Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Caracterización de la toxicidad deCaracterización de la toxicidad deinsecticidas piretroides en Triatomainsecticidas piretroides en Triatoma
infestans (Klug)infestans (Klug)
Alzogaray, Raúl A.
1996
Tesis presentada para obtener el grado de Doctor en CienciasBiológicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Alzogaray, Raúl A.. (1996). Caracterización de la toxicidad de insecticidas piretroides enTriatoma infestans (Klug). Facultad de Ciencias Exactas y Naturales. Universidad de BuenosAires. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_2815_Alzogaray.pdf
Cita tipo Chicago:
Alzogaray, Raúl A.. "Caracterización de la toxicidad de insecticidas piretroides en Triatomainfestans (Klug)". Tesis de Doctor. Facultad de Ciencias Exactas y Naturales. Universidad deBuenos Aires. 1996. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_2815_Alzogaray.pdf

UNIVERSIDAD DE BUENOS AIRES
Facultad de Ciencias Exactas y Naturales
Caracterización de la Toxicidad
de insecficidas pire‘rroides
en Tn'amma /m‘esfans (Klug)
POR
RAÚL A. ALZOGARAY
DIRIGIDO POR
EDUARDO N. ZERBA
Centro de Investigaciones de Plagas e Insecticidas(CIPElN - CITEFA/CONICET)
Tesis presentada para obtener el titulode Doctor en Ciencias Biológicas
MCMXCVI
¿6'14

CARACTERIZACION DE LA TOXICIDAD DE INSECTICIDAS PIRETROIDESEN Triatoma infestans (Klug)
Tesis Doctoral de Raúl A. AlzogarayDin'gida por Eduardo N. Zerba
Resumen
Se evaluaron los síntomas de intoxicación con deltametrina y cis-permetn'na en ninfaslll de T. infestans. Se estimaron los TE 50s de distintas dosis para dos efectos: (a)incoordinación, y (b) parálisis total. Deltametn‘nafue tanto o más rápida en producir elprimer efecto (según la dosis). cis-permetrina fue más rápida en producir el segundo.No hubo diferencia significativa entre las DE 50 a diferentes tiempos de los dospiretroides, y las ninfas tratadas con cualquiera de ellos mostraron una importanterecuperación en función del tiempo. La recuperación fue inhibida cuando se aplicósimultáneamente butóxido de piperonilo.
Se estudió Ia influencia de distintas variables sobre la toxicidad dedeltametrina y cis-permetrina. La temperatura tuvo un efecto importante sobre las CE505 y DE 50s de los piretroides estudiados. En general la toxicidad aumentó aldisminuir la temperatura; pero hubo excepciones que dependieron del insecticida, elestadio ninfal y el rango de temperatura. El efecto de la temperatura sobre los TE 505fue poco importante. En general la velocidad de acción aumentó al disminuir latemperatura; pero hubo excepciones que dependieron del insecticida, el estadioninfal y el rango de temperatura. En experimentos con ninfas de distintas edades yestados alimentarios. resultaron más sensibles los individuosde 1 día de edad. y mástolerantes los alimentados 7 días antes. La aplicación tópica en el tórax produjosíntomas de intoxicación más rápidamente que la aplicación tópica en el extremoabdominal. Deltametrina actuó más rápidamente que cis-pen'netn'na en un ampliorango de dosis (2-1.000 ngfinsecto).
En ninfas I, el isómero cis de la permetn’na fue más tóxico que el isómero transa 26 y 36°C, e igual de tóxico a 16°C. El pretratamiento con butóxido de piperonilosinergizó la toxicidad de cis-permetrina pero no la de trans-permetrina. Elpretratamiento con trifenilfosfato tuvo el efecto opuesto. La mezcla de los dosisómeros produjo un efecto de aditividad (ausencia de interacción). En ninfas lll, elisómero cis de la permetn'na fue más tóxico que el isómero trans a 16 y 26°C, e igualde tóxico a 36°C. Los pretratamientos con butóxido de piperonilo o trifenilfosfato nomodificaron las DE 50s La mezcla de los dos isómeros produjo un efecto deantagonismo.
Ambos piretroides produjeron hiperactividad locomotora en ninfas lll. Esteefecto estuvo influenciado por el modo y el lugar de aplicación de los insecticidas. Lahiperactividad fue inhibida por el pretratamiento con N-etilmaleimida y por eldescenso de la temperatura.
Ninguno de los dos insecticidas tuvo efectos subletales sobre la ingesta y lamortalidad de ninfas l y lll, y sólo deltametn’na produjo una disminución en la cantidadde ninfas l mudadas.
PALABRAS CLAVES
Tn'atomainfestans, deltametn'na, cis-permetn'na. piretroides, síntomas de intoxicación,toxicidad, efectos subletales, actividad locomotora, coeficientes de temperatura, acción
conjunta, antagonismo.

b.O...0...0...0.00000....0...OOOOOOOOOOOOOOOOOOOOI
CHARACTERIZATION OF THE TOXICITYOF PYRETHROIDS INSECTICIDESON Tríatoma infestans (Klug)
Dissertation by Raúl A. AlzogarayDirected by Eduardo N. Zerba
ABSTRACT
Symptoms of poisoning by deltamethrin and cis-pen'nethn'n in third instars of T.¡nfestans were described. The ET 505 for two effects, (a) incoordination, and (b) totalparalysis were calculated. Deltamethrin was equal or more rapid to produce the onsetof the first effect (according the dose), and cis-permethn'n to produce the onset of thesecond one. There was not significant differences between DE 50s at different timesfor the two pyrethroids. They showed equivalent initial knockdown performance, butinstars recovery was observed. The synergist piperonyl butoxide inhibited recovery.
The toxicities of deltamethn‘n and cís-permetrhn'n at different intn'nsec andextn'nsec conditions were compared. After treatment temperature had an importanteffect on ED 505 of both pyrethroids, which tended to decrease when temperaturedecreased (but excepts depending of the insecticide, the instar, and the temperaturerange were observed). A low influence of temperature on ET 505 was observed. At alltemperatures assayed deltamethrin was faster than cis-permethn’n to producesymptoms of poisoning. The symptoms rapidly desappeared when instars intoxicatedwere moved from 16 to 28°C. Instars exposed to low doses of pyrethroids at 28°C,which did not show symptoms of poisoning, were rapidly affected when they weremoved to 16°C. The toxicity of both pyrethroids was affected by the age and thenutn'tional estate of the instars. Topical application on the thorax was faster toproduce the onset of incoordination than on the abdomen.
The toxicity of pure and mixed cis- and trans-permethn‘n was evaluated. Thecis isomer was more active than the trans one in both first and third instars. When thetwo isomers were simultaneously applied an additive effect (lack of interaction) wasobserved in first instars, while an antagonistic effect in third instars occured. In firstinstars, the toxicity of cis-permethrin was increased by piperonyl butoxide; and thetoxicity of trans-pennethrin was increased by triphenyl phosphate. In third instars, thetoxicity of both isomers was not modified by neither piperonyl butoxide nor triphenylphosphate.
Deltamethrin and cis-pennethn'n caused an increase in the Iocomotor activity ofthird instars. This effect was inhibited by a temperature decrease, and by Nethylmaleimide pretreatment.
Sublethal effects on blood meaI ingestion and moulting were evaluated in bothfirst and third instars. Only deltamethrin yielded a substantial decrease in the numberof moulted nymphs
KEY WORDS
Tn'afoma ¡nfestans, deltamethrin, cis-pen'nethrin, pyrethroids, symptoms of poisoning, toxicity,sublethal effects, Iocomotor activity, temperature coefficients, joint action, antagonism.

AGRADECIMIENTOS
Mis padres, mi hermana, y mis tios y abuela materna, apoyaron mi educaciónincondicionalmente y de todas las formas imaginables, desde los lejanos díasdel Jardín de Infantes hasta ahora.
Eduardo fue jefe, maestro y compañero de trabajo, ejemplar en los tresroles, y siempre dispuesto a reimos juntos de lo bueno, Io malo y Iofeo.
Edgardo, Mariné, y Susana L., me enseñaron transmitiéndome susconocimientos y a través de sus actitudes.
Andrea, Adriana M., Adriana C., Claudia, Emilia, Héctor, Mary, Norma,Paola Andino, y Susana S., cada uno me ayudó a su manera a enfrentar losincontables problemas cotidianos.
Claudio Lazzari, consejero de estudios, con quien sé que puedo contar.José Monserrat, viejo amigo, resolvió mis balurdos estadísticos.EI trabajo experimental se realizó en el Centro de Investigaciones de
Plagas e Insecticidas (Cipein), dependiente del Centro de InvestigacionesCientificas y Técnicas de las Fuerzas Armadas (Citefa) y del Consejo Nacionalde Investigaciones Cientificas y Técnicas (Conicet). Durante el desarrollo de lamayor parte de la Tesis fui becario de Iniciación y de Perfeccionamiento delConicet. La financiación del trabajo provino de subsidios otorgados a nuestrolaboratorio por Chemotecnica Sintyal, UNDPNVorId Bank/WHO SpecialProgramme for Research and Trainning in Tropical Diseases, y el Conicet.

ÍNDICE
I. INTRODUCCION, 11.1.LOS PIRETROIDES, 2
1.1.1.Desarrollo histórico, 21.1.2.Inferacción con los insectos, 9
1.1.2.1. Toxicocinéfica, 91.1.2.1.1.Penefración. disfribución
y excreción, 91.1.2.1.2.Mefabolismo. 10
1.1.2.2. Toxicodinómica, 141.1.2.3. Pire't'roides sí, ¿of-ros no? 151.1.2.4. Los pirefroides y el confrol de la
vinchuca, 191.2. ESTA TESIS. OBJETIVOS Y MOTIVOS, 21
1.2.1. Objefivos, 211.2.2.¿Por qué c/s-permefrina? 221.2.3. Habla el Tesisfa, 23
2. MATERIALES Y METODOS, 262.1. INSECTOS. 262.2. SUSTANCIAS, 262.3. ESTIMACIÓN DE PARÁMETROS
TOXICOLÓGICOS, 272.3.1. Obfención de los daros, 27
2.3.1.1. CE 50, 272.3.1.2. DE 50. 282.3.1.3. TE 50, 282.3.1.4. Aplicación de sinergis‘ras, 292.3.1.5. Condiciones ambienfales, 302.3.1.6. Efec'ros evaluados, 30
2.3.2. Cálculo de los parámefros, 312.3.3. Coeficienfes de Temperatura. 322.3.4. Coeficien'res de inferacción, 32

2.4. SINTOMAS DE INTOXICACIÓN, 332.5. CAMBIOS DE TEMPERATURA, 332.6. ACTIVIDAD LOCOMOTORA, 352.7. EFECTOS SUBLETALES, 372.8. LISTA DE ABREVIATUR/XS, 38
. RESULTADOS Y DISCUSIÓN, 393.1. síNTOMAs DE INTOXICACIÓN, 39
3.2.
3.3.
3.4.
3.1.1. Resul‘rados, 393.1.1.1. Fases de infoxicación. 393.1.1.2. VoH'eo y recuperación, 41
3.1.2. Discusión, 433.1.3. Figuras, Tablas y Lámina, 46ALGUN/A6 VARIABLES QUE AFECTAN LATOXICIDAD, 563.2.1. Temperafura, 56
3.2.1.1. ResuHados, 563.2.1.1.1. Cianopire'rroides, 563.2.1.1.2. C/s-permeTrina
versus deH'arneh'ina, 583.2.1.2. Discusión, 593.2.1.3. Figuras, 63
3.2.2. Composiciónde isómeros, 723.2.2.1. Resuh'ados, 723.2.2.2. Discusión. 733.2.2.3. Figuras, 77
3.2.3. Edad y es'rado alimenfario, 833.2.3.1. Resul‘rados, 833.2.3.2. Discusión, 843.2.3.3. Figuras, 86
3.2.4. Dosis y lugar de aplicación, 883.2.3.1. Resulfados, 883.2.3.2. Discusión. 883.2.3.3. Figuras, 90
EFECTO SOBRE LA ACTIVIDAD LOCOMOTORA, 923.3.1. Resulfados, 923.3.2. Discusión, 933.3.3. Figuras y Tablas, 97EFECTOS SUBLETALES. 1043.4.1. Resulfados, 1043.4.2. Discusión, 104

3.4.3. Tablas, 106
4. CONCLUSIÓN, 108
5 BIBLIOGRAFIA, 111

INTRODUCCIÓN
La palabra Toxicología deriva del griego antiguo toxicon, que significa “vida de
amor" (Repetto, 1988). Esta contradicción se repite en otros idiomas como el
inglés y el alemán, donde gifl significa simultáneamente regalo y veneno. En
griego contemporáneo toxon significa arco y toxikon se refiere a algo propio
para el arco y la flecha, quizás en referencia a las sustancias usadas paraenvenenar a éstas últimas.
La Toxicología es “la ciencia que estudia las sustancias químicas y los
fenómenos físicos en cuanto son capaces de producir alteraciones patológicas
a los seres vivos, a Ia par que estudia los mecanismos de producción de tales
alteraciones y los medios para contrarrestarlas, así como los procedimientos
para detectar, identificar y determinar tales agentes y valorar su grado de
toxicidad" (Repetto, 1988).
Un tóxico es entonces una sustancia capaz de producir efectos nocivos
sobre los seres vivos. El concepto actual de toxicidad tiene un carácter
relativo: cualquier sustancia puede tener efectos tóxicos por encima de cierto
umbral de concentración en el organismo. Esto ya fue sabiamente señalado en
el siglo XVIpor el célebre protocientífico suizo Theophrastus Philipus Aureolus
Bombastus von Hohenheim, más conocido como Paracelsus, cuando escribió
(en alemán y seguramente en una agobiante caligrafía gótica) “loque mata es
la dosis” (Hayes, 1975).
El trabajo experimental de esta Tesis se enmarca en la Toxicología de
Insectos o Entomotoxicología. Las sustancias estudiadas fueron los
insecticidas piretroides; el organismo fue Tn'atoma ¡nfestans, principal vector
de la enfermedad de Chagas en Argentina.

1.1. LOS PIRETROlDES
1.1. l. Desarrollo histórico
En el siglo XVII los habitantes del lado asiático de los Montes Cáucaso
conocían las propiedades insecticidas de las flores de ciertos crisantemos, y
las usaban contra los piojos del cuerpo. Doscientos años después, Armenia y
Yugoslavia producían y comercializaban esas flores con fines insecticidas.
(McLaughlin, 1973). Estos dos países dominaron el mercado hasta Ia Primera
Guerra Mundial. Mientras tanto se averiguó que los solventes orgánicos
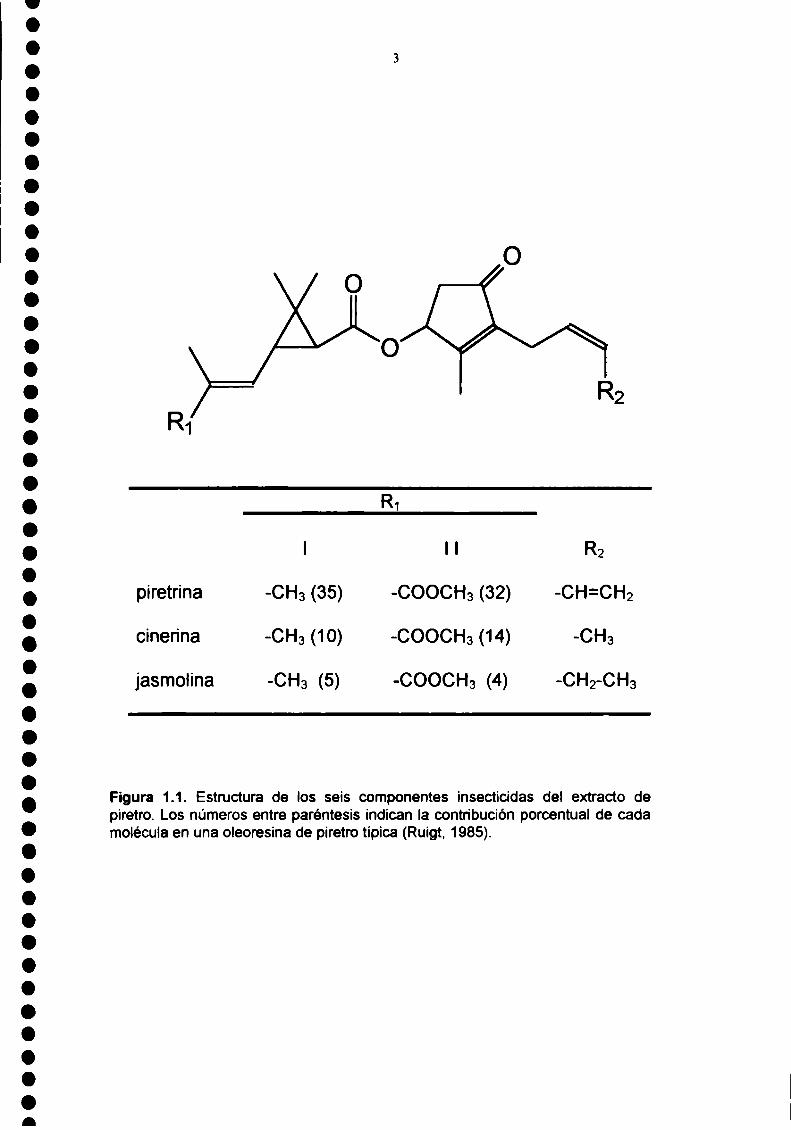
extraen de las flores secas del crisantemo una oleoresina que contiene seis
moléculas con propiedades insecticidas (Head, 1973). Estas moléculas, las
piretrinas, son ésteres provenientes de las distintas combinaciones entre dos
ácidos y tres alcoholes (Figura 1.1.). Los ácidos, crisantémico y pirétrico, son
ciclopropanocarboxilatos sustituidos. Los alcoholes, piretrolona, cinerolona y
jasmolona, son ciclopentenolonas sustituidas. Las principales características
estructurales de las piretrinas fueron esclarecidas entre 1910 y 1916 (pero
reveladas en 1924) por los quimicos suizos Hermann Staudinger y Leopold
Ruzicka, que años después recibieron sendos premios Nobel por sus aportes
en diversos campos de la química.
La planta que fabrica las piretrinas, Chrysanthemum cineranifo/¡um es
una herbácea perenne de la familia Compositae. Las piretrinas están
presentes en toda Ia planta, pero especialmente concentradas en las cabezas
florales (Head, 1973).
La Primera Guerra Mundial interrumpió el comercio con Dalmacia.
Entonces los japoneses, con una tradición de siglos en el cultivo de
crisantemos, dominaron el mercado. El auge japonés declinó durante los
primeros años ‘30, cuando se descubrió que el extracto de piretro producido en
Kenya contenía mayor concentración de piretrinas por unidad de peso seco
(McLaughlin, 1973). Y terminó de denumbarse cuando el emperador Hirohito
decidió atacar Pearl Harbour.
OO
O
R2R1
R1
I I l R2
piretrina -CH3 (35) -COOCH3 (32) -CH=CH2
cinen‘na -CH3 (10) -COOCH3 (14) -CH3
jasmolina -CH3 (5) -COOCH3 (4) -CH2-CH3
Figura 1.1. Estructura de los seis componentes insecticidas del extracto depiretro. Los números entre paréntesis indican la contribución porcentual de cadamolécula en una oleoresina de piretro típica (Ruigt, 1985).

En las décadas siguientes, el cultivo y producción de piretro se hizo
importante en Kenya, Tanzania, Ruanda, Ecuador y Nueva Guinea. El clima
ecuatorial de estos países permite que los crisantemos florezcan durante siete
a once meses al año (Casida, 1980). Hoy el piretro sigue ocupando un lugar
importante en el mercado de los plaguicidas. De acuerdo con la información
disponible, no presenta peligro de toxicidad aguda oral o dérmica en
mamíferos porque es rápidamente metabolizado (Elliot et al, 1972). En
trabajadores expuestos durante años, sólo se registraron unos pocos casos de
dermatitis, que desaparecieron al interrumpir la exposición (Barthel, 1973). No
hay evidencia de mutagénesis, carcinogénesis ni teratogénesis (Williams,
1973) y parece ser inocuo para la vida salvaje en general (Pillmore, 1973). Sin
embargo, el uso del piretro está retringido a las plagas domésticas y de granos
y frutas almacenados, porque la luz del sol lo degrada con facilidad (Mrak,
1973).
Por su utilidad contra los mosquitos transmisores de la malaria, Estados
Unidos declaró al piretro material estratégico de guerra en los años ‘40. Varios
intentos de cultivar los crisantemos en territorio norteamericano fracasaron. La
dificultad para obtener materia prima durante la guerra indujo en Estados
Unidos e Inglaterra la necesidad de sintetizar derivados de las moléculas
naturales. Unos años después aparecieron en el mercado los primeros
piretroides.
Además de esclarecer buena parte de la estructura de las piretrinas,
Staudinger y Ruzicka sintetizaron unos 100 análogos que no presentaron
actividad insecticida (Ruigt, 1985). Los esfuerzos iniciados durante Ia Segunda
Guerra Mundial permitieron obtener en 1948, en Estados Unidos, el primer
piretroide. Se llamó aletrina (su estructura y la de las moléculas mencionadas
en los párrafos siguientes están en la Figura 1.2.), y presentó una actividad
insecticida más rápida y más potente que las piretrinas en la mosca Musca
domestica. Alpoco tiempo se demostró que la aletrina era poco efectiva contra
otros insectos (Elliot, 1989). Esta matriarca de los piretroides se puede

()0
o o \O O \‘\ O
aletn'na resmetn'na
°Ü°© :W‘ÏC)permetrina deltametrina
9% °@C2H5O—©>—CICH20H2CH2‘@F
CH3
MTI - 800
Figura 1.2. Estructura de algunos piretroides.

conseguir hoy en cualquier supermercado, formulada en espirales y tabletas
termoevaporables.
El desafío que enfrentaban los quimicos era sintetizar moléculas que
reunieran las propiedades positivas de las piretrinas y que carecieran de sus
defectos. O sea: alta toxicidad en un amplio rango de insectos, acción rápida,
baja toxicidad en mamíferos y otros organismos no blanco, y estabilidad al aire
libre. En aquellos años, cuando se ignoraba casi todo acerca de la
toxicocinética y la toxicodinámica de las piretrinas, las nuevas moléculas se
buscaban introduciendo cambios empíricos en las estructuras conocidas. Se
tardaron veinte años en obtener la resmetrina, el primer piretroide con
actividad insecticida similar y toxicidad en mamíferos menor que las piretrinas
(Elliotet al, 1967). Unos años después se obtuvieron piretroides fotoestables
(Elliot et al. 1973). Moléculas como la pennetrina pudieron usarse al aire libre,
y ya nada impidióque los piretroides pasaran a encabezar las listas mundiales
de consumo de insecticidas.
AI incorporar un grupo cianuro a Ia estructura de los piretroides se
consiguió un aumento drástico de la acción insecticida (Elliot et aI., 1974). La
deltametrina es uno de los cianopiretroides más efectivos contra una
importante cantidad de plagas agrícolas, sanitarias y domésticas. La Tabla
1.1. ilustra la toxicidad y estabilidad en el ambiente de algunos piretroides.
Modificacionesestructurales posteriores demostraron que moléculas sin
la característica unión éster, como MTl-BOO, podían presentar un
comportamiento toxicológico similar al de las piretrinas (Udagawa et al, 1985).
El producto de la síntesis de muchos piretroides es una mezcla de
isómeros (Recuadro 1.1.) que en forma individual pueden presentar
toxicidades muy diferentes (Naumann, 1990). La tendencia actual es purificar o
sintetizar en forma dirigida los isómeros activos (Anónimo, 1988).
La producción mundial de piretroides se mide en miles de toneladas
anuales; el dinero que se destina a su compra en miles de millones de dólares
(Naumann, 1990). Seguramente esta situación se mantendrá mientras no
surjan alternativas más eficaces, veloces y baratas que la que ofrecen los
piretroides para el control de un amplio espectro de plagas.

DL 50‘l
(mg/k9)vida
Musca Tn'atoma media aIdomesticab ¡nfestans° rata aire libre
insecticida (contacto) (contacto) (oral) (días)
piretrina l 30 -— 420 < 1
aletrina 6 10,7 800 < 1
resmetrina 0,6 0,6 8000 < 1
permetrina 0,7 1,5 2000 > 20
deltametrina 0,02 0,3 100 > 20
a, Dosis Letal 50 (es Ia dosis que mata la mitad de la población expuesta); b, moscadoméstica; c, vinchuca.
Tabla 1.1. Actividad y estabilidad al aire libre de algunos piretroides (Elliot,
1989; Casabé et al, 1988),

Recuadro 1.1. lsomería de piretroides
Una molécula puede adoptardistintos arreglos espaciales. Estosarreglos se llaman isómeros yaunque tienen la misma fórmulaquímica, suelen difen'r en suspropiedades fisicas y químicas.Todos los piretroides presentanformas isoméricas y su actividadinsecticida es extremadamenteestereoespecifica.
lsomería ópficcCada átomo de carbono puedeunirse a otros cuatro átomos. Uncarbono unido a cuatro átomos ogrupos distintos se llama carbonoasimétrica o centro quirai. Losátomos unidos a un carbonosaturado siempre forman losvértices de un tetraedro regular.Bajo estas condiciones, sólo existendos ordenamientos posibles paraesos átomos, y cada uno es laimagen especular del otro. Lasmoléculas que reúnen estascondiciones se llamanenantiómeros. Una regla parareconocerlos es que uno de ellos nopuede ser transformado en el otrosin romperenlaces covalentes.
1 1
I 3
/c: 37K4 2 2 4
Si n es el número de carbonosasimétricos de una molécula.pueden existir 2” isómeros. Lamayoria de los piretroides tienentres carbonos asimétricos (el 1, el 3y el a) y entonces hasta ochoisómeros.
lsomería geomér'ïcaEstructuras químicas como el doble
enlgce (C= C) y el ciclopropano( ) forman un plano en elespacio. La isomería geométrica serefiere a la posición de lossustituyentes con respecto a eseplano. Si los dos sustituyentes seencuentran del mismo lado delplano, el isómero es cis; si seencuentran en lados opuestos delplano, el isómero es trans. Mirandola molécula de deltametn'na desdeel plano del ciclopropano, losisómeros geométricos se verían asi:
leórnero cls
r planodelciclopropano
:plano delciclopropanolsór'nerotrans
NomenclaturaLos isómeros se nombran deacuerdo con las reglas de Cahn,lngold y Pregold (CIP). Para lospiretroides se usa una simplificaciónde estas reglas. El carbono ‘l esllamado R cuando los sustituyentesestán ordenados en la dirección delas agujas del reloj, y S cuandoocurre lo contrario (para ordenar lossustituyentes existen a su vezciertas reglas). La estereoquimicade los carbonos 3 y a se definecomo cis o trans, según el arregloespacial de sus sustituyentesrespecto al carbono 1.De acuerdo con estas reglas, elnombre de la deltametn'na es (S)-aciano-3-fenoxibencil (1R)-cis-3-(2.2dibromovinil)-2,2-dimetilciclopropanocarboxilato.(información extraída de Alworth,1980; Zerba, 1988; Neumann, 1990)

1.1.2. Interacción con los insectos
1.1.2.1. Toxicocinéfica
El encuentro entre el insecto y el insecticida es el primer paso de la
interacción. A partir de ese momento, el insecticida ingresa al organismo y se
desencadena una serie de eventos: las moléculas son absorbidas, distribuidas,
almacenadas, metabolizadas y excretadas. Estudiar estos procesos es el
objetivo de la Toxicocinética (la segunda mitad de esta palabra compuesta
proviene del griego kinetíkos, que significa movimiento).
1.1.2.1.1.Penetración, distribución y excreclón.
La mayoría de los insecticidas sintéticos son lipofílicos y se disuelven con
facilidad en la capa serosa externa de la cutícula. Son rutas de ingreso al
organismo de los insecticidas de contacto los canales serosos que atraviesan
la cutícula (Welling and Paterson, 1985), las regiones donde Ia cutícula es más
delgada (membranas de las articulaciones, membranas intersegmentales)
(Ebeling, 1974; Fontán and Zerba, 1987), y el sistema traqueal, al cual
llegarían por desplazamiento lateral (Gerolt, 1983). La tasa de penetración
depende de las propiedades físicas y químicas de las sustancias y los
solventes usados para la aplicación (Brooks, 1976; Fontán and Zerba, 1987).
Cierta proporción del insecticida queda unido a la cutícula, donde puede ser
degradado por enzimas (Welling, 1979; Fontán and Zerba, 1984). La
disminuciónde la tasa de penetración por factores genéticos es un mecanismo
que confiere resistencia a insecticidas (Metcalf, 1989).
Atravesado el integumento, la hemolinfa distribuye el insecticida por
todo el organismo, unido a proteínas o dentro de partículas lipídicas (Brooks,
1976). Por sus características Iipofílicas, las moléculas insecticidas sólo son
excretadas después de sufrir modificaciones químicas que alteran su
polaridad. Estas transformaciones son hechas por las enzimas detoxicantes.

