Embed Size (px)
Citation preview

Elaborado por Miguel Ángel Gamboa- Gaitán (MAGG)
Capítulo 4: La célula vegetal
Introducción
La célula es la base funcional y estructural de la vida pues todo ser vivo se compone, como mínimo, de una célula. Dicho de
otra forma, la célula es la unidad vital mínima, dado que ninguno de sus componentes individuales posee vida ni puede
existir independientemente. La célula vegetal posee numerosas características que permiten diferenciarla perfectamente de
las células de otros seres vivos, como una pared celular compuesta mayoritariamente de celulosa y organelos especializados
en algunos aspectos metabólicos. Este capítulo está destinado a describir las características de la célula vegetal, enfatizando
en las que más la distinguen de las células halladas en otros seres vivos.
Contenido
4.1. La célula vegetal posee numerosos componentes de variada naturaleza y función.
4.2. La pared celular vegetal tiene funciones estructurales y metabólicas.
4.3. La membrana plasmática vegetal tiene las mismas funciones y estructura básica que en otros seres vivos.
4.4. El almacenaje del material genético y el control de la actividad celular son funciones propias del núcleo.
4.5. Los ribosomas son complejos supramoleculares involucrados en la síntesis de proteínas.
4.6. En la célula vegetal hay un complejo sistema de endomembranas.
4.7. El citoesqueleto provee rigidez interna y a la vez permite los movimientos intracelulares.
4.8. Los plastidios son organelos citoplasmáticos distintivos de las plantas.
4.9. La provisión de moléculas energéticas se lleva a cabo en las mitocondrias.
4.10. Los microcuerpos son organelos muy dinámicos en el metabolismo vegetal.
4.11. La vacuola es uno de los organelos más distintivos de la célula vegetal y posee funciones muy diversas.
4.12. La célula vegetal posee estructuras especializadas en el almacenamiento de varios tipos de sustancias.
4.13. Las plantas presentan varios tipos celulares altamente especializados.

MAGG - Botánica General: Introducción al estudio de las plantas
70
4.1. La célula vegetal posee numerosos componentes
de variada naturaleza y función.
Al visualizar una célula vegetal al microscopio de luz,
tres grandes componentes pueden ser identificados
fácilmente: el citoplasma, la pared celular y las sustancias
ergásticas (Figura 4.1). Si bien la célula vegetal es mucho
más compleja que lo indicado por esta descripción,
dividirla en tales componentes es útil para hacer una
introducción a su estudio. El citoplasma forma el grueso
de la masa celular y junto a la membrana plasmática que
lo limita, constituyen el protoplasto o parte viva de la
célula. En el interior del protoplasto se encuentra el
núcleo, el sistema de endomembranas, el citoesqueleto, la
vacuola, los ribosomas y los organelos. Este último
término quiere decir «órgano pequeño» y hace referencia
a los plastidios, mitocondrias y microcuerpos. De estos
componentes citoplasmáticos, sólo los cloroplastos y a
veces el núcleo, son visibles al microscopio óptico sin
usar tinción.
De otra parte, la pared celular es quizá la
característica más notoria y diferenciable de la célula
vegetal, particularmente ausente en células animales. Su
principal componente es la celulosa y, aunque parece una
estructura sin actividad, es un componente celular muy
dinámico, pues no sólo permite la interconexión celular
mediante plasmodesmos sino que posee actividad
enzimática vital para la célula vegetal. El tercer
componente visible de algunas células vegetales está
constituido por las sustancias ergásticas, que resultan de
la actividad metabólica del protoplasto y generalmente
están almacenadas en la vacuola, siendo ejemplo típico
de ellas los cristales de sales como el oxalato de calcio.
Los cristales más comunes y característicos de las células
vegetales se presentan en forma de rafidios y drusas.
En este capítulo se estudia cada uno de los
componentes de la célula vegetal de manera individual,
para entender su composición y funcionamiento dentro
del organismo vegetal. Ahora bien, es necesario aclarar
que dichos componentes no funcionan como entes
independientes ni se encuentran aislados en la naturaleza,
sino que por el contrario están constantemente
interrelacionados y sometidos a una estricta regulación de
conjunto.
4.2. La pared celular vegetal tiene funciones
estructurales y metabólicas.
La pared celular es muy dura debido a sus componentes
químicos y fue considerada durante mucho tiempo una
estructura protectora del protoplasto, a manera de
recipiente rígido e inerte. Sin embargo, la verdad es que a
pesar de su rigidez física, la pared es un componente
celular muy dinámico en cuanto a su funcionalidad
metabólica. Entre sus funciones está el ser una defensa
física contra invasores y contra la deshidratación, pero
también sirve para limitar el crecimiento del protoplasto,
además de ayudar en la absorción, secreción y transporte
de sustancias, e incluso se sabe que posee actividad
digestiva. La pared se encuentra constituida por dos
grandes componentes: una red de fibras de celulosa y una
matriz químicamente compleja de polisacáridos no
celulósicos, glicoproteínas y otras sustancias en la que
están inmersas las mencionadas fibras de celulosa (Figura
4.2).
El componente más abundante de la pared
primaria es la celulosa, que puede constituir hasta el 30%
de su masa seca, la cual se compone de monómeros de
glucosa unidos linealmente mediante enlaces β-1-4. La
molécula lineal de celulosa posee además numerosos
enlaces tipo puente de hidrógeno intra e intermoleculares,
los cuales le dan estabilidad al polímero y permiten que
se asocie con otras moléculas en microfibrillas de 10-25
nm de ancho. Éstas a su vez están asociadas entre sí
formando unas hebras llamadas macrofibrillas, de 0.5 µm

MAGG - Botánica General: Introducción al estudio de las plantas
71
de ancho, las cuales se entrelazan formando una
estructura similar a la de un lazo. La resistencia de la
celulosa es comparable a una estructura de acero del
mismo tamaño, lo que explica las propiedades de la pared
celular, al menos en lo referente a su rigidez. La celulosa
es insoluble en agua y muy resistente, y sólo hay unos
pocos microorganismos capaces de degradarla.
La matriz de la pared posee polisacáridos
pécticos, como los galactanos, los galacturonanos, los
arabinanos y los arabinogalactanos, además de
polisacáridos hemicelulósicos, como el xiloglucano, los
xilanos, la calosa y los mananos. Además, dicha matriz
posee proteínas estructurales como las extensinas, que
son muy ricas en el extraño aminoácido hidroxiprolina,
aunque también se sabe de la presencia de proteínas
enzimáticas como la peroxidasa y la glicosidasa, así
como endoglicanasas, oxidorreductasas, transglicosidasas
y esterasas.
Otro componente notable, sobre todo en paredes
secundarias, es la lignina, que está formada por polímeros
cuya composición varía entre especies. Tales polímeros
están constituidos por asociaciones de derivados de los
alcoholes p-cumaril, sinapil y coniferil. La lignina
aumenta la rigidez de la pared, así como la resistencia
física y química y puede constituir entre el 20 y 30 % de
la masa seca en fibras y vasos. También tiene cutina,
suberina y ceras, sustancias de naturaleza hidrófoba que
aumentan la impermeabilidad y evitan la pérdida de agua
y solutos. Por último, varios compuestos inorgánicos son
también abundantes en las paredes celulares, como
cationes (ej: Ca++) y óxidos de silicio.
