Embed Size (px)
Citation preview

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/261452295
Enigmas de la ciencia: el calamar gigante
Book · June 2006
CITATIONS
10READS
1,368
5 authors, including:
Some of the authors of this publication are also working on these related projects:
CEFAPARQUES View project
CAIBEX View project
Angel Guerra
Spanish National Research Council
382 PUBLICATIONS 5,749 CITATIONS
SEE PROFILE
Ángel F. González
Spanish National Research Council
242 PUBLICATIONS 3,155 CITATIONS
SEE PROFILE
Francisco Rocha
University of Vigo
83 PUBLICATIONS 1,797 CITATIONS
SEE PROFILE
All content following this page was uploaded by Angel Guerra on 18 January 2016.
The user has requested enhancement of the downloaded file.

Nota importante: la paginación es incorrecta.
Enigmas de la Ciencia:
El Calamar Gigante

Enigmas de la Ciencia:
El Calamar Gigante
Ángel Guerra, Ángel F. González, Francisco Rocha,
Joaquín Gracia y Luis Laria
Vigo, 2006

Queda rigurosamente prohibido, sin la autorización escrita de
los titulares del Copyright, bajo las sanciones establecidas en
las leyes, la reproducción total o parcial de esta obra por
cualquier medio o procedimiento, comprendidos la
reprografía y el tratamiento informático, y su distribución.
A efectos bibliográficos esta obra debe ser catalogada y
citada como sigue:
Guerra, A., González, A. F., Rocha, F., Gracia, J. y Laria, L.
2006. Enigmas de la Ciencia: El calamar gigante. Guerra et
al., (eds.). Instituto de Investigaciones Marinas (CSIC, Vigo),
España, 313 pp.
Distribuido por: CEPESMA, Aula del Mar, Luarca, España y
por el Instituto de Investiagaciones Marinas (CSIC, Vigo).
Este libro se ha realizado con el patrocinio de la Fundación
CAJA RURAL DE ASTURIAS.
© 2006 respecto a la primera edición.
ISBN: 84-609-999-8
Depósito Legal: C-752-2006
Diseño de cubierta y composición: Francisco Rocha
Impreso por Agencia Gráfica. Santiago de Compostela.
Impreso en España. Printed in Spai

ÍNDICE
INTRODUCCIÓN ………………………………………………………………. 9
Capítulo 1. El KRAKEN: DE LEYENDA A REALIDAD ……….. 19
La leyenda del KRAKEN …………………………………………….. 19
Un calamar gigante real e irrefutable ………………………. 38
La consolidación de los estudios científicos …………….. 44
Capítulo 2. LAS ESPECIES DE ARCHITEUTHIS QUE
EXISTEN EN EL MUNDO ………………………………………… 57
Nombres comunes y científicos ……………………………….. 57
El concepto de especie …………………………………………….. 61
El género Architeuthis ………………………………………………. 63
Johan Jappetus Steentrup ……………………………………….. 66
La historia de los nombres del calamar gigante ……… 69
Capítulo 3. LA DISTRIBUCIÓN GEOGRÁFICA DE
ARCHITEUTHIS …………………………………………………...... 77
El Atlántico y el Mediterráneo …………………………………… 77
El Pacífico …………………………………………………………………… 90
El Índico …………………………………………………………………….. 98
Comentarios sobre su distribución mundial …………….. 101
Distribución vertical en la columna de agua ……………. 103
Capítulo 4. PRIVILEGIOS EN LAS AGUAS ASTURIANAS .. 107
El Jurásico terrestre y marítimo asturiano ………………. 107
El árbol genealógico de los cefalópodos ………………….. 109
Algunos compañeros de los cefalópodos en su
escenario evolutivo ……………………………………………….. 111
Rasgos principales en la evolución de los
cefalópodos ……………………………………………………………. 114
Montañas en tierra y fosas en el mar ………………………. 117
Exploraciones acústicas ………...................…………….. 120
¿Por qué aparecen tantos calamares gigantes
en Asturias? …………………………………………………………… 127
Capítulo 5. LO QUE ACTUALMENTE CONOCEMOS SOBRE

LA VIDA DE LOS CALAMARES GIGANTES ……………. 133
Algunas características generales de los
Cefalópodos ………………..…………………………………………. 133
Forma y función ………………………………………………………… 135
Anatomía interna ………………………………………………………. 141
Estrategia reproductiva …………………………………………….. 162
Edad y crecimiento ……………………………………………………. 164
Estrategia trófica: depredadores y presas de
los cefalópodos ………………………………………………………. 174
Comportamiento ……………………………………………………….. 190
Capítulo 6. OTROS CEFALÓPODOS GIGANTES ………………. 201
No todos los cefalopodos gigantes son Architeuthis … 201
El calamar gato o pulpopota de profundidad,
Taningia danae …………………….……………………………….. 204
El calamar de Humbolt, Dosidicus gigas ………………….. 208
La cranquiluria antártica, Mesonychoteuthis
hamilton …………………………………………………………………. 216
Capítulo 7. EL CALAMAR GIGANTE Y EL GRAN PÚBLICO . 219
Museos y exposiciones ……………………………………………… 219
Literatura ………………………………………………………………….. 226
Películas, documentales y otros ………………………………. 233
Expediciones …………………………………………………………….. 242
Los peludines de Asturias ………….............………………. 252
El peludín del Canea ........................................... 256
Y el mudo habló ................................................ 258
Los peludines y los niños asturianos ..................... 259
Capítulo 8. ASPIRACIONES Y PERSPECTIVAS DE
FUTURO ……………………………………………………………………….. 265
Aumentar el conocimiento sobre las
profundidades submarinas ............................... 265
Equipar a España con instrumentos modernos
de exploración submarina ................................ 268

Los calamares gigantes: organismos
emblemáticos para inspirar la conservación
de los océanos ................................................ 272
BIBLIOGRAFÍA ………………………………………………………………… 275
LOS AUTORES …………………………………………………………………. 303
ÍNDICE SUBJETIVO …………………………………………………………. 309
INTRODUCCIÓN
Hace ahora unos veinte años que el primer autor de
este libro tuvo su primer encuentro con un calamar gigante.
Por aquel entonces, el ejemplar que se pudo estudiar fue un
Architeuthis de 38 Kg. capturado por el buque vigués
Farpesca IV en aguas de Namibia. Este animal quedó
apresado en las redes del pesquero en su última campaña de
merluza, fue congelado y trasladado posteriormente al
Instituto de Investigaciones Marinas de Vigo. Cinco años más
tarde llegaron otros tres ejemplares de la misma
procedencia, aunque pescados por distintos barcos: dos
hembras inmaduras de 55 y 40 Kg. y un macho de 41 Kg.
Los datos aportados por estos ejemplares contribuyeron a
incrementar los conocimientos sobre estos intrigantes
cefalópodos, describiéndose por vez primera su alimentación.
Pero además, esos hallazgos marcaron profundamente al
grupo de investigación sobre Ecología y Biodiversidad Marina
(ECOBIOMAR), de manera que los calamares gigantes
constituyeron un tema de estudio continuo, aunque
esporádico, debido, principalmente, a lo fortuito de las
muestras.
La colaboración entre ECOBIOMAR y la Coordinadora
para el Estudio y Protección de las Especies Marinas
(CEPESMA), con sede en Luarca, había comenzado a
mediados de los noventa con motivo de nuestro mutuo

interés por la investigación y protección de los mamíferos
marinos. ¿Quién iba a pensar entonces que esa relación
pondría al descubierto uno de los filones más importantes a
nivel mundial para el estudio de los calamares gigantes?
Pues así fue. El 9 de diciembre de 1999 un ejemplar de 148
Kg. fue capturado accidentalmente por una pareja de
arrastreros lucenses mientras pescaba bacaladilla entre 270
y 550 m de profundidad. El calamar lo compró un pescadero
en la lonja de Avilés y lo expuso en su puesto en el mercado.
De allí lo rescató Luis Laria, presidente de CEPESMA, quien lo
congeló y nos avisó del acontecimiento. El 25 de enero de
Fig. 1. Ejemplar de calamar gigante de 148 Kg. de peso disecado en enero de 2000 en Luarca (Asturias).
2000 íbamos bastante excitados camino de Luarca en el
vehículo todo terreno de nuestro Instituto. Desafiando al frío

reinante, disecamos el animal al aire libre en el patio del
colegio del Padre Galo, rodeados de los alumnos del colegio,
curiosos y periodistas.
Gracias al interés de Luis y de otros miembros de
CEPESMA, y con la colaboración económica de CAJASTUR, el
ejemplar quedó reconstruido, fijado y depositado en una
urna en el Aula del Mar de Luarca. Se iniciaba de esta forma
una colección de calamares gigantes que, con el tiempo, ha
llegado a ser la más importante del mundo. Esa fue la veta
que nos condujo a descubrir que esos enormes cefalópodos
eran bien conocidos en Asturias, donde los pescadores y
otras gentes del mar les habían dado el nombre vernáculo de
“peludines”, debido a su tendencia a pelarse después de ser
capturados. En la búsqueda histórica que iniciamos entonces
se encontraron registros de muchos años atrás en Asturias y
Galicia, remontándose hasta vivos recuerdos de pescadores
en los años cincuenta. Algunos de los encuentros entre
asturianos y Architeuthis quedaron registrados por escrito;
así, en el número 140 de la Hoja del Mar se encuentra la cita
de un ejemplar de 140 Kg. recogido en Figueras en
noviembre de 1979, y en el libro Asturias desde el Mar
(1986), escrito por el ovetense Jesús Ángel Ortea, aparece la
fotografía de un calamar gigante, de buen tamaño a juzgar
por la comparación con sus hijos que aparecen en la
ilustración, descargado en la lonja de Avilés en septiembre
de 1982. Luego se hallaron citas consecutivas procedentes
de la región occidental asturiana y la nororiental gallega,
cerrándose con ellas el arco de tiempo comprendido entre
mediados y finales del siglo XX.
A partir de aquella hembra, bautizada por los niños
del colegio Padre Galo como “Archi”, las llamadas de Laria y
nuestras visitas a Luarca para disecar y estudiar los
ejemplares se han sucedido ininterrumpidamente hasta
ahora. Además de especimenes de Architeuthis, tanto
machos como hembras, hemos podido estudiar cuatro
Taningia danae, otro calamar que alcanza grandes
dimensiones, y que mora también en el caladero de pesca de
Carrandi.

Cuando en ECOBIOMAR comenzamos las
investigaciones sobre el calamar gigante, su ciclo vital era
uno de los enigmas oceánicos por resolver y, pese a los
avances realizados, todavía quedan numerosos aspectos por
desentrañar.
La leyenda del calamar gigante que atrapa y hunde
barcos es muy antigua. Hasta Julio Verne se vio seducido por
estos animales en un inolvidable pasaje de su novela “20.000
leguas de viaje submarino”. Pero Verne, como buen escritor
de ciencia ficción, se había basado en el hallazgo realizado
hace un siglo y medio por el zoólogo danés Johan Japetus
Steenstrup, quien aportó un pico de calamar de 11,5 cm de
longitud como prueba irrefutable de la existencia de estos
gigantes marinos.
Estos cefalópodos constituyen el alimento básico de
numerosos depredadores de gran tamaño,
fundamentalmente cachalotes, en cuyos cuerpos dejan las
huellas de sus ventosas provistas de anillos córneos
dentados, como muestra de su lucha por intentar escapar de
las potentes mandíbulas de estos imponentes mamíferos
marinos. Los cachalotes son capaces de sumergirse a más de
mil doscientos metros de profundidad para capturar los
calamares gigantes usando técnicas de ecolocación. La
impresión de sus ventosas en la piel de los cachalotes ha
llevado a decir que los calamares gigantes pueden alcanzar
hasta cerca de una tonelada de peso, lo cual parece
exagerado aunque se trate sin duda de los mayores
invertebrados vivientes.
Estos auténticos Kraken o monstruos marinos nunca
han sido observados vivos, y todo lo que se conoce sobre
ellos es fruto del estudio de restos encontrados en los
estómagos de sus depredadores, de aquellos ejemplares que
fueron capturados ocasionalmente o de los hallados varados
en la costa. En este sentido, su estudio ha sido como un
juego detectivesco en el cual se han deducido aspectos de su
biología y ecología a partir de los datos obtenidos de
fragmentos de su cuerpo o de ejemplares moribundos o
muertos. Sin embargo, todavía son muchas las incógnitas
que precisan una contestación: ¿Cómo utilizan sus enormes

ojos, tan grandes como un balón de balonmano y los
mayores del reino animal? ¿Cómo capturan a sus presas?
¿Cómo encuentran a sus congéneres ya que no forman
cardúmenes o bancos? ¿Cuándo es su época de
reproducción? ¿Cómo es el apareamiento? ¿Dónde desovan?
¿Cómo usan los enormes anillos córneos dentados que tienen
en las ventosas de sus brazos y tentáculos? ¿Cambian de
color? ¿Pueden producir algún tipo de luz? Actualmente, ni
siquiera sabemos cuantas especies hay o si son agresivos,
pero de lo que tenemos certeza es que no son comestibles
por el hombre porque acumulan amoniaco en sus músculos
para poder permanecer flotando entre aguas sin apenas
gastar energía. Una adaptación metabólica que ha hecho,
entre otras cosas, que tengan unas aletas muy reducidas,
incapaces de actuar como herramienta para desplazarse y
sólo útiles para actuar como estabilizadores cuando el animal
se desplaza a reacción, pero ¿lo hace muy a menudo?, y si lo
hace: ¿cuál es la distancia máxima que puede alcanzar
desplazándose de esta forma? De nuevo aparecen numerosas
incógnitas.
Pese a que estos calamares no son comestibles y que
por lo tanto carecen de interés pesquero, su estudio, además
de interés científico y divulgativo, tiene importancia por su
potencial aplicación en numerosos campos de la Biomedicina
y Farmacología. No sólo los especialistas sienten una
creciente fascinación por estos gigantes marinos, sino
también el gran público. De esta forma, las páginas “web”
sobre este tema son muy visitadas y los museos ofrecen
elevadas cantidades de dinero por la adquisición de
ejemplares para sus exposiciones. Desde 1996 se viene
realizando un esfuerzo plurinacional en el que han colaborado
científicos, tecnólogos, expertos en filmación, barcos y
submarinos con objeto de obtener las primeras imágenes de
un calamar gigante vivo en su medio natural.
En la disección de la hembra capturada en Asturias en
enero de 2000, junto a los miembros de ECOBIOMAR y
CEPESMA, se encontraba tomando fotografías y filmando el
biólogo Fernando González-Sitges, director de cine y socio
fundador de la Productora de documentales TRANGLOBE

FILMS, que dirige Javier Linares. Pronto Fernando y Javier
comprendieron la importancia de estos hallazgos; el Kraken
estaba a nuestras puertas constituyendo un auténtico reto
tanto para la ciencia como para la industria de documentales
sobre Naturaleza. De ahí nació un proyecto enteramente
español cuyo objetivo era obtener, por primera vez,
imágenes de un calamar gigante en su entorno natural.
El Proyecto KRAKEN se planteó como una serie de
expediciones semejantes a los grandes viajes de
descubrimiento del siglo XIX. Viajes de revelación destinados
a abrir una brecha en los inexplorados ambientes marinos,
que nos proporcionasen una información nueva, capaz de
explicarnos viejos mitos marinos. Nuestro proyecto nacía
como una iniciativa en amistosa competencia con los
esfuerzos efectuados por las expediciones norteamericanas
realizadas desde 1996 en Nueva Zelanda y las Azores con el
mismo objetivo. La ventaja que teníamos sobre ellos era que
la zona donde se encuentra Architeuthis en el Cantábrico
queda a pocas millas de la costa asturiana, donde Gijón
ofrecía magníficas prestaciones como puerto base. La
desventaja estribaba en los tres años de adelanto que nos
llevaban y en la relativa facilidad con que ellos podían
obtener fondos para sufragar sus expediciones. España, en
cuestiones de patrocinio, no es como los Estados Unidos. En
nuestro país se necesita una ley de mecenazgo más efectiva,
ya que una buena legislación en ese sentido favorece el
desarrollo de las ideas y la aplicación de los resultados. Sin
embargo, todos nos pusimos a buscar financiación con
entusiasmo. En esa tarea nos llevamos bastantes
decepciones y algunas alegrías. Pensábamos, quizá
ingenuamente, que la idea sería entendida con prontitud,
especialmente por las autoridades e instancias sociales
asturianas, pero, pese a los esfuerzos, no fue así en todos los
casos. A excepción de CEPESMA, de las autoridades del
puerto de Gijón, de la Armada española y del actual Director
General de Pesca del Principado, Alberto Vizcaino Fernández,
que estuvieron a nuestro lado desde el primer momento,
encontramos poco eco y demasiadas puertas cerradas. A la
postre, el proyecto salió adelante y se realizaron dos

campañas en busca del Kraken, ambas a comienzos de los
otoños de 2001 y 2002. Con los equipos diseñados
específicamente para estas expediciones se organizó una
exitosa exposición en el Museo Nacional de Ciencias
Naturales (CSIC, Madrid). El material examinado en los
últimos cinco años sirvió para publicar varios trabajos
científicos en revistas internacionales. El 1 de marzo de
2004, la segunda cadena de Televisión Española, proyectó el
documental “Proyecto KRAKEN, en busca del calamar
gigante”, dirigido por Fernando, y que se está actualmente
distribuyendo entre varias cadenas de televisión nacionales y
extranjeras. Cuando se escribe esta introducción, a mediados
de junio de 2004, estamos a la espera de realizar la tercera
expedición del proyecto, sólo nos falta una parte de la
financiación, que tratará de cubrir la Consejería de Cultura
del Principado de Asturias. Confiamos en poder continuar la
sana competencia con nuestros colegas norteamericanos y
australianos, quienes estarán en septiembre en Kaikoura
(Nueva Zelanda) para tratar nuevamente de obtener
imágenes del elusivo calamar gigante. Esta historia está viva
y ahora, cuando se realizó la revisión de este capítulo en
noviembre de 2004, ya conocemos el resultado negativo de
nuestras gestiones para conseguir financiación, y el resultado
infructuoso de las expediciones de Kaikoura.
Aunque parte de lo aprendido sobre el calamar gigante se
ha publicado en revistas científicas, pronto comprendimos la
necesidad de recoger en un libro todo lo que habíamos
estudiado y aprendido sobre estos cefalópodos,
principalmente para intentar difundir el conocimiento
adquirido sobre estos organismos entre el público en general.
Muchas veces hablamos de este proyecto entre nosotros,
hasta que Luis Laria nos puso en relación con D. Román
Suárez Blanco, Presidente de la Caja Rural de Asturias. No
hicieron falta muchas explicaciones para que D. Román
apoyase la idea y nos proporcionase el soporte financiero
para realizarla. Después de una larga y amena conversación
en su casa de Luarca, a finales de 2003, el contenido del
trabajo estaba ya esbozado.

Este libro, escrito con rigor científico, pretende tener
un marcado carácter divulgativo con objeto de ofrecer a los
lectores un texto en el que se recojan los conocimientos
actuales sobre la biología y ecología de los calamares
gigantes, así como una relación de todos los registros
mundiales conocidos hasta la fecha y su distribución por
mares y océanos, analizándose, además, la influencia de los
fenómenos atmosférico-oceánicos sobre su distribución. Se
recoge también en este libro la secuencia histórica
transcurrida entre la aparición de los mitos sobre estas
enigmáticas criaturas y el afianzamiento de la realidad a
partir de los conocimientos científicos; los problemas que
encierran su sistemática y taxonomía y las técnicas
modernas que se están aplicando para solucionarlos; la
historia sobre las exposiciones y modelos realizadas y
diseñados para museos de todo el mundo; las principales
expediciones para filmarlo; los datos más relevantes sobre
otros calamares gigantes; las manifestaciones artísticas y
culturales, en forma de pinturas, grabados, libros,
documentales, películas y novelas, inspiradas por el Kraken;
y una extensa y cuidada bibliografía. Por último, aunque no
menos importante, hemos querido también recoger todas las
manifestaciones que relacionan a los calamares gigantes con
Asturias. Todo ello con una cuádruple esperanzada
pretensión: que este trabajo contribuya a que las autoridades
y la sociedad asturiana se aperciban de la importancia
científica, cultural y turística de lo que tienen en sus mismas
puertas y actúen en consecuencia; que se profundice en la
protección del medio y las especies marinas; que la
investigación marina se tome más en serio en España; y por
último, que la Ciencia penetre más profundamente en el
tejido social de nuestro país.
En una obra de esta índole, que supone la culminación
de una larga tarea, son muchas las personas que han
colaborado, por ello resulta prácticamente imposible referirse
a todas ellas, algunas de las cuales ya han sido citadas en
esta introducción. Sin embargo, aun conscientes que
podemos incurrir en omisiones, es obligado mostrar nuestro
agradecimiento a María Teresa Fernández Álvarez, entre

otras cosas por su colaboración técnica en la elaboración de
los ficheros informáticos bibliográficos, a D. Román Suárez
Blanco por su decisivo apoyo, a todos los participantes en las
expediciones del Proyecto KRAKEN, especialmente a José
Castro, Fernando González-Sitges, Javier Linares, José
Manuel Novoa, Jaime Otero, Gerardo Olivares y Oscar
Soriano, así como a Cándido Correa, armador de los buques
“Investigador” y “Científico”, a sus capitanes, Fernando
Santiáñez y Ramón Arguibay, y a los demás miembros de sus
tripulaciones. Estamos también en deuda con Alberto
Rodríguez Costa, cirujano pediátrico del Hospital Xeral de
Vigo, quien nos ayudó a preparar varios ejemplares de
Architeuthis y nos enseñó técnicas de reconstrucción de
órganos y de suturación. Estamos también agradecidos a
Alfredo López, Josep Mª Alonso Farré y a otros miembros de
la Coordinadora para el Estudio de los Mamíferos Marinos
(CEMMA), que nos acompañaron en la atención a varios
varamientos ofreciendo su apoyo técnico y colaboración, así
como a las tripulaciones de los barcos Elena María, Bautista
Pino, Minchos V y VI, Boer, Pino Ladra, Peña de Deva,
Luscinda, Travasía y Valdés Vega quienes nos allegaron los
calamares gigantes. Hay que sumar a estos reconocimientos
a varios establecimientos asturianos, como Pescaderías
Jiménez y Pescados Milagro, Paco y Somio, que tuvieron en
sus manos ejemplares, que luego nos fueron ofrecidos. Son
también muchos los particulares que merecerían aparecer en
este elenco de gratitudes, entre otros Celso Martínez Villar y
García Ramos. Por otra parte, ha sido esencial en numerosas
ocasiones la colaboración de la Guardia Civil, los Bomberos
de Asturias y Protección Civil 112, con quienes hemos
quedado en deuda, la cual debe hacerse extensible a la
Cofradía de pescadores de Avilés y a las lonjas Musel-Gijón y
Luarca. Por último, aunque no por ello menos importante,
vaya nuestro agradecimiento más sincero para todos los
voluntarios de CEPESMA, que tan útil y desinteresado apoyo
han prestado en las tareas de recolección, transporte,
disección y preparación de los calamares gigantes hallados
en Asturias.

Capítulo 1
EL KRAKEN: DE LEYENDA A REALIDAD
La leyenda del Kraken
La leyenda es una narración que deforma la historia, o
prescinde de ella, referida a personas, animales o cosas que
han existido realmente, o bien a figuras imaginarias ligadas a
un hecho y lugar concretos y reales, que acontecieron en un
determinado momento. En el origen de toda leyenda hay un
individuo que la crea, aunque aparezca anónima y se
atribuya a una colectividad. Es consustancial a toda leyenda
el carácter de cosa admirable y casi increíble, no
necesariamente fantástica, que produce asombro, y
veneración en algunos casos. Si en el origen de la leyenda
hubo un acontecimiento histórico, su evolución viene
conformada por factores que la amplían o simplifican, que la
fragmentan, o que la trasladan de lugar y época; en
definitiva, que la contaminan de alguna manera. Hasta
nuestros días han llegado leyendas de todo tipo,
generalmente originadas en la antigüedad, cuando los
métodos históricos no estaban todavía bien fijados, y
preponderaba la transmisión oral.
Leyenda y mito no son cosas equivalentes. Un mito es
una explicación sobrenatural de fenómenos naturales, donde
generalmente los personajes son dioses o semi-dioses y el
tiempo una ficción. Se trata de relatos transmitidos por una
tradición de antigüedad remota, donde se refieren y hacen
presente, en estrecha relación con el culto y el lenguaje
imaginativo, las acciones de dioses o personajes
sobrenaturales. El motivo de los mitos difiere con las
civilizaciones, pero en el caso de los griegos los temas
derivados de la Naturaleza son poco frecuentes, mientras
que en los escandinavos están bastante más ligados a su
culto, interviniendo frecuentemente animales.
Es bastante plausible que el calamar gigante
Architeuthis sea el responsable del mayor número de

leyendas, mitos e historias de ficción que cualquier otro
animal marino, y casi nos inclinamos por pensar que más
que todos ellos juntos. Las leyendas y el folklore de muchas
tierras mencionan monstruos que podrían estar relacionados
con los calamares gigantes, que podrían haber engendrado el
mito griego de Scylla.
En la mitología griega Scylla era un monstruo marino
que vivía debajo de una roca peligrosa en uno de las
márgenes del estrecho de Mesina, frente del torbellino
Caribdis. Scylla amenazaba el paso de las naves y en la
Odisea se relata como se comió a seis compañeros de Ulises.
Scylla era una ninfa, hija de Phorcys. El dios-pescador
Glaucus se enamoró de ella, pero Scylla huyó de él hasta
donde no podría seguirle. Glaucus acudió entonces a la bruja
Circe para pedirle una poción de amor que derritiese el
corazón de Scylla. Glaucus contó su historia de amor a Circe
y ésta se enamoró de él, cortejándole con palabras dulces y
miradas lascivas, pero Glaucus no atendió a su cortejo. Circe
se enfureció por ello y preparó un frasco con una poción
mágica de gran efecto, que vertió en la piscina donde Scylla
se bañaba. Tan pronto como la ninfa entró en el agua se
transformó en un monstruo espantoso. Sus doce patas eran
como grandes tentáculos parecidos a serpientes, sobre ellas
tenía seis cabezas con bocas provistas de tres filas de
dientes. Scylla quedó sumida en la más completa miseria,
incapaz de moverse del sitio, detestando y destruyendo todo
cuanto quedaba a su alcance con gran ferocidad. Se
alimentaba de delfines, tiburones y otras presas de gran
tamaño que se ponían a su alcance, y era un peligro para
todas las embarcaciones que pasaban cerca. Es muy
probable que la figura de serpiente en que se transformó la
ninfa fuese tomada de algún gran calamar o pulpo (polypus),
conocidos por los griegos desde la antigüedad. Entre los
libros que escribió el romano Cayo Plinio Segundo “El Viejo”
(79 al 23 antes de Cristo) únicamente nos ha llegado su
Naturalis Historia (Historia Natural) donde habla de un
“pólipo” que se apoderó de peces marinos en las pozas de
Carteia (en la costa Atlántica española), y “que se atrajo la
furia de los cuidadores, quienes, debido a la persistencia del

robo, lo consideraban lejos de todo límite admisible”. Los
guardas que rodearon a aquella bestia estaban: “atónitos
por su rareza; en primer lugar, porque su tamaño era
increíble, así como lo era su color; además, estaba
embadurnada de salmuera y exhalaba un olor nauseabundo
¿Quién habría esperado encontrar allí una criatura
semejante, o quien la habría reconocido en tales
circunstancias? Los guardas sintieron que se enfrentaban a
algo extraño, porque su horrible respiración también
atormentaba a los perros, que fueron azotados con la punta
de sus tentáculos y golpeados con sus largos brazos, que usó
como mazas. No sin gran dificultad consiguieron reducirla
usando varios tridentes”. Según la crónica de Plinio, la
cabeza de aquella criatura consistía en una gran bolsa con
capacidad para 400 litros; sus brazos (anudados como
mazas) median cerca de 9 metros de longitud y estaban
provistas de ventosas de más de un litro de capacidad, y sus
dientes correspondían a su tamaño. Sus restos pesaron cerca
de 320 Kg. Sus largos tentáculos identifican a aquella bestia
o pólipo como un calamar, más que como un pulpo. Pero
fuese lo que fuese, esa criatura debería constar como el
mayor de los cefalópodos que se haya acercado a la costa
por cualquier razón. Plinio comenta que: “estaba dentro de
los tanques descubiertos y que procedía del mar abierto, ello
quizás porque los tanques eran accesibles a ese habitante de
las profundidades”. Plinio dependió de relatos previos para
redactar esta crónica, pero, asumiendo que algo semejante a
lo relatado ocurrió realmente en la antigüedad e incluso
admitiendo exageraciones, este es el único hecho de tal
naturaleza recogido en toda la literatura antigua, donde,
junto con rasgos muy reales, se encuentran ya varios
aspectos de la leyenda del Kraken.
Habrá que esperar varios siglos para que la bestia
marina provista de varios brazos vuelva a aparecer en los
relatos. Una criatura semejante vuelve a citarse en el libro
Historia de gentibus septentrionalibus (Historia de los
pueblos del Norte) escrito por Olaus Magnus (1490-1557)
obispo católico de Upsala. Este tratado es una auténtica
fuente para el estudio de los monstruos marinos. En uno de

los pasajes del libro el obispo escandinavo se expresa de la
siguiente forma: "Aquellos que, para comerciar o pescar,
navegan por las costas de Noruega, coinciden en el relato de
una historia realmente admirable, la de una enorme
serpiente (Soe Orm) de una longitud de más de 200 pies, y
20 pies de diámetro que vive en las rocas y agujeros cerca
de la costa de Bergen; sólo sale de su caverna en las noches
de verano y con buen tiempo, para devorar terneros,
corderos y cerdos, o se sumerge en el mar para comer
pulpos, langostas y todo tipo de cangrejos. Tiene una hilera
de pelos de dos pies de largo que le cuelgan del cuello,
escamas afiladas de color oscuro, y ojos brillantes y
llameantes. Ataca a los barcos, se levanta fuera del agua, se
lleva a los hombres y los devora; y normalmente esto no
sucede sin que algo terrible acontezca en el reino, sin un
cambio próximo: o los príncipes van a morir o serán
exiliados, o una guerra va a estallar pronto."
Las descripciones y los dibujos que aparecen en este
libro establecieron con firmeza la existencia de muchas
criaturas fabulosas, y fueron reproducidos, en original o
retocadas, durante varios siglos, lo que aseguró al autor un
puesto preeminente en el campo de las leyendas zoológicas.
Fig. 2. La serpiente marina (Soe Orm) de Olaus Magnus, 1555.

La mayoría de las descripciones de Olaus Magnus
corresponden a animales marinos reales, aunque con claras
exageraciones, lo que puede explicarse porque escribía
basándose en testimonios directos de los hechos pero
distorsionados por los avatares de la transmisión oral. Sin
embargo, ¿cuál de esas descripciones correspondía a un
calamar gigante?
Los monstruos marinos son la parte principal de la
obra De Piscium et Aquatilium Animatum Natura (Sobre los
Peces y Animales Acuáticos de la Naturaleza) de Conrad
Gesner publicada en 1558. Uno de los dibujos más
interesantes de Gesner es una criatura con siete cabezas, un
largo cuerpo con escamas, dos pies con garras y una cola
curva. Quizás influido por las descripciones de la mitológica
hidra griega, ya que debía ilustrar un animal que no le era
familiar en absoluto, y tratando de mostrar a sus lectores
algo semejante a lo ya conocido, Gesner dibujó unas cabezas
con crestas como las de los pavos al final de cada cuello.
Algunos criptozoólogos consideran que pueda estar
refiriéndose a un calamar gigante, si se ve a los cuellos y las
cabezas de este monstruo como sus brazos y al cuerpo como
el de un cefalópodo de gran tamaño. Lo que no parece tener
una explicación adecuada son los pies con garras que ilustró
Gesner, que pudieran ser añadidos de su propia imaginación
o de la de sus confidentes.
Fig. 3. Monstruo marino dibujado por Gesner (1558)

Posteriormente, en Histoire of Foure-Footed Beastes
(Historia de las Bestias de Cuatro Patas) de Edward Topsell
(1607) se reproduce este, y otros dibujos, de Gesner. Con
relación al monstruo marino que se ilustra, y al que se
relaciona con la hidra matada por Hércules en sus trabajos,
Topsell reproduce la historia contada por Gesner sobre un
cadáver enorme de un organismo marino que fue
transportado desde Turquía hasta Venecia, y que, presentado
a las autoridades de la República, quisieron ver en el
monstruo una “mutación de las cosas del mundo que
señalaban la inminente llegada de un cataclismo”.
Introducidos de lleno en el siglo XVII, además de los
dibujos de Olaus Magnus, que aparecen en las obras de De
piscibus (Sobre los peces) de Ulisses Aldrovandi (1613) y en
la Historia naturalis de Joannes Jonstonus (1649), pueden
hallarse historias fantásticas incluyendo monstruos marinos
con características de calamares gigantes en las obras de
varios enciclopedistas del Renacimiento.
Cincuenta años más tarde, y gracias a la aparición y
conservación de algunas mandíbulas y tentáculos de
enormes dimensiones, Francesco Negri realizó los primeros
intentos de descripción de los calamares gigantes que fueron
registrados en su obra Viaggio settentrionale (1700).
Una historia llena de interés humano y científico es la
del clérigo protestante danés Hans Egede, quien llegó a ser
obispo de Groenlandia y vivió en esa isla durante muchos
años con el ánimo de convertir al cristianismo a los nativos.
Allí estableció dos asentamientos, en 1721 y 1723
respectivamente. Además de sus poco exitosas experiencias
misioneras y de maravillosas descripciones de la tierra y las
gentes de Groenlandia, en la obra “Der gamle Grøelands nye
perlustratio, éller natural-histoire” publicada en Copenhague
en 1741, el reverendo Egede cuenta un episodio bastante
ilustrativo sobre el tema que nos ocupa. Se trata de un
monstruo que se vio en la superficie del mar en el 6 de julio
de 1734 cerca de una de las colonias fundadas por él hacia
los 64º de latitud Norte. Según cuenta el obispo, se trataba
de un enorme animal cuya cabeza era tan alta como el palo
mayor del barco en que viajaba, su cuerpo tan corpulento

como el casco, y entre tres y cuatro veces más largo. Ese
monstruo presentaba un hocico puntiagudo y resoplaba como
una ballena, tenía aletas muy amplias, y todo él parecía estar
cubierto con pequeñas conchas, siendo su piel muy rugosa e
irregular. La parte inferior de su cuerpo se asemejaba a una
enorme serpiente, y cuando se sumergía lo hacía con la cola
hacia arriba, saliendo fuera del agua.
La personalidad del narrador, hombre de prestigio,
honesto y nada dado a exageraciones, nos inclina a aceptar
su declaración, incluso aunque no podamos explicar o
entender lo que vieron él y los demás tripulantes del barco,
que observaron algo previamente desconocido.
Afortunadamente, sin embargo, el dibujo realizado por el
señor Bing, otro de los misioneros en tierras groenlandesas,
nos ha permitido tener una idea de lo que observó el
reverendo Egede. Debemos recordar que se trata de un
esfuerzo por retratar el incidente, que debió acontecer en un
tiempo muy breve. Podemos reconocer de un vistazo el
dibujo de un viejo conocido, y afirmar casi sin vacilación que
el monstruo marino de Egede era un calamar gigante. Así lo
interpretó Henry Lee quien en su libro Sea Monster
Unmasked (Monstruos Marinos Desenmascarados), publicado
en Londres en 1883, sugirió una explicación bastante
plausible de lo que pudieron observar el reverendo Egede y
sus compañeros, y, desde luego, todo indica que pudieron
avistar un calamar gigante.
Fig. 4. Dibujo del Sr. Bing del monstruo observado por el reverendo Egede

Fig. 5. El animal que probablemente observó el reverendo Egede según sugerencia de Henry Lee (1883)
De las aproximadamente 570 citas de calamares
gigantes bien referenciadas en todo el mundo desde el siglo
XVI, 27 se produjeron en las costas de Noruega. La más
antigua en esas aguas aconteció en el siglo XVII.
Posteriormente, entre 1874 y 1950, hay referenciados 25
avistamientos, pero luego el número decayó abruptamente.
Es muy posible que en aquellas aguas hubiese en el pasado
abundantes calamares gigantes, alguno de cuyos cadáveres
vararon en sus costas y fueron vistos por bastante gente. Por
ello no es de extrañar que sea otro noruego, el obispo de
Bergen Erik Ludvigsen Pontoppidan quien, en su libro La
Historia Natural de Noruega publicado en 1752, haga
referencia a monstruos marinos, que en el lenguaje popular
de aquellas tierras se denominaban Kraken. Un árbol
arrancado de raíz pudiera ser una descripción bastante
adecuada para un calamar gigante, no obstante, un
monstruo de hasta 22 metros de longitud total es un pigmeo
en comparación con el Kraken tal como lo describe el obispo
Pontoppidan “el más grande y sorprendente de todos los
animales de la creación” e “indudablemente el monstruo
marino más grande del mundo”, ya que afirma que los
pescadores le contaron que vendría a medir hasta dos
kilómetros y medio de circunferencia. Esta enorme bestia
insular no parece poder conjugarse bien con un calamar
gigante, y muy posiblemente el buen obispo se creyó a pies
juntillas lo que en realidad podría ser una agrupación de
ballenas solazándose en la superficie del mar, mezclando ese
dato con las informaciones recibidas a partir de dos

cadáveres, posiblemente de calamares gigantes,
encontrados con la marea alta en Amunds Vaagen
(Nordfjord) y en la isla de Kramen, respectivamente. La
descripción de Pontoppidan contiene algunas características
sugerentes: “la criatura no se parece a una anguila o a una
serpiente, se agudiza gradualmente hacia un extremo, y su
cuerpo, que es tan grande como dos cabezas de cerdo, se
estrecha progresivamente justo donde comienza la cola. La
cabeza en todos los tipos tiene una frente alta y ancha, pero
en algunos hay un hocico puntiagudo, mientras que en otros
es chata, como la de un buey o un caballo, con amplios
orificios nasales, y varios pelos rígidos extendidos a cada
lado como bigotes”. Richard Ellis (1998) comenta que,
excepto los orificios nasales y los bigotes, que son
problemáticos, el resto de la descripción se ajusta con
bastante precisión a un Architeuthis, sobre todo si se
considera que lo está describiendo una persona que no tiene
ni idea de con que tipo de criatura está tratando. La cola que
se describe pueden ser los largos brazos del calamar
extendidos, a partir de los cuales sale el cuerpo, que el
obispo confunde con la cabeza, en cuya parte anterior se
sitúa la verdadera cabeza del calamar, y cuyo manto se
agudiza hacia el ápice dando lugar a la auténtica cola.
Pontoppidan también describe los ojos de la criatura que son
“muy grandes y de color azul, y se parecen a un par de
brillantes bandejas de peltre”, así como su color general
“castaño oscuro, pero moteado y abigarrado con franjas
luminosas y lunares que brillan como el caparazón de una
tortuga”. Desde luego que el buen obispo parece estar
describiendo los ojos y la piel de un calamar gigante fresco.
En la bibliografía sobre las serpientes marinas, que
durante siglos estuvieron ligadas a los calamares gigantes,
hay unanimidad en que Pontoppidan fue el primero en
describir “científicamente” una de estas criaturas. El obispo
había hecho leer ante el Tribunal de Justicia de Bergen una
carta del capitán Lorenz von Ferry en la que se describía con
todo lujo de detalles una serpiente marina que él y su
tripulación habían avistado mientras se dirigían a tierra en un
bote de remos, junto a la localidad de Molde (Noruega) en

1746. El capitán describía así a la serpiente: “tenía una
cabeza gris semejante a la de un caballo, grandes ojos
negros, boca negra y larga melena blanca. Detrás de la
cabeza del monstruo, pudieron apreciar hasta siete u ocho
promontorios que salían del agua, y el cuerpo de la bestia se
retorcía formando espirales”. Cuando el capitán von Ferry
ordenó hacer fuego contra la serpiente, ésta se sumergió en
el agua y no volvió a aparecer.
En el transcurso del siglo XVIII hubo otros
avistamientos de serpientes marinas, como el que describió
Charles Douglas, quien, a bordo del velero HMS Emerald en
1769, en una de las primeras campañas oceanográficas,
avistó uno de estos seres en las proximidades de las costa
finlandesa, dando cuenta del hallazgo y describiendo la
serpiente marina en la Royal Society de Londres en 1770. Sin
embargo, el peso cada vez mayor de la crítica racionalista y
del análisis científico determinó que los informes de los
marinos y pescadores que habían divisado monstruosas
bestias marinas fueran considerados exagerados y ridículos.
Por ejemplo, el científico noruego Peter Ascanius afirmó que
la hilera de jorobas, que generalmente divisaban los
hombres de la mar, no pertenecía a ningún descomunal
monstruo marino, sino a una comitiva de delfines haciendo
cabriolas. Y esta explicación tan endeble se convirtió desde
entonces en el recurso favorito de quienes pretendían
desacreditar los testimonios sobre la existencia de estos
monstruos marinos.
El 6 de agosto de 1848, el capitán M'Quhae y seis
miembros de la tripulación del Daedalus divisaron lo que
pensaran era una serpiente marina de 18 metros de largo en
aguas del Atlántico meridional, cerca del cabo de Buena
Esperanza. Un informe de prensa ilustrado sobre el suceso
desencadenó una cruda polémica, en la que el renombrado
naturalista Sir Richard Owen defendió que esos monstruos
marinos no existían y que lo observado probablemente se
trataba de un león marino, pero el capitán M'Quhae se
mantuvo firme en su convicción de que lo que había visto era
una serpiente marina, que ilustró en su informe a la Real
Sociedad de Londres.

En su libro sobre los calamares gigantes, Richard Ellis
presenta algunos argumentos y una ilustración bastante
ingeniosos con objeto de demostrar que lo que M’Quhae y su
tripulación observaron era un calamar gigante nadando en
superficie, pero a nuestro juicio esa es sólo una de las
posibilidades, ya que, existen bastantes organismos marinos
que podrían ser tomados como una serpiente.
Durante la primera mitad del siglo XIX se registraron
numerosas observaciones de “serpientes marinas” a lo largo
de la costa nororiental de América, sobre todo en torno al
puerto pesquero de Gloucester, en Massachusetts.
Constantin S. Rafinesque, profesor de Ciencias Naturales de
la Universidad de Transylvania, en Lexington, Kentucky,
examinó los informes, les concedió credibilidad y decidió
dividirlos en cuatro grupos, uno de los cuales denominó
Megophias, es decir, serpientes gigantescas. Pero los
investigadores de fenómenos inexplicados sobre las
apariciones de bestias marinas siguieron encontrando una
fuerte oposición entre los científicos. Esa controversia fue
recogida en varias publicaciones del siglo XIX entre las que
cabe destacar The Romance of Natural History del naturalista
británico Philip Henry Gosse publicada en 1861, y Mythical
Monsters de Charles Gould, miembro de la Real Sociedad de
Tasmania.
Una de las críticas más aceradas de Owen respecto a
la existencia de estos monstruos marinos era que nunca
habían sido observados por científicos. Sin embargo, esta
objeción iba a desaparecer en 1905 cuando dos naturalistas,
E. G. B. Meade-Waldo y M. J. Nicoll, vieron un extraño animal
frente a las costas de Brasil en el transcurso de una campaña
científica a bordo del yate Valhalla. Meade-Waldo publicó sus
observaciones bajo el título Description of an Unknown
Animal Seen at Sea off the Coast of Brazil en 1906 y Nicoll lo
hizo en su libro Three Voyages of a Naturalist aparecido en
1908. Aunque Ellis se empeña nuevamente, en su ya citado
libro sobre los calamares gigantes, en considerar que la
ilustración que aportó Nicoll, parecida a la M’Quhae, podría
considerarse como un calamar gigante nadando en
superficie, mostrando un brazo y una parte de su manto con

media aleta al aire, tampoco vemos aquí razones para
desechar que pudiera tratarse de cualquier otra criatura
marina de grandes dimensiones. De hecho, Bernard
Heuvelmans en su libro "Le grand serpent-de-mer: le
problème zoologique et sa solution", publicado en 1965,
realiza un estudio comparativo de varios centenares de
testimonios recogidos entre 1639 y 1965, concluyendo que la
serpiente de mar es en realidad un compendio de múltiples
animales, algunos ya conocidos en la actualidad, como el
calamar gigante (Architeuthis), el tiburón-ballena (Rhincodon
typus), el regaleco (Regalecus glesne), el gusano nemertino
gigante Lineus longissimus, y otros animales desconocidos,
que catalogó como varias especies de vertebrados.
La “serpiente marina” de 7 metros de longitud que
muestra la figura 6 es realmente un regaleco (Regalecus
glesne) capturado en Coronado Beach (San Diego, California)
en noviembre de 1996. Aparentemente murió tras haber sido
atrapado por las hélices de un buque, ya que muestra
múltiples cuchilladas en la mitad del cuerpo y pequeños
cortes en la cabeza. El animal fue disecado en el Centro
Especial de Guerra Naval por M. J. Walker, encargado de la
colección de peces del Instituto Oceanográfico de San Diego.
La cabeza y un metro del cuerpo se conservaron para ser
exhibidos en el Acuario Stephen Birch, mientras la Armada
enterró el resto del cuerpo en la playa.
Los avistamientos de Regalecus glesne documentados
pueden contarse con los dedos de ambas manos. El más
antiguo del que se tiene constancia es un pez serpentiforme
de 17 metros de longitud que fue hallado en las playas
escocesas en 1808, y que muy probablemente perteneciese a
esta especie. Cuatro regalecos se avistaron en los últimos
años en Baja California pero todos, excepto uno, estaban
muertos. A pesar de su feroz apariencia, por su cuerpo en
forma de cuchillo y sus grandes ojos, se trata de criaturas
inofensivas que se alimentan de pequeños crustáceos
planctónicos. Se cree que esta especie pasa la mayor parte
de su tiempo a profundidades que fluctúan entre los 300 y
900 metros y sólo por alguna razón desconocida asciende a

la superficie, donde generalmente se encuentran individuos
moribundos.
Roy P. Mackal propuso en 1980 otras dos nuevas
especies o formas gigantes de invertebrados como posible
identidad de algunos de los casos de “serpientes marinas”: el
pirosoma (Pyrosoma), una colonia tubular de tunicados con
un extremo abierto y el otro cerrado en forma de cono,
fosforescente, y que puede medir varios metros de longitud,
y el cinturón de Venus (Cestus veneris), que puede alcanzar
hasta un metro y medio de longitud.
Algunos de los casos de avistamientos de enormes
animales marinos parecen corresponder a calamares
gigantes, descritos o no como serpientes marinas, otros
consisten en distintos tipos de organismos como el regaleco,
pero hay bastantes que se podrían explicar sin necesidad de
recurrir a gigantescos animales o colonias de miles de
individuos. A veces se toma por una serpiente de mar lo que
sólo es una ballena, o enormes cocodrilos, o una gran
serpiente terrestre arrastrada por el mar. Un grupo de
marsopas nadando en hilera puede parecer también una gran
serpiente, ya que el movimiento encadenado de cada
individuo puede confundirse con la ondulación de un largo
cuerpo serpentiforme.
Fig. 6. Ejemplar de Regalecus glesne de 7 metros de longitud (Coronado Beach, California). Noticia publicada en el San Diego-
Union Tribune en diciembre de 1996

Un reciente ejemplo de este tipo de confusiones es el
caso del pulpo gigante (Octopus giganteus). El 23 de junio de
2003 apareció en los medios de comunicación la noticia de
que había varado en la Playa Pinuno cercana a la localidad de
los Muermos, a 1044 kilómetros al sur de Santiago de Chile,
una masa gelatinosa de más de 15 m de longitud y unas 13
toneladas de peso. Esta masa fue considerada desde el
primer momento como los restos de un pulpo de grandes
dimensiones por pescadores y gentes de la zona. Una
semana después la noticia era conocida por todo el mundo y
se realizaban las primeras conjeturas sobre si se trataba de
restos de una ballena, pulpo gigante o incluso un
monumental calamar. La confusión generada sobre la
naturaleza de estos restos se debió principalmente a que las
primeras personas que lo observaron carecían de
conocimientos biológicos. Por otra parte, las fotografías
tomadas buscaban más el sensacionalismo que la objetividad
y la zona era muy poco accesible y alejada, siendo difícil para
los especialistas llegar hasta el lugar.
Fig. 7. Restos hallados en la playa de Pinuno cercana a Muermos (Chile) el 23 de junio de 2003

