la parte superior de la célula), propia de las sudoríparas apocrinas y glándula mamaria;
Figura 1. Dibujo esquemático de la piel.
Tem
as g
ener
ales
4
Figura 1. Dibujo esquemático de la piel.
Figura 2. Foto histológica de piel y su división en capas.
Estrato córneo
Epidermis
Dermispapilar
Dermisreticular
Hipodermis
vasos y nervios, y se ve perforado por los anejos, unos glan-dulares (glándulas sebáceas y sudoríparas ecrinas y apocri-nas) y otros queratinizados (pelos y uñas).
Inmediatamente por debajo, y separada por la unióndermo-epidérmica, se encuentra la dermis, estratoconjuntivo 20 a 30 veces mayor que la capa anterior,que alberga en su interior los plexos vasculonervio-sos y sirve de sostén a la epidermis y a sus anejos.Está formada por un componente fibroso, queincluye fibras de colágeno (principal estructura de ladermis) y fibras elásticas. Sus células constitutivasson los fibroblastos, como las células más importantes,los mastocitos y los histiocitos. Estos dos componen-tes se encuentran dentro de una sustancia funda-mental, en la que predominan los mucopolisacári-dos hidratados, con gran capacidad para reteneragua.Por debajo de la dermis se encuentra la hipodermis,panículo adiposo o tejido celular subcutáneo, que,aparte de contener algunos elementos vasculonervio-sos, es un perfecto aislante térmico y sirve de pro tec-ción frente a los traumatismos a los órganos internos.Ya debajo existe una fascia fibrosa profunda, que seconsidera el límite cutáneo.
Figura 3. Esquema de las capas de la epidermis.
Capa córnea
Estrato lúcido
Capa granulosa
Capa espinosa
Capa basal
Dermis
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 4
Figura 2. Foto histológica de piel y su división en capas.
Tem
as g
ener
ales
4
Figura 1. Dibujo esquemático de la piel.
Figura 2. Foto histológica de piel y su división en capas.
Estrato córneo
Epidermis
Dermispapilar
Dermisreticular
Hipodermis
vasos y nervios, y se ve perforado por los anejos, unos glan-dulares (glándulas sebáceas y sudoríparas ecrinas y apocri-nas) y otros queratinizados (pelos y uñas).
Inmediatamente por debajo, y separada por la unióndermo-epidérmica, se encuentra la dermis, estratoconjuntivo 20 a 30 veces mayor que la capa anterior,que alberga en su interior los plexos vasculonervio-sos y sirve de sostén a la epidermis y a sus anejos.Está formada por un componente fibroso, queincluye fibras de colágeno (principal estructura de ladermis) y fibras elásticas. Sus células constitutivasson los fibroblastos, como las células más importantes,los mastocitos y los histiocitos. Estos dos componen-tes se encuentran dentro de una sustancia funda-mental, en la que predominan los mucopolisacári-dos hidratados, con gran capacidad para reteneragua.Por debajo de la dermis se encuentra la hipodermis,panículo adiposo o tejido celular subcutáneo, que,aparte de contener algunos elementos vasculonervio-sos, es un perfecto aislante térmico y sirve de pro tec-ción frente a los traumatismos a los órganos internos.Ya debajo existe una fascia fibrosa profunda, que seconsidera el límite cutáneo.
Figura 3. Esquema de las capas de la epidermis.
Capa córnea
Estrato lúcido
Capa granulosa
Capa espinosa
Capa basal
Dermis
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 4
Figura 3. Esquema de las capas de la epidermis.

5
Capítulo 1. Anatomía y fisiología de la piel
y holocrinas (secreción de la totalidad celular), representadas por las glándulas sebáceas y el propio epitelio epidérmico.
• Función inmunológica. Se ha demostrado que los queratinocitos intervienen de forma activa en el sistema inmune cutáneo o SALT (tejido linfoide asociado a la piel), tanto en las interacciones celula-res con las células de Langerhans y los linfocitos T epidermotrópicos, como en la producción de citocinas. Los histiocitos dérmicos también intervienen en la función defensiva cutánea.
Los péptidos antimicrobianos son un grupo de péptidos presentes en la superficie epidérmica que actúan como antibióticos naturales y par-ticipan en los procesos celulares de la defensa inmune y la reparación tisular. Hay dos grupos principales, las catelicidinas y las defensinas a y b. Normalmente se producen pequeñas cantidades de estos péptidos antimicrobianos en la epidermis, acumulándose alrededor de los folícu-los pilosos y las glándulas sudoríparas ecrinas, donde la función barrera está ausente o disminuida; cuando existe una infección o una herida, los queratinocitos incrementan rápidamente su producción, reclutando a los neutrófilos como parte de la respuesta inflamatoria aguda.
• Producción de vitamina D. La piel es el único órgano donde, en condiciones fisiológicas e inducida por la radiación UVB, se realiza la transformación completa del 7-dehidrocolesterol en calcitriol (1,25-di-hidroxivitamina D3). El calcitriol regula también el crecimiento y la diferenciación de los queratinocitos, por lo que se han introducido los análogos de la vitamina D en la terapéutica de las dermatosis hiper-proliferativas.
• Excreción. Hay que comentar que a través de la piel se eliminan muy pocas sustancias aunque, en determinadas situaciones patológicas, al producirse grandes cantidades de capa córnea, se pueden perder ele-mentos constitutivos del epitelio, especialmente azufre y proteínas. En la excreción cutánea también debemos considerar la perspiratio insensi-bilis, que es la pérdida de agua diaria a través de la superficie cutánea, sin relación con la secreción ecrina, y que para un varón de 70 kg, que se correspondería con una superficie de 1,80 m2, es de unos 350 ml.
EMBRIOLOGÍA DE LA PIEL
La epidermis, mucosas y anejos epidérmicos proceden del ectodermo, mientras que dermis e hipodermis del mesodermo. Aproximadamente en la tercera semana, el embrión está cubierto de una fina membrana uni-celular que, a partir de la quinta o sexta, se divide en dos: una superficial, o peridermo, y otra profunda, o estrato germinativo. Ya en el tercer mes
aparece un estrato intermedio como consecuencia de multiplicación del estrato germinativo. Entre el tercer y cuarto mes se diferencian desde el estrato germinativo las células basales que, al dividirse a lo largo del cuar-to y quinto mes, originan las capas espinosa, granulosa, lúcida y córnea, que acabarán sustituyendo al peridermo. La lámina densa de la unión dermo-epidérmica se observa en el segundo mes y los hemidesmosomas en el tercero.
Al mismo tiempo que del estrato germinativo surgen las células basales, se pueden observar los dos gérmenes epiteliales.
El germen epitelial primario surge como pregermen o germen piloso primitivo en el tercer mes, constituido únicamente por una concentración nuclear en la capa basal. Rápidamente las células basales se alargan y penetran en la dermis, formando el germen piloso, bajo el cual se acumulan numerosos núcleos mesodérmicos que formarán la futura papila. Las células basales se van multiplicando y penetrando de forma oblicua en la dermis, empujando al conjunto de núcleos mesodérmicos a los que va englobando poco a poco en su parte distal: es la etapa de clava pilosa. Al final de este proceso se observan dos protuberancias en la pared folicular: la superior, que es el esbozo de la glándula sebácea, y la inferior, llamado «bulge» o «protuberancia», zona donde se insertará el músculo erector. Se ha demostrado que en la protuberancia se encuentran células matriciales capaces de iniciar el anagen folicular estimulando las células de la papila dérmica.
Dos meses después, por encima de la glándula sebácea, brota un nue-vo engrosamiento, en el que se forma la glándula sudorípara apocrina. Posteriormente, en la etapa de diferenciación, parten desde la epidermis células para formar el canal del pelo y otras exteriores que dan lugar a la vaina epitelial externa y en la porción distal o bulbo piloso, que ya englo-ba las células de la papila, se forma la matriz, cuyas células se multiplican, dando lugar al pelo y vaina epitelial interna.
Desde aproximadamente el tercer mes de vida intrauterino, las células matriciales que se encuentran en el abultamiento superior del folículo dan lugar a la glándula sebácea. Entre el quinto y sexto mes prolifera el abultamiento superior, o de la glándula sudorípara apocrina, en forma
FUNCIONES DE LA PIEL
La piel es un órgano que presenta una amplia variedad de fun-ciones, incluyendo la protectora, la termorreguladora, la sen-sitiva, la secretora, la inmunológica, la producción de vita-mina D y la excretora.
� Protección. Mediante su especial textura y composiciónprotege a los órganos internos de traumatismos mecáni-cos, físicos y químicos, a la vez que evita la pérdida deagua y electrólitos desde el interior. De traumas mecáni-cos protege mediante los estratos dérmico e hipodérmico,que actúan a modo de cojinetes, y además con el creci-miento-engrosamiento epitelial, protege de los físicos,como radiaciones ultravioleta, mediante la pigmentaciónepidérmica y absorción de estas radiaciones a distintosniveles, y de los químicos impidiendo su paso a través deun epitelio celular compacto. Este mismo estrato, y por lamisma razón, evita las pérdidas internas.
� Termorregulación. Mediante los fenómenos de vasodila-tación y vasoconstricción en los plexos vasculares cutá-neos se aumenta o reduce la temperatura de la piel y, ensituaciones de calor exterior extremo, la secreción sudoralecrina refresca la superficie cutánea.
� Sensación. Tacto, presión, vibración, temperatura, dolory prurito son captados por receptores sensoriales libresy/o corpúsculos sensoriales que los transmiten al cerebropor los cordones medulares dorsales.
� Secreción. Las glándulas de secreción pueden ser ecrinas(ec = fuera; crinia = secreción), como sucede con las sud-oríparas ecrinas, y en este mismo orden podríamos consi-derar la citocrínia melánica desde el melanocito; apocrina(apo = fuera; secreción de la parte superior de la célula),propia de las sudoríparas apocrinas y glándula mamaria; yholocrinas (secreción de la totalidad celular), representa-das por las glándulas sebáceas y el propio epitelio epidér-mico.
� Función inmunológica. Se ha demostrado que los que-ratinocitos intervienen de forma activa en el sistemainmune cutáneo o SALT (tejido linfoide asociado a la piel),tanto en las interacciones celulares con las células de Lan-gerhans y los linfocitos T epidermotrópicos como en laproducción de citocinas. Los histiocitos dérmicos tambiénintervienen en la función defensiva cutánea.
Los péptidos antimicrobianos son un grupo de péptidospresentes en la superficie epidérmica que actúan comoantibióticos naturales y participan en los procesos celula-res de la defensa inmune y la reparación tisular. Hay dosgrupos principales, las catelicidinas y las defensinas α y β.Normalmente se producen pequeñas cantidades de estospéptidos antimicrobianos en la epidermis, acumulándosealrededor de los folículos pilosos y las glándulas sudorípa-ras ecrinas, donde la función barrera está ausente o dismi.C; cuando existe una infección o una herida, los querati-nocitos incrementan rápidamente su producción, reclu-tando a los neutrófilos como parte de la respuesta infla-matoria aguda.
� Producción de vitamina D. La piel es el único órganodonde, en condiciones fisiológicas e inducida por la radia-ción UVB, se realiza la transformación completa del 7-dehi-drocolesterol en calcitriol (1,25-dihidroxivitamina D3). Elcalcitriol regula también el crecimiento y la diferenciaciónde los queratinocitos, por lo que se han introducido losanálogos de la vitamina D en la terapéutica de las derma-tosis hiperproliferativas.
� Excreción. Hay que comentar que a través de la piel seeliminan muy pocas sustancias aunque, en determinadassituaciones patológicas, al producirse grandes cantidadesde capa córnea, se pueden perder elementos constitutivosdel epitelio, especialmente azufre y proteínas. En la excre-ción cutánea también debemos considerar la perspiratioinsensibilis, que es la pérdida de agua diaria a través de lasuperficie cutánea, sin relación con la secreción ecrina, yque para un varón de 70 kg, que se correspondería conuna superficie de 1,80 m2, es de unos 350 ml.
EMBRIOLOGÍA DE LA PIEL
La epidermis, mucosas y anejos epidérmicos proceden delectodermo, mientras que dermis e hipodermis del meso-
1.A
nato
mía
y f
isio
logí
a de
la p
iel
5
Figura 4. Esquema de la capa córnea.
Corneocitos
Figura 5. Esquema de la celularidad de la epidermis.
Célula deLangerhans
Queratinocito
Melanocito Célula de Merkel
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 5
Figura 4. Esquema de la capa córnea.
FUNCIONES DE LA PIEL
La piel es un órgano que presenta una amplia variedad de fun-ciones, incluyendo la protectora, la termorreguladora, la sen-sitiva, la secretora, la inmunológica, la producción de vita-mina D y la excretora.
� Protección. Mediante su especial textura y composiciónprotege a los órganos internos de traumatismos mecáni-cos, físicos y químicos, a la vez que evita la pérdida deagua y electrólitos desde el interior. De traumas mecáni-cos protege mediante los estratos dérmico e hipodérmico,que actúan a modo de cojinetes, y además con el creci-miento-engrosamiento epitelial, protege de los físicos,como radiaciones ultravioleta, mediante la pigmentaciónepidérmica y absorción de estas radiaciones a distintosniveles, y de los químicos impidiendo su paso a través deun epitelio celular compacto. Este mismo estrato, y por lamisma razón, evita las pérdidas internas.
� Termorregulación. Mediante los fenómenos de vasodila-tación y vasoconstricción en los plexos vasculares cutá-neos se aumenta o reduce la temperatura de la piel y, ensituaciones de calor exterior extremo, la secreción sudoralecrina refresca la superficie cutánea.
� Sensación. Tacto, presión, vibración, temperatura, dolory prurito son captados por receptores sensoriales libresy/o corpúsculos sensoriales que los transmiten al cerebropor los cordones medulares dorsales.
� Secreción. Las glándulas de secreción pueden ser ecrinas(ec = fuera; crinia = secreción), como sucede con las sud-oríparas ecrinas, y en este mismo orden podríamos consi-derar la citocrínia melánica desde el melanocito; apocrina(apo = fuera; secreción de la parte superior de la célula),propia de las sudoríparas apocrinas y glándula mamaria; yholocrinas (secreción de la totalidad celular), representa-das por las glándulas sebáceas y el propio epitelio epidér-mico.
� Función inmunológica. Se ha demostrado que los que-ratinocitos intervienen de forma activa en el sistemainmune cutáneo o SALT (tejido linfoide asociado a la piel),tanto en las interacciones celulares con las células de Lan-gerhans y los linfocitos T epidermotrópicos como en laproducción de citocinas. Los histiocitos dérmicos tambiénintervienen en la función defensiva cutánea.
Los péptidos antimicrobianos son un grupo de péptidospresentes en la superficie epidérmica que actúan comoantibióticos naturales y participan en los procesos celula-res de la defensa inmune y la reparación tisular. Hay dosgrupos principales, las catelicidinas y las defensinas α y β.Normalmente se producen pequeñas cantidades de estospéptidos antimicrobianos en la epidermis, acumulándosealrededor de los folículos pilosos y las glándulas sudorípa-ras ecrinas, donde la función barrera está ausente o dismi.C; cuando existe una infección o una herida, los querati-nocitos incrementan rápidamente su producción, reclu-tando a los neutrófilos como parte de la respuesta infla-matoria aguda.
� Producción de vitamina D. La piel es el único órganodonde, en condiciones fisiológicas e inducida por la radia-ción UVB, se realiza la transformación completa del 7-dehi-drocolesterol en calcitriol (1,25-dihidroxivitamina D3). Elcalcitriol regula también el crecimiento y la diferenciaciónde los queratinocitos, por lo que se han introducido losanálogos de la vitamina D en la terapéutica de las derma-tosis hiperproliferativas.
� Excreción. Hay que comentar que a través de la piel seeliminan muy pocas sustancias aunque, en determinadassituaciones patológicas, al producirse grandes cantidadesde capa córnea, se pueden perder elementos constitutivosdel epitelio, especialmente azufre y proteínas. En la excre-ción cutánea también debemos considerar la perspiratioinsensibilis, que es la pérdida de agua diaria a través de lasuperficie cutánea, sin relación con la secreción ecrina, yque para un varón de 70 kg, que se correspondería conuna superficie de 1,80 m2, es de unos 350 ml.
EMBRIOLOGÍA DE LA PIEL
La epidermis, mucosas y anejos epidérmicos proceden delectodermo, mientras que dermis e hipodermis del meso-
1.A
nato
mía
y f
isio
logí
a de
la p
iel
5
Figura 4. Esquema de la capa córnea.
Corneocitos
Figura 5. Esquema de la celularidad de la epidermis.
Célula deLangerhans
Queratinocito
Melanocito Célula de Merkel
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 5
Figura 5. Esquema de la celularidad de la epidermis.
Tabla I. Funciones de la piel
Protección
Termorregulación
Sensación
Secreción
Función inmunológica
Producción de vitamina D
Excreción

6
Manual de derMatología. Sección I: Temas generales
de cordón sólido, que avanza hasta un nivel bastante profundo, donde las células se separan y determinan la luz glandular.
El otro germen epitelial es el de las glándulas sudoríparas ecrinas, que profundiza en la dermis desde el tercer o cuarto mes, diferenciándose paulatinamente glomérulo secretor y conducto excretor, que se canaliza hacia el octavo mes, adoptando en ese momento el aspecto que poseen en el adulto.
La formación de la uña comienza a las siete semanas con un cúmu-lo de células muy activas, con abundantes mitosis y daño celular seguido de necrosis en el dorso del tercio distal de los dedos. La apoptosis de esas células epidérmicas permite una invaginación epidérmica cuyo resultado final es la formación de un surco transversal, que se convertirá en el pliegue proximal de la uña. A las 12 semanas, están formados los pliegues ungueales proximales y laterales. El pliegue transversal distal, correspondiente al hipo-niquio, se encuentra completamente queratinizado a los tres meses y medio. La producción de la lámina ungueal empieza a partir de las células de la matriz, siendo su presencia visible desde el quinto mes de vida intrauterina.
Los melanocitos, que se encuentran entre las células de la capa basal epidérmica y en los gérmenes epiteliales primarios, proceden de la cresta neural y, vía mesenquima y estructuras nerviosas, se trasladan a su situa-ción cutánea (además del tracto uveal, leptomeninges y oído interno), donde ya se observan en el tercer mes.
Las células de Langerhans se comprueban en la capa espinosa desde la decimocuarta semana, mientras que las células de Merkel aparecen en piel y mucosas sobre la semana 16.
La dermis deriva del mesodermo, donde en el segundo mes, se obser-van muchas células mesenquimatosas primitivas, y en el tercero, fibroblas-tos y fibras colágenas. Las fibras elásticas surgen en el quinto mes.
Los adipocitos, células específicas de la hipodermis, también proceden de las células mesenquimales primitivas y pueden observarse a partir del cuarto mes.
La red vascular comienza a formarse a partir del tercer mes y la ner-viosa desde la quinta semana. Las estructuras vasculares cutáneas proce-dentes de la mesenquima comienzan a diferenciarse en cúmulos de an-gioblastos que se canalizan y constituyen los capilares sanguíneos. Desde ellos, proceden las porciones arterial y venosa.
ESTRUCTURA DE LA EPIDERMIS (TABLA II)
La epidermis es un estrato celular compacto que mide 120-200 micras, con diferencias regionales según función a desarrollar.
Sus células principales, representando más del 95% del total, son los queratinocitos, los cuales por sucesiva multiplicación y diferenciación, van ascendiendo desde la capa basal o germinativa hasta la superficie cutánea constituyéndose, durante este tránsito, las otras cuatro capas: espinosa, granulosa, lúcida y córnea (Figs. 2, 3 y 6).
Es costumbre referirse al cuerpo mucoso de Malpighio como el estrato que comprende la capa basal y la capa espinosa, y el estrato precórneo al constituido por las capas granulosa y lúcida. Pero en esta definición general de la epidermis no podemos olvidar que, en realidad, se trata de un sistema celular binario compuesto por queratinocitos y melanocitos, aunque también se encuentran los otros dos tipos celulares, las células de
Langerhans, que son células dendríticas inmunocompetentes, y las células de Merkel, que son células neurosecretoras.
En la superficie inferior de la epidermis se encuentran unas prolonga-ciones digitiformes denominadas «crestas interpapilares» que se introdu-cen entre las «papilas dérmicas» que son proyecciones verticales cónicas de la dermis. La unión de ambas estructuras permite un incremento de la superficie de contacto entre la epidermis y la dermis proporcionando una mayor adhesión entre estas dos capas de la piel.
QueratinocitosLos queratinocitos que se encuentran en la capa basal forman una sola
hilera celular y son de forma cuboidea, poseyendo un gran núcleo oval, donde destaca una gran cantidad de cromatina y uno o dos nucleolos esféricos, lejanos de la membrana celular, y un gran citoplasma con mito-condrias, complejos de Golgi, ribosomas, tonofibrillas y abundante retícu-lo endoplásmico liso y rugoso. Rodeando toda la célula, una membrana lipoproteíca. Están unidos por desmosomas, que también se observan en las células de las capas superiores, donde se insertan los tonofilamentos de queratina, mientras que en su base, que reposa sobre la membrana basal, solo se observan hemidesmosomas, que sirven de elementos de unión epi-dermodérmicos (Figs. 7 y 8).
Se multiplican por mitosis siguiendo un «ritmo circadiano», aumen-tando por la noche. Conforme ascienden, las células cambian de morfo-
Tabla II. Estructura de la epidermis
Estructura de la epidermis
Capa córnea Estrato córneoEpidermis metabólicamente muertaEstrato lúcido
Estrato precórneoCapa granulosa
Capa espinosa Cuerpo mucoso de Malphigio Epidermis metabólicamente viva
Capa basal
Secreción de un cuerpo lamelar
Células queratinizadas
Célula granulosa
Célula espinosa
Célula basal
Envoltura celular
Cuerpos lamelares
Cuerpos lamelares
REr
Mitocondria
Tonofilamentos
Ribosomas
Lámina basal
Gránulos de queratohialina
Tonofilamentos
Aparato de Golgi
Figura 6. Esquema de la diferenciación de los queratinocitos.