La composición de la pared varía entre especies
y aún entre individuos conespecíficos dependiendo de su
estado de desarrollo. Sus componentes son sintetizados a
partir de sustancias precursoras en el citoplasma y son
enviados en vesículas a través del dictiosoma (complejo o
aparato de Golgi) y del retículo endoplásmico (RE), las
cuales se fusionan con la membrana plasmática y liberan
su contenido. Además de las variaciones en composición,
las paredes celulares poseen diferencias en su grosor
entre los diferentes tipos celulares (Figuras 4.3 y 4.5C), e
incluso a veces pueden estar completamente
interrumpidas en algunas caras de células vecinas. Un
ejemplo de la variación en el ancho de la pared celular
son las punteaduras del xilema, donde el grosor total está
dado sólo por la pared primaria, ya que la pared
secundaria está interrumpida. El colénquima es un tejido
que también presenta variaciones importantes en el ancho
de su pared celular, ya que las capas de depósito en ella
son más gruesas en las esquinas (cerca de los puntos de
contacto de varias células), que en la parte media (donde
sólo dos células se ponen en contacto).
Ahora bien, la pared celular también posee unos
poros de hasta unos 60 nm de diámetro que conectan las
células adyacentes y por los cuales las membranas
plasmáticas de dichas células se continúan. Estos canales
se denominan plasmodesmos y son variables en tamaño y
número entre los diferentes tipos de células. Algunos
poseen una estructura central denominada desmotúbulo,
el cual incluye proteínas en su composición y se puede
considerar una prolongación del RE. A través de los
plasmodesmos hay una activa y regulada comunicación
intercelular que permite la movilización de iones y
moléculas tan grandes como los ácidos nucleicos. Dado
que casi todas las células de la planta están conectadas a
través de estos canales, es viable pensar que el organismo
vegetal es realmente un sincicio, es decir, un solo
citoplasma gigantesco multinucleado. Cuando la planta
es herida, depósitos de calosa taponan los plasmodesmos
para evitar la pérdida de citoplasma y la entrada o
movilización de patógenos como los virus.

MAGG - Botánica General: Introducción al estudio de las plantas
72
4.3. La membrana plasmática vegetal tiene las mismas
funciones y estructura básica que en otros seres vivos.
La bicapa lipídica con proteínas asociadas que conforman
un mosaico fluido, es el modelo más aceptado para la
composición de las membranas celulares, incluyendo las
de las plantas (Figura 4.4). En este modelo la bicapa está
compuesta de fosfolípidos anfipáticos que exponen sus
extremos hidrofílicos al agua, mientras que sus extremos
hidrófobos están inmersos en el interior de la bicapa,
alejados de los ambientes acuosos. Inmersas en la bicapa
se hallan proteínas cuyos extremos pueden sobresalir
hacia el exterior de la célula o hacia el citoplasma, según
su función, a las cuales se denomina proteínas
intrínsecas. Anexas a la membrana plasmática (que
también es llamada plasmalema), hay proteínas
periféricas o extrínsecas, que poseen propiedades
enzimáticas pero que no forman parte integral de la
membrana.
La composición de las membranas vegetales
varía tanto en la calidad como en la cantidad de los
lípidos y proteínas entre sus dos capas, así como entre
células, tejidos y órganos, según el estado de desarrollo y
la función de la célula. En general, las proteínas
constituyen el 60% y los lípidos el 40% de la
composición del plasmalema y las variaciones en estos
porcentajes, junto a la composición química de lípidos y
proteínas, alteran el estado de fluidez de las membranas.
Algunas proteínas intrínsecas pueden desplazarse
lateralmente pero otras forman asociaciones llamadas
dominios membranales, los cuales constituyen sectores
discretos que son diferentes de sus alrededores (parches),
lo que le confiere a las membranas la propiedad de ser
heterogéneas. Algunos lípidos y proteínas poseen
cadenas cortas de glúcidos adheridas a sus extremos
externos, lo cual le confiere a las membranas importantes
propiedades de reconocimiento intercelular. Estas
moléculas se denominan glicolípidos y glicoproteínas,
respectivamente.
La membrana plasmática es el límite externo del
protoplasto (Figura 4.5C) y posee la capacidad de
seleccionar las sustancias que pasan a través de ella, por
lo que se dice que es selectivamente permeable. Las dos
caras de la membrana son diferentes entre sí,
particularmente en cuanto al tipo de proteínas que la
componen, ya que cada lado encara dos ambientes
totalmente diferentes: el lado interno está en contacto con
el citoplasma mientras que el externo enfrenta al
ambiente extracelular. Las proteínas de membrana
pueden tener variadas funciones, como la de constituir
canales o poros para el transporte facilitado de iones y
otras sustancias cargadas, o también pueden ser bombas
moleculares que ligan moléculas en un lado de la
membrana y las mueven hacia el otro lado usando
energía (ATP). También pueden formar receptores que
aceptan moléculas como hormonas y activan señales
intracelulares que desencadenan respuestas fisiológicas.
La membrana también cambia constantemente su forma,
tamaño y función debido a la continua incorporación y
remoción de sus componentes, lo cual se lleva a cabo por
la fusión (exocitosis) e invaginadión (endocitosis) de
vesículas. Tales vesículas son porciones pequeñas de la
membrana que adoptan una forma esférica. Estos
cambios de la membrana plasmática se dan de acuerdo
con las necesidades metabólicas de la célula.
4.4. El almacenaje del material genético y el control
de la actividad celular son funciones propias del
núcleo.
Toda la actividad metabólica de la célula está regulada,
en última instancia, por la síntesis de proteínas, la cual es
controlada por el núcleo. Este componente celular está
limitado por una membrana doble (denominada más
apropiadamente envoltura nuclear), la cual posee poros

MAGG - Botánica General: Introducción al estudio de las plantas
73
de 30-100 nm de diámetro y a menudo se continúa con el
retículo endoplásmico (RE) (Figura 4.5A). Entre las dos
capas de la envoltura nuclear hay un espacio de entre 20-
40 nm y se ha visto que hay proteínas asociadas a los
poros nucleares, que pueden estar involucradas en la
regulación del tránsito de sustancias a través de ellos. En
el núcleo se almacena y protege el material genético en
forma de ácido desoxirribonucleico (ADN), el cual está
asociado con proteínas básicas denominadas histonas,
constituyendo así la cromatina.
La cromatina es la principal sustancia del núcleo
y se encuentra inmersa en una matriz denominada
nucleoplasma. Como particularidades tiene que se tiñe
con colorantes básicos y que absorbe eficientemente la
luz ultravioleta a unos 260 nm. Cuando la cromatina se
condensa da origen a los cromosomas, estructuras en las
que el ADN experimenta su máximo grado de
compactación, en gran parte debido a las histonas con las
que se empaca. Cuando el material genético está
empacado en forma de cromosomas visibles en
cariotipos, no es posible que el núcleo efectúe la
replicación del ADN debido a su alto grado de
compactación. El número de cromosomas depende de la
especie y es muy variable en el reino vegetal, habiendo
valores tan disímiles como 4 en Haplopappus y 1250 en
Ophioglossum. Además del ADN y las proteínas
histonas, el nucleoplasma también contiene enzimas que
mantienen y reparan el ADN, varias clases de ácido
ribonucleico (ARN), agua y sustancias necesarias para el
metabolismo nuclear. Dentro del núcleo se encuentra
también el nucléolo, lugar asociado con la producción del
ARN y los ribosomas.
