Embed Size (px)
Citation preview

1
UNIVERSIDAD RICARDO PALMA
FACULTAD DE CIENCIAS BIOLOGICAS
DESARROLLO DE Verticillium chlamydosporium (Goddard ,1913) Y
EVALUACIÓN DE PATOGENICIDAD IN VITRO PARA EL CONTROL DEL
NEMATODO DEL NUDO DE LA RAIZ Melodoigyne incognita (Chitwood,
1949)
Tesis para optar el Titulo Profesional de
Licenciado en Biología
Claudia Sofía Picho Gómez
Bachiller en Biología
Lima - Perú
2006

2
Este trabajo se lo dedico a DIOS, por todas las cosas maravillosas que
me ha dado y por toda su energía que siempre me manda.
A mis padres Hilda y Heriberto, a Diego y Mónica, por su paciencia,
cariño, buenos consejos; por confiar mucho en mi y en todo lo que
puedo mostrar.
Por supuesto a mis amigos….y al Novoa, por todo...

3
MIS AGRADECIMIENTOS
Nuevamente a Dios, mi familia, mis seres queridos, mis amigos...
Al Doctor Tomas Agurto Sáenz
Por su amistad, tiempo y paciencia, en la orientación para la realización de
este trabajo.
A la Dra. Vera Allegan, el Dr. Alcides Guerra y la profesora Mercedes
Gonzáles, gracias por su tiempo empleado.
A la Mblga. Anny Zapata (Asesora externa), del Área de Producción de
Hongos entomopatogenos del Programa Nacional de Sanidad Agraria
SENASA, por la amistad, y orientación para la realización de este trabajo
Al Ing. Cosme Quispe, (Asesor externo) del Área de Nematologia del
Programa Nacional de Sanidad Agraria SENASA por su colaboración
desinteresada y por todas sus observaciones.
Al Biólogo Enrique Torres por toda la ayuda brindada en todas las etapas de
la Tesis y por todos los ánimos que echaba para la culminación de esta
También a todas aquellas personas que han colaborado indirectamente en
la realización de este trabajo.
GRACIAS

4
Resumen
El nemátodo del nudo de la raíz Meloidogyne incognita (Chitwood, 1949), es
un nemátodo que ataca a diferentes cultivos, tanto en campo como en
invernadero, causando grandes pérdidas económicas. Como alternativa de
control de este nemátodo, se logró estandarizar la producción masiva de
Verticillium chlamydosporium (Goddard, 1913) un hongo nematófago y se
evaluó la patogenicidad in vitro del bionematicida obtenido.
Para la conservación de la cepa de este hongo se probaron diferentes
medios de cultivo y para su desarrollo masivo, diferentes medios líquidos y
sustratos sólidos, también se determinaron las condiciones apropiadas para
el óptimo desarrollo del hongo.
El mejor medio sólido para la conservación de la cepa fue Médio 90, con lo
cual se obtuvo 4.31 x 108 con/ cm
2 a los 8 días de sembrado y a los 30 días
se obtuvieron clamidosporas, que fueron conservadas a 5 ºC.
Para el desarrollo masivo, el medio liquido Papa Dextrosa con un promedio
de 16.73 UFC/ 300 µl y el sustrato arroz fueron los mejores, obteniéndose a
los 30 días de sembrado el hongo un promedio de 4 x 108 con/g.
Las condiciones apropiadas para el desarrollo del hongo Verticillium
chlamydosporium son temperatura 24°C y humedad dentro de bolsa 30%
La patogenicidad de Verticillium chalmydosporium, observada en ootecas fue
de 32.25% y en huevos fue de 63.76%, esto demostró que el bionematicida
obtenido del desarrollo de Verticillium chlamydoporium fue un buen agente
controlador del nematodo del nudo Meloidogyne incognita

5
Abstract
The knoot root nematode Meloidogye incognita, (Chitwood, 1949), is one of
the main pest on differents crops, so much in field as in greenhouse, causing
big economic lost at the growers. As an alternative of control of this nematode,
to succeed in standarize the mass production of Verticillium chlamydosporium
(Goddard, 1913) a nematophagous fungi and evaluated its pathogenicity in
vitro of the bionematicide obtained.
In order to conservation the strain of this fungi, was tested differents culture
media and for its mass production differents liquid media and solid substrates,
also was determinated the appropriate conditions for the optimal fungus
development.
The best solid medium for the stored of the strain was Medium 90, obtained
4.31 x 108 con/cm
2 at the 8 days, at the 30 days was obtained
chlamydosporas, that was stored at 5 ºC
To the mass production, the liquid media Potato Dextrose with an average of
16.73 UFC/ 300 µl and the rice substrate were the best, obtaining at the 30
days of cultivated the fungi an average of 4 x 108 con/g.
The appropiated conditions for the development of Verticillium
chlamydosporium are, temperature 24ºC and a humity inside the bag of 30%
The pathogenicity observed in ootecas was 32.25% and in eggs was 63.76%,
this demonstrated that the bionematicide obtained of the development of fungi
Verticillium chalmydosporium was a good control agent of knoot root
nematode Meloidogye incognita

6
INDICE
Resumen 4
Summary 5
I. INTRODUCCIÓN 9
II. ANTECEDENTES 11
2.1 Antecedentes Nacionales 11
2.2 Antecedentes Internacional 11
2.3 Control Biológico de Meloidogyne incognita 13
2.4 Generalidades respecto al antagonista 14
2.4.1 Características morfológicas 15
2.4.2 Mecanismo de acción 15
2.5 Generalidades respecto al hospedante M. incognita 17
2.5.1 Características Morfológicas de M. incognita 18
2.5.2 Duración del ciclo de vida síntoma y daños 19
2.6 Métodos de Control de M. incognita 21
2.6.1 Control cultural 22
2.6.2 Control físico 23
2.6.3 Control Químico 23
2.7 Generalidades para la producción de H. entomopatógenos 23
III. MATERIALES Y MÉTODOS 27
3.1 Materiales 27
3.1.1 Material Biológico 27
3.2 Metodología 27
3.2.1 Obtención de material biológico de M. incognita 27
3.2.1.1 Identificación de M. incognita 28
3.2.2.1 Reactivación de cepa 28
3.2.2.2 Aislamiento del hongo 29
3.2.2.3 Cultivo monoesporico 29
3.2.2.4 Re-aislamiento del hongo 29
3.2.3 Producción del hongo V. chlamydosporium 30

7
3.2.3.1 Fase I 30
3.2.3.1.1 Preparación de inoculo liquido 31
3.2.3.1.2 Control de calidad del inoculo 32
3.2.3.2 Fase II 32
3.2.3.2.1 Preparación de sustrato sólido 32
3.2.3.2.2 Determinación de humedad y Temperatura 33
3.2.3.2.2.1 Humedad 33
Preparación de bolsas 33
Incubación 34
Secado 34
Control de calidad 34
3.2.3.2.2.2 Temperatura 34
Preparación de bolsas 35
Incubación 35
Secado 35
Control de calidad 35
3.2.3.2.3 Control de calidad 36
3.2.3.2.3.1 Concentración de conidias 36
3.2.3.2.3.2 Porcentaje de germinación o viabilidad 37
3.2.3.2.4 Prueba de Patogenicidad 38
3.2.3.2.4.1 Tratamiento de Ootecas 38
3.2.3.2.4.2 Tratamiento en huevos 38
3.2.3.2.4.3 Evaluación 39
IV. RESULTADOS 40
4.1 Desarrollo del hongo en medio de cultivo liquido para conservación de cepas 40
4.2 Desarrollo de medio líquido 40
4.2 Desarrollo de medio sólido 40
4.3 Porcentaje de germinación 41
4.4 Porcentaje de humedad 41
4.5 Porcentaje de parasitación en ootecas 42
4.6 Porcentaje de parasitación en huevos 42

8
V. DISCUSIÓN 44
VI. CONCLUSIONES 46
VII. REFERENCIAS BIBLIOGRAFICAS 47
VIII. ANEXOS 53

9
INTRODUCCIÓN
Los nemátodos del género Meloidogyne, son considerados como los
nemátodos fitoparásitos, más importantes debido a su amplio rango de
hospederos, en mayor parte cultivos alimenticios e industriales que son
susceptibles a esta plaga.
El promedio de las pérdidas a nivel mundial en hortalizas causado por
el género Meloidogine y otros nematodos se estima en un 12.3 %, en países
desarrollados, pudiendo llegar a 14.6% en países subdesarrollados, sin
embargo hay autores que han estimado pérdidas de hasta un 25%. Además
de un daño directo, Meloidogyne y otros nemátodos fitófagos, predisponen a
los cultivos a enfermedades fungosas y bacterianas causando pérdidas aun
mayores, no sólo en el rendimiento, sinó también en la calidad de la cosecha.
Actualmente, para el control de éste nemátodo se utilizan productos
químicos que son altamente tóxicos, estos son rápidos y eficientes, y son una
solución temporal al problema causado por el nematodo, pero la utilización
de estos de una manera inadecuada, pueden favorecer al desarrollo de
resistencia del nemátodo al producto, y ocasionar desequilibrios en el
ecosistema
La utilización de medidas de control biológico es una alternativa para
reducir sus poblaciones, a un bajo nivel de daño económico, sin riesgos de
contaminación del ambiente. De estas hay que destacar el control microbiano,
por la utilización de microorganismos entomopatógenos. El uso del control
microbiano minimiza el problema de la resistencia de plagas, y el exceso de
residuos químicos en los alimentos.
Entre las alternativas de control microbiano, se encuentra el uso del
hongo nematófago Verticillium chlamydosporium, (Goddard ,1913)
considerado como uno de los agentes de control biológico más promisorios

10
para el manejo de poblaciones de nemátodos formadores de agallas (Kerry y
Jaffe, 1997), en particular de huevos de Meloidogyne incognita (Atkins et. al.
2003, Kerry e Hidalgo, 2004).
En el Perú, la utilización de estos bioplaguicidas es incipiente,
desarrollando hace pocos años la producción masiva de hongos benéficos,
los cuales se realizan en forma semi - industrial, para lo cual es necesario
realizar trabajos de investigación que estén dirigidos al aislamiento,
selección de cepas nativas virulentas, medios adecuados para su producción
con la finalidad de obtener formulaciones comerciales patogénicos, capaces
de establecerse en el medio ambiente y sobrevivir a condiciones ambientales
adversas (Hidalgo, et al. 2004; Ciancio y Leonetti, 1999).
El objetivo de este trabajo fue de evaluar diferentes medios de cultivos
sólidos, líquidos y sustratos, y poder determinar las condiciones apropiadas
para el desarrollo de Verticillium chlamydosporium (Goddard ,1913) y
observar su capacidad patogénica en Meloidogyne incognita (Chitwood, 1949)
in vitro

11
ANTECEDENTES
En investigaciones efectuadas para el control biológico de nemátodos,
se ha demostrado que Verticillium chlamydosporium (Goddard
1913), es efectivo contra Meloidogyne arenaria (Neal), Meloidogyne
incógnita (Kofoid), Meloidogyne javanica, y Meloidogyne hapla.
(Chitwood). Así como también afecta a nemátodos del género
Heterodera y Globodera (De Leij et al. 1991). Este hongo nematófago
es un habitante del suelo, siendo un controlador natural de nemátodos.
En un estudio cuyo objetivo fue determinar que agente natural era
mejor como controlador de Melodoigyne incognita bajo condiciones de
laboratorio, se determinó que el hongo Paecilomyces. lilacinus era
mucho mas eficiente como controlador de Meloidogyne incognita, que
otros hongos como Verticillium lecanii y Verticillium chlamydisporium
(Limay ,1998)
López-Llorca, et al. (2002), realizaron ensayos con tres hongos
nematófagos, Pochonia rubens, P. chlamydosporia (V.
chlamydosporium) y Lecanicillium lecanii, observando infección en
huevos de Meloidogyne incógnita con porcentajes de 35%, 42% y
50% respectivamente.
Para la conservación y producción de hongos entomopatógenos se
desarrollaron diferentes metodologías, siendo una de ellas, las que ha
desarrollado el Laboratorio de Entomopatógenos del SENASA
(Gómez y Zapata 1998).
Camargo et al. (2002), realizaron un ensayo en el cual probaron cinco
medios de cultivo para el desarrollo de Verticillium lecanii encontrando
que el mejor medio para su desarrollo contiene peptona, extracto de
levadura y caseína hidrolizada.