7
Capítulo 1. Anatomía y fisiología de la piel
logía. En la capa basal son elongadas o columnares, con diámetro mayor perpendicular a la superficie cutánea, mientras que al ascender, se van haciendo poligonales y paulatinamente se aplanan hasta constituir un mosaico. Debido al aspecto que toma, en mosaico, con las uniones con desmosomas en forma de espículas o «espinas», recibe desde antiguo el nombre de capa espinosa, que tiene de 3 a 10 hileras de células.
El proceso de queratinización progresiva que van a sufrir los queratinocitos hace que en su citoplasma vayan apareciendo diversas estructuras que corresponderán a los precursores de la queratina. Así, los queratinocitos muestran abundantes «tonofibrillas» perinucleares, que se van haciendo más evidentes en las capas más altas, pues son los elementos iniciales de la queratina. Las células más altas de esta capa espinosa contienen, además, unas estructuras ovales laminadas, conocidas como queratinosomas, gránulos lamelares o cuerpos de Selby-Odland, recubiertos de una membrana bicapa y que contienen láminas paralelas orientadas según el eje menor del gránulo.
Estos corpúsculos de estructura laminar contienen grandes cantidades de lípidos (fosfolípidos, glucolípidos y esteroles libres) y enzimas hidro-líticas, que van a intervenir en la función barrera de la capa córnea y van a destruir los desmosomas, para que las células cornificadas puedan descamarse con facilidad.
Al seguir ascendiendo, las células se hacen cada vez más aplanadas, pierden su núcleo y muestran numerosos gránulos de queratohialina, partícu-las amorfas no recubiertas de membrana, que constituyen la matriz que engloba las tonofibrillas en el proceso de queratinización.
Estos gránulos son los que justifican el nombre de capa granulosa, que está compuesta por entre una y cuatro hileras celulares. Los cuerpos de Selby-Odland van migrando a la periferia de las células hasta descargar su contenido al espacio intercelular; los lípidos polares son remodelados a lípidos neutros en el espacio extracelular, por medio de las enzimas hi-drolíticas vertidas, formando una importante barrera a la permeabilidad cutánea.
Solo en palmas y plantas puede observarse con nitidez la capa lúcida, cuyas células, aplanadas y desprovistas de núcleo, forman, junto a la gra-nulosa, el estrato precórneo o de transición.
Por último, la capa córnea esta constituida por 15 a 25 hileras de célu-las queratinizadas (en las palmas y plantas puede llegar a más de cien), carentes de núcleo y con escasos desmosomas, que llegan a desaparecer en las últimas capas permitiendo su descamación espontánea. Esta capa córnea presenta una acentuada hidrofilia, especialmente por su envol-tura lipídica externa compuesta de ceramidas, esteroles libres y ácidos grasos libres.
El tiempo total desde que una célula germinativa de la capa basal comienza a multiplicarse y ascender hasta eliminarse en la capa córnea es de 52 a 75 días, pudiendo esta tasa de epidermopoyesis variar según las regiones corporales.
QueratinizaciónLa queratina (del griego keras = cuerno) está compuesta por una fibro-
proteína formada por cadenas de aminoácidos en secuencia constante y dispuestos helicoidalmente, y otra proteína globulosa que envuelve a la primera. Hay dos tipos: una blanda, procedente de la epidermis, y otra dura, que se observa en pelos y uñas.
La familia de las queratinas humanas comprende 54 miembros, 28 del tipo I (ácidas) y 26 del tipo II (básicas). Las ácidas son de bajo peso mole-cular (40-56 kDa) y las básicas de alto peso molecular (52-67 kDa). De los 28 miembros del tipo I, 17 son queratinas epiteliales y 11 son queratinas del pelo y, de las 26 queratinas del tipo II, hay 20 epiteliales y 6 pilosas. Conforme se produce la migración de los queratinocitos en la epidermis, se sintetizan queratinas de peso molecular creciente, que se agrupan for-mando pares constituidos por una queratina ácida y otra queratina básica. Los genes que codifican las queratinas de los tipos I (ácidas) y tipo II (básicas) se localizan, respectivamente, en los cromosomas 17 y 12.
En cuanto a su bioquímica, podemos señalar que es una escleroproteína integrada por cadenas paralelas de polipéptidos con numerosos enlaces perpendiculares, entre los que destacan los disulfuro (–S–S–), que pro-ceden de la conversión de dos moléculas de cisteína (–SH–SH–) en una de cistina. Estos puentes impiden el deslizamiento de unas cadenas de polipéptidos sobre las otras, limitando la extensibilidad. Normalmente las cadenas de polipéptidos están plegadas, formando lo que conocemos como a-queratina, y que, cuando se distienden y llegan a una posición casi recta, se denomina b-queratina. Los puentes estarán a tensión en la b-que-
Figura 7. Desmosomas. Imagen con microscopia electrónica de transmisión.
Es costumbre referirse al cuerpo mucoso de Malpighiocomo el estrato que comprende la capa basal y la capa espi-nosa, y el estrato precórneo al constituido por las capas gra-nulosa y lúcida. Pero en esta definición general de la epider-mis no podemos olvidar que, en realidad, se trata de unsistema celular binario compuesto por queratinocitos y mela-nocitos, aunque también se encuentran los otros dos tiposcelulares, las células de Langerhans, que son células dendríti-cas inmunocompetentes, y las células de Merkel, que soncélulas neurosecretoras.
En la superficie inferior de la epidermis se encuentran unasprolongaciones digitiformes denominadas «crestas interpapila-res» que se introducen entre las «papilas dérmicas» que sonproyecciones verticales cónicas de la dermis. La unión deambas estructuras permite un incremento de la superficie decontacto entre la epidermis y la dermis proporcionando unamayor adhesión entre estas dos capas de la piel.
QueratinocitosLos queratinocitos que se encuentran en la capa basal formanuna sola hilera celular y son de forma cuboidea, poseyendoun gran núcleo oval, donde destaca una gran cantidad de cro-matina y uno o dos nucleolos esféricos, lejanos de la mem-brana celular, y un gran citoplasma con mitocondrias, com-plejos de Golgi, ribosomas, tonofibrillas y abundante retículoendoplásmico liso y rugoso. Rodeando toda la célula, unamembrana lipoproteíca. Están unidos por desmosomas, que
también se observan en las células de las capas superiores,donde se insertan los tonofilamentos de queratina, mientrasque en su base, que reposa sobre la membrana basal, sólo seobservan hemidesmosomas, que sirven de elementos deunión epidermodérmicos (Figs. 7 y 8).
Se multiplican por mitosis siguiendo un «ritmo circa-diano», aumentando por la noche. Conforme ascienden, lascélulas cambian de morfología. En la capa basal son elonga-das o columnares, con diámetro mayor perpendicular a lasuperficie cutánea, mientras que al ascender, se van haciendopoligonales y paulatinamente se aplanan hasta constituir unmosaico. Debido al aspecto que toma, en mosaico, con lasuniones con desmosomas en forma de espículas o «espinas»,recibe desde antiguo el nombre de capa espinosa, que tienede 3 a 10 hileras de células.
El proceso de queratinización progresiva que van asufrir los queratinocitos hace que en su citoplasma
1.A
nato
mía
y f
isio
logí
a de
la p
iel
7
Figura 6. Esquema de la diferenciación de los queratinocitos.
Célulasqueratinizadas
Célulagranulosa
Célulaespinosa
Célulabasal
Envoltura celular
Secreción de uncuerpo lamelarCuerposlamelaresGránulos dequeratohialina
TonofilamentosAparato de Golgi
Cuerposlamelares
REr
Mitocondria
Tonofilamentos
RibosomasLámina basal
Figura 7. Desmosomas. Imagen con microscopia electrónica detransmisión.
Figura 8. Esquema de las uniones de los desmosomas.
Unión ajustada
Cinturón de adhesión
Desmosoma
Unión de tipo hendidura
IntegrinaSelectina
CAM
Adhesiónfocal
Hemi-desmosoma
Integrina Proteoglucanode la membrana
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 7
Figura 8. Esquema de las uniones de los desmosomas.

8
Manual de derMatología. Sección I: Temas generales
ratina por lo que, pasado el efecto de la distensión, volverán la queratina a su posición primitiva. A una situación de b-queratina se llega cuando la epidermis o capa córnea es calentada con agua a 85 ºC.
Respecto a su morfología, los gránulos de queratohialina intracitoplasmá-ticos, que son electrón-densos y tienen una morfología irregular, son muy evidentes en el estrato granuloso. Hay dos tipos de gránulos, los PF y los L. Los gránulos PF, que son los más importantes, contienen la profilagrina que posteriormente se desfosforiliza para formar filagrina, que es una proteína rica en histidina y responsable de la agregación de los filamentos inter-medios de queratina. Los gránulos L contienen un segundo polipéptido denominado loricrina que contribuye a la formación de la barrera intraci-toplasmática estable e insoluble conocida como «envoltura cornificada».
Los queratinocitos de la capa córnea o corneocitos son anucleados, están totalmente queratinizados, con pérdida de las organelas citoplasmáticas, ri-bosomas y otros componentes por hidrólisis enzimática fundamentalmente de origen lisosómico, y se disponen agrupados en forma compacta, «en ces-ta de mimbre» o laminados. Los corneocitos son aplanados y están totalmente repletos de queratina, unidos entre ellos por desmosomas. Esta queratina pa-rece disponerse en bandas, más que al azar, siendo la filagrina la responsable de esta agregación de los filamentos de queratina (Figs. 9 y 10).
Además, los corneocitos están protegidos por una envoltura proteica interna insoluble o envoltura cornificada, que está situada en la superficie interna de la membrana plasmática, que se engruesa por el depósito de un material denso.
Está compuesta por distintas proteínas, como involucrina, loricrina, elafina, cornifina, queratolinina, envoplaquina y periplaquina, unidas por puentes disulfuro y del isodipéptido glutamil-lisil y catalizadas por transglutaminasas, que son enzimas calcio-dependientes.
Su fijación a los filamentos intermedios de queratina le proporciona gran estabilidad. La loricrina parece ser el principal componente de esta envoltura celular cornificada, que interviene en las funciones barrera y de permeabilidad de la capa córnea. Y como antes señalamos, hay otra envoltura externa lipídica, compuesta por hidroxi-ceramidas y ácidos grasos libres, colesterol y ésteres de colesterol, que se liga a la involucrina en la parte proteica de la envoltura, que interviene en la cohesión entre los corneocitos.
Los corneocitos que se rompen liberan grasas, hidratos de carbono, aminoácidos, ácido úrico, urea y minerales y, por tanto, presentan una secreción holocrina. Hay que destacar de modo especial las primeras, ya que la piel es un tejido sintetizador de lípidos. Esta «excreción lipídica epitelial» forma parte del manto cutáneo ácido lipídico y está constituida por ácidos grasos libres, que derivan de fosfolípidos y ceramidas, que provie-nen de glucoesfingolípidos y esfingomielina. También hay ácidos grasos libres de cadena larga, colesterol y colesterol-sulfato, que son muy hidro-fílicos y, por tanto, son responsables de la hidratación de la capa córnea, además de la cohesión celular. Los triglicéridos y ácidos grasos insatura-dos, tipo oleico y linoleico, detectados en la capa córnea parecen provenir de la grasa sebácea y de la contaminación ambiental.
Uniones intercelularesExisten varios tipos de uniones celulares entre los queratinocitos ad-
yacentes, que son las responsables de las interacciones mecánicas y bio-químicas existentes entre ellos. Podemos referirnos a los desmosomas, las uniones adherentes, las uniones tipo hendidura (gap) y las uniones ajustadas.
Los desmosomas son los principales complejos de adhesión en la epider-mis, anclando los filamentos intermedios de queratina a la membrana ce-lular y permitiendo a las células resistir a los traumatismos. Al microscopio óptico se observan como puentes o espinas, lo que da nombre a la capa y a los tumores derivados, carcinomas espinocelulares.
vayan apareciendo diversas estructuras que correspon-derán a los precursores de la queratina. Así, los quera-tinocitos muestran abundantes «tonofibrillas» perinu-cleares, que se van haciendo más evidentes en las capasmás altas, pues son los elementos iniciales de la quera-tina. Las células más altas de esta capa espinosa contie-nen, además, unas estructuras ovales laminadas, cono-cidas como queratinosomas, gránulos lamelares ocuerpos de Selby-Odland, recubiertos de una mem-brana bicapa y que contienen láminas paralelas orien-tadas según el eje menor del gránulo.
Estos corpúsculos de estructura laminar contienen gran-des cantidades de lípidos (fosfolípidos, glicolípidos y estero-les libres) y enzimas hidrolíticas, que van a intervenir en lafunción barrera de la capa córnea y van a destruir los desmo-somas, para que las células cornificadas puedan descamarsecon facilidad.
Al seguir ascendiendo, las células se hacen cada vez másaplanadas, pierden su núcleo y muestran numerosos gránu-los de queratohialina, partículas amorfas no recubiertas demembrana, que constituyen la matriz que engloba las tonofi-brillas en el proceso de queratinización. Estos gránulos sonlos que justifican el nombre de capa granulosa, que está com-puesta por entre una y cuatro hileras celulares. Los cuerposde Selby-Odland van migrando a la periferia de las célulashasta descargar su contenido al espacio intercelular; los lípi-dos polares son remodelados a lípidos neutros en el espacioextracelular, por medio de las enzimas hidrolíticas vertidas,formando una importante barrera a la permeabilidad cutánea.
Sólo en palmas y plantas puede observarse con nitidez lacapa lúcida, cuyas células, aplanadas y desprovistas denúcleo, forman, junto a la granulosa, el estrato precórneo ode transición.
Por último, la capa córnea esta constituida por 15 a 25hileras de células queratinizadas (en las palmas y plantaspuede llegar a más de cien), carentes de núcleo y con escasosdesmosoma, que llegan a desaparecer en las últimas capas
permitiendo su descamación espontánea. Esta capa córneapresenta una acentuada hidrofilia, especialmente por suenvoltura lipídica externa compuesta de ceramidas, esteroleslibres y ácidos grasos libres.
El tiempo total desde que una célula germinativa de lacapa basal comienza a multiplicarse y ascender hasta elimi-narse en la capa córnea es de 52 a 75 días, pudiendo esta tasade epidermopoyesis variar según las regiones corporales.
QueratinizaciónLa queratina (del griego keras = cuerno) está compuesta poruna fibroproteína formada por cadenas de aminoácidos ensecuencia constante y dispuestos helicoidalmente, y otra pro-teína globulosa que envuelve a la primera. Hay dos tipos: unablanda, procedente de la epidermis, y otra dura, que seobserva en pelos y uñas.
La familia de las queratinas humanas comprende 54miembros, 28 del tipo I (ácidas) y 26 del tipo II (bási-cas). Las ácidas son de bajo peso molecular (40-56 kDa)y las básicas de alto peso molecular (52-67 kDa). De los28 miembros del tipo I, 17 son queratinas epiteliales y11 son queratinas del pelo y, de las 26 queratinas deltipo II, hay 20 epiteliales y 6 pilosas. Conforme se pro-duce la migración de los queratinocitos en la epidermis,se sintetizan queratinas de peso molecular creciente, quese agrupan formando pares constituidos por una quera-tina ácida y otra queratina básica. Los genes que codifi-can las queratinas de los tipos I (ácidas) y tipo II (bási-cas) se localizan, respectivamente, en los cromosomas17 y 12.
En cuanto a su bioquímica, podemos señalar que es unaescleroproteína integrada por cadenas paralelas de polipépti-
Tem
as g
ener
ales
8
Figura 9. Esquema de la estructura proteica de la queratina.
MicrofilamentosFilamentos intermedios
Actinaα-tubulina
β-tubulina Queratina
Microtúbulos
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 8
Figura 9. Esquema de la estructura proteica de la queratina.
dos con numerosos enlaces perpendiculares, entre los quedestacan los disulfuro (–S–S–), que proceden de la conversiónde dos moléculas de cisteína (–SH–SH–) en una de cistina.Estos puentes impiden el deslizamiento de unas cadenas depolipéptidos sobre las otras, limitando la extensibilidad. Nor-malmente las cadenas de polipéptidos están plegadas, for-mando lo que conocemos como α-queratina, y que, cuandose distienden y llegan a una posición casi recta, se denominaβ-queratina. Los puentes estarán a tensión en la β-queratinapor lo que, pasado el efecto de la distensión, volverán la que-ratina a su posición primitiva. A una situación de beta-quera-tina se llega cuando la epidermis o capa córnea es calentadacon agua a 85 ºC.
Respecto a su morfología, los gránulos de queratohialinaintracitoplasmáticos, que son electrón-densos y tienen unamorfología irregular, son muy evidentes en el estrato granu-loso. Hay dos tipos de gránulos, los PF y los L. Los gránulosPF, que son los más importantes, contienen la profilagrinaque posteriormente se desfosforiliza para formar filagrina,que es una proteína rica en histidina y responsable de laagregación de los filamentos intermedios de queratina. Losgránulos L contienen un segundo polipéptido denominadoloricrina que contribuye a la formación de la barrera intraci-toplasmática estable e insoluble conocida como «envolturacornificada». Los queratinocitos de la capa córnea o corneo-citos son anucleados, están totalmente queratinizados, conpérdida de las organelas citoplasmáticas, ribosomas y otroscomponentes por hidrólisis enzimática fundamentalmentede origen lisosómico, y se disponen agrupados en formacompacta, «en cesta de mimbre» o laminados. Los corneoci-tos son aplanados y están totalmente repletos de queratina,unidos entre ellos por desmosomas. Esta queratina parecedisponerse en bandas, más que al azar, siendo la filagrina laresponsable de esta agregación de los filamentos de quera-tina (Figs. 9 y 10).
Además, los corneocitos están protegidos por una envol-tura proteica interna insoluble o envoltura cornificada, queestá situada en la superficie interna de la membrana plasmá-tica, que se engruesa por el depósito de un material denso.Está compuesta por distintas proteínas, como involucrina,loricrina, elafina, cornifina, queratolinina, envoplaquina yperiplaquina, unidas por puentes disulfuro y del isodipép-tido glutamil-lisil y catalizadas por transglutaminasas, queson enzimas calcio-dependientes. Su fijación a los filamentosintermedios de queratina le proporciona gran estabilidad. Laloricrina parece ser el principal componente de esta envol-tura celular cornificada, que interviene en las funcionesbarrera y de permeabilidad de la capa córnea. Y como antesseñalamos, hay otra envoltura externa lipídica, compuestapor hidroxi-ceramidas y ácidos grasos libres, colesterol yésteres de colesterol, que se liga a la involucrina en la parteproteica de la envoltura, que interviene en la cohesión entrelos corneocitos.
Los corneocitos que se rompen liberan grasas, hidratos decarbono, aminoácidos, ácido úrico, urea y minerales y, portanto, presentan una secreción holocrina. Hay que destacarde modo especial las primeras, ya que la piel es un tejido sin-tetizador de lípidos. Esta «excreción lipídica epitelial» formaparte del manto cutáneo ácido lipídico y está constituida por
ácidos grasos libres, que derivan de fosfolípidos y ceramidas,que provienen de glicoesfingolípidos y esfingomielina. Tam-bién hay ácidos grasos libres de cadena larga, colesterol ycolesterol-sulfato, que son muy hidrofílicos y, por tanto, sonresponsables de la hidratación de la capa córnea, además dela cohesión celular. Los triglicéridos y ácidos grasos insatura-dos, tipo oleico y linoleico, detectados en la capa córnea pare-cen provenir de la grasa sebácea y de la contaminaciónambiental.
Uniones intercelularesExisten varios tipos de uniones celulares entre los queratino-citos adyacentes, que son las responsables de las interaccio-nes mecánicas y bioquímicas existentes entre ellos. Podemosreferirnos a los desmosomas, las uniones adherentes, lasuniones tipo hendidura (gap) y las uniones ajustadas.
Los desmosomas son los principales complejos de adhe-sión en la epidermis, anclando los filamentos intermedios dequeratina a la membrana celular y permitiendo a las célulasresistir a los traumatismos. Al microscopio óptico se obser-van como puentes o espinas, lo que da nombre a la capa y alos tumores derivados, carcinomas espinocelulares.
1.A
nato
mía
y f
isio
logí
a de
la p
iel
9
Figura 10. Estructura de la queratina en las diferentes capas de laepidermis.
Paso V (SC)
Paso IV (T)
Paso III (SS, SG)
Paso II (SS, SG)
Paso I (SB, SS, SG)
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 9
Figura 10. Estructura de la queratina en las diferentes capas de la epidermis.