En general las células contienen un núcleo, pero
en las plantas hay casos de células multinucleadas que se
originan por cariocinesis que no van acompañadas de
citocinesis, como en el desarrollo del endospermo de la
semilla. También se ha documentado la existencia de
células que, por el contrario, carecen de núcleo, ya que lo
pierden durante su proceso de diferenciación, como
sucede en los elementos cribosos del floema. Estas
células dependen para sobrevivir de células especiales,
denominadas acompañantes, las cuales controlan el
metabolismo del elemento criboso de tal forma que siga
siendo funcional.
4.5. Los ribosomas son complejos supramoleculares
involucrados en la síntesis de proteínas.
Más grandes que las macromoléculas pero inferiores en
tamaño a los organelos, estas estructuras intracelulares
son fundamentales para la producción de proteínas.
Pueden estar libres en el citoplasma o asociados
abundantemente al retículo endoplásmico, donde
sintetizan las proteínas celulares generalmente asociadas
a membranas. Están compuestos de proteínas y ARN
ribosomal (ARNr), el cual se compone de subunidades
ensambladas en el núcleo, concretamente en el nucléolo,
para luego ser exportadas hacia el citoplasma. Una vez
allí, los ribosomas se asocian al ARN mensajero
(ARNm), constituyendo una estructura denominada
polisoma que es fundamental en el proceso de síntesis
proteica. Es común que sobre una sola molécula de
ARNm se ubiquen varios ribosomas, por lo que varias
copias del polipéptido son sintetizadas simultáneamente.
Los ribosomas de los eucariotas, incluidos los de
la plantas, son bastante similares entre sí. En general, su
coeficiente de sedimentación es 80S con más de 4,000
kDa de masa molecular. Poseen dos subunidades,
denominadas mayor y menor, las cuales son 60S y 40S,
respectivamente. La mayor posee tres tipos de ARNr, uno
28S, uno 5.8S y un tercero 5S, mientras que la subunidad
menor posee un ARNr 18S. Es interesante tener presente
que en el cloroplasto hay también ribosomas, los cuales
son 70S y se pueden afectar por el cloranfenicol, por lo
que son más parecidos a los ribosomas de los procariotas
que a los de los eucariotas. Esto es de esperarse si la

MAGG - Botánica General: Introducción al estudio de las plantas
74
hipótesis de la endosimbiosis, que establece que los
cloroplastos eran microorganismos de vida libre que se
asociaron con otra célula, es correcta.
4.6. En la célula vegetal hay un complejo sistema de
endomembranas.
La envoltura nuclear se continúa con un sistema aplanado
o tubular de membranas denominado retículo
endoplásmico (RE), el cual está muy involucrado en el
metabolismo de las proteínas, ya que los ribosomas
asociados a él las sintetizan y luego las pasan a su lumen.
Si la proteína es de almacenamiento puede permanecer en
el RE como sucede en las células de semillas, pero si es
de secreción, como las proteínas enzimáticas o adhesivas,
se envuelve en una vesícula del RE y pasa al dictiosoma
(Figura 4.6), donde experimenta modificaciones. El RE
sin ribosomas se denomina retículo endoplásmico liso
(REL), mientras que el rugoso (RER), es el que posee
ribosomas sobre su superficie (Figura 4.8). El REL se
encarga de la síntesis de lípidos y del ensamblaje de las
membranas, ya que las vesículas que se liberan de él se
fusionarán con otras membranas y entrarán a formar parte
integral de ellas. El REL es abundante en células
vegetales que producen muchos ácidos grasos, como las
células de la epidermis que producen cutina y en células
que producen aceites y fragancias.
Los dictiosomas son vesículas membranosas
aplanadas que se agrupan formando apilamientos
denominados en conjunto el aparato o complejo de Golgi.
Es más común en animales que en plantas, pero en los
pelos radicales se observan complejos de Golgi muy
grandes. Esta estructura tiene una cara donde las
vesículas provenientes del RE se fusionan, denominada
cara de formación o Cis, así como otra cara por donde
dichas vesículas se liberan, llamada cara de maduración o
Trans. Los contenidos de dichas vesículas han sido
modificados durante su paso por los dictiosomas, que
comúnmente involucran la adición de azúcares a las
proteínas de dichas vesículas, generando glicoproteínas
que por lo general se fusionan con la membrana
plasmática. Al parecer algunos polisacáridos destinados a
la pared celular son también procesados en los
dictiosomas.
Cuando dos núcleos se han formado luego de la
mitosis, los dictiosomas son dirigidos por los
microtúbulos hacia la región central de la célula y
empiezan a fusionarse formando una estructura discoidea
llamada placa celular (Figura 4.7). Ésta se ubica en
posición equidistante de los núcleos hijos y se dispone de
forma perpendicular al eje en que se ubicó el huso
acromático en la célula madre. Dichos dictiosomas
poseen precursores de la nueva pared celular en su
interior y su fusión continúa hasta que alcanzan las
paredes laterales para completar la separación de las dos
células hijas.
4.7. El citoesqueleto provee rigidez interna y a la vez
permite los movimientos intracelulares.
Las células vegetales poseen un sistema filamentoso
intracelular de soporte denominado citoesqueleto, el cual
es muy importante en el crecimiento celular. El
citoesqueleto puede actuar reforzando ciertas áreas de la
célula mientras que otras permanecen más débiles para
permitir la expansión celular. Los movimientos internos
de los organelos y vesículas requieren del ensamblaje y
rompimiento del citoesqueleto, por lo que es un
componente muy importante para la dinámica celular. La
ciclosis, por ejemplo, que es el movimiento de organelos
y sustancia basal del citoplasma de manera circular,
depende también del citoesqueleto. Tanto el núcleo como
otros organelos pueden ser movilizados por este sistema
de microtúbulos, como también sucede con los
cromosomas durante la mitosis.

MAGG - Botánica General: Introducción al estudio de las plantas
75
El citoesqueleto está conformado básicamente
por microtúbulos de tubulina y filamentos de actina. Los
filamentos de actina, o microfilamentos, están
compuestos por la proteína globular actina y sólo
alcanzan 6 nm de diámetro. Los microtúbulos están
formados de tubulina, un dímero que está compuesto de
alfa y beta tubulinas, las cuales se polimerizan formando
un filamento largo, de unos 25 nm de diámetro. Cuando
el microtúbulo no se necesita, se puede despolimerizar y
sus componentes individuales se dispersan por el
citoplasma hasta que sean reutilizados. La colchicina,
alcaloide aislado de plantas del género Colchicum,
interrumpe el ensamblaje de los microtúbulos porque se
une a la tubulina e inhibe su polimerización, siendo una
droga de gran utilidad para el estudio de la dinámica
celular interna y del citoesqueleto.
Los microtúbulos también son componentes
fundamentales de los flagelos, estructuras involucradas
en el desplazamiento celular y que en los vegetales están
presentes sólo en las células espermáticas de algunas
plantas sin flores. Los flagelos están constituidos por
microtúbulos con el típico arreglo de 9+2 (Figura 4.9),
donde nueve pares de ellos se arreglan alrededor de dos
microtúbulos centrales. Cada par de microtúbulos
externos posee dos estructuras compuestas de una
proteína llamada dineína, la cual usa ATP para producir
energía cinética. Siempre hay asociado a la base del
flagelo una estructura conocida como cuerpo basal, que
se ve al microscopio igual a los centriolos de las células
animales, pero cuya función no ha sido aun clarificada.
4.8. Los plastidios son organelos citoplasmáticos
distintivos de las plantas.