Afortunadamente, un grupo de biólogos del Museo de
Historia Natural de Santiago, al frente del cual estaba Sergio
Letelier, pudo examinar los restos, realizar precisas
mediciones y tomar muestras. Sus análisis revelaron que no
se trataba de ningún cefalópodo, sino de un gran cetáceo,
probablemente de un cachalote, que debió morir en alta mar
donde flotó en superficie durante algún tiempo, por lo que su
cadáver se fue pudriendo paulatinamente, llenándose sus
tejidos del gas generado por la descomposición. Cuando ésta
alcanzó un determinado umbral, los tejidos comenzaron a
soltarse del esqueleto formando una gran masa gelatinosa,
que los vientos y las corrientes arrastraron hacia la playa
donde se depositó con la apariencia de un pulpo gigante.
En algunas de esas noticias aparecidas por entonces
se comentaba que se trataba sin duda de un pulpo
gigantesco, parecido al encontrado en un varamiento similar
ocurrido en Florida en 1896. Efectivamente, el 30 de
noviembre de 1896 se hallaron los restos de un cadáver, que
medían 5,4 m de largo, 2,1 m de ancho y 1 m de altura,
varados en los arenales de la isla Anastasia, a 18 Km. al sur
de la playa de San Agustín en la costa oriental de Florida. Los
dos jóvenes que lo encontraron pensaron que se trataba de
los restos de una ballena. Posteriormente, varios miembros
de la Sociedad Agustiniana de Ciencia, Literatura e Historia lo
examinaron, dibujaron y fotografiaron, concluyendo que se
trataba de un pulpo gigante. Ninguna de esas personas era
especialista en cefalópodos, y se pusieron en contacto con
A.E. Verrill, eminente especialista norteamericano de la
Universidad de Yale que había descrito varios calamares
gigantes, quien, a partir de esa información y de algunos
dibujos hechos por su propio hijo, lo identificó como un pulpo
al que dio el nombre de Octopus giganteus, información que
publicó en un artículo del American Journal of Science
aparecido en febrero de 1897, donde estimó que esa
gigantesca criatura tendría un peso de 20 toneladas y unos
50 m de longitud total. Verrill y W. Healey Dall, entonces
conservador de moluscos del Museo Nacional en Washington
DC, trataron de volver a examinar esos restos, e incluso
conservarlos, pero no fue posible por falta de financiación y

tiempo. El señor Webb, presidente de la Sociedad
Agustiniana, ayudado por algunos ciudadanos, movió
aquellos restos del lugar donde habían varado para evitar
que fuesen arrastrados por la marea, pero antes tomó varias
muestras de su probable manto, que conservó en formol.
Desgraciadamente, el cadáver se perdió y solo quedaron
esas muestras como prueba, las cuales recibió Verrill el 23 de
febrero. Al examinarlas con detenimiento se dio cuenta que
realmente no correspondían a un pulpo ni a ningún
cefalópodo sino que más parecía un tipo de grasa que
poseen algunos crustáceos, lo cual expuso en un articulo
aparecido en Science el 5 de marzo de 1897. Posteriormente,
el examen del profesor F.A. Lucas, del Museo Nacional,
corroboró que: “la sustancia parece grasa, huele como grasa
y es grasa, nada más y nada menos”. A partir de aquí se
abrió un intenso debate, en el que la mayoría defendía que
aquellos restos correspondían a un pulpo gigante y una
minoría que eran restos de una ballena. Entre esta minoría
se encontraba Verrill, quien se desdijo de su primer
diagnóstico aceptando que se trataba de restos de un
cetáceo. Pero incluso a finales del siglo XX y principios del
XXI el debate continúa. Aquellas muestras conservadas
fueron analizadas mediante mejores técnicas histológicas y
bioquímicas por Wood y Gennaro en 1971 y por Mackal en
1986, concluyéndose en ambos casos que se trataba de un
octópodo, o incluso de un calamar, pero nunca de un
vertebrado. Sus resultados se publicaron en las revistas
Natural History y Crytozoology, respectivamente.
Así permanecieron las cosas hasta que en mayo de
1988 apareció en Bahamas una masa similar a la recogida en
Florida casi un siglo antes, de la que pudieron recogerse
algunas muestras antes de que desapareciese por completo
arrastrada por el mar. El Biological Bulletin de la prestigiosa
institución oceanográfica norteamericana Woods Hole publicó
en abril de 1995 un artículo de S. K. Pierce, G. N. Smith Jr,
T. K. Maugel y E. Clark titulado “El pulpo gigante (Octopus
giganteus) y el “grumo gelatinoso” de Bermudas: Un
homenaje a A.E. Verrill”, donde, después de realizar
rigurosos análisis histológicos y bioquímicos de los tejidos de

los restos aparecidos en Florida y Bermudas, concluyeron
que el primero era el cadáver de un vertebrado de sangre
caliente, y el segundo correspondía a un vertebrado
poiquilotermo, pero ciertamente no a un molusco. Esos
investigadores indicaron también que no existían evidencias
de la existencia de un pulpo gigante en Florida o en
Bermudas. Este artículo produjo en los Estados Unidos una
reacción en otoño de 1995. En el número de primavera de
1996 de la revista Of Sea and Shore apareció publicado un
artículo de G. S. Mangiacopra, D. G. Smith, D. F. Avery, M.
Raynal, R. Ellis, J. R. Greenwell y B. Heuvelans quienes, en
colaboración con el Dr. Mackal, catedrático retirado de
Química de la Universidad de Chicago, defendieron que se
trataba de una especie desconocida de una forma gigante de
octópodo. En su artículo, además de exponer los principales
comentarios sobre este largo debate, muchos de ellos
procedentes de criptozoólogos, señalaban algunos defectos
en la metodología utilizada en los análisis químicos de Pierce
y sus colaboradores: en primer lugar la antigüedad de las
muestras, que podrían haberse desnaturalizado por su
conservación en formol durante cien años, pero también las
inadecuadas comparaciones con otros organismos marinos
de grandes dimensiones. Esto, unido a las observaciones,
fotos y dibujos existentes relativas al cadáver de Florida, a
varias historias de los pescadores nativos de Bahamas, y al
incidente del patrón de pesca, Sean Ingham, quien pescando
langosta con nasas a 1000 metros de profundidad en
Bahamas, observó en la sonda una criatura de forma
piramidal de 15 metros de longitud, que posteriormente se
enganchó en sus trampas y desplazó varios metros su
embarcación, parecen a Mangiacopra y colaboradores
evidencias suficientes para certificar que esos restos
pertenecen a un cefalópodo gigante.
La principal conclusión de todo este largo debate es
que actualmente se carece de datos fidedignos para afirmar
que los restos hallados en Florida y Bermudas corresponden
a un pulpo gigante. El reciente hallazgo en el sur de Chile,
identificado como restos de carne y grasa en descomposición
de un gran mamífero marino, decanta la cuestión hacia

considerar que la mayoría de esos restos no pertenecen a un
cefalópodo, sino a restos de piel y grasa de un gran cetáceo.
Sin embargo, la existencia de pulpos gigantes está
convenientemente comprobada. El mayor de todos ellos es
Enteroctopus dofleini del Pacífico Norte cuyo cuerpo de
consistencia musculosa alcanza más de 80 Kg. de peso y
hasta 3 m de longitud. Junto a este, también se puede citar
al pulpo pelágico Haliphron atlanticus, cuyo cuerpo es
totalmente gelatinoso, y que por lo que hasta ahora se
conoce, puede llegar a pesar hasta 60 Kg. y medir cerca de
tres metros de longitud total.
Las historias de Olaus Magnus, del obispo Pontoppidan
y de otros autores, dieron lugar a la leyenda del Kraken, que
en sus orígenes era una figura de la mitología escandinava.
Un animal de tamaño fabuloso que se dejaba ver raras veces
en la superficie del mar, armado de gruesos tentáculos,
voraz y agresivo. Temido por los marineros escandinavos, el
Kraken fue representado en algunas ocasiones como un
calamar y como un pulpo en otras. En su “Histoire naturelle
des mollusques”, el naturalista francés Pierre Denis de
Montfort basándose en un incidente ocurrido en la costa de
Angola y representado en un exvoto en la iglesia de Saint-
Malo (Francia), describió e ilustró un pulpo colosal, al que
llamó pulpo Kraken. Calamar o pulpo, el Kraken aparece
siempre en actitud beligerante, agarrando a los navíos en
busca de sus desesperadas tripulaciones.
Existen varias interpretaciones sobre el significado de
la palabra noruega Kraken. La versión más aceptada sostiene
que significa "árbol desraizado" por la similitud entre un
calamar gigante y un árbol con las raíces al aire. Para otros,
como para el lingüista y biólogo noruego Jan Haugum, se
trata de una palabra que apareció por primera vez en la
Historia Natural de Noruega de Pontoppidan en la que
significaba monstruo marino. Sea cual sea su origen, al paso
de los siglos, la palabra ha terminado por significar ni más ni
menos que calamar gigante.
La leyenda del Kraken, nacida en tiempos pretéritos
cuando eran insondables las oscuridades de nuestro
conocimiento sobre la naturaleza, no murió con el

racionalismo científico, sino que está viva todavía y
fecundando la imaginación de escritores y cineastas. Ese
gigante de las profundidades de cuerpo gelatinoso, enormes
ojos, poderosas mandíbulas en forma de pico de loro, lengua
rasposa, y largos tentáculos provistos de ventosas nos
produce terror. Y si ya horrorizan su tamaño y su figura,
mucho más lo logran las terribles cualidades que se le
atribuyen: una fuerza poderosa capaz de atrapar barcos
enteros, una tremenda agresividad y una voracidad sin
límites.
Fig. 8. El exvoto de Saint-Malo, reproducido por Pierre Denis de Montfort, 1801

Un calamar gigante real e irrefutable
La transformación del monstruo legendario llamado
Kraken en el muy real, aunque también fabuloso calamar
gigante Architeuthis, probablemente representa la secuencia
de anécdotas más instructiva de toda la historia de la cacería
de monstruos.
El registro más antiguo de un calamar gigante real
con el que se cuenta es el de un cadáver encontrado en
Øresund (entre Malmö y Copenhague) en 1545, que fue
llevado ante el rey de Dinamarca. Este hallazgo fue relatado
por el historiador danés Sørensen Vedel, quien describió al
animal como un curioso pez de unos dos metros y medio de
longitud con forma de monje.
Extraña, sin duda, que se compare la forma de un
calamar con la de un monje, pero así parece que fue pintada
para el rey, quien envió copias a diferentes cortes europeas,
haciendo del “monje marino” una figura famosa en toda
Europa. Ese dibujo fue recogido por el naturalista francés
Pierre Belon, quien, en su obra Nature et diversité des
poissons avec leurs pourtraicts représentez au plus près du
natural, publicada en 1551, describió un monstruo marino
ilustrado como un monje calvo revestido con ornamentos
sagrados (alba y casulla) y con los brazos extendidos, que
son postura y gesto frecuentes en la celebración de la Misa.
Posteriormente, en su obra Libri de piscibus marinis in quibus
verae piscium effigies expressae sunt (El libro de los peces
marinos), publicada en 1554-55, Guillaume Rondelet bautizó
a ese animal con el nombre de Monachus marinus,
incorporando también una ilustración del famoso clérigo
marino. También el naturalista suizo Gesner recibió un dibujo
y una descripción “del monje marino” en Zurich hacia 1558,
la cual incorporó a su libro De Piscium et Aquatilium
Animatum Natura, donde se describe al monje marino como
un hombre con el pelo arreglado como el de un monje de la
época, estando sus vestimentas hechas de escamas, provisto
de dos largas aletas en lugar de brazos y con la parte inferior
finalizando en una cola ancha.

J.J. Steenstrup demostró que el “monje marino” era
un calamar, y que, debido a su gran tamaño, debía tratarse
de un calamar gigante.
De acuerdo con los trabajos de Steenstrup, el
siguiente registro de calamar gigante del que se tiene
constancia ocurrió hacia finales del verano de 1639, cuando
se encontró un ejemplar de ente 7,5 y 9,4 metros de
longitud varado muerto en una playa de Islandia. El siguiente
registro corresponde a un animal de unos 5,70 metros de
longitud, que se encontró varado en la playa de la bahía de
Dingle, comarca de Kerry (Irlanda) en 1673. Este animal fue
expuesto al público, realizándose un tabloide que se imprimió
en Dublín. Desde entonces, y a intervalos irregulares, se
tienen registros de cadáveres de calamares gigantes varados
en playas de todo el mundo en diferente estado de
descomposición. Estos hallazgos fueron de alguna manera
conocidos por el naturalista sueco Linnaeus, quien los incluyó
bajo el nombre se Sepia microcosmus en la primera edición
de su Sistema Naturae publicada en 1735, aunque esa
especie no está recogida en la décima edición del libro, que
es la definitiva, ya que el autor dudó de su existencia y la
eliminó del registro.
Fig. 9. Dibujo realizado por Steenstrup para tratar de explicar el parangón entre el “monje marino” y un calamar.

A la par que acontecían estos primeros balbuceos
científicos en torno al calamar gigante entre los siglos XVI y
XVIII, y que, como ya vimos, crecía la leyenda del Kraken,
entre los balleneros europeos y norteamericanos existía un
conocimiento empírico de que los cachalotes regurgitaban
grandes trozos, e incluso calamares gigantes enteros. Sin
embargo, hubo de esperarse hasta el 30 de noviembre de
1861 para que se produjese el primer encuentro entre un
calamar gigante, entonces ya conocido como Architeuthis y el
hombre. Navegando cerca de las costas de Tenerife, el buque
de guerra francés Alecton avistó un calamar de unos 7,50
metros de longitud flotando en el mar. El comandante
Bouyer, capitán del barco, y su tripulación intentaron
recogerlo, pero sólo lo lograron a medias, rescatando la
sección posterior del manto, que fue trasladada a Tenerife.
La descripción del evento, realizada por Bouyer, es muy
significativa y demuestra que se trataba de un Architeuthis
moribundo. Ese informe fue presentado por Sabine Berthelot,
cónsul francés en las Canarias y naturalista aficionado, ante
la Academia Francesa de las Ciencias un mes más tarde.
Además, los naturalistas Henri Crosse y Paul Fischer
publicaron en 1862 el trabajo Nouveaux documents sur les
céphalopodes gigantesques basado en el informe de Bouyer.
Parece bastante seguro que estos trabajos fueron conocidos
por Julio Verne, quien los incorporó en 1869 a su novela
20.000 leguas de viaje submarino. Con Verne, y poco antes
con Victor Hugo quien trató el tema en su novela Les
travailleurs de la mer de 1866, el calamar gigante penetró
más amplia y profundamente en la imaginación de la gente
de la calle como un animal agresivo, peligroso y repulsivo.
En los años comprendidos entre 1871 y 1881 se
registraron cerca de 60 varamientos o avistamientos de
calamares gigantes en las costas atlánticas de América del
Norte, especialmente en Terranova (Canadá). El más
afamado de estos hallazgos fue el que ocurrió a finales de
octubre de 1873, cuando dos pescadores a bordo de un bote
hallaron un calamar gigante moribundo en Portugal Cove,
una pequeña cala situada en la bahía de Concepción. Desde
un punto de vista científico, lo más interesante de este

acontecimiento es que uno de los tentáculos del calamar fue
entregado por los pescadores al reverendo presbiteriano
Moses Harvey, quien, además de conservarlo en el museo de
San John y de publicar la historia en varios periódicos locales
y en dos revistas científicas, contactó con el zoólogo Addison
E. Verrill de la Universidad de Yale para que lo estudiase.
Fig. 10. Los tripulantes del Alecton tratando de capturar un calamar
gigante al NW de Tenerife en 1861

Fue así como Verrill inició una serie de trabajos sobre
los calamares gigantes, que le llevarían a publicar una
quincena de trabajos científicos sobre Architeuthis. Pero con
el suceso de Portugal Cove asistimos al comienzo de una
nueva leyenda. Existen varias canciones del folklore
canadiense que recogen la lucha que sostuvo un valiente
pescador, Tom Piccot, con un calamar gigante en una
pequeña cala de Terranova. La batalla contra el gigante, una
deformación del hecho histórico, se encuentra también
narrada por N. Duncan y recogida en la Antología de
literatura infantil (1940). Casi cincuenta y cinco años
después, la escritora Annie Prouxl cuenta ese
acontecimiento, ya legendario, en su novela Shipping News
(1993), por la que obtuvo el premio Pulitzer. Y dos años más
tarde, en la novela The Kraken, escrita por D. Reed para un
público juvenil, aparece nuevamente Piccot, un personaje
probablemente inventado por Harvey, como protagonista de
una heroica batalla que nunca existió.
En diciembre de 1873, mientras izaban las redes que
habían calado para pescar arenques, varios pescadores de la
bahía de Log (Terranova), hallaron un calamar gigante
enredado en ellas. Todavía estaba con vida cuando lo izaron
y tuvieron que matarlo con sus cuchillos. En la operación de
meterlo a bordo de la embarcación perdieron el cuerpo, pero
pudieron hacerse con la cabeza y los brazos, que llevaron al
reverendo Harvey, quien obtuvo la primera fotografía de un
calamar gigante casi completo, y permitió a Verrill hacer un
meticuloso examen.
Se debe sin duda a los trabajos sobre los calamares
gigantes publicados por este biólogo norteamericano entre
1874 y 1882, el notable incremento que experimentaron los
conocimientos científicos sobre Architeuthis a finales del siglo
XIX.
Se desconocen con exactitud las razones que podrían
explicar la gran cantidad de apariciones de calamares
gigantes en las proximidades de las costas atlánticas de
Canadá y del norte de los Estados Unidos durante esas
décadas del siglo XIX, pero a mediados XX el biólogo
canadiense Frederick Aldrich lo atribuyó al resultado de

fluctuaciones en la corriente de agua fría procedente del
Labrador. Sin embargo, esa hipotética causa no explicaría
adecuadamente los varamientos y avistamientos de
Architeuthis que se produjeron en las costas de Nueva
Zelanda entre 1879 y 1887. Los ejemplares allí encontrados
fueron estudiados por varios biólogos, entre los que destacan
los neozelandeses T. W. Krik, y C.W. Robson.
En el capítulo tercero de este libro, al tratar de la
distribución mundial de Architeuthis, se pasa revista a las
capturas, varamientos y avistamientos registrados en todo el
mundo desde el siglo XVI hasta nuestros días. En esa sección
queda bastante claro que hubo otros lugares, como las
costas noruegas, donde se registraron calamares gigantes a
finales del siglo XIX, y, además, que a partir de comienzos
del siglo XX se incrementaron notablemente sus registros y
que los núcleos geográficos de mayor frecuencia de aparición
se desplazaron de un lugar a otro. Es bastante probable que
ese incremento general de los registros se deba más a un
mayor esfuerzo de muestreo que a un aumento real de la
aparición de estos animales. Ese incremento del esfuerzo
vendría provocado por un mayor interés general por los
calamares gigantes, por una mayor frecuencia y atención del
rastreo de sus cadáveres en playas y roquedales costeros,
así como al aumento de las zonas visitadas por los
pescadores y por el también creciente tráfico marino
mundial. Tanto en las zonas costeras de Terranova como en
las de Nueva Zelanda debieron quedar varados durante
décadas numerosos calamares gigantes en zonas
deshabitadas o en áreas marinas muy poco frecuentadas por
pescadores y marinos mercantes, siendo muy probable que
fuesen mucho más frecuentes de lo que indican los registros
con los que contamos.
Los resultados obtenidos por aquellos científicos
pioneros contribuyeron notablemente a un mejor
conocimiento de Architeuthis, sobre todo en lo referente a su
morfología y anatomía, pero, debido fundamentalmente al
tipo y condición de las muestras (fragmentos o animales
muertos, muchas veces en avanzado estado de
descomposición), poco pudieron decir sobre otros aspectos

básicos de la biología de estos gigantes. Además,
introdujeron en la nomenclatura científica de ese género
numerosos nombres de especies cuya validez es bastante
discutible, aunque contribuyeron notablemente a que las
leyendas fuesen quedando bien acotadas, y a que la realidad
se fuese imponiendo.
La consolidación de los estudios científicos
Desde comienzos del siglo XX numerosos científicos
de varias nacionalidades han ido realizando una labor
detectivesca para comprender mejor al mayor invertebrado
existente. No obstante, el conocimiento que actualmente se
tiene de su ciclo vital, fisiología, ecología y comportamiento
es muy fragmentario y siempre condicionados por los
hallazgos esporádicos de Architeuthis, bien varados o
flotando cerca de la costas, en capturas accidentales o a
partir de restos encontrados en los estómagos de sus
principales depredadores. De hecho, todavía nadie ha visto o
filmado un solo ejemplar vivo en su elemento.
El primer ejemplar del que se tiene constancia en los
albores del siglo XX apareció en marzo de 1909 flotando en
superficie en las proximidades del cabo Cod (Massachusset,
EE.UU.). Sin embargo, es Noruega la zona con mayor
número de registros. Tal como se documentó en 1916,
cuando August Brinkman publicó un listado de todos los
registros existentes hasta entonces en ese país, junto con
varias observaciones sobre un calamar gigante hallado
flotando en superficie al norte de Bergen en noviembre de
1915. Esta información fue completada años después, en
1954, por Erling Sirvertsen, quien proporcionó un mapa con
los lugares donde se hallaron los 18 registros que
acontecieron en Noruega desde 1874 hasta entonces.
La publicación de J. Kjennerud de 1958 es muy
interesante porque en ella se cita uno de los primeros y
escasos ejemplares de macho de Architeuthis encontrados en
todo el mundo. Pero todavía lo es más la del dinamarqués J.
Knudsen, quien en 1957 publicó las primeras observaciones

de un macho maduro que apareció con esperma incrustado
en varios de sus brazos.
Otro de los ejemplares estudiados en Noruega iba a
proporcionar también una información sorprendente. El 23 de
agosto de 1982 se capturó en un anzuelo un Architeuthis
vivo de 10 metros de longitud y unos 220 Kg. de peso en
aguas someras (5 m) de la bahía de Radöy. El animal fue
transportado cuidadosamente a tierra por el pescador Rune
Ystebö, y Ole Brix, biólogo de la Universidad de Bergen, pudo
extraer una muestra de sangre de sus arterias branquiales.
Brix demostró, y publicó en la prestigiosa revista
Nature (1983), que la capacidad de transporte de oxígeno de
la hemocianina, que es el pigmento respiratorio de los
calamares, disminuía a un cuarto de lo normal cuando la
temperatura aumentaba de 6,4 a 15º C, lo cual sugería con
fuerza que los calamares gigantes pueden asfixiarse por
desaturación de la sangre arterial cuando la temperatura
ambiental aumenta.
Anton Bruun resumió en 1945 la situación de los
registros de Architeuthis en aguas de Islandia, donde se
habían producido seis varamientos. En su trabajo indicaba
que, a juzgar por la distribución de las localidades donde
aparecían en el Atlántico Norte, los calamares gigantes
debían ser especies propias de aguas subtropicales frías.
Para la consecución de muestras de calamares
gigantes ha sido siempre importante la aportación de los
naturalistas aficionados, y este es el caso de W.J. Clarke,
residente en Scarborough del condado de Cork en Inglaterra,
quien dedicó mucho tiempo a observar los cefalópodos de
Mar del Norte. Clarke estuvo en estrecho contacto con el
investigador del Museo Británico G.C. Robson, con quien
publicó en 1929 unas notas sobre un Architeuthis varado al
nordeste de las costas de Inglaterra, y suministró las
muestras de otro varado en enero de 1933, con las que
Robson describió una nueva especie, A. clarkei, dedicada al
celoso observador. Entre 1937 y 1957 ocurrieron algunos
varamientos en Escocia, que sirvieron a A. Stephen para
publicar varios artículos científicos, algunos de ellos
previamente presentados en las sesiones de la Real Sociedad

de Edimburgo. Semejantes tareas fueron realizadas
posteriormente por Peter R. Boyle, catedrático de Zoología
en la Universidad de Aberdeen.
Los biólogos marinos irlandeses Martin A. Collins y
Colm Lordan tuvieron la ocasión de examinar varios
calamares gigantes varados a mediados de los noventa en
las costas de Irlanda, todos ellos eran hembras excepto dos
que eran machos maduros. En sus estudios, además de
corroborar el dimorfismo sexual existente en los calamares
gigantes, de estudiar su dieta a partir de los contenidos
estomacales, y de estimar su edad en base a los incrementos
de deposición que se pueden leer en los estatolitos,
encontraron diferencias morfológicas tan significativas como
para sugerir que en el Atlántico Norte habría más de una
especie de Architeuthis.
Los registros y estudios franceses de calamares
gigantes se limitan a un trabajo de Jean Cadenat realizado
sobre un calamar cogido por el buque de arrastre Palombe de
La Rochelle (Francia) el 8 de junio de 1935 a unos 200
metros de profundidad en el Golfo de Vizcaya.
El interés de los científicos españoles por los
calamares gigantes es relativamente antiguo. Entre 1978 y
1990 llegaron congelados al puerto de Vigo cinco ejemplares,
cuatro hembras entre 220 y 38 Kg. de peso, y un macho
maduro de 41,5 Kg. provenientes de capturas accidentales
realizadas por barcos arrastreros españoles que faenaban
intensamente en aguas de Namibia en una pesquería dirigida
a la merluza,. Germán Pérez-Gándaras, Ángel Guerra y María
Teresa Fernández utilizaron este material para publicar la
primera cita sobre de la dieta de estos gigantes,
comparaciones anatómicas y morfológicas y varias
consideraciones biológicas y ecológicas de interés.
Con la desaparición de los arrastreros españoles de
esos caladeros, después de la independencia de Namibia,
desapareció también esa fuente de información. Sin
embargo, desde hace aproximadamente ocho años se abrió
la colaboración entre los miembros de CEPESMA y de
ECOBIOMAR, gracias a la cual ya se han examinado y
estudiado 18 ejemplares de Architeuthis procedentes de las

costas asturianas. Estos estudios, que han dado lugar a
varias publicaciones, han contribuido notablemente a
profundizar en el conocimiento de la biología y ecología de
los calamares gigantes. Entre otras cosas, los dos machos
hallados son los primeros capturados en la península Ibérica,
de los cuales uno mostraba esperma en sus brazos, tal como
había encontrado Kjennerud en el ejemplar varado en
Dinamarca en 1958. Estos hechos abren un interesante
debate sobre el comportamiento reproductor del calamar
gigante, en el sentido de si pueden autofecundarse
accidentalmente al copular con una hembra, o incluso si
existe o no homosexualismo entre los cefalópodos.
Por otra parte, como fruto de la colaboración entre la
productora Transglobe Films y ECOBIOMAR nació la idea de
realizar el primer documental español sobre los Architeuthis,
que se tradujo en el proyecto KRAKEN.
La disponibilidad de un material tan cuantioso y de
tanta calidad ha convertido la colección de calamares
gigantes depositada en el Aula del Mar que CEPESMA tiene
en Luarca, en la mejor dotada del mundo. De esta forma,
Asturias, junto con Nueva Zelanda, se ha convertido en un
polo de atracción para los científicos y en un lugar de
atractivo mundial en esta materia, que tanto interés
despierta entre los especialistas y el gran público.
Fig. 11. M. T. Fernández y A. Guerra. Vigo, 1987

Fig. 12. Algunos miembros del equipo de ECOBIOMAR (Vigo, 2001). De izquierda a derecha: Jaime Otero, Francisco Rocha, Ángel
Guerra, Joaquín Gracia, Ángel F. González y José Castro
Desde Luarca y Vigo se está irradiando el interés por
los calamares gigantes a toda Europa occidental. El 23 de
octubre de 2001, Ángel Guerra y Luis Laria realizaron la
primera disección de un Architeuthis en Francia. El ejemplar
disecado según una técnica desarrollada por ellos en
colaboración con el cirujano gallego Alberto Rodríguez Costa
está actualmente expuesto en el Museo del Mar de Biarritz.
En el Aula del Mar de Málaga hay conservado un
ejemplar hembra de calamar gigante que varó en una playa
de las proximidades de Fuengirola. Se trata de la primera
cita del Architeuthis para el mar Mediterráneo, que debió
ocurrir en primavera o verano de 2001. Otra hembra de 126
cm de longitud del manto apareció varada el 26 de junio de
2001, también en una playa de Fuengirola (Málaga). Ese
ejemplar fue recogido por miembros del Instituto Español de
Oceanografía y estudiado por María González y otros colegas
de esa institución. Fue disecado por miembros de
ECOBIOMAR y Oscar Soriano, el 21 del junio de 2002, en el
Museo Nacional de Ciencias Naturales de Madrid, donde está
conservado y expuesto. Precisamente fue uno de los
investigadores de ese museo (José Templado) quien nos
proporcionó información sobre un calamar gigante hallado
varado en la playa de Matalacañas (Huelva) el 2 de
septiembre de 1995. Se trataba de una hembra de 120 cm

de manto y cerca de 180 Kg de peso. Se encontró en
avanzado estado de descomposición echándose a un
vertedero de basuras cercano. Actualmente, miembros de
ECOBIOMAR y del Museo de Madrid, están desarrollando una
investigación para dilucidar el número de especies de
Architeuthis presentes en el Atlántico Norte utilizando
secuencias de ADN.
Hay dos registros más para la península Ibérica. El
primero se produjo el 24 de agosto de 2002, cuando se
encontró un ejemplar flotando entre la playa de Galé y la
laguna de Melides (38º 11’N-08º 48’W), a unos ochenta
kilómetros al sur de Lisboa. Se trataba de un macho maduro
de 60 Kg. de peso y 111 cm. de longitud del manto. El
ejemplar fue examinado por varios investigadores
portugueses. El segundo se halló varado en la playa del
Tejar, cerca de Tarifa (Cádiz), el 3 de julio de 2003. Se
trataba de una hembra inmadura, que actualmente está
depositada en el centro que el Instituto Español de
Oceanografía tiene en Málaga.
Fig. 13. Detalle de la disección de un calamar gigante en Biarritz. Luis Laria y Ángel Guerra. 2001.

Los avistamientos de calamares gigantes en aguas de
las islas Canarias, sobre todo de animales flotando en
superficie, son también frecuentes. Actualmente, un grupo
de veterinarios y biólogos canarios, encabezado por Antonio
Fernández y Luis Felipe Lópz Jurado se ha interesado por el
estudio de estos animales. Por el momento, tienen tres
cadáveres congelados para analizar y están intentando
conseguir información sobre registros históricos y lugares
donde son avistados con mayor frecuencia.
La presencia de Architeuthis en Madeira está
acreditada desde los estudios de W.J. Ress y G.E. Maul en
1956, así como por los trabajos de Malcolm R. Clarke sobre
contenidos estomacales de diferentes depredadores. Clarke
también señaló su presencia en las proximidades del
archipiélago de las Azores, desde cuyas aguas un pesquero
vigués trajo un ejemplar hembra de 80 Kg. en 2001, que
había capturado en un anzuelo de su palangre, y que midió el
biólogo marino Mario Rasero.
Las actividades de la flota pesquera de la antigua
Unión Soviética se extendieron por todo el mundo, y con
ellas la investigación relacionada con la pesca. Los
investigadores tuvieron así acceso a un gran número de
muestras de los mares de todo el globo, obteniendo una
visión muy amplia sobre los calamares gigantes. Entre los
biólogos marinos rusos dedicados a la teutología cabe
destacar a Kir N. Nesis, fallecido el 8 de enero de 2003, y a
su colega y amigo Chingis Nigmatullin, quien registró el
primer Architeuthis en aguas ecuatoriales de Atlántico.
Los calamares gigantes continuaron apareciendo en
Terranova a principios del siglo XX aunque con menor
frecuencia que en décadas anteriores. N. Frost estudió los
dos únicos Architeuthis de que se tiene constancia hasta
mediados del siglo. Por el contrario, entre 1952 y 1983 se
contabilizaron unos 17 registros, que en su mayoría fueron
estudiados por el biólogo marino canadiense Frederick Allen
Aldrich, quien realizó una extensa labor sobre la sistemática,
biología y ecología de estos grandes calamares en el
Atlántico Norte, en la que fue ayudado por su mujer

Margueritte. Ambos sugirieron que Architeuthis podría
regenerar sus brazos.
Hasta que Gilbert L. Voss, catedrático de Biología
Marina de la Universidad de Miami, examinó y describió en su
tesis doctoral dos calamares gigantes de este género
hallados en el golfo de México entre 1950 y 1956, siempre se
había pensado que Architeuthis era propio de aguas frías. El
primer registro de un juvenil de calamar gigante se debe a
dos discípulos de Voss, Clyde F. E. Roper y Richard E. Young.
Ambos ejemplares, de 57 y 45 mm de longitud de manto
respectivamente, estaban depositados en la colección de
cefalópodos de Instituto de Ciencias Marinas de la
Universidad de Miami. El primero procedía del estómago de
un ejemplar de pez sable (Alepisaurus ferox) capturado en la
Cámara de Lobos (Madeira), y el otro también era
procedente del estómago de un pez sable, pero esta vez
capturado en el Pacífico oriental cerca de las costas de Chile,
lo que constituyó el primer registro para el océano Pacífico
sudoriental.
Parece que fue por entonces cuando nació en Roper la
afición por el calamar gigante, al que dedicó un extenso
artículo, que se publicó en 1982 en la revista Scientific
American. En esa recopilación se recoge la información
proporcionada por R.B. Toll y S.C. Hess, según la cual los
machos de las especies de este género maduran muy
tempranamente, basada en la observación de un ejemplar de
menos de un metro de manto en el que se habían hallado
espermatóforos funcionales. Son numerosas las
contribuciones de Roper a la sistemática de los cefalópodos
desde su puesto de conservador de la colección de moluscos
del museo de la Smithsonian Institution en Washington D.C.
La más reciente, realizada cuando ya estaba a punto de
retirarse, en colaboración con su ayudante y amigo Michael J.
Sweeney. Entre ambos produjeron un listado de los registros
mundiales de Architeuthis.
Roper ha contribuido notablemente a la divulgación de
los conocimientos sobre Architeuthis a través de exposiciones
y conferencias en el Museo Nacional de Historia Natural de
los Estados Unidos y otros lugares del mundo. Desde

mediados de los noventa, Roper ha sido el líder e impulsor de
proyecto plurinacional en el que han colaborado científicos,
tecnólogos, expertos en filmación, barcos y submarinos con
objeto de obtener las primeras imágenes de un calamar
gigante vivo en su medio natural. Este esfuerzo ha
aglutinado científicos y técnicos de los EEUU, Canadá, Gran
Bretaña y Nueva Zelanda donde 10 instituciones, incluyendo
la Smithsonian Institution de Washington D.C, la National
Geographic Society y la National Geographic Television, han
realizado hasta la fecha cuatro expediciones en busca del
calamar gigante. Una de estas expediciones se realizó en las
Islas Azores y las restantes en el cañón de Kaikoura en
Nueva Zelanda. Sin embargo, todo este esfuerzo fue en vano
porque hasta la fecha no se han obtenido las imágenes
deseadas.
Las investigaciones sobre Architeuthis iniciadas por
Guerra y Pérez-Gándaras desde España fueron continuadas,
ya desde Sudáfrica, por Martina C. Roeleveld y Marek R.
Lipinski, quienes trabajaron fundamentalmente en aspectos
sistemáticos, morfológicos y biogeográficos, extendiendo
actualmente sus investigaciones hacia aspectos anatómicos.
Fig. 14. C.F. M. Roper, A.F. González y F. Rocha. Bayona, 1998

En las costas occidentales del Atlántico Sur los
registros de Architeuthis son escasos y recientes, habiéndo
sido estudiados principalmente por investigadores argentinos
dirigidos por Norma Brunetti y María Edith Ré.
Tampoco son frecuentes los registros en aguas
japonesas, donde, después de los estudios de comienzos de
siglo realizados por M. Sasaki, destacan los llevados a cabo
por M. Hamabe, T. Okutani, K. Tsuchiya y T. Kubodera.
Cuando R. K. Dell en 1952, resumió la situación de los
calamares gigantes en aguas de Nueva Zelanda, siguió las
observaciones que había hecho Bruun para aguas del
Atlántico Norte, e indicó que Architeuthis debería
concentrarse en dos zonas: los estrechos de Cook y Foveaux,
que se encuentran próximas a zonas oceánicas de gran
profundidad. En el trabajo de R.W Gauldie, I.F. West y E.C.
Forch se indica que los calamares gigantes recuperados en
aguas neozelandesas entre 1983 y 1994 fueron veinticuatro,
de los cuales, cinco fueron recogidos cerca del estrecho de
Foveaux y uno en las proximidades del estrecho de Cook,
mientras que el resto se repartió entre otras regiones de las
islas norte y sur así como en las islas Auckland, pero sobre
todo en el cañón submarino de Kaikoura. Esos ejemplares y
otros parecidos en aguas de Australia y Tasmania fueron
capturados principalmente de forma accidental por redes de
arrastre en pesquerías comerciales que faenaban
relativamente a gran profundidad. Gauldie y colaboradores
retomaron los estudios de George D. Jackson, C.C. Lu y
Malcolm Dunning de 1991, donde se estimaba la edad de los
Architeuthis a partir de la lectura de incrementos en sus
estatolitos, llegando a una parecida conclusión: que si los
incrementos se depositan diariamente, esos cefalópodos
alcanzan elevados pesos en muy breve tiempo, siendo muy
probablemente los animales marinos con la mayor tasa de
crecimiento de la naturaleza. Entre los calamares gigantes
hallados en Nueva Zelanda se encuentra una hembra de 21
metros de longitud total, con un peso aproximado de 275
Kg., que es el mayor registro mundial hasta la fecha. Por
otra parte, también en Nueva Zelanda, se ha cogido el único
ejemplar hembra fecundado que se conoce hasta ahora. Este

acontecimiento fue publicado por M. D. Norman y C.C Lu en
1997 en la prestigiosa revista Nature. Las paralarvas de
Architeuthis han sido también objeto de estudio en aguas de
Nueva Zelanda y Australia. Según parece, aunque su
identificación no está confirmada, C. C. Lu fue el primero en
registrar una paralarva de estos animales, que fue capturada
incidentalmente en 1986 en pescas de plancton realizadas en
aguas de Australia.
Hace apenas unos años comenzaron a aparecer,
principalmente en páginas de Internet, noticias de Steve
O´Shea sobre los calamares gigantes. Este investigador del
Instituto Nacional de Investigaciones del Agua y de la
Atmósfera de Wellington es quien más recientemente ha
tenido la oportunidad de estudiar los ejemplares de
Architeuthis capturados en aguas de Nueva Zelanda, así
como los ejemplares de la especie gigante Mesonychoteuthis
hamiltoni que aparecen con frecuencia en las redes de los
pescadores que faenan en las pesquerías desarrolladas en la
región subantártica, y cuyos mayores ejemplares alcanzan
hasta 150 Kg. de peso. O’Shea informó también de la
captura de varias paralarvas vivas, al parecer de
Architeuthis, en aguas superficiales de Nueva Zelanda hacia
finales del verano y comienzos de la primavera de 2002, las
cuales pudo mantener en cautividad durante 75 días. Bajo la
dirección de este científico neozelandés se han organizado
dos campañas internacionales de prospección en 2004 para
tratar de capturar nuevas paralarvas, así como filmar al
Kraken en su medio natural, pero parece que sin éxito.
Una de las figuras más emblemáticas en el campo de
la teutología es sin duda el inglés Malcolm R. Clarke,
miembro de la Royal Society. A él se debe la creación del
Cephalopod Internacional Advisory Council (CIAC), que desde
principios de los ochenta ha servido de marco para la
organización de numerosos congresos, simposios y reuniones
sobre diferentes aspectos de la sistemática, biología, ecología
y explotación de los cefalópodos.

Fig. 15. Banyuls sur Mer (Francia), 1985. Primer Comité de
Dirección del Cephalopod International Advisory Council
(CIAC). De pié y de izquierda a derecha: R. Toll, N.A. Voss,
G.L. Voss, J.B. Messenger, M. Nixon, E. Hochberg, R. Young
y M. R. Clarke. Sentados: S.v. Bolezky, K. Mangold, C.F.E.
Roper, A. Guerra, C.C. Lu y J. Wormuth
Su extensa revisión de la ecología de los cefalópodos
oceánicos publicada en 1966 es un punto de partida para el
estudio de numerosas familias, incluyendo la de los
Architeuthis. Sin embargo, fueron sus profundos estudios
sobre la dieta de los cachalotes y otros depredadores de los
cefalópodos en aguas de todo el mundo los que
contribuyeron notablemente a conocer la posición de los
cefalópodos, y entre ellos los calamares gigantes, en las
cadenas tróficas de los ecosistemas marinos. Para identificar
los restos de calamares, pulpos y sepias en los contenidos
estomacales de sus depredadores, Clarke estudió e impulsó
los trabajos sobre sus picos y estatolitos.
La cooperación internacional y la aplicación de
técnicas modernas van incrementando los conocimientos
sobre los calamares gigantes, como lo demuestra el reciente

trabajo de N.H. Landman y colaboradores, en el que se han
tratado de dilucidar varios aspectos referentes al hábitat y la
edad de Architeuthis sanctipali a partir de análisis de isótopos
en los estatolitos. Sus resultados corroboran que estos
cefalópodos viven preferentemente en aguas de entre 10,5 y
12,9ºC de temperatura, correspondiendo a profundidades
comprendidas entre 125 y 500 metros de profundidad. Sin
embargo, otros de sus datos son contradictorios con los
previamente conocidos, ya que sus estimaciones a partir de
los isótopos sugieren que la edad de ejemplares de
Architeuthis con 150 Kg. de peso promedio sería de unos 14
años aproximadamente, lo cual contradice todo lo publicado
hasta la fecha. Un nuevo e interesante tema de debate se
abre, por tanto, en el estudio de un aspecto básico de la
historia natural de los todavía tan ignotos calamares
gigantes.

Capítulo 2
LAS ESPECIES DE ARCHITEUTHIS QUE EXISTEN EN EL MUNDO
Nombres comunes y científicos
Una visita a diferentes puertos pesqueros cuando en
sus lonjas están expuestos los productos que los pescadores
extraen del mar, nos proporcionaría, al menos, dos
observaciones: una, que muchas especies se repiten, o que
hay muchas especies parecidas, y otra, que los habitantes de
diferentes localidades dan nombres distintos a la misma
especie, o bien, que un mismo nombre sirve para denominar
a especies diferentes. Esto último se debe a que los
habitantes de una región dan un “nombre común” a una
determinada especie asociándola con alguna característica
propia. Pero ese nombre común puede cambiar entre puertos
o de una zona a otra, ya que esa especie se asocia con
características que son allí más importantes o llamativas.
Esta confusión con los nombres comunes aumenta si
comparamos las denominaciones que recibe una misma
especie en varios países. Por ejemplo, hay una especie de
cefalópodo de gran tamaño que habita en el océano Pacífico
frente a la costa de América, entre México y Chile, que se
captura con frecuencia y recibe principalmente dos nombres
vernáculos: jibia y calamar humbolt. Pero esta especie se
parece muy poco a la sepia que habita en la costa española
del Mediterráneo y que en la zona meridional recibe el
nombre de jibia. La jibia del Pacífico pertenece a la misma
familia que las denominadas comúnmente voladores, potas y
potarros en la costa atlántica de la península Ibérica,
mientras que lo que en el Mediterráneo sur y golfo de Cádiz
se llama jibia, en Galicia se denomina choco y en otras
partes de España sepia.
A la gran cantidad y variedad de nombres comunes
que puede recibir una especie en un mismo idioma, se añade
que en cada idioma una misma especie puede recibir varios

nombres comunes, lo que aumenta todavía más la confusión
a la hora de tratar sobre una misma especie. De esta forma,
los científicos siempre encontraron difícil utilizar los nombres
comunes para designar una especie de manera unívoca. Así,
surge la necesidad de que cada especie estudiada o descrita
por la ciencia tenga un nombre científico, el cual debe ser
único, característico y universal, en el sentido de quesea cual
sea la nacionalidad del investigador o el idioma en que se
hable el nombre no cambie.
Al principio el nombre científico consistió en una
descripción abreviada que resaltara las características más
conspicuas de cada especie. Por ello, los nombres científicos
utilizados hasta mediados del siglo XVIII tendían a ser
largos, incómodos, inestables y, a veces, contradictorios. A
medida que se fueron identificando nuevas especies, los
nombres se fueron alargando y resultaban cada vez más
complejos e inmanejables. Aunque existieron varios intentos
por encontrar un sistema de nomenclatura adecuado para
denominar a las especies, el que finalmente fue aceptado por
la comunidad científica fue el sistema binomial de Linneo,
establecido en su obra Systema Naturae (1758). Este
sistema asigna a cada especie un nombre compuesto de dos
palabras, ambas escritas en cursiva o subrayadas. El nombre
científico se escribe en latín, lo cual es lógico ya que ese era
el idioma científico de la época; además, de esta forma, y
por ser el latín una lengua muerta, sea cual sea el idioma el
nombre científico no varía pudiendo utilizarse
universalmente. La primera palabra del binomio, cuya
primera letra siempre se escribe con mayúscula, corresponde
al nombre del género, es decir, al grupo o taxón de
características similares y bien definidas dentro del cual se
pueden diferenciar subgrupos, también claramente diferentes
entre sí, que están formados por individuos similares que
pueden cruzarse entre sí y producir descendencia fértil. El
segundo término, siempre escrito en minúsculas, es el
nombre específico que, normalmente, indica alguna
característica propia de la especie, aunque también se usan
patronímicos, gentilicios y otros tipos de denominaciones,
pero siempre latinizados. Este sistema, además de evitar la

confusión producida por los nombres vulgares que recibe una
determinada especie, permite el fácil y cómodo
entendimiento entre científicos de todo el mundo.
El nombre científico no se da al azar, sino que se
construye de tal forma que describe alguna característica
propia de la especie o, en menor medida, se dedica a una
persona o se refiere a un lugar determinado. También hay
nombres relacionados con la literatura, historia o mitologías
romana y griega, aunque éstos, generalmente, se pueden
asociar también a alguna característica propia del personaje
a que se refiere el nombre. Así, el nombre del grupo a que
pertenecen los calamares gigantes, Cephalopoda, indica la
principal característica del grupo (del griego “kephale”:
cabeza y “pous”: pies; es decir, con pies en la cabeza). Por
otra parte, Architeuthis significa “calamar antiguo” o
“calamar primigenio” (del griego “Archi”: ser el primero y
“teuthis”: calamar).
Por otra parte, los calamares comunes pertenecen al
género Loligo, dentro del cual hay cerca de cien especies,
como por ejemplo el calamar común europeo Loligo vulgaris
y el calamar de California Loligo opalescens. Sin embargo, no
muchos años después de comenzar a usarse este sistema, se
empezaron a encontrar problemas de diferente índole. El más
importante era que dos expertos diferentes hubiesen
denominado de manera diversa a la misma especie. Esta
dualidad se resolvió incorporando al nombre específico el del
autor y también el año de su publicación. Así, Loligo vulgaris
Lamarck, 1798 indica que esa especie fue nombrada por
primera vez por ese autor y en ese año. ¿Qué hacer cuando
se advertía que dos autores habían nombrado a una misma
especie con nombre diferente? Pues se adoptaba como válido
aquel con mayor antigüedad, siempre que se hubiese
publicado en un medio que ofreciese suficientes garantías de
haber podido ser conocido ampliamente, es decir, en una
revista conocida y de publicación periódica, desechándose
aquellos publicados en medios prácticamente inaccesibles.
Otra de las condiciones para la validez del nombre de una
especie es que ésta hubiese sido definida con claridad.
Entonces, el nombre más moderno pasaba a ser un sinónimo

del más antiguo. Así por ejemplo, Ommastrephes bartramii
(LeSueur, 1821) corresponde a la pota cárdena, y un
sinónimo de éste es Ommastrephes caroli Furtado, 1887.
¿Por qué está entre paréntesis el nombre y el año del autor
de esta especie? Esto se debe a que LeSueur la denominó
originalmente Loligo bartramii, siendo colocada
correctamente dentro del orden Ommastrephes por
d’Orbigny en 1885. Estos son sólo unos ejemplos de la
enorme casuística que genera la nomenclatura biológica, tan
compleja que ha precisado de la creación de un Código
Internacional de Nomenclatura Zoológica, como existen otros
para los demás reinos, y de un Comité Internacional de
expertos que resuelven los casos conflictivos y mantienen al
día dicho código.
Otro de los problemas con que se encuentran los
taxónomos es que muchas veces hallan diferencias
morfológicas, o de otro tipo, entre individuos muy parecidos
pero que no pueden ser considerados especies diferentes.
Entonces se suele optar por definir un subgrupo dentro de la
especie que se denomina subespecie. Así, el calamar común
Loligo vulgaris Lamarck, 1798 se divide en dos subespecies:
el propio de las costas europeas y del norte de África Loligo
vulgaris vulgaris Lamarck, 1798 y el correspondiente a las
costas sudafricanas Loligo vulgaris reynaudii d’Orbigny,
1845.
Algunas de las precauciones que se deben tomar para
que cualquier especie quede bien fijada y correctamente
definida, es que en su descripción se incluya un ejemplar que
se considere su tipo (holotipo), además de otros a los que se
denomina paratipos. También que se defina con claridad la
localidad donde se obtuvo el holotipo (localidad tipo). Por
último, que todo el material tipo quede almacenado en un
lugar adecuado donde pueda consultarse. Por desgracia, y
sobre todo en especies muy antiguas, estos requisitos no se
observan siempre, y a veces hay que considerar una especie
inválida por incumplir alguna o varias de estas condiciones.
En el trabajo Historical review of the genus
Architeuthis, que el zoólogo alemán George Pfeffer publicó en
1912, dentro de la magna obra donde se relatan los

descubrimientos de The Plankton-Expedition, se trató por
primera vez de clarificar la nomenclatura hasta entonces
existente referente a los calamares gigantes. En ese trabajo,
Pffefer señala que Architeuthis monachus de Steenstrup es
un nomen nudum, es decir un nombre vacío, y por lo tanto
que esa denominación específica es inválida y no merece
mantenerse en la nomenclatura zoológica. Pfeffer llegó a esa
conclusión luego de percatarse de que la denominación dada
por Steenstrup carece de una diagnosis o descripción
suficiente de la especie y que no hay material tipo de la
misma depositado en alguna parte.
Nuestro planeta está habitado por varios millones de
formas de vida y poder nombrarlas de una forma precisa,
utilizable por todo el mundo, es algo muy importante. Sin
embargo, dar un nombre científico a una especie no lo es
todo, sino que éste debe permitir clasificarla junto con otras
especies, agrupando aquellas que estén emparentadas. Así,
especies muy parecidas pueden tener el mismo nombre
genérico y un nombre específico distinto. Por ejemplo, las
especies de calamares gigantes propiamente dichas
pertenecen todas al género Architeuthis, pero hay descritas
varias especies: Architeuthis dux, Architeuthis martensi,
Architeuthis sanctipauli, etc. Es más, una buena clasificación
permite proporcionar una idea sobre las relaciones entre los
organismos y las características del grupo al que pertenecen,
así como reconstruir sus relaciones filogenéticas y, en cierta
medida, la historia evolutiva del grupo y la especie. En la
clasificación científica, cada organismo está asignado a una
categoría superior, que le agrupa con organismos
semejantes, y estas a su vez a otras de superior rango. La
mayor categoría es el Reino y dentro de ella se incluyen las
categorías inferiores. De esta manera, los calamares gigantes
tendrían la siguiente clasificación:
Reino: Animalia
Filo: Mollusca
Clase: Cephalopoda
Subclase: Coleoidea
Superorden: Decabrachia

Orden: Teuthida
Suborden: Oegopsina
Familia: Architeuthidae
Genero: Architeuthis
El concepto de especie
A uno le podría parecer que, ya que a todos los
organismos se les puede clasificar dentro de una especie, el
propio concepto de especie debería ser algo muy claro y
preciso. Sin embargo, la realidad dista mucho de ser así. A lo
largo de la historia se han ido dando diferentes criterios para
definir lo que es una especie, criterios que se han ido
perfilando a medida que el progreso científico ha facilitado
nuevos conceptos o posibilidades que era imposible
considerar con anterioridad.
El primer concepto de especie fue el morfológico. Este
concepto postula que los ejemplares de una misma especie
tienen la misma forma; por ello las especies se pueden
definir en base a caracteres formales, que representarían la
esencia de cada especie. De esta manera, todo ser vivo que
concuerde con la descripción original pertenecería a la misma
especie, siendo de distintas especies aquellos que difieran. La
aplicación estricta de este concepto implica que la variación
entre ejemplares de una misma especie es mínima, por lo
que la existencia de variaciones morfológicas entre
ejemplares de una misma especie es difícilmente explicable a
menos que sean especies diferentes. Este concepto
morfológico se ha enriquecido con el concepto biológico de
especie, que la define como el “conjunto de poblaciones que,
real y potencialmente, pueden reproducirse entre sí, pero
que están aisladas de otros grupos similares”. Para ello,
posteriormente, se desarrolló el concepto de reconocimiento
de especie, en el cual se la define como el “conjunto de
individuos y poblaciones que comparten un mismo sistema
de fertilización”. Finalmente, también ha aparecido el
concepto evolutivo de especie, donde se la define como el

conjunto de poblaciones que comparten un destino evolutivo
común a lo largo del tiempo.
En fin, para no complicarnos diremos que una especie
es un conjunto de individuos con características similares
entre sí, que poseen una serie de caracteres propios que les
diferencian de otras especies y pueden entrecruzarse
generando individuos fértiles semejantes a sus progenitores.
¿Cuales son los caracteres que nos permiten distinguir
una especie de otra? En general son características
morfológicas muy determinadas, que pueden ser observadas
en todas las especies de una misma familia, género o grupo
de especies, pero que son diferentes entre si. Estas
diferencias y similitudes permiten agrupar no sólo especies
sino grandes grupos de especies dentro de clases, órdenes y
familias. Por ejemplo, todos los cefalópodos del orden
Teuthida poseen, entre otras características, diez apéndices
alrededor de su boca, ocho brazos y dos tentáculos. Pero a
su vez se les puede subdividir en subórdenes Myopsina y
Oegopsina según tengan o no sus ojos cubiertos por una
membrana cornea. A su vez, dos especies pertenecientes a
un mismo género pueden diferir en unos pocos caracteres,
ya sean morfológicos (forma) o merísticos (número). Por
ejemplo, una de las diferencias entre los calamares Loligo
vulgaris y L. forbesi, que habitan las aguas de la península
Ibérica, es que en el primero las dos filas internas de las 4
que posee en la maza tentacular tienen ventosas de tamaño
mucho mayor, mientras que en la segunda especie, las 4
filas de ventosas son todas de tamaño semejante.
El género Architeuthis
Como veremos más adelante, no todos los
cefalópodos de gran tamaño son calamares gigantes. Los
cefalópodos pertenecientes al género Architeuthis poseen
una serie de caracteres que los diferencian de otras especies
de gran tamaño. Son animales con tentáculos muy largos
(2,5 a 6 veces la longitud de su manto), cuyas mazas
tentaculares tienen cuatro filas de ventosas, de las cuales las
dos filas centrales poseen ventosas de mayor tamaño que las

marginales. Una característica propia del género es que a lo
largo de todo el tentáculo se aprecian, espaciadas entre si,
dos filas longitudinales de ventosas y almohadillas dispuestas
alternativamente. Aparte de esto, los Architeuthis se
caracterizan por poseer un cartílago de cierre del sifón con el
manto sencillo y recto. Esta estructura es como un broche
que une el manto con el sifón del animal, que se abre cuando
el agua penetra al interior del manto para irrigar las
branquias y se cierra cuando es expulsada por la contracción
del manto, obligándola a salir por la abertura sifonal y, así,
impulsando al animal.
Otros caracteres que permiten identificar un calamar
gigante son el carecer de órganos luminosos o fotóforos y la
forma característica de sus pequeñas aletas que ocupan
menos del 40% de la longitud del manto. En el caso de los
machos, sus dos brazos ventrales (IV) están hectocotilizados,
es decir, modificados para la cópula con la hembra.
Fig. 16. Architeuthis dux hembra capturado en el caladero de Carrandi, frente a Asturias, en 2001. Detalle del tentáculo donde se
aprecian las dos filas de ventosas y almohadillas alternadas entre si.