9
Capítulo 1. Anatomía y fisiología de la piel
Los desmosomas forman una unión simétrica, tienen un diámetro de 0,1-0,5 nm y están formados por unos componentes transmembrana y otros de la propia placa electrón-densa, con un espacio intercelular central (Figs. 7, 8 y 11). Los componentes transmembrana son unas glucoproteínas, familias de las cadherinas, formando asociaciones heterófilas de desmogleínas y de desmocolinas; hay tres desmogleínas (desmogleínas 1-3) y otras tres desmocolinas (desmocolinas 1-3), y los de la placa son otras tres proteínas (desmoplaquina, placoglobina y placofilina), que son las que fijan las cadherinas a los tonofilamentos de queratina, que se disponen perpendicularmente a ella (Fig. 12).
Las uniones adherentes son estructuras transmembrana electrón-densas compuestas por cadherina E, la cual forma interacciones calcio-dependien-tes con la cadherina E de la célula adyacente. Estas interacciones se co-nexio nan con la membrana plasmática por medio de una red de proteínas adhesivas que incluyen a-catenina, b-catenina y p120ctn.
Las uniones tipo hendidura (gap) son un grupo de canales intercelulares conocidos como conexones, que forman conexiones entre los citoplasmas de los queratinocitos adyacentes. Los conexones se originan en el aparato de Golgi mediante el ensamblaje de seis subunidades de conexinas, sintetizadas en el retículo endoplásmico, siendo después transportados a la membrana plasmática, donde se agregan con otros conexones y, en combinación con los agregados del queratinocito adyacente, forman la unión gap. La fun-ción de estas uniones tipo hendidura es la transferencia de moléculas de bajo peso molecular (< 1000 Da) y de intercambio de iones entre células vecinas.
Las uniones ajustadas están compuestas de moléculas transmembrana e intracelulares, como la ocludina y las claudinas. Tienen como función la de controlar la permeabilidad epidérmica y la de mantener la polaridad celular.
Epidermopoyesis
En la cinética celular de la epidermis, el concepto más importante es el tiempo de regeneración celular (turnover time), que define el tiempo medio de una población celular para reproducirse a sí misma; es decir, el inter-valo entre dos mitosis sucesivas de las células germinativas epidérmicas.
Aunque turnover time también se emplea para definir el paso de una célula desde el estrato germinativo a la capa córnea epitelial, nos parece más lógico emplear el término tiempo de tránsito para denominar este pe-riodo. Ya hemos indicado que es de 52 a 75 días; sin embargo, un epitelio psoriásico solo necesita de 8 a 10 días. El hecho de que el tamaño de la epidermis permanezca constante, hace pensar que la tasa de producción celular en el estrato germinativo está equilibrada con la pérdida celular en el estrato córneo.
La capa basal tiene una serie de controles estimuladores y/o inhibido-res que permiten, en determinadas circunstancias, producir más o menos células.
Factores estimuladores• Familia del factor de crecimiento epidérmico (EGF, por las siglas
en inglés de epidermal growth factor). El EGF humano es un polipéptido que estimula la proliferación y diferenciación celular en un amplio rango de tejidos. Se encuentra en las glándulas salivales, plaquetas y algunas glándulas del duodeno. El EGF incrementa la epidermopoyesis unién-dose a unos receptores específicos de la superficie celular (EGFr) que se detectan en la capa basal de la epidermis humana. Los queratinocitos humanos sintetizan cuatro factores de crecimiento de la familia EGF, que son TGF-a (transforming growth factor-a), anfirregulina, HB-EGF (he-parin-binding EGF) y epirregulina, los cuales estimulan su crecimiento y diferenciación por vía autocrina, uniéndose al EGFr. Aunque la gran mayoría del crecimiento autocrino de los queratinocitos está mediado por el EGFr, hay otras citocinas sintetizadas por estas células que tam-bién estimulan su crecimiento, como las interleucinas 1 y 6 (IL-1, IL-6) y el factor estimulador de colonias granulocito-macrófago (GM-CSF).
• Factor de crecimiento de queratinocitos (KGF, keratinocyte growth factor). Está producido por los fibroblastos dérmicos y también estimula el crecimiento queratinocítico de una forma paracrina, uniéndose al receptor KGFr.
Los desmosomas forman una unión simétrica, tienenun diámetro de 0,1-0,5 nm y están formados por unoscomponentes transmembrana y otros de la propia placaelectrón-densa, con un espacio intercelular central (Figs.7, 8 y 11). Los componentes transmembrana son unasglicoproteínas, familias de las cadherinas, formando aso-ciaciones heterófilas de desmogleínas y de desmocoli-nas; hay tres desmogleínas (desmogleínas 1-3) y otrastres desmocolinas (desmocolinas 1-3), y los de la placason otras tres proteínas (desmoplaquina, placoglobinay placofilina), que son las que fijan las cadherinas a lostonofilamentos de queratina, que se disponen perpen-dicularmente a ella (Fig. 12).
Las uniones adherentes son estructuras transmembrana elec-trón-densas compuestas por cadherina E, la cual forma interac-ciones calcio-dependientes con la cadherina E de la célula adya-cente. Estas interacciones se conexionan con la membranaplasmática por medio de una red de proteínas adhesivas queincluyen α-catenina, β-catenina y p120ctn.
Las uniones tipo hendidura (gap) son un grupo de canalesintercelulares conocidos como conexones, que forman cone-xiones entre los citoplasmas de los queratinocitos adyacen-tes. Los conexones se originan en el aparato de Golgimediante el ensamblaje de seis subunidades de conexinas,sintetizadas en el retículo endoplásmica, siendo despuéstransportados a la membrana plasmática, donde se agregancon otros conexones y, en combinación con los agregados delqueratinocito adyacente, forman la unión gap. La función deestas uniones tipo hendidura es la transferencia de moléculasde bajo peso molecular (< 1000 Da) y de intercambio de ionesentre células vecinas.
Las uniones ajustadas están compuestas de moléculastransmembrana e intracelulares, como la ocludina y las clau-dinas. Tienen como función la de controlar la permeabilidadepidérmica y la de mantener la polaridad celular.
EpidermopoyesisEn la cinética celular de la epidermis, el concepto más impor-tante es el tiempo de regeneración celular (turnover time),que define el tiempo medio de una población celular parareproducirse a sí misma; es decir, el intervalo entre dos mito-sis sucesivas de las células germinativas epidérmicas.
Aunque turnover time también se emplea para definir elpaso de una célula desde el estrato germinativo a la capa cór-nea epitelial, nos parece más lógico emplear el términotiempo de tránsito para denominar este periodo. Ya hemosindicado que es de 52 a 75 días; sin embargo, un epitelio pso-riásico sólo necesita de 8 a 10 días. El hecho de que el tamañode la epidermis permanezca constante, hace pensar que latasa de producción celular en el estrato germinativo está equi-librada con la pérdida celular en el estrato córneo.
La capa basal tiene una serie de controles estimuladoresy/o inhibidores que permiten, en determinadas circunstan-cias, producir más o menos células.
Factores estimuladores� Familia del Factor de crecimiento epidérmico (EGF,
«Epidermal Growth Factor»). El EGF humano es un polipép-tido que estimula la proliferación y diferenciación celularen un amplio rango de tejidos. Se encuentra en las glándu-las salivales, plaquetas y algunas glándulas del duodeno.El EGF incrementa la epidermopoyesis uniéndose a unos
Tem
as g
ener
ales
10
Figura 11. Diferentes proteínas que constituyen las uniones celula-res en la epidermis.
Unión ajustadaTransmembrana: ocludina, claudinaPlaca: ZO, cingulina, simplequina, 7H6Conectada al citoesqueleto de actina
Unión adherenteTransmembrana: cadherinas E y PPlaca: cateninas a y b, placoglobinaConectada al citoesqueleto de actina
DesmosomaTransmembrana: desmocolinas, desmogleinasPlaca: desmoplaquinas, placoglobinaConectados al citoesqueleto intermedio
Unión de tipo hendiduraTransmembrana: conexón (6 conexinas)
Membrana basal
Hemidesmosoma ¿Adhesión focal?
Figura 12. Componentes de los desmosomas.
Desmocolina
Desmogleína
Placoglobina
Placofilina
Desmoplaquina
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 10
Figura 11. Diferentes proteínas que constituyen las uniones celulares en la epidermis.
Los desmosomas forman una unión simétrica, tienenun diámetro de 0,1-0,5 nm y están formados por unoscomponentes transmembrana y otros de la propia placaelectrón-densa, con un espacio intercelular central (Figs.7, 8 y 11). Los componentes transmembrana son unasglicoproteínas, familias de las cadherinas, formando aso-ciaciones heterófilas de desmogleínas y de desmocoli-nas; hay tres desmogleínas (desmogleínas 1-3) y otrastres desmocolinas (desmocolinas 1-3), y los de la placason otras tres proteínas (desmoplaquina, placoglobinay placofilina), que son las que fijan las cadherinas a lostonofilamentos de queratina, que se disponen perpen-dicularmente a ella (Fig. 12).
Las uniones adherentes son estructuras transmembrana elec-trón-densas compuestas por cadherina E, la cual forma interac-ciones calcio-dependientes con la cadherina E de la célula adya-cente. Estas interacciones se conexionan con la membranaplasmática por medio de una red de proteínas adhesivas queincluyen α-catenina, β-catenina y p120ctn.
Las uniones tipo hendidura (gap) son un grupo de canalesintercelulares conocidos como conexones, que forman cone-xiones entre los citoplasmas de los queratinocitos adyacen-tes. Los conexones se originan en el aparato de Golgimediante el ensamblaje de seis subunidades de conexinas,sintetizadas en el retículo endoplásmica, siendo despuéstransportados a la membrana plasmática, donde se agregancon otros conexones y, en combinación con los agregados delqueratinocito adyacente, forman la unión gap. La función deestas uniones tipo hendidura es la transferencia de moléculasde bajo peso molecular (< 1000 Da) y de intercambio de ionesentre células vecinas.
Las uniones ajustadas están compuestas de moléculastransmembrana e intracelulares, como la ocludina y las clau-dinas. Tienen como función la de controlar la permeabilidadepidérmica y la de mantener la polaridad celular.
EpidermopoyesisEn la cinética celular de la epidermis, el concepto más impor-tante es el tiempo de regeneración celular (turnover time),que define el tiempo medio de una población celular parareproducirse a sí misma; es decir, el intervalo entre dos mito-sis sucesivas de las células germinativas epidérmicas.
Aunque turnover time también se emplea para definir elpaso de una célula desde el estrato germinativo a la capa cór-nea epitelial, nos parece más lógico emplear el términotiempo de tránsito para denominar este periodo. Ya hemosindicado que es de 52 a 75 días; sin embargo, un epitelio pso-riásico sólo necesita de 8 a 10 días. El hecho de que el tamañode la epidermis permanezca constante, hace pensar que latasa de producción celular en el estrato germinativo está equi-librada con la pérdida celular en el estrato córneo.
La capa basal tiene una serie de controles estimuladoresy/o inhibidores que permiten, en determinadas circunstan-cias, producir más o menos células.
Factores estimuladores� Familia del Factor de crecimiento epidérmico (EGF,
«Epidermal Growth Factor»). El EGF humano es un polipép-tido que estimula la proliferación y diferenciación celularen un amplio rango de tejidos. Se encuentra en las glándu-las salivales, plaquetas y algunas glándulas del duodeno.El EGF incrementa la epidermopoyesis uniéndose a unos
Tem
as g
ener
ales
10
Figura 11. Diferentes proteínas que constituyen las uniones celula-res en la epidermis.
Unión ajustadaTransmembrana: ocludina, claudinaPlaca: ZO, cingulina, simplequina, 7H6Conectada al citoesqueleto de actina
Unión adherenteTransmembrana: cadherinas E y PPlaca: cateninas a y b, placoglobinaConectada al citoesqueleto de actina
DesmosomaTransmembrana: desmocolinas, desmogleinasPlaca: desmoplaquinas, placoglobinaConectados al citoesqueleto intermedio
Unión de tipo hendiduraTransmembrana: conexón (6 conexinas)
Membrana basal
Hemidesmosoma ¿Adhesión focal?
Figura 12. Componentes de los desmosomas.
Desmocolina
Desmogleína
Placoglobina
Placofilina
Desmoplaquina
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 10
Figura 12. Componentes de los desmosomas.

10
Manual de derMatología. Sección I: Temas generales
• Prostaglandinas. Algunos metabolitos del ácido araquidónico, como las prostaglandinas, especialmente PGE2, elevarían el AMPc celular, lo que parece reafirmarse por el hecho de que PGF2 y PGE2 estén aumentadas en la epidermis psoriásica. También se ha demos-trado que HPETE y leucotrienos, productos lipooxigenasa del ácido araquidónico, están aumentados en la psoriasis e inducen proliferación celular epidérmica in vitro.
• Poliaminas. Tanto putrescina como espermina y espermidina, contribuyen a la hiperepidermopoyesis, además de incrementar la neovasculariza-ción y la síntesis de proteínas de matriz extracelular en los mecanismos de cicatrización de heridas. Por supuesto, se encuentran elevadas en la epidermis psoriásica.
Factores inhibidores• Familia del factor transformador de crecimiento beta (TGF-b,
transforming growth factor-b): comprende dos polipéptidos, b-1 (TGF-b1) y b-2 (TGF-b2), sinterizados por los queratinocitos, y que son los fac-tores más importantes que inhiben su crecimiento, aunque estimulan los fibroblastos.
• Interferón alfa (IFN-a) e interferón gamma (IFN-g): tienen efectos citostáticos sobre los queratinocitos.
• Factor de necrosis tumoral alfa (TNF-a): producido por los que-ratinocitos, tiene un efecto citostático reversible sobre ellos mientras que estimula la proliferación de fibroblastos y la síntesis de citocinas.
• Las hormonas esteroideas, que pueden ser propias del individuo o venir del exterior, también controlarían parcialmente la epidermo-poyesis. Los andrógenos estimulan las mitosis epidérmicas y los gluco-corticoides las inhiben.
Unión o juntura dermoepidérmicaLa unión dermoepidérmica comprende un complejo multiproteínico
continuo, que forma un entramado que sustenta y fija los queratinocitos epidérmicos a la dermis subyacente y que regula el intercambio metabó-lico entre las dos capas.
Supone la zona de anclaje entre epidermis y dermis y está constituida por cuatro elementos fundamentalmente: a) tonofilamentos de queratina de las células basales, b) hemidesmosomas, c) membrana basal, y d) lámina fibroreticular.
Los hemidesmosomas se encuentran en la membrana celular y no están influenciados por la edad, sexo o región corporal. También se denominan máculas adherentes y se observan al microscopio óptico como un engrosa-miento («nódulo de Bizzozero»). Desde la porción intracitoplásmatica de los hemidesmosomas, o placa interna, parten hacia el citoplasma abun-dantes filamentos intermedios de queratina de los pares de queratinas K5 y K14, dispuestos perpendicularmente. La placa interna contiene el antí-geno mayor del penfigoide ampolloso de 230 Kd (BP230 o BPAg1) y la plectina. Exteriormente hay una placa externa o placa densa sub-basal, que es electrón-densa y que está separada de la membrana celular por una pequeña porción electrón-lúcida. La integrina a6b4 y el antígeno me-nor del penfigoide ampolloso de 180 Kd (BP180, BPAg2 o colágeno tipo XVII) son los componentes transmembrana de los hemidesmosomas. La integrina a6b4 es el receptor de las lamininas.
La membrana basal, auténtico límite entre epidermis y dermis, cuyo prin-cipal constituyente es el colágeno tipo IV. Consta de tres porciones: lámina
lúcida, lámina densa y lámina fibroreticular. La primera, conocida como espacio intermembranoso o lámina lúcida, por ser electrón-lúcida, es paralela a la membrana citoplasmática de las células basales, mide 30 nm (10-50 nm) y en ella se encuentran lamininas, fibronectina y colágeno tipo V, que facilitan su unión a la membrana plasmática. Esta lámina lúcida está atravesada por unos delgados filamentos de anclaje, orientados perpen-dicularmente, que unen la membrana plasmática a la lámina densa, y que constan de laminina 5 (predominantemente), laminina 6 y LAD1 (antígeno IgA lineal). La lámina densa es una capa electrón-densa que mide unos 35 nm (30-60 nm) y está compuesta por colágeno tipo IV, nidógeno, heparán-sulfato, condroitín-sulfato y perlecan. El perlecan es el principal proteoglucano de la membrana basal y en ella también se han detectado otras lamininas como la 1, 2 y 10.
La lámina fibroreticular, zona de debajo de la lámina densa, se conoce también como «sublámina densa» o «región de las fibras de anclaje» y consta de fibrillas de anclaje, microfibrillas elásticas y placas de anclaje. Sus principales componentes son las fibrillas de anclaje, compuestas de colá-geno tipo VII, que son estructuras curvadas que se insertan en la lámina densa y se extienden hasta la porción más superior de la dermis inser-tándose en unos cuerpos amorfos dérmicos denominados placas de anclaje, de colágeno tipo IV. Estas fibrillas de anclaje también pueden curvarse teniendo una segunda inserción en la lámina densa. Además, existen mu-chas microfibrillas elásticas (fibras oxitalánicas) que se extienden dentro de la dermis y pueden entremezclarse con el sistema microfibrilar dérmico. La principal microfibrilla elástica es la fibrilina.
MelanocitosLos melanocitos son células dendríticas que proceden de los melanoblas-
tos, precursores derivados de la cresta neural, encargadas de sintetizar las melaninas que dan color a la piel, pelos y ojos. Se encuentran entre los queratinocitos de la capa basal, de los que se distinguen por carecer de desmosomas y tonofilamentos, y con los que interaccionan vía cadherinas. Tienen prolongaciones citoplasmáticas dendríticas y un amplio citoplas-ma claro (por condensación perinuclear), por lo que también se las conoce como «células claras de Masson» (Figs. 13 y 14).
La migración de los melanoblastos y su diferenciación está influenciada por una serie de moléculas de señal producidas por las células vecinas. Entre ellas podemos referirnos a wnt, endotelina-3 (ET-3), las proteínas morfogé-nicas óseas (BMPs), el factor de crecimiento de los hepatocitos (HGF) y el
Figura 13. Imagen histológica de los melanocitos en la capa basal de la epidermis.
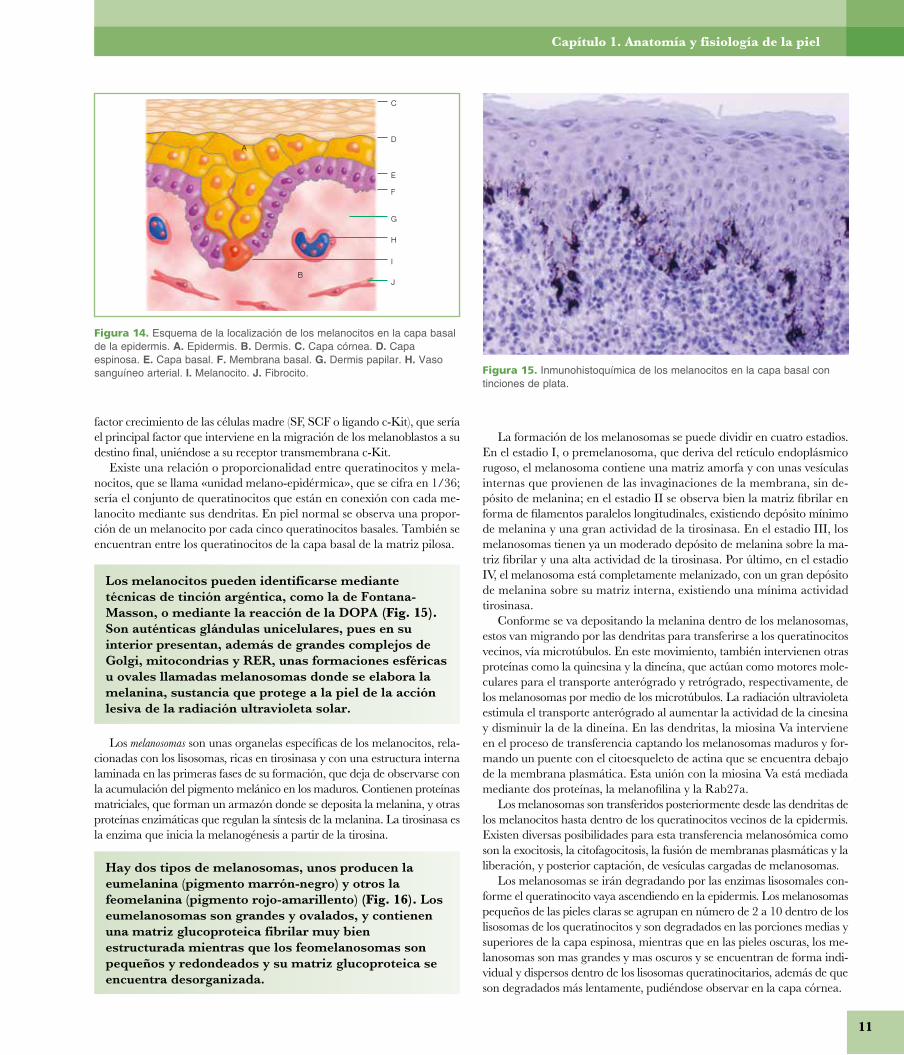
11
Capítulo 1. Anatomía y fisiología de la piel
factor crecimiento de las células madre (SF, SCF o ligando c-Kit), que sería el principal factor que interviene en la migración de los melanoblastos a su destino final, uniéndose a su receptor transmembrana c-Kit.
Existe una relación o proporcionalidad entre queratinocitos y mela-nocitos, que se llama «unidad melano-epidérmica», que se cifra en 1/36; sería el conjunto de queratinocitos que están en conexión con cada me-lanocito mediante sus dendritas. En piel normal se observa una propor-ción de un melanocito por cada cinco queratinocitos basales. También se encuentran entre los queratinocitos de la capa basal de la matriz pilosa.
Los melanocitos pueden identificarse mediante técnicas de tinción argéntica, como la de Fontana-Masson, o mediante la reacción de la DOPA (Fig. 15). Son auténticas glándulas unicelulares, pues en su interior presentan, además de grandes complejos de Golgi, mitocondrias y RER, unas formaciones esféricas u ovales llamadas melanosomas donde se elabora la melanina, sustancia que protege a la piel de la acción lesiva de la radiación ultravioleta solar.
Los melanosomas son unas organelas específicas de los melanocitos, rela-cionadas con los lisosomas, ricas en tirosinasa y con una estructura interna laminada en las primeras fases de su formación, que deja de observarse con la acumulación del pigmento melánico en los maduros. Contienen proteínas matriciales, que forman un armazón donde se deposita la melanina, y otras proteínas enzimáticas que regulan la síntesis de la melanina. La tirosinasa es la enzima que inicia la melanogénesis a partir de la tirosina.
Hay dos tipos de melanosomas, unos producen la eumelanina (pigmento marrón-negro) y otros la feomelanina (pigmento rojo-amarillento) (Fig. 16). Los eumelanosomas son grandes y ovalados, y contienen una matriz glucoproteica fibrilar muy bien estructurada mientras que los feomelanosomas son pequeños y redondeados y su matriz glucoproteica se encuentra desorganizada.
La formación de los melanosomas se puede dividir en cuatro estadios. En el estadio I, o premelanosoma, que deriva del retículo endoplásmico rugoso, el melanosoma contiene una matriz amorfa y con unas vesículas internas que provienen de las invaginaciones de la membrana, sin de-pósito de melanina; en el estadio II se observa bien la matriz fibrilar en forma de filamentos paralelos longitudinales, existiendo depósito mínimo de melanina y una gran actividad de la tirosinasa. En el estadio III, los melanosomas tienen ya un moderado depósito de melanina sobre la ma-triz fibrilar y una alta actividad de la tirosinasa. Por último, en el estadio IV, el melanosoma está completamente melanizado, con un gran depósito de melanina sobre su matriz interna, existiendo una mínima actividad tirosinasa.
Conforme se va depositando la melanina dentro de los melanosomas, estos van migrando por las dendritas para transferirse a los queratinocitos vecinos, vía microtúbulos. En este movimiento, también intervienen otras proteínas como la quinesina y la dineína, que actúan como motores mole-culares para el transporte anterógrado y retrógrado, respectivamente, de los melanosomas por medio de los microtúbulos. La radiación ultravioleta estimula el transporte anterógrado al aumentar la actividad de la cinesina y disminuir la de la dineína. En las dendritas, la miosina Va interviene en el proceso de transferencia captando los melanosomas maduros y for-mando un puente con el citoesqueleto de actina que se encuentra debajo de la membrana plasmática. Esta unión con la miosina Va está mediada mediante dos proteínas, la melanofilina y la Rab27a.
Los melanosomas son transferidos posteriormente desde las dendritas de los melanocitos hasta dentro de los queratinocitos vecinos de la epidermis. Existen diversas posibilidades para esta transferencia melanosómica como son la exocitosis, la citofagocitosis, la fusión de membranas plasmáticas y la liberación, y posterior captación, de vesículas cargadas de melanosomas.
Los melanosomas se irán degradando por las enzimas lisosomales con-forme el queratinocito vaya ascendiendo en la epidermis. Los melanosomas pequeños de las pieles claras se agrupan en número de 2 a 10 dentro de los lisosomas de los queratinocitos y son degradados en las porciones medias y superiores de la capa espinosa, mientras que en las pieles oscuras, los me-lanosomas son mas grandes y mas oscuros y se encuentran de forma indi-vidual y dispersos dentro de los lisosomas queratinocitarios, además de que son degradados más lentamente, pudiéndose observar en la capa córnea.
Melanocitos
Los melanocitos son células dendríticas que proceden de losmelanoblastos, precursores derivados de la cresta neural,encargadas de sintetizar las melaninas que dan color a la piel,pelos y ojos. Se encuentran entre los queratinocitos de la capabasal, de los que se distinguen por carecer de desmosomas ytonofilamentos, y con los que interaccionan vía cadherinas.Tienen prolongaciones citoplasmáticas dendríticas y unamplio citoplasma claro (por condensación perinuclear), porlo que también se las conoce como «células claras de Mas-son» (Figs. 13 y 14).
La migración de los melanoblastos y su diferenciación estáinfluenciada por una serie de moléculas de señal producidaspor las células vecinas. Entre ellas podemos referirnos a wnt,endotelina-3 (ET-3), las proteínas morfogénicas óseas (BMPs),el factor de crecimiento de los hepatocitos (HGF) y el factorcrecimiento de las células madre (SF, SCF o ligando c-Kit), quesería el principal factor que interviene en la migración de losmelanoblastos a su destino final, uniéndose a su receptortransmembrana c-Kit.
Existe una relación o proporcionalidad entre queratinoci-tos y melanocitos, que se llama «unidad melano-epidérmica»,que se cifra en 1/36; sería el conjunto de queratinocitos queestán en conexión con cada melanocito mediante sus dendri-tas. En piel normal se observa una proporción de un melano-cito por cada cinco queratinocitos basales. También seencuentran entre los queratinocitos de la capa basal de lamatriz pilosa.
Los melanocitos pueden identificarse mediante técni-cas de tinción argéntica, como la de Fontana-Masson,o mediante la reacción de la DOPA (Fig. 15). Sonauténticas glándulas unicelulares, pues en su interiorpresentan, además de grandes complejos de Golgi,mitocondrias y RER, unas formaciones esféricas u ova-les llamadas melanosomas donde se elabora la mela-nina, sustancia que protege a la piel de la acción lesivade la radiación ultravioleta solar.
Los melanosomas son unas organelas específicas de losmelanocitos, relacionadas con los lisosomas, ricas en tirosi-nasa y con una estructura interna laminada en las primerasfases de su formación, que deja de observarse con la acumu-lación del pigmento melánico en los maduros. Contienen pro-teínas matriciales, que forman un armazón donde se depositala melanina, y otras proteínas enzimáticas que regulan la sín-tesis de la melanina. La tirosinasa es la enzima que inicia lamelanogénesis a partir de la tirosina.
Hay dos tipos de melanosomas, unos producen la eumela-nina (pigmento marrón-negro) y otros la feomelanina (pig-mento rojo-amarillento) (Fig. 16). Los eumelanosomas songrandes y ovalados, y contienen una matriz glicoproteicafibrilar muy bien estructurada mientras que los feomelanoso-
mas son pequeños y redondeados y su matriz glicoproteicase encuentra desorganizada.
La formación de los melanosomas se puede dividir en cuatroestadios. En el estadio I, o premelanosoma, que deriva del retí-culo endoplásmico rugoso, el melanosoma contiene una matrizamorfa y con unas vesículas internas que provienen de las inva-ginaciones de la membrana, sin depósito de melanina; en elestadio II se observa bien la matriz fibrilar en forma de filamen-tos paralelos longitudinales, existiendo depósito mínimo demelanina y una gran actividad de la tirosinasa. En el estadio III,los melanosomas tienen ya un moderado depósito de melaninasobre la matriz fibrilar y una alta actividad de la tirosinasa. Porúltimo, en el estadio IV, el melanosoma está completamente
Tem
as g
ener
ales
12
Figura 13. Imagen histológica de los melanocitos en la capa basalde la epidermis.
Figura 14. Esquema de la localización de los melanocitos en la capabasal de la epidermis. A, epidermis; B, dermis; C, caja córnea; D,capa espinosa; E, capa basal; F, membrana basal; G, dermis papilar;H, vaso sanguíneo arterial; I, melanocito; J, fibrocito.
C
D
E
F
G
H
I
B
A
J
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 12
Figura 14. Esquema de la localización de los melanocitos en la capa basal de la epidermis. A. Epidermis. B. Dermis. C. Capa córnea. D. Capa espinosa. E. Capa basal. F. Membrana basal. G. Dermis papilar. H. Vaso sanguíneo arterial. I. Melanocito. J. Fibrocito. Figura 15. Inmunohistoquímica de los melanocitos en la capa basal con
tinciones de plata.
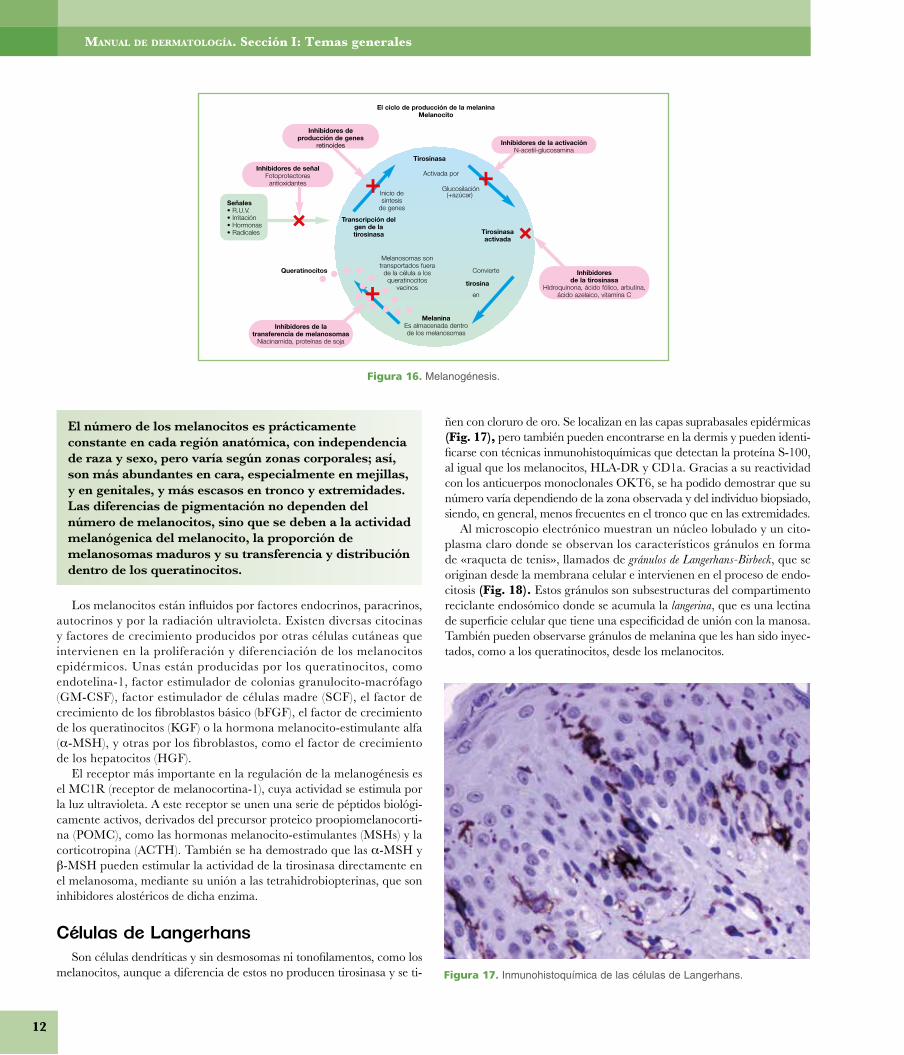
12
Manual de derMatología. Sección I: Temas generales
El número de los melanocitos es prácticamente constante en cada región anatómica, con independencia de raza y sexo, pero varía según zonas corporales; así, son más abundantes en cara, especialmente en mejillas, y en genitales, y más escasos en tronco y extremidades. Las diferencias de pigmentación no dependen del número de melanocitos, sino que se deben a la actividad melanógenica del melanocito, la proporción de melanosomas maduros y su transferencia y distribución dentro de los queratinocitos.
Los melanocitos están influidos por factores endocrinos, paracrinos, autocrinos y por la radiación ultravioleta. Existen diversas citocinas y factores de crecimiento producidos por otras células cutáneas que intervienen en la proliferación y diferenciación de los melanocitos epidérmicos. Unas están producidas por los queratinocitos, como endotelina-1, factor estimulador de colonias granulocito-macrófago (GM-CSF), factor estimulador de células madre (SCF), el factor de crecimiento de los fibroblastos básico (bFGF), el factor de crecimiento de los queratinocitos (KGF) o la hormona melanocito-estimulante alfa (a-MSH), y otras por los fibroblastos, como el factor de crecimiento de los hepatocitos (HGF).
El receptor más importante en la regulación de la melanogénesis es el MC1R (receptor de melanocortina-1), cuya actividad se estimula por la luz ultravioleta. A este receptor se unen una serie de péptidos biológi-camente activos, derivados del precursor proteico proopiomelanocorti-na (POMC), como las hormonas melanocito-estimulantes (MSHs) y la corticotropina (ACTH). También se ha demostrado que las a-MSH y b-MSH pueden estimular la actividad de la tirosinasa directamente en el melanosoma, mediante su unión a las tetrahidrobiopterinas, que son inhibidores alostéricos de dicha enzima.
Células de LangerhansSon células dendríticas y sin desmosomas ni tonofilamentos, como los
melanocitos, aunque a diferencia de estos no producen tirosinasa y se ti-
ñen con cloruro de oro. Se localizan en las capas suprabasales epidérmicas (Fig. 17), pero también pueden encontrarse en la dermis y pueden identi-ficarse con técnicas inmunohistoquímicas que detectan la proteína S-100, al igual que los melanocitos, HLA-DR y CD1a. Gracias a su reactividad con los anticuerpos monoclonales OKT6, se ha podido demostrar que su número varía dependiendo de la zona observada y del individuo biopsiado, siendo, en general, menos frecuentes en el tronco que en las extremidades.
Al microscopio electrónico muestran un núcleo lobulado y un cito-plasma claro donde se observan los característicos gránulos en forma de «raqueta de tenis», llamados de gránulos de Langerhans-Birbeck, que se originan desde la membrana celular e intervienen en el proceso de endo-citosis (Fig. 18). Estos gránulos son subsestructuras del compartimento reciclante endosómico donde se acumula la langerina, que es una lectina de superficie celular que tiene una especificidad de unión con la manosa. También pueden observarse gránulos de melanina que les han sido inyec-tados, como a los queratinocitos, desde los melanocitos.
Figura 17. Inmunohistoquímica de las células de Langerhans.
melanizado, con un gran depósito de melanina sobre su matrizinterna, existiendo una mínima actividad tirosinasa.
Conforme se va depositando la melanina dentro de losmelanosomas, éstos van migrando por las dendritas para
transferirse a los queratinocitos vecinos, vía microtúbulos. Eneste movimiento, también intervienen otras proteínas comola quinesina y la dineína, que actúan como motores molecula-res para el transporte anterógrado y retrógrado, respectiva-mente, de los melanosomas por medio de los microtúbulos.La radiación ultravioleta estimula el transporte anterógrado alaumentar la actividad de la quinesina y disminuir la de ladineína. En las dendritas, la miosina Va interviene en el pro-ceso de transferencia captando los melanosomas maduros yformando un puente con el citoesqueleto de actina que seencuentra debajo de la membrana plasmática. Esta unión conla miosina Va está mediada mediante dos proteínas, la mela-nofilina y la Rab27a.
Los melanosomas son transferidos posteriormente desdelas dendritas de los melanocitos hasta dentro de los querati-nocitos vecinos de la epidermis. Existen diversas posibilida-des para esta transferencia melanosómica como son la exoci-tosis, la citofagocitosis, la fusión de membranas plasmáticasy la liberación, y posterior captación, de vesículas cargadasde melanosomas.
Los melanosomas se irán degradando por las enzimaslisosomales conforme el queratinocito vaya ascendiendo enla epidermis. Los melanosomas pequeños de las pieles cla-ras se agrupan en número de 2 a 10 dentro de los lisosomas
1.A
nato
mía
y f
isio
logí
a de
la p
iel
13
Figura 15. Inmunohistoquímica de los melanocitos en la capa basalcon tinciones de plata.
Figura 16. Melanogénesis.
El ciclo de producción de la melaninaMelanocito
Inhibidores deproducción de genes
retinoides
Inhibidores de señalFotoprotectores
antioxidantes
Señales• R.U.V.• Irritación• Hormonas• Radicales
Inhibidores de latransferencia de melanosomas
Niacinamida, proteínas de soja
Inhibidores de la tirosinasa
Hidroquinona, ácido fólico, arbutina,ácido azelaico, vitamina C
Inhibidores de la activaciónN-acetil-glucosamina
MelaninaEs almacenada dentrode los melanosomas
Melanosomas sontransportados fuera
de la célula a losqueratinocitos
vecinosen
tirosina
Convierte
Tirosinasaactivada
Glucosilación(+azúcar)
Transcripción delgen de latirosinasa
Inicio desíntesis
de genes
Tirosinasa
Activada por
Queratinocitos
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 13
Figura 16. Melanogénesis.

