Embed Size (px)
Citation preview

4625
15
7
Volume 24 June 2015
AR
CH
AE
OFA
UN
AV
olu
me
24 -
Jun
e 2015
Índice / contents
issn - 1132-6891
150
51
1
Utilización de reptiles durante el período Agroalfarero de la región Chaco-Santiagueña, Argentina.Luis Manuel del Papa ...................................................................................................................
Micromammals, taphonomy, paleoenvironments and human occupation during the Holocene inArroyo Malo-3 (west-central Argentina). Fernando J. Fernández, Luciano J.M. De Santis &Gustavo A. Neme ...........................................................................................................................
Utilización de Rhea americana (Aves, Rheidae) en el sitio Paso Otero 4 (partido de Necochea, regiónpampeana). María Clara Álvarez ..................................................................................................
El rol de los pequeños animales en los estudios arqueofaunísticos de Argentina. Romina Frontini& Paula D. Escosteguy .................................................................................................................
Peces marinos, peces fluviales: explotación diferencial por grupos cazadores-recolectores del norestede Chubut (Patagonia central, Argentina). Ariadna Svoboda & Julieta Gómez Otero .................
Tendencias preliminares sobre el consumo de peces durante el Holoceno medio en el área de tran-sición Pampeano-Patagónica oriental (provincia de Buenos Aires). Luciana Stoessel.................
Primeros resultados de los análisis del conjunto de fauna menor del Sitio Arqueológico Zoko Andi1 (Transición Pampeano-Patagónica Oriental, Provincia De Buenos Aires, Argentina). Ana PaulaAlcaráz ..........................................................................................................................................
Explotación de peces en Lagunas de Guanacache. Altos de Melién II. M. Corbat, P. Cahiza, J. Gar-cía Llorca & A. Gil ........................................................................................................................
Análisis de la fauna menor del sitio Piedras Blancas (Valle de Ambato, Catamarca, Argentina). Ma-riana Dantas & Gabriela Srur ......................................................................................................
Patrón esquelético de roedores Cávidos en la secuencia arqueológica de Cueva Tixi, Tandilia Orien-tal (Pleistoceno Tardío - Holoceno Tardío) Argentina. Carlos A. Quintana .................................
Implicancias tafonómicas y paleoambientales de los pequeños vertebrados del sitio arqueológicoCampo Laborde (Centro de los Pastizales Pampeanos, Buenos Aires, Argentina). Nahuel A.Scheifler, Pablo G. Messineo & Ulyses F.J. Pardiñas...................................................................
Las investigaciones sobre preservación de huesos de aves y mamíferos grandes en Patagonia (Argen-tina). Isabel Cruz ...........................................................................................................................
Caza y extracción de pieles de Chinchillidae en la puna de Jujuy (Argentina): una aproximaciónarqueológica. Juan M. Maryañski & Axel E. Nielsen ...................................................................
Observaciones tafonómicas naturalistas sobre restos de armadillos (Dasypodidae) en Patagonia meri-dional. A. Sebastián Muñoz....................................................................................................................
Diversidad faunística en La Arcillosa 2: el aprovechamiento de aves por cazadores-recolectores delnorte de Tierra del Fuego. Angélica M. Tivoli & Mónica C. Salemme .........................................
Consumo de recursos animales de porte menor durante el Holoceno tardío en el sudoeste de la pro-vincia de Buenos Aires (Argentina). Romina Frontini & Cristina Bayón ....................................
Nuevos datos sobre fauna menor de la Depresión del río Salado (provincia de Buenos Aires, Argen-tina): el caso de San Ramón 7. Paula D. Escosteguy, María Isabel González & María MagdalenaFrère ..............................................................................................................................................
Archaeological applications of microvertebrate analysis: Reconstruction of natural and anthropicprocesses. Débora M. Kligmann ...................................................................................................
Fauna menor en sitios de momentos históricos. Matilde M. Lanza ...................................................
Announcements ..................................................................................................................................
7-26
27-52
53-65
67-85
87-101
103-117
119-133
135-151
153-171
173-185
187-208
209-224
225-237
239-251
253-269
271-293
295-313
315-339
341-362
363-367
UNAEOFRCHA A AINTERNATIONAL JOURNAL OF ARCHAEOZOOLOGY
El rol de los pequeños animales
en los estudios arqueofaunísticos
de Argentina
Assistant Editors:PAULA ESCOSTEGUY & ROMINA FRONTINI

ARCHAEOFAUNA
ARCHAEOFAUNA es una revista anual que publicatrabajos originales relacionados con cualquier aspecto delestudio de restos animales recuperados en yacimientosarqueológicos. Los manuscritos deben enviarse a:
Para la elaboración de manuscritos, que serán evaluadospor un mínimo de dos recensores externos, consultar las ins-trucciones de la contraportada. Todos los manuscritos noconformes con las normas de publicación serán automática-mente devueltos a los autores. Cada autor o grupo de autoresrecibirán un pdf de su trabajo.
ARCHAEOFAUNA is an annual journal that publishesoriginal papers dealing with all aspects related to the studyof animal remains from archaeological sites. Manuscriptsshould be sent to:
For preparation of manuscripts, that will be evaluated bya minimum of two external referees, please follow the ins-tructions to authors. All manuscripts not conforming tothese instructions will be automatically returned to theauthors. Each author (or group of authors) will receive a pdfof his/her (their) work.
EUFRASIA ROSELLó IZqUIERDOLaboratorio de Arqueozoología. Dpto. Biología. Universidad Autónoma de Madrid
28049 Madrid. España (Spain)
Director: ARTURO MORALES MUñIZLaboratorio de Arqueozoología. Dpto. Biología. Universidad Autónoma de Madrid.
28049 Madrid. España (Spain)
Comité editorial/Editorial board:
K. AARIS-SO/RENSEN: Zoologisk Museum, Ko/benhavn.Denmark.
J. ALTUNA ECHAVE. Sociedad de Ciencias Aranzadi, SanSebastián. Spain.
A. ANDERSON. Research School of Pacific and Asian Studies.The Australian National University, Canberra. Australia.
N. BENECKE. Deutsches Archäologisches Institut, Berlin.Germany.
A. ERVYNCK. Institute for the Archaeological Heritage of theFlemish Community. Belgium
A. GAUTIER. Laboratorium voor Paleontologie.Rijksuniversiteit, Gent. Belgium.
D. K. GRAYSON. Burke Memorial Museum. University ofWashington. U.S.A.
D. HEINRICH. Institut für Haustierkunde. Christian-Albrechts-Universität, Kiel. Germany.L. JONSSON. Central Boardof National Antiquities, Kungsbacka. Sweden.
F. B. LEACH. Archaeozoology Laboratory. Museum of NewZealand, Wellington. New Zealand.
C. LEFèVRE, Muséum National d’Histoire Naturelle, Paris.France.
M. LEVINE. Department of Archaeology. CambridgeUniversity. United Kingdom.
R. H. MEADOW. Zooarchaeology Laboratory. PeabodyMuseum of Archaeology and Ethnology. HarvardUniversity. U.S.A.
G. G. MONKS. University of Manitoba. Canada.W. VAN NEER. Musée Royal de L'Afrique Centrale, Tervuren.
Belgium.R. A. NICHOLSON. Department of Archaeological Sciences,
University of Bradford. Bradford. United Kingdom.S. PAYNE. Ancient Monuments Laboratory, English Heritage,
London. United Kingdom.C. A. POGGENPOEL. Department of Archaeology. University
of Cape Town. South Africa.H.P. UERPMANN. Universität Tübingen. Germany.E. WING. Department of Anthropology, University of
Florida, Gainesville. U.S.A.M. ZEDER. Smithsonian Institution. Washington D.C. U.S.A.
ARCHAEOFAUNALaboratorio de Arqueozoología. Depto. Biología.Universidad Autónoma de MadridCantoblanco 28049 Madrid. España
Editor: Eufrasia Roselló
ISSN: 1132 - 6891
Depósito Legal: M - 30872 - 1992
Imprime:LOKE CBc. Progreso, 2 - despacho 218Polígono Industrial Los Olivos28906 Getafe (Madrid)[email protected]
Revista incluida en las bases de datos ICYT (CINDOC), Catálogo Latindex, Zoological Record, The Arts & HumanitiesCitation Index y Current Contents / Arts & Humanities (JCR)
INFORMACIóN A LOS AUTORES
a) Los originales pueden redactarse en español,inglés, alemán o francés. Los editores pueden considerar,en determinadas circunstancias, la publicación de origi-nales en otros idiomas. En cualquier caso se proporcio-nará un resumen y palabras clave en español y en inglés.
b) Los originales no deberían sobrepasar 20 páginasA4 (29,5 x 21 cm) incluyendo tablas y figuras. En casode trabajos más extensos contáctese con el editor. Losmanuscritos deberán remitirse a [email protected].
c) Las figuras y tablas deberán ser originales y degran calidad. Las leyendas de figuras y de tablas debe-rán remitirse, numeradas, en ficheros independientes yserán concisas e informativas.
d) Estructuración del manuscrito. El orden requeri-do en los manuscritos de carácter experimental es elsiguiente: Título del trabajo; Autor(es) y Centro(s) detrabajo; Resumen y Palabras Clave; Abstract y key-words; Introducción; Discusión; Conclusiones;Agradecimientos (optativo); Referencias. Si el trabajoasí lo requiere, resultados y discusión pueden agruparseen el mismo epígrafe. En manuscritos no experimenta-les, la estructuración del trabajo se deja a la libre deci-sión del(de los) autor(es).
e) las citas bibliográficas en el texto incluirán autory año de publicación, por ejemplo (Smithm 1992) o(Smith & Jones, 1992). En trabajos con tres o más auto-res usar (Martín et al., 1993). En trabajos del(de los)mismo(s) autor(es) y año, se procederá a identificarcada trabajo con letras (a, b, c, etc…) tras la fecha.
f) Referencias. Sólo se incluirán aquellas citadas enel texto y se hará del siguiente modo:
PéREZ, C.; RODRíGUEZ, P. & DíAZ, J. 1960: Ecolo -gi can factors and family size. Journal of Bioethics 21:13-24.
RUIZ, L. 1980: The ecology of infectious diseases.Siglo XXI, Madrid.
g) Los autores son los únicos responsables de loscontenidos de sus artículos.
INFORMATION FOR AUTHORS
a) Manuscripts can be submitted in Spanish,English, German and French. Under certain circums-tances papers may also be published in other EuropeanCommunity languages. All papers will include an abs-tract and keywords in English and Spanish.
b) Manuscripts should usually not exceed 20 A4 prin-ted pages (29,5 x 21 cm), including figures and tables.For longer manuscripts, contact the editor. Manuscriptsshould be submitted to [email protected].
c) Figures and tables must be original and hightquality. Figure legends should be numbered with arabicnumerals and given on a separate file. Figure and tablelegends should be concise and informative.
d) Papers should be organized as follows: Title,name and mailing address(es) of author(s). Abstract,Keywords, Introduction, Materials and Methods,Results, Discussion, Conclusions, Acknowledgements,References. Results and Discussion may be treatedtogether if this is appropiate. Non-experimental workscan be organized in the way whicht th author(s)think(s) is the most appropiate one.
e) Citations in the text should be with author and dateof publication, e. g., (Smith, 1992) or (Smith & Jones,1992) with comma between author and date; for two-author papers, cite both authors; for papers by three ormore authors, use Martín et al., 1993. For two or morepapers with the same author(s) and date, use, a, b, c, etc.,after the date.
f) References: only papers cited in the text should beincluden; they should be arranged as indicated in point«f» of the other column.
g) Authors are responsible for the contents of theirmanuscripts.
PORTADA: Esqueletos de ñandúes, Rhea americana superpuestossobre fémures con fractura antrópica y marcas de carnívoro.FRONTISPIECE: Skeletons of Greater Rhea Rhea americana overfemora featuring man-made fractures and carnivore gnawing marks.