Fig. 17. Los cartílagos de cierre del manto con el sifón. En Architeuthis se caracteriza por ser simple y recto, a diferencia de lo
observado en otras especies.
No es un sólo carácter morfológico el que nos
permitirá diferenciar un Architeuthis de otros cefalópodos de
gran tamaño, sino el conjunto de estos caracteres.
Los Architeuthis son raramente capturados y cuando
aparecen suelen encontrarse en muy mal estado, por lo que
pocos ejemplares han logrado conservarse enteros en el
mundo. Además, muchos de los caracteres de valor
taxonómico que poseen se deterioran rápidamente o no se
conservan lo suficiente como para ser estudiados. Por
ejemplo, la coloración rojiza y las características de su
epidermis raras veces se aprecian debido a que la piel se
rompe con el roce con la arena o piedras de la costa o las
redes. Esto explica en gran parte las carencias que existen
en el estudio del calamar gigante. Así, actualmente hay
registradas unas 20 especies nominales, que están descritas
de forma inadecuada o insuficiente, por lo cual se desconoce
su número real. Al carecerse del suficiente número de
ejemplares para estudiar, o de descripciones adecuadas que
comparar, la situación es confusa. De todas formas, lo más

probable es que el número real de especies existente sea
menor, como se verá en el apartado dedicado a los nombres
del calamar gigante. Como la situación todavía es muy
confusa, la mayoría de los registros de calamares gigantes
figuran como Architeuthis sp., lo que significa que se
desconoce a que especie pertenece el ejemplar o los
individuos examinados.
Mucho queda por hacer en el caso de los calamares
gigantes. En primer lugar, clarificar cuantas especies existen
realmente, aspecto en el que las nuevas tecnologías como
técnicas de taxonomía numérica y los análisis genéticos
tendrán bastante que decir, pero ninguna de ellas estaban ni
tan siquiera esbozada cuando Steenstrup erigió el género
que engloba al más famoso de los calamares gigantes. Pero
¿quién era Steenstrup?
Johan Jappetus Steentrup
El naturalista Johan Jappetus Steenstrup nació en
1813 en la provincia de Thy, al noroeste de Dinamarca. Ya
desde pequeño, e influido por uno de sus tíos que era un
botánico aficionado, se interesó por las plantas, además de
complementar esta inquietud con el interés por la diversidad
faunística de la región. A los trece años se trasladó a
Aalborg, donde se incorporó a la escuela de gramática.
Durante su estancia en esta cuidad, Steenstrup convivió con
la familia Kaarsberg cuya hija Ida sería posteriormente su
esposa en 1841. En Aalborg colaboró con algunos amigos
naturalistas, familiarizándose con la bibliografía de la época,
entre la que se podrían citar el Sistema Naturae o la Flora
Danica. En 1832 comenzó sus estudios médicos en
Copenhague y entabló una fructífera relación con profesores
de Historia Natural, especialmente con el zoólogo J.H.
Reinhardt, e incluso con el famoso geólogo J.G.
Forchhammer. Un año más tarde tuvo que retornar a su
casa, fundamentalmente por motivos económicos,
permaneciendo allí durante dieciocho meses para volver a la
universidad en 1835. Durante los diez años siguientes
escribió numerosos artículos entre los que se encuentra su

interesante estudio sobre los pantanos daneses. En 1841
ejerció como conferenciante en mineralogía y botánica en la
Universidad de Soro para, posteriormente, llegar a ser
profesor de Zoología en la Universidad de Copenhague. J.J.
Steenstrup ejerció como profesor de Zoología durante casi
cuarenta años y se le puede considerar como un naturalista
en toda la extensión de la palabra, abarcando casi todos los
campos de la historia natural. De carácter extrovertido,
Steenstrup era una persona impulsiva e inquieta, lo que ha
quedado reflejado en algunos de sus artículos donde
arremetía contra aquellos investigadores que no apreciaban
los conceptos que para él eran evidentes. En Copenhague
trabajó hasta su retiro en 1885, muriendo en esta misma
cuidad en 1897.
Fig. 18. Retrato de J.J. Steenstrup. Litografía de 1855. Biblioteca Real. Copenhague.

Su labor científica sobre los calamares gigantes no
comenzó hasta 1847, cuando abordó el tema buscando las
referencias escritas que se poseían hasta esa época. Ese
mismo año impartió una conferencia en la Sociedad
Escandinava de Naturalistas, que pasó inadvertida. En ese
estudio se analizaban los varamientos de calamares gigantes,
acontecidos en Ørensud en 1545, en una playa de Islandia en
1639 y el hallazgo de un ejemplar de 12 metros flotando en
las proximidades de Aalbaekstrand (Islandia) en 1790. En
1853, después que un pescador capturase y trocease un
calamar gigante al noroeste de Dinamarca, Steenstrup
recuperó y estudió el pico, publicando sus conclusiones en
1855. A partir de esas mandíbulas y de algunas
características descritas referentes a los ejemplares
anteriores analizados en la bibliografía, Steenstrup denominó
a los calamares gigantes como Architeuthis en 1857,
erigiendo así un nuevo género dentro de la clase de los
cefalópodos.
A pesar de que los cefalópodos sólo constituyeron uno
de los numerosos campos de interés que cultivó Steenstrup
en sus trabajos zoológicos, se pueda afirmar que fue en este
grupo en el cual profundizó más y mejor. Este naturalista fue
pionero en el estudio de estos fascinantes moluscos durante
los años críticos, cuando muchas de las especies oceánicas
estaban siendo descubiertas y descritas por extraordinarios
investigadores, entre los que cabe resaltar a Vérany y Verrill.
Los teutólogos de esta época habían sido estimulados por la
anterior aparición del ingente trabajo de Férussac y
d’Orbigny “Histoire naturelle générale et particulaire des
Céphalopodes acétabuliferes vivant et fossiles”. A excepción
de Steenstrup, que publicó en danés, los demás
investigadores publicaron sus artículos en idiomas
comprensibles para un número elevado de lectores,
especialmente en inglés y alemán. Sin embargo, nadie duda
que Steenstrup realizó estudios muy importantes sobre
taxonomía y anatomía de cefalópodos, quizá más completos
que sus contemporáneos. Su artículo sobre la formación del
hectocotilo en Argonauta y Tremoctopus es un ejemplo de la
extraordinaria habilidad que desplegaba cuando lograba

centrarse en este tipo de trabajos. Algunos artículos sobre
especies de calamares oceánicos, especialmente sobre las
potas de la familia Ommastrephidae y los calamares gigantes
(Architeuthidae) reflejan fielmente el sello de un genio, quien
asimiló y utilizó con total precisión el concepto de especie
tipo, que es clave en la taxonomía moderna. Por otra parte,
aunque este danés conoció las obras de Darwin nunca aceptó
el concepto darwiniano de evolución.
Afortunadamente para aquellos que no son capaces de
comprender el danés, las obras de Steenstrup referidas a los
cefalópodos fueron traducidas al inglés por Volsoe, Knudsen
y Rees y publicadas en 1962. Existe también una traducción
muy cuidada de los artículos de Steenstrup relativos al
“monje marino” realizada por Roeleveld y Knudsen en 1980.
La historia de los nombres del calamar gigante
La historia del descubrimiento de diferentes especies
pertenecientes a un mismo género se puede seguir a través
de los nombres que se han ido dando a los ejemplares
estudiados. A continuación se expone la correspondiente a
los calamares del género Architeuthis.
En la reunión de la Sociedad Escandinava de
Naturalistas celebrada en Cristiana en 1856, Steenstrup
presentó una comunicación sobre un gran cefalópodo que
había varado en las costas danesas en 1853, y basándose en
sus mandíbulas, erigió la especie Architeuthis monachus. En
esa misma reunión se refirió a otro ejemplar capturado por el
capitán Hygom al norte de Bahamas, en el Atlántico
occidental (31ºN – 76ºW), que denominó Architeuthis dux. El
material tipo de ambas especies está depositado en el Museo
de Zoología de la Universidad de Copenhague. Sin embargo,
como posteriormente señaló Pfeffer (1912), Steenstrup
únicamente presentó en aquella ocasión un dibujo de las
mandíbulas de la primera especie, no ofreciendo una
auténtica descripción y diagnosis de la misma, por lo que A.
monachus, o monacus, que de las dos formas está escrito, es
un nombre vacío, como ya se indicó. Aparentemente,
Steenstrup tenía un texto y unos dibujos preparados para ser

publicados, junto con las ideas surgidas en la discusión que
se planteó en la sesión de la Sociedad, pero este trabajo no
se publicó en vida de Steenstrup sino en 1898, cuando ya
había fallecido. Lo que ocurrió entre ambas fechas, es que, al
menos desde 1860, una parte del texto y las láminas con las
figuras estuvo en manos de varios teutólogos como Karting,
Gervais y Verrill, que lo utilizaron y lo mencionaron en sus
propios estudios sobre calamares gigantes. Incluso dos de
ellos (Gervais y Verrill) pudieron discutirlo personalmente con
Steenstrup a través de la correspondencia que mantuvieron
entre sí. Por este motivo, los comentarios no publicados del
autor danés ocuparon un lugar permanente en la bibliografía
sobre Architeuthis, de parecida forma a como si hubiesen
sido publicados. Desde un punto de vista nomenclatural, por
lo tanto, ambas especies no eran válidas antes de ser
publicadas en 1860. Sin embargo, ignorar que ya desde 1857
eran conocidas porque únicamente estaban citadas en una
comunicación oral y en correspondencia privada, era faltar a
la verdad histórica, también a la justicia con Steenstrup y al
principio de la regla de prioridad, por ello se ha convenido en
considerar que A. dux es una especie válida y la especie tipo
del género Architeuthis, descrita por Steenstrup en 1857 en
la publicación Forhandlinger ved de Skandineviske
Naturforskeres, y de la que existe material tipo en el Museo
de Copenhague. Sin embargo, A. monachus fue considerada
inválida por la razón apuntada anteriormente. Por motivos
parecidos, se aceptó que el género Architeuthis había sido
erigido por Steenstrup también en 1857 y en la misma
publicación. La familia Architeuthidae fue, sin embargo,
creada por Pffefer en 1900.
Por otra parte, según señala Verrill en una nota a pie
de la página 238 de su publicación de 1881 en las actas de la
Academia de Ciencias de Conneticut, la especie Architeuthis
titan, tercera de las descritas por Steenstrup, debe
considerarse un nombre vacio.
Los naturalistas Crosse y Fischer dieron el nombre de
Loligo bouyeri en 1862 a un ejemplar hallado en las
proximidades de Tenerife en las islas Canarias, examinado
por Gervais en 1875, quien indicó que posiblemente se

trataba de un Architeuthis en la página 93 del cuarto
volumen del Journal de Zoologie. Sin embargo, como ni la
descripción es completa, ni hay material de este ejemplar
depositado en ningún sitio, esta especie debe considerarse
inválida.
Megaloteuthis harveyi fue la especie erigida por Kent
en 1874 a partir de un ejemplar hallado en la bahía de
Concepción de Terranova en 1873. El autor indica que hay
material tipo depositado en el Peabody Museum de la
Universidad de Yale con el número de registro 12600Y, pero
no se ha encontrado. Desde luego, si se tratase de un
calamar gigante, el género sería incorrecto y debería
cambiarse por Architeuthis, pero al no existir material de
referencia esta especie debe considerarse inválida por ser un
nombre vacío. El género Megaloteuthis debe ser por tanto
considerado como una sinonimia de Architeuthis.
Loligo hartingii es otro de los nombres específicos
dado por Verrill en 1875 a un probable calamar gigante. Este
mismo autor lo cambió por el de Architeuthis hartingii en
1880. Sin embargo, la erección de esa especie no lleva
aparejada la designación de una localidad tipo y tampoco hay
material tipo depositado, aunque el autor señala que lo dejó
en la Universidad de Utrecht, donde no se ha encontrado.
Estamos, pues, en presencia de otro nombre vacio, como lo
es también, por razones similares, Architeuthis mouchezi,
erigida por Vélain en 1875 a partir de un ejemplar
procedente de la isla de Saint Paul (Océano Índico).
Architeuthis princeps se debe también a Verrill, quien
la erigió en 1875 a partir de los restos hallados en el
contenido estomacal de un cachalote. El autor define como
localidad tipo los Grand Banks y el Atlántico Norte, lo cual es
muy vago. Por otra parte, el posible material depositado en
el Museo Nacional de Historia Natural de los Estados Unidos
no se ha encontrado hasta ahora. Por todo ello, esta especie
debe también considerarse inválida.
A partir de un ejemplar hallado en el condado de
Kerry en Irlanda, More erigió la especie Dinoteuthis
proboscideus en 1875 (el nombre significa calamar gigante
con una larga probóscide o nariz). Sin embargo, y aún

cambiando la especie de género, como no se depositó
material tipo en ningún lugar, esa especie debe considerarse
asimismo inválida. Dinoteuthis deviene por lo tanto en
sinónimo de Architeuthis.
De nuevo se debe a Vélain otra especie de calamar
gigante, que denominó en 1877 Mouchezis sanctipauli. El
material tipo, un ejemplar capturado en 1875, está
depositado en el Museo Nacional de Nueva Zelanda con el
número de registro MNHN 3-2-658+9, siendo la localidad
tipo de esta especie Saint Paul Island (Océano Índico).
Modificando el nombre genérico no exite motivo alguno para
considerar que Architeuthis sanctipauli (Vélain, 1877) sea
una especie inválida. El género Mouchezis debe ser
considerado, sin embargo, sinónimo de Architeuthis.
La última aportación de Verrill a este elenco de
especies válidas y nominales es Architeuthis megaptera, que
fue descrita en 1878 a partir de un ejemplar de la localidad
tipo de Nova Scotia, Canada, depositado en el Museo
Nacional de Ciencias de Canadá y registrado como NSMC
1870–Z-2.
Architeuthis verrilli fue erigida por Kirk en 1882 en
honor a Verrill. La localidad tipo de este ejemplar es el
estrecho de Cook en Nueva Zelanda, y el autor indica que el
material está depositado en el Museo Nacional de ese país,
aunque no proporciona número de registro. Steenstrupia
stockii Kirk, 1882 (un Architeuthis según Pfeffer en 1912) es
otra denominación para un ejemplar también del estrecho de
Cook, depositado con el registro NMNZ M.125405 +
M.125403 en el citado Museo.
Architeuthis longimanus se debe también a Kirk, quien
la erigió en 1888 a partir de material recolectado en la bahía
de Lyall en Nueva Zelanda en 1887. Parece ser que el autor
depositó ese material en el Museo Nacional de aquel país,
material que se ha perdido según comunicó Marshall en
1996.
El primer ejemplar procedente de Yedo, una lonja de
pescados japonesa, fue descrito por Hilgendorf en 1880 como
Megateuthis martensii. Ese material está depositado en las
colecciones del Museo de Zoología de la Universidad de

Hamburgo con el registro ZMB Moll. 34716 + 38980. Se trata
de un calamar gigante, por ello Architeuthis martensii
(Hilgendorf, 1880) debe mantenerse como una especie
válida.
La actividad de los naturalistas ingleses en relación
con los calamares gigantes se inicia con Owen, quien erigió la
especie Plectoteuthis grandis en 1881, sin señalar localidad
tipo. Owen encontró un brazo de más de dos metros y medio
de longitud en el Museo Británico de Historia Natural y,
después de describirlo, dejó ese material depositado allí,
pero sin número de registro, ni indicación sobre su
procedencia, quizás porque originalmente ya carecía de ella.
El nombre del género deriva del griego plectos y hace
referencia a las laminillas carnosas conspicuas que hay entre
las filas de ventosas, lo cual es bastante típico de un brazo
de Architeuthis. Posteriormente, Robson erigió Architeuthis
kirkii en 1887 a partir de un ejemplar capturado en Cape
Campbell (Nueva Zelanda), que está depositado en el Museo
Nacional de Nueva Zelanda con el número de registro NMNZ
M.125404. Esta especie se considera válida. Robson
describió y dió nombre a un ejemplar de calamar gigante
encontrado varado en Yorkshire (Inglaterra). La rádula y el
pico de Architeuthis clarkei Robson, 1933 están depositado
en el Museo Británico de Historia Natural con el registro
BMNH 1933.1.30.5 + 1926.3.31.24.
En el Museo Oceanográfico de Mónaco hay
depositados, sin número de registro, unos restos de un gran
calamar hallados en el estómago de un cachalote capturado
en las islas Azores. Estos restos sirvieron a Joubin para erigir
la especie Dubioteuthis physeteris en 1900. Tanto la
descripción como el material tipo son muy pobres y esta
especie, sobre cuya ubicación en un género nuevo planteó
dudas al propio autor, debe considerarse inválida.
Architeuthis japonica Pfeffer, 1912 es el nombre dado
a un ejemplar procedente de la bahía de Tokio, pero del cual
parece no existir material tipo en ningún museo del mundo.
La última de las especies de este género de la que se
tiene constancia es Architeuthis nawaji, que fue erigida por
Cadenat en 1935 a partir del único ejemplar encontrado

antes de la Segunda Guerra Mundial en la zona francesa el
Golfo de Vizcaya. No consta que se dejase material tipo
depositado en alguna colección de un museo, y por otra
parte, la descripción es bastante mediocre, lo que invalida
esta especie.
La historia hasta aquí descrita a partir de los nombres
que recibieron varios ejemplares y restos de calamares
gigantes, bien varados o hallados en colecciones de museos,
ofrece varias enseñanzas útiles y algunas incógnitas.
La primera de las enseñanzas es que no se debe a
proceder a erigir una nueva especie zoológica si no se cuenta
con suficiente y adecuado material tipo. Luego, que las
descripciones deben ser todo lo más completas y claras
posible, procurando cerciorarse, a partir de la comparación
del material que se está examinando con otros depositados
en las colecciones de los museos, de que lo hallado no
coincida con algo ya descrito anteriormente. De ahí la
importancia de depositar y conservar adecuadamente el
material tipo. También, que en la erección de una nueva
especie la localidad tipo debe ser indicada con la mayor
precisión posible. Por último, que la taxonomía de
Architeuthis es muy confusa.
Las incógnitas que se platean se refieren, en primer
término, a cuantos géneros de calamares gigantes existen.
Actualmente, parece existir un consenso general sobre la
existencia de un único género: Architeuthis. Este género
engloba adecuadamente las características propias más
relevantes de cuanto ejemplar o resto de calamar gigante se
ha encontrado. Asimismo, es claramente diferente de otros
géneros de la clase Cephalopoda con especies que alcanzan
también grandes dimensiones, tal y como se verá en el
capítulo sexto de este libro. Según esto, los géneros
Megaloteuthis, Dinoteuthis, Mouchezis, Steenstrupia,
Megateuthis, Plectoteuthis y Dubioteuthis deben considerarse
sinónimos de Architeuthis.
La siguiente incógnita se refiere a cual es el número
real de especies de Architeuthis que existen en el mundo.
Hasta la fecha, hay 20 especies nominales descritas, algunas
de las cuales son claramente inválidas, pero entre las

validamente erigidas ¿cuántas son verdaderas especies?
Sobre este particular no existe un consenso general entre los
especialistas, y la pregunta estará sin una respuesta clara
hasta que se consiga realizar un extenso y profundo estudio
con material y técnicas apropiadas, probablemente
genéticas. Sin embargo, la opinión más generalizada es que
parece existir una única especie, que por prioridad sería
Architeuthis dux Steenstrup, 1857, compuesta por diferentes
subespecies según su localidad geográfica: Architeuthis dux
dux Steenstrup, 1857 en el Atlántico, A. dux martensii
(Hilgendrof, 1880) en el Pacífico Norte y A. dux sanctipauli
(Vélain, 1877) en el hemisferio Sur. Otros especialistas, sin
embargo, consideran que estas tres subespecies deben
elevarse al rango de auténticas especies. Y por último, otros
expertos opinan que habría al menos cinco o seis especies
diferentes: A. dux en el Atlántico Norte; A. sanctipauli propia
de una extensa área que comprende la región subantártica y
antártica de los océanos Atlántico, Índico y Pacífico; A. kirkii
Robson, 1887 en aguas de Nueva Zelanda; A. martensii o A.
japonica Pfeffer, 1912 (aquí habría que definir cual de ellas
es la especie válida y cual la sinonimia) propia del Pacífico
Norte; y probablemente otras dos, todavía sin erigir, que
serían propias del Atlántico Sur e Índico y del Pacífico
meridional, respectivamente.
Hasta aquí lo que se refiere a la historia del nombre
científico de los calamares gigantes. En cuanto a los nombres
comunes o vernáculos que se les han dado desde muy
antiguo, en general, casi todos ellos hacen referencia al
enorme tamaño del animal. Así, en español se denominan
comúnmente calamares gigantes, aunque Guerra y Pérez-
Gándaras les dieron el nombre de megalurias para los
propósitos comerciales de FAO en la guía de especies
realizada por Roper, Sweeney y Nauen en 1984, que ahora
está en revisión. En francés, el calamar gigante recibe el
nombre de calmar géant o encornet monstre. En portugués
el de lula gigante, siendo en italiano calamaro gigante. Los
noruegos le denominan Kjemplblekksput, que literalmente
significa “calamar gigante que expulsa tinta”; nombre
parecido al de Kaemeblaeksprutt, que reciben en Dinamarca,

cuya etimología es similar a la que reciben en alemán, idioma
en que se denominan Riesentintenfish o Riesenkalmar. Los
rusos han acuñado el término de gigantskiy kalmar. Por
último, en Japón, el calamar gigante recibe el nombre común
de dai-oo-ika, que significa “gran rey de los calamares”,
mientras que en Nueva Zelanda las antiguas tradiciones
indígenas lo llaman Taninha, nombre que significa “serpiente
marina” en la mitología neozelandesa.

Capítulo 3
LA DISTRIBUCIÓN GEOGRÁFICA DE ARCHITEUTHIS
El Atlántico y el Mediterráneo
Los datos que se precisan para definir adecuadamente
el área de distribución geográfica de cualquier especie se
refieren a la distribución geográfica de sus individuos. La
Biogeografía es la parte de la Ecología que trata de encontrar
explicaciones a esa distribución, una vez que la información
proporcionada por aquellos datos ha sido sistematizada. Las
áreas de distribución de organismos sésiles, fácilmente
asequibles por alguna razón, o simplemente muy vistosos
(por ejemplo mariposas, moluscos gasterópodos y aves), o
de importancia comercial (especies de valor cinegético por
ejemplo), suelen estar bien respaldadas por una abundante
documentación y pueden aceptarse como base para los
estudios biogeográficos. Sin embargo, para numerosos
grupos de organismos, menos vistosos, menos asequibles, o
de menor interés, los conocimientos son muy incompletos,
hasta el punto que, para algunos de ellos, de amplia
distribución, las localidades en que han sido registrados
coinciden prácticamente con la distribución geográfica de sus
investigadores o de las regiones exploradas. Esto último es lo
que ocurre con Architeuthis, un organismo marino, oceánico,
meso-batipelágico y prácticamente cosmopolita, que cuenta
con el agravante de que el concepto de especie, basado en
características más o menos uniformes de los individuos,
presenta muchas dificultades de aplicación en este género
politípico, que podría estar compuesto por varias subespecies
o razas, o bien por varias especies diferentes que los
especialistas no han sido todavía capaces de definir.
Hasta ahora, el esfuerzo de muestreo aplicado a los
calamares gigantes ha sido muy pequeño y siempre
indirecto. Únicamente se cuenta con registros de capturas
accidentales en diferentes pesquerías, avistamientos

esporádicos de animales flotando en superficie, varamientos
en las costas, normalmente hallados por casualidad, y restos
encontrados en los estómagos de sus principales
depredadores. Estos datos, además de fraccionarios, son de
poca fiabilidad para definir con precisión la verdadera área de
distribución de los calamares gigantes. No obstante, son los
únicos con que contamos y con ellos hay que trabajar.
Aunque existen discrepancias, el número total de
registros de Architeuthis de los que se tiene constancia hasta
finales de 2004 en todo el Atlántico se puede evaluar en unos
306. Éstos se reparten muy desigualmente según regiones:
126 en el Atlántico Noroeste, 148 en el Atlántico nororiental,
tan sólo 6 en el Atlántico sudoeste, y 26 en la región
sudoriental de este océano.
Por razones desconocidas, pero que probablemente
estén relacionadas con cambios climáticos y oceanográficos,
aunque no se puede descartar una epidemia, docenas de
calamares gigantes aparecieron varados o flotando en
superficie en aguas de Terranova durante los años setenta
del siglo XIX (Tabla 1). Entre 50 y 60 de esos ejemplares,
según información de los pescadores, fueron recolectados por
buques pesqueros que faenaban en el Grand Bank y usados
como cebo para el bacalao. Fue posible recoger de las rocas
y playas otros 23 ejemplares, que fueron examinados por el
Verrill. Los varados y flotando en superficie aparecieron
principalmente en otoño e invierno. Aunque no se pudo
conocer el sexo de la mayoría de ellos, en aquellos en los que
si se supo este dato la desproporción a favor de las hembras
con relación a los machos es muy significativa. Estas
capturas y avistamientos desaparecieron bruscamente a
partir de finales del siglo XIX, volviendo a ser de nuevo
relativamente frecuentes en el periodo comprendido entre
1952 y 1977. Con todos estos datos históricos, el
investigador canadiense Frederick Aldrich había sugerido la
hipótesis de que el principal factor responsable de estos
acontecimientos eran las fluctuaciones que cada noventa
años experimentaba periódicamente la corriente fría de El
Labrador. Cuando una rama de agua de esa corriente,

conocida como Avalon Branch, chocaba con las tierras
septentrionales de Terranova, Tabla 1. Registros de Architeuthis en el Atlántico Noroeste. TN: Terranova. GL: Groenlandia. Mass: Massachusset (USA). Flor: Florida (USA). Bah: Bahamas. GM: Golfo de México. P: primavera. V: verano. O: otoño. I: invierno. M: macho. H: hembra. Va: varamiento. C: captura. FS: flotando en superficie. D:
depredadores. NC: no consta.
1785 a 1847
1848 a 1873
1874 a 1899
1900 a 1925
1926 a 1951
1952 a 1977
1978 a 2003
NC Total
Localidad TN 3 7 87 2 14 3 3 119 GL 1 1 Mass 1 1 2 Flor 2 2 Bah 1 1 GM 1 1 Periodo P 1 1 1 3 V 3 3 O 1 4 45 1 11 2 64 I 1 3 37 1 3 1 46 NC 2 2 6 10 Sexo M 1 1 2 3 7 H 23 1 8 2 34 NC 3 7 65 1 5 1 3 85 Registro Va 2 29 1 6 3 1 42 C 1 1 1 3 FS 1 4 54 2 1 4 66 D 1 1 2 NC 2 4 1 1 5 13 Total 3 7 88 1 2 14 5 6 126
los calamares se acercaban a las proximidades de la costa
siguiendo está masa de agua fría. Aldrich predijo que el
siguiente periodo en que los varamientos de Architeuthis
volverían a ser frecuentes ocurriría alrededor de los años
sesenta. Los hechos probaron que tenía razón, ya que entre
1961 y 1968 se hallaron nueve ejemplares de hembras

inmaduras varadas de nuevo en las costas del Labrador. Si
las predicciones de Aldrich son correctas, la próxima
aparición de calamares gigantes en estas costas debería
ocurrir alrededor del 2050. Sin embargo, y aun existiendo
esa coincidencia, la hipótesis de este investigador no está
probada. Por otra parte, mucho ha cambiado la situación en
los ecosistemas presentes en esas latitudes, principalmente
en lo referente a la abundancia de las poblaciones de peces
que podrían servir de alimento de los calamares gigantes,
drásticamente reducidas actualmente por múltiples efectos.
Los registros en el Atlántico nororiental se elevan a un
total de 148 (Tabla 2) abarcando un área muy extensa
comprendida entre Islandia y el estrecho de Gibraltar. La
mayoría (92,5%) de los animales registrados en Noruega
corresponden al periodo comprendido entre 1874 y 1977,
siendo casi nulos los avistamientos posteriores. Esta drástica
reducción no puede explicarse por una disminución en el
esfuerzo de observación ya que los calamares gigantes han
recibido mayor atención durante los años recientes y además
la pesca no ha disminuido en aquellas aguas. Este descenso
podría estar relacionado con acontecimientos oceanográficos.
Como demostraron Brix y sus colaboradores en 1989, la
capacidad de transporte de oxígeno de la hemocianina,
pigmento respiratorio de la sangre de los cefalópodos, que ya
es relativamente baja en este grupo, es muy inferior en los
calamares del tipo Architeuthis comparada con otros
cefalópodos pelágicos muy activos, como las potas de
diferentes especies. Por otra parte, a partir de las muestras
de sangre obtenidas en un calamar gigante que varó
prácticamente vivo en las proximidades de Bergen, Brix
demostró que la afinidad por el oxígeno era cuatro veces
inferior cuando la temperatura aumentaba de 6,4 a 15ºC, lo
cual sugiere con bastante fuerza que estos animales pueden
asfixiarse por desaturación de la sangre arterial cuando la
temperatura ambiente aumenta unos pocos grados. Este
hecho parece una explicación bastante plausible de la
aparición de calamares gigantes flotando moribundos en la
superficie del océano o varados en las costas de varios países
europeos, donde pueden interactuar con masas de aguas
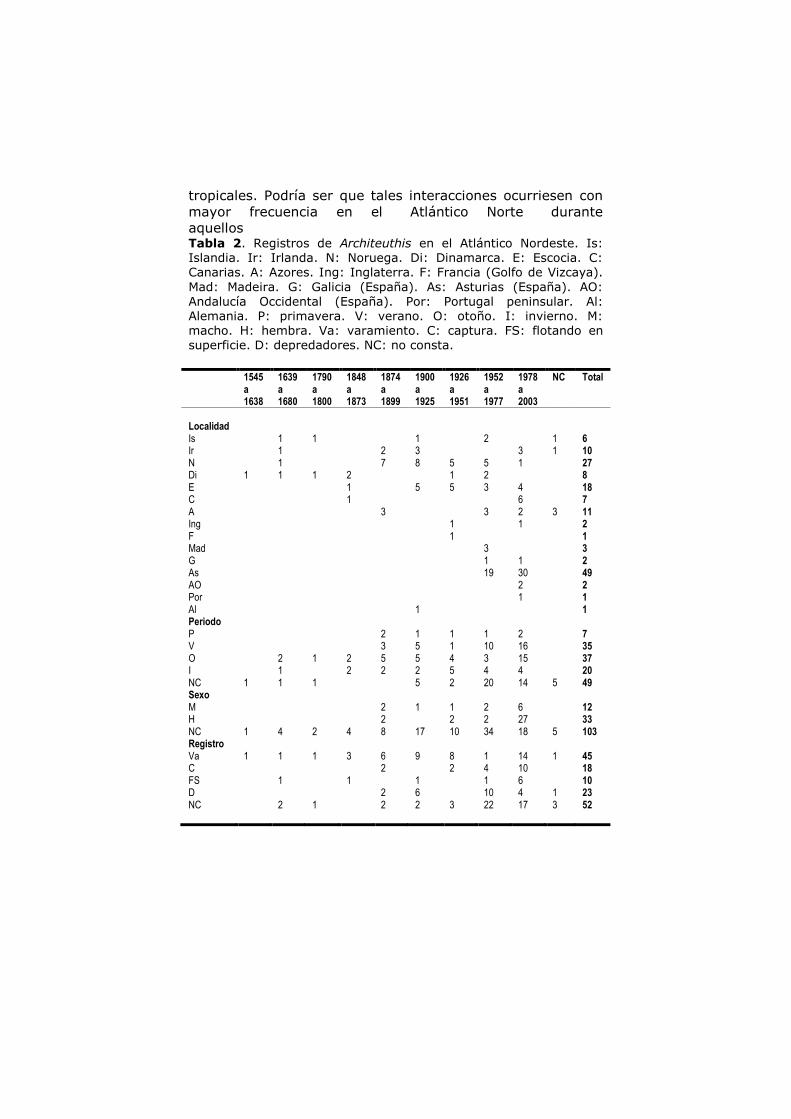
tropicales. Podría ser que tales interacciones ocurriesen con
mayor frecuencia en el Atlántico Norte durante
aquellos Tabla 2. Registros de Architeuthis en el Atlántico Nordeste. Is: Islandia. Ir: Irlanda. N: Noruega. Di: Dinamarca. E: Escocia. C: Canarias. A: Azores. Ing: Inglaterra. F: Francia (Golfo de Vizcaya).
Mad: Madeira. G: Galicia (España). As: Asturias (España). AO:
Andalucía Occidental (España). Por: Portugal peninsular. Al: Alemania. P: primavera. V: verano. O: otoño. I: invierno. M: macho. H: hembra. Va: varamiento. C: captura. FS: flotando en superficie. D: depredadores. NC: no consta.
1545 a 1638
1639 a 1680
1790 a 1800
1848 a 1873
1874 a 1899
1900 a 1925
1926 a 1951
1952 a 1977
1978 a 2003
NC Total
Localidad Is 1 1 1 2 1 6 Ir 1 2 3 3 1 10 N 1 7 8 5 5 1 27 Di 1 1 1 2 1 2 8 E 1 5 5 3 4 18 C 1 6 7 A 3 3 2 3 11 Ing 1 1 2 F 1 1 Mad 3 3 G 1 1 2 As 19 30 49 AO 2 2 Por 1 1 Al 1 1 Periodo P 2 1 1 1 2 7 V 3 5 1 10 16 35 O 2 1 2 5 5 4 3 15 37 I 1 2 2 2 5 4 4 20 NC 1 1 1 5 2 20 14 5 49 Sexo M 2 1 1 2 6 12 H 2 2 2 27 33 NC 1 4 2 4 8 17 10 34 18 5 103 Registro Va 1 1 1 3 6 9 8 1 14 1 45 C 2 2 4 10 18 FS 1 1 1 1 6 10 D 2 6 10 4 1 23 NC 2 1 2 2 3 22 17 3 52

Total 1 4 2 4 12 18 13 38 51 5 148
periodos en que, como resultado de una intensificación del
flujo de agua tropical procedente de la corriente del Golfo, se
incrementase la intensidad y la zona de influencia de las
corrientes cálidas de Irminger, de Noruega y de Canarias.
El gran número de registros correspondientes a
Asturias tiene una cuádruple explicación: por una parte, la
presencia de tres cañones submarinos que cortan la
plataforma continental aproximándose bastante a la línea de
costa; luego, el gran esfuerzo realizado por los
investigadores para recoger el mayor número de citas
posible; en tercer término, el aumento del esfuerzo de pesca
en los últimos años donde se supone que vive habitualmente
Architeuthis, de tal forma que la mayoría de los animales
aparecidos varados o flotando en superficie pueden atribuirse
a capturas accidentales; y, por último, las muertes
producidas por ondas acústicas en las prospecciones
geofísicas, realizadas al menos en los otoños de 2001 y
2003.
De nuevo se observa en esta área una desproporción
significativa entre el número de machos y de hembras. Es
difícil encontrar una explicación aceptable que permita
comprender la predominancia de hembras porque
prácticamente se desconoce casi todo sobre su ciclo vital y su
demografía. Este desequilibrio en la proporción sexual podría
también interpretarse como una real predominancia de las
hembras sobre los machos en las poblaciones naturales, lo
cual no es demasiado corriente en la naturaleza. Podría
también reflejar una mayor vulnerabilidad de las hembras
para la captura o el varamiento. Tal mayor vulnerabilidad
podría estar relacionada con una segregación espacial entre
ambos sexos. Por otra parte, la mayor frecuencia de
registros en otoño sugiere la existencia de algún
acontecimiento estacional que podría estar relacionado con
una mayor preponderancia de las hembras. Es posible, por
ejemplo, que su maduración sexual requiera mucha más
demanda de recursos somáticos y energéticos que la de los

machos, resultando ello en unos índices de condición (que
miden la “salud” del animal) más pobres en éstas que en
aquellos y, en consecuencia, de una mayor susceptibilidad a
las situaciones de estrés en las hembras que los machos. Sin
embargo, esta última hipótesis no parece resolver el
problema ya que la mayoría de las hembras registradas en el
Atlántico estaban sexualmente inmaduras o en maduración,
mientras que los machos estaban maduros.
Resulta evidente que el reducido número de registros
en el Atlántico Sudoeste (Tabla 3) bien podría deberse a un
sesgo de muestreo debido, entre otras cosas, a que muchos
animales varados o flotando en superficie no hayan sido
avistados por ocurrir en zonas prácticamente deshabitadas, o
bien, que aún habiendo sido vistos nadie los haya declarado.
Sin embargo, resulta algo sorprendente que siendo intensa
la actividad pesquera en varias zonas, sobre todo en la
plataforma Patagónica y alrededor de las islas Malvinas, haya
tan escasos registros. Esta carencia podría deberse a la
inexistencia de cañones submarinos en la extensa y
relativamente poco profunda plataforma Patagónica, lo que
parece ser, por lo observado en otras regiones, un factor
importante para asegurar la presencia de calamares
gigantes.
De los 26 registros procedentes del Atlántico
Sudoriental, 20 corresponden a las aguas de Namibia y
Sudáfrica (Tabla 4). Sin embargo, el número de registros
certificados de ambos países es una infravaloración de la
realidad ya que, como nos han comunicado los expertos
sudafricanos Martina Roeleveld y Marek Lipinski, son
numerosos los avisos que se reciben y que no pueden ser
atendidos por falta de medios, y es todavía mayor el número
de casos en que las capturas, los varamientos o los
avistamientos no se declaran. En esta zona, que comprende
toda la costa africana, hay dos áreas de afloramiento muy
características. Una de ellas está situada al norte y tiene
como centro geográfico cabo Blanco, la otra se halla al sur y
está estrechamente relacionada con la corriente de Benguela.
En ambas zonas hay importantes pesquerías que se explotan
intensamente desde hace muchos años.

El área comprendida entre cabo Bojador (26º10’N) y
el sur de cabo Verde (14ºN) puede dividirse en dos zonas
desde un punto de vista hidrográfico. La primera
comprendida entre cabo Bojador y punta Durnford (23º30’N),
y la segunda desde

157
Fig. 19. Principales corrientes oceánicas
84

91
esa latitud hasta 14ºN aproximadamente. Ambas zonas
tienen dos mecanismos de fertilización que se complementan
en distinto grado según la época del año, produciéndose el
afloramiento en la zona norte en verano, y en la sur
preferentemente en invierno, aunque persista con mayor o
menor intensidad durante todo el año, dando lugar a una
elevada producción primaria. La concentración de oxígeno
durante el afloramiento, cuyo ojo se sitúa entre 24 y 25ºN,
es siempre superior a 4 ml por litro a 100m de profundidad,
es decir el agua está saturada de ese gas. Otra de las
características de esta zona es la presencia de la corriente de
Canarias, que fluye hacia el sur a lo largo de la costa.
Entre 23º30’N y 14ºN también se pueden diferenciar
dos zonas: una comprendida entre cabo Barbas y cabo
Blanco, donde el afloramiento es constante durante todo el
año, y otra que abarca desde cabo Blanco hasta el sur de
cabo Verde, donde éste se produce en primavera y,
fundamentalmente, en invierno. Las aguas superficiales de
ambas zonas están prácticamente saturadas de oxígeno y
agotadas en nutrientes. Cuando las aguas profundas afloran
en superficie favorecen una elevada producción primaria.
Lo sorprendente de toda esta amplia región del
Atlántico Centro-Oriental, donde hay importantes pesquerías
de arrastre de varias especies de cefalópodos y peces desde
hace muchos años, es que nunca se han producido registros
de calamares gigantes. Quizá esto se deba a la inexistencia
de importantes cañones submarinos que se adentren en la
plataforma continental, o a que las características de las
masas de agua, especialmente su relativa elevada
temperatura, no constituyen un hábitat adecuado para estos
cefalópodos.
Desde la desembocadura del río Cunene (18º38’S)
hasta el cabo de Agulhas (34º50’S), la característica más
destacada es la presencia de la corriente de Benguela. Esta
corriente de aguas frías (8-10ºC) discurre hacia el norte con
cierta desviación hacia el oeste, y al remontar el talud y la
plataforma provoca un afloramiento. Se ha demostrado que
entre los paralelos 20 y 29ºS la intensidad del
afloramiento

91
Tabla 3. Registros de Architeuthis en el Atlántico sudoeste. Or: Orkney. SS: Shetland del sur. Br: Brasil. Arg: Argentina. P: primavera. V: verano. O: otoño. H: hembra. Va: varamiento. C: captura. FS: flotando en superficie. D: depredadores. NC: no consta.
1952 a 1977 1978 a 2003 Total
Localidad Or 1 1 SS 1 1 Brl 1 1 Arg 3 3 Periodo P 2 2 V 1 3 O 1 1 Sexo H 4 4 NC 2 2 Tipo registro Va 1 1 C 1 1 FS 2 2 D 2 2 NC 2 Total 2 4 6
es máxima entre septiembre y octubre, aunque este no
ocurre uniformemente a lo largo de toda la costa, sino que se
restringe a ciertos lugares, hallándose valores máximos entre
25 y 28ºS, existiendo núcleos de afloramiento entre 19º30’S
y 23ºS a los cuales están vinculadas áreas de estabilidad y
posible hundimiento.
Se han obtenido varios registros de calamares
gigantes en aguas de Namibia y de Sudáfrica (entre 17ºS y
35ºS aproximadamente) y aun más al sur, incluso hasta
cerca de la Antártica, aunque esos hallazgos proceden
principalmente de depredadores, siendo muy difícil concretar
el área donde fueron capturada sus presas. Estos registros
85
86
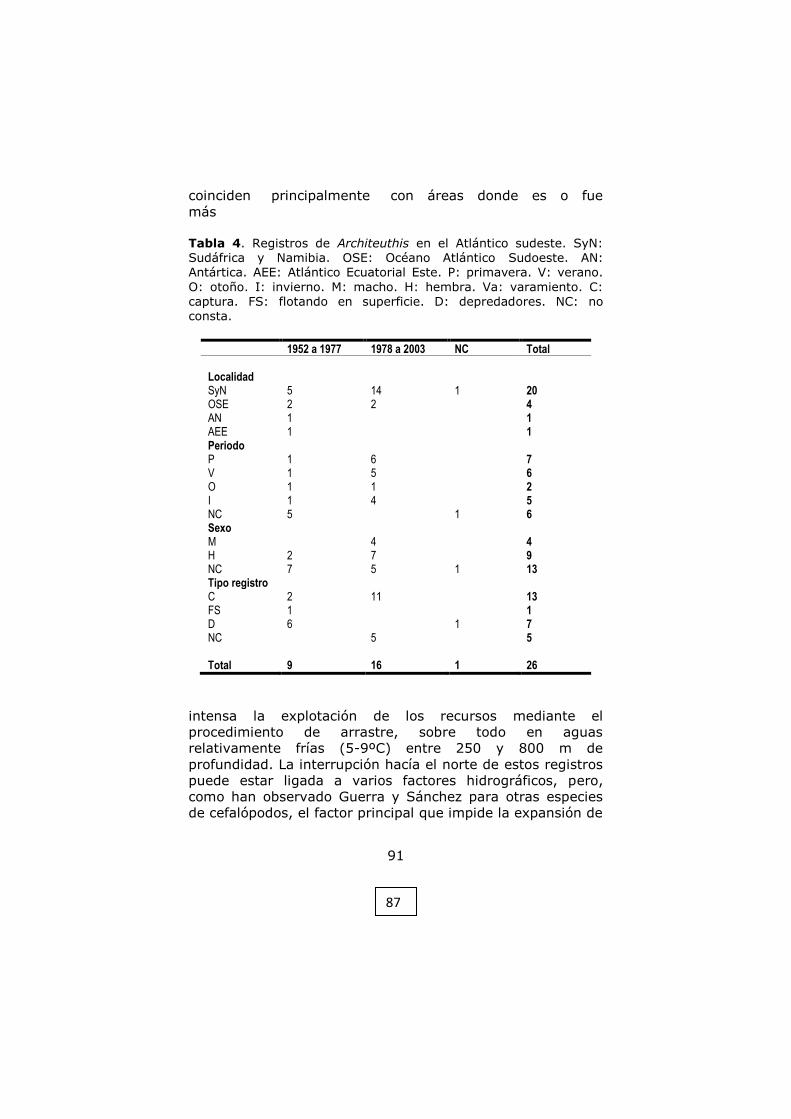
91
coinciden principalmente con áreas donde es o fue
más
Tabla 4. Registros de Architeuthis en el Atlántico sudeste. SyN: Sudáfrica y Namibia. OSE: Océano Atlántico Sudoeste. AN: Antártica. AEE: Atlántico Ecuatorial Este. P: primavera. V: verano.
O: otoño. I: invierno. M: macho. H: hembra. Va: varamiento. C:
captura. FS: flotando en superficie. D: depredadores. NC: no consta.
1952 a 1977 1978 a 2003 NC Total
Localidad SyN 5 14 1 20 OSE 2 2 4 AN 1 1 AEE 1 1 Periodo P 1 6 7 V 1 5 6 O 1 1 2 I 1 4 5 NC 5 1 6 Sexo M 4 4 H 2 7 9 NC 7 5 1 13 Tipo registro C 2 11 13 FS 1 1 D 6 1 7 NC 5 5 Total 9 16 1 26
intensa la explotación de los recursos mediante el
procedimiento de arrastre, sobre todo en aguas
relativamente frías (5-9ºC) entre 250 y 800 m de
profundidad. La interrupción hacía el norte de estos registros
puede estar ligada a varios factores hidrográficos, pero,
como han observado Guerra y Sánchez para otras especies
de cefalópodos, el factor principal que impide la expansión de
87

91
numerosas especies es la depleción de oxígeno y la
acumulación de materia orgánica en muchos fondos de áreas
de la plataforma continental. Lo cual sugiere, en el caso de
Architeuthis, que estos organismos están más relacionados
con el fondo de lo que piensan algunos investigadores. Por
otra parte, la llanura abisal del Cabo, comprendida entre la
costa y la dorsal de Walbis, está comunicada con el talud y la
plataforma continental por cañones submarinos en al
menos cinco zonas, lo que parece ser una constante en
todas las áreas donde son abundantes los registros de
calamares gigantes.
Dentro del sistema mundial de los océanos existen
masas de agua rodeadas por tierra que forman mares
periféricos, los cuales son particularmente interesantes para
la investigación biogeográfica. Entre ellos, el mar
Mediterráneo es la unidad más claramente diferenciada. El
Mediterráneo comprende una serie de fosas profundas
separadas por terrazas y rodeadas por áreas de plataforma
continental de extensión variable. En su extremo occidental,
el estrecho de Gibraltar constituye su conexión con el océano
Atlántico. El estrecho tiene un máximo de 14 Km. de anchura
y su mayor profundidad es de 260 m. Además de las fuertes
corrientes de marea generadas en el Atlántico, los
movimientos del agua a través de este cuello de botella que
significa el estrecho están gobernados por una masiva
entrada de agua oceánica por la superficie, y una
contracorriente, más débil y profunda, que discurre en
sentido contrario. Esta corriente transporta agua de elevada
salinidad hacia el Atlántico.

