Embed Size (px)
Citation preview


COLECCIÓN ESTUDIOS

LOS CROCODYLIA
DE VENEZUELA:
Ecología
y
Conservación
Andrés Eloy Seijas
2011

Los Crocodylia de Venezuela
Andrés Eloy Seijas
© Andrés Eloy Seijas, 2011
© Academia de Ciencias Físicas, Matemáticas y Naturales, 2011
Hecho el depósito de Ley
Depósito Legal: lf65920115914107
ISBN: 978-980-6195-19-6
Diseño y Diagramación: Antonio Machado- Allison
Impresión: Brima Color
Todos los derechos reservados, Ninguna parte de esta publicación puede ser
reproducida, por ningún medio, sin la previa autorización escrita del autor y
de la Academia.
Impreso en Venezuela- Printed in Venezuela

PRESENTACIÓN
"Aquella trompa feroz, toda negra y de duro hueso, con
quijadas, que las he medido, de cuatro palmos, y algunos
más; aquel laberinto de muelas, duplicadas las filas de
arriba y abajo, y tantas, no sé si diga navajas aceradas,
dientes o colmillos; aquellos ojos resaltados del casco, con
tal maña que sumida toda la corpulenta bestia bajo el agua,
saca únicamente la superficie de ellos para registrarlo todo
sin ser visto; aquel dragón de cuatro pies horribles,
espantosos en tierra y formidables en el agua, cuyas duras
conchas rechazan a las balas, frustrándoles el ímpetu, y
cuyo cerro de broncas y desiguales puntas, que les afecta el
lomo y la cola de alto bajo, publica que todo él es
ferocidad, saña y furor: por lo cual no hallo términos que
expliquen la realidad de las especies que de este infernal
monstruo retengo concebidas"
Esta descripción publicada sobre el "Caimán del Orinoco" por
Gumilla en el famoso libro El Orinoco Ilustrado y Defendido..... nos
permite interpretar la pléyade de reacciones por parte de los primeros
visitantes a nuestras tierras y el encuentro con nuestra fauna tropical.
Desde esos, hasta nuestros días hemos observado con suma
tristeza, como este grupo de animales silvestres ha sido diezmado y
colocado en situación cercana a su desaparición total en nuestro
mundo, ya sea por el temor que se desarrolló en las poblaciones de
colonos, o por el uso de sus pieles altamente cotizadas en el mercado.
El libro que tenemos la satisfacción de presentar nos otorga un
inmenso volumen de información sobre este grupo de animales
acuáticos desde aquellos temas de carácter científico taxonómico,
evolutivo y biológico, hasta otros, no menos interesantes y de rigurosa
investigación, como aquellos planes de manejo y cría en cautiverio que
ha permitido parcialmente la conservación de las especies y el
repoblamiento en algunos cuerpos de agua.

Este libro es el producto de toda una vida científica del autor, del
trabajo con sus alumnos y colegas, de la experiencia en campo y
laboratorio que inteligentemente ha colocado en ocho capítulos
ampliamente ilustrados y con una .extraordinaria base de datos.
Nos introduce con un pasaje general sobre los Crocodylia como
grupo zoológico al cual pertenecen en la actualidad 23 especies
mundialmente distribuidas, con 7 en América del sur y 5 de ellas en
nuestro país. Pasa posteriormente a describir detalladamente las
especies presentes en Venezuela. El Capítulo 3 lo dedica a los
aspectos de captura, manipulación y marcaje en el campo y el cuarto
a los aspectos morfométricos tradicionales usados para el reco-
nocimiento de las especies.
El Capítulo 5 lo dedica al análisis poblacional donde abunda en
datos sobre unidades de medida y determinación de la estructura
poblacional todo ello con amplias consideraciones metodológicas de
gran importancia para la formación de estudiantes.
El Capítulo 6 nos narra sobre aspectos de la dieta de estas
especies en ambientes naturales, en diferentes sitios y épocas del año.
Todo ello muy importante para poder manejar adecuadamente este
recurso, que es profundamente descrito en el Capítulo 7.
Finaliza con un capítulo dedicado a las experiencias nacionales de
la cría en cautiverio con fines de repoblamiento. Tres especies han
sido pioneras en este trabajo. La principal el "Caimán del Orinoco"
debido a la gran amenaza de desaparición. Le siguen el "Caimán de
la Costa", una especie rara, poco abundante y que vive en sistemas
asociados a la desembocadura de ríos cercanos al mar y por último
experiencias con la "Baba" habitante común de nuestros llanos y con
gran potencial económico.
Para la Academia es altamente reconfortante poder contribuir con
la difusión de esta experiencia de vida, en la seguridad que contribuirá
a la formación de nuestros estudiantes, generar un mayor aprecio por
esta fauna silvestre y a la conservación de nuestras especies.
Antonio Machado-Allison


7
CAPÍTULO 1
LOS CROCODYLIA
El presente libro trata sobre la ecología y la conservación de las
cinco especies de Crocodylia de Venezuela. Sin embargo, para
situar al lector en el contexto de lo que significa este grupo
zoológico, este primer capítulo se centra en sus características más
distintivas, su distribución y en aspectos generales compartidos por
todas las especies de este Orden respecto a la evolución, biología y
ecología. Con frecuencia se hará referencia al conjunto de animales
de este grupo como a los “crocodilios”, castellanizando de esta
manera el nombre del Orden. Términos como “cocodrilos”, “croco-
diloides” “cocodriloides” y “crocodiloideos”, pueden también consi-
derarse válidos. La expresión “crocodilianos”, muy en boga, pro-
viene del inglés “crocodilians” y su uso se ha extendido mucho.
Suramérica puede ser considerada como el continente de los
Crocodylia, ya que alrededor de un tercio de las 23 especies
reconocidas en el mundo de este grupo de animales habitan allí.
Cinco de las siete especies suramericanas están presentes en el
territorio venezolano: el caimán de la costa, Crocodylus acutus; el
caimán del Orinoco, Crocodylus intermedius; la baba o babo,
Caiman crocodilus; el babo negro, Paleosuchus trigonatus y el
babo morichalero, Paleosuchus palpebrosus. La supuesta pre-
sencia de una sexta especie en nuestro país (Melanosuchus niger)
no ha sido fehacientemente comprobada (ver, no obstante, Donoso-
Barros, 1966a y King, 1991). En el mundo, sólo Colombia y Brasil,
con seis, superan a Venezuela en número de especies de cro-
codilios.
En comparación con las especies presentes en los Estados
Unidos de América (Alligator misissippiensis y Crocodylus
Andrés Eloy Seijas
8
acutus) o en Australia (Crocodylus porosus y Crocodylus
johnsoni), los Crocodylia en Venezuela han sido poco estudiados.
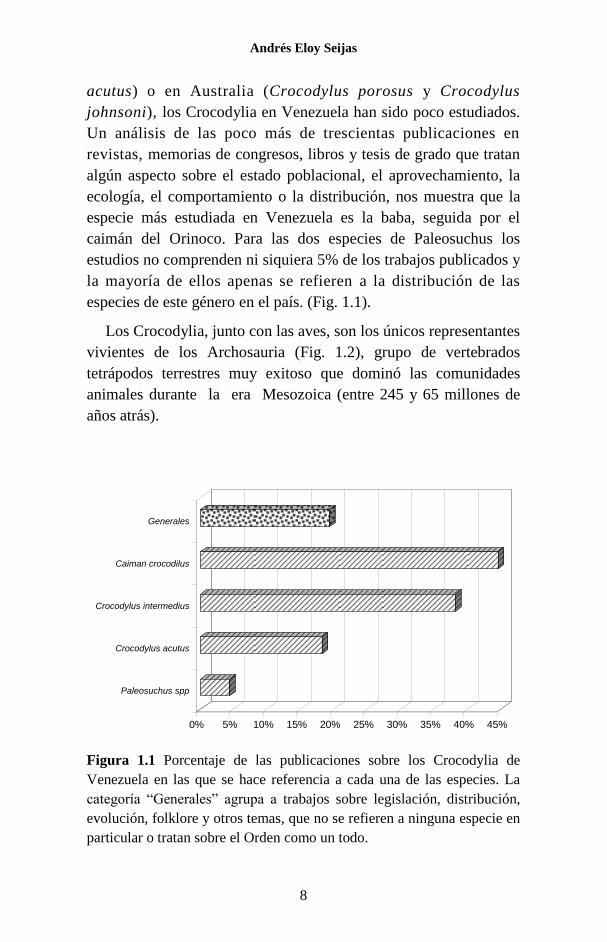
Un análisis de las poco más de trescientas publicaciones en
revistas, memorias de congresos, libros y tesis de grado que tratan
algún aspecto sobre el estado poblacional, el aprovechamiento, la
ecología, el comportamiento o la distribución, nos muestra que la
especie más estudiada en Venezuela es la baba, seguida por el
caimán del Orinoco. Para las dos especies de Paleosuchus los
estudios no comprenden ni siquiera 5% de los trabajos publicados y
la mayoría de ellos apenas se refieren a la distribución de las
especies de este género en el país. (Fig. 1.1).
Los Crocodylia, junto con las aves, son los únicos representantes
vivientes de los Archosauria (Fig. 1.2), grupo de vertebrados
tetrápodos terrestres muy exitoso que dominó las comunidades
animales durante la era Mesozoica (entre 245 y 65 millones de
años atrás).
Figura 1.1 Porcentaje de las publicaciones sobre los Crocodylia de
Venezuela en las que se hace referencia a cada una de las especies. La
categoría “Generales” agrupa a trabajos sobre legislación, distribución,
evolución, folklore y otros temas, que no se refieren a ninguna especie en
particular o tratan sobre el Orden como un todo.
0% 5% 10% 15% 20% 25% 30% 35% 40% 45%
Paleosuchus spp
Crocodylus acutus
Crocodylus intermedius
Caiman crocodilus
Generales

Los Crocodylia de Venezuela
9
Son animales altamente especializados para su modo de vida
semi-acuático, cuyos ancestros se remontan a unos 200 millones de
años. Los Crocodylia deben haber descendido de algún tipo de
theconodonte, reptiles Archosaurios cuyos dientes se alojaban en
alvéolos profundos (al igual que en los mamíferos actuales) en vez
de estar fusionados a las mandíbulas (Sues, 1989). Los crocodilios
modernos pertenecen a los Eusuchia, que aparecieron en el
Cretácico tardío hace unos 80 millones de años. Las 23 especies
de crocodilios actuales representan una pequeña fracción (10% de
acuerdo con una vieja estimación de Langston en 1973) de las que
se conocen que han existido con base en los registros fósiles. Ellas
son las sobrevivientes a los cambios climáticos globales desen-
cadenados por la colisión de un meteorito con la Tierra, los cuales
de acuerdo con la hipótesis en boga (Pough et al., 1996) causaron
la erradicación de muchas especies terrestres y marinas hace 65
millones de años.
Figura 1.2 Cladograma que muestra la relación filogenética de los Cro-
codylia con los otros vertebrados tetrápodos (Modificado a partir de
Pough et al., 1996). Los números indican los cambios o características
evolutivas claves que definen a cada grupo. Sarcopterygia es un antiguo
linaje de peces con aletas lobuladas (de las cuales evolucionaron las
extremidades) al que pertenecen los actuales peces pulmonados y el
celacanto.
Tetrápodos
Amniotas
Saurópsidos
AnuraAnura(ranas)(ranas)
UrodelaUrodela(salamandras)(salamandras)
GymnophionaGymnophiona(cecilias)(cecilias)
Testudomorpha(tortugas)
Lepidosauria(lagartos y serpientes)
CrocodyliaCrocodylia AvesMammaliaMammalia
Diápsidos
1
2
3
4
5
67
RELACIÓN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRÁPODOSRELACIRELACIÓÓN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRÁÁPODOSPODOS
AMNIOTASAMNIOTAS
ANFIBIOSANFIBIOS REPTILESREPTILES
1.- Par de extremidades pectorales y pélvicas.
3.- Huesos tabular y supratemporal pequeños o ausentes.
Centrum del atlas y el intercentrum del axis fusionados.
2.- Membranas que protegen el embrión (amnios,
alantoides, corion).
4.-Cráneo con par de aberturas temporales
5.- Presencia de una abertura craneal
anterior a la órbita ocular.
6.- Plumas. Pérdida de los dientes.
Homeotermia.
7.- Pelos. Mandíbula formada por
un solo hueso. Homeotermia.
SarcopterygiaSarcopterygia
Archosauria
Tetrápodos
Amniotas
Saurópsidos
AnuraAnura(ranas)(ranas)
UrodelaUrodela(salamandras)(salamandras)
GymnophionaGymnophiona(cecilias)(cecilias)
Testudomorpha(tortugas)
Lepidosauria(lagartos y serpientes)
CrocodyliaCrocodylia AvesMammaliaMammalia
Diápsidos
1
2
3
4
5
67
RELACIÓN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRÁPODOSRELACIRELACIÓÓN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRN EVOLUTIVA DE LOS CROCODYLIA CON OTROS TETRÁÁPODOSPODOS
AMNIOTASAMNIOTAS
ANFIBIOSANFIBIOS REPTILESREPTILES
1.- Par de extremidades pectorales y pélvicas.
3.- Huesos tabular y supratemporal pequeños o ausentes.
Centrum del atlas y el intercentrum del axis fusionados.
2.- Membranas que protegen el embrión (amnios,
alantoides, corion).
4.-Cráneo con par de aberturas temporales
5.- Presencia de una abertura craneal
anterior a la órbita ocular.
6.- Plumas. Pérdida de los dientes.
Homeotermia.
7.- Pelos. Mandíbula formada por
un solo hueso. Homeotermia.
SarcopterygiaSarcopterygia
Archosauria

Andrés Eloy Seijas
10
Las patas largas y delgadas que poseían los primitivos Croco-
dylia se toman como indicio de que eran probablemente depre-
dadores terrestres. El modo de vida anfibio fue desarrollado más
adelante en su historia evolutiva (Bellairs, 1987).
En contraste con la amplia diversidad exhibida por las aves (los
otros Archosauria vivientes, con más de 9.000 especies) la de los
Crocodylia es muy reducida, algo sorprendente para un grupo con
una historia geológica tan antigua (Buffetaut, 1989). Los registros
fósiles indican una mayor diversidad de especies en el pasado, pero
incluso en ese momento su número era relativamente escaso. Para
el caso de Venezuela, se han descrito varias especies fósiles
encontradas en el estado Falcón, algunas de ellas pertenecientes a
linajes no presentes actualmente en el país, como es el caso de los
gavialoides (Sill, 1970; Medina, 1976; Brochu y Rincón, 2004).
Los crocodilios viven en áreas tropicales y subtropicales del
mundo, aunque los aligatores pueden alcanzar latitudes de hasta los
35 grados al norte, en los E.U.A. (Alligator missisippiensis) y
China (A. sinensis).
CARACTERÍSTICAS FÍSICAS
¿Cuáles son las características distintivas que convierten a un
crocodilio en crocodilio?
Los Crocodylia poseen dos pares de patas (tetrápodos). Las
anteriores, más cortas, poseen cinco dedos. Las posteriores son
robustas y presentan cuatro dedos con membrana interdigital, al
menos parcial. Son animales alargados, con cuerpo semejante al de
los lagartos, con cola musculosa redondeada al inicio (en la
proximidad de las patas traseras) y comprimida lateralmente hacia
su parte final. La primera parte de la cola tiene una doble cresta,
mientras que la mitad posterior presenta solamente una (Fig. 2.1,
Capítulo 2).
Poseen mandíbulas armadas con numerosos dientes cónicos (de
30 a 40 en cada una, dependiendo de las especies) que encajan en

Los Crocodylia de Venezuela
11
alvéolos. No poseen labios y la lengua es plana y completamente
adosada al fondo de la boca.
Cuando los crocodilios están al acecho de sus presas y sumer-
gidos en el agua, lo único visible es la parte superior de la cabeza
(donde se ubican el oído, los ojos y los orificios nasales, Fig. 1.3) y,
a veces, algo del dorso.
Figura 1.3 Arriba: los crocodilios pueden permanecer sumergidos con sólo
los ojos y la punta del hocico en la superficie (fotografía de José de Jesús
Vargas); esa es la típica posición de acecho. Abajo: ubicación de las narinas
externas y el oído.
Narinas externas
Oído
Narinas externas
Oído

Andrés Eloy Seijas
12
Las aberturas nasales (narinas externas) de los Crocodylia están
ubicadas en posición dorsal en el extremo anterior del hocico. Ellas
conectan con canales internos que conducen el aire, a través de un
paladar secundario, desde el exterior hasta las llamadas coanas o
narinas internas, en la garganta, justo detrás de una válvula palatal
que impide el paso del agua hacia los pulmones cuando el animal
está sumergido (Fig. 1.4). Todo ello permite al animal respirar
apenas asomando la punta del hocico, aun cuando la boca perma-
nezca abierta dentro del agua. Las narinas externas se cierran de
manera refleja, impidiendo el paso del agua cuando el animal se
sumerge. Los oídos también poseen mecanismos de cierre, lo que
protege a estos órganos de los sentidos durante las inmersiones.
Figura 1.4 Válvula palatal al fondo de la boca de un caimán del
Orinoco. Esta válvula sirve de mecanismo para aislar la garganta de la boca.
Los ojos de los Crocodylia poseen membranas nictitantes, es
decir unos pliegues transparentes que cubren los globos oculares y
los separan del exterior con movimientos intermitentes. Esta mem-
brana protege el ojo y permite la visión dentro del agua. Sin
embargo, la captura de presas dentro del agua depende más de
órganos sensoriales en la piel que de la visión, que es más bien
pobre en ese medio (Fleishman, 1989).

Los Crocodylia de Venezuela
13
La piel de los crocodilios está formada por una gruesa capa
dérmica cubierta por escamas epidérmicas no superpuestas. Em-
bebidos en la piel se encuentran unas placas óseas, llamadas
osteodermos, que forman una especie de coraza. El grado de
desarrollo de esos huesos dérmicos varía entre las distintas
especies.
El cráneo de los Crocodylia tiene una configuración incon-
fundible. Detrás de las órbitas oculares aparecen dos pares de
perforaciones o aberturas denominadas fenestrae (ventana, en
latín). Un par redondeado en la parte superior de la tapa craneal y
el otro par ubicado en posición lateral, separadas de las órbitas por
una especie de puente o columna ósea (Fig. 1.5). Esta confi-
guración del cráneo, con dos pares de aberturas, recibe el nombre
técnico de “diápsido”. Los lagartos, serpientes, tuataras y aves
también son diápsidos, aunque con modificaciones en las cuales las
aberturas se han transformado o perdido durante el proceso
evolutivo.
Figura 1.5 Cráneo de Crocodylus intermedius. Observe la ubicación de los
dos pares de aberturas temporales.

Andrés Eloy Seijas
14
COMPORTAMIENTO
Las especies de crocodilios actuales son anfibias y pasan buena
parte de su vida en el agua, donde nadan mediante movimientos
rítmicos de la cola. En tierra, particularmente cuando escapan hacia
el agua ante la presencia de algún peligro, los crocodilios se
desplazan por medio de un rápido y poco elegante movimiento de
gateo. Cuando no hay apuro, también pueden caminar lentamente, a
cuatro patas, como los mamíferos.
No existen estudios detallados para cada una de las especies,
pero se conoce desde hace mucho tiempo que los crocodilios
pueden permanecer sumergidos durante largo tiempo. Cott (1961),
por ejemplo, aportó información de cocodrilos del Nilo adultos que
permanecieron por más de una hora bajo el agua sin salir a
respirar. Dicho autor, en un cruel experimento de acuerdo con las
normas de investigación actuales, mantuvo sumergidos de manera
forzada a cocodrilos pequeños por largos periodos. Algunos sobre-
vivieron más de media hora y uno de ellos (de 79 cm de longitud
total) hasta 57 minutos. En otro experimento con individuos
pequeños (entre 29 y 99 cm de longitud total), en los que no se
forzó a los animales a sumergirse se obtuvieron tiempos máximos
de 35 y 44 minutos, en los animales más grandes. Seebacher et al.
(2005) aportan datos de dos juveniles de Crocodylus johnsoni (de
9,2 y 14,8 kg) que permanecieron sumergidos casi dos horas. Para
las especies de Crocodylia de Venezuela, Gorzula (1978) reportó el
caso de una baba (no indicó la talla) que permaneció sumergida
durante 1 hora y 20 minutos.
Los crocodilios pasan parte de su tiempo fuera del agua. La
frecuencia, duración y momento en que lo hacen depende de la
especie, la latitud, la época del año, el tipo de ambiente que ocupan
y otros factores. Allí pueden permanecer por largos períodos con la
boca abierta, una conducta que ha sido asociada con la regulación
de la temperatura corporal (termorregulación). Este comportamiento
podría alterar la tasa de intercambio de calor de la cabeza con
respecto al cuerpo, por la exposición de las membranas húmedas
de la boca al aire, lo que produce enfriamiento por evaporación
(Spotila y Terpin, 1977; Mazzotti, 1989).

Los Crocodylia de Venezuela
15
En algunos aspectos de la anatomía interna y comportamiento,
los crocodilios se asemejan a las aves, sus parientes más cercanos.
Tienen un corazón con cuatro cámaras, son ovíparos y atienden el
nacimiento de sus crías, a las que protegen durante varias semanas
después de la eclosión. Al contrario de las aves, no poseen
mecanismos de control de la temperatura corporal, por lo que ésta
fluctúa con la del ambiente (poikilotermia). Sin embargo, por medio
del comportamiento, los crocodilios tienden a mantener una tem-
peratura corporal más o menos constante, exponiéndose al sol,
protegiéndose en la sombra o sumergiéndose en el agua de acuerdo
con su conveniencia. El comportamiento de termorregulación está
influenciado por el tamaño del individuo, su edad, su estado de
salud, el proceso de digestión de alimentos, las características del
ambiente que ocupa y otros factores.
REPRODUCCIÓN Y DESARROLLO
Los machos de los crocodilios son más grandes que las hembras
y durante la época reproductiva defienden territorios donde habitan
varias hembras. Los adultos muestran un elaborado ritual de cortejo
previo al apareamiento que incluye elementos visuales, acústicos y
táctiles, muy similares en todas las especies estudiadas
(Thorbjarnarson y Hernández, 1993b [caimán del Orinoco], Staton y
Dixon, 1977 y Ayarzagüena y Castroviejo, 2008 [baba]). La cópula,
que puede ocurrir repetidas veces durante la época reproductiva,
tiene lugar en el agua. Una vez finalizada la época de cortejo y
apareamiento la hembra comienza la preparación del nido.
Las hembras de los crocodilios ponen huevos cuyo número y
tamaño depende de la especie. En general se puede afirmar que
aquellas más grandes ponen mayor número de huevos, y de mayor
tamaño, que las especies pequeñas. Una vez depositados los
huevos, las hembras defienden el nido contra depredadores.
La forma de construir los nidos depende del hábitat y la época
en que se reproduce cada especie. Las que viven en ambientes
pantanosos y que se reproducen principalmente durante la época de
lluvias encierran los huevos en nidos en forma de montículos,

Andrés Eloy Seijas
16
formados en zonas no anegadas con material vegetal y suelo de los
alrededores. Las especies que habitan principalmente ríos, cons-
truyen nidos que consisten en simples excavaciones en las playas
arenosas donde depositan los huevos, que luego cubren con arena.
Los crocodilios no poseen cromosomas sexuales. El sexo de los
individuos está determinado por la temperatura a la cual se incuban
los huevos (Deeming y Ferguson 1989; Lang y Andrews, 1994).
Aunque existen pequeñas variaciones entre las especies, en aque-
llas que han sido estudiadas se ha demostrado que temperaturas de
incubación mayores a los 32 ºC producen machos y menores a 31 ºC
producen hembras. La exposición prolongada a temperaturas de
incubación por encima de 34 ºC o por debajo de 30 ºC puede ser
letal o provocar el nacimiento de crías con defectos congénitos
(Hutton y Webb, 1992).
Las hembras atienden el nacimiento de las crías y las defienden
por periodos que pueden prolongarse por varios meses en algunas
especies. En la etapa de huevos y primeros estadios de edad, los
individuos son presa de una amplia variedad de animales: garzas,
buitres, mamíferos carnívoros, lagartos depredadores, peces carní-
voros, serpientes, e, incluso, individuos mayores de su propia
especie. En la etapa adulta, en cambio, los crocodilios tienen pocos
enemigos naturales.
DIFERENCIAS ENTRE SEXOS
En general es difícil determinar el sexo de un crocodilio por sólo
su apariencia externa. Machos y hembras son prácticamente
indistinguibles, principalmente en la etapa de crías y juveniles. No
obstante, los machos alcanzan tallas bastante más grandes que las
hembras. En el caso de la baba en los llanos de Venezuela, por
ejemplo, las hembras raramente superan 1,80 m de longitud total,
mientras que los machos pueden exceder 2,40 m.
Los crocodilios machos poseen un pene (Fig. 1.6), a diferencia
de los lagartos y serpientes, que poseen dos hemipenes. La forma
más confiable de determinar el sexo de un individuo es examinar
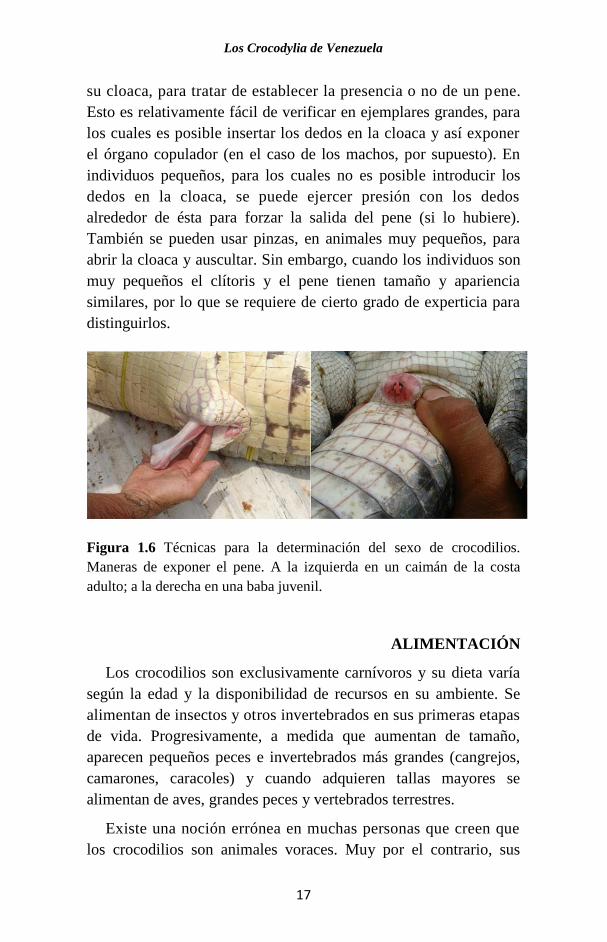
Los Crocodylia de Venezuela
17
su cloaca, para tratar de establecer la presencia o no de un pene.
Esto es relativamente fácil de verificar en ejemplares grandes, para
los cuales es posible insertar los dedos en la cloaca y así exponer
el órgano copulador (en el caso de los machos, por supuesto). En
individuos pequeños, para los cuales no es posible introducir los
dedos en la cloaca, se puede ejercer presión con los dedos
alrededor de ésta para forzar la salida del pene (si lo hubiere).
También se pueden usar pinzas, en animales muy pequeños, para
abrir la cloaca y auscultar. Sin embargo, cuando los individuos son
muy pequeños el clítoris y el pene tienen tamaño y apariencia
similares, por lo que se requiere de cierto grado de experticia para
distinguirlos.
Figura 1.6 Técnicas para la determinación del sexo de crocodilios.
Maneras de exponer el pene. A la izquierda en un caimán de la costa
adulto; a la derecha en una baba juvenil.
ALIMENTACIÓN
Los crocodilios son exclusivamente carnívoros y su dieta varía
según la edad y la disponibilidad de recursos en su ambiente. Se
alimentan de insectos y otros invertebrados en sus primeras etapas
de vida. Progresivamente, a medida que aumentan de tamaño,
aparecen pequeños peces e invertebrados más grandes (cangrejos,
camarones, caracoles) y cuando adquieren tallas mayores se
alimentan de aves, grandes peces y vertebrados terrestres.
Existe una noción errónea en muchas personas que creen que
los crocodilios son animales voraces. Muy por el contrario, sus

Andrés Eloy Seijas
18
requerimientos alimentarios son muy modestos si se comparan con
los de los mamíferos o las aves. En sus etapas tempranas, por
ejemplo, un cocodrilo requiere de 2 kg de alimento para incre-
mentar su peso en 1 kg (es decir, una eficiencia de 50%). El
ganado vacuno, las ovejas o los cerdos tendrían que comer
proporcionalmente de tres a cinco veces más para obtener la
misma ganancia en peso (NRC, 1983). En su ya clásico estudio
Cott (1961) reportó que cocodrilos del Nilo de mediano tamaño
(2,30 m y aproximadamente 45 kg de peso) consumieron en 36
días un promedio de 11,6 kg de alimento. En contraste, un pelícano,
ave con un peso de 8 kg, un poco menos de la quinta parte que la
de los cocodrilos mencionados, consumió 2,8 kg de pescado diario.
Es decir, extrapolando, el ave comería en los 36 días más de 100 kg
de alimento, nueve veces más que los cocodrilos de menor peso.
LONGEVIDAD
Los crocodilios son animales que pueden vivir muchos años,
pero no existen datos que permitan determinar la edad máxima
alcanzada por animales silvestres. Las referencias más confiables
sobre longevidad de los crocodilios provienen de individuos man-
tenidos en cautiverio. Un cocodrilo (especie no especificada) murió
a la edad de 110-115 años en un zoológico de Rusia en 1995
(Gorzula, 1995). Ese mismo año se reportó la muerte de un
Crocodylus porosus de 5,18 m en la granja de St. Augustine, en
el estado de Florida, EUA. La edad de ese cocodrilo se estimó,
con base en referencias sobre fecha y localidad de captura un
tanto imprecisas, en 70-80 años (http://www.flmnh.ufl.edu/cnhc/cbd-
faq-q3.htm). Pope (1960, citado por Coulson y Hernández, 1983)
aportó información sobre longevidad de crocodilios en zoológicos de
EUA, y mencionó a un Alligator mississippiensis que vivió 56
años y un A. sinensis que murió a los 50.
No se conoce con exactitud la edad de algunos grandes
caimanes mantenidos en cautiverio en Venezuela. Quizás el reporte
más confiable es el de la famosa caimana de José Faoro, comer-
ciante de origen italiano residenciado en San Fernando que participó

Los Crocodylia de Venezuela
19
en el negocio del comercio de pieles de caimanes del Orinoco.
Según datos recogidos por John Thorbjarnarson “La Negra”, como
denominaban a ese ejemplar, llegó a las manos de Faoro recién
nacida en 1922 y murió, sobreviviendo a su dueño, en 1992, a los
70 años de edad.
La edad de los caimanes adultos mantenidos en los zoocriaderos
de Venezuela en la mayoría de los casos sólo puede ser estimada.
Una excepción lo constituyen las caimanas llamadas colectivamente
como “chacachacares” del Fundo Pecuario Masaguaral. Esas
hembras provienen de una nidada colectada el 28 de febrero de
1978 por Robert Godshalk y Evelio Sosa en el río Cojedes
(Godshalk, 1978). Ellas nacieron el 11 de junio de ese año, por lo
que cumplieron 33 años en el 2011. El caimán más viejo de de ese
centro de cría llegó a las manos de Tomas Blohm en 1974 (Blohm,
1982) cuando medía 3,50 m. Si se le estima, de manera con-
servadora, una edad de 25 años para ese momento, en la
actualidad (2011) tendría cerca de 65 años y, aparentemente, en
plenitud de vigor reproductivo.
Manonerio, el C. intermedius más viejo del zoocriadero de la
UNELLEZ, ingresó a dicho centro en 1985, con una talla de 3,48
m. La edad de este ejemplar al momento de su muerte (15 mayo
2011) no debe haber sido menor de 50 años. Su compañera de
encierro, la hembra llamada “La Catira” ingresó en este centro de
cría en 1981, con una talla de 2,76 m y, quizá, unos 15 años de
edad, por lo que tendría, por lo menos, 45 años. Ambos ejemplares
mantenían, hasta la estación reproductiva de 2010, el vigor repro-
ductivo intacto, como lo demuestra el hecho de que en los últimos
25 años La Catira sólo dejó de poner huevos fertilizados una vez,
el año 2003.
Los caimanes más viejos de los zoocriaderos de Puerto Miranda
(Fig. 1.7) y El Frío también deben superar los 50 años. Como
sucede con los otros centros de cría se desconoce la edad en que
estos ejemplares ingresaron a dichos establecimientos, por lo que la
edad asignada es un tanto especulativa.

Andrés Eloy Seijas
20
Joselo, el caimán más viejo de la Estación Biológica El Frío, fue
llevado por José Ayarzagüena a ese centro de cría en 1977.
Estuvo cautivo desde su nacimiento en Achaguas y medía 3,90 m
al momento del traslado. Ayarzagüena cree que tendría 18-20 años
en esa época, por lo que su edad para 2011 sería de unos 55 años.
Figura 1.7 (A) Caimán del Orinoco macho del zoocriadero de Puerto Miranda,
estado Guárico. (B) Manonerio, caimán más viejo de la UNELLEZ (alrededor
de 50 años). Como sucede con la mayoría de los ejemplares en los centros de
cría, sus edades son difíciles de establecer con precisión ya que se desconoce
la edad al momento de ingreso.

Los Crocodylia de Venezuela
21
De acuerdo con el recién fallecido Tomás Blohm, el C. acutus
macho en el zoocriadero de Masaguaral estuvo en la casa de una
familia de apellido Goetz, en Caracas. Una foto de 1954 muestra a
dicho caimán con una talla alrededor de un metro, por lo que Don
Tomás estimaba que debió haber nacido en 1952. Para 2011 este
animal contaría con unos 59 años.
Es posible que la edad máxima alcanzable esté relacionada con
el tamaño de las distintas especies y que aquellas más pequeñas
tiendan a vivir menos años que las más grandes. Sería de esperar,
por lo tanto, que la longevidad de la baba y de los Paleosuchus sea
menor que la de los caimanes. En todo caso, no se dispone de
datos que permitan corroborar esta hipótesis.
CLASIFICACIÓN DE LOS CROCODYLIA
Tradicionalmente, siguiendo el sistema de Linneo, al Orden
Crocodylia se la ha dividido en tres familias: Gavialidae, Alli-
gatoridae y Crocodylidae y ese es el sistema de clasificación que
se seguirá en este libro (King y Burke, 1989). No obstante, hay
autores que consideran a los Crocodylia como una familia cons-
tituida por tres subfamilias: Gavialinae, Alligatorinae y Crocodylinae
(ver, por ejemplo, Ross y Magnusson, 1989).
Las relaciones filogenéticas entre las familias son todavía objeto de
controversia, así como la ubicación de Tomistoma schlegelii (entre los
gaviálidos o los Crocodílidos). En la Figura 1.8, se muestran dos
propuestas contradictorias entre las diversas que es posible encontrar
en la literatura. No es el objetivo de este libro tomar partido por
ninguna de ellas. Los lectores interesados pueden consultar los trabajos
en Grigg et al. (2001), así como otros artículos recientes (Harshman et
al., 2003; McAliley et al., 2006; Roos et al., 2007). Los rasgos
físicos principales que permiten distinguir a las especies de las tres
familias señaladas se resumen en la Figura 1.9.
En los Crocodylidae los dientes de las mandíbulas se entrelazan
cuando la boca está cerrada, por lo que quedan parcialmente a la

Andrés Eloy Seijas
22
vista. El cuarto diente de cada lado de la mandíbula inferior
sobresale al quedar sobre una escotadura lateral de la mandíbula
superior. La sínfisis mandibular, es decir, la unión de las dos piezas
de la mandíbula, se extiende al menos hasta el quinto diente.
Poseen Órganos Sensoriales Intertegumentarios (OSI) en las esca-
mas ventrales. Esas estructuras son papilas osmoreceptoras que
responden a cambios en la presión osmótica del agua y permiten a
los cocodrilos discriminar entre distintas salinidades (Jackson y
Brooks, 2007).
Figura 1.8 Genealogía de los Crocodylia propuesta por Norell (1989), a la
izquierda y Pough et al., 2004, a la derecha.
En los Alligatoridae, los dientes de la mandíbula inferior no son
visibles cuando el animal cierra la boca, ya que permanecen por
detrás de los dientes del maxilar superior. Por esta misma razón, el
cuarto diente de la mandíbula inferior, que es el que adquiere el
mayor desarrollo, queda oculto al encajar dentro de una oquedad de
la mandíbula superior. Además, no poseen OSI en las escamas
ventrales y la sínfisis mandibular generalmente no llega hasta el
quinto diente.
Los Gavialidae, que en la actualidad están representados por
una sola especie, Gavialis gangeticus, se caracterizan por poseer
un hocico muy largo, con una sínfisis mandibular que se extiende
entre los dientes 23 y 24. El hueso nasal, al contrario que en las
otras familias, no se conecta con el premaxilar. Esta última especie

Los Crocodylia de Venezuela
23
pudiera no estar sola en la familia, ya que con base en estudios
genéticos (Harshman et al., 2003) el falso gavial Tomistoma
schlegelii ha sido considerado como filogenéticamente más cercano
a Gavialis que a los Crocodylidae, familia en que ha sido
generalmente ubicado.
Figura 1.9 Clasificación de los Crocodylia vivientes. Se destacan algunos de
los caracteres que permiten diferenciar a las familias. Los OSI son pequeñas
papilas en las escamas ventrales. Dibujos de las cabezas tomados de CITES
(1995).
Existen también diferencias fisiológicas entre las familias. Los
Crocodylidae, al contrario de los Alligatoridae, poseen glándulas
especializadas en la lengua para la excreción de sales y
mecanismos desarrollados para conservar el agua del cuerpo
durante la eliminación de desechos metabólicos (Leslie y Taplin,
2001). Esto explicaría por qué varias especies de Crocodylidae
son capaces de vivir en ambientes de agua salada, e incluso
C r
o c
o d
y l i a
Gavialidae
Familias
Alligatoridae
El diente No. 4 del hueso dentario se
introduce en un alveolo de la sutura
premaxilo-maxilar. Queda oculto
cuando la boca está cerrada.
El diente No. 4 del hueso maxilar es el
más desarrollado. No posee Órganos
Sensoriales Intertegumentarios (OSI)
en las escamas ventrales.
4to diente
oculto
4to diente maxilar
más desarrolado
Crocodylidae
El diente No. 4 del hueso dentario
encaja en una escotadura lateral
del maxilar superior. Es visible
cuando la boca está cerrada.
El diente No. 5 del hueso maxilar
es el más desarrollado. Posee
OSI en escamas ventrales.
4to diente
visible
5to diente maxilar
más desarrolado
Hocico muy angosto y largo. Al
contrario de las otras familias el
hueso nasal no hace contacto con
los premaxilares. Posee OSI en
escamas ventrales
C r
o c
o d
y l i a
Gavialidae
Familias
Alligatoridae
El diente No. 4 del hueso dentario se
introduce en un alveolo de la sutura
premaxilo-maxilar. Queda oculto
cuando la boca está cerrada.
El diente No. 4 del hueso maxilar es el
más desarrollado. No posee Órganos
Sensoriales Intertegumentarios (OSI)
en las escamas ventrales.
4to diente
oculto
4to diente maxilar
más desarrolado
Alligatoridae
El diente No. 4 del hueso dentario se
introduce en un alveolo de la sutura
premaxilo-maxilar. Queda oculto
cuando la boca está cerrada.
El diente No. 4 del hueso maxilar es el
más desarrollado. No posee Órganos
Sensoriales Intertegumentarios (OSI)
en las escamas ventrales.
4to diente
oculto
4to diente maxilar
más desarrolado
Crocodylidae
El diente No. 4 del hueso dentario
encaja en una escotadura lateral
del maxilar superior. Es visible
cuando la boca está cerrada.
El diente No. 5 del hueso maxilar
es el más desarrollado. Posee
OSI en escamas ventrales.
4to diente
visible
5to diente maxilar
más desarrolado
Crocodylidae
El diente No. 4 del hueso dentario
encaja en una escotadura lateral
del maxilar superior. Es visible
cuando la boca está cerrada.
El diente No. 5 del hueso maxilar
es el más desarrollado. Posee
OSI en escamas ventrales.
4to diente
visible
5to diente maxilar
más desarrolado
Hocico muy angosto y largo. Al
contrario de las otras familias el
hueso nasal no hace contacto con
los premaxilares. Posee OSI en
escamas ventrales

Andrés Eloy Seijas
24
hipersalina, mientras que los Alligatoridae sólo pueden vivir en esos
ambientes de manera temporal o cuando el acceso a localidades
cercanas de agua dulce está garantizado.
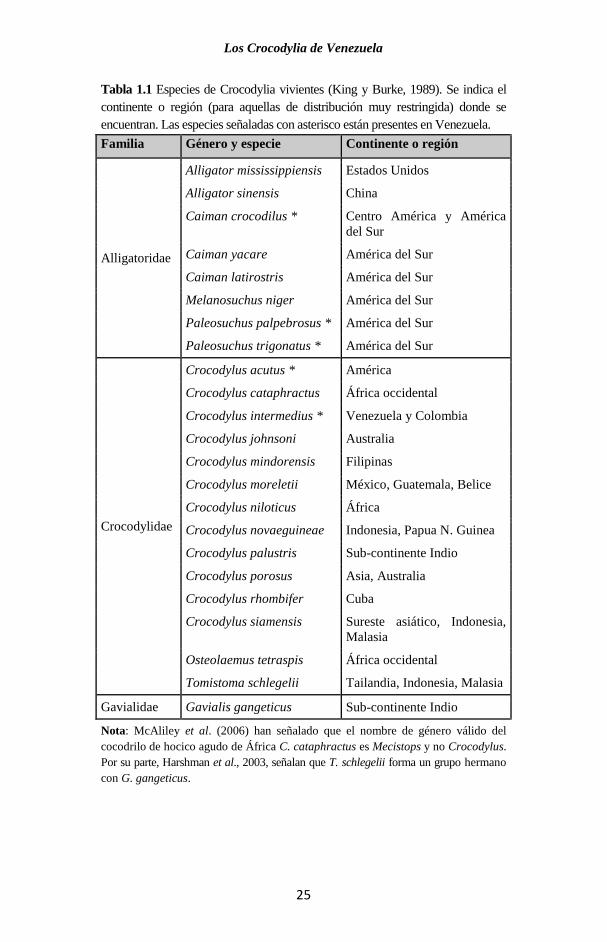
En la actualidad se conocen 23 especies de crocodilios vivientes
(Tabla 1.1), aunque hay discrepancia en esta cifra, ya que algunos
autores le dan rango de especies completas a los que otros
consideran sólo subespecies.
Este libro trata acerca de las especies de Crocodylia presentes
en Venezuela, por lo que se sugiere a lectores que quieran conocer
más detalles sobre este grupo o sobre especies de otras regiones,
revisar la literatura citada y las lecturas recomendadas.
BIBLIOGRAFIA RECOMENDADA
BELLAIRS, A. D´A.
1987. The Crocodylia. Pp. 5-7 in: Webb, G. J. W, C. Manolis y P. Whitehead.
(Eds.). Wildlife Management: Crocodiles and Alligators. Surrey Beatty
and Sons Pty Limited in Association with the Conservation Commission of
the Northern Territory. Chipping Norton, NSW, Australia.
GRIGG, G., F. SEEBACHER Y C. E. FRANKLIN. (EDS.).
2001. Crocodilian biology and evolution. Surrey Beatty and Sons PTY Limited,
Chipping Norton, NSW, Australia.
MEDEM, F.
1981. Los Crocodylia de Sur América, Vol. I. Los Crocodylia de Colombia.
Editorial Carrera 7a. Ltda, Bogotá. 354 pp.
ROSS, C. A.
1989. Crocodiles and Alligators. Golden Press Pty Ltd. Silverwater, NSW,
Australia.
Los Crocodylia de Venezuela
25
Tabla 1.1 Especies de Crocodylia vivientes (King y Burke, 1989). Se indica el
continente o región (para aquellas de distribución muy restringida) donde se
encuentran. Las especies señaladas con asterisco están presentes en Venezuela.
Familia Género y especie Continente o región
Alligatoridae
Alligator mississippiensis Estados Unidos
Alligator sinensis China
Caiman crocodilus * Centro América y América
del Sur
Caiman yacare América del Sur
Caiman latirostris América del Sur
Melanosuchus niger América del Sur
Paleosuchus palpebrosus * América del Sur
Paleosuchus trigonatus * América del Sur
Crocodylidae
Crocodylus acutus * América
Crocodylus cataphractus África occidental
Crocodylus intermedius * Venezuela y Colombia
Crocodylus johnsoni Australia
Crocodylus mindorensis Filipinas
Crocodylus moreletii México, Guatemala, Belice
Crocodylus niloticus África
Crocodylus novaeguineae Indonesia, Papua N. Guinea
Crocodylus palustris Sub-continente Indio
Crocodylus porosus Asia, Australia
Crocodylus rhombifer Cuba
Crocodylus siamensis Sureste asiático, Indonesia,
Malasia
Osteolaemus tetraspis África occidental
Tomistoma schlegelii Tailandia, Indonesia, Malasia
Gavialidae Gavialis gangeticus Sub-continente Indio
Nota: McAliley et al. (2006) han señalado que el nombre de género válido del
cocodrilo de hocico agudo de África C. cataphractus es Mecistops y no Crocodylus.
Por su parte, Harshman et al., 2003, señalan que T. schlegelii forma un grupo hermano
con G. gangeticus.

Andrés Eloy Seijas
26

Los Crocodylia de Venezuela
27
CAPÍTULO 2
LOS CROCODYLIA DE VENEZUELA
En la introducción ya se mencionaron las cinco especies de
Crocodylia presentes en Venezuela. Para referirse a este grupo se
usará el término “crocodilios”. Para las familias se emplearán las
expresiones aligatóridos y crocodílidos. A lo largo del libro se
usaran, en la medida de lo posible, los nombres vernáculos o
comunes con que se conocen a estas especies en el país, todos
ellos ya bien arraigados en la literatura técnica.
El término “caimán” se usará para referirse tanto a Crocodylus
intermedius como a Crocodylus acutus. Los añadidos “del
Orinoco” y “de la costa” que se usan en la literatura especializada
en Venezuela para separar a estas especies son relativamente
recientes y ya están bien arraigados en el habla popular. La
palabra “caimán” es de origen caribe, lo que explica que se use
ampliamente en países como Colombia, Panamá, República Domi-
nicana y Cuba para referirse a C. acutus. Los caimanes de
Venezuela son verdaderos cocodrilos, por lo cual también pueden
ser llamados de esa manera. En la literatura en inglés se usan los
nombres de “Orinoco crocodile” y “American crocodile” para
estas especies.
El término “baba” o “babo” se usará para señalar a Caiman
crocodilus. A esta especie se le denomina babilla en la parte
occidental de la cuenca del lago de Maracaibo, probablemente por
influencia colombiana, donde a este crocodilio se conoce con dicho
nombre. Las expresiones más frecuentes para mencionar a la baba
en inglés son “Spectacled caiman” y “Common caiman”.

Andrés Eloy Seijas
28
La coincidencia entre el nombre científico de la baba y el
nombre común de los Crocodylus es, en la actualidad, causa de
gran confusión. Los documentales y programas de entretenimiento
que transmiten a diario por la televisión, y que han sido origi-
nalmente producidos en inglés, hablan de “caimán” cuando se
refieren a la baba, al hacer una traducción textual del nombre que
se le da a este animal en esa lengua. La confusión se ha extendido
a tal punto que el diccionario de la RAE (Vigésima Segunda
Edición) dice que la palabra “caimán” viene del taíno kaiman, pero
por la descripción que hacen del animal se refieren, sin duda, a una
baba. Ahora bien, los taínos eran los aborígenes de las islas de
Cuba y La Española, lugares donde los únicos crocodilios autóc-
tonos son del género Crocodylus (C. acutus y C. rhombifer). No es
posible que los taínos le hayan dado nombre a un animal que no
conocían (la baba, en este caso).
Hay razones culturales y de identidad para insistir en el uso del
término “caimán” al hablar de los cocodrilos de Venezuela y del
Caribe. En todos los países que abarca el área de distribución
histórica de C. acutus y C. intermedius, existen numerosos
topónimos como “Punta caimán”, “Paso de caimán”, “Puerto
caimán” o, simplemente, “Caimán”. Muchos de esos nombres de
lugares son muy antiguos, con seguridad anteriores al momento en
que Johann Baptist Ritter von Spix, en 1825, tuviera la ocurrencia
de usar ese nombre como género para un Aligatórido. Los luga-
reños que dieron nombres a esos sitios, con seguridad se referían a
cocodrilos. Por otra parte, numerosos dichos y refranes mencionan
al caimán, con indiscutible referencia a los cocodrilos. Canciones
conocidísimas como la del “caimán que se va para Barranquilla” o
la del “caimán de más de una cuadra” que señala Simón Díaz, se
refieren también, sin duda a los grandes cocodrilos.
Por otro lado, el caimán conocido como “el espanto del
Bramador” que Pajarote y María Nieves, los personajes de la
novela Doña Bárbara de Rómulo Gallegos, matan en un caño de
Apure no era, en absoluto, una baba. En la versión al inglés de la
mencionada novela, traducida en 1948 por Robert Malloy, usan la
expresión “alligator” por caimán y “alligator hunting” por

Los Crocodylia de Venezuela
29
cacería de caimanes, lo cual es poco menos que una aberración; lo
correcto hubiera sido “crocodile” y “crocodile hunting”, respec-
tivamente.
La fuerza poderosa de la televisión obligará, en un futuro no
muy lejano, a usar la expresión “cocodrilo” al mencionar exclu-
sivamente a las especies del género Crocodylus, pero no será este
libro el que contribuya a acelerar ese proceso de pérdida cultural.
Los Paleosuchus son tan poco conocidos que no existen nom-
bres comunes de amplio uso con que referirse a ellos, como no
sean las denominaciones, por lo demás poco difundidas, que puedan
recibir de pueblos indígenas. La etnia Yaruro, por ejemplo, que
habita las márgenes del río Capanaparo, usa el nombre “aricué”
para la especie P. palpebrosus. La literatura científica ha impuesto
la expresión “babo morichalero” con que se refieren a esa especie,
según Godshalk (1982), algunos habitantes del sur de Apure.
También se usa la expresión “babo(a) negro(a)” la cual se usará
en este libro exclusivamente para P. trigonatus. En inglés se usan
las expresiones de “dwarf caiman” y “smooth fronted caiman”
para referirse a estas especies, así como “Cuvier´s smooth-
fronted caiman” para señalar a P. palpebrosus y “Schneider´s
smooth-fronted caiman” para P. trigonatus.
Para facilitar la descripción de las características morfológicas
de las especies de Venezuela, se presentan los dibujos de la Figura
2.1. Como fuente para la descripción de la escamación, se consultó
principalmente a Brazaitis (1973), Medem (1983) y CITES (1995),
así como anotaciones propias del autor.
SINOPSIS SOBRE LAS ESPECIES DE CROCODYLIA DE
VENEZUELA
La Baba (Caiman crocodilus)
Es probablemente la especie de Crocodylia más abundante del
mundo. Es, asimismo, una de las de más amplia distribución

Andrés Eloy Seijas
30
geográfica. Se encuentra desde la latitud 16º 20’ N en México
hasta la latitud 34º 00’ S en Paraguay y Argentina (Gorzula y
Seijas, 1989) aunque la mayoría de los autores señalan que las
poblaciones más al sur (Bolivia, Paraguay, Argentina) pertenecen a
Caiman yacare. La baba habita también en la isla de Trinidad y ha
establecido poblaciones al sur de la península de la Florida
(EE.UU), en Cuba y en Puerto Rico (Varona, 1976; Ellis, 1980;
Watlington, 2002).
Se han descrito distintas subespecies de C. crocodilus, de las
cuales C. c. crocodilus sería la de más amplia distribución en
Venezuela, mientras que C. c. fuscus estaría restringida en este
país a la cuenca del lago de Maracaibo y los ríos que drenan hacia
la costa de los estados Zulia, Falcón, Yaracuy y Carabobo
(Medem, 1981; Gorzula y Seijas, 1989). Prácticamente, no existen
estudios que apoyen con datos esta supuesta separación geográfica.
Se requiere de investigaciones que determinen las diferencias o
semejanzas entre las poblaciones de babas del país y que dibujen
de manera más clara la distribución de las subespecies. Por
ejemplo, no se tienen información sobre las poblaciones de
babas de la región costera oriental del país y, por lo tanto, sobre sus
afinidades con algunas de las subespecies mencionadas. Las
localidades para la que existen estudios ecológicos de C. crocodilus
con algún grado de detalle sobre la abundancia y estructura
poblacional se muestran en la Figura 2.2.
Características físicas
La baba es un crocodilio de pequeño tamaño. La talla al nacer
está alrededor de 22-25 cm de longitud total (LT). Se ha señalado
una talla máxima para los machos de 2,50-2,80 m (Brazaitis, 1973;
Ayarzagüena, 1983; Thorbjarnarson, 1990), mientras que las
hembras generalmente no superan 1,80 m (o 0,90 m de longitud
cabeza-cloaca, LCC). Todas estas dimensiones corresponden a las
babas de los llanos. Las babas de la región noroccidental del país
son más pequeñas. La hembra más grande capturada en el Zulia
por Barros (2003), por ejemplo, midió aproximadamente 1,50 m de
LT (0,74 m de LCC).

Los Crocodylia de Venezuela
31
Figura 2.1 Nombre de los diferentes grupos de escamas que se usarán para
describir a cada uno de los crocodilios de Venezuela. Se muestra además la
forma como se disponen los Órganos Sensoriales Intertegumentarios (OSI) en
las escamas ventrales, presentes en los crocodílidos pero no en los aligató-
ridos. Dibujos tomados de CITES (1995).
Por ser un aligatórido, sólo los dientes de la mandíbula superior
son visibles cuando la baba tiene la boca cerrada y no posee
órganos sensoriales inter-tegumentarios (OSI) en las escamas
ventrales. Presenta un característico borde transversal anterior a los
ojos, que se extiende entre las órbitas oculares, lo cual ha
justificado el apelativo de “caimán de anteojos” (Spectacled
caiman, en inglés) con que se le señala en la literatura (Ayarza-
güena, 1983) (Fig. 2.3).
Escamación
Con base en una muestra de babas de la región costera (desde
la cuenca del lago de Maracaibo hasta Yaracuy) y de individuos de
Andrés Eloy Seijas
32
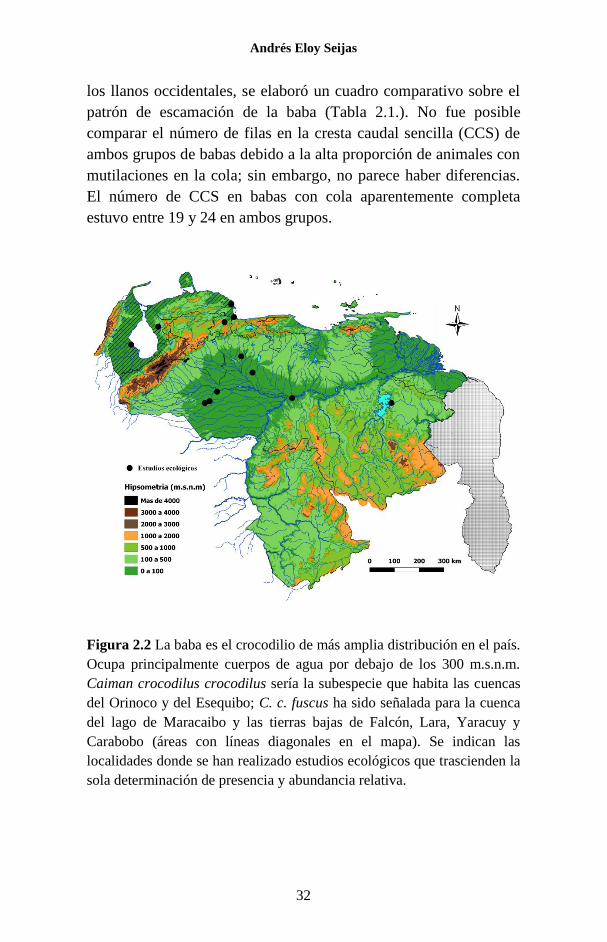
los llanos occidentales, se elaboró un cuadro comparativo sobre el
patrón de escamación de la baba (Tabla 2.1.). No fue posible
comparar el número de filas en la cresta caudal sencilla (CCS) de
ambos grupos de babas debido a la alta proporción de animales con
mutilaciones en la cola; sin embargo, no parece haber diferencias.
El número de CCS en babas con cola aparentemente completa
estuvo entre 19 y 24 en ambos grupos.
Figura 2.2 La baba es el crocodilio de más amplia distribución en el país.
Ocupa principalmente cuerpos de agua por debajo de los 300 m.s.n.m.
Caiman crocodilus crocodilus sería la subespecie que habita las cuencas
del Orinoco y del Esequibo; C. c. fuscus ha sido señalada para la cuenca
del lago de Maracaibo y las tierras bajas de Falcón, Lara, Yaracuy y
Carabobo (áreas con líneas diagonales en el mapa). Se indican las
localidades donde se han realizado estudios ecológicos que trascienden la
sola determinación de presencia y abundancia relativa.

Los Crocodylia de Venezuela
33
Figura 2.3 Cabeza de una baba donde se observa uno de los detalles más
distintivos de la especie: la arista entre los ojos. Esa característica es
suficiente para distinguirla de los otros crocodilios de Venezuela.
Ya se señaló que, de acuerdo con Donoso-Barros (1965) y
Medem (1983), las babas de la región costera, desde la desem-
bocadura del río Yaracuy hasta la cuenca del lago de Maracaibo,
pertenecerían a la subespecie fuscus, mientras que las de los llanos
pertenecerían a la subespecie crocodilus. Pese al desbalance en el
tamaño de la muestra analizada, los datos presentados en la Tabla
2.1, no permiten inferir que exista un rasgo que por sí solo permita,
de manera absoluta, ubicar a un ejemplar determinado en uno de
estos grupos de babas. No obstante, desde el punto de vista
estadístico se puede afirmar que, como grupo, las babas costeras
poseen menos filas de escamas dorsales y ventrales que las
llaneras, así como mayor número de CCD. Más adelante, en el

Andrés Eloy Seijas
34
Capítulo 4, se verá que las babas de la costa presentan un hocico
relativamente más ancho y corto que las de los llanos.
Hábitat
En Venezuela, la baba ocupa una amplia variedad de ambientes
acuáticos por debajo de 300 m.s.n.m pero hay registros de ejem-
plares localizados en cuerpos de agua ubicados a 800 m. de altitud
(Gorzula y Paolillo, 1986). Se ha señalado su presencia en ríos,
quebradas, madreviejas, lagos, planicies inundables, embalses, prés-
tamos y canales (Gorzula y Seijas, 1989). Es una especie esen-
cialmente dulceacuícola, pero que en ocasiones puede ocupar
ambientes de manglares con agua salobre. Durante el trabajo de
evaluación de las poblaciones del caimán de la costa en la región
costera de Venezuela, entre los años 1980-1986 (Seijas, 1986b) se
observaron babas en los Canales de río Chico, en aguas con
salinidad de 7-8 ppm y en Morrocoy, con salinidad de 25 ppm.
La distribución y abundancia actuales de la baba pueden
deberse a la expansión hacia zonas donde los grandes caimanes
(Crocodylus acutus y C. intermedius) fueron exterminados o
extremadamente reducidos en número debido a la cacería comercial
(Medem, 1981, 1983; Seijas, 1986b; Thorbjarnarson, 1990) y a la
creación de nuevos ambientes acuáticos para satisfacer necesidades
humanas (Rivero-Blanco, 1974; Seijas et al., 1989). En el primer
caso, la erradicación de los grandes cocodrilos debe haber signifi-
cado un alivio de la presión ejercida por esos formidables
competidores-depredadores que mantenían las poblaciones de C.
crocodilus reducidas. En el segundo caso, la construcción de
embalses, la modulación de sabanas, la interrupción de caños con
tapas y terraplenes para almacenar agua para el ganado y la creación
de líneas de préstamos a lo largo de las carreteras, ha puesto a la
disposición de esta especie nuevos hábitats que colonizar.
Reproducción
Como en todos los crocodilios, los machos de C. crocodilus
alcanzan mayor tamaño que las hembras. Este hecho produce una
Los Crocodylia de Venezuela
35
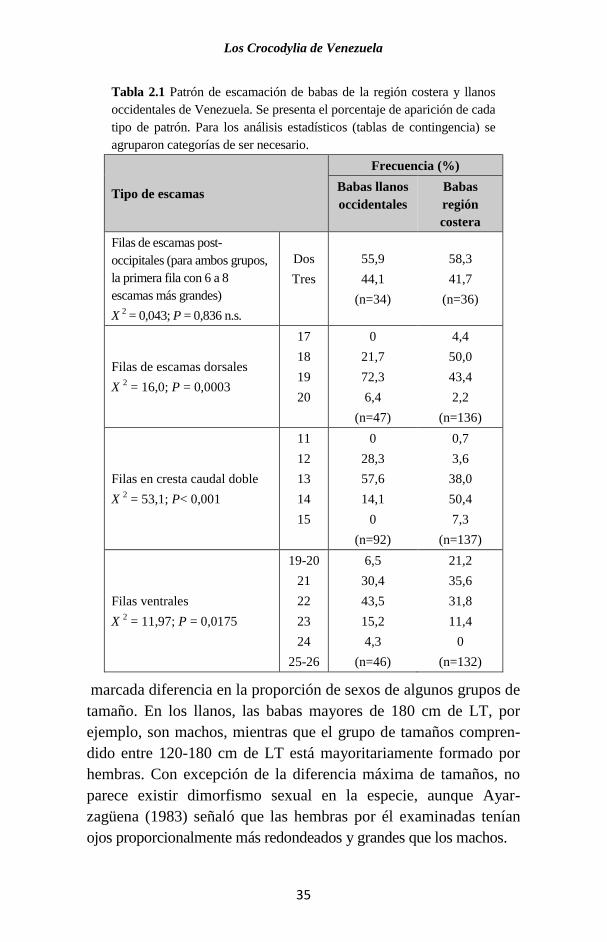
Tabla 2.1 Patrón de escamación de babas de la región costera y llanos
occidentales de Venezuela. Se presenta el porcentaje de aparición de cada
tipo de patrón. Para los análisis estadísticos (tablas de contingencia) se
agruparon categorías de ser necesario.
Tipo de escamas
Frecuencia (%)
Babas llanos
occidentales
Babas
región
costera
Filas de escamas post-
occipitales (para ambos grupos,
la primera fila con 6 a 8
escamas más grandes)
X 2 = 0,043; P = 0,836 n.s.
Dos
Tres
55,9
44,1
(n=34)
58,3
41,7
(n=36)
Filas de escamas dorsales
X 2 = 16,0; P = 0,0003
17
18
19
20
0
21,7
72,3
6,4
(n=47)
4,4
50,0
43,4
2,2
(n=136)
Filas en cresta caudal doble
X 2 = 53,1; P< 0,001
11
12
13
14
15
0
28,3
57,6
14,1
0
(n=92)
0,7
3,6
38,0
50,4
7,3
(n=137)
Filas ventrales
X 2 = 11,97; P = 0,0175
19-20
21
22
23
24
25-26
6,5
30,4
43,5
15,2
4,3
(n=46)
21,2
35,6
31,8
11,4
0
(n=132)
marcada diferencia en la proporción de sexos de algunos grupos de
tamaño. En los llanos, las babas mayores de 180 cm de LT, por
ejemplo, son machos, mientras que el grupo de tamaños compren-
dido entre 120-180 cm de LT está mayoritariamente formado por
hembras. Con excepción de la diferencia máxima de tamaños, no
parece existir dimorfismo sexual en la especie, aunque Ayar-
zagüena (1983) señaló que las hembras por él examinadas tenían
ojos proporcionalmente más redondeados y grandes que los machos.

Andrés Eloy Seijas
36
Rivero-Blanco (1974) y Pérez-Talavera (1999) reportaron hem-
bras reproductivamente activas con LT de 110 cm. Hembras
adultas con tallas menores que la señalada han sido reportadas o
sugeridas y pudiera tratarse de casos excepcionales o medidas de
individuos con la cola rota. Para evitar errores es preferible
presentar los datos de talla en LCC. La talla mínima reproductiva
encontrada por Staton y Dixon (1977) fue 67,7 cm de LCC, la
reportada por Ayarzagüena (1983) fue de 61,4 cm de LCC,
mientras que Thorbjarnarson (1990) indicó 60,0 cm de LCC. Una
hembra con esta última talla, capturada en el Módulo Experimental
de Mantecal (Apure) en 1978, mostró un desarrollo incipiente de
huevos (Seijas, 1979).
La cronología de anidación de C. crocodilus guarda una
estrecha relación con el régimen de lluvias de cada región. La
estrategia básica de reproducción en los llanos de Venezuela
consiste en anidar a mitad de la época lluviosa, de manera que las
crías nazcan al final de dicha estación o a comienzos del periodo
seco, cuando todavía permanecen muchos cuerpos de agua. Existen
pocos datos que permitan definir con claridad la cronología repro-
ductiva en regiones del país donde la estación de lluvias es muy
prolongada, como por ejemplo al sur del lago de Maracaibo o en
cuerpos de agua de zonas áridas con más de un pico de lluvias o
periodos lluviosos de menor duración, como ocurre en gran parte
de la región costera.
La baba construye sus nidos en lugares aledaños a los cuerpos
de agua, con materia vegetal viva o muerta, restos orgánicos en
descomposición y tierra que son arrastrados con las patas traseras
hasta formar un montículo. El material usado para construir el nido
depende de las características del área en que se encuentra la
baba. Hojas de gramíneas y otras plantas, pequeñas ramas,
fragmentos de corteza de árboles, entremezclado con el humus de
la capa superficial del suelo son los materiales más comunes de
encontrar.
En los llanos inundables la construcción de nidos comienza
cuando las aguas han alcanzado su máximo nivel. La temporada de
postura abarca agosto y septiembre en el hato El Frío, estado

Los Crocodylia de Venezuela
37
Apure (Ayarzagüena, 1983), de agosto a octubre en el hato
Masaguaral, estado Guárico (Staton y Dixon, 1977; Thorbjarnarson,
1990) y de septiembre a octubre en Caicara del Orinoco, estado
Bolívar (Pérez-Talavera, 1999). No existe información precisa sobre
la cronología de anidación de las babas en la región norte costera
de Venezuela. Es probable que el período de postura en la región
sur del lago de Maracaibo, con una muy corta estación seca, sea
mucho más amplio del que ocurre en los llanos. Barros (2003)
aportó información sobre tres nidadas en zonas aledañas a las
Ciénagas de Juan Manuel. Por el avance del estado de incubación
de los huevos observados, este último autor concluyó que el periodo
de desove ocurre entre julio y septiembre, coincidiendo con la
época de mayor precipitación en la zona. El comienzo de la
anidación podría iniciarse incluso antes, si tomamos en cuenta que
una hembra de esa misma localidad sacrificada en mayo de 1980
(Seijas, datos sin publicar), poseía huevos en formación en su oviducto.
El periodo de incubación dura de 70 a 75 días de acuerdo con
Staton y Dixon (1975). Thorbjarnarson (1990); sin embargo,
encontró que la duración promedio de este proceso, para 58 nidos
por él estudiados, fue de 81,8 días con pequeñas variaciones
dependiendo del tipo de microhábitat que ocupaban (ciénagas,
bosques, terraplenes o palmares). Dada la extensión del período de
apareamientos y anidación, la temporada de nacimiento de las crías de
babas en los llanos abarca desde mediados de octubre hasta diciem-
bre, con un pico en el mes de noviembre (Staton y Dixon, 1977).
Existe bastante información sobre el número de huevos por nido
(h/n). Rivero-Blanco (1974) señaló un promedio de 29,1 h/n
(n=164); Staton y Dixon (1977) 28,6 h/n (n= 30); Ayarzagüena
(1983) 30,4 h/n (n=45) y Pérez-Talavera (1999) encontró 29,6 h/n
(n=25). Velasco et al. (1996) reportaron promedios desde 26,6
hasta 32,4 para distintas regiones ecológicas de los llanos, con un
promedio global de 30,8 h/n (n=864).
En contraste con las cifras anteriores, Thorbjarnarson (1990)
reportó un promedio relativamente bajo de 22,2 h/n (n=154). Los
datos de este autor pueden ser comparados con los encontrados
por Rivero-Blanco (1974) (Fig. 2.4) quien trabajó en la misma
Andrés Eloy Seijas
38
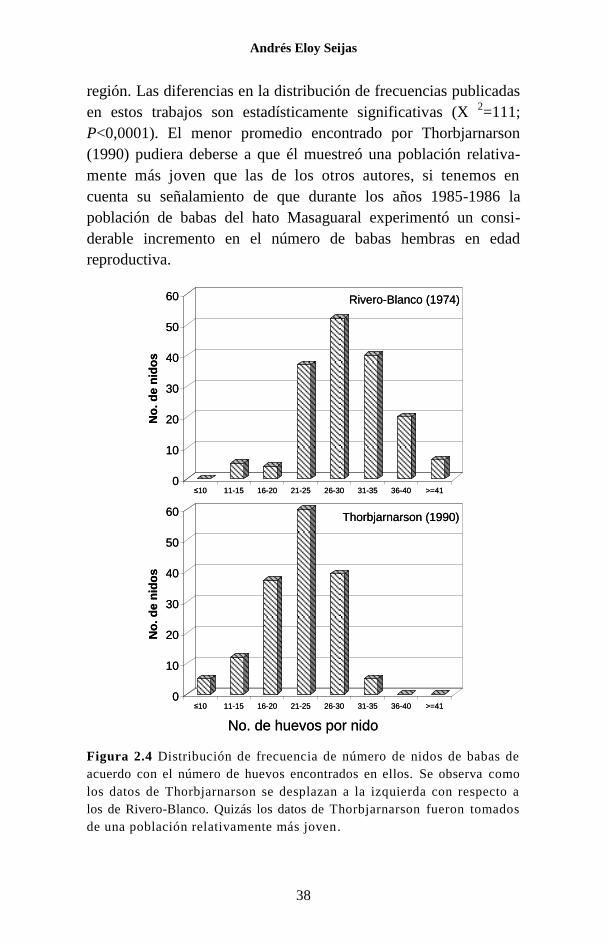
región. Las diferencias en la distribución de frecuencias publicadas
en estos trabajos son estadísticamente significativas (X 2=111;
P<0,0001). El menor promedio encontrado por Thorbjarnarson
(1990) pudiera deberse a que él muestreó una población relativa-
mente más joven que las de los otros autores, si tenemos en
cuenta su señalamiento de que durante los años 1985-1986 la
población de babas del hato Masaguaral experimentó un consi-
derable incremento en el número de babas hembras en edad
reproductiva.
Figura 2.4 Distribución de frecuencia de número de nidos de babas de
acuerdo con el número de huevos encontrados en ellos. Se observa como
los datos de Thorbjarnarson se desplazan a la izquierda con respecto a
los de Rivero-Blanco. Quizás los datos de Thorbjarnarson fueron tomados
de una población relativamente más joven.
0
10
20
30
40
50
60
No
. d
e n
ido
s
≤10 11-15 16-20 21-25 26-30 31-35 36-40 >=41
Rivero-Blanco (1974)
0
10
20
30
40
50
60
No
. d
e n
ido
s
≤10 11-15 16-20 21-25 26-30 31-35 36-40 >=41
Thorbjarnarson (1990)
No. de huevos por nido
0
10
20
30
40
50
60
No
. d
e n
ido
s
≤10 11-15 16-20 21-25 26-30 31-35 36-40 >=41
Rivero-Blanco (1974)
0
10
20
30
40
50
60
No
. d
e n
ido
s
≤10 11-15 16-20 21-25 26-30 31-35 36-40 >=41
Thorbjarnarson (1990)
No. de huevos por nido

Los Crocodylia de Venezuela
39
El menor número de huevos en un nido que ha sido reportado
es de cuatro (Thorbjarnarson, 1990) y el máximo 48 (Rivero-
Blanco, 1974). La diferencia en el tamaño de la nidada depende
del tamaño de la hembra, ya que hembras más grandes tienden a
poner mayor número de huevos (Thorbjarnarson, 1990).
El tamaño de la nidada pudiera depender también de las
características ecológicas de cada región. Velasco et al. (1996)
encontraron un promedio menor (26,6 h/n; n=137) en los llanos
boscosos y un promedio mayor (32,4 h/n; n=349) en el bajo Apure,
aunque no mencionaron la significación estadística de estas dife-
rencias. A pesar del menor promedio de huevos por nido, estos
autores señalaron que las zonas boscosas contienen mayor cantidad
de nidos que las sabanas abiertas, al disponer de microhábitats más
favorables para la anidación. Thorbjarnarson (1990) encontró que
el tipo de hábitat con mayor cantidad de nidos fue el de las sabanas
con palmas, con los bosques en un distante tercer lugar, después
de los médanos.
Respecto a las babas de la región noroccidental del país, sólo se
tienen los datos de Barros (2003), quien localizó tres nidadas en las
Ciénagas de Juan Manuel, estado Zulia, con 24, 29 y 30 huevos.
Los Paleosuchus
El género Paleosuchus posee dos especies: el babo morichalero
(P. palpebrosus) y el babo negro (P. trigonatus). Ambas se
encuentran en el país. Es tan poco lo que se ha investigado sobre
estas especies en Venezuela que gran parte de esta pequeña
sinopsis tiene que basarse en la información disponible para otros
países. Es sólo para los años 2008 y 2009 que se lleva a cabo la
primera investigación ecológica detallada para la primera de estas
especies, por parte de Andrés Pacheco y Gabriel González, quienes
trabajaron en un sector del sur del estado Anzoátegui.
El babo negro se localiza en las cuencas del Orinoco y del
Amazonas, así como en los ríos que drenan hacia el Atlántico entre

Andrés Eloy Seijas
40
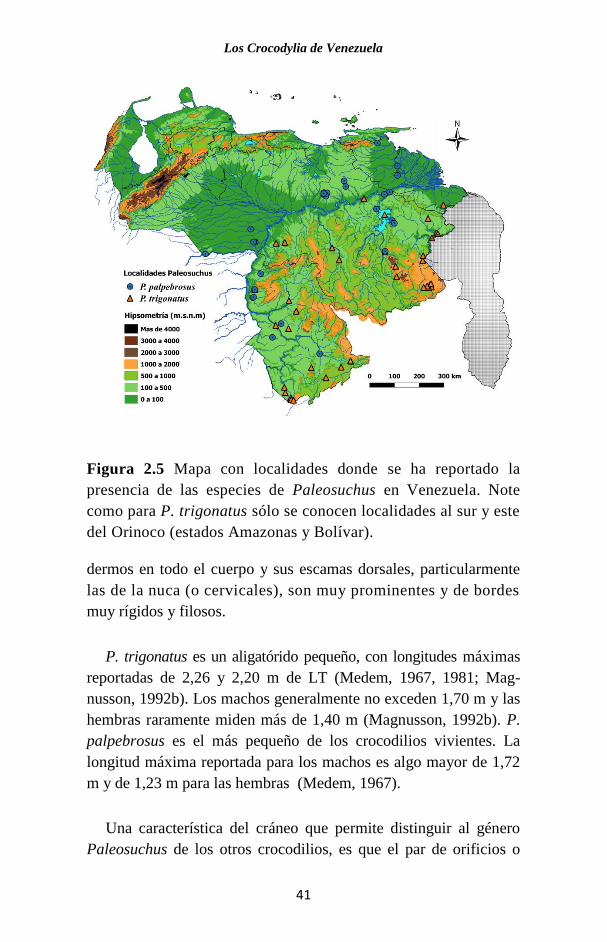
los grandes sistemas fluviales mencionados. De acuerdo con las
localidades reportadas de individuos conservados en museos, la
especie se localiza en Venezuela sólo al sur del Orinoco (Fig. 2.5)
(Seijas, 2007a). Existe un registro señalado por Godshalk (1982)
para el río Cinaruco. Desafortunadamente, este autor perdió los
ejemplares colectados, por lo que no es posible determinar si se
trató de un caso de identificación errónea (confundido con P.
palpebrosus). En un par de breves visitas al Cinaruco el 16 de
junio de 2001 y 18 de enero de 2002, se constató la presencia de
P. palpebrosus, pero no la de P. trigonatus.
El babo morichalero se encuentra en cuerpos de agua de las
tierras bajas de las cuencas del Orinoco y del Amazonas, así como
en los ríos que drenan hacia el Atlántico entre los grandes sistemas
fluviales mencionados, pero su distribución se extiende más al sur,
a lo largo del escudo brasileño, hasta las cuencas de los ríos
Paraná y Paraguay, así como en la cuenca del río San Francisco de
los estados de Bahía y Minas Gerais de dicho país (Magnusson,
1992a). El mapa de la Figura 2.5 se muestran los sitios donde la
especie ha sido reportada para Venezuela. Como se observa,
existen localidades tanto al norte como al sur del Orinoco. La
localidad más al norte de P. palpebrosus corresponde al río San
Juan en el estado Monagas (Donoso-Barros, 1966a), en una cuenca
no conectada directamente, en la actualidad, con el río Orinoco.
Características físicas
Los Paleosuchus, por ser aligatóridos, presentan un conjunto de
rasgos similares a los de la baba, entre los que vale la pena
destacar la ausencia de OSI en las escamas ventrales y la
disposición de los dientes. Las dos especies de este género son de
apariencia externa muy similar, por lo que su identificación no es
siempre sencilla. Se distinguen fácilmente de las babas por su
coloración más oscura (por lo que se les da en ocasiones el
nombre de “babos negros”) y por la ausencia de borde transversal
anterior a los ojos, entre las órbitas oculares (llamados por eso
“smooth-fronted caiman”, que prefiero traducir como babas de
frente lisa”). Poseen además un gran desarrollo de los osteo -
Los Crocodylia de Venezuela
41
Figura 2.5 Mapa con localidades donde se ha reportado la
presencia de las especies de Paleosuchus en Venezuela. Note
como para P. trigonatus sólo se conocen localidades al sur y este
del Orinoco (estados Amazonas y Bolívar).
dermos en todo el cuerpo y sus escamas dorsales, particularmente
las de la nuca (o cervicales), son muy prominentes y de bordes
muy rígidos y filosos.
P. trigonatus es un aligatórido pequeño, con longitudes máximas
reportadas de 2,26 y 2,20 m de LT (Medem, 1967, 1981; Mag-
nusson, 1992b). Los machos generalmente no exceden 1,70 m y las
hembras raramente miden más de 1,40 m (Magnusson, 1992b). P.
palpebrosus es el más pequeño de los crocodilios vivientes. La
longitud máxima reportada para los machos es algo mayor de 1,72
m y de 1,23 m para las hembras (Medem, 1967).
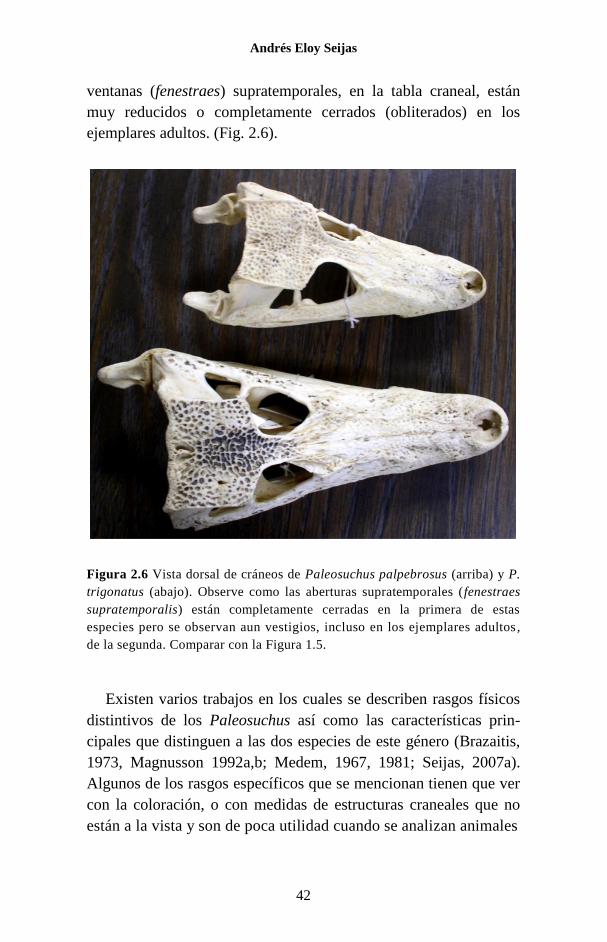
Una característica del cráneo que permite distinguir al género
Paleosuchus de los otros crocodilios, es que el par de orificios o
Andrés Eloy Seijas
42
ventanas (fenestraes) supratemporales, en la tabla craneal, están
muy reducidos o completamente cerrados (obliterados) en los
ejemplares adultos. (Fig. 2.6).
Figura 2.6 Vista dorsal de cráneos de Paleosuchus palpebrosus (arriba) y P.
trigonatus (abajo). Observe como las aberturas supratemporales (fenestraes
supratemporalis) están completamente cerradas en la primera de estas
especies pero se observan aun vestigios, incluso en los ejemplares adultos ,
de la segunda. Comparar con la Figura 1.5.
Existen varios trabajos en los cuales se describen rasgos físicos
distintivos de los Paleosuchus así como las características prin-
cipales que distinguen a las dos especies de este género (Brazaitis,
1973, Magnusson 1992a,b; Medem, 1967, 1981; Seijas, 2007a).
Algunos de los rasgos específicos que se mencionan tienen que ver
con la coloración, o con medidas de estructuras craneales que no
están a la vista y son de poca utilidad cuando se analizan animales
Los Crocodylia de Venezuela
43
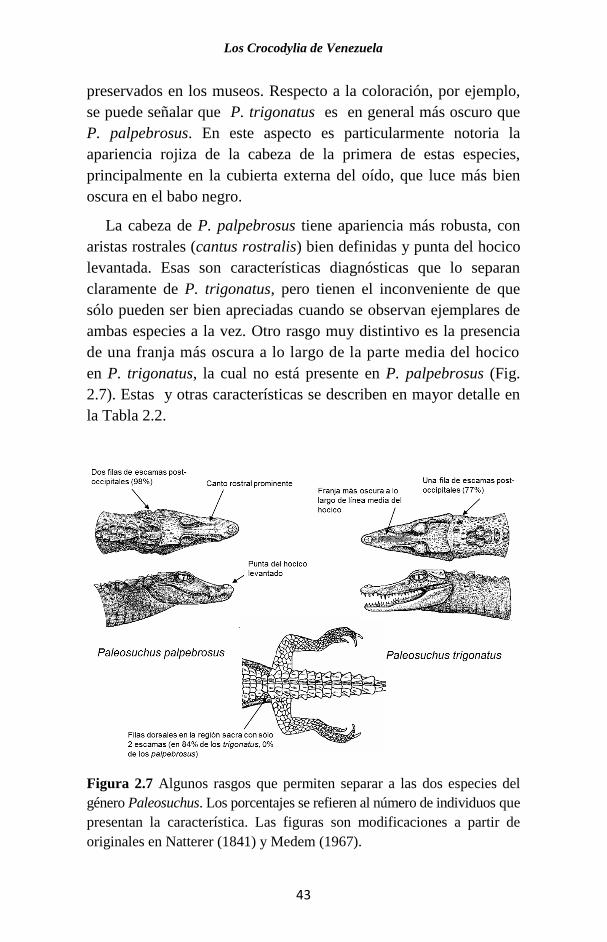
preservados en los museos. Respecto a la coloración, por ejemplo,
se puede señalar que P. trigonatus es en general más oscuro que
P. palpebrosus. En este aspecto es particularmente notoria la
apariencia rojiza de la cabeza de la primera de estas especies,
principalmente en la cubierta externa del oído, que luce más bien
oscura en el babo negro.
La cabeza de P. palpebrosus tiene apariencia más robusta, con
aristas rostrales (cantus rostralis) bien definidas y punta del hocico
levantada. Esas son características diagnósticas que lo separan
claramente de P. trigonatus, pero tienen el inconveniente de que
sólo pueden ser bien apreciadas cuando se observan ejemplares de
ambas especies a la vez. Otro rasgo muy distintivo es la presencia
de una franja más oscura a lo largo de la parte media del hocico
en P. trigonatus, la cual no está presente en P. palpebrosus (Fig.
2.7). Estas y otras características se describen en mayor detalle en
la Tabla 2.2.
Figura 2.7 Algunos rasgos que permiten separar a las dos especies del
género Paleosuchus. Los porcentajes se refieren al número de individuos que
presentan la característica. Las figuras son modificaciones a partir de
originales en Natterer (1841) y Medem (1967).
Andrés Eloy Seijas
44
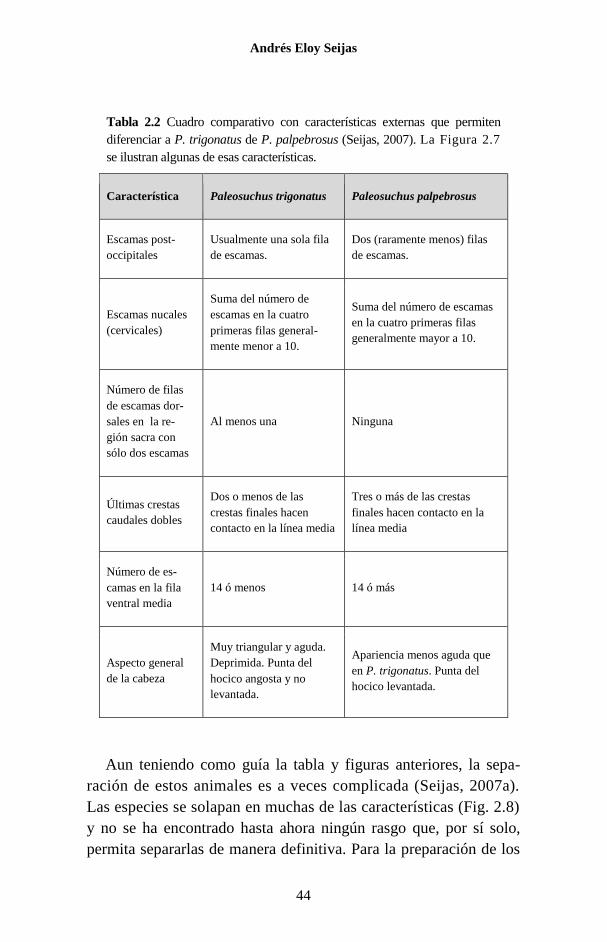
Tabla 2.2 Cuadro comparativo con características externas que permiten
diferenciar a P. trigonatus de P. palpebrosus (Seijas, 2007). La Figura 2.7
se ilustran algunas de esas características.
Característica Paleosuchus trigonatus Paleosuchus palpebrosus
Escamas post-
occipitales
Usualmente una sola fila
de escamas.
Dos (raramente menos) filas
de escamas.
Escamas nucales
(cervicales)
Suma del número de
escamas en la cuatro
primeras filas general-
mente menor a 10.
Suma del número de escamas
en la cuatro primeras filas
generalmente mayor a 10.
Número de filas
de escamas dor-
sales en la re-
gión sacra con
sólo dos escamas
Al menos una Ninguna
Últimas crestas
caudales dobles
Dos o menos de las
crestas finales hacen
contacto en la línea media
Tres o más de las crestas
finales hacen contacto en la
línea media
Número de es-
camas en la fila
ventral media
14 ó menos 14 ó más
Aspecto general
de la cabeza
Muy triangular y aguda.
Deprimida. Punta del
hocico angosta y no
levantada.
Apariencia menos aguda que
en P. trigonatus. Punta del
hocico levantada.
Aun teniendo como guía la tabla y figuras anteriores, la sepa-
ración de estos animales es a veces complicada (Seijas, 2007a).
Las especies se solapan en muchas de las características (Fig. 2.8)
y no se ha encontrado hasta ahora ningún rasgo que, por sí solo,
permita separarlas de manera definitiva. Para la preparación de los