Utilización de reptiles durante el período Agroalfarero de la región Chaco-Santiagueña,Argentina. Luis Manuel del Papa .........................................................................................
Micromammals, taphonomy, paleoenvironments and human occupation during the Holocenein Arroyo Malo-3 (west-central Argentina). Fernando J. Fernández, Luciano J.M. De San-tis & Gustavo A. Neme..........................................................................................................
Utilización de Rhea americana (Aves, Rheidae) en el sitio Paso Otero 4 (partido de Neco-chea, región pampeana). María Clara Álvarez .....................................................................
El rol de los pequeños animales en los estudios arqueofaunísticos de Argentina. RominaFrontini & Paula D. Escosteguy ...........................................................................................
Peces marinos, peces fluviales: explotación diferencial por grupos cazadores-recolectores delnoreste de Chubut (Patagonia central, Argentina). Ariadna Svoboda & Julieta GómezOtero......................................................................................................................................
Tendencias preliminares sobre el consumo de peces durante el Holoceno medio en el área detransición Pampeano-Patagónica oriental (provincia de Buenos Aires). Luciana Stoessel ...
Primeros resultados de los análisis del conjunto de fauna menor del Sitio Arqueológico ZokoAndi 1 (Transición Pampeano-Patagónica Oriental, Provincia De Buenos Aires, Argenti-na). Ana Paula Alcaráz .........................................................................................................
Explotación de peces en Lagunas de Guanacache. Altos de Melién II. M. Corbat, P. Cahiza,J. García Llorca & A. Gil ..........................................................................................................................................................
Análisis de la fauna menor del sitio Piedras Blancas (Valle de Ambato, Catamarca, Argenti-na). Mariana Dantas & Gabriela Srur .................................................................................
Patrón esquelético de roedores Cávidos en la secuencia arqueológica de Cueva Tixi, TandiliaOriental (Pleistoceno Tardío - Holoceno Tardío) Argentina. Carlos A. Quintana ...............
Implicancias tafonómicas y paleoambientales de los pequeños vertebrados del sitio arqueo-lógico Campo Laborde (Centro de los Pastizales Pampeanos, Buenos Aires, Argentina).Nahuel A. Scheifler, Pablo G. Messineo & Ulyses F.J. Pardiñas .........................................
Las investigaciones sobre preservación de huesos de aves y mamíferos grandes en Patagonia(Argentina). Isabel Cruz........................................................................................................
Caza y extracción de pieles de Chinchillidae en la puna de Jujuy (Argentina): una aproxima-ción arqueológica. Juan M. Maryañski & Axel E. Nielsen ...................................................
Observaciones tafonómicas naturalistas sobre restos de armadillos (Dasypodidae) en Patago-nia meridional. A. Sebastián Muñoz .....................................................................................
Diversidad faunística en La Arcillosa 2: el aprovechamiento de aves por cazadores-recolec-tores del norte de Tierra del Fuego. Angélica M. Tivoli & Mónica C. Salemme .................
Consumo de recursos animales de porte menor durante el Holoceno tardío en el sudoeste dela provincia de Buenos Aires (Argentina). Romina Frontini & Cristina Bayón ..................
Nuevos datos sobre fauna menor de la Depresión del río Salado (provincia de Buenos Aires,Argentina): el caso de San Ramón 7. Paula D. Escosteguy, María Isabel González &María Magdalena Frère .......................................................................................................
Índice/Contents
7-26
27-52
53-65
67-85
87-101
103-117
119-133
135-151
153-171
173-185
187-208
209-224
225-237
239-251
253-269
271-293
295-313
00. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 05/05/15 11:23 Página 5

Archaeological applications of microvertebrate analysis: Reconstruction of natural andanthropic processes. Débora M. Kligmann...........................................................................
Fauna menor en sitios de momentos históricos. Matilde M. Lanza ..........................................
Announcements..........................................................................................................................
315-339
341-362
363-367
00. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 05/05/15 11:23 Página 6

ariadna SvoBoda & Julieta GÓMez otero
Peces marinos, peces fluviales: explotación diferencial por grupos cazadores-recolectores del noreste de Chubut (Patagonia central, Argentina)

Archaeofauna 24 (2015): 87-101
Peces marinos, peces fluviales: explotación diferencialpor grupos cazadores-recolectores del noreste de Chubut
(Patagonia central, Argentina)
ARIADNA SVOBODA1 & JULIETA GÓMEZ OTERO2
1Centro Nacional Patagónico-CONICET. Puerto [email protected]
2Centro Nacional Patagónico-CONICET. Puerto Madryn. Universidad de la Patagonia San Juan Bosco. [email protected]
(Received 31 March 2014; Revised 31 May 2014; Accepted 2 June 2014)
RESUMEN: La incorporación de peces a la dieta de grupos cazadores-recolectores puede eva-luarse en función de su densidad, la disponibilidad de recursos de mayor tamaño corporal, latecnología disponible, entre otros. En este trabajo se aborda la variabilidad en el registro ictio-faunístico durante el Holoceno tardío en dos áreas: el valle inferior del río Chubut y su desem-bocadura y costa adyacente. De acuerdo a la productividad ambiental, la disponibilidad derecursos de alta jerarquía y las oportunidades de captura de peces, se genera un modelo de sub-sistencia que evalúa la diversidad e importancia relativa de los peces para las dos áreas. Lasexpectativas zooarqueológicas derivadas son discutidas a partir de los muestreos faunísticos delas localidades Barranca Norte y Los Cangrejales Sur (desembocadura y costa) y los sitiosLoma Grande 1 y Cinco Esquinas 1 (valle inferior), ubicados a unos 30 y 18 km del mar res-pectivamente. Los resultados arrojan diferencias en la riqueza taxonómica de peces: en el vallepredominan las percas, mientras que en la desembocadura y costa se explotó una amplia varie-dad. Se registró una importante contribución de peces en ambas áreas; sin embargo, en los cam-pamentos residenciales de la desembocadura los restos de peces son menos frecuentes.
PALABRAS CLAVE: SUBSISTENCIA, PECES, CAZADORES-RECOLECTORES, PATA-GONIA CENTRAL, ARGENTINA, HOLOCENO TARDÍO
ABSTRACT: The incorporation of fish into the diet of hunter-gatherers can be evaluated on thebasis of their densities, the abundance of high-ranked resources and available technology. Thispaper examines the variability of the late Holocene ichthyoarchaeological record in two environ-mental areas: the lower valley of the Chubut River, along with its mouth and adjacent marinecoast. According to environmental productivity, high-ranked resources availability and captureopportunities, a subsistence model is generated. This model evaluates diversity and relative impor-tance of fish for the two areas. Zooarchaeological expectations inferred are discussed on accountof faunal samples taken from different archaeological sites, such as Barranca Norte and Los Can-grejales Sur (located at the river mouth and marine coast), Loma Grande 1 and Cinco Esquinas 1(located on the valley, at 30 and 18 km distance from sea). Results show differences in fish taxo-nomic richness with perch dominant in the valley sites, whereas a high range of fish was found atthe river mouth and marine coast sites. Although an important contribution of fish registered inboth areas was recorded; in residential camps at the river mouth fish remains were scarce.
KEYWORDS: SUBSISTENCE, FISH, HUNTER-GATHERERS, CENTRAL PATAGONIA,ARGENTINA, LATE HOLOCENE
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 87