1.1.2.1.2. Metabolismo
“Ningún hombre es una isla", escribió en 1624 el poeta metafísico inglés
John Donne. Lo que sabemos hoy acerca del modo en que evolucionan los
organismos sobre Ia Tierra nos permite parafrasear a Donne: “ningún ser vivo
es una isla". Cada bacteria, hongo, planta y animal interactúa de incontables
maneras con otros individuos y con el ambiente. Estas interacciones son las
responsables de la evolución.
Un ejemplo de interacción es la que ocurre entre las plantas y los
insectos fitófagos. Las plantas fabrican moléculas que pueden resultar letales
para los insectos que las comen. Al mismo tiempo, en los insectos se
manifiestan mecanismos que confieren tolerancia o resistencia a esas
moléculas. Desde 1964, este proceso se llama coevolución (Berenbaum,
1986).
El principal mecanismo que protege a los insectos de la infinidad de
toxinas que ingieren continuamente es la actividad de las enzimas
detoxicantes. El uso del adjetivo detoxicante está bastante difundido en la
bibliografía sobre el tema, pero hay ejemplos donde su aplicación es discutible:
la actividad de ciertas enzimas consideradas detoxicantes genera productos
más tóxicos que el sustrato (ocurre con algunos insecticidas organofosforados
(Eto, 1974)). “Reacciones de activación y desactivación",
“biotransformaciones”, “metabolismo de xenobióticos", son términos usados
para nombrar el metabolismo de moléculas exógenas (Jacoby, 1980).
Las enzimas detoxicantes comparten dos características principales
(Brodie and Maickel, 1962): (a) convierten sustratos liposolubles en productos
hidrosolubles fáciles de excretar; (b) son muy inespecíficas. Su actividad está
dirigida hacia grupos funcionales (alquilos, arilos, ésteres), la estructura del
resto de la molécula no importa demasiado.
El metabolismo de las moléculas ajenas al organismo es bifásico (Sipes
and Gandolfi, 1991). Las reacciones de Fase 1 añaden o exponen grupos
funcionales. Son oxidaciones, reducciones e hidrólisis. Las reacciones de Fase
2 combinan esos grupos funcionales con metabolitos endógenos solubles en
agua (glucosa, aminoácidos, glutatión). Son conjugaciones.

ll
La actividad detoxicante varía entre especies, entre individuos, entre
momentos del desarrollo, con el sexo, con el estado nutricional y con Ia
naturaleza del ambiente (Terriere, 1984; Agosin, 1985; Dauterman, 1985;
Ruigt, 1985). La expresión exaltada de estas enzimas es uno de los
mecanismos que confieren resistencia a los insecticidas (Oppenoorth, 1985;
Devonshire, 1989).
Una manera de aumentar la toxicidad de un insecticida es aplicarlo
simultáneamente con un inhibidor de las enzimas que Io degradan. Los
inhibidores de enzimas detoxicantes se llaman sinergistas (Hodgson and Tate,
1976; Ruigt, 1985).
(a) HIDROLAGAS. Esta actividad consiste en la adición de una molécula de
agua a grupos carboxiéster, fosforoéster, carboxilamida y epóxido. Casi todos
los piretroides contienen un grupo carboxiéster que los convierte en sustratos
potenciales de las hidrolasas. Durante la hidrólisis se rompen el enlace éster y
una molécula de agua. Una mitad del éster se convierte en ácido, la otra en
alcohol.
RCOOR' —’ RCOOH+R'OH
La actividad hidrolasa en los insectos presenta un títqu en general bajo,
excepto en cepas resistentes a insecticidas (Dauterman, 1983). Su distribución
tisular suele coincidir con los puntos de ingreso, acumulación y acción de los
insecticidas en el organismo (Dauterman, 1985): cutícula, tubo digestivo,
cuerpo graso, sistema nervioso. En T. infestans se estudió la actividad de
hidrolasas de fosforoésteres durante el desarrollo embrionario (Villar et aI.,
1980), y en cutícula de adultos (Fontán and Zerba, 1984); se caracterizó y
determinó su localización subcelular (Casabé and Zerba, 1981); se aisló una
forma enzimática (Malkenson et al.,1984); y se desarrolló un método para
estudiar la actividad sobre análogos de la deltametrina (Wallace et aI., 1988).
La tasa de hidrólisis de los piretroides trans tiende a ser mayor que Ia
de los cis (Shono et al., 1979; lshaaya and Casida, 1983; Dowd and Sparks,

12
1988). La inhibición diferencial de estas actividades indica que existen
distintas enzimas y no una sola enzima con diferente especificidad hacia los
isómeros (Ruigt, 1985). EI metabolismo preferencial de los piretroides trans
puede ser la causa principal de la mayor toxicidad de los cis (Ahmad and
McCaffery, 1988; Dowd and Sparks, 1988; Toth and Sparks, 1988).
(b) OXIDASAB MICROSOMALES. La palabra microsoma deriva de dos
palabras griegas: micro, pequeño, y soma, cuerpo. Desde 1943 se la usa para
nombrar la fracción subcelular que sedimenta cuando se ultracentrifuga
(100.000 g) durante al menos una hora el sobrenadante posmitocondrial
(Agosin, 1985). En este sedimento, que contiene lípidos, proteínas y ácidos
ribonucleicos, se encuentran los restos del reticqu endoplásmico. En las
membranas reticulares existe un sistema enzimático involucrado en Ia
oxidación y reducción de moléculas lipofílicas. Sus componentes son la
NADPH - citocromo P450 reductasa, el citocromo P450 (una enzima con un
grupo hemo, cuyo nombre deriva del pico de absorbancia a 450 nm producido
por la forma reducida de la molécula cuando se une con el monóxido de
carbono) y un fosfolípido (Agosin, 1985; Sipes and Gandolfi, 1991). El
complejo está acompañado por otro citocromo, el bs, asociado a su vez con
otra reductasa. EI mecanismo de acción de las oxidasas microsomales implica
la ruptura de una molécula de oxigeno. Uno de los átomos es reducido a agua,
el otro oxida el sustrato. Los electrones provienen del NADPH.
NADPH NADP++ H+
RH + O2 ROH + HZC
La Figura 1.3. muestra algunos sitios de acción de la actividad oxidativa
sobre un piretroide.
Igual que las hidrolasas, la actividad de las oxidasas microsomales se
manifiesta con más intensidad en los lugares de acceso al organismo y tránsito
de los insecticidas: tubo digestivo y cuerpo graso (Nakatsugawa and Morelli,

O Ooxidación del O O oxidación delalcohol aldehído OH——>
HO OH
hidroxilaciónalifática
O X3
X1 o O
x rotura ’—_’2 del éster hidroxilación oaromática H
X3O
c"‘(“\OH + HO
Figura 1.3. Algunos productos de la acción de oxidasas microsomales sobre un
piretroide (Shono et aI._1979; Ruigt, 1985)

1976). En T. infestans se estudió la actividad de oxidasas microsomales hacia
el DDT (Agosin, et al. 1964; Agosin et al., 1969) y hacia el malatión (Wood et
a|., 1985).
(c) TRANSFER/36%. Son enzimas que conjugan moléculas exógenas con
metabolitos endógenos (aminoácidos, glutatión, glucosa, ácido glucurónico).
La conjugación de piretroides recibió hasta ahora mucha menos atención que
las actividades de hidrólisis y oxidación. Casi todos los metabolitos que
conservan intacto el enlace éster son excretados como conjugados (Soderlund
et al, 1983). La molécula endógena que se conjuga depende de la naturaleza
del metabolito exógeno y de la especie (Ruigt, 1985). En T. infestans se
estudió la actividad glutatión S-transferasa sobre insecticidas
organofosforados y su inhibicióncon diversas sustancias (Sívori, 1993)
1.1.2.2. Toxicodinámica
Una proporción generalmente pequeña del insecticida que entra en contacto
con el insecto alcanza un blanco molecular y ejerce su acción tóxica. Estudiar
esta interacción molecular y sus consecuencias es el objetivo de la
Toxicodinámica (la segunda mitad de esta palabra compuesta proviene del
griego dynamis, que significa fuerza).
Los venenos más poderosos que se conocen interfieren el
funcionamiento del sistema nervioso en vertebrados e invertebrados, y existen
en la naturaleza desde hace millones de años. Afectan al sistema nervioso las
toxinas del tétanos y el botulismo, las de la araña viuda negra y el pez globo,
las de los escorpiones y cobras, la nicotina y el piretro (Lund, 1985). Los
insecticidas sintéticos (clorados, fosforados, carbamatos y piretroides) son
también neurotóxicos.

15
electrofisiológicos, de flujo de iones y de unión a membranas señalan como
principales, y ya casi indiscutidos, sospechosos a los canales de sodio
dependientes de voltaje (Ruigt, 1985; Pauron et al. 1991, Osborne and Pipper,
1992). La evidencia farmacológica indica que los piretroides no se unen a
ninguno de los cinco sitios de unión a neurotoxinas identificados con otras
drogas, asi que se propuso la existencia de un sexto (Osborne and Pipper,
1992).
Algunos piretroides provocan descargas repetitivas en las membranas
nerviosas, otros bloquean la conducción de impulsos (Naumann, 1990). Por
éstas y otras caracteristicas, se los clasifica en Tipo l y ll (Tabla 1.2.). La
presencia o ausencia de un gmpo ciano en el carbono a, y las isomerías
ópticas y geométricas (Recuadro 1.1.) pueden determinar grandes cambios
cuantitativos y cualitativos en los efectos neurotóxicos (Ruigt, 1985; Coats,
1990). Aumentos en Ia densidad de canales de sodio o en la insensibilidad de
las membranas nerviosas para unirse a los piretroides, son mecanismos que
confieren resistencia a estos insecticidas (Osborne and Pipper, 1992).
Los piretroides afectan también la actividad de ATPasas, fosfatasas,
enzimas lisosomales, oxidasas microsomales, adenilciclasas, receptores
gabaérgicos y neurotransmisores (Ruigt, 1985; Vijverberg and van den
Bercken, 1990).
1.1.2.3.Pire'rroides sí. ¿otros no?
La síntesis de los cianopiretroides resolvió el desafío planteado en los
años '40: obtener moléculas que reunieran las ventajas de las piretrinas y no
presentaran sus desventajas. Los cianopiretroides son muytóxicos para un
amplio rango de insectos perjudiciales (Hirano, 1989; Carter, 1989), poco
tóxico para otros animales (lnglesfield, 1989; Hill, 1989), no tienen efectos
graves en trabajadores expuestos (Ecobichon, 1991), no se acumulan en los
seres vivos (Demoute, 1989), y su persistencia al aire libre es corta pero
suficiente para usarlos con éxito en el campo (Demoute, 1989) .

Tipo I Tipo ll
estructura química un grupo ciano en elcarbono a
síntomas en ratas - hiperexcitación- movimientos agresivosde las patas
- temblores en todo elcuerpo
- postración
síntomas en cucaracha - inquietud- incoordinación- postración- parálisis
efectos electrofisiológicos en vertebrados einvertebrados
dosis baias: descargasrepetitivas después deun único estímulo
dosis altas: despolarización permanente
aletrinaresmetrinapennetrina
ejemplos
sin grupo ciano
- sacudidas de cabeza- salivación abundante- convulsiones- coreoatetosis
- hiperactividad- incoordinación- convulsiones
despolarizaciónprolongada sindescargas repetitivas
cipennetrinadeltametrinafenvalerato
Tabla 1.2. Clasificación de los piretroides basada en los síntomas y efectos
electrofisiológicos que producen (Naumann, 1990; Ecobichon. 1991).

, chinches
Recuadro 1.2. La enfermedad de Chagas
Es una enfermedad crónicacausada por un protozoo flagelado,Trypanosoma cruzi. Las poblacionesde este parásito circulan entre loshospedadores, los vectores y losreservorios. Los vectores son
hematófagas queadquieren el parásito al alimentarsecon la sangre de un hospedadorinfestado (principalmentemamíferos). Después de laingestión, los parásitos migran alintestino medio de la chinche,sufren diversos cambiosmorfológicos y se dividen muchasveces. El vector queda infestadohasta la muerte. En el intestinomedio la morfología de los parásitosse modifica de nuevo. Las nuevasformas se desplazan al intestinoposterior.Las chinches hematófagas tienenhábitos nocturnos. Se posan sobrela piel de la presa dormida. la picany le suocionan la sangre. Mientrasse alimentan desalojan el contenidointestinal, liberando los parásitos. Através de escoriaciones,generalmente producidas por elrascado, los parásitos atraviesan lapiel y alcanzan los vasossanguíneos.La enfermedad de Chagas presentatres fases:(a) Fase aguda. Se produce unainflamación, chagoma. en el lugaren que los parásitos ingresaron alorganismo. El chagoma en el ojo esconocido como signo de Romaña.Los síntomas generales son fiebre,aumento de tamaño del higado y elbazo, y edemas generalizados. Untercio de los pacientes presenta
inegularidades cardíacas, que enniños menores de 2 años puedenser letales. Por debajo de esa edadtambién se producen complicaciones como la meningoencefalitis,que es letal el 50% de las veces. Enel resto de los casos, los sintomasdesaparecen espontáneamentedentro de las ocho semanas.(b) Fase indeterminada. No seobservan sintomas y las personaspueden desarrollar normalmenteactividades fisicas. Loselectrocardiogramas y radiografíasno presentan irregularidades. Lapresencia del parásito en la sangreno se detecta con metodos directos,pero sí por xenodiagnóstico(picadura de una chinche y posterioranálisis de su contenido intestinal)(c) Fase crónica. Se manifiestaentre 10 y 20 años después decontraer la enfermedad. Seproducen daños cardiacos,intestinales y neurológicos. Lasmuertes se deben principalmente afallas del corazón.Otras formas de adquirir laenfermedad son las transfusiones,el transplante de órganos y a travésde la placenta.Se estima que en latinoamérica 100millones de personas viven encondiciones de riesgo, 18 millonesestán infestadas, 3 millonespresentan complicaciones crónicas.Cada año 45.000 personas muerena causa de esta enfermedad. y1.000.000 la contraen.(Información extraída deCarlomagno et al.. 1989; WHO.1991; Moncayo, 1993)

Existen alternativas mucho más seguras para el ambiente y los
organismos no perjudiciales que los insecticidas sintéticos, pero con
limitaciones que impiden su uso masivo en el control de plagas. La técnica del
insecto estéril, las feromonas, los atractantes y repelentes de origen vegetal, el
control biológico, comparten haber despertado grandes entusiasmos al ser
concebidos como controladores de plagas, y alcanzado éxitos medianos o
nulos en los años posteriores. Esto se atribuye a que se usaron ensayos de
sensibilidad inadecuada, los bioensayos no concordaron con el uso en el
campo, no se tuvo en cuenta la variabilidad innata de los insectos, se
subestimó la complejidad de las interacciones entre los insectos y el ambiente
(Dent, 1991). En el caso de las feromonas hubo una tendencia a usarlas
apenas sintetizadas, sin estudiar sus limitaciones. En casos particulares Ia
especificidad del método es tan alta que no se justifica el esfuerzo necesario
para su desarrollo.
Algunos productos de la ingeniería genética son hoy candidatos a
ocupar puestos de privilegioentre las estrategias de control de las próximas
décadas. Plantas transgénicas productoras de toxinas insecticidas son ya una
realidad (Gasser and Fraley, 1992; Chilton, et al., 1993). La tasa y la
especificidad tisular de la expresión de los genes transgenizados todavía son
problemas por resolver (Gasser and Fraley, 1992). Otras cuestiones por ahora
sin respuesta son cómo se resolverá el surgimiento de resistencia a las
toxinas, y si la gente aceptará comer los frutos de las plantas transgénicas.
La acción rápida y efectiva, y la relación costo-beneficio justifica por
ahora el uso de los insecticidas sintéticos. En los programas de manejo
integrado de plagas, son una herramienta eficaz para reducir las poblaciones
de insectos antes de aplicar otros métodos de control. El uso racional puede
minimizar los riesgos. Mientras tanto habría que seguir buscando alternativas
que reduzcan los efectos indeseables de los insecticidas sintéticos.

l9
1.1.2.4. Los pire'rroides y el com-rol de la vinchuca
La enfermedad de Chagas (Recuadro 1.2.) es una endemia latinoamericana
que se extiende desde Méxicohasta Ia Patagonia. En la región ocupa el cuarto
lugar entre las patologías discapacitantes, después de las enfermedades
respiratorias, las diarreas y el SIDA(Schofield, 1994).
Loque puede hacer la medicina por un enfermo de Chagas es poco (Schofield,
1994). Sólo un par de drogas, nifurtimoxy benznidazol, curan la parasitemia si
se aplican los primeros días después de la infección. Estas drogas eliminan
también los sintomas de la fase aguda. La eliminación de los parásitos se hace
más improbable a medida que aumenta el tiempo entre el momento de la
infección y el comienzo del tratamiento. Además, estos remedios dan resultado
en dosis cercanas al límite de tolerancia: los efectos secundarios son
frecuentes y peligrosos. Los síntomas de la fase crónica pueden tratarse con
antiarrítmicos y vasodilatantes si son leves, y con marcapasos y cirugía si son
graves. Estas estrategias no siempre tienen éxito y significan un desembolso
que varía entre 700 y 1600 dólares anuales.
EIfracaso en el desarrollo de vacunas preventivas y la existencia de un
vector que, además de transmitir la enfermedad, se alimenta de la sangre de
las personas, completan el caracter funesto de esta desgracia latinoamericana.
Los expertos de la Organización Mundia de la Salud consideran que el
control de la enfermedad de Chagas es científica y tecnológicamente factible, y
que las herramientas y estrategias están disponibles desde hace más de 30
años (WHO, 1991). La ausencia de programas de control en la mayoría de los
países afectados se debe, según estos expertos, a causas políticas y
económicas, a la subestimación de la prevalencia de la enfermedad y del daño
social que causa. Es patético que las palabras de Carlos Chagas, escritas
hace 80 años, todavía tengan una triste vigencia: “Hay un designio nefasto en
el estudio de la tripanosomiasis. Cada trabajo, cada estudio, apunta un dedo
hacia una población mal nutrida, hacia un problema económico y social que a
los gobernantes les produce una tremenda desazón, pues es testimonio de
incapacidad para resolver un problema tremendo. Es un problema de

20
vinchucas que invaden y viven en habitaciones sucias, con habitantes
ignorantes, mal alimentados, pobres y envilecidos, sin esperanza y sin
horizonte social".
El control de los vectores parece ser la medida más rápida y eficaz para
interrumpir el ciclo del parásito (Metcalf, 1975). Aunque las chinches no
transmitieran enfermedades, su condición de chupadoras de sangre justifica
cualquier intento de erradicarlas de los lugares donde viven las personas.
Se conocen 105 especies de chinches hematófagas distribuidas en el
continente americano. En los intestinos de más de la mitad se detectó
Trypanosoma cruzi. El principal vector de la enfermedad de Chagas en
Argentina es T. infestans (Hemiptera: Reduviidae). Su nombre de entrecasa es
vinchuca, palabra quechua que significa “la que se deja caer" (de los techos
para picar a las personas).
En los años '50 y '60 el control químico de los vectores se basó en el
rociado de insecticidas clorados. En los años '70 se usaron carbamatos y
fosforados. Ensayos de laboratorio (Grupo de Entomotoxicología de CITEFA,
informe no publicado, 1978; de Licastro et al., 1982) y de campo (Gualtieri et
al, 1984) decidieron que los insecticidas de la década del '80 serían los
piretroides. Poco después se descubrió que la exposición de las vinchucas a
vapores subletales del insecticida clorado lindano y algunos piretroides
aumenta la velocidad de respiración y la incorporación de vapores por las vías
respiratorias (Zerba, et al., 1982). Es una manera de sinergizar la toxicidad de
insecticidas aplicados en forma de vapores o humos. Enseguida se
desarrollaron potes fumígenos con lindano mezclado con otros insecticidas,
incluidos los piretroides (Zerba et a|., 1988). El tipo de formulado y el tipo de
sinergismo no fueron las únicas novedades introducidas por los potes
fumígenos. Hasta ese momento casi todas las medidas de control eran hechas
por fumigadores profesionales estatales. Los potes permitieron la participación
de la comunidad y la descentralización de las actividades. Ahora dos
empresas comercializan tres versiones recientes de aquellos potes, todas con
piretroides. El Servicio Nacional de Chagas compra y distribuye los potes

2|
fumígenos, y existe la posibilidad de difundir su uso en otros países
latinoamericanos.
Un nuevo formulado no profesional de piretroides está siendo
desarrollado: telas impregnadas con insecticidas (Wood et a|., 1992; Zerba,
1995). Estas telas se colocan en los ranchos a modo de cielorraso y debajo de
los colchones de las camas (lugares de tránsito y refugio de las vinchucas), y
actúan como complemento de los rociados y los potes fumígenos. Ensayos
preliminares en domicilios de Gramilla (Dpto. Giménez, Santiago del Estero)
sugieren que las telas impregnadas con piretroides podrían prevenir la
reinfestación de las viviendas (Zerba, 1995).
El control químico de los vectores puede hacer mucho, pero no todo.
También deben fomentarse el saneamiento de los peridomicilios, la vigilancia
médica, el control de los bancos de sangre, el mejoramiento de las viviendas,
la educación sanitaria y la participación de las comunidades (WHO, 1991).
Imaginar América Latina sin Chagas suena utópico. Pero es una de esas
utopías a las que debemos acercarnos tanto como sea posible
1.2. ESTA TESIS. OBJETIVOS Y MOTIVOS
1.2.1.Objetivos
El objetivo general fue
- ampliar la escasa información disponible acerca de la toxicología de
piretroides de uso actual o potencial para el control de la vinchuca.
Los objetivos particulares fueron
- caracterizar y cuantificar los síntomas de intoxicación,
- estudiar la influencia de variables intrínsecas y extrínsecas sobre la toxicidad,
- evaluar el efecto sobre la actividad locomotora,
- evaluar efectos subletales.

22
Los experimentos se hicieron con ninfas l y lll de T. infestans. Se trabajó
principalmente con deltametrina y cis-permetrina, y en menor medida con
mezclas de isómeros de cipermetrina.
1.2.2. ¿Por qué c/s-perme‘rrina?
Durante años los principios activos y las formulaciones usadas en las
campañas de Chagas fueron seleccionados entre productos desarrollados por
fabricantes extranjeros. La idea que guía los objetivos de nuestro laboratorio
es que en la selección y el desarrollo de nuevos triatomicidas deben estar
involucrados laboratorios latinoamericanos (Zerba, 1989); y que son estos
laboratorios los que deben realizar una investigación racional que permita
diseñar productos especificos para controlar la vinchuca.
El desarrollo de nuevas moléculas insecticidas requiere un esfuerzo
económico impensable en regiones como la que habitamos. Se estima que el
desarrollo de un nuevo principio activo, desde su sintesis inicial hasta su
aplicación práctica, demanda unos 8 años y 100 millones de dólares
(Klassen, 1995). Una alternativa más apropiada para nuestra realidad es aislar
isómeros activos presentes en mezclas ya comercializadas. De esta manera se
acorta el camino para obtener nuevos productos. Buena parte de la
información toxicológica que hay que presentar para el registro ya está
disponible en el caso de los isómeros, porque fue requerida cuando se registró
la mezcla. Además, una vez que se identifican los isómeros inactivos, se
pueden desarrollar productos que no los posean y entonces evitar la aplicación
de moléculas sin actividad insecticida y con potencial impacto ambiental.
Teniendo en mente todo lo anterior, nuestro laboratorio decidió
investigar la toxicidad de isómeros aislados de piretroides. Y una de las
sustancias elegidas fue la permetrina.
Ya se discutió antes en esta Introducción que los piretroides, y en
particular deltametrina, son los insecticidas más usados en las campañas
argentinas para el control del vector de la Enfermedad de Chagas. La

23
perrnetrina comercial es una mezcla de isómeros que casi no se ha usado para
el control de la vinchuca. Estudios en campo sugirieron que se necesitan 0,5
g/cm2 para lograr un control efectivo, mientras que con 0,05 g/cm2 de
deltametrina puede obtenerse un resultado similar (Gualtieri et al, 1985). El
costo de la perrnetrina es similar al de la cipermetrina, pero la concentración
de cipermetrina recomendada para uso en Chagas es unas 4 veces menor
(Zerba, comunicación personal). Esta relación costo/concentración es la
principal causa por la cual no se usa perrnetrina para el control de la vinchuca
en Latinoamérica.
Los primeros ensayos realizados por nuestro laboratorio indicaron que
el isómero cis de la permetrina presenta una alta actividad vinchuquicida. Lo
interesante de este descubrimiento es que la perrnetrina se fabrica en
Argentina y la obtención de isómeros puros no implica técnicas particularmente
sofisticadas. Además, la cis-permetrina es sólida y puede formularse como
partículas micronizadas en agua con ayuda de un tensioactivo (formulado
floable). La perrnetrina, en cambio, es un aceite que sólo puede formularse
después de ser disuelto en un solvente orgánico (concentrado emulsionable).
El uso de solventes orgánicos en las formulaciones aumenta Ia alergenicidad
de los piretroides; en algunos casos el solvente solo es alergénico.
Estas circunstancias motivaron la ejecución de los experimentos que se
presentan en esta Tesis.
1.2.3. Habla el resista
Una fría tarde de invierno, hace algunos años, saqué de la cámara climatizada
unas ninfas de vinchucas tratadas con dosis bajas de piretroides. AI quedar
expuestas a la baja temperatura exterior las ninfas, que hasta ese momento se
veían normales, se incoordinaron en pocos minutos. No era novedad que el
descenso de temperatura suele aumentar la toxicidad de los piretroides, pero
yo no lo sabía. Por aquel entonces estaba terminando el Seminario de
Licenciatura y estábamos viendo qué sería de mi vida (quiero decir qué iba a

24
seguir investigando, por lo demás me sigo preguntando qué va a ser de mi
vida). En el laboratorio se estaba trabajando con isómeros de ciperrnetrina, y
como no había sido investigado antes, decidimos evaluar el efecto de la
temperatura sobre la toxicidad de estos piretroides. Asi empecé a hacer los
primeros experimentos de esta Tesis.
Casi enseguida descubrimos que la toxicidad en vinchuca de uno de los
isómeros geométricos de perrnetrina era equivalente a la toxicidad de
deltametrina. Decidimos que el resto de la Tesis estaria dedicado a comparar
distintos aspectos de la toxicología de estos dos piretroides. Los evaluamos en
distintas condiciones intrínsecas y extrínsecas, e investigamos si producían
efectos subletales.
Empíricamente desarrollamos un método para cuantificar la
incoordinación. Sabíamos que las ninfas tratadas con piretroides tardaban
varios días en morir, pero ¿qué pasaba mientras tanto?. Para averiguarlo
describimos y cuantificamos los sintomas de intoxicación.
Mientras tanto la Organización Mundial de la Salud aceptó nuestro
pedido de un analizador de imágenes y se abrió en nuestro laboratorio una
nueva línea de trabajo relacionada con el comportamiento y la actividad
sensorial de los insectos. A mí, que estaba estudiando síntomas y efectos
subletales de piretroides, me tocó estudiar la repelencia y el efecto sobre la
actividad locomotora. Parte de los resultados son presentados aquí.
No se acostumbra en estas circunstancias enumerar las cosas que se
quisieron pero no se pudieron hacer, así que no voy a mencionar que yo le
tenía muchas ganas a los estudios metabólicos de los isómeros de perrnetrina
y que el precio que nos pasaron para sintetizar las moléculas radiactivas me
quitó las ganas.
Ahora estoy entusiasmado con inminentes experimentos sobre los
efectos de los piretroides en los sistemas nerviosos central y periférico. Los
avatares politicos y económicos de la Argentina impiden prever si habrá vida
después de la Tesis. Uno viene sobreviviendo y habituándose a rumores de
interrupciones de sueldos y de becas, y pronósticos funestos sobre el futuro
inmediato de la investigación y los investigadores. Pero sigue metido en lo que

25
Io apasiona, con la esperanza de tiempos venideros mejores. Después de
todo, parafraseando al poeta García, “yo no quiero meterme en problemas, yo
no quiero asuntos que queman, yo tan solo les digo que es un bajón; yo no
quiero sembrar la anarquía, yo no quiero vivircomo digan, tengo algo que late
en mi corazón; yo no quiero vivir paranoico, yo no quiero ver chicos con odio,
yo no quiero sentir esta depresión; voy buscando el placer de estar vivo, no me
importa si soy un bandido, voy pateando basura en el callejón”.