13
Capítulo 1. Anatomía y fisiología de la piel
Hoy se consideran elementos mesenquimales que derivan de la médula ósea con funciones similares a los macrófagos (sistema mononuclear-fagocítico), te nien do a los monocitos sanguíneos como paso intermedio, y que actuarían como presentadores de antígenos y activadoras de células T en las reacciones de hipersensibilidad demorada, migrando desde la epidermis a los ganglios linfáticos regionales (Fig. 19).
Los antígenos capturados son reconocidos por las células de Lan-gerhans al interiorizarlos y procesarlos. Entonces, la langerina rápida-mente se interioriza desde la superficie celular hasta los gránulos de Birbeck.
Presentan múltiples receptores antigénicos que son capaces de respon-der a una gran variedad de antígenos, como alérgenos de contacto, mi-croorganismos y antígenos tumorales. Los queratinocitos producen una serie de citocinas, como GM-CSF y TNF-a, que influyen en la diferen-ciación de las células de Langerhans.
Células de MerkelSon células que se encuentran en la capa basal de la epidermis, espe-
cialmente de los pulpejos de los dedos, en las mucosas oral y labial y en la vaina epitelial externa de los folículos pilosos (Fig. 20). Presentan carac-terísticas neuroendocrinas y epiteliales. Lo más probable es que tengan un origen neural, derivando de la cresta neural.
Muestran un núcleo multilobulado y un gran citoplasma claro, con numerosos gránulos esféricos electrón-densos que contienen diferentes neuropéptidos, como el péptido intestinal vasoactivo (VIP) o la sinaptofisi-na. En su membrana plasmática pueden observarse algunos desmosomas, por donde se unen a los queratinocitos adyacentes. También presentan filamentos intermedios de queratina, siendo muy específica de ellas la citoqueratina 20, por lo que algunos autores piensan que se originarían en
la epidermis. No obstante, esta citoqueratina es característica de epitelios simples no estratificados.
Están en relación con nervios mielínicos que, al llegar cerca de la epi-dermis, pierden su vaina mielínica, continuando como axones amielínicos que terminan realizando una típica sinapsis química (Figs. 21 y 22).
Todavía existen muchas dudas sobre sus funciones; no obstante, pa-rece claro que actuarían como mecanorreceptores de adaptación lenta, estando en íntimo contacto con las fibras nerviosas amielínicas forman-do los «discos táctiles de la epidermis». Sin embargo, los numerosos péptidos que sintetizan y liberan hacen pensar que se comunican con otras células diferentes a las neuronas y que pueden intervenir en la fisiología cutánea.
Hoy se consideran elementos mesenquimales que deri-van de la médula ósea con funciones similares a losmacrófagos (sistema mononuclear-fagocítico), te nien -do a los monocitos sanguíneos como paso intermedio,y que actuarían como presentadores de antígenos yactivadoras de células T en las reacciones de hipersen-sibilidad demorada, migrando desde la epidermis a losganglios linfáticos regionales (Fig. 19). Los antígenoscapturados son reconocidos por las células de Lan ger-hans al interiorizarlos y procesarlos. Entonces, la lan-gerina rápidamente se interioriza desde la superficiecelular hasta los gránulos de Birbeck.
Presentan múltiples receptores antigénicos que son capa-ces de responder a una gran variedad de antígenos, comoalérgenos de contacto, microorganismos y antígenos tumora-les. Los queratinocitos producen una serie de citocinas, comoGM-CSF y TNF-α, que influyen en la diferenciación de las célu-las de Langerhans.
Células de Merkel
Son células que se encuentran en la capa basal de la epider-mis, especialmente de los pulpejos de los dedos, en las muco-sas oral y labial y en la vaina epitelial externa de los folículospilosos (Fig. 20). Presentan características neuroendocrinas yepiteliales. Lo más probable es que tengan un origen neural,derivando de la cresta neural.
Muestran un núcleo multilobulado y un gran citoplasmaclaro, con numerosos gránulos esféricos electrón-densos quecontienen diferentes neuropéptidos, como el péptido intesti-nal vasoactivo (VIP) o la sinaptofisina. En su membrana plas-mática pueden observarse algunos desmosomas, por dondese unen a los queratinocitos adyacentes. También presentanfilamentos intermedios de queratina, siendo muy específica deellas la citoqueratina 20, por lo que algunos autores piensanque se originarían en la epidermis. No obstante, esta citoque-ratina es característica de epitelios simples no estratificados.
Están en relación con nervios mielínicos que, al llegarcerca de la epidermis, pierden su vaina mielínica, continuandocomo axones amielínicos que terminan realizando una típicasinapsis química (Figs. 21 y 22).
Todavía existen muchas dudas sobre sus funciones; noobstante, parece claro que actuarían como mecanorrecepto-
1.A
nato
mía
y f
isio
logí
a de
la p
iel
15
Metabolito farmacológicoMetabolito
Citotoxicidad
Metabolito
QC
Bioactivación
Hapteno
Captación de antígeno
CL inmadura
MHC
HaptenoEpidermis
CL madura
Activación de la CL
DermisMigración de la CL alos ganglios linfáticos
CD80
CD28
Presentación del antígeno
Linfocito T
TCR
Proliferación del linfocito T
Migración de los linfocitos Ta los lugares de inflamación
Figura 19. Mecanismo de actuación de las células de Langerhans.
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 15
Figura 19. Mecanismo de actuación de las células de Langerhans.Figura 18. Corpúsculos de Birbek.
res de adaptación lenta, estando en íntimo contacto con lasfibras nerviosas amielínicas formando los «discos táctiles dela epidermis». Sin embargo, los numerosos péptidos que sin-tetizan y liberan hacen pensar que se comunican con otrascélulas diferentes a las neuronas y que pueden intervenir enla fisiología cutánea.
Dermis
Es la capa que sirve de sostén a la epidermis, a la que aportasus nutrientes, y que contiene los anejos y las estructuras vas-culonerviosas. Es una fascia superficial de tejido conjuntivocompuesta por células, fibras y sustancia fundamental, quetiene diferente textura según zonas del cuerpo y edad de lapersona, variando su grosor desde 1 mm en los párpadoshasta los 5 mm en la espalda. Es de 15 a 40 veces más gruesaque la epidermis. Al microscopio óptico muestra claramentedos partes: una superior, o dermis papilar, y otra inferior, odermis reticular.
a) Dermis papilar. Se llama así porque está com-puesta casi exclusivamente por la zona de las papi-las dérmicas ya que llega hasta donde las crestasinterpapilares epidérmicas penetran en la dermis.Tiene haces de colágeno y fibras elásticas, más del-gados que en la dermis reticular y orientados verti-calmente al epitelio, y, sobre todo, abundante sus-tancia fundamental, donde las finas fibras decolágeno forman una red. El diámetro de las fibrasde colágeno es de unos 50 nm y forman haces deunas 0,3-3 micras. En su interior está el plexo vas-cu lar superficial.
b) Dermis reticular. Es mucho más gruesa que la papi-lar, ocho o nueve veces más. Las bandas colágenasson más abundantes y más gruesas, de unos 63 nmde diámetro, se disponen en gruesos haces horizon-tales al epitelio, de unas 10-40 micras, y hay mayornúmero de fibras elásticas, también más gruesas yparalelas a la superficie cutánea. Proporcionalmentehay menos sustancia fundamental y fibrocitos queen la dermis papilar.
A pesar de dividir la dermis en estas dos porciones, tam-bién se acepta dividirla en «superficial, media y profunda»,que permite localizar mejor ciertas entidades, ya que la der-mis reticular es muy grande en proporción a la papilar.
FuncionesLas funciones de la dermis son deducibles de lo anterior-mente expuesto:
1) Protectora, ya que el tejido conjuntivo supondrá unasegunda línea de defensa frente a traumatismos. No escasualidad que la pongamos en primer lugar ya que setrata de la función más importante. Cuando se aplica unafuerza sobre la epidermis, ésta la transmite a la dermissuperficial y el gel fluido que la compone la disipahaciendo que no sea fácil de romper la cohesión epidér-
Tem
as g
ener
ales
16
Figura 20. Célula de Merkel (M). Su relación con los melanocitos(Mela) y la de Langerhans (Lang).
Figura 21. Inmunohistoquímica de células de Merkel en la epider-mis.
M
Lang
Mela
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 16
Figura 20. Célula de Merkel (M). Su relación con los melanocitos (Mela) y la de Langerhans (Lang).
14
Manual de derMatología. Sección I: Temas generales
DermisEs la capa que sirve de sostén a la epidermis, a la que aporta sus nu-
trientes, y que contiene los anejos y las estructuras vasculonerviosas. Es una fascia superficial de tejido conjuntivo compuesta por células, fibras y sustancia fundamental (Tabla III), que tiene diferente textura según zonas del cuerpo y edad de la persona, variando su grosor desde 1 mm en los párpados hasta los 5 mm en la espalda. Es de 15 a 40 veces más gruesa que la epidermis. Al microscopio óptico muestra claramente dos partes: una superior, o dermis papilar, y otra inferior, o dermis reticular.
a) Dermis papilar. Se llama así porque está compuesta casi exclu-sivamente por la zona de las papilas dérmicas ya que llega hasta donde las crestas interpapilares epidérmicas penetran en la dermis.
Tiene haces de colágeno y fibras elásticas, más delgados que en la dermis reticular y orientados verticalmente al epitelio, y, sobre todo, abundante sustancia fundamental, donde las finas fibras de colágeno forman una red. El diámetro de las fibras de colágeno es de unos 50 nm y forman haces de unas 0,3-3 micras. En su interior está el plexo vascu lar superficial.
b) Dermis reticular. Es mucho más gruesa que la papilar, ocho o nueve veces más. Las bandas colágenas son más abundantes y más gruesas, de unos 63 nm de diámetro, se disponen en gruesos haces horizontales al epitelio, de unas 10-40 micras, y hay mayor número de fibras elásticas, también más gruesas y paralelas a la superficie cutánea. Proporcionalmen-te hay menos sustancia fundamental y fibrocitos que en la dermis papilar.
A pesar de dividir la dermis en estas dos porciones, también se acepta dividirla en «superficial, media y profunda», que permite localizar mejor ciertas entidades, ya que la dermis reticular es muy grande en proporción a la papilar.
FuncionesLas funciones de la dermis son deducibles de lo anteriormente ex-
puesto:
1) Protectora, ya que el tejido conjuntivo supondrá una segunda línea de defensa frente a traumatismos. No es casualidad que la pongamos en primer lugar ya que se trata de la función más importante. Cuando se aplica una fuerza sobre la epidermis, esta la transmite a la dermis superficial y el gel fluido que la compone la disipa haciendo que no sea fácil de romper la cohesión epidérmica. Además, la estructura de la dermis reticular la hace muy resistente a los traumas.
2) De soporte, al mantener el sistema vasculonervioso y anexial.3) De almacenamiento, tanto en el sistema vascular como, a veces, en
la sustancia fundamental.
Celularidad• Fibrocitos. Proceden de unas células pluripotentes o primitivas del
mesenquima, de las que también derivan los adipocitos hipodérmi-cos. Los fibrocitos son las células específicas de la dermis, puesto que se encargan de producir los elementos del tejido conectivo dérmico, fibras y sustancia fundamental, o sus precursores, ya que son el origen del tropocolágeno y de la tropoelastina. Además, son las células más numerosas del tejido conectivo laxo. Su aspecto es fusiforme o estre-llado, con núcleo grande y citoplasma amplio donde se observan un retículo endoplásmico y un aparato de Golgi bien definidos junto con prominentes ribosomas. Estas son características de células con una activa síntesis y secreción (Fig. 23).
Tabla III. Componentes de la dermis
Componentes de la dermis
Células Fibras Sustancia fundamental
• Fibrocitos• Histiocitos• Mastocitos
• Fibras de colágeno• Fibras elásticas
• Agua• Electrolitos• Proteínas plasmáticas• Proteoglucanos
Figura 21. Inmunohistoquímica de células de Merkel en la epidermis.
mica. Además, la estructura de la dermis reticular la hacemuy resistente a los traumas.
2) De soporte, al mantener el sistema vasculonervioso y ane-xial.
3) De almacenamiento, tanto en el sistema vascular como, aveces, en la sustancia fundamental.
Celularidad� Fibrocitos. Proceden de unas células pluripotentes o
primitivas del mesenquima, de las que también derivanlos adipocitos hipodérmicos. Los fibrocitos son las célu-las específicas de la dermis, puesto que se encargan deproducir los elementos del tejido conectivo dérmico,fibras y sustancia fundamental, o sus precursores, yaque son el origen del tropocolágeno y de la tropoelas-tina. Además, son las células más numerosas del tejidoconectivo laxo. Su aspecto es fusiforme o estrellado,con núcleo grande y citoplasma amplio donde se obser-van un retículo endoplásmico y un aparato de Golgibien definidos junto con prominentes ribosomas. Estasson características de células con una activa síntesis ysecreción (Fig. 23).
� Histiocitos. Son células del sistema mononuclear fagocí-tico que se parecen mucho a los fibrocitos (núcleo grandebasófilo y citoplasma claro con abundantes lisosomas).
Dependiendo de la misión fagocitaria que efectúen, reci-ben diferente denominación. Así, cuando estos macrófa-gos captan lípidos, se denominan «lipófagos», si es mela-nina, «melanófagos», y si las partículas a fagocitar sonmuy grandes, se multiplican rápidamente sus núcleos obien se unen varios macrófagos, formando las «célulasgigantes de cuerpos extraños», con múltiples núcleos dedisposición anárquica. Y también, en ciertas infecciones(tuberculosis, lepra, leishmaniasis), los histiocitos adop-tan un aspecto semejante a las células del cuerpo mucosode Malpighi (núcleo voluminoso oval y citoplasma eosi-nófilo mal definido), por lo que se denominan «célulasepitelioides», que a su vez pueden unirse formando las«células gigantes tipo Langhans», con núcleos dispuestosen herradura en la periferia del citoplasma; así constitu-yen los folículos, como el de Köster en la tuberculosis.
� Mastocitos. Son células mononucleadas, voluminosas, deforma variada (ovoides, poliédricas, fusiformes, estrelladas),que tienen un origen en una célula madre hematopoyéticade la médula ósea. Son más numerosos en la dermis subpa-pilar, en la región del plexo vascular superficial. La activaciónde estas células induce la liberación de mediadores inflama-torios preformados, localizados en unos gránulos especiali-zados, y la síntesis de novo y secreción de citocinas, quimo-cinas y eicosanoides, que intervienen en los procesos de
inflamación tanto aguda como crónica. Su característica prin-cipal es la presencia de abundantes granulaciones redondea-das citoplásmicas metacromáticas, fácilmente revelables porlos colorantes básicos. Estos gránulos de 0,6 micras de diá-metro contienen heparina, ácido hialurónico, histamina,serotonina, bradiquinina, cisteinil-leucotrienos (anterior-mente denominados como SRS-A) y otros derivados del ácidoaraquidónico. Además, estos gránulos de los mastocitos dela dermis contienen triptasa y quimotriptasa (mastocitos-TQ)y se diferencian por su aspecto de las granulaciones de losmastocitos de mucosa intestinal y pulmón, que sólo contie-nen triptasa (mastocitos-T). Los mastocitos juegan un papelprincipal en las reacciones de fase inmediata, agudas, produ-ciéndose la liberación de mediadores, como la histamina y laserotonina, que aumentan la permeabilidad capilar y deter-minan la pápula dérmica típica de la urticaria. Los eicosanoi-des, las prostaglandinas y los leucotrienos son sintetizadosdespués de la activación de los mastocitos, contribuyendo alas reacciones de fase intermedia y, posteriormente, las cito-cinas proinflamatorias, que contribuyen a las reacciones defase tardía.
Fibras de la dermis� Fibras colágenas. Son el componente estructural más
importante de la dermis y las que le dan la fuerza de ten-sión. Están producidas por los fibrocitos y se agrupan engruesas bandas onduladas que, como ya indicamos, endermis media y profunda se disponen paralelas a la super-
1.A
nato
mía
y f
isio
logí
a de
la p
iel
17
Figura 22. Gráfico que representa la célula de Merkel.
Queratinocitos
Desmosoma
Gránulos densos
Terminación nerviosa
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 17
Figura 22. Gráfico que representa la célula de Merkel.