INTRODUCCIÓN
La fauna menor, incluyendo los peces, es untema que atrae a los zooarqueólogos porque ofre-ce información sobre el aporte nutricional, laamplitud del espectro alimenticio, la estacionali-dad de captura, las técnicas empleadas en la caza ypesca, y también es un indicador paleoecológico(Casteel, 1976; Smith, 1983; Wheeler & Jones,1989; Lindstrom, 1996; Malainey et al., 2001;Lupo & Schmitt, 2005; Scartascini & Volpedo,2013). Uno de los campos en que el registro defauna menor tomó mayor relevancia es la teoríadel forrajeamiento óptimo y el modelo derivado deamplitud de dieta (Bettinger, 1991), que permitenmonitorear cambios en la subsistencia de cazado-res-recolectores (Broughton, 1999; Stiner et al.,2000; Nagaoka, 2002; Butler & Campbell, 2004;Neme & Gil, 2008; entre otros). Dicho modeloecológico, que jerarquiza los recursos de acuerdo ala tasa de retorno energético –el cual suele eva-luarse en forma indirecta por el tamaño corporal(Lupo, 2007)– presume que la inclusión de faunamenor en la dieta dependerá de la abundancia yoportunidades de captura de los recursos de mayortamaño (Winterhalder et al., 1988; Bettinger,1991). No obstante, en ocasiones, algunos recursosde baja jerarquía, como la fauna menor, suelenobtenerse en grandes cantidades simultáneamente;por lo tanto, su rendimiento energético no está enfunción de su tamaño sino de la densidad y la tec-nología utilizada (Madsen & Schmitt, 1998; Gray-son & Cannon, 1999).
En referencia a los peces, estudios experimenta-les han permitido determinar que la captura enmasa en ambientes lagunares y lacustres y en arri-bazones marinos genera alto retorno energético(Limp & Reidhead, 1979; Perlman, 1980). De igualmodo, otras situaciones de pesca no dependientesde la densidad mantienen rendimientos productivos(Lindstrom, 1996). Más allá de la productividad,los peces (sobre todo los marinos) son un recursoaltamente previsible en el espacio y en el tiempopor su preferencia de hábitat y su disponibilidaddependiente de los ciclos de mareas, estacionales einteranuales (Bailey & Parkington, 1988). En otroorden, los peces constituyen una fuente de proteí-nas de mayor calidad que la provista por animalesterrestres ya que también aportan vitaminas yminerales (Stewart, 1994), por lo que su incorpora-ción a la dieta trae aparejados notables beneficiosnutricionales y menor morbilidad entre las pobla-ciones (Erlandson, 2001). En suma, si bien es cier-
to que la valorización de peces puede evaluarse enfunción de la disponibilidad de recursos de mayorjerarquía, es importante considerar que bajo dife-rentes situaciones puede resultar una presa previsi-ble y productiva obtenida mediante el empleo detecnologías relativamente sencillas, además deaportar complementos nutricionales con ampliosbeneficios para las poblaciones.
Entre los numerosos conjuntos arqueofaunísti-cos del noreste de Chubut (Patagonia central) sehan hallado restos de peces (Gómez Otero, 2006;Gómez Otero et al., 2009, 2013). Esta área fueocupada por grupos cazadores-recolectores demovilidad relativa y de baja densidad cuya subsis-tencia estuvo basada sobre una amplia gama derecursos terrestres (guanaco, plantas y animalespequeños) y marinos (moluscos, pinnípedos, y enmenor medida peces y aves marinas) (GómezOtero, 2006). Se pudo constatar que la explotaciónde peces se remite al Holoceno medio y continúahasta la época del contacto con los europeos(Gómez Otero et al., 2013).
Sobre la base de lo expuesto, en este trabajo sepropone indagar sobre la explotación de peces endos áreas de Patagonia central definidas por suvariabilidad ecológica: el valle inferior del río Chu-but por un lado; y, su desembocadura y costa mari-na inmediata, por el otro. Los ambientes ribereño-terrestres y la faja costera y estuarios son diferentesen cuanto a la calidad ambiental (Perlman, 1980;Bailey & Parkington, 1988). Principalmente en lacosta marina y estuarios se presenta alta biodiversi-dad y frecuencia de recursos que se traduce en unamayor biomasa disponible (Perlman, 1980; Yesner,1980; Bailey & Parkington, 1988). Si se tiene encuenta la productividad de los ambientes, la dispo-nibilidad de recursos de alta jerarquía y las oportu-nidades de captura de peces se espera hallar dife-rencias en el grado de intensidad de la pesca enrelación con otras prácticas de subsistencia para losgrupos cazadores-recolectores de cada uno de losambientes previamente definidos.
MARCO AMBIENTAL Y VARIABILIDADECOLÓGICA: CONDICIONESY EXPECTATIVAS PARA LA EXPLOTACIÓNDE PECES
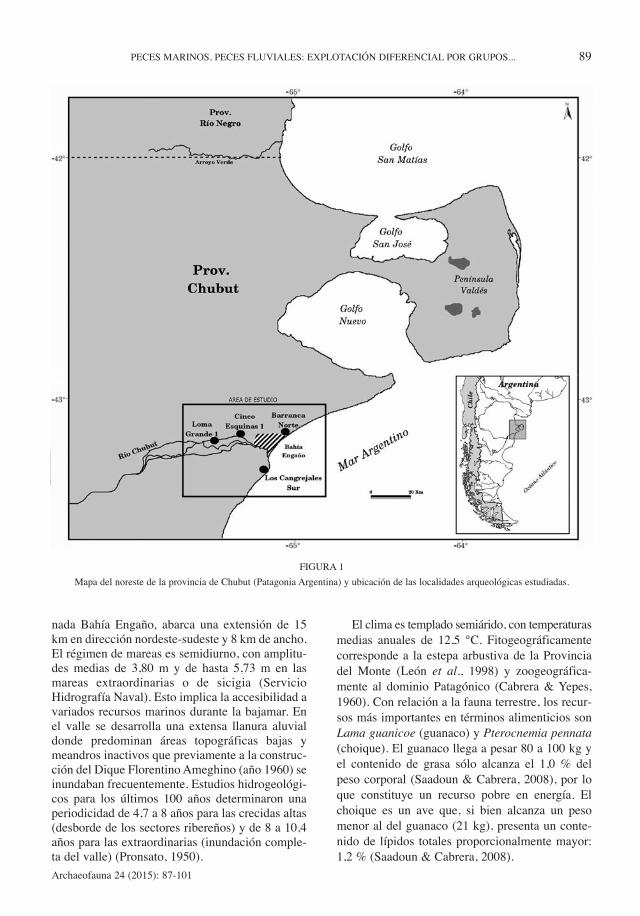
El área de estudio comprende el valle inferior ydesembocadura del río Chubut, y la costa inmedia-ta (Figura 1). La desembocadura del río, denomi-
88 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 88
nada Bahía Engaño, abarca una extensión de 15km en dirección nordeste-sudeste y 8 km de ancho.El régimen de mareas es semidiurno, con amplitu-des medias de 3,80 m y de hasta 5,73 m en lasmareas extraordinarias o de sicigia (ServicioHidrografía Naval). Esto implica la accesibilidad avariados recursos marinos durante la bajamar. Enel valle se desarrolla una extensa llanura aluvialdonde predominan áreas topográficas bajas ymeandros inactivos que previamente a la construc-ción del Dique Florentino Ameghino (año 1960) seinundaban frecuentemente. Estudios hidrogeológi-cos para los últimos 100 años determinaron unaperiodicidad de 4,7 a 8 años para las crecidas altas(desborde de los sectores ribereños) y de 8 a 10,4años para las extraordinarias (inundación comple-ta del valle) (Pronsato, 1950).
El clima es templado semiárido, con temperaturasmedias anuales de 12,5 °C. Fitogeográficamentecorresponde a la estepa arbustiva de la Provinciadel Monte (León et al., 1998) y zoogeográfica-mente al dominio Patagónico (Cabrera & Yepes,1960). Con relación a la fauna terrestre, los recur-sos más importantes en términos alimenticios sonLama guanicoe (guanaco) y Pterocnemia pennata
(choique). El guanaco llega a pesar 80 a 100 kg yel contenido de grasa sólo alcanza el 1,0 % delpeso corporal (Saadoun & Cabrera, 2008), por loque constituye un recurso pobre en energía. Elchoique es un ave que, si bien alcanza un pesomenor al del guanaco (21 kg), presenta un conte-nido de lípidos totales proporcionalmente mayor:1,2 % (Saadoun & Cabrera, 2008).
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 89
Archaeofauna 24 (2015): 87-101
FIGURA 1
Mapa del noreste de la provincia de Chubut (Patagonia Argentina) y ubicación de las localidades arqueológicas estudiadas.
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 89