2
MATERIALESY MÉTODOS
2. 1. INSECTOS
Se trabajó con ninfas I y IIIde T. infestans criadas en el insectario del Centro
de Investigaciones de Plagas e Insecticidas. La cria se realizó en una cámara
climatizada, a 28°C y un fotoperíodo 12: 12 (L: O). La alimentación se hizo
sobre paloma. Se usaron ninfas l de 7-10 días de edad, ayunadas desde la
eclosión; y ninfas lll de 11-16 días de edad, ayunadas desde la última muda
(excepto, para ambos estadios, cuando se indica otra cosa).
2. 2. SUSTANCI/XS
Todos los piretroides fueron de grado técnico (el origen se indica entre
paréntesis). Deltametrina (S)-a-ciano-3-fenoxibencil (1R)-c¡s-3-(2,2
dibromovinil)—2,2-dimetiIciclopropanocarboxilato (RousseI-Uclaf, Francia). Cis
y trans-permetrina 3-fenoxibencil (1RS)-cis, trans-3-(2,2-diclorovinil)-2,2
limetiIciclopropanocarboxilato (Environmental Protection Agency, USA;
Chemotecnica Sintyal, Argentina). Mezclas cis-permetrina: trans-permetrina 82:
18, 60: 40, 42: 58 (Chemotecnica Sintyal, Argentina), y 24: 76 (obtenida a
partir de la mezcla de los isómeros puros). Cipermetrina (RS)-a-ciano-3
fenoxibencil (1RS)-c¡s, trans-3-(2,2-diclorovinil)-2,2-dimetilciclopropanocarboxi
Iato (Chemotecnica Sintyal, Argentina). Mezclas de isómeros de Cipermetrina:

27
B-cipermetrina (38% /1Rcis/S+/1Sc¡s/R; 57,5% /1Rtrans/S+/1Strans/R)
(Chemotecnica Sintyal, Argentina); cipermetrina High cis (con alto contenido de
isómeros cis, en adelante llamada cipermetrina AC) (> 80%
/1Rcis/S+/1Scis/R+/1Sc¡s/S+/1Rcis/R) (Ciba Geigy, Suiza); alfametrina (95%
/1Rc¡s/S+I1Scis/R; 4,4% /1Sc¡s/S+/1Rcis/R) (Shell, Reino Unido).
Se usaron también butóxido de piperonilo (Chemotecnica Sintyal),
trifenilfosfato (Aldrich), N-etilmaleimida (Aldrich), y acetona para análisis
(Merck).
2. 3. ESTIMACIÓN DE PARÁMETROS TOXICOLÓGICOS
2.3.1. Obtención de los datos
2.3.1.1. CE 50
Los insectos fueron expuestos a filmes de los insecticidas sobre papel de filtro
Whatman Nro. 1. Los papeles fueron tratados con diluciones seriales de los
insecticidas en acetona (generalmente por un factor 2 ó 3) y se dejaron secar
30 minutos para eliminar el solvente por evaporación. La elección de
concentraciones que produjeran entre 0 y 100% de efecto se hizo en base a
los resultados de ensayos preliminares. Los controles fueron expuestos a
papeles tratados con acetona sola. La exposición se hizo en frascos de vidrio
con el fondo cubierto por los papeles tratados. La boca de los frascos fue
cubierta con una gasa sostenida por una banda elástica. Se expusieron al
menos 5 insectos por concentración, y se usaron al menos tres dosis para
estimar cada CE 50. Los resultados fueron evaluados a las 24 horas. Se
hicieron 2-3 réplicas independientes de cada experimento.

2.3.1.2. DE 50
Se hizo aplicación tópica de los insecticidas disueltos en acetona. Se usó una
microjeringa provista de pulsador, y cada insecto recibió 0,2 ul de solución en
la parte ventral del abdomen (excepto cuando se indica otra cosa). Se
aplicaron diluciones seriales de los insecticidas (generalmente por un factor 2
ó 3). La elección de dosis que produjeran entre 0 y 100% de efecto se hizo en
base a los resultados de ensayos preliminares. Los controles recibieron
acetona sola. Los insectos tratados fueron colocados en vasos de plástico con
un trozo de papel plegado en su interior. La boca de los frascos fue cerrada
con una gasa sostenida por una banda elástica. Se usaron al menos 5 insectos
por dosis, y al menos tres dosis para estimar cada DE 50. Los resultados
fueron evaluados a las 24 horas (excepto cuando se indica otra cosa). Se
hicieron 2-3 réplicas independientes de cada experimento.
2.3.1.3. TE 50
Se hizo exposición a filmes de insecticidas o aplicación tópica, como se
describió para las CE 505 y las DE 50s. Los controles fueron expuestos o
recibieron aplicación tópica de acetona sola. En cada experimento se aplicó
una única concentración o dosis de insecticida y se evaluó el resultado a
distintos tiempos.
Los insectos tratados fueron colocados en vasos de plástico con un
trozo de papel plegado en su interior. Se usaron al menos 5 insectos por
experimento, y al menos 5 tiempos para estimar cada TE 50. Se hicieron 2-4
réplicas independientes de cada experimento.
Se eligieron concentraciones o dosis que produjeran los efectos
buscados en tiempos relativamente cortos (pocas horas después de Ia
aplicación). A continuación se detallan las concentraciones y dosis usadas en
cada experimento y sus respectivas toxicidades.

29
concentración
experimento o dosis toxicidad
Figuras 3.2. y 3.3 0,02 pg/insecto DE 99 (deltametrina, 26°C, ninfas lll)
Figuras 3.2. y 3.3. 2 pg/insecto 100 x DE 99 (deltametrina, 26°C, ninfas III)
Figura 3. 6. 55,5 pg/cm2 10 x CE 99 (deltametrina, 28°C, ninfas Ill)
Figuras 3.7. y 3.8. 14 ng/cm2 0,01 x CE 5 (deltametrina, 28°C, III)
Figura 3.12. 0,1 pg/insecto 20 x DE 99 (deltametrina, 26°C, ninfas I)
Figura 3.13. 20 pg/insecto 100 x DE 99 (deltametrina, 26°C, ninfas Ill)
2.3.1.4. Aplicaciónde sinergistas
Se hizo exposición a filmes de los insecticidas sobre papel de filtro Whatman
Nro. 1. Los papeles fueron tratados con soluciones de sinergistas en acetona y
se dejaron secar 30 minutos para eliminar el solvente por evaporación. En los
experimentos de volteo y recuperación (Figura 3.4.) los insectos estuvieron
expuestos en forma permanente a 0,07 mg/cm2de butóxido de piperonilo. En
los experimentos con isómeros de permetrina (Figuras 3.18. y 3.19.),
estuvieron expuestos a butóxido de piperonilo o trifenilfosfato (0,7 mg/cm2 en
ambos casos) durante las 24 horas previas a la aplicación de los insecticidas.
En todos los experimentos se hicieron controles exponiendo insectos a
papeles tratados con acetona sola y con sinergistas disueltos en acetona.

2.3.1.5. Condiciones ambientales
Después de Ia aplicación de los insecticidas, los insectos fueron colocados en
una cámara climatizada a 28 ó 26°C (excepto cuando se indica otra cosa). El
fotoperíodo fue 12: 12 (L: O), y la humedad relativa varió entre 60 y 90%.
2.3.1.6. Efectos evaluados
Se evaluaron los tres siguientes efectos:
(a) Incapacidad de abandonar un disco de papel de filtro. Se colocó un
disco de papel de filtro (15 cm de diámetro) en el centro de una cuba
rectangular de plástico (21 x 33 cm). Los insectos fueron depositados en el
centro del disco de papel y estimulados con el toque suave de una pinza
entomológica de acero inoxidable. Los insectos controles (tratados con
acetona sola) abandonaban el disco de papel dentro de los 5 segundos
posteriores al estímulo, y muchas veces en ausencia de éste, dirigiéndose
rápidamente y en línea recta hacia las paredes de la cuba. Los insectos
tratados mostraron distintos comportamientos de acuerdo con el grado de
intoxicación. Insectos poco intoxicados demoraron más tiempo que los
controles en abandonar el círcqu de papel, porque sus movimientos eran
más lentos o porque se desplazaban en forma circular en vez de hacerlo en
línea recta. Insectos más intoxicados tenían paralizado el tercer par de
patas, se desplazaban con dificultad hasta el borde del papel pero no
podían abandonarlo porque las uñas de las patas posteriores quedaban
enganchadas en las irregularidades del papel. Insectos muy intoxicados con
incoordinación en todas la patas se arrastraban con dificultad, en forma
errática y sin llegar al borde del papel. Insectos altamente intoxicados
carecían por completo de actividad locomotora, aunque las patas seguían
agitadas por movimientos incoordinados.

3|
Se consideraron afectados aquellos insectos que no abandonaron el
disco de papel en 50 segundos, es decir en aproximadamente 10 veces el
tiempo empleado por los controles. Esta definición no se refiere a insectos
que muestran un síntoma determinado, abarca a insectos con distintos
sintomas. En otras palabras, se consideraron afectados todos los individuos
comprendidos en cierto rango de intoxicación y que tenían en común la
incapacidad de abandonar el circulo de papel. Se obró de esta manera
porque, a causa de la variabilidad biológica, en cualquier grupo de insectos
tratados con piretroides coexisten individuos con distintos grados de
afectación.
(b) Ausencia de actividad locomotora. Los insectos fueron observados
durante 50 segundos. Se consideraron afectados los que no mostraron
actividad locomotora (aunque continuaran agitando las patas en forma
incoordinada), ni siquiera después de ser estimulados con el toque suave
de una pinza entomológica.
(c) Ausencia de movimiento en patas, antenas y proboscis. Los insectos
fueron observados durante 2 minutos. Se consideraron afectados los que no
realizaron movimientoalguno de los apéndices, ni siquiera después de ser
estimulados con el toque suave de una pinza entomológica.
2.3.2. Cálculo de los parámel'ros
Se usó el programa Microprobit3.0 (Sparks and Sparks, no publicado), basado
en el método de Finney (1971). Este programa calcula CE 508, DE 503, o TE
503, con sus respectivos intervalos de confianza del 95%; y aplica factores de
corrección para hasta un 10% de mortalidad en los controles y para los casos
en que los datos son ligeramente heterogéneos. Los datos de las réplicas
fueron mezclados para calcular cada parámetro definitivo (Tabashnik et a|.,

32
1987). Se consideraron significativamente diferentes los valores cuyos
intervalos de confianza del 95% no se superpusieron (Toth and Sparks, 1990).
2.3.3. Coeficientes de Temperafura
Se calcularon dividiendo la CE 50 (o DE 50, o TE 50) mayor por Ia CE 50 (o
DE 50, o TE 50) menor (Toth and Sparks, 1988). Se asignó signo negativo a
los coeficientes cuando la CE 50 (o DE 50, o TE 50) a Ia mayor temperatura
fue mayor que Ia CE 50 (o DE 50, o TE 50) a la menor temperatura. De
acuerdo con esta definición, un coeficiente de temperatura puede ser positivo
o negativo, y tomar valores igual o mayores que 1. Los coeficientes iguales a 1
(neutros) corresponden a los casos en que la temperatura no afectó la
toxicidad. Se consideraron significativamente diferentes que 1 los coeficientes
calculados a partir de parámetros cuyos intervalos de confianza del 95% no se
superpusieron (Toth and Sparks, 1988).
2.3.4. Coeficientes de inferacción
Se usó el programa Bose-effect Analysis with Microcomputers (Chou and
Chou, 1987). Los coeficientes de interacción pueden ser menores
(sinergismo), iguales (aditividad), o mayores (antagonismo) que 1. Se
calcularon los coeficientes para cada réplica de los experimentos de mezclas
de cis- y trans-permetrina, se obtuvieron los promedios :l:DE y se determinó si
los valores eran significativamente distintos que 1 mediante la Prueba t para
una muestra del programa GraphPad Instat (Uchitel, 1990).

33
2.4. SÍNTOM/X‘S DE INTOXICACIÓN
Se hizo aplicación tópica, con una microjeringa provista de pulsador, en el lado
ventral del abdomen. Cada ninfa recibió 0,02 ó 2 pg de insecticida en 0,2 pl
de acetona. Se determinaron la naturaleza y el orden de aparición de los
síntomas de intoxicación por observación continua e individual de al menos 10
ninfas por dosis y por insecticida. Como controles se usaron ninfas tratadas
con acetona sola.
Para cada una de las dosis mencionadas se determinó la cantidad de
ninfas con el abdomen inflado y de ninfas recuperadas del volteo al cabo de 21
días. Se hicieron 4 réplicas independientes (n = 5). La idea inicial era someter
estos datos al análisis de la varianza. Algunos tratamientos no produjeron
efectos y la ausencia de varianza impidió usar esa prueba estadística.
Entonces se usó la Prueba t de Student del programa GraphPad lnstat
(Uchitel, 1990).
El efecto del tiempo sobre la DE 50 se analizó en la forma propuesta por
Sawicki (1962) con ligeras modificaciones. Se graficó DE 50 vs logaritmo del
tiempo para obtener una curva con 3 fases (fase1, descendente; fase 2,
ascendente; fase 3, meseta). El Punto Final del Volteo (PFV) es la mínima DE
50, es decir el punto de inflexión entre las fases 1 y 2. La Tasa de
Recuperación del Volteo (TRV) es Ia pendiente de la recta que se obtiene al
graficar las DE 503 de la fase 2 en función del logaritmo del tiempo. La TRV es
una medida de cuán rápidamente se recuperan del volteo los insectos. La
Proporción Efectividad: Volteo (PEV) se obtiene dividiendo la DE 50 en el PFV
por la DE 50 en Ia meseta, y es una medida de cuánta recuperación ocurre.
2.5. CAMBIOS DE TEMPERATURA
Se expusieron grupos de ninfas lIIa filmes de piretroides sobre papel de filtro
Whatman Nro. 1 (14 ng/cmz) durante 24 horas a 28°C. Los papeles fueron
tratados con soluciones de los insecticidas en acetona y se dejaron secar 30

34
minutos antes de exponer las ninfas. Después de la exposición las ninfas
fueron mantenidas otras 24 horas a la misma temperatura. La concentración
usada fue subtóxica, de modo que al término del segundo día ningún insecto
mostró síntomas de intoxicación. Se realizaron entonces tres tipos de
experimentos (en todos los casos el efecto evaluado fue Ia incapacidad de
abandonar un disco de papel de filtro):
(a) Las ninfas fueron transferidas de 28 á 16°C. AI disminuir la temperatura
aparecieron los sintomas de intoxicación.Entonces se determinó la cantidad
de ninfas afectadas en función del tiempo. Se calcularon los TE 505 como
se describió previamente. Se hicieron 2 réplicas independientes de cada
experimento (n = 20).
(b) Las ninfas fueron transferidas de 28 a 16°C. Dos y media ó 24 horas
después fueron regresadas a 28°C (en los dos casos, el 100% de las ninfas
estaba afectado al realizar el segundo cambio de temperatura). Entonces se
determinó la cantidad de ninfas recuperadas en función del tiempo (se
consideraron recuperadas las ninfas que abandonaron el disco de papel de
filtro en menos de 50 segundos). Se calcularon los Tiempos de
Recuperación del 50% de la población (TR 50) del mismo modo que se
describió antes para los TE 50s. Se hicieron 2 réplicas independientes de
cada experimento (n = 20).
(c) Las ninfas fueron transferidas de 28 a 16°C diariamente durante 2 horas.
Se determinó la cantidad de ninfas afectadas al cabo de ese tiempo y se las
regresó a 28°C. Se verificó que cada día, antes del primer cambio de
temperatura, ninguna ninfa mostró síntomas de intoxicación. Los datos
fueron analizados con el método de regresión lineal (Sokal and Rohlf,
1981).

2.6. ACTIVIDADLOCOMOTORA
Las ninfas fueron expuestas a filmes de los insecticidas sobre papel de filtro
Whatman Nro. 1. Los papeles fueron tratados con soluciones de los insecticida
en acetona y se dejaron secar 30 minutos para eliminar el solvente por
evaporación. Los controles fueron expuestos a papeles tratados con acetona
sola. Se trabajó con diluciones seriales (factor 10). Se eligió como mayor
concentración Ia que no produjo síntomas visibles de intoxicación dentro de los
30 minutos de exposición permanente, y como mínima Ia primera (en orden
descendente) que no tuviera un efecto significativo sobre la actividad
locomotora. La arena experimental fue un recipiente circular de plástico blanco
(7,5 cm de diámetro; 4 cm de alto) con velfondo cubierto con un disco de papel
de filtro tratado. Los insectos fueron depositados en la arena experimental e
inmediatamente se midió la actividad locomotora con un analizador de
imágenes.
Las imágenes fueron registradas con una cámara de video blanco y
negro (Sanyo) e ingresadas a un analizador de imágenes (Videomex-V,
Columbus) con una resolución de 256 x 192 pixels y una velocidad de
adquisición y procesado de 30 cuadros por segundo. Se usó el programa
Multiple Objects Motion for Videomex-V, que mide el movimiento de múltiples
objetos en una única zona. Para calcular la cantidad de movimiento durante
una determinada sesión, este programa compara Ia imagen actual con la
imagen previa y determina Ia cantidad de pixels que cambiaron de estado (de
encendido a apagado y viceversa). Al final de la sesión expresa la sumatoria
de los pixels que cambiaron de estado. También registra la cantidad de pixels
encendidos en cada imagen, los suma y los promedia por la cantidad de
imágenes procesadas durante Ia sesión. Este valor representa el área
promedio.
La arena experimental fue colocada a 15 cm de la cámara de video. Se
graduó el contraste de la imagen de tal manera que la cámara registrara
únicamente el tórax y el abdomen de las ninfas. La iluminación fue provista por
un tubo fluorescente (15 watts, estándar) colocado debajo del plano de la

36
arena experimental. Esta ubicación permitió eliminar por completo las sombras
producidas por los insectos, Ia cámara y las paredes del recipiente.
La actividad de las ninfas fue medida durante los 30 minutos siguientes
al comienzo de Ia exposición a los insecticidas (excepto cuando se indica otra
cosa). La información fue almacenada en una computadora IBMcompatible.
La actividad locomotora se expresó como pixels que cambiaron de
estado/ área promedio. De esta forma se relativizó el movimiento en función
del área, la cual no se mantuvo constante a lo largo de las sesiones. El área
disminuyó por ejemplo cada vez que una ninfa pasó por encima de otra, o se
irguió sobre sus patas intentando (infructuosamente) subir por las paredes del
recipiente. Este procedimiento experimental produjo resultados altamente
repetitivos.
Los experimentos se hicieron en un ambiente climatizado a 26 i 1°C. Se
hicieron 5 réplicas independientes de cada ensayo (n = 5). Los datos fueron
analizados con el método de regresión lineal (Sokal and Rohlf, 1981).
Después de cada experimento se verificóque las ninfas no presentaran
síntomas visibles de intoxicación, y se las mantuvo durante 24 horas en una
cámara climatizada a 26°C; entonces se determinó la cantidad de individuos
afectados. El efecto evaluado fue la incapacidad de abandonar un disco de
papel de filtro.
Se hicieron experimentos similares exponiendo las ninfas a una
concentración de 14 ug/cm2 de los insecticidas. En un caso se midió la
actividad a 16 y 36°C, en el otro a 26°C despues de tratar a las ninfas con N
etilmaleimida (aplicación tópica en el lado dorsal del abdomen de 20
ug/insecto 5 minutos antes de exponer a los piretroides).
También se hizo aplicación tópica de los insecticidas en el extremo
dorsal del abdomen o en el lado dorsal de Ia cabeza. Las dosis aplicadas se
eligieron con el mismo criterio usado para la elección de las concentraciones.
Cuando la aplicación se hizo en la cabeza de las ninfas, sólo se midió la
actividad durante 15 minutos, porque los síntomas de intoxicación se
manifestaron antes de 30 minutos. Igual que antes, las ninfas fueron

37
mantenidas durante 24 horas a 26°C, entonces se evaluó el efecto de los
piretroides.
Los datos de esto últimos experimentos fueron transformados a log x
para que reunieran los requisitos de normalidad y homocedasticidad, y
sometidos al análisis de Ia varianza (Sokal and Rohlf, 1981). Las medias
fueron separadas con la Prueba de rangos múltiples de Duncan (Duncan,
1955).
2.7. EFECTOS SUBLETALES
Se usaron ninfas l y IIIde 6 días de edad, ayunadas desde la eclosión o la
última muda. Se hizo exposición a filmes secos de piretroides (14 ng/cmz)
sobre tela (blanca, 100% algodón). Las telas fueron tratadas con soluciones de
los insecticidas en acetona y se dejaron secar 30 minutos antes de Ia
exposición. Los controles fueron expuestos a telas tratadas con acetona sola.
La elección de Ia concentración se realizó exponiendo grupos de ninfas a
diluciones seriales (factor 10) de deltametrina. Se eligió para el experimento la
máxima concentración que no afectó a los insectos al cabo de 7 días de
exposición.
Se trabajó con rectángulos de tela (6 x 6 cm) sobre los cuales se
colocaron aros de vidrio (5 cm de diámetro, 2,5 cm de alto). Se depositaron las
ninfas sobre las telas y se cubrió la parte superior de los aros con película
para transparencia. Al cuarto día de exposición las ninfas fueron alimentadas
sobre paloma. Se determinaron el peso antes y después de la ingesta, y la
cantidad de ninfas que no se alimentó. La exposición a los insecticidas fue
permanente durante 40 días. Entonces se determinó la cantidades de ninfas
que mudaron y que murieron. Se hicieron 5 réplicas independientes de cada
experimento (n = 15-20). Los resultados fueron sometidos al análisis de la
varianza (Sokal and Rohlf, 1981).

38
2.8. LISTA DE ABREVIATURAS
BP, butóxido de piperonilo.CE 50, concentración efectiva para el 50% de Ia población.Ci, coeficiente de interacción.Ciperrne'rrina AC, cipermetrina enriquecida en el isómero cis (> 80%)DE SO, dosis efectiva para el 50% de Ia población.NSIC, no superposición de los intervalos de confianza del 95%.PEV, proporción efectividadzvolteo.SIC, superposición de los intervalos de confianza del 95%.PF-‘V, punto final de volteo.TE 50, tiempo efectivo para el 50% de Ia población.TF, trifenilfosfato.
TR 50, tiempo de recuperación para el 50% de Ia población.TRV, tasa de recuperación del volteo.

3
RESULTADOS Y DISCUSIÓN
3. l. SÍNTOM/XS DE INTOXICACIÓN
3. l. 1. Resultados
3.1.1.1.Fases de intoxicación
Se determinaron los síntomas de intoxicación producidos por la aplicación
tópica de distintas dosis de deltametrina y cis-permetrina en ninfas III. Se
evaluaron para ambos piretroides el efecto sobre la actividad locomotora y los
TE 50 de distintos síntomas.
La Tabla 3.1. resume las fases de intoxicación, la variación de Ia
actividad locomotora y los síntomas en las patas de las ninfas intoxicadas.
La fase de latencia es el tiempo transcurrido desde que comienza la
exposición de las ninfas al insecticida hasta la aparición de los síntomas
visibles de intoxicación. En ese lapso, las moléculas insecticidas alcanzan un
blanco molecular e interaccionan con él.
El comienzo de la fase de excitación coincidió con la manifestación del
primer síntoma visible, que fue el aumento de la actividad locomotora o
hiperactividad. La Figura 3.1. muestra la variación de Ia actividad locomotora
en función del tiempo posterior al tratamiento. La aplicación tópica de 2
pg/insecto de deltametrina produjo un aumento inmediato de la actividad
locomotora. El efecto de igual dosis de cis-permetrina fue menos pronunciado.
La hiperactividad producida por estos piretroides en ninfas Ill de T. infestans

40
depende de Ia dosis, la forma de aplicación y Ia temperatura, y es afectada por
el tratamiento con N-etilmaleimida (un inhibidor de la quimiorrecepción). Los
resultados que fundamentan estas afirmaciones son presentados y discutidos
más adelante (3.3. Efecto sobre la actividad locomotora.).
Durante la fase de incoordinación la actividad locomotora disminuyó,
pero siguió siendo superior a la de los controles. En esta etapa se observaron
sintomas de intoxicación en los apéndices de la cabeza y el cuerpo. Los
síntomas en las patas fueron un buen marcador del desarrollo del proceso de
intoxicación. Los primeros síntomas fueron temblores leves y contracciones
momentáneas; por momentos las ninfas se desplazaban hacia atrás o de
costado. Al principio estos síntomas fueron intermitentes, y entre una y otra
manifestación las patas tenían un comportamiento normal. La frecuencia en la
aparición de los síntomas aumentó en función del tiempo y de la dosis de
insecticida. Otros síntomas esporádicos fueron el estiramiento de la proboscis
y las antenas rígidas apuntando hacia adelante. Avanzada la fase de
incoordinación se paralizó el tercer par de patas (Lámina 3.1.b.) y los
síntomas se manifestaron en forma continua. La Figura 3.2. muestra los TE
50s para 0,02 y 2 ug/insecto de deltametrina y cis-permetrina. El efecto
evaluado fue Ia incapacidad de abandonar un disco de papel de filtro. Los TE
503 de los dos piretroides fueron inversamente proporcionales a las dosis.
Cuando la dosis fue 0,02 ug/insecto no hubo diferencia significativa entre los
TE 505 de los dos insecticidas (SIC, P > 0,05); pero cuando fue 2 ug/insecto el
TE 50 de deltametrina fue significativamente menor que el de cis-permetrina
(NSIC, P< 0,05).
La fase de tetanización se caracterizó por violentos movimientos
convulsivos en todas las patas. La actividad locomotora fue menor que la de
los controles. En general las ninfas sólo podían arrastrarse con dificultad y las
patas incoordinadas formaban ángulos anormales con el cuerpo (Lámina
3.1.c.). Cerca del final de esta fase dejó de haber actividad locomotora, pero
las patas siguieron agitadas por convulsiones violentas.
En Ia fase de postración las ninfas yacían de costado o sobre la parte
dorsal del cuerpo. Los apéndices mostraban temblores y movimientos lentos y

41
esporádicos (Lámina 3.1.d.), cuya frecuencia disminuyó en función del tiempo
hasta desaparecer.
La Figura 3.3. muestra los TE 503 para el efecto ausencia de
movimiento en patas, antenas y proboscis. A las dos dosis aplicadas, la
ausencia de movimientos en los apéndices se produjo significativamente más
rápido en las ninfas tratadas con cis-permetrina (NSIC, P < 0,05). Las ninfas
tratadas con cualquiera de las dos dosis de deltametrina permanecieron unas
tres semanas en las fases de incoordinación o tetanización. La fase de
postración duró unas pocas horas y entonces se produjo la ausencia total de
movimientos. Las ninfas tratadas con cis-permetrina alcanzaron la fase de
postración dentro de las 24 horas después de la aplicación, y permanecieron
en ella alrededor de una semana antes que los movimientos desaparecieran.
Dentro de un mismo insecticida no hubo diferencia significativa entre los
tiempos efectivos de las dos dosis (SIC, P > 0,05).
Un efecto observado sólo en las ninfas tratadas con cis-permetrina fue
el inflamiento del abdomen, que ocurrió dentro de las 24 horas posteriores a la
aplicación (Lámina 3.1.e.). Las dos dosis de cis-permetrina aplicadas
produjeron este efecto (Tabla 3.2.), pero no hubo diferencia significativa entre
los porcentajes promedios de ninfas con el abdomen inflado para cada dosis
(Prueba t de Student; t = 0,7; g. I. = 6; P > 0.05). Las ninfas con el abdomen
inflado continuaron vivas varios días.
La muerte de las ninfas intoxicadas sólo fue evidente algún tiempo
después de ocurrida, cuando la deshidratación momificó a los insectos, la
cutícula se oscureció, y las patas y antenas se volvieron quebradizas al
tocarlas suavemente con una pinza de metal. Algunas ninfas adquirieron un
color rojizo debido al derramamiento del contenido intestinal en la hemolinfa.
3.1.1.2. Vol‘reo y recuperación
Se determinó la variación de las DE 505 de cada piretroide en función del
tiempo y el efecto del butóxido de piperonilo (inhibidor de Ia actividad de

42
oxidasas microsomales) sobre dicha variación. Se hizo aplicación tópica y el
efecto evaluado fue Ia incapacidad de abandonar un disco de papel de filtrode
15 cm de diámetro.
En la curva de DE 50 en función del logaritmo del tiempo para Ia cis
permetrina en ausencia de butóxido de piperonilo (Figura 3.4.b.), se
observaron tres fases: curva descendente (fase 1), curva ascendente (fase 2),
y meseta (fase 3). En Ia curva para deltametrina (Figura 3.4.a.) sólo se
observaron las dos primeras fases (el día 21 se interrumpió el experimento
porque Ia mortalidad en el grupo control superó el 10%).
En ninguno de los tiempos evaluados se encontró diferencia
significativa entre las DE 503 de deltametrina y cis-permetrina (NSIC, P >
0,05). El Punto Final del Volteo (PFV), es decir el tiempo para Ia mínima DE
50, fue el mismo para los dos insecticidas: 24 horas. La Tasa de Recuperación
del Volteo (TRV) de la deltametrina fue algo mayor que Ia de la cis-permetrina
(0,67 y 0,55; respectivamente). La Proporción Efectividad: Volteo (PEV) de Ia
deltametrina fue al menos tres veces mayor que la de la cis-permetrina (3 8,1 y
2,7; respectivamente). Estos resultados significan que ambos insecticidas
mostraron un poder de volteo similar, y que las ninfas tratadas con
deltametrina se recuperaron con mayor velocidad y en mayor cantidad que las
tratadas con cis-permetrina.
No se observó recuperación en ninfas expuestas a filmes de butóxido de
piperonilo después de Ia aplicación de los insecticidas. Las curvas de DE 503
en función del logaritmo del tiempo presentaron solamente las fases
descendente y meseta (Figuras 3.4.a. y 3.4.b.). Los PFVs para cada
insecticida no fueron significativamente diferentes de los respectivos valores
en ausencia de butóxido de piperonilo (SIC, P > 0,05).
Cuando se aplicaron dosis mayores, el porcentaje de ninfas
recuperadas dependió de la dosis para los dos piretroides (Tabla 3.3.).
Cuando la dosis fue 0,02 ug/insecto, no hubo diferencia significativa entre los
porcentajes promedio de ninfas recuperadas para deltametrina y cis-permetrina
(Prueba t de Student, t = 1, g. l. = 6, P > 0,05). Cuando la dosis fue 2
ug/insecto no se observó recuperación en ninguno de los dos tratamientos.

