Embed Size (px)
Citation preview

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
40
VIII. A. FOSFORILACIÓN OXIDATIVA
La FOSFORILACIÓN OXIDATIVA es el proceso por medio del cual se produce la mayor cantidad de ATP en las células tanto procariotas como eucariotas. La fosforilación oxidativa ocurre en la mitocondria en los eucariontes, pero la membrana mitocondrial interna y la matriz mitocondriales participan específicamente. En procariotes, la fosforilación oxidativa ocurre en la membrana plasmática.
.
La fosforilación oxidativa forma parte del catabolismo aerobio que se puede iniciar desde la oxidación de la glucosa hasta la la formación de piruvato y de ahí a Acetil-CoA, su subsecuente entrada al Ciclo de Krebs en donde se forman grandes cantidades de coenzimas reducidas, o sea ricas en electrones: NADH y FADH2, las cuales a su vez van a alimentar el proceso de la fosforilación oxidativa. La fosforilación oxidativa consta de dos partes que están acopladas entre sí: una oxidativa que comprende una serie de reacciones de transferencia de electrones a partir de la oxidación de sustratos y una fosforilante, en la cual el Pi se esterifica al ADP para formar ATP.
En la cadena respiratoria, los electrones son transferidos de complejo en complejo para generar ATP.
La parte oxidativa está integrada por una serie de reacciones de óxido-reducción que se llama cadena respiratoria o cadena transportadora de electrones. Esta cadena “recibe” a los electrones que vienen del resto del metabolismo y que vienen en la forma de las coenzimas reducidas NADH y FADH2. Los electrones son aceptados y transferidos secuencialmente por complejos de proteínas que están en la membrana interna mitocondrial. Al final, los electrones son aceptados por el oxígeno, que queda reducido a H2O. Como se consume este oxígeno, al proceso de transporte de electrones se le llama también respiración celular.
LA MITOCONDRIARIBOSOMAS
MATRIZ MITOCONDRIAL MEMBRANA EXTERNA
MEMBRANA INTERNA
ESPACIO INTERMEMBRANAL
CRESTAS MITOCONDRIALES
PORCIÓN Fo
PORCIÓN F1
Ácidos grasos
Ciclo de Krebs
Glucosa Piruvato
FOSFORILACIÓN OXIDATIVA
ELECTRONES

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
41
La cadena respiratoria está formada por: a) 4 complejos proteicos: los Complejos I, II, III y IV, embebidos a la membrana mitocondrial interna b) 2 transportadores móviles: la Coenzima Q o Ubiquinona que es una molécula orgánica, muy hidrofóbica solubilizada en la parte hidrofóbica de la membrana interna y el citocromo c que es una proteína periférica a la membrana mitocondrial interna por el lado de la membrana que encara al espacio intermembranal de la mitocondria. La ATP sintetasa es el Complejo V, pero no forma parte de la cadena respiratoria, sino que al fosforilar al ADP, es parte de la fosforilación oxidativa La cadena respiratoria o el transporte de electrones mitocondrial se lleva a cabo por complejos enzimáticos que están formados por muchos polipéptidos que tienen como grupos prostéticos moléculas orgánicas que les ayudan a acarrear los electrones y que por tanto se reducen y oxidan reversiblemente. Todos los complejos tienen grupos REDOX. Los 5 complejos de la fosforilación oxidativa se localizan en la membrana interna mitocondrial.
10-30 600 ATP sintasa o Complejo F1F0
V
Hemo a Hemo a3 CuA CuB
12 160 Citocromo c oxidasa IV
Hemo 1
12 Citocromo c
Citocromo c
Hemo b1 Hemo b2 Hemo c1 FeS
12 258 Complejo bc1 o CoQ citocromo c óxido reductasa
III
0.8636
Ubiquinona CoQ
FAD FeS
4 140 SUCCINATO DESHIDROGENASA O SUCCINATO CO Q REDUCTASA
II
FMN FeS
34 880 NADH deshidrogenasa o NADH Co Q oxido reductasa
I
GRUPO PROSTÉTICO
SUBUNIDADES
MASA MOLECULAR (Da)
NOMBRE COMPLEJO
COMPLEJOS DE LA FOSFORILACIÓN OXIDATIVA