15
Capítulo 1. Anatomía y fisiología de la piel
• Histiocitos. Son células del sistema mononuclear fagocítico que se parecen mucho a los fibrocitos (núcleo grande basófilo y citoplasma claro con abundantes lisosomas).
Dependiendo de la misión fagocitaria que efectúen, reciben diferente denominación. Así, cuando estos macrófagos captan lípidos, se denominan «lipófagos», si es melanina, «melanófagos», y si las partículas a fagocitar son muy grandes, se multiplican rápidamente sus núcleos o bien se unen varios macrófagos, formando las «células gigantes de cuerpos extraños», con múltiples núcleos de disposición anárquica. Y también, en ciertas infecciones (tuberculosis, lepra, leishmaniasis), los histiocitos adoptan un aspecto semejante a las células del cuerpo mucoso de Malpighi (núcleo voluminoso oval y citoplasma eosinófilo mal definido), por lo que se denominan «células epitelioides», que a su vez pueden unirse formando las «células gigantes tipo Langhans», con núcleos dispuestos en herradura en la periferia del citoplasma; así constituyen los folículos, como el de Köster en la tuberculosis.
• Mastocitos. Son células mononucleadas, voluminosas, de forma varia-da (ovoides, poliédricas, fusiformes, estrelladas), que tienen un origen en una célula madre hematopoyética de la médula ósea. Son más numero-sos en la dermis subpapilar, en la región del plexo vascular superficial. La activación de estas células induce la liberación de mediadores infla-matorios preformados, localizados en unos gránulos especializados, y la síntesis de novo y secreción de citocinas, quimocinas y eicosanoides, que intervienen en los procesos de inflamación tanto aguda como crónica. Su característica principal es la presencia de abundantes granulaciones redondeadas citoplásmicas metacromáticas, fácilmente revelables por los colorantes básicos. Estos gránulos de 0,6 micras de diámetro contienen heparina, ácido hialurónico, histamina, serotonina, bradiquinina, cis-teinil-leucotrienos (anteriormente denominados como SRS-A) y otros derivados del ácido araquidónico. Además, estos gránulos de los masto-citos de la dermis contienen triptasa y quimotriptasa (mastocitos-TQ) y se diferencian por su aspecto de las granulaciones de los mastocitos de mucosa intestinal y pulmón, que solo contienen triptasa (mastocitos-T). Los mastocitos juegan un papel principal en las reacciones de fase in-mediata, agudas, produciéndose la liberación de mediadores, como la histamina y la serotonina, que aumentan la permeabilidad capilar y de-terminan la pápula dérmica típica de la urticaria. Los eicosanoides, las
prostaglandinas y los leucotrienos son sintetizados después de la activa-ción de los mastocitos, contribuyendo a las reacciones de fase intermedia y, posteriormente, las citocinas proinflamatorias, que contribuyen a las reacciones de fase tardía.
Fibras de la dermis• Fibras colágenas. Son el componente estructural más importante
de la dermis y las que le dan la fuerza de tensión. Están producidas por los fibrocitos y se agrupan en gruesas bandas onduladas que, como ya indicamos, en dermis media y profunda se disponen paralelas a la superficie cutánea, mientras que en dermis papilar son verticales. El diámetro de estas fibras de colágeno se va incrementando de forma pro-gresiva desde la dermis superficial hasta la dermis media y la profun-da. Cada fibra de colágeno está compuesta por microfibrillas o fibrillas elementales formadas de tropocolágeno, el cual consta de tres cadenas de polipéptidos enrolladas unas a las otras formando una estructura de triple hélice. Cada cadena polipeptídica contiene grandes cantidades de hidroxiprolina, hidroxilisina y glicina. La alineación de las moléculas a lo largo de las microfibrillas se estabiliza por medio de uniones inter- e intramoleculares. Morfométricamente se comprueba que las bandas de colágeno representan el 24% del tejido de la dermis fetal. Aumentan hasta el 60% al nacer y alcanzan su máxima densidad, casi el 70%, du-rante la segunda década de la vida, para a partir de los 60 años disminuir hasta valores del 40%. El número de bandas de colágeno por unidad de área disminuye inmediatamente después de nacer y esta disminución sigue durante el resto de la vida aunque es menos evidente. Por el con-trario, el tamaño de las bandas de colágeno aumenta al nacer y tiende a seguir aumentando durante el resto de la vida. El colágeno se fabrica en el retículo endoplásmico rugoso de los fibroblastos, donde sus ribosomas, al polimerizar aminoácidos, sintetizan las cadenas del procolágeno. Tres de esas cadenas polipeptídicas se enrollan en forma de una triple hélice, convirtiéndose en tropocolágeno, el cual se almacena en el aparato de Golgi y después se libera al espacio extracelular, donde se ensambla en fibrillas elementales que, por último, constituyen, al agruparse también ellas, las fibras y bandas de colágeno. La hidroxilisina es crucial para las uniones inter- e intramoleculares de la fase extracelular. Estas bandas de colágeno mantienen la epidermis adherida a planos más profundos y, por tanto, son las responsables de la presencia de los «surcos cutáneos» visibles en la superficie externa del epitelio. Aunque los surcos se observan en forma de figuras geométricas cuadrangulares en todo el cuerpo, tienen especial configuración en las palmas de las manos (lineales) y, sobre todo, en la yema de los dedos, donde, al adop-tar una forma de líneas concéntricas, proporcionan carácter individual a las personas; son los denominados dermatoglifos, que van a formar la «huella dactilar».
En la dermis hay fundamentalmente colágeno tipo I (75-85%), tipo III (15%) y tipo V (2-4%), aunque también existen los tipos IV, VI y VII. El colágeno tipo I es el más abundante y se encuentra organizado en una densa red ortogonal en la dermis reticular. La fuerza tensora de la dermis se debe principalmente a este colágeno tipo I y, proporcionalmente en menor medida, al colágeno tipo III. Las fibras de colágeno tipo III, anteriormente denominadas reticulina, se encuentran principalmente situadas en la dermis papilar y perianexial (Fig. 23).
ficie cutánea, mientras que en dermis papilar son vertica-les. El diámetro de estas fibras de colágeno se va incre-mentando de forma progresiva desde la dermis superficialhasta la dermis media y la profunda. Cada fibra de colá-geno está compuesta por microfibrillas o fibrillas elemen-tales formadas de tropocolágeno, el cual consta de trescadenas de polipéptidos enrolladas unas a las otras for-mando una estructura de triple hélice. Cada cadena poli-peptídica contiene grandes cantidades de hidroxiprolina,hidroxilisina y glicina. La alineación de las moléculas a lolargo de las microfibrillas se estabiliza por medio de unio-nes inter e intramoleculares. Morfométricamente se com-prueba que las bandas de colágeno representan el 24% deltejido de la dermis fetal. Aumentan hasta el 60% al nacer yalcanzan su máxima densidad, casi el 70%, durante lasegunda década de la vida, para a partir de los 60 añosdisminuir hasta valores del 40%. El número de bandas decolágeno por unidad de área disminuye inmediatamentedespués de nacer y esta disminución sigue durante elresto de la vida aunque es menos evidente. Por el contra-rio, el tamaño de las bandas de colágeno aumenta al nacery tiende a seguir aumentando durante el resto de la vida.El colágeno se fabrica en el retículo endoplásmico rugosode los fibroblastos, donde sus ribosomas, al polimerizaraminoácidos, sintetizan las cadenas del procolágeno. Tresde esas cadenas polipeptídicas se enrollan en forma deuna triple hélice, convirtiéndose en tropocolágeno, el cualse almacena en el aparato de Golgi y después se libera alespacio extracelular, donde se ensambla en fibrillas ele-mentales que, por último, constituyen, al agruparse tam-bién ellas, las fibras y bandas de colágeno. La hidroxilisinaes crucial para las uniones inter e intramoleculares de lafase extracelular.
Estas bandas de colágeno mantienen la epidermis adhe-rida a planos más profundos y, por tanto, son las respon-sables de la presencia de los «surcos cutáneos» visibles enla superficie externa del epitelio. Aunque los surcos seobservan en forma de figuras geométricas cuadrangularesen todo el cuerpo, tienen especial configuración en las pal-mas de las manos (lineales) y, sobre todo, en la yema delos dedos, donde, al adoptar una forma de líneas concén-tricas, proporcionan carácter individual a las personas; sonlos denominados dermatoglifos, que van a formar la «hue-lla dactilar».
En la dermis hay fundamentalmente colágeno tipo I(75-85%), tipo III (15%) y tipo V (2-4%), aunquetambién existen los tipos IV, VI y VII. El colágeno tipoI es el más abundante y se encuentra organizado enuna densa red ortogonal en la dermis reticular. Lafuerza tensora de la dermis se debe principalmente aeste colágeno tipo I y, proporcionalmente en menormedida, al colágeno tipo III. Las fibras de colágenotipo III, anteriormente denominadas reticulina, seencuentran principalmente situadas en la dermis papi-lar y perianexial (Fig. 23).
Son más numerosas en la zona inmediatamente debajo dela unión dermoepidérmica, aunque también están presen-tes en toda la dermis, en íntima relación con las fibras decolágeno tipo I. El colágeno tipo V se encuentra localizadoentre las células dérmicas y el resto de fibras de colágenointersticiales y también alrededor de los vasos sanguí-neos.
Actualmente se han descrito 27 tipos de colágeno quese dividen en diversos subgrupos. Hay cinco miembros,los colágenos tipos I, II, III, V y XI, denominados colágenosfibrilares, que forman triples cadenas helicoidales perfec-tas. Por otro lado, estaría el grupo de los colágenos atípi-cos, o no fibrilares, que son más numerosos y más diver-sos. En éstos se encuadrarían los denominados colágenosrelacionados con la membrana basal, como el tipo IV y eltipo VII, que difieren del resto por su gran tamaño y quetienen triples hélices extendidas, y el colágeno tipo XVII oantígeno 2 del penfigoide ampolloso (BPAg2), los coláge-nos FACIT (colágenos tipos XII, XIV y XVI) y los colágenosde cadena corta, como los colágenos tipos VI, VIII y X.
El colágeno es relativamente inerte y persiste durantelargos periodos de tiempo, pero está sometido también auna remodelación. Los mecanismos de reabsorción sondiferentes según los tipos de colágeno y dependen de laregulación tisular específica. Los colágenos fibrosos sonespecíficamente degradados por las colagenasas. Las cola-genasas 1, 2 y 3 forman parte de las denominadas metalo-proteinasas de matriz, familia de más de 20 miembrosque, además de los colágenos, degradan proteoglicanos yotros componentes de la matriz extracelular. Tambiénexisten, al menos, tres inhibidores titulares de las metalo-proteinasas, que las bloquean reversiblemente.
� Fibras elásticas. Aunque son escasas, tienen una impor-tancia extrema, pues determinan la extensibilidad y elasti-cidad cutáneas. Están constituidas por un material amorfocentral consistente en elastina, que es una proteína fibrosainsoluble derivada de la tropoelastina, y unas microfibri-llas periféricas, más heterogéneas, formadas por variasglicoproteínas insolubles, como las fibrilinas 1 y 2 o lasfibulinas 1 y 2. Las propiedades de extensibilidad y elasti-cidad de las fibras elásticas se deben al contenido en elas-tina, mientras que las microfibrillas dan la estabilidad alconjunto de cada fibra. Se disponen orientadas perpendi-cularmente en el cuerpo papilar, donde son más delgadas,
Tem
as g
ener
ales
18
Figura 23. El Fibroblasto. Sustancia que produce.
Fibroblasto
Tropoelastina
Sustancia de sustentoFibra elástica
Fibrilla de colágeno (III)(fibra reticular)
Fibrilla decolágeno (I)
Tropocolágeno
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 18
Figura 23. El fibroblasto. Sustancia que produce.