43
3.1.2. Discusión
Las fases y los síntomas generales de intoxicación en ninfas de T. ¡nfestans
tratadas con piretroides son similares a los descritos en Ia mosca M. domestica
(Adams and Miller,1980), la cucaracha Periplaneta americana (Gammon et aI.,
1981; Benoit et al, 1985), y larvas de las mariposas Pieris brassicae (Tan,
1982), Plute/la xy/oste/Ia (Chen et al., 1985), y Tn'chop/usia ni (Toth and
Sparks, 1990). En P. americana se estableció una correlación entre síntomas
visibles de intoxicación y eventos neurofisiológicos (Gammon, 1978).
El inflamiento abdominal es el unico síntoma no mencionado en la
bibliografía disponible. Este evento puede originarse en la producción
endógena de compuestos gaseosos.
En dosis toxicológicamente altas, los síntomas de incoordinación
aparecieron con mayor velocidad en las ninfas tratadas con deltametrina
(Figura 3.2.). Se verá más adelante que esta situación se repitió a otras
temperaturas (Figura 3.13.), en otro estadio ninfal (Figura 3.12.) y en un
amplio rango de dosis (Figura 3.21.). Inversamente, las ninfas tratadas con
cis-permetrina alcanzaron más rápido la fase de postración. y los resultados de
la Figura 3.3. sugieren que también murieron antes que las tratadas con
deltametrina. En algunos casos se demostró que la velocidad de acción está
relacionada con las propiedades físicas de las sustancias y en particular con la
polaridad (Briggs et al.,1976). La interpretación de este fenómeno es que las
características de partición en medios no polares y polares gobiernan la tasa
de transporte al sitio de acción de los insecticidas. La polaridad óptima para
producir incoordinación sería ligeramente mayor que la óptima para producir la
muerte. Los piretroides más polares penetrarían más rápido y se equilibrarían
a mayores concentraciones en la hemolinfa; los menos polares actuarían más
lentamente y se equilibrarían a mayores concentraciones en las membranas
celulares. Los resultados presentados aquí coinciden con esta tendencia, ya
que deltametrina es más polar que cis-permetrina (los coeficientes de partición
octanol/ agua son 270.000 y 1.200.000, respectivamente). Otros autores
llegaron a la conclusión que las características estructurales son más

importantes que las físico-químicas, y que la presencia del grupo ciano en
moléculas como deltametrina es determinante de las diferencias toxicológicas
(Clements and May, 1977).
El volteo fue definido como un estado de parálisis rápida y reversible
producido por piretrinas y piretroides (Miller and Adams, 1982). Todavía no
está claro si el volteo es el comienzo de un proceso toxicológico que termina
con Ia muerte cuando la dosis Io permite, o si los mecanismos de volteo y
Ietalidad son independientes (Naumann, 1990).
Las tres fases en Ia curva de DE 50 en función del logaritmo del tiempo
fueron descritas para los principios activos del piretro (Sawicki, 1962) y varios
piretroides (Scott and Georghiu, 1984) en la mosca M. domestica. En este
insecto el volteo (fase 1) se manifestó en cuestión de minutos, la recuperación
(fase 2) en cuestión de horas; el fin de la recuperación (fase 3) fue alcanzado
en un día. Los datos presentados aquí indican que Ia cinética del proceso fue
muy diferente en ninfas Ill de T. infestans: el efecto insecticida fue máximo
recién 24 horas después de la aplicación, la recuperación se prolongó a Io
largo de dos o tres semanas. Cuando cada insecticida fue aplicado
simultáneamente con el sinergista butóxido de piperonilo (un inhibidor de la
actividad de oxidasas microsomales), no se modificaron los PFVs ni los valores
mínimos de las respectivas DE 50s. Sin embargo, la presencia del sinergista
impidió la recuperación. Resultados similares obtenidos en M. domestica
(Sawicki, 1962) fueron atribuidos a la degradación de las moléculas
insecticidas en el sitio de acción. Esta hipótesis fue apoyada por evidencia de
muy pobre recuperación de la actividad nerviosa normal en experimentos con
sistemas nerviosos aislados tratados con piretrinas (Camougis, 1973).
Se cree que la muerte producida por los piretroides, y otros insecticidas
que actúan sobre el sistema nervioso, se debe a un descontrol de los
mecanismos de homeostasis (Lund, 1985; Naumann, 1990). Hormonas que

45
controlan los metabolismos de carbohidratos y lípidos, y los balances de agua
y sales a través de la excreción, son sintetizadas, almacenadas y liberadas por
el sistema nervioso. La evidencia experimental indica que concentraciones
muy bajas de piretroides (10'7 - 10'11 M) bastan para excitar las células
neurosecretoras (Naumann, 1990) e inducir la liberación de neurohormonas
como la trihalosémica (en la mosca Cal/iphora vecinis), Ia hiperlipémica y la
adipocinética (en Rhodnius prolixus) (Miller and Adams, 1982; Naumann,
1990). Una disminución en el volumen de la hemolinfa en ninfas de R. prolixus
tratadas con piretroides fue atribuida a la liberación de hormona diurética
(Maddrell and Casida, 1971). Pequeños cambios en las concentraciones de
iones de la hemolinfa pueden ser incompatibles con el funcionamiento normal
de los tejidos y conducir a la muerte celular.

46
3.1.3. Figuras, Tablas y Lámina
actividadlocomotora(respecto a síntomas en las patasinsectos no (en orden aproximado de
fase tratados) aparición)
latencia igual
excitación mayor (creciente)
incoordinación mayor(decreciente)
tetanización menor
postración ausente
muerte
ql
9°
.h
UI
.‘1
a:
(D
. temblores levesdesplazamiento haciaatrás o de costadocontracción de los trespares por debajo delcuerpo (breve)
. elevación por encimadel cuerpo (breve)
. parálisis del tercer par
violentos movimientosconvulsivos en los trespares
parálisis de los trespares
. temblores, movimientoslentos y ocasionales
. ausencia de movimiento
Tabla 3.1. Fases y sintomas de la intoxicación con piretroides en ninfas IIIde T.infestans. Se hizo aplicación tópica en la parte ventral del abdomen de deltametrinao cis-permetn'na (0.02 ó 2 ugfinsecto). La naturaleza y el orden de aparición de lossíntomas fueron evaluados por observación individual de 10 ninfas por dosis paracada insecticida.

47
actividad locomotora (pixels/área)1100 _ 1100
1ooo 3 denannnfina € 1ooo
900 } { 900
aoo } g soo
7oo } Q 7oo
coo 3 { soo
soo l í sooZ cis-permetrina Z
400 } { 4oo
300 l ‘ 300: control
_ r l r l l l r l L L4 L L L 1 1 1 l 1 1 1 l L L ro zo 4o so ao 100 120
tiempo (min)
Figura 3.1. Efecto de piretroides sobre la actividad locomotora de ninfas lll. Sehizo aplicación tópica en el lado ventral del abdomen (2 ugfinsecto). La actividadlocomotora se midió durante los 100 minutos posteriores a la aplicación, en 40sesiones de 2,5 minutos cada una. Actividad locomotora = P/A; donde P es elnúmero de pixels que cambiaron de estado durante la sesión (de apagado aencendido y viceversa), y A es el promedio de pixels encendidos durante Ia sesión.Cada curva se obtuvo promediando los datos de 2 réplicas independientes (n = 5).

43
Lámina 3.1. Síntomas de intoxicación con piretroides en ninfas Ill.Se hizo tópicade distintas dosis de deltametnna o cis-permetrina en el lado ventral del abdomen. (a)ninfa no tratada; (b) parálisis del tercer par de patas; (c) incoordinación general; (d)postración (una de las patas del tercer par se agita convulsionada); (e) abdomeninflado (observado sólo en ninfas tratadas con cis-permetrina).

J.,.,. Íu‘ _.4..
Whoooooíooooooooooobvocooo.
_
y.3.,
_ - r“uvy.
ya“,
’LY
o_.,1“yUÓCOÓOÓÓÓCOÓÓÓÓÓ..A....,

50
TE 50 (min)600 600
í a a [:1 deltametrinat 'Iï' ' ' cis-pennetrina .
500 - - 500
400 — —400
300 - - 300
zooF _ 20°' = fu c
100 - - 100
0 0
0,02 ¡ig/insecto 2 pglinsecto
Figura 3.2. TE 50 de piretroides en ninfas III.Se hizo aplicación tópica en el ladoventral del abdomen. El efecto evaluado fue la incapacidad de abandonar un discode papel de filtro. Las lineas verticales representan los intervalos de confianza del95%. Las barras señaladas con la misma letra no presentan diferencia significativa(SIC, P > 0.05). Cada valor se calculó con los datos obtenidos en 4 réplicasindependientes (n = 5).

51
o TE 50 (días)
25
20
15
10
Figura 3.3. TE 50 de piretroides en ninfas III.Se hizo aplicación tópica en el ladoventral del abdomen. El efecto evaluado fue la ausencia de movimientos en patas,antenas y proboscis. Las líneas verticales representan los intervalos de confianza del95%. Las barras señaladas con la misma letra no presentan diferencia significativa(SIC. P > 0,05). Cada valor se calculó con los datos obtenidos en 4 réplicas
30:1 deltametrina
cis-permetrina-25
0,02 ng/insecto 2 ng/insecto
independientes (n = 5).

porcentaje de ninfas con el abdomen inflado(promedio :t DE)
dosis(ug/insecto) deltametrina cis-permetrina
0,02 0 25,0 t 19,1a
2 0 35,0 :t 19,1 a
Tabla 3.2. Porcentaje de ninfas lll con el abdomen inflado después deltratamiento con piretroides. Se hizo aplicación tópica en el lado ventral delabdomen. Los valores seguidos por la misma letra no presentan diferenciasignificativa (Prueba t de Student, P > 0.05). Cada valor es el promedio de 4réplicas independientes (n = 5).
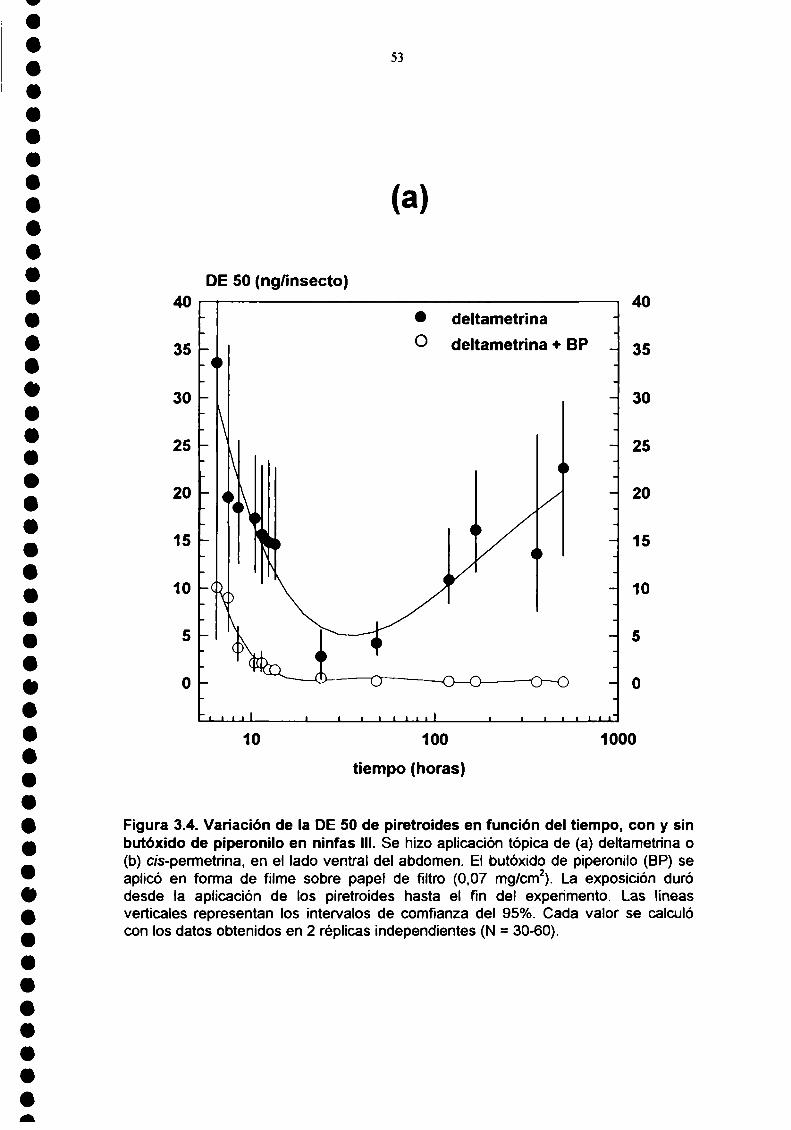
(a)
DE 50 (ng/insecto)40 40
C Ó deltametrina
35 _“ O deltametrina + BP _ 35
30 - - 30
25 - \ - 25
20% q, — 20
15 - “ d 15
1o 42‘ _. 10_ > .
5 F - 5
_ .1o— W do[Jl l l l l l l l ll I l l l l Ll_L-‘
10 100 1000
tiempo (horas)
Figura 3.4. Variación de la DE 50 de piretroides en función del tiempo, con y sinbutóxido de piperonilo en ninfas Ill. Se hizo aplicación tópica de (a) deltametrina o(b) cis-permetn'na, en el lado ventral del abdomen. El butóxido de piperonilo (BP) seaplicó en forma de filme sobre papel de filtro (0.07 mg/cmz). La exposición duródesde la aplicación de los piretroides hasta el fin del experimento. Las |¡neasvertica|es representan los intervalos de comfianza del 95%. Cada valor se calculócon los datos obtenidos en 2 réplicas independientes (N = 30-60).

50DE50 (ng/insecto)
(b)
45
40
35
30
25
20
15
10
ÏÏÍITITÏIÏÏÍÏÏIÏÏTIÏÏÏÏTÏIIIll]lllllïïïlllllllll
Figura 3.4.
JllllJ
Ó cis-permetrinaO cis-permetrina+ BP
Continuación.
tiempo (horas)
“50
45

porcentaje de ninfas recuperadas del volteo(promedio :t DE)
dosis(ug/insecto) deltarnetrina cís-permetrina
0,02 30,0 t 11,Sa 20,0 i 16,4a
2 0 0
Tabla 3.3. Recuperación del volteo en ninfas IIItratadas con piretroides.Se hizo aplicación tópica en el lado ventral del abdomen. Los resultados seevaluaron 21 días después del tratamiento. Los valores seguidos por lamisma letra no presentan diferencia significativa (Prueba t de Student, P >0,05). Cada valor es el promedio de 4 réplicas independientes (n = 5).

56
3. 2. ALGUN/06 VARIABLES QUE AFECTAN LA TOXICIDAD
3. 2. 1. Temperatura
3.2.1.1. Resultados
3.2.1.1.1. Cianopirefroides
Se evaluaron las CE 508 y los TE 505 a 16 y 28°C de deltametrina,
cipermetrina, B-cipermetrina, alfametrina y cipermetrina AC en ninfas Ill. Las
ninfas fueron expuestas a filmes de los insecticidas y el efecto evaluado fue la
incapacidad de abandonar un disco de papel de filtro.
La Figura 3.5. muestra las CE 503 a 16 y 28°C. A 28°C, la
deltametrina presentó Ia menor CE 50, y la cipermetrina la mayor. A 16°C, no
hubo diferencia significativa entre las CE 505 de los cinco cianopiretroides
(SIC, P > 0.05). Todos los insecticidas fueron significativamente más tóxicos a
16 que a 28°C (NSlC, P < 0,05), por lo tanto todos los coeficientes de
temperatura fueron negativos. La toxicidad de alfametrina fue Ia más afectada
por el descenso de la temperatura (su coeficiente fue —7,8), y Ia de B
cipermetrina la menos afectada (su coeficiente fue - 3,8).
La Figura 3.6. muestra los TE 50s a 16 y 28°C de los mismos
insecticidas de Ia figura anterior. A 28°C, la cipermetrina AC presentó el mayor
TE 50. Los TE 505 de los demás insecticidas no fueron significativamente
distintos unos de otros (SIC, P > 0.05). La cipermetrina AC tuvo el mayor TE
50 a 16°C, Ia alfametrina el menor. La acción de todos los insecticidas fue
significativamente más rápida a 28 que a 16°C (NSlC, P < 0.05), por Io tanto
los coeficientes de temperatura tuvieron signos positivos. El efecto de la
disminución de la temperatura sobre los TE 503 fue menos importante que
sobre los CE 50. Los coeficientes de temperatura variaron entre 1,4 para la
alfametrina y Ia cipermetrina AC, y 2,1 para la cipermetrina.
Para determinar si el efecto de Ia temperatura sobre Ia toxicidad de los
cianopiretroides era reversible, se hicieron tres tipos de experimentos. En los
tres las ninfas fueron tratadas durante 24 horas con una concentración

57
subtóxica de deltametrina o B-cipermetrina sobre papel de filtro (14 ng/cmz). La
temperatura fue 28°C. Después de interrumpir la exposición, las ninfas fueron
mantenidas otras 24 horas a la misma temperatura, entonces se realizó lo quese describe a continuación.
En el primer experimento (Figura 3.7.), ninfas intoxicadas que no
mostraban síntomas visibles fueron transferidas de 28 a 16°C. AI cambiar la
temperatura aparecieron los síntomas de intoxicación. Se evaluó la
incapacidad de abandonar un disco de papel de filtro. Cuando el cambio de
temperatura se hizo 1 día después de interrumpir la exposición a los
insecticidas, los síntomas de intoxicación aparecieron en pocos minutos y no
hubo diferencia significativa entre los TE 503 de la deltametrina y la B
cipermetrina (SIC, P > 0.05). Cuando el cambio de temperatura se hizo 6 días
después de interrumpirla exposición, los síntomas aparecieron en pocas horas
y se manifestaron significativamente más rápido en los insectos tratados con
deltametrina (NSIC, P < 0.05).
En el segundo experimento (Figura 3.8.), ninfas intoxicadas que no
mostraban sintomas visibles fueron transferidas de 28 a 16°C . Dos y media o
24 horas después las ninfas fueron llevadas de nuevo a 28°C (en ambos
casos, el 100% de las ninfas estaban afectadas al realizar el segundo cambio
de temperatura). Los TR 50 de insectos que estuvieron 2,5 o 24 horas a 16°C
no mostraron diferencia significativa (SIC, P > 0.05). En los dos casos, las
ninfas tratadas con deltametrina se recuperaron en forma significativamente
más lenta (NSIC, P< 0.05).
En el tercer experimento (Figura 3.9.), ninfas intoxicadas que no
mostraban síntomas visibles fueron transferidas diariamente de 28 a 16°C
durante 2 horas y entonces regresadas a 28°C. El porcentaje promedio de
ninfas afectadas al cabo de esas 2 horas disminuyó en forma lineal en función
del logaritmo del tiempo (deltametrina, y = 95,5 - 113,5 x, r2 = 0,97; B
cipermetrina, y = 54,9 —83,8 x, r2 = 0,97).

58
3.2.1.1.2. C/s-perrnerrina versus deltametrina
Se evaluaron las DE 505 y los TE 503 a 16, 26 y 36°C de deltametrina y cis
permetrina en ninfas I y Ill. Se hizo aplicación tópica y el efecto evaluado fue
ausencia de actividad locomotora.
La Figura 3.10. muestra las DE 50s a 16, 26 y 36°C de deltametrina y
cis-permetrina en ninfas I. La toxicidad de la deltametrina fue
significativamente mayor a 16°C que a las otras temperaturas (NSIC, P < 0.05).
Las DE 505 de este insecticida a 26 y 36°C no fueron significativamente
diferentes (SIC, P > 0.05). Los coeficientes de temperatura de Ia deltametrina
fueron negativos en los intervalos 16-26 y 16-36°C; en el intervalo 26-36°C el
coeficiente fue neutro (= 1).
La toxicidad de la cís-permetrina fue significativamente mayor a 16°C
que a las otras temperaturas (NSIC, P < 0,05), la menor toxicidad fue a 36°C.
Las DE 503 disminuyeron en forma significativa al disminuir la temperatura, por
lo tanto el signo de los tres coeficientes de temperatura fue negativo.
A 26°C la toxicidad de los dos insecticidas no fue significativamente
diferente (SIC, P > 0.05). A 16 y a 36°C la deltametrina fue significativamente
más tóxica que Ia cis-permetrina (NSIC, P < 0.05).
La Figura 3.11. muestra las DE 50s a 16, 26 y 36 °C de deltametrina y
cis-permetrina en ninfas lll. Las DE 505 de la deltametrina a 16 y 36°C no
fueron significativamente distintas (SIC, P > 0.05), pero la DE 50 a 26°C fue
significativamente mayor que las otras dos (NSIC, P < 0.05). Como
consecuencia, los coeficientes de temperatura fueron negativos en el rango
16-26°C y positivo en el rango 26-36°C. La DE 50 de la cis-permetrina no varió
en el intervan 16-26°C, pero aumentó en forma significativa en el intervan 26
36°C (NSIC, P < 0.05), y los coeficientes correspondientes a ese rango y a 16
36°C tuvieron signos negativos.
Las Figuras 3.12. y 3.13. muestran los TE 503 a 16, 26 y 36°C de
deltametrina y cis-permetrina en ninfas | y Ill, respectivamente. Los TE 50s de
la deltametrina en ninfas I disminuyeron en forma significativa al aumentar la
temperatura (NSIC, P < 0.05). Los coeficientes de temperatura tuvieron
entonces signo positivo. En ninfas Ill, los TE 505 a 16 y 36°C de deltametrina

59
no presentaron diferencia significativa (SIC, P > 0.05), pero sí fue
significativamente mayor que ambos el TE 50 a 26°C (NSIC, P < 0,05). Los TE
505 de Ia cis-permetrina no variaron significativamente al variar Ia temperatura
en ninguno de los dos estadios ninfales (SIC, P > 0.05), y los coeficientes de
temperatura fueron neutros.
3.2.1.2. Discusión
Clásicamente se considera que los piretroides presentan un coeficiente
negativo de temperatura. Esto significa que Ia toxicidad es mayor cuando Ia
temperatura disminuye (Ruigt, 1985). Pero una cantidad de contraejemplos
prueban que la temperatura puede afectar de otras maneras Ia toxicidad de
estos insecticidas.
Diversos piretroides presentaron coeficientes negativos de temperatura
en las moscas M. domestica (Ahn et a|., 1987) y Haematobía ¡m'tans (Schmidt
and Robertson, 1986); larvas de las mariposas T. ni (Sparks et al, 1982; Toth
and Sparks, 1988; Toth and Sparks, 1990), Spodoptera frugiperda, He/¡othis
virescens (Sparks et al, 1982), y Anthonomus grandis grandis (Sparks et al.,
1983); larvas de los escarabajos Leptinotarsa decemlíneata (Grafius, 1986), y
Tn’boliumcastaneum (Watters et aI., 1983); y la cucaracha Blate/Ia germanica
(Wadleigh et aI., 1991).
Se obtuvieron coeficientes positivos de temperatura en la mosca H.
¡rn'tans(Schmidt and Robertson, 1986); larvas de las mariposas Achaea janata,
Mythimna separata y Bombyx mori (Yadwad and Kallapur, 1988), S. frugiperda,
Heliothis virescens (Sparks et a|., 1982), Pectinophora gossypiella (Schouest
and Miller, 1988), y T. ni (Toth and Sparks, 1988); y larvas de los escarabajos
L. decemlíneata (Grafius, 1986), Sitophilus oryzae (Thaung and Collins, 1986),
y T. castaneum (Watters et al., 1983).
Y coeficientes neutros (= 1) en la mosca H. ¡rn'tans (Schmidt and
Robertson, 1986); larvas de las mariposas T. ni (Toth and Sparks, 1988; Toth
and Sparks, 1990), S. frugiperda, y H. virescens (Sparks et aI., 1982); y larvas
del escarabajo L. decemlíneata (Grafius, 1986).