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
42
FADH2
Espacio intermembranal
Membrana interna
Matriz
Membrana externa
Citocromo c oxidasa (bovino)
COMPLEJO I COMPLEJO II COMPLEJO III COMPLEJO IV COMPLEJO V
Coenzima Q o Ubiquinona Citocromo c
MATRIZ MITOCONDRIAL
ESPACIO INTERMEMBRANAL
ATP sintasa (E. coli)
Citocromo c oxidasa (bovino) Citocromo bc1
(bovino)
NADH deshidrogenasa (Thermus termophylus)
Membrana mitocondrial interna
Succinato Fumarato Fumarato reductasa (E.coli)
Los electrones que se transfieren entre los complejos respiratorios provienen del NADH o del FADH2 que se formó en Ciclo de Krebs y que entran a nivel de los complejos respiratorios I y II, respectivamente
La transferencia de electrones entre los complejos respiratorios sigue una secuencia específica que está de acuerdo a los potenciales de oxidoreducción (E°’) de los grupos prostéticos de los complejos. Como esta transferencia va siempre desde un grupo con E°’ más negativo a uno menos negativo (o más positivo), el proceso es espontáneo o exergónico

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
43
Los electrones son transferidos de complejo en complejo a través de los centros redox de cada uno. En cada una de las transferencias de electrones hay una molécula que pierde electrones, otra que los gana y luego ésta a su vez los pierde para entregarlos a otra molécula que a su vez los acepta para luego perderlos y cederlos a otra. Así se va realizando la transferencia de electrones en cadena.
Complejo I Complejo II
Succinato
Fumarato
Complejo III
Complejo IV

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
44
Los H del NADH y del FADH2 que van entrando a la cadena respiratoria se van “disociando” en e- y H+ al entrar a los Complejos I y II. Los e- siguen entonces una ruta “horizontal” dentro de las regiones membranales de los complejos respiratorios, mientras que los H+ se bombean por los complejos respiratorios también al otro lado de la membrana interna, hacia el espacio intermembranal, generándose así un gradiente de H+ a ambos lados de la membrana interna mitocondrial. Este gradiente tiene dos componentes uno de concentración de los H+, llamado componente químico u osmótico y uno eléctrico, por la carga positiva de los H+.
El gradiente electroquímico de H+ así generado constituye una fuente de energía, pues los H+ se acumulan en el espacio intermembranal y tenderían a moverse espontáneamente a una región de menor concentración de H+, pero no pueden hacerlo pues la membrana mitocondrial interna es impermeable a los H+. La forma de aprovechar esta energía almacenada en el gradiente de H+ es haciéndolos pasar por una “puerta” específica y al disiparse así la energía, es aprovechada para sintetizar ATP por la ATP sintasa, la cual es la puerta por la que los H+ ahora “regresan” a la matriz mitocondrial en un movimiento espontáneo pues van de una región de mayor concentración (el espacio intermembranal) a uno de menor concentración (la matriz). Se forman 3 ATP si los electrones entran en la NADH deshidrogenasa (Complejo I) y 2 ATP si entran en la succinato deshidrogenasa (Complejo II).
MATRIZ
MEMBRANA INTERNA
ESPACIO INTERMEMBRANAL
Complejo II Succinato-CoQ oxidoreductasa
Complejo III CoQH2-citocromo c oxidoreductasa
Complejo I NADH-CoQ oxidoreductasa
Fumarato Succinato
Ciclo Q Complejo IV
Citocromo oxidasa