Más allá que los tres ambientes (valle, desem-bocadura y costa marina) comparten la flora y lafauna terrestre antes mencionada, la diferencia encada uno está dada por la disponibilidad y oportu-nidades de captura de los recursos faunísticos dul-ceacuícolas y marinos. El ambiente ribereño ofre-ce principalmente aves y peces. Las avesdulceacuícolas de mayor importancia económicacorresponden a varias especies de Anátidos. Tam-bién habita un mamífero acuático, Myocastor
coypus (coipo), que es un roedor de entre siete ydiez kg de peso, cuyo contenido de lípidos totales(incluye ácidos grasos) alcanza 1,9 a 2,4% (Saa-doun & Cabrera, 2008). La fauna íctica autóctonaestá representada por Percichthys trucha (perca),Odontesthes hatcheri (pejerrey patagónico) ybagres chicos como Diplomystes mesembrinus yHatcheria macraei (Gosztonyi, 1988). Entre estasespecies, la perca es la que presenta mayor poten-cial de explotación económica por su accesibili-dad, oportunidad de captura y previsibilidad. Con-tiene altos porcentajes de grasa (López & Lipps,1988), y alcanza un peso de 1,5 kg, aunque enlagos del norte de Patagonia se han registradopesos de hasta 6 y 8 kg, (Amalfi 2009). Por sucapacidad de adaptación fisiológica, que le permi-te sobrevivir bajo condiciones de apoxia (MacDonagh, 1950), es la única especie fluvial quepuede perdurar en las lagunas ribereñas en proce-so de desecamiento. Esta situación es favorablepara la recolección y/o captura en masa de percas,estrategia que no podría practicarse en el caso deotras especies ícticas, cuya obtención sería indivi-dual (Martínez et al., 2005). Es interesante desta-car que los episodios de crecientes o grandes inun-daciones favorecen la colonización y reproducciónde la fauna dulceacuícola, al mismo tiempo gene-ran períodos de abandono y alejamiento de lafauna terrestre, que busca refugio en los sectoresaltos (Matthews, 1992).
En la costa marina la biomasa es mayor por laoferta conjunta de recursos del mar y de la tierra.Esto se potencia en la desembocadura, donde seagregan las especies dulceacuícolas. El mamíferomarino de mayor importancia corresponde a las dosespecies de pinnípedos: Arctocephalus australis
(lobo de dos pelos) y Otaria flavescens (lobo de unpelo). El peso promedio de A. australis es de 159kg para machos adultos, 48,9 kg para hembrasadultas y 3,5 a 5,5 kg en las crías (Schiavini,1994). El porcentaje de grasa subcutánea sobre elpeso total es de 12% en juveniles y adultos, y 31%en cachorros (Schiavini, 1994). Los tamaños O.
flavescens son aún mayores: los machos adultospesan 300-340 kg, las hembras adultas 144 kg ylas crías entre 10,2 y 14,8 kg (Vaz Ferreira, 1976).Las aves marinas de mayor importancia por sutamaño y abundancia son Phalacrocorax spp (cor-moranes) y Spheniscus magellanicus (pingüino deMagallanes) que no superan los 4,5 kg de pesocorporal. Los peces se destacan por su diversidadespecífica y pueden clasificarse de acuerdo a losrequerimientos de hábitat. Por un lado, en sustratosrocosos (v.g. plataformas de abrasión) se puede hallarpeces de arrecifes: Acanthistius patachonicus (mero),Patagonotothen spp (nototenias), Triathalossothia
argentina (pez sapo), y Austrolycus sp. (anguila)(Irigoyen & Galván, 2010). Por otro lado, en ellitoral somero frecuentan algunos condríctioscomo Callorhynchus callorhynchus (pez gallo),varias rayas (Rajidae) y Triakidae (tiburones).Entre los peces óseos se hallan varias especies delgénero Odontesthes spp (pejerreyes), Percophis
brasiliensis (pez palo), Engraulis anchoita
(anchoita) y Eleginops maclovinus (róbalo); esteúltimo frecuenta también los arrecifes. En la actua-lidad, durante el intermareal los peces de arrecifesuelen capturarse mediante uso de líneas o cañasen las pozas de marea o en los bordes de las res-tingas donde se accede al submareal. En las playasarenosas y de rodados se utilizan también redes otrasmallos. Finalmente, los moluscos son muyabundantes y variados en el área de estudio; losmás conocidos por su valor comercial real o poten-cial son Chlamys tehuelcha (vieyra), Mytilus edulis
(mejillón), Aulacomya ater (cholga), Venus antiqua
(almeja blanca), los géneros Buccinanops (nasas)y Trophon (trofones) y las familias Patellidae yFissurellidae (lapas) (Lasta et al., 1998).
En síntesis, en el área de estudio se definen dosunidades ambientales principales. Por un lado, elvalle donde el mayor aporte nutricional está poten-cialmente representado por guanacos y choiques,aunque con limitaciones en ciertos momentos (v.g.grandes crecientes). Con la llegada de las crecidasen este ambiente ecotonal se habrían generadocondiciones favorables para la explotación derecursos menores y la búsqueda de fuentes alter-nativas de alimento. Entre ellos, la perca presenta-ría mayor ventaja de captura mediante el empleode tecnologías sencillas. Por otro lado, en ladesembocadura y costa, además de los animalesterrestres, se hallan los mamíferos marinos quehabrían representado importantes aportes energéti-cos a la subsistencia humana, a los que se le sumanuna amplia diversidad de recursos menores entre
90 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 90
aves, peces y moluscos. De acuerdo a la estructu-ra ambiental antes presentada, se espera registrardiferencias en la composición y abundancia depeces entre los conjuntos zooarqueológicos delvalle inferior y la costa. A modo de herramientaheurística proponemos un modelo predictivo desubsistencia del cual deriva una serie de expectati-vas zooarqueológicas:
– En el valle, la menor diversidad faunística enrelación con la costa (en especial la ausenciade presas grandes de alto contenido calóricocomo los mamíferos marinos), habría propi-ciado un aprovechamiento más intensivo delos peces. Además, durante los periódicosepisodios de inundación del valle la explota-ción de presas dulceacuícolas, entre ellas lospeces, se habría incrementado. En consecuen-cia, se espera encontrar mayor abundanciarelativa de restos ícticos y baja riqueza taxo-nómica de peces en el registro zooarqueológi-co correspondiente a dichos sectores.
– En la desembocadura del río Chubut y costainmediata, la mayor biomasa y diversidadfaunística de recursos de alta jerarquía, nohabrían propiciado el aprovechamiento inten-sivo y regular de peces. Por lo tanto, se espe-ra registrar menor abundancia relativa de res-tos ícticos y una riqueza taxonómica acordecon la mayor disponibilidad específica depeces en la costa.
Para monitorear los cambios en los conjuntoszooarqueológicos a nivel general se utiliza unaserie de unidades que miden la abundancia relati-va de peces y la riqueza taxonómica (Grayson,1984; Zangrando, 2009).
MATERIALES Y MÉTODOS
El material faunístico proviene de cuatro locali-dades arqueológicas (Figura 1), y fue caracteriza-do y discutido en trabajos previos (Gómez Otero,2006; Gómez Otero et al., 2009; Svoboda &Gómez Otero, 2013). Para el presente trabajo secuenta con información ictiofaunística de nueveconjuntos recuperados en contextos estratigráficosasignados al Holoceno tardío (Tabla 1).
Del valle inferior se analizaron los sitios LomaGrande 1 (LG1) y Cinco Esquinas 1 (CE1). Estánubicados sobre antiguos albardones del río y fue-ron interpretados como ocupaciones residencialesdonde se detectaron numerosos relictos de fogónasociados a fauna menor dulceacuícola (GómezOtero, 1994; Gómez Otero et al., 2010). Ambossitios presentan antigüedades del Holoceno tardíofinal (Tabla 1). Las excavaciones fueron realizadasen aquellos sectores donde afloraban fogones.Cabe mencionar que parte de la muestra íctica deLG1 fue extraviada, aunque un análisis preliminarrealizado por el ictiólogo Dr. Atila Gosztonyi per-mitió estimar un MNI de 19 percas.
Los sitios de la desembocadura del río Chubut ycosta inmediata corresponden a las localidadesBarranca Norte (BN) (Gómez Otero, 2006) y LosCangrejales Sur (Gómez Otero et al., 2009). La pri-mera localidad está situada en Bahía Engaño, en lamargen norte de la desembocadura del río. Presen-ta diversidad y alta densidad de sitios que indicanocupaciones reiteradas y el desarrollo de múltiplesactividades (Gómez Otero, 2006). Es interesantedestacar que la distribución de los sitios corre de
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 91
Archaeofauna 24 (2015): 87-101
TABLA 1
Muestreos arqueofaunísticos analizados en este trabajo. Se presentan sus dimensiones, fechados radiocarbónicos y números de especí-menes de peces recuperados.
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 91