91
Fig. 20. Circulación oceánica superficial en el Mediterráneo. Tabla 5. Registros de Architeuthis en el Mar Mediterráneo. Ma: Málaga. I: invierno. H: hembra. Va: varamiento.
1978 a 2003 Total
Localidad Ma 2 2 Periodo I 2 2 Sexo H 2 2 Tipo registro V 2 2 Total 2 2
En su revisión de la fauna teutológica del
Mediterráneo realizada en 1988, Mangold y Boletzky
señalaban que a excepción de unas pocas especies
endémicas de sepiólidos, todos los cefalópodos que se
encuentran actualmente en el Mediterráneo se hallan
también en el Atlántico oriental. De alguna forma, parece que
el modelo de distribución seguido habría sido una invasión de
especies desde el océano hacia el mar, mas que un
movimiento en sentido opuesto. En 1988 no había todavía
citas de Architeuthis en el Mediterráneo. Sin embargo, como
se muestra en la Tabla 5, actualmente hay dos registros. El
primero de ellos, que tuvimos ocasión de examinar
conservado en formol en el Aula del Mar de CREMA en
Málaga, se trata de una hembra inmadura, cuyo manto mide
aproximadamente 125 cm. de longitud. Este ejemplar
apareció muerto, relativamente fresco y varado en una playa
próxima a Fuengirola (Málaga) a mediados de 1998. El
segundo ejemplar fue hallado muerto, relativamente freso y
varado en una playa de Fuengirola el 27 de junio de 2001. Se
trata de una hembra inmadura de 65 Kg., 125 cm. de manto
88

91
y 750 cm. de longitud total. Este ejemplar está conservado
en el Museo Nacional de Ciencias Naturales (CSIC, Madrid).
La hipótesis más plausible para explicar la presencia
de ambos cadáveres a unos cien kilómetros del estrecho de
Gibraltar es que, procedentes del Atlántico, fueron
arrastrados por la corriente superficial oceánica que penetra
en el Mediterráneo. Sin embargo, la captura de un macho
maduro de 47 Kg. por una arrastrero con base en Gandía el
19 de julio de 2005 entre 420 y 450 metros de profundidad
cuestiona esta hipótesis. Según recientes informaciones, las
grandes barcos de arrastre del Levante español están
pescando cada vez a mayor profundidad en busca,
principalmente, de recursos de gamba. Algunos de ellos
largan sus redes en fondos de hasta 1.000 metros de
profundidad, caladeros que nunca antes habían sido
explotados comercialmente. Cabe plantearse, por lo tanto,
que Architeuthis habitase en el Mediterráneo occidental, pero
que no hubiese sido detectado anteriormente,
consiguiéndose registros sólo cuando, recientemente, los
pesqueros han incrementado su rango batimétrico de
actuación, abarcando zonas que comprenden el hábitat
natural de estos gigantes.
El Pacífico
El Pacífico es el mayor y más profundo de los océanos
del globo con una superficie casi equivalente al Índico y
Atlántico juntos (Figura 19). En esta extensa área se han
encontrado, e identificado como tales, 264 Architeuthis.
Como ocurre en el océano Atlántico, los registros de la
especie se reparten de forma muy desigual: 28 en el Pacífico
noroeste, 43 en el Pacífico nordeste, 183 en el Pacífico
suroeste, y, finalmente, 10 en el Pacífico sudeste.
Es curioso que en un océano tan grande como es el
Pacífico no se registren más capturas de Architeuthis, sobre
todo considerando que allí se desarrollan importantes
pesquerías y habitan, como mínimo, dos subespecies de
calamar gigante: Architeuthis dux martensii en el Pacífico
norte y A. dux sanctipauli en el Pacífico sur, esto
90
89

91
considerando el último trabajo de Nesis sobre la distribución
de los cefalópodos. Sin embargo, y como se verá más
adelante, esta relativa escasez puede explicarse por la propia
extensión del océano y, en muchos casos, por la falta de
registros fiables. Salvo en las aguas de Nueva Zelanda,
Australia, Japón y Estados Unidos, dónde la correcta
identificación de los ejemplares es posible gracias a la
presencia de especialistas, en la mayoría de las zonas del
océano Pacífico su aparición pasa desapercibida o, en el caso
de ejemplares de pequeño tamaño, puede confundirse con
otras especies por lo que no existirían registros de su
presencia. Además, hay extensas zonas costeras en el
Pacífico muy poco habitadas, donde la aparición de un
calamar gigante puede pasar fácilmente inadvertida.
Las apariciones de Architeuthis en el Pacífico noroeste
(Tabla 6) están centradas principalmente entre invierno Tabla 6. Registros de Architeuthis en el Pacífico Noroeste. J: Japón. Chi: China. OP: Océano Pacífico. P: primavera. V: verano. I: invierno. M: macho. H: hembra. Va: varamiento. C: captura. FS:
flotando en superficie. D: depredadores. NC: no consta.
1850 a 1900
1900 a 1925
1925 a 1950
1950 a 1975
1975 a 2000
2000 - NC Total
Localidad J 2 1 3 6 7 1 2 22 Chi 1 1 OP 5 5 Periodo P 1 2 3 6 V 3 1 4 I 1 1 1 5 8 NC 1 1 1 5 2 10 Sexo M 1 1 H 1 1 2 NC 1 3 11 8 2 25 Registro Va 1 4 2 1 8 C 1 1 1 3

91
FS 1 1 D 4 5 9 NC 1 1 2 2 1 7 Total 2 1 3 11 8 1 2 28
y verano en aguas de Japón. Los registros de esta especie
han aumentado a partir de 1950, probablemente debido a la
existencia de mejores datos y a la presencia de especialistas.
De los 28 registros conocidos en toda el área, la mayoría
(60,7%) corresponden a varazones en la costa (28,6%) o a
la identificación de restos en estómagos de depredadores
(32,1%).
Esta región del Pacífico está controlada por el sistema
de corrientes del Japón, donde la corriente de Kuroshio, una
corriente cálida, estrecha y rápida que fluye hacia el
Noroeste a todo lo largo de la costa oriental japonesa, es la
más importante. Contraria a ésta, fluye la corriente fría de
Oyashio.
En esta zona del Pacífico existen una serie de fosas
como las Filipinas, Kuriles, Aleutianas y Challenger, la
mayoría con cerca de 10.000 m de profundidad. En estas
fosas el agua es fría, no superando normalmente los 2º C,
pero existe una elevada producción biológica, por lo que
constituye un hábitat adecuado para algunas especies
marinas. Como ya se ha mencionado, Architeuthis parece
habitar aguas frías, dónde su sangre puede transportar el
oxígeno suficiente para vivir. Es probable que la aparición de
estos ejemplares en aguas de Japón se produzca asociada a
estos cañones dónde las masas de agua fría y el abundante
alimento les facilitarían un hábitat adecuado.
En el Pacífico nordeste se han registrado 43
ejemplares de calamar gigante (Tabla 7), la mayoría
repartidos frente a las costas de California (44,2%) y en la
zona oceánica del Pacífico (44,2%). Existen registros a partir
de 1900, pero es después de 1950 cuando estos aumentan
sustancialmente. Architeuthis ha sido registrado en todas las
estaciones del año, con una mayor incidencia durante la
primavera (48,8%). En esta zona, las apariciones de la
especie corresponden principalmente a capturas con artes de

91
pesca (55,8%) y a registros de presencia en la dieta de sus
depredadores (32,6%).
Aparte de las Aleutianas, no existen grandes fosas
oceánicas en esta zona y el área está dominada en la parte
septentrional por la corriente Subantártica, y algo más al Sur
por la corriente del Pacífico Norte, que se desplaza en
dirección oeste a partir del meridiano 170º E. Una rama de
esta corriente se separa de la costa hacia oriente alcanzando
las islas Hawai, y otra, desplazándose en dirección Sur, se
Tabla 7. Registros de Architeuthis en el Pacífico Nordeste. Cal: California. VyO: Vancouver y Oregón. Ha: Hawai. OP: Océano Pacífico. P: primavera. V: verano. O: otoño. I: invierno. M: macho. H: hembra. Va: varamiento. C: captura. FS: flotando en superficie. D: depredadores. NC: no consta.
1900 a
1925
1925 a
1950
1950 a
1975
1975 a
2000
NC Total
Localidad Cal 2 11 3 3 19 VyO 2 1 3 Ha 2 2 OP 19 19 Periodo P 3 18 21 V 1 2 5 8 O 1 3 1 5 I 3 2 5 NC 4 4 Sexo M 4 4 H 8 8 NC 2 11 14 4 31 Registro Va 1 1 C 1 23 24 FS 1 1 D 11 3 14 NC 3 3
92

91
Total 2 0 11 26 4 43

Fig. 21. Relieve del fondo marino. (Tomado de B.C.Heezen y M, Tharp, 1977)

169
aproxima a la costa originando la corriente de California. La
tercera rama, que se desplaza hacia el Norte dando lugar a
las corrientes de Alaska y Behring.
La mayoría de los Architeuthis registrados en aguas
de California se hallaron en los contenidos gástricos de varios
tipos de depredadores, que perfectamente pudieron haber
ingerido esas presas lejos de la zona dónde fueron
capturados. Por el contrario, la mayoría de los ejemplares
capturados con redes, lo fueron en océano abierto y apenas
uno ha sido encontrado varado. Una explicación plausible de
esta situación es que Architeuthis se mantenga alejado de la
costa en esta zona, acercándose sólo ocasionalmente a ella
cuando las condiciones así lo permiten y las aguas
relativamente cálidas de la corriente de California no les
impiden el paso.
En el hemisferio sur, la zona suroeste del Pacífico es la
que presenta el mayor número de apariciones de
Architeuthis, con 183 ejemplares y registros que se
remontan a la segunda mitad del siglo XIX (Tabla 8). Pese a
ello, la mayoría de los registros corresponden al periodo
comprendido desde la década de 1950 hasta la fecha
(50,8%). Ya desde los primeros momentos son las costas de
Nueva Zelanda las que poseen la mayoría de las apariciones
de la especie (57,4%), seguidas por Tasmania (34,4%) y,
finalmente, las costas de Australia (8,2%). Aunque se
encuentran ejemplares durante todo el año, la mayor
incidencia parece ocurrir en otoño (39,3%), siendo las
capturas con redes (41,0%) y la presencia en la dieta de sus
depredadores (42,1%), las principales fuentes de registros
en la zona. Especial mención merecen, en este caso, las
fosas de Tonga y Kermadec al nordeste de Nueva Zelanda.
La fosa de las islas Tonga es el segundo punto más profundo
del planeta, con más de 10 kilómetros de profundidad.
Comienza en las proximidades de las islas Samoa y se
extiende hacia el Sur hasta unirse a la fosa de las Kermadec,
que continúa hasta Nueva Zelanda. Allí forma un sistema de
depresiones y cañones submarinos que conectan al cañón
submarino de Kaikoura con la fosa de Kermadec a través de
la depresión de Hikurangi. A esta zona con aguas y cañones
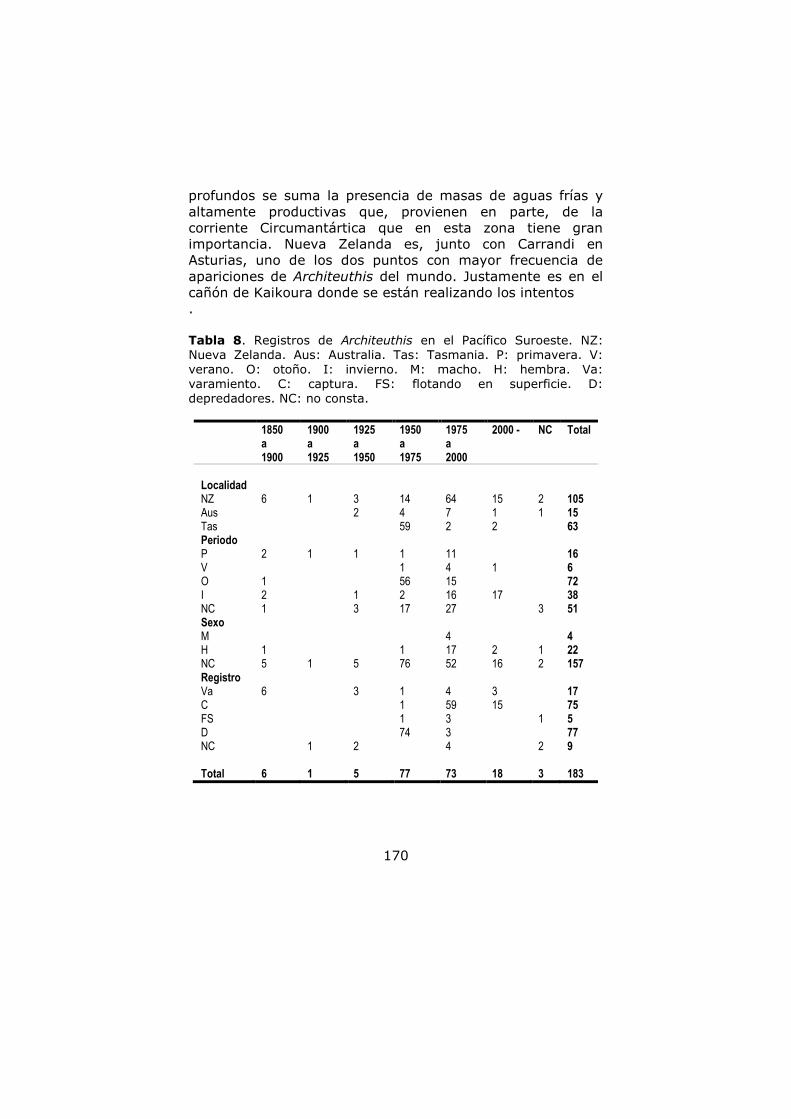
170
profundos se suma la presencia de masas de aguas frías y
altamente productivas que, provienen en parte, de la
corriente Circumantártica que en esta zona tiene gran
importancia. Nueva Zelanda es, junto con Carrandi en
Asturias, uno de los dos puntos con mayor frecuencia de
apariciones de Architeuthis del mundo. Justamente es en el
cañón de Kaikoura donde se están realizando los intentos
.
Tabla 8. Registros de Architeuthis en el Pacífico Suroeste. NZ: Nueva Zelanda. Aus: Australia. Tas: Tasmania. P: primavera. V: verano. O: otoño. I: invierno. M: macho. H: hembra. Va: varamiento. C: captura. FS: flotando en superficie. D: depredadores. NC: no consta.
1850 a 1900
1900 a 1925
1925 a 1950
1950 a 1975
1975 a 2000
2000 - NC Total
Localidad NZ 6 1 3 14 64 15 2 105 Aus 2 4 7 1 1 15 Tas 59 2 2 63 Periodo P 2 1 1 1 11 16 V 1 4 1 6 O 1 56 15 72 I 2 1 2 16 17 38 NC 1 3 17 27 3 51 Sexo M 4 4 H 1 1 17 2 1 22 NC 5 1 5 76 52 16 2 157 Registro Va 6 3 1 4 3 17 C 1 59 15 75 FS 1 3 1 5 D 74 3 77 NC 1 2 4 2 9 Total 6 1 5 77 73 18 3 183

171
más serios, aparte de los realizados en Asturias, para
localizar y filmar vivos a los calamares gigantes.
La zona del océano Pacífico sudeste es la que menos
registros presenta, con apenas 10 ejemplares entre 1950 y
2004 (Tabla 9). Esta cantidad se reparte entre dos
ejemplares aparecidos en la costa de Chile y ocho capturados
en aguas oceánicas
Esta zona presenta una larga fosa submarina, la fosa
Perú-Chile, con profundidades de más de 8000 m, que
bordea toda la costa de Sudamérica. El rasgo más
característico de esta zona corresponde a la corriente del
Perú, también llamada corriente de Humboldt. Se trata de
una corriente oceánica fría que fluye hacia el Norte a lo largo
de la costa occidental de sudamericana. Ocasionalmente, la
corriente de Humboldt es reemplazada por una masa de
agua cálida procedente del Pacífico central dando lugar al
evento conocido como El Niño. Resulta bastante
sorprendente que no se registren más ejemplares de
Architeuthis en la zona. Una explicación a este fenómeno
podría ser la falta de estudios profundos sobre los
cefalópodos del área y la carencia de información suficiente
para identificar estos ejemplares. Además, hay que
mencionar que a lo largo de casi toda la costa americana
habita una especie de pota o jibia de gran tamaño, Dosidicus
gigas. Esta especie puede alcanzar hasta 2 metros de
longitud y ha recibido también la denominación de “calamar
gigante”. Aparte de que D. gigas sustenta una importante
pesquería, es frecuente encontrar ejemplares varados en la
costa, por lo que distinguir entre las apariciones de éstos y
los Architeuthis, sobre todo si se trata de ejemplares
juveniles, es difícil para los lugareños. Como Architeuthis
habita aguas frías, parece más probable que pudiese
aparecer asociado a la corriente de Humboldt, sobre todo en
las costas de Chile, pero su aparición podría haber pasado

172
Fig. 22. Hembra de Architeuthis de 12 m de longitud total
capturada frente a Tasmania. (Foto de David Paul)
Tabla 9. Registros de Architeuthis en el Pacífico Sudeste. Ch: Chile. OP: Océano Pacífico. V: verano. I: invierno. M: macho. H: hembra. C: captura. D: depredadores. NC: no consta.
1950 a 1975
1975 a 2000
2000 - Total
Localidad Ch 1 1 2 OP 8 8 Periodo V 1 1 I 8 1 9 Sexo M 1 1 H 1 1 NC 8 8 Registro C 8 1 9 D 1 1 Total 1 8 1 10
totalmente desapercibida debido a que ese litoral posee
muchas zonas muy poco habitadas y a que los naturales del
lugar prestan escasa atención a la aparición de cefalópodos
en sus costas.

173
El Índico
El océano Índico es el menor en extensión de los tres
grandes océanos, con una superficie de 74.900.000 Km2,
extendidos mayormente al sur del Ecuador. Está limitado por
Australia al este, el continente asiático al norte, el africano al
oeste, y el océano Antártico por el sur. Su profundidad
promedio es de 3.900 metros, y la máxima de 7.450 metros,
situada en la llamada Fosa de la Sonda al suroeste de Java.
Cabe destacar que la región del Índico es relativamente
pobre en fosas y grandes profundidades, aunque si posee
importantes cordilleras submarinas, como la de Carlsberg
que divide el océano Índico desde el Mar Arábigo hasta los
20°S (latitud del Madagascar central). En este sistema, la
corriente marina más importante es la de Agulhas, que
transcurre frente a Mozambique, limitada al norte y sur por
las respectivas corrientes ecuatoriales, que discurren ambas
en dirección contraria a las agujas del reloj (Fig. 19).
Este océano, en el cual son famosos los monzones,
posee una zona norte dominada principalmente por masas de
aguas cálidas y tropicales, lo que no excluye la aparición de
sistemas de afloramientos en la costa africana, mientras que
en el sur se pueden observar masas de agua fría
provenientes de las corrientes antárticas.
El número conocido de citas de calamar gigante en el
océano Índico es escaso (Tabla 10), limitándose solamente a
20 registros. La mayoría proviene de datos de restos
encontrados en la dieta de grandes depredadores. Aunque la
escasez de registros podría indicar que la población de
calamar gigante en el Índico es reducida, no se puede
descartar que la falta de registros se deba más bien a que las
condiciones hidrográficas reinantes en la región son
desfavorables para el desplazamiento de los especimenes
hacia la costa. Tampoco puede ignorarse que las costas del
Índico poseen zonas poco pobladas y que existen escasos
medios para localizar y estudiar ejemplares varados en la
costa, lo que podría ser una explicación alternativa a la falta
de registros en la región. Pese a los pocos registros
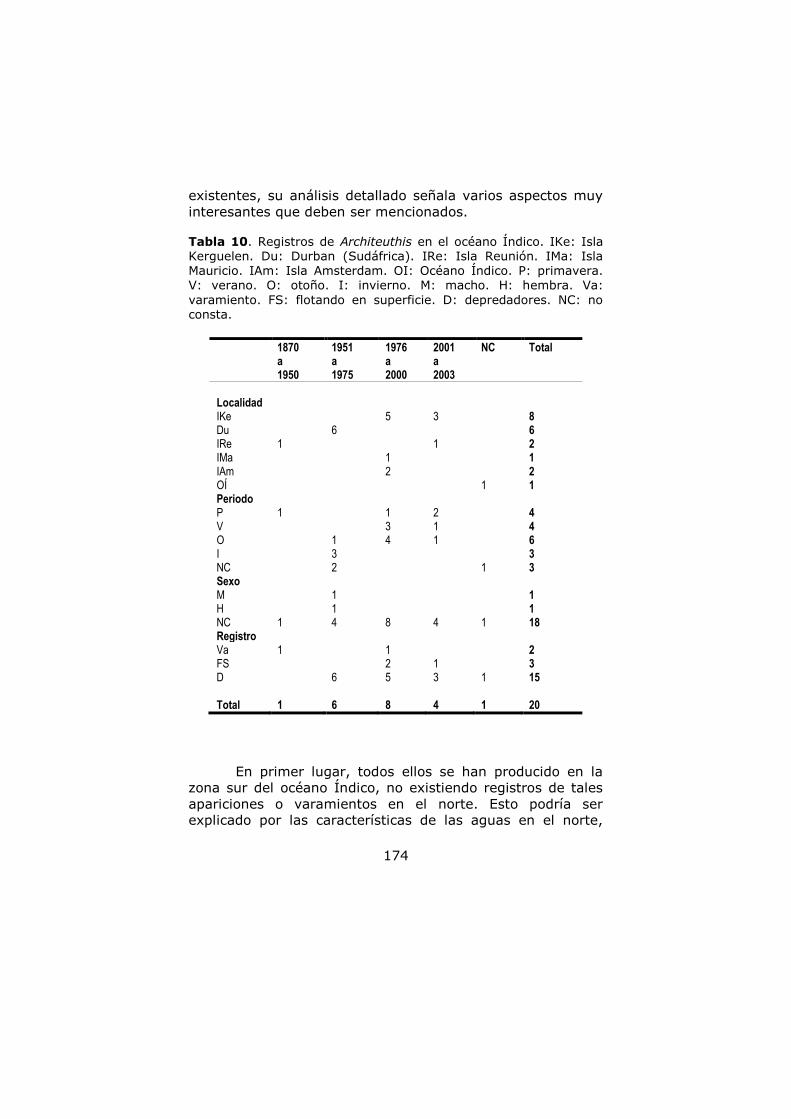
174
existentes, su análisis detallado señala varios aspectos muy
interesantes que deben ser mencionados. Tabla 10. Registros de Architeuthis en el océano Índico. IKe: Isla Kerguelen. Du: Durban (Sudáfrica). IRe: Isla Reunión. IMa: Isla Mauricio. IAm: Isla Amsterdam. OI: Océano Índico. P: primavera. V: verano. O: otoño. I: invierno. M: macho. H: hembra. Va:
varamiento. FS: flotando en superficie. D: depredadores. NC: no consta.
1870 a 1950
1951 a 1975
1976 a 2000
2001 a 2003
NC Total
Localidad IKe 5 3 8 Du 6 6 IRe 1 1 2 IMa 1 1 IAm 2 2 OÍ 1 1 Periodo P 1 1 2 4 V 3 1 4 O 1 4 1 6 I 3 3 NC 2 1 3 Sexo M 1 1 H 1 1 NC 1 4 8 4 1 18 Registro Va 1 1 2 FS 2 1 3 D 6 5 3 1 15 Total 1 6 8 4 1 20
En primer lugar, todos ellos se han producido en la
zona sur del océano Índico, no existiendo registros de tales
apariciones o varamientos en el norte. Esto podría ser
explicado por las características de las aguas en el norte,

175
mucho más cálidas, además de la escasa existencia de
cañones submarinos en la zona. Por otra parte,
prácticamente no hay especialistas en cefalópodos en los
países del área, las pesquerías de la zona son costeras y
escasamente explotan los recursos más pelágicos y,
finalmente, parece existir escaso interés local por publicar
este tipo de hallazgos.
En segundo lugar, muchos de los especimenes de
calamar gigante han sido encontrados en aguas más
asociadas al océano Antártico que al Índico. Los registros
correspondientes a ejemplares en contenidos gástricos de
depredadores (un 75,0% del total) se encontraron en
estómagos de cachalotes recolectados en Durban, Sudáfrica,
o en los del tiburón dormilón, una especie de profundidad, en
las aguas de las Islas Kerguelen en el Frente Polar. En ambos
casos, al tratarse de restos en el estómago y estar cerca del
límite del océano Índico, no se puede descartar que los
ejemplares fueran realmente ingeridos en zonas adyacentes
a este océano.
Al menos cinco ejemplares de Architeuthis han sido
hallados varados o flotando alrededor de las islas Reunión,
Mauricio y Amsterdam, cuyas aguas se consideran tropicales
o subtropicales. Estos registros son muy interesantes ya que
el calamar gigante parece más asociado a las aguas frías que
a las cálidas. Sin embargo, hay que recordar que esta
especie habita aguas profundas y la circulación de agua
profunda es diferente a la observada en la superficie.
Justamente por la zona donde se localizan estas islas circula
agua profunda fría procedente de la Antártica, dónde podría
habitar el calamar gigante.
Comentarios sobre su distribución mundial
Como se muestra en el mapa de la figura 23,
Architeuthis es un género prácticamente cosmopolita,
aunque parece estar ausente en las aguas polares y en el
área comprendida entre los trópicos de Cáncer (23º27’N) y
de Capricornio (23º27’S).

176
La práctica ausencia de registros de calamares
gigantes en las áreas ecuatorial y tropical no es indicio
suficiente para sugerir la total ausencia de Architeuthis en
estas zonas geográficas. Lo más probable es que en ellas
haya también calamares gigantes aunque en escasa
abundancia y más dispersos que en las zonas adyacentes. A
las profundidades en que suelen morar estos gigantes
marinos no hay realmente fronteras entre el hemisferio norte
y el sur. Como se ha demostrado en otras especies, como los
atunes, el cachalote y la pota cárdena (Stenoteuthis
pteropus), bien podría ser ésta una extensa área de
solapamiento entre dos poblaciones diferentes de
Architeuthis, una norteña y otra sureña, o bien una zona de
confluencia de dos ecotipos, subespecies o especies
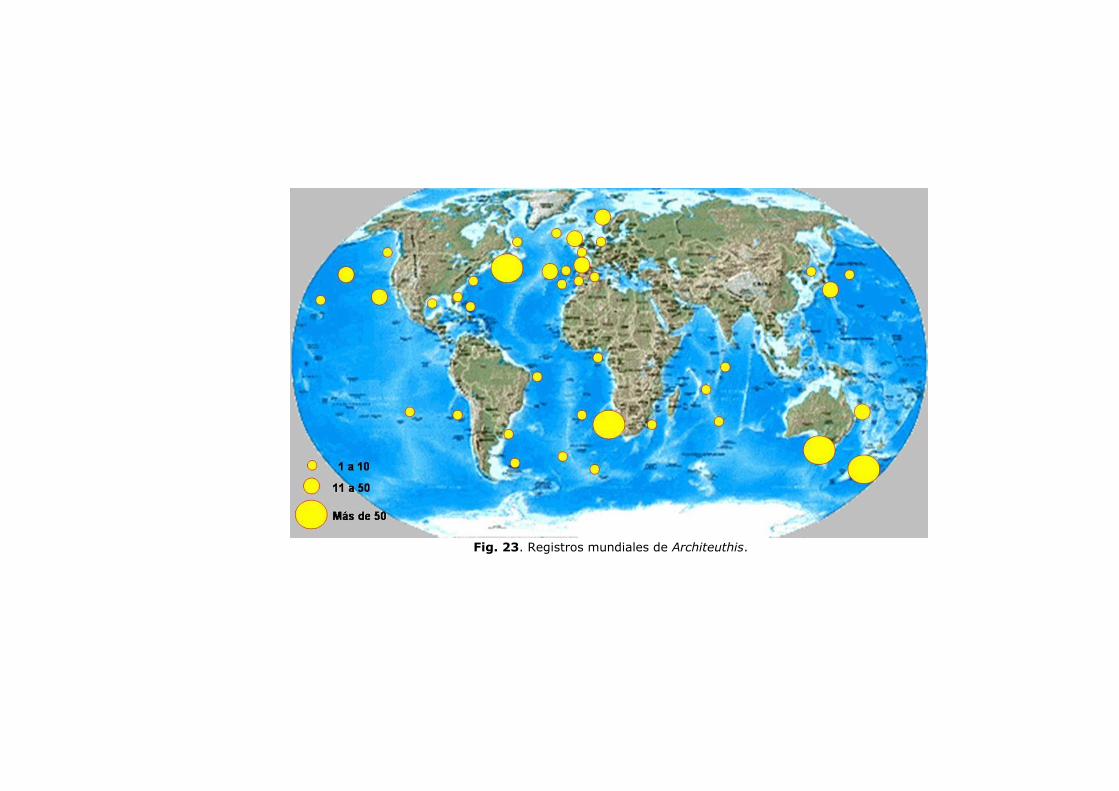
Fig. 23. Registros mundiales de Architeuthis.

107
diferentes. Esta suposición viene apoyada por algunos datos
procedentes de investigadores rusos (Nigmatullin,
comunicación personal) quienes encontraron tentáculos de
Architeuthis flotando en superficie mientras realizaban
trabajos de investigación a bordo de atuneros, que
habitualmente faenan en las proximidades del Golfo de
Guinea y el archipiélago de Cabo Verde. Por otra parte,
Nigmatullin tiene constancia de la presencia de 5 ó 6
ejemplares de Architeuthis poco digeridos en estómagos de
tiburón azul y de cachalotes capturados en las proximidades
de Cabo Verde. Si a esto sumamos los ejemplares
encontrados varados o flotando en aguas tropicales y
subtropicales del Índico, parece claro que la especie estaría
presente, pero dispersa, en los trópicos. Es muy posible que
en este caso, la presencia de agua fría profunda que circula
por todos los fondos oceánicos sea la que permite a estos
ejemplares sobrevivir en zonas donde las aguas superficiales
son exclusivamente cálidas y quizás no posean suficiente
oxígeno disuelto para permitir la vida de estos gigantes
marinos.
Muchas veces la distribución geográfica de una
especie o de un género con una amplia dispersión, como es
el caso del calamar gigante, pone de manifiesto
fundamentalmente la distribución geográfica de los
investigadores que trabajan en ella o indica las regiones más
explotadas, en lugar de mostrar su verdadera distribución.
Lamentablemente, esto es lo que ocurre con Architeuthis
donde no existen auténticos censos.
Distribución vertical
Aunque bastantes especies de cefalópodos llevan una
vida más o menos sedentaria viviendo durante todo su ciclo
vital en el mismo biotopo, hay muchas otras que efectúan
migraciones, ya sean estacionales o diarias, o, más
raramente, de ambos tipos. Las migraciones estacionales son
quizá los únicos desplazamientos horizontales que se realizan
de forma más generalizada durante la vida del cefalópodo, y
pueden abarcar distancias considerables. Sin embargo,

108
existen también numerosas especies que se desplazan
estacionalmente desde aguas costeras hacia más profundas y
viceversa por motivos ligados a la reproducción o a la
alimentación. En este caso, se trata de desplazamientos que
tienen una doble componente espacial: horizontal y vertical.
Desafortunadamente, en el caso de Architeuthis no existen
todavía suficientes datos que permitan inferir si realizan o no
este tipo de migraciones.
Se ha comprobado que hay muchas especies que se
desplazan de aguas superficiales a otras más profundas y
viceversa (desplazamiento batimétrico) a medida que
avanzan en su desarrollo y crecimiento. Cuando estos
desplazamientos están relacionados con la edad y el
crecimiento del animal se llaman migraciones ontogénicas, y
consisten en que generalmente las primeras fases del
desarrollo llamadas paralarvas en los cefalópodos y los
juveniles viven en zonas relativamente someras
(epipelágicas), mientras que los subadultos y adultos habitan
a mayor profundidad, ya sea entre aguas (meso y
batipelágicos) o en el fondo (bentónicos).
Fig. 24. Esquema que ilustra la migración ontogénica de un
cefalópodo. Las paralarvas nacen cerca de la costa y a medida que crecen, los juveniles migran a aguas más profundas donde crecen y
maduran para regresar a reproducirse, ya adultos, a la costa.
105

109
Quizás puede sorprender al lector que se utilice el
término paralarva para referirse a las etapas tempranas del
desarrollo y crecimiento de algunas especies de cefalópodos
en lugar de utilizar el término más clásico de larva. En
realidad el tema surge de un debate sobre la utilidad y
validez de emplear el término larva en los cefalópodos.
Young y Harman examinaron la cuestión en profundidad y
publicaron en 1988 un trabajo en el que concluyeron que el
término larva, tal y como se emplea en otros organismos,
necesitaría una ampliación tan grande para ser aplicado a los
cefalópodos, que se volvería operacionalmente inútil. Ello
debido principalmente a que el proceso por el que un
cefalópodo recién nacido se transforma en un subadulto no
requiere destrucción de tejidos ni cambios en la expresión
génica, es decir, una metamorfosis, sino meras
trasformaciones morfológicas. Así, ambos investigadores
propusieron el término paralarva para identificar a los
juveniles planctónicos que cumplen ciertos criterios
ecológicos y, en algunos casos, morfológicos. Por lo tanto,
una paralarva corresponde a aquel individuo de una especie
de cefalópodo que después de su eclosión y durante las
primeras etapas de su crecimiento es pelágico y propio de
aguas superficiales durante el día, llevando un tipo de vida
diferente al de los individuos más viejos de su propia
especie, y que carece de una metamorfosis verdadera.
Es muy poco lo que se conoce sobre las paralarvas y
los juveniles de Architeuthis. La información prácticamente
se reduce a una paralarva aparecida en muestras de
plancton, identificada por C.C. Lu como perteneciente a un
calamar gigante, pero cuya identificación no ha sido
refrendada, y a otras 14 paralarvas obtenidas en aguas de
Nueva Zelanda por S. O’Shea, empleando redes de plancton
cerca de superficie en los primeros meses de 2003, cuya
identificación tampoco ha sido refrendada por otros
especialistas.
Desde hace más de doscientos años se sabe que
bastantes especies de cefalópodos se acercan a la superficie
durante la noche desapareciendo de ella durante el día. Estos
desplazamientos verticales, a veces de varios cientos de

110
metros, han servido para agrupar a las especies según su
rango de distribución. De acuerdo con esa clasificación, que
se debe a Gilbert L. Voss (1968), los calamares gigantes del
género Architeuthis en sus fases tempranas (paralarvas y
juveniles) podrían considerarse epi y mesopelágicos; es
decir, vivirían en plenas aguas y entre la superficie y unos
700 m de profundidad, mientras que los adultos lo harían
generalmente a mayor profundidad, alcanzado cotas de hasta
1.500 metros (batipelágicos), aunque en las proximidades
del fondo y preferentemente en la cercanía de cañones
submarinos. Esto significaría que tienen una migración
ontogénica, lo que todavía está por probar.
Por otra parte, los datos proporcionados por los
registros actualmente disponibles parecen indicar que los
calamares gigantes adultos y subadultos ascienden desde
zonas relativamente profundas a capas entre 200 y 450 m de
profundidad para comer entre bancos de peces, que siguen a
su vez los movimientos diarios de sus presas planctónicas,
regidas por la luz solar. Sin embargo, este punto también es
otra incógnita.
Fig. 25. Paralarva de Architeuthis capturada en aguas superficiales de Nueva Zelanda. (S. O’Shea, 2002)

111
Capítulo 4
PRIVILEGIOS EN LAS AGUAS ASTURIANAS
El Jurásico terrestre y marítimo asturiano
El 31 de marzo de 2004 se inauguró el Museo Jurásico
de Asturias (MUJA), que está ubicado en la Rasa de San
Telmo del concejo de Colunga, a medio camino de la capital
del concejo y la localidad de Lastres. El MUJA se levanta en
un terreno alto desde el que se descubren unas vistas
magníficas del litoral oriental de la región. Desde esta
posición, en el exterior, contemplando la panorámica
marítima que se abre a nuestros ojos, pocos conocen lo que
encierran los fondos submarinos que hay tan próximos.
Los contenidos del MUJA abarcan la secuencia
temporal que transcurre por los tres periodos mayores del
Mesozoico: Triásico (250-205 millones de años antes de
nuestra era), Jurásico (205-140 millones de años) y
Cretácico (140-65 millones de años). Hay una especial
dedicación a las distintas faunas de dinosaurios durante el
Mesozoico y a las transformaciones de la geografía europea a
lo largo de esta era, con un módulo dedicado a explicar la
historia geológica de Asturias. Un periodo, el asturiano, que
contiene porciones de los 600 últimos millones de años de la
Fig. 26. Museo Jurásico de Asturias en Colunga
109

112
vida en la Tierra. Se hace hincapié en el Jurásico litoral,
desde los niveles marinos de la formación Gijón hasta los
canales representados en la formación Lastres. Entre otras
cosas, también tienen cabida los acontecimientos posteriores
al Mesozoico o los últimos 65 millones de años. Además, en
la colección fundacional del MUJA hay unos 8.000 ejemplares
de fósiles procedentes exclusivamente del Jurásico asturiano:
icnitas de dinosaurio (unas 150 huellas extraídas del litoral
de la región); restos de vertebrados (200 restos de
dinosaurios, cocodrilos, tortugas y peces); fósiles vegetales
(103 muestras de restos vegetales y 11 fragmentos de
troncos de gran tamaño); y, lo que es más interesante para
nuestra historia, elementos de invertebrados (más de 6.000
ejemplares de cefalópodos ammonoideos, y braquiópodos).
La costa jurásica asturiana venía reclamando desde
hace varios años una institución museística de este tipo. Los
investigadores también venían demandando más medios y
planificación para este tipo de estudios sobre el mejor
yacimiento Jurásico de Europa. De momento, la Costa
Jurásica está a la espera del distintivo de Patrimonio de la
Humanidad que concede la UNESCO. Sin embargo, esta
iniciativa quizá se quede corta porque cerca, entre los
cañones submarinos de Avilés, Lastres y Llanes, existen
poblaciones estables de calamares gigantes prácticamente
únicas en el mundo.
Según cuenta José Luis Sánchez Guillén, profesor de
Biología y Geología del Instituto de Enseñanza Media Pando
de Oviedo, la excavación realizada en el tramo de la autovía
del Cantábrico que une Colunga y Ribadesella bajo el Sueve
sacó a la luz un tesoro de alto valor geológico y
paleontológico. El hallazgo es de tal nivel que la Subcomisión
Internacional de Estratigrafía del Ordovícico, a propuesta de
su presidente Stan Finney, ha declarado el lugar de interés
geológico mundial. Incluso el túnel ha pasado ha llamarse
oficialmente Ordovícico del Fabar. El Ordovícico comenzó
hace 500 millones de años y finalizó hace unos 430 millones
de años. Juan Carlos Gutiérrez Marco, director del Instituto

113
de Geología Económica (CSIC-UCM) y descubridor del
yacimiento, lo califica como “el más importante del
Ordovícico del noroeste peninsular”. Fósiles invertebrados
marinos como trilobites, microfósiles y otros organismos hoy
desconocidos conforman, junto con la presencia de caolín y
del petróleo más antiguo de España, un valioso botín que ya
ha sido declarado de interés geológico internacional. Hace
460 millones de años Asturias estaba sumergida y formaba
parte de una plataforma continental de dimensiones
gigantescas que tenía una anchura de unos seiscientos
kilómetros. Aquel mundo perdido se extinguió casi en su
totalidad, desapareciendo el 90% de todas las especies que
lo habitaban, después de la glaciación de finales del
Ordovícico y de “la madre de todas las extinciones”, como
denominan los científicos a la acontecida en la tierra hace
250 millones de años, la cual dio paso a la vida tal como la
conocemos en la actualidad.
El árbol genealógico de los cefalópodos
¿Cómo eran los cefalópodos durante esas eras
geológicas? Los cefalópodos son un grupo de moluscos
bastante antiguo. Los Paleontólogos han situado su origen
hace unos 535 millones de años en el Cámbrico Medio o
Superior. Uno de los primeros grandes grupos en aparecer,
provistos de concha externa, fueron los nautiloideos, que
pudieron surgir muy tempranamente a partir de un
arqueogasterópodo de concha recta. El rasgo adaptativo
emergente más importante fue la formación de cámaras
llenas de gas dentro de la concha externa. Este desarrollo
evolutivo implicó el precintado hermético de la parte
posterior de la concha con cámaras calcáreas, que se
comunicaban con el resto del animal a través de un ramal
carnoso. Este ramal, el incipiente sifúnculo, fue fundamental
para permitir el llenado de las cámaras con un líquido. Los
restos fósiles han mostrado que los nautiloideos tuvieron una
gran radiación adaptativa en el Ordovícico (450 millones de
años), habiéndose identificado más de doscientos géneros.

114
Uno de los linajes exitosos fue el de los Bactrítidos, que
según la mayoría de los paleontólogos, fueron los
antecesores de los coleoideos y ammonoideos y cuyo primer
representante apareció hace unos 380 millones de años en el
Devónico. Los nautiloideos y ammonoideos tuvieron una
diversidad parecida durante aproximadamente 175 millones
de años en el Paleozoico, pero estos últimos fueron
superiores en riqueza de formas y abundancia durante el
Mesozoico (225-65 millones de años)
Los ammonoideos se extinguieron a finales del
Cretácico, hace unos 65 millones de años, mientras que los
nautiloideos, cuya diversidad decreció notablemente durante
el Terciario, tienen todavía un género (Nautilus) viviente que
cuenta con varias especies.
Se desconoce el origen de los grupos actuales de
coleoideos, taxón al que pertenecen todos los cefalópodos
actuales menos Nautilus, pero parece que surgieron entre el
Triásico superior y el Jurásico inferior (195-180 millones de
años). Aunque no se sabe cuando aparecieron las formas de
calamares gigantes del género Architeuthis o del género
Taningia, que habitan las fosas submarinas de Asturias, éstas
o sus predecesores directos son muy probablemente seres
del Jurásico que merecían ser considerados como auténticos
“dinosaurios marinos”.
Hay algunos zoólogos que consideran que la amplia
variación estructural de los cefalópodos vivientes, comparada
con la relativamente pequeña variación de las conchas fósiles

115
Fig. 27. Nautiloideo, (Cenoceras sp) de 11,5 cm y Ammonites
(Cruasiceras sp) de 39 cm de diámetro, ambos fósiles.
Fig. 28. Ejemplar viviente de Nautilus pompilius.
y combinado con la gran biomasa de los cefalópodos
actuales, son indicios que sugieren que actualmente nos
hallamos ante un máximo de su desarrollo evolutivo. No
obstante, esto es bastante especulativo si se considera que
hay identificadas a partir de los registros fósiles unas 10.500
especies de cefalópodos con concha externa y que se han

116
descrito al menos 28 órdenes fósiles, lo cual es mucho más
que las 700 especies y 4 órdenes de lo cefalópodos actuales.
Algunos compañeros de los cefalópodos en su
escenario evolutivo
Además de llamar la atención sobre una gran cantidad
de similitudes existentes entre los cefalópodos y los peces de
esqueletos óseos o teleósteos, Andrew Packard arguyó
persuasivamente en 1972 que su evolución ha estado
poderosamente influida por las presiones de competencia y
depredación ejercida por peces y reptiles marinos desde el
Mesozoico en adelante. Particularmente, Packard sugirió que
la radiación adaptativa de los peces y reptiles en las aguas
costeras forzó a los cefalópodos con concha, propios también
de esas áreas, a tratar de conquistar aguas más profundas y
lejanas del litoral. Una consecuencia de ello fue la pérdida de
la cocha externa con cámaras, debido a que la presión
hidrostática limitaba su desplazamiento pues la baja presión
en el interior de las conchas las hacía implosionar con la
profundidad. Subsecuentemente, algunos de los nuevos
coleoideos sin concha retornaron a aguas someras para
competir de nuevo con los teleósteos. Ciertamente, una
característica sobresaliente de la evolución de los cefalópodos
con concha externa, excepto Nautilus, es que todos se han
extinguido por lo que esta hipótesis había sido aceptada
ampliamente. Sin embargo, el descubrimiento de que los
coleoideos comenzaron a existir en el Devónico Inferior
mostró que cualquier presión por depredación sobre ellos
debió de ser ejercida por vertebrados muy primitivos, como
los peces acorazados o placodermos.
Basándose en esta y otras evidencias, en 1991
Aronson modificó el escenario descrito por Packard,
proponiendo que tanto los cefalópodos con concha externa
como los coleoideos primitivos vivieron en zonas someras
pero también en aguas profundas durante el Paleozoico y el
Mesozoico, y que posteriormente los cefalópodos con concha
externa fueron eliminados de las aguas poco profundas.
Incluso aunque el tiempo de la influencia de la depredación

117
en el escenario propuesto por Packard fuese erróneo, hay
evidencias bastante convincentes en las formas modernas de
Fig. 30. Los calamares gigantes (derecha) pueden considerarse
auténticos dinosaurios marinos, tal como los de tierra (izquierda).
los cefalópodos que la depredación puede ser un factor crítico
en la distribución de varias especies. La cuestión es que
cuando se trata de comprender las interrelaciones y
coevolución entre los depredadores y sus presas no sólo se
deben considerar características morfológicas sino también
etológicas. Tomando un ejemplo, la piel de los pulpos no sólo
implica elaborados mecanismos para el mimetismo y la
adopción de coloraciones de emergencia, sino que esos
instrumentos deben estar diseñados de manera que se
acoplen perfectamente al sistema visual de los vertebrados,
que han sido sus principales depredadores en el mar y con
los cuales han estado interactuando durante millones de
años.
La presión por competencia y depredación ejercida
por los saurios marinos desapareció prácticamente a finales
del Mesozoico, es decir hace unos 65 millones de años. Sin
embargo, los mamíferos marinos, cuyos primeros
representantes, derivaron de mamíferos terrestres parecidos
a mustélidos y úrsidos, durante el Eoceno (hace unos 55
millones de años), ocuparon paulatinamente su lugar. Las
reconstrucciones realizadas a partir de los restos fósiles
sugieren que estos animales vivían cerca de la costa y
nadaban de forma parecida a las focas actuales.

118
Fig. 30. Plesiousaurio capturando peces y cefalópodos
Rasgos principales en la evolución de los cefalópodos
El rasgo clave que aparece en la evolución de los
cefalópodos es el desarrollo de mecanismos de flotación de
baja presión. Es la inclusión de un espacio con aire dentro de
una concha lo que diferencia a los cefalópodos primitivos del
resto de los moluscos. Las consecuencias de esto fueron,
primariamente, la capacidad de regular su flotabilidad,
seguida por la interiorización y reducción de la concha, y el
desarrollo de la musculatura del manto para la locomoción
por propulsión a chorro, lo que les permitió desplazarse con
mayor rapidez y a mayores distancias que sus antecesores.
Asociado a ello aconteció el desarrollo de órganos de los
sentidos más eficaces, un aumento del tamaño y de la
complejidad del cerebro, cambios en la piel que permitieron
el mimetismo y la comunicación, todo lo que dio lugar a la
aparición de comportamientos complejos, que permitieron,
entre otras cosas, aumentar su supervivencia individual.
Los cefalópodos actuales han desarrollado un ciclo
vital con un amplio abanico de características, como sus
estrategias reproductivas, que muestran una flexibilidad
adaptativa muy importante. Todas ellas, excepto las de
Nautilus, pasan por un gradiente que va desde una freza o
desove simultáneo y masivo al final de su vida a una freza
continua durante una larga porción de su ciclo vital, pero
nunca hay ciclos múltiples de reposo y regeneración gonadal,
es decir, se reproducen una única vez en su vida, después de
la cual entran en un breve periodo de senectud y mueren,
ello debido a causas todavía no muy bien conocidas. Estas
estrategias reproductivas están adaptadas a un grado
determinado de estabilidad en el ecosistema, y representan

119
adaptaciones exitosas de cada especie a las características
de diferentes medios y presiones demográficas enfrentadas
durante los procesos evolutivos. Así, en ambientes
extremadamente inestables predominan las especies en las
que el desove es terminal y simultáneo, mientras que en los
ambientes extremadamente estables las especies presentes
tienen una freza prácticamente continua (Fig. 51).
La fecundidad de los cefalópodos, aunque muy
variable, es mucho más elevada en la mayoría de las
especies que en Nautilus, y su desarrollo embrionario y
larvario es mucho más breve. Por otra parte, las paralarvas o
estadios tempranos de los cefalópodos son bastante
diferentes de las larvas especializadas del resto de los
moluscos, que se denominan trocófora y veliger. De hecho,
los cefalópodos recién nacidos son, en esencia, adultos en
miniatura aunque sus proporciones corporales sean
diferentes. Esta ausencia de metamorfosis es otro rasgo
característico de los cefalópodos, que incrementa la
posibilidad de supervivencia individual y generacional.
Además, los cefalópodos actuales presentan unas
elevadas tasas de crecimiento y unas tasas de ingestión y de
eficiencia nutricional muy altas. Una tasa de crecimiento
elevada disminuye teóricamente la tasa de mortalidad
natural en las fases tempranas debidas a la depredación
porque los individuos son de tamaño pequeño durante menos
tiempo, reduciendo su vulnerabilidad. Por otra parte, la
supervivencia de los recién nacidos se beneficia por el
cuidado de la puesta hasta su eclosión, estrategia que han
desarrollado varias especies de octópodos. Finalmente, la
esperanza de vida de los cefalópodos actuales, excepto
Nautilus, es muy corta, no sobrepasando los dos años en la
mayoría de las especies, y nunca, aún en aquellas más
longevas, los cinco, aunque hay un trabajo reciente que
indica hasta 15 años de edad en Architeuthis sanctipauli de
unos 150 Kg. de peso corporal.
En definitiva, el estilo de vida que han adoptado la
mayoría de los cefalópodos vivientes es “vivir rápidamente y
morir jóvenes”, al contrario que la mayor parte de los peces
que crecen más lentamente y se reproducen varias veces a lo

120
largo de su vida. Es decir, mientras que los cefalópodos y la
mayoría de los peces actuales tienen estilos de vida
semejantes, poseen profundas diferencias en cuanto a su
ritmo vital.