Los Crocodylia de Venezuela
45
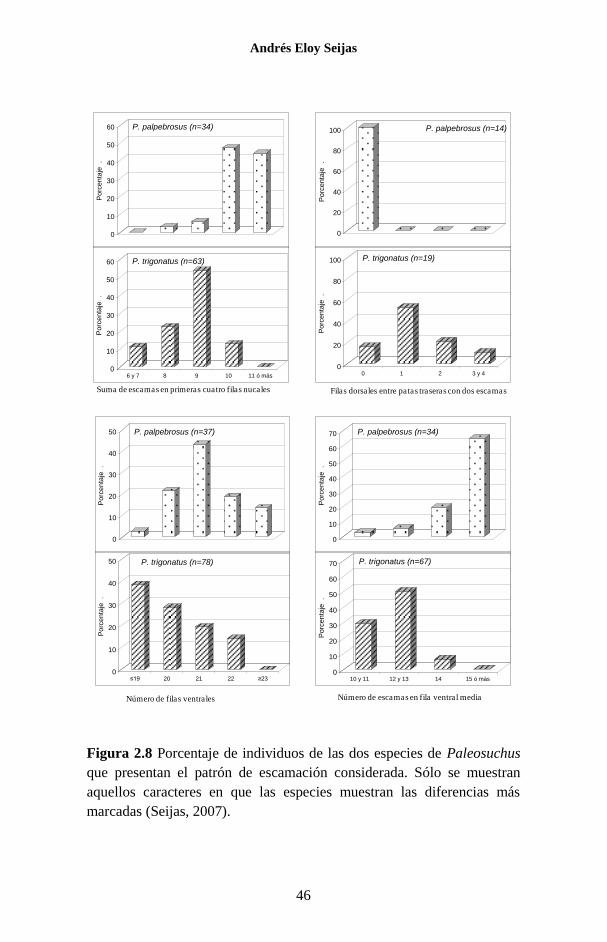
gráficos de la figura se analizaron en conjunto los datos aportados
por Medem (1981) con aquéllos recopilados en ejemplares depo-
sitados en distintos museos de Venezuela. Las diferencias más
notorias se encuentran en la sumatoria de escamas en las primeras
cuatro filas nucales, para las cuales P. trigonatus tiene gene-
ralmente nueve o menos y P. palpebrosus 10 ó más. Asimismo, el
babo negro frecuentemente presenta menos escamas en la fila
central del vientre que el babo morichalero. Aunque la muestra es
muy pequeña, ninguno de los P. palpebrosus analizados mostró
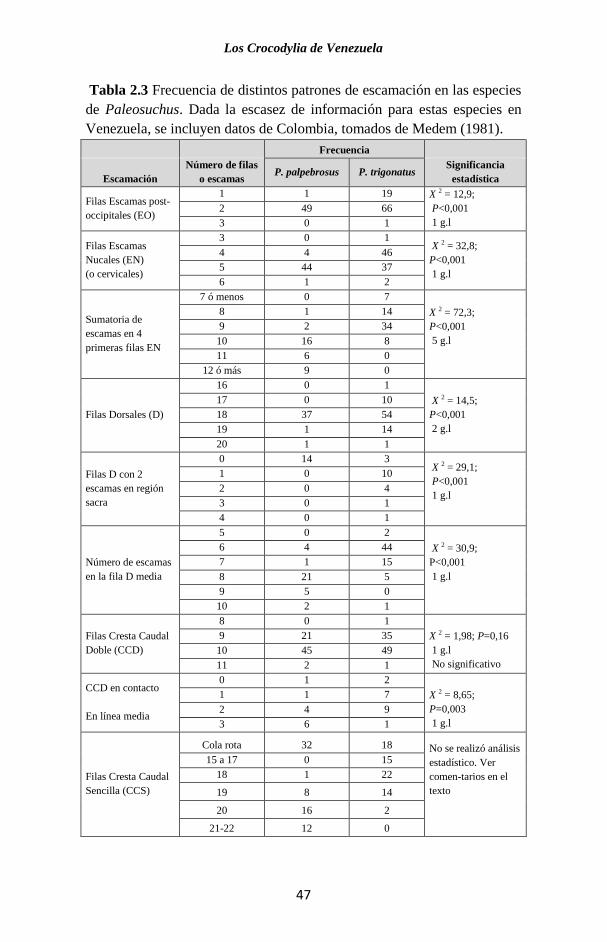
filas dorsales con sólo dos escamas en la región sacra. La Tabla
2.3 se compara otros patrones de escamación entre estas dos
especies.
Hábitat
Las dos especies de Paleosuchus habitan principalmente en ríos
y riachuelos en zonas boscosas. P. trigonatus parece estar más
restringido a ese tipo de ambientes que P. palpebrosus, el cual, de
acuerdo con Magnusson (1992a) también ocupa bosques inundados
cercanos a ríos principales, riachuelos de sabanas, lagos aislados y
canales. Ambas especies parecen encontrarse principalmente en
ríos de aguas claras y aguas negras pobres en nutrimentos. Varias
de las localidades de P. palpebrosus al norte del Orinoco corres-
ponden a zonas de bosques de palmares (Mauritia spp) conocidos
como morichales.
Aunque las especies de Paleosuchus son simpátricas en la
mayor parte de su área de distribución, el mapeo de las localidades
conocidas mostrado en la Figura 2.5 sugiere un patrón de sepa-
ración espacial y altitudinal. P. palpebrosus parece estar localizado
con más frecuencia en tierras más bajas que P. trigonatus. Tal
como lo discutieron Gorzula y Paolillo (1986), varios de los registros
de esta última especie corresponden a localidades por encima de
800 msnm y de aguas más frías (Gorzula el al., 1986). De esto se
desprende, que P. trigonatus tiende a ocupar ríos de menor orden
que P. palpebrosus. Investigaciones más detalladas permitirán
determinar si este aparente patrón es sólo aparente, con base en
una información todavía escasa y fragmentada.
Andrés Eloy Seijas
46
Figura 2.8 Porcentaje de individuos de las dos especies de Paleosuchus
que presentan el patrón de escamación considerada. Sólo se muestran
aquellos caracteres en que las especies muestran las diferencias más
marcadas (Seijas, 2007).
Suma de escamas en primeras cuatro filas nucales Filas dorsales entre patas traseras con dos escamas
Número de filas ventrales Número de escamas en fila ventral media
0
10
20
30
40
50
60
Porc
enta
je .
P. palpebrosus (n=34)
0
10
20
30
40
50
60
Porc
enta
je .
6 y 7 8 9 10 11 ó más
P. trigonatus (n=63)
0
20
40
60
80
100
Porc
enta
je .
P. palpebrosus (n=14)
0
20
40
60
80
100
Porc
enta
je .
0 1 2 3 y 4
P. trigonatus (n=19)
0
10
20
30
40
50
Porc
enta
je .
P. palpebrosus (n=37)
0
10
20
30
40
50
Porc
enta
je .
≤19 20 21 22 ≥23
P. trigonatus (n=78)
0
10
20
30
40
50
60
70
Porc
enta
je .
P. palpebrosus (n=34)
0
10
20
30
40
50
60
70
Po
rcen
taje
.
10 y 11 12 y 13 14 15 ó más
P. trigonatus (n=67)
Los Crocodylia de Venezuela
47
Tabla 2.3 Frecuencia de distintos patrones de escamación en las especies
de Paleosuchus. Dada la escasez de información para estas especies en
Venezuela, se incluyen datos de Colombia, tomados de Medem (1981).
Frecuencia
Escamación
Número de filas
o escamas P. palpebrosus P. trigonatus
Significancia
estadística
Filas Escamas post-
occipitales (EO)
1 1 19 X 2 = 12,9;
P<0,001
1 g.l
2 49 66
3 0 1
Filas Escamas
Nucales (EN)
(o cervicales)
3 0 1 X 2 = 32,8;
P<0,001
1 g.l
4 4 46
5 44 37
6 1 2
Sumatoria de
escamas en 4
primeras filas EN
7 ó menos 0 7
X 2 = 72,3;
P<0,001
5 g.l
8 1 14
9 2 34
10 16 8
11 6 0
12 ó más 9 0
Filas Dorsales (D)
16 0 1
X 2 = 14,5;
P<0,001
2 g.l
17 0 10
18 37 54
19 1 14
20 1 1
Filas D con 2
escamas en región
sacra
0 14 3 X 2 = 29,1;
P<0,001
1 g.l
1 0 10
2 0 4
3 0 1
4 0 1
Número de escamas
en la fila D media
5 0 2
X 2 = 30,9;
P<0,001
1 g.l
6 4 44
7 1 15
8 21 5
9 5 0
10 2 1
Filas Cresta Caudal
Doble (CCD)
8 0 1
X 2 = 1,98; P=0,16
1 g.l
No significativo
9 21 35
10 45 49
11 2 1
CCD en contacto
En línea media
0 1 2
X 2 = 8,65;
P=0,003
1 g.l
1 1 7
2 4 9
3 6 1
Filas Cresta Caudal
Sencilla (CCS)
Cola rota 32 18 No se realizó análisis
estadístico. Ver
comen-tarios en el
texto
15 a 17 0 15
18 1 22
19 8 14
20 16 2
21-22 12 0

Andrés Eloy Seijas
48
Reproducción
No hay un solo dato sobre los aspectos reproductivos de los
Paleosuchus en Venezuela. La información que se resume aquí
está basada en la también relativamente escasa literatura que existe
para estas especies en otros países.
Magnusson y Lima (1991) señalaron un inicio de la edad
reproductiva de 11 años para las hembras y de 20 años para los
machos de P. trigonatus. No existe información al respecto sobre
P. palpebrosus. Ambas especies construyen nidos de montículo
con material vegetal y lodo, de manera similar a la baba. El
número de huevos promedio por nidada de los Paleosuchus es
bajo, como se esperaría de especies de crocodilios de pequeño
tamaño: 15,1 y 13,3 huevos por nido para P. trigonatus y P.
palpebrosus respectivamente (Thorbjarnarson, 1996).
La fraccionada información disponible permite especular que las
estrategias reproductivas de las dos especies de Paleosuchus sean
distintas. Los datos aportados por Magnusson y Lima (1991) y
Magnusson (1992a) para zonas del Amazonas central, permiten
presumir que la época de anidación de P. trigonatus se inicia en
los períodos secos del año, para que las crías nazcan, después de
un periodo de incubación que supera los 100 días, en los primeros
meses de la época lluviosa. Esto contravendría el patrón supues-
tamente generalizado de que las crocodilios que depositan sus
huevos en montículos lo hacen en los períodos de máxima inun-
dación, para evitar la pérdida de nidadas debido a la elevación de
las aguas. Un aspecto importante que debe investigarse en el
futuro es como esta especie, a elevaciones tan altas como en las
que es posible encontrarla, logra conseguir una temperatura ade-
cuada para la incubación de los huevos y si existe alguna depen-
dencia a una fuente externa de calor, como por ejemplo montículos
de termitas, como han señalado Magnusson et al. (1990).
En el caso de P. palpebrosus, de los datos de Medem (1981)
para localidades de los llanos colombianos, se infiere que la
estrategia de esta especie es similar a la de la baba. Es decir, la

Los Crocodylia de Venezuela
49
postura de los huevos ocurriría a mediados del período lluvioso y el
nacimiento de las crías al final de las lluvias o comienzo del
período seco.
El caimán de la Costa (Crocodylus acutus)
Es una de los especies de la familia Crocodylidae de más
amplia distribución en el mundo. Los registros históricos de
distribución van desde 27º de latitud norte, en la Península de la
Florida, en los Estados Unidos de América, hasta 3º 30’ de latitud
sur en el río Tumbes, en Perú (LeBuff, 1957; Ross, 1998;
Thorbjarnarson, 1989). Dentro de esos amplios límites, la especie se
haya confinada principalmente a ambientes acuáticos cercanos a la
costa en la vertiente de los océanos Atlántico y Pacífico. Sin
embargo, se conoce que habitaba, y probablemente aún habita, en
algunos ríos a muchos kilómetros de la costa. Por ejemplo, existen
datos que indican que esta especie se localiza todavía en el río
Bogotá, un afluente del Magdalena en Colombia, a más de 700 km.
en línea recta desde la desembocadura de este río en el mar
Caribe (Barrera, 2004) y anteriormente mucho más arriba, hacia las
cercanías de la ciudad de Neiva, a unos 900 km del mar en el
mismo río (Medem, 1981). Por otra parte, Schmidt (1924) reportó
esta especie para el lago Yohoa en Honduras, sobre 600 m.s.n.m y
también existen registros de individuos observados a 1220 msnm en
México (Thorbjarnarson, 1988). C. acutus ha sido reportado para
17 países de la región, entre ellos los países islas de Cuba, Jamaica
y Haití-República Dominicana (Española). Esta amplia distribución
justificaría plenamente el nombre común de ‘cocodrilo americano’
con que se ha venido llamando a esta especie.
El caimán de la costa fue sometido durante el siglo XX a una
serie de factores adversos que provocaron su extinción en nume-
rosas localidades y disminuyeron sus poblaciones en el resto de su
área de distribución en la región norte-costera de Venezuela. La
disminución de la abundancia poblacional de C. acutus se debió, en
primer lugar, a la explotación irracional con fines comerciales a que
fue sometido entre 1929 y comienzos de los años sesenta del siglo

Andrés Eloy Seijas
50
pasado, periodo en que quedó completamente agotado como
recurso (Mondolfi, 1965; Medem, 1983). Sin embargo, la mayor
amenaza actual consiste en la persistente pérdida de hábitat. En la
región norte costera de Venezuela se concentra la mayoría de su
población humana. Un anárquico crecimiento industrial y urbanístico
ha traído consigo la destrucción de zonas de manglares, la
desecación de ciénagas y albuferas, la contaminación y disminución
de los caudales de los ríos costeros; todos ellos hábitats esenciales
del caimán de la costa.
Sobre la base de registros históricos y a datos obtenidos desde
1980 (Seijas, 1986c, 1990), se constata que el área de distribución
de C. acutus en Venezuela abarcaba toda la cuenca del lago de
Maracaibo, la cuenca caribeña (incluyendo la isla de Margarita) y
el sur de la península de Paria, en Sucre (Fig. 2.9).
La región que va desde la desembocadura del río Ocumare, en
el estado Aragua, hasta Cabo Codera, en el estado Miranda posee
ríos muy cortos y pocas áreas propicias para el establecimiento de
poblaciones grandes de caimanes, pero quizás en el pasado, cuando
las poblaciones humanas eran escasas, diversas localidades a lo
largo de esa franja costera eran ocupadas por pequeñas pobla-
ciones de caimanes, o por individuos errantes que garantizaban el
flujo genético entre las grandes áreas de distribución de la especie
en el oriente y el occidente del país. En apoyo a esta hipótesis
existe un registro en el museo de la Fundación La Salle en
Caracas, de una cría de unos 30 cm de LT, colectada por
Cornelius Vogl en enero de 1955 en Camurí Chico, en el litoral
central de Venezuela, lo que demuestra que todavía en ese año,
relativamente reciente, había individuos reproductores en esa región.
No existe mucha información sobre el estado poblacional del
caimán de la costa en Venezuela para el momento de la llegada de
los colonizadores españoles, pero la información reciente nos
permiten presumir que la especie era abundante en los ríos
mayores de la cuenca del lago de Maracaibo (Chama, Santa Ana,
Escalante, Catatumbo, por ejemplo) y en ríos que desembocan en

Los Crocodylia de Venezuela
51
la costa caribeña como el Tocuyo, Yaracuy, Tuy, Unare y Neverí,
por mencionar sólo algunos de los más largos. De las crónicas de
siglo XIX sólo tenemos los relatos de Alejandro de Humboldt
(1991) sobre la presencia de la especie en el río Neverí (Anzoá-
tegui) y Aguas Calientes (Carabobo) y de Karl Ferdinand Appun
(Appun, 1961), quien destacó la abundancia de caimanes tanto en la
desembocadura como aguas arriba de los ríos Yaracuy y San
Esteban, este último en el estado Carabobo.
En la década de los 80 del siglo XX se realizó un extenso
estudio para determinar la situación de C. acutus en Venezuela
(Seijas, 1986c) y se constató la existencia de 14 localidades que
todavía presentaban pequeñas poblaciones de la especie. Estudios
posteriores han profundizado en la evaluación del estado poblacional
y en la ecología de este cocodrilo (Seijas, 1988, 1996; Chávez,
1992; Arteaga, 1997, 1998; Arteaga y Gómez, 2000; Urdaneta
2002; Lander, 2003; Barros et al., 2005; Luzardo, 2005; Valeris,
2007; Lander et al., 2008). En el Capítulo 5 se discutirá en más
detalle la situación actual de las poblaciones de esta especie en
Venezuela y se presentará un listado de las localidades principales
con la cifra máxima de individuos reportada en la literatura.
Características físicas
El caimán de la costa es un crocodílido de gran tamaño, con
talla máxima sugerida (aunque nunca comprobada) de 7 m.
(Brazaitis, 1973, Álvarez del Toro, 1974, Schmidt 1924, Medem,
1981). Entre las características más distintivas de la especie resalta,
en primer lugar, su reducido e irregular arreglo de escamas
dorsales, donde usualmente no hay más de cuatro escamas en
ninguna de sus 13 a 18 filas transversales y, en segundo lugar, la
presencia de una elevación de la zona pre-ocular del hocico, que se
manifiesta mejor en los individuos de mayor tamaño (Fig. 2.10).
Estos dos rasgos son suficientes para distinguirlo de cualquier otro
cocodrilo del mundo. Además, presenta una amplia separación entre
las escamas nucales y las dorsales y la sínfisis mandibular se
completa al nivel del quinto diente.

Andrés Eloy Seijas
52
Respecto a la escamación, otra característica importante de C.
acutus es la gran variación en el patrón de escamas nucales (o
cervicales) (Seijas, 2002). En una muestra de 565 individuos
provenientes de distintas localidades de Venezuela, 440 (78%)
presentaban el patrón considerado típico de dos filas, la primera
con cuatro escamas grandes y la segunda con sólo dos. Los
caimanes restantes poseían 19 patrones distintos al típico, en los
que faltaban entre 1 y 4 escamas (Fig. 2.11). Un resumen de las
características de la escamación de 685 C. acutus, provenientes de
numerosas localidades del país, se presenta en la Tabla 2.4.
Figura 2.9 Localización de registros históricos y actuales de Crocodylus
acutus en Venezuela.

Los Crocodylia de Venezuela
53
Figura 2.10 Vista lateral del cráneo de un Crocodylus acutus adulto de la
colección del Museo de la Estación Biológica de Rancho Grande en
Maracay. Note el marcado desarrollo de la elevación pre -ocular, rasgo
que caracteriza a esta especie.
Hábitat
El hábitat principal u óptimo de C. acutus es el río, no en
balde Álvarez del Toro (1974) en su monografía sobre los
Crocodylia de México lo denominó “cocodrilo de río”, para distin-
guirlo de las otras especies de este grupo en ese país, que ocupan
con preferencia otros tipos de ambientes.
No obstante que C. acutus tenga como hábitat óptimo a los
ríos, es una especie de una gran adaptabilidad y ocupa también
estuarios, lagunas costeras y ciénagas de manglares, ambientes
estos de agua salobre, salina o incluso hipersalina (Thorbjarnarson
et al., 2006). Un ejemplo de esto último se presenta en una laguna
costera aledaña a la bahía de Turiamo, estado Aragua, donde
temporalmente las aguas pueden alcanzar salinidades muy altas. Allí
Seijas (1990) reportó, para marzo de 1987, valores de 80-90 partes
por mil, casi tres veces la concentración de sal de las aguas del
mar. En esa localidad de manera recurrente la salinidad puede
superar 60 ppm (Seijas, 1990, Vargas 1993, citada por González-
Fernández, 1999).
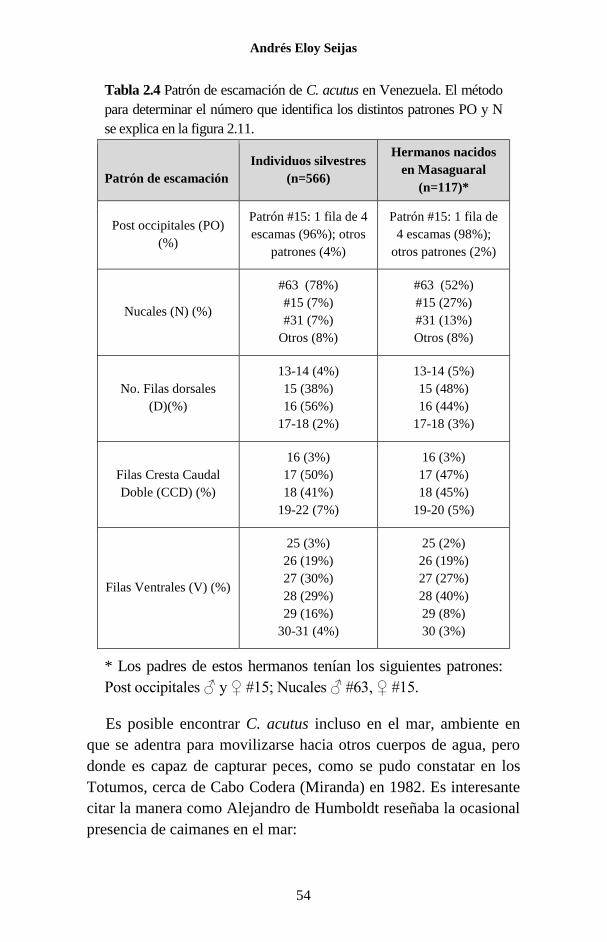
Andrés Eloy Seijas
54
Tabla 2.4 Patrón de escamación de C. acutus en Venezuela. El método
para determinar el número que identifica los distintos patrones PO y N
se explica en la figura 2.11.
Patrón de escamación
Individuos silvestres
(n=566)
Hermanos nacidos
en Masaguaral
(n=117)*
Post occipitales (PO)
(%)
Patrón #15: 1 fila de 4
escamas (96%); otros
patrones (4%)
Patrón #15: 1 fila de
4 escamas (98%);
otros patrones (2%)
Nucales (N) (%)
#63 (78%)
#15 (7%)
#31 (7%)
Otros (8%)
#63 (52%)
#15 (27%)
#31 (13%)
Otros (8%)
No. Filas dorsales
(D)(%)
13-14 (4%)
15 (38%)
16 (56%)
17-18 (2%)
13-14 (5%)
15 (48%)
16 (44%)
17-18 (3%)
Filas Cresta Caudal
Doble (CCD) (%)
16 (3%)
17 (50%)
18 (41%)
19-22 (7%)
16 (3%)
17 (47%)
18 (45%)
19-20 (5%)
Filas Ventrales (V) (%)
25 (3%)
26 (19%)
27 (30%)
28 (29%)
29 (16%)
30-31 (4%)
25 (2%)
26 (19%)
27 (27%)
28 (40%)
29 (8%)
30 (3%)
* Los padres de estos hermanos tenían los siguientes patrones:
Post occipitales ♂ y ♀ #15; Nucales ♂ #63, ♀ #15.
Es posible encontrar C. acutus incluso en el mar, ambiente en
que se adentra para movilizarse hacia otros cuerpos de agua, pero
donde es capaz de capturar peces, como se pudo constatar en los
Totumos, cerca de Cabo Codera (Miranda) en 1982. Es interesante
citar la manera como Alejandro de Humboldt reseñaba la ocasional
presencia de caimanes en el mar:
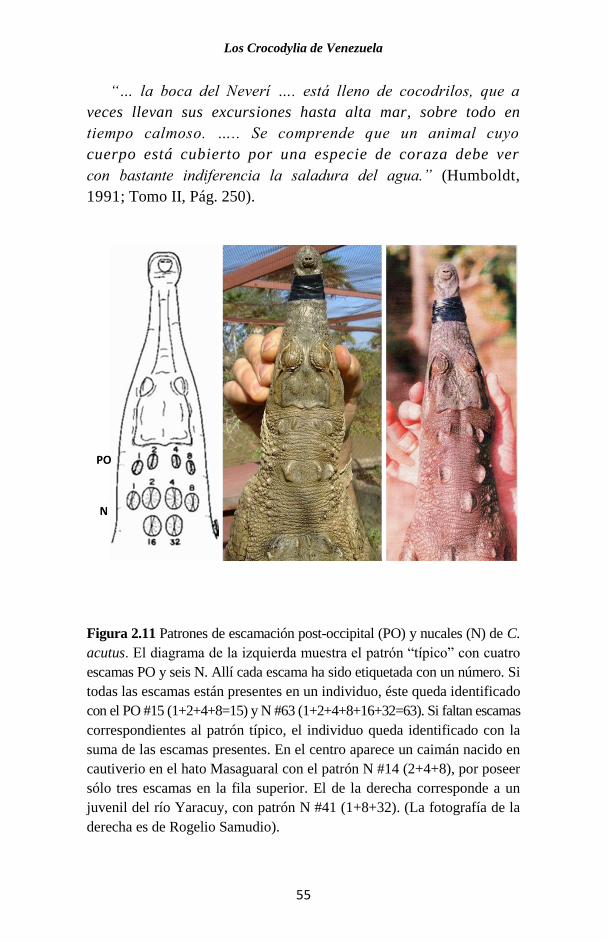
Los Crocodylia de Venezuela
55
“… la boca del Neverí …. está lleno de cocodrilos, que a
veces llevan sus excursiones hasta alta mar, sobre todo en
tiempo calmoso. ….. Se comprende que un animal cuyo
cuerpo está cubierto por una especie de coraza debe ver
con bastante indiferencia la saladura del agua.” (Humboldt,
1991; Tomo II, Pág. 250).
Figura 2.11 Patrones de escamación post-occipital (PO) y nucales (N) de C.
acutus. El diagrama de la izquierda muestra el patrón “típico” con cuatro
escamas PO y seis N. Allí cada escama ha sido etiquetada con un número. Si
todas las escamas están presentes en un individuo, éste queda identificado
con el PO #15 (1+2+4+8=15) y N #63 (1+2+4+8+16+32=63). Si faltan escamas
correspondientes al patrón típico, el individuo queda identificado con la
suma de las escamas presentes. En el centro aparece un caimán nacido en
cautiverio en el hato Masaguaral con el patrón N #14 (2+4+8), por poseer
sólo tres escamas en la fila superior. El de la derecha corresponde a un
juvenil del río Yaracuy, con patrón N #41 (1+8+32). (La fotografía de la
derecha es de Rogelio Samudio).

Andrés Eloy Seijas
56
El grado de adaptabilidad del caimán de la costa a distintos
ambientes se demuestra en el hecho de que prospera en distintos
embalses a lo largo de la costa de Venezuela (Pueblo Viejo en
Zulia; Jatira y Tacarigua en Falcón; Cumaripa en Yaracuy, entre
otros). Ha sido encontrado también en el sistema de canales
artificiales de las cercanías de la población de Río Chico en el
estado Miranda, entre las desembocaduras de los ríos Guapo y Tuy
(Seijas, 1986c, 1990; Babarro, 2011).
Aun cuando el caimán de la costa habita en diversidad de
ambientes, los estudios sobre crecimiento corporal de individuos
recién nacidos capturados en distintas localidades indicaron que en
aguas salobres o salinas, como Turiamo y la Laguna de Tacarigua
y Río Chico, en Miranda, los caimanes presentaron tasas de
crecimiento bajas (0,53 y 0,57 mm/día), en los de embalses como
Pueblo Viejo (Zulia) y Jatira-Tacarigua (Falcón) crecieron a tasas
intermedias (0,84 y 1,04 mm/día, respectivamente), mientras que los
del río Yaracuy crecieron a una tasa de 1,19 mm/día, es decir, más
del doble que en localidades de agua salada (Seijas, 1991a). Dichas
tasas de crecimiento pueden ser tomadas como indicadoras de la
calidad del hábitat para la especie de cada uno de esos ambientes
y apoyan la afirmación de que los ríos representan el hábitat óptimo.
Reproducción
Las hembras de C. acutus depositan sus huevos en huecos que
excavan en los bancos arenosos de las orillas de los ríos o en
playas u otras zonas elevadas cerca de las orillas de los cuerpos
de agua que habitan. En los embalses, donde el sustrato arenoso
es escaso o ausente, las hembras utilizan material más compacto y
duro, pero se desconoce la incidencia que este tipo de sustrato
tiene sobre el éxito de la eclosión. Se ha reportado que C. acutus
también construye nidos de montículo con detritus de material
vegetal, al igual que la baba (Ogden, 1978; Thorbjarnarson, 1990),
pero esto no ha sido observado en ninguna localidad de Venezuela.
Existe escasa información sobre el número de huevos por
nidada para C. acutus en Venezuela. Medem (1983) mencionó
datos anecdóticos de pobladores de la región de Cuare, Falcón,

Los Crocodylia de Venezuela
57
quienes afirmaron que el tamaño de las nidadas está en el orden
de los 20-25 huevos. Estos son precisamente los intervalos seña-
lados por Arteaga y Gómez (2000) para la laguna de Tacarigua.
Para el río Santa Rosa del estado Zulia hay datos de dos nidadas
con 22 y 38 huevos (Valeris, 2007). Thorbjarnarson (1999) aportó
datos de una hembra cautiva en el hato Masaguaral que puso 39 y
42 huevos en los años 1998 y 1999, respectivamente. Las cifras
reportadas por este último autor quizás se acerquen más a los
valores promedios de hembras grandes, si tomamos en cuenta que
en el río Yaracuy se han conseguido grupos de crías recién
nacidas que van de 29 a 34 individuos (Seijas y Chávez, 1991), lo
que sugiere nidadas aún mayores, ya que rara vez todos los huevos
de un nido eclosionan de manera satisfactoria. La cifra promedio que
reportó Thorbjarnarson (1996) para la especie es 35,6 huevos por nido.
Como ocurre en todos los crocodilios, la cronología reproductiva
de C. acutus está estrechamente asociada con el régimen climá-
tico. Sin embargo, debido a la escasez de información y a la gran
variedad de ambientes que esta especie ocupa en Venezuela no es
posible precisar cuáles son las épocas de cortejo, anidación y
eclosión para todas las localidades donde ha sido encontrada. Las
localidades para las que existe mayor información son río el río
Yaracuy, los embalses de Jatira y Tacarigua en Falcón, el Parque
Nacional Laguna de Tacarigua en Miranda, el embalse de Pueblo
Viejo en Zulia y Turiamo, en el estado Aragua.
La fecha de nacimiento de las crías en cada región se puede
inferir a partir de las tallas de individuos capturados en los primeros
meses de vida (Seijas et al., 2008). El método consistió en realizar
análisis de regresión lineal de la longitud total (en mm) de los
caimanes versus los días transcurridos después del 1 de enero del
año en que nacieron. Como en la literatura se señala que los
neonatos de C. acutus tienen una talla comprendida entre los 240
y 270 mm (Álvarez del Toro, 1974; Ogden, 1978; Seijas et al.,
1990), se calculó donde la recta de regresión talla vs fecha
obtenida, cortaba a la línea correspondiente a 255 mm, la talla
intermedia entre las cifras antes señaladas. La fecha obtenida se
consideró, para cada región, como el momento de inicio de la
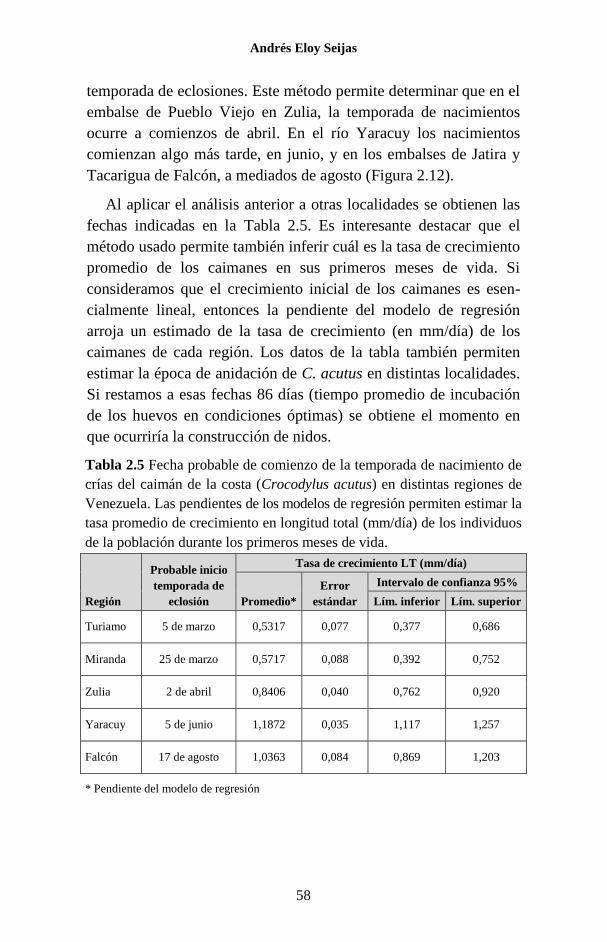
Andrés Eloy Seijas
58
temporada de eclosiones. Este método permite determinar que en el
embalse de Pueblo Viejo en Zulia, la temporada de nacimientos
ocurre a comienzos de abril. En el río Yaracuy los nacimientos
comienzan algo más tarde, en junio, y en los embalses de Jatira y
Tacarigua de Falcón, a mediados de agosto (Figura 2.12).
Al aplicar el análisis anterior a otras localidades se obtienen las
fechas indicadas en la Tabla 2.5. Es interesante destacar que el
método usado permite también inferir cuál es la tasa de crecimiento
promedio de los caimanes en sus primeros meses de vida. Si
consideramos que el crecimiento inicial de los caimanes es esen-
cialmente lineal, entonces la pendiente del modelo de regresión
arroja un estimado de la tasa de crecimiento (en mm/día) de los
caimanes de cada región. Los datos de la tabla también permiten
estimar la época de anidación de C. acutus en distintas localidades.
Si restamos a esas fechas 86 días (tiempo promedio de incubación
de los huevos en condiciones óptimas) se obtiene el momento en
que ocurriría la construcción de nidos.
Tabla 2.5 Fecha probable de comienzo de la temporada de nacimiento de
crías del caimán de la costa (Crocodylus acutus) en distintas regiones de
Venezuela. Las pendientes de los modelos de regresión permiten estimar la
tasa promedio de crecimiento en longitud total (mm/día) de los individuos
de la población durante los primeros meses de vida.
Región
Probable inicio
temporada de
eclosión
Tasa de crecimiento LT (mm/día)
Promedio*
Error
estándar
Intervalo de confianza 95%
Lím. inferior Lím. superior
Turiamo 5 de marzo 0,5317 0,077 0,377 0,686
Miranda 25 de marzo 0,5717 0,088 0,392 0,752
Zulia 2 de abril 0,8406 0,040 0,762 0,920
Yaracuy 5 de junio 1,1872 0,035 1,117 1,257
Falcón 17 de agosto 1,0363 0,084 0,869 1,203
* Pendiente del modelo de regresión
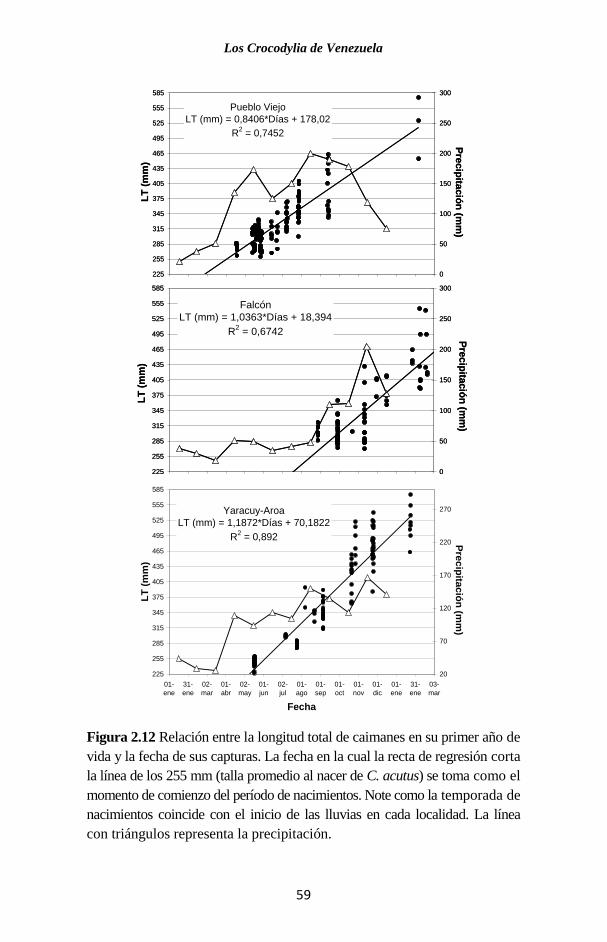
Los Crocodylia de Venezuela
59
Figura 2.12 Relación entre la longitud total de caimanes en su primer año de
vida y la fecha de sus capturas. La fecha en la cual la recta de regresión corta
la línea de los 255 mm (talla promedio al nacer de C. acutus) se toma como el
momento de comienzo del período de nacimientos. Note como la temporada de
nacimientos coincide con el inicio de las lluvias en cada localidad. La línea
con triángulos representa la precipitación.
Falcón
LT (mm) = 1,0363*Días + 18,394
R2 = 0,6742
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
0
50
100
150
200
250
300
Pre
cip
itac
ión
(mm
)
Yaracuy-Aroa
LT (mm) = 1,1872*Días + 70,1822
R2 = 0,892
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
20
70
120
170
220
270
Pre
cip
itació
n (m
m)
Pueblo Viejo
LT (mm) = 0,8406*Días + 178,02
R2 = 0,7452
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
0
50
100
150
200
250
300
Pre
cip
itació
n (m
m)
Falcón
LT (mm) = 1,0363*Días + 18,394
R2 = 0,6742
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
0
50
100
150
200
250
300
Pre
cip
itac
ión
(mm
)
Yaracuy-Aroa
LT (mm) = 1,1872*Días + 70,1822
R2 = 0,892
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
20
70
120
170
220
270
Pre
cip
itació
n (m
m)
Pueblo Viejo
LT (mm) = 0,8406*Días + 178,02
R2 = 0,7452
225
255
285
315
345
375
405
435
465
495
525
555
585
01-
ene
31-
ene
02-
mar
01-
abr
02-
may
01-
jun
02-
jul
01-
ago
01-
sep
01-
oct
01-
nov
01-
dic
01-
ene
31-
ene
03-
mar
Fecha
LT
(m
m)
0
50
100
150
200
250
300
Pre
cip
itació
n (m
m)

Andrés Eloy Seijas
60
Todas estas cifras y fechas deben ser tomadas como pre-
liminares y podrían ajustarse aún más cuando se recabe más
información. Por ejemplo, hay un dato de una hembra que anidó en
Cuare, en el estado Falcón, el 25 de agosto de 1982, lo cual
parece completamente extemporáneo para la zona. Pero, por otro
lado, Arteaga y Gómez (2000) indicaron que la temporada de
anidación en la laguna de Tacarigua en el estado Miranda, en 1999,
se inició el 23 de marzo, prácticamente idéntica a la señalada aquí.
El caimán del Orinoco (Crocodylus intermedius)
El caimán del Orinoco recibe ese nombre por ser endémico de
la cuenca de dicho río. Esta cuenca se extiende por un amplio
territorio (1.123.000 km2, Taphorn, 1992), que cubre casi tres
cuartas partes de la superficie de Venezuela y cerca de 35% de la
de Colombia (Fig. 2.13).
El área de distribución histórica de Crocodylus intermedius se
restringía principalmente a las tierras bajas, generalmente por
debajo de 300 m.s.n.m. Registros más recientes indican que la
distribución de la especie se extendía aguas arriba en muchos ríos
del piedemonte de los Andes (Ramo y Busto, 1986; Thorbjarnarson
y Hernández, 1992) y en la mayoría de los ríos de la margen
derecha del Orinoco, incluyendo aquellos que transcurren por áreas
de bosques cerrados (Hitchcock, 1948; Franz et al., 1985; Arteaga
et al., 1994). Poco se conoce sobre la distribución histórica del
caimán en el alto Orinoco. Humboldt (1991) indicó que los
caimanes eran comunes en el río Guaviare, afluente de la margen
izquierda de este río, pero que faltaban por completo en el Paragua
(Alto Orinoco) entre San Fernando de Atabapo y La Esmeralda.
Estas diferencias las explicaba por las distintas características de
las aguas: blancas las del Guaviare y claras las del Paragua. Hoy
sabemos que las primeras son más productivas que las segundas
(Sioli, 1984). Respecto al delta del Orinoco, el propio Humboldt se
refirió a caimanes en esa región (que nunca visitó). En tiempos
relativamente recientes Cardona (1959) señaló haber observado
“bastantes caimanes” en caños de Delta Amacuro, uno de ellos
muy grande cerca de Araguaimujo, es decir, no muy lejos de la

Los Crocodylia de Venezuela
61
costa en el océano Atlántico. No se ha podido establecer si las
mencionadas reseñas, y otras de distintos autores, correspondían a
caimanes del Orinoco o de la costa.
El caimán del Orinoco se encuentra en la actualidad en sólo una
pequeña fracción de su antigua área de distribución original. La
explotación comercial en el pasado reciente eliminó a la especie de
los sistemas de ríos antes mencionados. En los últimos 30 años se
han reportado individuos aislados y pequeñas poblaciones de la
especie en Venezuela (Godshalk, 1978; Franz et al., 1985; Ramo y
Busto, 1986; Thorbjarnarson y Hernández, 1992; Seijas, 1992;
Jiménez-Oraá, 2002). Las poblaciones mayores, y quizás las únicas
viables (Arteaga et al., 1997) de C. intermedius se localizan en el
río Capanaparo, en el estado Apure (Thorbjarnarson y Hernández,
1992; Llobet, 2002) y en el sistema Sarare-Cojedes (Ayarzagüena,
1987, 1990; Seijas, 1992, Seijas y Chávez, 2000). Habría que
agregar aquí a la población de lo que fue el hato El Frío y zonas
aledañas en el Caño Guaritico donde se restableció una población
gracias al esfuerzo de liberación de animales cautivos a partir de
1990 (Antelo, 2008).
Características físicas
El caimán del Orinoco es un cocodrilo de hocico alargado
(“longirrostrino”), para el que existen registros históricos de
individuos cercanos a los 6,50 metros de longitud total (Humboldt
1991; Medem, 1981; López-Corcuera, 1984) lo que lo convierte en
una de las especies de cocodrilos más grandes del mundo. De
acuerdo con información anecdótica, sin embargo, los machos más
grandes promedian tallas entre 4 y 5 m de longitud total (Medem,
1981, 1983; Whitney, 1912).
A diferencia de lo que ocurre con el caimán de la costa, la
escamación dorsal de C. intermedius exhibe un arreglo bastante
regular, con seis escamas en la mayor parte de sus 15 a 18 filas
de escamas dorsales. Presenta además una menor separación entre
las escamas nucales y las dorsales y la sínfisis mandibular se
completa al nivel del sexto y séptimo diente.
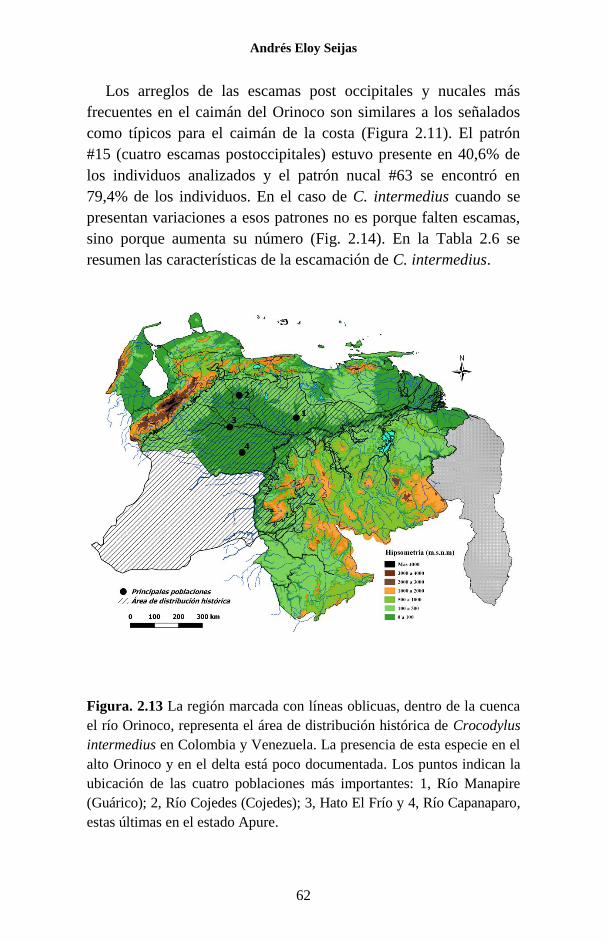
Andrés Eloy Seijas
62
Los arreglos de las escamas post occipitales y nucales más
frecuentes en el caimán del Orinoco son similares a los señalados
como típicos para el caimán de la costa (Figura 2.11). El patrón
#15 (cuatro escamas postoccipitales) estuvo presente en 40,6% de
los individuos analizados y el patrón nucal #63 se encontró en
79,4% de los individuos. En el caso de C. intermedius cuando se
presentan variaciones a esos patrones no es porque falten escamas,
sino porque aumenta su número (Fig. 2.14). En la Tabla 2.6 se
resumen las características de la escamación de C. intermedius.
Figura. 2.13 La región marcada con líneas oblicuas, dentro de la cuenca
el río Orinoco, representa el área de distribución histórica de Crocodylus
intermedius en Colombia y Venezuela. La presencia de esta especie en el
alto Orinoco y en el delta está poco documentada. Los puntos indican la
ubicación de las cuatro poblaciones más importantes: 1, Río Manapire
(Guárico); 2, Río Cojedes (Cojedes); 3, Hato El Frío y 4, Río Capanaparo,
estas últimas en el estado Apure.
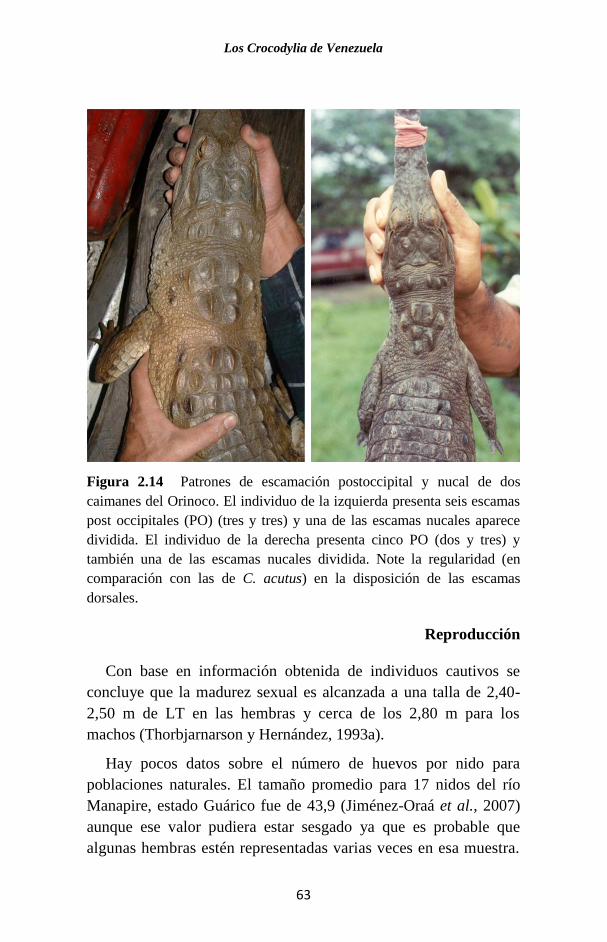
Los Crocodylia de Venezuela
63
Figura 2.14 Patrones de escamación postoccipital y nucal de dos
caimanes del Orinoco. El individuo de la izquierda presenta seis escamas
post occipitales (PO) (tres y tres) y una de las escamas nucales aparece
dividida. El individuo de la derecha presenta cinco PO (dos y tres) y
también una de las escamas nucales dividida. Note la regularidad (en
comparación con las de C. acutus) en la disposición de las escamas
dorsales.
Reproducción
Con base en información obtenida de individuos cautivos se
concluye que la madurez sexual es alcanzada a una talla de 2,40-
2,50 m de LT en las hembras y cerca de los 2,80 m para los
machos (Thorbjarnarson y Hernández, 1993a).
Hay pocos datos sobre el número de huevos por nido para
poblaciones naturales. El tamaño promedio para 17 nidos del río
Manapire, estado Guárico fue de 43,9 (Jiménez-Oraá et al., 2007)
aunque ese valor pudiera estar sesgado ya que es probable que
algunas hembras estén representadas varias veces en esa muestra.

Andrés Eloy Seijas
64
El mayor número de huevos en una nidada registrado en esa
localidad es de 66. Para el Río Cojedes han sido señalados valores
promedios de 41,0 (n=10) y 49,5 (n=8) huevos/nido (Navarro, 2007
y Espinosa-Blanco, 2010, respectivamente). Para 69 nidos del Hato
El Frío, los datos en Antelo et. al. (2010) permiten calcular un
promedio de 41.86 huevos/nido. El número de huevos promedio
colocados por cuatro hembras cautivas en el zoocriadero de la
UNELLEZ aparece en la Tabla 2.7. El número de huevos por postura
difiere de manera significativa entre estas hembras (F3,46=8,52;
P<0,001) lo que indica que cada hembra tiende a poner un número
de huevos que le es característico.
Tabla 2.6 Frecuencia de aparición de distintos patrones de escamación
de C. intermedius en Venezuela.
Patrón Post Occipitales
(PO)
(n = 244)
4 escamas (2 y 2): 40,6%
6 escamas (3 y 3): 25,4%
Otros patrones: 34%
Patrón Nucales (N)
(n = 243)
4 y 2: 79,4%
Otros patrones: 20,6%
No. Filas dorsales (D)
(n = 72)
15: 1,4%
16: 80,6%
17: 16,7%
18: 1,4%
Filas Cresta Caudal Doble
(CCD)
(n = 162)
18: 64,2%
19: 33,3%
20: 2,5%
Filas Ventrales (V)
(n = 30)
25: 6,7%
26: 36,7 %
27: 36,7%
28: 20,0%
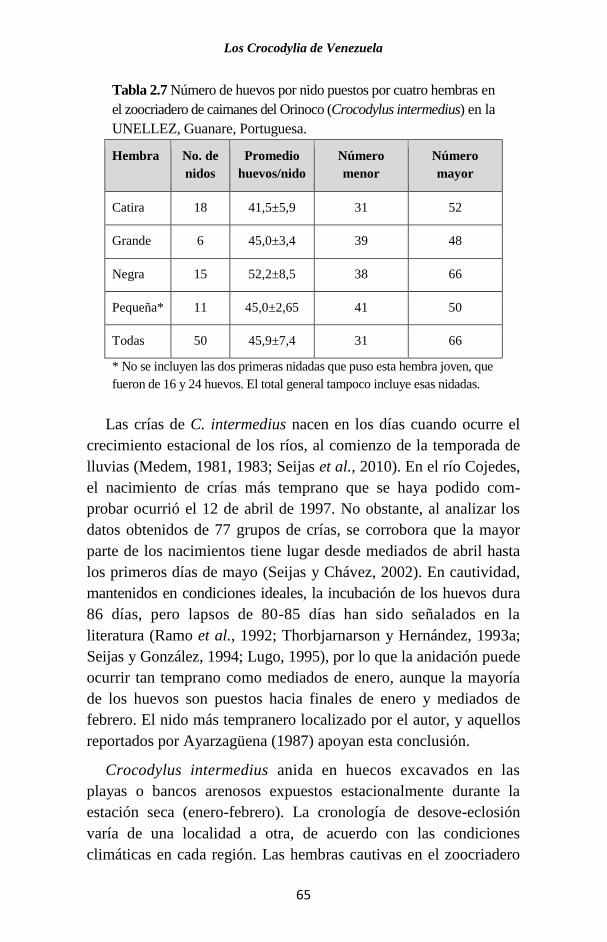
Los Crocodylia de Venezuela
65
Tabla 2.7 Número de huevos por nido puestos por cuatro hembras en
el zoocriadero de caimanes del Orinoco (Crocodylus intermedius) en la
UNELLEZ, Guanare, Portuguesa.
Hembra No. de
nidos
Promedio
huevos/nido
Número
menor
Número
mayor
Catira 18 41,5±5,9 31 52
Grande 6 45,0±3,4 39 48
Negra 15 52,2±8,5 38 66
Pequeña* 11 45,0±2,65 41 50
Todas 50 45,9±7,4 31 66
* No se incluyen las dos primeras nidadas que puso esta hembra joven, que
fueron de 16 y 24 huevos. El total general tampoco incluye esas nidadas.
Las crías de C. intermedius nacen en los días cuando ocurre el
crecimiento estacional de los ríos, al comienzo de la temporada de
lluvias (Medem, 1981, 1983; Seijas et al., 2010). En el río Cojedes,
el nacimiento de crías más temprano que se haya podido com-
probar ocurrió el 12 de abril de 1997. No obstante, al analizar los
datos obtenidos de 77 grupos de crías, se corrobora que la mayor
parte de los nacimientos tiene lugar desde mediados de abril hasta
los primeros días de mayo (Seijas y Chávez, 2002). En cautividad,
mantenidos en condiciones ideales, la incubación de los huevos dura
86 días, pero lapsos de 80-85 días han sido señalados en la
literatura (Ramo et al., 1992; Thorbjarnarson y Hernández, 1993a;
Seijas y González, 1994; Lugo, 1995), por lo que la anidación puede
ocurrir tan temprano como mediados de enero, aunque la mayoría
de los huevos son puestos hacia finales de enero y mediados de
febrero. El nido más tempranero localizado por el autor, y aquellos
reportados por Ayarzagüena (1987) apoyan esta conclusión.
Crocodylus intermedius anida en huecos excavados en las
playas o bancos arenosos expuestos estacionalmente durante la
estación seca (enero-febrero). La cronología de desove-eclosión
varía de una localidad a otra, de acuerdo con las condiciones
climáticas en cada región. Las hembras cautivas en el zoocriadero

Andrés Eloy Seijas
66
de la UNELLEZ, en Guanare, siguen un patrón muy similar al
descrito para el río Cojedes. Las hembras cautivas en el hato
Masaguaral, del estado Guárico, presentan un retardo de aproxi-
madamente un mes, al igual que las del río Manapire, en el mismo
estado, donde se ha reportado como época de desove la última
semana de febrero-primera de marzo, y la de eclosión entre finales
de mayo y comienzos de junio (Jiménez-Oraá, 2002).
Hábitat
En el pasado, el caimán del Orinoco se localizaba en una amplia
variedad de hábitats, incluidos ríos de bosques tropicales siempre-
verdes, ríos del piedemonte andino y lagunas temporales y perma-
nentes de los llanos de Venezuela y Colombia. Los registros
históricos permiten inferir que el hábitat principal de la especie lo
constituían los ríos mayores de la cuenca del Orinoco en los llanos
de estos países (Humboldt, 1975, 1991; Codazzi, 1940; Páez, 1980;
Medem, 1981, 1983), en particular el Casanare y en menor medida
el Guayabero-Vichada en Colombia, y el Apure, Portuguesa y el
propio Orinoco en Venezuela, así como también en los ríos
compartidos entre estos países como el Arauca y el Meta, por
mencionar sólo los más importantes.
Hoy en día se conoce que la especie puede ocupar embalses,
aunque en densidades muy bajas (Thorbjarnarson y Hernández,
1992; Seijas y Meza, 1994). Se puede afirmar que el hábitat óptimo
para esta especie son los ríos de aguas blancas con amplias playas
arenosas.
CONSERVACIÓN DE LAS ESPECIES DE
CROCODYLIA DE VENEZUELA
Los grandes caimanes (Crocodylus intermedius y C. acutus)
están consideradas especies “En Peligro” de acuerdo con la
clasificación del Libro Rojo de la Fauna de Venezuela (Rodríguez y
Rojas-Suárez, 2008). Están además en la lista de especies en
“peligro de extinción” por resolución presidencial No. 1486 de 1996
(Venezuela, 1996). Su caza fue vedada por un decreto del 4 de

Los Crocodylia de Venezuela
67
abril de 1973, decisión ratificada por resolución presidencial en 1979
(Venezuela, 1973, 1979). Ambas especies están incluidas en el
Apéndice I de la Convención sobre el Comercio Internacional de
Especies Amenazadas de Flora y Fauna Silvestres (CITES). No
obstante todas esas medidas protectoras, los caimanes están todavía
en una situación crítica, con niveles poblacionales bajos, en parte
como consecuencia de la explotación comercial a que fueron
sometidos hace ya varias décadas pero también porque persisten
actividades que impiden su recuperación: destrucción de hábitat,
saqueo de nidadas, venta de crías como mascotas, muertes acci-
dentales en redes de pescadores, caza para el consumo de su
carne o grasa y por el supuesto o real peligro que representan para
los animales domésticos o la gente.
En el caso del caimán del Orinoco existe un programa de
conservación en marcha desde mediados de los años 1980. Gran
parte de los esfuerzos iniciales para la conservación de C.
intermedius surgieron de la iniciativa personal de Cecilia y Tomás
Blohm y de la Fundación para la Defensa de la Naturaleza
(FUDENA), institución que coordinó el apoyo de instituciones
gubernamentales y privadas como el Ministerio del Ambiente y de
los Recursos Naturales (MARN), el Fondo Mundial para la
Conservación (WWF), La Sociedad Zoológica de Nueva York
(WCS-NYZS), la Agencia Española de Cooperación Internacional
(AECI) y muchas otras instituciones y personas (Thorbjarnarson y
Arteaga, 1995). Los detalles de uno de los componentes principales
de este programa, la cría en cautiverio con fines de repoblación,
serán discutidos en el Capítulo 8.
Un aspecto clave para la recuperación de los caimanes es la
conservación de sus hábitats. Las localidades legalmente protegidas
dentro del área de distribución histórica del caimán del Orinoco son
los parques nacionales Santos Luzardo (Cinaruco-Capanaparo) en el
estado Apure, el Aguaro-Guariquito y los Esteros de Camaguan, en
el estado Guárico. La guardería ambiental en esos parques es muy
deficiente y en la práctica no cumplen con la función de con-
servación que la especie requiere. Además del refugio “fantasma”
del Estero de Chiriguare, en el estado Portuguesa, existe otro

Andrés Eloy Seijas
68
refugio de fauna Silvestre, en los caños Guaritico y Setenta del
estado Apure. Esta localidad fue decretada como área protegida
específicamente para el caimán del Orinoco, pero padece de los
mismos problemas de vigilancia que los parques nacionales. Para el
río Cojedes, donde se encuentra la población más grande de C.
intermedius, no existe una figura jurídica que resguarde parte del
hábitat para la conservación de la especie en el largo plazo, pero
existen una propuesta orientada en ese sentido (Seijas, 2008).
El caimán de la costa no ha recibido en Venezuela la misma
atención que el caimán del Orinoco por parte de la comunidad
conservacionista internacional. Esto se debe en parte a que esta
especie tiene una amplia distribución en América, lo cual disminuye
su riesgo de extinción a escala continental. Sin embargo, en cierto
sentido, su situación en Venezuela es más preocupante que la de
C. intermedius, ya que para esta última especie su hábitat está
todavía en buena medida intacto, mientras que en el caso de C.
acutus, gran parte de su hábitat original ha desaparecido por
completo o está altamente degradado, situación que seguramente se
acentuará en el futuro. Varias localidades de protección estricta
(parques nacionales y refugios de fauna) se localizan a lo largo de
la costa venezolana, pero algunas de ellas poseen hábitats que
podrían catalogarse como marginales para el caimán, como es el
caso de los refugios de fauna de Los Olivitos (Zulia) y Cuare
(Falcón), así como el parque Nacional Morrocoy. Como ya se
señaló en otra parte, el hábitat óptimo del caimán de la costa son
los ríos y es precisamente en uno de ellos, el Yaracuy, donde se
ha encontrado la población más grande de la especie. No existen
políticas oficiales para la protección de esta localidad en particular,
ni para el fomento o restauración poblacional en otros ríos costeros
donde pequeñas poblaciones de este cocodrilo sobreviven a duras
penas (ríos Tocuyo, Neverí y Aroa) o han desaparecido por
completo (río Unare). El programa de restauración poblacional con
C. acutus a partir de animales criados en cautiverio ha sido más
bien modesto, como discutiremos en el Capítulo 8.
La baba es una especie muy abundante y el programa de
aprovechamiento que adelanta el MARNR no parece constituir una

Los Crocodylia de Venezuela
69
amenaza, siempre que se mantenga en los límites que ha mostrado
en los últimos años. Está incluida en el Apéndice II de CITES, lo
que indica que su comercio internacional está regulado. La cacería
para el consumo de su carne se ha intensificado en los últimos
años y esto pudiera ser un factor que afecte a varias poblaciones.
La información sobre los Paleosuchus es muy escasa, pero el
hecho de que habiten zonas por lo general despobladas hace
suponer que no están amenazadas de manera inminente. Se les
asigna la categoría “Insuficientemente conocido” en Libro Rojo de
la Fauna venezolana (Rodríguez y Rojas-Suárez, 1995). Los autores
mencionados señalan que la destrucción de hábitat por la minería
de oro y la contaminación mercurial asociada a esa actividad ha
causado un impacto severo en algunas áreas al sur del Orinoco. En
el caso particular de P. palpebrosus, la cacería de subsistencia y
las actividades de destrucción y contaminación de morichales en las
sabanas de Anzoátegui y Monagas, debido a la actividad petrolera,
son factores adversos a los que hay que prestarle atención en el
corto plazo.

Andrés Eloy Seijas
70

Los Crocodylia de Venezuela
71
CAPITULO 3
CAPTURA, MANIPULACIÓN Y MARCAJE
Algunos aspectos del comportamiento y la ecología de los
Crocodylia pueden ser estudiados sin necesidad de atraparlos, pero
hay cierta información que sólo es posible obtenerla con los
ejemplares al alcance de la mano. Aunque parezca obvio, es
necesario recalcar que los crocodilios pueden, si no son manejados
de manera adecuada, causar heridas a los investigadores que los
manipulan. Por otra parte, un mal manejo de los individuos
capturados puede provocarles daño o incluso la muerte.
Por otra parte, en muchos estudios, los individuos pueden o
deben ser capturados de manera repetida. En la mayor parte de
estos casos, la única manera de tener la certeza de que un animal
atrapado representa una recaptura, es que él esté identificado con
algún tipo de marca inconfundible. En este Capítulo se presentan un
conjunto de recomendaciones y consideraciones sobre las técnicas y
métodos para la captura, marcaje y manipulación de crocodilios.
CAPTURA
Existen muchas publicaciones que describen técnicas para la
captura de crocodilios (por ejemplo, Webb y Messel, 1977; Hutton
et al., 1987; Walsh, 1987 y Cherkiss et al., 2005). En esta
sección se enfatizará en los métodos usuales entre los inves-
tigadores que han trabajado en Venezuela. Sólo se describirán
técnicas para la captura de animales vivos.
Captura con la mano
El tamaño de los crocodilios varía mucho con las especies y,
sobre todo, con la edad de los ejemplares. Los recién nacidos de

Andrés Eloy Seijas
72
los Paleosuchus y de la baba apenas superan 20 cm; las crías de
C. acutus miden alrededor de 25 cm y las de C. intermedius
rara vez superan 30 cm. Para estos individuos pequeños la captura
directa con la mano no presenta ningún peligro, pero es necesario
tener siempre en cuenta que durante esa primera etapa de su vida
los crocodilios son cuidados y defendidos por sus madres, las
cuales en ocasiones pueden ser sumamente agresivas y peligrosas.
La protección de las crías por parte de las hembras se puede
prolongar mucho tiempo, meses en el caso de la baba, por lo que
la captura de animales pequeños, particularmente cuando todavía
forman grupos o “creches” debe llevarse a cabo después de
asegurarse que no se corre ningún riesgo de ataques por parte de
la madre o de algún otro adulto que se encuentre en las cercanías.
La captura con la mano, con un rápido movimiento al cuello,
todavía es relativamente sencillo de realizar con individuos cercanos
al metro en caso de ejemplares del género Crocodylus y de un
poco menos (70-80 cm) en el caso de la baba. Esta diferencia se
debe a que los caimanes son relativamente más delgados que las
babas y su cuello es más fácil de abarcar con los dedos para
asegurar el agarre. Por otra parte, C. crocodilus es una especie
más robusta y fuerte, y por lo tanto más difícil de manipular
inmediatamente después de la captura. Atrapar con la mano
animales más grandes que los señalados es todavía posible en
determinadas circunstancias favorables (profundidad del cuerpo de
agua, transparencia, posición del individuo con respecto a quien
quiera atraparlo, entre otras). Eso dependerá también de que tan
grandes sean las manos de la persona que intenta la tarea. En todo
caso, se debe tener en cuenta que ésta se torna más complicada y
riesgosa a medida que los animales son mayores. Gorzula, en su
estudio publicado en 1978, afirmó haber capturados todas las babas
por debajo de 1,2 m de LT con la mano. El record personal del
autor del presente libro es un caimán de la costa de 1,24 cm de
LT, capturado con ambas manos desde un bote en el embalse de
Tacarigua, en Falcón.
La captura con la mano es por lo general sólo posible durante
la noche, cuando se encandila al animal con la luz de una lámpara.
Esta técnica es más factible de llevar a cabo desde una embar-

Los Crocodylia de Venezuela
73
cación, ya que es poco probable que un ejemplar permita el
acercamiento suficiente cuando se le aproxima desde tierra, al
menos que el cuerpo de agua sea de pequeñas dimensiones y muy
somero, lo cual limitaría sus posibilidades de escape.
Captura con redes
La captura con redes de arrastre, los llamados “chinchorros” en
Venezuela, ha sido una técnica muy empleada en el caso de la
baba (Seijas, 1979; Ayarzagüena; 1983), especialmente en
préstamos y lagunas de pequeñas dimensiones donde esta especie
suele presentar densidades muy grandes hacia el final del periodo
de sequía. Seijas (1979) usó chinchorros con tamaño de trama de 1
cm. Las dimensiones de estas redes fueron de 50, 20 y 11 m de
largo por 2,50 m de altura. También usó una red de 12 m con
aberturas de malla de 5 cm, lo que evitaba captura innecesaria de
peces pequeños.
Los chinchorros son arrastrados a ras de fondo, preferiblemente
en cuerpos de agua que presentan orillas desprovistas de malezas,
lo que facilita el desplazamiento de las personas que llevan la red,
y áreas libres de vegetación acuática flotante o arraigada. De no
ser esta la situación, se requiere de un acondicionamiento previo de
las orillas del cuerpo de agua. Los préstamos de gran extensión y
profundidad sólo permiten un muestreo parcial, mientras los más
pequeños y someros pueden ser abarcados en su totalidad.
La técnica de captura con redes es muy efectiva en cuerpos de
agua que reúnan las características antes mencionadas. Tiene el
inconveniente de que en ciertas condiciones se produce la captura
no deseada de gran cantidad de peces, algunos de los cuales
pueden causar daños en las redes, como es el caso de los grandes
peces “caribe colorado” (Pygocentrus cariba) Los babos muy
grandes también pueden romper los chinchorros, por lo que el
método es más adecuado para ejemplares menores de 1,20 m. Otra
limitación del método es que requiere de la participación de varias
personas, particularmente cuando los chinchorros son muy largos.

Andrés Eloy Seijas
74
Captura con lazos y cuerdas
Los lazos corredizos, particularmente los fabricados con hilos de
alambre retorcido (las denominadas “guayas” en la jerga vene-
zolana) se han convertido en el método más usado y efectivo para
la captura de crocodilios. Existen varias compañías en los E.U.A. y
Canadá que las fabrican y vienen en distintas longitudes y grosores,
de acuerdo con las características (talla y peso) de los animales
que se pretenden atrapar. Las guayas son muy resistentes y
poseen un mecanismo de cierre que asegura al animal e impide su
escape.
La captura con guayas funciona muy bien de noche y cuando el
animal permite un acercamiento suficiente como para colocar el
lazo corredizo alrededor de su cuello (Fig. 3.1). Para aumentar el
alcance, los lazos se ajustan al extremo de una vara. En la mayoría
de las ocasiones una palo delgado de dos a tres metros de largo es
suficiente para llegar al crocodilio desde un bote cuando se trabaja
en un cuerpo de agua navegable, o desde tierra, cuando se trata de
préstamos u otro tipo de cuerpo de agua de pequeñas dimensiones
y poca profundidad. Cuando el dispositivo ha sido colocado alre-
dedor del cuello se levanta la vara con fuerza para provocar su
cierre. El extremo de la guaya opuesto al lazo debe estar ase-
gurado a una cuerda de varios metros de largo y resistente a la
tensión de los animales que se quieren capturar. El extremo libre
de la cuerda debe ser anudada al bote o al propio investigador para
impedir que el animal atrapado escape con el lazo ya ajustado a su
cuerpo. Es conveniente amarrar la guaya a la vara de manera que
se desprenda fácilmente. Esto evita que el palo se parta durante el
forcejeo del animal y pueda ser puesto de lado para facilitar el
maniobrar de la persona que lo atrapa.
Si el animal capturado es grande (por ejemplo más de 1,80 m)
es preferible dirigirse a una playa para arrastrarlo y manipularlo en
tierra. Individuos más pequeños pueden ser subidos al bote para la
toma de datos allí. Todo esto variará dependiendo que tan grande
sea el bote con el cual se trabaja y cuantas personas participan en
la captura.

Los Crocodylia de Venezuela
75
Figura 3.1 Captura de un caimán en el río Cojedes con un lazo corredizo
metálico (guaya).
Es posible preparar lazos corredizos con cuerdas, pero no son
tan efectivos como los metálicos. Para grandes cocodrilos cautivos,
como los adultos confinados en pequeños pozos en los zoocriaderos,
los lazos con cuerdas son efectivos, especialmente si se aseguran a
la mandíbula superior, donde los dientes facilitan el agarre.
Otros métodos de captura
Uno de los métodos más empleado para atrapar Crocodylia es
el de los arpones. Fue con este tipo de artefactos (Fig. 3.2a) que
se llevó a cabo la mayor parte de la caza de caimanes en la época
de explotación comercial masiva del siglo XX. Todavía se usa para

Andrés Eloy Seijas
76
la captura de babas con los mismos fines. Es sin duda el método
más afectivo para animales que van a ser sacrificados, ya que el
arpón puede ser arrojado desde cierta distancia, lo cual elimina la
necesidad de aproximación requerida en el caso del lazo corredizo
y el lugar de contacto no tiene que ser limitado al cuello. Para
capturas en las que se quiere causar el mínimo daño al animal
Webb y Messel (1977) recomiendan un tipo de arpón de doble
punta, el cual se ajusta al extremo de una lanza. El artefacto
contiene un orificio (Fig. 3.2b) por el que se introduce una cuerda,
para halar al animal una vez que ha sido atrapado. Los inves-
tigadores venezolanos no lo han usado, al menos no aparece
reseñado en la literatura.
También se reseñan en la literatura pinzas, trampas, dardos y
anzuelos para capturar crocodilios. No hay registro de su uso en
Venezuela con fines de investigación. Algunos de esos métodos
parecen muy engorrosos y de difícil aplicación en condiciones de
campo. Las trampas, por ejemplo, son generalmente muy grandes y
Figura 3.2 (a). Arpón usado para la caza de caimanes durante la época de
explotación comercial de mediados del siglo pasado (fotografía cortesía de
Robert Godshalk). (b): Arpón de doble punta mandado a fabricar por Jesús
Coty de acuerdo con las especificaciones de Webb y Messel (1977).

Los Crocodylia de Venezuela
77
necesitarían en ocasiones ser instaladas por varios días, ambas
cosas limitan su uso. Se han descrito también métodos que
producen parálisis por descargas eléctricas (Joanen y Perry, 1971;
Franklin et al., 2002) más apropiados para la captura de animales
en zoocriaderos o zoológicos.
MANIPULACIÓN
Una vez capturado un crocodilio, por cualquiera de los métodos
señalados en las secciones previas, es necesario someterlo para
tomar la información que se busca, para luego marcarlo y dejarlo
libre. Todo esto sin que se produzcan heridas entre las personas
que realizan el trabajo y que el animal retorne al ambiente sano y
salvo. Si la captura se produjo con la mano significa que el
individuo es lo suficientemente pequeño como para que la misma
persona que lo atrapó le cierre la boca con una cinta adhesiva o
una banda elástica. Debe tenerse cuidado de no tapar las aberturas
nasales del animal. Si no se van a tomar los datos de inmediato se
pueden colocar en sacos. No es necesario maniatarlos. Cuando se
manipulan para la toma de datos hay que agarrarlos siempre de
manera firme, para evitar que escape en un arranque súbito de
actividad. Cuando se toman medidas, es recomendable siempre
mantenerlos presionados contra una superficie plana y, prefe-
riblemente, con los ojos cubiertos.
La manipulación de animales un poco mayores, digamos entre
0,90 y 1,20 m en los caimanes y entre 0,80 y 1,20 en las babas y
Paleosuchus, es por lo general todavía muy sencilla y encierra
pocos riesgos, si se trabaja de la manera correcta. Cuando la
captura se realiza desde un bote, el ejemplar puede ser subido a la
embarcación evitando en lo posible que se golpee contra ésta y se
produzcan lesiones (pérdida de dientes, por ejemplo). La mejor
manera de subirlo al bote es que una de las personas hale de la
cuerda o lazo con que está sujeto al cuello en tanto que la otra le
toma la punta de la cola y entre ambas mantienen la tensión al
máximo. Luego se levanta y coloca sobre una superficie plana
donde se termina de dominar y se le amarra el hocico para

Andrés Eloy Seijas
78
mantenerlo cerrado. No se debe permitir nunca que el ejemplar
gire libremente su cuello, ya que podría morder a la persona que
sujeta la cola. El dominar a estos animales en tierra es mucho más
sencillo y sólo requiere que mientras una de las personas hala la
cuerda que sujeta al crocodilio, la otra le hala la cola en sentido
contrario, manteniendo la máxima tensión posible, hasta poder llegarle
al cuello y proceder a cerrar su boca con cuerdas o cintas adhesivas.
Para animales más grandes que los señalados, es preferible la
manipulación en tierra, aun cuando estos hayan sido enlazados
desde un bote. Mientras más grande sea el ejemplar mayor será el
número de personas requeridas para dominarlo (Fig. 3.3).
Cuando se trabaja con crocodilios hay que tratar que el tiempo
de manipulación sea lo más corto posible, particularmente con
ejemplares muy grandes. La lucha fuerte y prolongada durante la
captura puede provocarle la muerte, como consecuencia de una
severa acumulación de ácido láctico (Seymour et al., 1987).
Figura 3.3 Caimán del Orinoco de 4,30 m atrapado en el río Cojedes en
marzo de 2003. Para la captura de ese animal participaron seis personas.
Al frente aparece Brady Barr, de National Geographic, en el centro el
autor y atrás José Manuel Mendoza.

Los Crocodylia de Venezuela
79
El maniatar animales sólo se justifica cuando es necesario
improvisar un medio para inmovilizarlos (para trasladarlos a otro
sitio cercano, por ejemplo). Los amarres prolongados pueden causar
muertes de tejidos y, por lo tanto, mutilaciones. Para trasladar
grandes animales a lugares distantes, en viajes que requieren
muchas horas, es preferible preparar cajas donde estos puedan ser
introducidos y permanecer adentro con movilidad restringida, con
sólo el hocico amarrado y los ojos y oídos tapados (Fig. 3.4).
Figura 3.4 Cajón usado para el traslado de una caimana de 2,20 m desde la
ciudad de Barinas hasta el caño Guaritico, estado Apure, abril 2006. En
la fotografía aparecen Carlos Chávez y un trabajador del Hato Turagua.
Cuando se capturan varios animales para tomar sus datos
posteriormente, a veces después de varias horas, es necesario
colocarlos en lugares frescos y a la sombra. Si deben ser trasla-
dados en carro a lugares muy distantes se debe asegurar que los
individuos no queden sobre superficies que se recalienten y
esporádicamente deben ser rociados con agua para refrescarlos y
evitar la deshidratación.