Los plastidios tienen un sistema de membranas
limitantes, una interna y una externa, con un fluido
interno denominado estroma. Hay varias clases de
plastidios que se ocupan de importantes aspectos
metabólicos de la célula vegetal, como la síntesis de
NADPH, purinas, pirimidinas, aminoácidos y ácidos
grasos, además del almacenamiento de sustancias como
el almidón. Ellos son una de las características más
exclusivas de las plantas dado que están ausentes en
células animales, hongos y la mayoría de protistas, razón
por la cual algunos autores consideran que las plantas
debieran definirse como el conjunto de seres vivos que
poseen plastidios. Los tipos de plastidios existentes son:
cloroplastos (Figura 4.5D), amiloplastos, elaioplastos,
cromoplastos y leucoplastos. Los precursores de todos
ellos se denominan proplastidios y están presentes en
células en proceso de diferenciación. Los plastidios
pueden crecer, alterando la composición de sus
membranas y se reproducen por procesos similares a una
fisión binaria.
Los cloroplastos son los plastidios más
conspicuos (Figura 4.10) y están encargados de la
fotosíntesis. Poseen ADN propio, una doble membrana y
un sistema de endomembranas aplanadas, discoideas,
llamadas tilacoides, las cuales se agrupan formando
apilamientos denominados granas. Además, poseen
ribosomas parecidos a los de los microorganismos
procariotas, lo cual constituye, junto a las características
recientemente mencionadas, evidencia de su origen como
consecuencia de un evento endosimbiótico ancestral.
Es posible que los cloroplastos almacenen gotas
lipídicas y almidón, siendo éste el producto directo de la
fotosíntesis mediada por clorofilas a, b y pigmentos
accesorios del tipo carotenoide. Al interior de los granas
se acumulan iones H+ contra su gradiente debido al flujo
de electrones por la membrana tilacoidal, la cual posee
abundantes proteínas transmembranales que se ubican
diferencialmente según su función. La síntesis del
almidón se lleva a cabo por enzimas que se encuentran
solubles en el estroma del cloroplasto y es posible que se
acumule dentro de él si la tasa de producción es mayor
que la de consumo o exportación. Cuando una planta

MAGG - Botánica General: Introducción al estudio de las plantas
76
crece en oscuridad, los proplastidios foliares desarrollan
etioplastos, plastidios que almacenan un precursor de la
clorofila que es amarillo. Al ser expuestos a luz, se
induce su transformación en cloroplastos.
Los amiloplastos (Figura 4.11), almacenan los
productos de la fotosíntesis en forma de almidón,
preferencialmente en los órganos que no hacen
fotosíntesis, como las raíces. En su interior se depositan
grandes granos de almidón que prácticamente llenan todo
el estroma. Los leucoplastos carecen de pigmentos y su
función es sintetizar lípidos que exportan a otras partes
de la célula. Pueden diferenciarse en cloroplastos tras la
exposición a la luz. Otra aparente función de estos
plastidios es almacenar hierro, lo cual llevan a cabo
debido a la proteína fitoferritina, que también puede estar
presente en cloroplastos y amiloplastos. Los
cromoplastos guardan pigmentos diferentes de las
clorofilas y son particularmente abundantes en órganos
reproductores como flores y frutos. Pueden originarse por
transformación de cloroplastos y los pigmentos que
almacenan son de tipo lipídico, carecen de granas pero
poseen un sistema membranal ondulado que puede
contener los pigmentos, los cuales también pueden estar
en gotas denominadas plastoglóbulos.
4.9. La provisión de moléculas energéticas se lleva a
cabo en las mitocondrias.
Las mitocondrias (Figuras 4.5B y 4.12C), son unos
organelos muy plásticos y móviles, cuya forma y tamaño
varían considerablemente entre las células vegetales
según el estado funcional de ellas. Son capaces de
fusionarse y dividirse de nuevo, poseen su propio ADN y
se caracterizan por estar limitadas por dos membranas,
siendo la interna muy típica por poseer unos pliegues
internos denominados crestas, que aumentan su
superficie. Estos organelos se originaron aparentemente
por un proceso de endosimbiosis entre una proteobacteria
y una bacteria anaeróbica del tipo arqueobacteria, ambas
de vida libre que poseían su propio ADN y ribosomas. En
las mitocondrias actuales la mayoría de sus proteínas son
codificadas en el núcleo de la célula vegetal y no por su
propio ADN. En el interior de la mitocondria existe una
matriz mitocondrial, que es el contenido interno análogo
al estroma de los cloroplastos. En ellas se lleva a cabo
importantes procesos metabólicos, como la respiración
con la consecuente oxidación de los alimentos para
producir el ATP. Esta oxidación es mucho más eficiente
que la glicólisis anaerobia, por lo que la aparición de las
mitocondrias supuso un gran avance en la historia natural
de los eucariotas. A menudo ellas están concentradas en
zonas de altos requerimientos energéticos.
4.10. Los microcuerpos son organelos muy dinámicos
en el metabolismo vegetal.
Los microcuerpos son organelos esféricos de 0.5-1.5 µm
de diámetro, están limitados por una sola membrana y
comúnmente se les encuentra asociados al RE y a
cuerpos lipídicos. Carecen de material genético por lo
que todas las proteínas que poseen son importadas, lo
cual se logra porque ellas tienen una secuencia señal que
les permite la entrada a los microcuerpos. Están
involucrados en importantes aspectos del metabolismo
celular, entre los que se destaca la ruptura del peróxido
de hidrógeno (H2O2), que es una sustancia tóxica
producida por la oxidación de sustratos orgánicos por la
enzima catalasa. Dicha ruptura sucede en un tipo especial
de microcuerpo denominado peroxisoma (Figura 4.12B),
el cual está especializado en sostener recciones químicas
específicas, como la mencionada del peróxido de
hidrógeno o las que cataliza la enzima urato oxidasa. A
veces hay tantas copias de estas proteínas en su interior,
que es común encontrar estructuras sólidas, producto de
la cristalización de ellas. Los peroxisomas también están
asociados al metabolismo del ácido glicólico y a la
fotorrespiración, e incluso parece que pueden usar el
H2O2 para oxidar sustancias tóxicas como etanol y
nitritos. Otro grupo de microcuerpos importantes en las

MAGG - Botánica General: Introducción al estudio de las plantas
77
plantas son los glioxisomas, que convierten lípidos en
glúcidos debido a la producción de acetil-coA a partir de
dichos lípidos. Los glioxisomas están asociados al
proceso de germinación de las semillas, ya que extraen la
abundante energía almacenada en los lípidos de los
tejidos de reserva, para que la plántula la use en su
desarrollo inicial.
4.11. La vacuola es uno de los organelos más
distintivos de la célula vegetal y posee funciones muy
diversas.
La vacuola es un organelo típico de las células vegetales
y se entiende como una región del citoplasma rodeada
por una membrana simple llamada tonoplasto (Figura
4.5E). Su contenido es llamado savia celular, el cual
consta de agua, azúcares, sales, proteínas, iones, cristales
de oxalato de Ca, entre otras cosas. Cuando la célula es
inmadura hay muchas vacuolas de pequeño tamaño, las
cuales se fusionan para formar una sola y grande vacuola
central en la célula madura, que puede ocupar hasta el
90% del volumen celular. Está involucrada en el
crecimiento de la célula, ya que por el aumento de su
volumen presiona desde adentro (presión de turgencia),
para aumentar el volumen celular. También almacena
diversas sustancias como pigmentos (antocianinas), e
interviene en el rompimiento y reciclaje de
macromoléculas y de componentes celulares viejos.