12
Olivares-Bernabeu y Lopez-Llorca (2002), estudiaron factores como
temperatura, pH y humedad relacionados con el desarrollo y actividad
de diferentes hongos nematófagos, incluido Verticillium
chlamydosporium como agentes de control biológico de nemátodos
fitófagos en suelos españoles ( Melodoigne y Heterodera )
Atkins, et al. (2003), en un trabajo realizado determinaron el
parasitismo de Verticillium chlamidosporium, bajo condiciones
semi controladas en invernadero, observando un parasitismo de 68%
en huevos y 70% en ootecas del nematodo del nudo Meloidogne
incognita
Bharadaj y Trivedi (2004), evaluaron diferentes sustratos para la
multiplicación masiva de Verticillium chlamydosporuin y probaron
su eficacia sobre Heterodera cajani, ensayando diferentes
dosificaciones del hongo para el control de este nematodo, bajo
condiciones de invernadero.
Kerry e Hidalgo (2004), observaron 68 % de parasitismo en huevos de
Meloidogyne incógnita, utilizando el hongo Verticillium
chlamydosporium.
Soberanis et al. (2005), realizaron ensayos de multiplicación masiva
de hongos entomopatógenos para la obtención de producto seco,
utilizando temperaturas de 17 ºC ± 2 ºC para la etapa de secado,
elevando la concentración de conidias de 109 a 10
11 conidias por kilo,
para los hongos Beauveria.bassiana, Metharhizium anisopliae,
Trichoderma harzianum y Paecilomyces. fumosoroseus

13
2.3 Control biológico del género Meloidogyne
Los nemátodos del nudo de la raíz están ampliamente distribuidos y son
económicamente importantes, por lo tanto la mayor parte de estudios de
control biológico están enfocados a este grupo de nemátodos, sin
embargo su habito endoparásito es el principal impedimento para el
desarrollo de un control biológico exitoso (Stirling, 1991). La mayor parte
de su ciclo de vida, los nemátodos están completamente rodeados por
tejido radicular y están por lo tanto protegidos de los parásitos y
predadores del suelo (De Leij, et al 1992; Stirling, 1991).
Hasta hace poco el estado de desarrollo más utilizado para estudios de
control biológico de nemátodos endoparásitos sedentarios, ha sido el
segundo estadio juvenil (J2) (Stirling, 1991).
Sin embargo los J2 han sido un objetivo muy evasivo debido a que no se
presenta en forma constante en el suelo, el número de juveniles de
Meloidogyne varia de acuerdo a las condiciones ambientales de este, la
mayoría de J2 son encontrados luego de una lluvia o riego, cuando la
humedad del suelo es favorable para la eclosión (Davies et al. 1991;
Taylor and Sasser, 1988; Stirling, 1991).
Otra característica que vuelve a J2 en una difícil presa para los
antagonistas es su capacidad de escapar del parasitismo y predacion
invadiendo las raíces (Stirling, 1991), este proceso puede ocurrir
rápidamente y los J2 de Meloidogyne inoculados experimentalmente en
macetas o en agar pueden ser encontrados en las raíces después de 24
horas, además en 3 días los J2 pueden moverse hasta 50 cm para
establecerse en las raíces (Prot et al. 1979).
Debido a que las hembras de Meloidogyne están protegidas por las
raíces, y los juveniles son móviles, los huevos parecen ser el estado de
desarrollo más vulnerable para el ataque de los antagonistas (Stirling,
1991). Los huevos generalmente están localizados sobre la superficie de

14
las raíces y bajo condiciones ideales, en 10 días pueden completar su
desarrollo y pueden eclosionar, además ya que los huevos están
agrupados en masas, un parasito o predador efectivo localizado cerca,
podría ser capaz de eliminar muchos de los huevos (Stirling, 1991).
Sin embargo hay situaciones, donde los huevos, son puestos dentro del
tejido nodulado y los antagonistas del suelo son incapaces de ejercer un
control efectivo, por tener un acceso limitado a los huevos (Kerry et al.
1984; Stirling, 1991). Factores como la planta hospedante, la temperatura
y la densidad del inóculo inicial afectan la proporción de huevos puestos
dentro o fuera de los nódulos y por lo tanto influenciaran en la eficacia de
la infección de los huevos (De Leij y et al. 1992; Stirling, 1991).
El control biológico de nemátodos formadores de agallas abarca una gran
diversidad de organismos que viven en el suelo, como enemigos
naturales de los nemátodos que atacan a las plantas. Estos parásitos
incluyen hongos, bacterias, virus, protozoarios otros nemátodos e
invertebrados (Whitehead, 1997; Kerry, 2001).
Kerry (1995), presenta una relación de agentes microbianos para el
control biológico de nemátodos parásitos de plantas (ANEXOS, Tabla Nº
01)
2.4 Generalidades respecto al antagonista
Verticillium chlamydosporium (Goddard ,1913)
La identificación de esta especie se realizó en 1913, cuando Goddard
aisló de suelo de jardín a un hongo que identificó como Verticillium
chlamydosporium.

15
Según Agrios (1995), Verticillium chlamydosporium, es un hongo
imperfecto que pertenece:
Género: Verticillium
Especie: Verticillium chlamydosporium Goddard ,1913
2.4.1 Características morfológicas
Verticillium chlamydosporium (Goddard, 1913), presenta
conidióforos erectos, con ramificación verticilada, presentando una
protuberancia en forma de mazo en la punta del conidióforo que
sostiene una sola espora; fiálides cilíndricas de 15 – 30 m de longitud
en forma de espiral. Fialosporas hialinas, de 2 - 5 x 1,5 - 2 m de
diámetro, ovoides ligeramente cilíndricas, de paredes lisas.
Chlamydosporas muy comunes, que presentan de 4 a 9 celdas
globulares de paredes gruesas, miden de 15 –30 x 10 a 20 m,
muriformes, ligeramente lobulados, con contenido granular, naciendo
terminalmente sobre ramificaciones laterales cortas de 15 – 30 m de
longitud, son muy persistentes. El micelio maduro completamente
cubiertos de chlamydosporas, llegando a presentarse polvoso de color
crema u ocre (Cook and Bakes. 1989).
2.4.2 Mecanismo de acción
Verticillium chlamydosporium (Goddard, 1913), forma una red
miceliar muy ramificada la cual permanece en estrecho contacto con la
cutícula del huevo, luego el hongo produce órganos de penetración
especializados denominados apresorios, desarrollados a partir de la
hifa indiferenciada que permite la colonización de la superficie de los
huevos de los nemátodos. Sin embargo, la penetración, es el resultado
de una presión física y una actividad enzimático (Lopez-Llorca, 2002).

16
El hongo causa la desintegración de la capa vitelina de la pared del
huevo por una enzima proteasa serina alcalina (subtilasa), designada
como VCP1 y una dilución parcial de la capa quitinolitica y lipídica del
huevo (Lopez-Llorca, 1986 y 2002; Segers et al. 1996; Tikhonov, 2002).
Además de los diferentes efectos de parasitismo de Verticillium
chlamydosporium (Goddard ,1913), en el desarrollo embrionario, los
efectos enzimáticos en la pared del huevo incrementan su
permeabilidad y posiblemente facilita la entrada de toxinas que pudieran
estar presentes en el medio ambiente (Lopez- Llorca, 1986; Morgan,
1988).
Diferentes investigadores han sugerido que Verticillium
chlamydosporium (Goddard, 1913), podría producir toxinas a
nemátodos. Meyer (1990); Morgan –Jones, (1988), observaron que los
huevos no eclosionaron cuando el hongo estuvo cerca de ellos. Sin
embargo, Irving y Ferry (1986); Davies (1991), no obtuvieron ninguna
evidencia que sugiriera la producción de toxinas.
Debido a que Verticillium chlamydosporium (Goddard ,1913), parece
ser una especie que ha desarrollado cierta especialización en relación
con los nemátodos del nódulo de la raíz y los nemátodos quiste, en
1980, se comienza en Rothamsted, Inglaterra un programa de
investigación para desarrollar métodos de introducción del hongo en el
suelo con propósitos de control biológico (Kerry, 1984). El hongo puede
ser introducido en arena, cáscara de trigo, grano de trigo en polvo o
alícuotas de suspensión de clamidosporas (Davies, 1991; Bharadaj,
2004). Cuando las clamidosporas fueron utilizadas como inóculo, el
hongo fue capaz de establecerse en el suelo sin ninguna base
alimenticia, debido a que las esporas tienen suficientes reservas
alimenticias y no necesitaron de fuentes adicionales de nutriente.
(Crump, 1992). Las clamydosporas son el estado de sobrevivencia del
hongo de modo que es probable que pueda iniciar la infección (Crump,
1992)

17
2.5 Generalidades respecto al hospedante Meloidogyne incognita
Los nemátodos del nudo de la raíz de Meloidogyne incognita tienen una
amplia distribución a nivel mundial y son económicamente importantes
patógenos de plantas, son parásitos obligados que atacan a miles de
diferentes especies de plantas incluyendo monocotiledóneas,
dicotiledóneas, plantas herbáceas y leñosas, además algunas especies
de Meloidogyne incognita son patógenos importantes de cultivos
alimenticios, hortalizas, frutales y ornamentales. Taylor and Sasser
(1983), mencionan que el problema con nemátodos se encuentra en
todas las áreas del mundo donde los cultivos son de periodos largos, y
los mayores daños ocurren en áreas de climas tropicales y semi
tropicales y una de las especies que mas daño ocasiona a los cultivos
es la especie Meloidogyne incognita.
La sistemática para Meloidogyne incognita según Rafiq Siddiki (2000),
es la siguiente:
Género: Meloidogyne
Especie: Meloidogyne incognita (Kofoid & White 1919),
Chitwood, 1949
2.5.1 Caracteres morfológicos de Meloidogyne incognita
Ciclo biológico
Pre-parasítico:
El desarrollo del huevo comienza después de la ovoposición, resultando
una larva completamente formada, con un estilete, enrollada en la