Andrés Eloy Seijas
80
La manipulación de animales pudiera facilitarse y ser más
segura con el uso de drogas. No existen publicaciones sobre
experiencias de este tipo en el país, pero hay literatura abundante
sobre el tema (por ejemplo, Brisbin, 1966; Messel y Stephens, 1980;
Loveridge y Blake, 1987; Seashole et al., 2000).
MARCAJE
En la mayoría de las ocasiones en las que se atrapan crocodilios
es necesario, o recomendable, marcarlos para su reconocimiento en
caso de que ocurra una recaptura y ello permita obtener infor-
mación sobre los cambios experimentados en su condición física
(talla, peso, heridas) o su desplazamiento con respecto al lugar de
captura previa. El marcaje de los crocodilios es además una tarea
rutinaria en los zoocriaderos.
Lo ideal de un método de marcaje es que sea permanente y
fácil de reconocer por cualquiera que realice la recaptura. La
mutilación de escamas de las crestas de la cola cumple muy bien
la primera condición, siempre que los cortes sean lo suficientemente
profundos para que el recrecimiento, el cual siempre ocurre en una
pequeña medida, no permita que se confundan con crestas intactas.
Este tipo de marcaje se ha empleado ampliablemente tanto en
babas como en caimanes, pero es en estas últimas especies en las
que su uso se ha extendido.
Para facilitar la identificación de un individuo se le puede cortar
una o varias de las escamas de las crestas caudales dobles o
sencillas. Hay tres códigos que han sido usados en Venezuela,
cada uno de ellos implantado por los investigadores que han tenido
bajo su responsabilidad los primeros zoocriaderos de caimanes
establecidos en el país; a saber: Andrés E. Seijas (UNELLEZ),
John Thorbjarnarson (Masaguaral) y José Ayarzagüena (El Frío)
(Fig. 3.5).
Las mutilaciones de escamas funcionan bien cuando el número
de individuos por identificar es relativamente pequeño. Por ejemplo,
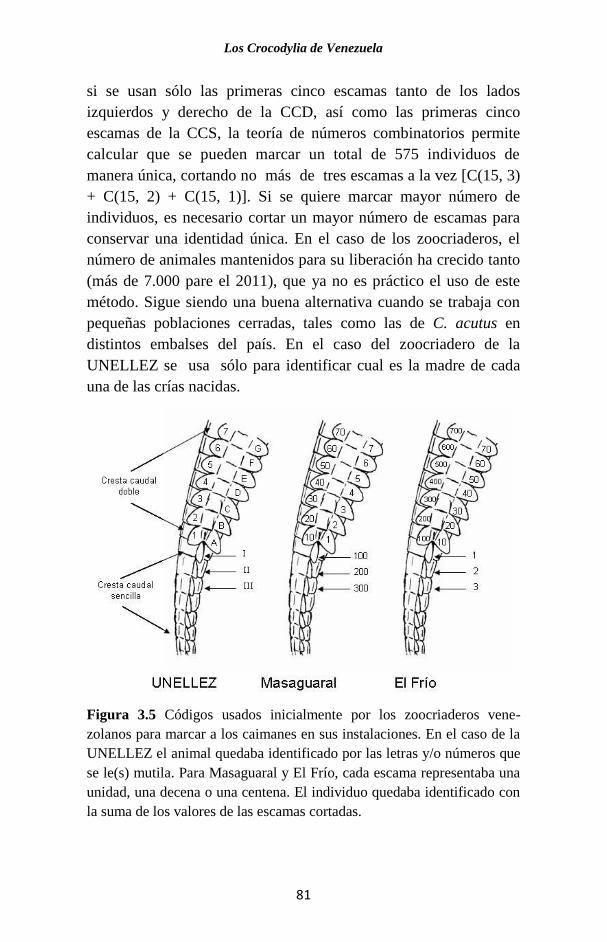
Los Crocodylia de Venezuela
81
si se usan sólo las primeras cinco escamas tanto de los lados
izquierdos y derecho de la CCD, así como las primeras cinco
escamas de la CCS, la teoría de números combinatorios permite
calcular que se pueden marcar un total de 575 individuos de
manera única, cortando no más de tres escamas a la vez [C(15, 3)
+ C(15, 2) + C(15, 1)]. Si se quiere marcar mayor número de
individuos, es necesario cortar un mayor número de escamas para
conservar una identidad única. En el caso de los zoocriaderos, el
número de animales mantenidos para su liberación ha crecido tanto
(más de 7.000 pare el 2011), que ya no es práctico el uso de este
método. Sigue siendo una buena alternativa cuando se trabaja con
pequeñas poblaciones cerradas, tales como las de C. acutus en
distintos embalses del país. En el caso del zoocriadero de la
UNELLEZ se usa sólo para identificar cual es la madre de cada
una de las crías nacidas.
Figura 3.5 Códigos usados inicialmente por los zoocriaderos vene-
zolanos para marcar a los caimanes en sus instalaciones. En el caso de la
UNELLEZ el animal quedaba identificado por las letras y/o números que
se le(s) mutila. Para Masaguaral y El Frío, cada escama representaba una
unidad, una decena o una centena. El individuo quedaba identificado con
la suma de los valores de las escamas cortadas.

Andrés Eloy Seijas
82
Ya se había señalado que el método de las mutilaciones de
escamas tiene la virtud de ser permanente, pero el uso de distintos
códigos ha causado algunas dificultades en la identificación de
individuos recapturados. Otro problema del método es que en
ocasiones pueden encontrarse individuos con mutilaciones naturales
que causan confusión.
Gorzula (1978) empleó un sistema de marcaje por remoción de
escama de la parte ventral de la cola, las cuales permanecieron
reconocibles por lo menos 26 meses.
El procedimiento más extendido para el marcaje de crocodilios
es el que consiste en la colocación de placas numeradas en las
membranas interdigitales. Este método tiene la gran ventaja de que
el número, a veces acompañado por unas siglas, es reconocible por
cualquier persona que efectúe la recaptura con poco riesgo de
equivocación. La gran limitación es que las placas pueden despren-
derse, aunque se conocen registros de animales recapturados que
las conservaban más de siete años después de la liberación
(Chávez, 2000; Llobet, 2002). Es conveniente colocar al menos dos
placas a los animales que se quiere identificar, ya que la pro-
babilidad de que pierda dos placas es menor que la de perder sólo
una. En ocasiones la pérdida de la placa se produce cuando el
crocodilio es mutilado por mordidas de otro animal, con frecuencia
un caribe (Fig. 3.6).
MARCAS NATURALES
Al igual que ocurre con los seres humanos, cada individuo de
una especie de crocodilio tiene un conjunto de rasgos que le son
exclusivos, únicos, que podrían permitirnos distinguirlos de manera
inequívoca de otros individuos de la misma especie. Esas carac-
terísticas distintivas pueden ser un patrón de coloración, algún tipo
de deformidad, alguna mutilación, algún patrón de escamación
peculiar o la combinación de varios de estos rasgos.
Cuando se trata de pocos animales, como por ejemplo los
escasos adultos en un zoocriadero, los rasgos distintivos de cada
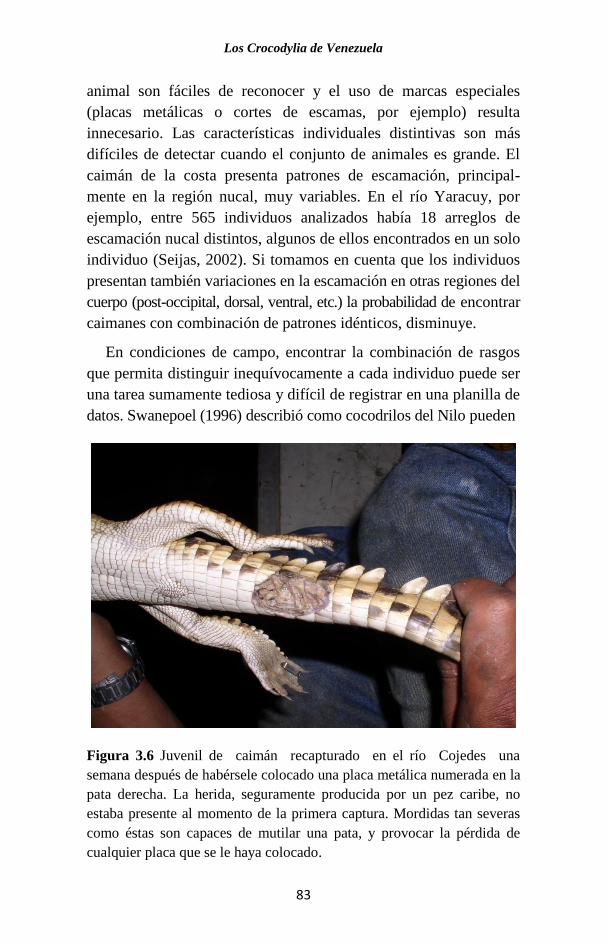
Los Crocodylia de Venezuela
83
animal son fáciles de reconocer y el uso de marcas especiales
(placas metálicas o cortes de escamas, por ejemplo) resulta
innecesario. Las características individuales distintivas son más
difíciles de detectar cuando el conjunto de animales es grande. El
caimán de la costa presenta patrones de escamación, principal-
mente en la región nucal, muy variables. En el río Yaracuy, por
ejemplo, entre 565 individuos analizados había 18 arreglos de
escamación nucal distintos, algunos de ellos encontrados en un solo
individuo (Seijas, 2002). Si tomamos en cuenta que los individuos
presentan también variaciones en la escamación en otras regiones del
cuerpo (post-occipital, dorsal, ventral, etc.) la probabilidad de encontrar
caimanes con combinación de patrones idénticos, disminuye.
En condiciones de campo, encontrar la combinación de rasgos
que permita distinguir inequívocamente a cada individuo puede ser
una tarea sumamente tediosa y difícil de registrar en una planilla de
datos. Swanepoel (1996) describió como cocodrilos del Nilo pueden
Figura 3.6 Juvenil de caimán recapturado en el río Cojedes una
semana después de habérsele colocado una placa metálica numerada en la
pata derecha. La herida, seguramente producida por un pez caribe, no
estaba presente al momento de la primera captura. Mordidas tan severas
como éstas son capaces de mutilar una pata, y provocar la pérdida de
cualquier placa que se le haya colocado.

Andrés Eloy Seijas
84
ser distinguidos individualmente por las marcas en su cola. A
manera de propuesta se presentan a continuación los resultados
preliminares de un intento de distinguir a caimanes del Orinoco
inspirado en los resultados del mencionado autor.
Todos los crocodilios presentan en los laterales de la cola
manchas oscuras sobre un fondo más claro. La manera exacta en
que se disponen estas manchas es probablemente única en cada
ejemplar. El describir en palabras el patrón de coloración de cada
individuo puede ser una tarea engorrosa. El registro en fotografías
es probablemente el método más confiable, pero si el número de
individuos es muy grande, el conseguir el patrón que se busca en
un catálogo de fotos puede requerir de mucho tiempo. Una manera
de facilitar el trabajo es convertir el patrón en un número, al crear
para ello una especie de “código de barras”. En la Figura 3.7 se
muestra la fotografía de parte de la cola de un caimán del
Orinoco. Las escamas en las primeras tres filas y seis columnas en
la vista lateral de la CCD, usadas para el desarrollo del código,
fueron delineadas con un color claro para facilitar su identificación.
Si una escama no posee una mancha oscura se le asigna un valor
de cero. Si la mancha está presente se le asigna el valor de 1, 2 ó
4, dependiendo si está en la primera, segunda o tercera fila,
respectivamente. Para cada columna de escamas se obtiene una
cifra equivalente a la suma del valor de sus tres escamas. Esta
cifra va desde cero, cuando ninguna de las escamas posee
manchas, hasta siete (1+2+4=7), cuando las tres la poseen. El
código final de un individuo es el que resulta de colocar en forma
sucesiva las cifras correspondientes a las seis columnas.
En una muestra de 190 caimanes del zoocriadero de la
UNELLEZ de la cohorte del 2002, se encontró que había sólo
cinco individuos (2,6%) con patrones similares, tanto si se consi-
deraba el lado derecho como el izquierdo de la cola. No hubo
individuos que compartieran los mismos patrones en ambos lados.
Aunque el método antes señalado se puede considerar como
promisorio, su utilidad quizá quede restringida al manejo de
poblaciones pequeñas y como una marca complementaria a la
suministrada por las placas metálicas y cortes de escamas.

Los Crocodylia de Venezuela
85
Figura 3.7 Método para asignarle una “etiqueta” a un caimán del
Orinoco de acuerdo con el patrón de manchas en la cola. La cifra se
antecede con la letra “D” para indicar que se trata del lado derecho de la
cola. Ver explicaciones en el texto. La flecha señala un corte en la escama
“V” (cinco romano) que, en el caso de la UNELLEZ, identifica al ejemplar
como hijo de la hembra llamada “La Negra”.

Andrés Eloy Seijas
86

Los Crocodylia de Venezuela
87
CAPÍTULO 4
MORFOMETRÍA
Los estudios morfométricos en crocodilios generalmente tienen
como propósito elaborar modelos matemáticos que permitan pre-
decir alguna medida corporal a partir de otra. En muchos casos,
por ejemplo, la cola de los ejemplares capturados en el campo está
rota y la longitud total (LT) no corresponde con la que el animal
tendría si tuviera su cola intacta. En este caso, y dependiendo de
los objetivos del investigador, la LT podría estimarse a partir de la
ecuación de regresión obtenida al relacionar otra medida corporal,
digamos la longitud de la cabeza o la longitud cabeza-cloaca
(LCC), con la LT de animales con cola completa. De la misma
manera, el peso de los animales muy grandes es difícil de medir en
el campo y, nuevamente, se puede estimar a partir de modelos
elaborados previamente a partir de animales con datos completos.
En ocasiones, durante las evaluaciones de campo, una impor-
tante fuente de información sobre la presencia, abundancia o
estructura poblacional de una especie es la que se obtiene a partir
de datos indirectos, como pudieran ser las huellas (Wilkinson y
Rice, 1996). Modelos de regresión que relacionan el tamaño de
estas huellas con la talla del animal son a veces la única manera
de determinar con cierta precisión el tamaño de algunos de los
animales presentes en un área.
Para análisis forenses, estimar la talla de un animal atrapado por
cazadores furtivos a partir de sus restos (cráneo, por ejemplo) o la
talla del animal adulto a partir de las dimensiones de parte del
cuerpo, puede facilitar los procesos de investigación de la guardería
ambiental y servir como pruebas en juicios por delitos ambientales.

Andrés Eloy Seijas
88
Las relaciones morfométricas entre distintas partes del cuerpo
varían entre especies y subespecies de crocodilios y pueden ser
usadas como caracteres diagnósticos para separarlas. Las dife-
rencias, particularmente en la forma del hocico, pueden ser usadas
incluso para efectuar inferencias ecológicas sobre el tipo de dieta y
tipo de ambiente en que habitan: las especies con hocicos muy
largos y delgados ocupan principalmente ríos y presentan una dieta
esencialmente piscívora y los de hocicos más anchos ocupan áreas
de pantano y consumen presas menos movibles y de cuerpo duro
(Ayarzagüena, 1984).
En los últimos años las relaciones morfométricas entre distintas
partes del cuerpo y el peso han sido usadas para elaborar índices
de condición física, los cuales permiten comparar la gordura relativa
de individuos y poblaciones que están en condiciones ecológicas
diferentes, o para detectar cambios de la gordura relativa de la
misma población en distintos períodos climáticos (Seijas et al.,
2003).
En este Capítulo se presentan y discuten, a la luz de las
consideraciones antes expuestas, algunos análisis morfométricos con
las cinco especies de crocodilios de Venezuela. Se usaron regre-
siones lineales para generar las ecuaciones que permiten estimar el
tamaño o dimensión de alguna estructura corporal a partir de otra,
es decir:
XbaY 11
Donde Y es el tamaño de la estructura del cuerpo que se quiere
predecir a partir del tamaño X de otra estructura; a es el
intercepto de la recta en el eje Y, y b es la pendiente de la recta,
es decir, cuantas veces se incrementa Y por cada incremento de
X.
Si se realizan los análisis de regresión con los logaritmos de las
variables, se obtiene una nueva ecuación lineal de la forma:
XbaY loglog 22

Los Crocodylia de Venezuela
89
Tomando los antilogaritmos, esta última ecuación se puede
convertir a su vez en:
2
3
b
XaY
La cual es la ecuación general del crecimiento (Le Cren, 1951;
Jolicoeur, 1963; Gould, 1966; Dodson, 1975). En esta última
ecuación, a3 es el antilogaritmo de a2; el exponente b2 nos indica si
las dos estructuras corporales que son analizadas cambian o no en
la misma proporción a medida que el individuo crece. Si b2 no
difiere estadísticamente de 1, se habla de isometría perfecta, es
decir: ambas partes del cuerpo crecen en la misma proporción.
Cuando b2 es distinto de 1, existe alometría, lo que significa que
las tasas de crecimiento de las partes del cuerpo analizadas no
cambian en la misma proporción a medida que el animal crece.
Cuando se relaciona una magnitud lineal con el peso (o el
volumen), el valor del exponente b2 para perfecta isometría es 3.
TOMA DE DATOS
Antes que nada es necesario precisar cómo se toman las
medidas que se usarán en cada uno de los análisis subsiguientes:
o Longitud total (LT), desde la punta del hocico hasta el
extremo de la cola. Para la toma de esta medida es reco-
mendable que se anote además el número de filas de la cresta
caudal sencilla (CCS), ya que eso nos indicará si el animal tiene la
cola completa o no. Asimismo se recomienda tomar la medida
colocando la cinta métrica por debajo del animal y en una
superficie plana.
o Longitud cabeza-cuerpo (LCC), desde la punta del hocico
hasta la parte posterior de la abertura cloacal. Al igual que para
la LT, se recomienda colocar la cinta métrica debajo del animal
y ladear ligeramente su cuerpo para anotar la medida donde
termina la cloaca.

Andrés Eloy Seijas
90
o Ancho de la cabeza (Acab). La parte más ancha de la
tabla craneal.
o Longitud de la cabeza, desde el borde exterior del hueso
supraoccipital hasta la punta del hocico.
o Longitud del hocico (Lhoc), desde el borde anterior de la
órbita ocular hasta la punta del hocico.
o Ancho de la punta del hocico (Aph), tomada en la parte
más ancha de los huesos maxilares, al nivel de los dientes
maxilares 4to
(para el caso de los aligatóridos) ó 5to
(para los
crocodílidos), dependiendo de la especie.
o Longitud de la planta de la pata trasera (LP), hasta la
punta del dedo más largo.
o Peso. La precisión del instrumento usado debe aumentar a
medida que disminuye el peso del individuo. Se recomienda una
precisión de un gramo para individuos menores de 100 g; 10 g
para individuos entre 100 y 1000 g; 50 g para aquellos entre
1001 g y 5 kg y 100 g para los comprendidos entre este último
peso y 20 kg. Una precisión de 0,5 kg es suficiente para
crocodilios grandes.
En los dibujos de la Figura 4.1, se describen gráficamente las
medidas señaladas. Al menos que se diga lo contrario, se expre-
sarán todas las longitudes en milímetros y el peso en gramos. Estas
medidas no necesariamente coinciden con las recomendadas por
otros investigadores. Medem (1981) y Ayarzagüena (1983), mi-
dieron la longitud total (LT) desde la punta del hocico hasta la
parte anterior de la cloaca; la mayor parte de los investigadores,
sin embargo, toman esa medida hasta el final de la cloaca. Es por
esa razón que se recomienda precaución a la hora de tratar de
comparar los análisis morfométricos reportados en distintas publi-
caciones, como se verá más adelante.
RELACIONES MORFOMÉTRICAS EN BABAS
Existen varios estudios sobre babas en Venezuela que presentan
y discuten algunos análisis morfométricos. Staton y Dixon (1975),

Los Crocodylia de Venezuela
91
Figura 4.1 Descripción de las medidas morfométricas tomadas a los
crocodilios. Los dibujos son modificaciones de originales tomados de
Natterer (1841) y Medem (1981).
por medio de regresiones lineales, calcularon las ecuaciones para
predecir la LT a partir de la LCC, así como para predecir ambas
medidas a partir de la longitud del hocico. Gorzula (1978), por su
parte, generó con datos de individuos de cola completa, modelos
para predecir la longitud de la cola a partir de la LCC. Seijas
(1979) realizó análisis similares y añadió uno, al usar las trans-
formaciones logarítmicas para relacionar el peso con la LCC.
Ayarzagüena (1983) comparó los modelos de regresión al tomar los
logaritmos de diversos pares de medidas para hembras y machos.
Muchos de los análisis mencionados por este último autor pre-
sentan coeficientes de determinación (R2) altos, generalmente por
encima de 0,95 y pueden ser usados para predecir las dimensiones
de alguna medida a partir de otra.
El estudio con babas de Ayarzagüena (1983) le llevó a concluir
que existe dimorfismo sexual en esta especie respecto a la forma
del ojo, el cual sería en las hembras más grande y redondeado. En
ninguno de los análisis morfométricos que se analizarán a

Andrés Eloy Seijas
92
continuación se buscó establecer la existencia de dimorfismo
sexual en Caiman crocodilus (ni en ninguna de las otras especies
estudiadas). La única obvia diferencia entre los sexos es que los
machos alcanzan dimensiones mayores que las hembras, pero hay
que advertir que la muestra estudiada sólo registró cuatro babas
llaneras por encima de los dos metros y apenas una de las babas
de la región costera superó 1,80 m. Por esa razón, en ninguno de
los análisis se discriminó por sexo. Es posible que algunas dife-
rencias morfológicas entre machos y hembras se manifiesten sólo
cuando se comparan los animales más grandes de ambos sexos,
como señalaron Webb y Messel (1978) para Crocodylus porosus.
Lo que sí quedó claro es que hay diferencias, por lo menos en
los tamaños relativos de algunas partes del cuerpo, entre las babas
de la región costera (supuestamente Caiman crocodilus fuscus) y
las de la de los llanos (Caiman crocodilus crocodilus) por lo que
los análisis de las babas de las distintas zonas se efectuaron por
separado.
Para los análisis en que una de las dimensiones corporales
usadas era la longitud total (LT) se excluyeron los individuos con
menos de 18 CCS (es decir, con cola rota). Un animal con la cola
completa tiene una longitud corporal (LCC) que equivale aproxi-
madamente a 50% de la LT. Esa relación aumenta en la medida
en la cual se encuentre mutilada la cola, al punto que para un
individuo que haya perdido todas sus CCS, la LCC representa
alrededor de 62% de su LT (Fig. 4.2).
En el Apéndice I se presentan las ecuaciones resultantes de los
análisis de regresión entre distintos pares de variables morfo-
métricas en babas. Los altos coeficientes de determinación (R2)
nos indican que los cambios en tamaño de las distintas medidas
corporales durante el crecimiento del animal, guardan una relación
esencialmente lineal. Cuando se realizan los análisis de regresión
con los datos transformados a logaritmos, en algunos casos el
ajuste mejora ligeramente. Este último procedimiento permite
además establecer si existe algún tipo de crecimiento alométrico, es
decir, si las distintas partes del cuerpo analizadas crecen a
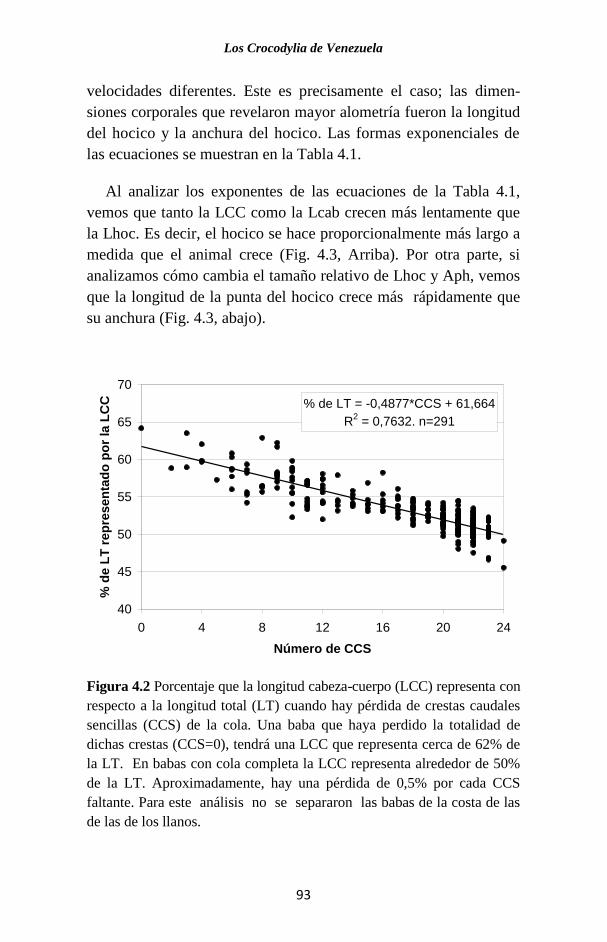
Los Crocodylia de Venezuela
93
velocidades diferentes. Este es precisamente el caso; las dimen-
siones corporales que revelaron mayor alometría fueron la longitud
del hocico y la anchura del hocico. Las formas exponenciales de
las ecuaciones se muestran en la Tabla 4.1.
Al analizar los exponentes de las ecuaciones de la Tabla 4.1,
vemos que tanto la LCC como la Lcab crecen más lentamente que
la Lhoc. Es decir, el hocico se hace proporcionalmente más largo a
medida que el animal crece (Fig. 4.3, Arriba). Por otra parte, si
analizamos cómo cambia el tamaño relativo de Lhoc y Aph, vemos
que la longitud de la punta del hocico crece más rápidamente que
su anchura (Fig. 4.3, abajo).
Figura 4.2 Porcentaje que la longitud cabeza-cuerpo (LCC) representa con
respecto a la longitud total (LT) cuando hay pérdida de crestas caudales
sencillas (CCS) de la cola. Una baba que haya perdido la totalidad de
dichas crestas (CCS=0), tendrá una LCC que representa cerca de 62% de
la LT. En babas con cola completa la LCC representa alrededor de 50%
de la LT. Aproximadamente, hay una pérdida de 0,5% por cada CCS
faltante. Para este análisis no se separaron las babas de la costa de las
de las de los llanos.
% de LT = -0,4877*CCS + 61,664
R2 = 0,7632. n=291
40
45
50
55
60
65
70
0 4 8 12 16 20 24
Número de CCS
% d
e L
T r
ep
res
en
tad
o p
or
la L
CC
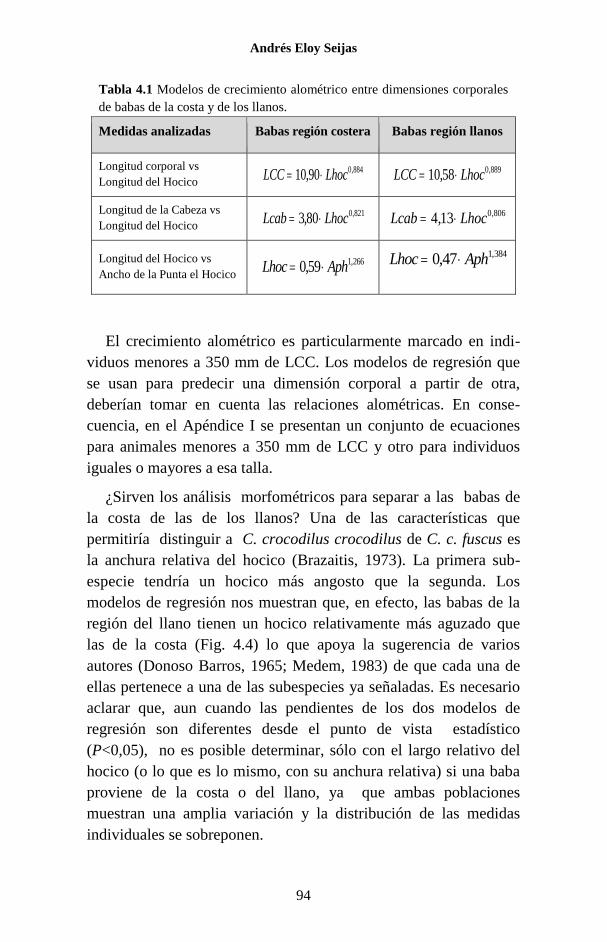
Andrés Eloy Seijas
94
Tabla 4.1 Modelos de crecimiento alométrico entre dimensiones corporales
de babas de la costa y de los llanos.
Medidas analizadas Babas región costera Babas región llanos
Longitud corporal vs
Longitud del Hocico LCC Lhoc 10 90 0 884, ,
LCC Lhoc 10 58 0 889, ,
Longitud de la Cabeza vs
Longitud del Hocico Lcab Lhoc 3 80 0 821, ,
Lcab Lhoc 4 13 0 806, ,
Longitud del Hocico vs
Ancho de la Punta el Hocico Lhoc Aph 0 59 1 266, ,
Lhoc Aph 0 47 1 384, ,
El crecimiento alométrico es particularmente marcado en indi-
viduos menores a 350 mm de LCC. Los modelos de regresión que
se usan para predecir una dimensión corporal a partir de otra,
deberían tomar en cuenta las relaciones alométricas. En conse-
cuencia, en el Apéndice I se presentan un conjunto de ecuaciones
para animales menores a 350 mm de LCC y otro para individuos
iguales o mayores a esa talla.
¿Sirven los análisis morfométricos para separar a las babas de
la costa de las de los llanos? Una de las características que
permitiría distinguir a C. crocodilus crocodilus de C. c. fuscus es
la anchura relativa del hocico (Brazaitis, 1973). La primera sub-
especie tendría un hocico más angosto que la segunda. Los
modelos de regresión nos muestran que, en efecto, las babas de la
región del llano tienen un hocico relativamente más aguzado que
las de la costa (Fig. 4.4) lo que apoya la sugerencia de varios
autores (Donoso Barros, 1965; Medem, 1983) de que cada una de
ellas pertenece a una de las subespecies ya señaladas. Es necesario
aclarar que, aun cuando las pendientes de los dos modelos de
regresión son diferentes desde el punto de vista estadístico
(P<0,05), no es posible determinar, sólo con el largo relativo del
hocico (o lo que es lo mismo, con su anchura relativa) si una baba
proviene de la costa o del llano, ya que ambas poblaciones
muestran una amplia variación y la distribución de las medidas
individuales se sobreponen.
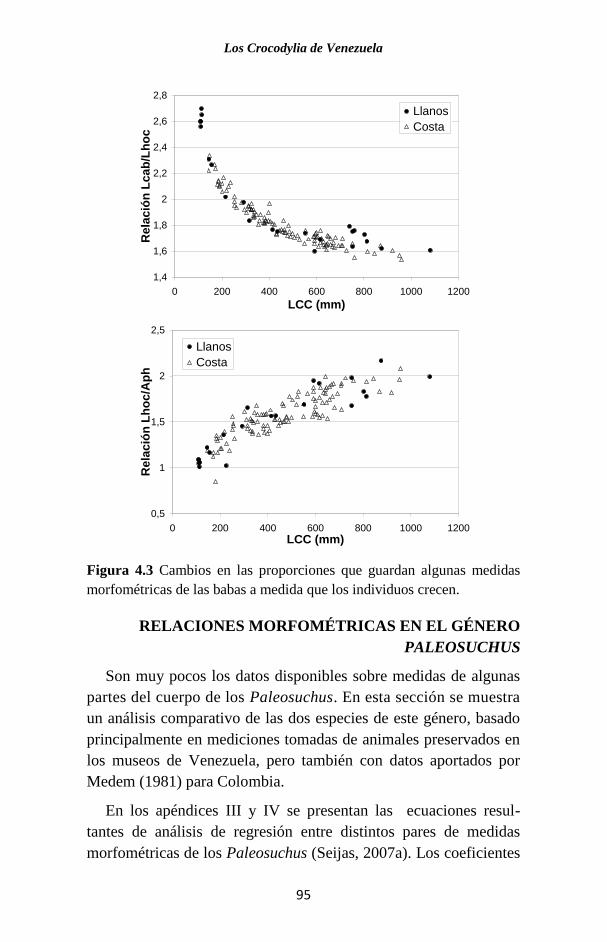
Los Crocodylia de Venezuela
95
Figura 4.3 Cambios en las proporciones que guardan algunas medidas
morfométricas de las babas a medida que los individuos crecen.
RELACIONES MORFOMÉTRICAS EN EL GÉNERO
PALEOSUCHUS
Son muy pocos los datos disponibles sobre medidas de algunas
partes del cuerpo de los Paleosuchus. En esta sección se muestra
un análisis comparativo de las dos especies de este género, basado
principalmente en mediciones tomadas de animales preservados en
los museos de Venezuela, pero también con datos aportados por
Medem (1981) para Colombia.
En los apéndices III y IV se presentan las ecuaciones resul-
tantes de análisis de regresión entre distintos pares de medidas
morfométricas de los Paleosuchus (Seijas, 2007a). Los coeficientes
1,4
1,6
1,8
2
2,2
2,4
2,6
2,8
0 200 400 600 800 1000 1200
LCC (mm)
Rela
ció
n L
ca
b/L
ho
c
Llanos
Costa
0,5
1
1,5
2
2,5
0 200 400 600 800 1000 1200
LCC (mm)
Rela
ció
n L
ho
c/A
ph
Llanos
Costa
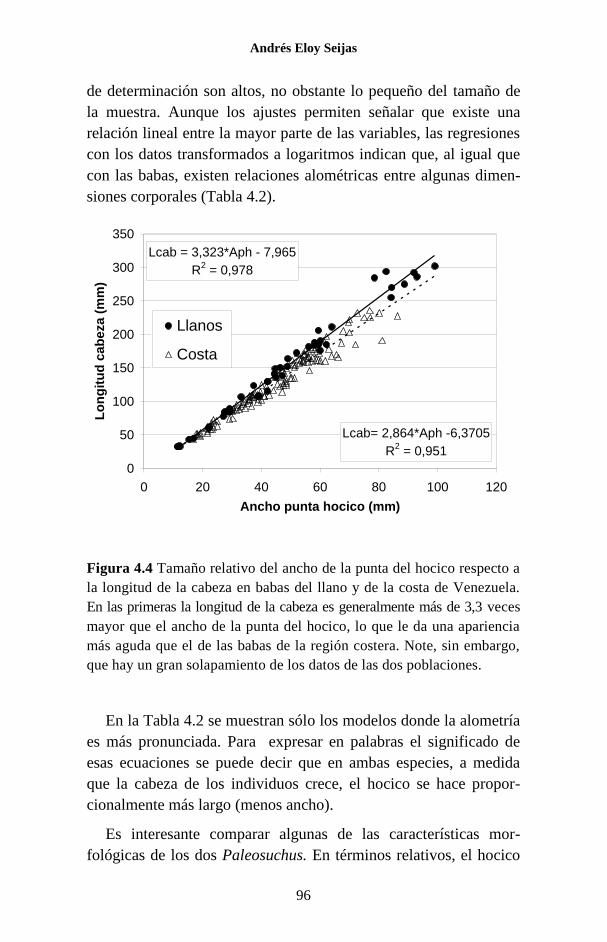
Andrés Eloy Seijas
96
de determinación son altos, no obstante lo pequeño del tamaño de
la muestra. Aunque los ajustes permiten señalar que existe una
relación lineal entre la mayor parte de las variables, las regresiones
con los datos transformados a logaritmos indican que, al igual que
con las babas, existen relaciones alométricas entre algunas dimen-
siones corporales (Tabla 4.2).
Figura 4.4 Tamaño relativo del ancho de la punta del hocico respecto a
la longitud de la cabeza en babas del llano y de la costa de Venezuela.
En las primeras la longitud de la cabeza es generalmente más de 3,3 veces
mayor que el ancho de la punta del hocico, lo que le da una apariencia
más aguda que el de las babas de la región costera. Note, sin embargo,
que hay un gran solapamiento de los datos de las dos poblaciones.
En la Tabla 4.2 se muestran sólo los modelos donde la alometría
es más pronunciada. Para expresar en palabras el significado de
esas ecuaciones se puede decir que en ambas especies, a medida
que la cabeza de los individuos crece, el hocico se hace propor-
cionalmente más largo (menos ancho).
Es interesante comparar algunas de las características mor-
fológicas de los dos Paleosuchus. En términos relativos, el hocico
Lcab = 3,323*Aph - 7,965
R2 = 0,978
Lcab= 2,864*Aph -6,3705
R2 = 0,951
0
50
100
150
200
250
300
350
0 20 40 60 80 100 120
Ancho punta hocico (mm)
Lo
ng
itu
d c
ab
eza (
mm
)
Llanos
Costa
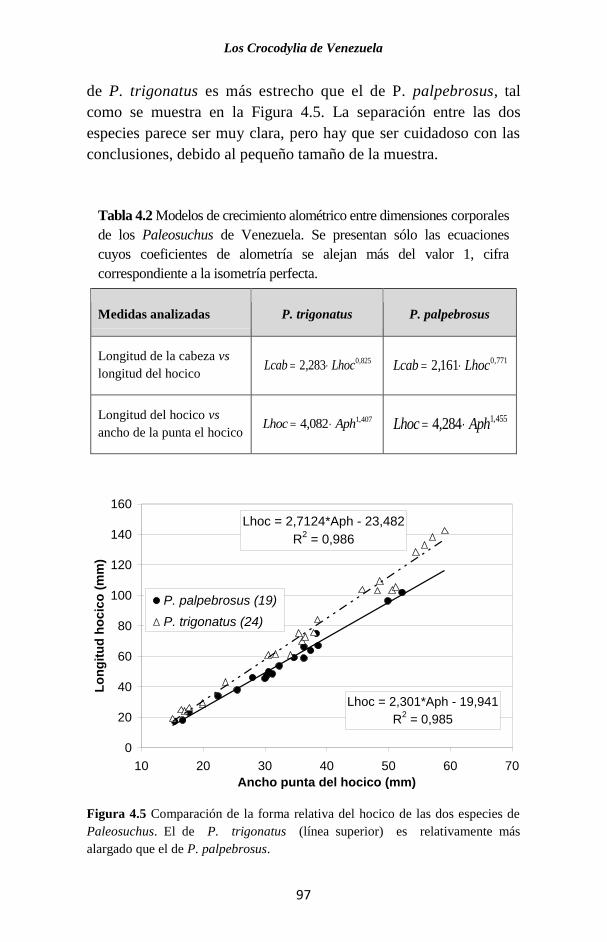
Los Crocodylia de Venezuela
97
de P. trigonatus es más estrecho que el de P. palpebrosus, tal
como se muestra en la Figura 4.5. La separación entre las dos
especies parece ser muy clara, pero hay que ser cuidadoso con las
conclusiones, debido al pequeño tamaño de la muestra.
Tabla 4.2 Modelos de crecimiento alométrico entre dimensiones corporales
de los Paleosuchus de Venezuela. Se presentan sólo las ecuaciones
cuyos coeficientes de alometría se alejan más del valor 1, cifra
correspondiente a la isometría perfecta.
Medidas analizadas P. trigonatus P. palpebrosus
Longitud de la cabeza vs
longitud del hocico Lcab Lhoc 2 283 0 825, , Lcab Lhoc 2 161 0 771, ,
Longitud del hocico vs
ancho de la punta el hocico Lhoc Aph 4 082 1 407, , Lhoc Aph 4 284 1 455, ,
Figura 4.5 Comparación de la forma relativa del hocico de las dos especies de
Paleosuchus. El de P. trigonatus (línea superior) es relativamente más
alargado que el de P. palpebrosus.
Lhoc = 2,301*Aph - 19,941
R2 = 0,985
Lhoc = 2,7124*Aph - 23,482
R2 = 0,986
0
20
40
60
80
100
120
140
160
10 20 30 40 50 60 70
Ancho punta del hocico (mm)
Lo
ng
itu
d h
oc
ico
(m
m)
P. palpebrosus (19)
P. trigonatus (24)

Andrés Eloy Seijas
98
Otra manera de ver las diferencias en la forma del hocico de
los Paleosuchus es elaborando un “índice de anchura relativa”
(IAR). Para ello se realizó un análisis de regresión entre la Lhoc y
el Aph, con los datos de ambas especies en conjunto (Fig. 4.6,
izquierda). El IAR se obtiene al dividir la Lhoc real entre la Lhoc
estimada por el modelo matemático obtenido en al análisis anterior.
Los animales representados por los puntos que quedan por encima
de la línea de regresión tienen un hocico más largo que lo que
predice el modelo, mientras que los representados por puntos
debajo de dicha línea los tienen más cortos. Este procedimiento es
análogo al que se obtendría si se toman los residuales del análisis
de regresión, sólo que los residuales se distribuyen alrededor del
valor cero (0), mientras que el IAR está centrado en uno (1). El
ancho relativo del hocico de los Paleosuchus no se mantiene
constante, sino que varía con el tamaño de los individuos (Fig. 4.6,
derecha).
RELACIONES MORFOMÉTRICAS EN EL GÉNERO
CROCODYLUS
Los caimanes de la costa y del Orinoco (como todos los
verdaderos cocodrilos) tienen una apariencia general del cuerpo
muy similar. Además de la utilidad práctica que puedan tener las
ecuaciones obtenidas por análisis de regresión de pares de variables
morfométricas (para estimar la dimensión de una medida a partir
de otra, por ejemplo), es interesante conocer aquellas medidas que
permitan diferenciar a estas especies.
De manera similar como se procedió con las babas y los
Paleosuchus, para los análisis en que se utilizó la LT se des-
cartaron aquellos individuos con menos de 17 CCS, a los cuales se
consideró con cola rota. La pérdida de parte de la cola trae como
consecuencia una disminución de la longitud total de los animales y
de cambios en la proporción relativa de los tamaños de esta
medida con las de otras partes del cuerpo (Fig. 4.7).
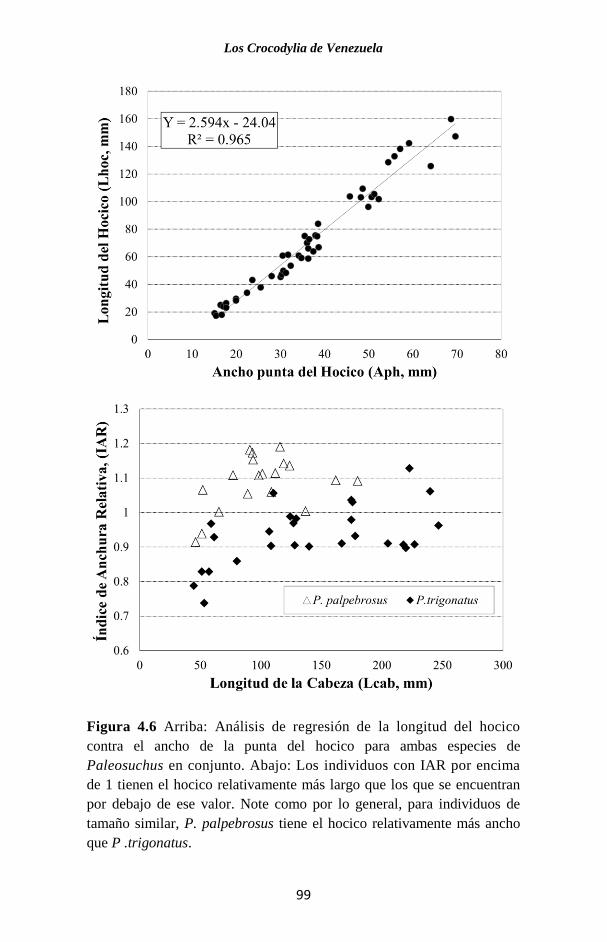
Los Crocodylia de Venezuela
99
Figura 4.6 Arriba: Análisis de regresión de la longitud del hocico
contra el ancho de la punta del hocico para ambas especies de
Paleosuchus en conjunto. Abajo: Los individuos con IAR por encima
de 1 tienen el hocico relativamente más largo que los que se encuentran
por debajo de ese valor. Note como por lo general, para individuos de
tamaño similar, P. palpebrosus tiene el hocico relativamente más ancho
que P .trigonatus.
Andrés Eloy Seijas
100
Figura 4.7 Cambios en el tamaño relativo de la longitud del cuerpo
(LCC) respecto a la longitud total (LT) en caimanes de la costa y del
Orinoco. En individuos con cola completa (CCS>=17) el cuerpo representa
alrededor de 53% de la longitud total. Note la poca frecuencia de C.
acutus con cola rota.
El conjunto de ecuaciones generadas a partir de los análisis de
regresión aparecen en los apéndices V y VI. Los coeficientes de
determinación R2
son en general superiores a 0,95; lo que indica un
buen ajuste a una línea recta entre los pares de variables morfo-
métricas que se analizan. Las regresiones con los datos trans-
formados a logaritmos indican también que existen relaciones
alométricas entre algunas dimensiones corporales (Tabla 4.3).
Algunos de los cambios en las proporciones de partes del
cuerpo se observan en la Figura 4.8. En primer lugar destaca que
el hocico de C. acutus es más ancho que el de C. intermedius,
aunque la muestra para esta última especie es más pequeña. Por
otra parte, se repite el patrón de las otras especies de Crocodylia
ya estudiadas, para las cuales el cambio en la anchura relativa del
hocico es especialmente marcado en los ejemplares más pequeños,
antes de alcanzar los 400 mm de LCC. A partir de esa medida el
crecimiento alométrico se atenúa.
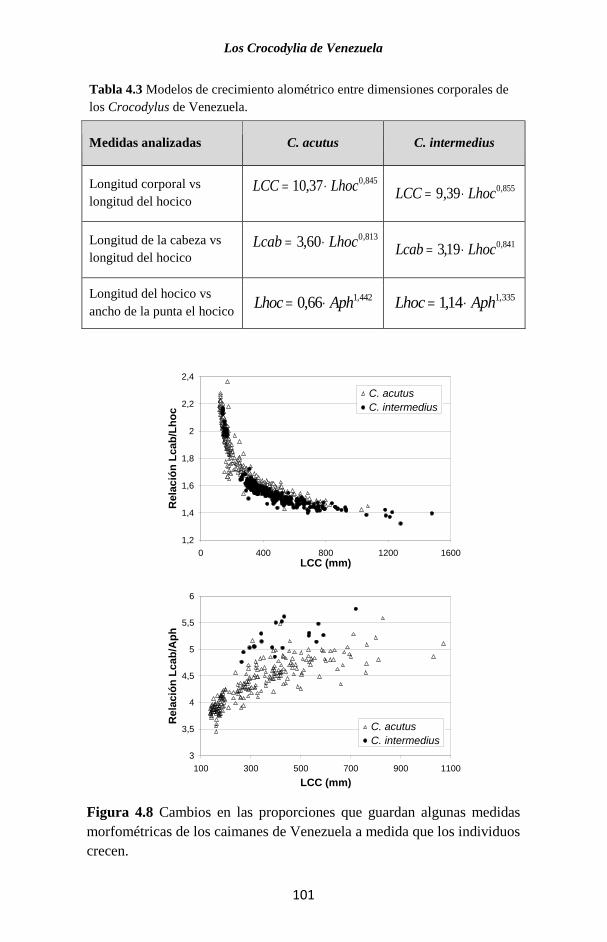
Los Crocodylia de Venezuela
101
Tabla 4.3 Modelos de crecimiento alométrico entre dimensiones corporales de
los Crocodylus de Venezuela.
Medidas analizadas C. acutus C. intermedius
Longitud corporal vs
longitud del hocico LCC Lhoc 10 37 0 845, ,
LCC Lhoc 9 39 0 855, ,
Longitud de la cabeza vs
longitud del hocico
Lcab Lhoc 3 60 0 813, ,
Lcab Lhoc 319 0 841, ,
Longitud del hocico vs
ancho de la punta el hocico Lhoc Aph 0 66 1 442, ,
Lhoc Aph 114 1 335, ,
Figura 4.8 Cambios en las proporciones que guardan algunas medidas
morfométricas de los caimanes de Venezuela a medida que los individuos
crecen.
1,2
1,4
1,6
1,8
2
2,2
2,4
0 400 800 1200 1600
LCC (mm)
Rela
ció
n L
ca
b/L
ho
c
C. acutus
C. intermedius
3
3,5
4
4,5
5
5,5
6
100 300 500 700 900 1100
LCC (mm)
Rela
ció
n L
ca
b/A
ph
C. acutus
C. intermedius

Andrés Eloy Seijas
102
Los cambios en la relación entre las dimensiones relativas del
cuerpo que se muestran en las figuras anteriores justifica la división
de la muestra en dos grupos: uno con los individuos menores a 400
mm de LCC y otro para aquellos mayores a esa talla, tal como se
presentan en los apéndices. Esto permite afinar el poder predictivo
de las ecuaciones.
Una ecuación de utilidad práctica es la que relaciona la longitud
de la planta (LP) de la pata trasera con la talla de los animales.
En algunas localidades donde los caimanes son muy escasos y
difíciles de observar, una manera de tener una primera aproxi-
mación de las tallas de los animales presentes es a través del
registro de sus huellas. Durante las fases iniciales del trabajo del
autor con el caimán de la costa entre 1980-1984, se midió la LP
de 78 animales. Los modelos de regresión para estimar la LT y la
LCC a partir de dichas medidas se muestran a continuación:
LT (mm) = -73,555 + 12,813 LP (mm) R2= 0,978
LCC (mm) = -49,936 + 6,978 LP (mm) R2= 0,977
Aunque los modelos anteriores fueron elaborados para C.
acutus, se pueden usar, como una aproximación para datos de C.
intermedius. En una evaluación sobre el estado poblacional del
caimán del Orinoco en el embalse de Tucupido (Portuguesa), Luis
Sánchez (com. pers.) sólo observó un ejemplar de esta especie,
pero también localizó tres huellas lo suficientemente alejadas entre
sí como para suponer que pertenecían a distintos individuos. Las
longitudes de esas huellas fueron 235, 258 y 264 mm. Estas huellas
pertenecerían a ejemplares de 2938, 3232 y 3307 mm de LT; es
decir, ejemplares adultos.
APLICABILIDAD DE LOS ANÁLISIS
MORFOMÉTRICOS
Predicciones a partir de los modelos
Como ejercicio para probar el ajuste y discutir las limitaciones
de algunos de los modelos que se mostraron en las secciones
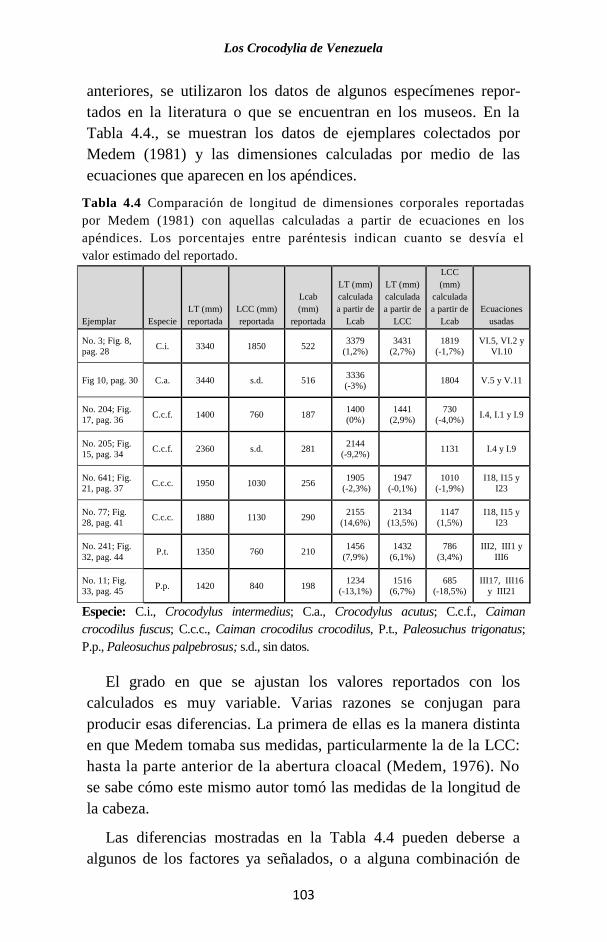
Los Crocodylia de Venezuela
103
anteriores, se utilizaron los datos de algunos especímenes repor-
tados en la literatura o que se encuentran en los museos. En la
Tabla 4.4., se muestran los datos de ejemplares colectados por
Medem (1981) y las dimensiones calculadas por medio de las
ecuaciones que aparecen en los apéndices.
Tabla 4.4 Comparación de longitud de dimensiones corporales reportadas
por Medem (1981) con aquellas calculadas a partir de ecuaciones en los
apéndices. Los porcentajes entre paréntesis indican cuanto se desvía el
valor estimado del reportado.
Ejemplar Especie
LT (mm)
reportada
LCC (mm)
reportada
Lcab
(mm)
reportada
LT (mm)
calculada
a partir de
Lcab
LT (mm)
calculada
a partir de
LCC
LCC
(mm)
calculada
a partir de
Lcab
Ecuaciones
usadas
No. 3; Fig. 8,
pag. 28 C.i. 3340 1850 522
3379
(1,2%)
3431
(2,7%)
1819
(-1,7%)
VI.5, VI.2 y
VI.10
Fig 10, pag. 30 C.a. 3440 s.d. 516 3336
(-3%) 1804 V.5 y V.11
No. 204; Fig.
17, pag. 36 C.c.f. 1400 760 187
1400
(0%)
1441
(2,9%)
730
(-4,0%) I.4, I.1 y I.9
No. 205; Fig.
15, pag. 34 C.c.f. 2360 s.d. 281
2144
(-9,2%) 1131 I.4 y I.9
No. 641; Fig.
21, pag. 37 C.c.c. 1950 1030 256
1905
(-2,3%)
1947
(-0,1%)
1010
(-1,9%)
I18, I15 y
I23
No. 77; Fig.
28, pag. 41 C.c.c. 1880 1130 290
2155
(14,6%)
2134
(13,5%)
1147
(1,5%)
I18, I15 y
I23
No. 241; Fig.
32, pag. 44 P.t. 1350 760 210
1456
(7,9%)
1432
(6,1%)
786
(3,4%)
III2, III1 y
III6
No. 11; Fig.
33, pag. 45 P.p. 1420 840 198
1234
(-13,1%)
1516
(6,7%)
685
(-18,5%)
III17, III16
y III21
Especie: C.i., Crocodylus intermedius; C.a., Crocodylus acutus; C.c.f., Caiman
crocodilus fuscus; C.c.c., Caiman crocodilus crocodilus, P.t., Paleosuchus trigonatus;
P.p., Paleosuchus palpebrosus; s.d., sin datos.
El grado en que se ajustan los valores reportados con los
calculados es muy variable. Varias razones se conjugan para
producir esas diferencias. La primera de ellas es la manera distinta
en que Medem tomaba sus medidas, particularmente la de la LCC:
hasta la parte anterior de la abertura cloacal (Medem, 1976). No
se sabe cómo este mismo autor tomó las medidas de la longitud de
la cabeza.
Las diferencias mostradas en la Tabla 4.4 pueden deberse a
algunos de los factores ya señalados, o a alguna combinación de
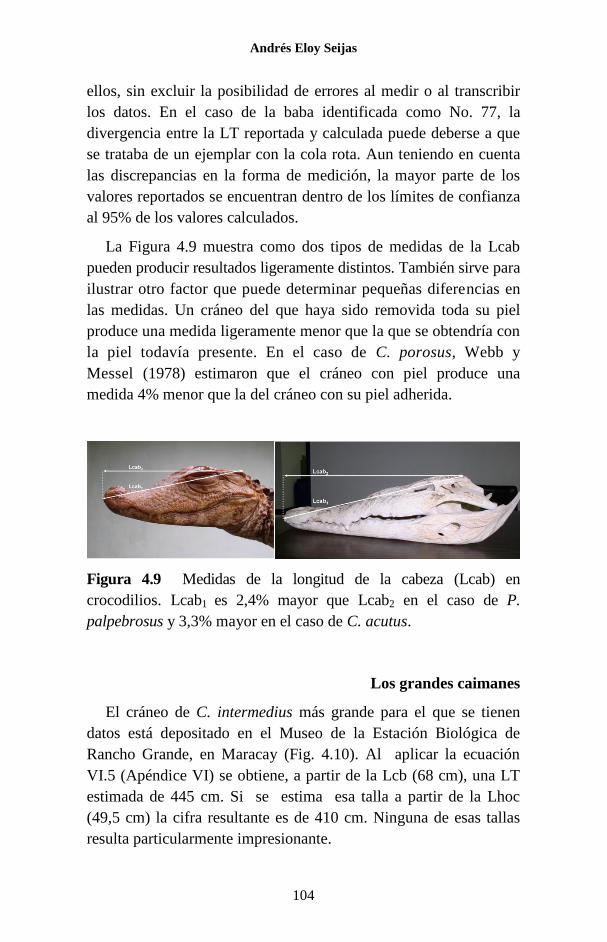
Andrés Eloy Seijas
104
ellos, sin excluir la posibilidad de errores al medir o al transcribir
los datos. En el caso de la baba identificada como No. 77, la
divergencia entre la LT reportada y calculada puede deberse a que
se trataba de un ejemplar con la cola rota. Aun teniendo en cuenta
las discrepancias en la forma de medición, la mayor parte de los
valores reportados se encuentran dentro de los límites de confianza
al 95% de los valores calculados.
La Figura 4.9 muestra como dos tipos de medidas de la Lcab
pueden producir resultados ligeramente distintos. También sirve para
ilustrar otro factor que puede determinar pequeñas diferencias en
las medidas. Un cráneo del que haya sido removida toda su piel
produce una medida ligeramente menor que la que se obtendría con
la piel todavía presente. En el caso de C. porosus, Webb y
Messel (1978) estimaron que el cráneo con piel produce una
medida 4% menor que la del cráneo con su piel adherida.
Figura 4.9 Medidas de la longitud de la cabeza (Lcab) en
crocodilios. Lcab1 es 2,4% mayor que Lcab2 en el caso de P.
palpebrosus y 3,3% mayor en el caso de C. acutus.
Los grandes caimanes
El cráneo de C. intermedius más grande para el que se tienen
datos está depositado en el Museo de la Estación Biológica de
Rancho Grande, en Maracay (Fig. 4.10). Al aplicar la ecuación
VI.5 (Apéndice VI) se obtiene, a partir de la Lcb (68 cm), una LT
estimada de 445 cm. Si se estima esa talla a partir de la Lhoc
(49,5 cm) la cifra resultante es de 410 cm. Ninguna de esas tallas
resulta particularmente impresionante.

Los Crocodylia de Venezuela
105
Figura 4.10 El autor sosteniendo el cráneo de Crocodylus intermedius
más grande depositado en la EBRG de Maracay, donado por el Walter
Arp. La Lcab de este cráneo es de 68 cm. El Lhoc es 49,5 cm. La longitud
total estimada del ejemplar en vida es 4,45 m.
El dato confiable sobre el caimán más grande que haya sido
medido alguna vez (22 pies 3 pulgadas) corresponde a Humboldt y
Bonpland. El transformar esa cifra al sistema métrico ha sido
objeto de controversia (Seijas, 2001). Si confiamos en Medem
(1983) la cifra exacta debería ser 6,75 m. ¿De qué tamaño debió
ser la cabeza de este caimán? Con los datos disponibles se obtiene
la siguiente ecuación de regresión:
(n=328; R2 = 0,984, E=12,634)
A partir de allí se obtiene una medida de la Lcab de 103,3 cm
(intervalo de confianza 95%, 100,8-105,8 cm).
Lcab LT 16 72 0 151, ,

Andrés Eloy Seijas
106
Los modelos que aquí se presentan tienen una finalidad funda-
mentalmente práctica, que permitan estimar las tallas y pesos de
ejemplares a partir de partes preservadas en museos o de infor-
mación publicada con datos incompletos.
Las predicciones que se puedan hacer a partir de las ecua-
ciones generadas tienen una serie de limitaciones y deben ser
usadas con precaución. En primer lugar, la base de datos disponible
tiene muy pocos ejemplares de gran tamaño. Esto es especialmente
cierto en el caso del caimán de la costa, especie para la cual el
ejemplar más grande con datos completos apenas alcanzó 2,12 m y
un peso de 27 kg. En el caso del caimán del Orinoco la muestra
contiene datos parciales de individuos de hasta 4,06 m y 369 kg,
pero esos animales provienen de zoocriaderos, lugares donde
generalmente los cocodrilos son relativamente más gordos que los
silvestres (ver Montague, 1984). Con esta última especie se usaron
también datos parciales obtenidos de cráneos depositados en
museos. Esto agrega otra fuente de error, ya que las medidas
tomadas en cráneos difieren ligeramente de las tomadas en
ejemplares vivos, debido a la pérdida de la piel (Hall y Portier,
1994; Mantague, 1984; Webb y Messel, 1978).
Otro inconveniente es que en este estudio no se discriminó a los
animales por sexo. Otros investigadores han demostrado
diferencias entre machos y hembras para distintas especies de
crocodilios (Ayarzagüena, 1983; Hall y Portier, 1994; Montague,
1984; Webb y Messel, 1978), aunque esas diferencias se
manifiestan princi-palmente en animales muy grandes.
Las limitaciones antes señaladas no le quitan el valor predictivo
a las ecuaciones presentadas, pero los modelos para estimar
medidas de longitud a partir de otras medidas lineales deben ser
usadas con ciertas precauciones para predecir las tallas de ani-
males muy grandes, ya que hay que tener en cuenta que los límites
de confianza para estimaciones de valores muy por encima de las
tallas de los animales con los cuales se trabajó para generar las
ecuaciones, pudieran producir subestimaciones. En el caso del peso,
por ejemplo, el análisis de los residuales de los modelos de

Los Crocodylia de Venezuela
107
regresión indica que el valor predictivo disminuye a medida que se
consideran individuos muy grandes. Estrictamente hablando, como
señaló Gould (1966) las regresiones alométricas sólo son aplicables
al intervalo de datos usados en su cómputo.
Este estudio es, por supuesto, preliminar. Es necesario recopilar
mucho más información que permitan subsanar sus lagunas. Se
espera que sirva de estímulo a otros investigadores para proseguir
con la tarea de conocer algo más sobre la biología de los croco-
dilios de Venezuela.

Andrés Eloy Seijas
108

Los Crocodylia de Venezuela
109
CAPÍTULO 5
ANÁLISIS DE POBLACIONES
Un aspecto básico que debe contemplar cualquier plan de
manejo de una especie animal es el referido a su abundancia y
estructura poblacional. Para conocer la situación de una especie, es
decir, si su número está declinando o aumentando, si se reproduce
o no, si su área de distribución aumenta o disminuye, lo primero
que hay que averiguar es su abundancia y distribución actual, para
compararla con datos sobre estos aspectos en el pasado. Es de los
resultados de esta primera evaluación en los que el manejador de
fauna se basa para programar investigaciones más detalladas (para
averiguar las razones que explican el estado poblacional encontrado,
por ejemplo) o para diseñar e implementar medidas que permitan
que el tamaño poblacional de la especie en un área determinada
aumente, disminuya o se estabilice, de acuerdo con objetivos
establecidos. Algunas consideraciones teóricas referidas a los
métodos de estimación poblacional con los Crocodylia se pueden
consultar en Bayliss (1987).
CONSIDERACIONES METODOLÓGICAS
Antes de emprender un estudio acerca del estado de una
población de algún crocodilio (su disposición espacial, abundancia,
estructura de tamaños, aspectos reproductivos y otros) es necesario
tener un conocimiento básico sobre su ecología y sobre las
características del área de estudio (extensión, vías de acceso,
características climáticas, navegabilidad, etc.). Este conocimiento
previo permitirá establecer cuál será el equipo de campo que se
utilizará y la mejor época del año para emprender el estudio.
Cuando se visita un sitio por primera vez es conveniente realizar un
reconocimiento general, durante el día, que permita al investigador

Andrés Eloy Seijas
110
familiarizarse con el entorno y las dificultades que se puedan
encontrar durante los recorridos nocturnos. Se debe tener muy
claros los objetivos que se persiguen y, por lo tanto, el tipo de
información que se quiere colectar y llevar planillas de campo para
anotar los datos obtenidos.
DETERMINACIÓN DE ABUNDANCIA
El método más común para determinar el tamaño en las
poblaciones de crocodilios es el basado en recorridos nocturnos a lo
largo de la orilla de cuerpos de agua, para detectar a los ejem-
plares gracias al reflejo que producen sus ojos al ser encandilados
con la luz de lámparas. Es bien conocido que diversos factores
afectan los resultados de este tipo de conteos (Woodward y
Marion, 1978; Hutton y Woolhouse, 1989). La época del año en
que se trabaja, la claridad de la noche por efecto de la fase lunar
al momento del conteo, las características del área de estudio, el
equipo de campo usado y otros aspectos pueden ocasionar dife-
rencias en los valores de abundancia y estructura poblacional
obtenidos (Fig. 5.1), situación que debe tenerse en cuenta a la hora
de sacar conclusiones, hacer comparaciones y tomar decisiones de
manejo. Otro factor muy importante es la pericia del investigador,
quien debe planificar su trabajo tratando de minimizar los efectos
distorsionadores antes señalados.
Por lo general no se conoce cuál es la fracción de la población
que se observa durante los conteos nocturnos y seguramente ésta
varía de acuerdo con los factores mencionados. El factor que
afecta de manera más pronunciada las estimaciones poblacionales
de crocodilios es el nivel del agua, el cual está asociado a la
estacionalidad climática. En el caso de la baba, por ejemplo, los
conteos en préstamos y lagunas a lo largo de caminos arrojan
resultados contrastantes dependiendo de la época del año en que se
realizan (Fig. 5.2). El mejor momento para contar es a finales de la
época seca, cuando la mayoría de las babas se concentran en los
pocos cuerpos de agua disponibles.
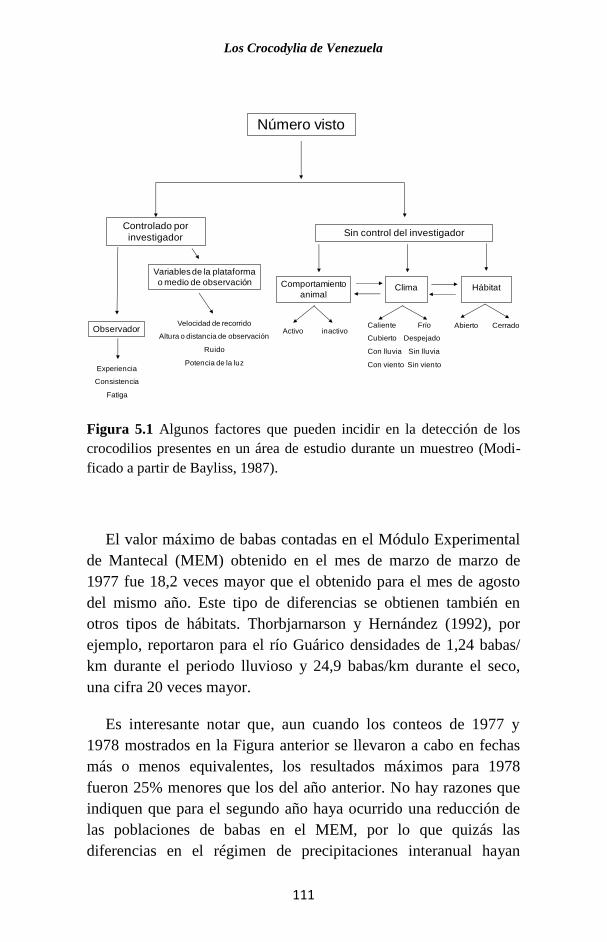
Los Crocodylia de Venezuela
111
Figura 5.1 Algunos factores que pueden incidir en la detección de los
crocodilios presentes en un área de estudio durante un muestreo (Modi-
ficado a partir de Bayliss, 1987).
El valor máximo de babas contadas en el Módulo Experimental
de Mantecal (MEM) obtenido en el mes de marzo de marzo de
1977 fue 18,2 veces mayor que el obtenido para el mes de agosto
del mismo año. Este tipo de diferencias se obtienen también en
otros tipos de hábitats. Thorbjarnarson y Hernández (1992), por
ejemplo, reportaron para el río Guárico densidades de 1,24 babas/
km durante el periodo lluvioso y 24,9 babas/km durante el seco,
una cifra 20 veces mayor.
Es interesante notar que, aun cuando los conteos de 1977 y
1978 mostrados en la Figura anterior se llevaron a cabo en fechas
más o menos equivalentes, los resultados máximos para 1978
fueron 25% menores que los del año anterior. No hay razones que
indiquen que para el segundo año haya ocurrido una reducción de
las poblaciones de babas en el MEM, por lo que quizás las
diferencias en el régimen de precipitaciones interanual hayan
Número visto
Controlado por
investigador Sin control del investigador
Observador
Experiencia
Consistencia
Fatiga
Variables de la plataforma
o medio de observación
Velocidad de recorrido
Altura o distancia de observación
Ruido
Potencia de la luz
Comportamiento
animalClima Hábitat
Activo inactivoCaliente Frío
Cubierto Despejado
Con lluvia Sin lluvia
Con viento Sin viento
Abierto Cerrado
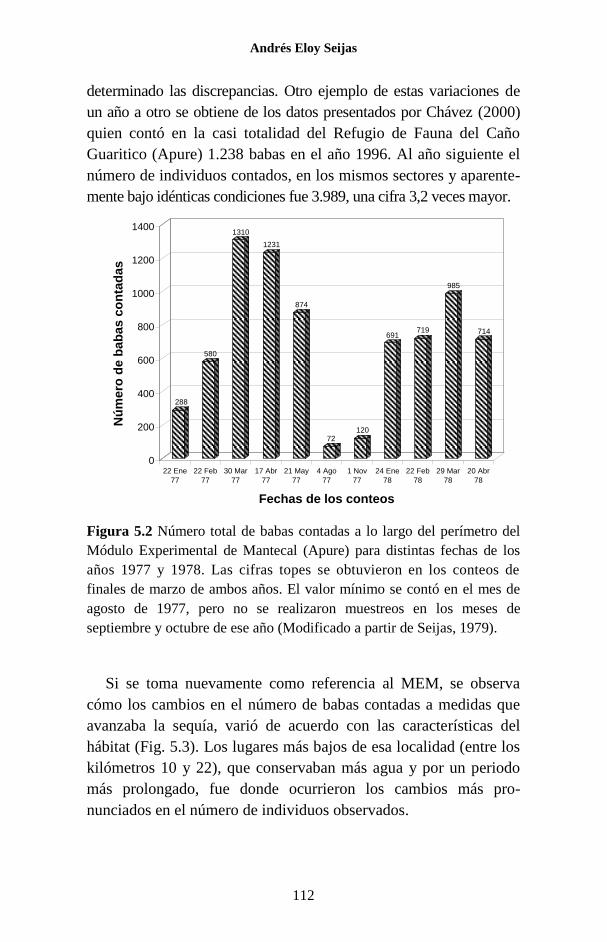
Andrés Eloy Seijas
112
determinado las discrepancias. Otro ejemplo de estas variaciones de
un año a otro se obtiene de los datos presentados por Chávez (2000)
quien contó en la casi totalidad del Refugio de Fauna del Caño
Guaritico (Apure) 1.238 babas en el año 1996. Al año siguiente el
número de individuos contados, en los mismos sectores y aparente-
mente bajo idénticas condiciones fue 3.989, una cifra 3,2 veces mayor.
Figura 5.2 Número total de babas contadas a lo largo del perímetro del
Módulo Experimental de Mantecal (Apure) para distintas fechas de los
años 1977 y 1978. Las cifras topes se obtuvieron en los conteos de
finales de marzo de ambos años. El valor mínimo se contó en el mes de
agosto de 1977, pero no se realizaron muestreos en los meses de
septiembre y octubre de ese año (Modificado a partir de Seijas, 1979).
Si se toma nuevamente como referencia al MEM, se observa
cómo los cambios en el número de babas contadas a medidas que
avanzaba la sequía, varió de acuerdo con las características del
hábitat (Fig. 5.3). Los lugares más bajos de esa localidad (entre los
kilómetros 10 y 22), que conservaban más agua y por un periodo
más prolongado, fue donde ocurrieron los cambios más pro-
nunciados en el número de individuos observados.
288
580
1310
1231
874
72120
691719
985
714
0
200
400
600
800
1000
1200
1400
Nú
me
ro d
e b
ab
as
co
nta
da
s
22 Ene
77
22 Feb
77
30 Mar
77
17 Abr
77
21 May
77
4 Ago
77
1 Nov
77
24 Ene
78
22 Feb
78
29 Mar
78
20 Abr
78
Fechas de los conteos
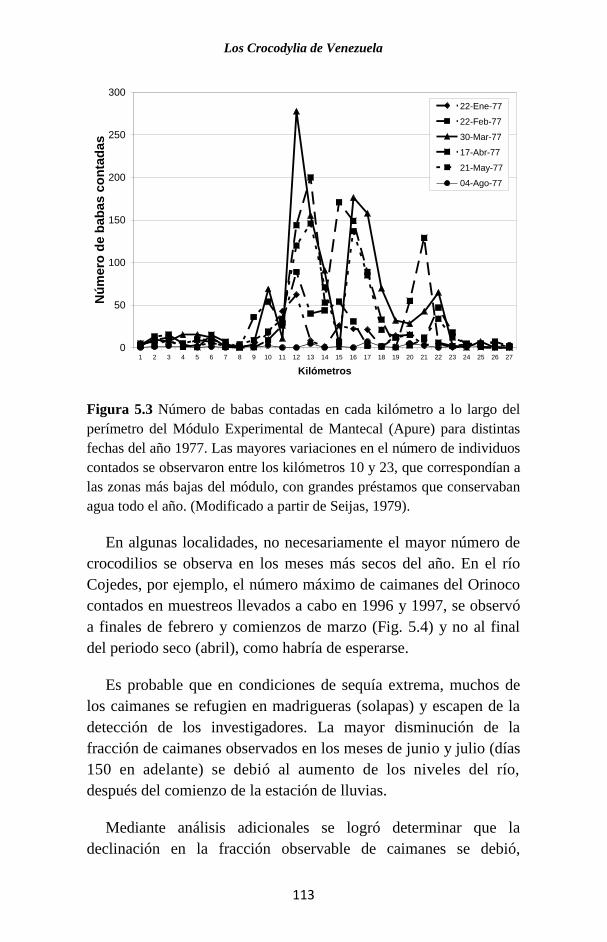
Los Crocodylia de Venezuela
113
Figura 5.3 Número de babas contadas en cada kilómetro a lo largo del
perímetro del Módulo Experimental de Mantecal (Apure) para distintas
fechas del año 1977. Las mayores variaciones en el número de individuos
contados se observaron entre los kilómetros 10 y 23, que correspondían a
las zonas más bajas del módulo, con grandes préstamos que conservaban
agua todo el año. (Modificado a partir de Seijas, 1979).
En algunas localidades, no necesariamente el mayor número de
crocodilios se observa en los meses más secos del año. En el río
Cojedes, por ejemplo, el número máximo de caimanes del Orinoco
contados en muestreos llevados a cabo en 1996 y 1997, se observó
a finales de febrero y comienzos de marzo (Fig. 5.4) y no al final
del periodo seco (abril), como habría de esperarse.
Es probable que en condiciones de sequía extrema, muchos de
los caimanes se refugien en madrigueras (solapas) y escapen de la
detección de los investigadores. La mayor disminución de la
fracción de caimanes observados en los meses de junio y julio (días
150 en adelante) se debió al aumento de los niveles del río,
después del comienzo de la estación de lluvias.
Mediante análisis adicionales se logró determinar que la
declinación en la fracción observable de caimanes se debió,
0
50
100
150
200
250
300
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Kilómetros
Nú
me
ro d
e b
ab
as
co
nta
da
s
22-Ene-77
22-Feb-77
30-Mar-77
17-Abr-77
21-May-77
04-Ago-77
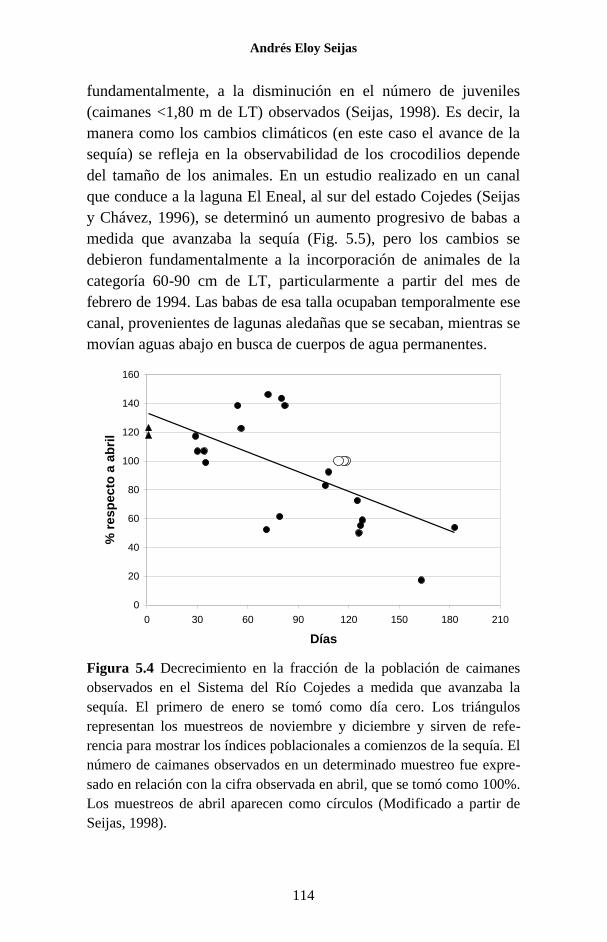
Andrés Eloy Seijas
114
fundamentalmente, a la disminución en el número de juveniles
(caimanes <1,80 m de LT) observados (Seijas, 1998). Es decir, la
manera como los cambios climáticos (en este caso el avance de la
sequía) se refleja en la observabilidad de los crocodilios depende
del tamaño de los animales. En un estudio realizado en un canal
que conduce a la laguna El Eneal, al sur del estado Cojedes (Seijas
y Chávez, 1996), se determinó un aumento progresivo de babas a
medida que avanzaba la sequía (Fig. 5.5), pero los cambios se
debieron fundamentalmente a la incorporación de animales de la
categoría 60-90 cm de LT, particularmente a partir del mes de
febrero de 1994. Las babas de esa talla ocupaban temporalmente ese
canal, provenientes de lagunas aledañas que se secaban, mientras se
movían aguas abajo en busca de cuerpos de agua permanentes.
Figura 5.4 Decrecimiento en la fracción de la población de caimanes
observados en el Sistema del Río Cojedes a medida que avanzaba la
sequía. El primero de enero se tomó como día cero. Los triángulos
representan los muestreos de noviembre y diciembre y sirven de refe-
rencia para mostrar los índices poblacionales a comienzos de la sequía. El
número de caimanes observados en un determinado muestreo fue expre-
sado en relación con la cifra observada en abril, que se tomó como 100%.
Los muestreos de abril aparecen como círculos (Modificado a partir de
Seijas, 1998).
0
20
40
60
80
100
120
140
160
0 30 60 90 120 150 180 210
Días
% r
esp
ecto
a a
bri
l
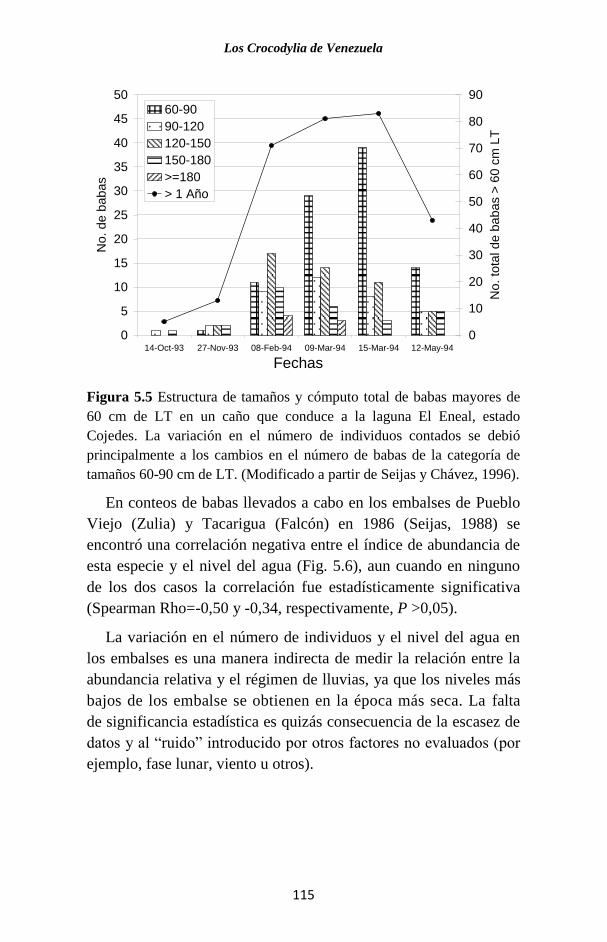
Los Crocodylia de Venezuela
115
Figura 5.5 Estructura de tamaños y cómputo total de babas mayores de
60 cm de LT en un caño que conduce a la laguna El Eneal, estado
Cojedes. La variación en el número de individuos contados se debió
principalmente a los cambios en el número de babas de la categoría de
tamaños 60-90 cm de LT. (Modificado a partir de Seijas y Chávez, 1996).
En conteos de babas llevados a cabo en los embalses de Pueblo
Viejo (Zulia) y Tacarigua (Falcón) en 1986 (Seijas, 1988) se
encontró una correlación negativa entre el índice de abundancia de
esta especie y el nivel del agua (Fig. 5.6), aun cuando en ninguno
de los dos casos la correlación fue estadísticamente significativa
(Spearman Rho=-0,50 y -0,34, respectivamente, P >0,05).
La variación en el número de individuos y el nivel del agua en
los embalses es una manera indirecta de medir la relación entre la
abundancia relativa y el régimen de lluvias, ya que los niveles más
bajos de los embalse se obtienen en la época más seca. La falta
de significancia estadística es quizás consecuencia de la escasez de
datos y al “ruido” introducido por otros factores no evaluados (por
ejemplo, fase lunar, viento u otros).
0
5
10
15
20
25
30
35
40
45
50
14-Oct-93 27-Nov-93 08-Feb-94 09-Mar-94 15-Mar-94 12-May-94
Fechas
No
. de b
abas
0
10
20
30
40
50
60
70
80
90
No
. to
tal de b
abas >
60 c
m L
T
60-90
90-120
120-150
150-180
>=180
> 1 Año
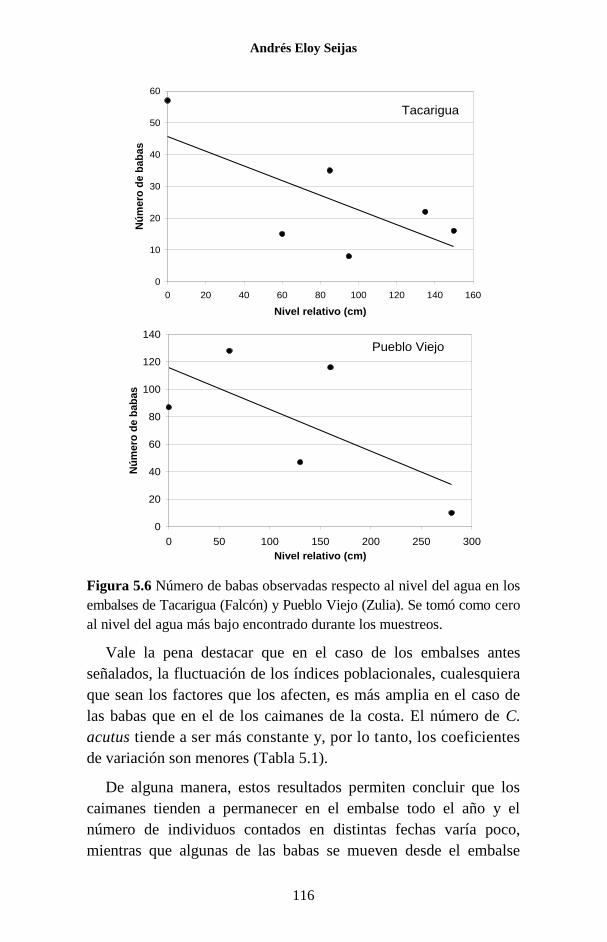
Andrés Eloy Seijas
116
Figura 5.6 Número de babas observadas respecto al nivel del agua en los
embalses de Tacarigua (Falcón) y Pueblo Viejo (Zulia). Se tomó como cero
al nivel del agua más bajo encontrado durante los muestreos.
Vale la pena destacar que en el caso de los embalses antes
señalados, la fluctuación de los índices poblacionales, cualesquiera
que sean los factores que los afecten, es más amplia en el caso de
las babas que en el de los caimanes de la costa. El número de C.
acutus tiende a ser más constante y, por lo tanto, los coeficientes
de variación son menores (Tabla 5.1).
De alguna manera, estos resultados permiten concluir que los
caimanes tienden a permanecer en el embalse todo el año y el
número de individuos contados en distintas fechas varía poco,
mientras que algunas de las babas se mueven desde el embalse
Tacarigua
0
10
20
30
40
50
60
0 20 40 60 80 100 120 140 160
Nivel relativo (cm)
Nú
mero
de b
ab
as
Pueblo Viejo
0
20
40
60
80
100
120
140
0 50 100 150 200 250 300
Nivel relativo (cm)
Nú
mero
de b
ab
as
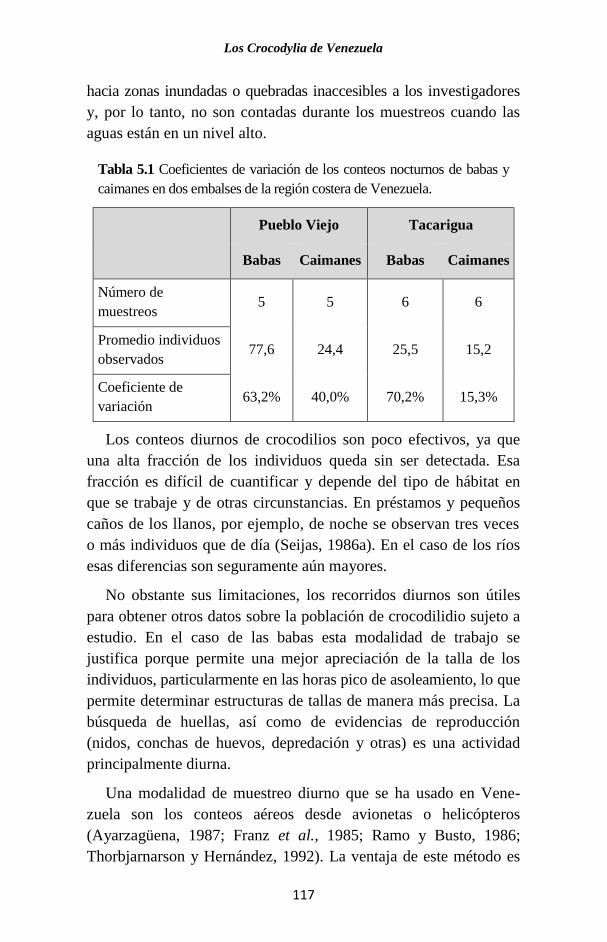
Los Crocodylia de Venezuela
117
hacia zonas inundadas o quebradas inaccesibles a los investigadores
y, por lo tanto, no son contadas durante los muestreos cuando las
aguas están en un nivel alto.
Tabla 5.1 Coeficientes de variación de los conteos nocturnos de babas y
caimanes en dos embalses de la región costera de Venezuela.
Pueblo Viejo Tacarigua
Babas Caimanes Babas Caimanes
Número de
muestreos 5 5 6 6
Promedio individuos
observados 77,6 24,4 25,5 15,2
Coeficiente de
variación 63,2% 40,0% 70,2% 15,3%
Los conteos diurnos de crocodilios son poco efectivos, ya que
una alta fracción de los individuos queda sin ser detectada. Esa
fracción es difícil de cuantificar y depende del tipo de hábitat en
que se trabaje y de otras circunstancias. En préstamos y pequeños
caños de los llanos, por ejemplo, de noche se observan tres veces
o más individuos que de día (Seijas, 1986a). En el caso de los ríos
esas diferencias son seguramente aún mayores.
No obstante sus limitaciones, los recorridos diurnos son útiles
para obtener otros datos sobre la población de crocodilidio sujeto a
estudio. En el caso de las babas esta modalidad de trabajo se
justifica porque permite una mejor apreciación de la talla de los
individuos, particularmente en las horas pico de asoleamiento, lo que
permite determinar estructuras de tallas de manera más precisa. La
búsqueda de huellas, así como de evidencias de reproducción
(nidos, conchas de huevos, depredación y otras) es una actividad
principalmente diurna.
Una modalidad de muestreo diurno que se ha usado en Vene-
zuela son los conteos aéreos desde avionetas o helicópteros
(Ayarzagüena, 1987; Franz et al., 1985; Ramo y Busto, 1986;
Thorbjarnarson y Hernández, 1992). La ventaja de este método es

Andrés Eloy Seijas
118
que permite recorrer grandes distancias en poco tiempo, pero es
extremadamente selectivo y sesgado hacia los grandes animales,
más fáciles de observar. Es por ello que se ha empleado exclu-
sivamente para detectar caimanes del Orinoco. Las horas más
adecuadas para este tipo de evaluación son las de la media mañana
(9-11 a.m.) cuando gran parte de los individuos se asolean.
Una razón para efectuar muestreo aéreos es la de ubicar
lugares donde posteriormente puedan realizarse trabajos más inten-
sivos, que incluyan recorridos nocturnos. La gran desventaja del
método es su costo, principalmente si se quisiera utilizar como
herramienta rutinaria. Por otra parte muchos ríos no presentan
características adecuadas para este tipo de muestreos, ya sea
porque poseen muchos meandros o porque están cubiertos por
bosques muy cerrados que impiden la detección de los animales.
UNIDADES DE MEDIDA DE LA DENSIDAD O
ABUNDANCIA
La manera como se expresa la densidad de crocodílidos
depende del tipo de ambiente en que se trabaja. En préstamos,
pequeñas lagunas y humedales es común expresarla como densidad
ecológica, es decir individuos por superficie de agua (o de humedal)
ocupada por la especie (ver, por ejemplo, Marcellini, 1979). La
unidad de densidad más conveniente en estos casos es la de ind/
m2, cuando se trata de áreas muy pequeñas o ind/ha, cuando se
agregan muchos ambientes como los señalados.
En el caso de la baba, en los llanos de Venezuela se ha
reportado con frecuencia la densidad bruta, es decir el número de
individuos por superficie total de alguna zona (un hato, por ejemplo)
o una región, la cual incluye áreas sin agua, o sólo temporalmente
inundadas. Ésto se ha justificado por razones de manejo y por el
hecho de que durante la época de lluvias las babas se dispersan en
la sabana inundada y ocupan gran parte de los predios o regiones
estudiadas (Ayarzagüena, 1983; Seijas, 1986a; Ramo y Busto, 1989;
Velasco y Ayarzagüena, 1995). La unidad más usada en estos
casos es ind/ha.