este a oeste, es decir en sentido paralelo al río. Estopodría asociarse con períodos en los que el niveldel mar estaba más elevado y había penetrado enBahía Engaño formando un amplio estuario(Monti, 2000). De acuerdo con cuatro datacionesde restos malacológicos, el último pulso de erosiónhabría ocurrido entre 3900 y 3200 años AP; a par-tir de entonces se produjo un ciclo de acreción cor-doniforme (Monti, 2000). Los muestreos corres-ponden a una lente de fogón en el perfil de unmédano (ca. 3060-2960 AP) y a un fogón dereciente exposición a la intemperie (1040 AP)(Tabla 1). Ambos presentan fauna malacológica(trofones y mejillones) y restos de vertebrados. Lalocalidad Los Cangrejales Sur (LC) se ubica en unaplaya de acumulaciones litorales, donde se regis-traron ocupaciones recurrentes y breves ligadas aactividades extractivas (Gómez Otero et al., 2009).Los muestreos fueron realizados en varias lentesestratificadas de fogones en los sectores 4 y 5, quecontenían predominantemente valvas de lapas,peces y aves. Entre ellos, los muestreos S4- L1 y S-5 L 0 inferior presentaron restos de peces en unafrecuencia menor a cuatro especímenes (GómezOtero et al., 2009: tabla 2), por lo cual no fueronconsiderados en el cálculo de los índices de abun-dancia (ver más abajo). Las edades radiocarbónicasobtenidas para depósitos culturales de circa 2000 a2200 AP sugieren que en esos momentos la línea decosta se encontraba entre 2, 50 y 5 m s.n.m.(Gómez Otero et al., 2009).
La metodología general de excavación consis-tió en cuadrículas de un metro de lado en el casode los fogones subsuperficiales y de muestreos de50 cm de lado por la potencia de la lente para losniveles de fogón estratificados (Tabla 1). Para evi-tar la pérdida de microrrestos en el campo, el tami-zado de la totalidad de sedimentos y materiales decada muestreo fue realizado en laboratoriomediante el empleo de zarandas finas de 1,3 mmde malla.
La identificación y cuantificación de los restosícticos se realizó de acuerdo a los procedimientoscomúnmente utilizados en el análisis ictiofaunísti-co (Wheeler & Jones, 1989; Zangrando, 2009).Para la identificación taxonómica aplicamos elmétodo de anatomía comparada con ejemplares dela colección de referencia del Laboratorio deArqueología del Centro Nacional Patagónico(Puerto Madryn, Argentina) y contamos con lacolaboración del Dr. A. Francisco Zangrando. Parala cuantificación de la abundancia taxonómica yanatómica se aplicaron las unidades NISP, MNI y
MNE de acuerdo con los lineamientos planteadospor Zangrando (2009). La estimación de talla depercas se basó sobre las ecuaciones de regresiónpara elementos craneales y de la cintura escapular(Svoboda, 2013a).
A los fines de conocer la variedad de especiesícticas explotadas y evaluar la distribución y abun-dancia relativa de los restos de peces aplicamosuna serie de unidades de medida que permitenmonitorear cambios en los conjuntos zooarqueoló-gicos: 1) el NTAXA o riqueza taxonómica (Gray-son, 1984) para evaluar la riqueza de especies ícti-cas; 2) el índice de equitatividad (V= H/ln S, Reitz& Wing, 1999: 235) para identificar el predominiode algún taxón entre los varios recursos registra-dos; y 3) el índice de abundancia de peces (∑ NISPpeces/ ∑ NISP total, Zangrando, 2009: 142) paraevaluar la contribución de estos en términos denúmeros de especímenes. En todos los casos sepuso especial cuidado en no contabilizar dos vecesel mismo taxón, así como se excluyeron del análi-sis aquellos especímenes, como huesos de peque-ños roedores (y en algunos casos placas de dasipó-didos), cuya presencia puede ser de origen natural.A su vez, para controlar que la cantidad de espe-cies halladas no estuviera mediada por el tamañode la muestra se correlacionó el NTAXA con elNISP (Grayson, 1984:132).
RESULTADOS
Los conjuntos ictioarqueológicos del valle inferior
del río Chubut
El conjunto ictiofaunístico proveniente de LG1esta conformado por un total de 389 restos óseos,de los cuales 226 (58,0%) pudo ser asignado aalguna categoría taxonómica y anatómica, 71(18,2%) a nivel anatómico y los restantes 92(23,6%) corresponden a fragmentos pequeñosindeterminados. La única especie representada esla perca (58,0 NISP%) con un MNI de seis indivi-duos calculado a partir de la primera vértebra pre-caudal. No obstante, el MNI se elevaría a 25 si sesuman los 19 individuos estimados por el Dr.Gosztonyi a partir del mismo elemento óseo(Tabla 2). En términos de abundancia anatómica seencuentran presentes elementos del esqueletocompleto con un predominio del sector axial(MNE= 155; MNE%= 79) (Svoboda & GómezOtero, 2013).
92 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 92
La muestra ictiofaunística de CE1-S3-F2 estáconformada por 1.580 restos de los cuales 337(21,3 %) pudieron ser asignados taxonómicamentey anatómicamente, 512 (32,4%) fueron asignados anivel anatómico y 731 (46,2 %) corresponden afragmentos indeterminados. Entre los taxa presen-tes predominan los peces fluviales representadosúnicamente por la perca (21,2 NISP%), cuyo MNIes de 10 individuos estimados a partir de la primervértebra precaudal. En menor abundancia se haregistrado la especie marina mero (0,1 NISP%),cuyo MNI fue de dos individuos calculado a partirdel otolito (Tabla 2). En términos de abundanciaanatómica de la perca, si bien se halla representadotodo el esqueleto, se observa predominio de ele-mentos axiales (MNE= 238; MNE%= 82).
En ambos conjuntos ninguno de los restos ana-lizados presentó huellas atribuibles al trozamiento;sí se registraron evidencias de combustión en 22especímenes (83%) de LG1 y en 14 restos (0,9 %)de CE1-S3-F2. Por otro lado, los especímenes pre-sentan buena preservación y no se registraron alte-raciones por agentes y procesos tafonómicos, porlo que se descarta que la ubicuidad taxonómica dela perca este afectada por factores posdepositacio-nales. Las evidencias de acción antrópica y lainformación contextual permiten sostener el ori-gen cultural de los depósitos estudiados (Svoboda& Gómez Otero, 2013).
En lo que respecta a las tallas de percas, en tér-minos generales los ejemplares de LG1 son máschicos que los de CE1 (Tabla 3) (véase Svoboda,2013a). No obstante, dado que la muestra LG1 seencuentra incompleta (por la pérdida de restos), esmuy probable que las tallas reales se encuentrensesgadas, por lo que no representa una muestra
confiable para discutir los rangos y distribución detamaños de perca. En el histograma de la Figura 2se presenta la distribución de tallas estimadas paraCE1. Se observa la ausencia de individuos meno-res a 240 mm de largo total y una distribución uni-modal de los datos que es estadísticamente signifi-cativa (Shapiro Wilk; W= 0,97, p > 0,05). Este testestadístico contribuye a la discusión del origen delos depósitos (natural o cultural), así como a lareconstrucción de la forma de captura de peces(Limp & Reidhead, 1979).
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 93
Archaeofauna 24 (2015): 87-101
TABLA 2
Número de especímenes identificados por taxón, Número mínimo de individuos y riqueza taxonómica por muestreo ictiofaunístico.Referencias: *: 25 individuos si se considera la muestra extraviada.
TABLA 3
Largo total (en mm) estimado para perca (P. trucha) de los sitiosLoma Grande 1 y Cinco Esquinas 1.
FIGURA 2
Distribución de la talla de percas (P. trucha) del sitio CincoEsquinas 1.
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 93