El signo y eI valor absoluto de los coeficientes de temperatura de los
piretroides dependen de Ia especie, el insecticida, eI modo de aplicación y el
rango de temperatura.
Los coeficientes de los ¡sómeros cis y trans de la permetrina
presentaron distintos signos y valores absolutos según se hiciera aplicación
tópica o incorporación en la dieta de T. ni (Toth and Sparks, 1988). Lo mismo
ocurrió con los coeficientes de la permetrina cuando se compararon los
resultados de aplicación tópica y exposición a filmes en H. ¡rn'tans (Schmidt
and Robertson, 1986).
En T. castaneum, los coeficientes de la cipermetrina y el fenvalerato
fueron negativos para el rango 20-30°C, y positivos para el rango 10-20°C
(Watters et aI., 1983). Los coeficientes de varios piretroides tuvieron distintos
signos según fueran evaluados en los rangos 14-23, 23-30, ó 30-35°C en L.
decem/ineata (Grafius, 1986).
El efecto de la temperatura sobre la velocidad de aparición de síntomas
ha sido menos estudiado. Se encontraron coeficientes positivos de
temperatura (mayor velocidad de acción a mayor temperatura) en larvas de las
mariposas A. janata, M. separata y B. mory (Yadwad and Kallapur, 1988) y T.
ni (Toth and Sparks, 1990)
Los datos presentados en esta Tesis confirman el comportamiento
impredecible de los piretroides en función de la temperatura. En T. infestans, el
signo y la magnitud de los coeficientes de temperatura de los piretroides
estudiados varió con el rango de temperatura, el estadio ninfal y el insecticida.
Todos los procesos fisiológicos, y por extensión los toxicológicos, son
influenciados por Ia temperatura. Se puede esperar entonces que cualquier
paso de la toxicocinética y Ia toxicodinámica de los insecticidas esté
involucrado en el efecto de la temperatura sobre la toxicidad.
En P. amen'cana, a igualdad de dosis, la penetración del DDT a través
de la cutícula ocurrió en mayor proporción a mayores temperaturas (Busvine,
1971). Pero la toxicidad del DDTen este insecto fue mayor a baja temperatura.

61
En M. domestica, la toxicidad y Ia penetración también fueron afectadas en
forma inversa por la temperatura (Ahn et al, 1987). Al menos en estos casos,
la penetración no pareció ser un factor crítico para el resultado final.
La velocidad de las reacciones catalizadas por enzimas se incrementa,
en general, al aumentar la temperatura (Lehninger, 1981). Se podría esperar
entonces una correlación directa entre la actividad de enzimas activantes y
coeficientes positivos, y la de enzimas desactivantes y coeficientes negativos
de temperatura. En la abeja Apis mellifera, el efecto negativo de la temperatura
sobre Ia toxicidad de carbamatos desapareció al inhibir Ia actividad de enzimas
detoxicantes (Busvine, 1971). Exactamente lo contrario se observó en M.
domestica: la inhibición de enzimas detoxicantes hizo que el coeficiente
positivo se volviera neutro y en algunos casos ligeramente negativo (Fahmy
and Gordon, 1965).
El efecto neurotóxico de los piretroides también muestra una fuerte
dependencia de la temperatura. En P. amen'cana, el tratamiento con aletrina
produjoen los nervios cercales descargas espontáneas más prolongadas pero
de menor frecuencia a 15 que a 32°C (Gammon, 1978). En M. domestica, Ia
deltametrina fue más efectiva en bloquear la actividad neuromuscular y en
generar descargas repetitivas a baja temperatura (Salgado et al, 1983); y la
neuroactividad de la permetrina disminuyó al pasar de 15 a 30°C, y aumentó
de nuevo a 35°C (Ahn et al. 1987). En axones gigantes y cordones nerviosos
de P. americana tratados con distintos piretroides se observó actividad
repetitiva por encima de 26°C, pero no por debajo; un decaimiento más lento
de la depolarización a 15 que a 23°C; actividad repetitiva después de un
estímqu a 32 pero no a 15°C (Millerand Adams, 1982; Ruigt, 1985).
Los resultados de los experimentos de cambio de temperatura (Figuras
3.7. a 3.9.) demuestran que el efecto de la temperatura sobre Ia toxicidad de
deltametrina y B-cipermetrina en ninfas Ill de T. infestans revierte en tiempos
relativamente cortos. Esto sugiere que los cambios de temperatura están
alterando la interacción de las moléculas insecticidas con algún tipo de blanco
molecular. Si la disminución de la toxicidad al cambiar Ia temperatura tuviera
un origen metabólico, el fenómeno no sería reversible.

62
Sí puede tener una explicación metabólica la disminución del efecto de
la temperatura a través del tiempo (Figura 3.9.). En la primera parte de esta
Tesis se demostró que Ia toxicidad de deltametrina disminuye en función del
tiempo, y que esa disminución se anula cuando se inhibe la actividad de
oxidasas microsomales.
De todas maneras, no hay razones para pensar que uno sólo de los
pasos toxicológicos mencionados en los párrafos anteriores sea el único
responsable del efecto de la temperatura sobre Ia toxicidad de los piretroides.
Un paso puede predominar en determinadas circunstancias, otro en otras. Y
probablemente Ia respuesta final es el resultado de complejas interacciones.

63
3.2.1.3. Figuras
CE 50 [cm24 (ug ) 14
i:l 16°C Z
12 ¡:3 m: -‘12
— 1o
—' a
—- 6
—' 4
—' 2
' l 5,707 , 5/7 0deltametrina alfametrlna cipermetrina AC
B-cipermetrina cipennetrina
coeficiente detemperatura
i_r_isecticida (16-28°C)
deltametn'na - 5,4‘
B-cipermetn'na - 3,8"
alfametn'na - 7,8‘
cipermetn‘na - 5,7"
cipermetn'na AC - 4'4'
Figura 3.5. CE 50 (24 hs) de cianopiretroides a dos temperaturas en ninfas Ill.Losinsectos fueron expuestos a filmes sobre papel de filtrode los insecticidas. El efectoevaluado fue la incapacidad de abandonar un papel de filtro. Coeficiente detemperatura = CE 50 mayor / CE 50 menor. El signo negativo significa que CE 50(28°C) > CE 50 (16°C). Las líneas verticales representan los intervalos de confianzadel 95%. Las barras señaladas con las mismas letras no presentan diferenciasignificativa (SIC, P > 0,05). Los coeficientes señalados con un asterisco sonsignificativamente distintos de 1,0 (NSIC, P < 0,05). Cada valor se calculó con losdatos obtenidos en 3 réplicas independientes (N = 72 - 187).

DOC...00......000......000......OOOOOOOOOOOOOOOOOG
64
TE so (min)120 120
l: 16°C '. [ZZ] 28°C d '
100 1 100
E rL 1
ao - —80
60 — — 60" 1
4° - - 4o
20 - / - 20
19W
deltametrina alfametrlna
B-cipermetrina clpermetrina
' 0cipermetrina AC
coeficiente detemperatura
insecticida (16-28’0)
deltametr‘ina 1,7'
B-cipermetn'na 1,9‘
alfametn’na 1,4‘
cipennetrina 2,1"
cipennetn’na AC 1,4'
Figura 3.6. TE 50 de cianopiretroides a dos temperaturas en ninfas lll. Losinsectos fueron expuestos a filmes sobre papel de filtro de los insecticidas (55,5ug/cmz). El efecto evaluado fue la incapacidad de abandonar un papel de filtro.Coeficiente de temperatura = TE 50 mayor / TE 50 menor. Los valores positivossignifican que TE 50 (28°C) < TE 50 (16°C). Las líneas verticales representan losintervalos de confianza del 95%. Las barras señaladas con las mismas letras nopresentan diferencia significativa (SIC, P > 0,05). Los coeficientes señalados con unasterisco son significativamente distintos de 1.0 (NSIC. P < 0,05). Cada valor secalculó con los datos obtenidos en 3 réplicas independientes (n = 10).

65
día 0 1 2 o 7comienzo fin_. cambio deexposición expo' ICIÓn temperatura
í í z. 28°C ; 16°C í
TE 50 (horas)5 15
* l:l deltametrina c '— B-cipermetrina e
12 — - 12
’ 'l
9 a — 9
6 - b - 6l
_ Ï _
3 r- - 3
t a a
O / O
1 día después 6 dlas despuésde la exposición de la exposición
Figura 3.7. TE 50 al disminuir la temperatura ambiental en ninfas Illtratadas conpiretroides. Los insectos fueron expuestos a filmes sobre papel de filtro de losinsecticidas (14 ng/cmz). EI esquema encima del gráfico representa el diseñoexperimental. El efecto evaluado fue la incapacidad de abandonar un disco de papelde filtro. Las líneas verticales representan los intervalos de confianza del 95%. Lasbarras señaladas con las mismas letras no presentan diferencia significativa (SIC, P >0,05). Cada valor se calculó con los datos obtenidos en 2 réplicas independientes (n= 20).

66
2+2,5 h
v camgio de cagao ded'a 1temperatura temperatura
comienzo , x ,exposición exposicion \\ ,'
g s í}; ;: 28°C :‘: 28°C Í
16°CJ
TR 50 (min)0 a 50
_ :1 deltametn'na H_ B-clpermetrlna _
40 - a 40
3° “ a —30
20 - - 20
_ b
10 * - 10
- -l
o CL“,¡3/ n/ /, z, o
2,5 hs a 16°C 24 hs a 16°C
Figura 3.8. TR 50 al aumentar la temperatura ambiental de ninfas lll afectadaspor piretroides. Los insectos fueron expuestos a filmes sobre papel de filtro de losinsecticidas (14 ng/cmz). El esquema encima del gráfico representa el diseñoexperimental. El efecto evaluado fue la incapacidad de abandonar un disco de papelde filtro. Las lineas verticales representan los intervalos de confianza del 95%. Lasbarras señaladas con las mismas letras no presentan diferencia significativa (SIC, P >0,05). Cada valor se obtuvo con los datos obtenidos en 2 réplicas independientes (n= 20).

67
_ 2+2h _ 3+ hd. o 1 cambio cambio cambio cam I0¡a de temp. de temp.de temp.de temp.
comienzo fin d \\ ,' \ ,'exposmion expo' ¡Clon \\ (I t‘ l,
l l l l l l
; i Li4!: 28°C : E 28°C 5¡28°CÏ
100 100- O deltametrina
- y=95,s-113,5x ' r’= 0,97 '
8° h Ó b-cipermetrina 80
_ y = 54.9 - 83.8 x '_ r2 = 0,97
60 - - 60
4o L - 40
r.
20 L - 20
0 - - 0L- q
l l I l l l l l l l 4 l J 4L i l l
0 0,2 0,4 0.6
log tiempo (dias)
Figura 3.9. Variación del efecto de la disminución de la temperatura en funcióndel tiempo en ninfas IIItratadas con piretroides. Los insectos fueron expuestos afilmes secos sobre papel de filtro de los insecticidas (14 ng/cmz). El esquema encimadel gráfico representa el diseño experimental. El efecto evaluado fue la incapacidadde abandonar un papel de filtro de 15 cm de diámetro. Las líneas verticalesrepresentan la desviación estándar. Cada valor se calculó con los datos obtenidos en2 réplicas independientes (n = 20).
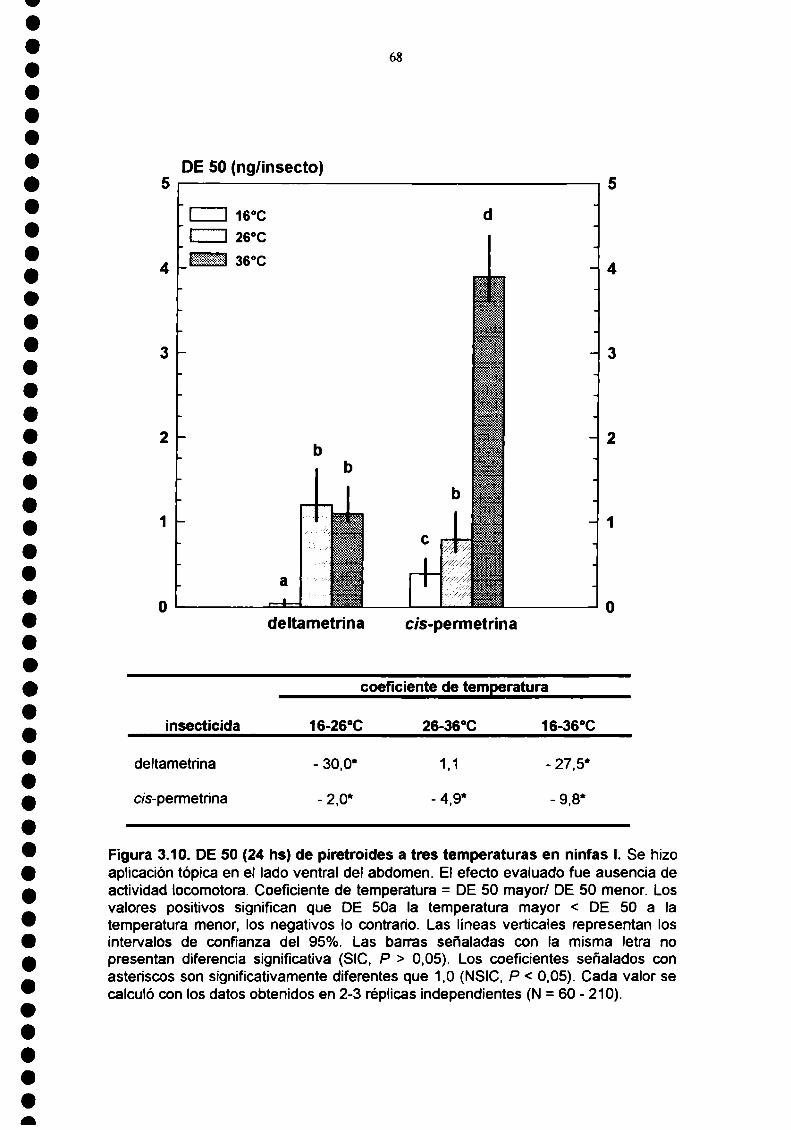
68
DE 50 (ng/insecto)5
:l:l 16°C d_E: 26°C
36°C j 4
3 ‘- _ 3
2 - - 2_ b _
b
r -i
1 - - 1’ 'l
. a _
o ' :L,;’/,/// o
deltametrina cis-pennetrina
coeficientede tempgratïrai
insecticida 16-26°C 26-36°C 16-36°C
deltametn’na - 30,0' 1,1 - 27,5‘
cis-permetrina - 2,0' - 4,9’ - 9,8‘
Figura 3.10. DE 50 (24 hs) de piretroides a tres temperaturas en ninfas l. Se hizoaplicación tópica en el lado ventral del abdomen. El efecto evaluado fue ausencia deactividad locomotora. Coeficiente de temperatura = DE 50 mayor/ DE 50 menor. Losvalores positivos significan que DE 50a la temperatura mayor < DE 50 a latemperatura menor, los negativos Io contrario. Las líneas verticales representan losintervalos de confianza del 95%. Las barras señaladas con la misma letra nopresentan diferencia significativa (SIC, P > 0,05). Los coeficientes señalados conasteriscos son significativamente diferentes que 1,0 (NSlC. P < 0,05). Cada valor secalculó con los datos obtenidos en 2-3 réplicas independientes (N = 60 - 210).

69
DE 50 (ng/insecto)0 450
’ b ¡:1 1s°c i400 — - 400
350 — ' - 350
300 — A 300
' ___ i250 — a 250
zoo — " ,Ï, — zoor- -i
150 L —150
100 e i a - 100
50 e
o ,
deltametrina cis-permetrina
coeficiente de temperatura
insecticida 16-26°C 26-36°C 16-36°C
deltametn’na - 6,8' 8,2' 1,2
cis-permetn'na - 1,3 - 10,0' - 13,1'
Figura 3.11. DE50 (24 hs) de piretroides a tres temperaturas en ninfas Ill.Se hizoaplicación tópica en el lado ventral del abdomen. El efecto evaluado fue ausencia deactividad locomotora. Coeficiente de temperatura = DE 50 mayor/ DE 50 menor. Losvalores positivos significan que DE 50 a la temperatura mayor < DE 50 a latemperatura menor, los negativos lo contrario. Las líneas verticales representan losintervalos de confianza del 95%. Las barras señaladas con la misma letra nopresentan diferencia significativa (SIC, P > 0,05). Los coeficientes señalados conasteriscos son significativamente diferentes que 1,0 (NSIC. P < 0,05). Cada valor secalculó con los datos obtenidos en 2-3 réplicas independientes (N = 80 - 132).

70
TE 50 (min)0
b [:1 16°C 160
deltametrina cis-pennetrlna
coeficiente de temperatura
insecticida 16-26°C 26-36°C 16-36°C
deltametn‘na 1,4“ 1,4' 2,0‘
cis-permetrina - 1,1 - 1,1 - 1,2
Figura 3.12. TE 50 de piretroides a tres temperaturas en ninfas I. Se hizoaplicación tópica en el lado ventral del abdomen (0,1 pgfinsecto). El efecto evaluadofue ausencia de actividad locomotora. Coeficiente de temperatura = TE 50 mayorlTE 50 menor. Los valores positivos significan que TE 50 a la temperatura mayor < TE50 a la temperatura menor, los negativos lo contrario. Las líneas verticalesrepresentan los intervalos de confianza del 95%. Las barras señaladas con la mismaletra no presentan diferencia significativa (SIC, P > 0,05). Los coeficientes señaladoscon asteriscos son significativamente diferentes que 1,0 (NSIC, P < 0,05). Cada valorse calculó con los datos obtenidos en 3 réplicas independientes (n = 10).

7l
5 TE 50 (min)l: 16°C26°C
25
deitametrina cis-permetrlna
coeficiente de temperatura
insecticida 16-26°C 26-36°C 16-36°C
deltametn'na - 1,6‘ 0,7' - 1.2
cis-permetrina - 1,3 - 1,0 - 1,3
Figura 3.13. TE 50 de piretroides a tres temperaturas en ninfas III. Se hizoaplicación tópica en el lado ventral del abdomen (20 pgfinsecto). El efecto evaluadofue ausencia de actividad locomotora. Coeficiente de temperatura = TE 50 mayorlTE 50 menor. Los valores positivos significan que TE 50 a la temperatura mayor < TE50 a la temperatura menor, los negativos lo contrario. Las líneas verticalesrepresentan los intervalos de confianza del 95%. Las barras señaladas con la mismaletra no presentan diferencia significativa (SIC, P > 0,05). Los coeficientes señaladoscon un asterisco son significativamente diferentes que 1,0 (NSlC, P < 0,05). Cadavalor se calculó con los datos obtenidos en 3 réplicas independientes (n = 10).

72
3. 2. 2. Composición de isómeros
3.2.2.1. Resultados
Se evaluó Ia toxicidad separada y conjunta de los isómeros cis y trans de
permetrina en ninfas I y lll. Se realizó aplicación tópica abdominal y el efecto
evaluado fue ausencia de actividad locomotora.
En ninfas I el isómero cis puro fue 2,9 veces más tóxico que el trans
puro (Figura 3.14.). La diferencia entre los respectivos valores de DE 50 fue
significativa (NSIC, P < 0,05). Se determinaron los Coeficientes de Interacción
(Ci) para 4 mezclas de isómeros, y ninguno fue significativamente distinto que
1,0 (Prueba t para una muestra, d.f.= 2, P > 0,05). Este resultado indica que la
mezcla se comportó en forma aditiva (ausencia de interacción).
En ninfas Ill el isómero cis puro fue 24,8 veces más tóxico que el
isómero trans puro (Figura 3.15.). La diferencia entre los valores de DE 50 fue
significativa (NSIC, P < 0,05). Se obtuvieron las DE 503 para 4 mezclas de
isómeros y se calcularon los respectivos Ci. Los Ci de todas las mezclas
fueron significativamente mayores que 1,0 (Prueba t para una muestra, g. l. =
2, P < 0,05). Esto significa que los isómeros actuaron en forma antagónica.
La Figura 3.16. muestra los valores de DE 50 de cis- y trans-pennetrina
a 16 y 36°C en ninfas l. A 16°C el isómero cis fue 1,3 veces más tóxico que el
trans, pero no hubo diferencia significativa entre las DE 508 (SIC, P > 0,05). A
36°C el isómero cis fue 1,8 veces más tóxico que el trans, y las DE 503 fueron
significativamente distintas (NSIC, P < 0,05). La DE 50 de trans-pennetrina
disminuyó significativamente al disminuir Ia temperatura (NSIC, P < 0,05), y los
coeficientes de temperatura tuvieron signo negativo.
La Figura 3.17. muestra los valores de DE 50 de cis- y trans-permetrina
a 16 y 36°C en ninfas III.A 16°C el isómero cis fue 33,3 veces más tóxico, y las
DE 505 fueron significativamente diferentes (NSIC, P < 0,05). A 36°C el
isómero cis fue 1,9 veces más tóxico, pero las DE 50s no fueron
significativamente diferentes (SIC, P > 0,05). La DE 50 de trans-permetrina no

73
varió significativamente con la temperatura (SIC, P > 0,05), y los coeficientes
de temperatura fueron neutros (= 1).
Para investigar una posible relación entre el metabolismo detoxicante y
las toxicidades diferenciales de los isómeros puros, éstos fueron aplicados en
ninfas previamente tratadas con los sinergistas butóxido de piperonilo
(inhibidor de la actividad de oxidasas microsomales) o trifenilfosfato (inhibidor
de la actividad de esterasas). Las ninfas estuvieron expuestas durante 24 hs a
filmes de los sinergistas sobre papel de filtro (0,7 mg/cm 2).
En ninfas l (Figura 3.18.) el pretratamiento con butóxido de piperonilo
redujo en forma significativa la DE 50 de la cis-permetrina (NSIC, P < 0,05),
pero no cambió en forma significativa la DE 50 de la trans-permetrina (SIC, P >
0.05). El pretratamiento con trifenilfosfato tuvo el efecto opuesto: redujo
significativamente Ia DE 50 de la trans-permetrina (NSIC, P < 0,05), pero no
cambió en forma significativa la DE 50 de la cis-permetrina (SIC, P > 0,05).
Estos resultados sugieren que en las ninfas I la actividad de oxidasas
microsomales estaría involucrada en el metabolismo del isómero cís, y Ia
actividad de esterasas en el metabolismo del isómero trans.
En ninfas Ill (Figura 3.19.) los pretratamientos con butóxido de
piperonilo y trifenilfosfato no produjeron cambios significativos en las DE 50 de
los isómeros (SIC, P > 0.05). Estos resultados sugieren que las actividades de
oxidasas microsomales y de esterasas no son relevantes en el metabolismo de
los isómeros de permetrina.
3.2.2.2. Discusión
Los efectos resultantes de la aplicación simultánea de dos o más sustancias
son llamados interacciones (Wilkinson, 1976). Si el efecto combinado de dos
sustancias es mayor que Ia suma de los efectos individuales, se trata de
sinergismo. Si el efecto combinado es menor que la suma de los efectos
individuales, es antagonismo. La aditividad es el caso en que el efecto
conjunto es igual a la suma de los efectos individuales, es decir cuando no hay

74
interacción. A veces se reserva el término sinergismo para el caso en que una
de las sustancias no es tóxica en Ia dosis usada, pero aumenta la toxicidad de
otra; y se usa potenciación cuando las dos sustancias son tóxicas en las dosis
usadas (Busvine, 1971). Otras veces, los términos sinergismo y potenciación
son usados en forma exactamente opuesta a la mencionada (Klaassen and
Eaton, 1991). Hay quien ha llegado a proponer 16 categorías para describir las
posibles interacciones entre dos sustancias (hay una revisión del tema en
Busvine, 1971). En esta Tesis se usa el término sinergismo para nombrar el
aumento de toxicidad de una sustancia producido por una dosis no tóxica de
otra. Se habría usado el término potenciación para nombrar el aumento de
toxicidad de una sustancia producido por una dosis tóxica de otra, pero no se
presentan casos de potenciación. Los términos antagonismo y aditividad se
usa en el mismo sentido en que todos los autores parecen estar de acuerdo:
cuando el efecto combinado de las sustancias es menor que Ia suma de los
efectos individuales y ausencia de interacción, respectivamente.
Como resultado de los métodos de síntesis, muchos piretroides de uso
comercial son una mezcla de isómeros ópticos y geométricos (ver el
Recuadro 1.1.). Algunos isómeros tienen actividad insecticida, otros no
(Naumann, 1990). La tendencia general entre los isómeros geométricos es que
la configuración cis es más tóxica que la trans (Naumann, 1990).
La toxicidad comparada de los isómeros cis y trans de la permetrina fue
estudiada en larvas de las mariposas Pseudoplusia inc/udens y H. virescens
(Dowd and Sparks, 1988; Naumann, 1990); y T. ni (Toth and Sparks, 1988); la
mosca M. domestica; el mosquito Culex quinquefasciatus quinquefasciatus; y la
cucaracha B. germanica) (Ruigt, 1985; Naumann, 1990). En estos insectos el
isómero cis fue entre 1,7 y 7,1 veces más tóxico que el trans. Los resultados
presentados aquí (Figuras 3.14. y 3.15.) coinciden con los antecedentes
citados: en ninfas I y III de T. infestans el isómero cis fue más tóxico que el
trans. Pero en ninfas III Ia diferencia en la toxicidad de los isómeros fue
inesperadamente alta.

75
Se conocen casos donde la diferencia en las toxicidades de los
isómeros cis y trans de la permetrina tiene una explicación metabólica. La tasa
de metabolismo de trans-permetrina fue mayor que la de cís-permetrina en M.
domestica (Shono et al. 1979), S. littoralisy T. ni (Ishaaya and Casida, 1983), y
P. includens y H. virescens (Dowd and Sparks, 1988).
Inhibidores de actividades enzimáticas detoxicantes inhibieron la
degradación de trans-permetrina ¡n vivo en S. littoralis y T. ni (Ishaaya and
Casida, 1983) e in vitro en homogenatos de P. includens (Dowd and Sparks,
1987). Los experimentos con sinergistas (Figuras 3.18 y 3.19.) revelaron que
en ninfas l de T. infestans la actividad de oxidasas microsomales esterasas
tiene relevancia en el metabolismo de cis-permetrina, y la actividad de
esterasas en el metabolismo de trans-permetrina. En ninfas III Ia toxicidad de
los isómeros no fue modificado por el tratamiento con sinergistas. Este
sinergismo diferencial en distintos estadios ninfales puede deberse a
variaciones en los títulos de enzimas detoxicantes a lo largo del desarrollo. Se
encontró este tipo de variaciones en la actividad hidrolítica hacia trans
permetrina en P. includens (Dowd and Sparks, 1984), y hacia permetrina en S.
frugiperda (Yu, 1983).
La ausencia de sinergismo en ninfas III sugiere que Ia actividad de
esterasas y de oxidasas microsomales no es importante en el metabolismo de
los isómeros de permetrina. Otras vías metabólicas, no inhibibles por los
sinergistas usados, e incluso otros pasos toxicocinéticos pueden ser
responsables de la toxicidad diferencial de estos insecticidas.
El análisis de las DE 503 de las mezclas de cis- y trans-permetrina
(Figuras 3.14. y 3.15.) indicó aditividad en ninfas I, y antagonismo en ninfas Ill.
En este último estadio, la relativamente baja toxicidad del isómero trans y el
efecto antagonístico de Ia aplicación conjunta enmascaran Ia alta toxicidad del
isómero cis. Como se mostró antes (Figura 3.4.), la cis-permetrina tiene en
ninfas lll una toxicidad comparable a la de deltametrina. Este resultado es
inusual: los cianopiretroides (como deltametrina) suelen ser más tóxicos que
los no ciano (como Ia cis-permetrina) (Elliot et al, 1974; Ruigt, 1985). Además,

76
deltametrina es uno de los piretroides más efectivos en vinchuca (Casabé et
al, 1988).
Cuáles son los isómeros de los piretroides con baja toxicidad y qué tipo
de interacciones ocurren en las mezclas son datos que deberían ser tenidos
en cuenta al programar el uso de insecticidas. Cuando se dispone de este
concimiento es posible por ejemplo evitar la aplicación inútil de moléculas sin
actividad o que disminuyen Ia toxicidad de otras.

77
3.2.2.3. Figuras
DE50 (ng/insecto)3
C
_ c
2 — - 2
l- b un
. a a _
1 — - 1
0100: 0 82: 18 60:40 42: 58 24: 76 0: 100
cls: trans
coeficiente de interaccióncis: trans (promedio :t DE)
82: 18 0,9 :l:0,1
60: 40 0.9 i 0,2
42: 58 1,0 :t 0,1
24: 76 2,0 :l:1,3
Figura 3.14. DE 50 (24 hs) a 26°C de ¡sómeros de pennetrina puros y mezcladosen ninfas l. Se hizo aplicación tópica en el lado ventral del abdomen. El efectoevaluado fue ausencia de actividad locomotora. Las líneas verticales representan losintervalos de confianza del 95%. Las banas señaladas con la misma letra nopresentan diferencia significativa (SIC, P > 0,05). Ningún coeficiente essignificativamente distinto que 1,0 (= ausencia de interacción) (Pmeba de t para unamuestra, P > 0,05). Cada valor se calculó con los datos obtenidos en 2-3 réplicasindependientes (N = 69 - 140).