18
membrana del huevo. Este es el primer estadio larval (Taylor and
Sasser, 1983).
La primera muda tiene lugar en el huevo, poco después, la larva
emerge a través de un agujero hecho en un extremo del cascaron
flexible del huevo, por medio de pinchazos repetidos con el estilete.
El J2 de Meloidogyne incognita es móvil, vermiforme e infectivo y se
desplaza en el suelo para penetrar en las raíces de una planta
hospedante (Eisenback et al. 1991; Hussey, 1985).
Parasítico:
Penetración en las raíces de hortalizas
Las larvas en el segundo estadio larval infectivo generalmente penetran
en la raíz justamente sobre la caliptra (punta de la raíz), se mueven
principalmente entre las células no diferenciadas de la raíz (Taylor and
Sasser, 1983).
Con sus estiletes perforan las paredes de las células e inyectan
secreciones de sus glándulas esofágicas. Esto da lugar a la formación
de células gigantes (también llamadas sincitos), formadas por una
agrandamiento de las células (hipertrofia), a la posible disolución de
paredes celulares, a un agrandamiento del núcleo y a cambios en la
composición de los contenidos celulares. Al mismo tiempo, hay una
intensa multiplicación de células vegetales (hiperplasia), alrededor de la
cabeza de la larva (Taylor and Sasser, 1983).
Mientras se están formando las células gigante y las agallas, aumente
el ancho de la larva, y hay una dilatación de las glándulas esofágicas.
Las glándulas del primordio genital se dividen y este se agranda

19
haciéndose notorias dos ramificaciones en la hembra, o formando un
cuerpo alargado en el macho.
Morfología:
La longitud promedio de las hembras adultas de las especies de
Meloidogyne fluctúa alrededor de 0.44 a 1.3 mm y el ancho promedio
esta entre 0.325 y 0.7 mm, las hembras de la mayoría de las especies
tienen cuerpo simétricos, es decir hay una línea que va desde la vulva
del estilete, atravesando la mitad del cuerpo (Taylor and Sasser, 1983).
2.5.2 Duración del ciclo de vida, Síntomas y daños
Depende de la temperatura, a una temperatura de 29º C las primeras
hembras adultas de Meloidogyne incognita (Chitwood, 1949), aparecen
a los 13 – 15 días después de la penetración en las raíces, las hembras
empiezan a ovipositar a los 19 – 21 días, después de la penetración, el
promedio de vida de una hembra es de 2 – 3 meses (Taylor and Sasser,
1983).
Los J2 penetran en las raíces detrás de la cofia, la penetración la
realizan por medio de acción mecánica del estilete como enzimático
(celulolitica o pectinolitica), a través de ciertas secreciones de la
glándula esofageal después de la penetración el J2 migra
intercelularmente desde la corteza a la región de diferenciación celular
donde se establece y empieza a alimentarse, la cabeza del J2 se
localiza en la periferia del tejido del parénquima vascular y el resto del
cuerpo en la corteza, paralelo al axis longitudinal de la raíz (Huang,
1985; Hussey 1985; Taylor and Sasser, 1983).
Los sitios preferidos de alimentación son principalmente el floema o las
células parénquimaticas indiferenciadas del periciclo adyacente.

20
En respuesta a la alimentación del J2 el tejido de la planta hospedante
soporta grandes cambios morfológicos y fisiológicos, algunas células
del parénquima se desarrollan en células de alimentación permanentes
para el nematodo. Estas células se transforman en unas células
hipertrofiadas y multinucleadas, células gigantes, posiblemente como
resultado de secreciones producidas por las células de la glándula
esofageal dorsal del J2 (Hussey 1985)
Los nemátodos consiguen su alimento a través de estas células
especializadas y no pueden continuar su desarrollo sin ellas (Eisenback
et al, 1991; Taylor and Sasser, 1983). Las células gigantes son
esenciales para una exitosa relación hospedante-parásito, su activo
metabolismo es mantenido a través de las secreciones de la glándula
esofageal dorsal y por la remoción de solutos por parte del nematodo
(Eisenback et al, 1991). Las células gigantes son esencialmente células
transportadoras de alimentos hacia el nematodo (Huang, 1985).
Los fotosintatos son movilizados hacia las células gigantes en las raíces
y como resultado el crecimiento como la producción de la planta puede
verse afectada (Eisenback y et al, 1991; Huang, 1985; Hussey, 1985).
Otros síntomas aéreos de las plantas infectadas incluyen clorosis en el
follaje y marchites temporal durante periodos de estrés hídrico, la
absorción de agua y nutrientes es reducida grandemente por los daños
y la nodulación del sistema radicular (Eisenback y et al, 1991; Huang,
1985, Taylor and Sasser, 1983).
El tejido radicular alrededor del nemátodo y de las células gigantes
sufre una hiperplágia e hipertrófia dando lugar al característico nódulo
radicular (Eisenback et al. 1991; Huang, 1985, Taylor and Sasser, 1983)
Los nódulos generalmente se desarrollan a los 2 días después de la
penetración del J2 (Eisenback et al. 1991), el tamaño del nódulo esta
generalmente relacionado al número de nemátodos presentes en el

21
tejido, pero también de la especie de planta parasitada (Dropkin, 1954;
Eisenback et al. 1991).
2.6 Métodos de control de Meloidogyne incognita.
Las decisiones para la implementación de medidas de control están
determinadas por la densidad mínima dañina de nemátodos (población
mínima del nemátodo capaz de causar daño económico).
La densidad mínima dañina varia dependiendo de la especie de
nemátodo, que varia en su patogenicidad y agresividad. En tomate
Meloidogyne incognita requiere de 25 juveniles/100 gramos de suelo para
causar daño económico, Rotylenchus requiere mas de 200 juveniles
/100gramos de suelo para causar daño económico (Quispe ,1995).
En conclusión la ¨ densidad mínima dañina ¨ es variable para cada
relación hospedero-parásito (Quispe, 1995).
2.6.1 Control cultural
Escape:
Consiste en sembrar en épocas en que las densidades poblacionales
son bajas o que no favorecen el desarrollo del nemátodo (Quispe, 1995).
Abonamiento:
El uso de abono orgánico es efectivo para reducir las poblaciones de
nemátodos, al descomponerse origina acido, gases, que tienen una
acción nematicida. La temperatura se eleva por la fermentación en
algunas ocasiones llega a 70ºC la desventaja es que se requiere
grandes volúmenes 20 – 30 toneladas/ha (Quispe, 1995).

22
Solarización:
Es un método de pasteurización del suelo que permite suprimir la
mayoría de las especies de nemátodos patógenos eficazmente. Sin
embargo solo es consistente en lugares con veranos calidos y calurosos.
La técnica consiste en poner una o dos laminas de plástico transparente
encima del suelo ligeramente humedecido durante el verano
aproximadamente de 6 a 8 semanas.
Plantas trampa
Plantas susceptibles que se siembran con la finalidad de que el
nemátodo penetre y estas se arrancan y se queman (Talavera, 2003).
Rotación de cultivos
Consiste en alternar hospederos eficientes con plantas no hospederas
esto reduce grandemente la población. Sin embargo son pocas las
plantas no hospederas y su rentabilidad baja en comparación con los
que generalmente se cultivan (Talavera, 2003; Quispe, 1995).
2.6.2 Control físico:
Temperatura:
Someter el suelo a temperaturas altas que sea letal para los nemátodos
y que no sean dañinos para el material vegetal. El suelo para almacigar
puede ser tratado a una temperara de 121 ºC 15 lb de presión atm
durante 20 horas (Quispe, 1995).

23
2.6.3 Control químico:
Desde que se eliminaron del mercado el bromuro de metilo, el DBCP y
otros fumigantes, la agricultura se ha encontrado desprotegida del
ataque de nemátodos pero en cambio se han desarrollado nematicidas
no fumigantes, que se aplican al suelo en forma granulada dentro de
estos tenemos ETHOPROP, CARBOFURAM, ALDCARBO, OXAMIL,
FEMIPHOS (Quispe, 1995).
2.7 Generalidades para la producción de hongos entomopatógenos
La producción de hongos entomopatógenos involucra la elección de
medios líquidos y sólidos apropiados, los cuales proporcionaran los
nutrientes necesarios para el buen desarrollo el hongo con la finalidad
de obtener un biopreparado que proporcionen al usuario un producto de
máxima eficacia en condiciones de campo.
Existen diversas tecnologías de producción de un bioplaguicida a partir
de hongos, las más usadas son a partir de soportes sólidos, en cultivos
líquidos sumergidos o combinaciones de ambos. Estos métodos se
consideran generalmente artesanales o semiartesanales atendiendo al
número de manipulaciones que involucran, cada método tienen ventajas
y desventajas, lo más importante es la calidad del producto final y su
factibilidad tecnológica y económica (Fernandéz-Larrea, 2001).
El proceso de producción semi-industrial se realiza en varias fases, que
van desde la obtención del cultivo puro hasta la formulación del
producto, en general el proceso está organizado en dos etapas, la etapa
de cepario y la etapa de producción:

24
Cepario
Aislamiento del hongo
Elaboración de cultivos puros
Producción
Preparación de matraces y bolsas
Inoculación e incubación de matraces
Inoculación e incubación en bolsas
Proceso de secado
Cosecha del hongo
Elaboración de formulaciones
Un requisito indispensable es contar con un buen aislamiento que
mantenga su virulencia y homogeneidad debiendo ser
cuidadosamente conservadas, libres de contaminantes en un medio
de cultivo adecuado (Gómez y Zapata, 1998; Soberanis et al. 2005).
El medio líquido debe estimular un rápido desarrollo micelial el cual
será usado para inocular el substrato sólido (Jenkins et al. 1998).
Debe contener fuentes de carbohidratos que le suministran energía y
nitrógeno en forma de proteínas o aminoácidos, los cuales son
esenciales para el desarrollo del hongo.
Existen diversos sustratos sólidos disponibles para usarlos en la
producción de hongos para el control biológico, los que provee un
soporte físico para que el hongo produzca conidias aéreas.
Generalmente el sustrato es un cereal como arroz, mjjo, maíz, trigo,
cebada, avena, sorgo. Los hongos utilizan ciertas proporciones de los
nutrientes proporcionados por los sustratos durante su desarrollo y
esporulación. La elección del sustrato depende de un número de
factores que incluyen la disponibilidad local, costo y preferencia del

25
aislado (Jenkins, et. al. 1998). Un sustrato ideal no solo deberá
contener partículas con las dimensiones correctas, sino también
mantener su integridad estructural durante la preparación de los
procesos de producción (Maheva et al. 1984; Bradley et al. 1992).
A nivel mundial, existen numerosos países en los cuales grupos de
investigadores y empresas productoras se concentran en el
desarrollo de productos comerciales a partir de hongos entre los que
se encuentran, Cuba, Brasil, Colombia, Australia, Italia, Inglaterra,
Holanda, Estados Unidos (Burges, 1998; Butt and Copping, 2000).