Los conjuntos ictioarqueológicos de la desembo-
cadura del río Chubut y la costa inmediata
En la mayoría de los conjuntos de BN y LC laidentificación se dificultó por la presencia deabundantes espinas que obstaculizaron la determi-nación taxonómica específica. Una excepción es elconjunto ictiofaunístico de BN1-F1 que presentóun total de 31 restos ícticos donde 23 especímenes(74,1%) pudieron ser asignados a nivel específicoy a una unidad anatómica, mientras que ocho (25,8%) únicamente a nivel anatómico. La abundanciataxonómica señala el predominio de pejerrey (41,9NISP%) y róbalo (32,2 NISP %), con un MNI deun individuo para cada especie (Tabla 2). Las par-tes esqueletales corresponden en su totalidad avértebras: el NME para pejerrey es 13 y para róba-lo 10.
El conjunto de BN2-N2 se compone de 272especímenes, entre los cuales 114 (41,5 %) fueronidentificados anatómica y taxonómicamente, 94(34,5%) sólo a nivel anatómico y 64 (23,3%)corresponden a fragmentos indeterminados. Lasespecies más abundantes son el pejerrey (21,6NISP%) y el róbalo (11,3 NISP%), siguiéndoles laanguila (4,7 NISP%), el mero (1,4 NISP%) y eltiburón (0,7 NISP%) (Tabla 2). A partir de la pri-mera vértebra precaudal o del recuento de vérte-bras (según la especie) se contabilizó un MNI detres pejerreyes, dos róbalos y un ejemplar paracada una de las restantes especies. En términos deabundancia anatómica hay un predominio absolu-to de la región axial en pejerrey (MNE= 58;MNE%= 100), róbalo (MNE= 39; MNE%= 100) yanguila (MNE= 13; MNE%= 100); con excepciónde la presencia de dos especímenes de merocorrespondientes al cráneo (maxila y premaxilar) yde dientes de la mandíbula del tiburón.
En LC-S4 los muestreos de las lentes L2a, L2b,L2c y L3 presentaron 790 especímenes, entre ellos152 (19,2 %) fueron identificados taxonómica yanatómicamente, los restantes 429 (54,3%) sólo anivel anatómico y 209 (26,4%) corresponden afragmentos indeterminados. En LC-S5 el conjuntoictiológico L1 se compone de 70 restos, de los cua-les sólo se identificaron taxonómicamente seis(3,6%); mientras que el conjunto L0-inf se compo-ne de cuatro especímenes, de los cuales dos fueronasignados a mero. Las especies con mayor abun-dancia relativa en LC-S4 y LC-S5 son róbalo (10,8 NISP%) y pejerrey (6,3 NISP%) (Tabla 2). Entotal se estimó un número mínimo de 16 indivi-
duos: nueve róbalos, cinco pejerreyes, un bagre demar y un mero. La abundancia de elementos ana-tómicos presenta una mayoría de vértebras enpejerrey (MNE=52; MNE%= 100%) y róbalo(MNE= 80; MNE%= 94%).
En los conjuntos de BN y LC no se registraronhuellas de corte, aunque sí evidencias de combus-tión en dos vértebras (6,4%) de BN1-F1, y en 84especímenes (10,6%) –mayormente vértebras– delas lentes LC-S4. Algunas modificaciones postde-positacionales fueron observadas en los muestreosde ambas localidades. Entre ellas, se registró cam-bio en la forma (generalmente aplastamiento) devértebras de róbalo de los conjuntos: BN2-N2 (n=9; 3,3%), LC-S4 L2a (n= 14; 3,5%), LC-S4 L3 (n=5; 3,0%) y LC-S4 L2c (n= 3; 3,6%); varias de ellas(n=7) se encontraban también carbonizadas. Cabemencionar que ningún espécimen presentó trazasdiagnosticas (hoyuelos y redondeamiento) produ-cidas por procesos digestivos (Butler & Schroeder,1998). Finalmente, una baja proporción de vérte-bras (n=34; 12,5%) correspondientes a BN2-N2presentaron exfoliaciones y agrietamientos que,según las observaciones de Svoboda & Moreno(2014), sugieren un estado avanzado de meteoriza-ción.
Abundancia taxonómica y riqueza íctica
En la Figura 3 se observa la distribución de losvalores de los índices de equitatividad y de abun-dancia de peces para las muestras faunísticas delvalle y de la desembocadura y costa inmediata. Encuanto al índice de equitatividad, los valores pró-ximos a 1 (uno) indican que los taxa presentes seencuentran en una proporción semejante entre sí yque los valores cercanos a 0 (cero) indican ladominancia de una especie. En este sentido, entodos los muestreos de LC-S4 y LC-S5 y en CE1-S3-F2 se observó el predominio marcado de unúnico taxón. Además de los restos de peces, en elúltimo conjunto se halló un espécimen asignable acoipo, mientras que en LC-S4 se hallaron ochoespecímenes de aves, uno de ellos correspondien-te a pingüino (Gómez Otero et al., 2009) (Tabla 4).En BN1-F1 y BN2-N2 los valores del índice seña-lan una importancia equitativa de varios taxa comopeces, aves, guanaco y pinnípedos; cabe mencio-nar además que en BN1-F1 se determinó un MNIde 13 pinnípedos, el más alto registrado en el áreade estudio (Gómez Otero, 2006) (Tabla 4). Por
94 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 94
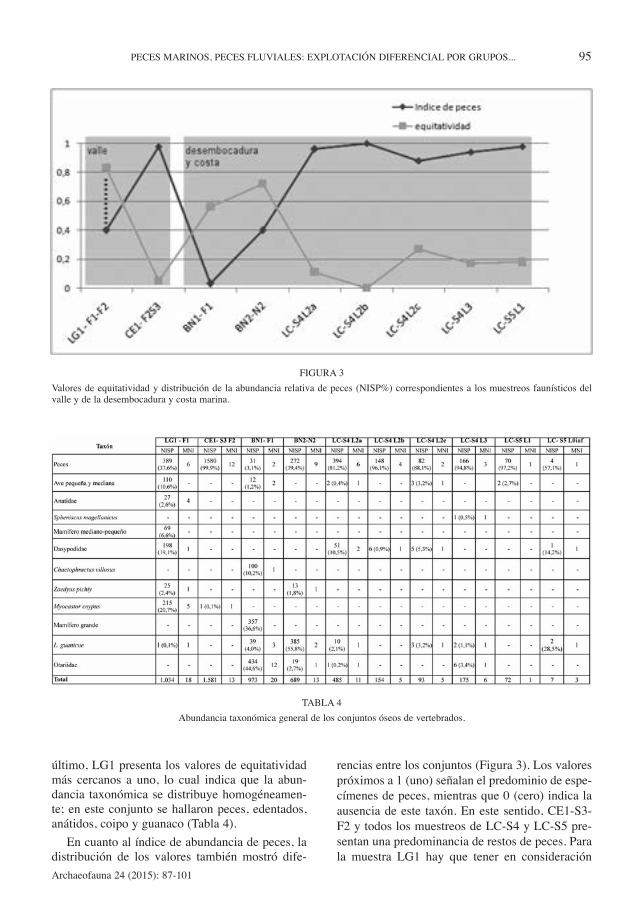
último, LG1 presenta los valores de equitatividadmás cercanos a uno, lo cual indica que la abun-dancia taxonómica se distribuye homogéneamen-te; en este conjunto se hallaron peces, edentados,anátidos, coipo y guanaco (Tabla 4).
En cuanto al índice de abundancia de peces, ladistribución de los valores también mostró dife-
rencias entre los conjuntos (Figura 3). Los valorespróximos a 1 (uno) señalan el predominio de espe-címenes de peces, mientras que 0 (cero) indica laausencia de este taxón. En este sentido, CE1-S3-F2 y todos los muestreos de LC-S4 y LC-S5 pre-sentan una predominancia de restos de peces. Parala muestra LG1 hay que tener en consideración
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 95
Archaeofauna 24 (2015): 87-101
FIGURA 3
Valores de equitatividad y distribución de la abundancia relativa de peces (NISP%) correspondientes a los muestreos faunísticos delvalle y de la desembocadura y costa marina.
TABLA 4
Abundancia taxonómica general de los conjuntos óseos de vertebrados.
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 95
que en el valor del índice debería ser más altodebido a que parte de la muestra fue extraviada(línea punteada en la Figura 3). Las muestras deBarranca Norte presentan los valores más bajos(Figura 3). Finalmente, al correlacionar los índicesde abundancia de peces y de equitatividad seobtiene una relación altamente significativa (r2= -0,85; p< 0,05), lo que implica que los conjuntoscon mayor abundancia de peces son los menosequitativos.
En suma, en los conjuntos de CE1-S3-F2(valle) y LC-S4 y LC-S5 (costa marina) hay unelevado predominio de peces por sobre los demásrecursos. En LG1 los recursos explotados sondiversos y la abundancia de peces es moderada,pero podría aumentar considerablemente (19 sonlos individuos que desaparecieron). En BN1-F1 elespectro faunístico es muy variado y los peces pre-sentan muy baja frecuencia relativa. Estos resulta-dos son similares a los de BN2-N2, aunque en esteúltimo aumenta la contribución numérica de peces.
En cuanto a la riqueza íctica, en el valle seregistraron solamente dos taxa de distinto origen:perca (hábitat fluvial) y mero (hábitat marino)(Tabla 2). En los sitios de la desembocadura y lacosta inmediata la mayor riqueza íctica, todasespecies marinas, varía entre cinco a un taxón.Como puede observarse en la Figura 4 la riquezataxonómica aumenta moderadamente en funciónde la cercanía al litoral marino. Sin embargo, estarelación no es estadísticamente significativa (r2 =0,1; p > 0,05). Para explorar variaciones tempora-les, y dado que en los conjuntos costeros se cuen-ta con varios fechados radiocarbónicos para una
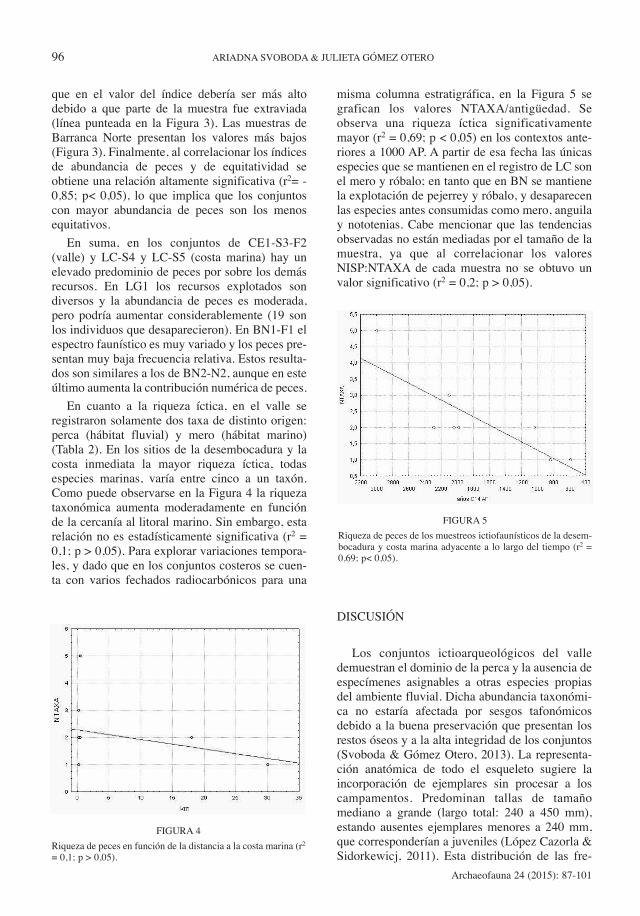
misma columna estratigráfica, en la Figura 5 segrafican los valores NTAXA/antigüedad. Seobserva una riqueza íctica significativamentemayor (r2 = 0,69; p < 0,05) en los contextos ante-riores a 1000 AP. A partir de esa fecha las únicasespecies que se mantienen en el registro de LC sonel mero y róbalo; en tanto que en BN se mantienela explotación de pejerrey y róbalo, y desaparecenlas especies antes consumidas como mero, anguilay nototenias. Cabe mencionar que las tendenciasobservadas no están mediadas por el tamaño de lamuestra, ya que al correlacionar los valoresNISP:NTAXA de cada muestra no se obtuvo unvalor significativo (r2 = 0,2; p > 0,05).
DISCUSIÓN
Los conjuntos ictioarqueológicos del valledemuestran el dominio de la perca y la ausencia deespecímenes asignables a otras especies propiasdel ambiente fluvial. Dicha abundancia taxonómi-ca no estaría afectada por sesgos tafonómicosdebido a la buena preservación que presentan losrestos óseos y a la alta integridad de los conjuntos(Svoboda & Gómez Otero, 2013). La representa-ción anatómica de todo el esqueleto sugiere laincorporación de ejemplares sin procesar a loscampamentos. Predominan tallas de tamañomediano a grande (largo total: 240 a 450 mm),estando ausentes ejemplares menores a 240 mm,que corresponderían a juveniles (López Cazorla &Sidorkewicj, 2011). Esta distribución de las fre-
96 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
FIGURA 4
Riqueza de peces en función de la distancia a la costa marina (r2
= 0,1; p > 0,05).
FIGURA 5
Riqueza de peces de los muestreos ictiofaunísticos de la desem-bocadura y costa marina adyacente a lo largo del tiempo (r2 =0,69; p< 0,05).
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 96

cuencias de talla se asemeja a las curvas de selec-tividad antrópica vinculadas con la captura enmasa (Greenspan, 1998: figura 11). En el valle delrío Chubut los lugares más favorables para la cap-tura en masa son las lagunas ribereñas. A modo deejemplo, Mac Donagh (1950) pudo pescar nume-rosas percas mediante una red de arrastre en unalaguna ubicada en el curso inferior del río Colora-do, a 500 km al norte del río Chubut. El autor con-signa tallas de entre 150 mm y 449 mm de largototal, lo cual se asemeja parcialmente a las obteni-das en CE1. Todo esto sugiere que en los casosarqueológicos aquí tratados se habría aplicado laestrategia de captura en masa. No obstante, hayque considerar la posibilidad de que el conjuntoictiofaunístico de CE1 haya sido acrecional yrepresente distintos episodios de captura indivi-dual. Por el momento no puede determinarse latécnica de captura ya que no se cuenta con ningúnotro tipo de evidencia arqueológica pesquera comolos pesos líticos recuperados en áreas marinas cer-canas (Gómez Otero, 2006; Scartascini & Cardi-llo, 2009, entre otros). De haber ocurrido la captu-ra en masa, podrían haberse aplicado otrosmétodos, como el uso de canastas, bolsas, ramas(Limp & Reidhead, 1979) e incluso las manos enel caso de peces que quedan encharcados (Stewart,1994). De todas formas, dado que estas tecnologí-as utilizan componentes orgánicos, son muy bajasa nulas las posibilidades de preservación de losmismos.
En lo que respecta al espectro de recursosexplotados, los contextos del valle se asemejanentre sí en cuanto al aprovechamiento casi exclu-sivo de fauna dulceacuícola (Svoboda & GómezOtero, 2013). No obstante, a partir de los índicesde abundancia aplicados a los conjuntos CE1-S3-F2 y LG1-F1 se observaron diferencias leves encuanto a la contribución numérica de los peces. EnCE1-S3-F2 hay un predominio casi absoluto depeces; sin embargo, en otros muestreos de estesitio se ha constatado la explotación de anátidos ycoipos en proporciones relevantes, aunque éstosno superan la abundancia relativa de las percas(Svoboda & Gómez Otero, 2013). Por su parte, enLG1-F1 se observó una importancia equivalentelos recursos dulceacuícolas (peces, aves, coipo), ymenor abundancia de peces con respecto a CE1;no obstante, esta proporción podría incrementarsesi se contabilizaran los restos extraviados.
En suma, por el momento los datos aquí pre-sentados permiten avalar la expectativa inicial deuna menor riqueza íctica y mayor registro de peces
grasos en el valle. La corroboración de que laperca fue la especie íctica de mayor importanciaeconómica, podría deberse a la búsqueda de fuen-tes genuinas de lípidos ante la ausencia de recursosde alto contenido calórico como los mamíferosmarinos. No obstante, tal como se propuso ante-riormente (Gómez Otero et al., 2010) tambiénpodría deberse a factores situacionales, en estecaso las periódicas grandes crecientes del río, quepropiciaban la formación y perduración de lagunasribereñas, y por ende, la colonización de faunadulceacuícola, pero el alejamiento de las grandespresas terrestres. En este sentido, es importantetener en cuenta que, a diferencia de otros pecesfluviales, la perca es la única especie que podríasobrevivir en condiciones de apoxia (sin oxígeno)durante períodos relativamente prolongados enambientes lagunares, lo que propiciaría su fácilcaptura. Cabe destacar que los primeros resultadossobre la estacionalidad estimada a partir de la lec-tura de anillos de crecimiento en otolitos indicanque los individuos fueron capturados en distintasépocas del año (Svoboda, 2013b).
Los conjuntos ictioarqueológicos de la desem-bocadura y costa presentan una mayor riqueza. Lasespecies más abundantes y recurrentes son el peje-rrey y el róbalo, y en menor medida se hallan otrasespecies litorales y de fondos rocosos. Entre ellas,el róbalo es la única que presentó deformaciónplástica en los elementos vertebrales. Esto, que seha observado en otros contextos costeros de Pata-gonia, se debería principalmente a la acción deprocesos digestivos de agentes ictiófagos comootros peces, aves y mamíferos marinos (Zangran-do, 2003; Scartascini, 2012). No obstante, dadoque algunas vértebras deformadas presentan ras-tros de combustión y que no se registraron otrasevidencias de procesos digestivos (hoyuelos,redondeamiento, pulimento), se infiere que elaplastamiento se relacionaría más con factorespostdepositacionales tales como el pisoteo o elpeso del sedimento. En lo que respecta al perfilanatómico de las especies más abundantes (peje-rrey y róbalo) hay un predominio de la porciónaxial. Este patrón anatómico se condice con elmayor rendimiento cárnico de la región poscrane-al (Butler, 1993), de modo que en estos sitios elespinazo se habría descartado luego de su coccióny consumo.
En cuanto a las formas de obtención de lospeces, en la desembocadura y la costa marina,podría haberse realizado mediante la captura indi-vidual (líneas, arpones) y/o en masa (redes, tram-
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 97
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 97

