Embed Size (px)
Citation preview

— 93 —
Productividad primaria, biomasa y tamaño del fitoplancton en canales y fiordos
6.2 Productividad primaria, biomasa y tamaño del fitoplanctonen canales y fiordos australes: patrones primavera-verano
Vivian Montecino1 & Gemita Pizarro2
1Facultad de Ciencias. Universidad de ChileE-mail: [email protected]; [email protected] de Fomento PesqueroE-mail: [email protected]
La productividad primaria de los océanos, ge-nerada principalmente por los diminutos organis-mos autotróficos que conforman el fitoplancton,es un elemento fundamental en los ecosistemasmarinos. Este es un proceso de dos etapas, foto-síntesis y biosíntesis. La fotosíntesis o fijación decarbono es mediada por la clorofila contenida enlos cloroplastos de las microalgas (Kirk, 1994). Laconcentración de clorofila-a (Cl-a) es utilizadauniversalmente como una medida de la biomasade fitoplancton. Estas microalgas forman asocia-ciones que interactúan con otros microorganismosconstituyendo una red microbiana que regula elreciclado de los nutrientes y del carbono in situ,su transferencia a los niveles tróficos superioreso su sedimentación hacia las aguas profundas.
El fitoplancton tiene un amplio espectro de for-mas y tamaños. En sistemas poco productivos,los organismos de menor tamaño (< 5 µm) sonmás frecuentes y abundantes; mientras que losorganismos de mayor tamaño (>20 µm) omicrofitoplancton dominan en aguas eutróficas,que corresponden a sistemas más productivos,ricos en fósforo y nitrógeno. En consecuencia, ladinámica del fitoplancton con relación al ambien-te local y a otros organismos, es de particular re-levancia para estimar la productividad biológicadel ecosistema. Como un componente de los pro-cesos biogeoquímicos, la productividad primaria,con valores de 1 g·m–2·d –1 en promedio, permitecomprender la función del fitoplancton en la bom-ba de carbono que reduce el CO
2 atmosférico.
La productividad primaria se estudia deacuerdo a la escala temporal en que ocurrenlos procesos fotosintéticos y el crecimiento delfitoplancton. Es así como a menor escala, es-tos experimentos se realizan, in situ o in vitro,con una duración de minutos-horas a horas-días
dependiendo de los objetivos planteados. Amayor escala, las interrogantes se enfocan ha-cia la variabilidad estacional, intraestacional einteranual en la columna de agua (vertical) y ho-rizontal (mesoescala). En el primer caso, lasincertezas son fisiológicas, mientras que en elsegundo son ecológicas (Marra, 2002).
Durante los cruceros CIMAR 2 a 4 Fiordosrealizados desde la boca del Guafo al cabo deHornos (Figs. 1 y 2), se ha estudiado una extensa
Avances en el conocimiento oceanográfico de las aguas interiores chilenas, Puerto Montt a cabo de Hornos.N. Silva & S. Palma (eds.)Comité Oceanográfico Nacional - Pontificia Universidad Católica de Valparaíso, Valparaíso, pp. 93-97, 2006.
Figura 1: Posición geográfica de las estaciones de muestreopara la determinación de productividad primaria ybiomasa fitoplanctónica en el crucero CIMAR 2Fiordos.
91
G. de Penas
C. Messier
S. Eyre
E. Falcon
S. Penguin
S. EuropaE. Peel
C. Smyth
C. Concepción
C. Ladrillero
E. de Magallanes
C. Baker
E. Steffen
E. Mitchel
S. Iceberg
A. Inglesa
C. Picton
C. Trinidad
E. Amalia
C. Fallos
CIMAR 2 Fiordos
Océ
ano
Pac
ífico
E. Calvo
92
17
75
90
85
86
87
88
81
84
82
77
7879
12
44
43
42
4548
50
61
67
74
73
70
75
71
29
2728
26
25
24
20
393836
35
32 3331
76
74°76° 73°W75°
52°
51°
50°
48°
49°
47°S
53°

— 94 —
Montecino, V. & G. Pizarro
área geográfica de canales y fiordos australes chile-nos, caracterizada por diferentes masas de agua(Silva et al., 1998; Guzmán & Silva, 2002; Valdenegro& Silva, 2003), donde se efectuaron mediciones debiomasa fitoplanctónica expresada como Cl-a, di-versidad específica estimada mediante el índice deShannon-Weaver (H’) y variedad de tamaños enmuestras superficiales obtenidas en la zona eufóticao iluminada. También se estimó la fijación de carbo-no utilizando un incubador con una fuente de luzartificial, según la metodología descrita por Pizarroet al. (2000).