16
Manual de derMatología. Sección I: Temas generales
Son más numerosas en la zona inmediatamente debajo de la unión dermoepidérmica, aunque también están presentes en toda la dermis, en íntima relación con las fibras de colágeno tipo I. El colágeno tipo V se encuentra localizado entre las células dérmicas y el resto de fibras de colágeno intersticiales y también alrededor de los vasos sanguíneos. Actualmente se han descrito 27 tipos de colágeno que se dividen en diversos subgrupos. Hay cinco miembros, los colágenos tipos I, II, III, V y XI, denominados colágenos fibrilares, que forman triples cadenas helicoidales perfectas. Por otro lado, estaría el grupo de los colágenos atípicos, o no fibrilares, que son más numerosos y más di-versos. En estos se encuadrarían los denominados colágenos relacionados con la membrana basal, como el tipo IV y el tipo VII, que difieren del resto por su gran tamaño y que tienen triples hélices extendidas, y el colágeno tipo XVII o antígeno 2 del penfigoide ampolloso (BPAg2), los colágenos FACIT (colágenos tipos XII, XIV y XVI) y los colágenos de cadena corta, como los colágenos tipos VI, VIII y X. El colágeno es relativamente inerte y persiste durante largos perio-dos de tiempo, pero está sometido también a una remodelación. Los mecanismos de reabsorción son diferentes según los tipos de colágeno y dependen de la regulación tisular específica. Los colágenos fibrosos son específicamente degradados por las colagenasas. Las colagenasas 1, 2 y 3 forman parte de las denominadas metaloproteinasas de matriz, familia de más de 20 miembros que, además de los colágenos, degra-dan proteoglucanos y otros componentes de la matriz extracelular. También existen, al menos, tres inhibidores titulares de las metalo-proteinasas, que las bloquean reversiblemente.
• Fibras elásticas. Aunque son escasas, tienen una importancia extrema, pues determinan la extensibilidad y elasticidad cutáneas. Están constituidas por un material amorfo central consistente en elas-tina, que es una proteína fibrosa insoluble derivada de la tropoelastina, y unas microfibrillas periféricas, más heterogéneas, formadas por varias glucoproteínas insolubles, como las fibrilinas 1 y 2 o las fibulinas 1 y 2. Las propiedades de extensibilidad y elasticidad de las fibras elásti-cas se deben al contenido en elastina, mientras que las microfibrillas dan la estabilidad al conjunto de cada fibra. Se disponen orientadas perpendicularmente en el cuerpo papilar, donde son más delgadas, y paralelas a la superficie cutánea en la dermis profunda, siendo más gruesas. La síntesis de las fibras elásticas por los fibrocitos consta de dos etapas. En la primera, se secretan al espacio extracelular las mi-crofibrillas agrupadas de forma lineal produciéndose una polimeriza-ción posteriormente. En la segunda, se produce la secreción de la tro-poelastina dentro de los cilindros preformados de microfibrillas; la elastina se une a ellas por medio de uniones covalentes conocidas como desmosinas e isodesmosinas. Estas fibras son muy numerosas al nacer, observándose como bandas de material amorfo rodeadas de un gran número de microfibrillas de elastina. Pocos meses después del nacimiento, aumenta la cantidad de elastina amorfa y ya se com-prueba la organización típica de las fibras elásticas maduras. Durante la vida adulta, las fibras elásticas muestran pocas variaciones, a ex-cepción de una discreta reducción de las microfibrillas y la presencia de algunas inclusiones electrón-densas y de márgenes irregulares, lo que se hace más evidente sobre los cincuenta años.
Morfométricamente, las fibras elásticas representan el 3-4% del total de la dermis en el recién nacido, no cambiando significativamente hasta la quinta década de la vida, a partir de la cual aumentan al 5% y en los ancianos llegan al 7-8%. Este aumento se debe al mayor diámetro de las fibras y al mayor número por unidad de área. Las mujeres ancianas muestran más fibras elásticas que los varones.
La degradación de la elastina por medio de las elastasas da lugar a la formación de unos fragmentos peptídicos denominados elastocinas, para remarcar sus propiedades de citocinas, ya que se ha visto que representan una importante señal de reparación tisular.
SUSTANCIA FUNDAMENTAL
Es un material extracelular amorfo que se encuentra entre las fibras propias de la dermis, siendo también producida por los fibrocitos. Está constituida por agua, electrolitos, proteínas plasmáticas y proteoglucanos. Los proteoglucanos se tratan de cadenas de polisacáridos aminados (glu-cosaminoglucanos) unidas a proteínas centrales.
En la piel, los glucosaminoglucanos más abundantes son derma-tán-sulfato y ácido hialurónico junto con pequeñas cantidades de con-droitín-6-sulfato, heparán-sulfato y heparina. Los glucosaminoglucanos son muy higroscópicos reteniendo gran cantidad de agua.
En la matriz extracelular también se encuentran diversas glucopro-teínas de adhesión como fibronectina, vitronectina, trombospondinas y tenascinas. Estas glucoproteínas se unen a las superficies celulares, a los glucosaminoglucanos y a las fibras de colágeno provocando unas interac-ciones estables.
Las fibronectinas que se unen a las fibras de colágeno tipo III, ante-riormente denominadas «fibras reticulares», son las responsables de las reacciones argirófilas de esas fibras.
ANEJOS CUTÁNEOS
Como señalamos antes, los hay queratinizados y glandulares; unos claramente separados y otros en íntima relación, conformando el deno-minado folículo pilosebáceo.
Anejos queratinizadosSon dos: el pelo, que forma parte del folículo piloso, y la uña.
Folículo pilosoEl folículo piloso tiene una porción distal conocida como «bulbo»,
compuesto por la matriz pilosa y la papila y un cuerpo cilíndrico en el que, de abajo arriba, se observa la inserción del músculo erector del pelo por medio del tendón elástico de Nagel, constituido por fibras elásticas elaunínicas y oxitalánicas, la glándula sebácea y la glándula sudorípara apocrina (Figs. 24, 25 y 26).
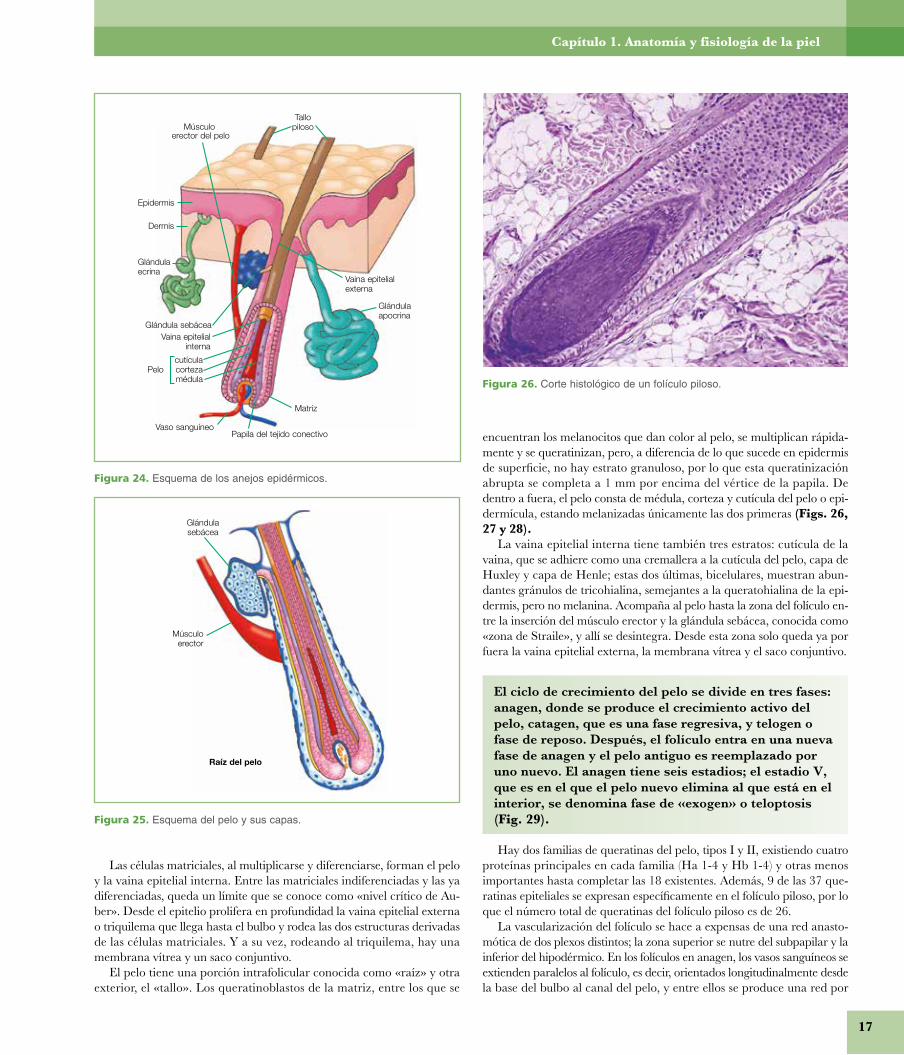
17
Capítulo 1. Anatomía y fisiología de la piel
Las células matriciales, al multiplicarse y diferenciarse, forman el pelo y la vaina epitelial interna. Entre las matriciales indiferenciadas y las ya diferenciadas, queda un límite que se conoce como «nivel crítico de Au-ber». Desde el epitelio prolifera en profundidad la vaina epitelial externa o triquilema que llega hasta el bulbo y rodea las dos estructuras derivadas de las células matriciales. Y a su vez, rodeando al triquilema, hay una membrana vítrea y un saco conjuntivo.
El pelo tiene una porción intrafolicular conocida como «raíz» y otra exterior, el «tallo». Los queratinoblastos de la matriz, entre los que se
encuentran los melanocitos que dan color al pelo, se multiplican rápida-mente y se queratinizan, pero, a diferencia de lo que sucede en epidermis de superficie, no hay estrato granuloso, por lo que esta queratinización abrupta se completa a 1 mm por encima del vértice de la papila. De dentro a fuera, el pelo consta de médula, corteza y cutícula del pelo o epi-dermícula, estando melanizadas únicamente las dos primeras (Figs. 26, 27 y 28).
La vaina epitelial interna tiene también tres estratos: cutícula de la vaina, que se adhiere como una cremallera a la cutícula del pelo, capa de Huxley y capa de Henle; estas dos últimas, bicelulares, muestran abun-dantes gránulos de tricohialina, semejantes a la queratohialina de la epi-dermis, pero no melanina. Acompaña al pelo hasta la zona del folículo en-tre la inserción del múscu lo erector y la glándula sebácea, conocida como «zona de Straile», y allí se desintegra. Desde esta zona solo queda ya por fuera la vaina epitelial externa, la membrana vítrea y el saco conjuntivo.
El ciclo de crecimiento del pelo se divide en tres fases: anagen, donde se produce el crecimiento activo del pelo, catagen, que es una fase regresiva, y telogen o fase de reposo. Después, el folículo entra en una nueva fase de anagen y el pelo antiguo es reemplazado por uno nuevo. El anagen tiene seis estadios; el estadio V, que es en el que el pelo nuevo elimina al que está en el interior, se denomina fase de «exogen» o teloptosis (Fig. 29).
Hay dos familias de queratinas del pelo, tipos I y II, existiendo cuatro proteínas principales en cada familia (Ha 1-4 y Hb 1-4) y otras menos importantes hasta completar las 18 existentes. Además, 9 de las 37 que-ratinas epiteliales se expresan específicamente en el folículo piloso, por lo que el número total de queratinas del folículo piloso es de 26.
La vascularización del folículo se hace a expensas de una red anasto-mótica de dos plexos distintos; la zona superior se nutre del subpapilar y la inferior del hipodérmico. En los folícu los en anagen, los vasos sanguíneos se extienden paralelos al folículo, es decir, orientados longitudinalmente desde la base del bulbo al canal del pelo, y entre ellos se produce una red por
y paralelas a la superficie cutánea en la dermis profunda,siendo más gruesas. La síntesis de las fibras elásticas porlos fibrocitos consta de dos etapas. En la primera se secre-tan al espacio extracelular las microfibrillas agrupadas deforma lineal produciéndose una polimerización posterior-mente. En la segunda se produce la secreción de la tropoe-lastina dentro de los cilindros preformados de microfibri-llas; la elastina se une a ellas por medio de unionescovalentes conocidas como desmosinas e isodesmosinas.Estas fibras son muy numerosas al nacer, observándosecomo bandas de material amorfo rodeadas de un grannúmero de microfibrillas de elastina. Pocos meses despuésdel nacimiento, aumenta la cantidad de elastina amorfa yya se comprueba la organización típica de las fibras elásti-cas maduras. Durante la vida adulta, las fibras elásticasmuestran pocas variaciones a excepción de una discretareducción de las microfibrillas y la presencia de algunasinclusiones electrón-densas y de márgenes irregulares, loque se hace más evidente sobre los cincuenta años.
Morfométricamente, las fibras elásticas representan el3-4% del total de la dermis en el recién nacido, nocambiando significativamente hasta la quinta décadade la vida, a partir de la cual aumentan al 5% y en losancianos llegan al 7-8%. Este aumento se debe almayor diámetro de las fibras y al mayor número porunidad de área. Las mujeres ancianas muestran másfibras elásticas que los varones.
La degradación de la elastina por medio de las elastasasda lugar a la formación de unos fragmentos peptídicos deno-minados elastocinas, para remarcar sus propiedades de cito-cinas, ya que se ha visto que representan una importanteseñal de reparación tisular.
SUSTANCIA FUNDAMENTAL
Es un material extracelular amorfo que se encuentra entre lasfibras propias de la dermis, siendo también producida por losfibrocitos. Está constituida por agua, electrolitos, proteínasplasmáticas y proteoglicanos. Los proteoglicanos se tratan decadenas de polisacáridos aminados (glicosaminoglicanos) uni-das a proteínas centrales.
En la piel, los glicosaminoglicanos más abundantes sondermatan-sulfato y ácido hialurónico junto con pequeñas can-tidades de condroitin-6-sulfato, heparan-sulfato y heparina.Los glicosaminoglicanos son muy higroscópicos reteniendogran cantidad de agua.
En la matriz extracelular también se encuentran diversasglicoproteínas de adhesión como fibronectina, vitronectina,trombospondinas y tenascinas. Estas glicoproteínas se unen alas superficies celulares, a los glicosaminoglicanos y a lasfibras de colágeno provocando unas interacciones estables.
Las fibronectinas que se unen a las fibras de colágeno tipoIII, anteriormente denominadas «fibras reticulares», son lasresponsables de las reacciones argirófilas de esas fibras.
ANEJOS CUTÁNEOS
Como señalamos antes, los hay queratinizados y glandulares;unos claramente separados y otros en íntima relación, confor-mando el denominado folículo pilosebáceo.
Anejos queratinizadosSon dos: el pelo, que forma parte del folículo piloso, y la uña.
Folículo pilosoEl folículo piloso tiene una porción distal conocida como«bulbo», compuesto por la matriz pilosa y la papila y uncuerpo cilíndrico en el que, de abajo arriba, se observa lainserción del músculo erector del pelo por medio del tendónelástico de Nagel, constituido por fibras elásticas elaunínicasy oxitalánicas, la glándula sebácea y la glándula sudoríparaapocrina (Figs. 24, 25 y 26).
Las células matriciales, al multiplicarse y diferenciarse, for-man el pelo y la vaina epitelial interna. Entre las matricialesindiferenciadas y las ya diferenciadas, queda un límite que se
1.A
nato
mía
y f
isio
logí
a de
la p
iel
19
Figura 24. Esquema de los anejos epidérmicos.
Epidermis
Dermis
Glándulaecrina
Músculoerector del pelo
Glándula sebáceaVaina epitelial
interna
Pelocutículacortezamédula
Vaso sanguíneoPapila del tejido conectivo
Matriz
Glándulaapocrina
Vaina epitelialexterna
Tallopiloso
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 19
Figura 24. Esquema de los anejos epidérmicos.
Figura 26. Corte histológico de un folículo piloso.
conoce como «nivel crítico de Auber». Desde el epitelio proli-fera en profundidad la vaina epitelial externa o triquilema quellega hasta el bulbo y rodea las dos estructuras derivadas delas células matriciales. Y a su vez, rodeando al triquilema, hayuna membrana vítrea y un saco conjuntivo.
El pelo tiene una porción intrafolicular conocida como«raíz» y otra exterior, el «tallo». Los queratinoblastos de lamatriz, entre los que se encuentran los melanocitos que dancolor al pelo, se multiplican rápidamente y se queratinizan,pero, a diferencia de lo que sucede en epidermis de superfi-cie, no hay estrato granuloso, por lo que esta queratinizaciónabrupta se completa a 1 mm por encima del vértice de la
papila. De dentro a fuera, el pelo consta de médula, corteza ycutícula del pelo o epidermícula, estando melanizadas única-mente las dos primeras (Figs. 26, 27 y 28).
La vaina epitelial interna tiene también tres estratos: cutículade la vaina, que se adhiere como una cremallera a la cutícula delpelo, capa de Huxley y capa de Henle; estas dos últimas, bicelu-lares, muestran abundantes gránulos de tricohialina, semejan-
Tem
as g
ener
ales
20
Figura 26. Corte histológico de un folículo piloso.
Figura 27. Detalle histológico de la zona matricial del pelo.
Figura 28. Detalle histológico de una sección transversal del folículopiloso.
Figura 25. Esquema del pelo y sus capas.
Glándulasebácea
Músculoerector
Raíz del pelo
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 20
Figura 25. Esquema del pelo y sus capas.
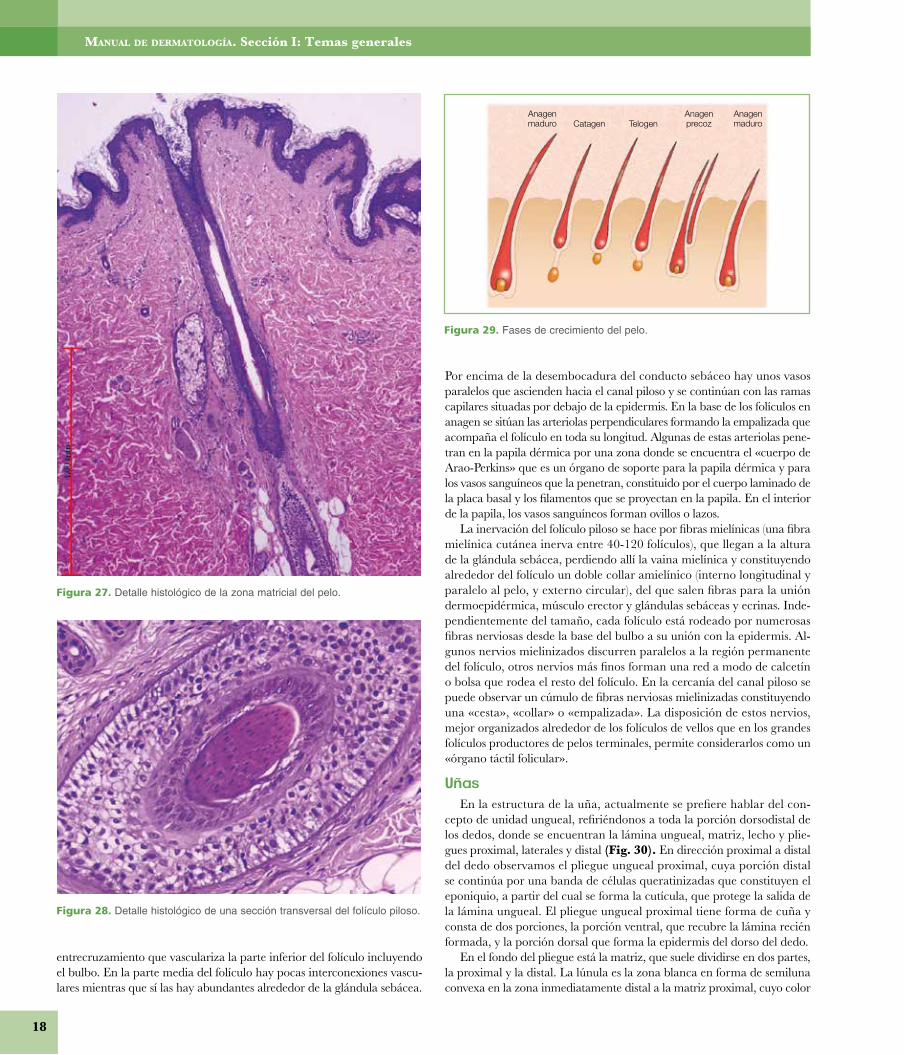
18
Manual de derMatología. Sección I: Temas generales
entrecruzamiento que vasculariza la parte inferior del folículo incluyendo el bulbo. En la parte media del folículo hay pocas interconexiones vascu-lares mientras que sí las hay abundantes alrededor de la glándula sebácea.
Por encima de la desembocadura del conducto sebáceo hay unos vasos paralelos que ascienden hacia el canal piloso y se continúan con las ramas capilares situadas por debajo de la epidermis. En la base de los folículos en anagen se sitúan las arteriolas perpendiculares formando la empalizada que acompaña el folículo en toda su longitud. Algunas de estas arteriolas pene-tran en la papila dérmica por una zona donde se encuentra el «cuerpo de Arao-Perkins» que es un órgano de soporte para la papila dérmica y para los vasos sanguíneos que la penetran, constituido por el cuerpo laminado de la placa basal y los filamentos que se proyectan en la papila. En el interior de la papila, los vasos sanguíneos forman ovillos o lazos.
La inervación del folículo piloso se hace por fibras mielínicas (una fibra mielínica cutánea inerva entre 40-120 folículos), que llegan a la altura de la glándula sebácea, perdiendo allí la vaina mielínica y constituyendo alrededor del folículo un doble collar amielínico (interno longitudinal y paralelo al pelo, y externo circular), del que salen fibras para la unión dermoepidérmica, músculo erector y glándulas sebáceas y ecrinas. Inde-pendientemente del tamaño, cada folículo está rodeado por numerosas fibras nerviosas desde la base del bulbo a su unión con la epidermis. Al-gunos nervios mielinizados discurren paralelos a la región permanente del folículo, otros nervios más finos forman una red a modo de calcetín o bolsa que rodea el resto del folículo. En la cercanía del canal piloso se puede observar un cúmulo de fibras nerviosas mielinizadas constituyendo una «cesta», «collar» o «empalizada». La disposición de estos nervios, mejor organizados alrededor de los folículos de vellos que en los grandes folículos productores de pelos terminales, permite considerarlos como un «órgano táctil folicular».
UñasEn la estructura de la uña, actualmente se prefiere hablar del con-
cepto de unidad ungueal, refiriéndonos a toda la porción dorsodistal de los dedos, donde se encuentran la lámina ungueal, matriz, lecho y plie-gues proximal, laterales y distal (Fig. 30). En dirección proximal a distal del dedo observamos el pliegue ungueal proximal, cuya porción distal se continúa por una banda de células queratinizadas que constituyen el eponiquio, a partir del cual se forma la cutícula, que protege la salida de la lámina ungueal. El pliegue ungueal proximal tiene forma de cuña y consta de dos porciones, la porción ventral, que recubre la lámina recién formada, y la porción dorsal que forma la epidermis del dorso del dedo.
En el fondo del pliegue está la matriz, que suele dividirse en dos partes, la proximal y la distal. La lúnula es la zona blanca en forma de semiluna convexa en la zona inmediatamente distal a la matriz proximal, cuyo color
Figura 27. Detalle histológico de la zona matricial del pelo.
Figura 28. Detalle histológico de una sección transversal del folículo piloso.
tes a la queratohialina de la epidermis, pero no melanina. Acom-paña al pelo hasta la zona del folículo entre la inserción del mús-cu lo erector y la glándula sebácea, conocida como «zona deStraile», y allí se desintegra. Desde esta zona sólo queda ya porfuera la vaina epitelial externa, la membrana vítrea y el saco con-juntivo.
El ciclo de crecimiento del pelo se divide en tres fases:anagen, donde se produce el crecimiento activo delpelo, catagen, que es una fase regresiva, y telogen o fasede reposo. Después, el folículo entra en una nueva fasede anagen y el pelo antiguo es reemplazado por unonuevo. El anagen tiene seis estadios; el estadio V, quees en el que el pelo nuevo elimina al que está en el inte-rior, se denomina fase de «exogen» o teloptosis (Fig.29).
Hay dos familias de queratinas del pelo, tipos I y II, exis-tiendo cuatro proteínas principales en cada familia (Ha 1-4 yHb 1-4) y otras menos importantes hasta completar las 18existentes. Además, 9 de las 37 queratinas epiteliales seexpresan específicamente en el folículo piloso, por lo que elnúmero total de queratinas del folículo piloso es de 26.
La vascularización del folículo se hace a expensas de unared anastomótica de dos plexos distintos; la zona superior senutre del subpapilar y la inferior del hipodérmico. En los folí-cu los en anagen, los vasos sanguíneos se extienden paralelosal folículo, es decir, orientados longitudinalmente desde labase del bulbo al canal del pelo, y entre ellos se produce unared por entrecruzamiento que vasculariza la parte inferior delfolículo incluyendo el bulbo. En la parte media del folículo haypocas interconexiones vasculares mientras que sí las hayabundantes alrededor de la glándula sebácea. Por encima dela desembocadura del conducto sebáceo hay unos vasos para-
lelos que ascienden hacia el canal piloso y se continúan conlas ramas capilares situadas por debajo de la epidermis. En labase de los folículos en anagen se sitúan las arteriolas per-pendiculares formando la empalizada que acompaña el folí-culo en toda su longitud. Algunas de estas arteriolas penetranen la papila dérmica por una zona donde se encuentra el«cuerpo de Arao-Perkins» que es un órgano de soporte para lapapila dérmica y para los vasos sanguíneos que la penetran,constituido por el cuerpo laminado de la placa basal y los fila-mentos que se proyectan en la papila. En el interior de lapapila, los vasos sanguíneos forman ovillos o lazos.
La inervación del folículo piloso se hace por fibras mielíni-cas (una fibra mielínica cutánea inerva entre 40-120 folícu-los), que llegan a la altura de la glándula sebácea, perdiendoallí la vaina mielínica y constituyendo alrededor del folículoun doble collar amielínico (interno longitudinal y paralelo alpelo, y externo circular), del que salen fibras para la unióndermoepidérmica, músculo erector y glándulas sebáceas yecrinas. Independientemente del tamaño, cada folículo estárodeado por numerosas fibras nerviosas desde la base delbulbo a su unión con la epidermis. Algunos nervios mieliniza-dos discurren paralelos a la región permanente del folículo,otros nervios más finos forman una red a modo de calcetín obolsa que rodea el resto del folículo. En la cercanía del canalpiloso se puede observar un cúmulo de fibras nerviosas mieli-nizadas constituyendo una «cesta», «collar» o «empalizada».La disposición de estos nervios, mejor organizados alrededorde los folículos de vellos que en los grandes folículos produc-tores de pelos terminales, permite considerarlos como un«órgano táctil folicular».
UñasEn la estructura de la uña, actualmente se prefiere hablar delconcepto de unidad ungueal, refiriéndonos a toda la porcióndorsodistal de los dedos, donde se encuentran la láminaungueal, matriz, lecho y pliegues proximal, laterales y distal(Fig. 30). En dirección proximal a distal del dedo observamos
1.A
nato
mía
y f
isio
logí
a de
la p
iel
21
Figura 29. Fases de crecimiento del pelo.
Anagenmaduro Catagen Telogen
Anagenprecoz
Anagenmaduro
Figura 30. Unidad ungueal.
Borde libreLámina ungueal
Lúnula
Lámina ungueal Hiponiquio
Matrizungueal
Eponiquio
Pliegue proximal
CutículaMatriz
ungueal
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 21
Figura 29. Fases de crecimiento del pelo.

19
Capítulo 1. Anatomía y fisiología de la piel
es debido a la queratinización incompleta de las células y a que la matriz no deja transparentar los vasos.
La matriz produce la lámina ungueal, que es convexa y translucida pero que presenta un color rosado debido a la riqueza vascular subya-cente, que se desliza y adhiere firmemente al lecho ungueal, soporte de la lámina, con su epidermis y dermis, debajo del cual está la falange distal a la que se fija. Distal al lecho queda un grupo de células queratinizadas que constituyen el hiponiquio, que protege al lecho de los agentes externos y al que sigue el pliegue distal, que se continúa hasta el pulpejo del dedo. Por encima es posible observar en la región distal de la lámina una fina banda amarillenta, convexa, la banda onicodérmica o porción dermoungueal de Terry, que parece tener una irrigación sanguínea diferente al resto del lecho ungueal. Marginando la uña se encuentran los pliegues ungueales laterales, que están separados de ella por los surcos ungueales laterales.
La lámina ungueal esta formada por tres capas: una fina capa dorsal, una capa gruesa intermedia y la capa ventral procedente del lecho. El 20% de su superficie total se localiza bajo la porción ventral del pliegue ungueal proximal. Al igual que en el pelo, la queratinización de la matriz y del lecho se produce en ausencia de capa granulosa.
El aporte sanguíneo ungueal se efectúa por los plexos arteriales proxi-mal y distal, formados por anastomosis entre las dos arterias digitales laterales; el proximal es paralelo a la lúnula y el distal a la punta de la falange distal subyacente.
La dermis subungueal presenta terminaciones nerviosas de Vater-Paci-ni y corpúsculos de Meissner. El hiponiquio y los pliegues laterales son las áreas que poseen mayor número de terminaciones nerviosas y corpúsculos de Meissner.
El crecimiento normal ungueal es de aproximadamente 0,1 mm al día, siendo más rápido en las manos que en los pies. El recambio completo de las uñas de las manos es de unos 6 meses mientras que en los pies, como la velocidad es la mitad o la tercera parte, será de 12-18 meses.
Anejos glandularesGlándulas sebáceas
Están situadas inmediatamente por encima del músculo erector, en la porción superior del folículo, donde desembocan por su conducto excre-tor, de epitelio poliestratificado. Están formadas por amplios lóbulos en cuya periferia se encuentran la membrana basal y las células matriciales
cuboides basófilas que, conforme van multiplicándose, se van llenando de vacuolas de lípidos que empujan y ahogan el núcleo, tornándose entonces claras. Estas células van aproximándose al conducto excretor y, al llegar a él, se rompe su membrana citoplasmática vertiéndose la to-talidad del contenido celular (Figs. 31 y 32), como corresponde a una auténtica secreción holocrina. Hay también glándulas independientes de los folícu los como las de Meibomio en párpados, de Tyson en prepu-cio, los tubérculos de Montgomery de la aréola mamaria y las de labios menores femeninos.
Durante la primera semana de vida se produce un aumento de la se-creción sebácea, que estaría mediado por los andrógenos maternos y por la síntesis esteroidea endógena, que lógicamente desaparecerá después. En la adrenarquia se produce un nuevo aumento de dicha secreción, que ya continúa hasta alcanzar los niveles del adulto.
tes a la queratohialina de la epidermis, pero no melanina. Acom-paña al pelo hasta la zona del folículo entre la inserción del mús-cu lo erector y la glándula sebácea, conocida como «zona deStraile», y allí se desintegra. Desde esta zona sólo queda ya porfuera la vaina epitelial externa, la membrana vítrea y el saco con-juntivo.
El ciclo de crecimiento del pelo se divide en tres fases:anagen, donde se produce el crecimiento activo delpelo, catagen, que es una fase regresiva, y telogen o fasede reposo. Después, el folículo entra en una nueva fasede anagen y el pelo antiguo es reemplazado por unonuevo. El anagen tiene seis estadios; el estadio V, quees en el que el pelo nuevo elimina al que está en el inte-rior, se denomina fase de «exogen» o teloptosis (Fig.29).
Hay dos familias de queratinas del pelo, tipos I y II, exis-tiendo cuatro proteínas principales en cada familia (Ha 1-4 yHb 1-4) y otras menos importantes hasta completar las 18existentes. Además, 9 de las 37 queratinas epiteliales seexpresan específicamente en el folículo piloso, por lo que elnúmero total de queratinas del folículo piloso es de 26.
La vascularización del folículo se hace a expensas de unared anastomótica de dos plexos distintos; la zona superior senutre del subpapilar y la inferior del hipodérmico. En los folí-cu los en anagen, los vasos sanguíneos se extienden paralelosal folículo, es decir, orientados longitudinalmente desde labase del bulbo al canal del pelo, y entre ellos se produce unared por entrecruzamiento que vasculariza la parte inferior delfolículo incluyendo el bulbo. En la parte media del folículo haypocas interconexiones vasculares mientras que sí las hayabundantes alrededor de la glándula sebácea. Por encima dela desembocadura del conducto sebáceo hay unos vasos para-
lelos que ascienden hacia el canal piloso y se continúan conlas ramas capilares situadas por debajo de la epidermis. En labase de los folículos en anagen se sitúan las arteriolas per-pendiculares formando la empalizada que acompaña el folí-culo en toda su longitud. Algunas de estas arteriolas penetranen la papila dérmica por una zona donde se encuentra el«cuerpo de Arao-Perkins» que es un órgano de soporte para lapapila dérmica y para los vasos sanguíneos que la penetran,constituido por el cuerpo laminado de la placa basal y los fila-mentos que se proyectan en la papila. En el interior de lapapila, los vasos sanguíneos forman ovillos o lazos.
La inervación del folículo piloso se hace por fibras mielíni-cas (una fibra mielínica cutánea inerva entre 40-120 folícu-los), que llegan a la altura de la glándula sebácea, perdiendoallí la vaina mielínica y constituyendo alrededor del folículoun doble collar amielínico (interno longitudinal y paralelo alpelo, y externo circular), del que salen fibras para la unióndermoepidérmica, músculo erector y glándulas sebáceas yecrinas. Independientemente del tamaño, cada folículo estárodeado por numerosas fibras nerviosas desde la base delbulbo a su unión con la epidermis. Algunos nervios mieliniza-dos discurren paralelos a la región permanente del folículo,otros nervios más finos forman una red a modo de calcetín obolsa que rodea el resto del folículo. En la cercanía del canalpiloso se puede observar un cúmulo de fibras nerviosas mieli-nizadas constituyendo una «cesta», «collar» o «empalizada».La disposición de estos nervios, mejor organizados alrededorde los folículos de vellos que en los grandes folículos produc-tores de pelos terminales, permite considerarlos como un«órgano táctil folicular».
UñasEn la estructura de la uña, actualmente se prefiere hablar delconcepto de unidad ungueal, refiriéndonos a toda la porcióndorsodistal de los dedos, donde se encuentran la láminaungueal, matriz, lecho y pliegues proximal, laterales y distal(Fig. 30). En dirección proximal a distal del dedo observamos
1.A
nato
mía
y f
isio
logí
a de
la p
iel
21
Figura 29. Fases de crecimiento del pelo.
Anagenmaduro Catagen Telogen
Anagenprecoz
Anagenmaduro
Figura 30. Unidad ungueal.
Borde libreLámina ungueal
Lúnula
Lámina ungueal Hiponiquio
Matrizungueal
Eponiquio
Pliegue proximal
CutículaMatriz
ungueal
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 21
Figura 30. Unidad ungueal.
Figura 31. Detalle histológico de la glándula sebácea.
Figura 32. Detalle de la glándula sebácea y su relación con el folículo piloso.
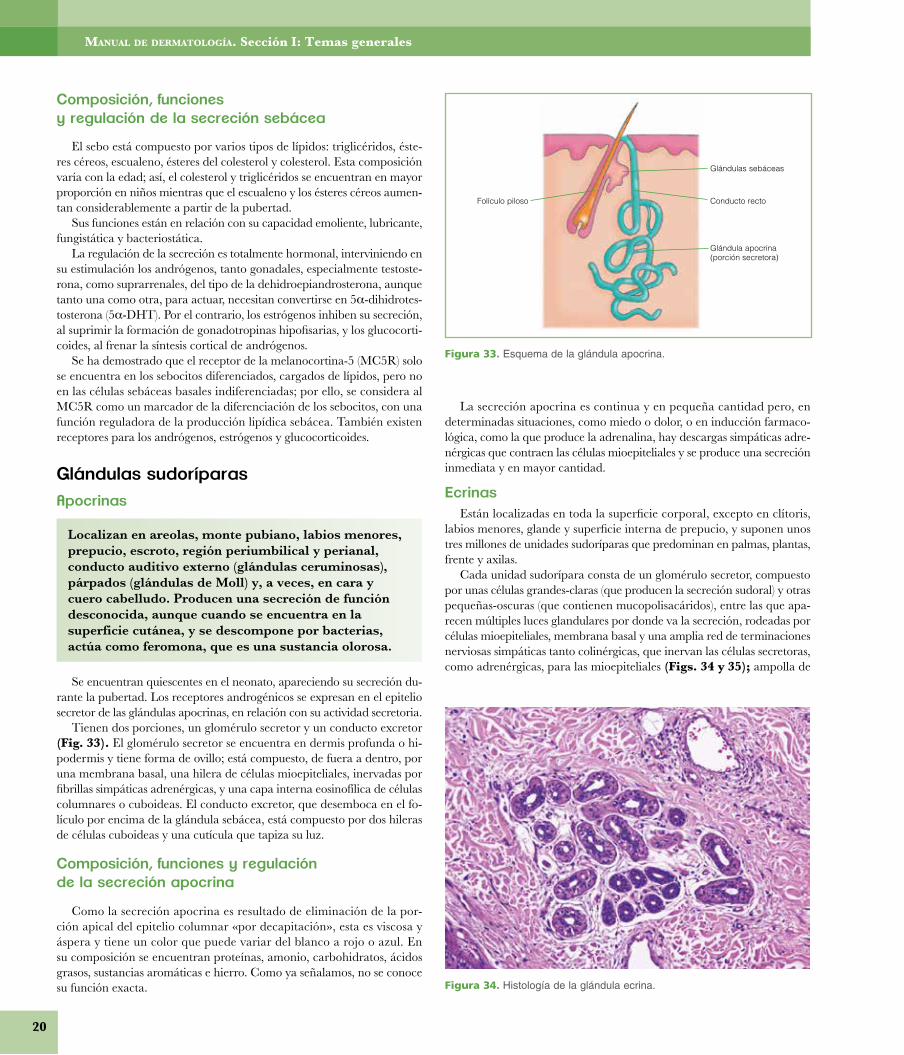
20
Manual de derMatología. Sección I: Temas generales
Composición, funciones y regulación de la secreción sebácea
El sebo está compuesto por varios tipos de lípidos: triglicéridos, éste-res céreos, escualeno, ésteres del colesterol y colesterol. Esta composición varía con la edad; así, el colesterol y triglicéridos se encuentran en mayor proporción en niños mientras que el escualeno y los ésteres céreos aumen-tan considerablemente a partir de la pubertad.
Sus funciones están en relación con su capacidad emoliente, lubricante, fungistática y bacteriostática.
La regulación de la secreción es totalmente hormonal, interviniendo en su estimulación los andrógenos, tanto gonadales, especialmente testoste-rona, como suprarrenales, del tipo de la dehidroepiandrosterona, aunque tanto una como otra, para actuar, necesitan convertirse en 5a-dihidrotes-tosterona (5a-DHT). Por el contrario, los estrógenos inhiben su secreción, al suprimir la formación de gonadotropinas hipofisarias, y los glucocorti-coides, al frenar la síntesis cortical de andrógenos.
Se ha demostrado que el receptor de la melanocortina-5 (MC5R) solo se encuentra en los sebocitos diferenciados, cargados de lípidos, pero no en las células sebáceas basales indiferenciadas; por ello, se considera al MC5R como un marcador de la diferenciación de los sebocitos, con una función reguladora de la producción lipídica sebácea. También existen receptores para los andrógenos, estrógenos y glucocorticoides.
Glándulas sudoríparas
Apocrinas
Localizan en areolas, monte pubiano, labios menores, prepucio, escroto, región periumbilical y perianal, conducto auditivo externo (glándulas ceruminosas), párpados (glándulas de Moll) y, a veces, en cara y cuero cabelludo. Producen una secreción de función desconocida, aunque cuando se encuentra en la superficie cutánea, y se descompone por bacterias, actúa como feromona, que es una sustancia olorosa.
Se encuentran quiescentes en el neonato, apareciendo su secreción du-rante la pubertad. Los receptores androgénicos se expresan en el epitelio secretor de las glándulas apocrinas, en relación con su actividad secretoria.
Tienen dos porciones, un glomérulo secretor y un conducto excretor (Fig. 33). El glomérulo secretor se encuentra en dermis profunda o hi-podermis y tiene forma de ovillo; está compuesto, de fuera a dentro, por una membrana basal, una hilera de células mioepiteliales, inervadas por fibrillas simpáticas adrenérgicas, y una capa interna eosinofílica de células columnares o cuboideas. El conducto excretor, que desemboca en el fo-lículo por encima de la glándula sebácea, está compuesto por dos hileras de células cuboideas y una cutícula que tapiza su luz.
Composición, funciones y regulación de la secreción apocrina
Como la secreción apocrina es resultado de eliminación de la por-ción apical del epitelio columnar «por decapitación», esta es viscosa y áspera y tiene un color que puede variar del blanco a rojo o azul. En su composición se encuentran proteínas, amonio, carbohidratos, ácidos grasos, sustancias aromáticas e hierro. Como ya señalamos, no se conoce su función exacta.
La secreción apocrina es continua y en pequeña cantidad pero, en determinadas situaciones, como miedo o dolor, o en inducción farmaco-lógica, como la que produce la adrenalina, hay descargas simpáticas adre-nérgicas que contraen las células mioepiteliales y se produce una secreción inmediata y en mayor cantidad.
EcrinasEstán localizadas en toda la superficie corporal, excepto en clítoris,
labios menores, glande y superficie interna de prepucio, y suponen unos tres millones de unidades sudoríparas que predominan en palmas, plantas, frente y axilas.
Cada unidad sudorípara consta de un glomérulo secretor, compuesto por unas células grandes-claras (que producen la secreción sudoral) y otras pequeñas-oscuras (que contienen mucopolisacáridos), entre las que apa-recen múltiples luces glandulares por donde va la secreción, rodeadas por células mioepiteliales, membrana basal y una amplia red de terminaciones nerviosas simpáticas tanto colinérgicas, que inervan las células secretoras, como adrenérgicas, para las mioepiteliales (Figs. 34 y 35); ampolla de
Figura 34. Histología de la glándula ecrina.
Glándulas sebáceas
Conducto rectoFolículo piloso
Glándula apocrina (porción secretora)
Figura 33. Esquema de la glándula apocrina.