26
III. MATERIALES Y METODOS
El presente trabajo de investigación se realizó en el Laboratorio de
Entomopatógenos de la Subdirección de Control Biológico (SCB) y en el
Laboratorio de Nematología del Centro de Diagnostico de Sanidad Vegetal
(CDSV), dependencias del Servicio Nacional de Sanidad Agraria – SENASA,
con sede en Vitarte y La Molina – Lima, durante los meses de enero del 2005
a julio del 2006.
3.1 MATERIALES 3.1.1 MATERIAL BIOLÓGICO
Cepa de Verticillium chlamydosporium (Goddard, 1913), obtenida
de la micoteca del Laboratorio de Entomopatógenos de la
Subdirección de Control Biológico (SCB) y de la Universidad Nacional
Agraria de La Molina, sin código, año de conservación 2002. Este
hongo nematófago ha sido introducido por el Dr. M. Canto, en el año
de 1996.
Huevos inmaduros y hembras de Meloidogyne incognita (Chitwood,
1949), a partir de raíces de plantas de tomate, obtenidas de la
empresa ICATOM S.A.
3.2 METODOLOGIA 3.2.1 Obtención del material biológico Meloidogyne incognita (Chitwood,
1949)
Se colectaron raíces de tomate de los campos de la empresa ICATOM
S.A. Ica, para lo cual se examinaron las raíces que presentaba

27
nodulaciones. Las raíces fueron colocadas individualmente en bolsas de
papel hasta su acondicionamiento en laboratorio
3.2.1.1 Identificación de Meloidogyne incognita (Chitwood, 1949)
La identificación de Meloidogyne incognita (Chitwood, 1949), se
realizó mediante cortes sagitales de hembras adultas, en machos
adultos se observó el estilete, la posición del bulbo esofágico, la parte
posterior del cuerpo y su medición corporal, confirmando la
identificación el Ing. Quispe, Nematólogo del Laboratorio de Sanidad
Vegetal del SENASA
3.2.2.1 Reactivación de cepa
Una vez obtenidos las raíces de tomate con las agallas formadas por
el nemátodo, fue llevado al laboratorio en el cual se realizó la
disección de las agallas para obtener masas de huevos del nemátodo.
Esta operación se realizó con sumo cuidado con la ayuda de un bisturí
tratando de no cortar las masas de huevos. Una vez obtenidas las
masas de huevo se colocaron en una placa de Petri estéril, estas
masas de huevos fueron lavadas con hipoclorito de sodio al 0.5 % por
espacio de 30 segundos, luego fueron lavadas con agua destilada
estéril por 4 veces. Se obtuvieron 30 masas de huevos de
Meloidogyne incognita Chitwood, 1949.
La reactivación consistió en: a las masas de huevos expuestos en la
placa de Petri se les asperjó una suspensión de conidias 1 x 108
conidias / mililitro y se les dejó con la suspensión de conidias por un
espacio de 2 minutos, luego con la ayuda de un pincel fueron
colocadas en una placa de Petri que contenía agar agua, se colocaron
10 masa de huevos por placa, se colocaron en la incubadora a una
temperatura constante de 25ºC y a una humedad de 65% por un
espacio de 8 días.

28
3.2.2.2 Aislamiento del hongo Verticillium chlamydosporium (Goddard ,1913)
Al observarse que las masas de huevo presentaban micelio, con la
ayuda de un asa de siembra se sembró por puntura en placas de Petri
que contenían medio PDA, luego se colocaron en la incubadora a una
temperatura constante de 25ºC y a una humedad de 65% durante 8
días.
3.2.2.3 Cultivo monoespórico
El cultivo monoespórico se realizó siguiendo la metodología propuesta
por Lecuona (1995). La metodología fue la siguiente: con un
sacabocado de un 1cm de diámetro se tomó una muestras del hongo
con medio y se realizaron diluciones (10-4
), luego se sembró en placas
de Petri que contenían agar agua en cuyo fondo se marcaron unas
líneas en forma de W, la siembra se realizó con la ayuda de un asa
de siembra por encima de las líneas marcadas, una vez listas las
placas, fueron rotuladas, selladas y se colocaron en la incubadora a
una temperatura constante de 25ºC y a una humedad de 65% por
espacio de cuatro días.
3.2.2.4 Re-aislamiento del hongo Verticillium chlamydosporium (Goddard ,1913)
Para el re-aislamiento del hongo se utilizaron cinco tratamientos: T1
(medio 90), T2 (Saboraud dextrosa agar), T3 (Papa dextrosa agar), T4
(Papa dextrosa agar + levadura) y T5 (Corn meal agar).

29
A los cuatro días de desarrollo del cultivo monoespórico, con la ayuda
de un bisturí se cortaron las colonias y se colocaron en placas de Petri
que contenían los diferentes tratamientos.
Por cada tratamiento se prepararon cinco placas colocándose una
colonia en el centro de la placa, se dejaron incubar por 12 días a una
Tº de 24 ºC a una humedad de 60%, durante 8 días.
Se realizó el control de calidad
3.2.3 Producción del hongo Verticillium chlamydosporium (Goddard ,1913)
Se realizó siguiendo la metodología empleada en el Laboratorio de
Entomopatógenos de la SCB, la cual consiste en una producción
bifásica, una fase líquida y una fase sólida
3.2.3.1 Fase I 3.2.3.1.1 Preparación de inóculo líquido
Se prepararon diferentes medios de cultivo líquido con la finalidad
de obtener el medio más adecuado en que se consiga el desarrollo
óptimo de Verticillium chlamydosporium (Goddard, 1913) Los
medios líquidos utilizados o tratamientos fueron: T1 (Czapeck), T2
(Papa dextrosa + levadura de cerveza), T3 (Medio 90), y T4 (Papa
- dextrosa)
El procedimiento fue el siguiente: se prepararon 300 ML de inóculo
de cada medio, para lo cual se utilizaron frascos erlenmeyer de 500
ML de capacidad. Una vez preparados los frascos se esterilizaron
a 121 ºC y 15 libras de presión durante 15 minutos, dejando enfriar a
medio ambiente. Una vez fríos, a cada frasco se le agregó 0.3 GR
de sulfato de estreptomicina y luego se inocularon con una placa

30
que contenía Verticillium Chlamydosporium (Goddard ,1913)
esporulada.
Por cada medio se prepararon de tres frascos, haciendo un total de
15 frascos.
Se dejó agitar por espacio de tres días en un agitador orbital a una
velocidad de 150 rpm a 24ºC
Los medios líquidos utilizados
M90:
1000 ML de agua destilada
5 GR de peptona universal
3 GR de extracto de malta
Czapeck:
1000 ML de agua destilada
35 GR de medio
PD + levadura
1000 ML de agua destilada
12 GR de levadura de cerveza
20 GR de dextrosa
PD
1000 ML de agua destilada
20 GR de dextrosa
200 GR de papa
Para preparar el medio PD + levadura y PD, se utilizó papa huayro,
la cual fue pelada, cortada y sancochada en el agua, luego el
líquido obtenido fue pasado por un tamiz cuidando de que no pase
ningún residuo sólido, se enrasó a 1000 ML con agua destilada
estéril y se esterilizó en autoclave a 15 libras de presión y 121 ºC
por espacio de 20 minutos.

31
3.2.3.1.2 Control de calidad del inóculo
Por cada tratamiento se tomó 1 ML de inóculo y se realizaron
diluciones hasta obtener la dilución 10-7
, con la ayuda de una
micropipeta se sembraron 300 microlitros por cada placa que
contenía agar agua, se realizaron 3 repeticiones por inóculo. Se dejó
incubar a una Tº constante de 24 ºC.
Para verificar el medio que presentaba mayor concentración de
conidias, se utilizó la metodología de unidades formadoras de
colonias UFC.
3.2.3.2 Fase II 3.2.3.2.1 Preparación de sustrato sólido
Como soporte sólido para la producción de conidias aéreas
(propágulos infectivos), se ensayaron diferentes cereales,
teniéndose siete tratamientos: T1 (sorgo), T2 (cebada), T3 (trigo), T4
(arroz + salvado de trigo), T5 (arroz seco), T6 (arroz húmedo), T7
(arroz seco + biodac). El arroz seco fue el tratamiento testigo, pues
es el que se utiliza para la producción de los diferentes hongos que
produce el Laboratorio de Entomopatógenos de la SCB.
Los granos (sorgo, cebada y trigo) se dejaron remojar por un tiempo
de 16 horas, luego se dejaron escurrir por 30 minutos. Para el arroz
húmedo, se dejó remojar por 2 horas y se dejó escurriendo por 30
minutos.
Para esta fase se prepararon bolsas de polipropileno que contenía
100 GR de sustrato. Por cada tratamiento se prepararon 5 bolsas,
haciendo un total de 35 bolsas (Tabla Nº 02).

32
Se procedió a autoclavar por 30 minutos a 121 ºC y 15 libras de
presión.
Cuando las bolsas estuvieron frías, se procedió a sembrarlas en una
cámara de flujo laminar, a cada bolsas se le agregó 4.5 ML del
inóculo preparado, luego se pasaron a la sala de incubación con una
temperatura constante de 24ºC y una humedad de 95%, por un
tiempo de cuatro días, luego de las cuales se abrieron las bolsas
para eliminar la humedad de las bolsas y favorecer la esporulación.
3.2.3.2.2 Determinación de humedad y temperatura
Una vez que se determinó que el mejor sustrato para la producción
de Verticillium chlamydosporium (Goddard, 1913), se procedió a
realizar los ensayos con la finalidad de determinar la humedad
óptima para el desarrollo del hongo.
Para determinar el porcentaje de humedad óptimo de desarrollo de
Verticillium chlamydosporium (Goddard, 1913), se ensayaron los
siguientes porcentajes de humedad: T1 (30%), T2 (46%), T3 (56%) y
T4 (66%), se tomó como referencia las humedades que utilizan en la
producción de otros hongos entomopatógenos. La humedad testigo
fue del 30%, que es la humedad utilizada en el Laboratorio de
Entomopatógenos de la SCB del SENASA.
3.2.3.2.2.1Humedad
Preparación de bolsas
Se utilizaron 5 bolsas que contenían 500 GR de arroz, por cada
tratamiento (humedad), con su respectivo testigo, la humedad del
testigo fue de 30%, en total se prepararon 20 bolsas. Una vez listas

33
las bolsas, se procedió a esterilizar por 30 minutos en una
autoclave a 121 ºC y 15 libras de presión. (Tabla Nº 03).
Incubación
Cuando las bolsas estuvieron frías, se procedió a inocularlas en una
cámara de flujo laminar, se agregó 30 ML de inóculo por bolsa, se
agitaron para homogenizar el inóculo con todo el sustrato, se
rotularon y se llevaron a la sala de incubación, las bolsas fueron
acondicionadas en la sala de incubación para su germinación y
esporulación, a una temperatura de 24 ± 2 ºC y una humedad
relativa ambiental, por espacio de tres días.
Secado
Luego las bolsas se removieron y se abrieron para eliminar la
humedad y favorecer la esporulación, manteniéndose así por
espacio de cinco días.
La cual mantienen una temperatura constante de 24 ºC y una
humedad ambiental de 95% por espacio de tres días. Después de
este tiempo, las bolsas se removieron y se llevaron a la sala de
secado en donde se procedió a abrirlas para retirar la humedad, en
donde permanecieron por espacio de ocho días a una temperatura
constante de 24 ºC y una humedad relativa ambiental de 60 – 65%
3.2.3.2.2.2 Temperatura
Para la fase de secado del los hongos entomopatógenos, la
temperatura utilizada es de 17 ºC en la cual se obtiene una mayor
cantidad de conidias. Para determinar la temperatura óptima de

34
secado de Verticillium chlamydosporium (Goddard, 1913), se
ensayaron dos temperaturas, 24 ºC y 17 ºC como testigo.
Preparación de bolsas
Se utilizaron 20 bolsas por tratamiento que contenían 500 GR de
arroz, las cuales se prepararon siguiendo la misma metodología ya
descrita anteriormente.
Incubación
Una vez inoculadas las bolsas fueron acondicionadas en la sala de
incubación para su germinación y esporulación, a una temperatura
de 24 ± 2 ºC y una humedad relativa ambiental, por espacio de
tres días, luego las bolsas se removieron y se abrieron para
eliminar la humedad y favorecer la esporulación, manteniéndose
así por espacio de cinco días.
Secado
Después de este tiempo fueron trasladadas a la sala de secado,
en que se mantiene una temperatura constante de 17 ºC. Las
bolsas en desarrollo fueron evaluadas diariamente, con la finalidad
de observar las características típicas del desarrollo del hongo en
estudio
Control de calidad
No se pudo realizar el control de calidad por estrés del hongo.