Los Crocodylia de Venezuela
119
En lagunas o lagos (por ejemplo embalses) una medida más
adecuada para expresar la densidad es la de número de individuos
por longitud del perímetro del cuerpo de agua. Esta medida se
justifica por el hecho de que en realidad los crocodilios habitan
principalmente los litorales y aquellas zonas donde las aguas no son
muy profundas (Seijas, 1988). Es raro, por tanto, localizar en una
laguna grande o en un embalse individuos muy alejados de la orilla.
La unidad más usada en estos casos es ind/km.
Cuando se trabaja en ríos, la unidad de medición de la abun-
dancia más usada es la de ind/km de recorrido. En este libro se usa
esa medida como una densidad, aunque quizás a los puristas de las
terminologías no les guste esa denominación. En muchos casos,
cuando se trata de ríos angostos, las cifras reportadas se refieren a
individuos encontrados en las dos márgenes del río, ya que las
lámparas que se usan para detectar a los animales tienen suficiente
potencia como para localizar los crocodilios en ambas orillas a
medida que se navega por él. En ríos muy anchos es reco-
mendable, o incluso inevitable, muestrear cada orilla de manera
independiente. Es necesario señalar en los resultados cuál de los
dos procedimientos se usó en el trabajo. En cualquier caso, la
unidad usada de ind/km.
Hay que ser cuidadoso a la hora de interpretar los valores de
densidad, cualquiera que sea la unidad usada, cuando se trata de
conteos realizados en cuerpos de agua de superficie o longitud muy
reducidos. Para ilustrar lo que se quiere decir, consideren una
situación en la que un crocodiliano cualquiera ocupe un pequeño
pozo de 100 m2. Esa densidad equivale a 100 individuos por
hectárea, una cifra muy grande, si se toma fuera de contexto. En el
río Sarare (Falcón) se reportó una densidad de 196,6 babas/km para
un pequeño segmento aislado de sólo 300 m. Esa densidad es 7,5
veces mayor que la del río Unare (Anzoátegui) para el que se
muestreó una longitud de 25 km (Seijas, 1986b). Sería un
exabrupto, sin embargo, concluir que hay más babas en el río
Sarare que en el río Unare.
De manera generalizada, en los estudios poblacionales de
crocodilios, cuando se reportan cifras de abundancia y densidad se

Andrés Eloy Seijas
120
excluyen de los análisis los individuos menores de un año, dado que
la tasa de mortalidad que ellos presentan en las primeras semanas
de vida es muy alta y, por tanto, su presencia en la población es
generalmente efímera.
DETERMINACIÓN DE LA ESTRUCTURA
POBLACIONAL
El cuerpo de los crocodilios crece de manera permanente
(durante toda la vida), pero su tasa de crecimiento, como se
discutirá en más detalle posteriormente, es muy variable y quizás
dependa principalmente de la calidad del hábitat que los individuos
ocupan (Seijas et al., 2008). Los ejemplares que habitan ambientes
muy productivos crecen rápido y alcanzan la madurez sexual en
menos tiempo que aquellos que viven en ambiente más pobres. En
apoyo de esta conclusión tenemos que Llobet (2002) recapturó en
el río Capanaparo en el año 2001 a tres C. intermedius que habían
sido liberados en esa localidad al año de edad en 1992 y 1993. La
talla (LT) de estos ejemplares fue 236 cm para el de nueve años y
204-223 para los de ocho. En contraste, Lugo (1998) reportó los
datos de tres caimanes del Orinoco liberados en el caño Matiyure
del estado Apure a los dos años de edad y recapturados en el
mismo lugar a la edad de cinco años y con tallas que iban desde
275 a 296 cm de LT. Igualmente, Chávez (2000) aportó datos de
dos C. intermedius recapturados en el Caño Guaritico (Apure) que
en poco menos de cuatro años superaban 240 cm de LT, es decir,
más grandes que los individuos de mayor edad recapturados en el
Capanaparo. Como conclusión se puede decir que el crecimiento de
los caimanes es relativamente lento en un río de aguas claras como
el Capanaparo y muy rápido en ríos de aguas blancas, más ricos
en nutrimentos, como el Matiyure y el Guaritico. También dife-
rencias genéticas entre individuos, interacciones intra e inter-
específicas y otros factores deben jugar un papel en la deter-
minación de la velocidad de crecimiento de cada animal. Todo esto
advierte sobre la imposibilidad de establecer la edad de un individuo
tomando como único criterio su tamaño.
La imposibilidad de asignarles una edad absolutamente confiable
a los animales observados durante los muestreos, ha llevado a la

Los Crocodylia de Venezuela
121
necesidad de determinar la estructura de las poblaciones que se
estudian con base en categorías de tallas o tamaños. En la mayor
parte de los casos es prácticamente imposible capturar a todos los
individuos de la población para medir sus tallas con exactitud.
Efectuar muestreos no sesgados que permitan inferir la estructura
de la población a partir de un conjunto de animales capturados es
también muy difícil de llevar a cabo, principalmente porque la
capturabilidad depende del tamaño y los individuos más grandes son
generalmente muy tímidos o requieren un esfuerzo de captura muy
grande en términos de tiempo u otros recursos, además que ello
representa en ocasiones una actividad riesgosa, tanto para los
animales como para las personas que tratan de capturarlos. La
manera más práctica de asignar a un individuo a una clase de
tamaño es, por lo tanto, estimando su talla cuando se observa a
distancia. El método requiere de pericia y no está exento de
errores (Magnusson, 1983). No es de extrañar que muchos
estudios se limiten a contar el número de individuos y no reportan
la estructura de tamaños de la población. Para corregir algunos de
los problemas mencionados se han propuesto técnicas fotográficas
para estimar las tallas de los individuos durante dichos conteos (Gorzula,
1984) pero el procedimiento es impracticable en la mayoría de las
ocasiones.
En un intento para cuantificar el grado de exactitud con que se
estima la talla de los animales observados, se llevó a cabo un
pequeño ensayo con babas en cuerpos de agua al sur del estado
Cojedes (Seijas y Chávez, 1996). Se capturaron 45 individuos, a los
cuales se les tomó un conjunto completo de datos y se elaboró un
modelo de regresión que permitiera predecir la longitud total (LT) a
partir de la longitud de la cabeza (Lcab). Para este modelo sólo se
tomaron en consideración animales con la cola completa (CCS>18)
y con una talla superior a 340 mm de LT. Posteriormente se
capturaron, desde un bote, 74 babas, a cada una de las cuales se
le estimaba en voz alta, justo antes de la captura (para que el
investigador acompañante tomara nota), la categoría de tamaño a la
que se creía que pertenecía. A estos individuos se les midió sólo la
Lcab y con el modelo de regresión señalado se les calculó la LT y
se determinó si ésta se encontraba entre los límites de la categoría

Andrés Eloy Seijas
122
de tamaños a que habían sido asignados en el instante previo a la
captura (Tabla 5.2). Los investigadores se turnaron en la tarea de
capturar y estimar la talla de las babas. Considerando solamente
las observaciones del investigador que usualmente realizó las
estimaciones poblacionales, en 18,8% de los casos la talla calculada
estuvo fuera de los límites de la categoría de tamaños estimada
antes de la captura. El porcentaje de desaciertos fue mayor en el
caso del segundo observador (38,1%). En un caso la talla de la
baba capturada estuvo 76,7 cm por encima del límite superior de la
categoría a la cual fue asignada, en los fallos restantes, sin
embargo, las tallas reales estuvieron en promedio 5,3 cm por
encima o por debajo del límite de la categoría a la cual habían sido
adjudicadas en el instante de la captura. En este ejercicio se
usaron categorías de tamaño con una amplitud de 30 cm. De
haberse usado categorías más estrechas, con seguridad el por-
centaje de fallos se hubiera incrementado.
Es necesario recordar que, en el ensayo descrito anteriormente,
las babas a las que se les estimó la talla fueron observadas desde
una lancha, de noche y a muy corta distancia, en general, menor a
los dos metros. En muchos estudios la talla de los crocodilios se
estima a la luz del día y a mucha mayor distancia que en las del
ensayo referido, a veces incluso con la ayuda de binoculares. Los
errores de apreciación en esas ocasiones pueden ser considerables.
Muchos de los conteos nocturnos de babas reportados en la
literatura se realizaron desde vehículos terrestres, generalmente a
distancias muy largas como para lograr una estimación precisa del
tamaño, por lo que habitualmente sólo se discrimina entre animales
menores de un año (que se distinguen porque tienden a permanecer
en grupos muy compactos, durante varias semanas después del
nacimiento) e individuos mayores de un año. Recorridos nocturnos
a pie a lo largo de caños o perímetros de préstamos y lagunas
permiten una aproximación suficiente como para obtener una buena
estimación de las tallas, pero este procedimiento es poco práctico
cuando el área de muestreo es muy extensa o las orillas de los
cuerpos de agua son difíciles de transitar.
Los investigadores en Venezuela han usado categorías de
tamaños diferentes a la hora de reportar sus resultados, lo cual ha
Los Crocodylia de Venezuela
123
dificultado un poco las comparaciones. Uno de los sistemas es el
establecido por Ayarzagüena (1983) de cuatro clases de tamaño, el
cual ha sido adoptado, con algunas modificaciones, por el Ministerio
del Ambiente en el programa de aprovechamiento de babas, como
discutiremos más adelante. En la literatura científica en inglés es
usual agrupar a los animales observados en categorías de longitud
total en pies, o su equivalente aproximado a 30 cm (Watson et al.,
1971; Chabreck, 1976; Messel et al., 1981; Read et al., 2004). Ese
es el tipo de clasificación que se ha venido usando en el país en
los últimos años, principalmente para el estudio de poblaciones de
caimanes del Orinoco y de la costa (Seijas y Chávez, 1991, 2000;
Llobet, 2002; Jiménez-Oráa, 2002; Urdaneta, 2002; Lander, 2003;
Barros et al., 2005).
Tabla 5.2 Aciertos y fallos de dos observadores en el intento de asignar
la categoría de tamaño de babas observadas a corta distancia desde un
bote. (A partir de datos en Seijas y Chávez, 1996).
Categoría Observador 1 Observador 2
aciertos fallos aciertos fallos
30-60 1 0 0 3
60-90 9 2 11 8
90-120 10 0 8 1
120-150 3 4 5 3
150-180 3 0 2 1
Todas 26 6 26 16
En localidades donde babas y caimanes coexisten es importante,
porque facilita el registro de datos, poseer un sistema de clasi-
ficación único que pueda emplearse tanto para babas como para
caimanes. La planilla mostrada en la Figura 5.7 es la que se ha
usado para recoger la información de campo en los ríos Cojedes,
Capanaparo y Manapire.
Aun cuando en la planilla se le asigna a cada animal observado
una de las categorías de tamaño preestablecidas, para los análisis
generalmente se agrupan éstas en clases más amplias, así:

Andrés Eloy Seijas
124
Clase I: Individuos menores de 60 cm de LT
Clase II: Individuos de 60 a 119,9 cm de LT
Clase III: Individuos de 120 a 179,9 cm de LT
Clase IV: Individuos de 180 a 239,9 cm de LT
Clase V: Individuos de 240 o más cm de LT
En el caso de las babas, la Clase I corresponde a individuos
menores a dos años. La Clase II incluye individuos juveniles, la III
contiene adultos (machos y hembras) y la IV está constituida casi
exclusivamente por machos. Esta clasificación está “inspirada” pero
no es exactamente equivalente a la establecida por Ayarzagüena
(1983) para las babas de los llanos, como veremos más adelante;
sólo que no hay babas de Clase V, ya que en esta especie son
muy raros los individuos que superan los 240 cm de LT. En el
caso de los caimanes la Clase I está conformada por individuos
menores de un año; la II y III por individuos juveniles; la IV
corresponde a subadultos y la V a adultos.
FACTORES DE CORRECCIÓN Y AJUSTES
Ya se señaló que en los conteos de crocodilios una fracción de
los animales presentes no es detectada. Staton y Dixon (1975)
estimaron que en los préstamos y lagunas en que trabajaron, en el
hato Masaguaral (Guárico), esa fracción alcanzaba 10% para los
conteos nocturnos. Esa cifra luce optimista ya que Thorbjarnarson
(1990), quien trabajó también en ese hato, reportó un valor
promedio de 19,5%.
En estudios realizados con cocodrilos del Nilo marcados en un
lago en Zimbabwe, se obtuvo que incluso en condiciones óptimas,
más de 37% de los individuos de la población permaneció sin ser
detectada, y la proporción de la verdadera población que es
observada durante los recorridos nocturnos varió entre 10 y 63%
(Hutton y Woolhouse, 1989). Bayliss (1987) discute los resultados
de metodologías basadas en el marcado y recaptura de Crocodylus
Los Crocodylia de Venezuela
125
Figura 5.7 Planilla para el registro de los datos de campo en
conteos nocturnos de crocodilios. Esta planilla se usa en recorridos
a lo largo de ríos y perímetros de embalses. Los datos anotados en
este caso no son reales y sólo pretenden ilustrar el tipo de
información que usualmente se vierte en ella.

Andrés Eloy Seijas
126
porosus, y concluye que la detectabilidad depende del tipo de
hábitat donde se trabaja y de la talla de los individuos. Combinando
marcaje con pintura, reconocimientos aéreos y recorridos nocturnos,
otros autores han calculado valores de detectabilidad aún más bajos
(Mourao et al., 1994; Woodward et al., 1996).
King et al. (1990) señalaron varias fórmulas para estimar la
fracción de individuos observables durante conteos nocturnos. Su
empleo requiere de muchas repeticiones, lo que limita su apli-
cabilidad en muchos programas de monitoreo, en los que se llevan
a cabo muy pocos conteos en la misma localidad o sector. A
continuación se presenta una fórmula que es una modificación del
modelo 3 de los mencionados autores:
sX
Xp
m
m
*3
Donde p representa la fracción observable de la población, Xm,
es el número promedio de individuos observados por conteo, s, es
la desviación estándar de la distribución de los conteos realizados.
Esta fórmula se fundamenta en el hecho de que para la
distribución normal, 99,73% de los valores quedan dentro del
intervalo de la media y más o menos tres veces su desviación
estándar. El denominador de la fórmula representa el extremo
derecho de ese intervalo, es decir, el valor máximo de animales
que se esperaría encontrar si se realizaran infinitos conteos.
Como ejercicio, se muestra el uso de la fórmula antes propuesta
con los datos aportados por Ávila-Manjón (2008) para el Río
Cojedes (Tabla 5.3).
Los datos en la Tabla 5.3 permiten calcular que sólo se
observaron alrededor de 62-63% de los caimanes presentes en el
segmento muestreado ¿Qué tanto se ajusta este valor a la realidad?
Los datos de Ávila-Manjón (2008) permiten otra manera de
calcular la fracción observable. Esta autora capturó durante el
desarrollo de su estudio 44 individuos <180 cm de LT en el sector
Confluencia-Caño Amarillo. El número máximo observado durante
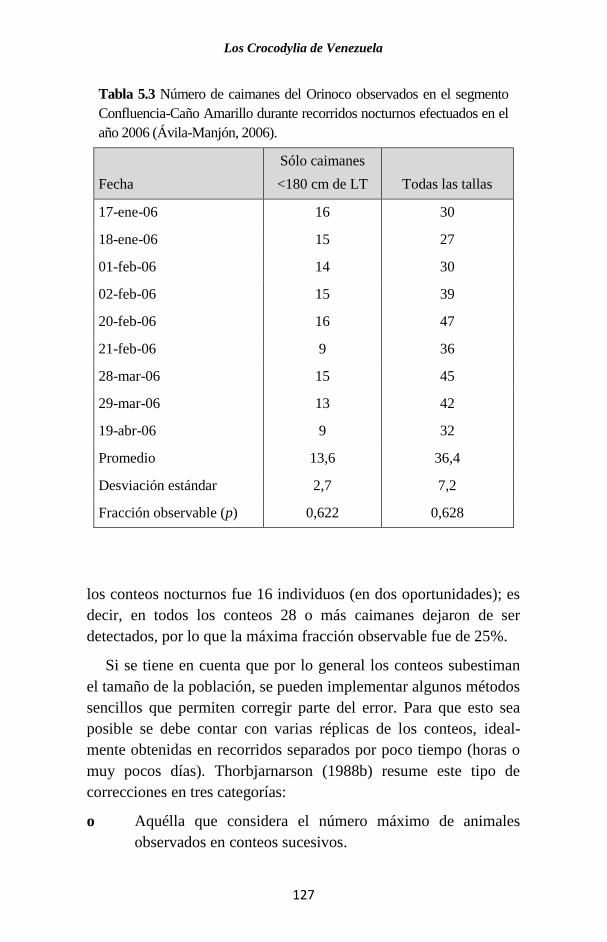
Los Crocodylia de Venezuela
127
los conteos nocturnos fue 16 individuos (en dos oportunidades); es
decir, en todos los conteos 28 o más caimanes dejaron de ser
detectados, por lo que la máxima fracción observable fue de 25%.
Si se tiene en cuenta que por lo general los conteos subestiman
el tamaño de la población, se pueden implementar algunos métodos
sencillos que permiten corregir parte del error. Para que esto sea
posible se debe contar con varias réplicas de los conteos, ideal-
mente obtenidas en recorridos separados por poco tiempo (horas o
muy pocos días). Thorbjarnarson (1988b) resume este tipo de
correcciones en tres categorías:
o Aquélla que considera el número máximo de animales
observados en conteos sucesivos.
Tabla 5.3 Número de caimanes del Orinoco observados en el segmento
Confluencia-Caño Amarillo durante recorridos nocturnos efectuados en el
año 2006 (Ávila-Manjón, 2006).
Fecha
Sólo caimanes
<180 cm de LT Todas las tallas
17-ene-06 16 30
18-ene-06 15 27
01-feb-06 14 30
02-feb-06 15 39
20-feb-06 16 47
21-feb-06 9 36
28-mar-06 15 45
29-mar-06 13 42
19-abr-06 9 32
Promedio 13,6 36,4
Desviación estándar 2,7 7,2
Fracción observable (p) 0,622 0,628

Andrés Eloy Seijas
128
o La que considera el número máximo de individuos en cada
clase de tamaño de un conjunto de conteos.
o La que toma en cuenta la talla y la ubicación espacial de
los animales observados.
El primero de estos métodos consiste simplemente en tomar, de
entre un conjunto de conteos repetidos en una localidad, aquel que
arroje el número más alto. Es decir, si contamos individuos en dos
o más ocasiones y suponemos que en el lapso transcurrido entre
conteos no han ocurrido ni entradas (nacimientos o inmigraciones)
ni salidas (muertes o emigraciones), entonces el resultado con la
cifra más alta es el que se aproxima más al tamaño poblacional
verdadero.
El segundo método se basa igualmente en el supuesto de que no
hay ni entradas ni salidas en la población, pero la corrección se
realiza para cada clase de tamaño. Es decir, el mínimo número de
animales presentes de una determinada clase de tamaño está
representado por el máximo número de ellos contado en cualquiera
de los conteos individuales. Se presume que la ubicación de cada
animal en una determinada clase de tamaños no tiene error. Este
método es llamado por Messel et al. (1981) “método de los
máximos-mínimos” (MM).
En el tercer método toma en cuenta la ubicación de los
individuos durante los conteos. Es particularmente útil para mues-
treos llevados a cabo a lo largo de ríos, sobre todo si las
densidades son relativamente bajas. En esencia, se trata de mapear
la ubicación de los individuos observados. Por ejemplo, si a lo largo
de un segmento del río aparecen en conteos sucesivos registros de
un crocodilio de una determinada talla en un determinado lugar (o a
una distancia no mayor que la fijada por el investigador), entonces
se supone que las observaciones corresponden al mismo individuo.
Si las observaciones están muy distantes entre sí, entonces se
presume que se trata de individuos distintos. En la actualidad, la
ubicación precisa de los individuos con geoposicionadores sate-
litarios (GPS) permite refinar esta técnica. Mendoza (2003) la
utilizó en el Cojedes Norte; el mapeo de los animales observados

Los Crocodylia de Venezuela
129
en distintos muestreos le permitió determinar que en el segmento
muestreado había, al menos, 25 caimanes mayores de un año, aun
cuando el número máximo de caimanes contados en cualquier
salida fue de siete. Note que con este método también se puede
calcular la fracción de la población que es observable durante los
conteos nocturnos. Con los datos reportados por este último autor
dicha fracción varió desde 0%, para recorridos en los que no se
observó ningún individuo, hasta 28% para aquellos en que se
observó el número máximo.
ABUNDANCIA Y ESTRUCTURA POBLACIONAL DE
LAS ESPECIES
BABAS
Abundancia: Llanos y Guayana
La especie de crocodilio de Venezuela más estudiada es la
baba. Es para esta especie, por tanto, que existe mayor infor-
mación sobre su abundancia poblacional. Es sin duda la especie
más numerosa del país, pero, al igual como ocurre con las otras
especies de crocodilios de Venezuela, no existen registros deta-
llados sobre su abundancia histórica. Para el observador despre-
venido, y probablemente para los llaneros de las nuevas gene-
raciones, la baba siempre ha sido tan numerosa como en la
actualidad. Las anotaciones de Humboldt permiten deducir una
imagen muy distinta. En la narrativa de este naturalista alemán
contrasta la frecuencia con que se menciona a los caimanes con
las escasísimas veces en que se refiere a las babas. Se puede
suponer que si Humboldt se hubiera topado con una de esas
grandes concentraciones de babas como las que se encuentran en
la actualidad, sin duda habría escrito sobre ella. Se puede afirmar
que, simplemente, eso nunca pudo haber ocurrido, no obstante que
este naturalista recorrió parte de los llanos durante el periodo seco,
el más propicio para observar grandes grupos de babas.
La actual abundancia de babas en los llanos es producto de dos
fenómenos que se conjugan. El primero de ellos es la erradicación

Andrés Eloy Seijas
130
de los caimanes entre los años 1929 y 1960. Ese hecho eliminó
uno de los más formidables competidores-depredadores de las
babas. Los caimanes seguramente mantenían a las babas con-
finadas en pequeños caños y lagunas, los cuales les servían de
refugios frente a una especie de mayor tamaño y más agresiva,
impidiéndoles además invadir los grandes caños y ríos llaneros,
hábitat principal del caimán. La otra razón que explicaría la actual
abundancia de babas es la ampliación del hábitat disponible para
esta especie como consecuencia de la construcción de préstamos,
tapas y módulos en gran parte de las sabanas llaneras, ya sea para
fomentar la actividad ganadera o como subproducto de la cons-
trucción de carreteras y terraplenes.
Aun cuando las babas se concentran en los reducidos cuerpos
de aguas que persisten al final de la época seca, ellas se dispersan
por la sabana inundada durante la estación lluviosa, lo que ha dado
pie a cálculos de la densidad bruta, que toma en consideración
también terrenos que no conservan agua durante la época seca.
Así, han sido estimadas densidades de 0,17 babas/ha en el hato El
Frío de 78.000 ha (Ayarzagüena, 1983), 0,21 babas/ha para el
mismo hato (D´Andria, 1980) y 0,19 para un conjunto de 16 hatos
que cubrían un área de 233.800 ha (Seijas, 1986a). Cifras más
bajas que las anteriores son las reportadas por Ramo y Busto
(1989) para el Módulo Fernando Corrales, en Apure (0,04 ind/ha).
Para los llanos boscosos de Cojedes, en el hato Piñero de 75.000
ha, se han reportado densidades de 1,11 y 1,01 babas/ha
(Woodward y David, 1985; Fitzgerald, 1988a). Para las sabanas
moduladas del estado Apure, las densidades para la estación seca
han sido estimadas en 3,10 baba/ha por superficie de agua, valor
que se redujo a 1,60 baba/ha en la estación lluviosa (Ramos et al.,
1981). Como se puede observar hay un alto grado de variabilidad
en los valores reportados. Es difícil establecer en qué grado estas
diferencias se deben a factores ecológicos o son consecuencias de
usos de distintas metodologías.
En un intento por encontrar un patrón en las diferencias en la
abundancia relativa de babas, Velasco y Ayarzagüena (1995)
dividieron a los llanos en regiones ecológicas. (Fig. 5.8). Estos
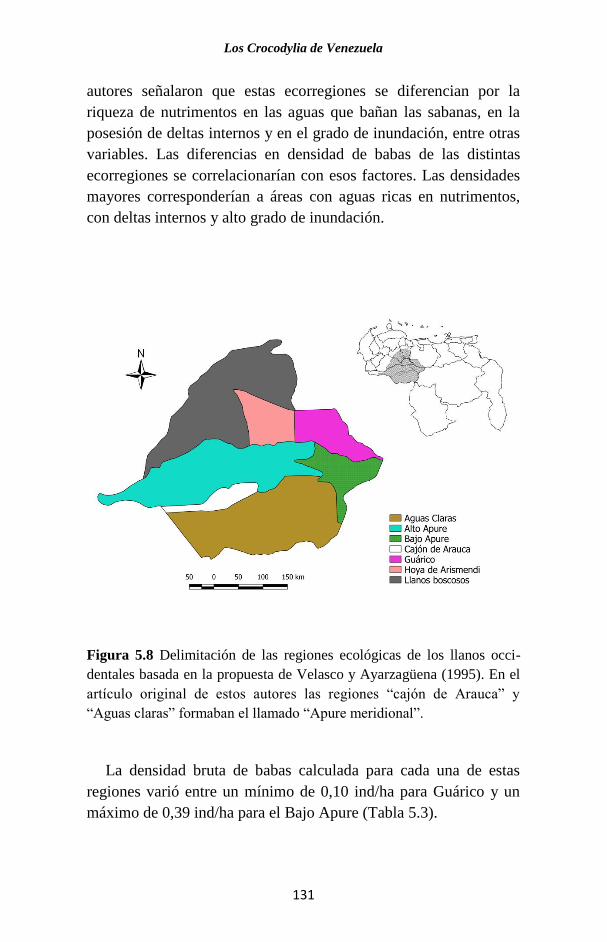
Los Crocodylia de Venezuela
131
autores señalaron que estas ecorregiones se diferencian por la
riqueza de nutrimentos en las aguas que bañan las sabanas, en la
posesión de deltas internos y en el grado de inundación, entre otras
variables. Las diferencias en densidad de babas de las distintas
ecorregiones se correlacionarían con esos factores. Las densidades
mayores corresponderían a áreas con aguas ricas en nutrimentos,
con deltas internos y alto grado de inundación.
Figura 5.8 Delimitación de las regiones ecológicas de los llanos occi-
dentales basada en la propuesta de Velasco y Ayarzagüena (1995). En el
artículo original de estos autores las regiones “cajón de Arauca” y
“Aguas claras” formaban el llamado “Apure meridional”.
La densidad bruta de babas calculada para cada una de estas
regiones varió entre un mínimo de 0,10 ind/ha para Guárico y un
máximo de 0,39 ind/ha para el Bajo Apure (Tabla 5.3).
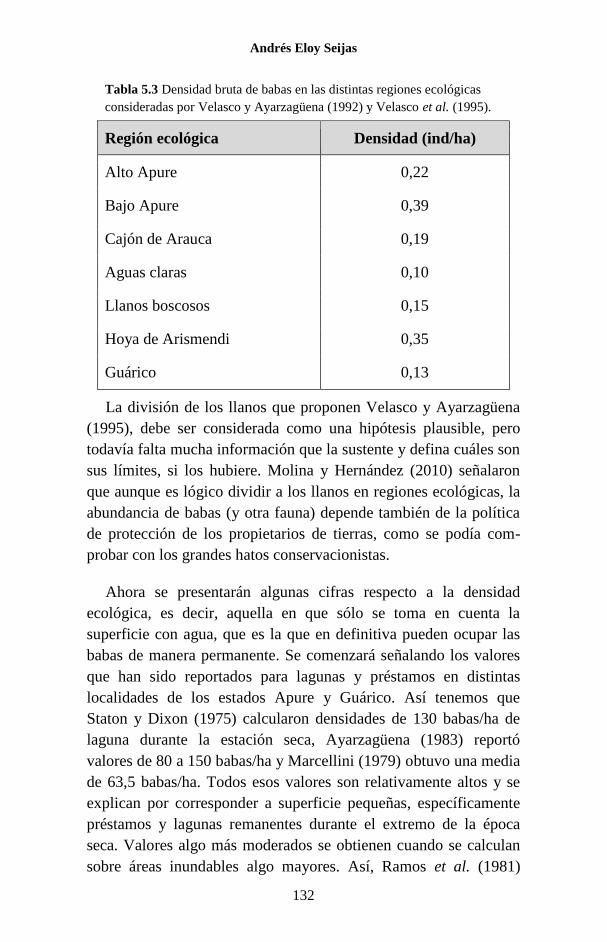
Andrés Eloy Seijas
132
Tabla 5.3 Densidad bruta de babas en las distintas regiones ecológicas
consideradas por Velasco y Ayarzagüena (1992) y Velasco et al. (1995).
Región ecológica Densidad (ind/ha)
Alto Apure 0,22
Bajo Apure 0,39
Cajón de Arauca 0,19
Aguas claras 0,10
Llanos boscosos 0,15
Hoya de Arismendi 0,35
Guárico 0,13
La división de los llanos que proponen Velasco y Ayarzagüena
(1995), debe ser considerada como una hipótesis plausible, pero
todavía falta mucha información que la sustente y defina cuáles son
sus límites, si los hubiere. Molina y Hernández (2010) señalaron
que aunque es lógico dividir a los llanos en regiones ecológicas, la
abundancia de babas (y otra fauna) depende también de la política
de protección de los propietarios de tierras, como se podía com-
probar con los grandes hatos conservacionistas.
Ahora se presentarán algunas cifras respecto a la densidad
ecológica, es decir, aquella en que sólo se toma en cuenta la
superficie con agua, que es la que en definitiva pueden ocupar las
babas de manera permanente. Se comenzará señalando los valores
que han sido reportados para lagunas y préstamos en distintas
localidades de los estados Apure y Guárico. Así tenemos que
Staton y Dixon (1975) calcularon densidades de 130 babas/ha de
laguna durante la estación seca, Ayarzagüena (1983) reportó
valores de 80 a 150 babas/ha y Marcellini (1979) obtuvo una media
de 63,5 babas/ha. Todos esos valores son relativamente altos y se
explican por corresponder a superficie pequeñas, específicamente
préstamos y lagunas remanentes durante el extremo de la época
seca. Valores algo más moderados se obtienen cuando se calculan
sobre áreas inundables algo mayores. Así, Ramos et al. (1981)

Los Crocodylia de Venezuela
133
reportan densidades de 3,1 y 1,6 ind/ha sobre superficies de 555,1
y 790,5 ha, respectivamente, del módulo experimental de Mantecal.
El valor calculados por Ramo y Busto (1989) en el módulo
Fernando Corrales, no muy lejos del lugar anterior, fue de 6,1 ind/
ha sobre una superficie de 85 ha de área encharcada.
Para la Guayana de Venezuela, (Gorzula, 1978) estimó una
densidad de 100 babas por ha de lagunas durante la estación seca
y de 10 babas/ha en los mismos cuerpos de agua para la estación
lluviosa. En estudios posteriores para esa región, Gorzula y Paolillo
(1986) estimaron una densidad global de 6,64 babas/ha de laguna.
Para la desembocadura del Orinoco, Velasco y Blanco (1996)
reportaron densidades observadas de 0,28 y 0,11 babas/ha para el
Alto y Medio Delta, respectivamente. Todas estas cifras son
difíciles de analizar para saber si existe algún tipo de patrón. La
densidad de babas en cada cuerpo de agua quizás dependa de
múltiples factores como extensión, profundidad, grado de
permanencia, cercanía a otros cuerpos de agua mayores, disponibilidad
de alimento, presión humana y otros.
Existe poca información sobre la abundancia de babas en caños
y ríos de los llanos de Venezuela. La mayor parte de los datos se
encuentran en trabajos cuyo objetivo principal era el de evaluar las
poblaciones de C. intermedius. Los ríos, y también los caños
mayores, se pueden clasificar de acuerdo con la productividad de
sus aguas en las categorías definidas por Sioli (1984) de aguas
blancas, aguas negras y aguas claras. Thorbjarnarson (1988b), en
parte apoyándose en dicho sistema de clasificación, divide a los
afluentes del Orinoco en dos grupos. El primero (I) contiene a los
tributarios que provienen de los llanos, que a su vez se pueden
dividir en: sub-grupo Ia, aquellos que se originan en las cordilleras
de los Andes o de la Costa, y sub-grupo Ib, los que forman el
drenaje interior de los propios llanos. El segundo grupo (II), lo
conforman los ríos de la Guayana, los cuales a su vez se dividen
en ríos de aguas negras (IIa) y ríos de Aguas Claras (IIb).
Los ríos que provienen de las montañas andinas y costeras (Ia)
son de aguas blancas, así denominados por el color que les

Andrés Eloy Seijas
134
confiere la alta carga de sedimentos que contienen; asimismo,
presentan una alta productividad. Los ríos de la red hídrica interna
de los llanos (Ib) poseen una menor carga de sedimentos y, en
muchos casos, son de aguas transparentes (aguas claras) y
relativamente poco productivos. Los tributarios guayaneses del Ori-
noco son también poco productivos. ¿Está relacionada la abundancia
de babas con las características ecológicas de los ríos en que se
encuentran? La Tabla 5.4 trata de responder esta pregunta.
Los datos presentados en la Tabla señalada anteriormente no
demuestran nada todavía, entre otras razones porque habría que
muestrear muchas otras localidades para comprobar si existe algún
tipo de patrón. La información para los ríos de la Guayana es
insuficiente; no existen datos para ríos y caños de algunas de las
regiones ecológicas propuestas por Velasco y Ayarzagüena (1995)
y, probablemente más importante que todo lo anterior, las meto-
dologías de trabajo de los distintos autores involucrados no nece-
sariamente son comparables. Por ejemplo, algunos reportes corres-
ponden a muestreos llevados a cabo temprano en la época seca,
mientras otros son más tardíos. No obstante, las cifras se corres-
ponden con lo esperado. Los valores mayores se encuentran en
ríos supuestamente más productivos de la vertiente andina y
costera del Orinoco. Los valores más bajos corresponden a ríos de
aguas claras y aguas negras del Apure meridional y la Guayana.
Otros factores que deben afectar la abundancia relativa de babas son
la presión de cacería y la presencia de caimanes (Thorbjarnarson y
Hernández, 1992; Seijas, 1998).
La productividad de las aguas debe ser uno de los factores
principales que pueden explicar la abundancia relativa de babas en
distintos ríos, pero ésta no es constante a lo largo de toda su
extensión y seguramente se incrementa aguas abajo, a medida que
el río aumenta su carga de sedimentos. Por lo demás, a medida que
avanzamos aguas abajo los ríos se hacen más anchos y más
profundos, y por tanto, más propicios para el establecimiento de
poblaciones de babas. Además discurren a través de áreas donde
los cuerpos de agua temporales propicios para el establecimiento de
poblaciones de babas son más frecuentes y extensos, cuyos
individuos se mueven hacia los ríos a medida que avanza la sequía.
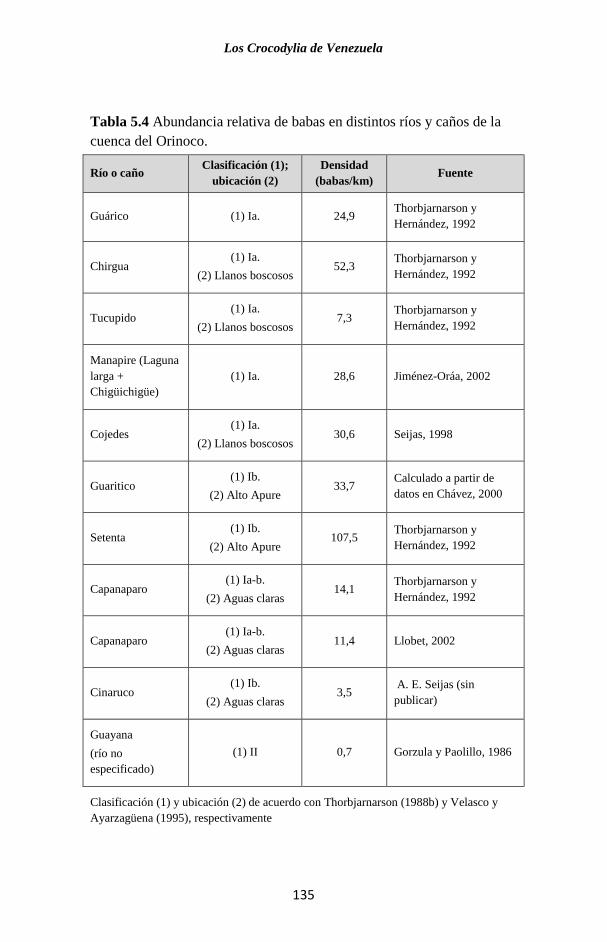
Los Crocodylia de Venezuela
135
Tabla 5.4 Abundancia relativa de babas en distintos ríos y caños de la
cuenca del Orinoco.
Río o caño Clasificación (1);
ubicación (2)
Densidad
(babas/km) Fuente
Guárico (1) Ia. 24,9 Thorbjarnarson y
Hernández, 1992
Chirgua (1) Ia.
(2) Llanos boscosos 52,3
Thorbjarnarson y
Hernández, 1992
Tucupido (1) Ia.
(2) Llanos boscosos 7,3
Thorbjarnarson y
Hernández, 1992
Manapire (Laguna
larga +
Chigüichigüe)
(1) Ia. 28,6 Jiménez-Oráa, 2002
Cojedes (1) Ia.
(2) Llanos boscosos 30,6 Seijas, 1998
Guaritico (1) Ib.
(2) Alto Apure 33,7
Calculado a partir de
datos en Chávez, 2000
Setenta (1) Ib.
(2) Alto Apure 107,5
Thorbjarnarson y
Hernández, 1992
Capanaparo (1) Ia-b.
(2) Aguas claras 14,1
Thorbjarnarson y
Hernández, 1992
Capanaparo (1) Ia-b.
(2) Aguas claras 11,4 Llobet, 2002
Cinaruco (1) Ib.
(2) Aguas claras 3,5
A. E. Seijas (sin
publicar)
Guayana
(río no
especificado)
(1) II 0,7 Gorzula y Paolillo, 1986
Clasificación (1) y ubicación (2) de acuerdo con Thorbjarnarson (1988b) y Velasco y
Ayarzagüena (1995), respectivamente
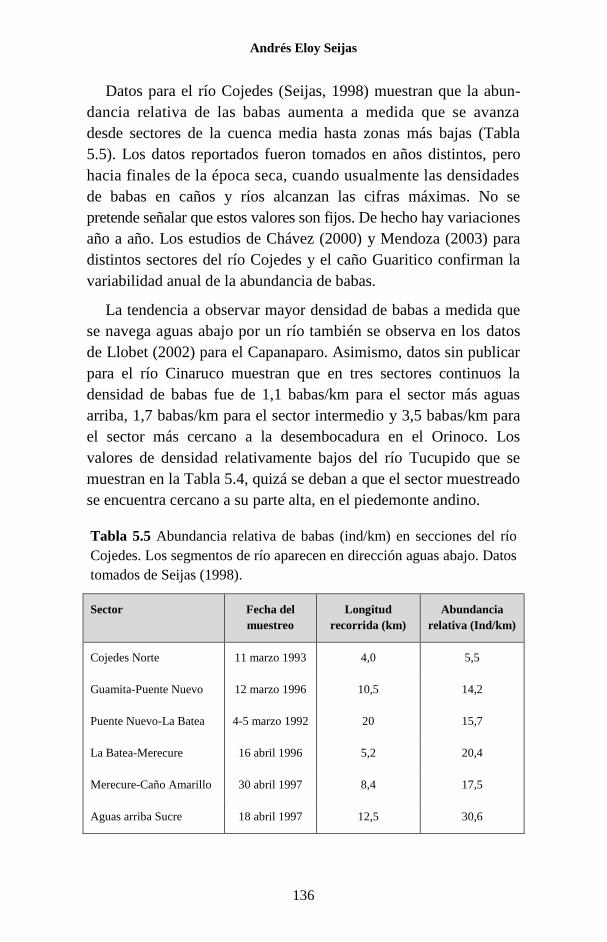
Andrés Eloy Seijas
136
Datos para el río Cojedes (Seijas, 1998) muestran que la abun-
dancia relativa de las babas aumenta a medida que se avanza
desde sectores de la cuenca media hasta zonas más bajas (Tabla
5.5). Los datos reportados fueron tomados en años distintos, pero
hacia finales de la época seca, cuando usualmente las densidades
de babas en caños y ríos alcanzan las cifras máximas. No se
pretende señalar que estos valores son fijos. De hecho hay variaciones
año a año. Los estudios de Chávez (2000) y Mendoza (2003) para
distintos sectores del río Cojedes y el caño Guaritico confirman la
variabilidad anual de la abundancia de babas.
La tendencia a observar mayor densidad de babas a medida que
se navega aguas abajo por un río también se observa en los datos
de Llobet (2002) para el Capanaparo. Asimismo, datos sin publicar
para el río Cinaruco muestran que en tres sectores continuos la
densidad de babas fue de 1,1 babas/km para el sector más aguas
arriba, 1,7 babas/km para el sector intermedio y 3,5 babas/km para
el sector más cercano a la desembocadura en el Orinoco. Los
valores de densidad relativamente bajos del río Tucupido que se
muestran en la Tabla 5.4, quizá se deban a que el sector muestreado
se encuentra cercano a su parte alta, en el piedemonte andino.
Tabla 5.5 Abundancia relativa de babas (ind/km) en secciones del río
Cojedes. Los segmentos de río aparecen en dirección aguas abajo. Datos
tomados de Seijas (1998).
Sector Fecha del
muestreo
Longitud
recorrida (km)
Abundancia
relativa (Ind/km)
Cojedes Norte 11 marzo 1993 4,0 5,5
Guamita-Puente Nuevo 12 marzo 1996 10,5 14,2
Puente Nuevo-La Batea 4-5 marzo 1992 20 15,7
La Batea-Merecure 16 abril 1996 5,2 20,4
Merecure-Caño Amarillo 30 abril 1997 8,4 17,5
Aguas arriba Sucre 18 abril 1997 12,5 30,6

Los Crocodylia de Venezuela
137
Abundancia: Región costera
Hay menos información sobre la abundancia de las babas en la
región costera de Venezuela. La mayor parte de los trabajos
publicados que aportan datos sobre esta especie han tenido como
objetivo principal el estudio de la situación del caimán de la costa,
crocodilio con el que coexiste en muchas localidades.
Para cinco embalses de esta región se reportaron densidades de
babas que van desde un mínimo de 2 ind/km en el embalse de
Pueblo Viejo del estado Zulia, hasta un máximo de 8,6 ind/km en el
embalse de Cumaripa, del estado Yaracuy (Seijas, 1986b). La
abundancia relativa de la baba parece estar correlacionada
negativamente con la del caimán de la costa en los tres embalses donde
esta especie también estuvo presente.
Seijas (1986b,c) mostró que en ríos y caños costeros las
densidades de babas resultaron mayores en localidades donde el
caimán de la costa estuvo ausente (7,1 babas/km a lo largo de 143
km de río) en comparación con hábitats donde estas especies eran
simpátricas (1,5 babas/km en 86,9 km de ríos muestreados). La
interpretación de estas relaciones se complica debido a la amplia
variación en las densidades, particularmente de las babas, a medida
que factores como el nivel de las aguas, cambian en el tiempo. Al
igual que sucedió con los ríos de la cuenca del Orinoco, en los ríos
Tocuyo (Falcón), Yaracuy (Yaracuy) y Neverí (Anzoátegui), las
densidades de babas se incrementaron a medida que se avanzaba
hacia la desembocadura (Seijas ,1986b; Seijas y Chávez, 1991). La
densidad más alta registrada (26,3 nd/km) correspondió al río Unare
(Anzoátegui) en su últimos 25 km. Cerca de la boca del río
Yaracuy se registró una densidad aun mayor (61 babas/km), pero a
lo largo de un segmento muy corto (1,7 km).
Estructura poblacional
En el estudio poblacional pionero de Staton y Dixon (1975), se
agruparon a las babas observadas en categorías de 10 cm de LCC.
La mayor parte de los estudios posteriores han adoptado el sistema
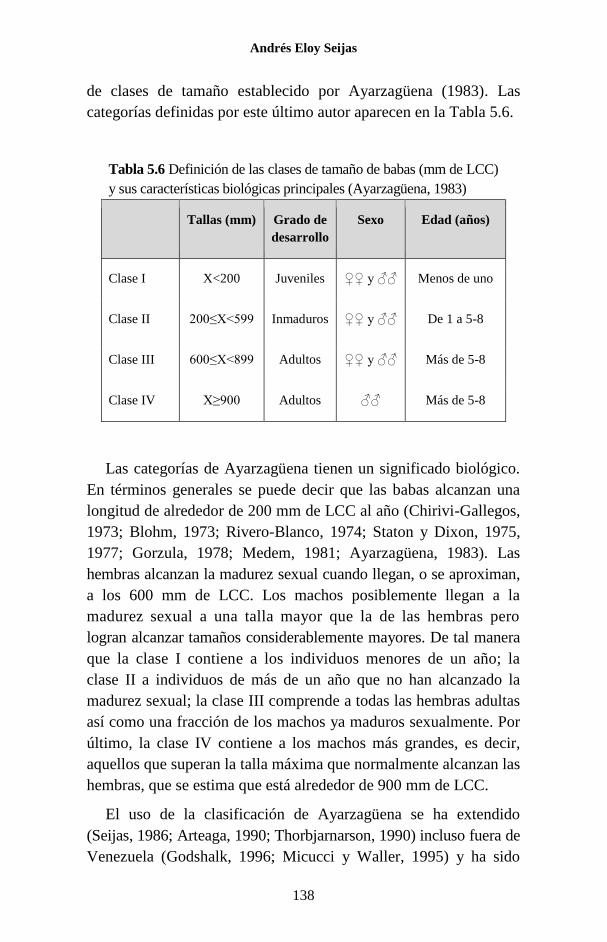
Andrés Eloy Seijas
138
de clases de tamaño establecido por Ayarzagüena (1983). Las
categorías definidas por este último autor aparecen en la Tabla 5.6.
Tabla 5.6 Definición de las clases de tamaño de babas (mm de LCC)
y sus características biológicas principales (Ayarzagüena, 1983)
Tallas (mm) Grado de
desarrollo
Sexo Edad (años)
Clase I X<200 Juveniles ♀♀ y ♂♂ Menos de uno
Clase II 200≤X<599 Inmaduros ♀♀ y ♂♂ De 1 a 5-8
Clase III 600≤X<899 Adultos ♀♀ y ♂♂ Más de 5-8
Clase IV X≥900 Adultos ♂♂ Más de 5-8
Las categorías de Ayarzagüena tienen un significado biológico.
En términos generales se puede decir que las babas alcanzan una
longitud de alrededor de 200 mm de LCC al año (Chirivi-Gallegos,
1973; Blohm, 1973; Rivero-Blanco, 1974; Staton y Dixon, 1975,
1977; Gorzula, 1978; Medem, 1981; Ayarzagüena, 1983). Las
hembras alcanzan la madurez sexual cuando llegan, o se aproximan,
a los 600 mm de LCC. Los machos posiblemente llegan a la
madurez sexual a una talla mayor que la de las hembras pero
logran alcanzar tamaños considerablemente mayores. De tal manera
que la clase I contiene a los individuos menores de un año; la
clase II a individuos de más de un año que no han alcanzado la
madurez sexual; la clase III comprende a todas las hembras adultas
así como una fracción de los machos ya maduros sexualmente. Por
último, la clase IV contiene a los machos más grandes, es decir,
aquellos que superan la talla máxima que normalmente alcanzan las
hembras, que se estima que está alrededor de 900 mm de LCC.
El uso de la clasificación de Ayarzagüena se ha extendido
(Seijas, 1986; Arteaga, 1990; Thorbjarnarson, 1990) incluso fuera de
Venezuela (Godshalk, 1996; Micucci y Waller, 1995) y ha sido

Los Crocodylia de Venezuela
139
tomada como base para el monitoreo de las poblaciones sometidas
a aprovechamiento por el Ministerio del Ambiente (Velasco y
Ayarzagüena, 1995, Velasco et al., 1995, Venezuela 1999). Por
razones que se desconocen el sistema adoptado por el MINAMB
modificó el límite de tamaño entre las clases I y II, llevándolo a
500 mm de LT, lo que determina que la clase I contenga individuos
de hasta, al menos, dos años.
El sistema de clasificación en Categorías de Tamaños es a
veces expresado en referencia a la LT en vez de la LCC. Es
precisamente esta modalidad la que adoptó el Ministerio del
Ambiente en Venezuela. En este caso los límites que definen a cada
clase duplican a los de Ayarzagüena (1983) al considerar, de manera
aproximada, que la LT es el doble de la LCC. Así, por ejemplo, la
clase IV comprende a los individuos mayores de 1800 mm de LT. Es
necesario aclarar que en realidad la LT es 1,864 veces mayor que la
LCC (ver Eq. I.15 en el Apéndice I) por lo que siguiendo de manera
estricta a Ayarzagüena (1983) la Clase IV estaría constituida por
individuos mayores de 1678 mm de LT. Asimismo, el límite de
tamaños que define la madurez sexual es de 1118 mm de LT, y no
1200 mm de LT como indica la estructura que estableció el MARNR.
Existe gran variabilidad en las estructuras de tamaño que muestran
las poblaciones de babas en distintas localidades de los llanos (Fig.
5.9), supuestamente debido a una combinación de factores que incluye
la presión de cacería, las características ecológicas de cada región y,
también, la experticia de las personas que toman los datos.
Los valores de la Figura 5.9 representan sólo una muestra
de las cifras disponibles en la literatura. No se puede hablar de
homogeneidad ni siquiera dentro de las regiones. En el caso del
Alto Apure, por ejemplo, Velasco y Ayarzagüena (1995) dividieron
el grupo de hatos muestreados en cuatro lotes. Un análisis a
posteriori con los datos de estos autores muestra que las estruc-
turas de tamaño para esta última región son estadísticamente
diferentes entre sí (X2=566, P<0,0001). Por otra parte, las estruc-
turas mostradas por los citados investigadores para Guárico son
notoriamente distintas a las de otras localidades en ese mismo
estado reportadas por Arteaga (1989) y Thorbjarnarson (1990).
Andrés Eloy Seijas
140
Figura 5.9 Estructura de tamaños de distintas poblaciones de babas de los
llanos de Venezuela. A: Ayarzagüena, 1983; B: Seijas, 1986a (sólo los
datos de Apure); C: Thorbjarnarson, 1990. D, E, F y G: Velasco y
Ayarzagüena, 1995. H: Seijas y Chávez, 1996. Es costumbre en estos
gráficos no reportar a los individuos de la Clase I, debido a que su
número es muy cambiante dependiendo de le época en que se realicen los
muestreos.
0
10
20
30
40
50
60
70
Po
rcen
taje
Clase II Clase III Clase IV
A: Apure, El Frío
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
B: Apure (Muñoz)
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
C: Guárico, Masaguaral
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
D: Guárico
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
E: Alto Apure
0
10
20
30
40
50
60
70
Po
rcen
taje
Clase II Clase III Clase IV
F: Llanos boscosos
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
G: Apure, aguas claras
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
H: Cojedes (El Eneal)
0
10
20
30
40
50
60
70
Po
rcen
taje
Clase II Clase III Clase IV
A: Apure, El Frío
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
B: Apure (Muñoz)
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
C: Guárico, Masaguaral
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
D: Guárico
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
E: Alto Apure
0
10
20
30
40
50
60
70
Po
rcen
taje
Clase II Clase III Clase IV
F: Llanos boscosos
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
G: Apure, aguas claras
0
10
20
30
40
50
60
70
Porc
enta
je
Clase II Clase III Clase IV
H: Cojedes (El Eneal)
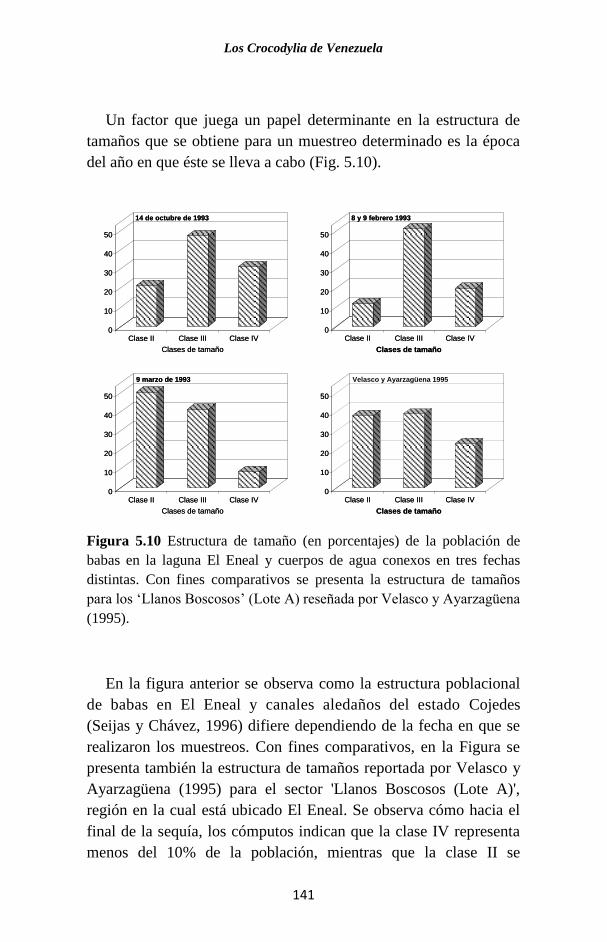
Los Crocodylia de Venezuela
141
Un factor que juega un papel determinante en la estructura de
tamaños que se obtiene para un muestreo determinado es la época
del año en que éste se lleva a cabo (Fig. 5.10).
Figura 5.10 Estructura de tamaño (en porcentajes) de la población de
babas en la laguna El Eneal y cuerpos de agua conexos en tres fechas
distintas. Con fines comparativos se presenta la estructura de tamaños
para los ‘Llanos Boscosos’ (Lote A) reseñada por Velasco y Ayarzagüena
(1995).
En la figura anterior se observa como la estructura poblacional
de babas en El Eneal y canales aledaños del estado Cojedes
(Seijas y Chávez, 1996) difiere dependiendo de la fecha en que se
realizaron los muestreos. Con fines comparativos, en la Figura se
presenta también la estructura de tamaños reportada por Velasco y
Ayarzagüena (1995) para el sector 'Llanos Boscosos (Lote A)',
región en la cual está ubicado El Eneal. Se observa cómo hacia el
final de la sequía, los cómputos indican que la clase IV representa
menos del 10% de la población, mientras que la clase II se
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
14 de octubre de 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
8 y 9 febrero 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
9 marzo de 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
Velasco y Ayarzagüena 1995
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
14 de octubre de 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
8 y 9 febrero 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
9 marzo de 1993
0
10
20
30
40
50
Clase II Clase III Clase IV
Clases de tamaño
Velasco y Ayarzagüena 1995

Andrés Eloy Seijas
142
convierte en la categoría más numerosa. La estructura de tamaños
para el 9 de marzo de 1993 es estadísticamente diferente a la
reportada por Velasco y Ayarzagüena (X²=41,02; P<0.001). Como
conclusión hay que señalar que la verdadera estructura de la
población es difícil de establecer y que es necesario reportar en
que época se realizaron los muestreos. Desafortunadamente esa
información no aparece en el trabajo de Velasco y Ayarzagüena
(1995), lo cual le quita valor a la comparación.
No sólo la fecha en que se hacen los conteos tiene incidencia
en los resultados, ya que incluso la hora en que se realizan éstos
puede ser un factor determinante. Thorbjarnarson (1990) presentó
estructuras de tamaños obtenidas en cuerpos de agua el mismo día
pero a diferentes horas (Fig. 5.11).
La estructura de tamaños también puede variar con el tipo de
hábitat. Hay ambientes que pueden ser propicios para el estable-
cimiento de los individuos de una determinada talla o edad, como
ocurre con las crías que ocupan principalmente cuerpos de agua
relativamente pequeños, poco profundos y con abundante vegeta-
ción acuática, donde por lo general permanecen protegidos por una
hembra adulta. Thorbjarnarson (1990) muestra gráficamente esta
situación, al comparar la abundancia relativa de babas, por cate-
goría de tamaños, en préstamos y lagunas de distintas dimensiones.
Aunque cualquier individuo de una especie de Crocodylia puede
ser ubicado en alguno de los estadios definidos por Ayarzagüena,
las tallas que delimitan esos estadios o clases de tamaño varían con
las especies, subespecies e, incluso, entre poblaciones de la misma
especie que habitan en condiciones ecológicas distintas. Las babas
de la Guayana Venezolana, por ejemplo, parecen ser más pequeñas
que las de los llanos y no se conoce cuál es la talla a la que las
hembras de esta región adquieren la madurez sexual (lo cual
delimitaría el comienzo de la clase III). En el río Yaracuy las
babas tampoco parecen alcanzar las grandes tallas comunes en los
llanos. Si se clasifica a la población de babas de esa localidad
(Seijas y Chávez, 1991) de acuerdo con las categorías del
MARNR, se obtiene una estructura en las que las babas Clase IV
están prácticamente ausentes.
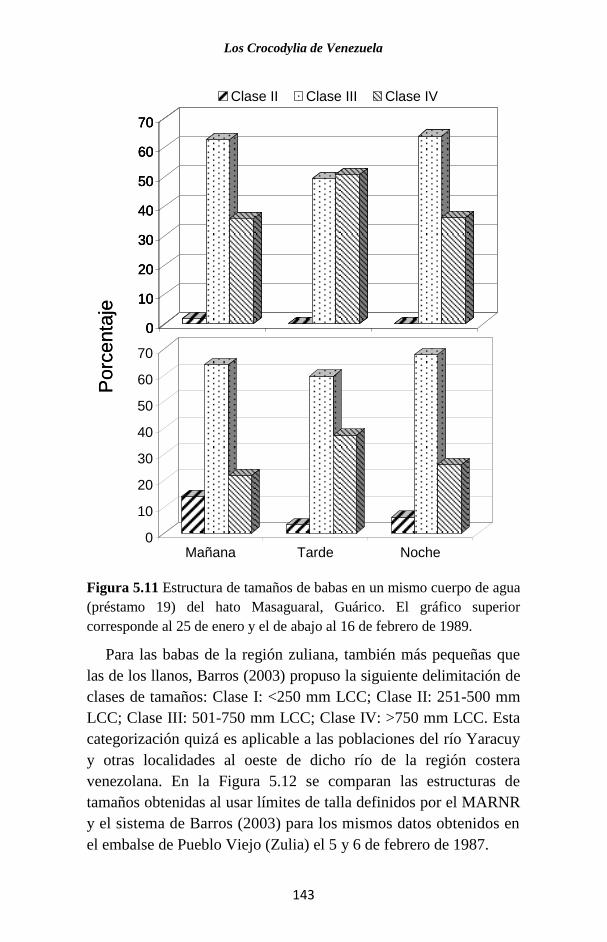
Los Crocodylia de Venezuela
143
Figura 5.11 Estructura de tamaños de babas en un mismo cuerpo de agua
(préstamo 19) del hato Masaguaral, Guárico. El gráfico superior
corresponde al 25 de enero y el de abajo al 16 de febrero de 1989.
Para las babas de la región zuliana, también más pequeñas que
las de los llanos, Barros (2003) propuso la siguiente delimitación de
clases de tamaños: Clase I: <250 mm LCC; Clase II: 251-500 mm
LCC; Clase III: 501-750 mm LCC; Clase IV: >750 mm LCC. Esta
categorización quizá es aplicable a las poblaciones del río Yaracuy
y otras localidades al oeste de dicho río de la región costera
venezolana. En la Figura 5.12 se comparan las estructuras de
tamaños obtenidas al usar límites de talla definidos por el MARNR
y el sistema de Barros (2003) para los mismos datos obtenidos en
el embalse de Pueblo Viejo (Zulia) el 5 y 6 de febrero de 1987.
0
10
20
30
40
50
60
70
Mañana Tarde Noche
Clase II Clase III Clase IV
0
10
20
30
40
50
60
70
Mañana Tarde Noche
Po
rce
nta
je
0
10
20
30
40
50
60
70
Mañana Tarde Noche
Clase II Clase III Clase IV
0
10
20
30
40
50
60
70
Mañana Tarde Noche
0
10
20
30
40
50
60
70
Mañana Tarde Noche
Clase II Clase III Clase IV
0
10
20
30
40
50
60
70
Mañana Tarde Noche
Po
rce
nta
je
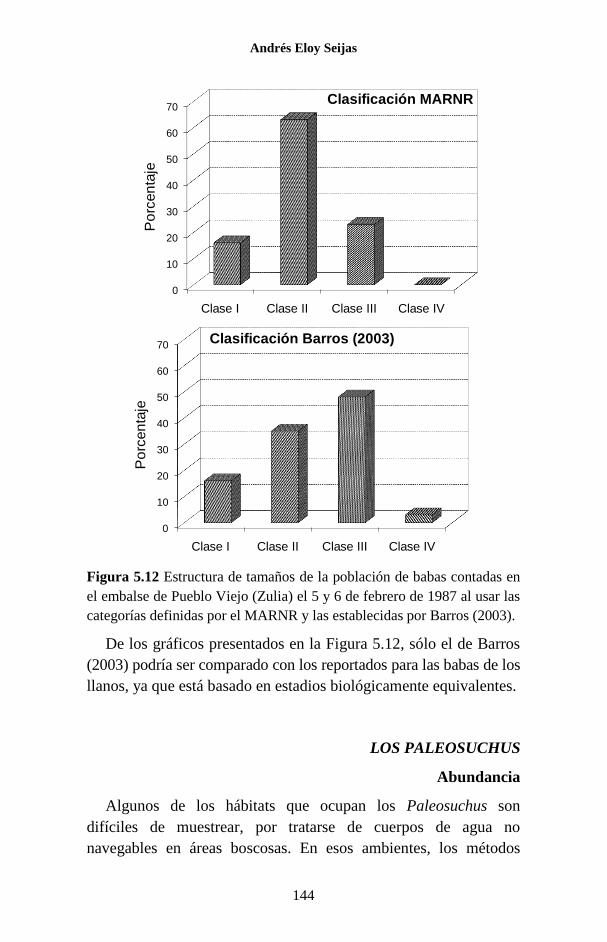
Andrés Eloy Seijas
144
Figura 5.12 Estructura de tamaños de la población de babas contadas en
el embalse de Pueblo Viejo (Zulia) el 5 y 6 de febrero de 1987 al usar las
categorías definidas por el MARNR y las establecidas por Barros (2003).
De los gráficos presentados en la Figura 5.12, sólo el de Barros
(2003) podría ser comparado con los reportados para las babas de los
llanos, ya que está basado en estadios biológicamente equivalentes.
LOS PALEOSUCHUS
Abundancia
Algunos de los hábitats que ocupan los Paleosuchus son
difíciles de muestrear, por tratarse de cuerpos de agua no
navegables en áreas boscosas. En esos ambientes, los métodos
0
10
20
30
40
50
60
70
Clase I Clase II Clase III Clase IV
Po
rcen
taje
Clasificación MARNR
0
10
20
30
40
50
60
70
Clase I Clase II Clase III Clase IV
Po
rcen
taje
Clasificación Barros (2003)

Los Crocodylia de Venezuela
145
tradicionales para el conteo de crocodílidos (recorridos nocturnos
con botes y linternas) son de poca aplicabilidad. Por eso no
sorprende la poca información sobre la abundancia de estas
especies en el país. Métodos de trabajos más intensivos, que
incluyan largos periodos de trabajo, recorridos a pie a lo largo de
las orillas, captura y marcaje de individuos y seguimiento por radio
telemetría, tal como lo hicieron Magnusson y Lima (1991) con P.
trigonatus en Brasil, serían los más recomendables.
Solamente Gorzula y Paolillo (1986) y Pacheco (2009) han
publicado información sobre la abundancia relativa de los
Paleosuchus en Venezuela. En la Tabla 5.7 se presentan los datos
de estos investigadores. Se incluyen en la tabla los resultados de un
conteo en el río Cinaruco (Seijas sin publicar).
Las densidades conocidas para los ríos nos indican que los
Paleosuchus son, comparados con las babas, relativamente escasos
(ver tablas 5.4 y 5.5). Gorzula y Paolillo (1986) reportan para C.
crocodilus densidades promedio de 2,5 ind/km. en ríos y ria-
chuelos, valor muy superiores a los mostrados en la Tabla 5.7 para
los babos negro y morichalero, con excepción de la cifra de 7,50
ind/km que es un valor exagerado provocado por la poca extensión
muestreada. En el trayecto del río Cinaruco señalado en la Tabla
5.7 se observaron 4 P. palpebrosus y 24 C. crocodilus, una
abundancia relativa seis veces mayor de esta segunda especie con
respecto a la primera. Pacheco (2009) por su parte encontró
durante la época seca siete veces más babas que P. palpebrosus
en el Río Canjilones del estado Anzoátegui, pero en el cercano Río
Negro el mismo autor reporta que el babo morichalero supera en
abundancia (2,8:1) a la baba, lo que indica que en cierto tipo de
hábitats Paleosuchus puede ser la especie dominante.
En el río Bita de Colombia, que se une al Orinoco muy cerca
de la desembocadura del Meta, Ríos y Trujillo (2004) reportaron
haber contado, en un recorrido total de 26,8 Km., 2 P.
palpebrosus y 235 C. crocodilus, una diferencia de abundancia
relativa considerable. A lo largo de ambas márgenes tanto del
Cinaruco como del Bita, como se puede constatar al analizar
imágenes de satélite Landsat TM, se encuentra un conjunto de
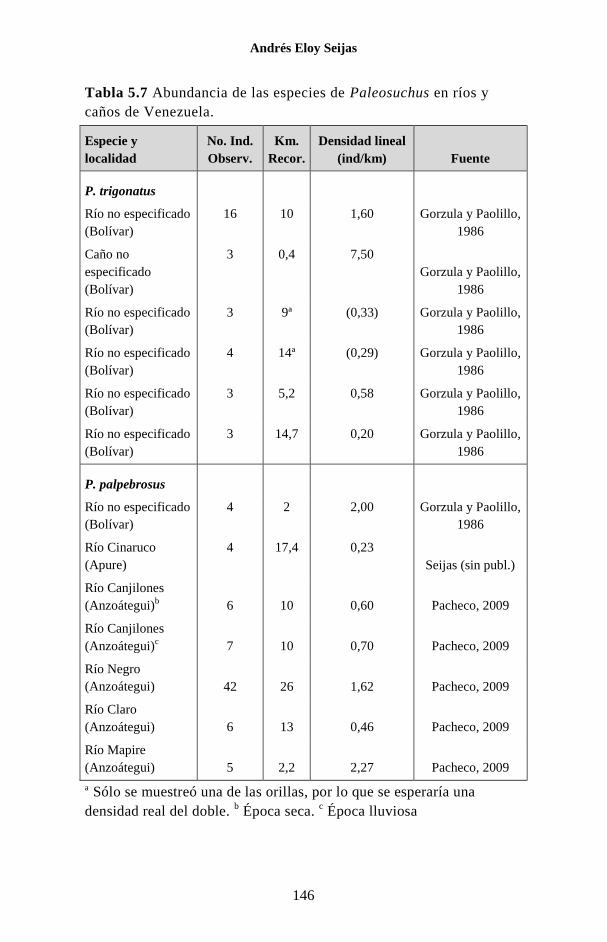
Andrés Eloy Seijas
146
Tabla 5.7 Abundancia de las especies de Paleosuchus en ríos y
caños de Venezuela.
Especie y
localidad
No. Ind.
Observ.
Km.
Recor.
Densidad lineal
(ind/km) Fuente
P. trigonatus
Río no especificado
(Bolívar)
16 10 1,60 Gorzula y Paolillo,
1986
Caño no
especificado
(Bolívar)
3 0,4 7,50
Gorzula y Paolillo,
1986
Río no especificado
(Bolívar)
3 9ª (0,33) Gorzula y Paolillo,
1986
Río no especificado
(Bolívar)
4 14ª (0,29) Gorzula y Paolillo,
1986
Río no especificado
(Bolívar)
3 5,2 0,58 Gorzula y Paolillo,
1986
Río no especificado
(Bolívar)
3 14,7 0,20 Gorzula y Paolillo,
1986
P. palpebrosus
Río no especificado
(Bolívar)
4 2 2,00 Gorzula y Paolillo,
1986
Río Cinaruco
(Apure)
4 17,4 0,23
Seijas (sin publ.)
Río Canjilones
(Anzoátegui)b 6 10 0,60 Pacheco, 2009
Río Canjilones
(Anzoátegui)c 7 10 0,70 Pacheco, 2009
Río Negro
(Anzoátegui) 42 26 1,62 Pacheco, 2009
Río Claro
(Anzoátegui) 6 13 0,46 Pacheco, 2009
Río Mapire
(Anzoátegui) 5 2,2 2,27 Pacheco, 2009
a Sólo se muestreó una de las orillas, por lo que se esperaría una
densidad real del doble. b Época seca.
c Época lluviosa
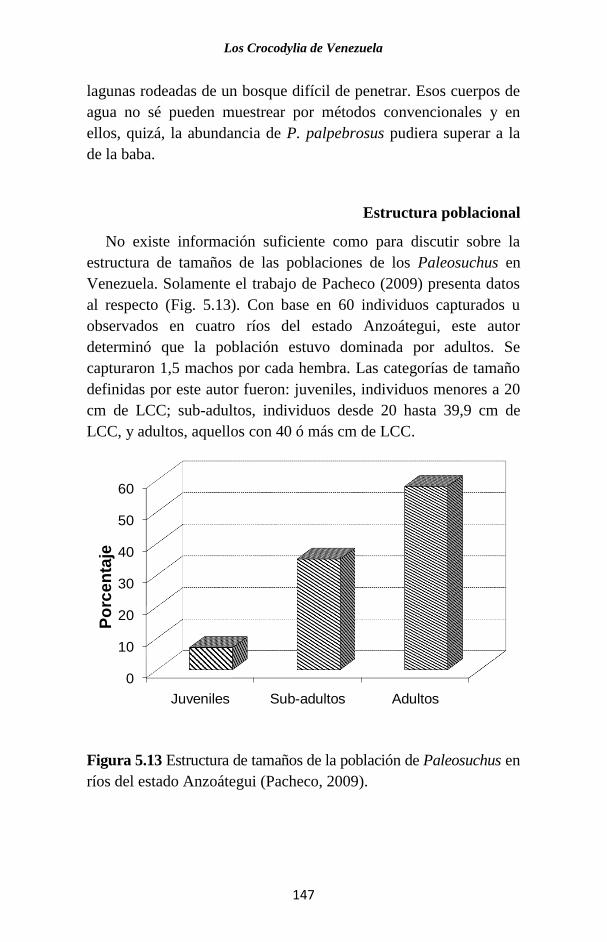
Los Crocodylia de Venezuela
147
lagunas rodeadas de un bosque difícil de penetrar. Esos cuerpos de
agua no sé pueden muestrear por métodos convencionales y en
ellos, quizá, la abundancia de P. palpebrosus pudiera superar a la
de la baba.
Estructura poblacional
No existe información suficiente como para discutir sobre la
estructura de tamaños de las poblaciones de los Paleosuchus en
Venezuela. Solamente el trabajo de Pacheco (2009) presenta datos
al respecto (Fig. 5.13). Con base en 60 individuos capturados u
observados en cuatro ríos del estado Anzoátegui, este autor
determinó que la población estuvo dominada por adultos. Se
capturaron 1,5 machos por cada hembra. Las categorías de tamaño
definidas por este autor fueron: juveniles, individuos menores a 20
cm de LCC; sub-adultos, individuos desde 20 hasta 39,9 cm de
LCC, y adultos, aquellos con 40 ó más cm de LCC.
Figura 5.13 Estructura de tamaños de la población de Paleosuchus en
ríos del estado Anzoátegui (Pacheco, 2009).
0
10
20
30
40
50
60
Juveniles Sub-adultos Adultos
Po
rce
nta
je

Andrés Eloy Seijas
148
CAIMÁN DE LA COSTA
Abundancia
Muy poco se conoce sobre la distribución y abundancia del
caimán de la costa en Venezuela en épocas muy anteriores a la
explotación comercial de mediados del siglo XX que diezmó sus
poblaciones. Las únicas referencias confiables son las Alejandro de
Humboldt y las de Karl Ferdinan Appun, otro naturalista alemán
que visitó el país a mediados del siglo XIX (Appun, 1961). De lo
anotado por estos autores se desprende que C. acutus era muy
abundante en los ríos Neverí y Yaracuy. Las pocas referencias de
mediados del siglo XX (Röhl, 1956; Donoso-Barros, 1966b), así
como algunas relativamente más recientes (Maness, 1982; Medem,
1983; Seijas, 1988, 1991b, 1996) permiten suponer que el caimán
de la costa abundaba en todos los ríos de la cuenca del Lago de
Maracaibo y en los ríos más importantes que drenan hacia la costa
del Caribe, entre otros el Tocuyo, El Aroa, el Tuy y el Unare,
además de los ya mencionados Yaracuy y Neverí. Asimismo, el
caimán de la costa probablemente se encontraba en grandes
números en los manglares del río Limón (Zulia), río Hueque, Cuare
y Morrocoy (Falcón), Buche y Laguna de Tacarigua (Miranda) por
sólo mencionar las localidades más grandes y conocidas.
En un extenso recorrido por la región costera venezolana entre
los años 1980 y 1984, el cual incluyó más de 50 localidades, sólo
se pudo constatar la presencia de caimanes en 14 lugares (Seijas,
1986c). Exploraciones posteriores y datos obtenidos por información
de otros investigadores han ampliado la lista hasta una veintena de
sitios (Seijas, 1988). En la Tabla 5.8 se presentan los datos sobre
la abundancia de C. acutus en las localidades conocidas, colocando
en cada caso la cifra más alta reportada desde 1980, cuando se
iniciaron los estudios detallados con esta especie.
Otras localidades, no incluidas en la Tabla 5.8, donde se ha
confirmado en los últimos 30 años la presencia de C. acutus son
los ríos Aricuaisá, Santa Rosa, Torondoy y Chama en la cuenca
del Lago de Maracaibo; embalses Machango y Agua Viva, en los

Los Crocodylia de Venezuela
149
estados Zulia y Trujillo, respectivamente; Golfete de Coro, río
Mitare y embalse El Camare del estado Falcón (Seijas, 1986c,
1990; Lander et al., 2008; Tito Barros y Luis Sánchez, com.
pers.). Quizás con la excepción del río Santa Rosa, las poblaciones
de caimán de la costa en esas localidades son muy pequeñas.
No existe un programa nacional de monitoreo continuo de las
poblaciones de C. acutus. La mayor parte de las cifras reportadas
en la Tabla 5.8 son ya un poco viejas, pero sirven de línea base
con la cual comparar los resultados de estudios futuros. Pero la
información disponible permite, para algunas localidades, comparar
la situación de la especie en distintos años para así determinar si
han ocurrido cambios. De esta manera, se puede constatar que
existió un considerable incremento en el número de animales
observados respecto a estudios previos para el embalse de Pueblo Viejo
(109%), Parque Nacional Morrocoy (280%), Turiamo (100%)
y Parque Nacional Laguna de Tacarigua (112%). En otras loca-
lidades ha ocurrido una considerable disminución. Esto último es el
caso del Refugio de Fauna de Cuare, donde Manness (1982) contó
23 individuos adultos en 1975. Los muestreos en esta localidad a
comienzos de los años 80 arrojaron una cifra máxima de 2
individuos (Seijas, 1986c), aunque Arteaga (1997) reportó 23 indivi-
duos en los primeros años de la década de los 90, cuando ya un
programa de reforzamiento poblacional se había iniciado.
Las poblaciones de caimán de la costa en Venezuela se
encuentran en su mayoría aisladas entre sí. Como se señaló hace
unos cuantos años (Seijas, 1991b) en la actualidad es casi imposible
que ocurra, por ejemplo, el intercambio genético necesario para
reducir la endogamia entre los caimanes de la Laguna de Taca-
rigua, en Miranda, y los escasos individuos que hayan podido
sobrevivir en el río Neverí. Si algún caimán intentara ese viaje, a
través del mar, se encontraría con un ambiente degradado y hostil,
lleno de playas contaminadas, construcciones portuarias y seres
humanos -pescadores y turistas- dispuestos a matarlo en la primera
oportunidad que se les presente (Fig. 5.14). El flujo genético entre
los caimanes de las localidades mencionadas estaba garantizado en
el pasado, cuando estos cocodrilos abundaban a lo largo de toda la
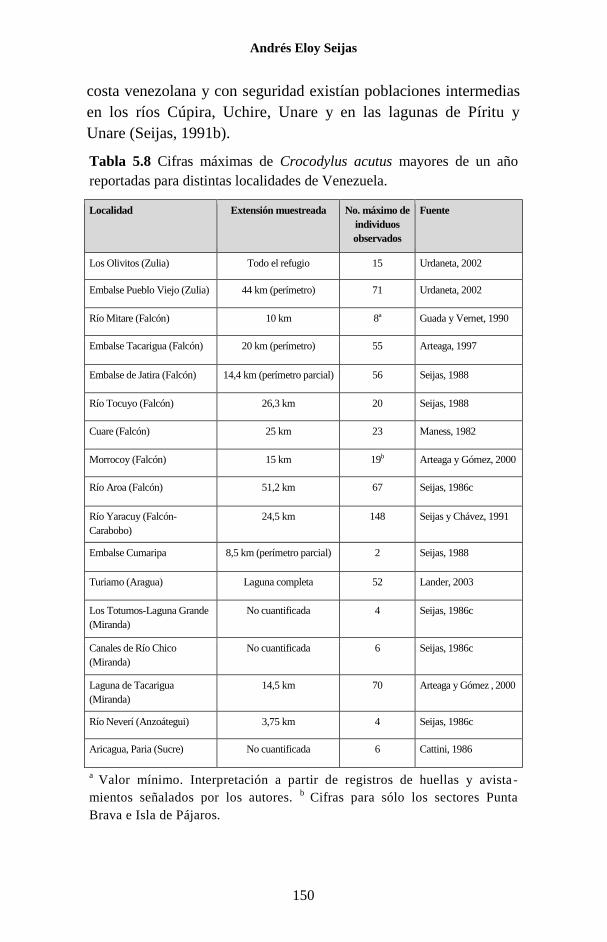
Andrés Eloy Seijas
150
costa venezolana y con seguridad existían poblaciones intermedias
en los ríos Cúpira, Uchire, Unare y en las lagunas de Píritu y
Unare (Seijas, 1991b).
Tabla 5.8 Cifras máximas de Crocodylus acutus mayores de un año
reportadas para distintas localidades de Venezuela.
Localidad Extensión muestreada No. máximo de
individuos
observados
Fuente
Los Olivitos (Zulia) Todo el refugio 15 Urdaneta, 2002
Embalse Pueblo Viejo (Zulia) 44 km (perímetro) 71 Urdaneta, 2002
Río Mitare (Falcón) 10 km 8a Guada y Vernet, 1990
Embalse Tacarigua (Falcón) 20 km (perímetro) 55 Arteaga, 1997
Embalse de Jatira (Falcón) 14,4 km (perímetro parcial) 56 Seijas, 1988
Río Tocuyo (Falcón) 26,3 km 20 Seijas, 1988
Cuare (Falcón) 25 km 23 Maness, 1982
Morrocoy (Falcón) 15 km 19b Arteaga y Gómez, 2000
Río Aroa (Falcón) 51,2 km 67 Seijas, 1986c
Río Yaracuy (Falcón-
Carabobo)
24,5 km 148 Seijas y Chávez, 1991
Embalse Cumaripa 8,5 km (perímetro parcial) 2 Seijas, 1988
Turiamo (Aragua) Laguna completa 52 Lander, 2003
Los Totumos-Laguna Grande
(Miranda)
No cuantificada 4 Seijas, 1986c
Canales de Río Chico
(Miranda)
No cuantificada 6 Seijas, 1986c
Laguna de Tacarigua
(Miranda)
14,5 km 70 Arteaga y Gómez , 2000
Río Neverí (Anzoátegui) 3,75 km 4 Seijas, 1986c
Aricagua, Paria (Sucre) No cuantificada 6 Cattini, 1986
a Valor mínimo. Interpretación a partir de registros de huellas y avista -
mientos señalados por los autores. b
Cifras para sólo los sectores Punta
Brava e Isla de Pájaros.