Fig. 31. Evolución de los cefalópodos

122
Montañas en tierra y fosas en el mar
El macizo central asturiano es la región occidental de
lo que normalmente se denomina cordillera Cantábrica, que
surgió en vísperas de los fenómenos geológicos que
originaron los Pirineos. Es el sector de mayor variedad
litológica, en donde se encuentran materiales blandos, como
las calizas, y materiales cristalinos, como los granitos, las
cuarcitas y las pizarras. Esta variedad se debe a la mezcla de
elementos que se produjo en la zona de contacto entre el
zócalo y la fosa sedimentaria. La presión sufrida aquí por los
materiales fue máxima y provino del oriente. De esta manera
se formó la “rodilla asturiana”, más doblada cuanto más
hacia el este. Pero esa intensa presión no sólo dobló los
materiales, sino que los fracturó, formándose entonces un
sector de bloques elevados y hundidos que complican aún
más el relieve. Los ríos son cortos y de gran poder erosivo,
tendiendo a cortar perpendicularmente las estructuras,
aunque terminan por seguirlas. En el occidente encontramos
las sierras de Gamoniteiro, Aramo, Rañadoiro y la Bobia. En
el oriente podemos distinguir la cordillera, con sierras como
las de Pontón, Somiedo, Vegarada o Cebolledo, la fosa
prelitoral del Nalón y las sierras prelitorales de Naranco,
Suevo y Fito, y en el extremo oriental los Picos de Europa,
que son un gran bloque elevado de caliza, que ha sido
intensamente afectado por los procesos de glaciación. El
conjunto también presenta importantes formaciones
cársticas, no sólo en las cumbres sino también en las zonas
costeras. En el litoral se pueden observar hasta cuatro
superficies de abrasión. La rasa es el único elemento de
continuidad con el sector occidental. Si hacia el norte el
macizo central asturiano es escarpado, hacia el sur es
tendido, y se comunica con la meseta a través de las rañas
del norte de la cuenca sedimentaria. Los ríos más
importantes son cinco: Navia, Narcea, Nalón, Cares y Sella. Y
las mayores alturas se alcanzan en los picos de Torre
Cerredo (2.648 m) y Peña Vieja (2.618 m).
Los materiales más antiguos de Asturias se localizan
en la zona occidental. En el macropliegue con alabeamientos

123
del Narcea existen formaciones del Precámbrico, cuya
antigüedad varía entre 570-2.500 millones de años. También
se observan materiales más modernos, entre 400 y 570
millones de años, del Paleozoico inferior, que describen el
arco o rodilla astúrica, donde se encuentran los afloramientos
graníticos de Salave, Boal y El Pato.
Desde el macizo central asturiano hasta el mar el
territorio es muy estrecho y cae desde más de 2.000 metros
hasta la línea costera cuya longitud es de 334 Km. A
continuación, y ya sumergida, se extiende una estrecha y
reducida plataforma continental. Su máxima extensión es a
la altura de Tapia de Casariego, donde llega a los 50 Km. En
otros lugares es más estrecha, como en Ribadesella, San
Esteban de Pravia y Avilés, donde apenas se alcanza los 16
Km. A partir de los 200 m termina la plataforma y empieza el
talud continental, que se extiende hacia zonas abisales de
hasta 4.200 metros de profundidad.
Una innegable peculiaridad del fondo oceánico que se
extiende frente a Asturias es la existencia de tres cañones
submarinos: Avilés, Lastres y Llanes. Estas formaciones
cortan no sólo el talud continental sino que su cabecera
incide claramente en la parte externa de la plataforma,
aproximadamente para profundidades comprendidas entre
150 y 4.000 metros. El perfil transversal de esos cañones
varía desde formas en V en la parte superior hasta formas en
U en la parte inferior.
Por lo tanto, considerando los relieves terrestre y
submarino de Asturias, en un espacio de aproximadamente
50 Km. existe un desnivel de más de 6.500 metros. Los
materiales que forman las paredes de los cañones
submarinos proceden de diferentes períodos geológicos, y
tienen parangón con los que conforman el relieve emergido:
esquistos blandos, cuarcitas y granitos. Se trata de una
amplia zona donde es posible que existan bolsas de gas y de
petróleo. Al extenderse desde el borde de la plataforma
existen cambios en los fenómenos que controlan su
morfología, pasando a dominar los procesos sedimentarios
frente a los erosivos con el aumento de profundidad.

Fig. 32. Mapa donde se indican los tres cañones que se adentran en la plataforma de la costa asturiana y
el centro geográfico del caladero de pesca de Carrandi.

133
Entre el cañón de Avilés, que discurre en dirección
NO-SE, y el de Lastres, que lo hace en dirección opuesta, se
extiende una playa de pesca conocida por los pescadores
como “caladero de Carrandi”, cuyo centro geográfico queda
prácticamente en la vertical de Colunga. Este caladero
discurre durante unos 45 Km. a profundidades comprendidas
entre los 400 y 1.100 metros, sorteando pozos como el de la
Vaca. En ese caladero faenan arrastreros asturianos y
gallegos, muchos de ellos a la modalidad de pareja, en una
pesquería dirigida principalmente a la bacaladilla, que
constituye uno de los alimentos más importantes en la dieta
de Architeuthis. Son precisamente esos barcos los que
habitualmente capturan calamares gigantes, como aconteció,
por ejemplo, en septiembre de 2002, cuando la pareja de
arrastreros gallega formada por los barcos “Helena María” y
“Bautista Pino” capturó accidentalmente un macho maduro
de 42 Kg. y 98 cm de manto en las proximidades del Pozo de
la Vaca (43º54.26’N-5º28.38’W) a una profundidad
comprendida entre 450 y 475 metros.
Exploraciones acústicas
Desde 1962 se han observado 47 registros de
calamares gigantes en la costa occidental de Asturias. De
éstos, casi el 75% corresponden a animales vivos capturados
por arrastreros que faenan entre 400 y 800 m de
profundidad en el extenso caladero de Carrandi.
El ritmo natural de apariciones de calamares gigantes
en aguas de Asturias es de un registro por año. Sin embargo,
esto ha cambiado significativamente en dos ocasiones en los
últimos años. El primero de estos fenómenos aconteció entre
el 13 de septiembre y el 23 de octubre de 2001, cuando
cinco calamares gigantes se hallaron varados cerca de la
zona donde los barcos Barracuda y Nina Hay 502, trabajaron
desde principios de septiembre hasta finales de octubre,
realizando prospecciones geofísicas en busca de bolsas de
gas natural o de petróleo mediante el uso de dispositivos de
cañones de aire comprimido. Dos años más tarde, entre el 13
y 17 de septiembre de 2003, otros cuatro calamares gigantes

134
vararon o fueron recolectados moribundos flotando en
superficie en las proximidades de la costa. En esta ocasión
los cadáveres se recogieron en las cercanías de los lugares
donde se desarrolló una campaña del proyecto internacional
Marconi. Esta campaña de investigaciones geofísicas discurrió
entre el 30 de agosto y el 18 de septiembre de 2003, y en
ella se utilizaron también dispositivos con 10 cañones de aire
comprimido para producir ondas acústicas de baja frecuencia
(<100 hertzios) y de alta intensidad (200 decibelios por
cañón). En ambos casos, el objetivo de esos cañones de aire
comprimido era generar ondas sónicas capaces de atravesar
la columna de agua y varios kilómetros de subsuelo marino,
para luego recoger las ondas reflejas en los sismógrafos del
barco, y así conocer las naturaleza de los materiales de la
corteza terrestre, y la presencia o no de bolsas de gas
natural o petróleo.
Todos los ejemplares aparecidos en ambas ocasiones
pudieron ser estudiados y conservados; seis se congelaron a
-20º C inmediatamente después de su recolección y el resto
fueron fijados en formol al 10% en agua de mar. Se
realizaron las necropsias de estos nueve ejemplares de A.
dux, encontrándose que ocho de ellos eran hembras
inmaduras o en maduración de entre 67 y 140 Kg. de peso,
mientras el noveno era un macho maduro de 66 Kg. Siete de
estos ejemplares están conservados en el Aula del Mar de
CEPESMA en Luarca, otro se conserva en la sede de la
Sociedad Gallega de Historia Natural en Ferrol, y el restante
en el Museo del Mar que la Dirección General de Pesca del
Principado de Asturias tiene en Gijón.
La hembra más grande entre las estudiadas (140
Kg.), varó en la playa de Colunga el 13 de septiembre de
2003. Este ejemplar tenía un manto cuyo grosor era de 35
mm y mostraba una banda de 43 cm de ancho en la zona
central del manto con graves lesiones tisulares visibles a
simple vista. Las túnicas externa e interna de colágeno que
envuelven la musculatura del manto estaban intactas,
mientras que, por el contrario, las fibras musculares y
radiales que hay entre ambas túnicas estaban destrozadas y
convertidas en pequeños trozos. El grosor de la zona

135
afectada era de 20 mm y mostraba intacto el reticulado
romboidal de fibras de colágeno que hay entre las capas
musculares del manto.
Fig. 33. a) Lesiones en las paredes del manto y en los órganos internos de la hembra de mayor tamaño; b) Las capas de fibras musculares circulares y radiales de la pared del manto entre las túnicas externa e interna de colágeno de este ejemplar estaban
destrozadas y convertidas en pequeñas trozos.

136
Fig. 34. Fotografía mostrando una visión lateral del estatocisto de
un Architeuthis dux de 157 cm de longitud total; mac: mácula; cr:
crista y anticristas (numerados de 1 a 7); h: hámulos numerados de
1 a 5 (J.Z. Young, 1989)
Por otra parte, se observó que todos los trozos de
fibras musculares estaban embebidos en un líquido cuya
concentración de amonio era elevada. Además de la
destrucción diferencial de las paredes del manto, este animal
mostraba la mayoría de los órganos internos prácticamente
deshechos y formando un amasijo de tejidos dentro de la
cavidad paleal: los órganos del tracto digestivo tenían varias
largas y profundas llagas en distintas zonas, especialmente
en el ciego; la glándula digestiva estaba reventada, y su
contenido, un fluido parduzco de consistencia oleosa, estaba
esparcido sobre el resto de la masa informe de tejidos; la
capa externa del epitelio en columna de ambos corazones
branquiales estaba desgarrada en varias zonas, dando la
impresión de haber explosionado; las branquias estaban muy
magulladas y mostraban varios capilares rotos; el ovario
estaba roto en varios pedazos y numerosos ovocitos
desprendidos se habían esparcido por toda la cavidad del
manto. Aunque se buscaron, no se observaron signos de
congestión vascular o microhemorragias. Ello podría deberse
al relativamente pequeño volumen sanguíneo
(aproximadamente un 5% del peso corporal) y a la dificultad
de observar hemorragias en animales cuya sangre carece de
células y donde el pigmento respiratorio-la hemocianina-es
de color azul claro. Además, se observó que este animal
mostraba daños importantes en su sistema receptor del
equilibrio o estatocistos. En los receptores de gravedad había
algunas células foliculares receptoras de la mácula
gravemente dañadas, y también estaban dañadas algunas
células foliculares secundarias de las cristas del sistema de la
recepción angular de ambos estatocistos.
125

137
Las lesiones halladas en las cinco de las seis hembras
restantes y en el macho consistían principalmente en daños
severos en sus estatocistos y en el sistema microvascular de
sus branquias. Las concentraciones de Cd, Zn, Co, Cr y Ni en
el músculo del manto, glándula digestiva, corazón y
branquias de tres de estos ejemplares estuvieron dentro de
los rangos encontrados en otros cefalópodos pelágicos de
zonas poco contaminadas. En ninguno de los ejemplares
estudiados se observó una carga parasitaria de metazoos
excesiva. Finalmente, ninguno de los ejemplares mostró
signos o lesiones atribuibles a microorganismos, ni se
aislaron bacterias patogénicas en los cultivos de tejidos
obtenidos de los tres cadáveres frescos.
Fig. 35. Macho de Architeuthis dux de 66 Kg. varado en la costa
occidental de Asturias

138
Considerando el estado de madurez de los
ejemplares, la mayoría inmaduros o madurando, quedó
patente que su muerte no se produjo debido a una
mortalidad natural posterior a su único ciclo reproductivo y
tampoco se observaron indicios de otras posibles causas de
muerte natural o de captura accidental. Por el contrario,
tanto las lesiones observadas en los clamares gigantes como
la concurrencia entre el sustancial incremento de los
varamientos y la presencia de barcos realizando
prospecciones geofísicas con cañones de aire comprimido,
sugieren que la muerte de estos animales pudo estar
relacionada con el daño subletal o letal producido por el
impacto de ondas acústicas.
El impacto de la tecnología acústica marina sobre el
medio ambiente y los organismos marinos es un tema
candente, sobre todo en lo referente al efecto letal que las
ondas de presión de diferente origen y naturaleza pueden
causar en los oídos y en la activación del gas sobresaturado
presente en la sangre de los mamíferos marinos y en sus
células de diferentes tejidos, al formar microburbujas que
pueden causar embolismos en los órganos vitales. Este
asunto ha dado lugar, entre otros, al trabajo de P.D. Jepson
y colaboradores, publicado en Nature a finales de 2003, con
réplica y contrarreplica muy recientes. No obstante, como se
ha constatado en numerosas reuniones internacionales de
expertos, se conoce muy poco sobre el impacto de la
tecnología acústica marina tanto sobre los ecosistemas como
los organismos marinos, y mucho menos en el caso de los
cefalópodos.
Hasta la fecha, además de los nueve ejemplares
anteriormente citados, hemos examinado otros diecisiete
varados en diferentes partes de la costa española, además
de otros seis procedentes de Namibia, y esta es la primera
vez que se han observado el tipo de lesiones encontradas en
la mayor de las hembras. Lesiones que, por su impacto
diferencial en tejidos de distinta composición química y
microestructura, así como por la ausencia de llagas o heridas
externas en el manto, no pueden explicarse como
consecuencia de varazones o capturas accidentales.
127

139
Por otra parte, la presencia de daños en los receptores
de ondas del sistema del equilibrio nos permite plantear el
siguiente mecanismo para explicar su muerte: esas lesiones
podrían haber provocado fácilmente la desorientación en los
animales, así como una disfunción en la información sensorial
que les permite regular su posición general y la de algunas
partes de su cuerpo, en particular ante los giros. Además de
esa desorientación, las ondas acústicas podrían también
haberlos aturdido, debido a que son bastante sensibles a
vibraciones de baja frecuencia. Desorientados y aturdidos,
estos cefalópodos, moderadamente activos y con flotabilidad
neutra, podrían haber ascendido hacia la superficie, pasando
desde aguas profundas, cuya temperatura varia entre 10 y
12,5ºC, a capas de aguas más superficiales, donde la
temperatura puede superar los 15º C en la época del año en
que se produjeron los acontecimientos. Como la capacidad de
transporte de la hemocianina disuelta en la sangre de los
calamares gigantes disminuye cuatro veces cuando la
temperatura pasa de 6,4 a 15ºC, parece plausible pensar que
nuestros calamares podrían haber muerto asfixiados.
Como los calamares gigantes flotan después de
muertos y son suficientemente grandes como para que sus
cadáveres tarden en ser devorados por los carroñeros, su
presencia podría ser un indicador del impacto que ejercen las
ondas de presión acústica sobre el ambiente y la fauna
marina de profundidad. Por otra parte, los cadáveres
hallados en Asturias quizá sólo sean la punta de un iceberg
que esté señalando efectos importantes sobre otras especies
marinas, algunas de ellas de importancia comercial.
Es indudable que para conocer a fondo los efectos de
los pulsos acústicos de diferente intensidad y frecuencia en
los organismos marinos, es necesario incrementar los
estudios físicos, fisiológicos y etológicos. Sin embargo, ante
la ausencia de suficientes conocimientos y la presencia de
muchas incertidumbres, es recomendable adoptar una
actitud precautoria. No se trata tanto de prohibir este tipo de
técnicas, cuya utilidad es grande, sino que todos los agentes
implicados acepten y afronten el problema. Consideramos
que se debería ser muy cauto a la hora de utilizar tecnología

140
acústica hasta que su impacto en los ecosistemas y
organismos se conozca suficientemente. También se debería
abrir un debate transparente y profundo, concienciando lo
más posible a la sociedad, con el mejor lenguaje posible, y
principalmente dirigido a que se cree la capacidad necesaria
para que se produzca el desarrollo científico apropiado en el
uso de estas técnicas. Todo ello orientado a que se
establezcan los mecanismos adecuados para que los
gobiernos correspondientes presten la debida atención a este
asunto.
¿Por qué aparecen tantos calamares gigantes en
Asturias?
Es difícil hallar explicaciones precisas a esta pregunta.
Una respuesta obvia es porque Architeuthis dux habita en
las proximidades de las costas asturianas. Lo que todavía se
desconoce es si efectúa allí todo su ciclo vital, o, por el
contrario, únicamente pasa una fase de su vida. Hasta ahora
no se han encontrado puestas, ni recién nacidos, ni juveniles
sino hembras inmaduras o en maduración (nunca
completamente maduras) y dos machos maduros. El motivo
de que no se hayan capturado paralarvas o ejemplares de
menor tamaño puede ser triple: por una parte que no se han
buscado, por otra que los aparejos de pesca de los
arrastreros son excesivamente grandes, y finalmente que los
pesqueros no faenan en los lugares donde moran esos
individuos. Otros tres factores a considerar son que la
mayoría de las capturas y subsiguientes varamientos se
produjeron principalmente en otoño, que se capturaron en
copos con abundante bacaladilla juvenil, y que los animales
presentaban estómagos vacíos o con muy escasos restos de
contenidos, siendo estos fundamentalmente de peces
teleósteos.
La proximidad de los cañones submarinos parece ser
un factor importante para explicar la presencia de
Architeuthis en aguas asturianas. Esto mismo es lo que
acontece en Nueva Zelanda, donde la mayoría de los

141
Fig. 36. Hembra inmadura de 70 Kg. de A. dux capturada por un
pesquero en la zona más occidental del caladero de Carrandi en
octubre de 2002. (Foto de Eloy Alonso)
calamares gigantes son capturados en las proximidades del
cañón de Kaikoura. Por otra parte, hay que considerar que el
hábitat debe ser idóneo para Architeuthis en lo referente a
las condiciones oceanográficas reinantes así como en la
abundante presencia de las presas potenciales con
aceptables niveles de capturabilidad.
Conocer algo de la biología de la que parece ser la
presa principal de los calamares gigantes en las aguas
asturianas nos puede llevar a una mejor comprensión de los
motivos por los que están allí estos cefalópodos. La
bacaladilla o lirio (Micromesistius poutassou) es un pez
marino de la misma familia que la merluza y el bacalao, es
decir, un gádido.
Esta especie alcanza hasta 50 cm de longitud total y
un peso máximo de 850 g. pudiendo llegar a vivir hasta 20
años. Es un pez pelágico cuyo rango de profundidad varía
entre 150 y 3000 m, siendo sobre todo frecuente entre 300 y
400 metros de profundidad. Su dieta se compone
fundamentalmente de pequeños crustáceos, pero los
individuos grandes también comen peces y cefalópodos.

142
Efectúa migraciones diarias verticales, apareciendo cerca de
la superficie durante la noche y más próxima al fondo
durante el día. Esta especie es propia de zonas templadas y
su área de distribución es muy amplia extendiéndose desde
79°N hasta 26°N en latitud y desde 67°W a 40°E de
longitud, o lo que es lo mismo: en el Atlántico nororiental
desde la parte sur del mar de Barents y las costas Noruegas
hasta alrededor de Islandia por el norte, y por el sur hasta el
cabo Bojador en las costa africana; en el Atlántico
noroccidental abarca desde el sur de Groenlandia hasta la
costa noreste de los Estados Unidos de América; es también
común en el Mediterráneo y el mar Negro.
El área de desove más importante en el Atlántico
noreste se localiza al noroeste de Irlanda, y la freza acontece
en primavera. Las larvas y los juveniles se dispersan por una
gran área, siendo la zona meridional del golfo de Vizcaya una
zona de alimentación y crecimiento, donde abundan los
juveniles de entre 18 y 22 cm de longitud, que forman
cardúmenes de varios kilómetros de extensión, distribuidos
en agregados discretos a lo largo de toda la costa. Tiene
importancia comercial ya que sólo en las aguas europeas se
Fig. 37. La bacaladilla, Micromesistius poutassou.
131

143
pescan más de un millón y medio de toneladas anuales. Se
captura al arrastre con artes de pesca cuya apertura de boca
puede alcanzar hasta unos 4.000 metros cuadrados. Esas
artes trabajan próximas al fondo o entre aguas, y la
operación puede hacerse con una o dos embarcaciones,
como en el caso de las parejas en Asturias. Suele darse un
lance de pesca diario de entre 8 y 10 horas de duración a 2,5
nudos de velocidad. El volumen de agua que pasa a través
de la red en cada lance es enorme, variando entre 120 y 160
millones de metros cúbicos.
La presencia de juveniles de bacaladilla frente a las
costas asturianas está relacionada con, al menos, dos
fenómenos: en primer término, con la existencia de
corrientes de agua templada que transportan las larvas y
juveniles desde el área de puesta hasta estas costas, y en
segundo lugar a la presencia de abundante alimento, lo que
está ligado a la existencia de un afloramiento de aguas
profundas y ricas en nutrientes que se produce en
primavera-verano. Este enriquecimiento de las aguas
favorece la producción, primero de fitoplancton, y luego, de
presas abundantes y asequibles para la bacaladilla juvenil,
Fig. 38. Esquema de una red de pesca de arrastre tirada por un
solo barco.

144
fundamentalmente crustáceos pelágicos. Esto acontece hacia
el otoño, que es la época del año en que más intensa es la
explotación de la especie y cuando se producen el mayor
número de capturas de Architeuthis.
Un escenario plausible de lo que ocurre con los
calamares gigantes, independientemente de que tengan o no
un área de desove en aguas asturianas, es que estos
animales pueden vivir ocultos y protegidos en zonas
profundas de los cañones submarinos (1.000-1.500 m) y
ascender por ellos hasta la plataforma (250-600 m) para
alimentarse. Aunque todavía no existen imágenes de cómo
se nutren, en razón de sus características anatómicas y de lo
observado en otros calamares, se supone que éstos gigantes
deben situarse próximos o dentro de un banco de bacaladilla
alargando sus tentáculos para capturar a sus presas una por
una. Luego de apresado, el pez es llevado hacia la boca,
donde es matado de una fuerte y certera dentellada en las
proximidades de su bulbo raquídeo, y después, engullido en
pequeños trozos, nunca entero, con la ayuda de la rádula.
Esta operación debe llevarles algún tiempo al que además
hay que agregarle que deben comer aproximadamente entre
un 5 y un 10% de su peso diariamente. La consecuencia es
que deben gastar bastante tiempo en alimentarse. Los
calamares gigantes deben ser bastante certeros a la hora de
capturar a sus presas, ya que cualquier demora o deficiencia
en su captura representa un gasto energético inaceptable,
por lo que estos cazadores deben valerse muy
probablemente, de su gran agudeza visual, muy necesaria en
aguas de luz tenue y difusa, y de su capacidad de detectar la
luz polarizada, que les proporciona una imagen muy precisa
de sus potenciales presas.

145
Capítulo 5
LO QUE ACTUALMENTE CONOCEMOS SOBRE LA VIDA DE LOS CALAMARES GIGANTES
Algunas características generales de los cefalópodos
Los cefalópodos se sitúan entre los animales más
hermosos de la naturaleza y su comportamiento es complejo
y fascinante. Actualmente existen unas 700 especies
vivientes, muy pocas si se las compara con las miles de
especies de otros grupos zoológicos. Sin embargo, su bajo
número es compensado por su gran abundancia en los
océanos. Los cefalópodos poseen una enorme capacidad de
adaptación y elasticidad para ajustarse a todo tipo de
ambientes y ecosistemas marinos, por lo cual su forma
corporal y sus estilos de vida son muy variados. El
cefalópodo adulto más pequeño (Idiosepius) mide
aproximadamente 8 mm de longitud, mientras que el más
grande (Architeuthis) alcanza hasta cerca de dos y medio
metros de longitud corporal, hasta 22 de longitud total,
siendo el mayor invertebrado que existe actualmente. Por
otra parte, hay un pulpo pigmeo, Octopus micropyrsus, que
escasamente alcanza los 50 gramos de peso, mientras que el
pulpo gigante del Pacífico septentrional, Octopus dofleini,
crece hasta alcanzar cerca de los 100 Kg. El típico pulpo que
uno acostumbra ver, y en muchas ocasiones comer,
corresponde a especies que viven en hábitat intermareales y
submareales, pero existen otras especies como los
denominados pulpos cirrados (Opisthoteuthis y Cirrothauma,
por ejemplo), así como Vampyroteuthis, que viven en las
aguas profundas de los océanos, generalmente por debajo de
los 3.000 m de profundidad, adaptados completamente a
dichos ambientes. También hay cefalópodos, como el pulpo
Vulcanoctopus hydrothermalis, que moran en ambientes
extremos como lo son las fuentes hidrotermales oceánicas

146
que se encuentran a más de 2.000 m de profundidad y
donde la actividad volcánica genera un ecosistema único con
temperaturas que oscilan entre los 2ºC y los 300ºC y aguas
tan llenas de metales pesados y elementos volcánicos que
otras especies peor adaptadas morirían rápidamente.
Fig. 39. Cuadro esquemático mostrando la clasificación de los cefalópodos actuales (Clase Cephalopoda).

147
Aunque los cefalópodos habitan desde el Ecuador
hasta ambas regiones polares, no se ha encontrado ninguno
en el mar Báltico, el mar Negro, ni en aguas dulces. Esto se
debe a que los cefalópodos no están capacitados a vivir en
aguas de baja salinidad. Sin embargo, gracias a su enorme
adaptabilidad, algunas especies como el calamar Lolliguncula
brevis han colonizado aguas salobres, soportando salinidades
de hasta 17 partes por mil. En cuanto a su forma de vida,
muchos cefalópodos, como por ejemplo los omastréfidos o
potas, viven en las capas superiores de los océanos, donde
compiten con los peces por el alimento y son presa de aves y
mamíferos marinos. Estos cefalópodos, tienen el cuerpo en
forma de torpedo, son rápidos nadadores y activos
depredadores a diferencia de las especies que viven a
profundidades intermedias, cuya naturaleza es delicada y
poco activa al grado de tener que retener cloruro amónico en
algunas cavidades o en los tejidos de su cuerpo para poder
flotar sin esfuerzo. Esto ocurre en gran cantidad de especies
de la región subantártica, donde son tan abundantes que
constituyen el alimento básico de los cachalotes. En otras
regiones, los cefalópodos constituyen pesquerías de gran
importancia económica.
Los calamares gigantes del género Architeuthis son
cefalópodos pertenecientes al filo Mollusca, es decir,
invertebrados parientes de organismos tan conocidos como el
mejillón, las ostras, las almejas o el caracol. A su vez, entre
los cefalópodos pertenecen a la subclase que agrupa a
aquellos que tienen concha interna (Coleoidea), y dentro de
ella se ubican en el orden de los verdaderos calamares
(Teuthida) y del suborden que engloba a los que carecen de
córnea en sus ojos (Oegopsida), que es un suborden
extremadamente polimórfico y compuesto de numerosas
familias que ocupan una gran variedad de hábitats.
Forma y función
El cuerpo de todos los cefalópodos puede dividirse en
dos partes principales: la anterior, que se denomina
cefalopáleo y está formada por la cabeza, una corona de

148
apéndices móviles (los brazos y tentáculos) y el sifón, y la
posterior o visceropáleo, que consiste en un saco muscular o
manto, en cuyo interior se alojan las branquias y las
vísceras; el espacio vacío que existe entre ellas recibe el
nombre de cavidad paleal.
La cabeza está bien separada del manto por una
constricción nucal y en su base, por el lado ventral, se
localiza el sifón, que es un embudo cónico, estrecho por la
parte anterior y ancho en la posterior, que corre paralelo al
eje longitudinal del cuerpo. En Architeuthis, los márgenes
laterales inferiores del sifón están conectados al manto
mediante un cartílago de cierre del sifón con el manto recto y
sencillo, que funciona como un broche.
Fig. 40. Esquema de un Architeuthis (A), el brazo modificado
para la reproducción en machos, llamado hectocotilo (B) y la
maza tentacular (C).

149
En la cabeza, y rodeando a la boca, el calamar gigante
tiene diez extremidades: ocho brazos y dos tentáculos. Los
brazos se numeran desde el par dorsal al ventral: el primer
par es el dorsal, el segundo el dorsolateral, el tercero el
ventrolateral y el cuarto el ventral. En ellos se pueden
observar dos filas de ventosas pedunculadas que poseen
anillos de dientes córneos para lograr un mejor agarre de su
presa. Los brazos derecho e izquierdo del mismo par suelen
ser semejantes en longitud.
Los tentáculos se localizan entre los brazos
ventrolateral y ventral y consisten en un largo pedúnculo o
funículo que termina en una expansión apical, la maza
tentacular, que posee cuatro filas de ventosas, las dos de en
medio de mayor tamaño que las laterales. En esta última se
distinguen tres partes: la anterior o dáctilo, la media o mano
y la posterior o carpo. En el carpo hay numerosas ventosas y
almohadillas. A lo largo del funículo se alternan almohadillas
y ventosas en dos filas, lo que representa una de las
principales características de este género.
Fig. 41. Maza tentacular de Architeuthis.

150
Fig. 42. Tentáculo de Architeuthis donde se aprecian las dos filas de ventosas y almohadillas alternadas entre si que hay a lo largo del funículo.
El manto es un saco muscular fusiforme que contiene
los órganos internos. Está constituido por tres capas
musculares y la piel, todas bajo control nervioso. La
musculatura del manto es la responsable de los movimientos
respiratorios y del sistema de propulsión a chorro de los
cefalópodos. La piel tiene numerosas bolsas y células con
pigmentos responsables de la coloración y de los cambios de
color del animal.
Los calamares gigantes poseen un par de aletas en el
extremo del manto, de pequeño tamaño con los márgenes
posteriores cóncavos. Probablemente estas aletas cumplan
una función más bien de estabilización que de propulsión.
Como en todos los cefalópodos, los sexos en el
calamar gigante están separados: siempre hay machos y
hembras. Aunque en larvas y juveniles es difícil diferenciar
entre ambos sexos, a medida que el animal crece y va
madurando aparecen diferencias morfológicas claras entre
machos y hembras. Estas diferencias, que permiten
distinguirles externamente, se denominan dimorfismo sexual
y se deben al desarrollo de los caracteres sexuales

151
secundarios. Las hembras de Architeuthis alcanzan tamaños
más grandes que los machos, y, por otra parte, ambos
brazos ventrales de éstos experimentan modificaciones
morfológicas con la maduración, convirtiéndose en
estructuras especializadas para el traspaso de los
espermatóforos (paquetes de esperma que produce el
macho) durante la cópula. Cada una de las porciones
modificadas de estos brazos se denomina hectocotilo. En
Architeuthis, sin embargo, la hectocotilización característica
de los machos es muy leve, consistiendo en que algunas
ventosas apicales se transforman en laminillas casi cubiertas
por los márgenes del brazo que se ensanchan y se engrosan.
Esto se debe muy posiblemente a que en Architeuthis el pene
está muy desarrollado, sobresaliendo varios centímetros por
fuera del manto en los machos adultos, por lo cual parece
probable que lo utilicen para traspasar directamente los
espermatóforos a la hembra, función realizada por el
hectocotilo en numerosas especies.
Algo que llama la atención inmediatamente es que
Architeuthis posee unos tentáculos extremadamente largos y
que estos no son retráctiles, es decir que no pueden meterse
en el interior de una bolsa, como ocurre por ejemplo en las
sepias. Considerando que una hembra adulta puede llegar a
tener un cuerpo que mide unos dos metros y medio de
manto, más una cabeza de 50-80 cm. de largo, pero que su
longitud total puede alcanzar de 18 a 20 metros, esto
significa que el animal poseería unos tentáculos de entre 14
y 16 m. de longitud.
¿Por qué el calamar gigante posee unos tentáculos tan
largos? Como todos los cefalópodos, Architeuthis es un
depredador y lo poco que se conoce sobre su dieta indica que
consume peces y otros cefalópodos. Como todos sus demás
congéneres, Architeuthis utiliza sus tentáculos como
extensiones con las que atrapan a sus presas. Una vez que
las tienen sujetas con los tentáculos, las atraen y las
envuelven con los brazos para comenzar a comerlas. Debido
a su gran tamaño y al enorme gasto energético que debe
representar desplazar un cuerpo tan grande como el suyo, lo
más probable es que Architeuthis no sea un buen nadador. Si

152
esto es así, los tentáculos serían utilizados para capturar a
sus presas a distancia, evitando así el tener que moverse
rápido para acercarse a ellas. Esta sería una razón de
economía energética, pero se pueden aducir también razones
de estrategia cazadora, según la cual el cuerpo del cazador
permanece relativamente lejos de la presa, que no sospecha
ser alcanzada por tan largas extremidades. Pero todo esto
son solo suposiciones, ya que nadie todavía ha visto a un
calamar gigante cazando.
La forma de los cefalópodos está relacionada con su
hábitat y su conducta. De ella se deduce que los Architeuthis
son organismos que, pese a ser relativamente poco activos,
se desplazan por la columna de agua. En este sentido, la
forma hidrodinámica del calamar gigante le permite
desplazarse por el agua con la mínima resistencia. El manto
del calamar gigante es fusiforme, y se estabiliza por las
aletas durante el desplazamiento. Además, cuando nadan,
los cefalópodos juntan sus brazos y tentáculos
extendiéndolos a lo largo del eje del cuerpo para reducir la
resistencia al agua. Una vez conseguida la forma
hidrodinámica, ¿cómo se impulsa el Architeuthis? Sus aletas
son demasiado pequeñas como para ser eficientes
propulsores, por lo que el impulso principal para la natación
lo proporciona la expulsión de agua a presión por el sifón.
Architeuhis, como la mayoría de los cefalópodos utilizan la
propulsión a chorro como medio de propulsión desde
millones de años antes que el hombre siquiera existiera. El
agua penetra en la cavidad paleal por el hueco que hay entre
la cabeza y el manto, siendo luego expulsada a través del
sifón, cuya función durante la natación es la de concentrar y
dirigir el chorro de agua, logrando un impulso efectivo en
sentido contrario al que el agua es expulsada. Para ello, la
musculatura del manto se contrae y, junto con comprimir el
agua que hay dentro de la cavidad paleal, cierra los bordes
de manto mediante dos broches que se denominan cartílagos
de cierre del sifón, lo que impide que el agua salga por donde
había entrado obligándola a salir por el sifón, cuya forma de
embudo flexible y la gran lengüeta interna que hay en
Architeuthis regulan tanto la presión como la dirección del

153
chorro permitiendo al animal desplazarse en la dirección
deseada.
Anatomía interna
Las descripciones de los órganos internos de
Architeuthis existentes en la bibliografía son muy escasas. El
sistema digestivo de Architeuthis se abre en la boca, situada
en el bulbo bucal, en la base de los brazos. La boca se halla
provista de dos mandíbulas semejantes al pico de los loros y
de una rádula. Las mandíbulas, o pico, son dos, una inferior
y otra superior, y están constituidas por un material fibroso
muy duro (queratina). Sus bordes libres son afilados y
cortantes, mientras que los demás están insertados a
poderosos músculos. Pueden cortar fácilmente carne e
incluso romper caparazones de crustáceos, conchas y huesos
de los animales que constituyen sus presas.
La rádula es una larga cinta sobre la que están fijados
dientes minúsculos dispuestos en hileras transversales de
siete dientes cada una. Su función es parecida a la de una
lengua rasposa: arrancar partes carnosas de estructuras
duras y arrastrarlas hasta la entrada del esófago.
En el bulbo bucal desembocan tres tipos de glándulas
salivales: sublingual, anteriores y posteriores. La función de
la glándula salival sublingual se desconoce todavía con
precisión. El par de glándulas salivales anteriores secretan
principalmente mucosidad, mientras que las glándulas
salivales posteriores, que aquí son muy pequeñas, secretan
enzimas digestivos además de sustancias tóxicas con las que
paralizan, o matan, a sus presas.
La boca se continúa en el esófago, cuya luz es
estrecha y no se puede dilatar porque pasa a través del
cerebro y del cartílago craneal. Por esa razón, estos gigantes
no pueden tragar grandes presas sino que deben trocearlas
previamente, como ocurre en el resto de los cefalópodos. A
continuación del esófago se abre el estómago, que es muy
largo, extendiéndose hasta el final de la cavidad paleal, y
cuyo color externo es blanco. Sus paredes son musculosas
y

154

Fig. 43. Anatomía general interna de una hembra de Architeuthis según Roper y Voss, 1982

219
Fig. 44. Mandíbula superior (a) e inferior (b) de un Architeuthis.
está tapizado internamente por una cutícula plisada, igual
que acontece en el esófago. Después del estómago existe un
ciego que es un saco arrollado en espiral, cuyas paredes
carecen de cutícula y son un epitelio mucoso y ciliado. Entre
el estómago y el ciego desembocan dos conductos provistos
de apéndices -los conductos digestivos- provenientes de la
glándula digestiva. A continuación del ciego viene el
intestino, que se abre en el ano, situado en la parte ventral
anterior de la cavidad del manto, próximo al sifón.
En conjunto, el digestivo está dispuesto en forma de
U. En el ano hay dos lóbulos grandes o dos conspicuas
papilas anales. Forma parte también de este sistema la bolsa
de la tinta. Esta estructura es una especialización de la
glándula rectal que se ha transformado en una glándula y
bolsa de tinta, que se abre cerca del ano. En Architeuthis la
bolsa de la tinta es relativamente pequeña. La tinta está
formada por melanina, pero también contiene otras
sustancias, como tirosina, que irrita los ojos y paraliza
temporalmente los órganos olfatorios de los depredadores.
Apenas se sabe nada de cómo es y cuanto dura la
digestión en los calamares gigantes. Sin embargo, como
ocurre en otros cefalópodos oceánicos, se conoce que son
carnívoros y muy voraces, pudiendo comer al día hasta un
10% de su peso corporal. Las enzimas digestivas son

220
Fig. 45. Fotografía electrónica de la rádula de un
Architeuthis formada por 7 filas de dientes y dos filas de
placas marginales.
elaboradas principalmente en la glándula digestiva, y la
digestión se realiza en el estómago, aunque puede
completarse de forma intracelular en dicha glándula, que
representa entre el 3 y 5% de peso total del animal. La
duración de la gestión a temperaturas entre 10º y 12ºC, que
son las reinantes en el hábitat propio de estos animales,
debe durar más de doce horas. La absorción se realiza sobre
todo en el ciego y en la glándula digestiva. En el intestino
apenas hay absorción y su función principal consiste en la
formación de los excrementos; como ocurre en otras
especies, los restos deben ser envueltos en abundante
mucosidad.
Como todos los cefalópodos, Architeuthis debe tener
un catabolismo proteico, es decir, queman principal y
primariamente proteínas, en lugar de usar lípidos, que dan
más energía por masa consumida. Este sistema es pues
antieconómico y también peligroso ya que las proteínas son
el componente somático más importante, pero tiene una
ventaja que es aprovechada por estos, y otros calamares,
que consiste en producir amoniaco como producto final de la
combustión. El amoniaco unido a otros iones, como los de
cloro, da lugar a sustancias menos densas que el agua de
mar. La acumulación de esas substancias ricas en amonio en
determinadas cavidades del cuerpo o en el músculo
proporciona una flotabilidad neutra a los cefalópodos que las
poseen. Y eso es lo que acontece en Architeuthis, donde el
músculo del manto y de los brazos presenta un sistema
vacuolar lleno de productos de esa naturaleza,
proporcionándoles un sistema de flotación económico. Por
otra parte, la glándula digestiva exuda una sustancia líquida
de color parduzco con una elevada concentración de lípidos.
El amonio, los lípidos y que el cartílago de la cabeza sea
reticulado en lugar de compacto, como ocurre en otras
especies de cefalópodos, sugieren con relativa contundencia

221
que estas grandes criaturas pueden flotar durante largo
tiempo entre aguas sin apenas esfuerzo muscular. Sin
embargo, la alta concentración de amonio en los tejidos
musculares hace que su carne sea incomestible, y también
que su putrefacción sea muy rápida y con un olor
característico.
Esta condición de animales con flotabilidad neutra
conduce a pensar que se trata de criaturas con poca
capacidad para la natación. Lo que parece corroborar el
pequeño tamaño de sus aletas, así como la relativa debilidad
del aparato cartilaginoso de cierre del sifón con el manto,
que es indispensable para la natación a reacción, cuando el
animal expulsa el agua contenida en la cavidad del manto a
través del sifón.
Architeuthis tiene, al igual que la mayoría de los
cefalópodos, un sistema circulatorio evolucionado y con una
organización compleja, que difiere del sistema circulatorio de
otros moluscos (como caracoles, ostras, babosas, almejas,
etc.) por la existencia de una extensa red de arterias y de
venas y por presentar un gran número de capilares, en lugar
de lagunas abiertas donde se mezcla la sangre arterial con la
venosa produciendo un intercambio poco eficaz,
característico de la mayoría de los moluscos. Este sistema
circulatorio consta de un corazón principal o sistémico con
dos aurículas y un ventrículo. En las aurículas desembocan
las venas branquiales eferentes, mientras que del ventrículo
parte la vena aorta, con ramificaciones a las vísceras,
órganos de la cabeza y brazos. El corazón sistémico, muy
musculoso, impulsa la sangre oxigenada en las branquias a
través de las arterias. La sangre vuelve de los capilares por
un sistema venoso complejo: las venas abdominales llevan la
sangre procedente de las vísceras; las venas paleales, la del
manto; y la vena cava anterior, la de la cabeza. Todas estas
venas desembocan en los vasos branquiales eferentes, que
confluyen a su vez en un par de corazones branquiales, que
impulsan la sangre a las branquias, donde es oxigenada por
un mecanismo de contra-corriente entre la sangre y el agua
inhalada en la cavidad paleal. Las branquias de Architeuthis
son muy largas, alcanzando un tercio de la longitud del

222
manto del animal. Cada branquia está formada por entre 50
y 70 laminillas branquiales por hemibranquia. Estas
corresponden a finas láminas altamente capilarizadas donde
se produce el intercambio gaseoso. La sangre vuelve a las
aurículas a través de las venas branquiales eferentes. Dado
que la sangre circula por venas, arterias y corazones, se
trata de un sistema circulatorio cerrado, que es característico
de los cefalópodos y, a la vez único, pues no existe en
ningún otro grupo de moluscos.
El oxígeno es transportado por la hemocianina, una
proteína con cobre que circula en la sangre en forma de
agregados moleculares (no hay hematíes) y se origina en las
glándulas branquiales, que están adheridas al par de
corazones branquiales. Aunque la capacidad de absorción del
oxígeno es alta (superior al 50%), la de transporte en la
sangre es baja (inferior al 4,5%), por lo que Architeuthis,
como los demás cefalópodos, precisa bombear un gran
volumen de sangre por unidad de tiempo. En comparación
con un pez de su tamaño, necesita bombear ocho veces más
sangre para moverse la mitad de rápido. Aun así, y pese a
que debe tener unas tasas metabólicas muy altas, el oxígeno
no suele ser un factor limitante de la actividad. Sin embargo,
como ya se dijo, la capacidad de transporte de la
hemocianina, que disminuye cuatro veces de lo normal
cuando la temperatura aumentaba de 6,4 a 15º C, si lo
puede ser.
En Architeuthis no hay verdaderos riñones ya que
como en todos los cefalópodos, su sistema excretor está
compuesto de varias estructuras relacionadas con los
sistemas respiratorio y circulatorio: los apéndices de los
sacos renales, las glándulas pericárdicas anejas a los
corazones branquiales, los apéndices renopericárdicos, los
apéndices renales de la vena cava, las branquias, y, en
alguna medida, la glándula digestiva. El producto principal de
la excreción es el amoníaco y las superficies branquiales
constituyen una vía importante para la eliminación de los
desechos nitrogenados. La orina se origina por un proceso de
filtrado, y parte de sus componentes son reabsorbidos en los
apéndices renales de la vena cava y en las branquias. El

223
líquido renal de Architeuthis, como el de otros cefalópodos,
es un medio idóneo para la proliferación de diferentes tipos
de parásitos.
Todos los órganos internos de Architeuthis están
cubiertos por una membrana gruesa de color rojizo que está
unida a la cara interna del manto. Además de encerrar y
proteger los órganos, esa membrana encierra una cavidad
muy peculiar, que se denomina celoma, cuya función es
proteger al corazón, almacenar tanto los productos de
desecho como los genitales.
El celoma se divide en dos secciones: la pericárdica y
la genital, unidas ambas por el conducto gonopericárdico. En
la primera sección se alojan básicamente el corazón
sistémico, los branquiales y las glándulas pericárdicas,
mientras que la segunda contiene el estómago, el ciego,
parte del intestino y las gónadas.
El sistema reproductor de las hembras de Architeuthis
consta de un ovario, dos oviductos y dos glándulas
nidamentarias. El ovario se sitúa en la parte posterior de la
cavidad del manto y en el se forman los ovocitos. Se han
observado ejemplares de Architeuthis de Asturias
(probablemente A. dux) en los que el ovario representaba el
0,4% del peso total de la hembra cuando ésta era todavía
inmadura, mientras que éste se incrementó hasta un 2,1%
en tres hembras en maduración y en hembras maduras de
otras regiones el ovario alcanza entre el 5 y 8% de su peso.
La función principal de las glándulas nidamentarias es la
formación de las envolturas que rodean a los huevos tras la
cópula.
La fecundidad potencial de la hembra en maduración
de 81 Kg capturada en Asturias se estimó en nueve millones
doscientos mil ovocitos. Esta estima concuerda con las
realizadas en una hembra casi madura de 168 Kg hallada en
las costas de Escocia, cuyo ovario pesaba 7676 g donde se
calcularon diez millones de ovocitos. El tamaño de los
ovocitos en las hembras de Asturias todavía en proceso de
maduración varió entre 0,2 y 0,4 mm, valores inferiores a los
observados en una hembra de 200 Kg capturada en
Sudáfrica, donde la dimensión de los ovocitos varió entre 0,5
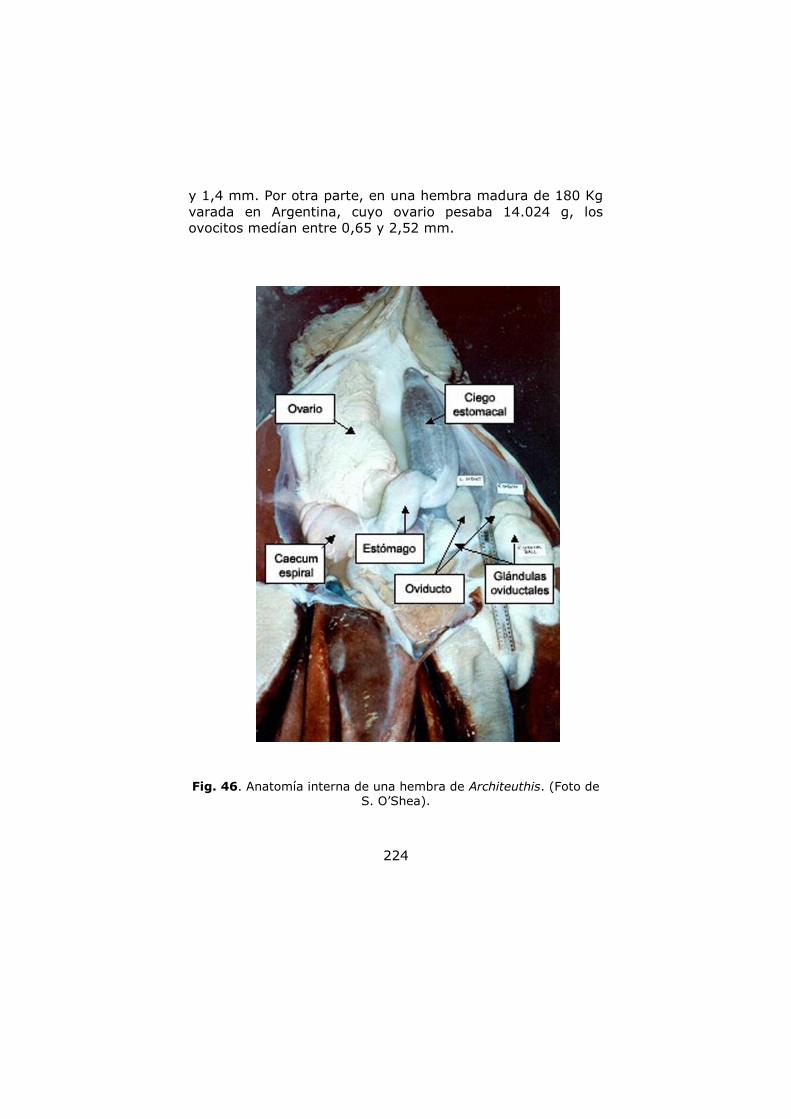
224
y 1,4 mm. Por otra parte, en una hembra madura de 180 Kg
varada en Argentina, cuyo ovario pesaba 14.024 g, los
ovocitos medían entre 0,65 y 2,52 mm.
Fig. 46. Anatomía interna de una hembra de Architeuthis. (Foto de S. O’Shea).