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
45
La reacción de síntesis de ATP es un proceso endergónico. La energía necesaria es proporcionada por el gradiente electroquímico de H+, que fue formado por la actividad de transporte de electrones de la cadena respiratoria. La ATP sintasa lleva a cabo la formación del enlace anhídrido éster entre el ADP y el Pi, “descargando” el gradiente de H+, por lo que también es capaz de transportar H+.
Esta enzima tiene un mecanismo rotacional de síntesis de ATP en el que la enzima funciona como un molino de agua impulsado por una corriente de iones H+, que desde la parte inferior base de la enzima (Fo) hace girar a la parte central de la estructura de la F1, mientras la gran masa formada por las subunidades α y β son fijadas a la membrana por el estator y no giran. TEORÍA QUIMIOSMÓTICA.- LOS POSTULADOS DE MITCHELL Peter Mitchell concibió que la forma en la que se acoplaban la síntesis de ATP y el transporte de electrones por la cadena respiratoria era a través del gradiente electroquímico de H+. Sus ideas están contenidas en la Hipótesis Quimiosmótica, cuyos postulados son: 1) Las cadenas transportadoras de electrones, respiratoria y fotosintética deben translocar H+. 2) La ATP sintasa debe funcionar como una ATPasa que transloca H+ reversiblemente. 3) Las membranas transductoras de energía deben tener una baja conductancia efectiva de H+. 4) Las membranas transductoras de energía deben poseer acarreadores de intercambio específico, para permitir la permeación de metabolitos y mantener la estabilidad osmótica, en presencia de un alto potencial de membrana. Se puede entender claramente que estos postulados reflejan los conceptos explicados anteriormente para describir el funcionamiento de la fosforilación oxidativa. BALANCE TOTAL DE PRODUCCIÓN DE NADPH/ FADH2/ ATP DESDE QUE ENTRA GLUCOSA A GLUCÓLISIS Y TERMINA EN FOSFORILACIÓN OXIDATIVA
Región F0: Es hidrofóbica y cruza la membrana, tiene de 4 a 12 diferentes cadenas polipeptídicas. Forma el canal que conduce los protones.
COMPLEJO V o ATP SINTASA o F1F0
Región F1: Es hidrofílica pero está unida a la región F0. Da Hacia la matriz mitocondrial y consta de 5 diferentes tipos de cadenas polipeptídicas. Lleva a cabo la síntesis de ATP, o sea tiene el sitio catalítico.
H
H
H
HH+
H
HH
H
H
H
HH+
H
H H
H
H
[H+]
[H+]
ADP +Pi
ATP CADENA RESPIRATORIA
ATP SINTETASA
MEMBRANA INTERNA
MEMBRANA EXTERNA

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
46
Asumimos que cada NADH que se forma puede llegar a la mitocondria y ahí oxidarse en la cadena respiratoria. Por cada NADH así incorporado, generaríamos 3 ATP en la fosforilación oxidativa y por cada FADH2 generaríamos 2 ATP
Glucosa
2 Piruvato
Balance para una molécula de glucosa que se convierte en 2 piruvatos, luego en 2 Acetil-CoA y luego a CO2 en la vía el ciclo del ácido tricarboxílico , con todo el NADH y el FADH2 convertidos en ATP por la respiración: 1 glucosa + 38 ADP + 38 Pi -------> 6 CO2 + 38 ATP Nota: 2 de los NADH son formados en el citoplasma durante la glicólisis. Para ser transportados a la matriz mitocondrial para ser posteriormente oxidado por la cadena transportadora de electrones, tienen que pasar por medio de transporte activo al interior de la mitocondria. Esto "cuesta" 1 ATP por NADH. Por lo tanto el balance final resulta en 36 ATP por glucosa y no 38 ATP.

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
46
VIII. B. FOTOFOSFORILACIÓN
El otro proceso, además de la fosforilación oxidativa, que es muy eficiente para formar ATP es la fotosíntesis. En la fosforilación oxidativa, los hidrógenos que alimentan de electrones a la cadena respiratoria vienen en el NADH y el FADH2, producidos principalmente de la oxidación de sustratos en el Ciclo de Krebs. En la fotosíntesis, los electrones provienen de clorofilas excitadas por la luz. Aquí también el transporte de electrones genera un gradiente de H+ que sirve para sintetizar ATP y por eso aquí se llama fotofosforilación.
Los principales productos de la fotosíntesis son O2, ATP, NADPH y Carbohidratos. Estos productos se generan en dos fases: la luminosa y la oscura. En las reacciones de la fase luminosa se produce ATP, O2 y NADPH y en las reacciones de la fase oscura (Ciclo de Calvin), se producen carbohidratos y se consumen NADPH y ATP (por tanto consume energía y poder reductor y es una vía anabólica que genera al NADP+ que es un oxidante).
ESTRUCTURA DEL CLOROPLASTO
Membrana interna Membrana externa
Espacio tilacoidal
Espacio intermembranal Estroma
lamelar
Estroma Grana
La fotosíntesis ocurre en los cloroplastos en las algas y plantas y en la membrana plasmática de las bacterias fotosintéticas El cloroplasto tiene el sistema membranal para hacer la transducción de energía, a semejanza de lo que sucede en la mitocondria y crear así compartimentos que puedan sostener la formación de gradientes de H+
.
Tilacoide (cada “bolsita”) Rodeado por la membrana tilacoidal