Algunas sustancias tóxicas de desecho son
permanentemente almacenadas allí, siendo liberadas sólo
cuando la célula se rompe, en lo que se supone es un
sistema de defensa contra los herbívoros. La vacuola
también sirve como regulador de los niveles
protoplásmicos de calcio, un segundo mensajero muy
importante al cual puede almacenar asociándolo a otras
sustancias.
4.12. La célula vegetal posee estructuras
especializadas en el almacenamiento de varios tipos de
sustancias.
Las células vegetales tienen la capacidad de almacenar
una gran variedad de compuestos ya que los vegetales
carecen de sistemas de excreción. Esto quiere decir que
hasta las sustancias de desecho se pueden guardar de
manera indefinida, la mayoría de las veces en la vacuola
central. Sin embargo, también hay estructuras
especializadas en el almacenamiento de sustancias que
serán útiles para la planta, tal como sucede en los
amiloplastos, que son plastidios encargados de almacenar
el almidón que resulta de la fotosíntesis. Otra estructura
especializada en almacenamiento son las gotas lipídicas,
que almacenan sustancias grasas y por su pequeño
tamaño le dan una apariencia granular a la célula.
También han sido llamadas esferosomas y es común
hallarlas en semillas, por ejemplo en la capa de aleurona
de las monocotiledóneas, donde constituyen un
reservorio energético muy importante, de gran utilidad en
el momento de la germinación y durante los primeros
estadios de la plántula.
4.13. Las plantas presentan varios tipos celulares
altamente especializados.
Las células vegetales adoptan numerosas variantes de
forma y función dentro de la planta según el tejido en el
que se encuentren. Se sabe de la existencia de al menos
diez tipos celulares en las plantas (Figura 4.13), los
cuales se describen brevemente a continuación y están
bien ilustrados en el apartado dedicado a los tejidos
(Capítulo 5). El tipo más básico de célula vegetal es el
parenquimático, el cual se caracteriza por poseer una
pared celular delgada, generalmente primaria, aunque
puede haber algunas con pared secundaria; sus células
son poliédricas y adoptan variadas formas. Están
presentes en todo el cuerpo vegetal, razón por la cual se
creyó que sólo servían para constituir la masa corporal, es

MAGG - Botánica General: Introducción al estudio de las plantas
78
decir, para servir de relleno entre los tejidos funcionales.
Sin embargo, se descubrió que se encargan de muy
diversas funciones, como protección, almacenamiento,
conducción, fotosíntesis, reparación y regeneración, e
incluso a menudo acompañan a otras células para
complementar sus funciones según el tipo de tejido. Un
tipo especial de célula de parénquima está conformado
por células fotosintetizadoras, al cual se denomina
parénquima asimilador o clorénquima. Este tejido está
generalmente en las hojas y puede tener dos
presentaciones, el parénquima de células esponjosas y el
parénquima en empalizada.
Otro tipo celular importante es el de las células
meristemáticas, que se caracteriza por poseer pocos
espacios intercelulares y la capacidad de
autorregenerarse. Estas células sólo poseen pared celular
primaria y están restringidas a los meristemos, lugares
del cuerpo vegetal por donde éste crece, generalmente
situados en los extremos de las ramas y las raíces.
También existe en las plantas un tipo de células
especializadas en cubrir el cuerpo vegetal, constituyendo
la <<piel>> de las plantas, por lo que están
especializadas en protección y reciben el nombre de
células epidermales. Entre ellas hay varios subtipos como
las células guardas de los estomas y las células de los
tricomas, los cuales cumplen funciones como la
protección contra herbívoros y el mantenimiento de la
temperatura. Al igual que las meristemáticas, las
epidermales poseen muy pocos espacios intercelulares.
Existe otro tipo importante de células vegetales
que son las que proveen sostén. Entre ellas están las
células de colénquima, que son generalmente alargadas,
con pared celular primaria desigualmente engrosada y no
presentan lignificación. Generalmente se ubican hacia la
periferia de los órganos jóvenes, en cilindros o en parches
o bordes que sobresalen en las venas o en el tallo. El otro
tipo importante de células de sostén son las
esclerenquimáticas, las cuales son de dos tipos: fibras y
esclereidas. Las fibras son muy largas, con pared
secundaria lignificada y a menudo están muertas en su
madurez funcional. Casi siempre están asociadas a tejidos
vasculares, en la corteza de los tallos y en hojas de
monocotiledóneas. Las esclereidas son más variables en
forma, generalmente mucho más cortas que las fibras,
poseen pared secundaria lignificada y algunas mueren en
la madurez. Proveen soporte en tejidos como hojas de
plantas flotantes y en la epidermis de frutos y otros
órganos. Hay varios subtipos de ellas de acuerdo con su
estructura y función particulares, entre las que se destaca
a las artroesclereidas y braquiesclereidas.
Las células conductoras constituyen un tipo
especial muy importante para las plantas. Las traqueidas
son delgadas y alargadas, con punteaduras y pared celular
secundaria lignificada, algunos autores las consideran
como un tipo especial de fibras esclerenquimáticas. Están
muertas cuando son funcionales y su principal función es
el transporte de agua y solutos por el xilema, siendo el
principal sistema conductor de las gimnospermas y las
plantas vasculares inferiores, aunque también pueden
estar presentes en las angiospermas. Los elementos de
vaso son también células conductoras pero más cortas y
anchas que las traqueidas, con paredes celulares
secundarias lignificadas que presentan punteaduras y
perforaciones, están muertas en la madurez y forman
parte del xilema de las angiospermas. Son mucho más
eficintes que las traqueidas para transportar el agua.
Continuando con las células vasculares, es
necesario pasar al floema. Allí se encuentra la célula
cribosa, que es ancha y un poco alargada y forma parte
del floema de las gimnospermas y de las plantas
vasculares inferiores. Su pared celular es primaria, con
áreas cribosas a través de las cuales se comunica con sus
vecinas y carece de núcleo en la madurez. Su función es
la de conducir los fotosintatos desde los órganos que los

MAGG - Botánica General: Introducción al estudio de las plantas
79
producen (fuentes), hasta los órganos que los consumen o
almacenan (sumideros). Junto a ella siempre hay una
célula albuminosa, la cual es alargada y nutre mediante
numerosas conexiones a la célula cribosa, dado que ésta
carece de núcleo. Su origen está en una célula madre
diferente a la de la célula cribosa. El otro tipo celular
vascular altamente especializado en el floema es el
elemento de tubo criboso. Se trata del principal
componente conductor de fotosintatos en las
angiospermas, el cual, además de las áreas cribosas,
presenta placas cribosas en las paredes terminales, por las
cuales se conecta con sus vecinas constituyendo
alargadas estructuras tubulares llamadas tubos cribosos.
En la madurez el elemento del tubo criboso está vivo
pero carece de núcleo y otros organelos, razón por la cual
siempre está asociado a una célula acompañante que
dirige su metabolismo. Ambas células se derivan de una
misma célula madre y sus paredes celulares primarias
están atravesadas por numerosos plasmodesmos que las
conectan ampliamente, lo cual es apenas lógico dada su
estrecha dependencia fisiológica.
BIBLIOGRAFÍA
Alberts B, Bray D, Lewis J, Raff M, Roberts K &
Watson JD. 1994. Molecular biology of the cell, 3 ed.
Garland Publishing, Inc., Nueva York, E.U.A.
Kragler F, Lucas WJ & Monzer J. 1998.
Plasmodesmata: dynamics, domains and patterning.