225
Se desconoce cómo es el desarrollo ovárico y también
si la hembra realiza una o varias puestas. Sin embargo, dada
su alta fecundidad potencial y las limitaciones de capacidad
del oviducto, es de suponer que se realicen varias puestas en
vez de una sola. Los ovocitos maduros migran del ovario a
los oviductos y durante su desplazamiento desde el extremo
próximal al distal pasan por las glándulas oviductales. Es
muy probable que las glándulas secreten substancias
químicas que induzcan la activación y atracción de la
esperma a los ovocitos. Al llegar al extremo distal del
oviducto los ovocitos son, probablemente, expulsados a la
cavidad paleal donde las secreciones de las glándulas
nidamentarias los recubren con una gelatina, formando la
puesta. Teóricamente, debería ser en este momento cuando
se produce la fecundación de los ovocitos. Una vez formada
la puesta, es expulsada de la cavidad paleal por la
contracción del manto.
La forma de la puesta de Architeuthis es totalmente
desconocida hasta la fecha, pero se supone que será una
masa flotante de huevos, semejante a una gran esfera o a un
cilindro transparente. Como la gelatina absorbe agua, la
puesta podría dilatarse y llegar a tener un diámetro cercano
a uno o dos metros. Esta puesta flotaría a media agua
arrastrada por las corrientes marinas hasta la eclosión de las
paralarvas. Aunque hay limitada información sobre la
especie, es muy probable que la puesta tenga estas
características ya que se han observado puestas similares
pertenecientes a otras especies de cefalópodos oceánicos.
Las especies asociadas a la plataforma y talud continental,
como los pulpos y los calamares, realizan puestas que son
fijadas al fondo, pero parece poco probable que Architeuthis,
aunque habite ambientes asociados al talud y cañones
submarinos, tenga esta estrategia en particular. Sea como
sea, es muy poco lo que se conoce sobre este aspecto de la
biología del calamar gigante y hablar más al respecto es pura
especulación.
150

226
En la figura 25 se ilustró un ejemplar de tres
centímetros que parece ser un juvenil de Architeuthis. Sin
embargo, los recién eclosionados de esta especie, a juzgar
por el tamaño de los ovocitos maduros no superaría los 2,5
mm de longitud. Estas paralarvas, parecidas al adulto
aunque diferentes en sus proporciones corporales, deben
formar parte del zooplancton durante algún tiempo, siendo
arrastradas pasivamente por las corrientes. A medida que el
animal crece, aumenta su capacidad para migrar en la
columna de agua y para nadar activamente contra las
corrientes, pasando a ser un juvenil y adoptando
paulatinamente el hábitat de los adultos. Aunque se
sospecha que la mortalidad natural en los estadios
tempranos del desarrollo de Architeuthis debe ser muy
elevada, este es otro aspecto que se desconoce por
completo.
Un último punto interesante es que parece existir una
notoria desproporción entre el número de machos y de
hembras que se han encontrado en todos los océanos:
generalmente hay tres veces más hembras que machos. Ello
puede indicar que, o hay efectivamente una mayor
proporción de hembras que de machos, o que ambos sexos
viven en hábitat diferentes durante una parte de su ciclo
vital, siendo el hábitat de las hembras más expuesto a las
capturas que el de los machos, pero aquí también nos
adentramos en terrenos especulativos.

227
Fig. 47. Macho maduro Architeuthis de 60 Kg. de peso capturado
en Asturias. (Foto Eloy Alonso).
En los machos, los espermatozoides producidos por el
testículo son empaquetados y rodeados por membranas,
dando lugar a los espermatóforos. La longitud de los
espermatóforos depende del tamaño del macho, pero está
comprendida entre 110 y 150 mm. Las glándulas
espermatofóricas y accesorias son las responsables de las
secreciones que sirven de cemento para aglutinar los
espermatozoides así como de la formación de sus
membranas. Estas glándulas se unen al testículo a través
del conducto seminal deferente. Los espermatóforos
completamente formados se almacenan en el saco
espermatofórico o bolsa de Needham, de la cual salen a
través del conducto seminal aferente y del pene. Cuando
desaparece la cápsula del espermatóforo, su órgano
eyaculador se evagina y se liberan los espermatozoides para
fecundar a los ovocitos.
Los machos maduran a tamaños muy inferiores que
las hembras; se han registrado machos completamente
maduros entre 61 y 110 cm de LM. Las hembras alcanzan la
madurez en un amplio rango de tamaño.
En el Atlántico nordeste únicamente se han registrado
ocho machos desde 1952 hasta la fecha, todos ellos
funcionalmente maduros, y tres de ellos (Dinamarca, 1957,
Asturias, 2002 y Portugal, 2002) con espermatóforos
embebidos en la piel en varios de sus brazos y en el manto.
Actualmente se desconoce cual es el origen de estos
espermatóforos. Podría tratarse de una inyección de esperma
de otro macho, que en las obscuras aguas donde moran
estos animales, se confundió de sexo. Podría deberse
también, a una auto-inseminación accidental producida
cuando el macho copulaba con una hembra. Esta última es la
explicación más plausible porque los machos de esta especie,
mucho más pequeños que las hembras, poseen un verdadero
y conspicuo pene, que utilizan para colocar los paquetes de
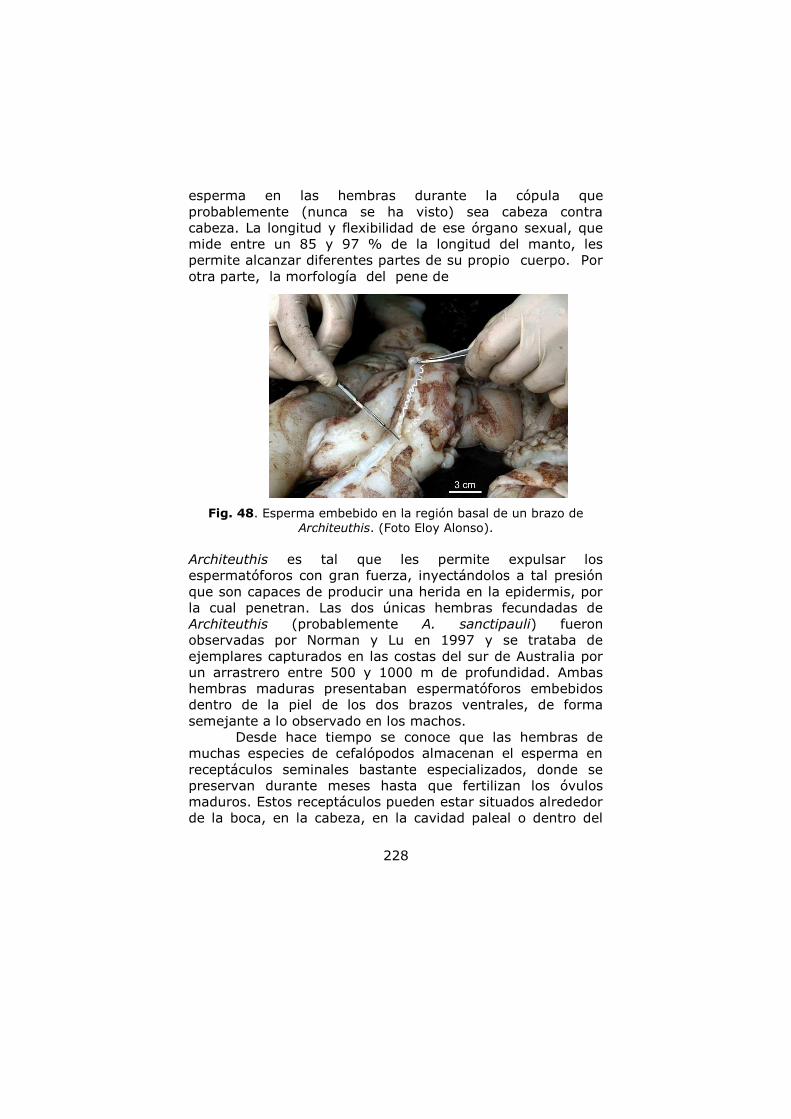
228
esperma en las hembras durante la cópula que
probablemente (nunca se ha visto) sea cabeza contra
cabeza. La longitud y flexibilidad de ese órgano sexual, que
mide entre un 85 y 97 % de la longitud del manto, les
permite alcanzar diferentes partes de su propio cuerpo. Por
otra parte, la morfología del pene de
Fig. 48. Esperma embebido en la región basal de un brazo de
Architeuthis. (Foto Eloy Alonso).
Architeuthis es tal que les permite expulsar los
espermatóforos con gran fuerza, inyectándolos a tal presión
que son capaces de producir una herida en la epidermis, por
la cual penetran. Las dos únicas hembras fecundadas de
Architeuthis (probablemente A. sanctipauli) fueron
observadas por Norman y Lu en 1997 y se trataba de
ejemplares capturados en las costas del sur de Australia por
un arrastrero entre 500 y 1000 m de profundidad. Ambas
hembras maduras presentaban espermatóforos embebidos
dentro de la piel de los dos brazos ventrales, de forma
semejante a lo observado en los machos.
Desde hace tiempo se conoce que las hembras de
muchas especies de cefalópodos almacenan el esperma en
receptáculos seminales bastante especializados, donde se
preservan durante meses hasta que fertilizan los óvulos
maduros. Estos receptáculos pueden estar situados alrededor
de la boca, en la cabeza, en la cavidad paleal o dentro del

229
tracto reproductor. Por otra parte, hay otras especies en las
que los machos producen en el manto de la hembra heridas
externas y depositan allí sus espermatóforos. Esas heridas
pueden ser producidas por las mandíbulas o por los garfios
en que se transforman algunas ventosas de los brazos o de
las mazas tentaculares. Cuando esto ocurre, los machos de
estas especies tienen normalmente penes largos y
musculosos, cuya morfología sugiere que pueden usarse para
insertar los espermatóforos a presión en esas heridas. Sin
embargo, en Architeuthis no se han observado este tipo de
heridas externas en el cuerpo o brazos. De tal forma que el
mecanismo de inserción del esperma en las especies de
Architeuthis resulta bastante particular.
Otro aspecto importante de la reproducción de estos
calamares es cómo se realiza la fecundación de los ovocitos.
En la mayoría de los cefalópodos éstos son fecundados
cuando salen del oviducto (o de los oviductos, que a veces
son pares). En estos casos el esperma se encuentra
almacenado en receptáculos especiales próximos a la boca o
los espermatóforos están insertados, no embebidos, en la
cara interna del manto de la cavidad paleal. En esos lugares,
la hembra manipula el esperma y lo aproxima a los ovocitos,
y es entonces cuando el sistema eyaculador del
espermatóforo expulsa los espermatozoides de la cápsula
que conforma el espermatóforo, penetrando uno de ellos en
cada ovocito antes de que las sustancias secretadas por las
glándulas nidamentarias que lo rodean se vuelvan
impermeables al contacto con el agua. En Architeuthis el
problema es cómo llega el esperma embebido en los
tegumentos de los brazos o del manto de la hembra a entrar
en contacto con los ovocitos. En 1997, Norman y Lu
propusieron varias hipótesis para explicar este
mecanismo:

230
Fig. 49. Esquema de un espermatóforo. a: aparato eyaculador. ft:
filamento terminal. glc: glándula de cimentación. me: masa espermática. te: tubo eyaculador.
primero que la propia hembra usa sus mandíbulas, o las
ventosas, para desgarrar la piel sobre los espermatóforos y
liberarlos; en segundo lugar, que la piel de la hembra podría
experimentar algún proceso degenerativo en los lugares de la
implantación justo antes de la puesta, lo suficientemente
intenso como para que los espermatóforos queden al
descubierto; en tercer lugar, que los espermatóforos tengan
capacidad de migrar hacia la superficie de la piel, e incluso
desplazarse hasta el interior de la cavidad paleal cerca de la
salida del oviducto para encontrarse allí disponibles cuando
salen los ovocitos y fecundarlos.
El sistema nervioso de los cefalópodos es el más
desarrollado de todos los invertebrados y, en consecuencia,
origina unos patrones de comportamiento bastante
complejos, aunque variables según el hábitat al que cada
especie está adaptada. Se pueden reconocer masas
comparables a los ganglios cerebral, pedial, pleural y visceral
del resto de los moluscos, pero éstos han perdido su
integridad como ganglios individuales, se han subdividido y
se han reunido en un centro nervioso que rodea al esófago,
el cual funciona como un cerebro o sistema nervioso central
relativamente complejo. Comparado con su tamaño, el
cerebro de Architeuthis es relativamente pequeño; en un
ejemplar de 96 cm de longitud del manto y unos 90 Kg. su
cerebro apenas representó el 0,01% de su peso, presentando
además dos lóbulos ópticos de peso similar (0,01%). En otro
ejemplar (157 cm de longitud y unos 110 Kg. de peso) el
volumen del cerebro apenas representó el 0,008 % del
volumen del cuerpo, mientras que el volumen de los lóbulos
ópticos fue también del 0,008%.
En el cerebro existe una verdadera capa de neuronas
corticales que recubren un complejo neuronal más profundo.
Se pueden distinguir varias vías y conductos organizados,

231
también hay centros o lóbulos definidos que ejercen control
sobre determinadas actividades. Las respuestas a los
estímulos externos son parcialmente determinadas gracias a
las asociaciones neuronales específicas, pero los centros de
asociación de la corteza cerebral son capaces de
realizar
Fig. 50. Esquema del sistema nervioso central de Octopus vulgaris
en visión dorsal (A) y lateral (B). bm: lóbulo basal medio. bi: ganglio bucal inferior. bs: lóbulo bucal superior. cs: lóbulo
cromatofórico posterior. fim: lóbulo frontal inferior medio. fi: lóbulo frontal inferior. fs: lóbulo frontal superior. ma: masa subesofágica
anterior. mag: lóbulo magnocelular. nb: nervios braquiales. nia: nervio infundibular (sifonal) anterior. noc: nervio ocular. nof: nervio
olfatorio. np: nervio paleal. ns: nervio del estatocisto. nv: nervio visual. olf: lóbulo olfatorio. op: glándula óptica. ped: lóbulo
pedunculado. to: tracto óptico. vas: lóbulo vasomotor. vert: lóbulo vertical. (Young, 1971).
discriminación y aprendizaje. El cerebro consta de cuatro
partes principales: supraesofágica, subesofágica, lóbulos
ópticos y ganglio de la boca y de los brazos, todas ellas
interconectadas por numerosas comisuras. En Architeuthis la
parte que se sitúa por encima del esófago representa un 35,5
% del volumen total del cerebro y consta principalmente de

232
los ganglios cerebrales y de sus subdivisiones (lóbulo frontal
inferior, superior y vertical); no obstante, un par de nervios
pasan hacia el complejo bucal, que consta de un anillo
nervioso que rodea ganglios bucales superiores e inferiores
unidos entre sí. La región del cerebro que se halla por debajo
del esófago consta de ganglios (o lóbulos) pedios, pleurales
magnocelular y viscerales pares y de sus correspondientes
subdivisiones. Los lados derecho e izquierdo se han soldado
parcialmente formado una masa simétrica lobulada. Los
ganglios de los brazos y del sifón derivan de los pedios,
mientras que los pleurales y viscerales se han fusionado
formado una masa simple, generalmente denominada
ganglio visceral. Los nervios de los brazos, sifón, manto y
vísceras parten de la región subesofágica, donde también se
encuentran los centros de control de los músculos del ojo, el
iris, la pupila, de la actividad de los cromatóforos (lóbulo
cromatofórico), de los movimientos respiratorios inhalantes y
expelentes, de los movimientos de los brazos, manto y aletas
y de la actividad de distintos órganos viscerales.
Mientras que la región subesofágica es sobre todo un
centro motor, la supraesofágica está compuesta
predominantemente por centros de retención y transmisión
de estímulos sensoriales. Los nervios olfatorios y ópticos
entran en este nivel y los centros sensoriales que provocan
respuestas específicas a los estímulos sensoriales también se
hallan implantados en esta región del cerebro. La porción
cerebral supraesofágica contiene, además, una zona de
asociación. En Architeuthis, los lóbulos de los brazos
(braquiales) constituyen el 29,4 % del volumen total
cerebral, el lóbulo pedio el 14,1% y los lóbulos
paleoviscerales el 18,4%. El lóbulo magnocelular contiene
neuronas gigantes y recibe los estímulos sensoriales que las
activan. Cada una de estas neuronas se entrecruza con la del
otro lado y establece uniones con las neuronas gigantes de
segundo orden del ganglio visceral. Estas a su vez pasan a lo
largo de los nervios paleales hasta los ganglios estrellados
del manto, donde forman una unión sináptica con las
neuronas gigantes de tercer orden, cuyos axones inervan los
músculos del manto. En Architeuthis estos axones, o fibras

233
nerviosas, son ocho troncos que se dividen en nervios
menores inervando la parte anterior del manto, mientras la
musculatura paleal central y la posterior está inervada por
unas 18 ramificaciones del nervio mediano, que llega hasta
las aletas.
Las fibras gigantes de esos nervios no son
especialmente largas, pero existen fibras nerviosas gigantes
que proceden directamente del cerebro. En las fibras
gigantes el impulso nervioso se transmite a una velocidad
cinco veces mayor que las fibras nerviosas normales. La
conexión nerviosa entre cabeza y manto se produce a través
de los ganglios estrellados que están ubicados a unos 150
mm del borde anterior del manto y a 65 mm de su línea
media. Estos tienen forma de riñones alargados y en un
ejemplar adulto miden 22 x 12 mm, lo que es un tamaño
relativamente pequeño en comparación con el animal.
Al cerebro se conectan distintos órganos sensoriales.
Los ojos, que por su similitud con los de los vertebrados son
un claro ejemplo de convergencia adaptativa, son los
órganos de los sentidos más patentes y los mayores de
cualquier animal conocido. En los ejemplares de mayor
tamaño pueden alcanzar hasta 25 cm de diámetro. En el
caso de Architeuthis carecen de córnea (ojo oegópsido),
disponiendo de un cristalino, una pupila ajustable en el
centro de un iris pigmentado y de células pigmentadas
retinianas.
Los ojos del calamar gigante están adaptados a un
hábitat donde la luz es muy tenue o inexistente. Por
experimentos realizados en el laboratorio se ha determinado
que los calamares comunes son ciegos a los colores, pero se
desconoce si esto rige o no para los calamares gigantes. Los
lóbulos ópticos junto con sus correspondientes cuerpos
blancos, cuya función está relacionada con el sistema
inmunológico de estos animales, están embebidos en una
masa grande de tejido situada detrás de los ojos.
El hábitat donde mora el Architeuthis es un mundo
acuático submarino situado a gran profundidad donde apenas
llega luz solar. Es un mundo oscuro donde la supervivencia
de muchas especies depende de su capacidad de detectar o

234
generar luz y de permanecer camuflado en la escasa luz
ambiental. En este hábitat, el calamar gigante ha
desarrollado uno de los más completos y eficientes sensores
visuales del reino animal.
La luz penetra muy poco en los océanos, ya sea
porque es reflejada o absorbida. Cuando la luz del sol incide
sobre la superficie del mar, un 5% es reflejada y el resto se
transmite siendo rápidamente absorbida por el agua y por los
componentes químicos y orgánicos en suspensión. El agua
absorbe todas las ondas luminosas, excepto de las
correspondientes al color azul y verde, que corresponden a la
escasa luz que llega a las profundidades.
Los ojos del calamar gigante representan un
sofisticado sistema óptico para localizar a sus presas. Este
ojo, además, es el de mayor tamaño entre los animales del
planeta, hecho que podría explicarse en parte por el hábitat
del calamar gigante. Así, el tamaño de los ojos en
animales

235
Fig. 51. Fibras nerviosas gigantes del manto de Architeuthis (Nixon y Young, 2003)
Fig. 52. Estructura del ojo de un Architeuthis.
con actividad diurna, que viven en lugares iluminados, es
pequeño aunque el animal sea de grandes dimensiones.
Sencillamente, no es necesario un tamaño mayor para
satisfacer las necesidades de estos animales ya que la
densidad de elementos sensores en la retina es suficiente en
un ojo de unos pocos centímetros de diámetro para obtener
resoluciones altísimas. Por otra parte, en animales de
actividad nocturna o hábitat con muy poca luz, se han
desarrollado ojos un poco más grandes que en los animales
diurnos para desenvolverse perfectamente en la oscuridad.
En las grandes profundidades marinas apenas llega la
luz solar existiendo la débil luz generada por la
bioluminiscencia. Estos niveles de luz pueden haber
favorecido el desarrollo del ojo de Architeuthis, el cual es un
extraordinario órgano receptor fotomultiplicador. Su retina,

236
que tiene unos 25 cm. de diámetro, sirve para detectar la
escasa luz existente, pues la energía captada por varias
células se suma para obtener una respuesta significativa que
permite al animal ver con niveles de luz que representan una
total oscuridad para la mayoría de las especies que se
encuentren en el mismo nivel oceánico.
La retina del Architeuthis contiene aproximadamente
mil millones de unidades receptoras, frente a los ciento
treinta y dos millones de la retina humana. Esto le
permite
Fig. 53. La luz es absorbida al pasar por entre las capas de agua y
sólo las longitudes de onda azules llegan a las profundidades.
multiplicar su sensibilidad, respecto a nuestra retina, en casi
ocho veces manteniendo la misma resolución, lo cual le
resulta muy útil para cazar a las profundidades en que
habita.
Otros órganos sensoriales muy importantes en los
cefalópodos, que se localizan en la región occipital del
cráneo, son los estatocistos, los cuales corresponden a un
par de estructuras complejas en cuyo interior se alojan los
estatolitos, dos estructuras calcáreas duras de cristales de

237
aragonito además de materia orgánica. Los estatolitos tienen
unos 2 mm de longitud y su interior muestra anillos de
deposición rítmica, gracias a los que se puede determinar la
edad de estos organismos. Los estatocistos de Architeuthis
son de un tamaño relativamente pequeño en relación con el
animal. En los estatocistos (Fig. 35) hay dos sistemas
mecanorreceptores: mácula-estatolito y crista-cúpula. El
receptor de aceleraciones lineales o de la gravedad del
estatocisto -el sistema mácula-estatolito- es bastante amplio
debido a lo alargado de la mácula, pero también estrecho.
Este sistema utiliza información sobre la dirección de la
gravedad y la aceleración lineal para regular la orientación de
la cabeza, ojos y cuerpo, así como para controlar los reflejos
de la coloración de contra-sombreado producida por los
cromatóforos ventrales que tienen muchas especies de vida
pelágica. El sistema crista-cúpula proporciona al cerebro
información sobre la aceleración rotacional necesaria para
regular la posición de la cabeza, el sifón y los ojos del
animal. El sistema crista-cúpula de Architeuthis está
constituido por siete crestas protuberantes y cinco
oquedades. La estructura peculiar de este estatocisto es
parecida a la de otros cefalópodos con flotabilidad neutra, y
también sugiere que Architeuthis realiza los giros
lentamente. Nada se sabe sobre si los calamares gigantes
pueden oír o no, aunque esa capacidad parece demostrada
en otros cefalópodos y se realiza en los estatocistos, capaces
de detectar sonidos de baja frecuencia.
Se conoce muy poco sobre otros órganos sensoriales
de estas especies, aunque deben existir numerosos
receptores de presión, quimiorreceptores, receptores de la
posición de sus órganos internos y también fotorreceptores
distribuidos en diferentes partes del cuerpo.
Otra estructura rígida y dura presente en los
calamares gigantes es el gladio o pluma. Se ubica en la
región central más interna de la cavidad del manto, estando
adosada por el interior a los músculos de su pared dorsal y
envuelta en una membrana traslúcida delgada.
El grosor de las paredes del manto hacen que el
gladio no se note dorsalmente, pero confiere cierta rigidez a

238
todo el cuerpo en sentido longitudinal. Se extiende desde una
pequeña expansión triangular presente en el margen dorsal
del manto hasta la punta de la pequeña prolongación en
forma de cola que hay en su parte posterior. Su naturaleza
es quitinosa, con una zona central o raquis no muy
pronunciado ni robusto. La pluma posee dos anchas
expansiones laterales o palas en la zona media, que se
aguzan hacia los dos extremos, con el posterior terminado en
un endocono poco conspicuo. Esta concha residual interna es
bastante frágil sobre todo cuando se deshidrata, y aparece
con mucha frecuencia rota en los animales varados.
Estrategia reproductiva
Los cefalópodos son un grupo con una gran diversidad
de estrategias y comportamientos reproductivos. En muchas
especies, sobre todo costeras, el macho y la hembra adoptan
aptitudes exhibicionistas con importantes cambios en los
componentes cromáticos, texturales y posturales que
determinan la elección y la formación de la pareja para la
cópula. Aunque en la mayoría de las especies se forman
parejas para la reproducción esto no es óbice para que, en
muchos casos, las parejas cambien y una misma hembra
copule con varios machos. En varias especies, además de las
típicas disputas entre dos machos por una misma hembra, se
ha observado que durante el cortejo nupcial hay machos
intrusos o furtivos, que están a la espera que el macho
dominante abandone a su pareja para copular con la hembra,
eliminando previamente el semen del macho anterior. Este
comportamiento, que asegura que sea ese macho el que
realmente fecunde a la hembra, se denomina competencia
espermática.
Para ser un grupo con un número relativamente
pequeño de especies, los cefalópodos presentan una gran
diversidad de estrategias reproductivas que, en el fondo, son
un indicativo de su plasticidad a la hora de adaptarse a sus
diferentes hábitats. Así, cada especie posee adaptaciones
propias en su reproducción las que, atendiendo a factores

239
como la maduración de los ovocitos, el tipo de puesta y si
hay o no crecimiento entre las diferentes tandas de huevos,
se pueden agrupar dentro de cinco tipos de estrategias
reproductoras: 1) Cefalópodos que ponen una única vez en
su vida con una puesta terminal y simultánea, donde la
maduración de los ovocitos es sincrónica, la puesta
monocíclica, y los huevos son frezados en una sola tanda. En
este caso, todos los individuos frezantes morirían casi al
mismo tiempo después de una auténtica orgía sexual de toda
la población. Esto se ha observado, entre otras especies, en
el calamar de California Loligo opalescens. 2) Cefalópodos
que ponen varias veces durante su vida: a) Puesta policíclica,
donde hay varios ciclos de maduración de los ovocitos a lo
largo de la vida del animal. Esta estrategia reproductora se
ha observado únicamente en las especies de Nautilus; b)
Puesta múltiple, en la cual los ovocitos maduran por tandas y
la puesta es monocíclica, pero los huevos son puestos en
varias tandas distribuidas a lo largo de un periodo que
representa sólo una fracción de la vida del cefalópodo,
durante el cual se produce crecimiento somático entre cada
tanda de huevos. Esta estrategia reproductora es típica de
varios omastréfidos como la pota Sthenoteuthis oualaniensis;
c) Puesta terminal intermitente, la cual es muy semejante al
caso anterior pero sin que se produzca crecimiento somático
entre las tandas de huevos. Esta estrategia es típica de la
pota voladora Illex coindetii y de los calamares Loligo
vulgaris y L. forbesi, entre otras especies; d) Puesta
continua, en la cual se produce la maduración asincrónica de
los ovocitos, por lo cual la hembra,

240
Fig. 54. Estrategias reproductivas en cefalópodos y su relación con
el ambiente. (Rocha et al., 2001)
una vez que ha madurado, realiza una puesta monocíclica en
la cual va poniendo continuamente huevos a lo largo de un
extenso periodo de tiempo, que representa una importante
fracción de su ciclo de vida, y durante el cual hay crecimiento
somático. Esta estrategia se produce en especies que habitan
ambientes muy estables y es típica en pulpos de profundidad
como Opisthoteuthis agassizii.
En el caso del calamar gigante, hay una escasa
evidencia que indique el tipo de estrategia reproductiva que
este posee. La elevada fecundidad de las hembras y el
hábitat relativamente estable en que viven podrían indicar un
tipo de estrategia de desove múltiple o terminal fraccionada.
Sin embargo, careciendo de mayores datos sobre su biología
y ecología, intentar definir mejor su tipo de estrategia
reproductiva es hipotetizar.
Edad y crecimiento
Entre las cuestiones que más intrigan de los
calamares gigantes están las relacionadas con su edad y su
crecimiento. ¿Cuánto tiempo tarda un calamar gigante en

241
alcanzar longitudes de 20 metros y pesos de hasta media
tonelada? La cronobiología, o rama de la biología que estudia
la edad y el crecimiento de los seres vivos, tiene como uno
de sus principales retos el desarrollar técnicas y métodos que
permitan determinar la edad de organismos que son, en
muchos casos, difíciles de mantener en cautividad debido a
su particular ciclo de vida, como es el caso de los
cefalópodos. Estos estudios han sido motivo de infinidad de
investigaciones en los cefalópodos, no solamente en los
denominados gigantes, sino en muchas de las
aproximadamente 700 especies vivientes que habitan
nuestros mares en la actualidad. La determinación de la edad
y del tipo de crecimiento de cualquier organismo permite,
entre otras cosas, comparar las diferentes estrategias
reproductivas, alimenticias y ecológicas de una especie en las
diferentes fases de su ciclo biológico. Además, ambos
parámetros son necesarios para evaluar los cambios que se
producen en la biomasa de las poblaciones, ya sea por
causas naturales o por efecto de la pesca, es decir, son
necesarios para conocer la estructura demográfica de las
poblaciones.
En todos los organismos se observan cambios
morfológicos y fisiológicos a lo largo de su ciclo vital, que
suelen ser más bruscos en las etapas iniciales de su
desarrollo, y los cefalópodos se rigen por esas mismas
pautas. Después de la fecundación, el huevo que se forma
tiene abundante vitelo o alimento y el embrión ocupa un
pequeño disco en su polo animal. El tipo de segmentación
embriogénica de los cefalópodos es espiral, a diferencia de la
que se produce en los demás moluscos, lo que constituye un
caso único de desarrollo dentro de este grupo zoológico.
Después de un período más o menos extenso, dependiente
de la temperatura, el embrión del cefalópodo se ha
desarrollado por completo, siendo capaz de romper la capa o
capas externas del huevo y eclosionar. El recién nacido suele
ser como un adulto en miniatura, aunque difiere de él en sus
proporciones corporales, es decir, en los cefalópodos el
desarrollo embrionario es directo, no existiendo estadíos
larvarios que requieran una verdadera metamorfosis para

242
llegar a ser adultos, sino únicamente un crecimiento
diferencial de sus partes corporales o crecimiento alométrico.
Por ello, el término clásico de larva no es aplicable a este
grupo de moluscos, habiéndose acuñado, como ya se dijo, el
de paralarva para denominar a los estadíos postembrionarios
de numerosas especies, entre ellas las de Architeuthis.
Los cefalópodos son moluscos de crecimiento rápido,
regulado por factores bióticos y abióticos. Dichos factores
influyen notablemente en su crecimiento, dando lugar a
distintas tasas de crecimiento individual. Las diferentes
condiciones ambientales, sobre todo la temperatura, varían
el ritmo de crecimiento entre individuos de la misma edad,
acentuándose esta influencia en zonas en las que existen
marcadas diferencias estacionales, dependiendo de la época
del año en la que se realice la puesta.
El cálculo de la edad a partir de estudios detallados en
los cefalópodos puede decirse que comenzaron a
desarrollarse a finales de los años sesenta. Actualmente, a
pesar de que se ha avanzado en gran medida en las
estimaciones directas e indirectas de la edad en estos
moluscos, es mucho todavía lo que queda por conocer, sobre
todo en relación a los métodos a emplear y en cuanto a las
conclusiones generales sobre el crecimiento que se extrae de
las diferentes técnicas empleadas.
Para determinar la edad y el crecimiento de los
cefalópodos se han utilizado dos tipos de metodologías
diferentes: En primer lugar los métodos indirectos, que son
aquellos que se basan en el desplazamiento de los valores
modales de las distribuciones de tallas en el tiempo (método
de Petersen). Este método fue el único empleado
sistemáticamente en cefalópodos durante casi 90 años. Sin
embargo, la técnica sólo es útil cuando la especie tiene uno ó
varios períodos de desove cortos, bien definidos y cuando
además no acontecen cambios poblacionales debidos a
migraciones, por lo que dadas las peculiares características
biológicas de los cefalópodos, este método indirecto no
resulta de gran utilidad en el estudio de sus poblaciones
cuando se trata de estimar su edad y crecimiento. En
segundo término están los métodos directos, donde se

243
distinguen tres tipos diferentes según la técnica utilizada: a)
aquellos que se basan en observaciones directas de
individuos mantenidos en cautividad bajo condiciones
controladas. Aquí se suman otras técnicas que se están
utilizando actualmente como métodos para estimar la tasa
instantánea de crecimiento en un organismo, mediante el
cálculo de la razón ARN/ADN en tejidos, técnica que hasta
ahora únicamente se ha realizado con animales en
cautividad; b) los que se basan en datos de captura-
recaptura empleando técnicas de marcado en la naturaleza;
y c) los que se realizan a partir del análisis de estructuras
que presentan marcas de crecimiento. Estos últimos se basan
en la asunción de que estas estructuras se forman
rítmicamente, y que la distancia entre las marcas de
deposición o incrementos, es proporcional al crecimiento del
animal. En peces estos métodos se han aplicado en
vértebras, escamas, huesos, espinas y, sobre todo, en
otolitos. Para la determinación de la edad a partir de
estructuras duras con incrementos de crecimiento periódico
se requiere que dichas estructuras constituyan un registro
permanente de los diferentes acontecimientos de la vida del
animal, manifestados por la deposición diferencial rítmica de
materia orgánica e inorgánica.
Las estructuras duras disponibles en los cefalópodos
coleoideos actuales son escasas, y no todas son útiles, como
es el caso de la rádula que es una estructura donde la
determinación de la edad no es posible porque se desgasta
continuamente en un extremo, reemplazándose por el otro.
Otras estructuras son las mandíbulas ó picos, las cuales han
recabado notable atención al observarse y contarse en ellas
ciclos regulares de incrementos en varias especies, como los
de la enoploluria Moroteuthis ingens, aunque todavía no se
ha encontrado ninguna periodicidad en su formación.
Estudios recientes han indicado que se podía calcular en el
pulpo común Octopus vulgaris, una edad aproximada a partir
de la longitud de la cresta de su mandíbula superior, ya que
se descubrieron en ella marcas depositadas con cierta
periodicidad acompasadas con el crecimiento. Sin embargo,
la utilización de estas estructuras como un parámetro fiable

244
que estime la edad se encuentra todavía en un estado
bastante incipiente. En tercer lugar está la pluma o gladio,
que es una estructura quitinosa de soporte con una
morfología variable según la especie, situada en el músculo a
lo largo de la línea dorsal del manto en los calamares. Varios
investigadores, principalmente rusos, desarrollaron una
técnica basada en diferentes cortes del gladio, mediante la
cual se evidenció la existencia de líneas periódicas de
crecimiento en varias especies, entre ellas en Kondakovia
longimana e Illex argentinus. A partir de estas líneas de
crecimiento pudieron estimarse el número de incrementos,
que, relacionados con la longitud dorsal del manto,
proporcionaron altas y positivas correlaciones.
El problema remanente era conocer la periodicidad
con que se formaban las líneas de crecimiento observadas en
las plumas. Fue a principios de los años noventa cuando se
comenzó a pensar que esos incrementos tenían una
periodicidad diaria, pero la validación no se realizó en el
gladio sino en otra estructura dura, el estatolito, como
más
Fig. 55. Incrementos de crecimiento en la pluma de Sthenoteuthis
oualaniensis. (Bizikov, 1991)
adelante se verá. Posteriormente, se realizó un estudio
comparativo de la edad, analizando los estatolitos y el gladio
de diferentes especies de calamares (Loligo y Sepioteuthis)

245
observándose que había una significativa concomitancia, por
lo que el estudio de los incrementos de deposición en el
gladio pasó a considerarse un método útil y adecuado para la
determinación de la edad en los cefalópodos. El gladio está
considerablemente modificado en algunas especies, así en la
familia Sepiidae (sepias o chocos) es de naturaleza calcárea
y recibe el nombre de sepión. Este se trata de una concha
calcárea interna que sirve, entre otras cosas, para regular la
flotabilidad del animal. En un estudio realizado en 1960 se
encontró un período de nueve días en la formación de dos
lamelas sucesivas en el sepión de Sepia esculenta. Otros
estudios posteriores postularon la existencia de una
periodicidad diaria de deposición en diferentes especies de
choco (Sepia officinalis, S. subaculeata y Sepiella maindroni).
Sin embargo, un reciente y cuidadoso trabajo realizado con
Sepia officinalis mantenidas en cultivo a diferentes
temperaturas demostró que el número de lamelas del sepión
no se corresponde con la edad de los individuos y que la
periodicidad de la deposición varía con la temperatura de
cultivo, lo que invalida esta estructura para estimar la edad.
Los ojos de la mayoría de los cefalópodos están provistos de
cristalinos. Aunque estas estructuras crecen por añadido de
capas concéntricas, y se observó que el peso del cristalino
del calamar Loligo pealei se incrementaba linealmente con
respecto al peso corporal y a la edad del animal, todavía no
se ha desarrollado una técnica adecuada que permita usar
los cristalinos en la estimación de la edad.
La última estructura dura que tienen los cefalópodos
son los estatolitos, que consisten en un par de concreciones
calcáreas formadas por cristales de aragonito alojadas en dos
cavidades adyacentes llamadas estatocistos, las cuales se
encuentran situadas ventro-posteriormente en el interior del
cartílago cefálico y contienen un complicado sistema de
receptores. Estas cavidades se pueden definir como órganos
de situación espacial o equilibrio en invertebrados, aunque en
los cefalópodos sirven además para detectar aceleraciones
lineales y angulares. La primera ilustración publicada sobre la
forma de un estatolito de cefalópodo pertenecía

246
precisamente a la de un calamar gigante del género
Architeuthis.
Fig. 56. Estatolitos dentro de sus estatocistos. (Clarke, 1998)
La microestructura y composición química de los
estatolitos se conocen relativamente bien y son las
estructuras más utilizadas para la determinación de la edad y
del crecimiento en los cefalópodos. Los cristales que
componen la mayoría de los estatolitos de los cefalópodos se
depositan de dos formas diferentes: el primer patrón de
deposición se localiza en el ala, en la porción ventral de la
cresta dorsal y en la porción media del rostro. El segundo
patrón tiene una orientación casi paralela al eje longitudinal
de los cristales que tienen el núcleo como origen, y se
encuentran en la cúpula, en el lóbulo lateral, la mayor parte
del rostro y la región central. El primer tipo de cristalización,
altamente desordenado, dificulta la observación de bandas
de crecimiento, mientras que en el segundo es posible
observar marcas que corresponden a incrementos de

247
deposición periódica, donde se alternan materia orgánica y
carbonato cálcico.
Los estatolitos son las estructuras más utilizadas para
la determinación de la edad y crecimiento en los cefalópodos.
El primer intento que determinó satisfactoriamente la edad a
partir de los estatolitos fue realizado en el calamar Loligo
opalescens de California, encontrándose unos incrementos
que se consideraron de periodicidad diaria, y otros más
espaciados y próximos al rostro de los estatolitos que se
consideraron de periodicidad mensual. Desde esta primera
aproximación, realizada hace ya casi treinta años, la técnica
de preparación y lectura de los incrementos de crecimiento
en los estatolitos se ha perfeccionado notablemente y, por
otra parte, se han efectuado validaciones utilizando como
marcadores el estroncio y la tetraciclina en varias especies
mantenidas en cautividad, comprobándose que la
periodicidad de las bandas de los estatolitos es diaria.
Se definen como bandas de deposición ó incrementos
a los depósitos periódicos de carbonato cálcico y materia
orgánica presentes en los estatolitos, cada uno de los cuales
se distingue como una banda clara rodeada por una banda
oscura al observarse al microscopio óptico. Se ha
comprobado que los cefalópodos pelágicos tienen estatolitos
con incrementos periódicos más fácilmente visibles que los
demersales y éstos que los bentónicos.
Es mucho lo que todavía queda por conocer sobre los
mecanismos fisiológicos específicos que provocan la
deposición diferencial de carbonato cálcico y de proteínas,
pero un trabajo reciente ha evidenciado que la variación del
pH de la endolinfa del estatocisto, relacionado con el ciclo
día-noche, podría ser el desencadenante de este fenómeno.
Lo cual parece ser una explicación bastante plausible.
De lo expuesto se deduce que las estructuras más
fiables para estimar la edad de la mayoría de los
cefalópodos, y en este caso la de los calamares gigantes, son
los estatolitos.

248
Fig. 57. Una de las formas de validar la determinación de la edad en un cefalópodo es mantenerlo en cautividad y marcarlo en dos
momentos diferentes con una sustancia fluorescente no nociva. En
la figura se muestra el estatolito de un Todarodes pacificus con dos marcas de tetraciclina inyectadas con un intervalo de 30 días, que
son los incrementos o marcas que se visualizan entre las dos bandas fluorescentes.
Actualmente, sólo se ha podido estimar la edad y las
tasas de crecimiento en seis ejemplares de Architeuthis: tres
machos maduros con tallas comprendidas entre 97,5 y 108
cm de longitud del manto, capturados en Irlanda en 1998, y
tres hembras inmaduras con tallas de 42, 103 y 161 cm de
longitud del manto, capturados en Australia, Asturias y
Nueva Zelanda, respectivamente. Estos estudios se
realizaron a partir del análisis de las bandas de deposición de
sus estatolitos, y lo primero que se advirtió es que las
secciones de estas estructuras tienen una apariencia similar
a las de otros calamares de menor tamaño, tanto en lo que
concierne a la anchura de los incrementos como a su
orientación. El número de incrementos en los estatolitos
varió entre los 153 de una hembra inmadura de 43 cm de
longitud del manto y los 435 de un macho maduro de 108

249
cm de longitud del manto. Si, tal como acontece en otros
cefalópodos, la deposición de estos incrementos es diaria,
entonces uno de
Fig. 58. Foto donde se muestran las bandas de crecimiento en un estatolito de Architeuthis.
los aspectos más relevante de estos trabajos sería el
evidenciar la elevada tasa de crecimiento de los calamares
gigantes y su breve esperanza de vida. La tasa de
crecimiento variaría entre 2,60 y 4,68 mm de manto al día
para un macho y para una hembra respectivamente. Esto
implicaría que los calamares gigantes tienen la mayor tasa de
crecimiento conocida hasta la fecha de todos los organismos
marinos vivientes. Por otra parte, clarificaría la incógnita de
su edad ya que, por extrapolación de lo que ocurre con los
peces, se había supuesto que los calamares gigantes
necesitarían varias décadas para completar su ciclo biológico.
Una vez conocida la edad de los animales y por cálculo
retrospectivo, se estimó la fecha de nacimiento de los seis
ejemplares estudiados, la cual se extendería desde marzo

250
hasta junio para los Architeuthis del Atlántico, y desde abril
hasta agosto en los ejemplares del Pacífico.
En un trabajo muy reciente publicado por Landman y
colaboradores en 2004, se ha utilizado por primera vez para
el estudio de la edad y el hábitat del calamar gigante
Architeuthis sanctipauli el análisis de isótopos de carbono y
oxígeno. En este estudio se utilizaron los estatolitos de tres
hembras, dos en incipiente estado de madurez y una
madura, cuyas longitudes del manto estaban comprendidas
entre 159 y 240 cm. Estos ejemplares fueron capturados en
Tasmania y Australia durante el verano austral de 1996. Uno
de los estatolitos de una hembra de 191 cm de longitud del
manto fue utilizado para la lectura de bandas de deposición,
evidenciándose unos resultados similares a los encontrados
en los otros estudios anteriores, pues en este estatolito se
observaron 351 bandas, lo que implicaría una edad estimada
de 1 año. Sin embargo, los resultados derivados del análisis
de isótopos sugerirían una edad próxima a los 14 años, lo
cual trastoca todo lo que anteriormente se pensaba del ciclo
biológico de estos gigantes. No obstante, tal y como indican
los propios autores, estos resultados deben ser tomados con
precaución, ya que entre otras razones, para efectuar los
cálculos de edad es preciso adoptar demasiadas asunciones
sobre el cambio isotópico en la columna de agua, el rango de
profundidades en el que viven estos calamares y el patrón de
crecimiento de los estatolitos, como para que las
estimaciones con isótopos sean aceptadas sin condiciones.
Los datos presentados, aunque incompletos, sugieren que los
machos maduran a menor edad que las hembras y que las
paralarvas del calamar gigante eclosionan durante un período
prolongado, probablemente durante todo el año, aunque con
un pico de eclosión cercano a la primavera y el verano, tal y
como sucede con otras especies de cefalópodos. En cuanto a
la duración de su vida y crecimiento, parece que existiría un
marcado dimorfismo sexual ya que los machos tendrían un
ciclo de vida de alrededor de un año y que además no
alcanzarían las tallas de las hembras. Por su parte, las
hembras poseerían un tamaño mayor y madurarían a una
edad más tardía, por lo que se presupone que su ciclo de

251
vida excedería el año de vida, probablemente dos o tres
años, pero parece improbable que alcancen edades
superiores a las mencionadas. Estos datos solo representan
los resultados de los escasos estudios existentes, que deben
ser completados y contrastados en el futuro, cuando se
pueda contar con más ejemplares de esta especie o con la
posibilidad de estudiarles en vivo mediante cámaras o
submarinos. Se podría decir que los mimbres están puestos,
pero que es necesario terminar el cesto para tener un
conocimiento más preciso de la edad y el crecimiento de este
kraken que habita todos los océanos de nuestro planeta.
Estrategia trófica: depredadores y presas de los
cefalópodos
Todos los cefalópodos, tanto los fósiles como los
vivientes, son activos depredadores. En lo referente a los
cefalópodos fósiles, los contenidos estomacales de los
ammonites son bien conocidos y están representados
principalmente por pequeños fragmentos de invertebrados.
Así, los restos de crustáceos, equinodermos y foraminíferos
encontrados en sus tractos digestivos fosilizados sugieren
que los ammonites tendían a depredar sobre animales
pequeños con poca capacidad natatoria. Estos estudios
también indican que los ammonites pelágicos solían depredar
sobre el zooplancton más frecuente en aquella época. Por
otra parte, los garfios que tamizaban los brazos de los
belemnites y otras especies de cefalópodos fósiles sugieren
un comportamiento depredador más activo, probablemente
basado en la captura de pequeños peces y otros cefalópodos.
Trasladándonos a la época actual, el papel de los
cefalópodos como depredadores en los ecosistemas marinos
les ha llevado a competir activamente con los vertebrados.
Así, el debate sobre la evolución convergente de los
cefalópodos y los peces ha sido un tema que ha generado
discusiones interesantes durante las últimas décadas. A
principios de los setenta se argumentó que los cefalópodos
eran un equivalente funcional a los peces. Esta observación
se hizo en base a la competencia que existe entre ambos

252
grupos por el mismo nicho ecológico. Por otra parte, aunque
los cefalópodos están limitados por su herencia de moluscos
en lo que concierne a anatomía, fisiología y bioquímica, estos
animales han tenido que adaptar su forma, sistema nervioso
y características biológicas para poder sobrevivir en el medio
marino. Los posteriores estudios sobre el ciclo vital de los
cefalópodos sugieren la existencia de factores que les
permiten competir exitosamente con los peces. Así, los peces
carecen del sistema de propulsión de los cefalópodos y de la
capacidad de cambiar de dirección rápidamente, que requiere
adaptaciones fisiológicas, bioquímicas y bioenergéticas que
optimicen las tasas metabólicas. No obstante, esta
adaptación no ha sido la única desarrollada por los
cefalópodos, los cuales poseen unos ojos muy especializados
y, además, muchas especies poseen cromatóforos que les
permiten cambiar de color y un sistema defensivo que
consiste en la expulsión de tinta cuando se ven amenazados.
Para capturar a sus presas, sus brazos y sus ventosas
(garfios en algunas especies) les permiten asirlas de tal
forma que difícilmente pueden escapar, lo que acentúa su
ventaja adaptativa respecto a otros depredadores que tienen
limitada la captura de sus presas a la capacidad de sus
mandíbulas. Por éstos y por otros motivos (entre otros las
interacciones depredador-presa y la competencia) es
importante estudiar las relaciones

253
Fig. 59. Esquema que muestra la red trófica dentro de la cual se
encuentran las potas Illex coindetii y Todaropsis eblanae en el ecosistema de las aguas de Galicia.
de los cefalópodos con otros componentes del ecosistema
marino, además de con los peces, con objeto de obtener una
comprensión más profunda de cuales son sus interrelaciones.
Es indispensable destacar que los cefalópodos juegan
un papel importantísimo en las redes tróficas de los
ecosistemas marinos. Para subrayar su papel bastaría con
señalar las estimaciones que sugieren que la población
mundial de cachalotes consumió alrededor de 100 millones
de toneladas de cefalópodos en 1972. A menudo, los
vertebrados oceánicos son mejores recolectores de
cefalópodos que las artes utilizadas por los biólogos para

254
llevar a cabo los muestreos biológicos, por ello el estudio de
los restos de cefalópodos encontrados en sus contenidos
estomacales, a menudo aporta más información que derivada
de los muestreos con redes.
La mayoría de los cefalópodos actuales son
depredadores activos y rápidos, que se alimentan de presas
vivas y tienen un ciclo de vida corto, cuya esperanza de vida,
generalmente, no sobrepasa el año. Para soportar esta
estrategia vital, los cefalópodos poseen tasas de crecimiento
elevadas y un metabolismo proteico muy alto, por lo cual
depredan vorazmente sobre un amplio rango de presas,
convirtiéndose así en depredadores oportunistas de
diferentes ecosistemas.
En los ecosistemas marinos, las poblaciones de
cefalópodos responden rápidamente a cambios
medioambientales de origen natural y antropogénico. Por
este motivo, los cefalópodos experimentan fluctuaciones
anuales de su abundancia, relacionadas tanto con su
explotación pesquera como con cambios en las condiciones
atmosférico-oceánicas que acontecen a escala global y
regional. Las apariciones repentinas de cefalópodos en
grandes abundancias están bien documentadas, existiendo
evidencias que algunas poblaciones de peces que han sido
sobreexplotadas son rápidamente reemplazadas por otras de
cefalópodos que ocupan su lugar en el nicho ecológico de
esos ecosistemas marinos. Un ejemplo cercano se puede
encontrar en las aguas de Marruecos y Mauritania donde los
cefalópodos ocuparon el nicho ecológico de diferentes
poblaciones de peces teleósteos que se explotaron
intensivamente a mediados del siglo XX.
En cefalópodos con ciclo anual, muchos de los cuales
son altamente migratorios, el impacto sobre las poblaciones
de sus presas es muy variable y altamente estacional. Un
ciclo anual breve implica que el impacto de los cefalópodos
en los ecosistemas marinos experimente importantes
variaciones interanuales. Así, cuando una cohorte poderosa
pasa a través de un sistema se producen elevadas tasas de
depredación que, en el caso de afectar a los primeros
estadíos de desarrollo de una especie de teleósteo

255
comercialmente importante, afectarán al reclutamiento y
abundancia de dicha población.
Los cefalópodos son únicos en el reino animal por
poseer unas modificaciones adaptativas que les facilitan la
captura de las presas y les permiten depredar sobre un rango
amplísimo de ellas. Los cefalópodos poseen una corona
braquial formada, en el caso de Architeuthis, por ocho brazos
y dos tentáculos que rodean su boca. Los pulpos carecen de
tentáculos y algunos calamares los pierden durante la
ontogénesis, como en el caso del calamar gato Taningia
danae. En Architeuthis, los tentáculos son extremadamente
largos y parecen desempeñar un papel muy importante en la
captura de sus presas. La corona braquial rodea la boca de
los cefalópodos, y les sirve para capturar, asir e inmovilizar
presas de un tamaño relativamente grande. Comparado con
la boca de los peces, la corona braquial de los cefalópodos es
considerablemente más versátil como mecanismo de captura
y manejo de las presas.
Los brazos y tentáculos de los cefalópodos están
armados con ventosas, suaves y musculares en los
octópodos, y provistas de un anillo quitinoso con pequeños
dientes en los calamares y sepias. En varias especies de
calamares y potas, aunque no en Architeuthis, algunas
ventosas se han modificado formando ganchos que facilitan
el agarre de presas como peces u otros cefalópodos. Un
ejemplo cercano lo constituye Taningia danae, que habita en
aguas de la vertiente noroccidental española, especie que
potentes garfios a lo largo de sus brazos.
La boca, que es relativamente pequeña, contiene un
par de picos o mandíbulas quitinosas que trocean la comida
antes de ser ingerida. Las diferencias mandibulares entre
especies no han sido relacionadas con la dieta, pero su forma
dicta el área disponible para la inserción de los músculos
mandibulares y, por lo tanto, la fuerza que puedan hacer
para morder y trocear la presa. El aparato bucal posee una
rádula adaptada para raspar el alimento, a modo de lija. Las
glándulas salivares posteriores de algunas especies
producen

256
Fig. 60. Gracias a la corona braquial, los cefalópodos son capaces
de capturar presas de gran tamaño debido a que ésta, unida a la boca, es un mecanismo de captura y manejo de las presas
tremendamente efectivo.
cefalotoxinas, que paralizan las presas al morderlas y son
responsables de una previa digestión externa de la misma.
Su tracto digestivo está adaptado para digerir rápidamente y
con una eficiente tasa de absorción. Las presas son
localizadas primariamente mediante la vista ya que los
cefalópodos poseen unos ojos bastante desarrollados, cuya
máxima expresión está representada por el calamar gigante.
Además, recientemente se ha descubierto que los
cefalópodos poseen un sistema de “línea lateral” compuesto
por mecanorreceptores que les permite captar las ondas de
presión provocadas por el movimiento de sus presas, y que
les ayuda en la localización de las mismas durante la noche o
cuando la visibilidad es baja.
Los escasos registros de calamares gigantes del
género Architeuthis a lo largo de su distribución mundial y su
carácter aparentemente solitario impiden un conocimiento
profundo de su comportamiento migratorio. Sin embargo, de
ocurrir este tipo de desplazamientos, las rutas migratorias
deben facilitar el encuentro de suficiente alimento capaz de
satisfacer las elevadas tasas de alimentación que requieren
estos gigantes.