pas). Una vía para evaluar las técnicas de pescatiene que ver con la etología de los peces captura-dos (Morales Muñiz, 2008). En este sentido, losróbalos y pejerreyes que forman cardúmenespodrían ser capturados con redes, pero tambiénotras técnicas individuales podrían ser empleadas.Por otro lado, los peces de arrecife, como el mero,tienen un comportamiento solitario y se escondenen cuevas, por lo cual la captura individual resultala estrategia más adecuada. En efecto, en un nivel(sector 5- L0 inferior) de Los Cangrejales Surdatado en 590 ± 70 AP se halló un peso lítico aso-ciado con cuatro restos correspondientes a mero.Cabe tener en cuenta también otra modalidad decaptura que no requiere tecnologías, como la reco-lección manual de anguilas o nototenias en elintermareal rocoso.
Paralelamente, los muestreos de la desemboca-dura y costa marina muestran una leve tendenciaen la disminución de los taxa explotados luego del1000 AP: sólo mero y róbalo en Los CangrejalesSur y pejerrey y róbalo en Barranca Norte. Estamenor diversidad de especies ícticas podría rela-cionarse con distintas causas, como cambios en ladisponibilidad de estos recursos a través del tiem-po y/o decisiones humanas.
Con referencia a la contribución numérica depeces en los sitios de la desembocadura y costamarina se observaron diferencias entre los mues-treos de ambas localidades. En BN los conjuntosBN1-F1 y BN2-N2 sugieren la explotación de unaamplia variedad de recursos, entre ellos pinnípe-dos y guanaco, pero sólo en el segundo los pecestuvieron un aporte moderado. Cabe recordar queen BN1-F1 los pinnípedos (13 individuos) fueronla principal fuente energética. Además, no se halla-ron peces en los restantes muestreos de esta loca-lidad (Gómez Otero, 2006). Contrariamente, enLC todos los muestreos señalan la explotación casiexclusiva de peces. En síntesis, en BN se pudoconstatar bajo aprovechamiento y en LC explota-ción intensiva y regular. Las diferencias observa-das se relacionarían con la funcionalidad de estaslocalidades dentro del sistema de asentamiento. Eneste sentido, BN fue interpretada como una locali-dad de actividades múltiples con ocupaciones pro-longadas, donde se habrían explotado variedad demoluscos y mamíferos, entre ellos los pinnípedosque aportan lípidos en abundancia. En cambio enLC las ocupaciones fueron recurrentes, breves yvinculadas con actividades extractivas en las quese habrían aprovechado principalmente peces y
moluscos (Gómez Otero et al., 2009). Por lo tanto,solamente en Barranca Norte se cumple la expec-tativa inicial para los sitios de la desembocadura ycosta inmediata.
CONCLUSIONES
El objetivo principal fue indagar sobre la explo-tación de peces durante el Holoceno tardío en dosáreas de Patagonia central definidas por su varia-bilidad ecológica. En el valle inferior del río Chu-but la pesca de percas parece haber tenido un rolimportante, aunque por el momento el registroarqueológico no permite determinar con certeza sisu explotación fue una práctica regular o situacio-nal, aprovechando las ventajas para su obtencióndurante las inundaciones (Gómez Otero et al.,2010). Por su parte, en la costa marina cercana a ladesembocadura, tal como era esperado, se explotóuna amplia variedad de especies ícticas, aunque elgrado de contribución de los peces a la dieta pare-ce vincularse más a la funcionalidad de las ocupa-ciones que a la oferta de recursos de alto valorcalórico, como los mamíferos marinos. Comoagenda para el futuro, queda entonces ampliar elestudio a otros contextos ictiofaunísticos de laregión para continuar explorando la validez delmodelo de subsistencia y si se corrobora la ten-dencia de menor diversidad íctica en los sitios mástardíos.
AGRADECIMIENTOS
A Francisco Zangrando por la colaboración enel análisis de los conjuntos y sus comentarios a unaversión preliminar. A los evaluadores SergioBogan y un anónimo por sus comentarios y suge-rencias. Este trabajo fue realizado con el financia-miento del Consejo Nacional de InvestigacionesCientíficas y Técnicas (Argentina) y de los subsi-dios PIP-CONICET 6470 y 11401000100210. Elcontenido de este artículo fue presentado en elsimposio «La fauna menor en los conjuntos arque-ofaunísticos sudamericanos: ¿agentes disturbado-res o recursos económicos?», organizado porPaula Escosteguy y Romina Frontini en el marcodel III Congreso Nacional de ZooarqueologíaArgentina.
98 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 98

REFERENCIAS
AMALFI, M.N. 2009: Consideraciones sobre las percas(Percichthys colhuapiensis y P. trucha) de la Patago-nia Norte. Años 1955 a 1957. Comparaciones conmaterial de años recientes. Probiota 10: 1-39.
BAILEY, G. & PARKINGTON, J. 1988: The Archaeology ofPrehistoric Coastlines: an Introduction. In: The
Archaeology of Prehistoric Coastlines: 1-10, Cam-bridge University Press, New Directions in Archaeol-ogy, Nueva York (Traducido por Luis Abel Orquera).
BETTINGER, R.L. 1991: Hunter-Gatherers. Archaeologi-
cal and Evolutionary Theory. Interdisciplinary Con-
tributions to Archaeology. Plenum Press, New York.
BROUGHTON, J.M. 1999: Resource Depression and Inten-
sification during the Late Holocene, San Francisco
Bay: Evidence from the Emeryville Shellmound Ver-
tebrate Fauna. University of California Press, Berke-ley.
BUTLER, V.L. 1993: Natural versus cultural salmonidremains: origin of the Dalles Roadcut Bones, Colum-bia River, Oregon, U.S.A. Journal of Archaeological
Science 20: 1-24.
BUTLER, V.L. & CAMPBELL, S.K. 2004: Resource Inten-sification and Resource Depression in the PacificNorthwest of North America: A ZooarchaeologicalReview. Journal of World Prehistory 18(4): 327-405.
BUTLER, V.L. & SCHROEDER, R.A. 1998: Do digestiveprocesses leave diagnostic traces on fish bones?Journal of Archaeological Science 25: 957-971.
CABRERA, A.L. & YEPES, J. 1960: Mamíferos Sudameri-
canos. Editorial Ediar, Buenos Aires.
CASTEEL, R.W. 1976: Fish Remains in Archaeology and
Paleo-environmental Studies. Academic Press, Lon-don.
ERLANDSON, J.M. 2001: The Archaeology of AquaticAdaptations: Paradigms for a New Millennium.Journal of Archaeological Research 9(4): 287-350.
GÓMEZ OTERO, J. 1994: Sitio Loma Grande. In: Guía de
Campo de la VII Reunión de Campo del CADIN-
QUA: 66-67. Centro Nacional Patagónico, PuertoMadryn.
GÓMEZ OTERO, J. 2006: Dieta, uso del espacio y evolu-ción en poblaciones cazadoras-recolectoras de lacosta centro-septentrional de Patagonia durante elHoloceno medio y tardío. Tesis Doctoral inédita,Universidad de Buenos Aires.
GÓMEZ OTERO, J.; WEILER, N. & MORENO, E. 2009:Localidad arqueológica Los Cangrejales Sur: eviden-cias de ocupaciones humanas y de variaciones de lalínea de costa en el Holoceno tardío. En: Salemme,M.; Santiago, F.; Álvarez, M.; Piana, E.; Vázquez, M.& Mansur, E. (eds.): Arqueología de la Patagonia,
una mirada desde el último confín: 1023-1036.CADIC. Ushuaia.
GÓMEZ OTERO, J.; MORENO, E. & SCHUSTER, V. 2010:Ocupaciones tardías en el valle inferior de río Chu-but: primeros resultados del sitio Cinco Esquinas.En: Actas del XVII Congreso Nacional de Arqueolo-
gía Argentina: 1917-1922. Universidad Nacional delCuyo, Mendoza.
GÓMEZ OTERO, J.; WEILER, N.; BANEGAS, A. & MORENO,E. 2013: Ocupaciones del Holoceno medio en BahíaCracker, costa atlántica de Patagonia Central. En:Zangrando, A.F.; Barberena, R.; Gil, A.; Neme, G.;Giardina, M.; Luna, L.; Otaola, C.; Paulides, S.; Sal-gán, L. & Tivoli, A. (eds.): Tendencias teórico-meto-
dológicas y casos de estudio en la arqueología de la
Patagonia: 177-186. Museo de Historia Natural deSan Rafael, San Rafael.
GOSZTONYI, A.E. 1988: Peces del río Chubut inferior,Argentina. Physis. Secc. B 46(110): 41-50.
GRAYSON, D.K. 1984: Quantitative Zooarchaeology.Academic Press, Orlando.
GRAYSON, D.K. & CANNON, M. 1999: Human paleoecol-ogy and foraging theory in the Great Basin. In: Beck,C. (ed.): Models for the Millennium: Great Basin
Anthropology Today: 141-150. University of UtahPress, Salt Lake City.
GREENSPAN, R.L. 1998: Gear Selectivity Models, Mor-tality Profiles and the Interpretation of Archaeologi-cal Fish Remains: a Case Study from the HarneyBasin, Oregon. Journal of Archaeological Science
25: 973-984.
IRIGOYEN, A.J. & GALVÁN, D. 2010: Peces de arrecife
argentinos. Proyecto arrecife. Puerto Madryn.
LASTA, M.L.; CIOCCO, N.F.; BREMEC, C. & ROUX, A.1998: Moluscos bivalvos y gasterópodos. En: El Mar
Argentino y sus recursos pesqueros: 115-143. INI-DEP, Mar del Plata.
LEÓN, R.; BRAN, D.; COLLANTES, M.; PARUELO, J.M. &SORIANO, A. 1998: Grandes unidades de vegetaciónde la Patagonia extra andina. Ecología Austral 8(2):75-308.
LIMP, W.F. & REIDHEAD, V.A. 1979: An Economic Eval-uation of the Potential of Fish Utilization in RiverineEnvironments. American Antiquity 44(1): 70-78.
LINDSTROM, S. 1996: Great Basin fisherflok: optimaldiet breadth modeling the Truckee river aboriginalsubsistence fishery. In: Plew, M.G. (ed.): Prehistoric
hunter-gatherer fishing strategies: 114-179. BoiseState University, Boise.
LÓPEZ, G.R. & LIPPS, E.F. 1988: Análisis de composi-ción química de especies ícticas patagónicas de aguadulce. In: Segunda Reunión Argentina de Acuicultu-
ra: 40. Puerto Madryn.
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 99
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 99