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
47
FASE LUMINOSA DE LA FOTOSÍNTESIS Las reacciones básicas de la fase luminosa de la fotosíntesis incluyen:
1. Absorción de luz por los pigmentos cosechadores de luz 2. Excitación de dos clorofilas especiales que emiten un electrón 3. Transporte de electrones y por tanto óxidoreducciones (se forman NADPH y O2) 4. Formación de un gradiente de H+ 5. Fosforilación de ADP para formar ATP
Por tanto, la energía luminosa se convierte en energía química del enlace ADP~Pi. A esto se le llama transducción de energía.
Las clorofilas y los pigmentos accesorios forman trampas de luz que colectan a los fotones y los dirigen hacia un par de clorofilas especiales para excitar su estado y producir la emisión de electrones que van a ser transportado por una cadena de transporte de electrones. Tanto las clorofilas como los pigmentos accesorios contienen muchos enlaces dobles conjugados por lo que se excitan fácilmente, absorbiendo la luz de diferentes longitudes de onda para cubrir el espectro de la luz solar.
Las reacciones de la fase luminosa se llevan a cabo en los fotosistemas I y II (PSI, PSII). Ambos funcionan secuencialmente. Estos fotosistemas contienen tres grandes tipos de componentes: •Clorofilas: tipo a, b •Pigmentos accesorios o antena: carotenoides y ficobilinas •Proteínas: integrales de membrana y periféricas •Otros grupos prostéticos como grupos hemo, átomos de Cu, centros Fe-S
carotenoide
clorofilas
ficobilinas
Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
48
Los fotosistemas que emiten electrones están “conectados” a una serie de transportadores de electrones. Todos los componentes de los fotosistemas I y II y los del transporte de electrones y de la síntesis de ATP están embebidos en la membranas tilacoidales. Los transportadores de electrones y la enzima que sintetiza ATP son complejos parecidos a los del transporte de electrones de la fosforilación oxidativa.
º
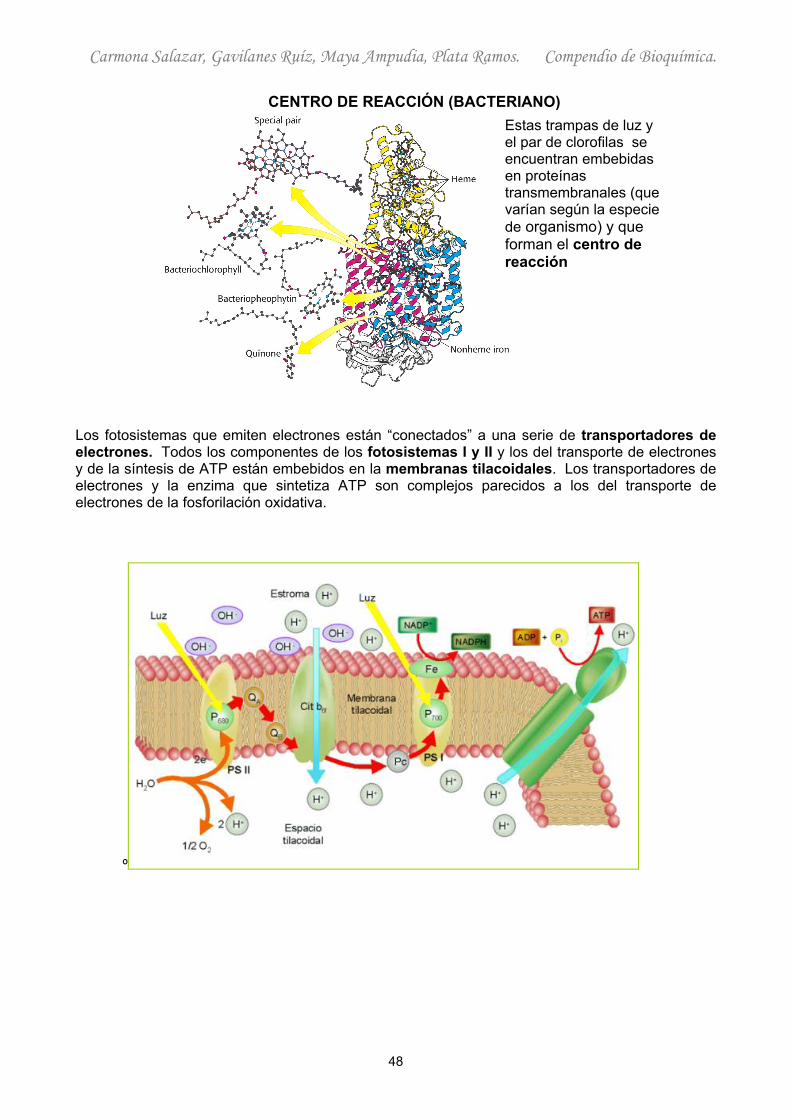
CENTRO DE REACCIÓN (BACTERIANO) Estas trampas de luz y el par de clorofilas se encuentran embebidas en proteínas transmembranales (que varían según la especie de organismo) y que forman el centro de reacción