En el área analizada, el fraccionamiento de labiomasa total mostró que los organismos fitoplanc-tónicos mayores de 20 µm (microfitoplancton) fue-ron recurrentes a meso y macroescala. Además, sedeterminó que la riqueza de especies fue de 17-27para los valores máximos de diversidad H’ y de 5-10para los valores mínimos de H’. La abundancia su-perficial de microfitoplancton y las especies más re-currentes (> 45%) en las tres zonas de estudio fue-ron: Skeletonema costatum (67%) en octubre de1998 y Guinardia delicatula (65%) en febrero de 1999entrre la boca del Guafo y la laguna San Rafael (ZonaNorte); Thalassiosira minuscula (91%) en agosto de1995 y Chaetoceros cinctus (36%) en octubre de
1996 en la zona desde el golfo de Penas al estrechode Magallanes (Zona Central); y Chaetoceros sp.(56%), en octubre de 1998 en la zona del estrechode Magallanes al cabo de Hornos (Zona Sur).
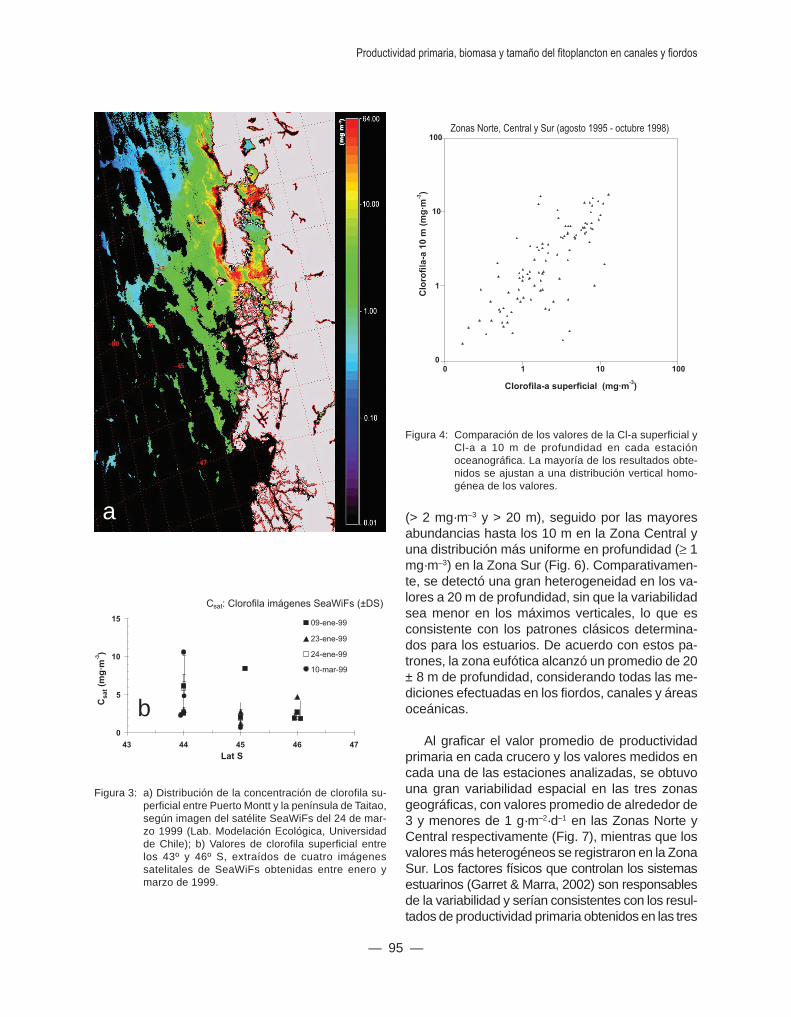
El patrón de distribución más frecuente mostróque las especies numéricamente dominantes fue-ron las mismas en pocos sitios, mientras que lasmás escasas se encontraron en la mayoría de lossitios. Algo similar se observó en la biomasa, quemostró una distribución heterogénea de la clorofilasatelital (Cl-sat), con sitios específicos con altas con-centraciones (> 10 mg Cl-a·m–3) (Fig. 3).