35
3.2.3.2.3 Control de calidad
3.2.3.2.3.1 Concentración de conidias
Las bolsas con crecimiento de Verticillium chlamydosporium
(Goddard, 1913), se mezclaron para homogenizar las conidias con
el sustrato, se tomó 1 GR y se puso en un tubo de prueba que
contenía 10 ML de agua destilada estéril + tween, se procedió a
agitar en el Vortex por 2 minutos, se tomó 1 ML para llevar a otro
tubo de prueba conteniendo 9 ML de agua destilada esteril
(dilución 10-1
), así sucesivamente hasta obtener la dilución 10-3
.
De esta ultima y mediante una pipeta se llenó la Cámara de
Neubauer, y se hizo la lectura con la ayuda de un microscopio.
Para obtener la concentración de conidias se utilizo la siguiente
formula:
X
C = ----------
Fc (d). N
Donde:
C = Concentración de conidias
Fc = Factor de la cámara
d = dilución de la suspensión
N = Número de cuadrados secundarios tomados e el cuadrado
principal central. En este se contó en cinco cuadrados secundarios
de un total de 25 existentes en la cámara, esto es 5/25 o 1/5.
X = es el promedio de conidias contadas en los cinco cuadrados
secundarios.

36
3.2.3.2.3.2 Porcentaje de germinación o viabilidad
La viabilidad se estimó mediante el porcentaje de germinación de
conidias. De cada bolsa, al igual que para la concentración de
conidias se tomó 1 GR de muestra y se depositó en un tubo de
prueba que contenía 10 ML de agua destilada estéril + tween, se
agitó en un vortex, se tomó 1 ML y se agregó a un tubo de prueba
conteniendo 9 ML de agua destilada estéril + tween, y así
sucesivamente hasta obtener la dilución 10-3
, de esta última con la
ayuda de una pipeta se tomó alicotas de 2 ML y se depositó sobre
laminas delgadas de PDA contenidos en placas de Petri. Luego
se extendió por toda la placa con la ayuda de una asa de siembra
y se incubó a 25ºC durante 15 horas; transcurrido el tiempo, con
un bisturí estéril se cortó cuadraditos de 1cm2 y se colocaron sobre
láminas portaobjeto, luego se agregó 1 gota de azul de lactofenol
para colorear las estructuras del hongo
Se registró el numero de conidias germinadas y no germinadas,
tomando cinco campos de lectura por cada cm2 y tres cm
2 por
placa
Para calcular el porcentaje de germinación se utilizo la siguiente
relación:
% de germinación = a x 100
a + b
Donde:
a = Numero de conidias germinadas
b = Numero de conidias no germinadas

37
3.2.3.2.4 Prueba de patogenicidad
Para las pruebas de patogenicidad se utilizó el método de inmersión,
utilizando una solución de esporas de una concentración conocida,
utilizándose ootecas y huevos de Meloidogyne incognita (Chitwood,
1949)
3.2.3.2.4.1 Tratamiento de ootecas
Se tomaron 100 ootecas de Meloidogyne incognita (Chitwood,
1949), de raíces de tomate. Las ootecas se extrajeron con la
ayuda de dos estiletes y un estereoscopio, fueron lavadas con
hipoclorito de sodio al 0.5% por un minuto, luego fueron
enjuagadas por tres veces con agua destilada estéril. Luego fueron
sumergidas en una solución del hongo Verticillium
chlamydosporium (Goddard, 1913), desarrollado en M90 por un
espacio de 10 días, a una concentración de 2.42 x 109 conidias /
mililitro por 5 minutos, luego con la ayuda de un pincel fueron
colocadas en placas de Petri que contenía agar agua, se
colocaron 20 ootecas por placa, se colocaron en la incubadora a
una temperatura constante de 25ºC y a una humedad de 65% por
espacio de 8 días.
3.2.3.2.4.2 Tratamiento de huevos
Se tomaron 5 GR de raíces con presencia de ootecas, se les
colocó en un frasco con hipoclorito de sodio al 1%, se tapó el
frasco y se agitó durante cinco minutos, con lo cual se logró
desintegrar la masa gelatinosa que recubre los huevos,
obteniéndose una suspensión de huevos, los que fueron lavados

38
tres veces con agua estéril con la ayuda de un tamiz de 38
micrones. Una vez obtenido los huevos se sumergió en una
solución de esporas a una concentración de 2.42 x 109 conidias /
mililitros por 2 minutos, luego con la ayuda de un gotero se
colocaron 5 gotas de la suspensión de huevos tratada en placas
que contenían M90, cada gota contenía aproximadamente 35
huevos, los lugares donde fueron colocados los huevos fueron
marcados con un plumón indeleble por el envés de la placa. Una
vez listas las placas, se colocaron en una incubadora a 25 ºC por
espacio de siete días.
3.2.3.2.4.3 Evaluación
Antes de la evaluación se colocaron cubreobjetos esterilizados en
los medios de cultivo donde se habían colocado los huevos y se
procedió a evaluarlos con la ayuda de un microscopio. El
parasitismo fue reconocido cuando se observó invasión del micelio,
en las partes internas del huevo y la consiguiente destrucción del
mismo, los resultados se expresaron como % de huevos
parasitados
Se evaluaron cinco placas petri, cada de unas de las cuales se
consideraba un repetición y por cada placa se evaluó 100 huevos,
se consideró como testigo a los huevos sin el tratamiento con el
hongo Verticillium chlamydosporium (Goddard, 1913).
Para determinar el porcentaje de mortalidad sobre las ootecas y
huevos de Meloidogyne incógnita, se utilizó la formula de Abbot
modificada:
% Mortalidad = mortalidad en el testigo - mortalidad en el ensayo x 100
100 – mortalidad en el testigo

39
IV. RESULTADOS
4.1 Desarrollo del hongo en medio de cultivo para conservación de cepas
Se observó que el mejor tratamiento a los 10 días de evaluación, fue el
T1 (Medio 90), en el que se obtuvo 4.31 x 108 conidias / mililitro, seguido
del T3 (PDA) con 4.65 x 107
conidias / mililitro. El tratamiento en que
obtuvo menor número de conidias fue el T5 (CMA), con 5.93 x 106
conidias / ML, pero estadísticamente no hubo diferencias significativas
entre los tratamientos evaluados. (ANEXOS Tabla Nº 04) (ANEXOS Fotos
Nº 1, 2, 3, 4,5)
4.2. Desarrollo en medio liquido
Se observó que el mejor medio líquido para la multiplicación masiva del
hongo Verticillium chlamydosporium (Goddard, 1913) es PD con un
promedio de 16,73% UFC/250µl, seguida de Czapeck con 12,23%
UFC/250µl. El medio que presentó la mínima cantidad de UFC/250µl
fue el Medio 90 con 6.63%, pero estadísticamente no hubo diferencias
significativas entre los tratamientos. (ANEXOS Tabla Nº 05).
Para el control de calidad de los medios líquidos se utilizó el parámetro
de UFC/250µl, debido a que los inóculos presentaban aglomeraciones
de micelio, lo que no permitió realizar el conteo de conidias, esto puede
deberse a la característica del hongo que presenta desarrollo sumergido
(ANEXOS Tabla Nº 05). (ANEXOS Foto Nº 8)
4.3 Desarrollo en medio sólido
En la tabla Nº 06, se observa que a los 15 días de inoculados los
tratamientos presentaban diferencias significativas, siendo los mejores
tratamientos el T4 (Arroz seco) con 8.2 x 108
con/g y T7 (Arroz + Biodac)

40
con 7.62 x 108 conidias / gramos siendo el sustrato Sorgo el que
presentó menor número de conidias con 2.74 x 108
conidias / gramos.
Entre los demás tratamientos no hubo diferencias significativas.
(ANEXOS Tabla Nº 06).
A los 23 días se observa las mismas diferencias significativas siendo los
mejores tratamientos T4 y El T7 con 4,5 x 108
conidias / gramos
respectivamente. Con relación a la evaluación a los 15 días, el
tratamiento que presentó menor número de conidias fue cebada con 1.8 x
108
conidias / gramos. Así mismo a los 23 días se observó una
disminución del número de conidias con relación a lo observado a los 15
días. (ANEXOS Tabla Nº 06) (ANEXOS Fotos Nº 10, 11, 12, 13)
A los 30 días no hubo diferencias significativas entre los tratamientos (AENEXOS Tabla Nº 06).
4.3.1 Porcentaje de Germinación
Se obtuvo en todos los sustratos una germinación del 100%, a las 16
horas de haber sido sembrado el hongo en placa, ya que al hacer las
observaciones al microscopio todas las conidias estaban germinadas.
4.4 Porcentaje de Humedad
En cuanto al % de humedad se observó que el testigo, con una humedad
de 30% presentó un número mayor de conidias, de 2.16 x 108
conidias /
mililitro, lo que concuerda con lo utilizado por el Laboratorio de
Entomopatógenos del SENASA, para la producción masiva de los
diferentes hongos entomopatógenos.
Según la prueba de Duncan existe diferencias significativas entre los
tratamientos, aunque numéricamente sean relativamente iguales.
(ANEXOS Tabla Nº 07)

41
4.4.1 Porcentaje de Germinación
Se obtuvo en todos los sustratos una germinación del 100%, a las 16
horas de haber sido sembrado el hongo en placa, al hacer las
observaciones al microscopio todas las conidias presentaban sus tubos
germinativos.
4.5 Porcentaje (%) de Parasitación en Ootecas
De acuerdo a la fórmula de mortalidad corregida de Abbot, se observó
una mortalidad de 32.25% en ootecas por el hongo Verticillium
chlamydosporium (Goddard, 1913)
En donde hubo una mortalidad en el tratamiento (Verticillium
chlamydosporium) de 79% y una mortalidad en el testigo de 69%
4.7 Porcentaje (%) de Parasitación en Huevos
De acuerdo a la formula de mortalidad corregida de Abbot , se observó
una mortalidad del 63.76%, sobre huevos de Meloidogyne incognita
(Chitwood, 1949) por el hongo Verticillium chlamydosporium
(Goddard, 1913). (ANEXOS Foto Nº 17)
En donde hubo una mortalidad en el tratamiento (Verticillium
chlamydosporium) de 73.4% y una mortalidad en el testigo de 26.6%