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
49
feofitina y luego la plastoquinona, tiene que ser “repuesto” a la clorofila. Esto lo hace un complejo proteico que tiene Mn2+ y que obtiene esos electrones del H2O, produciéndose así el O2 de la fotosíntesis. De la plastoquinona, los electrones reducen al complejo b6f que es análogo al bc1 mitocondrial. De aquí, los electrones son transferidos a la plastocianina, una proteína que tiene Cu2+ como grupo prostético. Los electrones son aceptados por el PSI que había perdido un electrón y que es también un centro de reacción con trampas de luz y un par de clorofilas especiales, a las que la luz había excitado para emitir un electrón que fluyó hacia la ferredoxina. La ferredoxina es una proteína soluble que transfiere electrones a la ferredoxina-NADP-reductasa, enzima que transfiere los electrones al NADP+ para formar NADPH. Durante las transferencias de electrones se produce un gradiente de H+ generado por un bombeo neto de H+ a nivel del complejo b6f y también por los H+ producidos por la oxidación del H2O y la reducción que sufre la plastoquinona cuando toma los electrones de la feofitina (ver 2 figuras atrás). La energía del gradiente de H+ se disipa o utiliza para formar ATP por la ATP sintasa del cloroplasto, que es muy parecida a la mitocondrial y es la que fosforila al ADP para dar ATP. En el cloroplasto se le llama también CF1FO. Así se lleva a cabo la fotofosforilación. FASE OSCURA DE LA FOTOSÍNTESIS En esta fase de la fotosíntesis se utilizan el NADPH y el ATP formados en las reacciones de la fase luminosa. A las reacciones de la fase oscura de la fotosíntesis se le llama también reacciones del Ciclo de Calvin En el Ciclo de Calvin se llevan a cabo reacciones que tienen como fines:
Fijar el CO2 del aire en moléculas orgánicas sencillas. Producir esqueletos carbonados sencillos que se usan como precursores en la síntesis
de otros compuestos. Producir grandes cantidades de NADP+
. La vía es por tanto sintética (anabólica) y es reductora (tiene reacciones que consumen poder reductor o NADPH).
Una vez que la clorofila del PS I o el PSII se excita, su potencial redox pasa de ser positivo a ser muy negativo gracias a los fotones excitadores de la luz. Los electrones pueden entonces fluir cuesta abajo energéticamente por una cadena que va a transportarlos usando centros redox de potenciales más negativos a menos negativos (y que son transferencias espontáneas, con ∆G negativos). En el PSII, el electrón de la clorofila que se transfiere al siguiente aceptor, que es la
FOTOSISTEMA I
FOTOSISTEMA II
Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
50
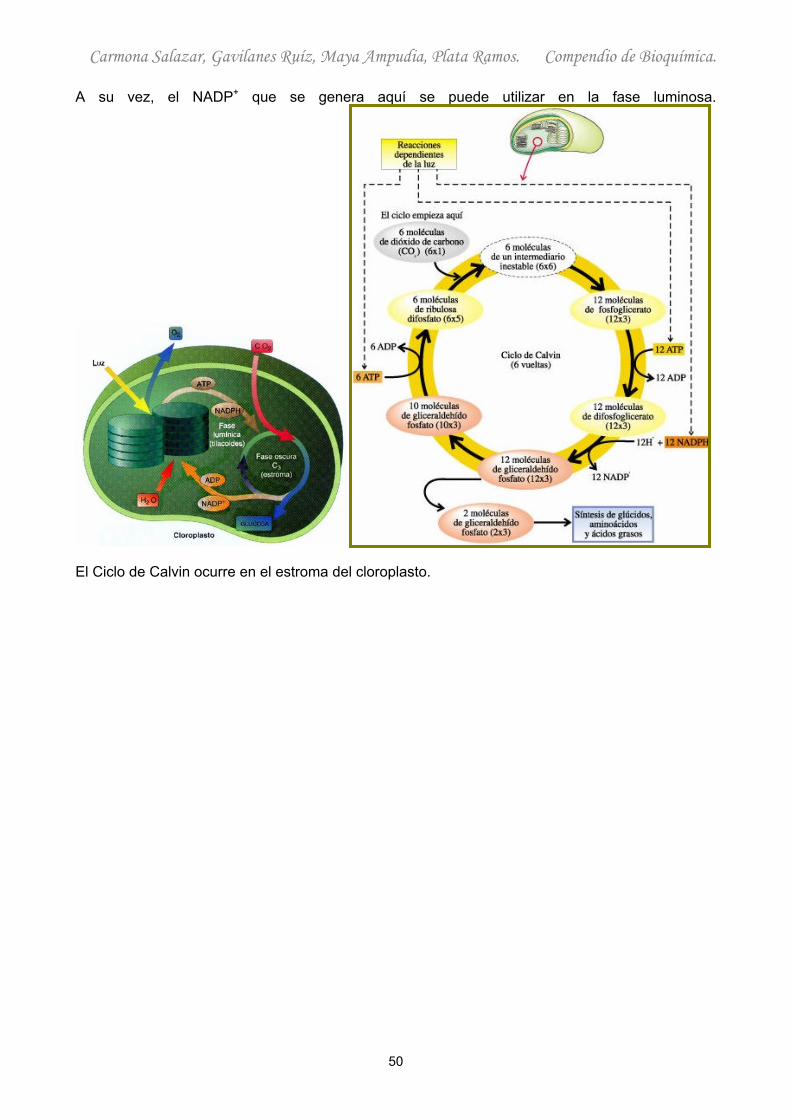
A su vez, el NADP+ que se genera aquí se puede utilizar en la fase luminosa.
El Ciclo de Calvin ocurre en el estroma del cloroplasto.