La distribución vertical de la biomasa fitoplanctóni-ca mostró una relación significativa entre la Cl-a super-ficial (0-5 m) y la Cl-a a 10 m de profundidad (Fig. 4),siendo relativamente escasos los sitios con diferen-cias de un orden de magnitud entre estas dos profun-didades. En concentraciones superiores a 1 mg·m–3,el fitoplancton estuvo constituido mayoritariamente porla fracción superior a 20 µm (Fig. 5).
Si se considera los perfiles verticales de Cl-apromedio para las tres zonas, es aparente que enla Zona Norte hubo mayores concentraciones yque alcanzaron también a mayores profundidades
Figura 2: Posición geográfica de las estaciones de muestreo para la determinación de productividad primaria y biomasafitoplanctónica en los cruceros CIMAR 3 (Etapa 2) y 4 Fiordos.
66°W68°70°72°74°
52°S
53°
56°
55°
54°
75º 74° 73º
47º
46º
45º
44º
43ºS
72°W
Océano
Atlántico
I. WollastonC. de Hornos
CIMAR 3 Fiordos (Etapa 2)
Océano Pacífico
C. Cockburn
S. Otway
S. Almirantazgo
1 Angosturaa
2 Angosturaa
C. Beagle
C.Deseado
I. Navarino
E. d
eM
agal
lane
s
B. Inútil
C. Ballenero
C. Froward
P. Dungeness
C. Whiteside
B. Nassau
11
44
42
4140
47
48
20
21
25
7
6
56
54
51
27
3234 35
37Fiordo Aysén
Estero Cupquelán
Estero Quitralco
Canal Puyuguapi
Canal Jacaf
Golfo Elefantes
I.C
hilo
é
BocadelGuafo
Canal Moraleda
Isla Meninea
CIMAR 4 Fiordos
5
27
211917
22
14
4
10
9
8
7
1312
3418
1615
24
23
3032
21A
17A
26
2529
28
3133
6
12A
— 95 —
Productividad primaria, biomasa y tamaño del fitoplancton en canales y fiordos
Figura 4: Comparación de los valores de la Cl-a superficial yCl-a a 10 m de profundidad en cada estaciónoceanográfica. La mayoría de los resultados obte-nidos se ajustan a una distribución vertical homo-génea de los valores.
Figura 3: a) Distribución de la concentración de clorofila su-perficial entre Puerto Montt y la península de Taitao,según imagen del satélite SeaWiFs del 24 de mar-zo 1999 (Lab. Modelación Ecológica, Universidadde Chile); b) Valores de clorofila superficial entrelos 43º y 46º S, extraídos de cuatro imágenessatelitales de SeaWiFs obtenidas entre enero ymarzo de 1999.
(> 2 mg·m–3 y > 20 m), seguido por las mayoresabundancias hasta los 10 m en la Zona Central yuna distribución más uniforme en profundidad ( 1mg·m–3) en la Zona Sur (Fig. 6). Comparativamen-te, se detectó una gran heterogeneidad en los va-lores a 20 m de profundidad, sin que la variabilidadsea menor en los máximos verticales, lo que esconsistente con los patrones clásicos determina-dos para los estuarios. De acuerdo con estos pa-trones, la zona eufótica alcanzó un promedio de 20± 8 m de profundidad, considerando todas las me-diciones efectuadas en los fiordos, canales y áreasoceánicas.