257
Los ejemplos más elocuentes de cefalópodos con un
comportamiento migratorio relevante se encuentran entre los
omastréfidos. Un caso típico se halla en el género Illex, cuyas
especies realizan migraciones de alimentación y reproducción
de miles de kilómetros, y cuyo desove tiene lugar en zonas
tropicales o subtropicales de elevada productividad. Sus
paralarvas, denominadas Rhynchoteution, crecen en zonas
de retención o en las corrientes que fluyen de regreso hacia
los polos. Una vez que los juveniles se acercan a la fase de
maduración y están preparados para reproducirse existe una
migración contraria hacia las áreas de puesta. De este modo,
la tasa de migración de especies como Illex illlecebrosus es
de unos 20 Km. diarios. Los cálculos realizados demuestran
que la alimentación debe continuar durante la migración, ya
que sus reservas metabólicas son insuficientes para
mantener el gasto energético durante las distancias que
recorren. Por ello, los circuitos de estas especies deben
discurrir por áreas de alta producción. Por otra parte, se ha
observado que tanto I. illecebrosus como otras especies
altamente migratorias son caníbales si las condiciones de
alimentación son insuficientes durante esos desplazamientos.
Este comportamiento puede significar un mecanismo de
supervivencia para, al menos, una fracción de la población,
que se mueve formando bancos numerosos. La existencia de
canibalismo en Architeuthis es todavía materia de debate
entre los expertos. Hasta la fecha únicamente se han
encontrado restos de calamar gigante en los estómagos de
dos Architeuthis, uno capturado en aguas australianas y otro
en las neozelandesas. Los despojos estaban constituidos por
una maza tentacular completa y restos de ventosas; ello
parece evidenciar que Architeuthis es caníbal. No se descarta
que existan luchas entre los calamares gigantes, ya que se
han encontrado cicatrices producidas por los anillos córneos
de las ventosas en el cuerpo y los brazos de calamares
gigantes examinados en Terranova, y que en esas batallas
un contendiente ingiera restos del otro, pero podría tratarse
también de autofagia. Lo que sí parece probado es que,
como acontece en la mayoría de los cefalópodos, Architeuthis

258
sufre mutilaciones en sus extremidades que pueden
regenerarse.
En cuanto a la dieta de los calamares gigantes, pese a
la escasa información disponible, parece muy probable que
sus paralarvas sean planctónicas y, por ello, su alimentación
estaría basada en organismos del zooplancton,
principalmente crustáceos planctónicos. Por otra parte, la
más que probable ausencia de gregarismo en los juveniles y
adultos de Architeuthis afectaría su eficacia para capturar
presas, lo que haría que su comportamiento alimentario
fuese diferente de aquellas especies que forman bancos. Este
comportamiento solitario se infiere, en primer lugar, por la
escasez de registros, los cuales nunca han supuesto más de
tres capturas o avistamientos simultáneos en una misma
zona. Debido a su corpulencia y al reducido tamaño de sus
aletas, parece inviable que los calamares gigantes realicen
grandes migraciones en busca de alimento.
Un cefalópodo gasta aproximadamente ocho veces
más energía que un pez teleósteo para realizar un mismo
desplazamiento. La energía necesaria procede de quemar
proteínas y carbohidratos, lo que es propio de todos
moluscos. Por el contrario, el combustible básico de los peces
son los lípidos. Quemar un gramo de grasa significa obtener
6 kilocalorías, mientras que un gramo de proteínas o
carbohidratos produce 4 kilocalorías. El sistema metabólico
de los cefalópodos está bien adaptado a este tipo de
combustibles, cuyos principales productos finales son
compuestos nitrogenados. Parte de estas sustancias de
desecho son eliminadas por la orina, pero otras se acumulan
en diferentes formas amoniacales en distintas partes del
cuerpo de algunos cefalópodos. En el calamar gigante lo
hacen en su masa muscular y en forma de cloruro amónico,
lo que les proporciona una flotabilidad neutra.
Atendiendo al principio de optimización del gasto
energético, según el cual un depredador debe gastar menos
energía para localizar y capturar una presa que la que vaya a
obtener de la misma, junto al hecho de que la natación a
chorro es muy costosa desde el punto de vista energético, no
parece que los calamares gigantes realicen importantes

259
migraciones horizontales. Sin embargo, como la gran
mayoría de los cefalópodos oceánicos, Architeuthis, parece
realizar migraciones verticales diarias de cierta magnitud, lo
que le permitiría maximizar la amplitud de sus nichos
ecológicos mientras minimiza el riesgo de depredación en las
zonas superficiales de la columna de agua.
Los patrones de migración difieren entre especies y
cambian con la edad y la madurez sexual. Por ejemplo, en el
Atlántico sur, los inmaduros y maduros de Illex argentinus se
alimentan cerca del fondo en la plataforma patagónica
durante el día y migran hacia la superficie durante la noche.
A medida que se aproxima la madurez, se desplazan hacia
aguas más profundas en el talud continental, donde
permanecen cerca del fondo durante el día y se mueven
hacia profundidades comprendidas entre los 200-300 m
durante la noche, para alimentarse de mictófidos. Estos
aspectos son prácticamente desconocidos en Architeuthis,
por lo que cualquier aseveración sería mera especulación. Sin
embargo, los ejemplares capturados en las costas asturianas
lo han sido en redes que faenan para pescar bacaladilla entre
unos 250 y 800 metros de profundidad durante el día. Hasta
ahora sólo han aparecido hembras inmaduras o madurando y
un par de machos maduros, nunca hembras maduras, lo cual
podría estar más relacionado con una posible segregación
espacial que con la migración diaria de una fracción de la
población a zonas más accesibles a los arrastreros.
Existen varios problemas específicos en los análisis de
la dieta de los cefalópodos. Como ya se ha comentado, el
diámetro del esófago apenas puede ampliarse debido a que
se encuentra rodeado por el cerebro, que está envuelto por
un cráneo cartilaginoso. Por ello, aunque los cefalópodos son
capaces de capturar grandes presas, las mandíbulas deben
trocear la carne en pequeños trozos para que puedan ser
ingeridas, y sus partes duras, primordiales para la
identificación de las presas, tales como las vértebras,
otolitos, tegumentos de crustáceos, etc, son frecuentemente
rechazadas. Este rechazo selectivo además de dificultar la
identificación, sesga los datos sobre sus presas y sus
tamaños. Además, su voracidad no disminuye cuando son

260
capturados, habiéndose observado que pueden comer dentro
de las redes de pesca, por lo que sus contenidos estomacales
pueden no reflejar su dieta natural.
Por otra parte, los cefalópodos pelágicos, como los
calamares gigantes, son difíciles de observar directamente en
la naturaleza y más aún bajo condiciones de confinamiento
debido a la necesidad de enormes infraestructuras como
acuarios de gran tamaño. Incluso las especies de aguas
litorales, que son frecuentemente observadas por los
buceadores, son también bastante crípticas, lo que dificulta
la observación de su comportamiento depredador en su
hábitat natural. Por último, las altas tasas de digestión
provocan que muchos ejemplares carezcan de contenidos
estomacales, o que el material hallado en sus estómagos sea
virtualmente inidentificable.
Aunque existen numerosos problemas para estudiar la
alimentación de un cefalópodo, gracias al paciente trabajo de
los investigadores se ha llegado a tener un gran
conocimiento de la dieta de algunas especies. Una revisión
de las presas de los cefalópodos basada en los contenidos
estomacales y observaciones directas, cubriendo un gran
número de especies que representaban varias familias,
demostró que son animales oportunistas, habiéndose
encontrado en sus tractos digestivos representantes de la
mayoría de los grupos de animales marinos, incluyendo
formas inesperadas de cnidarios. No puede obviarse, sin
embargo, que algunos despojos hallados en los estómagos
de estos animales correspondan a presas de sus presas, en
lo que gráficamente se ha denominado “efecto de las
muñecas rusas”, donde las más pequeñas están encerradas
en otras progresivamente mayores.
En general los cefalópodos pelágicos y demersales
neríticos depredan principalmente sobre crustáceos,
poliquetos, peces y cefalópodos. Los juveniles y subadultos
se alimentan principalmente de crustáceos, cambiando sus
hábitos alimentarios hacia peces y cefalópodos con la edad.
Los adultos de especies pelágicas oceánicas comen
principalmente peces, crustáceos y cefalópodos, en orden
decreciente de importancia, mientras los recién eclosionados

261
y juveniles se alimentan en su mayoría de crustáceos
planctónicos, especialmente de copépodos.
Los calamares gigantes pueden considerarse como
cefalópodos oceánicos, ya que, pese a que pueden acercarse
hacia la plataforma continental, su hábitat natural reside en
la columna de agua sobre el talud y los cañones submarinos.
Se conocen muy pocos casos de ejemplares de Architeuthis
que hayan sido capturados o hallados con un estómago
repleto de comida recién ingerida. En varios de los individuos
examinados en Asturias se encontraron restos digeridos
(escamas y otolitos) de bacaladilla en sus contendidos
estomacales, junto con restos de crustáceos y otros de
cefalópodos.
En el caso de las paralarvas y juveniles de
Architeuthis, apenas hay datos sobre su dieta pues hasta
ahora se han obtenido sólo tres ejemplares de pequeño
tamaño (18, 45 y 57 mm de longitud de manto) y, sólo en el
juvenil de mayor talla se encontraron restos de comida en el
estómago, correspondientes a carne digerida y restos de un
teleósteo de pequeño tamaño. Este hallazgo se puede
considerar un hecho que diferencia a los primeros estadíos
del calamar gigante con los de otros cefalópodos oceánicos,
ya que, estas especies se alimentan principalmente de
crustáceos y son muy pocas las citas de restos de peces en
su régimen alimentario durante los primeros estadíos del
desarrollo.
En los calamares gigantes adultos de otras áreas,
como Irlanda, Namibia y Nueva Zelanda, la dieta observada
es parecida aunque, lógicamente, las especies no coinciden.
Los ejemplares capturados en Irlanda se alimentaron
principalmente de jurel (Trachurus trachurus), anchoa de
fondo (Maurolicus muelleri) y bacaladilla (Micromesistius
poutassou) entre los peces; pulpo cabezón (Eledone cirrhosa)
y potarro (Todarodes sagittatus) entre los cefalópodos, y
también de cigala (Nephrops norvegicus), moluscos bivalvos
(Modiolus phaseolinus) y ascidias; estos tres últimos tipos de
presa resultan algo sorprendente para un calamar de este
tamaño que parece tener su hábitat natural en la columna de
agua. Los contenidos estomacales de los ejemplares de

262
Namibia mostraban fundamentalmente restos de
cefalópodos, como joyelurias (Histioteuthis spp) y potas
(Ommastrephes bartramii), especie que es una excelente
nadadora, como lo son las potas neozelandesas (Nototodarus
spp), cuyos despojos se hallaron en los estómagos de
Architeuthis capturados en Nueva Zelanda, junto con restos
de al menos ocho especies de peces, muchos de ellos
macrúridos y peces hacha (Hoplostetus atlanticus). Estos
datos sobre la dieta del calamar gigante corroboran dos
cosas y sugieren una pregunta. En primer lugar, su carácter
oportunista; en segundo término, que, además de depredar
sobre peces y cefalópodos pelágicos y demersales, también
se nutre de especies que se fijan o se entierran en el fondo.
Por último, ¿cómo actúa Architeuthis para capturan especies
de cefalópodos que son notables nadadoras?
Para comprender la ecología trófica de los cefalópodos
resulta indispensable analizar no sólo su comportamiento
depredador y su régimen alimenticio sino también sus
potenciales depredadores. Estos no se encuentran solamente
en los niveles tróficos superiores sino que también pueden
estar en niveles tróficos similares, alimentándose de las
paralarvas y juveniles. Así, los potenciales depredadores de
los cefalópodos se pueden catalogar en tres categorías:
mamíferos marinos, peces y aves marinas. Por supuesto, la
importancia de cada uno de ellos es diferente, ya que su
rango de presas está determinado por su hábitat.
Circunscribiéndonos a los calmares gigantes, es obvio que no
existe un amplio elenco de posibles depredadores, al menos
si nos referimos a los animales de mayor tamaño. Sin
embargo, sus paralarvas y juveniles deben constituir parte
de la dieta de peces y otros organismos de tamaños
inferiores. Tal y como acontece en otros aspectos de la
bioecología de Architeuthis, su aparición en la dieta de la
mayoría de sus depredadores potenciales es relativamente
escasa. Sin embargo, señalar cuales son los potenciales
depredadores de los calamares gigantes es necesario, ya que
esto nos permitirá una mejor comprensión de su posición en
la trama trófica del ecosistema.

263
Fig. 61. Los mamíferos marinos son los principales
depredadores de los cefalópodos. En la foto, un delfín trata
de capturar un pulpo.
Entre los mamíferos marinos, los principales
depredadores de cefalópodos son los cetáceos. El término
cetáceo se utiliza para nombrar, en forma general, a las 78
especies vivientes de ballenas, delfines y marsopas. El orden
de los cetáceos se divide en dos subórdenes: misticetos o
cetáceos con barbas y odontocetos o cetáceos con dientes y
sin barbas. Los estudios realizados hasta la fecha no revelan
que los cefalópodos sean una parte constitutiva importante
del régimen alimentario de las ballenas o misticetos y
únicamente se han citado en los contenidos estomacales de
algunos rorcuales. Pero ninguno de estos es considerado un
depredador de calamares gigantes. Por el contrario, la dieta
del 80% de los odontocetos, entre los que se encuentran
cachalotes, calderones, zífidos y delfines, incluye
regularmente a los cefalópodos como presa. Aquí, el calamar
gigante desempeña un papel fundamental en la dieta de uno
de los depredadores de cefalópodos más voraces, el

264
cachalote (Physeter macrocephalus). La relación entre estos
dos colosos marinos se hace evidente no solamente por la
presencia de picos y restos de Architeuthis en los estómagos
de los cachalotes, sino también por las marcas de ventosas
que aparecen frecuentemente cerca de la boca de estos
cetáceos. Estas marcas o cicatrices pueden llegar a ser de
gran tamaño, lo que ha llevado en el pasado a
extrapolaciones que apuntan hacia la idea de que los
calamares gigantes podrían llegar a pesar varias toneladas.
Sin embargo, actualmente se considera que estas cicatrices
han podido ser producidas años antes de la captura del
cachalote, cuando este tenía un tamaño menor y así, con el
paso del tiempo y el crecimiento del animal, las marcas
también habrían crecido de tamaño. Lo que nadie duda es
que los calamares gigantes se resisten a ser capturados
pero, a pesar de la mitología y las leyendas sobre una lucha
encarnizada, parece obvio que ésta es a todas luces desigual,
y tiene con un claro favorito: los cachalotes.
Fig. 62. Marcas de ventosas en la piel de un cachalote (A);
fotografía de un ataque de un cachalote a un Architeuthis
(B); anillos quitinosos de calamar gigante (C y D).

265
En todos los océanos del mundo los estudios sobre el
régimen alimentario de los cachalotes muestran que
depredan sobre 19 familias de cefalópodos incluyendo los
Architeuthis. Además. Parece ser que estos animales son la
presa más frecuente de los ejemplares que habitan las aguas
del Atlántico norte y de Hawaii. Se ha estimado que los
cachalotes consumen entre 200 y 300 millones de toneladas
anuales de cefalópodos. Centrándonos por ejemplo en las
Azores, las estimaciones realizadas indican que los
cachalotes que habitan esta zona del Atlántico consumirían
unas 373.000 toneladas anuales de cefalópodos, de la cuales
45.000 toneladas pertenecerían a Architeuthis.
Entre los odontocetos de menor tamaño, se han
encontrado restos de calamares gigantes en algunas especies
de delfines de los géneros Stenella y Ziphius.
Los peces constituyen alrededor de la mitad de las
especies de vertebrados existentes, totalizando
aproximadamente 21.700 especies vivientes, de las que el
58% son marinas. Muchas de estas especies se alimentan de
cefalópodos en mayor o menor medida, pero su escasa
importancia relativa en relación con otros componentes de la
dieta hace que estos moluscos marinos no sean citados
frecuentemente como una fracción importante de su régimen
alimentario. Desafortunadamente, el estudio de la dieta de
los peces no representa un importante avance para la
comprensión de la ecología de los calamares gigantes, ya
que raramente se han encontrado restos de estos
cefalópodos en sus contenidos estomacales. Residuos de
Architeuthis se han encontrado en los estómagos de varias
especies de tiburones, entre los que destacan el tiburón
portugués (Centroscymnus coelolepis), el alicorto (Isurus
oxyrinchus), el tiburón azul (Prionace glauca), el tiburón
dormilón antártico (Somniosus antarcticus) y el pez martillo
(Sphyrna zygaena).
En el caso de los atunes, depredadores oportunistas
que ingieren cualquier presa abundante presente en el área
donde viven, se han observado despojos de Architeuthis en
la albacora (Thunnus alalunga) y el patudo (Thunnus

266
obesus). El pez sable, Alepisaurus ferox, prefiere
cefalópodos gelatinosos bastante pasivos, como
histiotéutidos, batitéutidos y cránchidos, habiéndose
encontrado también restos de calamar gigante en sus
contenidos estomacales. Otro importante depredador de
cefalópodos es el pez espada (Xiphias gladius), que depreda
sobre peces y cefalópodos, siendo los primeros el
componente principal de su dieta. Entre los cefalópodos, los
omastréfidos constituyen la fracción más importante en peso
de su dieta, pero también se han hallado restos de individuos
de otras diez familias, incluidos los Architeuthidae.
Es bastante probable que cuando se profundice en el
estudio de la dieta de peces demersales de profundidad se
encuentren restos de Architeuthis de pequeño tamaño, ya
que suelen capturar presas pelágicas que realizan
migraciones verticales diarias en la columna de agua y se
acercan al fondo. Por otra parte, las paralarvas y juveniles de
los calamares gigantes deben ser presa de numerosas
especies pelágicas de peces, siendo su ausencia explicable
por la escasez de estudios, la relativa poca abundancia de
estas, su gran dispersión y la errónea identificación de sus
restos.
Existe muy pocas evidencias que indiquen que las
aves marinas consuman selectivamente cefalópodos. Por ello,
aunque muchas especies son frecuentes en sus contenidos
estomacales, ello probablemente refleja una cuestión de
accesibilidad por tamaño o distribución más que una
preferencia específica. Existen muy pocos registros de
calamares gigantes en la dieta de las aves marinas, lo que no
representa una sorpresa debida a que el hábitat de los
calamares gigantes no es alcanzable por ningún ave marina.

267
Fig. 63. Pez espada Xiphias gladius
Hay que considerar el tamaño, naturaleza y
características de los cefalópodos depredados por las aves.
Es difícil creer que las aves son capaces de capturar sin
ayuda grandes cefalópodos, algunos mucho mayores que el
ave misma. Por todo ello, lo más probable es que, al menos
en el caso de los cefalópodos de gran tamaño, las aves
consuman restos flotantes dejados por otros depredadores,
como peces y cetáceos, o bien que sean carroñeras,
alimentándose de individuos gastados y moribundos que
flotan en la superficie después de haber efectuado la puesta.
Sobre este último punto, hay observaciones realizadas en el
área de Benguela, que evidencian que algunas aves marinas
consumieron cefalópodos que se encontraban flotando o se
habían hundido después de su muerte.
Las frecuentes asociaciones entre aves marinas y
cetáceos en aguas tropicales y templadas indican la clara
posibilidad de la existencia de una alimentación carroñera
donde las aves se beneficien de la actividad de los mamíferos
marinos y de los grandes teleósteos, que puedan desplazar a
los cefalópodos hacia la superficie. Por ello, no se puede
descartar que algunas especies, sobre todo albatros, petreles
y fragatas, se alimenten de individuos moribundos o
cadáveres que flotan ocasionalmente a la deriva.

268
Fig. 64. Las aves marinas son probablemente depredadores
oportunistas y carroñeros de los cefalópodos.
Actualmente, se está aplicando al estudio de la
ecología trófica de los cefalópodos una nueva técnica. Esta
técnica consiste en el estudio de la variación de la cantidad
de ciertos isótopos (principalmente Carbono 13 y Nitrógeno
15) en determinados tejidos. Como los picos son los restos
más frecuentes, y más resistentes, en los estómagos de los
depredadores de los cefalópodos, éstos se han usado para
conocer las relaciones tróficas y las pautas migratorias de
varias especies. En el caso de los calamares gigantes
Architeuthis dux y Mesonychoteuthis hamiltoni de las islas
Kerguelen, se ha demostrado que viven en ecosistemas
diferentes a lo largo de su vida. A. dux migra a las aguas de
esas islas desde zonas subtropicales, mientras que M.
hamiltoni lo hace desde la Antártica. Las variaciones del N15
sugieren, además, la existencia de un cambio en la dieta de
A. dux desde niveles tróficos inferiores a superiores a medida
que el animal se desarrolla. Los registros cíclicos de este
isótopo en el pico de esta especie, probablemente asociados
a la estacionalidad de la producción de plancton, sugieren
que la esperanza de vida del calamar gigante es de dos años.
Por otra parte, la composición isotópica del carbono indica un
comportamiento sedentario al menos en la fase de adulto,
mientras que los perfiles de fósforo y selenio en los picos
sugieren un descenso gradual hacia aguas más profundas a
lo largo de su ciclo de vida.
El conocimiento de la concentración de determinados
metales pesados, como plata, cadmio y mercurio, en
diferentes tejidos de Architeuthis dux está aportando nueva
información sobre su ecología.

269
Comportamiento
El comportamiento de cualquier animal depende,
entre otras cosas, del tipo de información que recibe a través
de sus órganos sensoriales, de la naturaleza de su efectores
(fundamentalmente los músculos) y de la organización de su
cerebro. Los cefalópodos pertenecen al filo de los moluscos,
un grupo de organismos muy antiguos al que también
pertenecen los mejillones, las ostras, los caracoles y las
babosas. Tienen en común con ellos la organización general
de su cuerpo en tres partes: cabeza, pie y masa visceral. Sin
embargo, en la mayoría de los cefalópodos el proceso
evolutivo ha llevado a la internalización o pérdida total de la
típica concha externa de los demás moluscos, al desarrollo
de unos excelentes órganos de los sentidos, a la agrupación
de varios ganglios nerviosos en un auténtico cerebro
protegido por un cráneo cartilaginoso, a la aparición de
neuronas gigantes, a la modificación del pie en un sifón por
donde, entre otras cosas, proyectan el agua que les permite
nadar a reacción, a la aparición de apéndices (brazos y
tentáculos) alrededor de la boca, al desarrollo de una
circulación sanguínea cerrada con auténticas venas, arterias
y capilares, y a la construcción de elaborados sistemas
excretores, digestivos y reproductores. Todos estos órganos
y sistemas son el soporte somático de un comportamiento
tan complejo, diverso y versátil, que no tiene parangón entre
los demás invertebrados marinos. Ello hace que muchos
investigadores consideren que los cefalópodos son peces
funcionales. Teniendo en cuenta únicamente el cerebro, el
índice de cefalización de los cefalópodos es parecido al de los
peces más avanzados, aunque su complejidad varía según
sea el hábitat que ocupan: mayor número de neuronas y de
conexiones neuronales en animales que viven en ambientes
abigarrados y complejos, como los arrecifes de coral, y
menor en los que viven en hábitats más sencillos, como los
ambientes pelágicos. De cualquier forma, la mayoría de los
cefalópodos son capaces de adoptar numerosos cambios
cromáticos y posturales y también de aprender.

270
En un cefalópodo como el calamar gigante se
distinguen cinco tipos de centros o lóbulos en el sistema
nervioso central: 1) Los centros nerviosos inferiores, situados
en los ganglios acetabularios y braquiales de la región
subesofágica del cerebro, cuya estimulación sólo genera
respuestas motoras de un efector concreto. 2) Los centros
motores intermedios, ubicados en diferentes zonas de la
región subesofágica, cuya estimulación provoca el
movimiento de numerosas partes del cuerpo, como por
ejemplo la eyección de los tentáculos para capturar una
presa, pero los movimientos que controlan no se organizan
en secuencias de comportamiento. 3) Los centros nerviosos
superiores, situados en la región subesofágica, compuestos
por diferentes lóbulos cuya estimulación provoca la actividad
de numerosos órganos efectores. Así por ejemplo, el lóbulo
pedunculado recibe información de las fibras ópticas y de los
estatocistos y manda señales que se relacionan con el control
motor visual y estático del animal. Su ablación impide
cualquier tipo de movimiento. 4) Los centros de análisis de
los órganos receptores o centros sensoriales primarios,
situados en la región supraesofágica del cerebro, están
compuestos por lóbulos que analizan la información recibida
por los órganos de los sentidos y elaboran las respuestas en
función de los cambios de situación que se presenten. 5) Los
centros auxiliares de la memoria, situados en los lóbulos
superiores, verticales y subfrontales de la región
supraesofágica, pueden considerarse como el equivalente de
las regiones asociativas del cerebro de los vertebrados.
La conducta de los cefalópodos implica una gran
cantidad de partes corporales cuyos constituyentes básicos
son tres: elementos, constituidos por los cromatóforos, las
células reflectantes (iridóforos y leucóforos), los fotóforos, los
músculos y los órganos internos. Unidades o suma de
distintos elementos. Y componentes, que son la unión de
diferentes unidades. Los componentes pueden ser cromáticos
(bandas, ocelos, etc.), texturales (piel lisa o rugosa),
posturales (brazos extendidos o cabeza y brazos
sobresalientes) y locomotores (en reposo, enterrándose en la
arena, nadando a propulsión a chorro o por impulso de las

271
aletas, etc.). De esta forma, cualquier pauta corporal puede
definirse por la adición de componentes cromáticos,
texturales, posturales y locomotores. Estas pautas pueden
ser crónicas (constantes) o agudas (pasajeras). Todas estas
pautas corporales son, en sí mismas, componentes de las
secuencias de conducta que presentan los cefalópodos. Así
por ejemplo, la secuencia que conforma el ataque visual de
un choco a una presa, está constituida primero por una pauta
o etapa de atención, después por un posicionamiento del
depredador respecto a la presa y, por último, del ataque con
apresamiento, y cada una de esas pautas tiene sus
componentes cromáticos, posturales, texturales y
locomotores específicos.
Otro aspecto fundamental de la conducta de los
cefalópodos es su comportamiento defensivo ante un
depredador. Lo primero que se manifiesta cuando se
produce
Fig. 65. El comportamiento de un cefalópodo implica, cambios posturales y de coloración. Un ejemplo es el comportamiento durante la caza, donde el cefalópodo no sólo se mueve de una
forma determinada (1 y 2), sino que cambia de color (1).

272
el ataque es una cripsis (camuflaje u ocultación), que
consiste en la adopción de una coloración protectora que
imita el ambiente circundante. Los tipos de cripsis son muy
variados, dos de ellos son el contrasombreado u ocultación
de la sombra que proyecta el animal hacia abajo cuando
nada de día entre aguas, y la coloración disruptiva o parecido
engañoso, que rompe la unidad corporal, lo que provoca que
el depredador se decepcione y no ataque. Si la cripsis falla,
entonces el cefalópodo puede desaparecer del escenario bien
sea escapando (nadando a reacción, enterrándose o
"volando" fuera del agua) o mediante un comportamiento
deimático, es decir, bravucón y amenazante. Este tipo de
conducta intenta espantar al depredador. Las actitudes
deimáticas más comunes son la flamígera o llamativa, la
formación de bandas de diferentes colores, la presentación
de ocelos, y los cambios súbitos de coloración que recorren
todo el cuerpo del animal modo de ondas. Este tipo de
actitudes son tan impredecibles y erráticas que se denominan
proteúsicas, nombre que deriva del dios griego Proteus, que
variaba constantemente de forma y color. Al producirse un
polimorfismo tan cambiante el depredador se confunde y su
potencial ataque queda inhibido. Un cefalópodo amenazado
puede también escapar de un depredador formando falsas
imágenes o fantasmas por expulsión de su tinta, o bien
puede dejar al depredador una parte de su cuerpo
(normalmente un brazo), que se autotomiza, para
entretenerlo. Cuando el cefalópodo es social, puede formar
un cardumen, con lo que el depredador pierde la noción de
individualidad y duda al atacar. Las defensas agresivas son,
sin embargo, raras en los cefalópodos.
En la reproducción los comportamientos son también
muy diversos como se vio en la sección dedicada a ello
dentro de este capítulo.
Un aspecto muy interesante de los cefalópodos es que
pueden comunicarse entre sí mediante señales posturales,
locomotoras, texturales y cromáticas. Las primeras pueden
ser del cuerpo entero (aplanamiento, extensión o despliegue
del cuerpo, orientación hacia el receptor, etc.) o únicamente

273
de los brazos (de uno sólo, de un par, de todos juntos,
bajados, extendidos, separados, juntos, presentando las
ventosas, o el hectocotilo, etc.). Las señales locomotoras
varían desde un posicionamiento determinado (por ejemplo
nadando en paralelo) hasta la persecución. Las texturales
consisten básicamente en mostrar el cuerpo liso o rugoso.
Por el contrario, las señales cromáticas son muy variadas:
palidecimiento y obscurecimiento general, destellos, oleada
de nubes obscuras, moteado, franjas, barras o bandas claras
y obscuras, lunares, manchas brillantes, anillos iridiscentes,
bordes de las ventosas obscuros, mitad del cuerpo de un
color y la otra de un color distinto, gónadas de color blanco
acentuado, glándulas accesorias rojas, emisión de luz por los
fotóforos internos o externos, etc.
Los cefalópodos además de ver excepcionalmente no
son sordos, habiéndose observado que son capaces de
detectar sonidos de baja frecuencia. Esto, ligado con la
existencia de receptores de presión (análogos a los
existentes en la línea lateral de los peces teleósteos), hace
que nos hallemos en presencia de unos animales que reciben
numerosísimas y complejas señales del exterior, cuyo
cerebro analiza y asocia, y del que parten órdenes hacia los
efectores, de todo lo cual se guarda memoria, más o menos
persistente según sea la importancia de la situación para la
supervivencia del individuo y su reiteración.
Realmente es muy poco lo que se conoce de la
etología de los calamares gigantes del género Architeuthis.
La principal razón de esta carencia de conocimientos es que
nunca se les ha podido observar vivos, sino, todo lo más,
moribundos. Sin embargo, a partir de los datos que
proporcionan sus registros se ha podido realizar una
actuación detectivesca que permite ofrecer algunas
suposiciones y sugerencias. La estructura y organización de
las criaturas pertenecientes a este género muestran ciertas
características paradójicas. Son sin duda los invertebrados de
mayor tamaño, pudiendo alcanzar hasta media tonelada de
peso o incluso más, pero la musculatura de su manto y
brazos no está particularmente bien desarrollada. Por otra
parte, hay muchos espacios rellenos de cloruro amónico que

274
les confiere flotabilidad neutra. Este hecho lo conocen desde
hace mucho tiempo los pescadores de muchos lugares, que
los recogían flotando y los utilizaban como un buen cebo. Las
aletas no son especialmente anchas ni fuertes y las fibras
nerviosas gigantes son mucho menores que las encontradas
en el calamar común (250 micras de diámetro frente a cerca
de 1000). Por otra parte, la morfología y el tamaño de los
estatocistos tienen las características propias de cefalópodos
poco activos y que flotan, con el cuerpo probablemente
inclinado y con la cabeza hacia abajo cuando están entre
aguas. Este cuadro nos presenta a una criatura que se debe
mover lentamente y no a un cazador agresivo. Sin embargo,
en sus estómagos se han hallado restos de peces que son
relativamente buenos nadadores, así como crustáceos y
otros cefalópodos, que no son precisamente nadadores
lentos. Por otra parte, el calamar gigante tiene un pico bien
desarrollado y una musculatura bucal poderosa, junto a
ventosas grandes con anillos córneos denticulados tanto en
brazos como tentáculos, los cuales son muy largos.
Finalmente, el cerebro posee un lóbulo braquial bien
desarrollado, el cual controla el movimiento de los apéndices.
Estas características, propias de cazadores muy activos,
introducen numerosas incógnitas sobre la forma como el
Kraken captura a sus presas. Por una lado, existen algunas
opiniones que los equiparan a carroñeros inactivos, que
estarían flotando a la espera de sus presas cerca del fondo,
más que a activos depredadores. Pudiera ser que
Architeuthis acechase verdaderamente, en solitario y
flotando pasivamente a media agua, el paso de un banco de
peces o de un grupo de cefalópodos o crustáceos. El ataque
se desencadenaría cuando estuviese a la distancia adecuada,
cálculo para el que precisaría distinguir perfectamente cada
presa dentro del cardumen y sobre el fondo marino. Si
Architeuthis es capaz, como hacen otros cefalópodos
pelágicos, de discriminar la luz polarizada, tendría resuelto
ese problema. Luego de situarse a cierta distancia, por
economía energética, debiera capturar sus presas una a una
y con gran precisión, para lo que utilizaría sus largos
tentáculos, mientras su enorme cuerpo quedaba a prudente

275
distancia y dentro de una envolvente y segura penumbra.
Pero nada de esto se ha visto todavía y todo son
suposiciones.
En términos absolutos, los ojos de Architeuthis son los
mayores del reino animal, pero esto no es así en términos
relativos ya que otros cefalópodos que viven en ambientes
abigarrados, como fondos coralígenos o en áreas de roca,
arenales cubiertos por vegetación y otras zonas fangosas,
poseen ojos que en proporción con su cuerpo o su sistema
nerviosos, son mayores. Ello es consecuencia de que la
capacidad y complejidad cerebral está relacionada con la
complejidad del hábitat. Esto implica también una elevada
variedad y riqueza de patrones de comportamiento, debida
principalmente, pero no sólo, al número y variedad de
componentes cromáticos que despliega el cefalópodo. Las
funciones de las pautas corporales incluyen, entre otras, el
camuflaje o coloración críptica y la comunicación.
Las especies activas durante las horas diurnas y que
moran en ambientes complejos han desarrollado ricos
repertorios de pautas corporales. Por el contrario, aquellas
que viven en ambientes simples o monótonos, como las
aguas pelágicas profundas donde moran los calamares
gigantes y son preferentemente nocturnas, tienen repertorios
más pobres. En el caso que nos ocupa, y como se deduce a
partir de los cadáveres estudiados de Architeuthis, su cuerpo
parece poseer pocos elementos cromáticos ya que no se han
observado fotóforos y los cromatóforos de las capas
tegumentarias que envuelven el cuerpo, la cabeza y los
brazos, así como los internos son básicamente de un único
tipo. Por otra parte, parecen carecer de iridóforos o
leucóforos y los músculos no son apropiados para
proporcionar a los individuos importantes cambios texturales
(piel lisa o rugosa). Además, el repertorio de pautas
posturales y locomotoras parece también limitado en
Architeuthis. En este limitado rango de información en que
nos movemos algo pueden ayudarnos las observaciones de
personas que tuvieron ocasión de estar junto a un calamar
gigante moribundo o recién muerto. Estas personas
describen su color variando entre el rojizo oscuro al castaño
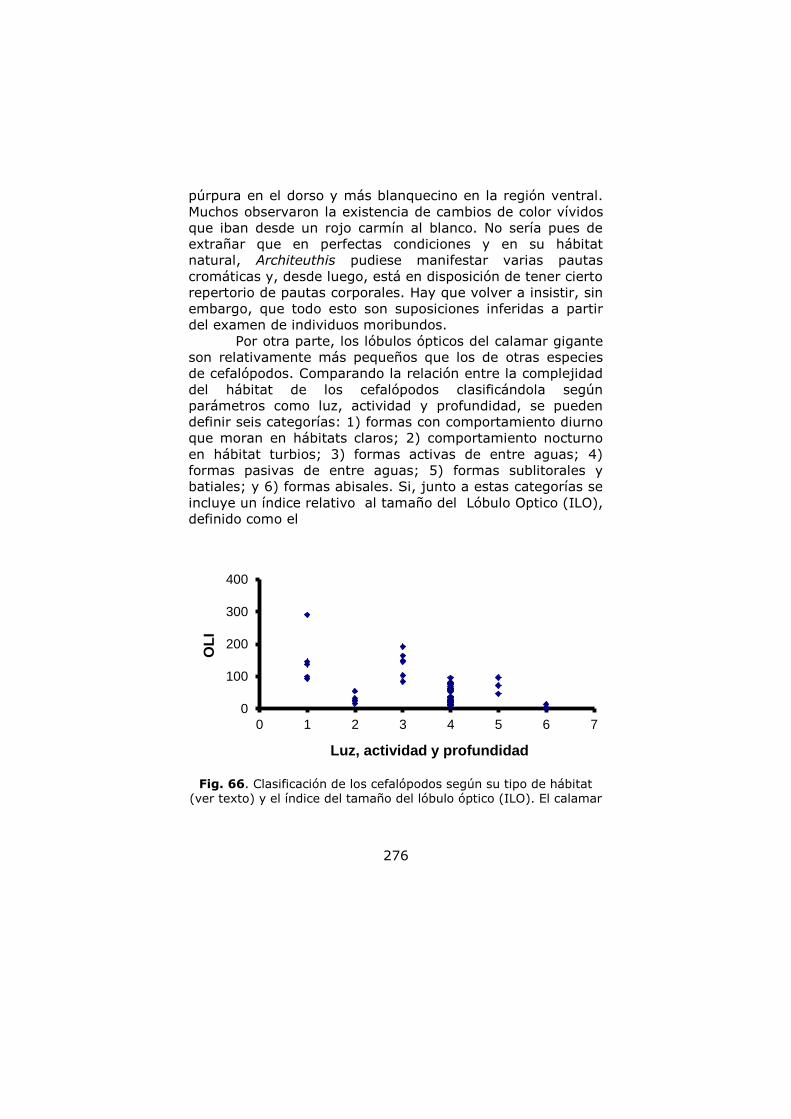
276
púrpura en el dorso y más blanquecino en la región ventral.
Muchos observaron la existencia de cambios de color vívidos
que iban desde un rojo carmín al blanco. No sería pues de
extrañar que en perfectas condiciones y en su hábitat
natural, Architeuthis pudiese manifestar varias pautas
cromáticas y, desde luego, está en disposición de tener cierto
repertorio de pautas corporales. Hay que volver a insistir, sin
embargo, que todo esto son suposiciones inferidas a partir
del examen de individuos moribundos.
Por otra parte, los lóbulos ópticos del calamar gigante
son relativamente más pequeños que los de otras especies
de cefalópodos. Comparando la relación entre la complejidad
del hábitat de los cefalópodos clasificándola según
parámetros como luz, actividad y profundidad, se pueden
definir seis categorías: 1) formas con comportamiento diurno
que moran en hábitats claros; 2) comportamiento nocturno
en hábitat turbios; 3) formas activas de entre aguas; 4)
formas pasivas de entre aguas; 5) formas sublitorales y
batiales; y 6) formas abisales. Si, junto a estas categorías se
incluye un índice relativo al tamaño del Lóbulo Optico (ILO),
definido como el
Fig. 66. Clasificación de los cefalópodos según su tipo de hábitat
(ver texto) y el índice del tamaño del lóbulo óptico (ILO). El calamar
0
100
200
300
400
0 1 2 3 4 5 6 7
OL
I
Luz, actividad y profundidad

277
gigante estaría entre las especies pasivas que habitan entre aguas de escasa luminosidad.
porcentaje entre el volumen de los lóbulos ópticos y el
volumen total cerebral, se observa (Fig. 66) que las especies
de cefalópodos que son activas y viven entre aguas, junto a
las las sublitorales y batiales tienen mayores valores del ILO
que las nocturnas, pasivas de entre aguas y abisales, hábitat
donde se integraría Architeuthis. Si a esto añadimos que el
calamar gigante carece de fotóforos, sus vesículas
fotosensibles están moderadamente desarrolladas y que su
lóbulo basal lateral y sus lóbulos cromatofóricos son muy
pequeños, todo parece indicar que estas criaturas pasan
mucho tiempo en zonas de penumbra o de total oscuridad.
Sin embargo, estas consideraciones surgidas a partir
de datos morfológicos están sujetas a debate, sobre todo por
los nuevos datos que se van obteniendo respecto al rango de
distribución vertical de Architeuthis, la posible existencia de
migraciones verticales, y la carencia de conocimientos sobre
sus diferentes fases del desarrollo. Se desconoce si realiza
migraciones de origen ontogénico, es decir, si las larvas y
juveniles viven próximas a la superficie mientras que los
individuos en maduración y los adultos descienden a medida
que crecen. Tantas incógnitas y conjeturas están reclamando
a gritos poder obtener imágenes de estos gigantes vivos y
desenvolviéndose en su medio natural.

278
Capítulo 6
OTROS CEFALÓPODOS GIGANTES
No todos los cefalópodos gigantes son Architeuthis
Este libro versa sobre el calamar gigante, un
cefalópodo perteneciente al género Architeuthis; el monstruo
mitológico de los mares que, como ya hemos visto, aparece
en muchas leyendas, la literatura e, incluso, en las películas.
Pero, ¿realmente todos los calamares gigantes son
Architeuthis? La respuesta es negativa, puesto que existen
varias especies de cefalópodos cuyo tamaño puede alcanzar
varios metros de longitud total, aunque, por lo que sabemos
hasta ahora, no parece que puedan alcanzar los 22 metros a
los que puede llegar Architeuthis.
En la clase de los cefalópodos hay varias familias que
poseen especies de gran tamaño, dos de ellas son
Ommastrephidae y Onychoteuthidae, que además
corresponden a grupos con especies explotadas
comercialmente. Los primeros son conocidos como las
grandes potas oceánicas, entre las cuales cabe destacar
Dosidicus gigas con sus casi 4 metros de longitud total
máxima; Ommastrephes bartramii, que alcanza los 2 metros,
y Todarodes sagittatus, que puede llegar a medir 1,5 metros
de tamaño total. Los Onychoteuthidae, o potas ganchudas ya
que tienen ganchos en sus tentáculos, también poseen
ejemplares de gran tamaño, como Moroteuthis robusta que
puede alcanzar una longitud total cercana a los 5 metros,
Moroteuthis ingens, Kondakowia longimana y Onykia sp, que
pueden llegar a tener algo más de 2 metros de longitud total
máxima.
Entre los Cranchiidae existe un verdadero gigante que
es Mesonychoteuthis hamiltoni, capaz de superar fácilmente
los 4 metros de longitud. Sobre este verdadero coloso de las
aguas antárticas se tratará con mayor detalle
posteriormente.

279
Fig. 67. Ejemplar de la cranquiluria Onykia sp.
Otro cefalópodo de gran tamaño perteneciente a la
familia Octopoteuthidae es Taningia danae, que también
recibirá una particular atención más adelante. También
podría considerarse como un gigante al calamar losange,
Thysanoteuthis rhombus, una especie cosmopolita de la
Fig. 68. Ejemplares de Thysanoteuthis rhombus capturados en
Asturias en 2002. (Foto de Eloy Alonso)

280
familia Thysanoteuthidae, que posee grandes aletas
romboidales y que alcanza tamaños de hasta 2 metros de
longitud total.
Sin embargo, no todos los cefalópodos gigantes son
calamares o potas, sino que también existen pulpos
gigantescos, como Enteroctopus dofleini y Haliphron
atlanticus. El primero se considera el pulpo gigante por
antonomasia. Se trata de una especie relativamente común,
que habita las aguas del Pacífico Norte que bañan América
del Norte y el continente asiático hasta Japón, alcanzando
hasta 3 metros de longitud total y unos 80 Kg de peso,
aunque hay citas no refrendadas de hasta 200 Kg y cerca de
9 metros de longitud total.
Recientemente se ha capturado un gran ejemplar de
pulpo pelágico de profundidad, cuya longitud total era de casi
3 metros y pesaba 61 Kg. Este ejemplar corresponde a la
especie Haliphron atlanticus, un pulpo gelatinoso de
considerables dimensiones que, hasta esa fecha, se pensaba
sólo podía alcanzar los 2 metros de longitud total. Debido a
la escasez de material se desconoce cual puede ser el
tamaño máximo que alcanza esta especie, así como
numerosos aspectos de su historia natural.
Fig. 69. Enteroctopus dofleini, el pulpo gigante del Pacífico.