42
V. DISCUSION
El uso de un agente de control biológico con fines de control de plagas
depende del mantenimiento de las cepas, de la obtención de medios
adecuados para su desarrollo, permitiendo su conservación por un largo
periodo de tiempo, tanto en cepario como en anaquel hasta su utilización.
En este estudio se determinó que para la conservación del hongo
nematófago Verticillium chlamydosporium (Goddard, 1913) el mejor medio
de cultivo para su conservación es el Medio 90 (peptona, extracto de
levadura) lo que concuerda con lo observado por Camargo et al. (2002), que
encontraron que el mejor medio para el desarrollo de Verticilium lecanii, fue
el medio completo (peptona, extracto de levadura y caseína hidrolizada),
manifestando que los resultados obtenidos se debían a que la peptona y el
extracto de levadura contienen varios macro y microelementos que favorecen
el desarrollo del hongo.
El medio líquido en el que se obtuvo mayor cantidad de UFC fue Papa
Dextrosa, lo cual difiere con De Leij y kerry (1991), que utilizaron el medio
Czapeck Dox con lo cual obtuvieron mayor cantidad de conidias
El hongo Verticillium chlamydosporium, tuvo un mejor desarrollo en arroz
seco, lo cual concuerda con Bharadaj y Trivedi (1998), Mendonca (1992),
Ibrahim and Low (1993), Milner et al. (1993), quienes manifiestan que el
sustrato comúnmente seleccionado para la producción de conidias fungales
es el arroz blanco, debido a una combinación de factores incluyendo un
balance nutricional, costo, disponible a nivel mundial, características físicas
tales como el tamaño del grano y su forma, propiedades de hidratación e
integridad estructural aún después de su colonización por el hongo.
La temperatura óptima para todas las etapas de desarrollo de Verticillium
chlamydosporium fue de 24ºC ± 2ºC, lo cual concuerda con Olivares-
Bernebeu, (2002), quien manifiesta que la temperatura óptima para el
desarrollo de hongos parásitos de nematodos fluctúa entre 25 - 30 ºC,

43
mientras que Soberanis et. al. (2005), utiliza 17ºC de temperatura para la
etapa de secado, de los diferentes hongos entomopatógenos y antagonistas
que produce el Laboratorio de Entomopatógenos del SENASA
La humedad óptima del sustrato para el desarrollo de Verticillium
chlamydosprium, fue de 30% en la etapa de germinación, el cual permite la
formación de gran cantidad de conidias. Esto se diferencia de los demás
hongos entomopatógenos, en que se utiliza una humedad de 50%, para esta
etapa (Gómez y Zapata, 1998).
El porcentaje de parasitismo de Verticillim chlamydosporium observado en
huevos de Meloidogyne incognita, fue de 63.76%, lo cual concuerda con
Kerry e Hidalgo (2004), Atkins, et. al. (2003), López-Llorca, et. al. (2002) y
Limay (1998), quienes observaron un parasitismo promedio entre 60 – 70%,
en huevos de Meloidogyne incognita.

44
VI. CONCLUSIONES
1. El mejor medio de cultivo para la conservación de Verticillium
chlamydosporium es el M90, ya que a los 30 días el hongo llega a
formar estructuras de resistencia que son las clamydosporas, las
cuales son ideales para su conservación.
2. El mejor medio líquido a utilizarse para la producción es el Caldo
Papa Dextrosa, en el cual se formaron mayor cantidad de Unidades
formadoras de Colonias (UFC).
3. El sustrato con el cual se obtuvo mayor concentración de conidias y
un porcentaje de germinación del 100% en este estudio fue el sustrato
arroz
4. La patogenicidad de Verticillium chlamydosporium in vitro en
huevos fue de 63.76%, lo cual indica que es un buen controlador de
Meloidogyne incognita
5. En la fase de secado el hongo Verticillium chlamydosporium
necesita una temperatura de 24 ºC a bolsa abierta.
6. En este estudio se observó que hasta los 30 días el hongo
Verticillium chlamydosporium mantiene su calidad en cuanto a
concentración de conidias y germinación permitiendo su estabilidad
por mas tiempo lo cual permite mayor facilidad para su
comercialización.

45
VII. REFERENCIAS BIBLIOGRAFICAS
Agrios, G. (1995) Fitopatología. 2da ed. Ed. Limusa S.A. México. 838.
Atkins, S.D., Hidalgo, L., Kalisz, H., Muchline, T., Hirsch, P. y Kerry B.
(2003) Development of a new management strategy for the control
of toot knot nematode (Meloidogyne spp) in organic vegetable
production. Pest. Management 59: 183-89
Bharadaj, J.P. and Trivedi P. (1998) Biocontrol of Heterodera cajani
using fungus Verticillium chlamydosporium. Jaipur, India.
Bradley, C.A., Black, W., Kearns, R. and Wood P. (1992) Role of
production technology in mycoinsecticides development. In: Frontiers
Industrial Mycology (Leatham, G.F., Ed): 160-73, Chapman and Hall,
London.
Burges, H.D. (1998) Formulation of microbial Pesticides. Kluwer
Academic Publishers, Dordrecht, Netherlands.
Butt, T.M. and Copping, L. (2000) Fungal biological control agents.
Pesticide Outlook 11: 186-191.
Camargo, C., Monteiro, C.A., Bracéelos-Correia, A. y Pereira G.
(2002) Crescimento e esporulacao de isolados de Verticillium lecanii
sob diferentes condicoes nutricionais. Pesq. Agropec bras 6: 821-29.
Ciancio, A. and Leonetti, P. (1999) Isolation and selection of the
nematode parasitic fungus Verticillium chlamydosporium from
perennial and horticultural corps in Italy. Resumenes de la XXXI
Reunion de ONTA, San Juan.

46
Cook, R.J. and Bakes K. (1989) The nature and practice of biological
control of plant pathogens. The American Phytopathological Society.
Crump, D.H. and Irving, I. (1992) Selection of isolates and methods of
culturing V. chlamydosporium and its efficacy as a biological control
agent of beet on Potato cyst nematode. Nematologica 38: 367-374.
Davies, F.A., De Leij, F.A. and Kerry, B. (1991) Microbial agents for
the biological control of plant parasitic nematodes. Tropical agriculture
pest Management 37: 302-320
De Leij, F.A. and Kerry, B.R. (1991) The nematophagus fungus V.
chlamydosporium as a potential biological control agent for M. arenaria.
Revue Nematologica 14: 157-164.
De Leij, F.A., Kerry, B. and Dennehy, J.A. (1992) The effect of fungal
applications rate and nematode density of the effectiveness of V.
chlamydosporium as biological control agent for M. incognita.
Nematologica 38: 112 –122.
Dropkin, V.H. (1954) Infectivity and galls size in tomato and cucumber
seedling infected with. M. incognita var. acriita (root knot nematode)
Phytopathology 24: 453 – 489
Eisenback, J.D. and Triantaphylloum, H. (1991) Root knot Nematodes
Melodoigyne Incognita and Races. In: Manual of agricultural
Nematology Marcel De Kkeir (Nickle, W.R., Ed): 191 – 274, New York.
Fernandez-Larrea, O. (2001) Temas interesantes acerca del control
microbiológico de plagas. Instituto de Investigaciones de Sanidad
Vegetal (INISAV), La Habana.

47
Gómez, H. y Zapata, A. (1998) Guía de producción de hongos
entomopatógenos. PNCB, SENASA.
Hidalgo, L., Montes de Oca, N., Correa, H. y Hernández, M. (2004)
Manual para la investigación y desarrollo de bionematicidas a partir de
una cepa Vcc-108 de Pochonya chlamydosporia var. Catenulata.
CENSA. OCIC. CENSA.
Huang, C.S. (1985) Formation, anatomy and Physiology of giant cells
induced by root- know nematode. In: Advance treatise on
Molodoigyne Biology and control. (Sasser, J.N. and Carter, C.C., Ed)
155-164, North Carolina State.
Hussey, R.S. (1985) Host parasite relationships associated
physiological changes. In Advance treatise on Molodoigyne Biology
and control. (Sasser, J.N. and Carter, C.C., Ed) 143 – 153, North
Carolina State.
Ibrahim, Y.B. and Low, W. (1993) Potential of mass production and
fields efficacy of isolates of the entomopathogenic fungi Beauveria
bassiana and Paecilomyces fumosoroseus against Plutella xyllostella.
International Journal of Pest Management 39: 288-292.
Jenkins, E.N., Haviefo, G., Langewald, J., Cherry, A. y Lomer, C.
(1998) Development of mass production technology for aerial conidia
for use as mycpesticides. Biocontrol News and Information 19 (1): 21-
31.
Kerry, B.R., Simon, A. and Rovira, A.D. (1984) Observations of the
introduction of Verticillium chlamydosporium and other parasitic fungi
into soil for control of the cereal cyst-nematode Heterodera avenae.
Annals of applied biology 105: 509-516.

48
Kerry, B.R. and Bourne, J. (1995) The importance of rizhosphera
interactions in the biological control of plant parasitic nematodes-a
case study using Verticillium chlamydosporium. Pesticide Science 47:
69-75
Kerry, B.R. and Jaffee, B.A. (1997) Fungi as biological control agents
for plant parasitic nematodes. In: The Mycota. IV Environmental and
Microbial Relationships. ( Wicklow and Sodestrom., Ed) Springer-
Verlag, Heidelberg, Berlin.
Kerry, B.R. (2001) Exploitation of the Nemathophagous Fungis
Verticilium chlamydpsorium Goddard for the biological control of Root-
knot nematodes (Meloidogyne spp.) In: Fungi as Biocontrol Agents:
Progress, Problems and Potential. (Butt, T.M., Jackson, C., Magan, N.,
Ed) 155-168.
Kerry, B.R. and Hidalgo, L. (2004) Applications of Pochonia
chlamidosporia in the integrated control of root-knot nematode on
organically grown vegetable crops in Cuba. En Prensa. IOBC.Bulletin.
Lecuona, R. (1995) Microorganismos patógenos empleados en el
control microbiano de insectos plaga. Buenos Aires, Argentina.
Limay, V. (1998) Control de Melodoigyne incognita (Kofoid & white
1919) Chitwood, con seis agentes naturales. (Tesis Ingeniero
agrónomo). Agraria La Molina. Lima: Perú.
Lopez-LLorca, L.V. and Duncan, G.H. (1986) A study fungal
endoparasitsm of the cereal cyst nematode (Heterodera avenae) by
scanning electron microscopy. Canadian Journal of Microbiology 34:
613-619