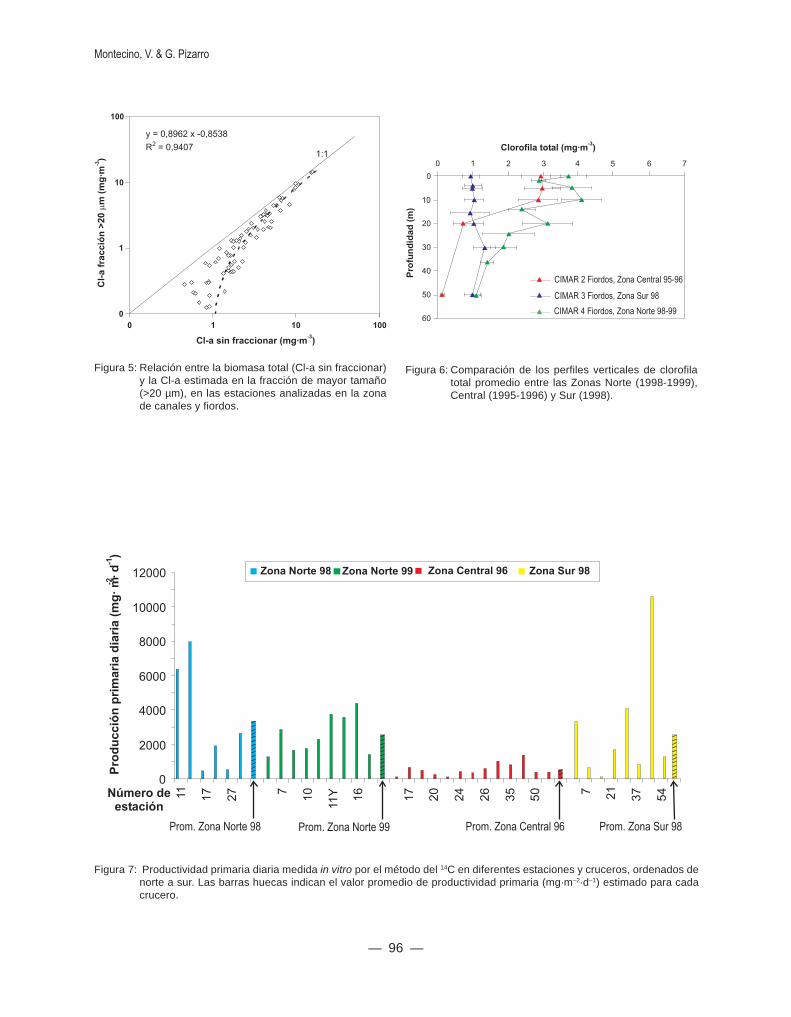
Al graficar el valor promedio de productividadprimaria en cada crucero y los valores medidos encada una de las estaciones analizadas, se obtuvouna gran variabilidad espacial en las tres zonasgeográficas, con valores promedio de alrededor de3 y menores de 1 g·m–2·d–1 en las Zonas Norte yCentral respectivamente (Fig. 7), mientras que losvalores más heterogéneos se registraron en la ZonaSur. Los factores físicos que controlan los sistemasestuarinos (Garret & Marra, 2002) son responsablesde la variabilidad y serían consistentes con los resul-tados de productividad primaria obtenidos en las tres
0
1
10
100
0 1 10 100
Clorofila-a superficial (mg·m )-3
Clo
rofi
la-a
10
m(m
g·m
)-3
Zonas Norte, Central y Sur (agosto 1995 - octubre 1998)
a
Csat: Clorofila imágenes SeaWiFs (±DS)
0
5
10
15
43 44 45 46 47
10-mar-99
09-ene-99
23-ene-99
24-ene-99
Lat S
C(m
g·m
)s
at
-3
b
— 96 —
Montecino, V. & G. Pizarro
Figura 7: Productividad primaria diaria medida in vitro por el método del 14C en diferentes estaciones y cruceros, ordenados denorte a sur. Las barras huecas indican el valor promedio de productividad primaria (mg·m–2·d–1) estimado para cadacrucero.
Figura 5: Relación entre la biomasa total (Cl-a sin fraccionar)y la Cl-a estimada en la fracción de mayor tamaño(>20 µm), en las estaciones analizadas en la zonade canales y fiordos.
Figura 6: Comparación de los perfiles verticales de clorofilatotal promedio entre las Zonas Norte (1998-1999),Central (1995-1996) y Sur (1998).
0
10
20
30
40
50
60
0 1 2 3 4 5 6 7
Clorofila total (mg·m )-3
Pro
fun
did
ad
(m)
CIMAR 3 Fiordos, Zona Sur 98
CIMAR 4 Fiordos, Zona Norte 98-99
CIMAR 2 Fiordos, Zona Central 95-96
y = 0,8962 x -0,8538
R2
= 0,9407
0
1
10
100
0 1 10 100
Cl-a sin fraccionar (mg·m )-3
Cl-
afr
ac
ció
n>
20
m(m
g·m
)�
-3
1:1
0
2000
4000
6000
8000
10000
12000
11
17
27 7
10
11Y
16
17
20
24
26
35
50 7
21
37
54Número de
estación
Pro
du
cció
np
rim
ari
ad
iari
a(m
g·m-2
·d
-1)
Prom. Zona Sur 98Prom. Zona Central 96Prom. Zona Norte 98 Prom. Zona Norte 99
Zona Norte 98 Zona Norte 99 Zona Central 96 Zona Sur 98

— 97 —
Productividad primaria, biomasa y tamaño del fitoplancton en canales y fiordos
zonas analizadas. Así los patrones de distribución ver-tical de la biomasa fitoplanctónica se pueden atribuira diferencias locales en la intensidad de los procesosde mezcla y estratificación; y consecuentemente, conlos procesos de fotoaclimatización de los organismosautotróficos. Las similitudes entre las tres zonas, serefieren a que la abundancia de la Cl-a está determi-nada por la estructura de tamaño de los organismosfitoplanctónicos (Montecino, 2001).