Los Crocodylia de Venezuela
151
Figura 5.14 Izquierda. Fotografía en el diario El Nacional (17-11-2004) de
un caimán de la costa capturado en Playa Los Cocos en el litoral central.
El animal, con múltiples heridas, fue trasladado al zoológico de Caricuao
donde murió unas tres semanas después. La autopsia reveló que tenía en
el estómago el anzuelo mostrado en la fotografía de la derecha
(información aportada por Ricardo Babarro).
Mucho más distantes entre sí se encuentran las localidades del
estado Miranda que aún conservan caimanes (Laguna de Taca-
rigua, por ejemplo) y la bahía de Turiamo, en Aragua. Pero en el
pasado con seguridad había poblaciones intermedias que facilitaban
el flujo genético. En la colección del Museo de la Fundación La
Salle, en Caracas, hay una cría de C. acutus que fue colectada en
enero de1955 en Camurí Chico, estado Vargas, lo que demuestra
que todavía en un pasado relativamente reciente existían pobla-
ciones de esta especie en el litoral central del país.
Estructura poblacional
En los estudios sobre la estructura de tamaños de las pobla-
ciones de caimanes de la costa en Venezuela se ha utilizado el
sistema de 5 clases ya discutido anteriormente. En la mayor parte
de las localidades estudiadas hasta ahora, las poblaciones del C.
acutus aparecen dominadas por individuos juveniles (clases II y
III). La únicas excepciones corresponden a los muestreos llevados a
cabo en el embalse de Jatira el 15 de marzo de 1987 (Seijas,
1988), y a los resultados reportados por Arteaga y Gómez (2000)
(Fig. 5.15).

Andrés Eloy Seijas
152
Figura 5.15 Estructura de tamaños de distintas poblaciones de C. acutus
en Venezuela. Sólo se muestran gráficos basados en la observación de 50
o más individuos. Clases II y III, juveniles; Clase IV sub-adultos y Clase
V, adultos. Las Fuentes para estos gráficos son: Lander, 2003 (Turiamo),
Urdaneta, 2002 (Pueblo Viejo), Arteaga y Gómez, 2000 (PN Lag.
Tacarigua), Seijas, 1988 (Jatira) y Seijas y Chávez, 1991 (Yaracuy).
Vale la pena discutir las posibles causas de las diferencias en
las estructuras de tamaño presentadas. La dominancia de individuos
pequeños quizás refleje la fuerte presión que ejerce la gente sobre
los animales adultos que, por ser grandes y conspicuos, son
frecuentemente cazados. Los caimanes más pequeños, al pasar
más fácilmente inadvertidos, permanecen en la población, al menos
mientras crecen. Aunque parezca contradictorio, otro factor que
seguramente influye en determinar la estructura observada es la
timidez de los caimanes mayores. Los animales grandes tienden a
hundirse muy temprano cuando un bote se aproxima, y escapan a
la detección (Seijas, 1988). Por otra parte, los animales grandes
tienden a ocupar pozos más profundos, donde les es más fácil
sumergirse por completo. En los conteos nocturnos es frecuente
incluir a los individuos para los que no fue posible estimar la talla
en una categoría denominada “sin datos”. Muy frecuentemente se
trata de animales observados a mucha distancia, muy proba-
blemente animales grandes.
0
10
20
30
40
50
60
70P
orc
en
taje
Clase II Clase III Clase IV Clase V
Pueblo Viejo (n=50)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
Jatira (n=68)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clases IV-V
Yaracuy (n=148)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
Turiamo (n=55)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
PN Lag. Tacarigua (n=80)
0
10
20
30
40
50
60
70P
orc
en
taje
Clase II Clase III Clase IV Clase V
Pueblo Viejo (n=50)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
Jatira (n=68)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clases IV-V
Yaracuy (n=148)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
Turiamo (n=55)
0
10
20
30
40
50
60
70
Po
rce
nta
je
Clase II Clase III Clase IV Clase V
PN Lag. Tacarigua (n=80)

Los Crocodylia de Venezuela
153
Los resultados reportados por Arteaga y Gómez (2000) para la
Laguna de Tacarigua en Miranda, muestran muy pocos clase V
(adultos) y una importante representación de caimanes de la clase
IV (subadultos). Estos autores señalan que la población de C.
acutus estaba creciendo durante los años de estudio (1997 a 1999),
lo cual se reflejaba principalmente en el número de individuos de
esa última categoría de tamaño. En este caso se puede comprobar
que la proporción de individuos clase V es una subestimación, ya
que ellos reportaron la localización de 50 nidos en el año 1999, lo
que por supuesto significa la existencia de al menos igual número
de hembras adultas. Sin embargo, sólo 8 de los individuos obser-
vados durante los conteos nocturnos fueron asignados a esa
categoría. El sesgo pudiera haber ocurrido al momento de estimar
las tallas durante los conteos nocturnos y que algunos de los 48
caimanes que ellos colocaron en la clase IV, pertenecieran en
realidad a la clase V.
En el caso de Jatira ejemplifica también las dificultades en
establecer de manera precisa la estructura de tamaños de una
población de crocodilios. El gráfico para esta localidad mostrado en
la Figura 5.14, representa un ajuste basado en tres conteos
realizados entre marzo y junio de 1987. En uno de esos conteos se
observaron 42 ejemplares adultos, mientras que en los otros dos
conteos se observaron sólo 6 individuos de esa talla. La gran
diferencia radica en que en el primero de esos conteos se logró
recorrer una zona nunca antes explorada, ya que el nivel del agua
en el embalse permitió la navegación como en ninguna otra
ocasión. De no haberse presentado esa afortunada circunstancia,
los resultados habrían sido completamente diferentes.
CAIMÁN DEL ORINOCO
Abundancia
Las anotaciones de Humboldt (1975, 1991) constituyen la fuente
antigua más confiable que permiten formarnos una imagen de la
abundancia de los caimanes en una época muy anterior a la

Andrés Eloy Seijas
154
cacería comercial que los llevó al borde de la extinción en el siglo
XX. Son frecuentes las referencias de este insigne alemán en las
que muestra asombro por la cantidad de caimanes con que “hier-
ven los ríos”. Unas pocas citas permiten formarse una idea de
cuan numerosas eran las poblaciones de C. intermedius:
“Más abajo de la desembocadura del Arauca aparecieron
los caimanes en mayor número que hasta entonces. Los
indios nos dijeron que los saurios provenían del interior del
país, donde habían pasado su período de letargo en el
barro seco de las sabanas. Tan pronto como vuelven a la
vida con las primeras lluvias, se reúnen en manadas y se
dirigen al río [Orinoco] donde se dispersan nuevamente.”
(Humboldt, 1975, p. 206)
“Los indios nos aseguraban que los cocodrilos nuevos
prefieren las charcas y los ríos menos anchos y profundos.
Se acumulan sobre todo en los caños, y está uno por decir
de ellos lo que Abd-Allatif dice de los cocodrilos del Nilo:
”que hormiguean como gusanos en las aguas bajas del río
y al abrigo de las islas inhabitadas” (Humboldt, 1991, Tomo
III, p. 324).
“Cuando las playas son de una anchura considerable [en
el Apure] … vence los cocodrilos, a menudo en número de
8 ó 10, tendidos sobre la arena, inmóviles, … Se han
multiplicado de tal manera estos reptiles monstruosos, que
a todo lo largo del río hemos tenido a la vista, casi a cada
instante, cinco o seis de ellos.” (Humboldt, 1991, Tomo III, p.
293 y 294).
Hoy en día se pudiera recorrer al río Apure en toda su
extensión sin contar, en total, el número de caimanes del Orinoco
que Humboldt pudo observar en una sola playa. La primera
evaluación de la situación de esta especie, luego de que su cacería
comercial fue prohibida, fue llevada a cabo por Robert Godshalk,
con los auspicios de la Fundación para la Defensa de la Naturaleza
(FUDENA) entre los años 1976 y 1977 (Godshalk, 1978, 1982).
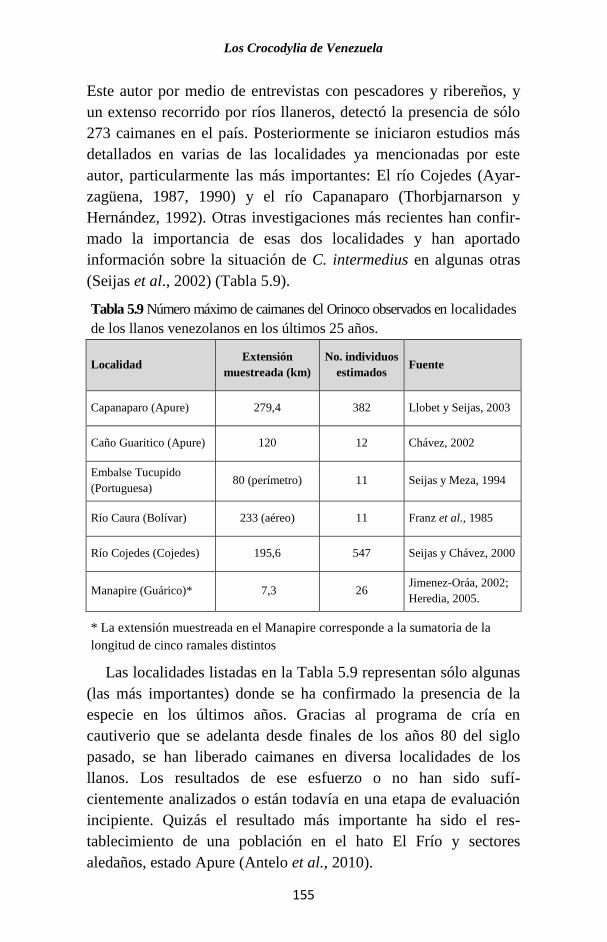
Los Crocodylia de Venezuela
155
Este autor por medio de entrevistas con pescadores y ribereños, y
un extenso recorrido por ríos llaneros, detectó la presencia de sólo
273 caimanes en el país. Posteriormente se iniciaron estudios más
detallados en varias de las localidades ya mencionadas por este
autor, particularmente las más importantes: El río Cojedes (Ayar-
zagüena, 1987, 1990) y el río Capanaparo (Thorbjarnarson y
Hernández, 1992). Otras investigaciones más recientes han confir-
mado la importancia de esas dos localidades y han aportado
información sobre la situación de C. intermedius en algunas otras
(Seijas et al., 2002) (Tabla 5.9).
Tabla 5.9 Número máximo de caimanes del Orinoco observados en localidades
de los llanos venezolanos en los últimos 25 años.
Localidad Extensión
muestreada (km)
No. individuos
estimados Fuente
Capanaparo (Apure) 279,4 382 Llobet y Seijas, 2003
Caño Guaritico (Apure) 120 12 Chávez, 2002
Embalse Tucupido
(Portuguesa) 80 (perímetro) 11 Seijas y Meza, 1994
Río Caura (Bolívar) 233 (aéreo) 11 Franz et al., 1985
Río Cojedes (Cojedes) 195,6 547 Seijas y Chávez, 2000
Manapire (Guárico)* 7,3 26 Jimenez-Oráa, 2002;
Heredia, 2005.
* La extensión muestreada en el Manapire corresponde a la sumatoria de la
longitud de cinco ramales distintos
Las localidades listadas en la Tabla 5.9 representan sólo algunas
(las más importantes) donde se ha confirmado la presencia de la
especie en los últimos años. Gracias al programa de cría en
cautiverio que se adelanta desde finales de los años 80 del siglo
pasado, se han liberado caimanes en diversa localidades de los
llanos. Los resultados de ese esfuerzo o no han sido sufí-
cientemente analizados o están todavía en una etapa de evaluación
incipiente. Quizás el resultado más importante ha sido el res-
tablecimiento de una población en el hato El Frío y sectores
aledaños, estado Apure (Antelo et al., 2010).

Andrés Eloy Seijas
156
A la luz de los resultados mostrados en la Tabla 5.9 se puede
concluir que la situación de C. intermedius en Venezuela no es tan
desfavorable como la que reportó Godshalk en 1978. No se puede
afirmar, sin embargo, que las cifras que se conocen en la actua-
lidad signifiquen que la especie se ha recuperado un poco. Las
valores más altos de la actualidad pueden deberse a un mejor
conocimiento de la real situación de la especie, producto de
estudios más detallados e intensivos. La especie sigue estando en
una situación crítica y en algunos sectores hay datos que indican
que existe un deterioro de su situación, como es el caso de
diversos sectores del río Cojedes, donde la población ha disminuido
de manera notoria (Mendoza, 2003; Seijas et al., 2010). Asimismo,
la población que quedó encerrada en el embalse de Tucupido
(Seijas y Meza, 1994) es mucho menor que la que se esperaría de
acuerdo con estimaciones previas a la construcción de este cuerpo
de agua (Ramo y Busto, 1986; Thorbjarnarson y Hernández, 1992).
Otras localidades donde parece haber habido un deterioro de la
población son los ríos Meta y Cinaruco (Seijas, 2007b).
Estructura poblacional
Las poblaciones de caimanes del Orinoco son tan pequeñas que
sólo tiene sentido describir las estructuras de tamaños de las
poblaciones del Capanaparo y del Cojedes. En el primero de estos
ríos los datos más recientes son de Llobet (2002). En el caso del
Cojedes existen datos mucho más abundantes, que corresponden a
varios años de muestreos, desde los iniciales reportados por
Ayarzagüena (1987) hasta los de Ávila Manjón (2008) y Espinosa
(2010). Se mostrarán sólo algunos gráficos que permitan discutir las
particularidades más resaltantes de la estructura poblacional de C.
intermedius en esas localidades (Fig. 5.16).
En el Capanaparo los caimanes adultos (Clase V) representan
un porcentaje importante de la población (entre 19,8 y 30,6%). En
el caso del Cojedes, solamente en la sección de La Batea-
Merecure, los adultos representan un porcentaje destacable. Una
relativa abundancia de adultos puede ser indicativa de poca presión
de cacería, la cual afecta principalmente a los adultos, pero las
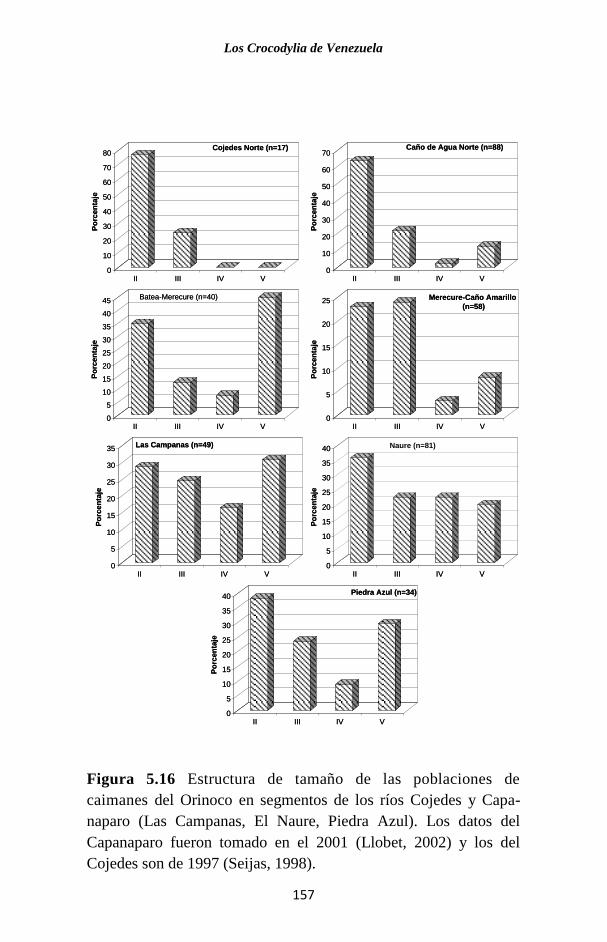
Los Crocodylia de Venezuela
157
Figura 5.16 Estructura de tamaño de las poblaciones de
caimanes del Orinoco en segmentos de los ríos Cojedes y Capa-
naparo (Las Campanas, El Naure, Piedra Azul). Los datos del
Capanaparo fueron tomado en el 2001 (Llobet, 2002) y los del
Cojedes son de 1997 (Seijas, 1998).
0
10
20
30
40
50
60
70
80
Po
rcen
taje
II III IV V
Cojedes Norte (n=17)
0
10
20
30
40
50
60
70
Po
rcen
taje
II III IV V
Caño de Agua Norte (n=88)
0
5
10
15
20
25
30
35
40
45
Po
rcen
taje
II III IV V
Batea-Merecure (n=40)
0
5
10
15
20
25
Po
rcen
taje
II III IV V
Merecure-Caño Amarillo
(n=58)
0
5
10
15
20
25
30
35
Po
rcen
taje
II III IV V
Las Campanas (n=49)
0
5
10
15
20
25
30
35
40
Po
rcen
taje
II III IV V
Naure (n=81)
0
5
10
15
20
25
30
35
40
Po
rcen
taje
II III IV V
Piedra Azul (n=34)
0
10
20
30
40
50
60
70
80
Po
rcen
taje
II III IV V
Cojedes Norte (n=17)
0
10
20
30
40
50
60
70
Po
rcen
taje
II III IV V
Caño de Agua Norte (n=88)
0
5
10
15
20
25
30
35
40
45
Po
rcen
taje
II III IV V
Batea-Merecure (n=40)
0
5
10
15
20
25
Po
rcen
taje
II III IV V
Merecure-Caño Amarillo
(n=58)
0
5
10
15
20
25
30
35
Po
rcen
taje
II III IV V
Las Campanas (n=49)
0
5
10
15
20
25
30
35
40
Po
rcen
taje
II III IV V
Naure (n=81)
0
5
10
15
20
25
30
35
40
Po
rcen
taje
II III IV V
Piedra Azul (n=34)

Andrés Eloy Seijas
158
diferencias entre secciones del mismo río respecto a la importancia
relativa de alguna clase de tamaño en particular puede también ser
un reflejo de contrastes en la calidad del hábitat. La sección La
Batea-Merecure del río Cojedes, por ejemplo, posee quizás las
mejores condiciones para la anidación de C. intermedius (Seijas,
1998) lo cual explicaría la presencia de numerosos individuos
adultos allí. La sección inmediata aguas abajo (Merecure-Caño
Amarillo) posee escaso hábitat de anidación, por lo que la población
está constituida principalmente de juveniles y sub-adultos. Hay que
hacer notar, en todo caso, que las estructuras de tamaños no son
una característica fija de las poblaciones. En el caso del último
segmento señalado del río Cojedes, los datos de Ávila (2008)
indican que la proporción de animales adultos en la población se ha
incrementado.

Los Crocodylia de Venezuela
159
CAPÍTULO 6
DIETA
Un aspecto relevante en el estudio ecológico de cualquier
especie animal es el referido a su dieta. Los estudios sobre la
dieta nos indican: 1) qué tipo de presa caza determinada especie de
crocodilio; 2) cuándo y dónde las captura y 3) cuáles son sus
presas más importantes. Toda esa información es relevante desde
el punto de vista de la conservación y el manejo. La disponibilidad
y calidad de los recursos alimentarios son indicadores de la
idoneidad del hábitat para una especie, pero solamente podemos
evaluar esa calidad si sabemos de qué se alimenta la especie cuyo
hábitat queremos evaluar.
Por otra parte, el conocimiento de la dieta de una especie
aporta información directa acerca de sus relaciones tróficas, sobre
el tipo de interacciones que establece con las otras especies de la
comunidad. Así se puede inferir su papel ecológico en el eco-
sistema que ocupa, el impacto que su erradicación podría causar y
la ruta que algunos nutrientes o contaminantes pudieran seguir
hasta llegar a alcanzarla. Al mismo tiempo orienta sobre el tipo de
actividades de manejo que pudieran implementarse para favorecerla
o perjudicarla, si alguna circunstancia particular así lo requiriera.
La dieta de tres de las especies de crocodilios de Venezuela ha
sido estudiada en diferentes grados de detalles. Para la baba se
tienen los estudios realizados en distintas localidades de los llanos
(Staton y Dixon, 1975; Ayarzagüena, 1983, 1988; Seijas y Ramos,
1980; Fitzgerald, 1988a; Thorbjarnarson, 1990), en la región de
Guayana (Gorzula, 1978) y en la región costera (Seijas 1988, 1996;
Chávez, 1992). La dieta de caimán de la costa ha sido estudiada

Andrés Eloy Seijas
160
en tres embalses (Pueblo Viejo, Jatira y Tacarigua) por Seijas
(1988, 1996), Chávez (1992) y Arteaga (1998), y en el río Yaracuy
(Seijas y Chávez, 1991). Para el caimán del Orinoco sólo hay
información recabada en el río Cojedes (Seijas, 1998). Sólo existe
un trabajo sobre la dieta de una de las especies de Paleosuchus (P.
palpebrosus) realizado en Venezuela (González Orihuela, 2010).
Métodos
Los métodos que han sido usados para los estudios de la dieta
han sido diversos. En las investigaciones iniciales (Gorzula, 1978;
Seijas y Ramos, 1980; Ayarzagüena, 1983) el procedimiento usado
para obtener la información consistió, al menos parcialmente, en el
sacrificio de los animales. Como anécdota vale la pena referir que
esta forma de trabajo fue criticada públicamente por el australiano
William Magnusson durante la reunión del Grupo de Especialistas
en Cocodrilos de la UICN que se realizó en Caracas en 1984, al
referirse en particular al trabajo de Seijas y Ramos ya citado.
Quizás en la mente de Magnusson estaba todavía fresca la imagen
de la carnicería de millares de crocodilios, sacrificados con fines de
“investigación”, que había sido denunciada años antes por Gans y
Pooley (1976). Aun cuando sin duda crueles, estos métodos tienen
la ventaja de permitir la obtención de la totalidad del contenido
estomacal, eliminando errores en análisis comparativos. Sin embar-
go, esta forma de trabajo debe ser descartada en la actualidad y
sólo se justificaría si los animales deben ser inevitablemente
sacrificados por alguna otra razón. En este sentido, es sorprenden-
temente que no existan publicaciones sobre dieta de la baba que
estén basados en el abundantísimo material disponible que ha
significado la cosecha autorizada por el MARN de cerca de
millón y medio de individuos adultos desde el año 1983.
Parte de la información sobre la dieta, particularmente de babas,
ha sido obtenida al revisar los contenidos estomacales de animales
atropellados en las carreteras (Staton y Dixon, 1975; Ayarzagüena,
1983; Thorbjarnarson, 1990). Existen también, y en la actualidad son
ampliamente usados, métodos de extracción por lavado del conte-
nido estomacal de individuos vivos. El primero en usar un proce-

Los Crocodylia de Venezuela
161
dimiento de este tipo en el país fue José Ayarzagüena (1983),
quien luego de inmovilizar a las babas con amarras y mantenerles
la boca abierta con trozos de madera, introducía una manguera y
expulsaba el contenido estomacal con la presión del agua. Un mé-
todo más refinado es el descrito por Fitzgerald (1988b) quien usó,
además del agua a presión, la denominada “maniobra de Heimlich”,
la cual consiste en ejercer presión con las manos sobre el
estómago del animal para facilitar la expulsión de su contenido. El
procedimiento de Fitzgerald ha sido empleado también con C.
acutus (Seijas, 1988, 1996; Chávez, 1992; Arteaga, 1998) y con C.
intermedius (Seijas, 1998). En cualquiera de las modalidades de
lavado para la remoción del contenido de los estómagos, el agua
junto al material extraído son recogidos en un recipiente. Con un
colador se separan del agua las presas o sus restos y se preservan
para su posterior análisis.
Un inconveniente del método de Fitzgerald es que en con-
diciones de campo es a veces difícil de implementar un mecanismo
para introducir agua a presión en el estómago de los animales.
Para solventar este problema se han usado bombas manuales, o
recipientes con agua colocadas a cierta altura (techo de los
vehículos, por ejemplo) para que el agua baje a través de la
manguera por gravedad. La presión del agua en estos casos no es
muy grande y no se ha evaluado su incidencia en la eficiencia del
método. Por otra parte, el equipo que se requiere no es portátil, por
lo que se necesita instalar un campamento base para llevar los
animales, a veces desde una distancia considerable.
Otro método, descrito por Taylor (1979), consiste en introducir
agua al estómago a través de una manguera y un embudo (Fig.
6.1), para luego extraer el contenido estomacal con la ayuda de un
tipo de cuchara. Ese sistema de muestreo ha sido ensayado en
Venezuela por distintos investigadores (Fitzgerald, 1988a; Alfredo
Arteaga com. pers.) pero de manera esporádica. Fitzgerald (1988b)
encontró, al comparar el contenido estomacal remanente en grupos
de babas sacrificadas después de removerles el contenido esto-
macal por alguno de los dos métodos ya señalados, que la técnica
del lavado con la introducción de agua a presión y la aplicación de

Andrés Eloy Seijas
162
la maniobra de Heimlich fue mucho más eficiente que el descrito
por Taylor (1979).
Un aspecto que hay que tener en cuenta a la hora de analizar
los resultados de todos estos métodos es el tiempo transcurrido
entre la captura del animal y el momento cuando se le remueve el
contenido estomacal. De manera ideal, la extracción debería
realizarse inmediatamente después de la captura, lo cual no es
siempre posible. Una pronta extracción reduce el tiempo de
digestión y posibilita que presas recién capturadas por el crocodilio
aparezcan más completas, lo que facilita tanto su identificación
como la determinación de su peso y volumen.
En estudios de la dieta de babas y caimanes en la región
costera de Venezuela (Seijas, 1988, 1996; Seijas y Chávez, 1991) las
capturas de los animales se realizaban en la noche y la extracción
se efectuaba a primera hora del día siguiente, por lo que en
algunos casos podían transcurrir hasta 10 horas entre la captura y
la colecta del contenido estomacal. Los resultados obtenidos de
esta manera pueden estar sesgados a favor de renglones cuya
digestión es más lenta, teniendo en cuenta que la carne es digerida
más rápidamente que los huesos, y estos a su vez se digieren más
rápidamente que el componente quitinoso de algunos invertebrados.
Para acortar el tiempo entre captura y extracción de contenido
estomacal es recomendable atrapar a los animales en las últimas
horas de la madrugada y efectuar el lavado apenas amanezca, o
antes, si las condiciones lo permiten. El lavado estomacal inmedia-
tamente después de la captura es posible de realizar, pero significa
por lo general trabajar en el bote y con una serie de inconvenientes
y limitaciones que incomodan el desempeño de los investigadores.
Por lo general, los contenidos estomacales son preservados en
formol al 10% o, preferiblemente, alcohol etílico al 70%, para su
posterior análisis en el laboratorio. Allí, los renglones de cada
muestra son separados y ligeramente presionados con papel absor-
bente para retirar el exceso de líquido. Luego se les determina su
volumen (en mililitros) por desplazamiento de agua en un cilindro
graduado. Por lo general se intenta una identificación de las presas

Los Crocodylia de Venezuela
163
consumidas al nivel taxonómico más bajo posible. Se recomienda
recolectar muestras de las presas potenciales en el área de estudio
para preparar una colección de referencia y facilitar la identi-
ficación de los fragmentos encontrados en los estómagos. El
número, tamaño y volumen original de las presas capturadas
pueden ser estimados a partir de los fragmentos encontrados.
Figura 6.1 Preparación de una baba para proceder a extraerle el con-
tenido estomacal.
Para determinar cambios ontogénicos en la dieta, los contenidos
estomacales se agrupan de acuerdo con la talla de los crocodilios
de los cuales se extrajeron. Es preferible trabajar siempre con la
longitud corporal (LCC) de los crocodilios, tomada desde la punta
del hocico hasta el extremo posterior de la abertura cloacal, ya que
en algunas localidades una alta proporción de los animales tienen la
cola rota, por lo que la longitud total (LT) no refleja fielmente la
talla del animal.

Andrés Eloy Seijas
164
No existe un modo perfecto para cuantificar la dieta de los
crocodilios. Los análisis basados en la frecuencia de aparición de
las presas probablemente sobreestimen la importancia de aquellos
renglones que poseen estructuras de difícil digestión, como por
ejemplo los élitros de los coleópteros o los opérculos de los
caracoles, que permanecen mucho tiempo en el estómago. En
ocasiones no es posible conocer el número de presas a partir de
los restos encontrados en el estómago, como sucede cuando se
encuentran pelos sin ningún otro tipo de fragmentos del mamífero a
que pertenecían. Esto puede provocar una subestimación del
número de individuos consumidos. Si se toma en consideración sólo
el número de presas capturadas se sobreestima la importancia de
animales pequeños, que suelen ser consumidos en grandes canti-
dades y, por lo tanto, se subestima la relevancia de presas grandes
que sólo aparecen ocasionalmente. Por último, el volumen medido de
algún tipo de renglón puede ser engañoso, particularmente si la presa
fue comida mucho tiempo antes de la extracción y ha sido digerida en
su casi totalidad. En este caso, se produciría una subestimación de su
importancia. Para corregir el sesgo, en ocasiones se puede estimar el
tamaño (y volumen) original de la presa al momento de ser atrapada
por el crocodilio a partir de los fragmentos encontrados en el
estómago (Staton y Dixon, 1975; Webb et al., 1983).
Los datos que usualmente se toman en cualquier estudio de
dieta de Crocodilios son:
o Frecuencia de aparición de un renglón determinado, es
decir, el porcentaje (o proporción) de estómagos analizados
con el tipo de presa en consideración.
o Número de individuos de cada presa en particular, tanto por
estómago analizado como en el total de crocodilios
estudiados.
o Volumen (por estómago y total) de cada uno de las presas
encontradas.
o Peso seco. En este caso se extrae la humedad en una es-
tufa (55-60 ºC). Este procedimiento es necesario si se quiere
determinar el contenido calórico de las presas consumidas.

Los Crocodylia de Venezuela
165
o Es usual encontrar en los contenidos estomacales renglones
considerados “no alimenticios”. Es el caso de los gastrolitos
(piedras), restos vegetales y parásitos del tracto digestivo,
especialmente nematodos. Éstos también se cuantifican.
Debido a la amplia variedad de especies que en ocasiones apa-
recen en los contenidos estomacales, algunas de ellas representadas
por muy pocos individuos, es conveniente discriminarlas por cate-
gorías. Es usual agruparlas de acuerdo a criterios taxonómicos
(artrópodos, moluscos, peces, mamíferos, por ejemplo). Pero este
tipo de categorización es muy artificial y aporta poca información
sobre los hábitos alimentarios del crocodilio estudiado, ya que
dentro de una misma categoría taxonómica puede haber especies
que ocupan diversos microhábitats en los ambientes terrestres o
acuáticos. Una primera alternativa de categorización es el de
segregar a las presas de acuerdo con el tipo de ambiente en que
se encuentran, por ejemplo en acuáticas o terrestres. Esto nos
permite inferir con qué frecuencia una determinada especie se
alimenta en estos ambientes. Otras categorizaciones son posibles,
como por ejemplo agrupar las presas de acuerdo por el tipo de
microhábitat que ocupan (habitantes de aguas abiertas, organismos
frecuentes en la vegetación acuática, organismos anfibios). En todo
caso, son los objetivos que se buscan con la investigación los que
determinan el tipo de categorización más adecuada.
Por lo general, los investigadores toman sólo algunos de los
datos señalados anteriormente. Para contrarrestar los sesgos que se
producen se han usado índices de abundancia relativa, que toman en
consideración tanto el número total de individuos de cada presa en
toda la muestra como su frecuencia de aparición (Staton y Dixon,
1975). Asimismo, se han asignado rangos de importancia a cada presa
de acuerdo con su tamaño (Fitzgerald, 1988a; Thorbjarnarson, 1990).
No existe una metodología perfecta y correcta para todas las
situaciones. Cada investigador debe estudiar con detalle los distintos
procedimientos y conocer sus limitaciones. Seguramente, todavía
queda campo para innovar y proceder de acuerdo con las nece-
sidades y objetivos de las investigaciones que se planteen.

Andrés Eloy Seijas
166
ANÁLISIS COMPARATIVO DE DIETAS DE
CROCODILIOS
A partir del trabajo pionero de Cott (1961), en numerosos
estudios se ha mostrado que los crocodilios son depredadores
oportunistas con un cambio ontogenético de sus dietas (Fig. 6.2).
También se ha señalado que no hay datos que permitan demostrar
una marcada preferencia de los crocodilios por determinadas presas
(Webb et al., 1982), es decir, que la frecuencia y abundancia con
que aparece un renglón alimentario determinado en sus estómagos
es sólo un reflejo de la disponibilidad con que éste se encuentra en
el medio. Por otra parte, algunos investigadores han señalado que
existe una relación entre la morfología bucal y el tipo de dieta de
la cual depende cada una de las especies de crocodílidos
(Iordansky, 1973; Langston, 1973; Ayarzagüena, 1984). Esta
relación establecería límites al “oportunismo” en la captura de las
presas.
No obstante la diversidad de metodologías que se han empleado,
los estudios llevados a cabo en Venezuela permiten comparar la
dieta de poblaciones de la misma especie pero que ocupan
ambientes distintos. Asimismo, existen estudios en los cuales es
posible comparar la dieta de especies distintas que ocupan la
misma localidad. Estas investigaciones ponen a prueba las ase-
veraciones sobre la condición de cazadores oportunistas de los
crocodilios, así como la de que estos animales no muestran
preferencias por presa alguna.
DIETA DE LA MISMA ESPECIE EN DISTINTAS
LOCALIDADES
Baba
Una primera oportunidad de comparación se presenta con los
estudios de Seijas y Ramos (1980) y Ayarzagüena (1983). Esas
investigaciones fueron realizadas casi de manera simultánea, a
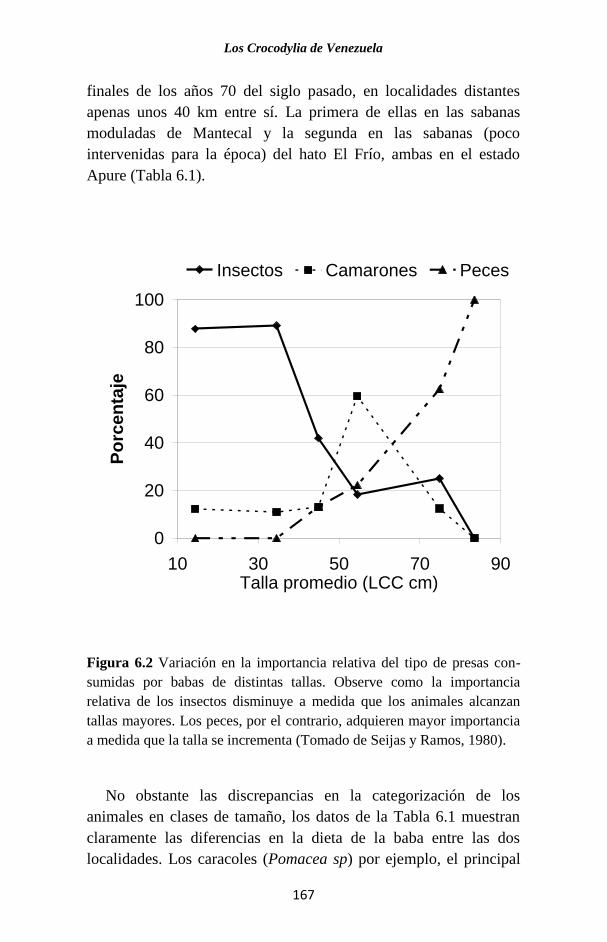
Los Crocodylia de Venezuela
167
finales de los años 70 del siglo pasado, en localidades distantes
apenas unos 40 km entre sí. La primera de ellas en las sabanas
moduladas de Mantecal y la segunda en las sabanas (poco
intervenidas para la época) del hato El Frío, ambas en el estado
Apure (Tabla 6.1).
Figura 6.2 Variación en la importancia relativa del tipo de presas con-
sumidas por babas de distintas tallas. Observe como la importancia
relativa de los insectos disminuye a medida que los animales alcanzan
tallas mayores. Los peces, por el contrario, adquieren mayor importancia
a medida que la talla se incrementa (Tomado de Seijas y Ramos, 1980).
No obstante las discrepancias en la categorización de los
animales en clases de tamaño, los datos de la Tabla 6.1 muestran
claramente las diferencias en la dieta de la baba entre las dos
localidades. Los caracoles (Pomacea sp) por ejemplo, el principal
0
20
40
60
80
100
10 30 50 70 90Talla promedio (LCC cm)
Po
rcen
taje
Insectos Camarones Peces
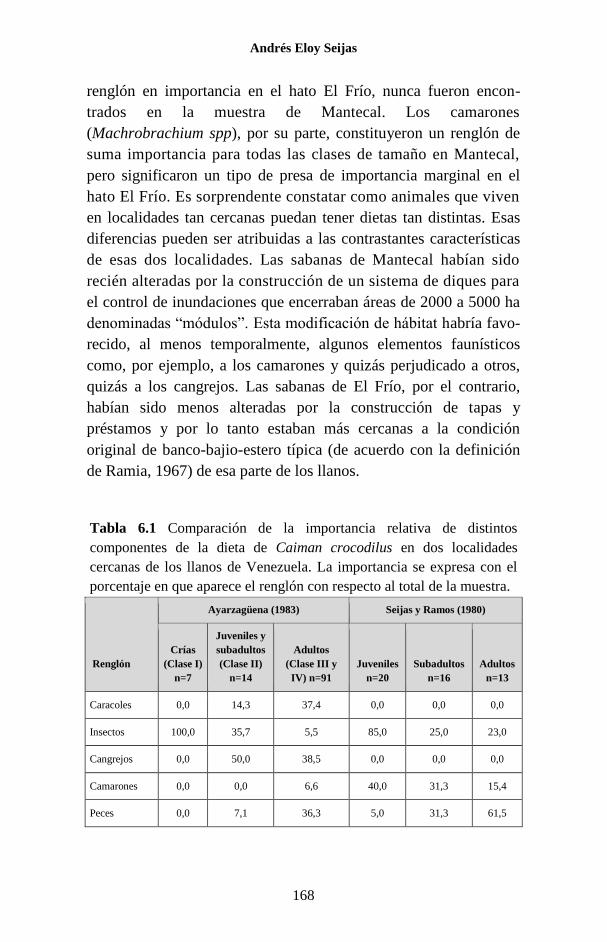
Andrés Eloy Seijas
168
renglón en importancia en el hato El Frío, nunca fueron encon-
trados en la muestra de Mantecal. Los camarones
(Machrobrachium spp), por su parte, constituyeron un renglón de
suma importancia para todas las clases de tamaño en Mantecal,
pero significaron un tipo de presa de importancia marginal en el
hato El Frío. Es sorprendente constatar como animales que viven
en localidades tan cercanas puedan tener dietas tan distintas. Esas
diferencias pueden ser atribuidas a las contrastantes características
de esas dos localidades. Las sabanas de Mantecal habían sido
recién alteradas por la construcción de un sistema de diques para
el control de inundaciones que encerraban áreas de 2000 a 5000 ha
denominadas “módulos”. Esta modificación de hábitat habría favo-
recido, al menos temporalmente, algunos elementos faunísticos
como, por ejemplo, a los camarones y quizás perjudicado a otros,
quizás a los cangrejos. Las sabanas de El Frío, por el contrario,
habían sido menos alteradas por la construcción de tapas y
préstamos y por lo tanto estaban más cercanas a la condición
original de banco-bajio-estero típica (de acuerdo con la definición
de Ramia, 1967) de esa parte de los llanos.
Tabla 6.1 Comparación de la importancia relativa de distintos
componentes de la dieta de Caiman crocodilus en dos localidades
cercanas de los llanos de Venezuela. La importancia se expresa con el
porcentaje en que aparece el renglón con respecto al total de la muestra.
Renglón
Ayarzagüena (1983) Seijas y Ramos (1980)
Crías
(Clase I)
n=7
Juveniles y
subadultos
(Clase II)
n=14
Adultos
(Clase III y
IV) n=91
Juveniles
n=20
Subadultos
n=16
Adultos
n=13
Caracoles 0,0 14,3 37,4 0,0 0,0 0,0
Insectos 100,0 35,7 5,5 85,0 25,0 23,0
Cangrejos 0,0 50,0 38,5 0,0 0,0 0,0
Camarones 0,0 0,0 6,6 40,0 31,3 15,4
Peces 0,0 7,1 36,3 5,0 31,3 61,5

Los Crocodylia de Venezuela
169
En los estudios de la dieta de la baba en los hatos Masaguaral
y Piñero (Staton y Dixon, 1975; Fitzgerald, 1988a; Thorbjarnarson,
1990), en los llanos de Guárico, relativamente más secos que los de
Apure, los camarones estuvieron ausentes o fueron muy escasos.
Los cangrejos, por el contrario, fueron por lo general muy fre-
cuentes. En dichas localidades, los caracoles del género Pomacea
también constituyeron un importante renglón en la dieta, al igual
que en el hato El Frío.
En algunos de los estudios de la dieta de la baba antes
mencionados, los anfibios no parecen tener una relevancia especial.
Pero eso podría ser un indicativo de que el muestreo no se llevó a
cabo en la época en que éstos son más abundantes. En contraste,
en el trabajo de Gorzula (1978) en la Guayana venezolana, las
ranas representaron 66% de las presas consumidas durante la
estación lluviosa.
Con respecto a las babas de la región norte costera de
Venezuela, se pueden comparar los resultados de los estudios
disponibles para localidades en Zulia y Falcón (Tabla 6.2. Seijas,
1988, 1996). Dos componentes importantes en la dieta de C.
crocodilus en el embalse de Pueblo Viejo fueron los cangrejos y
los invertebrados terrestres. En los embalses de Jatira-Tacarigua,
los componentes más importantes fueron los caracoles y los
insectos acuáticos. Estas diferencias se explican por las caracte-
rísticas contrastantes de estas localidades. En Pueblo Viejo, las
aguas eran turbias y carecían por completo de macrofitas acuá-
ticas, condiciones poco propicias para la existencia de caracoles, lo
que explicaría la poca relevancia (tanto en frecuencia de aparición
como en número de individuos presentes) de esta presa en los
estómagos analizados. Las babas de esta localidad parecían depen-
der en gran medida de presas que habitaban la zona litoral (dipló-
podos, ortópteros, cangrejos y pequeños vertebrados terrestres). En
los embalses de Jatira-Tacarigua, las aguas eran claras y las
macrofitas acuáticas muy abundantes, particularmente en Jatira,
siendo Eichhornia sp, Chara sp y Najas sp las especies domi-
nantes. Esto explicaría la mayor importancia de animales acuáticos
(caracoles, invertebrados acuáticos, peces) en estas localidades.
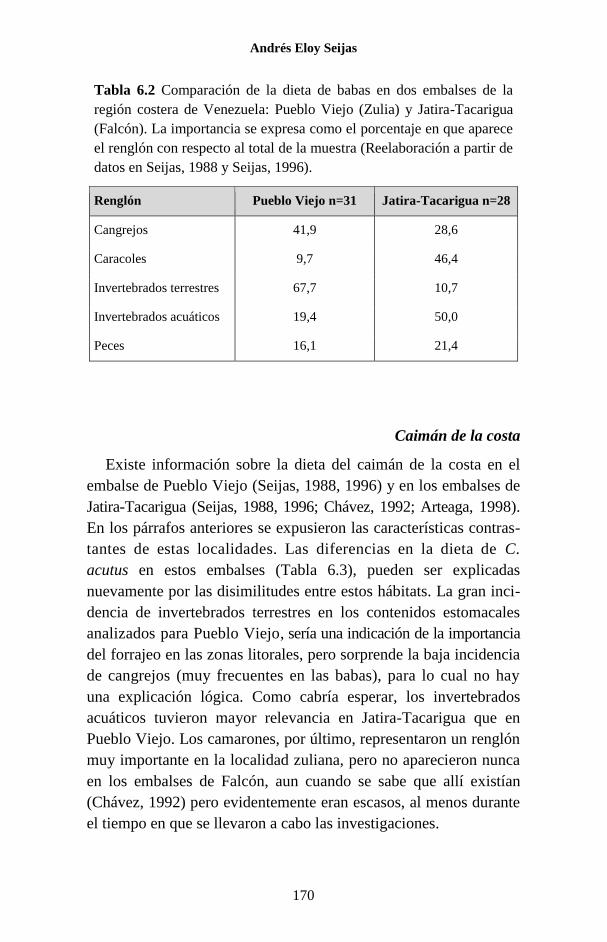
Andrés Eloy Seijas
170
Tabla 6.2 Comparación de la dieta de babas en dos embalses de la
región costera de Venezuela: Pueblo Viejo (Zulia) y Jatira-Tacarigua
(Falcón). La importancia se expresa como el porcentaje en que aparece
el renglón con respecto al total de la muestra (Reelaboración a partir de
datos en Seijas, 1988 y Seijas, 1996).
Renglón Pueblo Viejo n=31 Jatira-Tacarigua n=28
Cangrejos 41,9 28,6
Caracoles 9,7 46,4
Invertebrados terrestres 67,7 10,7
Invertebrados acuáticos 19,4 50,0
Peces 16,1 21,4
Caimán de la costa
Existe información sobre la dieta del caimán de la costa en el
embalse de Pueblo Viejo (Seijas, 1988, 1996) y en los embalses de
Jatira-Tacarigua (Seijas, 1988, 1996; Chávez, 1992; Arteaga, 1998).
En los párrafos anteriores se expusieron las características contras-
tantes de estas localidades. Las diferencias en la dieta de C.
acutus en estos embalses (Tabla 6.3), pueden ser explicadas
nuevamente por las disimilitudes entre estos hábitats. La gran inci-
dencia de invertebrados terrestres en los contenidos estomacales
analizados para Pueblo Viejo, sería una indicación de la importancia
del forrajeo en las zonas litorales, pero sorprende la baja incidencia
de cangrejos (muy frecuentes en las babas), para lo cual no hay
una explicación lógica. Como cabría esperar, los invertebrados
acuáticos tuvieron mayor relevancia en Jatira-Tacarigua que en
Pueblo Viejo. Los camarones, por último, representaron un renglón
muy importante en la localidad zuliana, pero no aparecieron nunca
en los embalses de Falcón, aun cuando se sabe que allí existían
(Chávez, 1992) pero evidentemente eran escasos, al menos durante
el tiempo en que se llevaron a cabo las investigaciones.
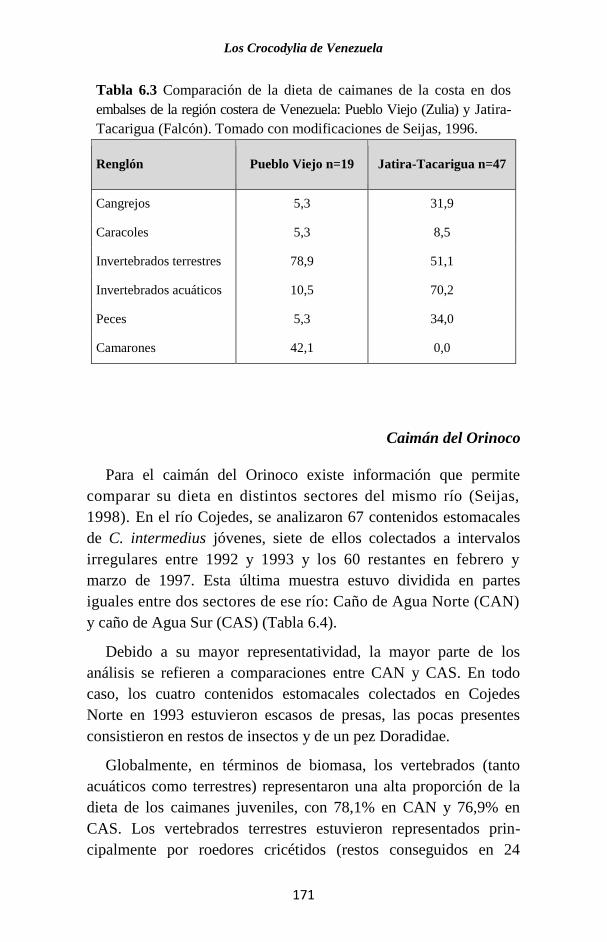
Los Crocodylia de Venezuela
171
Tabla 6.3 Comparación de la dieta de caimanes de la costa en dos
embalses de la región costera de Venezuela: Pueblo Viejo (Zulia) y Jatira-
Tacarigua (Falcón). Tomado con modificaciones de Seijas, 1996.
Renglón Pueblo Viejo n=19 Jatira-Tacarigua n=47
Cangrejos 5,3 31,9
Caracoles 5,3 8,5
Invertebrados terrestres 78,9 51,1
Invertebrados acuáticos 10,5 70,2
Peces 5,3 34,0
Camarones 42,1 0,0
Caimán del Orinoco
Para el caimán del Orinoco existe información que permite
comparar su dieta en distintos sectores del mismo río (Seijas,
1998). En el río Cojedes, se analizaron 67 contenidos estomacales
de C. intermedius jóvenes, siete de ellos colectados a intervalos
irregulares entre 1992 y 1993 y los 60 restantes en febrero y
marzo de 1997. Esta última muestra estuvo dividida en partes
iguales entre dos sectores de ese río: Caño de Agua Norte (CAN)
y caño de Agua Sur (CAS) (Tabla 6.4).
Debido a su mayor representatividad, la mayor parte de los
análisis se refieren a comparaciones entre CAN y CAS. En todo
caso, los cuatro contenidos estomacales colectados en Cojedes
Norte en 1993 estuvieron escasos de presas, las pocas presentes
consistieron en restos de insectos y de un pez Doradidae.
Globalmente, en términos de biomasa, los vertebrados (tanto
acuáticos como terrestres) representaron una alta proporción de la
dieta de los caimanes juveniles, con 78,1% en CAN y 76,9% en
CAS. Los vertebrados terrestres estuvieron representados prin-
cipalmente por roedores cricétidos (restos conseguidos en 24
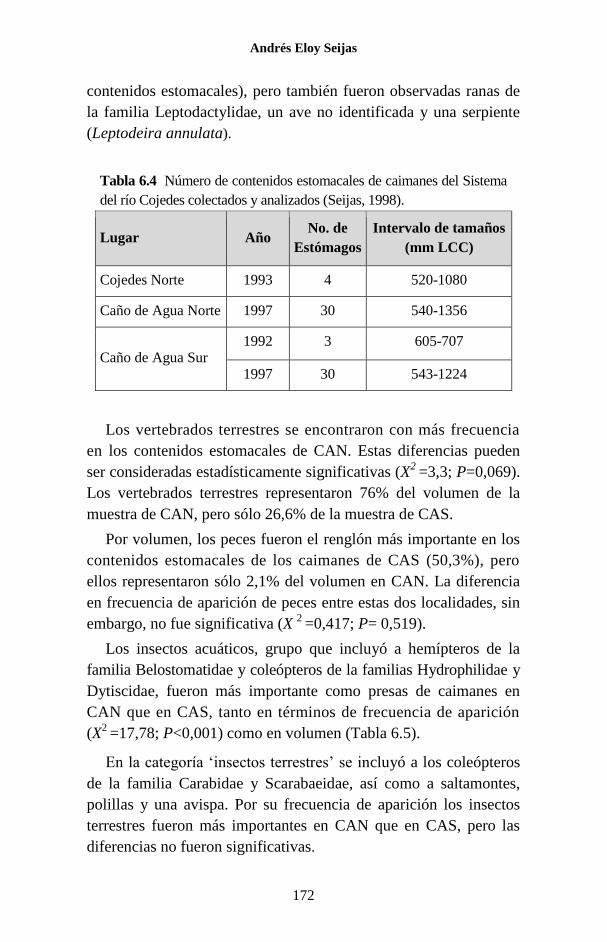
Andrés Eloy Seijas
172
contenidos estomacales), pero también fueron observadas ranas de
la familia Leptodactylidae, un ave no identificada y una serpiente
(Leptodeira annulata).
Tabla 6.4 Número de contenidos estomacales de caimanes del Sistema
del río Cojedes colectados y analizados (Seijas, 1998).
Lugar Año No. de
Estómagos
Intervalo de tamaños
(mm LCC)
Cojedes Norte 1993 4 520-1080
Caño de Agua Norte 1997 30 540-1356
Caño de Agua Sur 1992 3 605-707
1997 30 543-1224
Los vertebrados terrestres se encontraron con más frecuencia
en los contenidos estomacales de CAN. Estas diferencias pueden
ser consideradas estadísticamente significativas (X2
=3,3; P=0,069).
Los vertebrados terrestres representaron 76% del volumen de la
muestra de CAN, pero sólo 26,6% de la muestra de CAS.
Por volumen, los peces fueron el renglón más importante en los
contenidos estomacales de los caimanes de CAS (50,3%), pero
ellos representaron sólo 2,1% del volumen en CAN. La diferencia
en frecuencia de aparición de peces entre estas dos localidades, sin
embargo, no fue significativa (X 2
=0,417; P= 0,519).
Los insectos acuáticos, grupo que incluyó a hemípteros de la
familia Belostomatidae y coleópteros de la familias Hydrophilidae y
Dytiscidae, fueron más importante como presas de caimanes en
CAN que en CAS, tanto en términos de frecuencia de aparición
(X2
=17,78; P<0,001) como en volumen (Tabla 6.5).
En la categoría ‘insectos terrestres’ se incluyó a los coleópteros
de la familia Carabidae y Scarabaeidae, así como a saltamontes,
polillas y una avispa. Por su frecuencia de aparición los insectos
terrestres fueron más importantes en CAN que en CAS, pero las
diferencias no fueron significativas.
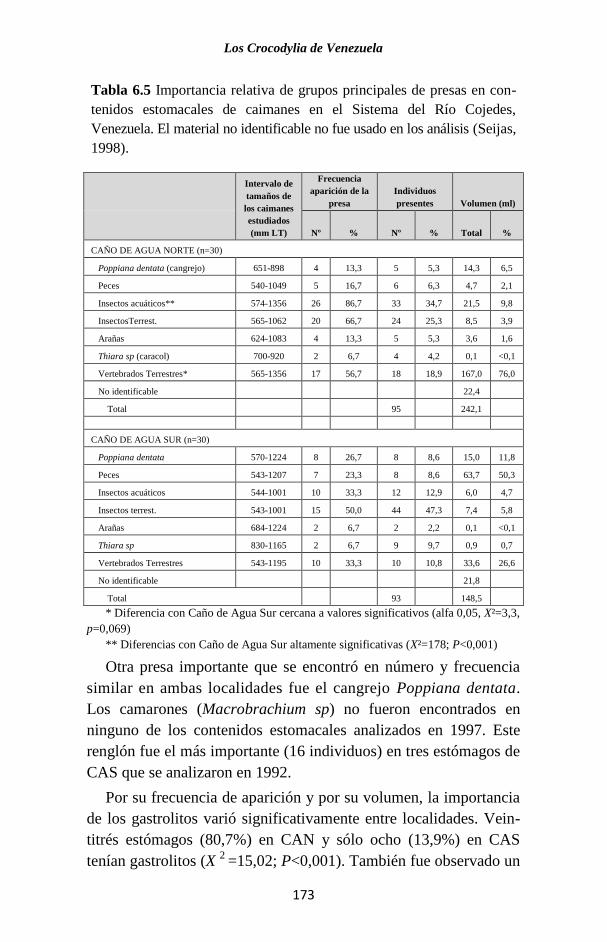
Los Crocodylia de Venezuela
173
Tabla 6.5 Importancia relativa de grupos principales de presas en con-
tenidos estomacales de caimanes en el Sistema del Río Cojedes,
Venezuela. El material no identificable no fue usado en los análisis (Seijas,
1998).
Intervalo de
tamaños de
los caimanes
estudiados
(mm LT)
Frecuencia
aparición de la
presa
Individuos
presentes Volumen (ml)
Nº % Nº % Total %
CAÑO DE AGUA NORTE (n=30)
Poppiana dentata (cangrejo) 651-898 4 13,3 5 5,3 14,3 6,5
Peces 540-1049 5 16,7 6 6,3 4,7 2,1
Insectos acuáticos** 574-1356 26 86,7 33 34,7 21,5 9,8
InsectosTerrest. 565-1062 20 66,7 24 25,3 8,5 3,9
Arañas 624-1083 4 13,3 5 5,3 3,6 1,6
Thiara sp (caracol) 700-920 2 6,7 4 4,2 0,1 <0,1
Vertebrados Terrestres* 565-1356 17 56,7 18 18,9 167,0 76,0
No identificable 22,4
Total 95 242,1
CAÑO DE AGUA SUR (n=30)
Poppiana dentata 570-1224 8 26,7 8 8,6 15,0 11,8
Peces 543-1207 7 23,3 8 8,6 63,7 50,3
Insectos acuáticos 544-1001 10 33,3 12 12,9 6,0 4,7
Insectos terrest. 543-1001 15 50,0 44 47,3 7,4 5,8
Arañas 684-1224 2 6,7 2 2,2 0,1 <0,1
Thiara sp 830-1165 2 6,7 9 9,7 0,9 0,7
Vertebrados Terrestres 543-1195 10 33,3 10 10,8 33,6 26,6
No identificable 21,8
Total 93 148,5
* Diferencia con Caño de Agua Sur cercana a valores significativos (alfa 0,05, X²=3,3,
p=0,069)
** Diferencias con Caño de Agua Sur altamente significativas (X²=178; P<0,001)
Otra presa importante que se encontró en número y frecuencia
similar en ambas localidades fue el cangrejo Poppiana dentata.
Los camarones (Macrobrachium sp) no fueron encontrados en
ninguno de los contenidos estomacales analizados en 1997. Este
renglón fue el más importante (16 individuos) en tres estómagos de
CAS que se analizaron en 1992.
Por su frecuencia de aparición y por su volumen, la importancia
de los gastrolitos varió significativamente entre localidades. Vein-
titrés estómagos (80,7%) en CAN y sólo ocho (13,9%) en CAS
tenían gastrolitos (X 2
=15,02; P<0,001). También fue observado un

Andrés Eloy Seijas
174
pequeño número de conchas del caracol Thiara sp, quizá ingeridas
junto a los gastrolitos.
El río Cojedes está altamente contaminado lo cual quedó de
manifiesto con algunos de los restos encontrados en algunos estó-
magos: un pequeño pedazo de tela en un contenido estomacal de
CAS, restos de material plástico en otro de CAN y partes de un
guante de goma localizados en un estomago de Cojedes Norte.
Otro renglón que es importante mencionar es el de los nemá-
todos. Estos fueron mucho más abundantes y frecuentes (X 2
=8,3;
P<0,01) en los contenidos estomacales de CAS, en comparación
con aquellos de CAN.
Con la excepción de peces y cangrejos, todos los otros renglo-
nes alimentarios se encontraron con mayor frecuencia y abundancia
en los contenidos estomacales de los caimanes de CAN que en los
de CAS. En el caso de los insectos y vertebrados terrestres, esas
diferencias fueron estadísticamente significativas.
La importancia relativa de las presas de origen terrestre varió
entre localidades, seguramente un reflejo de las características
contrastantes entre estos segmentos de río y de sus alrededores.
En este sentido, la mayor importancia relativa de presas de origen
terrestres (particularmente vertebrados) en CAN, se podría explicar
por el hecho de que en este sector del Sistema del Río Cojedes (al
contrario de lo que ocurre en CAS donde las orillas están casi
siempre desnudas) los bordes del río están profusamente cubiertos
de vegetación graminosa y pequeños arbustos, lo cual probable-
mente representa un mejor hábitat para insectos y otros inverte-
brados, los que a su vez atraen a pequeños vertebrados (por ejem-
plo, ranas).
El comportamiento tanto de los caimanes como de sus presas, y
las características del río en cada lugar, deben jugar un papel de
importancia, determinando que es accesible y que no lo es.

Los Crocodylia de Venezuela
175
DIETA DE DISTINTAS ESPECIES EN LA MISMA
LOCALIDAD
La baba y el caimán de la costa
Un análisis más interesante, por sus implicaciones ecológicas, es el
que se produce al comparar la dieta de dos especies de crocodilios
coexistiendo en la misma localidad, es decir, en sintopía. Ésto se llevó
a cabo con las babas y caimanes tanto en el embalse de Pueblo Viejo,
como en los embalses de Jatira-Tacarigua, en Falcón (Seijas, 1988,
1996). En las tablas 6.6 y 6.7 se detallan la frecuencia y abundancia
(en volumen y en número de individuos) en que las distintas presas
fueron encontradas en los crocodilios de esas localidades.
Para facilitar las comparaciones, los contenidos estomacales de
crocodilios de tallas similares se analizaron en conjunto. Los
renglones de la dieta fueron agrupados en amplias categorías, a
saber: invertebrados acuáticos, invertebrados terrestres, arañas,
caracoles (Pomacea sp), cangrejos, camarones (no encontrados en
Tacarigua-Jatira), peces, y otros vertebrados (Tabla 6.8).
Los invertebrados terrestres (T) incluyen varias especies del orden
Coleoptera, Orthoptera y Diplopoda. Los invertebrados acuáticos (A)
incluyen a insectos de las familias Hydrophilidae (Coleoptera) y
Belostomatidae (Hemiptera) y larvas del orden Odonata. En Verte-
brados están incluidos todos los vertebrados con excepción de peces.
Los invertebrados acuáticos, distintos a camarones o cangrejos,
fueron escasos e infrecuentes tanto en babas como en caimanes, lo
cual quizá refleje la poca abundancia de esos animales en un
embalse donde la vegetación acuática estaba prácticamente ausen-
te. Esto también explicaría la escasez de aparición de caracoles,
incluso en los estómagos de las babas.
Los cangrejos sólo aparecieron en estómagos de crocodilios
mayores a 300 cm LCC. Si se considera sólo estas tallas, la
frecuencia de aparición en babas (13 estómagos) resultó estadís-
ticamente distinta a la encontrada en los caimanes (1 estómago)
(Prueba Exacta de Fisher, PEF, P = 0,012).
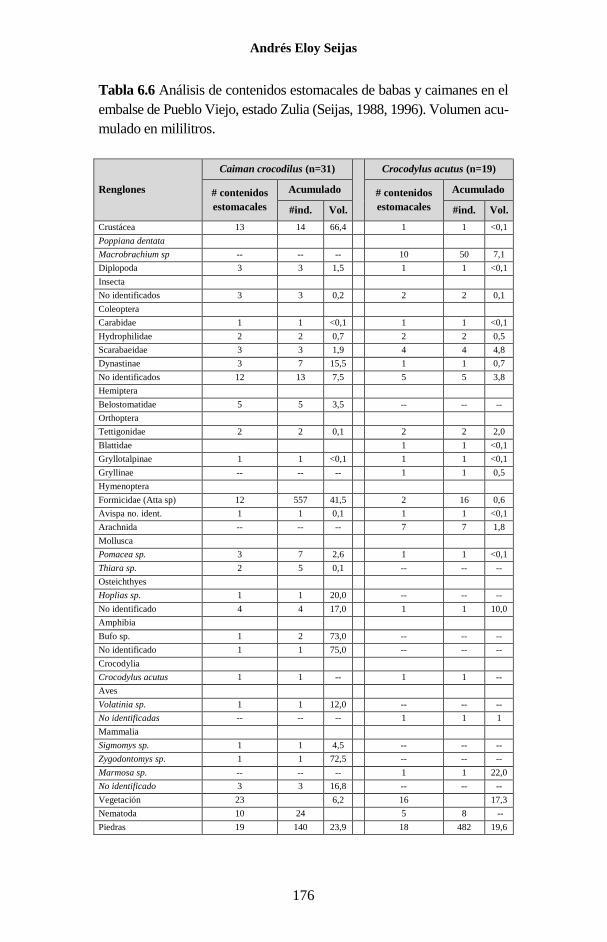
Andrés Eloy Seijas
176
Tabla 6.6 Análisis de contenidos estomacales de babas y caimanes en el
embalse de Pueblo Viejo, estado Zulia (Seijas, 1988, 1996). Volumen acu-
mulado en mililitros.
Renglones
Caiman crocodilus (n=31)
Crocodylus acutus (n=19)
# contenidos
estomacales
Acumulado # contenidos
estomacales
Acumulado
#ind. Vol. #ind. Vol.
Crustácea 13 14 66,4 1 1 <0,1
Poppiana dentata
Macrobrachium sp -- -- -- 10 50 7,1
Diplopoda 3 3 1,5 1 1 <0,1
Insecta
No identificados 3 3 0,2 2 2 0,1
Coleoptera
Carabidae 1 1 <0,1 1 1 <0,1
Hydrophilidae 2 2 0,7 2 2 0,5
Scarabaeidae 3 3 1,9 4 4 4,8
Dynastinae 3 7 15,5 1 1 0,7
No identificados 12 13 7,5 5 5 3,8
Hemiptera
Belostomatidae 5 5 3,5 -- -- --
Orthoptera
Tettigonidae 2 2 0,1 2 2 2,0
Blattidae 1 1 <0,1
Gryllotalpinae 1 1 <0,1 1 1 <0,1
Gryllinae -- -- -- 1 1 0,5
Hymenoptera
Formicidae (Atta sp) 12 557 41,5 2 16 0,6
Avispa no. ident. 1 1 0,1 1 1 <0,1
Arachnida -- -- -- 7 7 1,8
Mollusca
Pomacea sp. 3 7 2,6 1 1 <0,1
Thiara sp. 2 5 0,1 -- -- --
Osteichthyes
Hoplias sp. 1 1 20,0 -- -- --
No identificado 4 4 17,0 1 1 10,0
Amphibia
Bufo sp. 1 2 73,0 -- -- --
No identificado 1 1 75,0 -- -- --
Crocodylia
Crocodylus acutus 1 1 -- 1 1 --
Aves
Volatinia sp. 1 1 12,0 -- -- --
No identificadas -- -- -- 1 1 1
Mammalia
Sigmomys sp. 1 1 4,5 -- -- --
Zygodontomys sp. 1 1 72,5 -- -- --
Marmosa sp. -- -- -- 1 1 22,0
No identificado 3 3 16,8 -- -- --
Vegetación 23 6,2 16 17,3
Nematoda 10 24 5 8 --
Piedras 19 140 23,9 18 482 19,6
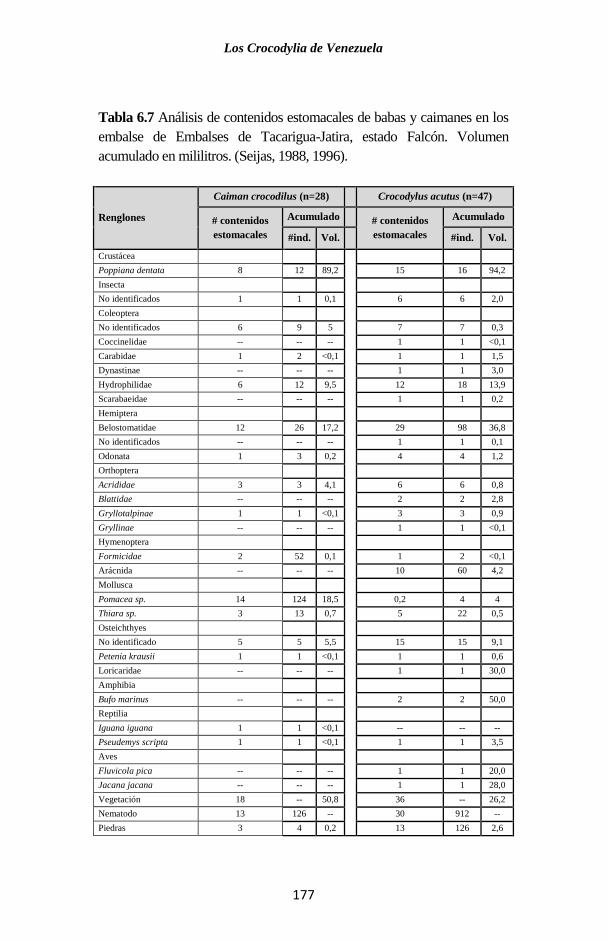
Los Crocodylia de Venezuela
177
Tabla 6.7 Análisis de contenidos estomacales de babas y caimanes en los
embalse de Embalses de Tacarigua-Jatira, estado Falcón. Volumen
acumulado en mililitros. (Seijas, 1988, 1996).
Renglones
Caiman crocodilus (n=28) Crocodylus acutus (n=47)
# contenidos
estomacales
Acumulado # contenidos
estomacales
Acumulado
#ind. Vol. #ind. Vol.
Crustácea
Poppiana dentata 8 12 89,2 15 16 94,2
Insecta
No identificados 1 1 0,1 6 6 2,0
Coleoptera
No identificados 6 9 5 7 7 0,3
Coccinelidae -- -- -- 1 1 <0,1
Carabidae 1 2 <0,1 1 1 1,5
Dynastinae -- -- -- 1 1 3,0
Hydrophilidae 6 12 9,5 12 18 13,9
Scarabaeidae -- -- -- 1 1 0,2
Hemiptera
Belostomatidae 12 26 17,2 29 98 36,8
No identificados -- -- -- 1 1 0,1
Odonata 1 3 0,2 4 4 1,2
Orthoptera
Acrididae 3 3 4,1 6 6 0,8
Blattidae -- -- -- 2 2 2,8
Gryllotalpinae 1 1 <0,1 3 3 0,9
Gryllinae -- -- -- 1 1 <0,1
Hymenoptera
Formicidae 2 52 0,1 1 2 <0,1
Arácnida -- -- -- 10 60 4,2
Mollusca
Pomacea sp. 14 124 18,5 0,2 4 4
Thiara sp. 3 13 0,7 5 22 0,5
Osteichthyes
No identificado 5 5 5,5 15 15 9,1
Petenia krausii 1 1 <0,1 1 1 0,6
Loricaridae -- -- -- 1 1 30,0
Amphibia
Bufo marinus -- -- -- 2 2 50,0
Reptilia
Iguana iguana 1 1 <0,1 -- -- --
Pseudemys scripta 1 1 <0,1 1 1 3,5
Aves
Fluvicola pica -- -- -- 1 1 20,0
Jacana jacana -- -- -- 1 1 28,0
Vegetación 18 -- 50,8 36 -- 26,2
Nematodo 13 126 -- 30 912 --
Piedras 3 4 0,2 13 126 2,6
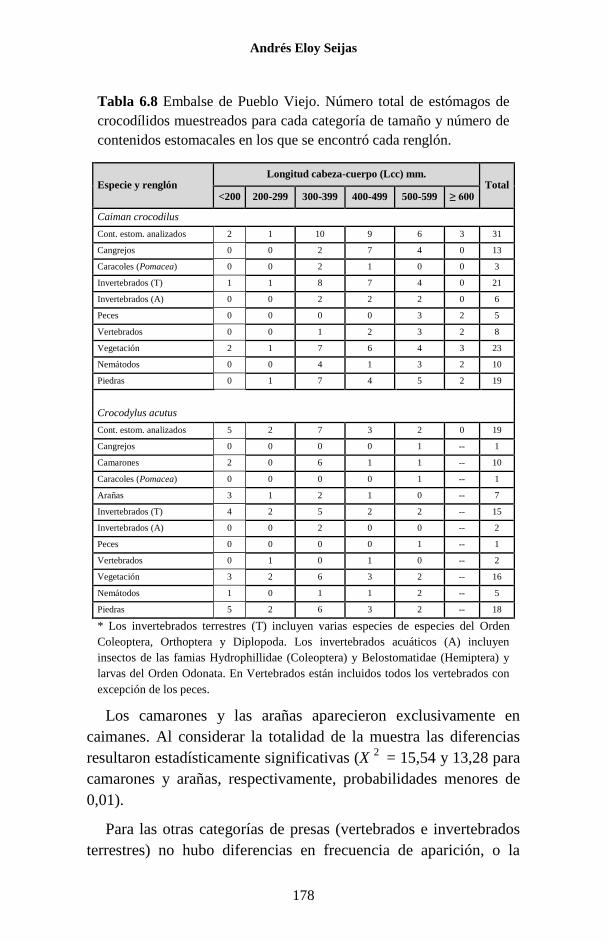
Andrés Eloy Seijas
178
Tabla 6.8 Embalse de Pueblo Viejo. Número total de estómagos de
crocodílidos muestreados para cada categoría de tamaño y número de
contenidos estomacales en los que se encontró cada renglón.
Especie y renglón Longitud cabeza-cuerpo (Lcc) mm.
Total <200 200-299 300-399 400-499 500-599 ≥ 600
Caiman crocodilus
Cont. estom. analizados 2 1 10 9 6 3 31
Cangrejos 0 0 2 7 4 0 13
Caracoles (Pomacea) 0 0 2 1 0 0 3
Invertebrados (T) 1 1 8 7 4 0 21
Invertebrados (A) 0 0 2 2 2 0 6
Peces 0 0 0 0 3 2 5
Vertebrados 0 0 1 2 3 2 8
Vegetación 2 1 7 6 4 3 23
Nemátodos 0 0 4 1 3 2 10
Piedras 0 1 7 4 5 2 19
Crocodylus acutus
Cont. estom. analizados 5 2 7 3 2 0 19
Cangrejos 0 0 0 0 1 -- 1
Camarones 2 0 6 1 1 -- 10
Caracoles (Pomacea) 0 0 0 0 1 -- 1
Arañas 3 1 2 1 0 -- 7
Invertebrados (T) 4 2 5 2 2 -- 15
Invertebrados (A) 0 0 2 0 0 -- 2
Peces 0 0 0 0 1 -- 1
Vertebrados 0 1 0 1 0 -- 2
Vegetación 3 2 6 3 2 -- 16
Nemátodos 1 0 1 1 2 -- 5
Piedras 5 2 6 3 2 -- 18
* Los invertebrados terrestres (T) incluyen varias especies de especies del Orden
Coleoptera, Orthoptera y Diplopoda. Los invertebrados acuáticos (A) incluyen
insectos de las famias Hydrophillidae (Coleoptera) y Belostomatidae (Hemiptera) y
larvas del Orden Odonata. En Vertebrados están incluidos todos los vertebrados con
excepción de los peces.
Los camarones y las arañas aparecieron exclusivamente en
caimanes. Al considerar la totalidad de la muestra las diferencias
resultaron estadísticamente significativas (X 2
= 15,54 y 13,28 para
camarones y arañas, respectivamente, probabilidades menores de
0,01).
Para las otras categorías de presas (vertebrados e invertebrados
terrestres) no hubo diferencias en frecuencia de aparición, o la

Los Crocodylia de Venezuela
179
muestra resultó muy pequeña para realizar comparaciones esta-
dísticas. Como dato anecdótico hay que resaltar que entre el 16 y
el 20 de mayo de 1987 aparecieron abundantes reinas y zánganos
de hormigas del género Atta en los contenidos estomacales
muestreados. Dos babas tenían cada una no menos de 250 de
estos insectos en sus estómagos. Debido a la presencia efímera
(durante el vuelo nupcial al comienzo de las lluvias) y al hecho de
que la muestra de esos días estuvo muy desviada a favor de las
babas (22 estómagos de babas contra sólo 6 de caimanes) estas
hormigas no se tomaron en cuenta en los análisis.
Vale la pena destacar que en los análisis de los contenidos
estomacales en Pueblo Viejo se obtuvo evidencias tanto de
depredación interespecífica como de canibalismo. En una opor-
tunidad se consiguió dentro del estómago de un caimán juvenil (372
mm de LCC) una placa metálica numerada, que había sido
colocada a una cría de C. acutus. Asimismo se consiguieron dos
de esas placas, con las que se habían marcado otras crías de
caimán, en el estómago de una baba (432 mm de LCC). Todas las
crías habían sido marcadas un mes antes.
En Jatira-Tacarigua los invertebrados acuáticos se encontraron
con mayor frecuencia en caimanes que en babas para tres de las
cinco categorías de tamaño consideradas, aunque sólo para la
categoría 200-299 mm Lcc, la diferencia fue estadísticamente
significativa (PEF, P < 0,018). Estos invertebrados también se
encontraron con mayor abundancia en caimanes que en babas. No
hubo diferencias significativas en la frecuencia de aparición de
invertebrados terrestres entre estos crocodilios (Tabla 6.9). Sólo se
encontraron peces en crocodilios mayores de 200 mm de LCC. La
ocurrencia de peces solamente se puede considerar mayor en
caimanes que en babas, para animales de más de 600 mm de Lcc
(PEF, P = 0,063).
Tanto para la baba como para el caimán, la frecuencia de
aparición de cangrejos aumentó con la talla. En los animales
mayores de 600 mm de LCC, la frecuencia de aparición de
cangrejos fue mayor para los caimanes que para las babas (PEF, P

Andrés Eloy Seijas
180
= 0,034). Las arañas solamente se encontraron en los estómagos
de los caimanes. Estos invertebrados fueron particularmente comu-
nes en animales menores de 300 mm de Lcc. Para estas tallas, la
diferencia en frecuencia de aparición entre babas y caimanes resultó
estadísticamente significativa (X 2
= 5,27; P = 0,021). Las arañas no
sólo resultaron comunes, sino en ocasiones muy abundantes. Dos
contenidos estomacales mostraron 21 ejemplares cada uno.
Los caracoles del género Pomacea aparecieron con mucho más
frecuencia y abundancia en las babas que en los caimanes. Se
encontró un solo opérculo de caracol en cada uno de los cuatro
contenidos estomacales de caimanes en donde esta presa estuvo
presente; en contraste, el número acumulado de opérculos de Poma-
cea en los 14 estómagos de babas con este renglón alimentario alcanzó la
cifra de 124 (intervalo 1-50), diferencias que son estadísticamente
significativas (t = 2,23; P = 0,044). Para otros renglones alimentarios la
muestra fue muy pequeña para intentar análisis estadísticos.
Las diferencias parciales en la dieta de babas y caimanes son
probablemente consecuencia de formas distintas de usar el hábitat
por estas especies. El caimán usa una franja más ancha desde el
borde del agua y se encuentra con más frecuencia en aguas
abiertas. Las babas, por su parte, se localizan por lo general en
aguas someras y más cerca de la orilla y se adentran un poco en
el ambiente terrestre que bordea a los cuerpos de agua. El
contraste en el uso del hábitat entre babas y caimanes es menos
evidente en localidades estructuralmente complejas como los embal-
ses de Jatira-Tacarigua, lugares donde la vegetación acuática es
muy abundante, que en otra más simple como el embalse de
Pueblo Viejo, donde la vegetación acuática estaba ausente para el
momento en que se realizó el estudio. En Jatira-Tacarigua la
vegetación acuática parece separar físicamente a las dos especies,
mientras que en Pueblo Viejo el hábitat ofrece menos posibilidades
para esa separación. No obstante, en la represa de Pueblo Viejo,
las babas fueron localizadas predominantemente en ensenadas,
bocas de caño y otros lugares estrechos, mientras que C. acutus
se encontró con mayor frecuencia en orillas abiertas hacia el
cuerpo principal del embalse (Seijas, 1988). El tipo de micro-
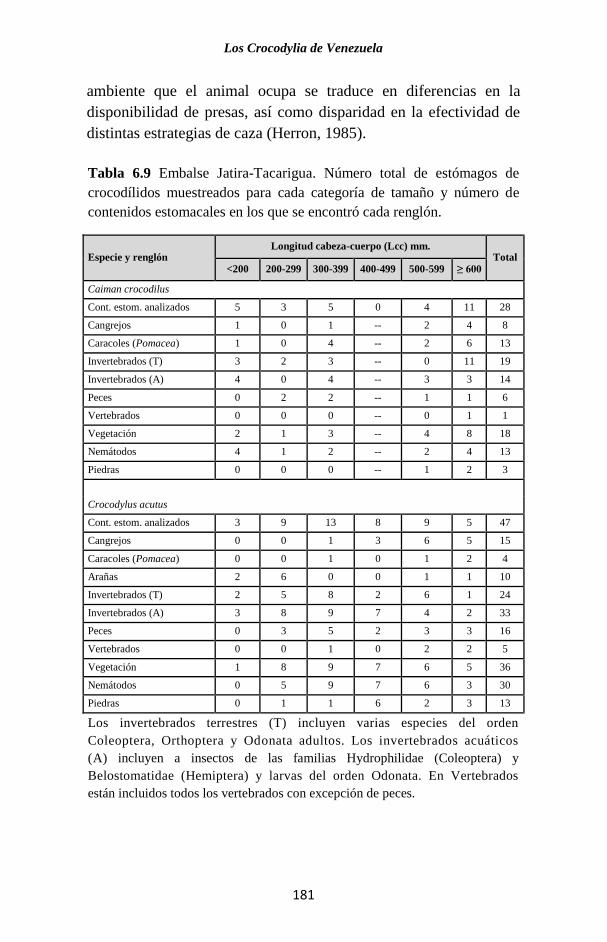
Los Crocodylia de Venezuela
181
ambiente que el animal ocupa se traduce en diferencias en la
disponibilidad de presas, así como disparidad en la efectividad de
distintas estrategias de caza (Herron, 1985).
Tabla 6.9 Embalse Jatira-Tacarigua. Número total de estómagos de
crocodílidos muestreados para cada categoría de tamaño y número de
contenidos estomacales en los que se encontró cada renglón.
Especie y renglón Longitud cabeza-cuerpo (Lcc) mm.
Total <200 200-299 300-399 400-499 500-599 ≥ 600
Caiman crocodilus
Cont. estom. analizados 5 3 5 0 4 11 28
Cangrejos 1 0 1 -- 2 4 8
Caracoles (Pomacea) 1 0 4 -- 2 6 13
Invertebrados (T) 3 2 3 -- 0 11 19
Invertebrados (A) 4 0 4 -- 3 3 14
Peces 0 2 2 -- 1 1 6
Vertebrados 0 0 0 -- 0 1 1
Vegetación 2 1 3 -- 4 8 18
Nemátodos 4 1 2 -- 2 4 13
Piedras 0 0 0 -- 1 2 3
Crocodylus acutus
Cont. estom. analizados 3 9 13 8 9 5 47
Cangrejos 0 0 1 3 6 5 15
Caracoles (Pomacea) 0 0 1 0 1 2 4
Arañas 2 6 0 0 1 1 10
Invertebrados (T) 2 5 8 2 6 1 24
Invertebrados (A) 3 8 9 7 4 2 33
Peces 0 3 5 2 3 3 16
Vertebrados 0 0 1 0 2 2 5
Vegetación 1 8 9 7 6 5 36
Nemátodos 0 5 9 7 6 3 30
Piedras 0 1 1 6 2 3 13
Los invertebrados terrestres (T) incluyen varias especies del orden
Coleoptera, Orthoptera y Odonata adultos. Los invertebrados acuáticos
(A) incluyen a insectos de las familias Hydrophilidae (Coleoptera) y
Belostomatidae (Hemiptera) y larvas del orden Odonata. En Vertebrados
están incluidos todos los vertebrados con excepción de peces.