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
51
IX. METABOLISMO DEL GLUCÓGENO
ESTRUCTURA DEL GLUCÓGENO El glucógeno es un polímero de moléculas de glucosa unidas por enlaces glucosídicos α(1→4) y con ramificaciones formadas por enlaces α(1→6). Es la reserva de carbohidratos en animales y se almacena principalmente en hígado y músculo. En las células se encuentra como gránulos citosólicos que tienen asociadas a las enzimas responsables de su síntesis y degradación. Su degradación se realiza liberando unidades de glucosa de los extremos no reductores. Su alto grado de ramificación permite que su ruptura sea rápida a través de la liberación simultánea de glucosa de los varios extremos no reductores.

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
52
GLUCOGENÓLISIS O DEGRADACIÓN DEL GLUCÓGENO
La degradación del glucógeno inicia con la fosforólisis del glucógeno por la glucógeno fosforilasa para producir glucosa-1-P. Después, la fosfoglucomutasa convierte a la glucosa-1-P en glucosa-6-P, que puede seguir la vía glucolítica (en músculo) o ser hidrolizada a glucosa por la glucosa fosfatasa para ser liberada al torrente sanguíneo desde el hígado. Las ramificaciones del glucógeno se rompen por la enzima desramificadora del glucógeno que transfiere un trisacárido con uniones α(1→4) hacia otro extremo no reductor, originando así más enlaces α(1→4) para la acción de la fosforilasa. La glucógeno fosforilasa es la enzima reguladora de la degradación del glucógeno, y es modulada por alosterismo y modificación covalente.
Glucógeno ( Glucosa n )
Glucógeno fosforilasa
Glucosa - 1P + Glucosa n -1
fosfoglucomutasa
Glucosa - 6P (a glucólisis en músculo)
Glucosa - 6 - fosfatasa (hígado)
Glucosa + Pi (a torrente sanguíneo)