49
Lopez-LLorca, L.V., Linares-Bernebeu, C.A. and Salinas, J. (2002)
Pre-penetration in fungal parasitism of nematodes eggs. Mycology
4:499-506.
Maheva, E., Laroche, C. and Gross, C.B. (1984) Sporulation of
Penicillium requeforti in solid substrate fermentation. Biotechnology
letters 6: 97-102.
Mendonca, A.F. (1992) Mass production, application and formulation
of Metarhizium anisopliae for control of sugar cane froghopper,
Manaharva postica, en Brazil. In: Biological control of locust ands
grasshoppers. (Lomer, C.J. and Prior, C., Ed) 239-244, Wallingford,
UK.
Meyer, S.L., Huettel, R.N. and Sayre, R.M. (1990) Isolation of fungi
from Heterodera glycines in vitro bioassays for their antagonism to
eggs. Journal of Nematology 22: 532-537.
Milner, R.J., Rogers, D.J., Macrae, C.M., Huppatz, R.J. and Brier, H.
(1993) Preliminary evaluation of the use of Metarhizium anisopliae as
a mycopesticide for control of peanut scarabs. An: Pest control in
sustainable agriculture. Melbourne, Australia; CSIRO. 253-255.
Morgan-Jones, G. and Rodriguez-Kabana, R. (1988) Fungi colonizing
cyst and eggs. In Disease of nematodes. (Painar, G.O. and Jannson,
H., Ed) 39-58, Boca de Raton.
Olivares-Bernebeu, CMª. and Lopez-LLorca, L. (2002) Fungal eggs
parasite of plant parasitic nematodes from Spanish soils. Mycology 19:
104-110.
Prot, J.C. and Netscher (1979) Influence of movement of juveniles on
detection of fields, infested with Melodoigyne, root knot nematode

50
(Melodoigyne especies) systematic biology and control. (Lamberti, F.
and Taylor, C.E., Ed) 193 – 203, New York.
Quispe, C. (1995) Manejo Integrado de plagas. INIA, Huaral.
Siddiki, M.R. (2000) Parasites of Plants and Insects. 2nd
edit, St Albans.
UK.
Seggers, R.T., Butt, T.M., Kerry, B. and Peberdy, J.F. (1994) The
nematophagus V. chlamydosporium produces a chymoetestasa-like
protease wich hydrolyses host nematodes proteins in situ.
Microbiology 140: 2715-2723.
Soberanis, W. (2005) Informe anual PNCB-SENASA.
Stirling, G.R. (1991) Biologycal control of plant Parasitic Nematodes.
Progress, Problems and Prospect. CAB, Wallingfor, UK, 283.
Talavera, M.R. (2003) Métodos culturales. In: Manual de Nematologia
Agrícola.
Taylor, A.L. and Sasser, J.N. (1983) Biología, identificación y control
de los nematodos del nódulo de la raíz (especies de Melodoigyne)
North Carolina State; (traducido al español por el centro internacional
de la papa) 111.
Tikhonov, V.E., Lopez-LLorca, L., Salinas, J. and Jansson, H-B. (2002)
Fungal genetics and Biology 35: 67-78
Whitehead, A.G. (1997) Plant Nematodo Control. CAB Internacional,
Wallingford.

51
ANEXOS

52
Tabla Nº 01
Agentes microbianos para el control biológico de nemátodos parásitos de
plantas. (Kerry, 1995)
Grupos Agentes potenciales Probable modo de acción
Parásitos obligados
Bacterias
Pasteuria penetrans
Pasteuria thornei
Pasteuria rishizanome
Parásitos verdaderos que colonizan estados
vermiformes y sedentarios de nematodos y
los cuales no tienen un modo de acción
toxica
Hongos Hirsutella rhossiliensis
Drechmeria coniospora
Parásitos
facultativos
Hongos parasito de
nematodos
Paecylomices lilacinus
Verticillium lecanii
Verticillium chalmydosporium
Parásitos de nematodos que pueden
también colonizar materia orgánica en
el suelo y la rizosfera.
Hongos atrapadores
de nematodos
Arthrobotrys oligospora
Hongos endefiticos Fusarium oxysporum
Cylindrocarpon destructans
Mycorrhizae
Compiten en las raíces de las plantas y
destruyen las células alimenticias de
los nematodos sedentarios o producen
toxinas, los nematodos también pueden
ser colonizados
Bacterias de la
rizosfera
Agrobacterium radiobacter
Bacillus aphaericus
Bacillus subtilis
Reducen la invasión de Las raíces por
nematodos mediante la producción de
toxinas, cambios en los exudados
radiculares o por una supresión
reducida
Hongos del suelo Trichoderma harzianum
Gliocladium virens
Producen metabolitos tóxicos en el
suelo y o en la rizosfera
53
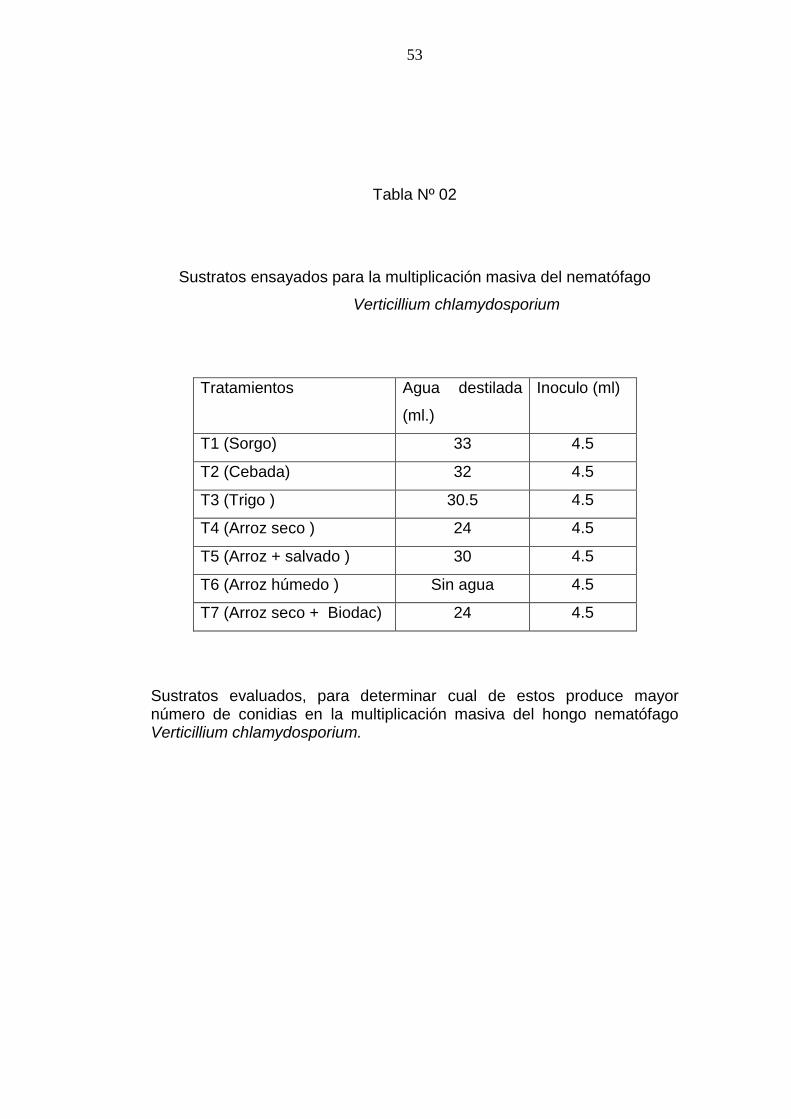
Tabla Nº 02
Sustratos ensayados para la multiplicación masiva del nematófago
Verticillium chlamydosporium
Tratamientos Agua destilada
(ml.)
Inoculo (ml)
T1 (Sorgo) 33 4.5
T2 (Cebada) 32 4.5
T3 (Trigo ) 30.5 4.5
T4 (Arroz seco ) 24 4.5
T5 (Arroz + salvado ) 30 4.5
T6 (Arroz húmedo ) Sin agua 4.5
T7 (Arroz seco + Biodac) 24 4.5
Sustratos evaluados, para determinar cual de estos produce mayor número de conidias en la multiplicación masiva del hongo nematófago Verticillium chlamydosporium.

54
Tabla Nº 03.
Porcentaje de humedad del sustrato arroz, para la multiplicación masiva
del nematófago Verticillium chlamydosporium
Tratamiento Cantidad de agua agregada para
500gr de arroz
Cantidad de Inóculo
T1 30% 120 ml. de agua 30 ml
T2 46% 200 ml de agua 30 ml
T3 56% 250 ml de agua 30 ml
T4 66% 300 ml de agua 30 ml
Porcentaje de humedad evaluados, para determinar cual de estos produce mayor número de conidias en la multiplicación masiva del hongo nematófago Verticillium chlamydosporium.