Andrés Eloy Seijas
182
El comportamiento para la búsqueda y captura del alimento por
parte de los crocodílidos ha sido poco estudiado. La mayor parte
de los investigadores señalan que estos reptiles son oportunistas y
que no hay datos que muestren una marcada preferencia por
determinadas presas (Webb et al., 1982) y que la frecuencia y
abundancia con que aparece un renglón determinado es sólo un
reflejo de la abundancia de éste en el medio. De acuerdo con esto,
si consideramos exclusivamente los contenidos estomacales de
babas, deberíamos concluir que las arañas son escasas en las
localidades estudiadas. Mientras que si nos limitáramos a analizar
los contenidos estomacales de los caimanes llegaríamos a una
conclusión opuesta. Asimismo, si tratáramos de inferir la abundan-
cia de camarones o caracoles a partir de los análisis de los
contenidos estomacales, se llegaría a conclusiones contradictorias
dependiendo de si se trabaja con babas o con caimanes. Más
ajustada parece la conclusión de Taylor (1979, aun cuando ella se
refería a la importancia de ingestión secundaria de ciertos renglones
en C. porosus) de que la composición de la dieta es el resultado
tanto del comportamiento o modo de captura de las presas como
de las oportunidades de alimentación que le ofrece el hábitat.
Las discrepancias en la dieta entre babas y caimanes (tanto del
Orinoco como de la costa) probablemente se basan más en dife-
rencias en conducta de forrajeo entre estas especies que en distinta
disponibilidad de alimento para cada una de ellas. Se puede señalar
que, en términos relativos C. acutus es un cazador más activo que
C. crocodilus (Seijas, 1988). Esta última especie tiene como
estrategia principal la caza al acecho (Fitzgerald, 1988a; Seijas,
1988), aunque esto no quiere decir que en ciertas circunstancias las
babas no sean capaces de utilizar otros tipos de estrategias de
caza, como por ejemplo perseguir a sus presas (Medem, 1981;
Ayarzagüena, 1983; Thorbjarnarson, 1993). De acuerdo con Schaller y
Crawshaw (1982), los yacaré (Caiman crocodilus yacare) del
Pantanal en Brasil, acechan a la presa en vez de perseguirla.
En la región costera de Venezuela la baba parece permanecer
más tiempo en tierra que el caimán, al menos de noche cuando las
observaciones han sido realizadas (Seijas, 1988). Por esta razón

Los Crocodylia de Venezuela
183
sería de esperar encontrar más presas de hábitos terrestres en
babas que en caimanes. Los datos apoyan esta observación,
aunque las diferencias no fueron estadísticamente significativas. Por
otra parte, la presencia de presas duras como cangrejos y
caracoles se esperaba que fuera mayor en babas que en caimanes,
ya que en teoría (Iordansky, 1973; Ayarzagüena, 1984; Pooley,
1989) las especies de hocico ancho (como la baba) están mejor
adaptadas para la captura de ese tipo de presas. Los datos para la
región costera venezolana (Seijas, 1988) demuestran, sin duda, que
los caracoles son consumidos con más frecuencia y abundancia por
las babas. Los cangrejos aparecieron con mayor frecuencia en
babas que en caimanes en el embalse de Pueblo Viejo, pero la
frecuencia y abundancia de este renglón fueron prácticamente las
mismas en Tacarigua-Jatira. El cangrejo Dilocarcinus dentatus
(=Poppiana dentata) puede ser encontrado dentro o fuera del
agua. En las aguas turbias de Pueblo Viejo, esta presa pudiera ser
más fácilmente cazada en tierra que en agua, lo que explicaría su
relativa mayor importancia como parte de la dieta de las babas.
En las investigaciones sobre la dieta de babas y caimanes de la
costa, no hay datos que demuestren que los recursos alimentarios
fueran, ni siquiera temporalmente, escasos en las localidades
estudiadas, una condición necesaria para la ocurrencia de compe-
tencia por explotación de recursos. Sin embargo, la competencia
por interferencia podría ser importante. Para otras localidades, y
con otras especies de crocodilios, se han documentado, o han sido
sugeridas, interacciones negativas directas entre distintas especies
de crocodílidos (Álvarez del Toro, 1974; Herron, 1985; Webb et
al., 1983; Scott et al., 1990). La dominancia de una especie que
alcanza gran tamaño sobre otra más pequeña podría significar que
la primera impide a la segunda el acceso a determinado recurso,
por ejemplo algún tipo de presa. En este sentido, los grandes
caimanes de la costa podrían excluir a las babas, por medio de
agresión directa, de ambientes con recursos alimentarios más
abundantes y de mejor calidad. Pero al mismo tiempo, una baba
grande puede dominar, en interacciones directas, a un caimán
pequeño. Es difícil demostrar cómo estos tipos de interacciones
negativas pudieran contribuir a determinar la característica de la

Andrés Eloy Seijas
184
dieta de dos especies de crocodilios que viven en sintopía. En el
embalse de Pueblo Viejo se pudo demostrar que las interacciones
entre crocodilios que viven en una misma localidad pueden ser muy
directas, ya que se encontraron evidencias de que una baba de 643
mm de LT se comió a una cría de C. acutus (Seijas, 1988).
En conclusión, se puede afirmar que aunque los crocodilios son
depredadores oportunistas, la disponibilidad de determinada presa no
depende exclusivamente de la abundancia en que ésta se encuentra
en el medio: la complejidad estructural del medio, el tamaño del
depredador, su comportamiento y la forma de su hocico deben
jugar un papel en la delimitación de sus dietas. Es posible que la
dieta de una especie de crocodilio en una localidad determinada se
vea influenciada por la presencia de otra especie del mismo grupo,
posibilidad que vale la pena investigar en estudios futuros.

Los Crocodylia de Venezuela
185
CAPÍTULO 7
LOS CROCODYLIA COMO RECURSO
Los crocodilios forman parte del conjunto de los muchos
animales silvestres que han servido de alimento a los seres
humanos desde tiempos antiquísimos. No existen, sin embargo,
datos precisos que permitan cuantificar dicha importancia antes de
la conquista de las tierras americanas por los europeos. Algunas
referencias aisladas de la época de la colonia, así como otras del
siglo pasado, permiten formarnos una idea del papel que estos
reptiles jugaron en las economías de subsistencia de los pueblos
aborígenes de Venezuela.
Usos tradicionales
El padre Gumilla en su crónica de la primera mitad del siglo
XVIII (citado por Medem, 1981) señaló que los indios Otomacos y
Guamos, por ejemplo, sólo cazaban caimanes para comer cuando la
pesca era difícil o escasa pero que, en cambio, apreciaban mucho
la carne de la “babilla” aun cuando el pescado estuviera disponible.
En su recopilación sobre El Caimán en el folklore venezolano
Cardona (1959) transcribió algunas de las observaciones de José
Luis de Cisneros, escritas en 1764 (Cisneros J. L., 1950.
Descripción exacta de la provincia de Benezuela. Edit. Ávila
Gráfica Caracas), parte de las cuales citaré a continuación:
“Asimismo se crían en todos los ríos de los llanos
innumerables caimanes,… tienen mucha grasa, en especial
en la cola; su carne es muy blanca, aunque algo dura. Los
indios la comen muy bien. … sus colmillos tienen virtud
contra el veneno, y una ochava de polvos de la Verga de

Andrés Eloy Seijas
186
este animal, es especial curación para el pasmo; la manteca
es delgada y purgante; los indios la beben y con ella se
limpian el vientre. … Hay otro animal de la misma figura,
aunque mucho más pequeño que los indios llaman Baba: no
es carnicero, solo se mantiene de peces. Yo he comido su
carne, es muy blanca y gustosa”
Es de suponer que el consumo de huevos, carne y otros pro-
ductos de crocodilios era una actividad común para muchos otros
pueblos que vivían en las tierras bajas donde se encontraban (y
todavía existen) estos animales. En ese sentido, Muñoz (1986)
recoge algunos testimonios de cronistas de la época colonial. Datos
colectados principalmente por antropólogos en tiempos relativamente
recientes permiten cuantificar dicho consumo. Hames (1985), por
ejemplo, comparó la eficiencia de caza de los Yanomami y los
Ye´kwana y reportó que las babas representaron el animal de caza
más importante (30% en peso del total de las capturas) en el
segundo grupo aborigen, pero con sólo una importancia marginal
(2%) en el primer grupo indígena. Estas diferencias las explicaba
por el uso de distintos métodos de caza entre estos pueblos y
porque los Ye´kwana estudiados vivían principalmente a lo largo de
los ríos. Por otra parte, para el momento del estudio usaban armas
de fuego y lanchas con motores fuera de borda, mientras que los
Yanomami eran gente del bosque y dependían principalmente del
arco y la flecha para cazar. Lizot (1979) reportó que las babas
representaban 5% en peso de los animales cazados por los Yano-
mami por él estudiados. Estos reportes parecen sugerir que los
crocodilianos jugaban un papel relativamente pequeño como fuente
de proteínas para las comunidades indígenas previo al contacto con
los colonizadores europeos, al menos si se compara con la impor-
tancia de las tortugas acuáticas, los mamíferos y las aves. Quizá
en apoyo de esta posibilidad se puede mencionar que Roze (1964)
señaló que los indios Piaroas mataron 18 caimanes grandes en una
sola noche durante la temporada de anidación de la tortuga de río
(Podocnemis expansa) y agregó que esa matanza pudo haber
aliviado la presión de estos depredadores sobre las tortugas, a las
cuales los indígenas apreciaban mucho más por ser su fuente de
alimento preferida.

Los Crocodylia de Venezuela
187
El consumo de carne de crocodilios por comunidades rurales no
indígenas tampoco está muy bien documentado. Ojasti et al. (1986)
resumieron información colectada por varios autores en la que
estos animales no aparecen como uno de los renglones cazados por
comunidades criollas.
Los crocodilios han sido cazados tradicionalmente no sólo por su
carne. La grasa, por ejemplo, fue usada como agente fijador en la
industria de perfumes o como combustible (mezclada con kerosén)
para el alumbrado, o como agente curativo de heridas y rasguños
(Medem, 1981, 1983; Godshalk, 1978). A los dientes y huevos
también se le han adjudicado supuestas propiedades curativas y
mágicas (Gumilla, 1963; López-Corcuera, 1984).
Aprovechamiento comercial
Las referencias previas sobre el aprovechamiento de crocodilios
para el consumo de su carne u otros productos no pretenden ser
una recopilación exhaustiva. Con ellas se quiere poner de mani-
fiesto que el uso con fines no comerciales de estas especies
seguramente no causó una merma considerable en sus poblaciones,
en contraste con lo ocurrido con la explotación de sus cueros.
Una antigua referencia sobre el valor comercial de los caimanes
la recogió Tablante-Garrido (1961), al reseñar una monografía del
año 1831 del gobernador Gral José Cornelio Muñoz, de la Provincia
de Apure. En ella el gobernador señalaba:
"de tres años a esta parte, se hace un gran consumo de
la manteca del cayman que se consigue en abundancia. El
atraso de nuestra industria no ha permitido que esta grasa
tan buena como las otras, reciba las preparaciones que son
indispensables para liberarla de la fetidez que exhala. Sin
embargo por un proceder muy sencillo y brusco, con el
auxilio del ácido del limón se obtiene ya manteca del propio
anfibio, que sirve para los alumbrados sin molestia casi del
olfato"

Andrés Eloy Seijas
188
El valor comercial de estas especies también fue destacado más
tarde por Adolfo Ernts, el científico alemán radicado en Venezuela
a finales del siglo XIX, quien en su libro La Exposición Nacional
de Venezuela en 1983 indicaba:
“Pocos ríos y caños hay en el interior de la República
donde no se encuentre una que otra de las diferentes
especies llamadas caimán en el país. Tenemos por lo menos
cinco, sin contar la baba,… y casi todas las hay en número
muy considerable. Siendo animales eminentemente dañinos,
debería hacerse todo lo posible por destruirlos, y esto tanto
más cuando es posible sacar provecho de sus cadáveres. En
los Estados Unidos es muy usado el cuero de Aligator,…
[por lo que] … no sería absurdo no hacer un ensayo
industrial con los cueros de los caimanes en nuestro país.
… Se sabe que [la baba]… es el más pequeño de los
crocodílidos. … [por lo que] no daría mucha ventaja en
cuanto á su cuero; pero su destrucción sería importante
para la piscicultura, porque la baba es uno de los enemigos
más rapaces de los peces.” (Adolfo Ernts. Obras completas.
Compilación de Blas Bruni Celli. Fundación Venezolana para la
Salud y la Educación, Caracas 1983. Pág. 297-298).
Más adelante Ernts se refirió en su libro a otros productos
aprovechables de los caimanes en los siguientes términos:
“La manteca de caimán… es de color amarillento,
semifluida á la temperatura ordinaria y de un olor muy poco
pronunciado. Si realmente es siempre de ese color, creemos
que bien pudiera servir para muchos propósitos industriales,
lo que sería un aliciente más para la caza y la destrucción
de estos reptiles dañinos”.
“No son menos grandes las propiedades medicinales
que el pueblo atribuye a los colmillos de caimán… La
creencia debe ser muy general, puesto que el gran número de
estos dientes presentados en la exposición encontró
rapidísima salida.

Los Crocodylia de Venezuela
189
“Con el nombre de almizcle de caimán había en la
colección formada por la Sociedad Patriótica de San
Fernando una muestra… de glándulas anales de este saurio:
forman pedazos algo irregulares, aunque bastante redondos,
de color grís amarillento, con cavidades interiores, y de un
olor muy fuerte á almizcle. Los pedazos más grandes tienen
3 ó 4 cm. de diámetro. Leemos en la última Revista del
Mercado de Drogas, publicado por Gehe & Ca. Dresden
(Set. 1884) que se ha empezado á exportar el almizcle del
aligator de los Estados Unidos como artículo usado en la
perfumería, y creemos que bien podría hacerse lo mismo
respecto a los caimanes de nuestros ríos”. (Adolfo Ernts.
Obras completas. Compilación de Blas Bruni Celli. Fundación
Venezolana para la Salud y la Educación, Caracas 1983. Pág. 309).
Conocidas las recomendaciones, muy autorizadas para la época,
de Adolfo Ernts, no sorprende, como veremos más adelante, la
saña con que estos caimanes fueron cazados en décadas poste-
riores. La percepción despectiva sobre los crocodilios persistía
todavía en 1933, en pleno apogeo de la cacería con fines comer-
ciales de estos animales. En una breve nota del 7 de julio
aparecida en El Nuevo Diario, de Caracas, firmada por Ceferino
R. Avecilla, ese articulista ironizaba sobre los gustos culinarios de
un Club de Gastrónomos Franceses, que alardeaban de haber
comido “colas de cocodrilos” entre otras extravagancias. Agregaba
Avecilla que
“Los cocodrilos son una de las bestias que tienen menos
derecho a la vida. Hasta ahora no ha podido utilizarse de
ellos más que su piel,…”.
Debido a sus grandes tamaños y la calidad de sus cueros
(denominados “pieles clásicas” en la jerga del comercio de pieles
de crocodilios) las primeras especies explotadas en gran escala
fueron los caimanes (Crocodylus acutus y C. intermedius). Los
intentos iniciales de comercializar las pieles del caimán en Vene-
zuela fueron realizados por una compañía norteamericana, que tenía
su centro de operaciones en El Yagual, estado Apure, en los años
1894-1895 (Verstraeten, 1939, citado por Medem 1983; Calzadilla,

Andrés Eloy Seijas
190
1948). Los caimanes eran cazados con armas de fuego durante el
día, un método ineficiente en el que muchos animales muertos o
heridos se perdían al no poder ser recuperados. André (1904)
señaló que dos hermanos noruegos, de apellido Christianssen, se
dedicaban a la cacería de caimanes hacia 1897. Ellos, o sus em-
pleados, cazaban de noche con la ayuda de reflectores adaptados a
linternas. Entre 60 y 70 animales eran cazados con armas de fuego
en una sola noche. Los cueros salados, que eran luego procesados
para la manufactura de ‘bolsos y maletas durables y refinados’
eran vendidos a un dólar la pieza. Esos primeros intentos no
prosperaron. Los gastos de preparación y transporte de las pieles
fueron muy altos y por lo tanto el trabajo tuvo que ser abandonado
(Mozans, 1910; Calzadilla, 1948).
A pesar de estos intentos iniciales de explotación comercial,
durante los primeros 25 años del siglo XX los caimanes eran pro-
bablemente tan abundantes como cuando Humboldt y otros natura-
listas del siglo XIX (Appun, 1961; Páez, 1980) se sorprendieron por
su número. Whitney (1912) y Calzadilla (1948) se refirieron a la
abundancia de caimanes en esos primeros años del siglo pasado en
ríos como el Apure y el Matiyure.
Una nueva fase de explotación comercial se inició a finales de
los años 20 (Medem, 1983). El empleo de nuevos métodos de ca-
cería (arpones y linternas eléctricas) y un incremento en la deman-
da internacional por las pieles de los grandes reptiles, se combi-
naron para llevar al borde de la extinción, en menos de tres
décadas, a unas especies cuyos individuos originalmente podían ser
contados por millones, al menos en el caso del caimán del Orinoco.
Esta situación era inimaginable, de acuerdo con la opinión que tenía
Humboldt cuando visitó a Venezuela en el año 1800:
“Es poco probable que se llegue un día a librar de los
cocodrilos a un país en el cual el laberinto de los ríos sin
número conduce todo los días nuevas bandadas desde las
vertientes orientales de los Andes, por medio del Meta y el
Apure, hacia las costas de la Guayana española.” (Humboldt,
1991, Tomo IV, p. 485).

Los Crocodylia de Venezuela
191
Medem (1981, 1983) presentó el registro más detallado sobre la
explotación de C. intermedius en Venezuela (y Colombia). Sólo las
cifras más resaltantes serán repetidas aquí. El pico de la explo-
tación ocurrió entre los años 1930 y 1931, cuando entre 3.000 y
4.000 cueros eran negociados diariamente en San Fernando de
Apure. Entre 1933 y 1935, Venezuela exportó más de 900,000
pieles de caimanes. La explotación a gran escala finalizó en 1947-
1948, debido esencialmente a la escasez del recurso para ese
momento. Algunos cazadores persistieron en esa actividad por
varios años, pero la exportación de pieles de caimanes desde 1950 a
1963 fue mínima (Mondolfi, 1965). Cuando los caimanes se
volvieron escasos en las áreas donde históricamente eran más
abundantes, las poblaciones más distantes, en la periferia de su
área de distribución, comenzaron a ser cazadas. Hitchkock (1948),
por ejemplo, señaló que una partida de cazadores mató 58 caima-
nes durante la estación seca de 1947 en el río Ventuari, uno de los
extremos sureños de la distribución de la especie.
No existen registros que permitan discriminar en cual proporción
las cifras de cueros de caimanes exportados por Venezuela entre
1929 y 1963 correspondieron a C. intermedius o a C. acutus, ya
que no se distinguió nunca entre esas dos especies. Es de suponer
que la cacería fue igualmente intensa en el caso el caimán de la
costa. No obstante la falta de estadísticas oficiales al respecto, con
base en conversaciones con pescadores, lugareños y personas
conocedoras, se puede establecer que la explotación comercial fue
particularmente fuerte en la Laguna de Tacarigua en Miranda y en
el río Catatumbo y otros ríos de la cuenca del lago de Maracaibo.
Lo mismo debió haber ocurrido en ríos como el Tocuyo, Aroa,
Yaracuy, Unare y Neverí.
Todas las actividades de explotación previamente mencionadas
fueron ejecutadas legalmente, como lo comprueban las resoluciones
y decretos publicados en la prensa del país, principalmente en el
periódico gomecista El Nuevo Diario. Como ejemplo transcribo el
texto de una resolución del Ministerio de Fomento, aparecida en el
mencionado diario el 10 de octubre de 1930:

Andrés Eloy Seijas
192
Hacia finales de los años 40, se empezó a manifestar preocu-
pación por la escasez de caimanes como consecuencia de la
actividad comercial (Blohm, 1948; Mondolfi, 1957; Medina-Padilla,
1960). El estado extremadamente deprimido de las poblaciones de
las especies fue materia de preocupación en publicaciones poste-
riores (Mondolfi, 1965; Donoso-Barros, 1966b; Rivero-Blanco, 1968,
King, 1973, Blohm, 1973). Irónicamente, los primeros instrumentos
legales para proteger la especie fueron emitidos entre 1968-1970 en
Colombia (Donadío, 1981) y entre 1970-1971 en Venezuela (Vene-
zuela 1970), cuando ya quedaba poco que resguardar.
A comienzos de los años 60, una vez que los grandes cocodrilos
habían sido agotados como recurso, se incorporó a la baba
(Caiman crocodilus) al comercio de pieles (Medem 1981, 1983).
La explotación de esta especie, simpátrica de los dos grandes
caimanes, de piel menos valiosa pero ya para ese momento más
abundante, seguramente contribuyó con la completa erradicación de
los caimanes de muchas áreas. Como ha sido señalado por King
(1978) la especie menos valiosa permitió a los cazadores y a los
teneros y comerciantes en pieles, tanto nacionales como interna-
cionales, continuar con sus operaciones cuando las especies más
LA CAZA DA CAIMANES
PERMISO CONCEDIDO
Por Resolución del Ministerio de
Fomento y llenados los requisitos de
Ley, se le ha concedido al señor Er-
nesto Sánchez Lacruz, comerciante,
domiciliado en esta ciudad, permiso
para cazar caimanes en el río Unare,
jurisdicción del Municipio Píritu, Dis-
trito Peñalver del Estado Anzoátegui.
Nota: Se mantiene el error encontrado en el original
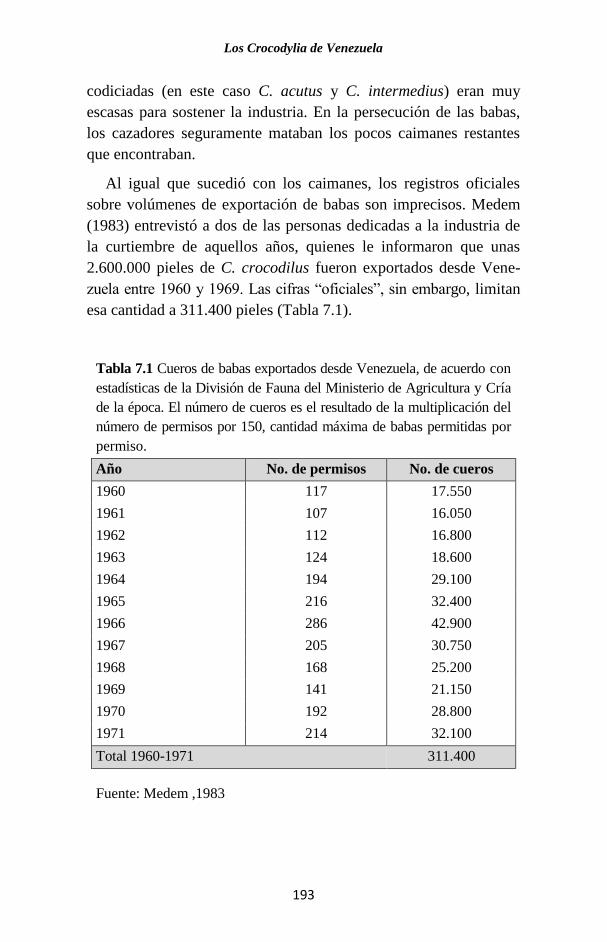
Los Crocodylia de Venezuela
193
codiciadas (en este caso C. acutus y C. intermedius) eran muy
escasas para sostener la industria. En la persecución de las babas,
los cazadores seguramente mataban los pocos caimanes restantes
que encontraban.
Al igual que sucedió con los caimanes, los registros oficiales
sobre volúmenes de exportación de babas son imprecisos. Medem
(1983) entrevistó a dos de las personas dedicadas a la industria de
la curtiembre de aquellos años, quienes le informaron que unas
2.600.000 pieles de C. crocodilus fueron exportados desde Vene-
zuela entre 1960 y 1969. Las cifras “oficiales”, sin embargo, limitan
esa cantidad a 311.400 pieles (Tabla 7.1).
Tabla 7.1 Cueros de babas exportados desde Venezuela, de acuerdo con
estadísticas de la División de Fauna del Ministerio de Agricultura y Cría
de la época. El número de cueros es el resultado de la multiplicación del
número de permisos por 150, cantidad máxima de babas permitidas por
permiso.
Año No. de permisos No. de cueros
1960 117 17.550
1961 107 16.050
1962 112 16.800
1963 124 18.600
1964 194 29.100
1965 216 32.400
1966 286 42.900
1967 205 30.750
1968 168 25.200
1969 141 21.150
1970 192 28.800
1971 214 32.100
Total 1960-1971 311.400
Fuente: Medem ,1983
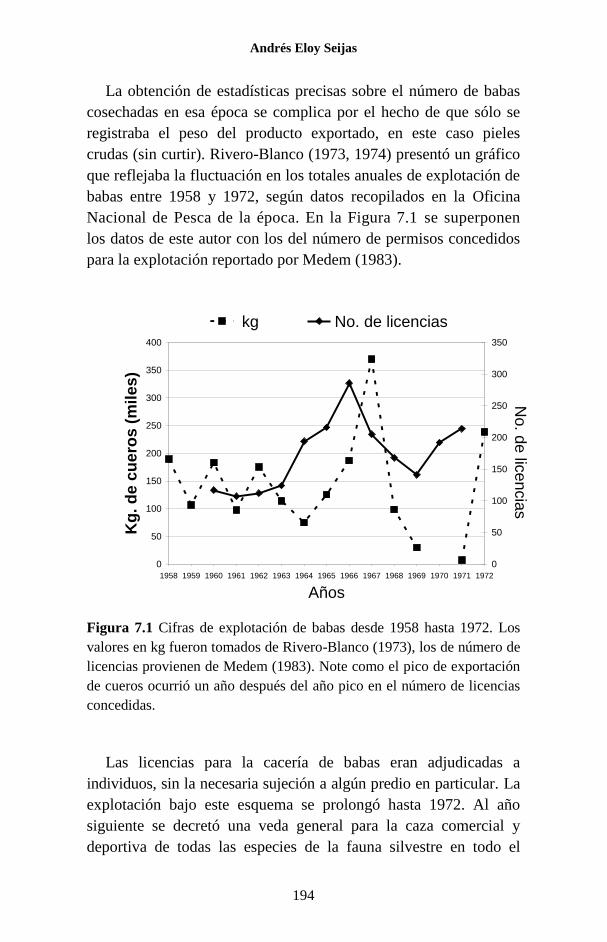
Andrés Eloy Seijas
194
La obtención de estadísticas precisas sobre el número de babas
cosechadas en esa época se complica por el hecho de que sólo se
registraba el peso del producto exportado, en este caso pieles
crudas (sin curtir). Rivero-Blanco (1973, 1974) presentó un gráfico
que reflejaba la fluctuación en los totales anuales de explotación de
babas entre 1958 y 1972, según datos recopilados en la Oficina
Nacional de Pesca de la época. En la Figura 7.1 se superponen
los datos de este autor con los del número de permisos concedidos
para la explotación reportado por Medem (1983).
Figura 7.1 Cifras de explotación de babas desde 1958 hasta 1972. Los
valores en kg fueron tomados de Rivero-Blanco (1973), los de número de
licencias provienen de Medem (1983). Note como el pico de exportación
de cueros ocurrió un año después del año pico en el número de licencias
concedidas.
Las licencias para la cacería de babas eran adjudicadas a
individuos, sin la necesaria sujeción a algún predio en particular. La
explotación bajo este esquema se prolongó hasta 1972. Al año
siguiente se decretó una veda general para la caza comercial y
deportiva de todas las especies de la fauna silvestre en todo el
0
50
100
150
200
250
300
350
400
1958 1959 1960 1961 1962 1963 1964 1965 1966 1967 1968 1969 1970 1971 1972
Años
Kg
. d
e c
ue
ros (
mil
es
)
0
50
100
150
200
250
300
350
No. d
e lic
en
cia
s
kg No. de licencias

Los Crocodylia de Venezuela
195
territorio de la República (Venezuela, 1973) la cual se prolongaría,
para el caso de la baba, por unos 10 años.
En 1983 se inició en Venezuela un nuevo programa de aprove-
chamiento, esta vez sólo para la baba, único crocodilio con
poblaciones relativamente numerosas en el país para ese momento
(Seijas, 1986). Este programa ha estado restringido casi exclusiva-
mente a los llanos occidentales. Entre 1993 y 1997 también se llevó
a cabo un programa de aprovechamiento de las babas del delta del
Orinoco, que tuvo unos alcances limitados, debido al relativamente
pequeño número de ejemplares que se cosecharon (Tabla 7.2).
La información sobre los aspectos legales que rigen el programa
de aprovechamiento comercial de la baba en los llanos occidentales
de Venezuela es muy abundante. Se remite a los lectores a
algunas de las fuentes de referencia más importantes (Velasco y
Ayarzagüena, 1995; Velasco et al., 2003; De Sola et al., 2000;
Villarroel et al., 2000; Colomine et al., 2000; Thorbjarnarson, 1991;
Thorbjarnarson y Velasco, 1998a,b). Una crítica pormenorizada de
los aspectos técnicos y administrativos del programa ha sido
publicada por Molina y Hernández (2010).
El programa de aprovechamiento de la baba ha tenido marchas
y contramarchas. Hasta 2006 se habían publicado 13 resoluciones
que establecían las normas que lo rigen y cerca de 30 resoluciones
en las que, dadas las circunstancias particulares de cada tempo-
rada, se efectuaron cambios en el calendario de ejecución. La
norma vigente para el momento de la preparación de este libro es
la Nº 195 del 05/04/2006 en GORBV Nº 38412 del 21/04/2006.
Ésta derogó a la Nº 23 del año 99 (R. De Sola, com. pers.).
El ámbito de ejecución del programa de aprovechamiento de la
baba son las tierras de propiedad privada o aquellas que estaban
bajo la propiedad y posesión del Instituto Agrario Nacional hoy
suprimidas y traspasadas al Instituto Nacional de Tierras (INTI)
por mandato de la Ley de Tierras y Desarrollo Agrario. El ámbito
de ejecución del programa abarca a los estados Apure, Barinas,
Guárico, Cojedes y Portuguesa. Originalmente, también estuvo
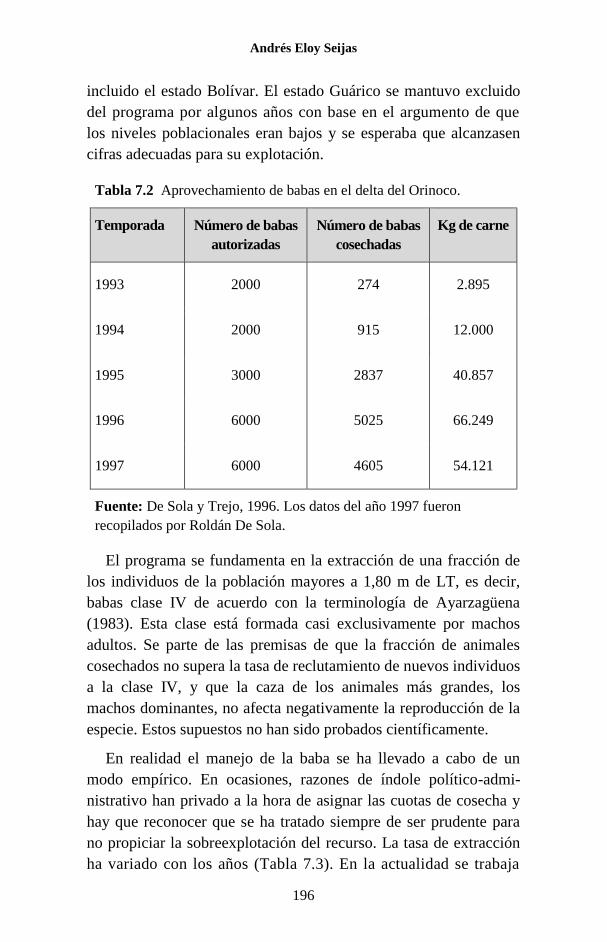
Andrés Eloy Seijas
196
incluido el estado Bolívar. El estado Guárico se mantuvo excluido
del programa por algunos años con base en el argumento de que
los niveles poblacionales eran bajos y se esperaba que alcanzasen
cifras adecuadas para su explotación.
Tabla 7.2 Aprovechamiento de babas en el delta del Orinoco.
Temporada Número de babas
autorizadas
Número de babas
cosechadas
Kg de carne
1993 2000 274 2.895
1994 2000 915 12.000
1995 3000 2837 40.857
1996 6000 5025 66.249
1997 6000 4605 54.121
Fuente: De Sola y Trejo, 1996. Los datos del año 1997 fueron
recopilados por Roldán De Sola.
El programa se fundamenta en la extracción de una fracción de
los individuos de la población mayores a 1,80 m de LT, es decir,
babas clase IV de acuerdo con la terminología de Ayarzagüena
(1983). Esta clase está formada casi exclusivamente por machos
adultos. Se parte de las premisas de que la fracción de animales
cosechados no supera la tasa de reclutamiento de nuevos individuos
a la clase IV, y que la caza de los animales más grandes, los
machos dominantes, no afecta negativamente la reproducción de la
especie. Estos supuestos no han sido probados científicamente.
En realidad el manejo de la baba se ha llevado a cabo de un
modo empírico. En ocasiones, razones de índole político-admi-
nistrativo han privado a la hora de asignar las cuotas de cosecha y
hay que reconocer que se ha tratado siempre de ser prudente para
no propiciar la sobreexplotación del recurso. La tasa de extracción
ha variado con los años (Tabla 7.3). En la actualidad se trabaja

Los Crocodylia de Venezuela
197
con un sistema de asignación de cosechas que trata de tomar en
cuenta las diferencias ecológicas de las regiones en que se
localizan los hatos.
El sistema de regiones ecológicas establece que cada una de
ellas tiene una densidad y estructura poblacional de babas que le
es característico (Fig. 7.2). Se remite al lector a que consulte la
discusión que aparece en el Capítulo 2 respecto a la estructura de
tamaños de las poblaciones de babas.
Tabla 7.3 Cambios en la cuota de aprovechamiento que ha sufrido el
programa de aprovechamiento de babas.
Resolución Fecha Temporadas Cuota asignada Condiciones
445 14-Dic-82 1983; 1984 7% - 12% población total
122 21-Dic-84 1985, 1986 y
1987 7% población total
73 29-Dic-87 1988 7% población total*
60 12-Sep-88 1989 25% población clase IV % IV 32; 8% población
total
138 06-Dic-89 1990 25% población clase IV % IV 32; 8% población
total
144 26-Dic-90 1991 25% población clase IV % IV 32; 8% población
total
139 13-Dic-91 1992 25% población clase IV % IV 23,3; 3,5 % población
total. Ajuste a 30.000.
177-A 25-Nov-92 1993; 1994 MARNR - CITES Regiones ecológicas
1 04-Ene-94 1995; 1996 MARNR - CITES Regiones ecológicas
1 04-Ene-97 1997;1998 MARNR - CITES Regiones ecológicas
23 25-Ene-99 1999
(Vigente) MARNR - CITES Regiones ecológicas
Datos aportados por Roldán De Sola. * En 1988, no obstante que la cifra
acordada fue 7%, sólo se asignó 4%.

Andrés Eloy Seijas
198
Es importante hacer notar que, dada la extensión de los llanos y
las insuficiencias de personal con que siempre ha contado el
Ministerio del Ambiente, es imposible una supervisión detallada,
predio por predio, del programa de aprovechamiento (Molina y
Hernández, 2010). Es factible, y probable, que en algunas loca-
lidades las babas hayan sido sobreexplotadas. Muchas veces las
babas no son cazadas en los predios donde corresponde, hay
cacería furtiva, se caza en lugares prohibidos (áreas protegidas y
ríos navegables). Pero en términos globales, tomando a los llanos
como una totalidad, la especie sigue siendo común y abundante.
La asignación de cuotas de aprovechamiento tiene en cuenta la
ubicación de las propiedades en el sistema de ecorregiones ya
señalado, pero también la extensión de los predios para los cuales
se realiza la solicitud. Éstos se categorizan así: Grandes Hatos
(GH), aquellos mayores de 25.000 ha; Hatos (HH), posesiones con
superficies comprendidas desde 12.500 hasta 25.000 ha; Fundos,
propiedades con extensiones desde 3.500 hasta 12.500 ha; Funditos
(FI), con superficies desde 1.000 hasta 3.500 ha, y, finalmente,
Pequeños Funditos (PF), posesiones con superficies menores a 1.000 ha.
Figura 7.2 Densidad bruta y porcentaje de la población de babas repre-
sentada por la clase IV en cada una de las regiones ecológicas de
acuerdo con Velasco y Ayarzagüena (1995).

Los Crocodylia de Venezuela
199
Hasta el año 2004, el total de babas aprovechadas legalmente
en los llanos occidentales de Venezuela alcanzó la cifra de
1.307.095 individuos (Tabla 7.4).
La cifra total no parece muy grande si la comparamos con las
señaladas para el período previo de explotación comercial de los
años 60. La cantidades anuales han fluctuado considerablemente y
tal vez la del año 1985 representó una sobreexplotación, lo que
sensibilizó a la opinión pública y obligó al MARN a decretar una
suspensión (“pausa ecológica” fue denominada) en 1986. En los
últimos 10 años la cifra de explotación se puede considerar
modesta y, probablemente, sustentable. Si tomamos como cierta la
cifra de 2.600.000 babas explotadas entre 1960 y 1969 (Medem
1983), cuando los sistemas de control por parte del estado eran
extremadamente laxos o inexistentes, entonces la cifra total del
periodo 1983-2004 (más de 20 años) representa una tasa de
extracción cuatro veces menor. En cualquier caso, hay que man-
tener una vigilancia constante sobre la manera como se maneja el
recurso y mejorar los sistemas de monitoreo de las poblaciones que
posibiliten la ejecución de respuestas rápidas ante cualquier señal
de deterioro.
ZOOCRIADEROS COMERCIALES
Desde finales de los años 80, paralelo al programa de aprove-
chamiento de las poblaciones de babas silvestres, se inició un pro-
grama de cría en cautiverio apoyado en el rancheo de huevos
(Martínez, 1996). Gran parte de la información que aparecerá en
los párrafos siguientes se generó a partir de una importante
recopilación efectuada por el Lic. Roldan De Sola, funcionario del
Ministerio del Ambiente.
El establecimiento de los zoocriaderos de babas se inició con
fines “experimentales” (nunca quedó claro en qué consistía el
experimento). La primera resolución que regulaba el funcionamiento
de éstos fue la No. 79, publicada en la Gaceta Oficial de la
República de Venezuela No. 34.490, del 15 de junio de 1990
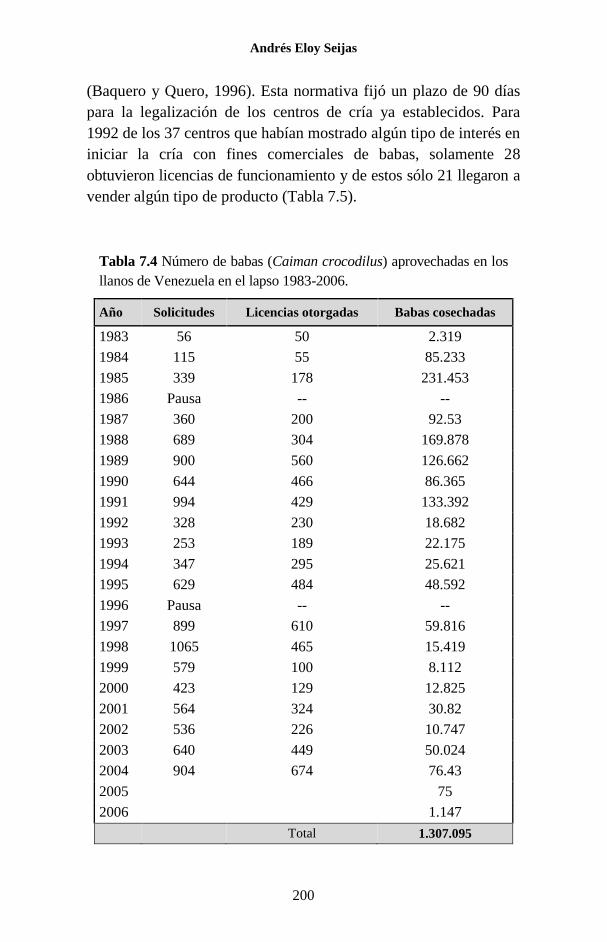
Andrés Eloy Seijas
200
(Baquero y Quero, 1996). Esta normativa fijó un plazo de 90 días
para la legalización de los centros de cría ya establecidos. Para
1992 de los 37 centros que habían mostrado algún tipo de interés en
iniciar la cría con fines comerciales de babas, solamente 28
obtuvieron licencias de funcionamiento y de estos sólo 21 llegaron a
vender algún tipo de producto (Tabla 7.5).
Tabla 7.4 Número de babas (Caiman crocodilus) aprovechadas en los
llanos de Venezuela en el lapso 1983-2006.
Año Solicitudes Licencias otorgadas Babas cosechadas
1983 56 50 2.319
1984 115 55 85.233
1985 339 178 231.453
1986 Pausa -- --
1987 360 200 92.53
1988 689 304 169.878
1989 900 560 126.662
1990 644 466 86.365
1991 994 429 133.392
1992 328 230 18.682
1993 253 189 22.175
1994 347 295 25.621
1995 629 484 48.592
1996 Pausa -- --
1997 899 610 59.816
1998 1065 465 15.419
1999 579 100 8.112
2000 423 129 12.825
2001 564 324 30.82
2002 536 226 10.747
2003 640 449 50.024
2004 904 674 76.43
2005 75
2006 1.147
Total 1.307.095
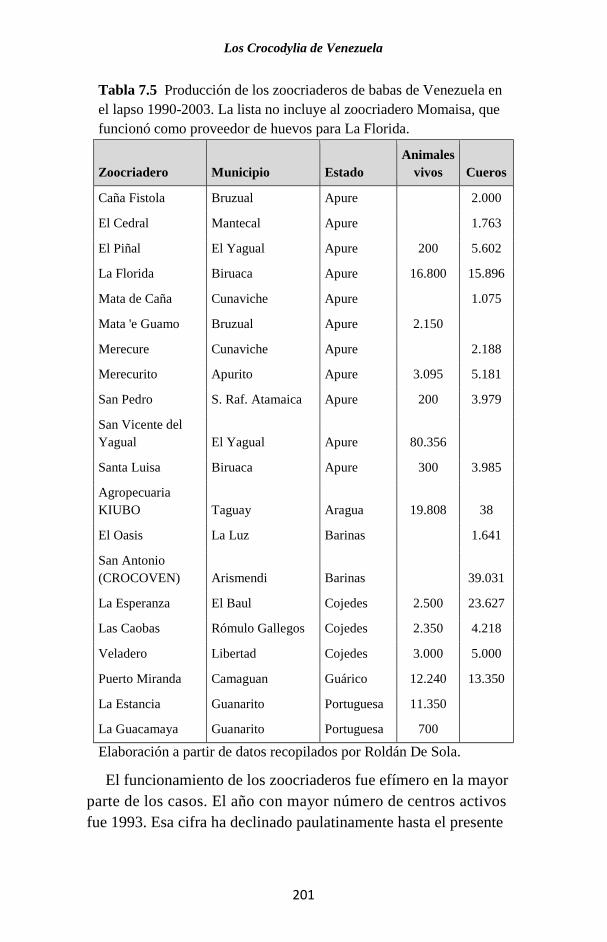
Los Crocodylia de Venezuela
201
Tabla 7.5 Producción de los zoocriaderos de babas de Venezuela en
el lapso 1990-2003. La lista no incluye al zoocriadero Momaisa, que
funcionó como proveedor de huevos para La Florida.
Zoocriadero Municipio Estado
Animales
vivos Cueros
Caña Fistola Bruzual Apure 2.000
El Cedral Mantecal Apure 1.763
El Piñal El Yagual Apure 200 5.602
La Florida Biruaca Apure 16.800 15.896
Mata de Caña Cunaviche Apure 1.075
Mata 'e Guamo Bruzual Apure 2.150
Merecure Cunaviche Apure 2.188
Merecurito Apurito Apure 3.095 5.181
San Pedro S. Raf. Atamaica Apure 200 3.979
San Vicente del
Yagual El Yagual Apure 80.356
Santa Luisa Biruaca Apure 300 3.985
Agropecuaria
KIUBO Taguay Aragua 19.808 38
El Oasis La Luz Barinas 1.641
San Antonio
(CROCOVEN) Arismendi Barinas 39.031
La Esperanza El Baul Cojedes 2.500 23.627
Las Caobas Rómulo Gallegos Cojedes 2.350 4.218
Veladero Libertad Cojedes 3.000 5.000
Puerto Miranda Camaguan Guárico 12.240 13.350
La Estancia Guanarito Portuguesa 11.350
La Guacamaya Guanarito Portuguesa 700
Elaboración a partir de datos recopilados por Roldán De Sola.
El funcionamiento de los zoocriaderos fue efímero en la mayor
parte de los casos. El año con mayor número de centros activos
fue 1993. Esa cifra ha declinado paulatinamente hasta el presente
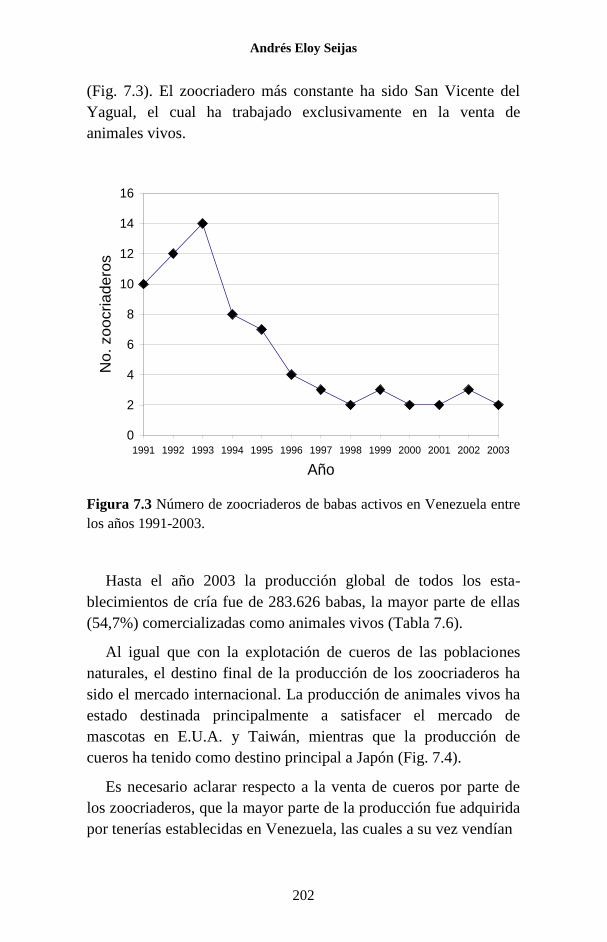
Andrés Eloy Seijas
202
(Fig. 7.3). El zoocriadero más constante ha sido San Vicente del
Yagual, el cual ha trabajado exclusivamente en la venta de
animales vivos.
Figura 7.3 Número de zoocriaderos de babas activos en Venezuela entre
los años 1991-2003.
Hasta el año 2003 la producción global de todos los esta-
blecimientos de cría fue de 283.626 babas, la mayor parte de ellas
(54,7%) comercializadas como animales vivos (Tabla 7.6).
Al igual que con la explotación de cueros de las poblaciones
naturales, el destino final de la producción de los zoocriaderos ha
sido el mercado internacional. La producción de animales vivos ha
estado destinada principalmente a satisfacer el mercado de
mascotas en E.U.A. y Taiwán, mientras que la producción de
cueros ha tenido como destino principal a Japón (Fig. 7.4).
Es necesario aclarar respecto a la venta de cueros por parte de
los zoocriaderos, que la mayor parte de la producción fue adquirida
por tenerías establecidas en Venezuela, las cuales a su vez vendían
0
2
4
6
8
10
12
14
16
1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003
Año
No.
zoocriadero
s
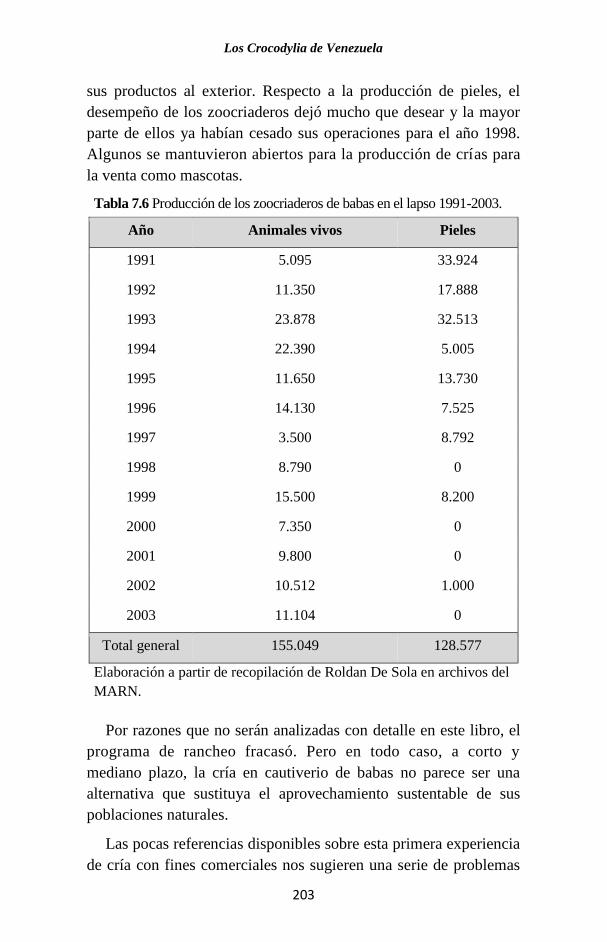
Los Crocodylia de Venezuela
203
sus productos al exterior. Respecto a la producción de pieles, el
desempeño de los zoocriaderos dejó mucho que desear y la mayor
parte de ellos ya habían cesado sus operaciones para el año 1998.
Algunos se mantuvieron abiertos para la producción de crías para
la venta como mascotas.
Tabla 7.6 Producción de los zoocriaderos de babas en el lapso 1991-2003.
Año Animales vivos Pieles
1991 5.095 33.924
1992 11.350 17.888
1993 23.878 32.513
1994 22.390 5.005
1995 11.650 13.730
1996 14.130 7.525
1997 3.500 8.792
1998 8.790 0
1999 15.500 8.200
2000 7.350 0
2001 9.800 0
2002 10.512 1.000
2003 11.104 0
Total general 155.049 128.577
Elaboración a partir de recopilación de Roldan De Sola en archivos del
MARN.
Por razones que no serán analizadas con detalle en este libro, el
programa de rancheo fracasó. Pero en todo caso, a corto y
mediano plazo, la cría en cautiverio de babas no parece ser una
alternativa que sustituya el aprovechamiento sustentable de sus
poblaciones naturales.
Las pocas referencias disponibles sobre esta primera experiencia
de cría con fines comerciales nos sugieren una serie de problemas
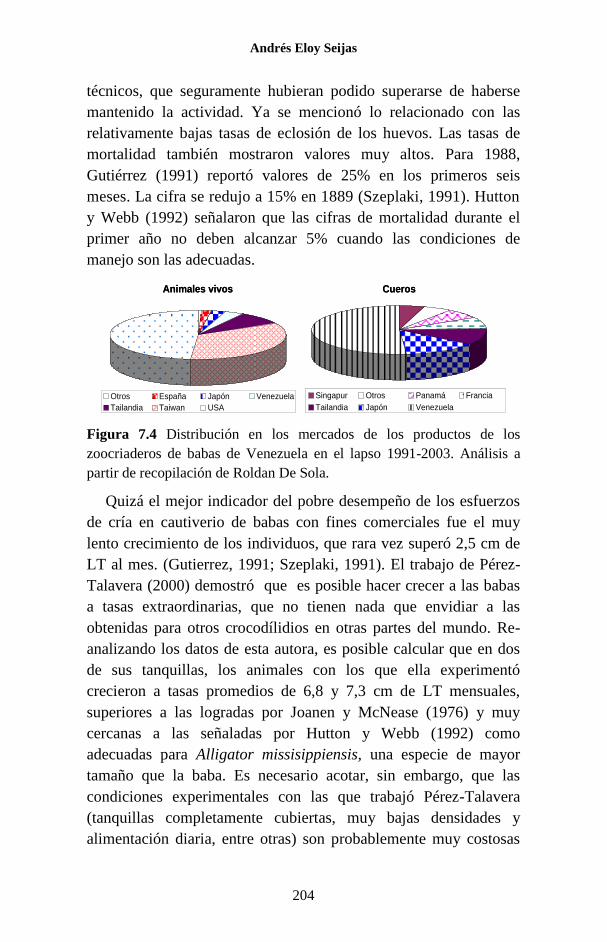
Andrés Eloy Seijas
204
técnicos, que seguramente hubieran podido superarse de haberse
mantenido la actividad. Ya se mencionó lo relacionado con las
relativamente bajas tasas de eclosión de los huevos. Las tasas de
mortalidad también mostraron valores muy altos. Para 1988,
Gutiérrez (1991) reportó valores de 25% en los primeros seis
meses. La cifra se redujo a 15% en 1889 (Szeplaki, 1991). Hutton
y Webb (1992) señalaron que las cifras de mortalidad durante el
primer año no deben alcanzar 5% cuando las condiciones de
manejo son las adecuadas.
Figura 7.4 Distribución en los mercados de los productos de los
zoocriaderos de babas de Venezuela en el lapso 1991-2003. Análisis a
partir de recopilación de Roldan De Sola.
Quizá el mejor indicador del pobre desempeño de los esfuerzos
de cría en cautiverio de babas con fines comerciales fue el muy
lento crecimiento de los individuos, que rara vez superó 2,5 cm de
LT al mes. (Gutierrez, 1991; Szeplaki, 1991). El trabajo de Pérez-
Talavera (2000) demostró que es posible hacer crecer a las babas
a tasas extraordinarias, que no tienen nada que envidiar a las
obtenidas para otros crocodílidios en otras partes del mundo. Re-
analizando los datos de esta autora, es posible calcular que en dos
de sus tanquillas, los animales con los que ella experimentó
crecieron a tasas promedios de 6,8 y 7,3 cm de LT mensuales,
superiores a las logradas por Joanen y McNease (1976) y muy
cercanas a las señaladas por Hutton y Webb (1992) como
adecuadas para Alligator missisippiensis, una especie de mayor
tamaño que la baba. Es necesario acotar, sin embargo, que las
condiciones experimentales con las que trabajó Pérez-Talavera
(tanquillas completamente cubiertas, muy bajas densidades y
alimentación diaria, entre otras) son probablemente muy costosas
Animales vivos
Otros España Japón Venezuela
Tailandia Taiwan USA
Cueros
Singapur Otros Panamá Francia
Tailandia Japón Venezuela
Animales vivos
Otros España Japón Venezuela
Tailandia Taiwan USA
Cueros
Singapur Otros Panamá Francia
Tailandia Japón Venezuela
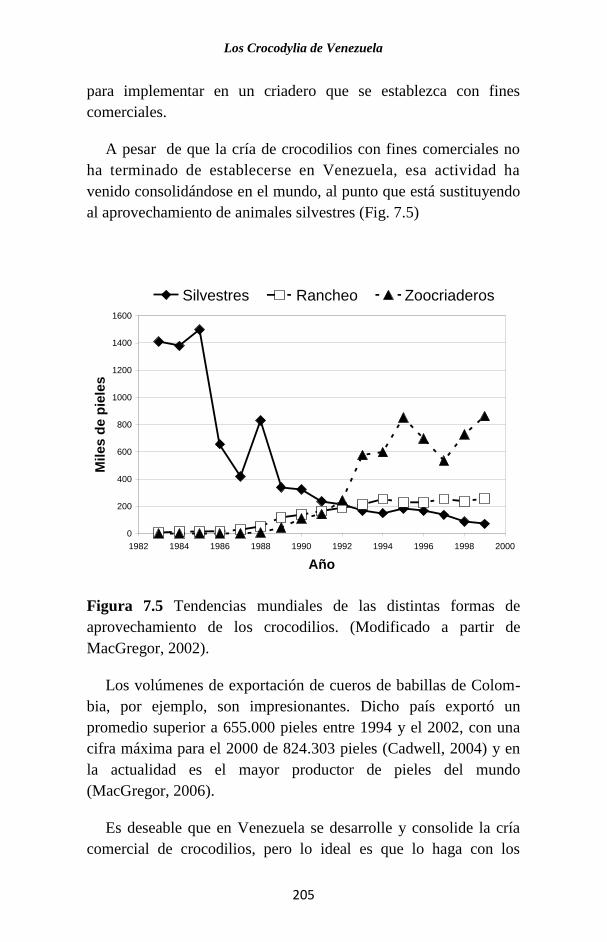
Los Crocodylia de Venezuela
205
para implementar en un criadero que se establezca con fines
comerciales.
A pesar de que la cría de crocodilios con fines comerciales no
ha terminado de establecerse en Venezuela, esa actividad ha
venido consolidándose en el mundo, al punto que está sustituyendo
al aprovechamiento de animales silvestres (Fig. 7.5)
Figura 7.5 Tendencias mundiales de las distintas formas de
aprovechamiento de los crocodilios. (Modificado a partir de
MacGregor, 2002).
Los volúmenes de exportación de cueros de babillas de Colom-
bia, por ejemplo, son impresionantes. Dicho país exportó un
promedio superior a 655.000 pieles entre 1994 y el 2002, con una
cifra máxima para el 2000 de 824.303 pieles (Cadwell, 2004) y en
la actualidad es el mayor productor de pieles del mundo
(MacGregor, 2006).
Es deseable que en Venezuela se desarrolle y consolide la cría
comercial de crocodilios, pero lo ideal es que lo haga con los
0
200
400
600
800
1000
1200
1400
1600
1982 1984 1986 1988 1990 1992 1994 1996 1998 2000
Año
Miles
de p
iele
s
Silvestres Rancheo Zoocriaderos

Andrés Eloy Seijas
206
grandes cocodrilos C. acutus y C.intermedius (Ayarzagüena,
1991; Seijas, 1992) especies que poseen una piel mucho más
valiosa que la de la baba. Es necesario advertir, sin embargo, que
el desarrollo de la cría en cautiverio de los grandes cocodrilos de
Venezuela no puede ir en detrimento de la conservación de las
poblaciones naturales. Muy por el contrario, esta actividad debe
estar condicionada al mantenimiento de saludables y numerosas
poblaciones silvestres. Los zoocriaderos cerrados tienen muy poco
(si alguno) valor conservacionista (MacGregor, 2006).
Tal como señaló Thorbjarnarson (2002), se ha vendido la idea
de que el aprovechamiento comercial de la fauna es una especie
de panacea que contribuirá a la conservación del recurso. Sin
embargo, ha sido el esfuerzo mundial en proteger las poblaciones
naturales y el establecimiento de límites al comercio lo que ha
permitido el espectacular retorno de las poblaciones de algunas
especies, como el aligátor (Alligator mississippiensis) y ciertas po-
blaciones del cocodrilo del Nilo (Crocodylus niloticus) y del de
agua salobre (Crocodylus porosus). No se debe llegar tampoco al
extremo de creer que el aprovechamiento comercial y la conser-
vación de los crocodilios son actividades antagónicas. El uso con
fines de consumo debe considerarse como una herramienta comple-
mentaria, que provea incentivos a propietarios o habitantes locales
para que protejan a unos animales que, de otra manera, no son
muy bien vistos (Hutton et al., 2001).
También se ha señalado que el aprovechamiento de las pobla-
ciones naturales de crocodilios es un incentivo que tienen los
propietarios de tierras para proteger sus hábitats. Por los momentos
se puede señalar, que no hay evidencias de que el aprovechamiento
de las babas en Venezuela haya influenciado la toma de decisiones
relacionadas con el uso de la tierra (Thorbjarnarson y Velasco,
1998a,b). La principal actividad económica en los llanos es la gana-
dería y en la búsqueda de mecanismos para mejorarla, los dueños
de tierra han creado préstamos y aumentado la capacidad de
retención de agua de las sabanas, lo que ha beneficiado circuns-
tancialmente a la baba. En el caso de los caimanes, especialmente
con respecto a C. intermedius, el rancheo de huevos y neonatos

Los Crocodylia de Venezuela
207
para ser suministrados luego a zoocriaderos con fines comerciales
puede ser una actividad que beneficie a la gente que vive a lo
largo de los ríos y pudiera convertirse en un incentivo para la
conservación de sus poblaciones y su hábitat. Este es un esfuerzo
que vale la pena intentar.

Andrés Eloy Seijas
208

Los Crocodylia de Venezuela
209
CAPÍTULO 8
CRÍA EN CAUTIVERIO
Comparado con otros países (EE.UU., Australia y Colombia, por
ejemplo), la experiencia en la cría en cautiverio de crocodilios en
Venezuela ha sido muy limitada. El que se puede considerar como
el trabajo pionero en este campo fue llevado a cabo por Carlos
Rivero-Blanco (Rivero-Blanco 1973, 1974) quien colectó 170 nidos
de babas en el hato La Guanota, cerca de San Fernando de Apure,
para intentar su incubación en condiciones controladas y el levante
posterior de las crías. Debido a la inexperiencia el éxito de eclosión
fue muy bajo (alrededor de 15%). No hay información publicada
sobre el levante posterior de los animales nacidos.
También a comienzos de los años 1970 se empezó a recopilar
información sobre el crecimiento de babas en cautiverio (Blohm
1973). Los registros corresponden a ejemplares mantenidos en
tanquillas improvisadas en la casa del mencionado autor, en Cara-
cas. La tasa promedio de crecimiento de los individuos durante los
primeros 35 meses fue de 2,04 cm/mes. En el artículo citado,
Tomás Blohm también aportó datos sobre los primeras experiencias
de cría con caimanes, específicamente con C. intermedius.
Respecto al caimán de la costa Seijas et al. (1990) realizaron
entre 1984 y 1985 un ensayo de levante con 17 individuos recién
nacidos (de un grupo original de 37) provenientes del río Yaracuy y
los Canales de Río Chico, en el estado Miranda. El estudio se llevó
a cabo en las instalaciones del Servicio Nacional de Fauna Silvestre
del MARNR, en El Limón, estado Aragua. Durante los 15 meses
en que se llevó el registro, los caimanes crecieron a tasas que iban
desde 1,42 a 2,07 cm de longitud total por mes.

Andrés Eloy Seijas
210
Como se discutirá más adelante, tanto el porcentaje de eclosión
de los huevos, así como el incremento en la talla de los crocodilios
en los primeros ensayos de cría fueron extremadamente bajos.
Ellos sirven para ilustrar los alcances poco auspiciosos de los
primeros pasos en la cría de crocodilios en Venezuela. En lo que
se refiere al crecimiento corporal de los crocodilios, el factor que
probablemente jugó un papel determinante en los malos resultados
obtenidos fue la muy baja temperatura de los lugares donde se
mantenía a los animales. Como ejemplo se puede señalar que en el
ensayo con C. acutus ya mencionado, durante los meses más fríos
del año, de diciembre a marzo, las temperaturas mínimas fueron
menores de 17 ºC (Seijas et al., 1990), muy por debajo de los 32 ºC
señalados por Coulson y Hernández (1983) como óptimos para
obtener un crecimiento máximo en Alligator mississppiensis. Igual-
mente, las bajas temperaturas de Caracas deben haber tenido un
efecto determinante en el bajo crecimiento de los caimanes men-
cionados por Blohm (1973) y Rivero-Blanco (1973).
Fue asimismo en la década de los 70 del siglo pasado que se
adquirieron las primeras experiencias con la reproducción en
cautiverio del caimán del Orinoco (Ramírez et al., 1977; Blohm,
1982). Con ellas se demostró que la cría en condiciones contro-
ladas era posible, pero los esfuerzos no tuvieron mayor tras-
cendencia, principalmente porque las instalaciones donde se llevaron
a cabo no eran las más adecuadas (Thorbjarnarson y Blohm,
1986).
Una década más tarde se establecen ya unos centros de cría
para C. intermedius, mejor ubicados geográficamente y con un
manejo más riguroso. Ellos estaban orientados específicamente a la
producción de individuos para la restauración poblacional de la
especie (Thorbjarnarson y Blohm, 1986; Ramo et al., 1992). En el
año 1987 se inició un programa “experimental” de cría de babas
(Espinoza, 1988, 1989; Gutiérrez, 1991) con los objetivos básicos de
repoblar áreas naturales sujetas a explotación comercial y deter-
minar la factibilidad económica de la cría de este crocodilio. Esta
actividad creció de manera explosiva y para 1988 ya había al
menos 36 establecimientos en los que se había tomado alguna
iniciativa para iniciar sus funciones (Espinoza, 1988).

Los Crocodylia de Venezuela
211
TIPOS DE ZOOCRIADEROS
Para la cría de crocodilios en cautiverio en Venezuela se han
manejado todas las alternativas posibles: 1.- Las de los zoocriaderos
cerrados, es decir, aquellos donde los reproductores están en
cautiverio y son la fuente de donde provienen huevos y crías para
el levante; 2.- zoocriaderos abiertos, que son los que mantienen una
vinculación directa con las poblaciones naturales, las cuales son la
fuente proveedora de huevos o neonatos (rancheo) para el levante
en condiciones controladas. En este caso, los reproductores están
libres en su ambiente natural; y 3.- Los zoocriaderos mixtos, que
combinan las dos modalidades anteriores.
Los tipos de zoocriaderos en Venezuela han dependido de las
especies con las que se trabaja. En el caso del caimán del Orinoco
la cría en cautiverio se inició con zoocriaderos cerrados, con
esporádicas colectas de huevos y neonatos, principalmente de los
ríos Capanaparo y Cojedes, localidades con las poblaciones más
grandes de la especie. Con el tiempo, sin embargo, el rancheo de
huevos y recién nacidos se ha convertido en una opción muy
aprovechada al punto de que es prácticamente la única que se
lleva a cabo en los zoocriaderos del hato El Frío y en el más
reciente de todas las instalaciones, la de FUDECI en Puerto
Ayacucho.
En el caso del caimán de la costa, la cría en cautiverio se ha
apoyado principalmente en el rancheo, aun cuando existe una
pareja cautiva en Masaguaral la cual ha producido la mayoría de
los animales levantados en cautiverio en Venezuela. En lo con-
cerniente a las babas, los zoocriaderos que han existido han
trabajado exclusivamente con la modalidad del rancheo.
Desde el punto de vista económico el rancheo es la alternativa
más adecuada, en especial cuando la especie con que se trabaja es
relativamente numerosa y los huevos o neonatos son fáciles de
colectar, que es precisamente lo que ocurre con la baba. Con esta
modalidad no se incurre en gastos de mantenimiento de individuos
adultos cautivos, los cuales requerirían de espacios amplios, cons-

Andrés Eloy Seijas
212
trucción de corrales, suministro de agua abundante, alimentación
apropiada y, posiblemente, eventual atención veterinaria. Lo impor-
tante en estos casos es proteger las poblaciones silvestres o, incluso,
manejarlas para que incremente el número de reproductores.
A continuación se analizarán detalladamente las experiencias de
cría en cautiverio de crocodilios en Venezuela. Sólo se consi-
derarán los aspectos técnicos, es decir aquellos que tienen que ver
con el mantenimiento de los reproductores, la colecta e incubación
de los huevos en condiciones controladas y el levante de animales.
En todos estos puntos se recomienda al lector avanzado que
consulte el excelente material preparado por Hutton y Webb (1992)
sobre el manejo de zoocriaderos de crocodilios. Otro asunto para
discutir es el relacionado con los resultados obtenidos en la
liberación de animales criados en cautiverio con fines de restau-
ración o fomento de poblaciones. Lo referente a los aspectos
comerciales de la cría fue analizado de manera superficial en el
Capítulo anterior.
1. Mantenimiento de adultos
Como se indicó en párrafos anteriores, los zoocriaderos abiertos
dependen de reproductores que vivan libres en su ambiente natural
y que, por lo tanto, no requieran de cuidados especiales, aunque sí
de protección. En ocasiones el mantenimiento de adultos en los
criaderos es recomendable, o inevitable, como ha ocurrido con el
caimán del Orinoco y, en menor medida con el caimán de la costa.
Para estas especies han existido individuos en manos de parti-
culares, que han permanecido cautivos por muchos años y ya no se
considera conveniente su liberación al medio natural. Dado la
situación crítica en que se encuentran estos cocodrilos, cada
individuo sobreviviente es una fuente de genes importante para
conservar y transmitir a las poblaciones futuras. Las recomen-
daciones que se presentan a continuación se refieren a los
caimanes, ya que con las numerosas poblaciones de babas silves-
tres que existen en Venezuela, el mantenimiento de reproductores
de esa especie en cautiverio no tiene ningún tipo de justificación
económica, al menos en la región de los llanos.

Los Crocodylia de Venezuela
213
1.1. No obstante el gran tamaño que alcanzan los caimanes
adultos, se pueden mantener en espacios relativamente reducidos,
siempre que éstos contenga un área seca, donde puedan salir a
asolearse, y un pozo lo suficientemente profundo como para que se
puedan sumergir por completo. La pareja reproductora de C.
acutus del hato Masaguaral, por ejemplo, está ubicada en un corral
de menos de 60 m2, con un pozo circular de unos 4 m de
diámetro. Es necesario recordar que los crocodilios copulan en el
agua y que, por lo tanto, estanques muy someros pudieran impedir
que el apareamiento se lleve a cabo de manera efectiva.
1.2. La parte con agua del encierro puede ser una simple
excavación en el terreno, si las características del suelo lo per-
miten. En los llanos este tipo de pozos se mantienen llenos en el
período de lluvias pero tienden a secarse en la época seca, por lo
cual es necesario mantener una fuente de agua para reponer la
que se pierde por evaporación e infiltración. En terrenos que no
permiten la retención de agua hay que construir estanques de
concreto o alguna alternativa que supla de manera constante el
agua que se pierde.
1.3. El cercado que rodea a los encierros debe ser a prueba de
escapes. Las cercas tipo “ciclón”, con unos 1,80 m de altura
cumplen perfectamente con estos requerimientos (Fig. 8.1).
1.4. El número de adultos por corral puede variar dependiendo
del tamaño de éste. No es recomendable mantener más de un
macho por encierro, ya que estos son extremadamente territoriales
y se pelean hasta la muerte. Cuando los corrales son muy
pequeños hay que mantener sólo una pareja, ya que cuando el
espacio es muy reducido las hembras también pelean entre si con
riesgos de muerte.
1.5. Contrario a lo que el común de la gente piensa, la cantidad
de comida necesaria para mantener a un caimán adulto es muy
pequeña, si se compara con la que requeriría un mamífero de su
mismo peso. Una o dos gallinas a la semana, por ejemplo, es
suficiente, dependiendo del tamaño del ejemplar. En el caso de la
UNELLEZ los adultos se alimentan con los animales muertos que
aparecen eventualmente en otras unidades de producción de la

Andrés Eloy Seijas
214
institución (conejos, cochinos, ovejas) o con becerros nonatos
conseguidos en el matadero que allí funciona. Otros criaderos de
caimanes alimentan a sus adultos con caballos y burros de
descarte. Otras alternativas son posibles (pescado, chigüires, cerdos
silvestres, entre otros) de acuerdo con la disponibilidad de recursos
que ofrezca el área donde se encuentran las instalaciones.
Figura 8.1 Encierro tipo “préstamo” en el zoocriadero de la UNELLEZ.
1.6. Se debe acondicionar, en algún lugar cercano al cuerpo de
agua, una pequeña “playa” que sirva como lugar de anidación para la
hembra. Será necesario preparar tantos nidos artificiales como hem-
bras haya en el encierro, para evitar la competencia entre ellas por
lugares de anidación. El nido artificial consiste en un hueco excavado
en el terreno (mínimo 1 m2) con una profundidad de al menos 50 cm.
Dicha cavidad se rellena con arena fina, idealmente recogida de la
playa de un río cercano. Hay que colocar arena suficiente como para
formar un montículo que sobresalga 30-40 cm. sobre el terreno. La
excavación donde se vacía la arena debe estar por encima del nivel
del agua del pozo aledaño, para evitar que se inunde.

Los Crocodylia de Venezuela
215
1.7. Los lugares para la anidación deben ser acondicionados por
lo menos con un mes de anticipación al momento esperado del
desove, para que la hembra tenga tiempo de reconocerlo.
COLECTA E INCUBACIÓN DE HUEVOS
La colecta de huevos de crocodilios para su posterior incubación
es un trabajo delicado, que requiere de una serie de precauciones.
Cuando el trabajo se realiza de la manera adecuada, el éxito de
eclosión debe superar 90% (Joanen y McNease, 1991). La infor-
mación disponible para Venezuela indica que el éxito promedio de
la incubación de huevos de Crocodylus colectados en el medio
natural ha sido bajo (Jiménez-Oraá, 2007, Barros et al., 2010),
aunque Antelo et al. (2010) reportan un aceptable 84,35% de éxito,
una vez descartados para los cálculos 24,6% de los huevos
originales por considerarlos infértiles. En el caso de reproductores
mantenidos en cautiverio, en el zoocriadero de la UNELLEZ el
éxito de eclosión total desde el 2003 hasta el 2008 fue 64,4%.
En Masaguaral, para las temporadas 1998 y 1999, el éxito global
de eclosión alcanzó 66,1% (Thorbjarnarson, 1999). En descargo de
estos malos resultados hay que señalar que en ninguno de los
zoocriaderos del país se realiza una revisión minuciosa de los
huevos que se colocan en la incubadora, para descartar los
infértiles. No obstante el relativo pobre desempeño señalado,
algunos eventos aislados demuestran que es posible lograr valores
óptimos (Seijas y González, 1994).
En el caso de la baba existen estadísticas sobre éxito de
eclosión de nidadas para algunos zoocriaderos. En 1987 los hatos
afiliados a ASOBABA (Asociación para la Cría y Conservación de
la Baba) recolectaron un total de 12.002 huevos, de los cuales
eclosionaron de manera exitosa 7.642 (68,3%; Espinoza, 1989). Al
año siguiente, el éxito se elevó hasta 87,0% (Gutiérrez, 1991;
Szeplaki, 1991). Los valores reportados son mejores que los antes
señalados para los caimanes, pero todavía lejos de las cifras
óptimas.

Andrés Eloy Seijas
216
Los aspectos elementales que se deben tener en cuenta para
obtener los mejores resultados en la incubación de huevos de
crocodilios en condiciones controladas, tienen que ver con el mo-
mento de la colecta, manipulación de los huevos, material usado
para el mantenimiento de los huevos, y la temperatura y humedad
relativa del ambiente donde se incuban. Los bajos resultados
obtenidos en Venezuela nos indican que algunos de los factores
mencionados no han sido los adecuados. A continuación se enu-
meran los pasos que deben seguirse, al tomar como base el trabajo
de Joanen y McNease (1991), un manual preparado para el hato
Masaguaral (Thorbjarnarson y Rodríguez, 1992), el artículo de
Martínez (1996) y la experiencia adquirida en el zoocriadero de la
UNELLEZ.
2. Colecta de los huevos
2.1. Las hembras de los crocodilios protegen a sus nidos. Esto
es particularmente cierto con las caimanas de los zoocriaderos. Es
por ello que el primer paso en la localización del nido (cuando se
trata de poblaciones silvestres) y llegar a los huevos que se quieren
recoger, es tener las mayor precaución para evitar heridas de los
colectores y de los mismos animales. Para las hembras cautivas en
algunos casos es necesario amarrarlas, para mantenerlas alejadas
del nido, pero generalmente se puede trabajar sin recurrir a ese
extremo. En este caso a las hembras se les empuja al agua con la
ayuda de varas largas, propinándole pequeños golpes en la punta
del hocico y en los costados. La idea no es maltratar al animal,
sino mantenerlo a raya. Se recomienda un mínimo de tres personas
para realizar esta labor (Fig. 8.2).
2.2. En todo momento la manipulación de los huevos debe ser
cuidadosa y evitar que éstos roten o sufran movimientos bruscos.
Idealmente, deben colectarse en las primeras 24 horas después de
la puesta. En ese momento los embriones no se han fijado a la
membrana del huevo y son menos susceptibles a la manipulación.
Si la colecta se efectúa después de las 24 horas hay que ser
mucho más cuidadosos en su manejo, para reducir la probabilidad
de muerte de los embriones.