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
53
REGULACIÓN DE LA DEGRADACIÓN DEL GLUCÓGENO
GLUCOGÉNESIS O SÍNTESIS DE GLUCÓGENO La síntesis del glucógeno inicia con la activación de la glucosa-1-P con UTP catalizada por la UDP-glucosa pirofosforilasa. La Glucosa-UDP tiene un estado de mayor energía que le permite ser un buen donador del grupo glucosilo. Esta reacción está cercana al equilibrio; sin embargo, está acoplada a la hidrólisis del pirofosfato por una pirofosfatasa inorgánica. Ésto convierte a la reacción en una muy exergónica (∆G°’=-31 kJmol-1). Después, la glucógeno sintasa transfiere el grupo glucosilo a un extremo no reductor del glucógeno para formar un enlace α(1→4). Esta reacción tiene ∆G°’=-13.4 kJmol-1, por lo que es espontánea. Las ramificaciones en las cadenas del glucógeno se generan por la enzima ramificadora (amilo–(1,4 →1,6)-transglicosilasa).
La conformación activa de la glucógeno fosforilasa se promueve por dos mecanismos: 1) por conformaciones alostéricas inducidas por el AMP (que indica deficiencia energética) y 2) por la fosforilación de la enzima catalizada por la fosforilasa b cinasa, dicha cinasa se activa también por fosforilación a través de la PKA que fue activada por glucagón (en hígado) o por epinefrina llamada también adrenalina (en músculo). La forma inactiva de la glucógeno fosforilasa también se genera por cambios conformacionales en la estructura de la enzima con los inhibidores alostéricos ATP y Glucosa-6-P (G6P) y por desfosforilación de la misma. Cuando la glucógeno fosforilasa se encuentra activa (alta degradación del glucógeno), la enzima que la desfosforila (fosfoproteína fosfatasa) debe estar inactiva. Ésto se logra por la interacción de la fosfatasa con un inhibidor fosforilado (también por la PKA). Cuando el inhibidor se desfosforila pierde afinidad por la fosfatasa y ésta se activa, defosforilando a la glucógeno fosforilasa y por tanto disminuyendo la glucogenólisis.
UDP-glucosa
Glucógeno sintasa : +Glu6P
Glucosa -1P
Glucosa -UDP
Glucosan-1
Glucosa nGlucógeno
UTP
PPi*
UDP
* PPi + H 2
O
Pirofosfatasa inorgánica ∆ G°’= -33.5 kJ/mol
2 HOPO 3
2 -
pirofosforilasa

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.
54
REGULACIÓN DE LA SÍNTESIS DEL GLUCÓGENO La glucógeno sintasa se regula por alosterismo y por modificación covalente. La forma fosforilada es inactiva y se inhibe alostéricamente por ATP, ADP y Pi, ya que éstos son indicio de abundancia energética, por lo que promueven el almacenamiento de la glucosa en forma de glucógeno. La fosforilación de esta enzima se puede llevar a cabo por diversas cinasas como la PKA, la cinasa de proteína dependiente de calmodulina y la glucógeno sintasa cinasa 3.
m-GlucógenoSintasaInactiva
O-Glucógenosintasaactiva
OH
O-P
Cinasa Fosfatasa
SÍNTESIS DEGLUCOGENO
SÍNTESIS DEGLUCOGENO
-ATP-ADP
-PI
´
´
La síntesis y degradación del glucógeno están reguladas de manera coordinada. Por ejemplo, cuando el glucagon o la epinefrina incrementan el AMPc, se incrementa la fosforilación por PKA de la glucógeno fosforilasa (que se activa) y de la glucógeno sintasa (que se inactiva). De esta forma. Se incrementa la glucogenólisis y se disminuye la glucogenogénesis.

Carmona Salazar, Gavilanes Ruíz, Maya Ampudia, Plata Ramos. Compendio de Bioquímica.



![[PPT]LA MEMBRANA PLASMÁTICA - Biologia Blog | … · Web viewTEMA 8. LA MEMBRANA PLASMÁTICA Y LAS ENVOLTURAS EXTERNAS 1. LA MEMBRANA PLASMÁTICA 1.1. MEMBRANA UNITARIA Membrana](https://img.pdfslide.es/doc/110x75/5bcd58cf09d3f206798d90a2/pptla-membrana-plasmatica-biologia-blog-web-viewtema-8-la-membrana-plasmatica.jpg)