Los patrones de productividad primaria y abun-dancia de Cl-a, son congruentes también con los es-tudios sobre la cantidad de materia orgánica en lossedimentos y el efecto de los glaciares en la zona(Silva et al., 1998). Estos últimos pueden liberar ma-terial inorgánico denominado “glacial silt”, que en al-gunos sectores produciría una “dilución” del conteni-do orgánico de los sedimentos, como también unaatenuación de la productividad primaria, debido a ladisminución de la penetración de luz.
Sin embargo, la limitación de la luz estaría ocu-rriendo también por causa endógena debido a suabsorción por los pigmentos fotosintéticos (Pizarroet al., 2005). Esta condición de limitación de luz,unida a la escasez de otros recursos comonutrientes disueltos, tiende a favorecer a las frac-ciones de menor tamaño del fitoplancton, comocomponente predominante en la biomasa total. Lospatrones descritos son una herramienta para cuanti-ficar la variabilidad de estos ecosistemas a mesoy macroescala.
Referencias
Garrett, A. & J. Marra. 2002. Effects of upper oceanphysical processes (turbulence, advection and air-seainteraction) on oceanic primary production. En: A.Robinson, J. Mc Carthy & B. Rotschild (eds.). The Sea,12: 19-49.
Guzmán D. & N. Silva. 2002. Caracterización física y quí-mica y masas de agua en los canales australes de
Chile entre boca del Guafo y golfo Elefantes (CruceroCIMAR 4 Fiordos). Cienc. Tecnol. Mar, 25(2): 45-76.
Kirk, J. T. O. 1994. Light and photosynthesis in aquaticecosystems. Cambridge University Press, London, 509pp.
Marra, J. 2002. Approaches to the measurements ofplankton production. En: P. J. le B. Williams, D. N. Tho-mas & C. S. Reynolds (eds.). Phytoplankton producti-vity: carbon assimilation in marine and freshwaterecosystems. Blackwell Science, New York, pp. 78-108.
Montecino, V. 2001. Alometría y biodiversidad enfitoplancton en relación con la productividad primariaen ecosistemas pelágicos. En: K. Alveal & T. Antezana(eds.). Sustentabilidad de la biodiversidad, un proble-ma actual, bases científico-técnicas, teorizaciones yproyecciones. Universidad de Concepción, Concep-ción, pp. 199-215.
Pizarro, G., J. L. Iriarte, V. Montecino, J. L. Blanco & L.Guzmán. 2000. Distr ibución de la biomasafitoplanctónica y productividad primaria máxima defiordos y canales australes (47°-50°S) en octubre1996. Cienc. Tecnol. Mar, 23: 25-48.
Pizarro, G., V. Montecino, L. Guzmán, V. Muñoz, V. Cha-cón, H. Pacheco, M. Frangópulos, L. Retamal & C.Alarcón 2005. Patrones locales recurrentes del fito-plancton en fiordos y canales australes (46º-56º S) enprimavera y verano. Cienc. Tecnol. Mar, 28(2): 63-83.
Silva, N., J. Maturana, J. Sepúlveda & R. Ahumada. 1998.Materia orgánica, C y N, su distribución y estequiometría,en sedimentos superficiales de la región norte de losfiordos y canales australes de Chile (Crucero CIMAR-Fiordo 1). Cienc. Tecnol. Mar, 21: 49-74.
Valdenegro, A. & N. Silva. 2003. Caracterización física y quí-mica de la zona de canales y fiordos australes de Chileentre el estrecho de Magallanes y cabo de Hornos(CIMAR 3 Fiordos). Cienc. Tecnol. Mar, 26(2): 19-60.





