Annals of Botany 81: 1-10.
Márquez O, Waliszewski KN, Oliart RM & Pardio
VT. 2008. Purification and characterization of cell wall-
bound peroxidase from vanilla bean. LWT – Food
Science and Technology 41: 1372-1379.
Paniagua R, Nistal M, Sesma P, Álvarez-Uría M,
Fraile B, Anadón R & Sáez FJ. 2007. Citología e
histología vegetal y animal. Volumen 2: Histología
vegetal y animal, 4 ed. McGraw-Hill Interamericana,
Madrid, España.
Raven PH, Evert RF & Curtis H. 1981. Biology of
plants, 3 ed. Worth Publishers, Inc., Nueva York, E.U.A.
Taiz L & Zeiger E. 2002. Plant physiology, 3 ed.
Sinauer Associates, Inc., Sunderland, E.U.A.
Zarra I & Revilla G. 1993. Pared celular. Estructura y
función, págs. 1-24 en: Azcón-Bieto J & Talón M (Eds.).
Fisiología y bioquímica vegetal. Interamericana
McGraw-Hill, Madrid, España.
Figura de la portada del capítulo
80
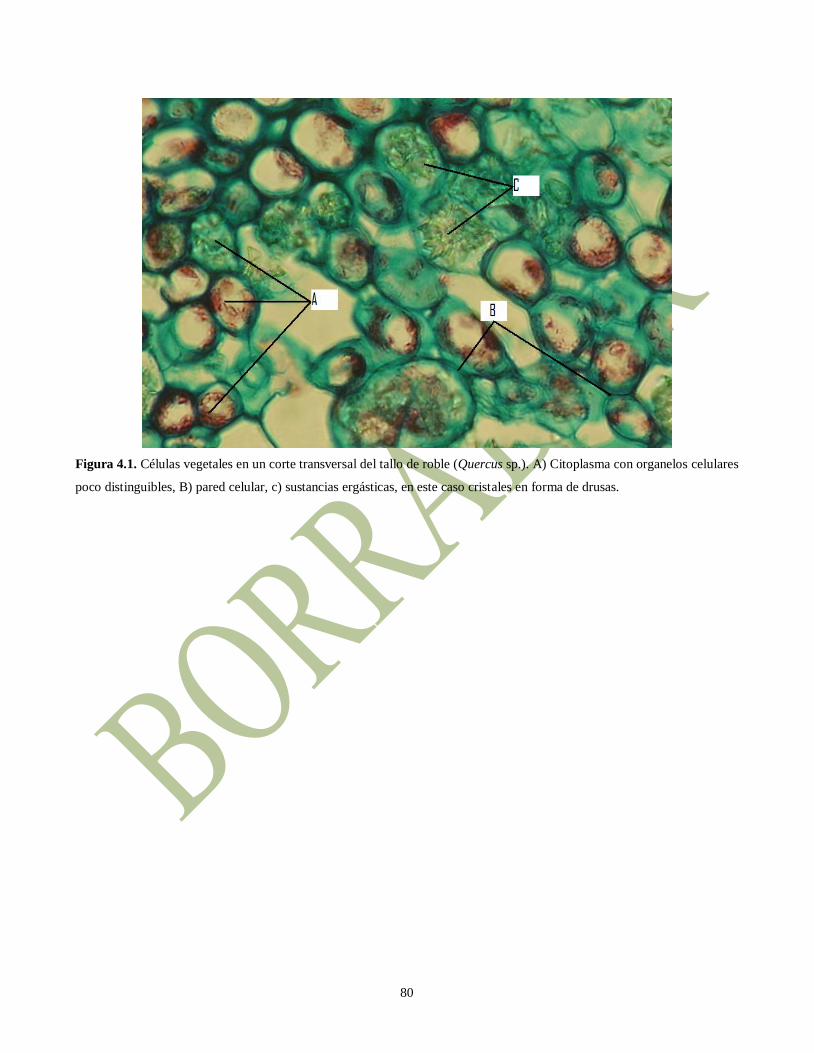
Figura 4.1. Células vegetales en un corte transversal del tallo de roble (Quercus sp.). A) Citoplasma con organelos celulares
poco distinguibles, B) pared celular, c) sustancias ergásticas, en este caso cristales en forma de drusas.
81
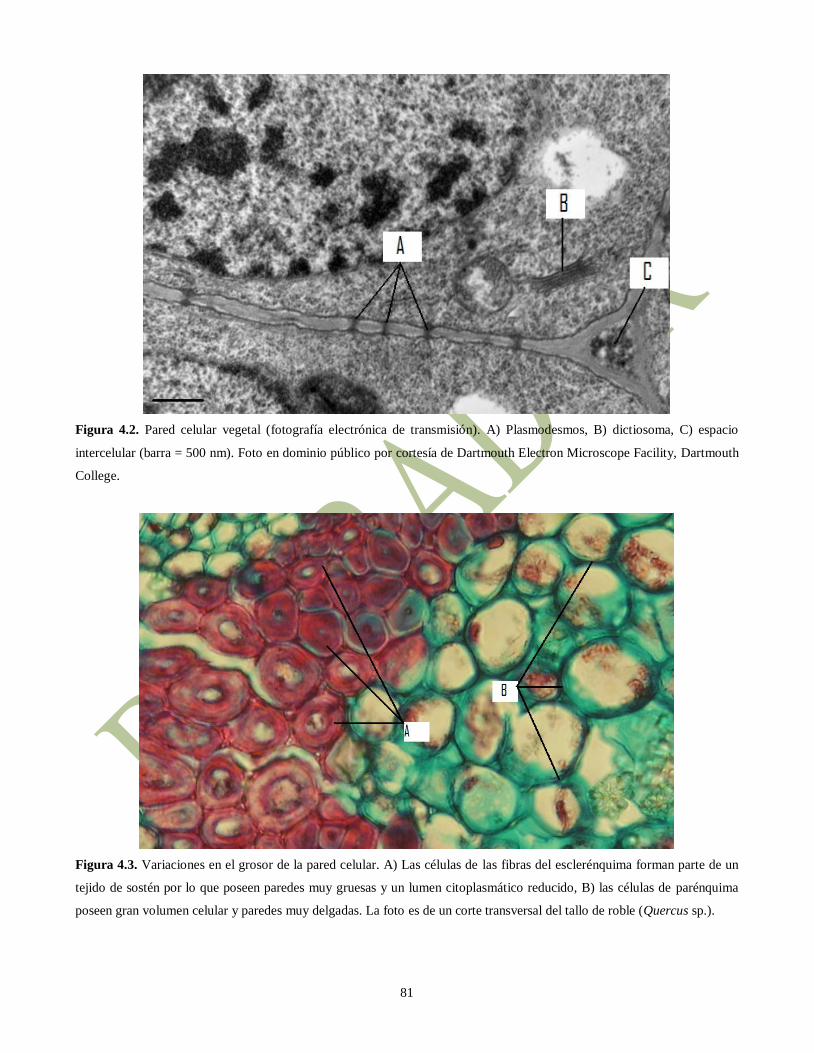
Figura 4.2. Pared celular vegetal (fotografía electrónica de transmisión). A) Plasmodesmos, B) dictiosoma, C) espacio
intercelular (barra = 500 nm). Foto en dominio público por cortesía de Dartmouth Electron Microscope Facility, Dartmouth
College.