78
DE 50 (ng/insecto)
225 d 225
200 - —200
175 — d - 175
150 — d _ — 150
125 F — 125i.
100 b — 100
75 - — 75
50 F c — 50
25 r b W —25. a I .
0 I l 0100: 0 82: 18 60:40 42: 58 24: 76 0: 100
cís: trans
coeficiente de interaccióncis: trans (promedio :i:DE)
82:18 1.710,1'
60: 40 3,0 :t 0,6'
42: 58 8,711,1'
24: 76 8,8 11,0'
Figura 3.15. DE50 (24 hs) a 26°C de isómeros de pennetrina puros y mezcladosen ninfas III. Se hizo aplicación tópica en el lado ventral del abdomen. El efectoevaluado fue ausencia de actividad locomotora. Las líneas verticales representan losintervalos de confianza del 95%. Las banas señaladas con la misma letra nopresentan diferencia significativa (SIC, P > 0,05). Los coeficientes señalados con unasterisco son significativamente distintos que 1,0 (coeficiente de interacción > 1,0significa antagonismo) (Prueba t para una muestra, P < 0,05). Cada valor se calculócon los datos obtenidos en 2-3 replicas independientes (59 - 121).

79
8 DE 50 (ng/insecto) 8
Z 16°C :7 - 36°C - 7
6 ¡- __ 6
5 -_ L 5
4 L —_4
: j3 — - 3
2 l 3 2
1 _- a 1 1
_ _ W 7
o - W //////% ._ ocIs-pennetrlna trans-permetrlna
coeficiente de temperaturarango de temperaïra (°C) para trans-pennetrina
16-26 - 4.6'
26-36 - 3,0‘
16-36 -13.8‘
Figura 3. 16. DE 50 (24 hs) de cis- y trans-permetn'na a 16 y 36°C en ninfas l. Sehizo aplicación tópica en el lado ventral del abdomen. El efecto evaluado fueausencia de actividad locomotora. Coeficiente de temperatura = DE 50 mayor/ DE 50menor. El signo negativo significa que la DE 50 a la mayor temperatura > DE 50 a lamenor temperatura. Los coeficientes para los rangos 16-26°C y 26-36°C secalcularon con datos de la Figura 3.14. Las líneas verticales representan losintervalos de confianza del 95%. Las banas señaladas con las mismas letras nopresentan diferencia significativa (SIC. P > 0,05). Los coeficientes señalados conasten‘scos son significativamente diferentes que 1,0 (NSlC, P < 0,05). Cada valor secalculó con los datos obtenidos en 3 réplicas independientes (N = 101 - 210).

so
DE 50 (ngllnsecto)0 180
c b c- [:1 16°C —
150 L 36°C 1 150
cis-permetrlna tran -permetrlna
coeficiente de temperaturaMo de temperaïtura(°C) para trans-permetrina
16-26 1.0
26-36 1,3
16-36 1,4
Figura 3.17. DE 50 de cls- y trans-pemetrina a 16 y 36°C en ninfas lll. Se hizoaplicación tópica en el lado ventral del abdomen. El efecto evaluado fue ausenciade actividad locomotora. Coeficiente de temperatura = DE 50 mayor/ DE 50 menor.El signo positivo significa que la DE 50 a la mayor temperatura < DE 50 a Ia menortemperatura. Los coeficientes para los rangos 16-26°C y 26-36°C se calcularon condatos de la Figura 3.15. Las líneas verticales representan los intervalos deconfianza del 95%. Las barras señaladas con las mismas letras no presentandiferencia significativa (SIC, P > 0,05). Ningún Coeficiente de Temperatura fuesignificativamente distinto que 1,0 (SIC, P > 0,05). Cada valor se calculó con losdatos obtenidos en 3 réplicas independientes (N = 101 - 124).

8]
DE 50 (ng/insecto)
|:l sin tratamientoprevio/ ” ' tratamiento previo: BP
V - tratamiento previo: TF
Figura 3.18. DE 50 (24 hs) de isómeros de permetrina con y sin sinergistas enninfas I. Los insectos fueron expuestos durante 24 horas a filmes sobre papel defiltro de los sinergistas (0,7 mg/cmz). Entonces se hizo aplicación tópica de losinsecticidas en el lado ventral del abdomen. BP: butóxido de piperonilo; TF:trifenilfosfato. Las líneas verticales representan los intervalos de confianza del 95%.Las barras señaladas con las mismas letras no presentan diferencia significativa(SIC, P > 0,05). Cada valor se calculó con los datos obtenidos en 2-3 réplicasindependientes (N = 50 - 80).

82
o DE 50 (ng/lnsecto) 250|:] sintratamientoprevio
’ tratamiento previo: BP _
tratamiento previo: TF — zoo
L 1
150 - r - 150
1oon /1oo/ /
25 - - 25
l a 1a a
o IM 2 ocis-perlna tmns-pennetrlna
Figura 3.19. DE 50 (24 hs) de isómeros de permetn'na con y sin sinergistas enninfas lll. Los insectos fueron expuestos durante 24 horas a filmes sobre papel defiltro de los sinergistas (0,7 mg/cmz). Entonces se hizo aplicación tópica de losinsecticidas en el lado ventral del abdomen. BP: butóxido de piperonilo; TF:tn’fenilfosfato.Las líneas verticales representan los intervalos de confianza del 95%.Las barras señaladas con las mismas letras no presentan diferencia significativa(SIC, P > 0,05). Cada valor se calculó con los datos obtenidos en 2-3 réplicasindependientes (N = 64 - 145).

83
3. 2. 3. Edad y estado alimentario
3.2.3.1. Resultados
Se determinaron las DE 50s de deltametrina y cis-permetrina en ninfas l de
distintas edades y estados nuticionales. Se hizo aplicación tópica y el efecto
evaluado fue Ia ausencia de actividad locomotora.
La Figura 3.20.a. muestra los valores de DE 505 para deltametrina. No
hubo diferencia significativa entre las DE 508 de ninfas ayunadas de 7 y 21
días de edad (SIC, P > 0,05), pero la DE 50 de ninfas ayunadas de 1 día de
edad fue significativamente menor que ambas (NSIC, P < 0,05). Las ninfas de
21 días de edad alimentadas 1 hora antes del tratamiento fueron
significativamente más susceptibles que las alimentadas 7 días antes (NSIC, P
< 0,05). A una misma edad (21 días), las ninfas ayunadas fueron
significativamente más susceptibles que las alimentadas 7 días antes (NSIC, P
< 0,05), pero su DE 50 no fue significativamente diferente que la de las ninfas
alimentadas 1 hora antes (SIC, P > 0,05). El perfil de toxicidad se mantuvo al
expresar los datos en ng/mg.
La Figura 3.20.b. muestra los valores de DE 50 para cis-permetrina. No
hubo diferencia significativa entre las DE 505 en ninfas ayunadas de 1 y 21
días de edad (SIC, P > 0,05), pero la DE 50 de ninfas ayunadas de 7 días de
edad fue significativamente mayor que ambas (NSIC, P < 0,05). Igual que para
deltametrina, las ninfas de 21 días de edad alimentadas 1 hora antes del
tratamiento fueron significativamente más susceptibles que las alimentadas 7
días antes (NSIC, P < 0,05). A una misma edad (21 días), las ninfas ayunadas
fueron significativamente más susceptibles que las alimentadas (NSIC. P <
0,05). El perfil de toxicidad se mantuvo al expresar los datos en ng/mg.

84
3.2.3.2. Discusión
Las diferencias en Ia toxicidad de deltametrina y cis-permetrina en ninfas I de
distintas edades y estados nutricionales pueden ser atribuidas a variaciones
en Ia actividad de enzimas detoxicantes y a las características de Ia cutícula.
La variación de la actividad de oxidasas microsomales en función de Ia
edad y el desarrollo fue demostrada en varias especies de insectos (revisión
del tema en Agosin, 1985). En ninfas V de T. infestans de distintas edades se
encontró una correlación entre la variación de Ia toxicidad del malatión y la
actividad de enzimas detoxicantes (Wood et al, 1993). En el último estadio
larval de la mariposa P. ¡nc/udens la actividad de esterasas que hidrolizaban
trans-permetrina varió en función de Ia edad, distintos tejidos mostraron
distintos patrones de variación y los valores de DL 50 a distintos tiempos
reflejaron los cambios en la actividad hidrolítica (Dowd and Sparks, 1984).
Durante Ia ingesta la cutícula de los insectos hematófagos se modifica
química y físicamente, y adquiere propiedades elásticas que permiten su
estiramiento (Hackman and Goldberg, 1987). La cutícula abdominal de ninfas
V de R. pro/¡xus se adelgazó considerablemente como consecuencia del
estiramiento, y su peso seco se modifica en función del tiempo posterior a la
ingesta (Hillerton, 1978). No se demostró con exactitud Ia naturaleza de los
cambios químicos que acompañan el estiramiento, pero se sugirieron
variaciones en el pH (Reynolds, 1975) y en el grado de hidratación (Maddrell,
1966). Se encontró que la velocidad de penetración de los insecticidas
depende de la composición y las propiedades físicas del integumento (Ebeling,
1974; Fontán and Zerba, 1987).
La relación entre toxicidad y estado nutricional de la cis-permetrina
presentada aquí, fue similar a la encontrada para el DDT en ninfas II de T.
infestans (Fontán and Zerba, 1992). En ese trabajo se demostró que la
velocidad de penetración del DDT era mucho más lenta en ninfas ayunadas
que en ninfas alimentadas 1 hora antes, y que este aumento en la velocidad de

85
penetración estaba correlacionado con un aumento de la actividad insecticida.
La mayor permeabilidad al DDT en las ninfas alimentadas fue atribuida a los
cambios que cuticulares que acompañan el estiramiento.
Varios autores encontraron que los insectos son más susceptibles a la
acción de los insecticidas inmediatamente después de la muda, y más
tolerantes inmediatamente antes (Busvine, 1971). Estas diferencias en Ia
toxicidad fueron atribuidas a cambios en el grosor de Ia cutícula. En el
presente trabajo las ninfas de T. infestans de un día de edad fueron las más
susceptibles a los dos piretroides. Veinticuatro horas después de la eclosión la
esclerotización de la cutícula es todavía incompleta en este estadio ninfal, a
juzgar por su coloración. La cutícula semiesclerotizada puede ser una barrera
más fácilmente penetrable para los insecticidas que la cutícula totalmente
esclerotizada.
La alta tolerancia de las ninfas alimentadas 7 días antes puede estar
relacionada con los cambios fisiológicos previos a la muda (Agrell and
Lundquist, 1973; Gilbert and King, 1973). Las ninfas l de T. infestans
comienzan a mudar unos 11 días después de la ingesta (Alzogaray, datos no
publicados). La nueva cutícula en formación puede constituir además una
barrera mecánica al ingreso de los insecticidas.
Los valores de DE 503 pueden ser expresados por unidad anatómica
(por ejemplo ng/insecto) o por unidad de peso corporal (por ejemplo ng/mg).
En general se considera que ambas formas de expresión son
aproximadamente equivalentes (Klaassen and Eaton, 1991), pero esto
depende de la homogeneidad del peso corporal (Busvine, 1971). Los
resultados de la Figura 3.20 indican que en ninfas IIIde T. infestans el perfil
de toxicidad de deltametrina y cis-perrnetrina fue eI mismo expresando los
datos en ng/insecto y ng/mg.

DOC...00......OOO...OOOOOOOOOOOOOÓOOOOOOOOOOOOOOOG
86
3.2.3.2. Figuras
(a)
DE 50
c 7:1 ayunadas' (edad: 1 día)_ ayunadas _
6 (edad: 7 días) 6e ayunadas
(edad: 21 días)5 ' - alimentadas- 5
1hora antes' (edad: 21 días)
4 _ ¡:1 alimentadas _ 47 dias antes- (edad: 21 dias)
3 e “ 3
2 .. - 2
_ b b .1 — b — 1
0 ‘ 0
nglinsecto nglmg
Figura 3.20. DE 50 (24 hs) de piretroides en ninfas I de distintas edades yestados alimentarios. Se hizo aplicación tópica en el lado ventral del abdomen de(a) deltametrina o (b) cis-permetn'na. EI efecto evaluado fue ausencia de actividadlocomotora. Las líneas verticales representan los intervalos de confianza del 95%.Dentro de cada grupo, las barras señaladas con Ia misma letra no presentandiferencia significativa (SIC, P > 0,05). Cada valor se calculó con los datos obtenidosen 2-3 réplicas independientes (N = 46 - 142).

87
(b)
DE 50
[:1 ayunadas(edad: 1 día)
/ "7 ayunadas(edad: 7 días)
ayunadas (edad: 21 días) _
- alimentadas1hora antes(edad: 21 dias) ‘
[:1 alimentadas7 días antes (edad: 21 días) .
-(
o unglinsecto
Figura 3.20. (Continuación)

88
3. 2. 4. Dosis y lugar de aplicación
3.2.4.1. ReSuH'ados
Se evaluaron los TE 505 de distintas dosis de deltametrina y cis-permetrina en
ninfas l, y los TE 505 de una misma dosis (20 nglinsecto) aplicada en distintos
lugares del cuerpo. En ambos casos se hizo aplicación tópica en el lado
ventral del abdomen, y el efecto evaluado fue la incapacidad de abandonar un
disco de papel de filtro.
La Figura 3.21. muestra Ia variación de los TE 50s en función de Ia
dosis. Se usaron dosis comprendidas en el rango 2 - 1.000 nglinsecto. Los TE
503 de deltametrina fueron significativamente menores que los de cis
permetrina en todas las dosis aplicadas (NSlC, P < 0,05), y dejaron de
disminuir en forma significativa a partir de los 20 nglinsecto. EI TE 50 de cis
perrnetrina decreció en todo el rango de dosis estudiado.
Los resultados de la Figura 3.22 demuestran que el TE 50 depende del
lugar de aplicación. El efecto tóxico se manifestó significativamente más rápido
cuando los insecticidas fueron aplicados en el tórax (entre el segundo y el
tercer par de patas) que cuando fueron aplicados en los últimos terguitos
(NSlC, P < 0,05). En los dos lugares, la deltametrina actuó unas dos veces
más rápido que la cis-permetrina.
3.2.4.2. Discusión
En 3.1. Síntomas de intoxicación se demostró que las DE 505 de
deltametrina y cis-permetrina no presentaron diferencia significativas para el
efecto incapacidad de abandonar un disco de papel de filtro (Figura 3.4.). Las
diferencias en los TE 503 de estos dos piretroides deben originarse entonces
en diferencias en los pasos toxicocinéticos. Los datos presentados son
insuficientes para determinar en cuál o cuáles pasos toxicocinéticos radican

89
esas diferencias (penetración, distribución, acumulación, acceso al blanco,
metabolismo).
T. ¡nfestans tiene dos ganglios en el tórax y ninguno en el abdomen
(lnsausti, 1994). De los ganglios torácicos parten los nervios cuyas
ramificaciones inervan los músculos de las patas. La aplicación de los
insecticidas en el lado ventral del tórax, es decir sobre los nervios que inervan
los músculos locomotores, tuvo un efecto más rápido que Ia aplicación en el
extremo dorsal del abdomen. La interpretación más simple del efecto del sitio
de aplicación sobre los TE 503 es que cuánto más lejos del sitio de acción son
aplicados los insecticidas, más tardan en manifestarse los efectos tóxicos.
Todavia no está claro si el efecto sobre el sistema nervioso central es el
responsable del volteo, pero sí está ampliamente demostrada la acción de los
piretroides sobre el sistema nervioso central de los insectos (hay revisiones en
Ruigt, 1985; Naumann, 1990). En sistema nervioso aislado de cucaracha (P.
americana) bastó una concentración 0,1 nM de piretrina I para causar una
violenta actividad espontánea en el sexto ganglio abdominal (Burt and
Goodchild, 1971). Efectos sobre el cordón nervioso se manifestaron con
concentraciones al menos 3 órdenes de magnitud mayores. Se encontró una
correlación entre el efecto a nivel ganglionar y los síntomas de intoxicación.
Otros trabajos sugirieron que el cordón nervioso no es un sitio crítico para Ia
acción de piretroides (Burt and Goodchild, 1974; Gammon, 1978).
La variación de los TE 505 en función del lugar de aplicación puede
tener una explicación más compleja y diferentes pasos de Ia toxicocinética
pueden estar influenciandoestos cambios. En 3.3. Efecto sobre la actividad
locomotora se discutiránotros aspectos de este tema.

90
3.2.4.3. Figuras
TE50(mnn14o _ 14o
120 '_ + deltametrina _“120Z . ' I ' cís-permetrina Z
100 3 “ { 1ool ‘ íl.
ao Ï - ao
so 3 { so
4o } 3 4o
20 l J 20- -l
o I- 41 4L l l l l l l l l l l l l J l ll l l l l l l l ll o
1 1o 100 1ooo
dosis (ng/lnsecto)
Figura 3.21. Variación del TE 50 en función de la dosis en ninfas I. Se hizoaplicación tópica en el lado ventral del abdomen. El efecto evaluado fue incapacidadde abandonar un disco de papel de filtro. Las líneas verticales representan losintervalos de confianza del 95%. Se consideran significativamente distintos losvalores cuyos intervalos de confianza no se superponen (P < 0,05). Cada valor secalculó con los datos obtenidos en 3 réplicas independientes (n = 10).

91
TE 50 (min)120 120
: b :1 deltametrina 7_ í cls-permetrina _
100 - — 100
ao - - 80
60 L 60
4o — 40
20 — 20
o 0aplicación aplicaclón
en el abdomen en el tóráx
Figura 3. 22. TE 50 de piretroides aplicados en el abdomen y en el tórax deninfas lll. Se hizo aplicación tópica (20 ngfinsecto) sobre los últimos terguitos o en ellado ventral del tórax (entre el primer y el segundo par de patas). El efecto evaluadofue la incapacidad de abandonar un disco de papel de filtro. Las líneas verticalesrepresentan los intervalos de confianza del 95%. Las banas señaladas con lasmismas letras no presentan diferencia significativa (SIC, P > 0.05). Cada valor secalculó con los datos obtenidos en 4 réplicas independientes (n = 5).

92
3. 3. EFECTO SOBRE LA ACTIVIDADLOCOMOTORA
3. 3. 1. Resultados
Se evaluó con un analizador de imágenes el efecto de deltametrina y cis
permetrina sobre Ia actividad locomotora de ninfas Ill. Se estudiaron las
influencias del modo de aplicación de los insecticidas, el pretratamiento con N
etilmaleimida y la temperatura sobre dicho efecto.
La Figura 3.23. muestra cómo la actividad locomotora de ninfas
expuestas a filmes de piretroides aumentó en forma lineal en función del
logaritmo de Ia concentración (deltametrina, y = 2210,? + 1280,? x, r2 = 0,92;
cis-permetrina, y = - 498,7 + 1479,4 x, r2 = 0,99). La Tabla 3.4. muestra la
toxicidad para cada insecticida de las concentraciones usadas para construir
las rectas de regresión. Concentraciones subtóxicas de los dos piretroides
aumentaron la actividad locomotora de las ninfas en forma significativa
(Prueba de Duncan, P < 0,05),
Cuando se hizo aplicación tópica de los piretroides en el extremo dorsal
del abdomen de las ninfas (Figura 3.24.a.), la deltametrina produjo un
aumento significativo de la actividad locomotora (Anova; F = 4,3; g. l. = 3, 16; P
< 0,05), pero al intentar estimar una recta de regresión se encontró que su
pendiente no era significativamente distinta de cero (Anova; F = 69,0; g. I. = 1,
2; P > 0,05). La actividad locomotora no fue afectada en forma significativa por
dosis subtóxicas de deltametrina (Prueba de Duncan, P > 0,05) (Tabla 3.5.).
Ninguna de las dosis de cis-permetrina aplicadas afectó significativamente la
actividad locomotora (Figura 3.24.a.) (Anova, F = 2,8; g. I. = 3, 19; P> 0,05).
Cuando se hizo aplicación tópica en la cabeza de las ninfas (Figura
3.24.b.), hubo diferencia significativa entre los efectos de los distintos
tratamientos (Anova; F = 7,2; g. l. = 2, 12; P < 0,01), y ambos piretroides
aumentaron en forma significativa la actividad locomotora (la misma dosis de
cis-permetrina aplicada en el extremo dorsal del abdomen de las ninfas no
había modificado significativamente dicha actividad).

93
La Figura 3.25. muestra cómo el pretratamiento tópico con N
etilmaleimida (20 ug/insecto) modificó el efecto de los piretroides sobre Ia
actividad locomotora. En los insectos pretratados con N-etilmaleimida, ninguno
de los dos piretroides produjo cambios significativos en la actividad locomotora
(Anova; F = 1,3; g. l. = 2, 12; P > 0,05). Las mismas concentraciones de cada
insecticida produjeron un efecto significativo en los insectos pretratados con
acetona sola (Anova; F = 14,0; g. l. = 2, 12; P < 0,001).
La Figura 3.26. muestra el efecto de los piretroides sobre la actividad
locomotora a dos temperaturas. A 16°C ninguno de los insecticidas afectó
significativamente la actividad locomotora (Anova; F = 0,7; g. I. = 2, 9; P >
0,05); a 36°C ambos la afectaron en forma significativa (Anova; F = 20,8; g. l. =
2, 9; P < 0,0005).
3. 3.2. Discusión
En 3.1. Síntomas de intoxicación se mencionó el aumento de la actividad
locomotora (hiperactividad) como el primer síntoma de la intoxicación con
piretroides en insectos. Este efecto es responsable del fenómeno de expurgue,
es decir la capacidad de hacer que los insectos abandonen sus refugios
(Pinchin et al, 1980; Wood et al., 1993). El expurgue tiene al menos dos usos
prácticos: (a) la aplicación simultánea de una sustancia expurgante y una
sustancia tóxica, para que los insectos abandonen sus refugios y se expongan
a la acción insecticida (en el caso de los piretroides, una misma sustancia
puede cumplir las dos funciones) (Gualtieri et al, 1985); (b) como herramienta
para muestrear poblaciones de insectos antes y después de un tratamiento
insecticida, y así evaluar la eficacia del tratamiento (WHO, 1991; Oliveira
Filho, 1994).
Los resultados de las Figuras 3.23 y 3.24. indican que la hiperactividad
producida por deltametrina y cis-permetrina en ninfas III de T. infestans fue

94
mayor cuando se hizo exposición a filme que cuando se hizo aplicación tópica;
y Ia aplicación tópica en la cabeza produjo un efecto mayor que Ia aplicación
tópica en el extremo del abdomen. Como se discutió previamente (3.2.4. Dosis
y lugar de aplicación.) la variación de la toxicidad en función del lugar de
aplicación puede estar reflejando la distancia al sitio de acción.
Estudios de comportamiento, electrofisiología y bioquímica demostraron
que los grupos sulfhidrilos intervienen en los mecanismos de quimiorrecepción
de los insectos. En estos trabajos, distintos reactivos de sulfhidrilos (N
etilmaleimida, p-cloromercuribenzoato, azoester, ácido iodoacético, 1,4
ditiotreitol) bloquearon los comportamientos alimentario en insectos
hematófagos (Galun et al, 1969) y en la mosca Boettchen'sca peregn’na
(Koyama and Kurihara, 1971), y el de agregación en la cucaracha B.
germanica (Koyama and Kurihara, 1971), disparados por estímulos químicos;
inhibieron las respuestas electrofisiológicas en quimiorreceptores de antenas
de las mariposas B. mory y Antheraea pernyi (Villet, 1974) y de boca de la
mosca B. peregrina (Shimada et al. 1972); y compitieron por
quimiorreceptores antenales de quinonas de Ia cucaracha P. amen'cana (Norris
et aI., 1971; Singer et aI., 1975). En T. infestans se obtuvo evidencia de
efectos antialimentario y anticópula producidos por el tratamiento con N
etilmaleimida (Picollo et al, 1993). Estos efectos fueron atribuidos a un
bloqueo de la quimiorrecepción.
En este trabajo se encontró que la N-etilmaleimida inhibe Ia
hiperactividad producida por los piretroides (Figura 3.25.). Este resultado
apoya la posibilidad de que Ia hiperactividad inducida por deltametrina y cis
permetrina está asociada con un mecanismo de quimiorrecepción.
Los órganos sensoriales son muy susceptibles a la acción de los
piretroides (Ruigt, 1985). Se encontró que diversos piretroides generaron
potenciales de acción en el nervio cruraI de Ia langosta Schistocerca gregan'a,
no asociados con actividad muscular (Clements and May, 1977). La actividad
nerviosa no se manifestó en individuos a los que previamente se les extirpó el

95
órgano cordotonal (el mayor órgano sensorial en las patas de la langosta). Se
observó actividad repetitiva provocada por piretroides en quimiorreceptores de
la mosca Phormia regina (Holan et al., 1978). La tetrametrina, en Ia baja
concentración de 0,3 pM, causó actividad repetitiva en el nervio sensorial
cercal de la cucaracha P. americana (Gammon et al, 1981). En experimentos
con cucarachas P. americana vivas y libres, tratadas con aletrina, se
estudiaron simultáneamente los síntomas de intoxicación y los efectos
neurotóxicos (Gammon, 1978). Durante la fase de excitación se detectó
actividad aferente espontánea en el nervio cercal, pero no en el cordón
nervioso abdominal ni en nervios cercales eferentes. Estos resultados hicieron
pensar que el aumento de la actividad locomotora inducido por el piretroide
podía ser consecuencia de un efecto sobre el sistema nervioso periférico.
Podía estar ocurriendo por ejemplo un bloqueo de la inervación eferente
(inhibitoria).
El efecto de Ia temperatura sobre la acción de los piretroides se discutió
en Ia 3.2.1. Temperatura. Con respecto a la quimiorrecepción, la respuesta
electrofisiológica de quimiorreceptores bucales de mosca (P. regina) disminuyó
hasta un 80% cuando Ia temperatura descendió de 28 a 12°C (Uehara and
Morita, 1972). En Drosophila melanogaster el comportamiento alimentario
mediado por quimiorreceptores tarsales disminuyó al disminuir la temperatura
(Napolitano et al, 1986).
Una misma concentración de los piretroides estudiados produjo
hiperactividad en ninfas lll de T. infestans a 26 y 36°C, pero no a 16°C (Figura
3. 26.). Este efecto de la temperatura es opuesto al efecto sobre otros
síntomas de intoxicación presentados en esta Tesis. En las Figuras 3.5. y
3.11. se observa que deltametrina y cis-permetrina fueron significativamente
más tóxicas a 16 que a 26°C en ninfas Ill. Estos resultados sugieren que los
mecanismos responsables de la hiperactividad y la incoordinación son
distintos. Esta es una cuestión que todavía no fue resuelta. Se propuso que el
volteo resulta de un exceso de estimulación producido por las piretrinas y sus

96
derivados sintéticos sobre los nervios sensoriales superficiales (Camougis,
1973; Clements and May, 1977; Gammon, 1981). Esto interrumpiría el flujo de
información hacia el sistema nervioso central y los insectos intoxicados serían
incapaces de coordinar los movimientos de las patas. Sin embargo, otros
autores no encontraron efectos neurotóxicos en células sensoriales, y
propusieron que el sitio primario de acción para el volteo es el sistema
nervioso central (Ruigt, 1985).
Los datos obtenidos son evidencias indirectas de una posible
interacción entre los piretroides y el sistema nervioso periférico a nivel de
quimiorreceptores. Habría que realizar experimentos de electrofisiología para
obtener una evidencia más contundente.
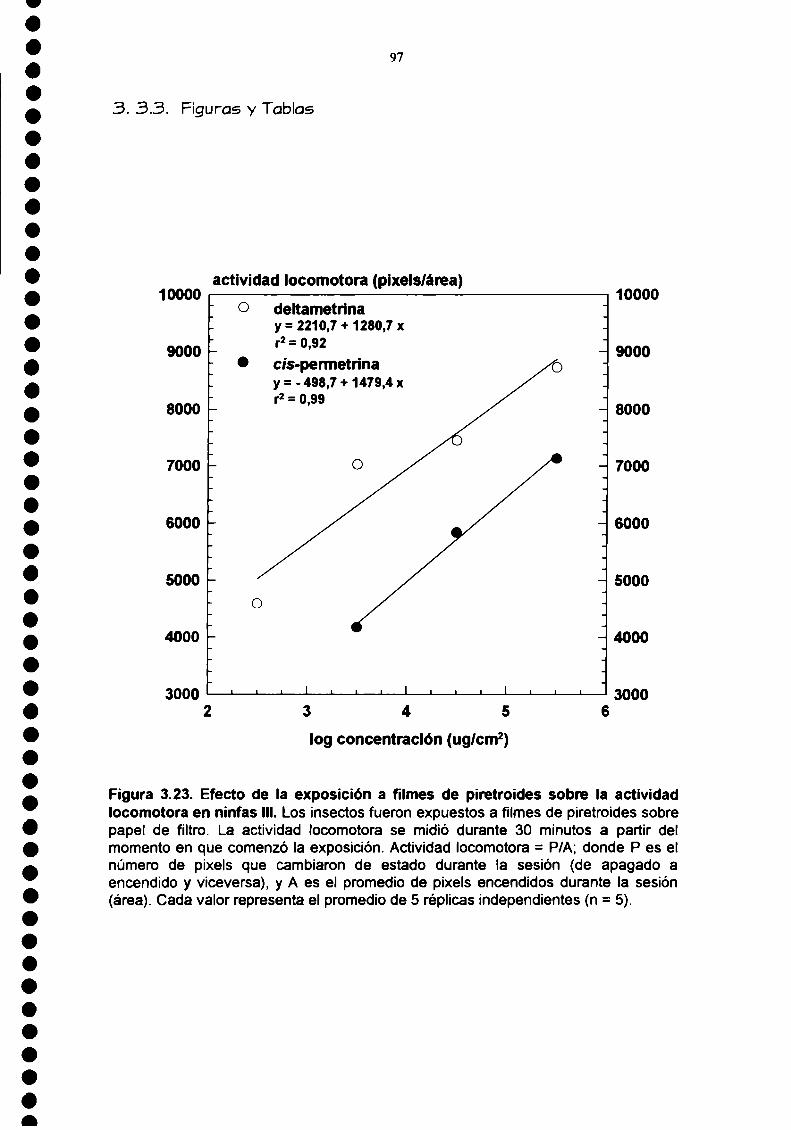
97
3. 3.3. Figuras y Tablas
actividad locomotora (pixels/area)10000 10000
O deltametrina j_ y = 2210.7 + 1280,7 x i- 2 =
9000 — '_ “2 —9000z 0 els-pennetrina o j- y = - 498,7 + 1479,4 x _ 2 =
aooo _— ' °'9° —_aooo
7000 l 7ooo
E I
6000 5 —_6000
5000 -' sooo: O '
4000 í 4000
_ 1 l 1 L L L J l l l l I l l l2 3 4 5 6
log concentración (ug/cm!)
Figura 3.23. Efecto de la exposición a filmes de piretroides sobre la actividadlocomotora en ninfas lll. Los insectos fueron expuestos a filmes de piretroides sobrepapel de filtro. La actividad locomotora se midió durante 30 minutos a partir delmomento en que comenzó la exposición. Actividad locomotora = P/A; donde P es elnúmero de pixels que cambiaron de estado durante la sesión (de apagado aencendido y viceversa), y A es el promedio de pixels encendidos durante la sesión(área). Cada valor representa el promedio de 5 réplicas independientes (n = 5).