55
Tabla Nº 04
Promedio de concentración de conidias de Verticillium chlamydosporium, evaluados a los 10 días.
Tratamientos X ± DE Sig.
T1 (Medio 90) 4.31 x 108
1.19 a
T2 (SDA) 3.27 x 107
2.90 a
T3 (PDA) 4.65 x 107
3.65 a
T4 (PDA + levadura) 3.26 x 107
2.87 a
T5 (CMA) 5.93 x 106
3.37 a
Según Duncan (α = 0,05) letras iguales en la misma columna no difieren en los porcentajes estadísticamente entre si. X= Promedio. DE = Desviación estándar Sig. = Significancia
Se observó que el mejor tratamiento a los 10 días de evaluación, fue el T1
(Medio 90), en el que se obtuvo 4.31 x 108 conidias / mililitro, seguido del T3
(PDA) con 4.65 x 107 conidias / mililitro. El tratamiento en que obtuvo menor
número de conidias fue el T5 (CMA), con 5.93 x 106
conidias / ML, pero
estadísticamente no hubo diferencias significativas entre los tratamientos
evaluados.
56
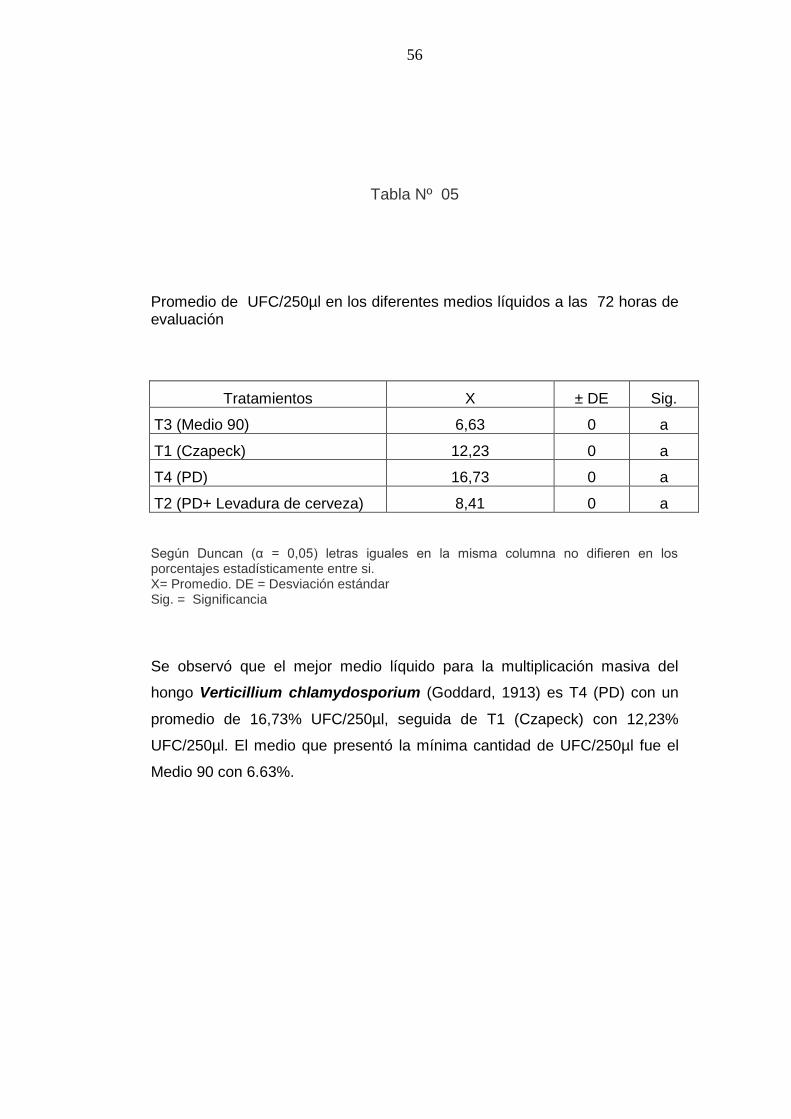
Tabla Nº 05
Promedio de UFC/250µl en los diferentes medios líquidos a las 72 horas de evaluación
Tratamientos X ± DE Sig.
T3 (Medio 90) 6,63 0 a
T1 (Czapeck) 12,23 0 a
T4 (PD) 16,73 0 a
T2 (PD+ Levadura de cerveza) 8,41 0 a
Según Duncan (α = 0,05) letras iguales en la misma columna no difieren en los porcentajes estadísticamente entre si. X= Promedio. DE = Desviación estándar Sig. = Significancia
Se observó que el mejor medio líquido para la multiplicación masiva del
hongo Verticillium chlamydosporium (Goddard, 1913) es T4 (PD) con un
promedio de 16,73% UFC/250µl, seguida de T1 (Czapeck) con 12,23%
UFC/250µl. El medio que presentó la mínima cantidad de UFC/250µl fue el
Medio 90 con 6.63%.
57
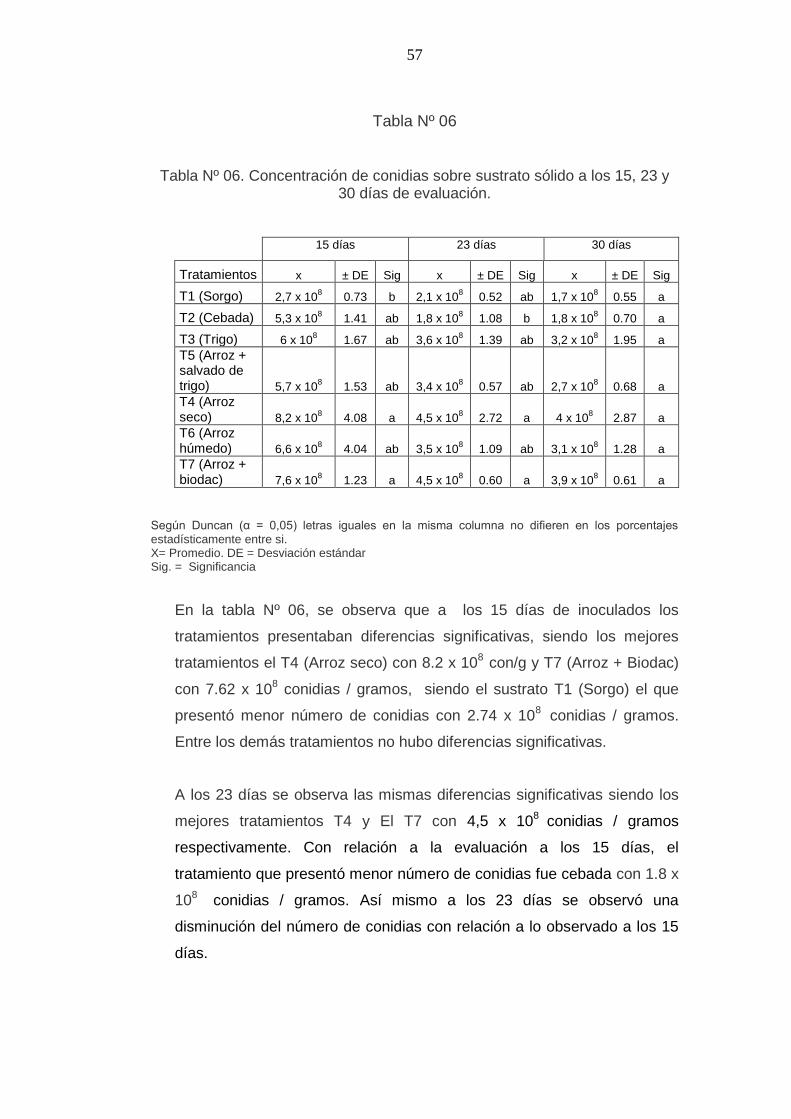
Tabla Nº 06
Tabla Nº 06. Concentración de conidias sobre sustrato sólido a los 15, 23 y 30 días de evaluación.
15 días 23 días 30 días
Tratamientos x ± DE Sig x ± DE Sig x ± DE Sig
T1 (Sorgo) 2,7 x 108
0.73 b 2,1 x 108 0.52 ab 1,7 x 10
8 0.55 a
T2 (Cebada) 5,3 x 108 1.41 ab 1,8 x 10
8 1.08 b 1,8 x 10
8 0.70 a
T3 (Trigo) 6 x 108 1.67 ab 3,6 x 10
8 1.39 ab 3,2 x 10
8 1.95 a
T5 (Arroz + salvado de trigo) 5,7 x 10
8 1.53 ab 3,4 x 10
8 0.57 ab 2,7 x 10
8 0.68 a
T4 (Arroz seco) 8,2 x 10
8 4.08 a 4,5 x 10
8 2.72 a 4 x 10
8 2.87 a
T6 (Arroz húmedo) 6,6 x 10
8 4.04 ab 3,5 x 10
8 1.09 ab 3,1 x 10
8 1.28 a
T7 (Arroz + biodac) 7,6 x 10
8 1.23 a 4,5 x 10
8 0.60 a 3,9 x 10
8 0.61 a
Según Duncan (α = 0,05) letras iguales en la misma columna no difieren en los porcentajes estadísticamente entre si. X= Promedio. DE = Desviación estándar Sig. = Significancia
En la tabla Nº 06, se observa que a los 15 días de inoculados los
tratamientos presentaban diferencias significativas, siendo los mejores
tratamientos el T4 (Arroz seco) con 8.2 x 108
con/g y T7 (Arroz + Biodac)
con 7.62 x 108 conidias / gramos, siendo el sustrato T1 (Sorgo) el que
presentó menor número de conidias con 2.74 x 108
conidias / gramos.
Entre los demás tratamientos no hubo diferencias significativas.
A los 23 días se observa las mismas diferencias significativas siendo los
mejores tratamientos T4 y El T7 con 4,5 x 108
conidias / gramos
respectivamente. Con relación a la evaluación a los 15 días, el
tratamiento que presentó menor número de conidias fue cebada con 1.8 x
108
conidias / gramos. Así mismo a los 23 días se observó una
disminución del número de conidias con relación a lo observado a los 15
días.

58
A los 30 días no hubo diferencias significativas entre los tratamientos

59
Tabla Nº 07
Tabla Nº 07. Influencia de la humedad en el desarrollo de V. chlamydosporium a los 12 días de inoculados.
Tratamientos X ± DE Sig.
T3 (56%) 1.38 x 108 0.09 c
T2 (46%) 1.68 x 108 0.18 bc
T4 (66%) 1.98 x 108 0.19 ab
T1 (30% testigo)) 2.16 x 108 0.23 a
Según Duncan (α = 0,05) letras iguales en la misma columna no difieren en los porcentajes estadísticamente entre si. X= Promedio. DE = Desviación estándar Sig. = Significancia
En cuanto al % de humedad se observó que el testigo, con una humedad
de 30% presentó un número mayor de conidias, de 2.16 x 108
conidias /
mililitro, lo que concuerda con lo utilizado por el Laboratorio de
Entomopatógenos del SENASA, para la producción masiva de los
diferentes hongos entomopatógenos.

60
Re- aislamiento del hongo Verticillium chlamydosporium
Foto 1. Vista anterior y posterior de placa que contiene medio Papa dextrosa agar + levadura de cerveza con el hongo Verticillium chlamydoporium a los 6 días de haber sido traspasada la colonia de un cultivo monoesporico (TABLA 04).
Foto 2. Vista anterior y posterior de placa que contiene medio Papa dextrosa agar con el hongo Verticillium chlamydoporium a los 6 días de haber sido traspasada la colonia de un cultivo monoesporico (TABLA 04)

61
Foto 3. Vista anterior y posterior de placa que contiene medio 90 con el hongo Verticillium chlamydoporium a los 6 días de haber sido traspasada la colonia de un cultivo monoesporico (TABLA 04)
Foto 4. Vista anterior y posterior de placa que contiene medio Saboraud dextrosa agar con el hongo Verticillium chlamydoporium a los 6 días de haber sido traspasada la colonia de un cultivo monoesporico (TABLA 04)

62
Foto 5. Vista anterior de placa que contiene medio Corn meal agar con el hongo Verticillium chlamydoporium a los 6 días de haber sido traspasada la colonia de un cultivo monoesporico (TABLA 04)
Foto 6. Placa que contiene M90 después de 30 días de haber sido sembrado el hongo Verticillium chlamydodporium, de un cultivo monoesporico, se logro la obtención de clamydosporas

63
Foto 7. Clamidosporas con cuatro células (estructuras de resistencia) del hongo Verticillium chlamydporium.
Foto 8. Placas con Medio agar agua el cual han sido sembrados con 300 microlitros de diferentes medios líquidos, después de tres días de agitación orbital a 150 rpm. De izquierda a derecha, Medio 90, Czapeck, Papa dextrosa y Papa dextrosa + levadura. Nótese la mayor cantidad de UFC en el medio líquido Papa dextrosa. (TABLA 05)
Clamidosporas

64
Foto 9. Sustratos con Verticilium chlamydosporium en sala de esporulación
Foto 10. De izquierda a derecha Arroz seco y arroz húmedo con el hongo Verticillium chlamydoporium (TABLA 06)

65
Foto 11. De izquierda a derecha Biodac y Sorgo con el hongo Verticillium chlamydoporium (TABLA 06)
Foto 12. De izquierda a derecha Cebada y Trigo con el hongo Verticillium chlamydoporium (TABLA 06)

66
Foto 13. Arroz + salvado de trigo con el hongo Verticillium chlamydoporium (TABLA 06)
Foto 14. Sustrato Arroz con Verticillium chlamydosporium a los 15 días de
sembrado. (Producto Final)

67
Foto 15. El daño causado por Meloidogyne incognita en tomate forma
nodulaciones en las raíces impidiendo la absorción de nutrientes
(Lycopersicum esculetum).
Foto 16. Masa de huevo de Meloidogyne incognita en raíz de tomate
(Lycopersicum esculetum)

68
Foto 17. Parasitacion en ootecas de Meloidogyne incognita (inmersión de
ootecas) en una suspensión de conidias de Verticillium chlamydosporium
Foto 18. Ootecas en Agar agua

69
Foto 19. Hifa de Verticillium chlamydosporium emergiendo de huevo de
Meloidigyne incognita.
Hifa emergiendo de huevo
de Meloidogyne incognita





