Los Crocodylia de Venezuela
217
2.3. Los huevos deben ser trasladados desde el nido hasta la
bandeja de colecta, la cual debe estar preparada con material
adecuado, generalmente colectado del mismo nido natural. Los
huevos deben ser colocados en la misma posición en que se
encontraron en el nido, impidiendo que rueden o sufran cualquier
tipo de desplazamiento súbito.
Figura 8.2 Colecta de huevos de una de las hembras del zoocriadero de
la UNELLEZ. Una persona permanece vigilante con una vara para evitar
que la hembra suba hasta el nido.
2.4. A medida que se sacan los huevos, se cuentan y se anota
el número de ellos que aparecen con fracturas o con cualquier otro
detalle que pudiera afectar su futuro desarrollo. Si la colecta se
realiza en las primeras 24 horas después de la puesta, es posible
que la mayor parte de ellos no presenten la mancha o banda típica
que señala el punto de fijación del embrión, por lo que no es
posible saber si el huevo está fertilizado o no. Aun en esta etapa
es posible detectar la presencia del embrión, si se observa el huevo
al trasluz en un cuarto oscuro. Para colectas más tardías la banda

Andrés Eloy Seijas
218
de fijación del embrión debe ser visible y su anchura puede ser
usada para determinar qué tan avanzado está el proceso de
desarrollo embrionario (Joanen y McNease, 1991). En este último
caso, la ausencia de banda puede ser usada como indicadora de
que el huevo no ha sido fertilizado.
2.5. Todos los pasos anteriores deben llevarse a cabo en el
tiempo más corto que la prudencia indique. Hay que evitar que los
huevos permanezcan mucho tiempo descubiertos y expuestos a la
deshidratación y el sobrecalentamiento por efecto de la brisa y el
sol.
2.6. Luego los huevos son trasladados a la incubadora. Si
ésta está muy distante del sitio de colecta, es preferible colocar los
huevos en recipientes herméticos (cavas), cubrirlos con material del
nido y taparlos para evitar pérdida excesiva de humedad. El
traslado a largas distancias, particularmente si las vías de comu-
nicación no están en buenas condiciones, pone en riesgo la
viabilidad de los huevos.
3. Cuarto de incubación
3.1. De acuerdo con la experiencia en el zoocriadero de la
UNELLEZ con C. intermedius, se puede aseverar que la mejor
alternativa para la incubación es colocar cada nidada en un
recipiente cerrado (hielera o cava) con arena. Los otros zoo-
criaderos han construidos “canteros” llenos de arena, donde se
entierran los huevos simulando las condiciones naturales. En el
caso de los aligatóridos habría que colocar materia vegetal en el
nido, preferiblemente recogida en el mismo nido natural. Los
huevos deben permanecer completamente cubiertos por una capa
que supere 10 cm. El material del nido tiene que contener suficiente
humedad como para garantizar una saturación cercana a 100%
durante todo el período de incubación. Debe evitarse, sin embargo,
exceso de agua.
3.2. La cava cerrada o el cantero debe colocarse en un cuarto
donde sea posible mantener una temperatura constante. Como
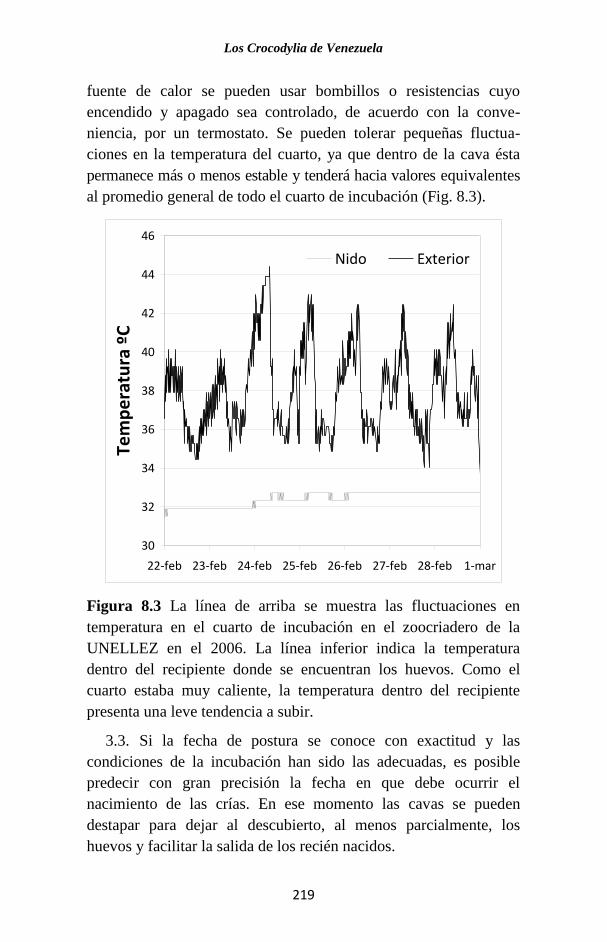
Los Crocodylia de Venezuela
219
fuente de calor se pueden usar bombillos o resistencias cuyo
encendido y apagado sea controlado, de acuerdo con la conve-
niencia, por un termostato. Se pueden tolerar pequeñas fluctua-
ciones en la temperatura del cuarto, ya que dentro de la cava ésta
permanece más o menos estable y tenderá hacia valores equivalentes
al promedio general de todo el cuarto de incubación (Fig. 8.3).
Figura 8.3 La línea de arriba se muestra las fluctuaciones en
temperatura en el cuarto de incubación en el zoocriadero de la
UNELLEZ en el 2006. La línea inferior indica la temperatura
dentro del recipiente donde se encuentran los huevos. Como el
cuarto estaba muy caliente, la temperatura dentro del recipiente
presenta una leve tendencia a subir.
3.3. Si la fecha de postura se conoce con exactitud y las
condiciones de la incubación han sido las adecuadas, es posible
predecir con gran precisión la fecha en que debe ocurrir el
nacimiento de las crías. En ese momento las cavas se pueden
destapar para dejar al descubierto, al menos parcialmente, los
huevos y facilitar la salida de los recién nacidos.
30
32
34
36
38
40
42
44
46
22-feb 23-feb 24-feb 25-feb 26-feb 27-feb 28-feb 1-mar
Tem
pe
ratu
ra º
C
Nido Exterior

Andrés Eloy Seijas
220
En la Figura 8.4 se muestran fotografías que ilustran distintas
etapas del proceso de colecta e incubación de los huevos de C.
intermedius en el zoocriadero de la UNELLEZ.
4. Crianza de individuos
4.1. Es recomendable mantener los individuos recién nacidos en
recipientes pequeños, con un poco de agua, por unas 24 a 48 horas
antes de trasladarlos a las tanquillas o estanques de crianza. El
agua debe ser renovada con frecuencia. Es conveniente, incluso,
mantenerlos durante ese periodo en el cuarto de incubación, a
temperaturas cercanas a 32 ºC, para acelerar la absorción del saco
vitelino. Esta medida facilita la inspección de los individuos para la
atención inmediata de cualquier problema que se presente en los
primeros días después de la eclosión.
4.2. Inmediatamente después del paso anterior, los animales deben
ser trasladados a las tanquillas. Se les debe colocar comida molida o
cortada en trozos que puedan ser tragados por ellos sin dificultad. No
todos los animales comenzaran a comer de inmediato. Se puede forzar
la alimentación en unos cuantos animales, para acelerar el aprendizaje
en el consumo del alimento. A las dos semanas ya todos los animales
deberían estar alimentándose por si solos.
4.3. Las características de la dieta a ser suministrada dependerá
del tamaño de los animales y del tipo de comida disponible en el
área donde se encuentre el zoocriadero: Carne de burro, caballo o
ganado vacuno, pescado (de mar o de río), restos de aves
(mollejas, pescuezos). Se debe evitar, sin embargo, la monodieta y
el exceso de grasa. Es indispensable suministrar complementos
minerales y vitamínicos para evitar la aparición de enfermedades
por deficiencias nutricionales (Boede y Sogbe 2000).
4.4. La cantidad de alimento proporcionado a la semana debe
equivaler a 15-20% del peso de los individuos (Joanen y McNease,
1976; Hutton y Webb, 1992). Esa cifra puede variar dependiendo
de la temperatura ambiental y la época de año. Los animales
tienden a comer más en los meses calurosos, por lo que la

Los Crocodylia de Venezuela
221
cantidad debe ser ajustada frecuentemente. Se recomienda una
frecuencia de alimentación diaria (lunes a viernes) durante los
primeros seis meses, que puede ser reducida a tres veces por semana
después de esa edad. Hay que destacar que Pérez-Talavera (1999,
2000, 2005) logró tasas de crecimiento extraordinarias con un
régimen de alimentación mucho más intensivo (todos los días,
incluyendo sábados y domingos) que el recomendado aquí.
4.5. La limpieza periódica de las tanquillas o piletas de levante
es un elemento clave para evitar enfermedades. La frecuencia con
que ésta se realiza dependerá de las características de las
instalaciones, pero se recomienda un mínimo de tres veces por
semana. Se debe ofrecer el alimento a los animales de manera de
evitar al máximo el despilfarro y, en particular, que éste caiga en el
agua (donde por lo general no es comido). La acumulación de
restos de comida en el agua constituye una fuente de conta-
minación y hace necesario una mayor frecuencia en la limpieza. En
la UNELLEZ se coloca el alimento en bandejas de aluminio, lo
cual reduce la caída de éste al agua. Cuando se efectúa la
limpieza de los estanques los animales tienden a amontonarse y
frecuentemente, especialmente si el agua no les cubre el cuerpo,
estallan peleas que pueden producir heridas profundas y fracturas
en la mandíbula.
4.6. Pérez-Talavera (1999, 2000) y Pérez-Talavera y Rodríguez
(2005) comprobaron que el crecimiento de los crocodilios es mayor
en tanquillas completamente cubiertas, donde es posible mantener el
agua a temperaturas más altas, lo que acelera la absorción del
alimento suministrado y su conversión en masa corporal. Es posible
que el aislamiento de perturbaciones externas que permiten estos
recintos ayude a disminuir el estrés y se traduzca en tasas de
crecimiento mayores. Como la construcción de encierros comple-
tamente cerrados es costosa, se puede manejar, como alternativa
para reducir el estrés, la colocación de láminas de zinc unos 10 a
20 cm. por arriba de la superficie del agua. Estas estructuras
sirven de refugio a los animales y los aísla un poco de pertur-
baciones externas y, quizá, permiten mantener temperaturas del
agua más altas y constantes.

Andrés Eloy Seijas
222
Figura 8.4 Fotografías de distintas etapas del proceso de colecta e
incubación de huevos de C. intermedius en el zoocriadero de la
UNELLEZ. A, apariencia de una nidada al momento de exponerla para su
colecta. B, Huevo con banda incipiente. C y D, huevos colocados en la
bandeja. E, Huevos en el recipiente dentro del cuarto de incubación; se
observan los envases donde se colocaron los termómetros que registraron
la temperatura durante todo el proceso. F, Neonato saliendo del huevo.
B
D

Los Crocodylia de Venezuela
223
4.7. El diseño de los encierros para el levante queda abierto a
la creatividad del criador. En Venezuela se ha trabajado con
encierros redondos, cuadrados y rectangulares. No obstante, se
pueden adelantar algunas recomendaciones.
4.7.a. La superficie de los encierros debe estar dividida
en una parte con agua y otra seca. La proporción entre ellas
puede ser 50:50. Tanquillas rectangulares, con una parte seca
formando una franja en el medio y dos franjas con agua que la
flanquean, ofrecen la ventaja de que los animales se pueden
refugiar en una de las franjas con agua mientras la otra es
limpiada. Esto disminuye la probabilidad de que los animales se
peleen entre sí cuando se amontonan en un lugar seco y se
inflijan heridas, a veces fatales.
4.7.b. La profundidad máxima de la parte con agua
dependerá de hasta qué tamaño se pretenda levantar a los
animales. Para animales de hasta dos años es suficiente una
profundidad de 40 cm.
4.7.c. La parte con agua debe contener un rampa, que
disminuya gradualmente la profundidad desde la parte más honda
hasta la parte seca del encierro. Esto facilita el acceso a la
comida y permite a los animales descansar con el cuerpo
cubierto de agua y apoyados sobre el fondo, con gasto mínimo
de energía.
4.7.d. El sistema de drenaje de las piletas es un aspecto
clave. Es preferible construir encierros sobre la superficie del
terreno (no en excavaciones) y con sistemas de drenaje también
superficiales, lo que facilita su mantenimiento y reparación
4.8. Se debe evitar el hacinamiento. Sin embargo, no existe una
cifra exacta a la hora de recomendar la densidad óptima de
animales por tanquillas. La especie con que se trabaja, la forma de
la tanquilla, la relación superficie con agua/superficie seca, la
profundidad del agua y el manejo, pueden tener algún tipo de
incidencia sobre el número de animales que se puede mantener por

Andrés Eloy Seijas
224
metro cuadrado. Pérez-Talavera (1999) obtuvo crecimientos muy
buenos en caimanes del Orinoco mantenidos cerca de un año a
una densidad de 0,71 individuos por m2. En el caso de la baba, la
misma autora (Pérez-Talavera, 2000) logró los mejores resultados
con densidades de 1,10 ind/m2, aunque no muy alejados de aquellos
obtenidos con densidades de 1,80 ind/m2.
Las recomendaciones enumeradas anteriormente no pretenden
ser rígidas ni absolutas, existe excelente documentación sobre la
cría de crocodílidos con fines comerciales, una actividad en
expansión a escala mundial pero que, por razones que no se
pretenden analizar en profundidad aquí, ha fracasado en Venezuela.
La cría de crocodilios que ha tenido mayor continuidad en
Venezuela es la que se ha desarrollado con fines de conservación,
para la producción de individuos para la restauración o fomento de
poblaciones. Es este tipo de experiencias el que será objeto de
discusión en las siguientes secciones.
CRIANZA DE CROCODILIOS CON FINES DE
REPOBLACIÓN
El levante o crianza de crocodilios en cautiverio para su
posterior liberación en el ambiente natural ha sido una estrategia de
conservación varias veces propuesta y practicada en Venezuela
(Blohm, 1973; Chávez, 2000; Espinoza, 1988; Lugo, 1998; Muñoz
y Thorbjarnarson, 2000; Ramo et al., 1992; Rivero-Blanco, 1974;
Seijas, 1995; Seijas et. al., 1990). Esta estrategia se fundamenta en
el hecho de que en su medio natural todos los crocodilios presentan
una tasa de mortalidad muy alta (mayor de 95%) en los primeros
meses de vida, etapa en que los recién nacidos son fáciles presas
de un gran número de depredadores.
El levante en condiciones controladas de individuos nacidos en
cautividad, o colectados en el ambiente natural en las primeras
horas después de la eclosión, aumenta la probabilidad de sobre-
vivencia de éstos. Los ejemplares son mantenidos en los centros de

Los Crocodylia de Venezuela
225
levante durante un tiempo que les permite alcanzar una talla y peso
adecuados para su liberación, es decir, suficientemente grandes
como para reducir su vulnerabilidad ante gran parte de sus
depredadores naturales.
CAIMÁN DEL ORINOCO
Ya para el año 1986 existían en Venezuela tres zoocriaderos de
caimanes, en los hatos El Frío (Apure) y Masaguaral (Guárico) y
en la UNELLEZ (Portuguesa). No obstante, la primera liberación
formal de individuos criados en cautiverio se efectuó el 17 de abril
de 1990, cuando se introdujeron al medio natural, en el hato El
Frío, los primeros 31 individuos. En 1991 se incorpora un nuevo
centro de cría, la Agropecuaria Puerto Miranda (APM, Guárico) y
en el 2004 inicia operaciones un último zoocriadero en Puerto
Ayacucho, de la Fundación para el Desarrollo de las Ciencias
(FUDECI). El Ministerio del Ambiente ha criado también caimanes
del Orinoco en las instalaciones de la Estación Biológica de
Rancho Grande (El Limón, Maracay), lugar poco propicio para esa
tarea. Hasta junio de 2010 se habían liberado poco más de 7.600
individuos provenientes de esos zoocriaderos en un total de 14
localidades de los llanos (Fig. 8.5).
La talla promedio de los primeros individuos liberados fue
cercana a 150 cm. de LT, por tratarse de caimanes de tres o más
años de edad (Fig. 8.6). De ese año en adelante las tallas al liberar
han disminuido, ya que corresponden principalmente a ejemplares de
alrededor de un año de edad. Sólo el hato El Frío ha liberado, de
manera rutinaria y mayoritaria, individuos de dos o más años.
La caída en la talla promedio para 1992, fue consecuencia de la
liberación de 259 caimanes de sólo seis meses de edad en el río
Capanaparo ese año. El menor promedio de 1997 pudiera ser
consecuencia de falta de datos, ya que uno solo de los zoo-
criaderos (Masaguaral) reportó haber liberado caimanes ese año. El
gráfico muestra que la talla de liberación en los últimos años ha
declinado de manera progresiva, tendencia que es necesario corregir.
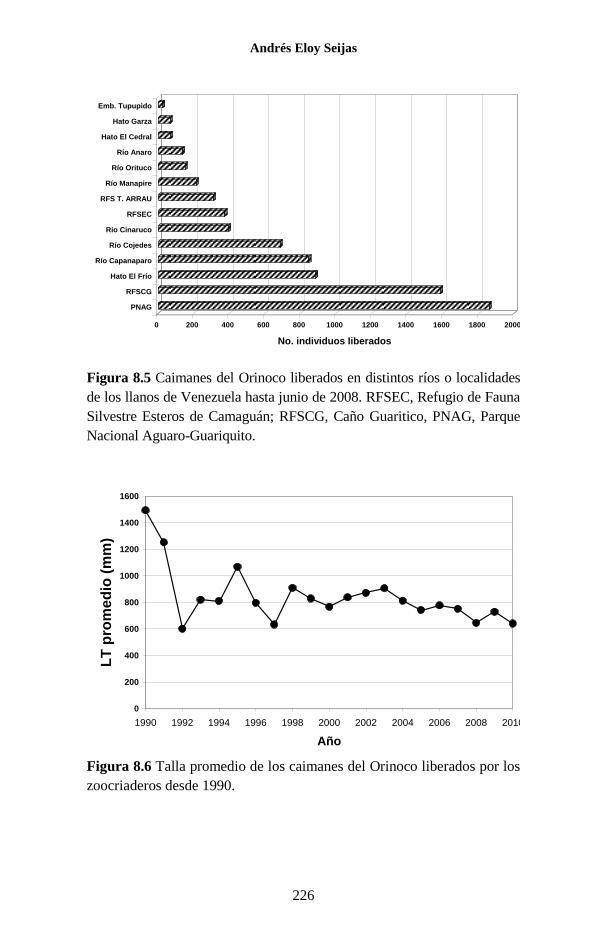
Andrés Eloy Seijas
226
Figura 8.5 Caimanes del Orinoco liberados en distintos ríos o localidades
de los llanos de Venezuela hasta junio de 2008. RFSEC, Refugio de Fauna
Silvestre Esteros de Camaguán; RFSCG, Caño Guaritico, PNAG, Parque
Nacional Aguaro-Guariquito.
Figura 8.6 Talla promedio de los caimanes del Orinoco liberados por los
zoocriaderos desde 1990.
0 200 400 600 800 1000 1200 1400 1600 1800 2000
No. individuos liberados
PNAG
RFSCG
Hato El Frío
Río Capanaparo
Río Cojedes
Río Cinaruco
RFSEC
RFS T. ARRAU
Río Manapire
Río Orituco
Río Anaro
Hato El Cedral
Hato Garza
Emb. Tupupido
0
200
400
600
800
1000
1200
1400
1600
1990 1992 1994 1996 1998 2000 2002 2004 2006 2008 2010
Año
LT
pro
me
dio
(m
m)
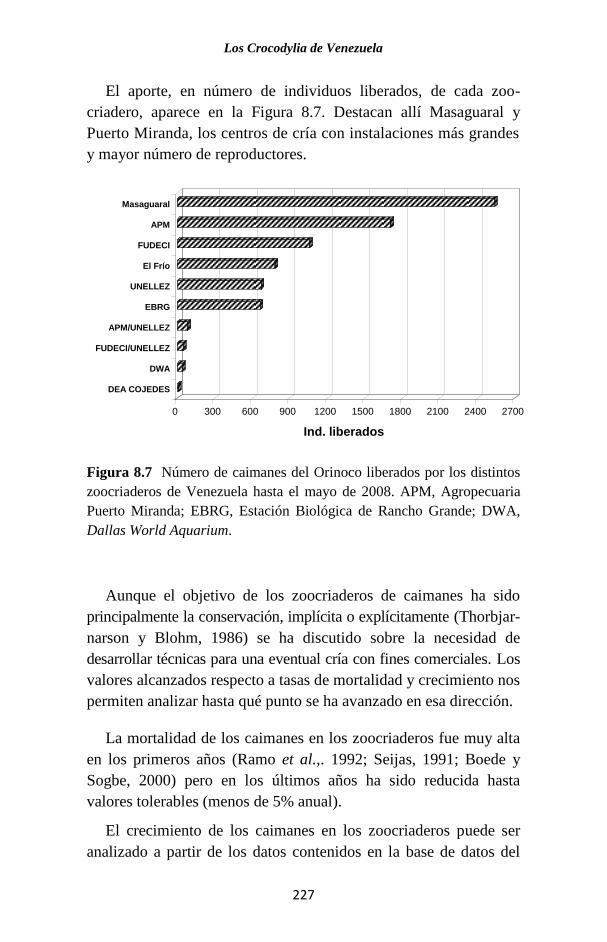
Los Crocodylia de Venezuela
227
El aporte, en número de individuos liberados, de cada zoo-
criadero, aparece en la Figura 8.7. Destacan allí Masaguaral y
Puerto Miranda, los centros de cría con instalaciones más grandes
y mayor número de reproductores.
Figura 8.7 Número de caimanes del Orinoco liberados por los distintos
zoocriaderos de Venezuela hasta el mayo de 2008. APM, Agropecuaria
Puerto Miranda; EBRG, Estación Biológica de Rancho Grande; DWA,
Dallas World Aquarium.
Aunque el objetivo de los zoocriaderos de caimanes ha sido
principalmente la conservación, implícita o explícitamente (Thorbjar-
narson y Blohm, 1986) se ha discutido sobre la necesidad de
desarrollar técnicas para una eventual cría con fines comerciales. Los
valores alcanzados respecto a tasas de mortalidad y crecimiento nos
permiten analizar hasta qué punto se ha avanzado en esa dirección.
La mortalidad de los caimanes en los zoocriaderos fue muy alta
en los primeros años (Ramo et al.,. 1992; Seijas, 1991; Boede y
Sogbe, 2000) pero en los últimos años ha sido reducida hasta
valores tolerables (menos de 5% anual).
El crecimiento de los caimanes en los zoocriaderos puede ser
analizado a partir de los datos contenidos en la base de datos del
0 300 600 900 1200 1500 1800 2100 2400 2700
Ind. liberados
DEA COJEDES
DWA
FUDECI/UNELLEZ
APM/UNELLEZ
EBRG
UNELLEZ
El Frío
FUDECI
APM
Masaguaral

Andrés Eloy Seijas
228
Grupo de Especialistas en Cocodrilos de Venezuela (GECV) y si
se toman en cuenta sólo las tasas de crecimiento (cm/mes) de los
individuos liberados de edad conocida y menores de dos años
(Tabla 8.1). El centro de crianza con el mejor desempeño en este
aspecto ha sido la Agropecuaria Puerto Miranda, en el cual los
caimanes han crecido a una tasa promedio de 4,62 cm/mes.
No hay elementos para discriminar entre los distintos aspectos
que pudieran haber determinado las diferencias entre las cifras de
la Tabla 8.1. Probablemente las características de las instalaciones
sea el factor más importante. La APM posee las instalaciones más
adecuadas y amplias, lo que les ha permitido mantener a los
animales en bajas densidades. Ellos fueron además los primeros en
implementar un sistema de refugios dentro de las tanquillas, con
láminas de zinc colocadas sobre la parte con agua, para reducir el
estrés que produce en los animales las labores rutinarias del
zoocriadero o la presencia esporádica de visitantes. Los muy bajos
valores de FUDECI en su primer año de funcionamiento pudieran
reflejar una etapa de aprendizaje por parte de los encargados del
mantenimiento, pero también que el origen de los caimanes allí
mantenidos fue muy diverso, y muchos de los individuos perma-
necieron relativamente hacinados o sin la adecuada atención en
otros lugares antes de su traslado a Puerto Ayacucho.
El crecimiento de los crocodilios en cautiverio depende de
diversos factores como calidad y cantidad del alimento suministrado,
manejo sanitario, temperatura del agua, densidad de individuos en
las tanquillas, sexo, carga genética y quizá algunos otros. Incluso
las condiciones en que se desarrolla el embrión durante la
incubación del huevo, tienen consecuencias tanto en el sexo como
en la potencialidad de crecer después del nacimiento (Ferguson
1987; Ferguson y Joanen, 1983; Joanen et al., 1987; Hutton y
Webb, 1992). No es fácil controlar todas las variables señaladas,
pero un análisis más detallado de los resultados obtenidos por los
zoocriaderos permite establecer que, para al menos una fracción de
la población, los resultados fueron adecuados. Así se tiene que 84
individuos de la APM (7,7%) crecieron a tasas iguales o superiores
a 6 cm/mes. Esa cifra fue de sólo 4,5% para el zoocriadero de
Masaguaral, mientras que los otros centros de cría no alcanzaron
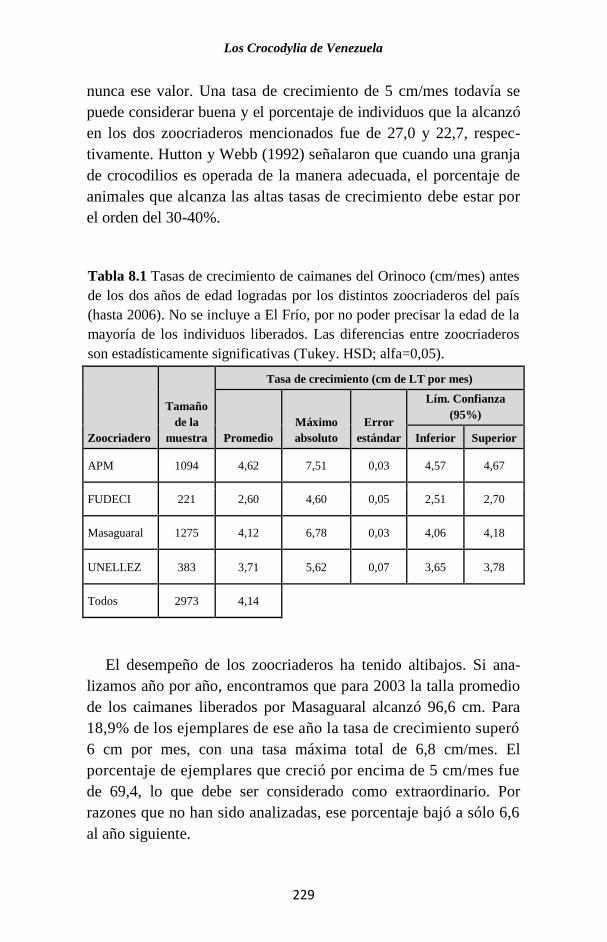
Los Crocodylia de Venezuela
229
nunca ese valor. Una tasa de crecimiento de 5 cm/mes todavía se
puede considerar buena y el porcentaje de individuos que la alcanzó
en los dos zoocriaderos mencionados fue de 27,0 y 22,7, respec-
tivamente. Hutton y Webb (1992) señalaron que cuando una granja
de crocodilios es operada de la manera adecuada, el porcentaje de
animales que alcanza las altas tasas de crecimiento debe estar por
el orden del 30-40%.
Tabla 8.1 Tasas de crecimiento de caimanes del Orinoco (cm/mes) antes
de los dos años de edad logradas por los distintos zoocriaderos del país
(hasta 2006). No se incluye a El Frío, por no poder precisar la edad de la
mayoría de los individuos liberados. Las diferencias entre zoocriaderos
son estadísticamente significativas (Tukey. HSD; alfa=0,05).
Zoocriadero
Tamaño
de la
muestra
Tasa de crecimiento (cm de LT por mes)
Promedio
Máximo
absoluto
Error
estándar
Lím. Confianza
(95%)
Inferior Superior
APM 1094 4,62 7,51 0,03 4,57 4,67
FUDECI 221 2,60 4,60 0,05 2,51 2,70
Masaguaral 1275 4,12 6,78 0,03 4,06 4,18
UNELLEZ 383 3,71 5,62 0,07 3,65 3,78
Todos 2973 4,14
El desempeño de los zoocriaderos ha tenido altibajos. Si ana-
lizamos año por año, encontramos que para 2003 la talla promedio
de los caimanes liberados por Masaguaral alcanzó 96,6 cm. Para
18,9% de los ejemplares de ese año la tasa de crecimiento superó
6 cm por mes, con una tasa máxima total de 6,8 cm/mes. El
porcentaje de ejemplares que creció por encima de 5 cm/mes fue
de 69,4, lo que debe ser considerado como extraordinario. Por
razones que no han sido analizadas, ese porcentaje bajó a sólo 6,6
al año siguiente.

Andrés Eloy Seijas
230
Los valores óptimos discutidos anteriormente nos indican cual
sería el potencial de levante en condiciones ideales. Con cifras
como esas se levantarían animales de hasta un metro o más al
año. Como el crecimiento en los dos primeros años es esencial-
mente lineal, se podrían obtener animales entre 1,40 y 1,60 metros
en dos años, una talla adecuada quizá para el aprovechamiento
comercial.
Pérez-Talavera (1999) reportó datos de crecimiento para C.
intermedius en condiciones de cuidado sanitario, alimentación y
temperatura difícilmente superables. La manera como ella presentó
sus datos, sin embargo, no permite una comparación directa con los
valores alcanzados por los otros zoocriaderos. Al re-analizar los
resultados presentados por esta autora se pueden calcular tasas de
crecimiento mensual que van desde 4,53 hasta 6,15 cm/mes. Esta
última cifra fue alcanzada para un lote de 40 caimanes mantenidos
con un régimen de alimentación diaria, en una tanquilla comple-
tamente cerrada y por un lapso de 5,5 meses. Una tasa de cre-
cimiento ligeramente menor (6,07 cm/mes) fue obtenida para un
grupo de 18 animales mantenidos en condiciones similares a las
señaladas, pero por un periodo de 11,5 meses.
Tanto los valores más altos de la APM y Masaguaral, como los
resultados de Pérez-Talavera (1999) nos indican que es posible
hacer crecer a los caimanes a tasas muy altas, comparables a las
mejores reportadas en la literatura (Hutton y Webb, 1992). Inves-
tigaciones futuras tendrán que determinar la factibilidad de lograrlo
de manera económicamente rentable, si el objetivo principal del
zoocriadero es el de producir pieles y carne para el mercado.
Monitoreo de las liberaciones
Con la excepción de ocho caimanes liberados en el río Capa-
naparo en 1991, los cuales fueron seguidos (radio-telemetría) de
manera continua durante once meses (Muñoz y Thorbjarnarson
2000), la mayor parte de las liberaciones no han recibido un
monitoreo adecuado. Se han realizado sólo tres estudios con un
esfuerzo de muestreo relativamente intenso, dos de ellos en el RFS

Los Crocodylia de Venezuela
231
de Caño Guaritico y zonas aledañas del hato El Frío (Lugo, 1998;
Chávez, 2000) y el tercero en el río Manapire del estado Guárico
(Jiménez-Oraá, 2005). Los indicadores de éxitos de estas libera-
ciones se pueden obtener a partir de la fracción de individuos
sobrevivientes, de las tasas de crecimiento corporal que experi-
mentaron en su nuevo ambiente, y en la eventual reproducción de
los caimanes liberados.
Sobrevivencia en el ambiente natural
En primer lugar hay que aclarar que aunque se usará el término
“sobrevivencia”, quizá en algunos casos la expresión “permanencia”
podría ser más adecuada, ya que es posible que algunos individuos
abandonen el área de estudio y escapen a la detección de los
investigadores.
El seguimiento realizado por Muñoz y Thorbjarnarson (2000)
mostró que los individuos liberados (con tallas comprendidas entre
los 103,3-139,3 de LT) se movieron considerablemente durante los
primeros meses (hasta 11,6 km. del lugar de liberación) para luego
ser más sedentarios y establecerse en un zona. Sin contar un
ejemplar para el cual el radiotransmisor falló a los 253 días, sólo
uno de los individuos liberados murió (cazado por un indígena)
durante los 352 días del estudio. Esto indica una sobrevivencia sor-
prendente de, al menos 75%. Una sobrevivencia de 100% cuatro
años después de la liberación, aunque basada en sólo cuatro
animales, se comprobó en caimanes liberados en el caño Matiyure
del hato El Cedral, Apure (Lugo, 1998).
Lugo (1998), con base en el número de caimanes observados
durante conteos diurnos y nocturnos, calculó fracciones de perma-
nencia desde 8,9 hasta 12,2% en el caño Guaritico y zonas
aledañas. En las mismas localidades Chávez (2000), utilizando una
metodología similar, calculó una permanencia de 7% para caimanes
liberados entre dos y ocho años antes de su estudio, y de 16%
para caimanes liberados entre 6 y ocho meses previos a su eva-
luación. El método para calcular estas permanencias-sobrevivencias
es mucho menos eficiente que el de la radio-telemetría empleado

Andrés Eloy Seijas
232
por Muñoz y Thorbjarnarson (2000) y por lo tanto, los valores
señalados probablemente representan sub-estimaciones
En el río Capanaparo (Apure) se liberaron 456 caimanes con
talla promedio de 65 cm de LT en los años 1992 y 1993. Con los
datos obtenidos por Llobet (2002) se pudo determinar que los
caimanes sobrevivientes de esas liberaciones representaban, por lo
menos, 26% de los individuos mayores a 1,80 m observados en
sectores de dicho río muestreados en el año 2001.
Crecimiento en el ambiente natural
El crecimiento corporal experimentado por los caimanes en su
nuevo ambiente, constituye otra manera de medir el éxito de las
liberaciones. Muñoz y Thorbjarnarson (2000) reportaron que la tasa
de crecimiento promedio de los caimanes liberados en el río Capa-
naparo (0,079 cm LT/día) fue similar a los de los animales
silvestres. Los datos aportados por varios investigadores permiten
comparar el crecimiento de los caimanes recapturados en distintas
localidades (Tabla 8.2).
De los datos presentados en la Tabla 8.2 hay que destacar su
gran variabilidad. A ella contribuye que los lapsos agrupados son
muy amplios, además de que no se discriminó a los animales de
acuerdo con el sexo. Sin embargo, parte de las diferencias pueden
estar relacionadas con las características ecológicas de los ríos y la
calidad de hábitat que ellos representan para los caimanes. Las
tasas máximas de crecimiento de Guaritico y Macanillal, por
ejemplo, son mucho mayores que la de los ríos Cinaruco-
Capanaparo. Esto es de esperar si se considera que estos últimos
ríos son de aguas claras y, por lo tanto, poco productivos, como se
discutió en el Capítulo 5.
Los valores de crecimiento para el caño Matiyure del hato El
Cedral son extraordinarios, y quizás sean parcialmente consecuencia
del hecho de que esos caimanes fueron alimentados eventualmente
por turistas (J. Thorbjarnarson, com, pers.). Con tasas de cre-
cimiento como las reportadas, los individuos liberados pueden
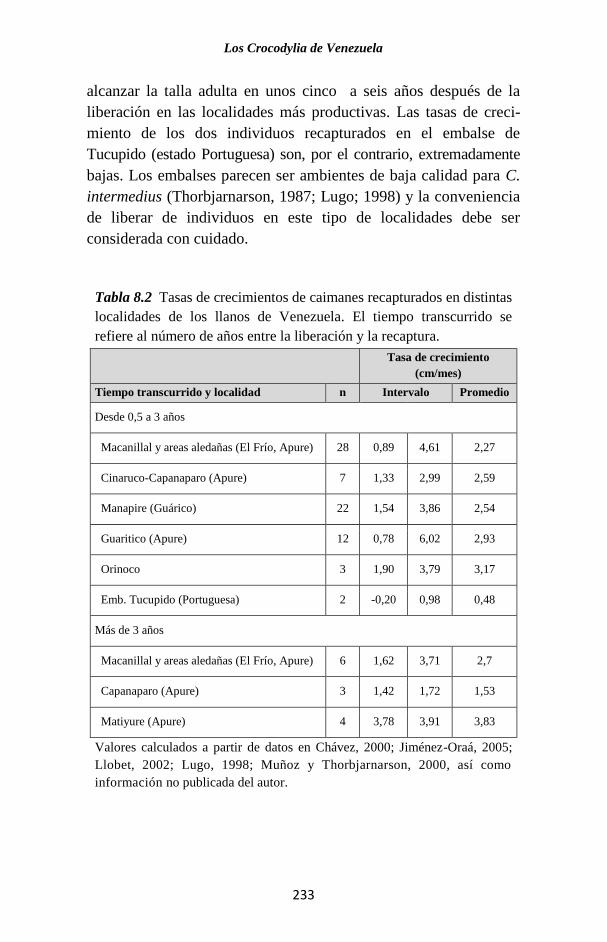
Los Crocodylia de Venezuela
233
alcanzar la talla adulta en unos cinco a seis años después de la
liberación en las localidades más productivas. Las tasas de creci-
miento de los dos individuos recapturados en el embalse de
Tucupido (estado Portuguesa) son, por el contrario, extremadamente
bajas. Los embalses parecen ser ambientes de baja calidad para C.
intermedius (Thorbjarnarson, 1987; Lugo; 1998) y la conveniencia
de liberar de individuos en este tipo de localidades debe ser
considerada con cuidado.
Tabla 8.2 Tasas de crecimientos de caimanes recapturados en distintas
localidades de los llanos de Venezuela. El tiempo transcurrido se
refiere al número de años entre la liberación y la recaptura.
Tasa de crecimiento
(cm/mes)
Tiempo transcurrido y localidad n Intervalo Promedio
Desde 0,5 a 3 años
Macanillal y areas aledañas (El Frío, Apure) 28 0,89 4,61 2,27
Cinaruco-Capanaparo (Apure) 7 1,33 2,99 2,59
Manapire (Guárico) 22 1,54 3,86 2,54
Guaritico (Apure) 12 0,78 6,02 2,93
Orinoco 3 1,90 3,79 3,17
Emb. Tucupido (Portuguesa) 2 -0,20 0,98 0,48
Más de 3 años
Macanillal y areas aledañas (El Frío, Apure) 6 1,62 3,71 2,7
Capanaparo (Apure) 3 1,42 1,72 1,53
Matiyure (Apure) 4 3,78 3,91 3,83
Valores calculados a partir de datos en Chávez, 2000; Jiménez-Oraá, 2005;
Llobet, 2002; Lugo, 1998; Muñoz y Thorbjarnarson, 2000, así como
información no publicada del autor.

Andrés Eloy Seijas
234
Reproducción en el ambiente natural de los
individuos liberados
Como consecuencia de las tasas de crecimiento ya discutidas en
párrafos anteriores, gran parte de los caimanes liberados en los
primeros años (entre 1990 y 1995) ya han alcanzado la talla
reproductiva. La primera evidencia de reproducción se obtuvo en
1997 (Lugo, 1998) en el hato El Frío. Durante la temporada
reproductiva de 2001 se comprobó la anidación de 11 hembras en
los alrededores del caño Macanillal del hato El Frío (M. Trillo,
com. pers.) y de tres hembras en el hato El Cedral (C. Chávez,
com. pers.). Antelo et al. (2010) reportaron un total acumulado de
69 nidos para las temporadas 2003-2007, con un máximo de 19
nidos en el año 2006. Existe también información de que ya hay
hembras reproductivamente activas en el Caño Guaritico (Rafael
Antelo, com. pers.) lugar donde se han liberado, hasta el 2011,
cerca de 1600 de los caimanes, por lo cual se deberían intensificar las
actividades de monitoreo en esta localidad en los próximos años.
CAIMÁN DE LA COSTA
Los esfuerzos de cría de C. acutus con fines de restauración o
fomento poblacional son más modestos que los que han sido
adelantados con el caimán del Orinoco. La primera liberación de
individuos de esta especie se llevó a cabo el 26-feb-1986, con
animales cercanos a los dos años que habían sido capturados
recién nacidos en el río Yaracuy en 1984 (Seijas et al., 1990).
Estos 19 primeros individuos se soltaron en los embalses de
Tacarigua y Jatira, del estado Falcón. Al año siguiente se liberaron
nueve caimanes en el embalse de Pueblo Viejo, pero en este caso
los individuos provenían de esta misma localidad, donde habían sido
capturados como crías en 1986.
A partir de 1991 se intensifica un poco el programa de
liberación cuando se comienzan a soltar caimanes hijos de una
pareja de C. acutus mantenida en el hato Masaguaral. Aunque la
base de datos del GECV es un poco imprecisa, se puede asegurar
Los Crocodylia de Venezuela
235
que al menos 312 de los 704 caimanes de la costa (44,3%)
liberados hasta el 2011, son hijos de la misma pareja antes
mencionada. En el año 1995 el MARN inició un programa de
captura de neonatos y juveniles en la laguna de Turiamo, en el
estado Aragua (González-Fernández, 1999; Lander, 2003) para su
levante en cautiverio y posterior liberación. La cifra de individuos
soltados en el medio natural provenientes de esta localidad es de
124 (17,6% respecto al total).
La localidad que ha recibido la mayor parte de los individuos
liberados es el Refugio de Fauna Silvestre de Cuare, en el estado
Falcón (37,5%), seguido por el Refugio de Fauna Silvestre de los
Olivitos en el estado Zulia (19,0%) y el Río Santa Rosa, también
en el estado Zulia (16,8%) (Fig. 8.8).
Figura 8.8 Caimanes de la costa liberados en distintas localidades de la
región costera de Venezuela hasta el año 2011.
0 50 100 150 200 250 300
RFS Cuare (Falcón)
RFS Los Olivitos (Zulia)
Río Santa Rosa (Zulia)
Emb. Jatira-Tacarigua (Falcón)
Emb. Machango (Zulia)
PN Laguna Tacarigua (Miranda)
RFS Tucurere (Falcón)
Turiamo (Aragua)
Río Negro (Zulia)
Emb. Pueblo Viejo (Zulia)
No. individuos liberados

Andrés Eloy Seijas
236
El programa de fomento y restauración poblacional con el
caimán de la costa ha sido poco evaluado. Cuatro de los primeros
animales liberados fueron recapturados en 1987 y 1988 y mostraron
tasas de crecimiento similares a la de los animales silvestres y
condición física aceptables (Seijas et al., 1990). Arteaga (1991)
encontró que siete de 25 caimanes recapturados en el embalse de
Tacarigua mostraron tasas de crecimiento promedio de 3,63 cm. al
mes, superiores a las que habían experimentado en la etapa previa
en cautiverio (2,72 cm/mes). Urdaneta (2002), en recorridos
nocturnos en Los Olivitos contó un máximo de 15 ejemplares, una
cifra baja si se tiene en cuenta que para el momento de su
evaluación se habían liberado 45 individuos en el refugio, pero hay
que considerar que la fracción de cocodrilos observados en reco-
rridos nocturnos puede ser tan baja como 10% (ver Capítulo 5).
Un solo ejemplar ha sido recapturado en esta última localidad, no
obstante el esfuerzo importante de muestreo que se ha llevado a
cabo (Barros et al., 2005).
El aspecto más crítico del programa con C. acutus es el
altísimo porcentaje de animales hermanos que se han liberado, lo
cual podría traer consecuencias genéticas indeseables en el futuro.
Otro punto que hay que revisar, para evitar que ocurra de nuevo,
es que en el programa que se desarrolló en Turiamo varios de los
animales capturados en dicha localidad, para su traslado al criadero
de El Limón y de allí a otros lugares del país, eran en su mayoría
animales de tallas comprendidas entre 60 y 95 cm de LT
(González-Fernández, 1999; Lander, 2003), es decir, caimanes que
habían sobrevivido su etapa más vulnerable, algunos de ellos con
seguridad mayores de dos años y por lo tanto ya bien establecidos
en la población. Tal vez se partió de una idea preconcebida de que
los caimanes de Turiamo debían ser “rescatados”, cuando lo
apropiado era su protección e, incluso, el fomento de su población.
BABA
En el marco del programa de aprovechamiento comercial de
babas silvestres que comenzó en Venezuela en 1982, se planteó la

Los Crocodylia de Venezuela
237
necesidad de establecer estrategias para mitigar el impacto que la
extracción de individuos pudiera causar sobre las poblaciones
naturales. De esta manera, se estableció en la normativa legal que
cada zoocriadero debería liberar a la vida silvestre una cuota anual
(entre 2% y 10%) de los animales levantados en sus instalaciones,
provenientes de neonatos o huevos colectados en el medio natural
(Baquero y Quero, 1996). El fundamento de este requerimiento era
el de resarcir, de alguna manera, el impacto provocado por el
manejo de la población silvestre. Esa normativa se cumplió de
manera parcial. Aunque no hubo un seguimiento sistemático de sus
resultados, las primeras experiencias reportadas mostraron resul-
tados desastrosos. Espinoza (1989) indicó, por ejemplo, que una
primera liberación de babas criadas en cautiverio sufrió de manera
inmediata una alta mortalidad (estimada en 5%) por ataques de
caribes y babas adultos. Una segunda liberación se llevó a cabo en
un préstamo que tenía babas adultas, una de las cuales fue
posteriormente cazada para el aprovechamiento del cuero. En su
estómago se encontraron las placas de, al menos, 70 de los
individuos liberados en meses anteriores.
Una evaluación más detallada se llevó a cabo con 1287 babas
de dos o más años liberadas en el hato El Cedral, estado Apure
(Seijas et al., 2003). Un grupo de ellas fue soltado antes del
comienzo de la época seca (noviembre-diciembre de 1992) y otro a
comienzos del período de lluvias (abril-julio 1993) (Tabla 8.3).
La recaptura de 210 individuos, en lapsos comprendidos entre
los 26 y los 523 días, permitió determinar cambios en la gordura
relativa, incidencia de heridas y tasas de crecimiento en su hábitat
natural. Ciento quince (115) babas silvestres capturadas en el
mismo hato se usaron como grupo control. Un análisis con un
índice de gordura relativa (IGR) mostró que tanto las babas
silvestres (F=25,4; P<0,001) como las liberadas (F=15,4; P<0,001)
perdieron peso y crecieron a una tasa muy baja (0,257 cm. LCC/
mes) durante la época seca, comparado con el de aquellas
liberadas al comienzo del periodo de lluvias (0,65 cm. LCC/mes).
Las tasas de crecimiento fueron muy bajas, comparadas con las
que se obtienen en condiciones de cautiverio.
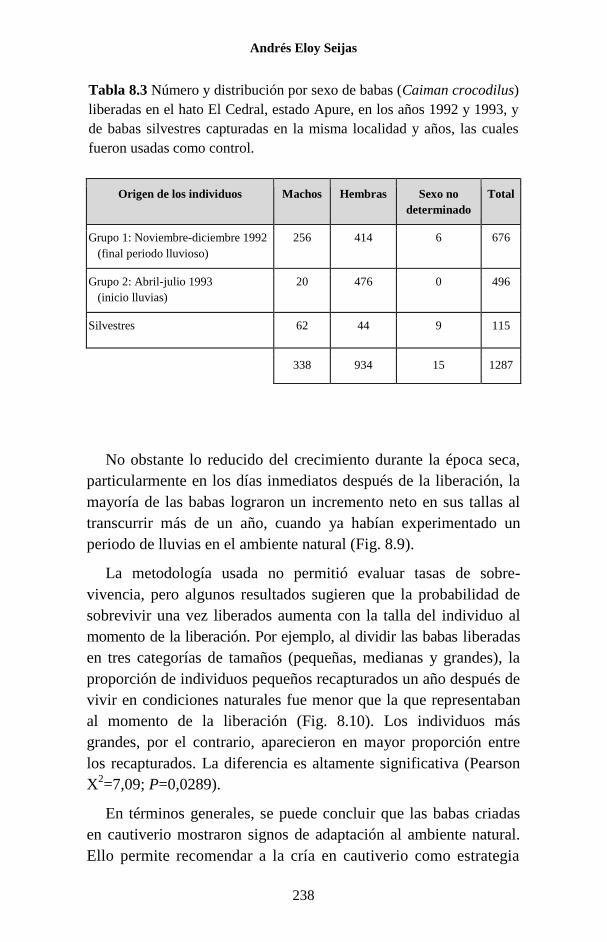
Andrés Eloy Seijas
238
Tabla 8.3 Número y distribución por sexo de babas (Caiman crocodilus)
liberadas en el hato El Cedral, estado Apure, en los años 1992 y 1993, y
de babas silvestres capturadas en la misma localidad y años, las cuales
fueron usadas como control.
Origen de los individuos Machos Hembras Sexo no
determinado
Total
Grupo 1: Noviembre-diciembre 1992
(final periodo lluvioso)
256 414 6 676
Grupo 2: Abril-julio 1993
(inicio lluvias)
20 476 0 496
Silvestres 62 44 9 115
338 934 15 1287
No obstante lo reducido del crecimiento durante la época seca,
particularmente en los días inmediatos después de la liberación, la
mayoría de las babas lograron un incremento neto en sus tallas al
transcurrir más de un año, cuando ya habían experimentado un
periodo de lluvias en el ambiente natural (Fig. 8.9).
La metodología usada no permitió evaluar tasas de sobre-
vivencia, pero algunos resultados sugieren que la probabilidad de
sobrevivir una vez liberados aumenta con la talla del individuo al
momento de la liberación. Por ejemplo, al dividir las babas liberadas
en tres categorías de tamaños (pequeñas, medianas y grandes), la
proporción de individuos pequeños recapturados un año después de
vivir en condiciones naturales fue menor que la que representaban
al momento de la liberación (Fig. 8.10). Los individuos más
grandes, por el contrario, aparecieron en mayor proporción entre
los recapturados. La diferencia es altamente significativa (Pearson
X2=7,09; P=0,0289).
En términos generales, se puede concluir que las babas criadas
en cautiverio mostraron signos de adaptación al ambiente natural.
Ello permite recomendar a la cría en cautiverio como estrategia
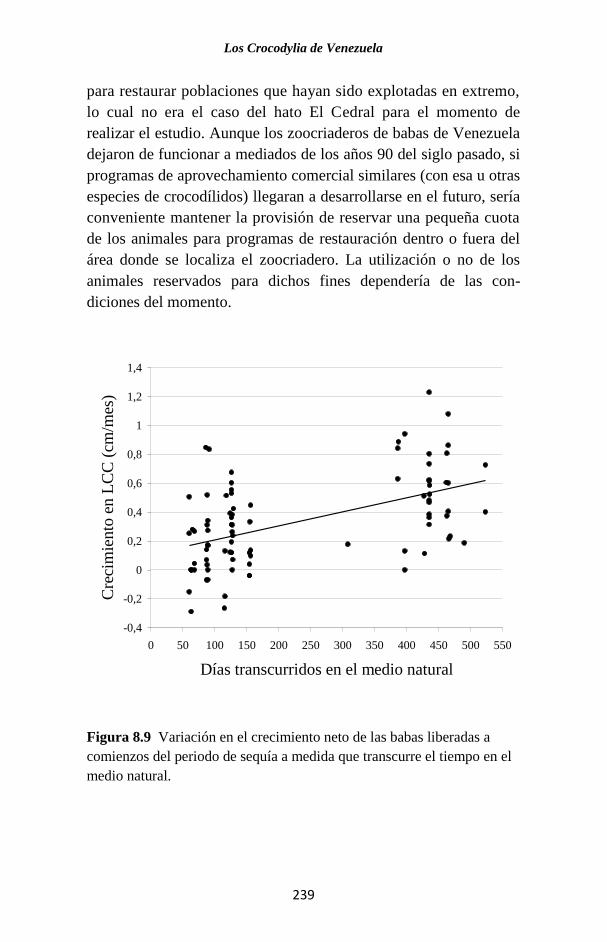
Los Crocodylia de Venezuela
239
para restaurar poblaciones que hayan sido explotadas en extremo,
lo cual no era el caso del hato El Cedral para el momento de
realizar el estudio. Aunque los zoocriaderos de babas de Venezuela
dejaron de funcionar a mediados de los años 90 del siglo pasado, si
programas de aprovechamiento comercial similares (con esa u otras
especies de crocodílidos) llegaran a desarrollarse en el futuro, sería
conveniente mantener la provisión de reservar una pequeña cuota
de los animales para programas de restauración dentro o fuera del
área donde se localiza el zoocriadero. La utilización o no de los
animales reservados para dichos fines dependería de las con-
diciones del momento.
Figura 8.9 Variación en el crecimiento neto de las babas liberadas a
comienzos del periodo de sequía a medida que transcurre el tiempo en el
medio natural.
-0,4
-0,2
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 50 100 150 200 250 300 350 400 450 500 550
Días transcurridos en el medio natural
Cre
cim
iento
en L
CC
(cm
/mes
)
Andrés Eloy Seijas
240
Figura 8.10 Proporción de babas en cada una de las categorías de
tamaño (mm de LCC) en que se dividió la muestra. La estructura de
tamaño de las babas capturadas un año después de la liberación es
distinta a la del momento de liberación, lo cual indica que las más
grandes tienen mayor probabilidad de sobrevivir.
0
10
20
30
40
50
60
Po
rcen
taje
de b
ab
as
Liberadas Más de un año
<300 mm LCC 300-399 mm LCC >=400 mm LCC

Los Crocodylia de Venezuela
241
REFERENCIAS
ÁLVAREZ DEL TORO, M.
1974. Los Crocodylia de México (Estudio comparativo). México, D.F., Instituto
Mexicano de los Recursos Naturales Renovables, A.C.
ANDRÉ, E.
1904. A naturalist in the Guianas. Thomas Nelson & Sons, London.
ANTELO, R.
2008. Biología del cocodrilo o caimán del Orinoco (Crocodylus intermedius) en la
Estación Biológica El Frío, Estado Apure (Venezuela). Tesis doctoral.
Facultad de Ciencias. Universidad Autónoma de Madrid, Madrid. 336 pp.
ANTELO, R., J. AYARZAGÜENA Y J. CASTROVIEJO
2010. Reproductive ecology of Orinoco crocodiles (Crocodylus intermedius) in
a newly established population at El Frío Biological Station, Venezuela.
Herpetol. J. 20:51-58.
APPUN, K. F.
1961 [1871]. En los trópicos. Universidad Central de Venezuela. Ediciones de la
Biblioteca. Universidad Central de Venezuela. Caracas.
ARTEAGA, A.
1989. Estudio prospectivo de las poblaciones de Caiman crocodilus Linneaus,
1758 (Crocodylia, Alligatoridae) del estado Guárico. Tesis de Licenciatura.
Escuela de Biología, Facutad de Ciencias. UCV, Caracas.
1997. Actualización de la situación poblacional de Crocodylus acutus y Caiman
crocodilus spp en las costas de Venezuela. Pp. 6-16 in: 4ta Reunión del
Grupo de Especialistas en Cocodrilos de América Latina y el Caribe,
Villahermosa, Tabasco, México.
1998. Aspectos bioecológicos de Crocodylus acutus liberados en el embalse
Tacarigua (Falcón, Venezuela), Tesis de Maestría. Universidad Simón
Bolívar, Caracas.

Andrés Eloy Seijas
242
ARTEAGA, A. Y F. GÓMEZ
2000. Recuperación de Crocodylus acutus en parques nacionales de Venezuela.
Pp. 336-352 in: Crocodiles. Proceedings of the 15th Working Meeting of
Crocodile Specialist Group-SSC. IUCN-The World Conservation Union
Gland, Switzerland.
ARTEAGA, A., A. E. SEIJAS, C. CHÁVEZ, Y J. THORBJARNARSON
1994. Status and conservation of the Orinoco crocodile. An update. Pp. 143-150 in:
Crocodiles. Proceedings of the 12th Working Meeting of the Crocodile
Specialist Group (Volumen 1) IUCN-The World Conservation Union,
Gland, Switzerland.
ARTEAGA, A., I. CAÑIZALES, G. HERNÁNDEZ, M. C. LAMAS, A. DE LUCA, M.
MUÑOZ, A. OCHOA, A. E. SEIJAS, J. B. THORBJARNARSON, A. VELASCO, S. ELLIS
Y U. SEAL
1997. Taller de análisis de la viabilidad poblacional y del hábitat del caimán del
Orinoco (Crocodylus intermedius) UICN/SSC Conservation Breeding
Specialist Group. Apple Valley, MN.
ÁVILA-MANJÓN, P. M.
2008. Estado poblacional del caimán del Orinoco (Crocodylus intermedius) en el
río Cojedes, Venezuela. Tesis de Maestría. Vicerrectorado de Producción
Agrícola. UNELLEZ, Guanare.
AYARZAGÜENA, J.
1983. Ecología del caimán de anteojos o baba (Caiman crocodilus L.) en los llanos
de Apure (Venezuela). Doñana Acta Vertebrata. 10(3): 1-134.
1984a. Variaciones en la dieta de Caiman sclerops. La relación entre morfología
bucal y dieta. Mem Soc Cienc Nat La Salle 44(122):123-140.
1987. Conservación del caimán del Orinoco (Crocodylus intermedius) en
Venezuela. Parte I. Río Cojedes. Informe para FUDENA, WWF-US,
Proyecto 6078.
1988. Cambios en la diversidad en sabanas modulares en los llanos de Apure,
medidos con la dieta de un depredador oportunista (Caiman crocodilus).
Mem. Soc. Cienc. Nat. La Salle 48(129): 163-166.
1990. An update on the recovery program for the Orinoco crocodile. Croc. Spec.
Gr. News. 9(3):16-18.
1991. Zoocriaderos: Una alternativa válida en el aprovechamiento y manejo de
cocodrilos. Relatoría III Taller sobre Conservación y Manejo de la Especie
Baba (Caiman crocodilus). Caracas, 28 al 30 de noviembre de 1990.

Los Crocodylia de Venezuela
243
AYARZAGÜENA, J., Y J. CASTROVIEJO
2008. Las babas (Caiman crocodilus L.) de la Estación Biológica EL Frío (Estado
de Apure), Llanos del Orinoco, Venezuela. Pp 181-294: in J. Castroviejo, J.
Ayarzagüena, and A. Velasco, editors. Contribución al conocimiento de los
caimanes del género Caiman de Suramérica. Publicaciones de la
Asociación Amigos de Doñana.
BABARRO G. R.
2011. Censo preliminar de caimanes de la costa (Crocodylus acutus) en el sector
de los canales de Río Chico y Paparo 30-mayo al 01 de junio 2011.
Caracas: MPPPA. Oficina Nacional de Diversidad Biológica. 10 p.
BAQUERO, B. Y M. QUERO
1996. Manejo del programa de zoocriaderos de la especie baba (Caiman
crocodilus) en Venezuela. Zoocriaderos 1(1):1-6.
BARRERA, L. F.
2004. Estado actual de un relicto poblacional del caimán agujo (Crocodylus acutus
Cuvier, 1807) en una zona del Magdalena medio. Fundación Omacha. En:
www.proaves.org.
BARROS, T.
2003. Evaluación del recurso Caiman crocodilus fuscus (Babilla) en un sector del
P. N. Ciénagas de Juan Manuel, Estado Zulia. Tesis de Maestría. Postgrado
en Manejo de Fauna Silvestre. UNELLEZ, Guanare, Venezuela.
BARROS, T., A. URDANETA, A. LANDER, R. LÓPEZ Y T. GUTIÉRREZ
2005. Reforzamiento y seguimiento de la población de Caimanes de la Costa
(Crocodylus acutus) en la Ciénaga de Los Olivitos, Estado Zulia,
Venezuela. Ciencia 13(2):162-181.
BAYLISS, P.
1987. Survey methods and monitoring within crocodile management programs. Pp.
157-175 in: Webb, G. J. W, C. Manolis y P. Whitehead. (Eds.). Wildlife
Management: Crocodiles and Alligators. Surrey Beatty and Sons Pty
Limited in Association with the Conservation Commission of the Northern
Territory. Chipping Norton, NSW, Australia.
BELLAIRS, A. D´A.
1987. The Crocodylia. Pp. 5-7 in: Webb, G. J. W, C. Manolis y P. Whitehead.
(Eds.). Wildlife Management: Crocodiles and Alligators. Surrey Beatty and
Sons Pty Limited in Association with the Conservation Commission of the
Northern Territory. Chipping Norton, NSW, Australia.

Andrés Eloy Seijas
244
BLOHM, T.
1948. Observaciones sobre los caimanes traídos del río Orinoco en abril de 1946.
Mem. Soc. Cienc. Nat. La Salle 8(22):129-132.
1973. Conveniencia de criar cocodrilos en Venezuela con fines económicos y para
prevenir su extinción. Primer Simposio Internacional sobre la Fauna
Silvestre y Pesca Fluvial y Lacustre Amazónica. Manaus, Brazil.
Noviembre 26-Diciembre 1, 1973.
1982. Husbandry of Orinoco crocodiles (Crocodylus intermedius) in Venezuela.
Pp. 267-285 in: Crocodiles. Proceedings of the 5th Working Meeting of the
Crocodile Specialist Group. IUCN- The World Conservation Union. Gland,
Switzerland.
BOEDE, E. Y E. SOGBE
2000. Enfermedades de caimanes del Orinoco (Crocodylus intermedius) y
caimanes de la costa (Crocodylus acutus) mantenidos en zoocriaderos
venezolanos. Revista Científica, FCV-LUZ 10(4): 328-338.
BRAZAITIS, P.
1973. The identification of living crocodilians. Zoologica 58:59-101.
BRISBIN, L.
1966. Reactions of the American alligator to several inmobilizing drugs. Copeia
1966(1): 129-130.
BROCHU, C. A. Y A. D. RINCÓN
2004. A gavialoid Crocodylian from the lower Miocene of Venezuela. Special
Papers in Paleontology 71: 61-79.
BUFFETAUT, E.
1989. Evolution. In Crocodiles and Alligators: 26-41. Ross, C. (Ed.) Silverwater,
NSW, Australia: Golden Press Pty. Ltd.
CALDWELL, J.
2004. World Trade in Crocodilian Skins, 2000-2002. UNEP World Conservation
Monitoring Centre, Cambridge, UK
CALZADILLA V., F.
1948. Por los llanos de Apure. Ediciones del Ministerio de Educación Nacional,
Dirección de Cultura. Caracas, Venezuela.
CARDONA, M.
1959. El Caimán en el folklore venezolano. Arch. Venez. Folk. V-VI(6): 61-80.

Los Crocodylia de Venezuela
245
CATTINI, A.
1987. Expedición a la Península de Paria. El Natural 24. (Mimeografiado).
CHABRECK, R.
1976. Cooperative surveys of population trends in the American alligator. in:
Crocodiles. 3rd Working Meeting Crocodile Specialist Group/SSC, IUCN-
The World Conservation Union. Maningrida, Northern Territory,
Australia. Abril 5-12.
CHÁVEZ, C.
1992. Separación del nicho entre la población del caimán de la costa (Crocodylus
acutus) y baba (Caiman crocodilus) en el embalse de Jatira, estado Falcón.
Tesis de Licenciatura, Escuela de Biología, Facultad de Ciencias. UCV,
Caracas.
2000. Conservación de las poblaciones del caimán del Orinoco (Crocodylus
intermedius) en Venezuela. Caracas, Informe Profauna-Corpoven, MARN
CHERKISS, M. S., H. E. FLING, F. MAZZOTTI, K. RICE Y M. D. CONILL
2005. Contando y capturando cocodrílidos, University of Florida, IFAS Extension.
(Disponible en http://edis.ifas.edu).
CHIRIVI-GALLEGOS, H.
1973. Contribución al conocimiento de la babilla o yacaré tinga (Caiman
crocodilus) con notas acerca de su manejo y de otras especies de
Crocodylia neotropicales. Primer Simposio Internacional sobre la Fauna
Silvestre y Pesca Fluvial y Lacustre Amazónica. Manaus, Brasil.
Noviembre 26-Diciembre 1, 1973.
CITES
1995. Guía de identificación de los cocodrilos protegidos por la Convención sobre
el Comercio Internacional de Especies Amenazadas de Fauna y Flora
Silvestres. Una iniciativa del Ministerio del Medio Ambiente del Canadá.
Minister of Supply and Services, Canada.
CODAZZI, A.
1940 [1841]. Resumen de la Geografía de Venezuela. Venezuela en 1841. Tomo I.
Geografía Física. Biblioteca Venezolana de Cultura. Taller de Artes
Gráficas Escuela Técnica Industrial, Caracas.
COLOMINE, G., A. VELASCO, R. DE SOLA, G. VILLARROEL, A. LANDER, W.
VÁSQUEZ, T. PINO Y J. CORAZZELLI
2000. Evaluation of the density and structure of size class of the spectacled caiman
(Caiman crocodilus) in the Alto and Bajo Apure ecological regions,
Venezuela. Pp. 304-309 in: Crocodiles. Proceedings of the 15th Working
Meeting of Crocodile Specialist Group, SSC, IUCN-The World
Conservation Union Gland, Switzerland.

Andrés Eloy Seijas
246
COTT, H. B.
1961. Scientific results of an inquiry into the ecology and economic status of the
Nile crocodile (Crocodylus niloticus) in Uganda and Northern Rhodesia.
Transactions of the Zoological Society of London 29(4):211- 358.
COULSON, R. A. Y T. HERNÁNDEZ
1983. Alligator metabolism: Studies of chemical reactions in vivo. Oxford,
Pergamon Press.
D'ANDRIA, B.
1980. Estructuración de un modelo matemático sobre la dinámica poblacional de la
baba (Caiman crocodilus) con miras a su posible explotación comercial.
Tesis de Licenciatura, Universidad Simón Bolívar, Caracas.
DE SOLA, R., G. COLOMINE, A. VELASCO, A. LANDER Y G. VILLARROEL
2000. Potential and current use of the populations of spectacled caiman subject to
commercial use in Venezuela. Pp. 285-297 in: Proceedings of the 15th
Working Meeting of Crocodile Specialist Group, SSC. IUCN - The World
Conservation Union, Gland, Switzerland.
DE SOLA, R. Y V. A. TREJO
1996. Aprovechamiento Comercial de la baba (Caiman crocodilus) en el estado
Delta Amacuro. Una visión retrospectiva. Temporadas 1993-1996.
MARNR-PROFAUNA. Caracas.
DEEMING, D. Y M. FERGUSON
1989. The mechanism of temperature sex determination in crocodilians. A
hypothesis. American Zoologist 29: 973-985.
DODSON, P.
1975. Functional and ecological signifficance of relative growth in Alligator. J.
Zool. Lond. 175:315-355.
DONADIO, R.
1981. Legislación para la fauna silvestre en Colombia. Pp. 193-198 in: Medem, F.
1981. Los Crocodylia de Sur América, Vol. I. Los Crocodylia de Colombia.
Editorial Carrera 7a. Ltda, Bogotá.
DONOSO-BARROS, R.
1965. Contribución al conocimiento de los cocodrilos de Venezuela. Physis
25(70): 387-400.
1966a. Contribución al conocimiento de los cocodrilos de Venezuela. Parte 2.
Physis 26:15-32.

Los Crocodylia de Venezuela
247
DONOSO-BARROS, R.
1966b. Contribución al conocimiento de los cocodrilos de Venezuela. Conclusión.
Physis 26:263-274.
ELLIS, T. M.
1980. Caiman crocodilus: an established exotic in South Florida. Copeia 1980(1):
52-54.
ESPINOSA-BLANCO, A. S.
2010. Colecta de huevos como estrategia de conservación del cocodrilo del
Orinoco en el Sistema del río Cojedes, Venezuela. Tesis de Maestría.
Postgrado en Manejo de Fauna Silvestre. Unellez-Guanare. 94 pp.
ESPINOZA, E.
1988. Desarrollo de Criaderos de Babas Caiman crocodilus, en los Llanos
Venezolanos. Pp. 127-134 in: II Simposio Nacional Sobre Biotecnologia,
Maracaibo, 12 al 15 de julio de 1988.
1989. La cría de Baba, Caiman crocodilus, en Venezuela: Estado Actual y Futuro.
II Taller Sobre Conservación y Manejo de la Baba (Caiman crocodilus) en
Venezuela. UNELLEZ, FUNDAFAUNA, FUNDACITE, MARNR.
Guanare 21 al 23 de agosto de 1989.
FERGUSON, M.
1987. Post-laying stages of embryonic development for crocodilians. Pp. 427-444
in: Wildlife management: Crocodiles and Alligators. G. Webb, C. Manolis
and P. Whitehead. Chipping Norton, Australia, Surrey Beatty and Sons Pty
Limited.
FERGUSON, M. Y T. JOANEN
1983. Temperature-dependent sex determination in Alligator mississipiensis. J.
Zool. Lond.: 143-177.
FITZGERALD, L.
1988a. Dietary patterns of Caiman crocodilus in the Venezuelan llanos. Tesis de
Maestría. University of Alburquerque, New Mexico.
1988b. An evaluation of stomach flushing techniques for crocodilians. J. Herpetol.
23(2):170-172.
FLEISHMAN, L.
1989. Caiman crocodilus does not require vision for underwater prey capture. J.
Herpetol. 23(3):296.

Andrés Eloy Seijas
248
FRANKLIN C. E., B. M. DAVIS, Y S. K. J. PEUCKER
2002. Stress response to noosing and restraint versus capture by electrical stunting
in captive Crocodylus porosus. Pp. 157 in: Crocodiles. Proceedings of the
16th Working Meeting of the Crocodile Specialist Group. IUCN- The
World Conservation Union. Gland, Switzerland.
FRANZ, R., S. REID, Y C. PUCKETT
1985. Discovery of a population of Orinoco crocodile Crocodylus intermedius in
Southern Venezuela. Biol. Conserv. 32:137-147.
GANS, C. Y A. POOLEY
1976. Commentary-Research on crocodiles? Ecology 57(5): 839-840.
GODSHALK, R.
1978. El caimán del Orinoco, Crocodylus intermedius, en los Llanos Occidentales
de Venezuela con observaciones sobre su distribución en Venezuela y
recomendaciones para su conservación. FUDENA, Caracas, 58 pp.
1982. Status and conservation of Crocodylus intermedius in Venezuela. Pp. 39-53
in: Crocodiles. Proceedings of the 5th Working Meeting of the IUCN/SSC
Crocodile Specialist Group, SSC. Gainesville, FL. IUCN Publ. N.S., Gland,
Switzerland.
1996. Conservation and Sustainable Use of the Yacare Caiman, Caiman yacare, in
Bolivia. Pp. 1341-150 in: Crocodiles. Proceedings of the 13th Working
Meeting of Crocodile Specialist Group, SSC. IUCN-The World
Conservation Union, Gland, Switzerland.
GONZÁLEZ FERNÁNDEZ, M. J.
1999. Conservación del caimán de la costa (Crocodylus acutus) en la bahía de
Turiamo, estado Aragua, Venezuela. Caracas, MARN, Servicio Autónomo
de Fauna (PROFAUNA): 36 pp.
GONZÁLEZ ORIHUELA, G. A.
2010. Caracterización del hábitat y composición de la dieta del babo morichalero
(Paleosuchus palpebrosus) en los Llanos Orientales, estado Anzoátegui.
Tesis de Grado. Escuela de Biología, Facultad de Ciencias, Universidad
Central de Venezuela, Caracas.
GORZULA, S.
1978. An ecological study of Caiman crocodilus crocodilus inhabiting Savanna
lagoons in the Venezuelan Guayana. Oecologia (Berl.) 35:21-34.
1984. Proposal for a photographic method for size estimates of crocodilians. Herp.
Rev. 15: 38-39.
1995. Russia´s oldest crocodile dies. Croc. Spec. Gr. News. 14(1):8.

Los Crocodylia de Venezuela
249
GORZULA, S. Y A. E. SEIJAS
1989. The common caiman. Pp 44-61 in: Crocodiles, Their Ecology, Management
and Conservation. Special Publication of the Crocodile Specialist Group.
IUCN Publications, N.S., Gland, Switzerland.
GORZULA, S. Y A. PAOLILLO
1986. La ecología y estado actual de los Alligatoridos de la Guayana Venezolana.
Pp. 37-54 in: Crocodiles. Proceedings of the 7th Working Meeting of
Crocodile Specialist Group, SSC. IUCN - The World Conservation Union,
Gland, Switzerland.
GORZULA, S., J. PAOLINI Y J. THORBJARNARSON
1986. Some Hydrochemical and hydrological characteristics of crocodilian
habitats. Trop. Freshw. Biol. (0795-0101/87).
GOULD, S. J.
1966. Allometry and size in ontogeny and phylogeny. Biol. Rev. 41:587-640.
GRIGG, G. C., F. SEEBACHER Y C. E. FRANKLIN (EDS.)
2001. Crocodilian Biology and Evolution. Surrey Beatty & Sons. Chipping Norton,
NSW, Australia.
GUADA, H. J. Y P. VERNET
1990. Informe de inspección de campo al Parque Nacional Médanos de Coro y
áreas propuestas de ampliación. Instituto Nacional de Parques
(INPARQUES). Dirección General Sectorial de Parques Nacionales.
División de Estudios Básicos, Caracas, 14 pp.
GUMILLA, J.
1963 [1745]. El Orinoco Ilustrado y Defendido. Biblioteca Nacional de la Historia.
No. 68. Caracas.
GUTIERREZ, S.
1991. Situación actual de los zoocriaderos de baba en Venezuela. III Taller sobre
Conservación y Manejo de la Especie Baba (Caiman crocodilus), Caracas.
HALL, P. M. Y K. M. PORTIER.
1994. Cranial morphometric of the New Guinea crocodile (Crocodylus
novaeguineae): ontogenetic variation in relative growth of the skull and an
assessment of is utility as a predictor of the sex and size of individuals.
Herpetol. Monog. 8:203-225.

Andrés Eloy Seijas
250
HAMES, R. B.
1979. A Comparison of the efficiencies of the shotgun and the bow in neotropical
forest hunting. Hum. Ecol. 7(3): 219-251.
HARSHMAN, J., J. C. HUDDLESTON, J. P. BOLLBACK, T. J. PARSONS Y M. J.
BRAUN.
2003. True and False Gharials: A Nuclear Gene Phylogeny of Crocodylia. Syst.
Biol. 52(3):386–402.
HEREDIA, H. J.
2005. Estado poblacional del caimán del Orinoco (Crocodylus intermedius) en tres
sectores del río Manapire, Guárico, Venezuela. Aplicación de
Conocimientos. Ing. Rec. Naturales Renovables, UNELLEZ, Guanare,
Venezuela.
HERRON, J. C.
1985. Population status, spatial relations, growth, and injuries in black and
spectacled caiman in Cocha Cachu. Tesis pregrado, Princeton University.
New Jersey.
HITCHCOCK, C. B.
1948. La región Orinoco-Ventuari, Venezuela. El Compás, Caracas.
HUMBOLDT, A.
1975 [1859-1869]. Del Orinoco al Amazonas. Viaje a las regiones equinocciales
del Nuevo Continente. Traducción de la segunda edición alemana por
Francisco Payarols. Edt. Labor, Barcelona. 429 p.
1991. Viaje a las Regiones Equinocciales del Nuevo Continente. Traducción de
Lisandro Alvarado, Eduardo Röhl y José Nucete-Sardi. Monte Ávila
Editores.
HUTTON, J. M. Y M. E. J. WOOLHOUSE
1989. Mark-recapture to assess factors affecting the proportion of a Nile crocodile
population seen during spotlight counts at Ngezi, Zimbabwe, and the use of
spotlight counts to monitor crocodile abundance. Journal of Appl. Ecol. 26:
381-395.
HUTTON, J. Y G. J. W. WEBB
1992. An introduction to the farming of crocodilians. Pp. 1-39 in: R. Luxmoore
(Ed.). Directory of Crocodile Farms. 2da. Ed. IUCN, Gland, Switzerland.

Los Crocodylia de Venezuela
251
HUTTON, J., J. P. LOVERIDGE Y D. K. BLAKE
1987. Capture methods for the Nile crocodile in Zimbabwe. Pp. 243-247 in: Webb,
G. J. W, C. Manolis y P. Whitehead. (Eds.). Wildlife Management:
Crocodiles and Alligators. Surrey Beatty and Sons Pty Limited in
Association with the Conservation Commission of the Northern Territory.
Chipping Norton, NSW, Australia.
HUTTON, J., J. P. ROSS Y G. WEBB
2001. Using the market to create incentives for the conservation of crocodilians.
IUCN/SSC Crocodile Specialist Group: 1-28.
IORDANSKY, N.
1973. The skull of the Crocodilia. Pp. 201-261 in: Gans, C. (ed.). Biology of the
Reptilia, Vol. 4. London and New York. Academic Press London and New
York.
JACKSON, K. Y D. R. BROOKS
2007. Do crocodiles co-opt their sense of “touch” to “taste”? A possible new type
of vertebrate sensory organ. Amphibia-Reptilia 28:277-285.
JIMÉNEZ-ORAÁ, MAGDDY
2002. El caimán del Orinoco (Crocodylus intermedius) en los sectores Laguna
larga y Chigüichigüe del río Manapire, Guárico, Venezuela. Tesis de
Maestría. Postgrado en Manejo de Fauna Silvestre. UNELLEZ, Guanare,
Venezuela.
JIMÉNEZ-ORAÁ, MAGDIEL
2005. Evaluación del programa de liberación de caimán del Orinoco (Crocodylus
intermedius) en el río Manapire, Guárico, Venezuela. Aplicación de
Conocimientos. Ing. Rec. Naturales Renovables, UNELLEZ, Guanare,
Venezuela.
JIMÉNEZ-ORAÁ, MAGDDY, A. E. SEIJAS, MAGDIEL JIMÉNEZ-ORAÁ Y H.
HEREDIA AZUAJE
2007. Colecta de huevos como estrategia de conservación del Caimán del Orinoco
(Crocodylus intermedius) en el río Manapire, Guárico, Venezuela. Pp. 33-
39 in: Seijas, A. E. (Ed.). Conservación del Caimán del Orinoco.
Memorias del III Taller para la Conservación del Caimán del Orinoco. San
Carlos (Cojedes, Venezuela), 18 y 19 de enero de 2007.
JOANEN, T. Y G. PERRY
1971. A new method for capturing alligators using electricity. Annual Conference
Southeastern Association of Game and Fish Commissioners 25: 124-130.

Andrés Eloy Seijas
252
JOANEN, T. Y L. MCNEASE
1976. Culture of immature American alligators in controlled environmental
chambers. Proceedings of the Annual Meeting World Mariculture Society
7: 201-211.
1991. Incubación de huevos de lagartos. Pp. 25-31 in: Crianza de cocodrilos:
Información de la literatura científica. CSG/SSC IUCN, Gland,
Switzerland.
JOANEN, T., L., MCNEASE Y M. FERGUSON
1987. The effects of eggs incubation temperature on post - hatching growth of
American alligator. Pp. 533-537 in: Webb, G. J. W, C. Manolis y P.
Whitehead. (Eds.). Wildlife Management: Crocodiles and Alligators.
Surrey Beatty and Sons Pty Limited in Association with the Conservation
Commission of the Northern Territory. Chipping Norton, NSW, Australia.
JOLICOEUR, P.
1963. The multivariate generalization of the allometry equation. Biometrics 19:
497-499.
KING, F. W.
1978. The wildlife trade. Pp. 253-271 in: H. P. Brokaw (ed.). Wildlife and
America. U. S. Govt. Printing Office, Washington, DC.
1991. The first record of Melanosuchus niger from Venezuela. Croc. Spec. Gr.
News. 10(4): 24-25.
KING, F. W. Y R. L. BURKE
1989. Crocodilian, tuatara and turtle species of the world. A taxonomic and
geographic reference. Assoc. Syst. Collect., Washington D. C.
KING, F. W., M. ESPINAL Y C.A. CERRATO
1990. Distribution and status of the Crocodilians of Honduras. Pp. 313-354 in:
Proceedings of the Tenth Working Meeting of Crocodile Specialist Group.
IUCN - Gland, Swizerland. 354p.
LANDER, A.
2003. Seguimiento de la población del caimán de la costa (Crocodylus acutus
Cuvier) en la bahía de Turiamo, Estado Aragua, Venezuela. Maracay,
MARN: 15.
LANDER, A., S. BERMÚDEZ, Y T. BARROS
2008. Monitoring populations of American crocodile (Crocodylus acutus) in the
states of Aragua, Falcón, Trujillo and Zulia, Venezuela. Pp. 73. 19th
Working Meeting of the Crocodile Specialist Group, IUCN/SSC (Libro de
Resúmenes), Santa Cruz, Bolivia.

Los Crocodylia de Venezuela
253
LANG, J. W. Y H. V. ANDREWS
1994. Temperature-dependent sex determination in Crocodilians. J. Exp. Zoo. 270:
28-44.
LANGSTON, W., JR.
1973. The Crocodilian skull in historical perspective. Pp. 263-284 in: Gans, C.
(ed.). Biology of The Reptilia. London and New York, Academic Press.
LE CREN, E. D.
1951. The length-weight relationship and seasonal cycle in gonad weight and
condition in the perch (Perca fluviatilis). J. Anim. Ecol. 20:201-219.
LEBUFF, C.
1957. The range of Crocodylus acutus along the Florida Gulf Coast. Herpetologica
13: 188.
LESLIE, J. A. Y L. E. TAPLIN
2001. Recent developments in osmoregulation in crocodilians. Pp. 265-279 in:
Grigg, G. C., F. Seebacher y C. E. Franklin (Eds.). Crocodilian Biology
and Evolution. Surrey Beatty & Sons. Chipping Norton, NSW, Australia.
LIZOT, J.
1979. On food taboos and Amazon cultural ecology. Curr. Anthr. 20: 150-151.
LLOBET, A.
2002. Estado poblacional y lineamientos de manejo del caimán del Orinoco
(Crocodylus intermedius) en el río Capanaparo (Apure, Venezuela). Tesis
de Maestría. Postgrado en Manejo de Fauna Silvestre. UNELLEZ,
Guanare, Venezuela.
LLOBET, A. Y A. E. SEIJAS
2003. Estado poblacional y lineamientos de manejo del caimán del Orinoco
(Crocodylus intermedius) en el río Capanaparo, Venezuela. Pp. 117-129 in:
Polanco-Ochoa, R. (ed.) Manejo de Fauna Silvestre en Amazonía y
Latinoamérica. Selección de Trabajos V Congreso Internacional. Bogotá,
CITES, Fundación Natura.
LÓPEZ-CORCUERA, G.
1984. Fauna legendaria: Rayas, tembladores, caimanes y culebras de agua.
Editorial Arte, Caracas.

Andrés Eloy Seijas
254
LOVERIDGE, J. P. Y D. K. BLAKE
1987. Crocodile inmovilization and anaesthesia. Pp. 259-267 in: Webb, G. J. W, C.
Manolis y P. Whitehead. (Eds.). Wildlife Management: Crocodiles and
Alligators. Surrey Beatty and Sons Pty Limited in Association with the
Conservation Commission of the Northern Territory. Chipping Norton,
NSW, Australia.
LUGO, M.
1995. Cría del caimán del Orinoco (Crocodylus intermedius) en la Estación de
Biología Tropical "Roberto Franco", Villavicencio, Meta. Rev. Acad.
Colomb. Cienc. Exac. Fís. Nat. 19(74): 601-606.
1998. Evaluación del programa de liberación del caimán del Orinoco (Crocodylus
intermedius) en el Refugio de Fauna Silvestre “Caño Guaritico” y
alrededores. Estado Apure. Tesis de Maestría. Postgrado en Manejo de
Fauna Silvestre. UNELLEZ, Guanare, Venezuela.
LUZARDO, A. L.
2005. Crecimiento sobrevivencia y dispersión del caimán de la costa (Crocodylus
acutus) en el embalse de Pueblo Viejo, estado Zulia. Tesis de licenciatura.
Facultad Experimental de Ciencias. Universidad del Zulia. Maracaibo,
Venezuela.
MACGREGOR, J.
2002. International trade in crocodilian skins: review and an analysis of the trade
and industry dynamics for market-based conservation. Pp. 12-18 in:
Proceedings of the 16th Working Meeting of the Crocodile Specialist
Group, SSC. IUCN- The World Conservation Union. Gland, Switzerland.
2006. Captive crocodilian production and the shaping of conservation incentives.
TRAFFIC Online Report Series No. 12.
MAGNUSSON, W. E.
1983. Size estimates of crocodilians. J. Herpetol. 17(1): 86-88.
1992a. Paleosuchus palpebrosus. Catalogue of American Amphibians and
Reptiles: 554.1-554.2.
1992b. Paleosuchus trigonatus. Catalogue of American Amphibians and Reptiles:
555.1-555.3.
MAGNUSSON, W. E. Y A. LIMA
1991. The Ecology of a Cryptic Predator, Paleosuchus trigonatus, in a Tropical
Rainforest. J. Herpetol. 25(1): 660-666.

Los Crocodylia de Venezuela
255
MAGNUSSON, W. E., A. LIMA, J. HERO, T. SANAIOTTI Y M. YAMAKOSHI
1990. Paleosuchus trigonatus Nests: Sources of Heat and Embryo Sex Ratios. J.
Herpetol. 24(4): 397-400.
MANESS, S. J.
1982. Status of Crocodylus acutus, Caiman crocodilus fuscus, and Caiman
crocodilus crocodilus in Venezuela. Pp. 117-120 in: Crocodiles.
Proceedings of the 5th Working Meeting of Crocodile Specialist Group,
SSC. IUCN - The World Conservation Union , Gland, Switzerland.
MARCELLINI, D. L.
1979. Activity patterns and densities of Venezuelan caiman (Caiman crocodilus)
and pond turtles (Podocnemis vogli). Pp. 263-271 in: J. F. Eisemberg (Ed.).
Vertebrate Ecology in the Northern Neotropics. Smithsonian Institution
Press. Washington, DC.
MARTÍNEZ, E.
1996. Diseño y operación de zoocriaderos de babas (Caiman crocodilus
crocodilus). Zoocriaderos 1(2):11-20.
MAZZOTTI, F.
1989. Structure and function. Pp. 42-52 in: Ross, C. A. Crocodiles and Alligators.
Golden Press Pty Ltd. Silverwater, NSW, Australia.
MCALILEY, L. R., R. E. WILLIS, D. A. RAY, P. SCOTT WHITE, C. BROCHU Y L. D.
DENSMORE III.
2006. Are crocodiles really monophyletic?—Evidence for subdivisions from
sequence and morphological data. Mol. Phylog. Evol. 39(1)16-32..
MEDEM, F.
1967. El género "Paleosuchus" en Amazonia. Atas Simposio sobre a Biota
Amazonica: 141-162.
1976. Recomendaciones respecto a contar el escamado y tomar las dimensiones de
nidos, huevos, y ejemplares de los Crocodylia y Testudines. Lozanía 20:1-
17.
1981. Los Crocodylia de Sur América, Vol. I. Los Crocodylia de Colombia.
Editorial Carrera 7a. Ltda, Bogotá. 354 pp.
1983. Los Crocodylia de Sur América, Vol. II. Editorial Carrera 7a. Ltda, Bogotá.
270 pp.
MEDINA, C.
1976. Crocodilians from the late tertiary of northwestern Venezuela:
Melanosuchus fisheri sp. nov. Breviora (438): 1-14.