Figura 4.3. Variaciones en el grosor de la pared celular. A) Las células de las fibras del esclerénquima forman parte de un
tejido de sostén por lo que poseen paredes muy gruesas y un lumen citoplasmático reducido, B) las células de parénquima
poseen gran volumen celular y paredes muy delgadas. La foto es de un corte transversal del tallo de roble (Quercus sp.).
82
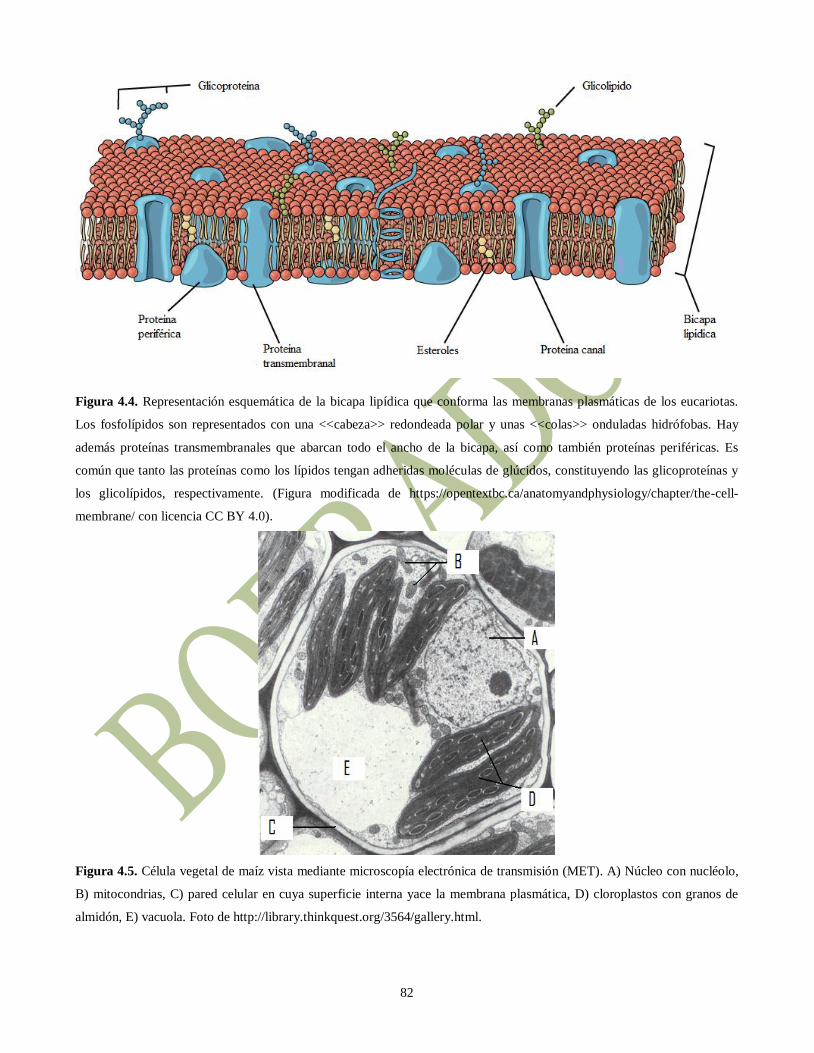
Figura 4.4. Representación esquemática de la bicapa lipídica que conforma las membranas plasmáticas de los eucariotas.
Los fosfolípidos son representados con una <<cabeza>> redondeada polar y unas <<colas>> onduladas hidrófobas. Hay
además proteínas transmembranales que abarcan todo el ancho de la bicapa, así como también proteínas periféricas. Es
común que tanto las proteínas como los lípidos tengan adheridas moléculas de glúcidos, constituyendo las glicoproteínas y
los glicolípidos, respectivamente. (Figura modificada de https://opentextbc.ca/anatomyandphysiology/chapter/the-cell-
membrane/ con licencia CC BY 4.0).
Figura 4.5. Célula vegetal de maíz vista mediante microscopía electrónica de transmisión (MET). A) Núcleo con nucléolo,
B) mitocondrias, C) pared celular en cuya superficie interna yace la membrana plasmática, D) cloroplastos con granos de
almidón, E) vacuola. Foto de http://library.thinkquest.org/3564/gallery.html.

83
Figura 4.6. Detalle de una célula vegetal fotografiada con microscopía electrónica de transmisión. A) Plasmodesmo en la
pared celular, B) dictiosomas apilados (barra = 50 nm). Foto en dominio público por cortesía de Dartmouth Electron
Microscope Facility, Dartmouth College.
Figura 4.7. Formación de la placa celular en citocinesis. A) Núcleo de célula hija luego de mitosis, B) placa celular
formada por la fusión de vesículas (barra = 500 nm). Foto en dominio público por cortesía de Dartmouth Electron
Microscope Facility, Dartmouth College.
84
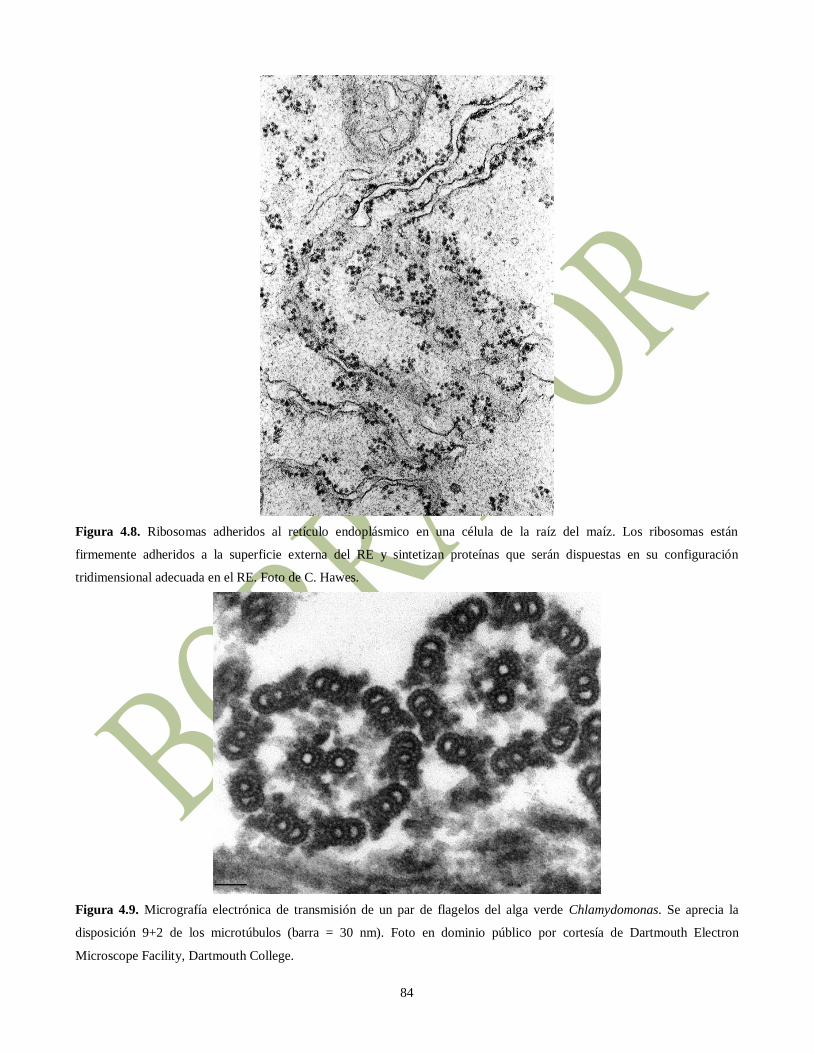
Figura 4.8. Ribosomas adheridos al retículo endoplásmico en una célula de la raíz del maíz. Los ribosomas están
firmemente adheridos a la superficie externa del RE y sintetizan proteínas que serán dispuestas en su configuración
tridimensional adecuada en el RE. Foto de C. Hawes.