21
Capítulo 1. Anatomía y fisiología de la piel
Loewenthal, que es un ensanchamiento fusiforme que se encuentra entre la porción secretora y el conducto excretor, donde las células fusiformes forman un esfínter; conducto excretor dérmico, longitudinal y formado por dos capas de células cuboides basófilas y una cutícula tapizando su luz; y conducto excretor epidérmico o acrosiringio, en forma de espiral, lo que le permite adaptarse a las modificaciones epiteliales, ya sean de atrofia o de acantosis, constituido por queratinocitos que se queratinizan de forma independiente a los propiamente epiteliales, a los que están unidos por desmosomas.
Composición, funciones y regulación de la secreción ecrina
Es una secreción hipotónica, inodora e incolora, de pH ácido (4,5 a 5,5), compuesta por agua (un 99%) y sodio, potasio, cloro, urea, proteínas, lípidos, aminoácidos, calcio, fósforo e hierro, aunque todos los electrolitos del plasma se encuentran en mayor o menor proporción.
Tiene como funciones mantener el pH de la superficie cutánea y la tem-peratura corporal. El principal estímulo para la secreción sudoral es el calor (un aumento de temperatura en sangre pone en marcha los mecanismos estimuladores hipotalámicos), aunque hay otros mecanismos nerviosos que producen respuestas localizadas, como el sudor reflejo facial o gustativo (con los alimentos picantes), sudor psíquico (el que se produce con pensar que al salir a la calle va a exponerse a una temperatura elevada), en tensiones emo-cionales (como miedo o dolor, que localiza fundamentalmente en palmas de manos), por reflejo axónico (como sucede en úlceras), etc.
Se ha demostrado la expresión de receptores para mineralocorticoides, que median las acciones de la aldosterona en el balance hidrosalino cor-poral, en los conductos de las glándulas ecrinas.
Su «regulación» es nerviosa, mediante fibras simpáticas, segregando sudor de modo intermitente bajo estímulos colinérgicos, mediante recep-tores muscarínicos, y, en situaciones de tensión psíquica, por contracción adrenérgica de las células mioepiteliales, vaciando totalmente la glándula.
Aunque el volumen de secreción individual es mínimo, en conjunto es muy grande y puede llegar a poner en peligro la vida en climas tórridos, pues en estas situaciones alcanza los tres litros por hora y los doce por día, y no hay que olvidar que cada litro de sudor evaporado elimina 540 calorías.
MANTO CUTÁNEO ÁCIDO LIPÍDICOSupone la unión de las secreciones epidérmicas y anexiales que recu-
bren, hidratan y protegen la epidermis.
Se llama así porque dominan los lípidos cutáneos y sebáceos, de función hidratante y lubricante respectivamente, y el pH ácido del sudor ecrino (de 5,5), que también protege de irritaciones primarias y sensibilizaciones.
Este manto sería una emulsión oleoacuosa donde el agua proviene del sudor ecrino y la retención de este agua se efectúa gracias al componente oleoso, especialmente epitelial (colesterol y sus ésteres).
Las funciones del manto ácido-lipídico son las siguientes:
a) Hidratación de la piel. La retención de agua en el estrato cór-neo depende principalmente de dos componentes: 1) la presencia de agentes higroscópicos naturales dentro de los corneocitos, colectiva-mente denominados como «factor hidratante natural», y 2) los lípidos intercelulares situados ordenadamente para formar una barrera a la pérdida de agua transepidérmica. Además, las fibras de queratina de los corneocitos tienen propiedades hidrofílicas y contienen también una fracción soluble en agua que permite aumentar su capacidad para retenerla.
– La composición de los lípidos de la epidermis varía dependiendo de diversos factores: i) Diferenciación epitelial: el estrato córneo con-tiene fundamentalmente ceramidas, esteroles libres y ácidos grasos libres, que son hidrofílicos. ii) Estación del año: en invierno y prima-vera las proporciones de ceramidas, colesterol y ácidos grasos son mucho más reducidas que en verano, y especialmente reducidas en invierno las ceramidas esterificadas a ácido linoleico. iii) Región del organismo, sexo y edad.
– Los lípidos hidrofílicos dan «plasticidad» o «reblandecen» la super-ficie cutánea y regulan la deshidratación al formar emulsiones con el agua. Pero determinadas situaciones, como la excesiva humedad, por aumento de secreción ecrina o contacto prolongado con agua, hacen que esta adopte un aspecto blanco-lechoso característico de la hidratación excesiva. Y la escasa producción lipídica, sequedad am-biental o empleo de de ter gen tes/sol ven tes orgánicos, que destruyen los lípidos cutáneos, la resecan tornándola escamosa o agrietada.
– El «factor hidratante natural» (FHN), que supone el 15-20% del peso total del estrato córneo, es una mezcla compleja de iones y sustancias de bajo peso molecular, solubles en agua. La mayor parte de este FHN está formado por la degradación de la filagrina, pro-teína rica en histidina. Su composición final, que se expone en la Tabla IV, es a base de aminoácidos o sus derivados, como ácido pirrolidon-carboxílico (PCA) y ácido urocánico, ácido láctico, urea, citrato y azúcares. La señal que inicia la proteólisis de la filagrina es el gradiente de agua dentro del estrato córneo, ya que la deshidrata-ción desencadena la conversión de filagrina en FHN. No obstante, el lactato deriva de las glándulas ecrinas.
– El contacto prolongado con el agua y los jabones pueden reducir el FHN y alterar las propiedades de hidra tación de la capa córnea. También existen cambios estacionales, del verano al invierno, en
las células secretoras, como adrenérgicas, para las mioepite-liales (Figs. 34 y 35); ampolla de Loewenthal, que es unensanchamiento fusiforme que se encuentra entre la porciónsecretora y el conducto excretor, donde las células fusiformesforman un esfínter; conducto excretor dérmico, longitudinal yformado por dos capas de células cuboides basófilas y unacutícula tapizando su luz; y conducto excretor epidérmico oacrosiringio, en forma de espiral, lo que le permite adaptarsea las modificaciones epiteliales, ya sean de atrofia o de acan-tosis, constituido por queratinocitos que se queratinizan deforma independiente a los propiamente epiteliales, a los queestán unidos por desmosomas.
Composición, funciones y regulación de la secreción ecrina
Es una secreción hipotónica, inodora e incolora, de pH ácido(4,5 a 5,5), compuesta por agua (un 99%) y sodio, potasio,cloro, urea, proteínas, lípidos, aminoácidos, calcio, fósforo ehierro, aunque todos los electrólitos del plasma se encuen-tran en mayor o menor proporción.
Tiene como funciones mantener el pH de la superficiecutánea y la temperatura corporal. El principal estí-mulo para la secreción sudoral es el calor (un aumentode temperatura en sangre pone en marcha los mecanis-mos estimuladores hipotalámicos), aunque hay otrosmecanismos nerviosos que producen respuestas locali-zadas, como el sudor reflejo facial o gustativo (con losalimentos picantes), sudor psíquico (el que se producecon pensar que al salir a la calle va a exponerse a unatemperatura elevada), en tensiones emocionales (comomiedo o dolor, que localiza fundamentalmente en pal-mas de manos), por reflejo axónico (como sucede enúlceras), etc.
Se ha demostrado la expresión de receptores para minera-locorticoides, que median las acciones de la aldosterona en elbalance hidrosalino corporal, en los conductos de las glándu-las ecrinas.
Su «regulación» es nerviosa, mediante fibras simpáticas,segregando sudor de modo intermitente bajo estímulos coli-nérgicos, mediante receptores muscarínicos, y, en situacionesde tensión psíquica, por contracción adrenérgica de las célu-las mioepiteliales, vaciando totalmente la glándula.
Aunque el volumen de secreción individual es mínimo, enconjunto es muy grande y puede llegar a poner en peligro lavida en climas tórridos, pues en estas situaciones alcanza lostres litros por hora y los doce por día, y no hay que olvidarque cada litro de sudor evaporado elimina 540 calorías.
MANTO CUTÁNEO ÁCIDO LIPÍDICO
Supone la unión de las secreciones epidérmicas y anexialesque recubren, hidratan y protegen la epidermis.
Se llama así porque dominan los lípidos cutáneos ysebáceos, de función hidratante y lubricante respecti-vamente, y el pH ácido del sudor ecrino (de 5,5), quetambién protege de irritaciones primarias y sensibili-zaciones.
Este manto sería una emulsión oleoacuosa donde el aguaproviene del sudor ecrino y la retención de este agua se efec-túa gracias al componente oleoso, especialmente epitelial(colesterol y sus ésteres).
Las funciones del manto ácido-lipídico son las siguientes:
a) Hidratación de la piel. La retención de agua en el estratocórneo depende principalmente de dos componentes: 1)la presencia de agentes higroscópicos naturales dentro de
Tem
as g
ener
ales
24
Figura 35. Esquema de la glándula ecrina.
Epidermis
Nervio simpáticocolinérgico
Conductoexcretor
Dermis
Figura 34. Histología de la glándula ecrina.
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:31 Página 24
Figura 35. Esquema de la glándula ecrina.

22
Manual de derMatología. Sección I: Temas generales
las propiedades físicas de la capa córnea asociándose una disminu-ción significativa de los niveles de lactato, potasio, sodio y cloro en el FHN. Por otro lado, se ha identificado en los queratinocitos la acuaporina-3, que es una proteína transmembrana transportadora de agua, relacionada con el glicerol, el cual se ha encontrado en la capa córnea actuando como un humectante endógeno natural. Ambos intervienen en la hidratación de la epidermis humana.
b) Permeabilidad o función barrera de la piel. Los lípidos segregados por el epitelio se disponen en forma de «mosaico» que impide la pérdi-da de agua a través del epitelio y permite la hidratación de los corneo-citos. La integridad del estrato córneo va a determinar el porcentaje de la «pérdida transepidérmica de agua», que se conoce con el acrónimo inglés de TEWL. Esta TEWL muestra variaciones entre los distintos individuos y las diferentes partes del cuerpo. Otra función importante de la piel es «impedir la entrada de agentes ambientales»; no obstante, una sustancia logrará penetrar el epitelio dependiendo de la integridad de la capa córnea y del peso molecular y lipofilia que posea.
– En cualquier caso, la permeabilidad de la piel dependerá de cua-tro factores: 1) contenido de lípidos en el estrato córneo, 2) grado de hidratación de las capas cutáneas, 3) tamaño de los corneocitos, 4) grosor del estrato córneo. De ellos, el más importante en la «fun-ción barrera» son los lípidos.
c) Acción bacteriostática/bactericida. Para dicha acción es nece-saria la presencia de todo el manto ácido-lipídico ya que los lípidos sebáceos tienen propiedades antibacterianas y los glucofosfolípidos y los ácidos grasos libres del estrato córneo tienen efectos bacteriostáti-cos. Sin duda, también influye el pH ácido y, de su acción conjunta, se logra eliminar de la superficie cutánea a Streptococcus pyogenes en un día y a Staphylococcus aureus en tres.
d) Acción fungistática. Propia de los lípidos sebáceos, puesto que a partir de la pubertad desaparece la tinea capitis, ya que en cuero cabe-lludo hay muchas glándulas sebáceas.
HIPODERMIS
También conocida como tejido celular subcutáneo o panículo adiposo, está constituida por lipocitos o adipocitos que son células encargadas de fabricar y almacenar grasas por lo que, al ir llenándose del material lipídico, van rechazando su núcleo a la periferia adoptando el aspecto de «células en anillo de sello» (Fig. 36).
Estas células adiposas se disponen en microlóbulos primarios, que a su vez se agregan en lóbulos secundarios, de mayor tamaño, que están rodeados de tejido fibroso, en forma de trabéculas, por donde discurren las arterias, venas, linfáticos y nervios. Los septos fibrosos que rodean a los microlóbulos no son visibles en las secciones histológicas habituales.
Cada adipocito está rodeado de un capilar para facilitar el transporte de los productos de la lipólisis, ácidos grasos y glicerol, a la circulación general. El aporte vascular de cada microlóbulo es terminal, no existiendo capilares que atraviesen los septos entre microlóbulos adyacentes.
El espesor de los lóbulos varía según las regiones corporales. Así, por ejemplo, en abdomen, muslos, palmas y plantas hay una hipodermis muy densa, mientras que en labios menores vulvares prácticamente no existen. En los adultos también hay una «configuración ginoide» y otra «configu-ración androide» de la hipodermis. Microscópicamente, los lóbulos de las mujeres son más grandes y más rectangulares mientras que los de los hombres son más pequeños y más poligonales.
Las funciones de la hipodermis son: 1) mantener la temperatura orgá-nica actuando como aislante, 2) proteger frente a los traumatismos mecá-nicos, 3) servir de reserva y depósito de calorías, en forma de triglicéridos, y su liberación, en forma de ácidos grasos no esterificados, cuando sea necesario un aporte de energía (Fig. 37). Las zonas que normalmente no se afectan cuando el individuo está hipoalimentado son las que cumplen misiones de protección como, por ejemplo, palmas y plantas.
Tabla IV. Composición del factor hidratante natural (FHN)
Elementos Porcentaje
Aminoácidos 40%
Ácido pirrolidon carboxílico 12%
Lactato 12%
Azúcares 8,5%
Urea 7%
Cloro 6%
Sodio 5%
Potasio 4%
NH3, ácido úrico, glucosamina y creatina 1,5%
Calcio 1,5%
Magnesio 1,5%
Fosfato 0,5%
Citrato y formato 0,5%
Figura 36. Esquema del adipocito.

23
Capítulo 1. Anatomía y fisiología de la piel
Por debajo de la hipodermis se encuentra la fascia profunda que está constituida por tejido fibroso. En algunas zonas del cuerpo pueden ob-servarse músculos en el interior de la hipodermis: cara (músculo de la expresión), cuello (platisma), escroto (dartros) y areola mamaria.
VASOS Y NERVIOS CUTÁNEOS
Sistema vascular de la piel
Desde un punto de vista topográfico hay tres plexos vasculares en la piel: subpapilar, dermohipodérmico y subcutáneo, de menor a mayor grosor y con numerosas interconexiones entre ellos (Fig. 38), lo que permite la importante función del control de la temperatura corporal.
La pared de los capilares está constituida a veces por una sola célula endotelial, con un citoplasma muy elongado que margina la luz. A su alre-dedor se encuentra el peritelio, capa discontinua y unicelular de pequeñas células llamadas pericitos.
Las arteriolas poseen una capa muscular lisa que llega a formar un esfínter precapilar y determina que su luz sea redondeada. Regulan el aporte sanguíneo a la piel, conduciendo la sangre desde las arterias me-diante contracciones o dilatación de sus paredes y esfínter precapilar, por lo que tienen gran trascendencia en la termorregulación.
Las vénulas, de diámetro mayor, luz fusiforme y pared más delgada que las arteriolas, al estar constituida por fibras conjuntivas, tienen como misión retornar la sangre. En su revestimiento periendotelial existen más de un pericito y no contienen células musculares lisas. Solo algunas volu-minosas venas tienen musculatura lisa (Fig. 39).
Los glomus son anastomosis arteriovenosas que se encargan de permi-tir la rápida circulación entre estos vasos, por lo que representan un factor importantísimo en la regulación térmica. Su estructura es entre arteriola y vénula y están rodeados de células de aspecto epitelioide (células glómicas) que, en realidad, son de naturaleza muscular y funcionan como esfínter. La parte arterial del glomus se llama «canal de Suquet-Hoyer».
La inervación de los vasos cutáneos está representada por finas fibras que van por las paredes de las arteriolas, incluyendo las glómicas, y no inervan capilares y vénulas. En consecuencia, las paredes arteriolares van a responder a gran variedad de estímulos, aunque normalmente están en contracción tónica debido a los impulsos nerviosos simpáticos. De elimi-narse estos impulsos se produciría vasodilatación como resultado de la presión intravascular. Todo lo anterior explica que, por ejemplo, en situa-ciones de estrés intenso, por excitación masiva del simpático, se produzca vasoconstricción generalizada y palidez cutánea.
No hay fibras vasodilatadoras para las arteriolas; sin embargo, pue-de producirse vasodilatación cuando actúan sobre ellas reflejos axónicos locales. Es decir, que los nervios sensitivos liberan acetilcolina en vez de adrenalina, aunque no se conoce bien el por qué de esta reacción. Los músculos de la pared arteriolar pueden también ser estimulados por el frío y el calor; el primero actuaría produciendo vasocons tricción, y des-pués, cuando el enfriamiento es muy intenso, se estimulan los centros hipotalámicos que aumentan los impulsos autónomos vasoconstrictores. Si el enfriamiento es acentuado ocurre un reflejo axónico que determina vasodilatación, con lo que ejerce una acción protectora, reduciendo las lesiones que se pudieran producir en los tejidos como consecuencia de la baja temperatura.
estrato córneo tienen efectos bacteriostáticos. Sin duda,también influye el pH ácido y, de su acción conjunta, selogra eliminar de la superficie cutánea a Streptococcuspyogenes en un día y a Staphylococcus aureus en tres.
d) Acción fungistática. Propia de los lípidos sebáceos, puestoque a partir de la pubertad desaparece la tinea capitis, yaque en cuero cabelludo hay muchas glándulas sebáceas.
HIPODERMIS
También conocida como tejido celular subcutáneo opanículo adiposo, está constituida por lipocitos o adi-pocitos que son células encargadas de fabricar y alma-cenar grasas por lo que, al ir llenándose del materiallipídico, van rechazando su núcleo a la periferiaadoptando el aspecto de «células en anillo de sello»(Fig. 36).
Estas células adiposas se disponen en microlóbulos prima-rios, que a su vez se agregan en lóbulos secundarios, demayor tamaño, que están rodeados de tejido fibroso, enforma de trabéculas, por donde discurren las arterias, venas,linfáticos y nervios. Los septos fibrosos que rodean a losmicrolóbulos no son visibles en las secciones histológicashabituales.
Cada adipocito está rodeado de un capilar para facilitar eltransporte de los productos de la lipólisis, ácidos grasos y gli-cerol, a la circulación general. El aporte vascular de cadamicrolóbulo es terminal, no existiendo capilares que atravie-sen los septos entre microlóbulos adyacentes.
El espesor de los lóbulos varía según las regiones corpora-les. Así, por ejemplo, en abdomen, muslos, palmas y plantashay una hipodermis muy densa, mientras que en labios meno-res vulvares prácticamente no existen. En los adultos tambiénhay una «configuración ginoide» y otra «configuraciónandroide» de la hipodermis. Microscópicamente, los lóbulosde las mujeres son más grandes y más rectangulares mientrasque los de los hombres son más pequeños y más poligonales.
Las funciones de la hipodermis son: 1) mantener la tempe-ratura orgánica actuando como aislante, 2) proteger frente alos traumatismos mecánicos, 3) servir de reserva y depósitode calorías, en forma de triglicéridos, y su liberación, enforma de ácidos grasos no esterificados, cuando sea necesa-rio un aporte de energía (Fig. 37). Las zonas que normalmenteno se afectan cuando el individuo está hipoalimentado sonlas que cumplen misiones de protección como, por ejemplo,palmas y plantas.
Por debajo de la hipodermis se encuentra la fascia pro-funda que está constituida por tejido fibroso. En algunaszonas del cuerpo pueden observarse músculos en el interiorde la hipodermis: cara (músculo de la expresión), cuello (pla-tisma), escroto (dartros) y areola mamaria.
Tem
as g
ener
ales
26
Figura 37. Funciones del adipocito.
Factorestisulares
Leptina
Estrógeno
Adiposina
Otros
Agutina
Visfatina
Adiponectina
Resistina
Citocinas
Figura 38. Plexo vascular de la piel.
Figura 39. Detalle histológico de una arteria de la piel.
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:32 Página 26
Figura 37. Funciones del adipocito.
Figura 38. Plexo vascular de la piel. Figura 39. Detalle histológico de una arteria de la piel.