Andrés Eloy Seijas
256
MEDINA-PADILLA, G.
1960. Caza y conservación de fauna en Venezuela. El Agricultor Venezolano
22(216):22-28.
MENDOZA, J. M.
2003. Calidad de hábitat y viabilidad poblacional del caimán del Orinoco en el río
Cojedes Norte, Venezuela. Tesis de Maestría. Postgrado en Manejo de
Fauna Silvestre. UNELLEZ, Guanare, Venezuela.
MESSEL, H. Y D. STEPHENS
1980. Drug immobilization of crocodiles. J. Wildl. Manag. 44(1): 395-396.
MESSEL, H., G. C. VORLICEK, A. G. WELLS, Y W. J. GREEN
1981. Surveys of tidal rivers system study in the Northern Territory of Australia
and their crocodile populations. Monograph 1. The Blyth-Cadell rivers
systems study and the status of Crocodylus porosus in tidal waterways of
Northern Australia. Oxford and Sydney.
MICUCCI, P. A. Y T. WALLER
1995. Los yacares en Argentina: Hacia un aprovechamiento sustentable. Pp. 81-
112 in: Larriera, A. y L. M. Verdade (eds.). La conservación y el manejo de
caimanes y cocodrilos en América Latina. Vol. 1. Fundación Banco Bica.
Santo Tomé, Santa Fe, Argentina.
MONDOLFI, E.
1957. La conservación de nuestra fauna autóctona. Conservación 3:3-18.
1965. Nuestra fauna. El Farol 214:2-13.
MONTAGUE, J. J.
1984. Morphometric analysis of Crocodylus novaeguineae from the Fly River
Drainage, Papua New Guinea. Aust. Wildl. Res. 11: 395-414.
MOURAO G. M., P. BAYLISS, M. E. COUTINHO, C. L. ABERCROMBIE Y A. ARRUDA
1994. Test of an aerial survey for caiman and other wildlife in Pantanal, Brazil.
Wildlife Society Bulletin 22:50-56.
MUÑOZ, I.
1986. El caimán de la costa. Bases para su conservación. Editorial Primicia,
Barquisimeto.
MUÑOZ, M. Y J. THORBJARNARSON
2000. Movement of captive-released Orinoco crocodiles (Crocodylus intermedius)
in the Capanaparo River, Venezuela. J. Herpetol. 34(3): 397-403.

Los Crocodylia de Venezuela
257
NATTERER, J. VON
1841. Beitrag zur näheren Kenntnis der südamerikanischen Alligatoren, nach
gemeinschaftlichen Untersuchungen mit L. J. Fitzinger. Ann. Wien. Mus.
Naturgesch., 2:313-324.
NAVARRO, M.
2007. Estado poblacional y reproductivo del caimán del Orinoco (Crocodylus
intermedius) en el río Cojedes. Tesis de Licenciatura, Universidad Simón
Bolívar, Caracas.
NORELL, M. A.
1989. The higher level relationship of the extant Crocodylia. J. Herpetol. 23(4):
325-335.
NRC (NATIONAL RESEARCH COUNCIL)
1983. Crocodiles as a resource for the tropics. National Academy Press,
Washington, D.C.
OGDEN, J. C.
1978. Status and nesting biology of the American crocodile, Crocodylus acutus,
(Reptilia, Crocodilidae) in Florida. J. Herpetol. 12:183-196.
OJASTI, J., G. FEBRES Y M. COVA
1987. Consumo de fauna por una comunidad indígena en el estado Bolivar,
Venezuela. Pp. 45-50 in: Actas del 9no Congreso Latinoamericano de
Zoología. Arequipa, Perú.
PACHECO, A. D.
2009. Distribución, abundancia y estructura poblacional del Babo Morichalero
(Paleosuchus palpebrosus) en los Llanos Orientales del Estado Anzoátegui.
Tesis de Licenciatura. Escuela de Biología,Universidad Central de
Venezuela (UCV), Caracas.
PÁEZ, R.
1980 [1868]. Escenas rústicas en Sur América o Vida en los Llanos de Venezuela.
Ediciones Centauro. Caracas.
PÉREZ TALAVERA, A. T.
1999. Estrategia de nidificación del Caiman crocodilus crocodilus (Baba) en
Caicara del Orinoco, estado Bolívar, Venezuela. Mem. Soc. Cienc. Nat. La
Salle 59(152): 105-114.
2000. Crecimiento del Caiman crocodilus en cautiverio. Interciencia 25: 9.

Andrés Eloy Seijas
258
PÉREZ TALAVERA A. T. Y J. C. RODRÍGUEZ
2005. Influencia de la temperatura del aire y del agua en el crecimiento de
Crocodylus intermedius en dos condiciones de cautiverio. Boletín Centro
Invest Biol 39(1):15-26.
POOLEY, A.
1989. Food and feeding habits. Pp. 79-91 in: Ross C. (Ed.). Crocodiles and
Alligators. Weldon Owen Pty Limited. Silverwater, SNW, Australia.
POUGH, F. H., J. B. HEISNER Y W. N. MCFARLAND
1996. Vertebrate life.1996. (4ta edición). Prentice Hall, New Jersey.
POUGH, F. H., R. M. ANDREWS, J. E. CADLE, M. L. CRUMP, A. H. SAVITZKY Y K.
D. WELLS
2004. Herpetology (Third Edition). Pearson Prentice Hall, New Jersey.
RAMIA, M.
1967. Tipos de sabanas en los llanos de Venezuela. Boletín de la Sociedad
Venezolana de Ciencias Naturales 27(112): 264-288.
RAMÍREZ, Y., C. CASTILLO Y S. GORZULA
1977. Proyecto venezolano sobre cocodrilos. Herpetologica Review 8(4): 130.
RAMO, C. Y B. BUSTO
1986. Censo aéreo de caimanes (Crocodylus intermedius) en el río Tucupido
(Portuguesa, Venezuela) con observaciones sobre su actividad de
soleamiento. Pp. 109-119 in: Crocodiles. Proceeding of the 7th Working
Meeting of the Crocodile Specialist Group Meeting. IUCN-The World
Conservation Union. Gland, Switzerland.
1989. Anfibios y Reptiles del Modulo Fernando Corrales (Apure). Mem. Soc.
Cienc. Nat. La Salle 49:131-132.
RAMO, C., B. BUSTO, Y J. UTRERA
1992. Breeding and Rearing the Orinoco crocodile Crocodylus intermedius in
Venezuela. Biol. Conserv. 60:101-108.
RAMOS, S., S. DANIELEWSKI Y G. COLOMINE
1981. Contribución a la ecología de los vertebrados acuáticos en esteros y bajíos de
Sabanas Moduladas. Boletín de la Sociedad Venezolana de Ciencias
Naturales 35(139): 79-103.
READ, M. A., J. D. MILLAR, I. P. BELL Y A. FELTON
2004. The distribution and abundante of the estuarine crocodile, Crocodylus
porosus, in Queensland. Wildlife Research 31:527-534.

Los Crocodylia de Venezuela
259
RÍOS, M. Y F. TRUJILLO
2004. Censo preliminar de Crocodylia en los ríos Meta y Bita, Departamento del
Vichada (Colombia). Pp. 229-242 in: Diaz Granados, M. C. y Trujillo, F.
(Ed.). Fauna Acuática en la Orinoquia Colombiana. Bogotá, Instituto de
Estudios Ambientales para el Desarrollo, Departamento de Ecología y
Territorio (IDEADE-DET).
RIVERO-BLANCO, C.
1968. El caimán y su valiosa piel. Líneas: 11-13.
1973. Sobre los hábitos reproductivos de la baba o Jacare-tinga, Caiman
crocodilus, en los Llanos de Venezuela y las posibilidades de manejo en
semi-cautividad, con sugerencias sobre las posibles aplicaciones de la
metodología de manejo al caimán negro o jacare-acu Melanosuchus niger
de las cuencas del Amazonas y Río Negro. Primer Simposio Internacional
sobre la Fauna Silvestre y Pesca Fluvial y Lacustre Amazónica. Manaus,
Brasil. Noviembre 26-Diciembre 1, 1973.
1974. Hábitos reproductivos de la baba en los llanos de Venezuela. Natura
52:24-29.
RODRÍGUEZ, J. P. Y F. ROJAS-SUÁREZ
2008. Libro Rojo de la fauna Venezolana. Provita y Shell de Venezuela. S.A.
Caracas. 364 pp, Caracas.
RÖHL, E.
1956. Fauna descriptiva de Venezuela. Caracas, Nuevas Gráficas S. A.
ROOS, J., R. K. AGGARWAL Y A. JANKE
2007. Extended mitogenetic phylogenetic analyses yield new insight into
crocodylian evolution and their survival of the Cretaceous-Tertiary
boundary. Molecular Phylogenetics and Evolution. 45(2):663-673.
ROSS, C. A.
1989. (Ed.). Crocodiles and Alligators. Golden Press Pty Ltd. Silverwater, NSW,
Australia.
ROSS, C. A. Y W. E. MAGNUSSON
1989. Living crocodilians. Pp. 58-73 in: Ross, C. A. (Ed.). Crocodiles and
Alligators. Golden Press Pty Ltd. Silverwater, NSW, Australia.
ROSS, J. P.
1998. Crocodiles: Status survey and conservation action plan. Gland, Switzerland,
IUCN/SSC Crocodile Specialist Group.

Andrés Eloy Seijas
260
ROZE, J. A.
1964. Pilgrims of the river. Nat. Hist. 73:35-41.
SCHMIDT, K.
1924. Notes on Central American Crocodiles. Zoology 12(6): 79-92.
SCHALLER, G. Y P. CRAWSHAW
1982. Fishing behavior of Paraguayan caiman (Caiman crocodilus). Copeia
1982(1):66-72.
SEASHOLE, S. G., S. SWIESZ, C. ARMELL, C. ABERCROMBIE, Y C. HOPE
2000. A cost effective method for chemical restraint in crocodilians: initial
observations and species specific responses. Pp. 124-126 in: Proceedings of
the 15th Working Meeting of Crocodile Specialist Group, IUCN - Gland,
Switzerland.
SCOTT, N., A. AQUINO Y L. FITZGERALD
1990. Distribution, Habitats, and Conservation of the Caimans (Alligatoridae) of
Paraguay. V. Silv. Neotrop. 2(2):43-51.
SEEBACHER, F., C. E. FRANKLIN Y M. READ
2005. Diving behaviour of a reptile (Crocodylus johnstoni) in the wild: interactions
with heart rate and body temperature. Physiological and Biochemical
Zoology 78:1-8.
SEIJAS, A. E.
1979. Aspectos fundamentales sobre la alimentación y el metabolismo de la baba
(Caiman crocodilus) en las sabanas moduladas del estado Apure,
Venezuela. Tesis de Licenciatura. Escuela de Biología, Facultad de
Ciencias, UCV. Caracas.
1986a. Estimaciones poblacionales de babas (Caiman crocodilus) en los llanos
occidentales de Venezuela. V. Silv. Neotrop.1(1):24-30.
1986b.Situación actual de las poblaciones de babas y babillas (Caiman crocodilus)
en la región norte costera de Venezuela. Pp. 28-36 in: Crocodiles.
Proceeding of the 7th Working Meeting of the Crocodile Specialist Group
Meeting. IUCN-The World Conservation Union. Gland, Switzerland.
1986c. Situación actual del caimán de la costa, Crocodylus acutus, en Venezuela.
Pp. 96-108 in: Crocodiles. Proceeding of the 7th Working Meeting of the
Crocodile Specialist Group Meeting. IUCN-The World Conservation
Union. Gland, Switzerland

Los Crocodylia de Venezuela
261
SEIJAS, A. E.
1988. Habitat use by the American crocodile and the spectacled caiman coexisting
along the Venezuelan coastal region. Tesis de Maestría. School of Forest
Resources and Conservation. Universidad de Florida, Gainesville.
1990. Status of the American crocodile (Crocodylus acutus) in Venezuela. A
review. Pp. 144-156 in: Crocodiles. Proceedings of the 9th Working
Meeting of Crocodile Specialist Group (Volume 2). IUCN - The World
Conservation Union, Gland, Switzerland.
1991a. Crecimiento de caimanes de la costa (Crocodylus acutus) en condiciones
naturales. Pp. 36 in: VIII Jornadas Técnicas de Investigación UNELLEZ.
Guanare, 3 al 5 de abril de 1991 (Resumen).
1991b. El Caimán de la Costa no Encuentra Refugio. Carta Ecológica 56: 16-19.
1992. Status of Orinoco crocodile populations in some localities in the Venezuelan
llanos. Pp 79 in: Crocodiles. Proceedings of the 11th Working Meeting of
the Crocodile Specialist Group (Volume 2). IUCN-The World
Conservation Union, Gland, Switzerland.
1995. Captive breeding and rearing as conservation tool for the Orinoco crocodile.
Pp. 596-598 in: J. A. Bissonette y P. R. Krausman, eds. Integrating People
and Wildlife for a sustainable future. Proceedings of the First International
Wildlife Management Congress. The Wildlife Society, Bethesda, Md.
1996. Coexistencia de babas y caimanes en la región costera venezolana. Pp. 217-
253 in: Péfaur, J. E. (Ed.). Herpetología Neotropical. Actas II Congreso
Latinoamericano de Herpetología. Vol. II. Universidad de Los Andes.
Consejo de Publicaciones. Consejo de Desarrollo Científico, Humanístico y
Tecnológico, Mérida, Venezuela.
1998. The Orinoco crocodile (Crocodylus intermedius) in the Cojedes River
System, Venezuela: population status and ecological characteristics. Tesis
doctoral.Universidad de Florida. Gainesville.
2001. Caimanes y babas en la narrativa de Humboldt. Pp. 224-233 in: Aymar, G.
(Ed.). Alexander von Humboldt: Homenaje al Bicentenario de su llegada a
tierras venezolanas. Guanare, Portuguesa, Biollania, Edición Especial No.
7.
2002. Scale patterns of American crocodiles (Crocodylus acutus) from several
Venezuelan localities. Revista UNELLEZ de Ciencia y Tecnología 20: 118-
134.
2007a. Características distintivas y estado del conocimiento de las especies del
género Paleosuchus (Crocodylidae; Alligatoridae) en Venezuela. Mem.
Soc. Cienc. Nat. La Salle 166:27-44.

Andrés Eloy Seijas
262
SEIJAS, A. E.
2007b. Tendencias de las poblaciones del Caimán del Orinoco (Crocodylus
intermedius) en Venezuela: Balance de las investigaciones de los últimos
30 años. Pp.11-21 in: Seijas, A. E. (Ed.). Conservación del Caimán del
Orinoco. Memorias del III Taller para la Conservación del Caimán del
Orinoco. San Carlos (Cojedes, Venezuela), 17 al 19 de enero de 2007.
Biollania, Ed. Especial No. 8.
2008. Reserva de Fauna Silvestre Río Cojedes. Propuesta de Creación. Oficina
Nacional de Diversidad Biológica. Ministerio del Ambiente. Caracas.
SEIJAS, A. E. Y C. CHÁVEZ
1991. Conservación del caimán de la costa (Crocodylus acutus) en el río Yaracuy y
el Parque Nacional Laguna de Tacarigua, Informe para FUDENA. Caracas.
1996. Potencial de la laguna El Eneal, Estado Cojedes, para la conservación de la
fauna silvestre, especialmente de las especies baba (Caiman crocodilus) y
caimán (Crocodylus intermedius). Proyecto Convenio CONICIT
Gobernación del estado Cojedes No. PC-CO-009. UNELLEZ, Guanare,
Venezuela.
2000. Population status of the Orinoco crocodile (Crocodylus intermedius) in the
Cojedes river system, Venezuela. Biol. Conserv. 94(2000): 353-361.
2002. Reproductive status and nesting ecology of the Orinoco crocodile
(Crocodylus intermedius) in the Cojedes River System, Venezuela. V. Silv.
Neotrop.11(1-2):23-32.
SEIJAS, A. E., A. CHANG Y D. G. CORDERO.
1990. Cría en cautiverio de caimanes de la costa (Crocodylus acutus) con fines de
repoblamiento. Biollania 7: 13-26.
SEIJAS, A. E., A. LLOBET, MAGDDY JIMÉNEZ-ORÁA, J. M. MENDOZA, F.
GARAVITO Y Y. TERÁN
2002. Estado de las poblaciones del caimán del Orinoco (Crocodylus intermedius)
en Venezuela. Pp. 7-15 in: Velasco, A., Colomine, G., Villarroel, G. y
Quero, M. (Eds.). Memorias del Taller para la Conservación del Caimán
del Orinoco (Crocodylus intermedius) en Colombia y Venezuela. Facultad
de Ciencias, UCV. Caracas.
SEIJAS, A. E., C. RAMO, Y B. BUSTO
1989. Manejo de hábitat para la baba: ¿Utopía o Realidad? II Taller Sobre
Conservación y Manejo de la Baba (Caiman crocodilus) en Venezuela.
UNELLEZ, FUNDAFAUNA, FUNDACITE, MARNR. Guanare 21 al 23
de agosto de 1989.

Los Crocodylia de Venezuela
263
SEIJAS, A. E., E I. GONZÁLEZ
1994. Incubación artificial de huevos de caimán del Orinoco. Biollania 12(2):36-
41.
SEIJAS, A. E., H. CUEVAS Y N. GONZÁLEZ
2003. Adaptación al medio natural de babas (Caiman crocodilus) criadas en
cautiverio. Interciencia 28(6): 340-346.
SEIJAS A. E.,J. M. MENDOZA Y P. M. ÁVILA-MANJÓN
2010. Tendencias poblacionales a largo plazo del caimán del Orinoco (Crocodylus
intermedius) en el Sistema del Río Cojedes, Venezuela. Pp. 149-160 in:
Machado Allison A, Hernández O, Aguilera M, Seijas AE, Rojas-Suarez F,
editors. Simposio Investigación y Manejo de Fauna Silvestre en Venezuela
en homenaje al Dr Juhani Ojasti. Caracas: FUDECI, IZET, UNELLEZ,
USB, PROVITA, FLSCN, PDVS y JB "Dr. Tobias Lasser".
SEIJAS, A. E., Y P. MEZA
1994. El caimán del Orinoco (Crocodylus intermedius) en el embalse del río
Tucupido, estado Portuguesa, Venezuela. Biollania 10:42-51.
SEIJAS, A. E., Y S. RAMOS
1980. Caracterización de la dieta de la baba, Caiman crocodilus, durante la
estación seca en las sabanas moduladas del Estado Apure, Venezuela. Acta
Biol. Venez. 10(4):373-389.
SEIJAS, A. E., A. URDANETA Y T. BARROS
2008. Tasas de crecimiento durante el primer año de vida y la temporada de
eclosión de caimán de la costa (Crocodylus acutus) en Venezuela Boletín
del Centro de Investigaciones Biológicas 42(4): 445-459.
SEYMOUR, R. S., G. WEBB, A. F. BENNETT Y D. F. BRADFORD
1987. Effect of capture on the physiology of Crocodylus porosus. Pp. 253-257 in:
Webb, G. J. W, C. Manolis y P. Whitehead. (Eds.). Wildlife Management:
Crocodiles and Alligators. Surrey Beatty and Sons Pty Limited in
Association with the Conservation Commission of the Northern Territory.
Chipping Norton, NSW, Australia.
SILL, W.
1970. Nota preliminar sobre un nuevo Gavial del Plioceno de Venezuela y una
discusión de los gaviales sudamericanos. Ameghiniana. 7(2): 151-159.

Andrés Eloy Seijas
264
SIOLI, H.
1984. The Amazon and its main affluents: Hydrography, morphology of the
courses and river types. Pp. 127-165 in: Sioli, H. (ed.). The Amazon:
Limnology and landscape ecology of a mighty tropical river and its basin.
Dr. W. Junk Publishers, Dordrecht, The Neederlands.
SPOTILA, J. R., Y K. M. TERPIN
1977. Mouth gaping as an effective thermoregulatory device in alligators. Nature
265:235-236.
STATON, M. Y J. M. DIXON
1975. Studies on the dry season biology of Caiman crocodilus crocodilus from the
Venezuelan Llanos. Memorias de la Sociedad de Ciencias Naturales La
Salle. 35:237-265.
1977. Breeding biology of the spectacled caiman (Caiman crocodilus crocodilus)
in the Venezuelan Llanos. U.S. Fish and Wildlife Service Report 5:1-21.
SUES, H. D.
1989. The place of crocodilinas in the living World. Pp. 14-25 in: Ross, C. A.
Crocodiles and Alligators. Golden Press Pty Ltd. Silverwater, NSW,
Australia.
SWANEPOEL, D. G. J.
1996. Identification of the Nile crocodile Crocodylus niloticus by the use of natural
tail marks. Koedoe 39(1): 113-115.
SZEPLAKI, E.
1991. Desarrollo de zoocriaderos abiertos de la especie baba (Caiman crocodilus
crocodilus) en la región de los llanos de Venezuela. Pp. 196-207 in:
Zoocría de los Crocodylia. Memorias de la I Reunión del CSG, Grupo de
especialistas en Cocodrilos de la IUCN, The World Conservation Union.
Gland, Switzerland.
TABLANTE-GARRIDO, P. N.
1961. Provincia de Apure. Monografía del Gobernador General José Cornelio
Muñoz [1831]. Cuadernos Geográficos No. 1. Instituto de Geografía y de
Conservación de Recursos Naturales. Universidad de Los Andes.
Venezuela.
TAPHORN, D. C.
1992. The Characiform fishes of the Apure river drainage, Venezuela. Biollania,
Ed. Esp. No. 4. Guanare, Venezuela.

Los Crocodylia de Venezuela
265
TAYLOR, J. A.
1979. The food and feeding habits of subadult Crocodylus porosus Schneider in
Northern Australia. Aust. Wildl. Res. 6:347-359.
THORBJARNARSON, J. B.
1987. Status, ecology, and conservation of the Orinoco crocodile. Informe para
FUDENA. 74 p.
THORBJARNARSON, J. B.
1988. The status and ecology of the American crocodile in Haiti. Bulletin of
Florida State Museum Biologica Sciences 33(1): 1-86.
1989. Ecology of the American crocodile, Crocodylus acutus. Pp. 228-258 in:
Crocodiles, Their Ecology, Management and ConservationA Special
Publication of the Crocodile Specialist Group. IUCN Publications, N.S.,
Gland, Switzerland.
1990. Ecology and behavior of the spectacled caiman (Caiman crocodilus) in the
central Venezuelan llanos. Tesis doctoral. School of Forest Resources and
Conservation. Universidad de Florida. Gainesville.
1991. An analysis of the spectacled caiman (Caiman crocodilus) harvest program
in Venezuela. Pp. 217-235 in: Robinson, J. G. y Redford, K. H. (Ed.)
Neotropical Wildlife Use and Conservation. University of Chicago Press,
Chicago and London.
1996. Reproductive characteristics of the Order Crocodylia. Herpetologica
52(1):8-24.
1999. Fundo Pecuario Masaguaral. Crocodile Nesting 1998-1999 Year Summary.
Manuscrito sin publicar.
2002. Commercial consumptive use of crocodilians: A conservation Panacea or
Pitfall. Pp. 11 in: Crocodiles. Proceedings of the 16th Working Meeting of
the Crocodile Specialist Group, SSC. IUCN- The World Conservation
Union. Gland, Switzerland.
1993. Fishing behavior of the spectacled caiman in the Venezuelan llanos. Copeia
1993:1166-1171.
THORBJARNARSON, J. B. Y A. VELASCO
1998a. Venezuela´s caiman harvest program. An historical perspectiva and
analysis of its conservation benefits. Wildlife Conservation Program.
Working Paper No. 11.
1998b. Economic incentives for management of Venezuelan caiman. Conserv.
Biol. 13(2): 397-406.
THORBJARNARSON, J. Y G. HERNÁNDEZ
1992. Recent investigations on the status and distribution of Orinoco crocodile
Crocodylus intermedius in Venezuela. Biol. Conserv. 62:179-188.

Andrés Eloy Seijas
266
THORBJARNARSON, J. Y G. HERNÁNDEZ
1993a. Reproductive ecology of the Orinoco crocodile (Crocodylus intermedius) in
Venezuela. I. Nesting ecology and egg and clutch relationship. J. Herpetol.
27(4):363-370.
1993b. Reproductive ecology of the Orinoco crocodile (Crocodylus intermedius) in
Venezuela. II. Reproductive and social behavior. J. Herpetol. 27(4):371-
379.
THORBJARNARSON, J. Y M. RODRIGUEZ
1992. Manual del zoocriadero de caimán del Orinoco para personal del Hato
Masaguaral, Estado Guárico. Manuscrito sin publicar.
THORBJARNARSON, J. Y T. BLOHM
1986. Captive rearing of Orinoco crocodile on hato Masaguaral, Venezuela. Pp.
120-123 in: Crocodiles. Proceeding of the 7th Working Meeting of the
Crocodile Specialist Group Meeting. IUCN-The World Conservation
Union, Gland, Switzerland.
THORBJARNARSON, J., F. MAZZOTTI, E. SANDERSON, F. BUITRAGO, M.
LAZCANO, K. MINKOWSKI, M. MUÑIZ, P. PONCE, L. SIGLER, R. SOBERON, A. M.
TRELANCIA Y A. VELASCO
2006. Regional habitat conservation priorities for the American crocodile. Biol.
Conserv. 128 (2006): 26-36.
URDANETA, A.
2002. Evaluación poblacional del caimán de la costa (Crocodylus acutus) en el
R.F.S.R.P Ciénaga de Los Olivitos y el embalse Pueblo Viejo, Estado
Zulia, Tesis de licenciatura. Facultad Experimental de Ciencias.
Universidad del Zulia. Maracaibo, Venezuela.
VALERIS, C.
2007. Estado poblacional, distribución espacial y hábitat del caimán de la costa
(Crocodylus acutus) en el rio Santa Rosa, Zulia, Venezuela. Tesis de
licenciatura. Facultad Experimental de Ciencias. Universidad del Zulia.
Maracaibo, Venezuela.
VARONA, L.
1976. Caiman crocodilus (Reptilia: Alligatoridae) en Cuba. Miscelánea Zoológica
(5): 2.
VELASCO, A. Y J. AYARZAGÜENA
1995. Situación actual de las poblaciones de babas (Caiman crocodilus) sometidas
a aprovechamiento comercial en los Llanos venezolanos. Publ. Asoc.
Amigos de Doñana 5: 71 pp.

Los Crocodylia de Venezuela
267
VELASCO, A. Y V. BLANCO
1996. Population evaluation of the spectacled caiman (Caiman crocodilus) in the
Orinoco Delta. Pp. 107-110 in: Crocodiles. Proceedings of the 13th
Working Meeting of Crocodile Specialist Group, SSC. IUCN –The World
Conservation Union. Gland, Switzerland.
VELASCO A., R. DE SOLA Y M. QUERO
1995. Programa de manejo de la baba (Caiman crocodilus) de Venezuela. Pp. 213-
220 in: Larriera A, Verdade LM, editors. La conservación y manejo de
caimanes y cocodrilos de América Latina. Santa Fe, Argentina: Fundación
Banco Bica. p 213-220.
VELASCO, A., G. COLOMINE, G. VILLARROEL, O. CAMACARO, R. DE SOLA, N.
LEÓN, E. OROPEZA, R. PÉREZ-HERNÁNDEZ, T. PINO, M. QUERO, J. RAMOS, S.
RAMOS, G. SÁNCHEZ Y W. VÁSQUEZ
1996. Surveying nests of spectacled caiman (Caiman crocodilus) in Ecological
Areas of Venezuelan Llanos. Pp. 111-116 in: Crocodiles. Proceedings of
the 13th Working Meeting of Crocodile Specialist Group, SSC. IUCN –
The World Conservation Union. Gland, Switzerland.
VELASCO, A., G. COLOMINE, R. DE SOLA Y G. VILLARROEL
2003. Effects of sustained harvest on wild populations of Caiman crocodilus
crocodilus in Venezuela. Interciencia 28(9): 544-548.
VENEZUELA.
1970. Ley de Protección a la Fauna Silvestre. Gaceta Oficial de la República de
Venezuela No. 29289.
1973. Veda general para todas las especies de la fauna silvestre en todo el territorio
de la República. Resolución Nº RNR-5-143. Gaceta Oficial de la
República de Venezuela No. 30.072.
1979. Veda el todo el Territorio Nacional para la Caza. Gaceta Oficial de la
República de Venezuela No. 31.901.
1996. Especies de fauna en peligro de extinción (Decreto No. 1486). Gaceta
Oficial de la República de Venezuela No. 36.062.
1999. Normas para el manejo racional de la especie baba (Caiman crocodilus).
Resolución Nº 23. Gaceta Oficial de la República de Venezuela Nº 36.653.
VILLARROEL, G., A. VELASCO, R. DE SOLA, G. COLOMINE, A. LANDER, T. PINO,
W. VASQUEZ Y J. CORAZZELLI
2000. Effect of sustained harvest in the spectacled caiman populations (Caiman
crocodilus) in the Alto and Bajo Apure regions. Pp. 298-303 in: Crocodiles.
Proceedings of the 15th Working Meeting of Crocodile Specialist Group, SSC.
IUCN –The World Conservation Unio. Gland, Switzerland.

Andrés Eloy Seijas
268
WALSH, B.
1987. Crocodile capture methods used in the Northern Territory of Australia. Pp.
249-252 in: Webb, G. J. W, C. Manolis y P. Whitehead. (Eds.). Wildlife
Management: Crocodiles and Alligators. Surrey Beatty and Sons Pty
Limited in Association with the Conservation Commission of the Northern
Territory. Chipping Norton, NSW, Australia.
WATLINGTON, F.
2002. Stranger in a lost paradise: Caiman crocodilus, Puerto Rico´s own
aliengator. Pp. 239-243 in: Crocodiles. Proceedings of the 16th Working
Meeting of the Crocodile Specialist Group, SSC. IUCN- The World
Conservation Union. Gland, Switzerland.
WATSON, R. M., A. D. GRAHAM, R. H. V. BELL Y I. S. C. PARKER
1971. A comparison of four east African crocodile (Crocodylus niloticus Laurenti)
populations. East Afric. Wildl. J. 1971(9): 25-34.
WEBB, G. J. W. Y H. MESSEL
1978. Morphometric analysis of Crocodylus porosus from the north coast of
Arnhem Land, Northern Australia. Aust. J. Zool. 26: 1-27.
1977. Crocodile capture techniques. J. Wildl. Manag. 41(3): 228-231.
WEBB, G., C. MANOLIS Y R. BUCKWORTH
1982. Crocodylus johnstoni in the McKinlay river area, N.T. 1. Variation in diet,
and a new method of assesing the relative importance of prey. Austr. J.
Zool. 30: 877-899.
WEBB, G., C. MANOLIS, Y G. SACK
1983. Crocodylus johstoni and C. porosus coexisting in a tidal river. Austr. Wildl.
Res. 10:639-650.
WHITNEY, C.
1912. The flowing road. Adventuring on the great rivers of South America. J. B.
Lippincott Company, Philadelphia & London.
WILKINSON, P. M. Y K.G. RICE
1996. Hind-foot track length: a method for determining the size of American
alligators. Pp. 429-436 in: Crocodiles. Proceedings of the 13th Working
Meeting of Crocodile Specialist Group, SSC. IUCN –The World
Conservation Union. Gland, Switzerland.

Los Crocodylia de Venezuela
269
WOODWARD, A. R., Y W. R. MARION
1978. An evaluation of factors affecting night-light counts of alligators. Proc. Ann.
Conf. S. E. Assoc. Fish & Wildl. Agencies 32:291-302.
WOODWARD, A. Y D. DAVID
1985. A Study of the Baba (Caiman crocodilus crocodilus) Population on the Hato
Piñero in Venezuela. Manuscrito sin publicar.
WOODWARD, A., K. G. RICE Y S. B. LINDA
1996. Estimating sighting proportions of American alligator during night-light and
aerial helicopter surveys. Proc. Annu. Conf. SEAFWA 50:509-519.

Andrés Eloy Seijas
270
Los Crocodylia de Venezuela
271
APÉNDICES
Apéndice I. Valores necesarios para calcular el tamaño estimado de una variable morfométrica, y sus
límites de confianza, a partir de otra variable en babas (Caiman crocodilus). En "Selección" se indica
cuales individuos fueron usados para generar la ecuación de regresión: Grupo 1, individuos menores a
350 mm de LCC; Grupo 2, individuos mayores o iguales a 350 mm de LCC; n, tamaño de la muestra.
Los significados de Xm, Sxx y s, se explican en el texto.
Babas región costera
Ec. Variables Selección n Intercepto Pendiente Xm Sxx s R2
I.1 LT vs LCC Todos 73 34,7 1,850 404,8 4977018,4 24,90 0,994
I.2 LT vs Lcab Todos 69 -36,8 7,624 109,3 129998,1 39,29 0,986
I.3 Lt vs Lcab Grupo 1 31 3,5 7,095 70,5 8055,8 29,29 0,940
I.4 Lt vs Lcab Grupo 2 38 -80,4 7,915 141,0 37145,7 44,91 0,970
I.5 Lt vs Lhoc Todos 71 99,8 11,374 60,7 57176,9 34,40 0,989
I.6 Lt vs Aph Todos 64 -97,7 23,155 39,1 12089,4 74,43 0,950
I.7 LCC vs Lcab Todos 107 -36,8 4,073 126,1 2446621,1 20,86 0,988
I.8 LCC vs Lcab Grupo 1 35 -8,9 3,747 71,7 9774,0 10,32 0,975
I.9 LCC vs Lcab Grupo 2 72 -68,8 4,269 152,5 81072,6 22,99 0,976
I.10 LCC vs Lhoc Todos 110 40,5 6,007 71,9 112098,7 19,74 0,990
I.11 LCC vs Aph Todos 103 -67,5 12,364 44,5 23497,2 46,69 0,942
I.12 Lcab vs Lhoc Todos 132 20,2 1,457 74,5 142333,2 3,10 0,996
I.13 Lcab vs Aph Todos 125 -3,6 2,938 45,9 31377,9 10,58 0,952
I.14 Lhoc vs Aph Todos 128 -15,3 1,989 45,8 31762,6 7,97 0,940
Babas región llanera
Ec. Variables Selección n Intercepto pendiente Xm Sxx s R2
I.15 LT vs LCC Todos 94 27,3 1,864 382,8 1739299,9 20,07 0,993
I.16 LT vs Lcab Todos 95 -32,1 7,779 98,7 92415,9 26,71 0,998
I.17 Lt vs Lcab Grupo 1 40 -30,5 7,667 70,6 12207,6 23,66 0,971
I.18 Lt vs Lcab Grupo 2 55 23,9 7,349 119,2 25665,7 26,54 0,974
I.19 Lt vs Lhoc Todos 15 68,5 12,272 46,3 18932,4 60,33 0,984
I.20 Lt vs Aph Todos 15 -60,0 23,109 23,5 2060,5 44,18 0,977
I.21 LCC vs Lcab Todos 135 -28,7 4,148 105,3 170273,0 17,13 0,987
I.22 LCC vs Lcab Grupo 1 83 -17,0 3,909 72,1 13275,6 11,20 0,972
I.23 LCC vs Lcab Grupo 2 87 -21,4 4,029 123,6 74827,1 23,89 0,972
I.24 LCC vs Lhoc Todos 20 33,8 6,484 65,0 40958,9 42,41 0,982
I.25 LCC vs Aph Todos 19 -63,2 13,439 32,5 7832,0 28,36 0,99
I.26 Lcab vs Lhoc Todos 47 18,6 1,474 92,8 198762,6 4,40 0,997
I.27 Lcab vs Aph Todos 47 -7,8 3,313 47,5 47331,7 12,02 0,978
I.28 Lhoc vs Aph Todos 45 -17,1 2,235 48,7 25172,0 9,41 0,971
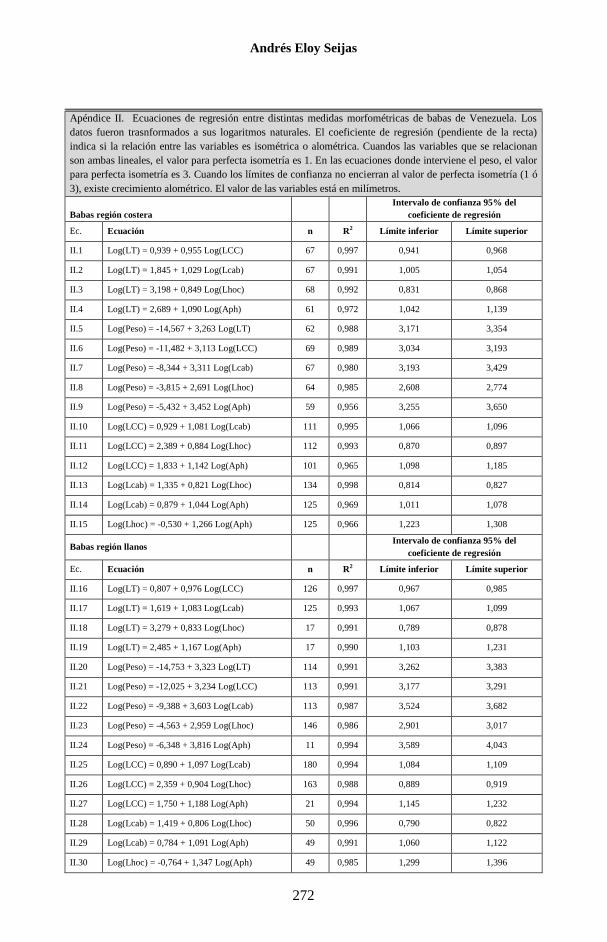
Andrés Eloy Seijas
272
Apéndice II. Ecuaciones de regresión entre distintas medidas morfométricas de babas de Venezuela. Los
datos fueron trasnformados a sus logaritmos naturales. El coeficiente de regresión (pendiente de la recta)
indica si la relación entre las variables es isométrica o alométrica. Cuandos las variables que se relacionan
son ambas lineales, el valor para perfecta isometría es 1. En las ecuaciones donde interviene el peso, el valor
para perfecta isometría es 3. Cuando los límites de confianza no encierran al valor de perfecta isometría (1 ó
3), existe crecimiento alométrico. El valor de las variables está en milímetros.
Babas región costera
Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación n R2 Límite inferior Límite superior
II.1 Log(LT) = 0,939 + 0,955 Log(LCC) 67 0,997 0,941 0,968
II.2 Log(LT) = 1,845 + 1,029 Log(Lcab) 67 0,991 1,005 1,054
II.3 Log(LT) = 3,198 + 0,849 Log(Lhoc) 68 0,992 0,831 0,868
II.4 Log(LT) = 2,689 + 1,090 Log(Aph) 61 0,972 1,042 1,139
II.5 Log(Peso) = -14,567 + 3,263 Log(LT) 62 0,988 3,171 3,354
II.6 Log(Peso) = -11,482 + 3,113 Log(LCC) 69 0,989 3,034 3,193
II.7 Log(Peso) = -8,344 + 3,311 Log(Lcab) 67 0,980 3,193 3,429
II.8 Log(Peso) = -3,815 + 2,691 Log(Lhoc) 64 0,985 2,608 2,774
II.9 Log(Peso) = -5,432 + 3,452 Log(Aph) 59 0,956 3,255 3,650
II.10 Log(LCC) = 0,929 + 1,081 Log(Lcab) 111 0,995 1,066 1,096
II.11 Log(LCC) = 2,389 + 0,884 Log(Lhoc) 112 0,993 0,870 0,897
II.12 Log(LCC) = 1,833 + 1,142 Log(Aph) 101 0,965 1,098 1,185
II.13 Log(Lcab) = 1,335 + 0,821 Log(Lhoc) 134 0,998 0,814 0,827
II.14 Log(Lcab) = 0,879 + 1,044 Log(Aph) 125 0,969 1,011 1,078
II.15 Log(Lhoc) = -0,530 + 1,266 Log(Aph) 125 0,966 1,223 1,308
Babas región llanos Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación n R2 Límite inferior Límite superior
II.16 Log(LT) = 0,807 + 0,976 Log(LCC) 126 0,997 0,967 0,985
II.17 Log(LT) = 1,619 + 1,083 Log(Lcab) 125 0,993 1,067 1,099
II.18 Log(LT) = 3,279 + 0,833 Log(Lhoc) 17 0,991 0,789 0,878
II.19 Log(LT) = 2,485 + 1,167 Log(Aph) 17 0,990 1,103 1,231
II.20 Log(Peso) = -14,753 + 3,323 Log(LT) 114 0,991 3,262 3,383
II.21 Log(Peso) = -12,025 + 3,234 Log(LCC) 113 0,991 3,177 3,291
II.22 Log(Peso) = -9,388 + 3,603 Log(Lcab) 113 0,987 3,524 3,682
II.23 Log(Peso) = -4,563 + 2,959 Log(Lhoc) 146 0,986 2,901 3,017
II.24 Log(Peso) = -6,348 + 3,816 Log(Aph) 11 0,994 3,589 4,043
II.25 Log(LCC) = 0,890 + 1,097 Log(Lcab) 180 0,994 1,084 1,109
II.26 Log(LCC) = 2,359 + 0,904 Log(Lhoc) 163 0,988 0,889 0,919
II.27 Log(LCC) = 1,750 + 1,188 Log(Aph) 21 0,994 1,145 1,232
II.28 Log(Lcab) = 1,419 + 0,806 Log(Lhoc) 50 0,996 0,790 0,822
II.29 Log(Lcab) = 0,784 + 1,091 Log(Aph) 49 0,991 1,060 1,122
II.30 Log(Lhoc) = -0,764 + 1,347 Log(Aph) 49 0,985 1,299 1,396
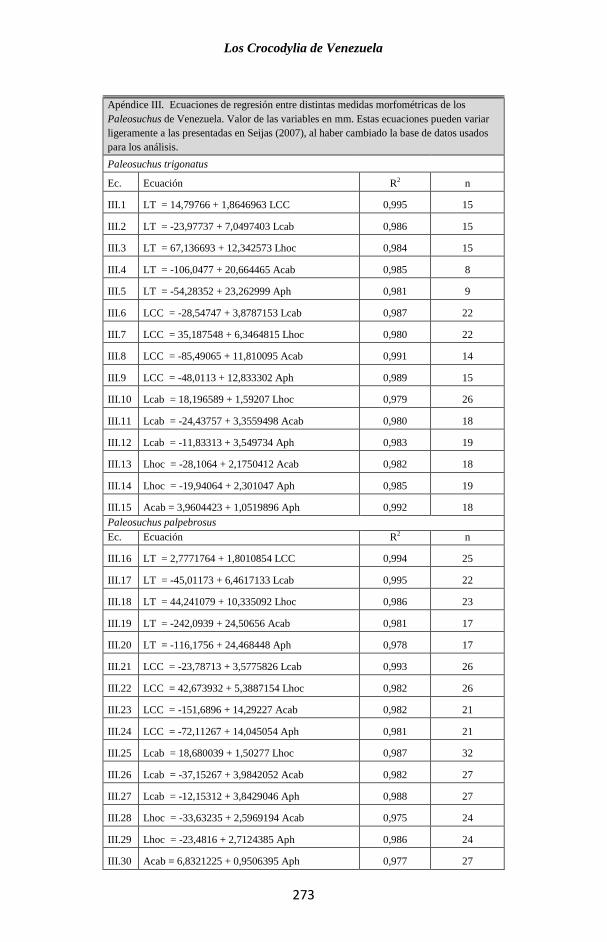
Los Crocodylia de Venezuela
273
Apéndice III. Ecuaciones de regresión entre distintas medidas morfométricas de los
Paleosuchus de Venezuela. Valor de las variables en mm. Estas ecuaciones pueden variar
ligeramente a las presentadas en Seijas (2007), al haber cambiado la base de datos usados
para los análisis.
Paleosuchus trigonatus
Ec. Ecuación R2 n
III.1 LT = 14,79766 + 1,8646963 LCC 0,995 15
III.2 LT = -23,97737 + 7,0497403 Lcab 0,986 15
III.3 LT = 67,136693 + 12,342573 Lhoc 0,984 15
III.4 LT = -106,0477 + 20,664465 Acab 0,985 8
III.5 LT = -54,28352 + 23,262999 Aph 0,981 9
III.6 LCC = -28,54747 + 3,8787153 Lcab 0,987 22
III.7 LCC = 35,187548 + 6,3464815 Lhoc 0,980 22
III.8 LCC = -85,49065 + 11,810095 Acab 0,991 14
III.9 LCC = -48,0113 + 12,833302 Aph 0,989 15
III.10 Lcab = 18,196589 + 1,59207 Lhoc 0,979 26
III.11 Lcab = -24,43757 + 3,3559498 Acab 0,980 18
III.12 Lcab = -11,83313 + 3,549734 Aph 0,983 19
III.13 Lhoc = -28,1064 + 2,1750412 Acab 0,982 18
III.14 Lhoc = -19,94064 + 2,301047 Aph 0,985 19
III.15 Acab = 3,9604423 + 1,0519896 Aph 0,992 18
Paleosuchus palpebrosus
Ec. Ecuación R2 n
III.16 LT = 2,7771764 + 1,8010854 LCC 0,994 25
III.17 LT = -45,01173 + 6,4617133 Lcab 0,995 22
III.18 LT = 44,241079 + 10,335092 Lhoc 0,986 23
III.19 LT = -242,0939 + 24,50656 Acab 0,981 17
III.20 LT = -116,1756 + 24,468448 Aph 0,978 17
III.21 LCC = -23,78713 + 3,5775826 Lcab 0,993 26
III.22 LCC = 42,673932 + 5,3887154 Lhoc 0,982 26
III.23 LCC = -151,6896 + 14,29227 Acab 0,982 21
III.24 LCC = -72,11267 + 14,045054 Aph 0,981 21
III.25 Lcab = 18,680039 + 1,50277 Lhoc 0,987 32
III.26 Lcab = -37,15267 + 3,9842052 Acab 0,982 27
III.27 Lcab = -12,15312 + 3,8429046 Aph 0,988 27
III.28 Lhoc = -33,63235 + 2,5969194 Acab 0,975 24
III.29 Lhoc = -23,4816 + 2,7124385 Aph 0,986 24
III.30 Acab = 6,8321225 + 0,9506395 Aph 0,977 27
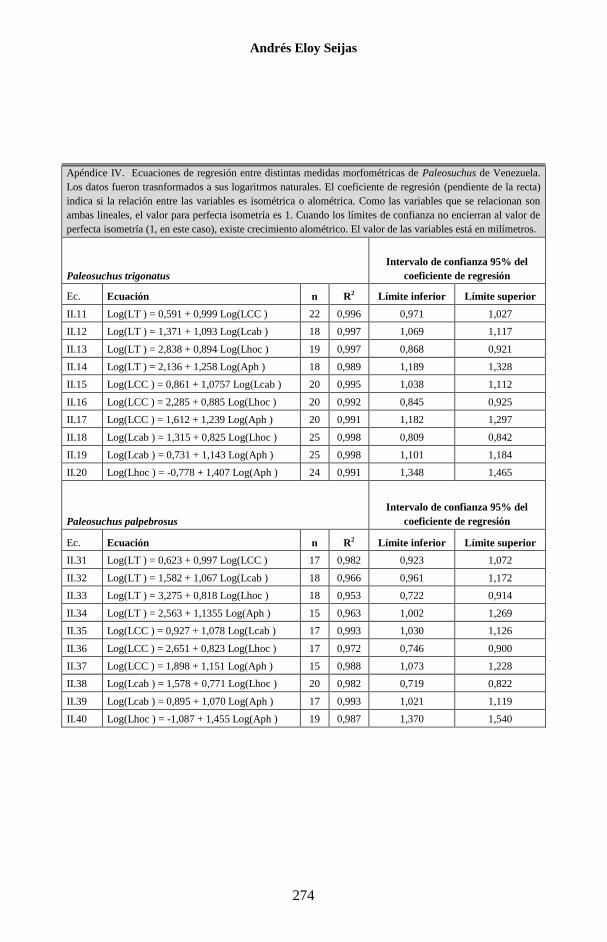
Andrés Eloy Seijas
274
Apéndice IV. Ecuaciones de regresión entre distintas medidas morfométricas de Paleosuchus de Venezuela.
Los datos fueron trasnformados a sus logaritmos naturales. El coeficiente de regresión (pendiente de la recta)
indica si la relación entre las variables es isométrica o alométrica. Como las variables que se relacionan son
ambas lineales, el valor para perfecta isometría es 1. Cuando los límites de confianza no encierran al valor de
perfecta isometría (1, en este caso), existe crecimiento alométrico. El valor de las variables está en milímetros.
Paleosuchus trigonatus
Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación n R2 Límite inferior Límite superior
II.11 Log(LT ) = 0,591 + 0,999 Log(LCC ) 22 0,996 0,971 1,027
II.12 Log(LT ) = 1,371 + 1,093 Log(Lcab ) 18 0,997 1,069 1,117
II.13 Log(LT ) = 2,838 + 0,894 Log(Lhoc ) 19 0,997 0,868 0,921
II.14 Log(LT ) = 2,136 + 1,258 Log(Aph ) 18 0,989 1,189 1,328
II.15 Log(LCC ) = 0,861 + 1,0757 Log(Lcab ) 20 0,995 1,038 1,112
II.16 Log(LCC ) = 2,285 + 0,885 Log(Lhoc ) 20 0,992 0,845 0,925
II.17 Log(LCC ) = 1,612 + 1,239 Log(Aph ) 20 0,991 1,182 1,297
II.18 Log(Lcab ) = 1,315 + 0,825 Log(Lhoc ) 25 0,998 0,809 0,842
II.19 Log(Lcab ) = 0,731 + 1,143 Log(Aph ) 25 0,998 1,101 1,184
II.20 Log(Lhoc ) = -0,778 + 1,407 Log(Aph ) 24 0,991 1,348 1,465
Paleosuchus palpebrosus
Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación n R2 Límite inferior Límite superior
II.31 Log(LT ) = 0,623 + 0,997 Log(LCC ) 17 0,982 0,923 1,072
II.32 Log(LT ) = 1,582 + 1,067 Log(Lcab ) 18 0,966 0,961 1,172
II.33 Log(LT ) = 3,275 + 0,818 Log(Lhoc ) 18 0,953 0,722 0,914
II.34 Log(LT ) = 2,563 + 1,1355 Log(Aph ) 15 0,963 1,002 1,269
II.35 Log(LCC ) = 0,927 + 1,078 Log(Lcab ) 17 0,993 1,030 1,126
II.36 Log(LCC ) = 2,651 + 0,823 Log(Lhoc ) 17 0,972 0,746 0,900
II.37 Log(LCC ) = 1,898 + 1,151 Log(Aph ) 15 0,988 1,073 1,228
II.38 Log(Lcab ) = 1,578 + 0,771 Log(Lhoc ) 20 0,982 0,719 0,822
II.39 Log(Lcab ) = 0,895 + 1,070 Log(Aph ) 17 0,993 1,021 1,119
II.40 Log(Lhoc ) = -1,087 + 1,455 Log(Aph ) 19 0,987 1,370 1,540
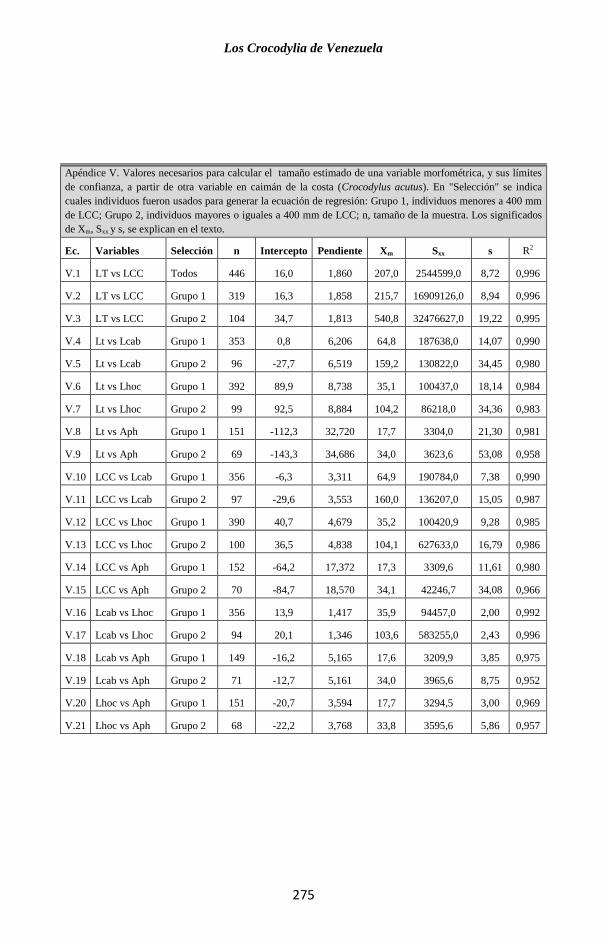
Los Crocodylia de Venezuela
275
Apéndice V. Valores necesarios para calcular el tamaño estimado de una variable morfométrica, y sus límites
de confianza, a partir de otra variable en caimán de la costa (Crocodylus acutus). En "Selección" se indica
cuales individuos fueron usados para generar la ecuación de regresión: Grupo 1, individuos menores a 400 mm
de LCC; Grupo 2, individuos mayores o iguales a 400 mm de LCC; n, tamaño de la muestra. Los significados
de Xm, Sxx y s, se explican en el texto.
Ec. Variables Selección n Intercepto Pendiente Xm Sxx s R2
V.1 LT vs LCC Todos 446 16,0 1,860 207,0 2544599,0 8,72 0,996
V.2 LT vs LCC Grupo 1 319 16,3 1,858 215,7 16909126,0 8,94 0,996
V.3 LT vs LCC Grupo 2 104 34,7 1,813 540,8 32476627,0 19,22 0,995
V.4 Lt vs Lcab Grupo 1 353 0,8 6,206 64,8 187638,0 14,07 0,990
V.5 Lt vs Lcab Grupo 2 96 -27,7 6,519 159,2 130822,0 34,45 0,980
V.6 Lt vs Lhoc Grupo 1 392 89,9 8,738 35,1 100437,0 18,14 0,984
V.7 Lt vs Lhoc Grupo 2 99 92,5 8,884 104,2 86218,0 34,36 0,983
V.8 Lt vs Aph Grupo 1 151 -112,3 32,720 17,7 3304,0 21,30 0,981
V.9 Lt vs Aph Grupo 2 69 -143,3 34,686 34,0 3623,6 53,08 0,958
V.10 LCC vs Lcab Grupo 1 356 -6,3 3,311 64,9 190784,0 7,38 0,990
V.11 LCC vs Lcab Grupo 2 97 -29,6 3,553 160,0 136207,0 15,05 0,987
V.12 LCC vs Lhoc Grupo 1 390 40,7 4,679 35,2 100420,9 9,28 0,985
V.13 LCC vs Lhoc Grupo 2 100 36,5 4,838 104,1 627633,0 16,79 0,986
V.14 LCC vs Aph Grupo 1 152 -64,2 17,372 17,3 3309,6 11,61 0,980
V.15 LCC vs Aph Grupo 2 70 -84,7 18,570 34,1 42246,7 34,08 0,966
V.16 Lcab vs Lhoc Grupo 1 356 13,9 1,417 35,9 94457,0 2,00 0,992
V.17 Lcab vs Lhoc Grupo 2 94 20,1 1,346 103,6 583255,0 2,43 0,996
V.18 Lcab vs Aph Grupo 1 149 -16,2 5,165 17,6 3209,9 3,85 0,975
V.19 Lcab vs Aph Grupo 2 71 -12,7 5,161 34,0 3965,6 8,75 0,952
V.20 Lhoc vs Aph Grupo 1 151 -20,7 3,594 17,7 3294,5 3,00 0,969
V.21 Lhoc vs Aph Grupo 2 68 -22,2 3,768 33,8 3595,6 5,86 0,957
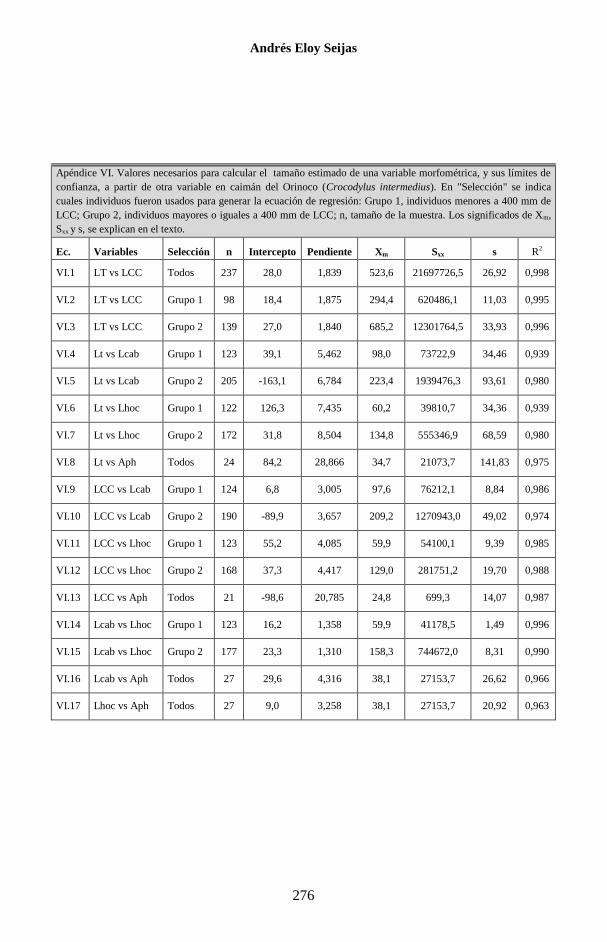
Andrés Eloy Seijas
276
Apéndice VI. Valores necesarios para calcular el tamaño estimado de una variable morfométrica, y sus límites de
confianza, a partir de otra variable en caimán del Orinoco (Crocodylus intermedius). En "Selección" se indica
cuales individuos fueron usados para generar la ecuación de regresión: Grupo 1, individuos menores a 400 mm de
LCC; Grupo 2, individuos mayores o iguales a 400 mm de LCC; n, tamaño de la muestra. Los significados de Xm,
Sxx y s, se explican en el texto.
Ec. Variables Selección n Intercepto Pendiente Xm Sxx s R2
VI.1 LT vs LCC Todos 237 28,0 1,839 523,6 21697726,5 26,92 0,998
VI.2 LT vs LCC Grupo 1 98 18,4 1,875 294,4 620486,1 11,03 0,995
VI.3 LT vs LCC Grupo 2 139 27,0 1,840 685,2 12301764,5 33,93 0,996
VI.4 Lt vs Lcab Grupo 1 123 39,1 5,462 98,0 73722,9 34,46 0,939
VI.5 Lt vs Lcab Grupo 2 205 -163,1 6,784 223,4 1939476,3 93,61 0,980
VI.6 Lt vs Lhoc Grupo 1 122 126,3 7,435 60,2 39810,7 34,36 0,939
VI.7 Lt vs Lhoc Grupo 2 172 31,8 8,504 134,8 555346,9 68,59 0,980
VI.8 Lt vs Aph Todos 24 84,2 28,866 34,7 21073,7 141,83 0,975
VI.9 LCC vs Lcab Grupo 1 124 6,8 3,005 97,6 76212,1 8,84 0,986
VI.10 LCC vs Lcab Grupo 2 190 -89,9 3,657 209,2 1270943,0 49,02 0,974
VI.11 LCC vs Lhoc Grupo 1 123 55,2 4,085 59,9 54100,1 9,39 0,985
VI.12 LCC vs Lhoc Grupo 2 168 37,3 4,417 129,0 281751,2 19,70 0,988
VI.13 LCC vs Aph Todos 21 -98,6 20,785 24,8 699,3 14,07 0,987
VI.14 Lcab vs Lhoc Grupo 1 123 16,2 1,358 59,9 41178,5 1,49 0,996
VI.15 Lcab vs Lhoc Grupo 2 177 23,3 1,310 158,3 744672,0 8,31 0,990
VI.16 Lcab vs Aph Todos 27 29,6 4,316 38,1 27153,7 26,62 0,966
VI.17 Lhoc vs Aph Todos 27 9,0 3,258 38,1 27153,7 20,92 0,963
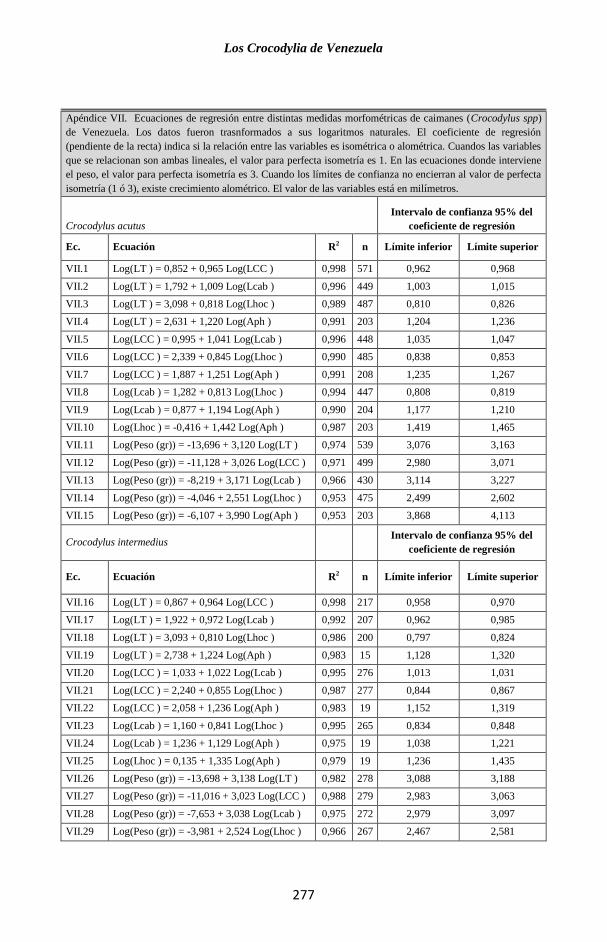
Los Crocodylia de Venezuela
277
Apéndice VII. Ecuaciones de regresión entre distintas medidas morfométricas de caimanes (Crocodylus spp)
de Venezuela. Los datos fueron trasnformados a sus logaritmos naturales. El coeficiente de regresión
(pendiente de la recta) indica si la relación entre las variables es isométrica o alométrica. Cuandos las variables
que se relacionan son ambas lineales, el valor para perfecta isometría es 1. En las ecuaciones donde interviene
el peso, el valor para perfecta isometría es 3. Cuando los límites de confianza no encierran al valor de perfecta
isometría (1 ó 3), existe crecimiento alométrico. El valor de las variables está en milímetros.
Crocodylus acutus
Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación R2 n Límite inferior Límite superior
VII.1 Log(LT ) = 0,852 + 0,965 Log(LCC ) 0,998 571 0,962 0,968
VII.2 Log(LT ) = 1,792 + 1,009 Log(Lcab ) 0,996 449 1,003 1,015
VII.3 Log(LT ) = 3,098 + 0,818 Log(Lhoc ) 0,989 487 0,810 0,826
VII.4 Log(LT ) = 2,631 + 1,220 Log(Aph ) 0,991 203 1,204 1,236
VII.5 Log(LCC ) = 0,995 + 1,041 Log(Lcab ) 0,996 448 1,035 1,047
VII.6 Log(LCC ) = 2,339 + 0,845 Log(Lhoc ) 0,990 485 0,838 0,853
VII.7 Log(LCC ) = 1,887 + 1,251 Log(Aph ) 0,991 208 1,235 1,267
VII.8 Log(Lcab ) = 1,282 + 0,813 Log(Lhoc ) 0,994 447 0,808 0,819
VII.9 Log(Lcab ) = 0,877 + 1,194 Log(Aph ) 0,990 204 1,177 1,210
VII.10 Log(Lhoc ) = -0,416 + 1,442 Log(Aph ) 0,987 203 1,419 1,465
VII.11 Log(Peso (gr)) = -13,696 + 3,120 Log(LT ) 0,974 539 3,076 3,163
VII.12 Log(Peso (gr)) = -11,128 + 3,026 Log(LCC ) 0,971 499 2,980 3,071
VII.13 Log(Peso (gr)) = -8,219 + 3,171 Log(Lcab ) 0,966 430 3,114 3,227
VII.14 Log(Peso (gr)) = -4,046 + 2,551 Log(Lhoc ) 0,953 475 2,499 2,602
VII.15 Log(Peso (gr)) = -6,107 + 3,990 Log(Aph ) 0,953 203 3,868 4,113
Crocodylus intermedius Intervalo de confianza 95% del
coeficiente de regresión
Ec. Ecuación R2 n Límite inferior Límite superior
VII.16 Log(LT ) = 0,867 + 0,964 Log(LCC ) 0,998 217 0,958 0,970
VII.17 Log(LT ) = 1,922 + 0,972 Log(Lcab ) 0,992 207 0,962 0,985
VII.18 Log(LT ) = 3,093 + 0,810 Log(Lhoc ) 0,986 200 0,797 0,824
VII.19 Log(LT ) = 2,738 + 1,224 Log(Aph ) 0,983 15 1,128 1,320
VII.20 Log(LCC ) = 1,033 + 1,022 Log(Lcab ) 0,995 276 1,013 1,031
VII.21 Log(LCC ) = 2,240 + 0,855 Log(Lhoc ) 0,987 277 0,844 0,867
VII.22 Log(LCC ) = 2,058 + 1,236 Log(Aph ) 0,983 19 1,152 1,319
VII.23 Log(Lcab ) = 1,160 + 0,841 Log(Lhoc ) 0,995 265 0,834 0,848
VII.24 Log(Lcab ) = 1,236 + 1,129 Log(Aph ) 0,975 19 1,038 1,221
VII.25 Log(Lhoc ) = 0,135 + 1,335 Log(Aph ) 0,979 19 1,236 1,435
VII.26 Log(Peso (gr)) = -13,698 + 3,138 Log(LT ) 0,982 278 3,088 3,188
VII.27 Log(Peso (gr)) = -11,016 + 3,023 Log(LCC ) 0,988 279 2,983 3,063
VII.28 Log(Peso (gr)) = -7,653 + 3,038 Log(Lcab ) 0,975 272 2,979 3,097
VII.29 Log(Peso (gr)) = -3,981 + 2,524 Log(Lhoc ) 0,966 267 2,467 2,581

Andrés Eloy Seijas
278
CONTENIDO
Pag.
Capítulo 1
LOS CROCODYLIA…………………………………. 7
Características físicas…………………………………… 10
Comportamiento……………………………………….. 14
Reproducción y desarrollo ……………………………. 15
Diferencia entre sexos……………………………….... 16
Alimentación………………………………………...... 17
Longevidad…………………………………………..... 18
Clasificación de los Crocodylia……………………… 21
Capítulo 2
LOS CROCODYLIA DE VENEZUELA……………. 27
Sinopsis sobre las especies de Crocodylia de Venezuela…. 29
La Baba (Caiman crocodilus)…………………………… 29
Los Paleosuchus……………………………………………… 39
El Caimán de la Costa (Crocodylus acutus)……………. 49
El Caimán del Orinoco (Crocodylus intermedius)……… 60
Conservación de las especies de Crocodylia de
Venezuela ……………………………………………….. 66
Capítulo 3
CAPTURA, MANIPULACIÓN Y MARCAJE……… 71
Captura………………………………………………….. 71
Manipulación……………………………………………. 77
Marcaje………………………………………………….. 80
Capítulo 4
MORFOMETRÍA……………………………………. 87
Toma de datos…………………………………………... 89
Relaciones Morfométricas en babas……………………. 90
Relaciones Morfométricas en el género Paleosuchus…… 95
Relaciones Morfométricas en el género Crocodylus……. 98
Aplicabilidad de los Análisis Morfométricos…………... 102

Los Crocodylia de Venezuela
279
CONTENIDO
Pag.
Capítulo 5
ANÁLISIS DE POBLACIONES…………………….. 109
Consideraciones metodológicas…………………………. 109
Determinación de abundancia………………………….. 110
Unidades de medida de la densidad o abundancia……. 118
Determinación de la estructura poblacional…………… 120
Factores de corrección y ajustes……………………….. 124
Abundancia y estructura poblacional de las especies… 129
Capítulo 6
DIETA………………………………………………… 159
Métodos…………………………………………………… 160
Dieta de la misma especie en distintas localidades……… 166
Dieta de distintas especies en la misma localidad……….. 175
Capítulo 7
LOS CROCODYLIA COMO RECURSO……………. 185
Usos tradicionales……………………………………….. 185
Aprovechamiento comercial…………………………….. 187
Zoocriaderos comerciales………………………………... 199
Capítulo 8
CRÍA EN CAUTIVERIO…………………………..… 209
Colecta e incubación de huevos…………………………. 215
Crianza de Crocodilios con fines de repoblación……….. 224
Caimán del Orinoco………………………………….. 225
Caimán de la Costa…………………………………… 234
Baba………………………………………………….. 236
REFERENCIAS…………………………… 241
APÉNDICES………………………………. 271

NOTAS DEL AUTOR PARA LA EDICIÓN DIGITAL
Para esta edición digital agregué, como apéndices adicionales,
1) una Fe de Errata; 2) Aclaratoria de imprecisiones; 3) Cálculo de
límites de confianza para el estimado de una medida corporal a
partir de otra; 4) Un Resumen Curricular y 5) Agradecimientos.
Estos dos últimos puntos se presentan tal como fueron redactados
en el 2011.
Fe de erratas
Para esta edición digital se corrigieron algunos errores presentes
en la edición impresa, la mayor parte de ellos referentes a unos
pocos problemas de acentuación y a inconsistencias en el uso de las
comas (“,”) y puntos y comas (“;”) en el momento que se citan las
referencias en el texto. Se corrigieron algunas otras pocas
inconsistencias o errores, fácilmente reconocibles como tales por
cualquier lector, y que no provocaban ningún tipo de confusión
sobre lo que se quería decir, por lo que no vale la pena detallarlas
en esta nota. Ninguna de esas correcciones cambia el sentido
original de las oraciones en las que aparecen. Otras modificaciones
relevantes se señalan a continuación.
En la primera fila de la Tabla 2.3 (página 47) donde decía
“Escamas Occipitales (EO)” se escribió “Escamas Post-
Occipitales”, que es lo que en realidad corresponde.
En versión impresa se omitió por error de la Figura 2.11 (página
55), el dibujo de los patrones de escamación de Crocodylus acutus
que debió aparecer a la izquierda de las dos fotografías
incorporadas. Sin ese dibujo la leyenda de la figura pierde
completamente su sentido. En esta versión digital se agrega el
dibujo referido.
Dos tablas con distintos contenidos fueron identificadas con el
mismo número (Tabla 5.3) (páginas 127 y 132). Se mantuvo el
error, porque su corrección hubiera significado numerosos cambios
tanto en el texto como en la numeración de todas las tablas
sucesivas, lo que me pareció inconveniente.
Se completó la nota al pie de la Tabla 6.8 (página 178),
agregando al final del texto “con excepción de los peces”.

Por lo demás, el significado del texto de cada página, cada
párrafo y cada línea se corresponde de manera fiel con el texto
original impreso, aunque pueden existir pequeñas diferencias de
forma (tamaño de las letras, tamaño de las tablas y otras).
Imprecisiones
Vale la pena mencionar algunos errores u omisiones de otra
índole. En el segundo párrafo de la página 7 se dice que son siete
las especies de Crocodylia que existen en Sur América, cuando en
realidad son ocho, como queda claro en la Tabla 1.1 (página 25).
Por otra parte, en el último párrafo de la página 104 se afirma
que el cráneo más grande de Crocodylus intermedius del que se
tenga noticias (68 cm) se encuentra en el Museo de Rancho
Grande, en Maracay, cuando en realidad Antelo (2008) menciona a
un ejemplar con un cráneo de 74 cm (el Caimán de Pariaguán).
Ambas imprecisiones se mantienen en esta versión digital.
Cálculo límites de confianza
Los modelos de regresión que aparecen en los apéndices pueden
ser usados para estimar el tamaño de una parte del cuerpo (que se
desconoce) a partir de otra (conocida). La ecuación general de estos
modelos es:
𝒀 = 𝒂 + 𝒃 ∙ 𝒙
Donde Y es valor predicho por el modelo, a es el intercepto de la
recta en el eje de las abscisas, b es la pendiente de la línea de
regresión, y, por último, x es el tamaño de la parte del cuerpo que
se usa para predecir Y. Los valores de “Xm”, “Sxx” y “s” que
aparecen en los señalados apéndices I, V y VI sirven para calcular
los límites de confianza de Y. En la versión impresa se dice que sus
significados aparecían en el texto, lo cual no fue el caso, por lo que
se hace a continuación:
Xm = 𝒙, el valor promedio de la variable morfométrica (tomada
como variable independiente) utilizada para el análisis de
regresión.

𝑺𝒙𝒙 = ∑ (𝒙 − 𝒙)𝟐𝒏𝒊=𝟏 , es la suma de las diferencias al cuadrado
de cada observación respecto a la media general.
𝒔 = √𝟏
𝒏−𝟐∑ (𝒚𝒊 − 𝒀𝒊)𝟐𝒏
𝒊=𝟏
𝟐; es la desviación estándar de los
residuales del modelo de regresión, donde yi es el valor observado
y Yi es el valor predicho por el modelo. El valor de s puede ser
obtenido a partir de distintos análisis de algunos programas
estadísticos (Por ejemplo, s es el root mean square error, RMSE,
que aparece en el análisis de varianza con JMP In; o la raíz
cuadrada de Dispersion Phi en el GLM con Past, por citar dos
ejemplos).
Con todos esos valores es posible calcular los límites de
confianza de un valor estimado (Y), de acuerdo a la ecuación:
𝑳í𝒎𝒊𝒕𝒆𝒔 𝒅𝒆 𝒄𝒐𝒏𝒇𝒊𝒂𝒏𝒛𝒂 = 𝒀 ∓ 𝒕𝒔 {𝟏 +𝟏
𝒏+
(𝒙 − 𝒙)𝟐
𝑺𝒙𝒙}
𝟏𝟐
Donde, n es el número de datos usados en el modelo de
regresión y t es el valor tabulado de la distribución de Student para
el nivel de probabilidad deseado (con n-2 grados de libertad).
Para más detalles, consultar:
CAMPBELL, R. C.
1967. Statistics for biologists (3ra ed.). Cambridge University Press, New
York (Pag. 341-342).
HAMMER, Ø.
2017. Past. Paleontological Statistics Reference Manual. Version 3.15.
SALL, J., A. LEHMAN Y L CREIGHTON.
2001. JMP Start statistics. A guide to statistics and data analysis using JMP
and JMP In Software. (2da ed.).Duxbury, Pacific Grove, CA. (Pag.
138).
Un ejemplo servirá para ilustrar mejor el uso de las ecuaciones
de los apéndices I, V y VI.
El 11-MAY-1987 se capturó una baba (Caiman crocodilus
fuscus) en el embalse de Tacarigua, estado Falcón, con la cola
mutilada. Le faltaban la totalidad de las CCS y poseía solo 11 CCD

(por lo general poseen entre 13 y 14 CCD). La LT medida fue 1005
mm. ¿Cuál sería la LT de este animal con la cola completa? Se
estimará a partir de tres medidas disponibles: LCC=645 mm;
Lcab=169 mm, y Lhoc=101.7 mm. Utilizando los valores
reportados en el Apéndices I, se calcula la LT estimada (Y) y se
sustituyen los otros valores en la ecuación anterior: Los resultados
son:
LT(LCC) = 1228 mm ± 49.8 mm (Eq. I.1)
LT(Lcab) = 1257 mm ± 92.9 mm (Eq. I.4)
LT(Lhoc) = 1256 ± 69.2 mm (Eq. I.5)
El intervalo se calculó para el 95% de confianza. Los valores de
t-student´s varían en cada ecuación, dependiendo del tamaño de la
muestra.
El animal en cuestión había perdido entre 15.9 a 34.5 cm de su
cola.
Andrés Eloy Seijas Y.
4 sep. 2017

AGRADECIMIENTOS
Mi relación con los Crocodylia comienza cuando buscaba un
tema para mi tesis de Licenciatura en Biología en el año 1976.
Después de explorar varias posibilidades, que iban desde el estudio
del “periphyton” en cuerpos de agua en el Módulo Experimental de
Mantecal (MEM) hasta la de ahondar en la ecología de la tortuga
Kinosternon scorpioides en los llanos, me topé, casi por descarte,
con la propuesta que me hiciera el ya fallecido profesor Jesús
María Pacheco de trabajar bajo la dirección de Santiago Ramos,
recién ingresado profesor del Instituto de Zoología Tropical (IZT).
El objetivo era estudiar aspectos de la ecología y el metabolismo de
la baba (Caiman crocodilus) integrante conspicuo de la fauna de
los llanos inundables de Venezuela. La biología de esta especie era
apenas conocida en aquel momento y los primeros estudios en el
país, de Carlos Rivero-Blanco y Mark Staton, eran todavía muy
recientes.
Mi trabajo en el MEM contó con la entusiasta colaboración de
todo el equipo del IZT que coordinaba el profesor Pacheco. Quiero
agradecer en particular a mi tutor Santiago y a quienes se
constituyeron en mis ayudantes de campo: Tomás Pino, Gustavo
Villarroel, Gregory Colomine (††), Eduardo “El Pavo” Miranda
(††), Wilman “Picosa” Vásquez, Franklin Michelangelli, Napoleón
León, Eglis Méndez, José Rodríguez, Oswaldo Travieso y Gerardo
García.
Una vez graduado, en 1979, entré a trabajar en la Oficina de
Fauna del Ministerio del Ambiente y de los Recursos Naturales
Renovables-MARNR (ahora MINAMB). Mi jefe, Gonzalo Medina
Padilla, apoyó mis iniciativas de investigación sobre el caimán de
la costa (Crocodylus acutus) primero en la bahía de Turiamo,
apenas a una hora de recorrido desde nuestra oficina en Rancho
Grande, y luego al resto del país. Numerosos colegas y compañeros
de trabajo participaron en las expediciones que planificamos entre
1980 y 1984. Quiero darle un reconocimiento especial a Luis

Caballero, Pedro Liendo, Gerardo Cordero, Arnaldo Ferrer, Ramón
Rivero, Sergio Bermúdez, Alberto Wilinski, José Ochoa, Arelis
Chang, Edgar Useche, Frank Espinoza, Luis Machado, Francisco
Bisbal y José Daniel Pérez Bora. En esa época tuve el privilegio de
organizar una actividad de campo en la compañía de Tomás
Blohm, Peter Brazaitis y Myrna Watanabe.
Para el trabajo de campo de mi tesis de maestría (con el caimán
de la costa) me acompañaron en ocasiones algunos de los amigos
del MARNR ya mencionados y también Betty Baquero, F. Wayne
King (mi tutor), Dante Vides, John Thorbjarnarson (††) y, en
especial, Carlos Chávez, quien me ayudó también años más tarde
con el trabajo de campo con el caimán del Orinoco en el río
Cojedes.
Otras personas con las que he compartido actividades de campo
relacionadas con los crocodilios, o que me han suministrado
gentilmente datos, son Alfredo Arteaga, Alfredo Morales, Álvaro
Velasco, Bethina Vargas, Carlos Cáseres, Carlos Sánchez, Daría
Pirela, Esteban López, Ernesto Boede, Gustavo Hernández,
Gustavo Spinetti, Hedelvy Guada, Ildemaro González, Jesús Rivas,
José Ayarzagüena, María Muñoz, Marianne Navarro, Martín
Correa-Viana, Néstor Windelboxel, Omar Hernández, Oscar Brull,
Ramón Leal, Ricardo Babarro y Tibisay Escalona.
A mis estudiantes: Alfonso Llobet, Ariel Espinosa Blanco,
Fredy Garabito, Heide Heredia, Joaquín Salazar, José Adrianza,
José Manuel Mendoza, Juan Manrique, Keila Matos, Luis Sánchez,
Magddy Jiménez-Oraá, Magdiel Jiménez-Oraá, Manuel González-
Fernández, Pablo Meza, Pamela Ávila, Rogers Barreto, Tito
Barros, Yamilé Terán, Yubane Rosales, Yohiner Delfín. En ellos
ha recaído gran parte del trabajo pesado en los últimos años.
A las instituciones y personas que facilitaron mis estudios de
postgrado y han apoyado mi trabajo de campo: MARNR (Gonzalo
Medina Padilla, Douglas Figueroa, Enrique Dorta), World Wildlife
Fund (Curtis Freese, Byron Swift), Universidad de Florida (F.
Wayne King, John Robinson, Kent Redford, Richard Bodmer, J.
Perran Ross), Wildlife Conservation Society (Stuart Strahl, John
Thorbjarnarson), UNELLEZ, Fundación Fullbright, CONICIT,

FUDENA (Cecilia Blohm, Glenda Medina) y PROVITA, FUDECI
(Omar Hernández, Antonio Machado Allison).
A mi esposa, Lemnie Falkenhagen, y mis hijos: Andrés y Sara,
quienes con frecuencia me asistieron en la toma de datos y en el
trabajo sucio de mantenimiento del zoocriadero de la UNELLEZ.
Con seguridad hay más personas con las que estoy en deuda por
la ayuda que me han prestado, lamento no poder recordarlas a
todas. Mis sinceras disculpas a todas ellas.
La versión final de este libro mejoró sustancialmente con las
correcciones y sugerencias de Jorge Méndez, Lemnie Falkenhagen,
Andrés Seijas F., Sara Seijas F., Aldeima Pérez, Roldán De Sola, y
los miembros del jurado que lo evaluó como trabajo de ascenso a la
categoría de profesor titular de la UNELLEZ (Rony Tejos, Alexis
Arends y, muy especialmente, Martín Correa-Viana). Como ya se
acostumbra a decir en estos casos, los errores y limitaciones que
permanecen son de mi entera responsabilidad.
Andrés E. Seijas
Guanare, Junio 2011.

RESUMEN BIOGRÁFICO DEL AUTOR
Andrés E. Seijas Yanes. Biólogo (Escuela de Biología, UCV-
1979) con maestría (1988) y Doctorado (1998) de la Universidad
de Florida (Gainesville-EE.UU). Trabajó (1979-1989) en el
Ministerio del Ambiente (MARNR) en investigaciones
relacionadas con inventarios de fauna, anillado de aves y
evaluaciones poblacionales del caimán de la costa (Crocodylus
acutus) y la baba (Caiman crocodilus). Ingresó a la Universidad de
los Llanos (UNELLEZ) en 1989, institución donde impartió
docencia en Zoología, Manejo de Fauna y Biología de la
Conservación. Encargado del zoocriadero de Crocodylus
intermedius de esa institución por 20 años. Pertenece al Comité de
Supervivencia de las Especies (SSC-UICN) desde 1984. Fue
presidente del Grupo de Especialistas en Cocodrilos de Venezuela
(GECV). Es autor o coautor de más de 50 publicaciones.






