Figura 4.9. Micrografía electrónica de transmisión de un par de flagelos del alga verde Chlamydomonas. Se aprecia la
disposición 9+2 de los microtúbulos (barra = 30 nm). Foto en dominio público por cortesía de Dartmouth Electron
Microscope Facility, Dartmouth College.
85
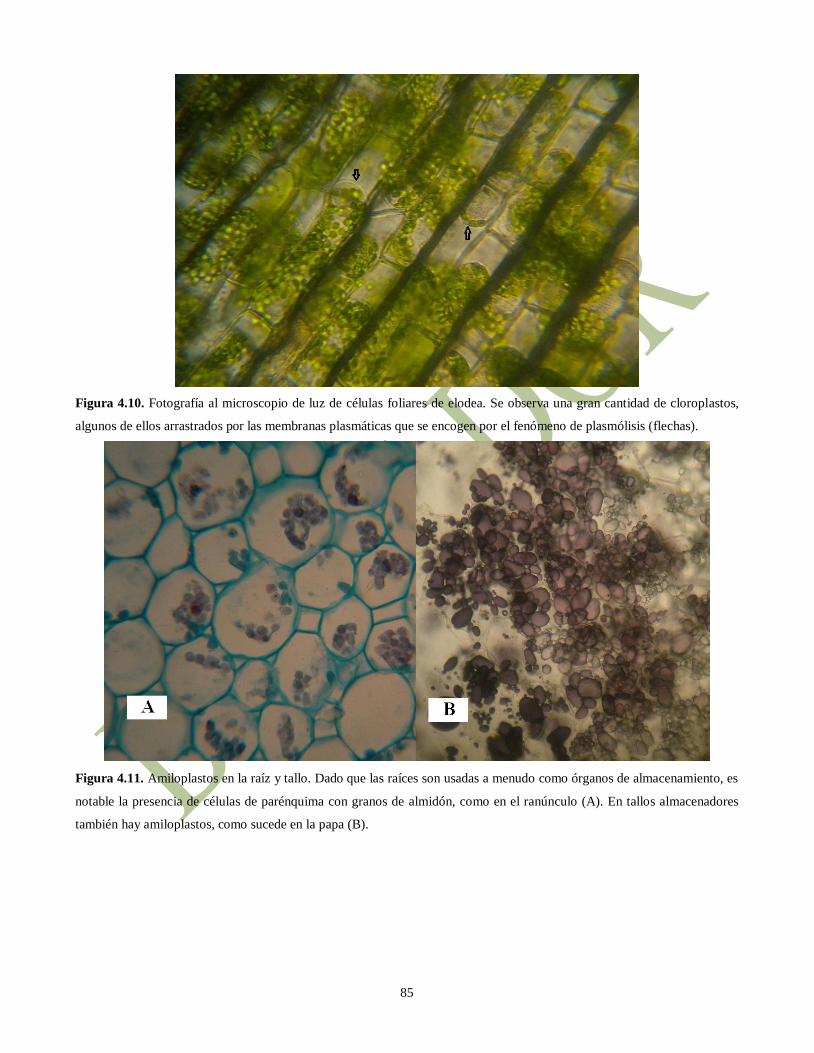
Figura 4.10. Fotografía al microscopio de luz de células foliares de elodea. Se observa una gran cantidad de cloroplastos,
algunos de ellos arrastrados por las membranas plasmáticas que se encogen por el fenómeno de plasmólisis (flechas).
Figura 4.11. Amiloplastos en la raíz y tallo. Dado que las raíces son usadas a menudo como órganos de almacenamiento, es
notable la presencia de células de parénquima con granos de almidón, como en el ranúnculo (A). En tallos almacenadores
también hay amiloplastos, como sucede en la papa (B).
86
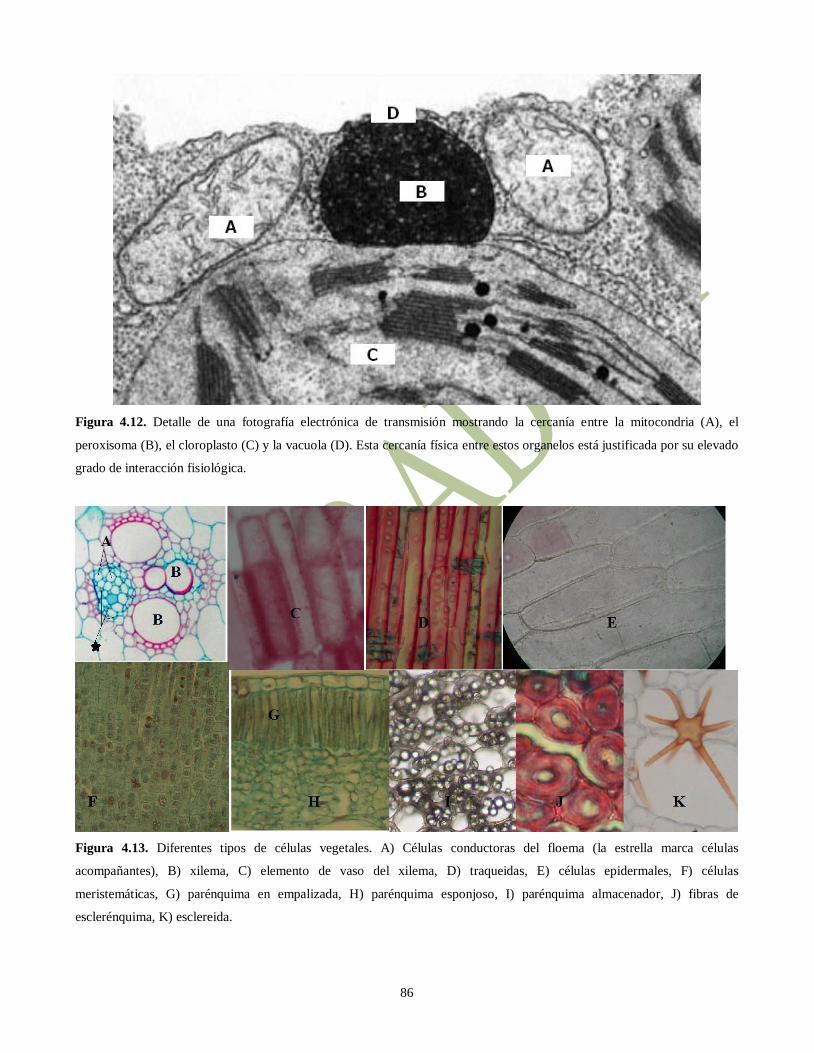
Figura 4.12. Detalle de una fotografía electrónica de transmisión mostrando la cercanía entre la mitocondria (A), el
peroxisoma (B), el cloroplasto (C) y la vacuola (D). Esta cercanía física entre estos organelos está justificada por su elevado
grado de interacción fisiológica.
Figura 4.13. Diferentes tipos de células vegetales. A) Células conductoras del floema (la estrella marca células
acompañantes), B) xilema, C) elemento de vaso del xilema, D) traqueidas, E) células epidermales, F) células
meristemáticas, G) parénquima en empalizada, H) parénquima esponjoso, I) parénquima almacenador, J) fibras de
esclerénquima, K) esclereida.