98
%de ninfas afectadas(promedio :t DE)
concentración(ug/cmz) deltametn’na cis-pen'netrina
0,14 o‘ o‘
1,4 0 o.
14'0 0 8,0 :l:11,0
140.0 92,0 111,0 100
Tabla 3.4. Toxicidad de las concentraciones de piretroides usadas para evaluarel efecto sobre la actividad locomotora en ninfas Ill.Estos resultados se evaluaronen los mismos insectos usados en los experimentos de medición de la actividadlocomotora, 24 horas después de la aplicación de los insecticidas. El efecto evaluadofue la incapacidad de abandonar un disco de papel de filtro. Los asteriscos indicanconcentraciones que no afectaron significativamente Ia actividad locomotora de lasninfas (Prueba de Duncan, P > 0,05). Cada valor es el promedio de 5 réplicasindependientes (n = 5).

99
(a)
actividad locomotora"2 (pixels/área)120 , 120
2 ng/insecto ' 20 nglinsecto
200 ngllnsectob ‘100 M
ao
60
4o
20
deltametrina cis-pennetrlna
Figura 3.24. Efecto de la aplicación tópica de piretroides sobre la actividadlocomotora en ninfas Ill. Se hizo aplicación tópica en (a) el extremo dorsal delabdomen (2-200 nglmsecto), y (b) el lado dorsal de la cabeza (20 ngfinsecto). Laactividad locomotora se midió durante los (a) 30 ó (b) 15 minutos posteriores a laaplicación. Actividad locomotora = P/A; donde P es el número de pixels quecambiaron de estado durante la sesión (de encendido a apagado y viceversa), y A esel promedio de pixels encendido durante la sesión (área). En (a), dentro de cadainsecticida, las barras señaladas con la misma letra no son significativamentediferentes (Prueba de Duncan, P > 0,05); en (b) las barras señaladas con la mismaletra no son significativamente diferentes (Pmeba de Duncan, P > 0,05). Cada valores el promedio de 5 réplicas independientes :t DE (n = 5).
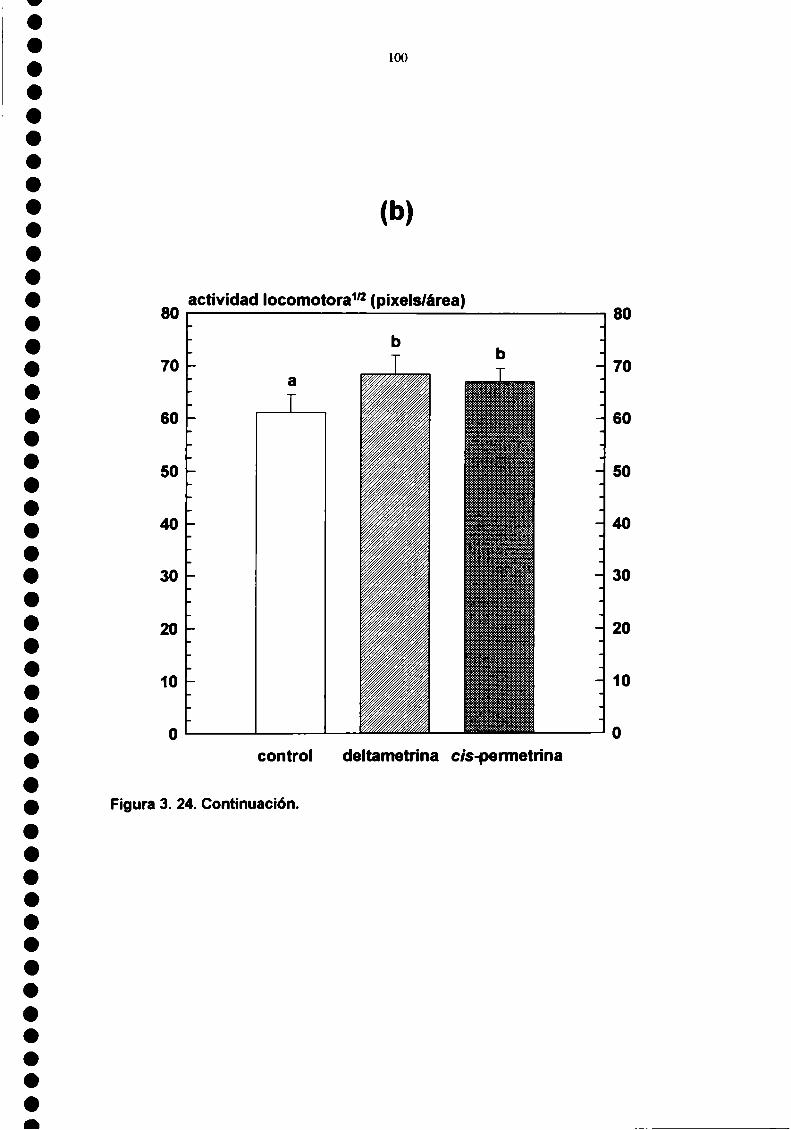
0000m00o
8765321.onq-—q-q—q-u—uq-———--nqq--qq
/
(b)
o0oooo0o
865w321

lOl
%de ninfas afectadas(promedio :l:DE)
dosis(ng/insecto) deltametrina cis-pennetrina
2 10,0 :t 24,7' o'
20 80,0 i 14,1 100'
200 100 100.
Tabla 3.5. Toxicidad de las dosis de piretroides usadas para evaluar el efectosobre la actividad locomotora en ninfas III.Estos resultados se evaluaron en losmismos insectos usados en los experimentos de medición de la actividad locomotora,24 horas después de la aplicación de los insecticidas. El efecto evaluado fue laincapacidad de abandonar un disco de papel de filtro. Los asteriscos indican dosisque no afectaron significativamente la actividad locomotora de las ninfas (Prueba deDuncan, P > 0,05). Cada valor es el promedio de 5 réplicas independientes (n = 5).

102
actividad locomotora"2 (pixels/área)0 120
¡:1 control -i" deltametrina
100 _ b cis-permetrina A 10°
80 "
60
40
20
pretratamlento conacetona sola
pretratamiento conN-etllmaleimida
Figura 3.25. Efecto de piretroides sobre la actividad locomotora en ninfas llltratadas con N-etilmaleimida. Se hizo aplicación tópica de N-etilmaleimida (20pgfnnsecto) sobre el lado dorsal del tórax 5 minutos antes de la exposición a losinsecticidas. Los insectos fueron expuestos a filmes de piretroides sobre papel defiltro (14 ug! cmz). La actividad locomotora se midió durante los 30 minutosposteriores al comienzo de la exposición. Actividad locomotora = P/A; donde P es elnúmero de pixels que cambiaron de estado durante Ia sesión (de apagado aencendido y viceversa), y A es el promedio de pixels encendidos durante la sesión(área). Dentro de cada pretratamiento. las barras señaladas con la misma letra nopresentan diferencia significativa (Prueba de Duncan, P > 0.05). Cada valorrepresenta el promedio de 4 réplicas independientes t DE (n = 5).

103
o actividad locomotora"2 (pixels/área)r {:1 control' ’? deltametrina
100 _ cis-permetrina
80
60
40
20
0
Figura 3.26. Efecto de piretroides sobre la actividad locomotora en ninfas lll a 16y 36°C. Los insectos fueron expuestos a filmes de piretroides sobre papel de filtro (14ug/cmz). La actividad locomotora se midió durante los 30 minutos posteriores alcomienzo de Ia exposición. Actividad locomotora = P/A; donde P es el número depixels que cambiaron de estado durante la sesión (de apagado a encendido yviceversa), y A es el promedio de pixels encendidos durante la sesión (área). Dentrode cada temperatura. las barras señaladas con la misma letra no presentan diferenciasignificativa (PnJeba de Duncan, P > 0.05). Cada valor representa el promedio de 4réplicas independientes :l:DE (n = 5).

104
3. 4. EFECTOS SUBLETALES
3. 4.2. Resultados
Se evaluó el efecto de concentraciones subletales de deltametrina y cis
permetrina sobre Ia ingesta, la muda y la mortalidad de ninfas | y III. Los
insectos estuvieron expuestos durante 40 días a filmes secos de los piretroides
(14 ng/cmz).
La Tabla 3.6. muestra los resultados en ninfas l. Ninguno de los
piretroides tuvo efectos significativos sobre la relación peso preingesta/ peso
posingesta (Anova; F = 1,5; g. l. = 2, 12; P > 0,05), el porcentaje promedio de
ninfas que no se alimentaron (Anova; F = 0,7; g. I. = 2, 12; P > 0,05), ni el
porcentaje promedio de mortalidad (Anova; F = 0,2; g. I. = 2, 12; P > 0,05).
Hubo diferencia significativa entre los tratamientos en el porcentaje promedio
de insectos mudados (Anova; F = 10,3; g. l. = 2, 12; P < 0,05). La deltametrina
produjo una disminución significativa en dicho porcentaje (Prueba de Duncan,
P < 0,05), la cis-permetrina no lo afectó significativamente (Prueba de Duncan,
P > 0,05).
La Tabla 3.7. muestra los resultados en ninfas III. Ninguno de los
piretroides tuvo efectos significativos sobre la relación peso preingesta/ peso
posingesta (Anova; F = 2,1; g. l. = 2, 12; P > 0,05), el porcentaje promedio de
ninfas que no se alimentaron (Anova; F = 0,4; g. l. = 2, 12; P > 0,05), el
porcentaje promedio de mortalidad (Anova; F = 1,2; g. I. = 2, 12; P > 0,05), ni
el porcentaje promedio de muda (Anova; F = 0,1; g. I. = 2, 12; P > 0,05).
3. 4.3. Discusión
Las estrategias de control de plagas se basan en la aplicación de dosis letales
de insecticidas. La aplicación incorrecta, la cobertura incompleta debido a la
existencia de refugios y otros lugares a donde no llegan los insecticidas, la
degradación ambiental y la reinfestación pueden provocar la exposición de los

l05
insectos a dosis subletales. El término subletal se usa comunmente para
referirse a una cantidad de insecticida insuficiente para matar. Aquí lo estamos
usando en un sentido amplio para mencionar dosis que no producen síntomas
visibles de intoxicación.
Algunos efectos subletales conocidos de piretroides en insectos son
inhibición de Ia alimentación (Ruscoe, 1977; Tan, 1981; Kumar and Chapman,
1984; Armstrong and Bonner, 1985; lftner et aI., 1986) y del aumento de peso
(ishaaya et al, 1983); prolongación de los períodos Iarval (Tan, 1981) y pupal
(Kumar and Chapman, 1984); alteración de la tasa reproductiva (Kumar and
Chapman, 1984; Hodges and Meik, 1985; Jackson and Wilkins, 1985);
aumento o disminución de distintas actividades enzimáticas (Saleem and
Shakoori, 1985; Bounias et al, 1985; Saleem and Shakoori, 1986), alteración
de las concentraciones en hemolinfa de azúcares, hormonas y lípidos (Bounias
et al. 1985; Saleem and Shakoori, 1986), repelencia (Ruscoe, 1977; Kumar
and Chapman, 1984; Hodges and Meik, 1986); e inhibición de la comunicación
química (Linn and Roelofs, 1984; Haynes and Baker, 1985; Moore, 1988).
Se propusieron tres mecanismos posibles para explicar los efectos
subletales de los insecticidas (Croft, 1990): (a) efectos directos sobre el
sistema nervioso, (b) efectos indirectos causados por perturbaciones en el
sistema hormonal con la consiguiente alteración de los títulos de secreciones
endocrinas (c) efectos directos sobre blancos no neurológicos, por ejemplo
relacionados con la sintesis de proteínas. Es probable que estos tres tipos de
efectos contribuyan al resultado final.
Los efectos subletales complementan la actividad insecticida de los
piretroides y pueden dar un control superior al esperado en base a los valores
de DE 50 obtenidos en laboratorio (Ruscoe, 1977). Una disminución de Ia
proporción de insectos que mudan, como el que se presenta aquí en ninfas l
de T. ¡nfestans, puede mejorar el control de las poblaciones tratadas.

3.4.3.Tablas
%insectosqueno°/oinsectosque
pesopreingestacomieronmudaron%mortalidad
tratamientopesoposingesta(promedio:i:DE)(promedio:i:DE)(promedio:tDE)
control6,6i0,710,0:t7,173,0i4,514,0i7,4
deltametrina6,1:i:0,711,0:i:6,549,6114,2"13,0:i:5,6
106
cis-permetrina6,2:t0,46,0i8,277,0i9,711,0:t8,9
Tabla3.6.lngesta,mudaymortalidadenninfasltratadasconconcentracionessubletalesdepiretroides.Losinsectosfueron expuestosafilmesdelosinsecticidas(14ng/cm’)durante40días.AIcuartodíadeexposiciónfueronalimentadossobrepaloma.Elvalor señaladoconunasteriscoessignificativamentediferentedelcontrol(PruebadeDuncan,P>0,05).Cadavaloreselpromediode5réplicasindependientes(n=20).

pesopreingesta
tratamientopesoposingesta
%insectosqueno
comieron
(promedio:l:DE)
%insectosque
mudaron
(promedio:l:DE)
%mortalidad
(promedio1:DE)
control6,0i0,8
deltametrina
6,7:l:0,8
cis-permetrina6,8i0,7
15,4i12,9 9,8:t13,68,4i11,1
74,4i17,6 78,6i20,7 78,4:l:12,7
3,2i4,4
107
1,4i3,3
Tabla3.7.lngesta,mudaymortalidadenninfasllltratadasconconcentracionessubletalesdepiretroides.Losinsectosfueron expuestosafilmesdelosinsecticidas(14ng/cmz)durante40días.Alcuartodíadeexposiciónfueronalimentadossobrepaloma.Enningún casoseencontrarondiferenciassignificativasentrelosgruposcontrolytratados(PruebadeDuncanyPmebatparaunamuestra,P>0,05). Cadavaloreselpromediode5réplicasindependientes(n=15).

4
CONCLUSIÓN
Los resultados presentados aqui indican que
_l
M
9’
:5
. Las DE 50s de deltametrina y cis-permetrina en ninfas Ill de T. ¡nfestans
variaron en función del tiempo, pero en ningún momento fueron
significativamente diferentes. Los minimos valores de DE 505 se
manifestaron 24 horas después de la aplicación de los insecticidas.
. En ninfas Ill tratadas con cualquiera de los dos piretroides se manifestó el
fenómeno de recuperación del volteo. Esta recuperación fue inhibida con Ia
aplicación simultánea de butóxido de piperonilo.
En general, los TE 503 de deltametrina fueron menores que los de cis
permetrina para los efectos (a) incapacidad de abandonar un disco de papel
de filtro, (b) ausencia de actividad locomotora. Esto fue así en distintos
estadios ninfales, a distintas temperaturas y en un amplio rango de dosis.
Cuando se hizo aplicación tópica en el extremo abdominal de las ninfas, los
TE 505 de ambos piretroides fueron mayores que cuando Ia aplicación se
hizo en el tórax.
Sólo ei tratamiento con cis-permetrina produjo el síntoma de inflamiento
abdominal.
La temperatura tuvo un efecto importante sobre las CE 503 y DE 503 de los
piretroides estudiados. En general Ia toxicidad aumentó al disminuir la

.05
CO
¡09
temperatura; pero hubo excepciones que dependieron del insecticida, el
estadio ninfal y el rango de temperatura.
EI efecto de la temperatura sobre los TE 50s de los piretroides estudiados
fue poco importante. En general Ia velocidad de acción aumentó al disminuir
la temperatura; pero hubo excepciones que dependieron del insecticida, el
estadio ninfal y el rango de temperatura.
Ninfas Ill intoxicadas con deltametrina o B-cipermetrina sin síntomas visibles
se incoordinaron al descender Ia temperatura, y ninfas incoordinadas
dejaron de mostrar síntomas visibles al aumentar la temperatura. El primero
de estos fenómenos disminuyó en función del tiempo después de la
aplicación de los insecticidas.
. En ninfas l, el isómero cis de la permetrina fue más tóxico que el isómero
trans a 26 y 36°C, e igual de tóxico a 16°C. El pretratamiento con butóxido
de piperonilo sinergizó la toxicidad de cis-permetrina pero no la de trans
permetrina. El pretratamiento con trifenilfosfato tuvo el efecto opuesto. La
mezcla de los dos isómeros produjo un efecto de aditividad (ausencia de
interacción). En ninfas III,el isómero cis de la permetrina fue más tóxico que
el isómero trans a 16 y 26°C, e igual de tóxico a 36°C. Los pretratamientos
con butóxido de piperonilo o trifenilfosfato no modificaron las DE 50s. La
mezcla de los dos isómeros produjo un efecto de antagonismo.
En experimentos con ninfas I de distintas edades y estados alimentarios,
resultaron en general más sensibles los individuos de 1 día de edad, y más
tolerantes los alimentados 7 días antes.
10. Ambos piretroides produjeron hiperactividad locomotora en ninfas lll. Este
efecto estuvo influenciado por el modo y el lugar de aplicación de los
insecticidas. La hiperactividad fue inhibida por el pretratamiento con N
etilmaleimiday por el descenso de la temperatura.

110
11. Ninguno de los dos insecticidas tuvo efectos subletales sobre Ia ingesta y
la mortalidad de ninfas I y III.Sólo deltametrina produjo una disminución en
la cantidad de ninfas I mudadas.
Las propiedades toxicológicas de la cis-permetrina la convierten en una
herramienta potencial para el control de la vinchuca. En la introducción se
mencionó que existe Ia posibilidad de producir este piretroide en Argentina.
Una de las premisas que impulsó nuestros objetivos fue que Latinoamérica
debe desarrollar la tecnología necesaria para resolver sus problemas locales.
Esperamos que este trabajo sea el punto de partida de logros exitosos en el
área del desarrollo de nuevas estrategias para el control de vectores de la
Enfermedad de Chagas.
Los resultados presentados aquí nos hacen plantear un montón de
preguntas. ¿Cuáles son los mecanismos involucrados en la hiperactividad, el
volteo y la muerte?, ¿o se trata de un único mecanismo?; las variaciones de
temperatura ¿afectan principalmente la toxicocinética o Ia toxicodinámica?;
¿qué causa Ia toxicidad diferencial de los isómeros de permetrina?, ¿a qué
nivel ocurre el antagonismo entre los isómeros?; ¿dónde se encuentran los
blancos moleculares cuya interacción con los piretroides genera la
hiperactividad locomotora? Cada una de estos interrogantes es un importante
desafío. Alguien dijo una vez que sólo puede hacer la pregunta correcta quien
conoce buena parte de la respuesta. En los días por venir intentaremos
obtener resultados que ayuden a comprender y replantear mejor estascuestiones.

5
BIBLIOGRAFÍA
- Adams, M. E., and T. A. Miller. 1980. Neural and behavioral correlates of pyrethroidand DDT-type poisoning in the house fly, Musca domestica L. Pestic. Biochem.Physiol. 13: 137-147.
- Agosin, M. 1985. Role of microsomal oxidations in insecticide degradation. In: G. A.Kerkut and L. I. Gilbert (Eds.); Comprehensive Insect Physiology, Biochemistry andPhan'nacology; Vol. 12, pp. 647-712. Pergamon Press, Oxford, UK.
- Agosin, M; A. Morello, and N. Scaramelli. 1964. Partial Characten’zation of the ¡nvivo metabolites of DDT-C" in Tn'atoma infestans. J. Econ. Entomol. 57: 974-977.
- Agosin, M.; N. Scaramelli; L. Gil, and M. E. Letelier. 1969. Some properties of themicrosomal system metabolizing DDT in Tn'atoma ¡nfestans Comp. Biochem.Physiol., 29: 785-793.
- Agrell, Y. P. S., and A. M. Lundquist. 1973. Physiological and biochemical changesduring ¡nsect development. ln: M. Rockstein (Ed.), The physiology of insecta, vol. 1,pp. 159-247. Academic Press, New York, USA.
- Ahmad, M. and A. R. McCaffery. 1988. Resistance to insecticides in a thailand strainof Heliothis annigera (Hübner) (Lepidoptera: Noctuidae). J. Econ. Entomol., 81: 4548.
- Ahn, Y. J.; T. Shono, and J-l. Fukami. 1987. Effect of temperature on pyrethroidaction to kdr-type house fly adults. Pestic. Biochem. Physiol., 28: 301-307.
- Aiworth, W. L. 1980. Estereoquímica y su aplicación en Bioquímica. Alhambra,Barcelona, España.
- Anónimo. 1988. Pyrethroid insecticides in public health. Parasitol. Today, 4: 60-61.
- Armstrong, K. F. , and A. B. Bonner. 1985. lnvestigation of a permethn’n-inducedantiffeding effect in Dmsophila melanogaster. an ethological approach. Pestic. Sci.16: 641-650.
- Barthel, W. F. 1973. Toxicity of pyrethrum ans ¡ts constituents to mammals. In: J. E.Casida (Ed.); Pyrethrum, pp. 123-142. Academic Press, New York, USA.
- Benoit, M; P. Bruneau, and C. Meinard. 1985. Comparative activity of tralomethrinand deltamethnn on Periplaneta americana. Pestic. Biochem. Physiol. 26: 284-291.

D.O...0...0...00.0.00...OOOOOCOOOOOOOOOOOOOOOOOOOi
112
- Berenbaum, M. 1986. Postingestive effects of phytochemicals on insects: onParacelsus and plant products. In: J. R. Millerand T. A. Miller(Eds); Insect-plantinteractions, pp. 121-153. Springer-Verlag, New York, USA.
- Bounias, M; N. Dujin, and D. S. Popeskovic. 1985. Sublethal effects of a syntheticpyrethroid, deltamethn‘n, on the glycemia, the Iipemia and the gut alkalinephosphatases of honeybees. Pestic. Biochem. Physiol. 24: 149-160.
- Bn'ggs, G. G.; M. Elliot; A. W. Farnham; N. F. Janes; P. H. Needham; D. A. Pulman,and S. R. Young. 1976. lnsecticidal activity of the pyrethrins and relatedcompounds. VIII.Relation of polarity with activity in pyrethroids. Pestic. Sci. 7: 236240.
- Brodie, B. B. and R. P. Maickel. 1962. Comparativa biochemistry of drugmetabolism. Proc. First lnternat. Pharmacol. Meet, vol.6, pp. 299-324. PergamonPres, New York, USA.
- Brooks, G. T. 1976. Penetration and distribution of insecticides. In: C. F. Wilkinson(Ed,); Insecticide biochemistry and physiology, pp. 3-58. Plenum Press, New York,USA.
- Burt, P. E., and R. E. Goodchild. 1971. The site of action of pyrethrin I in thenervous system of the cockroach Pen'planeta americana. Entomol. Exp. Appl. 14:179-189.
- Burt, P. E., and R. E. Goodchild. 1974. Knockdown by pyrethroids: its role in theintoxication process. Pestic. Sci. 5: 625-633.
- Busvine, J. R. 1971. A critical review of the techniques for testing insecticides.Commonwealth Agricultural Bureaux, London, UK.
- Camougis. G. 1973. Mode of action of pyrethrum on arthropod nerves. ln: J. E.Casida (Ed.); Pyrethrum, pp. 211-222. Academic Press, New York, USA.
- Cariomagno, M.; E. Cura; A. C. Pérez y E. Segura. Informe sobre Chagas. CienciaHoy, 1: 36-44.
- Carter, S. W. 1989. A review of the use of synthetic pyrethroids in public health andvector pest control. Pestic. Sci. 27: 361-374.
- Casabé, N.; F. Melgar; E. J. Wood, and E. N. Zerba. 1988. Insecticidal activity ofpyrethroids against Tn'atoma infestans. Insect Sci. Applic., 9: 233-236.
- Casabé, N and E. N. Zerba. 1981, Esterases of Tn‘atoma infestans and itsrelationship with the metabolism of organophosphorus insecticides. Comp.Biochem. Physiol., 68: 255-258.
- Casida, J. E. 1980. Pyrethrum flowers and pyrethroid insecticides. Environ. HealthPerspect, 34: 189-202.
- Chen, J. 8.; C. J. Lee, M. G. Yao, and C. N. Sun. 1985. Effects of pyrethroids onknockdown and lack of coordination responses of susceptible and resistantdiamondback moth (Lepidoptera: Noctuidae). J. Econ. Entomol. 78: 1198-1202.