24
Manual de derMatología. Sección I: Temas generales
La respuesta al calor local es la vasodilatación que, si llega a ser sufi-ciente como para aumentar la temperatura sanguínea, inhibe los impulsos vasoconstrictores simpáticos, produciendo vasodilatación generalizada. También hay sustancias químicas, como la histamina, que provocan va-sodilatación localizada.
La piel también posee vasos linfáticos que, al ser de estrechísima luz tapizada por una delgada pared endotelial, no se observan al microscopio óptico. Sirven para transportar partículas y materiales líquidos desde la matriz extracelular dérmica.
La red linfática muestra variaciones según edad, grosor de la piel o localización, aunque el plexo dérmico es uniforme en toda la dermis. Hay una red subpapilar de vasos linfáticos interconectados entre sí de una forma hexagonal y, a través de unos precolectores, con los linfáticos de la dermis profunda. Desde allí pasan a la hipodermis e interconexionan con los existentes por debajo de la fascia, que son ya mucho más grue-sos pero con menor densidad de red. A partir de aquí, ya los linfáticos son contráctiles, pues tienen músculo liso, y ya se vehiculizan los fluidos linfáticos a través de colectores a los ganglios linfáticos y a la circulación central; es decir, al conducto torácico. Los linfáticos cutáneos no solo es-tán asociados a arteriolas, como lo hacen en el músculo esquelético, sino también a los folículos pilosebáceos. Tienen una porción terminal ciega y las dimensiones de su luz son más grandes que las de los capilares ya que miden unas 50 micras.
En la piel no hay linfáticos contráctiles, sino que estos drenan en los contráctiles más profundos. Tienen una luz irregular tapizada por células endoteliales muy delgadas con membrana basal discontinua y uniones celulares no contiguas. Tienen dos tipos de válvulas, unas a nivel de las uniones interendoteliales en la pared de los linfáticos y otras, en forma de válvulas intralinfáticas, en la luz. Los linfáticos colectores contráctiles tienen músculo liso y peristaltismo, que es lo que hace que, a partir de ellos, la linfa se mueva independientemente.
Nervios cutáneosLa piel es un exquisito órgano sensorial, pues mediante su inervación
permite la relación del hombre con el mundo exterior. En efecto, gracias a las fibras sensitivas, mielínicas o amielínicas, que llegan a las raíces me-
dulares dorsales y de allí al cerebro, transmiten las sensaciones de tacto, vibración, presión, temperatura, dolor y prurito (Figs. 40 y 41).
Y a través de fibras motoras, procedentes de los ganglios simpáticos, inervan diversas estructuras cutáneas; la mayoría son motoras adrenér-gicas, que activan arteriolas, porción arteriolar del «glomus», músculos erectores y glándulas sudo ríparas, mientras que las colinérgicas solo ac-túan en las glándulas sudoríparas ecrinas.
Los troncos nerviosos gruesos penetran en la hipodermis donde se dividen en nervios más pequeños que van ascendiendo, generalmente acompañando a los vasos sanguíneos, hasta la dermis superficial; algunos penetran en la zona de la unión dermo-epidérmica, pero ninguno atra-viesa hasta la epidermis.
Nervios sensorialesLas sensaciones de tacto, presión, vibración, temperatura, dolor y pru-
rito son recibidas de dos formas principales: los elementos corpusculares, que pueden contener tejido no nervioso, y las terminaciones nerviosas libres. Las terminaciones corpusculares pueden ser subdivididas en recep-
lar del «glomus», músculos erectores y glándulas sudo ríparas,mientras que las colinérgicas sólo actúan en las glándulassudoríparas ecrinas.
Los troncos nerviosos gruesos penetran en la hipodermisdonde se dividen en nervios más pequeños que van ascen-diendo, generalmente acompañando a los vasos sanguíneos,hasta la dermis superficial; algunos penetran en la zona de launión dermo-epidérmica, pero ninguno atraviesa hasta la epi-dermis.
Nervios sensorialesLas sensaciones de tacto, presión, vibración, temperatura,dolor y prurito son recibidas de dos formas principales: los
elementos corpusculares, que pueden contener tejido no ner-vioso, y las terminaciones nerviosas libres. Las terminacionescorpusculares pueden ser subdivididas en receptores encap-sulados, dérmicos, y los receptores no encapsulados, comolas células de Merkel. Los nervios sensoriales pueden serfibras A mielínicas, que son más gruesas, o fibras C amielíni-cas de conducción lenta.
� Receptores sensoriales libres. Son recibidos por del-gadas fibras amielínicas que terminan en dermis papilar yunión dermoepidérmica, en forma de penachos, y alrede-dor de los folículos pilosos. Se encargan de recibir las sen-saciones de tacto, temperatura, dolor y prurito. Esteúltimo es transmitido al SNC por las fibras C (fibras amielí-nicas lentas) mientras que el dolor lo hace también por lasfibras A mielínicas.Estos receptores libres son más numerosos en zonas depiel lampiña como palmas, plantas y dedos y en áreasmucocutáneas erógenas: boca, labios menores, clítoris,pene y pezones. En las zonas pilosas, los mecanorrecep-tores predominantes son los receptores de los folículospilosos.En cuanto a la temperatura, que antes se pensaba transmi-tida por los corpúsculos de Krause (frío) y órganos de Ru f-fini (calor), hoy está claro que también se recibe a través determinaciones libres, existiendo en la superficie cutáneadeterminados «puntos de calor y frío», que son termore-ceptores, que pueden cambiar en situaciones patológicas.El dolor es transmitido por los nociceptores, que respon-den selectivamente a estímulos que pueden provocar dañotisular. Hay tres tipos de nociceptores, los mecánicos, lostérmicos y los polimodales (responder a varios tipos deestímulos).
� Receptores encapsulados. Estos corpúsculos sensorialesreciben las sensaciones de tacto, presión y vibración. Eltacto es captado por los corpúsculos de Meissner, que
Tem
as g
ener
ales
28
Figura 41. Diferentes tpos de terminaciones nerviosas en la piel. C, Vater-Pacini; D, Meissener; E, Krause; F, Ruffini.
Terminaciones libresdel axón aferente
Disco terminaldel axónaferente
Célula deMerkel
Terminal delaxón
aferente
Cápsulamulticapa
Ramas terminalesdel axón aferente
A B C
D E F
Cápsula
Terminalesespirales del
axón aferente
Papila dérmica
Células deSchwanntortuosas
Cápsula
Ramasterminales delaxón aferente
Cápsula
Figura 40. Terminaciones nerviosas en la piel. Corpúsculos cutá-neos.
Terminación libre
Meissner
Krause
Pacini
Ruffini
Epidermis
01_anatomia.qxp:_Dermatologia_01.qxd 3/12/09 11:32 Página 28
Figura 40. Terminaciones nerviosas en la piel. Corpúsculos cutáneos.
Terminaciones libres del axón aferente
A
Disco terminal del axón aferente
Célula de Merkel
B
Terminales espirales del axón aferente
Papila dérmica
Células de Schwann tortuosas
Cápsula
E
Ramas terminales del axón aferente
Cápsula
D Ramas terminales del axón aferente
Cápsula
F
Terminal del axón aferente
Cápsula multicapa
C
Figura 41. Diferentes tipos de terminaciones nerviosas en la piel. C. Vater-Pacini; D. Meissner; E. Krause; F. Ruffini.

25
Capítulo 1. Anatomía y fisiología de la piel
tores encapsulados, dérmicos, y los receptores no encapsulados, como las células de Merkel. Los nervios sensoriales pueden ser fibras A mielínicas, que son más gruesas, o fibras C amielínicas de conducción lenta.
• Receptores sensoriales libres. Son recibidos por delgadas fibras amielínicas que terminan en dermis papilar y unión dermoepidérmica, en forma de penachos, y alrededor de los folículos pilosos. Se encargan de recibir las sensaciones de tacto, temperatura, dolor y prurito. Este último es transmitido al SNC por las fibras C (fibras amielínicas lentas) mientras que el dolor lo hace también por las fibras A mielínicas.
Estos receptores libres son más numerosos en zonas de piel lampi-ña como palmas, plantas y dedos y en áreas mucocutáneas erógenas: boca, labios menores, clítoris, pene y pezones. En las zonas pilosas, los mecanorreceptores predominantes son los receptores de los folículos pilosos.
En cuanto a la temperatura, que antes se pensaba transmitida por los corpúsculos de Krause (frío) y órganos de Ru ffini (calor), hoy está claro que también se recibe a través de terminaciones libres, existiendo en la superficie cutánea determinados «puntos de calor y frío», que son termorreceptores, que pueden cambiar en situaciones patológicas.
El dolor es transmitido por los nociceptores, que responden selecti-vamente a estímulos que pueden provocar daño tisular. Hay tres tipos de nociceptores, los mecánicos, los térmicos y los polimodales (respon-der a varios tipos de estímulos).
• Receptores encapsulados. Estos corpúsculos sensoriales reciben las sensaciones de tacto, presión y vibración. El tacto es captado por los corpúsculos de Meissner, que actuarían como mecanorreceptores de adaptación rápida y están situados en la dermis papilar de las manos y pies; están constituidos por tejido conectivo cilíndrico, dentro del cual ramifican numerosas fibras mielínicas y amielínicas.
La presión y la vibración se reciben en los corpúsculos de Vater-Pa-cini, elementos de gran tamaño que están formados por numerosas laminillas de tejido conjuntivo dispuestas concéntricamente, al igual que una cebolla, alrededor de filamentos nerviosos mielínicos, que lo-calizan en dermis profunda e hipodermis. Los corpúsculos de Ruffini también son grandes y profundos, tienen una estructura fusiforme y actúan como mecanorreceptores de adaptación lenta.
Los corpúsculos de Krause son terminaciones redondeadas de fi-bras mielínicas encapsuladas localizadas en las capas superficiales de la dermis y los corpúsculos de Ru ffini están formados por ramificaciones terminales de una fibra mielínica que están directamente relacionadas con las fibras de colágeno.
• Receptores no encapsulados. Ya hemos comentado que las células de Merkel funcionan como mecanorreceptores de adaptación lenta.
Nervios motoresExisten de tipos muy diversos.
Hay fibras simpáticas adrenérgicas que inervan las arterias, produciendo vasoconstricción, los músculos erectores, provocando su contracción y la consiguiente «cutis anserina» o «piel de gallina», y las células mioepiteliales de las glándulas sudoríparas, determinando la excreción sudoral; y parasimpáticas colinérgicas, que, aunque inervan las glándulas sudoríparas, solo son trascendentes en las ecrinas.
Por tanto, las glándulas sudoríparas ecrinas reciben doble inervación: la colinérgica, que será responsable del sudor en toda la superficie cor-poral, importante en la regulación de la temperatura cutánea, y la adre-nérgica, que determina la hiperhidrosis en los periodos de mayor tensión nerviosa. El hecho de no mencionar las glándulas sebáceas es porque su control no es nervioso sino hormonal.
TENDENCIAS EN LA INVESTIGACIÓN DEL DESARROLLO Y ESTRUCTURA DE LA PIEL
La pérdida de la integridad de la piel y sus funciones puede poner en peligro la vida de los pacientes como ocurre en los grandes quemados. La ingeniería tisular constituye un conjunto de técnicas y métodos de base biotecnológica que permiten diseñar y generar en laboratorio sustitutos tisulares, tejidos artificiales o constructos de origen autólogo o heterólogo a partir de células madre y biomateriales.
La ingeniería tisular representa una gran esperanza para pacientes en espera de una solución a sus problemas orgánicos, lo cual supone un enorme avance en el trasplante de órganos y en la medicina regenerati-va. De hecho, las células madre a utilizar en ingeniería tisular se pueden obtener a partir de pequeñas muestras tisulares (biopsias) obtenidas del propio paciente y, por tanto, los tejidos generados a partir de estas células presentarían carácter autólogo, no existiendo posibilidad de rechazo in-munológico ni ningún tipo de problema ético o legal.
La utilización de piel artificial autóloga supone una alternativa te-rapéutica muy importante para estos pacientes, disminuyendo las com-plicaciones asociadas a la obtención de autoinjertos como las cicatrices, el dolor o riesgo de infección.
La bioingeniería cutánea surgió por la necesidad clínica de pro-porcionar un tratamiento alternativo a los grandes quemados, aunque posteriormente se ha empleado en otras patologías como úlceras cróni-cas, tumores de gran tamaño, fascitis necrotizantes, extirpación de nevus gigantes, enfermedad injerto contra huésped y fines de investigación. El equivalente cutáneo ideal debería reproducir las características fisiológicas de la piel humana normal, ser resistente, fácil de manipular, coste-eficiente y no inducir rechazo inmunológico en el receptor. Se han desarrollado diferentes modelos de equivalentes cutáneos, inicialmente constituidos por láminas de queratinocitos y posteriormente se demostró que la utilización de una estructura o matriz que se comportara como un tejido conectivo mejoraba mucho las características funcionales. Como moldes tridimen-sionales sobre los que se embeben los fibroblastos (dermis) se han utilizado la fibrina, el colágeno tipo I, el ácido poliláctico-poliglicólico, el quitosán, gelatina, dermis acelular y matrices sintéticas. Algunos de estos materiales presentan varios inconvenientes como que pueden ser heterólogos o de origen animal, la contracción de las matrices de colágeno o la dificultad para el manejo de las matrices de fibrina por no ser suficientemente resis-tentes. Además también se han mostrado efectivos los sustitutos dérmicos alogénicos constituidos por una matriz doble de ácido hialurónico y colá-geno con fibroblastos sin incluir células epidérmicas para el tratamiento de las úlceras crónicas refractarias.

26
Manual de derMatología. Sección I: Temas generales
Tabla V. Estructura, función y patología de la piel
Estructura Función Proceso biológico Patología Enfermedad
Capa córnea Protección Descamación Alteración de la descamación Psoriasis
Queratinocitos Síntesis de queratina Queratinogénesis Queratinogénesis alterada Ictiosis
Melanocitos Síntesis de melanina Melanogénesis Aumento o disminución de la melanización
Enfermedad de AddisonAlbinismo
Células de Langerhans Inmunológica Reconocimiento y procesamiento de antígenos
Alteración o proliferación de células de Langerhans
Eczema de contacto alérgicoHistiocitosis langerhansianas
Dermis Protección sosténDepósito de agua
Síntesis de colágenoReacción inflamatoria
Degeneración solar de colágenoInflamación edema
Elastosis solarUrticaria
Vasos TermorregulaciónNutrición
Reacción inflamatoria DilataciónConstricción
Fenómeno de Raynaud
Glándulas sudoríparas ecrinas Termorregulación Secreción de sudor ecrino Aumento o disminución de la producción de sudor
HiperhidrosisAnhidrosis
Glándulas sudoríparas apocrinas
¿? Secreción de sudor apocrino Inflamación de glándulas apocrinas
Hidrosadenitis
Glándulas sebáceas Lubricación Secreción de lípidos Formación de comedones Acné
Pelo Protección Formación de pelo Producción alterada AlopeciaHirsutismo
Hipodermis Aislante de calor Síntesis de grasa Inflamación Eritema nudoso
Aunque todavía no se ha conseguido un sustituto cutáneo perfecto, se ha desarrollado recientemente en laboratorio un sustituto dermo-epi-dérmico que reúne muchas de las características deseadas y que ha de-mostrado su viabilidad clínica (animales de experimentación) para la regeneración cutánea. Esta piel consta de fibroblastos humanos viables embebidos en una matriz de fibrina agarosa sobre los que se siembran los queratinocitos favoreciendo la reparación y la migración celular de origen epitelial y mesenquimal. La fibrina constituye un reservorio de di-ferentes factores de crecimiento y la agarosa proporciona estabilidad y consistencia adecuada favoreciendo el transporte y la manipulación en el quirófano. Además los fibroblastos humanos inducen la proliferación de los queratinocitos que se encuentran en las capas superiores. Las carac-terísticas histológicas, bioquímicas y reológicas de la piel artificial creada en laboratorio eran a nivel epidérmico y dérmico muy similares a la piel humana normal y además mostró una adecuada biointegración en los animales de experimentación en los que se utilizó.
En resumen, el desarrollo de la ingeniería tisular cutánea permitirá disponer de sustitutos cutáneos autólogos para su uso clínico y para la investigación.
Una vez descrita y analizada la piel en su conjunto y cada uno de sus componentes, entenderemos mucho mejor sus diferentes funciones y al-teraciones que sustentan diversas patologías (Tabla V).
A lo largo del capítulo hemos comprobado que la piel además de ser una frontera eficaz que nos protege del exterior, es una sensibilísima envoltura que nos permite sentir numerosas sensaciones del mundo exterior y a la vez nos permite expresar numerosos sentimientos como la vergüenza o timidez mediante el rubor; el temor, frío o agresividad mediante la palidez; la emoción o miedo mediante la «piel de gallina», etc.Para finalizar podríamos decir que la piel nos ayuda a sentir, a amar, a vivir y a morir.

27
Capítulo 1. Anatomía y fisiología de la piel
Abd E, Yousef SA, Pastore MN, Telaprolu K, Mohammed YH, Namjoshi S, Grice JE, Roberts MS. Skin models for the testing of transdermal drugs. Clin Pharmacol 2016 Oct 19; 8: 163-176.
Alaminos M, Garzón I, Sánchez-Quevedo MC, Moreu G, González Andrades M, Fernández-Montoya A, Campos A. Time-course study of histological and genetic patterns of differentiation in human engineered oral mucosa. Journal of Tissue Engineering and Regenerative Medicine 2007; 1: 350-359.
Baddour JA, Sousounis K, Tsonis PA. Organ repair and regeneration: an overview. Birth Defects Res C Embryo Today 2012 Mar; 96: 1-29.
Brölmann FE, Eskes AM, Goslings JC, Niessen FB, de Bree R, Vahl AC, Pierik EG, Vermeulen H, Ubbink DT; REMBRANDT study group. Randomized clinical trial of donor-site wound dressings after split-skin grafting. Br J Surg 2013 Apr; 100: 619-27.
Carriel V, Garzón I, Jiménez JM, Oliveira AC, Arias-Santiago S, Campos A, Sánchez-Quevedo MC, Alaminos M. Epithelial and stromal developmental patterns in a novel substitute of the human skin generated with fibrin-agarose biomaterials. Cells Tissues Organs 2012; 196: 1-12.
Carriel V, Garzón I, Jiménez JM, Oliveira AC, Arias-Santiago S, Campos A, et al. Epithelial and stromal developmental patterns in a novel substitute of the human skin generated with fibrin-agarose biomaterials. Cells Tissues Organs. 2012; 196: 1-12.
Chu, DH. Generalidades de la biología, el desarrollo y la estructura de la piel. En: Wolf K, Goldsmith LA,
Katz SI, Gilchrest BA, Paller AS, Leffell DJ. Fitzpatrick Dermatología en Medicina General. Tomo I. 7ª edición. Ed. Médica Panamericana; 2009; 57-72.
Gibot L, Galbraith T, Huot J, Auger FA. Development of a tridimensional microvascularized human skin substitute to study melanoma biology. Clin Exp Metastasis 2013 Jan; 30: 83-90.
Hu DH, Zhang ZF, Zhang YG, Zhang WF, Wang HT, Cai WX, Bai XZ, Zhu HY, Shi JH, Tang CW. A potential skin substitute constructed with hEGF gene modified HaCaT cells for treatment of burn wounds in a rat model. Burns 2012 Aug; 38: 702-12.
Kim HL, Lee JH, Lee MH, Kwon BJ, Park JC. Evaluation of electrospun (1,3)-(1,6)-β-D-glucans/biodegradable polymer as an artificial skin for full-thickness wound healing. Tissue Eng Part A. 2012; 18: 2315-22.
Montagna W, Camacho FM. Embriología y anatomía del folículo piloso. Ciclos de crecimiento del pelo. Desarrollo de los folículos pilosos humanos. Anatomía de la glándula sebácea, glándula apocrina, musculo erector. Vascularización e inervación. En: Camacho FM, Tosti A. Tricología. Enfermedades del folículo pilosebáceo. Volumen I. 3ª edición. Aula médica 2013; 3-37.
Naves LB, Dhand C, Almeida L, Rajamani L, Ramakrishna S. In vitro skin models and tissue engineering protocols for skin graft applications. Essays Biochem 2016 Nov 30; 60: 357-369.
Nerem, R.M. Regenerative medicine: the emergence of an industry. J R Soc Interface 2010; 7: S771-S775.
Orlando G, Wood KJ, De Coppi P, Baptista PM, Binder KW, Bitar KN, et al. Regenerative medicine as applied to general surgery. Ann Surg 2012 May; 255: 867-80.
Papa G, Pangos M, Renzi N, Ramella V, Panizzo N, Marij AZ. Five years of experience using a dermal substitute: indications, histologic studies, and first results using a new single-layer tool. Dermatol Surg 2011 Nov;37: 1631-7.
Sánchez-Quevedo MC, Alaminos M, Capitán LM, Moreu G, Garzón I, Crespo PV, Campos A. Histological and histochemical evaluation of human oral mucosa constructs developed by tissue engineering. Histology and Histopathology 2007; 22: 631-40.
Sánchez-Quevedo MC, M Alaminos, G Moreu, M González-Jaranay, LM Capitán, I Labrot, A. Campos. In-vitro cultivation of normal human oral fibroblasts and keratinocytes. Histology and Histopathology 2005; S1: 152-153.
Vandergriff TW, Bergstresser PR. Anatomy and Physiology. En: Bolonia JL, Jorizzo JL, Schaffer JV. Dermatology. Elsevier. Estados Unidos. 2012; 43-54.
Yamada N, Uchinuma E, Kuroyanagi Y. Clinical trial of allogeneic cultured dermal substitutes for intractable skin ulcers. J Artif Organs 2012 Jun; 15: 193-9.
Zhou H, You C, Jin R, Wu P, Li Q , Han C. The progress and challenges for dermal regeneration in tissue engineering. J Biomed Mater Res A. 2017 Apr; 105: 1208-1218.
BIBLIOGRAFÍA