LÓPEZ CAZORLA, A. & SIDORKEWICJ, N. 2011: Age,growth and reproduction in creole perch (Percichthys
trucha) in the Negro River, Argentinean Patagonia.Journal of Applied Ichthyology 27: 30-38.
LUPO, K.D. 2007: Evolutionary Foraging Models inZooarchaeological Analysis: Recent applications andFuture Challenges. Journal of Archaeological
Research 15: 143-189.
LUPO, K.D. & SCHMITT, D. 2005: Small prey huntingtechnology and zooarchaeological measures of taxo-nomic diversity and abundance: Ethnoarchaeologicalevidence from Central African forest foragers. Jour-
nal of Anthropological Archaeology 24: 335-353.
MAC DONAGH, E.J. 1950: Las razas de percas o truchascriollas (Percichthys) y su valor para la repoblaciónpesquera. Revista del Museo de La Plata 6: 71-170.
MADSEN, D.B. & SCHMITT, D.N. 1998: Mass collectingand the diet-breadth model: A Great Basin example.Journal of Anthropological Archaeology 25: 445-55.
MALAINEY, M.E.; PRZYBYLSKI, R. & SHERRIFF, B.L.2001: One Person’s Food: How and Why Fish Avoid-ance May Affect the Settlement and Subsistence Pat-terns of Hunter-Gatherers. American Antiquity 66(1):141-161.
MARTINEZ, G.; ZANGRANDO, F. & STOESSEL, L. 2005:Sitio El Tigre (Pdo. de Patagones, Pcia. de BuenosAires, Argentina): evidencias sobre la explotación depeces en el curso inferior del Río Colorado e impli-caciones para los sistemas de subsistencia. Magalla-
nia 33: 127-142.
MATTHEWS, A. 1992: Crónica de la colonia galesa de la
Patagonia. Editorial El Regional, Rawson.
MONTI, A.J. 2000: Edades 14C y ciclicidad de la acre-ción en depósitos costeros elevados. Bahía Engaño,Chubut. Revista de la Asociación Geológica Argenti-
na 55(4): 403-406.
MORALES MUñIZ, A. 2008: De los peces a las redes: lasartes de pesca desde una perspectiva arqueoictiológi-ca. Archaeobios 2: 40-63.
NAGAOKA, L. 2002: The effects of resource depressionon foraging efficiency, diet breadth, and patch use insouthern New Zealand. Journal of Anthropological
Archaeology 21: 419-442.
NEME, G. & GIL, A. 2008: Biogeografía humana en losAndes meridionales: tendencias arqueológicas en elsur de Mendoza. Chungara 40(1): 5-8.
PERLMAN, S.M. 1980: An Optimum Diet Model, CoastalVariability and hunter-gatherer Behavior. In: Schif-fer, M.B. (ed.): Advances in archaeological Method
and Theory: 257- 310. Vol. 3. Academic Press, NewYork.
PRONSATO, A.D. 1950: Estudio geohidrológico del RíoChubut. Parte II. Dirección General de Agua y Ener-gía Eléctrica. Revista Agua y Energía 29 y 30.
REITZ, E. & WING, E. 1999: Zooarchaeology. CambridgeUniversity Press, Cambridge.
SAADOUN, A. & CABRERA, M.C. 2008: A review of thenutritional content and technological parameters ofindigenous sources of meat in South America. Meat
Science 8(3): 570-681.
SCARTASCINI, F.L. 2012: Primeras tendencias ictioarque-ológicas en la localidad Bajo de La Quinta, RíoNegro, Argentina. Intersecciones en Antropolo-
gía 13: 315-326.
SCARTASCINI, F.L. & CARDILLO, M. 2009: Explorando lavariabilidad métrica y morfológica de las «pesas líti-cas» recuperadas en el sector norte de la costa delgolfo San Matías. En: Palacios T.; Palacios, O.; Váz-quez, C. & Cabanillas, E. (eds.): Arqueometría lati-
noamericana: Segundo Congreso Argentino y Prime-
ro Latinoamericano: 162-168. Comisión Nacional deEnergía Atómica, Buenos Aires.
SCARTASCINI, F.L. & VOLPEDO, A. 2013: White croaker(Micropogonias furnieri) paleodistribution in theSouthwestern Atlantic Ocean. An archaeological per-spective. Journal of Archaeological Science 40(2):1059-1066.
SCHIAVINI, A. 1994: Los lobos marinos como recursopara cazadores-recolectores marinos: el caso de Tie-rra del Fuego. Latin American Antiquity 4: 346-366.
SMITH, E.A. 1983: Anthropological Applications ofOptimal Foraging Theory. Current Anthropology 24:625-651.
STEWART, K.M. 1994: Early hominid utilisation of fishresources and implications for seasonality and behav-iour. Journal of Human Evolution 27: 229-245.
STINER, M.C.; MUNRO, N.D. & SUROVELL, T.A. 2000:The tortoise and the hare: small-game uses, thebroad-spectrum revolution and Paleolithic demogra-phy. Current Anthropology 41: 39-73.
SVOBODA, A. 2013a: Disponibilidad cárnica, rendimien-to energético y estimación de la talla de Percichthys
trucha (perca criolla) a partir de la morfometría dehuesos diagnósticos y su aplicación a los conjuntosictioarqueológicos de Patagonia Central. Cuadernos
de Antropología 9: 251-266.
SVOBODA, A. 2013b: Método para determinar la estacio-nalidad de ocupación de sitios arqueológicos deambientes fluviolacustres de Patagonia a partir de laobservación de otolitos de percas. La Zaranda de
Ideas, Revista de Jóvenes Investigadores en Arqueo-
logía 9(2): 145-153.
SVOBODA, A. & GÓMEZ OTERO, J. 2013: Explotación defauna dulceacuícola en el valle inferior del río Chu-but (Patagonia Central) durante el Holoceno tardío.Intersecciones en Antropología. En prensa.
SVOBODA, A. & MORENO, J. 2014: Experimentaciónsobre los efectos de la meteorización en la supervi-vencia de elementos óseos de Percichthys trucha:
100 ARIADNA SVOBODA & JULIETA GÓMEZ OTERO
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 100

implicaciones ictioarqueológicas para el sitio DV1,Lago Musters (Prov. de Chubut, Argentina). Revista
Chilena de Antropología 29: 60-67.
VAZ FERREIRA, R. 1976: South American sea lion. Mam-
mals in the seas: 9-11. Vol. II. FAO, Roma.
WHEELER, A. & JONES, A.K. 1989: Fishes. CambridgeUniversity Press, Cambridge.
WINTERHALDER, B.; BAILLARGEON, W.; CAPELLETTO, F.;DANIEL, I.R. & PRESCOTT, C. 1988: The PopulationEcology of Hunter-Gatherers and their Prey. Journal
of Anthropological Archaeology 7: 289-328.
YESNER, D. 1980: Maritime Hunter-Gatherers: Ecologyand Prehistory. Current Anthropology 21(6): 727-750. (Traducción L. A. Orquera).
ZANGRANDO, A.F. 2003: Ictioarqueología del canal Bea-
gle, explotación de peces y su implicación en la sub-
sistencia humana. Sociedad Argentina de Antropolo-gía, Buenos Aires.
ZANGRANDO, A.F. 2009: Historia Evolutiva, tiempos y
subsistencia humana en la región del Canal Beagle.
Una aproximación zooarqueológica. SociedadArgentina de Antropología, Buenos Aires.
PECES MARINOS, PECES FLUVIALES: EXPLOTACIÓN DIFERENCIAL POR GRUPOS... 101
Archaeofauna 24 (2015): 87-101
05. ARCH. VOL. 24 (2ª)_ARCHAEOFAUNA 04/05/15 13:25 Página 101





