113
- Chilton, M-D; M. Koziel; T. C. Currrier, and B. J. Miflin. 1993. Bt maize for control ofeuropean corn borer. In: R. D. Lumsden and J. L. Vaughn (Eds.); Pestmanagement: biologically based technologies, pp. 370-380. American ChemicalSociety, Washington, USA.
- Chou, J., and T.-C Chou. 1987. Dese-effect analysis with microcomputers. Elsevier,Cambn’dge, UK.
- Clements, A. N., and T. E. May. 1977. The actions of pyrethroids upon the peripheralnervous system and associated organs in the locust. Pestic. Sci. 8: 661—680.
- Coats, J. R. 1990. Mechanisms of toxic action and structure-activity relationships fororganochlorine and synthetic pyrethroid insecticides. Environ. Health Perspec, 87:255-262.
- Croft, B. A. 1990. Arthropod biological control agents and pesticides. Wiley, NewYork, USA.
- Dauterman, W. C. 1983. Role of hydrolases and glutathione S-transferases ininsecticide resistance. In: G. P Georghiou and T. Saito (Eds.); Pesticide resistanceto Pesticides: challenges and prospects, pp. 229-247. Plenum Press, New York,USA.
- Dauterman, W. C. 1985. Insect metabolism: extramicrosomal. ln: G. A. Kerkut and L.I. Gilbert (Eds.); Comprehensive Insect Physiology. Biochemistry andPharmacology; Vol. 12, pp. 713-730. Pergamon Press, Oxford, UK.
- de Licastro, S. A.; E. N. Zerba; E. Wood, and M. I. P. de Villar. 1982. The insecticidaland anticholinesterase activities of 0,0-diethyI-0,4-(2,2-disubstituted vinyl)phenylphosphates and phosphorothioates. Pestic. Sci, 13:505-512.
- Demoute, J.-P. 19891 A brief review of the environmental fate and metabolism ofpyrethroids. Pestic. Sci, 27: 375-386.
- Dent, D. 1991. Insect pest management. Cab, Wallingford, UK.
- Devonshire, A. L. 1989. lnsecticide resistance in Myzus persicae: from field to geneand back again. Pestic. Sci, 26: 375-382.
- Dowd, P. F., and T. C. Sparks. 1984. Developmental changes in trans-pennethn’nand a-naphthyl acetate ester hydrolysisduring the last larval instar of Pseudoplusiaincludens. Pestic. Biochem. Physiol. 21: 275-282.
- Dowd, P. F., and T. C. Sparks. 1987. lnhibition of trans-pen'nethn'n hydrolysis inPseudoplusia includens (Walkers) and use of inhibitors as pyrethroid synergists.Pestic. Biochem. Physiol. 27: 237-245.
- Dowd, P. F. and T. C. Sparks. 1988. Relative toxicity and ester hydrolysis ofpyrethroids in the soybean Iooper and Tobacco budwonn (Lepidoptera: Noctuidae).J. Econ. Entomol, 81: 1014-1018.
- Duncan, D. B. 1955. Multiple range and multiple F tests. Biometrics 11: 1-42.

ll4
- Gammon, D. W.; M. A. Brown, and J. E. Casida. 1981. Two classes of pyrethroidaction in the cockroach. Pestic. Biochem. Physiol. 15: 181-191.
- Ebeling, W. 1974. Permeability of insect cuticle. In: M. Rockstein (Ed.); Thephysiology of insecta, vol. 6, pp. 271-343.
- Ecobichon, D. J. 1991. Toxic effects of pesticides. In: M. O. Amdur, J. Doull and C.D. Klaassen (Eds.); Casarett and Doull's toxicology, pp. 565-622. Pergamon Press,New York, USA.
—Elliot, M. 1989. The pyrethroids: early discovery, recent advances and the future.Pest. Sci, 27: 337-351.
- Elliot, M; A. C. Famham; N. F. Janes; P. H. Needham, and B. C. Pearson. 1967. 5BenzyI-3-furylmethylchrysanthemate: a new potent insecticide. Nature, 213: 493494.
- Elliot, M; A. C. Famham; N. F. Janes; P. H. Needham, and D. A. Pulman. 1974.Synthetic insecticide with a new order of activity. Nature, 248: 71-711.
- Elliot, M.; A. C. Famham; P. H. Needham; D. A. Pulman. and J. H. Stevenson. 1973.A photostable pyrethroid. Nature, 246: 169-170.
- Elliot, M.; N. F. Janes; E. C. Kimmel, and J. E. Casida. 1972. Metabolic fate ofpyrethn'n IIand allethrin orally to rats. J. Agr. Food Chem., 20: 300-313.
- Eto, M. 1974. Organophosphorus pesticides: organic and biological Chemistry. CRCPress, Ohio, USA.
- Fahmy, M. A., and H. T. Gordon. 1965. Selective synergism of carbamateinsecticides on house flies by aryloxyalkylamines. J. Econ. Entomol, 58: 451-455.
- Finney, D. J. 1971. Probit Analysis. Cambridge University Press, New York, USA.
- Fontán, A. and E. N. Zerba. 1984. lntegumental esteratic activity in Tn'atoma¡nfestans and its contribution to the degradation of organophosphorus insecticides.Comp. Biochem. Physiol., 79: 183-188.
- Fontán, A. and E. N. Zerba. 1987. Mode of entry of insecticides in Tn'atoma¡nfestans. Arch. Insect Biochem. Physiol. 4: 313-323.
- Fontán, A. and E. N. Zerba. 1992. Influence of the nutn‘tional state of T. ¡nfestansover the insecticidal activity of DDT. Comp. Biochem. Physiol 101C: 589-591.
- Galun, R.; E. M. Kosower, and N. S. Kosower. 1969. Effect of MethylPhenyldiazenecarboxylate (Azoester) on the feeding behaviour of blood suckinginvertebrates. Nature 224: 181-182.
- Gammon, D. W. 1978. Neural Effects of Allethn'n on the free walking cockroachPen'planeta americana: an investigation using defined doses at 15 and 32°C.Pestic. Sci. 9: 79-91.

115
- Gammon, D. W.; M. A. Brown, and J. E. Casida. 1981. Two classes of pyrethroidaction in the cockroach. Pestic. Biochem. Physiol. 15: 181-191.
- Gasser, C. S., and R. T. Fraley. 1992. Cultivos transgénicos. Investigación y Ciencia191: 64-70.
- Gerolt, P. 1983. lnsecticides: their routes of entry, mechanism of transport and modeof action. Biol. Rev., 58: 233-274.
- Gilbert, L. Y., and King, D. S. 1973. Physiology of growth and development:endocnne aspects. In: M. Rockstein (de), The physiology of insecta, vol. 1, pp.249-370. Academic Press, New York, USA.
- Grafius, E. 1986. Effects of temperature on pyrethroid toxicity to colorado potatobeetle (Coleoptera: Chrysomelidae). J. Econ. Entomol, 79: 588-591.
- Gualtien‘, J. M.; M. Nelson, and J. A. Cichero. 1985. Presente y perspectiva delcontrol químico. En: R. U. Carcavallo; J. E. Rabinovich, y R. J. Tonn; Factoresbiológicos y ecológicos en la Enfermedad de Chagas, pág. 319-329. CentroPanamericano de Ecología Humana y Salud, Organización Panamericana de laSalud, Servicio Nacional de Chagas y Ministerio de Salud y Acción Social de laRepública Argentina; Buenos Aires, Argentina
- Gualtien‘, J. M.; C. H. Rios; J. A. Cichero; R. Váez, and R. U. Carcavallo. 1984.Ensayo de campo con decametrina en su formulación liquido emulsionable yfloable en el control del Tn'atoma infestans en la provincia de Córdoba. Chagas, 1:17-20.
- Hackman, R. H., and Goldberg, M. 1987. Comparativa study of some expandingcuticles: the relation between composition. structure and function. J. Insect Physiol.33: 39-50.
- Hayes, W. J. 1975 Toxicology of pesticides. Williams and Wilkins, Baltimore, USA.
- Haynes, K. F., and T. C. Baker. 1985. Sublethal effects of pen'nethrin on thechemical communication system of the pink bollwon'n moth, Pectinophoragossypiella. Arch. Insect Biochem. Physiol., 2: 283-293.
- Head, S. W. 1973. Composition of pyretthm extract and analysis of pyrethrins. In:J. E. Casida (Ed); Pyrethrum, pp. 25-53. Academic Press, New York, USA.
- Hill,I. R. 1989. Aquatic organisms and pyrethroids. Pestic. Sci, 27: 429-458.
- Hillerton, J. E. 1978. Changes in the structure and composition of the extensiblecuticle of Rhodnius prolixus through the 5th larval instar. J. Insect Physiol. 24: 399412.
- Hirano, M. 1989. Characteristics of pyrethroids for insect pest control in agriculture.Pestic. Sci, 27: 353-360.
- Hodges, R., and J. Meik. 1986. Lethal and sublethal effects of pennethn'n onTanzanian strains of Tn'boliumcastaneum (Herbst), Gnatocerus maxillosus (F.),

ll6
Sitophilus oryzae (L.) and Sitophilus zeamais Motschulsky. Insect Sci. Applic. 7:533-537.
- Hodgson, E., and L. G. Tate. 1976. Cytochrome P450 interactions. ln: C. F.Wilkinson (Ed,); Insecticide biochemistry and physiology, pp. 115-148. PlenumPress, New York, USA.
- Holan, G.; D. F. O'Keefe; C. Virgona, and R. Walser. 1978. Stmctural and biologicallinkbetween pyrethroids and DDTin new insecticides. Nature, 272: 734-736.
- Iftner, D. C.; F. R. Hall, and M. M. Stun'n. 1986. Effects of residues of fenvalerateand pennethrin on the feeding behavior of Tetranychus urticae (Koch). Pestic. Sci.17: 242-248.
- Inglesfield, C. 1989. Pyrethroids and terrestn'al non-target organisms. Pestic. Sci.,27: 387-429.
- Insausti, T. 1994. Nervous system of T. infestans. J. Morphol. 221: 343-359.
- lshaaya, I., and J. Casida. 1983. Pyrethroid detoxification and synergism in insects.In: J. Miyamoto and P. C. Keamye (Eds.); Pesticide Chemistry, human welfare andthe environment; vol. 3, pp. 307-310. Pergamon Press, Oxford, UK.
-Ishaaya, I.; A. Elsner; K. R. Simon Ascher, and J. Casida. 1983. Syntheticpyrethroids: toxicity and synergism on dietary exposure of Tn'boliumcastaneum(Herbst) Iarvae. Pestic. Sci. 14: 367-372.
- Jacoby, W. B. 1980. Detoxication enzymes. In: W. B. Jacoby (Ed.); Enzymatic basisof detoxication, pp. 1-6. Academic Press, New York, USA.
- Jackson, A. E. M., and R. M. Wilkins. 1985. The effect of sub-Iethal dosages of thesynthetic pyrethroid fenvalerate on the reproductive rate of the aphid Myzuspersicae. Pestic. Sci. 16: 364-368.
- Klaassen, C. D., and D. L. Eaton. 1991. Principles of toxicology. In: M. O. Amdur, J.Doull and C. D. Klaassen (Eds.); Casarett and Doull's toxicology. pp. 565-622.Pergamon Press, New York, USA.
- Klassen, W. 1995. World Food Security up to 2010 and the global pesticidesituation. In: N. N. Ragsdale; P. C. Keamey, and J. R. Plimmer; Eighth internationalcongress of pesticide Chemistry; pp. 1-32. American Chemical Society,Washington, USA.
- Koyama, N., and K. Kurihara. 1971. Modification by chemical reagents of proteins inthe gustatory and olfactory organs of the fleshfly and cockroach. J. Insect Physiol.17: 2435-2440.
—Kumar, K., and R. B. Chapman. 1984. Sublethal effects of insecticides on thediamondback moth Plutella xylostella (L.). Pestic. Sci. 15: 344-352.
- Lehninger, A. L. 1981. Bioquímica. Omega, Barcelona, España.

ll7
- Linn, C. E., Jr., and W. L. Roelofs. 1984. Sublethal effects of neuroactivecompounds on pheromone response thresholds in male oriental fruit moths. Arch.Insect Biochem. Physiol. 1: 331-344.
- Lund, A. E. 1985. Insecticides effects on the nervous system. ln: G. A. Kerkut and L.l. Gilbert (Eds); Comprehensive Insect Physiology, Biochemistry andPharrnacology; Vol. 12, pp. 203-216. Pergamon Press, Oxford, UK.
- Maddrell, S. H. P. 1966. Newous control of the mechanical properties of theabdominal wall at feeding in Rhodnius. J. Exp. Biol. 44: 59-68.
- Maddrell, S. H. P., and J E. Casida. 1971. Mechanism of insecticide-induced diuresisin Rhodnius. Nature. 231: 55-56.
- Malkenson, N. C. de; E. J. Wood, and E. N. Zerba. 1984. lsolation andcharactenzation of an esterase of Tríatoma infestans with a critical role in thedegradation of organophosphoan esters. Insect Biochem., 14: 481-486.
- Matsumura, F. 1975. Toxicology of insecticides. Plenum Press, New York, USA.
- McLaughlin, G. A1973. History of pyrethrum. ln: J. E. Casida (Ed); Pyrethrum, pp.3-15. Academic Press, New York, USA.
- Metcalf, R. L. 1975. Pest management strategies for the control of insects affectingman and domestic animals. In: R. L. Metcalf and W. H. Luckman (Eds.);Introduction to Insect pest management, pp. 529-564. Wiley, New York, USA.
- Metcalf, R. L. 1989. Insect resistance to insecticidas. Pestic. Sci, 26: 333-358.
- Miller,T. A., and M. E. Adams. 1982. Mode of action of pyrethroids. ln: J. R. Coats(Ed); Insecticide mode of action, pp. 3-27. Academic Press, New York, USA.
- Moncayo, A. 1993. Chagas disease. ln: World Health Organization (Ed); Tropicaldisease research, Progress 1991-1992; pp. 67-75. World Health Organization,Geneva, Switzerland.
- Moore, R. F. 1988. Inhibition of chemical communication between male and femalebollworrns (Lepidoptera: Noctuidae) by sublethal amounts of pen'nethn'n. J. Econ.Entomol. 81: 78-82.
- Mrak, E. M. 1973. Advantages and disadvantages of pyrethmm. ln: J. E. Casida(Ed); Pyretthm, pp. 307-311. Academic Press, New York, USA.
—Nakatsugawa, T. and M. A. Morelli. 1976. Microsomal oxidation and insecticidemetabolism. In: C. F. Wilkinson (Ed,); Insecticide biochemistry and physiology, pp.61-114. Plenum Press, New York, USA.
- Napolitano, L.M.; S. C. Ford, and L. Tompkins. 1986. The effects of developmentaland experimental temperature on responses of Drosophila melanogaster adults tosucrose. J. Insect Physiol. 11: 937-940.

ll8
- Naumann, K. 1990. Synthetic pyrethroid insecticide. ln: W. S. Bowers, W. Ebing, D.Martin and R Wegler (Eds.); Chemistry of plant protection, Vol. 4. Springer-Venag,Berlin, Germany.
- Norris, D. M.; S. M. Ferkovich; J. E. Baker; J. M. Rozental, and T. K. Borg. 1971.Energy transduction in quinone inhibition of insect feeding. J. Insect Physiol. 17:85-97.
- Oliveira Filho, A. M. 1994. Techniques for the entomological evaluation of the controlof chagas disease vectors. Acta Toxicol.Argentina 2: 52- 55.
- Oppenhoorth, F. J. 1985. Biochemistry and genetics of insecticide resistance. ln: G.A. Kerkut and L. I. Gilbert (Eds.); Comprehensive Insect Physiology. Biochemistryand Pharmacology; Vol. 12, pp. 731-773. Pergamon Press, Oxford, UK.
- Osborne, M. P., and D. R. Pipper. 1992. Mechanisms of kdr and super-kdrresistance. In: C. A. Mullin and J. G. Scott (Eds.); Molecular mechanisms ofinsecticide resistance, pp. 71-89. American Chemistry Society, Washington, USA.
- Pauron, D.; M.Amichot, and J-B. Berge. 1992. Voltage-dependent sodium channelsin susceptible and pyrethroid-resistant Drosophila strains. ln: I. Denholm; A. L.Devonshire, and D. W. Hollomon;Resistance 91, Achievements and developmentsin combating perticide resistance, pp. 228-239. Elsevier, London, UK.
- Picollo, M. Y.; E. Seccacini; C. Vassena, and E. Zerba. 1993. Feeding and matingdeterrency by sulphydryl reagents in Tn'atomainfestans. Acta Tropica 52: 297-307.
- Pillmore, R. E. 1973. Toxicity of pyrethrum to fish and wildlife. ln: J. E. Casida (Ed);Pyrethrum, pp. 143-165. Academic Press, New York, USA.
- Pinchin, R.; A. M. de Oliveira Filho, and A. C. B. Pereira. 1980. The flushing-outactivity of pyrethrum and synthetic pyrethroids on Pastrongilus megistus, a vectorof Chagas's disease. Trans. Roy. Soc. Trop. Med. Hyg. 75: 691-694.
- Repetto, M1988. Toxicología fundamental. Científico-médica, Madrid, España.
- Reynolds, S. E. 1975. The mechanism of plasticization of the abdominal cuticle inRhodnius. J. Exp. Biol. 62: 81-98.
- Ruigt, G. S. F. 1985. Pyrethroids. ln: G. A. Kerkut and L. l. Gilbert (Eds.);Comprehensive Insect Physiology. Biochemistry and PharmacoIOQY;vol. 12, pp183-262. Pergamon Press, Oxford, UK.
- Ruscoe, C. N. E. 1977. The new NRDC pyrethroids as agricultural insecticides.Pestic. Sci. 8: 236-242.
- Saleem, M. A., and A. R. Shakoon‘. 1985. Effects of permethn'n and deltamethrin onsome biochemicals components of Tn'boliumcastaneum larvae. Pakistan J. Zool.17: 321-328.
- Saleem, M. A., and A. R. Shakoori. 1986. Biochemical effects of sublethal doses ofcypermethn'n on the sixth-instar Iarvae of Tn'bolium castaneum (Herbst). Arch.Insect Biochem. Physiol. 27: 101-105.

H9
- Salgado, V. L.; S. N. lrving, and T. A. Miller. 1983. The importance of nerve terminaldepolan'zation in pyrethroid poisoning of insects. Pestic. Biochem. Physiol. 20: 169182
- Sawicki, R. M. 1962. Insecticidal activity of pyrethmm extract and its four insecticidalconstituents against house fly. III. Knock-down and recovery of flies treated withpyrethrum extract with and without piperonyl butoxide. J. Sci. Food Agn'c. 13: 283292.
- Schmidt, C. D., and J. L. Robertson. 1986. Effects of treatment technique onresponse of horn flies (Diptera: Muscidae) to pennethn'n at different temperatures.J. Econ. Entomol, 79: 684-687.
- Schofield, C. J. 1994. Triatominae, biología y control. Eurocommunica,Sussex, UK.
- Schouest, L. P., Jr., and T. A. Miller. 1988. Factors influencing pyrethroid toxicity inpink bollworm (Lepidoptera: Gelechiidae): implications for resistance management.81: 431-436.
- Scott, J. G.; and G. P. Georghiu. 1984. Influence of temperature on knockdown,toxicity, and resistance to pyrethroids in the house fly, Musca domestica. Pestic.Biochem. Physiol. 21: 53-62.
- Shimada, Y.; A. Shiraishi; H. Kijima, and H. Mon'ta. 1972. Effects of sulphydrylreagents on the labellar sugar receptor of the fleshfly. J. lnsect Physiol. 18: 18451855.
- Shono, T; K. Ohsawa, and J. Casida. 1979. Metabolism of trans- and cis-pennethn‘n,trans- and cis-cypennethrin, and decamethn'n by microsomal enzymes. J. Agn'c.Food Chem., 27: 316-325.
- Singer, G.; J. M. Rozental, and D. M. Norris. 1975. Sulphydryl groups and thequinone receptor in insect olfaction and gustation. Nature 256: 222-223.
- Sipes, l. G. and A. J. Gandolfi. 1991. Biotransfonnation of toxicants. In: M. O.Amdur, J. Doulland C. D. Klaassen (Eds.); Casarett and Doull's toxicology, pp. 88126. Pergamon Press, New York, USA.
- Sívon', J. L. 1993. Regulación de la glutatión S-transferasa de Tn'atoma infestans ysu importancia en el proceso de intoxicación por insecticidas organofosforados.Tesis doctoral. Universidad de Buenos Aires.
- Soderlund, D. M.; J. R. Sanbom, and P. W. Lee. 1983. Metabolism of pyrethrins andpyrethroids in insects. ln: D. H. Hutson and T. R. Roberts (Eds.); Progress inpesticide biochemstry and toxicology, vol. 3, pp. 401-435. Wiley, New York, USA.
- Sokal, R. R., and F. J. Rohlf. 1981. Biometry. Freeman, San Francisco, USA.
- Sparks, T. C.; Pavloff, A. M.; R. L. Rose, and D. F. Clower. 1983. Temperaturetoxicity relationship of pyrethroids on Heliothis virescens (F.) (Lepidoptera:

120
Noctuidae) and Anthonomus grandis grandis Boheman (Coleoptera:Curculionidae). J. Econ. Entomol., 76: 243-246.
- Sparks, T. C.; Shour. M. H.; and E. G. Wellemeyer. 1982. Temperature-toxicityrelationships of pyrethroids on three Iepidopterans. J. Econ. Entomol., 75: 643-646.
- Tabashnik. B. E.; N. L. Cushing, and M. W. Johnson. 1987. Diamond moth(Lepidoptera: Plutellidae) resistance to insecticides in Hawaii: intra-island variationand cross resistance. J. Econ. Entomol. 80: 1091-1099.
- Tan, K. -H. 1981. Antifeeding effect of cypennethn'n and pennethn'n at sub-lethallevels against Pien's brassicae larvae. Pestic. Sci. 12: 619-626.
- Tan, K. -H. 1982. Irn'tance response to temperature after sub-Iethal action ofpyrethroids against cabbage white caterpillars Pien's Brassicae. Entomol. Exp.Appl. 32: 151-154.
- Terriere, L. C. 1984. Induction of detoxication enzymes in insects. Ann. Rev.Entomol. 29: 71-88.
- Thaung, M._and P. J. Collins. 1986. Joint effects of temperature and insecticides onmortality and fecundity of Sitophilus oryzae (coleoptera: Curculionidae) in wheatand maize. J. Econ. Entomol._79: 909-914.
- Toth, Jr; S. J.. and T. C. Sparks. 1988. Influence of treatment technique on thetemperature-toxicity relationships of cis- and trans-pennethn'n in the cabbageIooper (Lepidoptera: Noctuidae). J. Econ. Entomo|., 81: 115-118.
- Toth, Jr; S. J.. and Sparks, T. C. 1990. Effect of temperature on toxicity andknockdown activity of cis-permethn'n, esfenvalerate. and x-cyalothn'n in thecabbage Iooper (Lepidoptera: Noctuidae). J. Econ. Entomol., 83: 342-346.
- Uchitel, D. P. 1990. GraphPad lnstat 2.0. GraphPad Software, lrvine, USA.
- Udagawa, T; S. Numata; K. Oda; S. Shiraishi; K. Kodaka. and K. Nakatani. 1985. Anew type of synthetic pyrethroid insecticide. In: N. F. Janes (Ed.); Recent advancesin the Chemistry of insect control, pp 192-204. Royal Society of Chemistry,Dorchester. UK.
- Uehara, S., and H. Mon'ta. 1972 The effects of temperature on the labellarchemoreceptors of the blowfly.J. Gen. Physiol. 59: 213-226.
- Vijverberg, H. P. M. and J. van den Bercken. 1990. Neurotoxicological effects andthe mode of action of pyrethroid insecticides. Cn'tical Rev. Toxicol., 21: 105-126.
- Villar, M. l. P. de; E. J. Wood; E. N. Zerba; S. A. de Licastro, and N. Casabé. 1980.Cholinesterase and esen‘ne resistant esterases in the developing embryo ofTn'atoma infestans and its role as targets for inhibition in the ovicide action ofparathion. Comp. Biochem. Physiol. 67: 55-59.
- Villet, R. H. 1974. Involvement of amino and sulphydryl groups in olfactorytransduction in silkmoths. Nature 248: 707-709.

121
- Wadleigh, R. W.; P. G. Koeheler; H. K. Preisler; R. S. Patterson, and J. L.Robertson. 1991. Effect of temperature on the toxicities of ten pyrethroids togerman cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol., 84: 1433-1436.
- Wallace, G.; N. Casabé; E. Wood, and E. Zerba. 1988. Assay of pyrethroidhydrolising esterases using (1,R)-cis-3-(2,2-dibromovyniI)-2,2-dimethylcyclopropanecarboxylates as substrates. Xenobiotica, 18: 351-355.
- Watters, F. L.; N. D. G. White, and D. Coté. 1983. Effect of temperature on toxicityand persistence of three pyrethroid insecticides applied to fir plywood for thecontrol of the red flour beetle (Coleoptera: Tenebn'onidae). J. Econ. Entomol., 76:11-16.
- Welling, W. 1979. Toxicodynamics of the insecticidal action: an introduction. Pestic.Sci, 10: 540-546.
- Welling, W. and G. D. Paterson. 1985. Toxicodynamics of insecticides. In: G. A.Kerkut and L. I. Gilbert (Eds.); Comprehensive Insect Physiology, Biochemistry andPharmacology; vol. 12, pp. 603-645. Pergamon Press, Oxford, UK.
- [WHO] World Health Organization. 1991. Control of Chagas disease, Report of aWHO Expert Commitee. World Health Organization, Geneva, Switzenand.
- Wilkinson, C. F. 1976. Insecticide interactions. In: C. F. Wilkinson (Ed,); Insecticidebiochemistry and physiology, pp. 605-647. Plenum Press, New York, USA.
- Williams,C. H. 1973. Tests for possible teratogenics, carcinogenics, mutagenics andallergenics effects of pyrethrum. ln: J. E. Casida (Ed); Pyrethrum, pp. 167-176.Academic Press, New York, USA.
- Wood, E. J.; M. I. P. de Villar, and E. N. Zerba. 1985. Role of a microsomalcarboxylesterase in reducing the action of malathion in eggs of Tn'atoma infestans.Pest. Biochem. Physiol., 23: 24-32.
- Wood, E.; S. A. de Licastro, N. Casabé, J. Sivori, and E. Zerba. 1993. Evalua-tion ofthe flushing-out activity of pyrethroids on Tn'atoma infestans. Insect Sci. Applic. 14:651-655.
- Wood, E. J.; E. N. Zerba; M. l. Picollo y N. Casabé. 1992. TIP: formulado deinsecticidas no contaminante y de alta seguridad para el control de vinchucas.Resúmenes del VIII Congreso Argentino de Toxicología, p. 84. Buenos Aires,Argentina.
- Wood, E. J.; M. I. Picollo de Villar, y E. N. Zerba. 1993. Comparación entre lavariación de la capacidad detoxificante y la diferente susceptibilidad al insecticidamalatión entre ninfas V de distinta edad de T. ¡nfestans An. Asoc. Quim. Argent.81: 153-162.
- Yadwad, V. B., and V. L. Kallapur. 1988. Influence of temperature on knock-downand mortality to fenitrothion in the three Iepidopten’an species of insects. Insect Sci.Applic., 9: 531-534.

122
- Yu, S. J. 1983. Age variation in insecticide susceptibility and detoxification capacityof fall armyworm (Lepidoptera: Noctuidae) Iarvae. J. Econ. Entomol. 76: 219-222.
- Zerba, E. 1988. Insecticidal activity of pyrethroids on insects of medical importance.Parasitol. Today 4: 53-57.
- Zerba. E. N. 1989. Chemical control of Chagas Disease Vectors. Biomed. Environ.Sci. 2:24-29.
- Zerba. E. 1995. Fumigant canister and other novel insecticide formulations. PublicHealth 12: 62-68.
- Zerba, E. N.; F. Melgar, and E. J. Wood. 1982. Sublethal effect of Iindane in vaporphase on the respiratory activity of Tn'atoma ¡nfestans (Hemiptera) as a cause ofsynergism in fumigants. Abstracts of the Fifth international congress of pesticideChemistry (IUPAC), Iva-11. Kyoto, Japan.
- Zerba. E. N.; F. Melgar; G. Wallace; E. Wood; S. A. de Licastro, y M. l. P. de Villar.1988. Nuevas formulaciones fumígenas para el control de vectores de Iaenfermedad de Chagas. Chagas, 5: 2-7.
mil 511057
























![Insecticidas[1] N10](https://img.pdfslide.es/doc/110x75/54a221c2ac7959d2348b45a1/insecticidas1-n10.jpg)