281
Por lo tanto, junto al Architeuthis existen otras
especies de calamares gigantes que podrían ser objeto de
confusión para el neófito. A continuación se presenta una
breve visión sobre la biología y ecología de las especies más
relevantes, así como los principales caracteres que las
diferencia del genuino calamar gigante.
El calamar gato o pulpopota de profundidad, Taningia
danae
Entre los calamares de gran tamaño que habitan las
aguas de la mayoría de los océanos de nuestro planeta y que
no deben ser confundidos con Architeuthis, destaca Taningia
danae, vulgarmente denominado en castellano como calamar
gato, debido a la presencia de dos filas de garfios a lo largo
de sus brazos. También se le llama pulpopota debido a que
los adultos únicamente tienen ocho brazos. Esta especie fue
erigida por el naturalista francés Joubin en 1931. Taningia
danae pertenece a la familia Octopoteuthidae que, a
diferencia de Architeuthis, el cual posee largos tentáculos,
comprende aquellos cefalópodos que los pierden cuando
alcanzan los estadíos de paralarva o juvenil. Los juveniles de
esta especie presentan características peculiares, por
ejemplo: las aletas de estos animales son extremadamente
anchas en estos primeros estadíos de su desarrollo y sus
paralarvas poseen unas ventosas agrandadas características
situadas sobre brazos más gruesos y los ápices de los brazos
dorso-laterales (II), que están engrosados debido al
desarrollo de un fotóforo de gran tamaño. Los adultos poseen
un manto grueso y muscular y los fotóforos compuestos del
segundo par de brazos se hacen más patentes y están
recubiertos por un pliegue cutáneo que les permite mostrar u
ocultar el órgano luminoso. Aunque Taningia carece de
fotóforos en los ápices de los restantes brazos, si posee un
fotóforo redondeado interior, ubicado cerca de la bolsa de la
tinta. Las aletas de los adultos no son tan anchas como en
las paralarvas y juveniles, pero en ellos su longitud alcanza
alrededor del 85% de la del manto, siendo su anchura de

282
Fig. 70. A la izquierda (A) se muestra un momento de la disección
del ejemplar de Taningia danae de mayor tamaño capturado en el mundo (124 Kg. de peso), que fue pescado en el caladero de
Carrandi (Asturias) el 30 de octubre de 2000. A continuación se representa el rango de distribución de la especie (B), y el fotóforo
compuesto que hay en el ápice de cada segundo par de brazos de un adulto: cerrado (C) y abierto (D)
hasta el 135% del ancho del mismo. La longitud máxima del
manto de esta especie se aproxima a 170 cm.
Del mismo modo que ocurre con otros calamares de
gran tamaño, como Architeuthis, las capturas de paralarvas,
juveniles o adultos, o los estudios en laboratorio de Taningia
danae son ocasionales y, por lo tanto, su bioecología es
desconocida en gran medida. Sin embargo, esto no ha
impedido se hayan podido inferir algunos aspectos
interesantes sobre su ciclo vital.
Se trata de una especie cosmopolita, oceánica y
mesopelágica, es decir, que habita en la mayoría de los
océanos. En la columna de agua parece encontrarse desde la

283
superficie, sobre todo en el caso de los juveniles, hasta más
de 1000 m de profundidad (adultos). No obstante, estos
animales también parecen desplazarse hasta cerca del fondo
marino, tal y como sugiere la captura de la mayoría de los
ejemplares de Asturias, realizada por barcos que operan con
artes de arrastre demersal y bentónico. Los adultos de esta
especie poseen una coloración entre rojo burdeos y pardo
castaña distribuida homogéneamente a lo largo de todo el
animal. Se alimentan de peces pelágicos, crustáceos
decápodos y cefalópodos. Entre sus principales depredadores
se pueden citar grandes peces, como varias especies de
atunes, el pez sable (Alepisaurus ferox), y el tiburón
portugúes (Centroscimnus coelopis). También es parte de la
dieta de varios mamíferos marinos, como los elefantes
marinos (Mirounga angustirostris), el delfín de frente plana
(Hyperoodon planifrons) o el cachalote (Physeter
macrocephalus), constituyendo además alimento de de
varias aves marinas como el albatros errante (Diomedea
exulans). Estos depredadores capturan ejemplares de
Taningia danae en diferentes estadíos de su ciclo vital, desde
juveniles a adultos, y en océanos tan distantes como el
Índico, el Pacífico tropical, el Atlántico noreste y sudoeste o
el Antártico.
Externamente, el dimorfismo sexual se manifiesta
únicamente en que las hembras adultas son mucho mayores
que los machos, y en que éstos poseen un pene que
sobresale del manto. La fecundidad potencial de una hembra
madura es cercana a los cinco millones de ovocitos, lo que no
quiere decir que todos ellos se conviertan en huevos o
paralarvas viables, por lo que su fecundidad real es
desconocida aunque podría acercarse al millón de huevos. El
tamaño de los ovocitos de una hembra madura se
desconoce, sin embargo los de una hembra en avanzado
estado de madurez midieron entre 0,4 y 1,0 mm, lo que
sugiere que los recién eclosionados deben tener tallas
comprendidas entre 2 y 3 mm. Además, los estudios
reproductivos realizados indican que las hembras de esta
especie tienen una estrategia reproductiva de tipo desove
múltiple, lo cual implica que los huevos de la hembra son

284
puestos en diferentes episodios de desove, es decir, se
realizaría por tandas y en etapas durante un periodo que
puede llegar a durar uno o varios meses, y que durante este
tiempo la hembra sigue creciendo. Este tipo de estrategia
reproductiva es característica de ecosistemas relativamente
estables que presentan alteraciones periódicas. La captura de
la mayoría de los ejemplares en la columna de agua, pero
próximos al fondo, sugiere que el desove se realice cerca del
mismo, aunque se desconoce si la puesta se fija a algún
sustrato, o por el contrario las masas de huevos son
pelágicas y flotan libremente en la columna de agua.
La primera referencia de T. danae en aguas españolas
data del año 1966, cuando se encontraron 10 picos de varios
ejemplares en el contenido estomacal de un cachalote
capturado frente a la costa de Vigo. Posteriormente, se
encontraron restos de otros ejemplares en el estómago de un
cachalote capturado frente a la costa de Galicia en 1974. Sin
embargo, tuvieron que transcurrir más de 25 años para
obtenerse el primer ejemplar capturado aguas gallegas, que
se trasladó posteriormente a la lonja de Vigo. De este
animal, que pesaba unos 45 Kg, desafortunadamente, sólo
se pudo recuperar algunas vísceras (incluido el estómago, el
ovario y los oviductos), junto a la cabeza y los brazos, pero
el resto del cuerpo se perdió. Los registros recientes de la
especie en España vienen de aguas asturianas donde cuatro
ejemplares se capturaron en el año 2000, y otros tres más
entre 2002 y 2004.
Los ejemplares capturados en el año 2000 en aguas
asturianas eran un macho de 19 Kg. y dos hembras de 124 y
66 Kg. capturados en el caladero de Carrandi, a
profundidades comprendidas entre 400 y 600 m durante el
otoño de ese año. Los otros animales obtenidos
posteriormente se capturaron entre Carrandi y las aguas
situadas frente a Ribadesella a una profundidad similar a los
anteriores, variando sus pesos entre 12 y 104 Kg. Seis de los
siete ejemplares estudiados, todos ellos en excelente estado,
eran hembras inmaduras y no fecundadas, siendo una de
ellas el ejemplar más grande capturado hasta la fecha (124
Kg.). El espécimen macho capturado estaba completamente

285
maduro y pesaba tan solo 19 Kg. Los rasgos más
sobresalientes de este macho fueron la presencia de un pene
de 73 cm, que sobresalía 17 cm por fuera de la apertura del
manto, y cuya longitud corresponde al 102% de la longitud
de su manto, y la presencia de varios espermatóforos con
una longitud promedio de 170 mm. Considerando la carencia
de hecocotilización y la presencia de un pene de gran tamaño
en los machos maduros de Taningia danae, parece que la
reproducción de esta especie podría asemejarse a lo
observado en la familia Architeuthidae, donde el macho
inyecta a presión los espermatóforos en el cuerpo de la
hembra.
La edad de los ejemplares capturados en Asturias se
estudió a partir de los estatolitos de dos hembras de 66 y
124 Kg. de peso fresco. Estas estructuras duras se pulieron
por ambas caras y se estudiaron utilizando un microscopio
óptico con la ayuda de un sistema de análisis de imagen con
objeto de poder visualizar las bandas o anillos de deposición.
Es importante destacar que la deposición diaria no está
validada en esta especie, aunque sí lo está en otros
cefalópodos. Sin embargo, si se asume que los anillos se
depositan diariamente, la edad estimada para estos
ejemplares sería de 21 y 33 meses, respectivamente. Esta
estimación es importante si se compara con las realizadas
para ejemplares de Architeuthis utilizando la misma
metodología, pues los calamares gigantes, que eran también
hembras inmaduras de gran tamaño (más de 100 Kg.),
tenían una edad máxima estimada de un año. Según esto, la
edad del calamar gigante es sensiblemente inferior a los tres
años observados en Taningia danae, lo que indica que el
calamar gato crece a un ritmo inferior y es más longevo que
el calamar gigante.
El calamar de Humbolt, Dosidicus gigas
El calamar gigante de la corriente de Humboldt,
Dosidicus gigas, conocido en la bibliografía anglosajona como
Jumbo Flying Squid y en la española como calamar de
Humbolt, es el mayor y uno de los más abundantes

286
cefalópodos nectónicos de la familia Ommastrephidae que,
además, corresponde a una especie intensamente explotada
por flotas pesqueras especializadas en las costas de México y
Perú. Los omastréfidos son uno de los grandes nadadores del
océano pues poseen una poderosa musculatura y una forma
hidrodinámica tal que les permite nadar a gran velocidad y
ser activos depredadores, realizando migraciones de miles de
kilómetros. Su forma es la de un gran calamar con grandes
aletas, que se caracteriza por poseer un cartílago de cierre
del sifón con el manto con forma de T invertida y el que sus
ojos están expuestos al agua de mar, es decir, no recubiertos
por una membrana cornea. Una particularidad de estos
animales es que todos ellos poseen un tipo de paralarva muy
característica denominada Rhynchoteuthion, que se distingue
por tener los tentáculos fusionados en una probóscide, que
se van separando durante el crecimiento.
Fig. 71. A la izquierda, dibujo de un ejemplar adulto de Dosidicus gigas; y a la derecha un ejemplar capturado en aguas de Chile.
(Foto Pablo Reyes & Mathias Hüne)

287
Dosidicus gigas se caracteriza además por tener un
manto muy robusto, unas anchas y musculosas aletas
romboidales, que ocupan entre el 45 y 56% de la longitud
del manto, y poseer los extremos de sus brazos de forma
aguzada con entre 100 y 200 diminutas ventosas. Su
coloración es rojo-violeta y carece de grandes fotóforos en la
piel, si bien pueden existir algunos de pequeño tamaño
durante algún periodo de su ciclo de vida. Aunque pueden
alcanzar entre 1,2 y 1,5 m de longitud del manto, unos 4
metros de longitud total y hasta 150 Kg. de peso,
generalmente su tamaño oscila entre los 20 y 50 cm. de
longitud del manto, lo que corresponde a 1 ó 2 metros
totales, con pesos que varían entre 10 y 35 Kg.
La especie fue descrita por d’Orbigny en 1835 sobre
ejemplares capturados en las costas de Chile y es el “calamar
gigante” más conocido de toda la costa americana del
Pacífico. Dosidicus gigas habita el Pacífico oriental y se
considera una especie nerítico oceánica de aguas tropicales y
subtropicales que se puede encontrar frente a la costa de
América entre el norte de California (40-45ºN) y el sur de
Chile (47-47ºS), aunque es más abundante entre los 30º N y
los 25ºS. Su distribución longitudinal es máxima en el
ecuador, abarcando desde la costa americana hasta los 125-
140ºW, desde donde se va estrechando hacia el norte y el
sur, y siempre hacia la costa. En general, parece habitar
zonas de alta productividad marina dominadas por los giros
oceánicos y bajo la influencia de las corrientes de California y
de Humboldt.
Dosidicus gigas habita un rango de profundidades que
varía entre la superficie y los 1200 metros. Recientes
estudios realizados colocando transmisores en ejemplares
vivos y monitorizando sus desplazamientos, indican que los
adultos suelen desplazarse en la capa de agua de 0 a 200
metros de profundidad durante la noche y entre los 800 y
más de 1000 metros de profundidad durante el día. A
diferencia de Architeuthis, que parece ser una especie de
aguas frías, el calamar gigante de Humboldt habita
principalmente aguas cálidas entre 15 y 28ºC, aunque

288
Fig. 72. Distribución geográfica de Dosidicus gigas
también se le puede encontrar en zonas con aguas de baja
temperatura, hasta los 4,0-4,5ºC.
Esta especie es conocida por sus amplias migraciones,
que realiza de norte a sur y viceversa. Las puestas,
paralarvas y juveniles migran pasivamente arrastrados por
las corrientes, pero los adultos se desplazan activamente
desde sus zonas de alimentación a las de puesta a
velocidades que varían entre 5 y 25 Km. por hora. Estas
migraciones son masivas ya que se puede observar el
desplazamiento de las poblaciones de Dosidicus gigas
migrando hacia el norte y noreste desde zonas oceánicas al
golfo de California durante invierno y primavera, para luego
regresar en sentido opuesto durante el verano y otoño del
hemisferio norte. Desplazamientos similares se observan
desde el Pacífico ecuatorial hacia las costas de Perú durante
primavera y verano del hemisferio sur. Ocasionalmente y
asociados con fenómenos oceanográficos como El Niño,
cuando aguas tropicales irrumpen por la costa americana, se
producen verdaderas invasiones de este calamar que arriban
a las costas de Norteamérica (norte de los Estados Unidos) y
Sudamérica (centro y sur de Chile) en áreas periféricas de su
distribución. Estos fenómenos generalmente causan serios
problemas en las pesquerías locales ya que la especie es un
depredador muy agresivo que, aparte de experimentar

289
grandes varazones en las playas, colapsa las pesquerías
locales constituidas por sus presas. Las últimas veces que se
han producido estas invasiones en la costa chilena, durante
la década de 1990 y en los primeros años del 2000, su efecto
sobre los ecosistemas y en las pesquerías locales de Chile, ha
sido importante.
Las varazones de Dosidicus gigas son periódicas,
aunque su intensidad es muy variable. En Chile se tienen
registros históricos sobre este fenómeno, que indican que en
marzo de 1930 tuvo tal magnitud que bloqueó parte del
puerto de Talcahuano (centro-sur de Chile). Por el contrario,
durante la década de 1970 la especie prácticamente
desapareció de las costas chilenas. Posteriormente, ya por la
década de 1990 y 2000, se han observado grandes
concentraciones de D. gigas frente a las costas de Chile y se
han repetido, aunque no a gran escala, las varazones en sus
costas. Debido a que casi cualquier calamar grande que
aparece en las costas pacíficas de América a priori es
catalogado como perteneciente a esta especie a menos que
un experto tenga la oportunidad de estudiarlo, no es de
extrañar que existan muy pocos registros de Architeuthis,
sobre todo en Centro y Sudamérica, donde hay pocos
teutólogos.
Fig. 73. La aparición de un enorme cardumen de Dosidicus gigas bloqueó el puerto chileno de Talcahuano en marzo de 1930. Estas enormes varazones de la especie eran muy comunes en las costas
de Chile a principios del siglo XX. (Wilhelm, 1930)

290
Como se trata una especie explotada comercialmente,
gran parte del conocimiento sobre su biología y ecología
proviene del estudio de sus pesquerías. Al parecer, la especie
está dividida en dos poblaciones separadas, una asociada a
la corriente de Humboldt y otra al Golfo de California. A su
vez, en cada población se reconocen tres grupos de talla:
uno de ejemplares de pequeño tamaño (130-340 mm de
manto), otro de ejemplares medianos (240-600 mm de
manto) y un tercer grupo de ejemplares gigantes (400-1500
mm de manto). La presencia de varios grupos de talla es una
característica ya observada en otras especies de
omastréfidos, como en el caso de Sthenoteuthis oualaniensis,
y podría corresponder a la existencia de subpoblaciones
dentro de la propia población. Los tres grupos habitarían las
mismas zonas, aunque los gigantes parecen ser más
comunes en los extremos norte y sur de la distribución de la
especie.
Dosidicus gigas está entre los omastréfidos con
mayores tasas de crecimiento observadas, ya que diversos
estudios realizados utilizando seguimiento de tallas en la
población, incrementos en los estatolitos y marcas de
crecimiento en el gladio indican que este animal crece muy
rápidamente y vive generalmente un año. El crecimiento de
las paralarvas y juveniles es extremadamente rápido ya que
desde su eclosión, con 1,1 mm de manto, alcanzan entre 100
y 270 mm de manto en 45 a 55 días. Parece que el
crecimiento se reduce en los adultos cuando comienza la
maduración, pero no se detiene, ya que pasa de 0,8-1,5% en
juveniles a 0,2-0,4% al día en adultos.
El calamar gigante de la corriente de Humboldt es uno
de los cefalópodos con mayor fecundidad potencial que se
conoce, ya que el ovario de una hembra puede llegar a tener
hasta 32 millones de ovocitos, aunque esta cantidad
generalmente oscila entre 0,3 y 13 millones. Por su parte, los
machos pueden acumular entre 300 y 1200 espermatóforos
de entre 25 y 35 cm de longitud para la cópula. Como la
hembra no puede evacuar por los oviductos más del 10% de
los ovocitos maduros que posee el ovario a la vez, lo más
probable es que esta especie tenga una puesta múltiple

291
dentro de un ciclo reproductivo único. Aunque hay
ejemplares maduros con capacidad para reproducirse todo el
año, el principal periodo de reproducción parece acontecer
durante la primavera y principios del verano. Las zonas de
reproducción de Dosidicus gigas están situadas sobre el talud
continental o en aguas oceánicas adyacentes, asociadas a
áreas de alta productividad primaria y localizadas entre los
25ºN y 20-25ºS y a una distancia de entre 50-150 a 200-250
millas náuticas de la costa.
La cópula parece ocurrir cerca de la superficie y ha
sido observada en una ocasión por el investigador A.V.
Parfenjuk, quien vio a dos ejemplares nadando en la
superficie durante la noche: en un momento determinado el
macho se puso a nadar lentamente cerca de la hembra por
alrededor de unos 20 segundos para luego aproximarse y
copular con ella enlazando los tentáculos cabeza contra
cabeza durante unos 50 segundos y manteniéndose a flote
mediante el movimiento suave de sus grandes aletas. Una
vez completada la cópula ambos ejemplares se separaron.
Se desconoce cómo es la puesta de esta especie,
aunque se supone que es pelágica y flota a media agua
llevada por las corrientes. Varios experimentos en laboratorio
indican que el desarrollo embrionario es muy rápido,
pudiéndose completar en 6 a 9 días a 18º C, después de lo
cual nace una paralarva Rhynchoteution característica.
Durante su crecimiento, los ejemplares pasan por las etapas
de paralarva (1-10 mm de manto), juvenil (10-100 mm),
subadulto (100-350 mm) y adulto (más de 350 mm), que
implican cambios en su morfología, dieta y hábitat. Por
ejemplo, los juveniles y subadultos desarrollan varios
fotóforos oculares y dos intestinales, que producen luz
azulada-verdosa, los cuales están ausentes en las paralarvas
y los adultos.
Los juveniles de la especie son más activos que los
adultos y se desplazan generalmente en cardúmenes de 20 a
50 ejemplares nadando muy rápido y pudiendo incluso llegar
a saltar fuera del agua. Los subadultos se agrupan en
cardúmenes de 20 a 1200 ejemplares, pero son menos

292
activos. Finalmente, los adultos se mueven solos o en grupos
pequeños de entre 2 y 12 ejemplares.
Dosidicus gigas es un activo y muy agresivo
depredador cuyas presas son copepodos, amfipodos,
eufausidos, crustáceos pelágicos, el cangrejo rojo
(Pleuroncodes planipes), cefalópodos, otros moluscos y
peces. Este amplio espectro y la importancia de cada tipo de
presa va cambiando según Dosidicus crece. Así, en
ejemplares juveniles los invertebrados macroplanctónicos
son muy importantes mientras que en los adultos los peces y
los cefalópodos son el principal componente de su dieta. En
los adultos se ha observado que casi un 30% de la dieta
corresponde a cefalópodos y que, de estos, la mayoría son
casos de canibalismo. Tal es el grado a agresividad de esta
especie que se ha observado que muchos ejemplares son
atacados por otros de la misma especie al ser pescados e
izados a bordo y, en más de un caso, sólo a llegado la cabeza
al barco ya que el cuerpo se lo habían comido durante la
maniobra de izarlo a bordo. Incluso se han documentado de
ataques de estos calamares a buzos, aunque se trata de
casos relativamente aislados y bajo circunstancias especiales.
Los principales depredadores de Dosidicus gigas son
peces como los atunes, tiburones, pez espada, etc, aunque
también es consumido por otros cefalópodos, ya sea de su
misma especie u otros omastréfidos (Sthenoteuthis
oualaniensis por ejemplo), aves marinas como las gaviotas y,
finalmente, por mamíferos marinos.
El calamar de Humbolt es uno de los recursos con
mayor biomasa estimada, ya que se calcula que ésta alcanza
unas 7 a 10 millones de toneladas. Este es el motivo por el
cual la especie es fuertemente explotada en pesquerías
especializadas de aguas de México y, sobre todo, Perú, que
llegan a extraer hasta 200 mil toneladas anuales. Como la
especie experimenta importantes migraciones entre el norte
y sur, estas pesquerías fluctúan dependiendo de la
abundancia del recurso en cada zona, situación que se
agrava cuando ocurren fenómenos como El Niño, que alteran
completamente las distribuciones y abundancias de la
especie en toda su área de distribución.
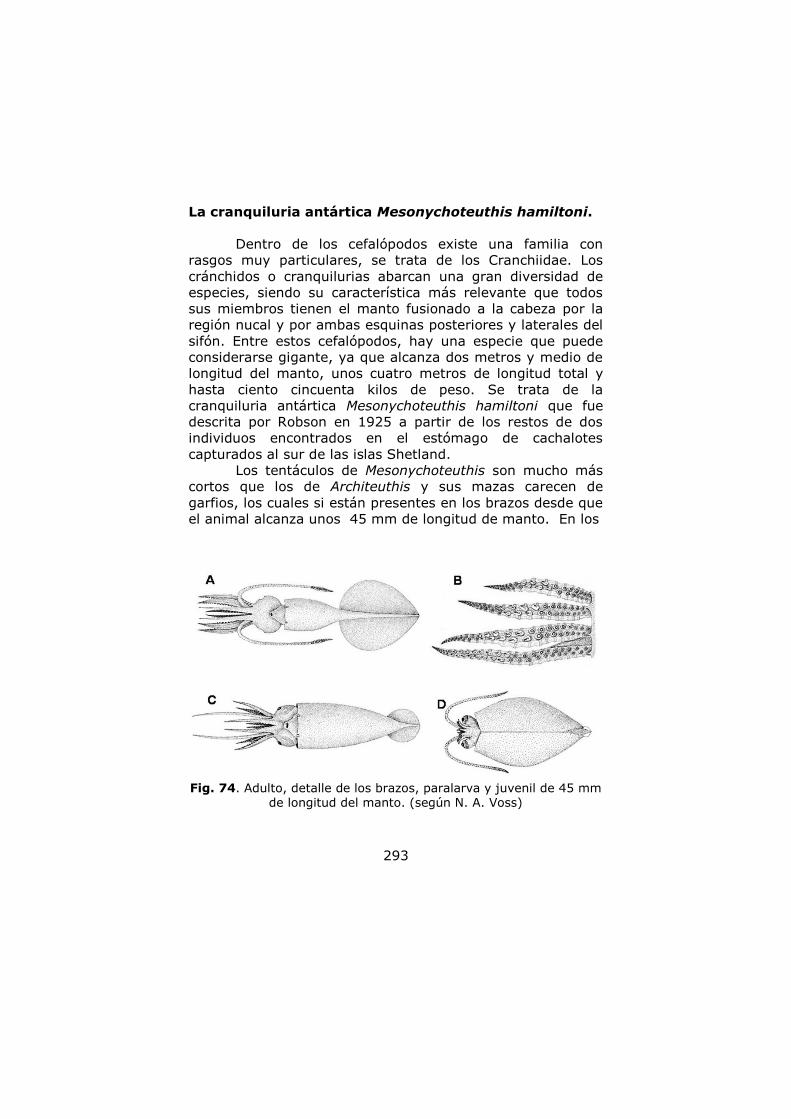
293
La cranquiluria antártica Mesonychoteuthis hamiltoni.
Dentro de los cefalópodos existe una familia con
rasgos muy particulares, se trata de los Cranchiidae. Los
cránchidos o cranquilurias abarcan una gran diversidad de
especies, siendo su característica más relevante que todos
sus miembros tienen el manto fusionado a la cabeza por la
región nucal y por ambas esquinas posteriores y laterales del
sifón. Entre estos cefalópodos, hay una especie que puede
considerarse gigante, ya que alcanza dos metros y medio de
longitud del manto, unos cuatro metros de longitud total y
hasta ciento cincuenta kilos de peso. Se trata de la
cranquiluria antártica Mesonychoteuthis hamiltoni que fue
descrita por Robson en 1925 a partir de los restos de dos
individuos encontrados en el estómago de cachalotes
capturados al sur de las islas Shetland.
Los tentáculos de Mesonychoteuthis son mucho más
cortos que los de Architeuthis y sus mazas carecen de
garfios, los cuales si están presentes en los brazos desde que
el animal alcanza unos 45 mm de longitud de manto. En los
Fig. 74. Adulto, detalle de los brazos, paralarva y juvenil de 45 mm
de longitud del manto. (según N. A. Voss)

294
dos tercios distales del funículo de los tentáculos hay varias
series de ventosas y almohadillas. Dentro de las
características más llamativas de esta especie están la
presencia de un órgano luminoso o fotóforo sobre cada ojo
en forma de media luna, dentro de cuya concavidad hay otro
más pequeño y oblongo; y el que sus aletas sean terminales
y ovaladas, careciendo de lóbulos anteriores y extendiéndose
a lo largo de toda la región posterior del manto, que es
bastante acuminado. Esta especie posee un manto cuyas
paredes son muy gruesas (6 cm) y de consistencia algo
gelatinosa.
Muy poco se conoce sobre la biología y ecología de
este calamar gigante antártico, que tampoco nunca ha sido
observado vivo en su ambiente natural. La distribución de la
cranquiluria antártica es circumpolar, ubicándose al sur de la
convergencia Antártica. Se alimenta de peces y cefalópodos,
siendo sus principales depredadores los cachalotes, elefantes
marinos, varias especies de focas, albatros, pingüinos y el
Fig. 75. Steven O’Shea con un gran ejemplar de M. hamiltoni capturado en el mar de Ross (Antártica).

295
bacalao de profundidad Dissostichus eleginoides. Los
juveniles viven preferentemente entre 20 y 600 m de
profundidad, mientras que los adultos lo hacen a mayores
profundidades, llegando hasta los 2.200 m. Se captura al
arrastre y también aparece en los palangres de profundidad,
pero siempre como captura incidental ya que, aunque es
comestible y parece tener una biomasa de cerca de 90
millones de toneladas, todavía no se ha desarrollado una
pesquería dirigida a esta especie.

296
Capítulo 7
EL CALAMAR GIGANTE Y EL GRAN PÚBLICO
Museos y exposiciones
Desde hace siglos, los calamares gigantes han sido
objeto de innumerables historias que han servido para
alentar nuestra imaginación, creando aventuras que incluyen
a estos colosos marinos o seres basados en ellos en las que
cualquiera de nosotros podría tener cabida. Por eso, es
frecuente que los niños que acaban de visitar una exposición
sobre los calamares gigantes hagan comentarios exagerados
sobre su apariencia o voracidad y comiencen a relatar
historias que les gustaría protagonizar, o que ellos creen
haber vivido, en sueños, claro está. Pero estas fantasías no
sólo las tienen los niños, sino también los adultos. Así, y a
modo de anécdota, solemos comentar lo que nos ocurrió en
una de las disecciones de un calamar gigante en Asturias, la
del más grande que se puede observar el la exposición que
CEPESMA tiene en Luarca, una hembra de 148 Kg. Durante
su disección se acercó una señora de mediana edad que, al
ver el ejemplar que estábamos analizando, nos dijo que no
era tan grande como ella se imaginaba. Discernir si ella creía
que se encontraría un monstruo marino de al menos 500 Kg.
debido a su desbordante imaginación, o si su experiencia le
había permitido ver un ejemplar más grande hace décadas,
fue algo imposible a la luz de la conversación que
mantuvimos. Lo que parece claro es que, a veces, nuestra
mente nos hace ver más allá de la realidad.
Desde un punto de vista objetivo, el calamar gigante
Architeuthis es el mayor invertebrado que habita el planeta
tierra. Por ello, tiene un gran poder evocador que se traduce
en numerosos símbolos, los cuales están representados de
múltiples formas, tales como tatuajes, collares, pinturas, etc.

297
Sinceramente, la pasión que mueve a los teutólogos en el
estudio de estos colosos es superada a veces por la que
sienten algunas personas anónimas por estos moluscos
marinos, como las que deciden representar obras de un valor
artístico innegable tatuadas en su cuerpo.
Las conversaciones mantenidas con personas
relacionadas o no con el campo de las ciencias marinas
revelan el creciente interés por conocer la realidad sobre los
calamares gigantes. Este interés se incrementa con cada
noticia sobre la aparición de alguno de estos extraordinarios
animales en algún lugar. Si el hallazgo se produce cerca de
nuestras costas, lo primero que se plantea el público en
general es que los calamares gigantes no son un tema tan
lejano como pensaban. En este sentido, las exposiciones en
las que se pueden observar calamares gigantes son
importantes por múltiples razones, tanto para los que
piensan que estos grandes invertebrados no existen, como
para los que imaginan que su tamaño podría superar límites
inimaginables. Desafortunadamente, existen muy pocos
ejemplares de Architeuthis en las colecciones de los museos
de ciencias naturales de todo el mundo. Las razones de esta
mínima representación se deben, principalmente, al escaso
número de animales registrados a nivel mundial, así como al
mal estado en que se encontraron. Por otra parte, la
preparación y conservación de animales de gran tamaño no
resulta sencilla y los materiales de preservación y recipientes
especiales para mantenerlos necesitan de presupuestos, que,
aunque no inalcanzables, representan un desembolso
importante.
La primera vez que nos enfrentamos a la preparación
de un ejemplar de estas características para un museo, una
hembra de 148 Kg. de peso, nos planteamos numerosos
interrogantes. Nuestro primer objetivo se centró en buscar
un impacto visual para que el público pudiera hacerse una
idea lo más real posible de cómo son estos animales en la
naturaleza. Para ello, teníamos que huir de una mera
introducción del calamar eviscerado en una urna, que con el
paso del tiempo convirtiese al animal en un ser plano,
deforme y poco atractivo. Por ello, lo primero que debíamos

298
solucionar era como proporcionar una sensación de volumen
al animal. Esto se resolvió con dos procedimientos: en primer
lugar, sustituyendo la pluma del calamar, generalmente rota
por varias partes, por una estructura rígida de madera o de
plástico; en segundo término, mediante la introducción de
material de relleno plástico dentro del manto eviscerado, que
fue luego cuidadosamente suturado; y en tercero, por la
introducción de un cilindro de plástico dentro del sifón, que le
proporcionaba la apariencia de estar expeliendo agua. La
naturaleza de los materiales utilizados, además de dar el
volumen deseado a la pieza, le proporcionaban cierta
flotabilidad dentro del líquido conservante, separando al
espécimen del fondo de la urna. Lo que mayor complicación
planteó fue la reconstrucción de los ojos, que en el calamar
gigante son de grandes dimensiones, y que en la mayoría de
los ejemplares aparecen totalmente reventados debido al
brusco cambio de presión entre la profundidad a las que se
capturan estos animales y la superficie, así como al daño
producido por las redes. La mejor solución fue introducir
focos del tamaño semejante al ojo y con una superficie
convexa de color grisáceo que simulase el cristalino. El
resultado fue satisfactorio y después de suturar los tejidos
oculares, parecía que el gran calamar estuviese
observándote. También se suturaron los trozos de brazos y
tentáculos rotos con objeto de reconstruirlos en su mayor
extensión posible.
En lo que referente a la fijación y conservación del
animal, después de ser introducido en la urna diseñada y
construida ex profeso para su conservación, se llenó esta con
una solución de formaldehído al 10% en agua de mar
destinada a fijar los tejidos, que al cabo de 15 días, fue
reemplazada por otra de alcohol al 70% en agua dulce, que
es el líquido en el cual se conservan actualmente todos lo
calamares gigantes que se pueden observar en el Museo Aula
del Mar de CEPESMA.
Resulta innegable afirmar que la colección que se
encuentra en este pequeño museo de Luarca es la más
importante del mundo en lo que respecta a calamares
gigantes, ya que, además de varias hembras de Architeuthis,

299
se encuentran dos de los ocho machos hallados hasta la
fecha en aguas del Atlántico nororiental, uno de los cuales
presenta espermatóforos en la base de sus brazos. También
se encuentran allí expuestos cuatro ejemplares del calamar
gato Taningia danae recolectados en aguas asturianas, uno
de los cuales es el de mayor peso nunca antes observado.
Por ello, es realmente inaudito que esta exposición
permanente no tenga la difusión que se merece, máxime
cuando se compara con los escasos ejemplares que se
conservan a lo largo y ancho de los museos más importantes
del mundo.
Una de las colecciones más importantes de calamares
gigantes de allende nuestros mares se encuentran en la
sección de moluscos de la Smithsonian Institution de
Washington D.C. Esta institución, que posee más de 142
millones de muestras en su colección, es también un centro
puntero dedicado a la investigación pública y a subvencionar
becarios en los campos del arte, la ciencia y la historia. La
visita a cualquiera de sus instalaciones impresiona por el
amplísimo elenco de material disponible para el estudio y
observación en sus decenas de exposiciones. Esta institución
Fig. 76. Primer ejemplar de Architeuthis que se expuso en el Aula
del Mar de Luarca.

300
se estableció en 1846 con fondos legados por el científico
británico James Smithson, y tiene como objeto el aumento y
la difusión del conocimiento. Es en el Museo Nacional de
Ciencias Naturales de los EE.UU. anejo a la Smithsonian
Institution, donde se conserva uno de los primeros
ejemplares de Architeuthis expuestos al público. Comparar
los miles de organismos marinos expuestos en este museo
con el material de Luarca no tiene sentido debido a la
abismal desproporción en casi todos los aspectos favorable a
la Smithsonian. Sin embargo, hay un punto en el que Luarca
es muy superior, y este es la cantidad y calidad de los
ejemplares de calamar gigante que se pueden ver en este
pequeño pueblo asturiano.
Además del Architeuthis que se puede observar en la
Smithsonian, existen otros ejemplares expuestos
permanentemente en diferentes museos de todo el mundo,
como por ejemplo dos que se conservan en el Museo
Americano de Historia Natural de Nueva York y en el Museo
Nacional de Escocia, respectivamente. Es tanta la
importancia que se le concede a estos calamares que, en
numerosas ocasiones, se les dan arbitrariamente apelativos
tan pomposos como: “el ejemplar mejor conservado del
mundo”. Así se afirma, por ejemplo, del ejemplar de 114 Kg.
expuesto en el museo de Nueva York, el cual llegó a su sala
de biodiversidad el 12 de octubre de 1999 procedente del
Instituto de Investigación Atmosférica y Marina de Nueva
Zelanda. Este animal fue capturado accidentalmente a finales
de 1997, congelado inmediatamente y transportado a Nueva
York en avión, pagándose por él cerca de diez mil dólares.
Además de este ejemplar, también se exponen en
este museo dos maquetas de calamar gigante, teniendo una
de ellas 110 años de antigüedad, ya que fue comprada en
1895. El otro modelo representa la lucha de un cachalote con
un calamar gigante en las profundidades del océano.
Desafortunadamente, en Europa no existen muchos
ejemplares de Architeuthis expuestos en museos. Así, hasta
donde alcanza nuestro conocimiento, además de los
calamares gigantes exhibidos en Luarca, solamente se puede
observar otro ejemplar en el Museo de Biarritz, con una

301
longitud total superior a los siete metros, que fue cedido por
CEPESMA. Por otra parte, en el Museo de Historia Natural de
Viena se conserva un brazo de largas proporciones, así como
otros tentáculos intactos en Noruega y Dinamarca.
Aunque Australia, Nueva Zelanda y Sudáfrica son tres
de los lugares del mundo donde han aparecido más
Architeuthis, no existen hasta ahora en esos países
exposiciones que muestren alguno de los muchos ejemplares
que han aparecido flotando o han sido capturados en sus
aguas.
Lo que se puede encontrar en otros museos, además
del material conservado en colecciones para el estudio de
especialistas, son maquetas, que en algunos casos no
guardan siquiera las proporciones adecuadas. Sin embargo,
no es nuestra intención analizar la adecuación a la realidad
de esas reproducciones, sino constatar donde se pueden
observar, si es que tenemos la suerte de viajar a lugares tan
distantes como Sudáfrica, EE.UU. o Canadá. Así, tal y como
se muestra en la fotografía, una bonita reproducción de un
calamar gigante se puede observar en una bahía de
Terranova.
Fig. 77. Maqueta de un Architeuthis expuesta en una bahía de Terranova (Canadá).

302
La historia de las maquetas de calamares gigantes se
remonta al siglo XIX. La primera que se expuso al público fue
la realizada por J.H. Emerson, bajo la dirección científica de
A. E. Verrill, en 1883, que se expuso en el Museo Peadbody
de la Universidad de Yale (EE.UU.). Luego vinieron otras
como la que existía en la universidad de Harvard, o la que se
mostró en el Museo de Historia Natural de Londres en 1906,
que fue destruida en 1940 por los bombardeos alemanes
durante la Segunda Guerra Mundial. También hubo otras dos
en el Museo de la Naturaleza de Salzburgo (Austria) y en el
Museo de Zoología de la Universidad de Copenhague. Una de
las más modernas, realizada en 1997 por el artista R. Ellis,
se encuentra expuesta en el Museo Nacional de Escocia en
Edimburgo. Los materiales con los que se confeccionaron
estas maquetas han variado con el transcurso del tiempo,
realizándose actualmente con plásticos de diferentes tipos.
Actualmente, se puede encontrar maquetas de
calamar gigante en las exposiciones de los Museos de
Historia Natural de Londres y Chicago. Otra maqueta
espectacular es la que se exhibe en el Museo de Historia
Natural de Ciudad el Cabo que representa a un calamar
gigante hembra de 1,84 m de longitud del manto varado en
Soetwater (Sudáfrica) después de una fuerte tormenta en
1991. El modelo tiene una longitud total de 9.3 m.
Finalmente, cabe destacar varias maquetas que se pueden
contemplar en Luarca, que reflejan fielmente la forma del
Architeuthis.
La leyenda del Kraken se difundió en inglés gracias al
popular poema The Kraken de Lord Alfred, Tennyson,
publicado en 1830. Además de pánico, el Kraken produce
una abyección ligada a la posibilidad de ser absorbido a
través de sus ventosas. En su novela Les travailleurs de la
mer, publicada en 1866, Víctor Hugo nos hace experimentar
el horror que se puede sentir cuando un cefalópodo gigante
nos aprisiona entre sus tentáculos “Una dentellada es
horroroso; pero menos temible que una succión. [...] la
ventosa, eres tú mismo que penetra en la bestia, tus fibras
se retuercen, tu piel revienta bajo un peso inmundo, tu

303
Fig. 78. Maquetas de Architeuthis en el Aula del Mar de Luarca.
sangre brota y se mezcla horrorosamente con la linfa del
molusco. La bestia se superpone a ti por miles de bocas
infames; la hidra se incorpora al hombre; el hombre se
amalgama con la hidra. [...] el pulpo, ¡que horror!, te
absorbe incorporándote a sí mismo, y te engulle, impotente,
y te sientes lentamente vacío en ese espantoso saco, que es
el monstruo. Aún más terrible que es ser comido vivo está lo
indescriptible de ser bebido vivo”.
Literatura
En la ciencia ficción los calamares, o entes fantásticos
basados en ellos, han sido protagonistas de un amplio
número de relatos, novelas y películas, debido
probablemente a que se trata de un depredador con cierta

304
inteligencia y a las leyendas del Kraken. Uno de los episodios
de la literatura clásica mejor conocidos es el relato del
encuentro del submarino Nautilus del capitán Nemo con
cefalópodos gigantes, que incorpora Julio Verne en su libro
Veinte mil leguas de viaje submarino (1870). Verne
demuestra tener bastantes conocimientos de la leyenda del
Kraken, citando los exvotos de Saint-Malo, las descripciones
de Olas Magnus y del obispo Pontoppidan, así como el
encuentro de la tripulación del Alécton con un calamar de
enormes dimensiones al nordeste de Tenerife en 1861. Verne
imagina a esos cefalópodos como animales notablemente
repulsivos, malévolos y perversos. A través de uno de los
ventanales del submarino los protagonistas de la novela
observan un calamar gigante, que el profesor Aronnax
describe de la forma siguiente: “era un calamar de
dimensiones colosales, y de ocho metros de longitud, que
nadaba en la dirección del Nautilus. Estaba mirando con sus
enormes ojos fijos de matices verdosos. Sus ocho brazos, o
por mejor decir sus ocho pies, implantados en la cabeza, lo
cual ha hecho dar a estos animales el nombre de
cefalópodos, tenían un desarrollo de su cuerpo y se retorcían
como la cabellera de las furias. Se veía con toda claridad las
doscientas cincuenta ventosas dispuestas en la cara interna
de los tentáculos en forma de cápsulas semiesféricas.
Algunas veces las ventosas se aplicaban sobre los cristales
del salón, adhiriéndose a él por medio del vacío que
practicaban. La boca de este monstruo, pico córneo
semejante al del papagayo, se abría y cerraba verticalmente.
Su lengua, sustancia córnea también, y armada con varias
hileras de agudos dientes, salía estremecida de aquella
verdadera cizalla ¿Qué capricho de la Naturaleza!¡Un pico de
ave para un molusco! Su cuerpo fusiforme y abultado en su
parte media, formaba una masa carnosa, que debía pesar de
veinte a veinticinco mil kilos. Su color inconstante,
cambiando con extraordinaria rapidez, según la irisación del
animal, pasaba del gris lívido al pardo rojizo”. Más adelante,
siete de esos “pulpos” (ahora los llama así Verne) atacan al
submarino, deteniendo su marcha porque uno de ellos agarra
una de las aletas de la hélice con sus mandíbulas. Como esos

305
monstruos, cuya descripción contiene tanto elementos
extraños a los auténticos calamares gigantes como otros
bastante ajustados a la realidad, no son afectados por los
campos eléctricos que defienden al submarino, su tripulación
y los tres invitados por Nemo, el profesor Aronnax, su criado
Conseil y el pescador de ballenas canadiense Ned Land,
luchan contra ellos con hachas y arpones, hasta que los
ahuyentan no sin antes haber sufrido la pérdida de un
tripulante y haber estado seriamente en peligro la vida de
Aronnax.
La novela de Verne se hizo todavía más famosa a raíz
de la proyección en todo el mundo de la película Veinte mil
leguas de viaje submarino producida por los estudios de Walt
Disney en 1954, donde James Mason interpretó el papel de
capitán Nemo, y Paul Lukas, Peter Lorre y Kirk Douglas los
de sus tres invitados.
La tradición literaria se continúa en obras como Deep-
Sea Plunderings (Los saqueadores de las profundidades)
donde, junto con relatos reales, su autor, Frank Bullen
(1902), mezcla algunos de ciencia ficción, como el titulado La
última etapa de los Decápodos. Sorprendentemente, ese
relato no trata sobre personas asesinadas por los calamares
gigantes o embarcaciones engullidas por estas aviesas
criaturas, sino sobre la rivalidad entre el Kraken y los
cachalotes.
En The Sea Raiders (Los invasores marinos),
publicada por H. G. Wells en 1905, el autor parece estar
relativamente familiarizado con alguno de los varamientos de
calamares gigantes acontecidos en Terranova unos años
antes de la publicación de su novela, pero sitúa la acción en
la comarca de Devon al sur de Inglaterra. El autor inventa la
especie Haploteuthis ferox, que puede traducirse como
calamar feroz, animal que llega a atacar al Sr. Fison
persiguiéndole incluso por tierra, y que es considerado
responsable de la muerte de numerosas personas.
Ian Fleming, en su novela Doctor No de 1958, coloca
a James Bond en la tesitura de tener que escapar de un
tanque trampa donde el malévolo doctor ha encerrado un
agresivo calamar gigante que intenta matarlo. El sagaz y

306
temerario agente 007 logra, por supuesto, escapar con éxito
de los tentáculos y mandíbulas del monstruo.
Uno de los más famosos escritores de ciencia ficción
del siglo XX, Arthur C. Clarke, estuvo desde niño cautivado
por el Architeuthis, tratándolo e incorporándolo en
numerosos de sus relatos. En Childhood’s End (El final de la
infancia) publicada en 1953, los calamares gigantes son
traídos a colación, aunque la historia, trata sobre el fin del
mundo y poco tiene que ver con ellos. En The Shining Ones
(1962), traducido como Criaturas abisales, se juega con la
idea de la posible inteligencia de los calamares gigantes, que
serían capaces, entre otras cosas, de comunicarse entre sí
por medio de señales luminosas. En el corto relato Big Game
Hunt (El gran juego de la caza) aparecido en 1957, Clarke
cuenta la historia de un biólogo que intenta, fallidamente,
aplicar la pesca eléctrica a los calamares gigantes. Ese
mismo año, se publicó el relato de ficción The Deep Range
(En las profundidades) donde el autor, pese a conocer bien el
nombre científico correcto del calamar gigante, le nombra
como Bathyteuthis maximus, que traducido significa el
calamar más grande de las profundidades abisales. En esta
novela, que transcurre en el sugerente ambiente de ficción
de una granja submarina de ballenas y cachalotes, varios
científicos y granjeros tratarán de capturar a un monstruo -
Percy- que deja enormes marcas de sus ventosas en los
lomos y cabezas de los cetáceos y que puede ahogarlos al
mantenerlos en las profundidades marinas sujetándolos con
sus largos brazos. Un famoso centro donde se exhiben
animales marinos ofrece bastante dinero por un calamar
gigante vivo para sus exposiciones. Con objeto de capturarlo
se realizan inmersiones con un submarino que desciende
hasta cerca de 1200 metros de profundidad. En el primer
encuentro con el gigante, además de filmarlo, los tripulantes
del submarino logran clavarle un dardo con un narcótico,
“que no le resultará dañino”. Luego se narra la aventura para
localizar y tratar de llevar a la superficie al anestesiado Percy
hasta que es encerrado en un gran tanque. Aquí, como
posteriormente en el artículo publicado en 1992 en la revista
Omni cuyo título es: ¡Los Calamares! En defensa de una

307
noble criatura, Clarke trata de mostrar a los lectores que
Percy no es un ser malévolo sino un animal inteligente y
tímido que huye de los cachalotes y mora en las
profundidades marinas. Lejos de ser un peligro para el
hombre, se trata de una criatura que debe protegerse, igual
que debe hacerse con su hábitat natural. Clarke quiere así
salir al paso de la influencia negativa de la novela The Beast,
luego convertida en película, escrita por Peter Benchley,
donde un calamar de más de treinta metros es un cazador
inteligente, poderoso, vengativo y antropófago que provoca
el pánico, horror y odio entre los humanos. Se trata de una
historia de ficción del mismo autor que se hizo famoso con
las novelas y películas Jaws, que es anterior a The Beast, y
The White Shark (Tiburón Blanco) publicada en 1994. La
temática del Kraken volverá a aparecer en otros relatos de
Clarke como Challenge of the Deep (El reto de las
profundidades) de 1966, o en The Ghost from the Grand
Bank (El fantasma del Gran Banco) publicada en 1990.
La batalla entre el hombre y el calamar gigante, una
deformación de los hechos históricos ocurridos en la cala de
Portugal (Terranova) de 1871, y recogidos por el reverendo
Harvey, es narrada por N. Duncan en la Antología de
literatura infantil publicada en Canadá en 1940. Casi
cincuenta y cinco años después, la escritora estadounidense
Annie Prouxl cuenta ese acontecimiento, ya legendario, en su
novela Shipping News (1993), por la que obtuvo el premio
Pulitzer. Y dos años más tarde, en la novela The Kraken,
escrita por D. Reed para un público juvenil, aparece
nuevamente Tom Piccot, un personaje probablemente
inventado por Harvey, como protagonista de una heroica
batalla que nunca existió.
Architeuthis ha sido usado en varias novelas de
ciencia ficción relativas al espacio, tanto interior como
exterior, y a los viajes interestelares. Así ocurre en la novela
Kraken acecha de John Wyndham, publicada en inglés en
1957 y traducida al español en 1965, donde aparecen
criaturas interestelares semejantes a calamares gigantes,
que son capaces de devorar de un bocado enormes naves
espaciales. Charles Sheffield en La Teleraña entre los

308
mundos (1979) crea un mundo futuro donde un millonario
posee un calamar gigante inteligente en el interior de una
estación espacial. Este ser responde al nombre de Calibán,
famoso en la literatura de ficción. Y en la novela de ficción
Esfera de Michael Crichton, autor de Parque Jurásico,
publicada en 1987, un científico transformado por efecto de
una esfera extraterrestre, exterioriza sus miedos en forma de
calamar gigante. En El final de la tierra, de Frederik Pohl y
Jack Williamson (1979), aparecen calamares adiestrados
para actuar como obreros de los humanos. Por otra parte, el
carácter depredador del calamar gigante es fuente de
inspiración para Dan Simmons, autor contemporáneo de
terror y ficción, para quien los centinelas de Matrix y a las
naves éxter del Hyperion son calamares. Como ejemplo de
traducción curiosa, cabe mencionar al calamar que porta el
delfín militar cibernético de Johnny Mnemónic, relato de
ficción escrito por William Gibson en 1981. Este calamar no
es otra cosa que un cefalópodo que permite al delfín tanto
detectar submarinos a larga distancia como leer y manipular
las señales de delicados equipos electrónicos.
No se acaban aquí los relatos que tratan de los
calamares gigantes y si usted se decide a buscar en Internet
encontrará, entre otros, títulos como: Tentacles de S.R.
Redmon, Giant squid de J. Dussling y P. Johson, Outside and
Inside Giant Squid de S. Markle, The incredible hunt for the
giant squid de B. Matsen, y el libro Giant squid: mystery of
the Deep de J. Dussling y P. Johson.
En Caneliñas, un lugar de la costa norte de Galicia
donde hasta hace pocos años existió una factoría de
ballenas, todavía quedan marineros que recuerdan la caza de
esos magníficos animales a su paso por el Finisterre europeo.
No es infrecuente oír contar a esas personas que muchos de
los cachalotes que eran cazados regurgitaban grandes trozos
de calamares, que, a juzgar por su tamaño, debían ser de
grandes dimensiones. Parecidas historias podían escucharse
no hace demasiados años en otros puertos con tradición
ballenera. Esos relatos, transmitidos oralmente, han sido
recogidos en algunas publicaciones. Quizás las dos más
antiguas se deben a Herman Neville en su famosa novela

309
Moby Dick (1851) y a Charles Nordhoff en su libro Whaling
and Fishing de 1856, pero que se encuentran también en The
Whale and his Captors de Henry Cheever, escrita en 1859, y
en The Cruise of the Cachalot y Denizens of the Deep de Fran
Bullen, escritas en 1989 y 1904, respectivamente.
La relación entre una presa de grandes dimensiones
(Architeuthis) y su principal depredador (el cachalote,
Physter macrocephalus) ha excitado la imaginación de
numerosos escritores. Las grandes marcas de ventosas que
aparecían en la piel del cuerpo de los cachalotes, de hasta 37
mm de diámetro, provocaron las exageraciones sobre el
tamaño de los calamares gigantes, ya que la extrapolación
de esas marcas, exageradas por la deformación,
proporcionaba calamares de varias toneladas.
Esas marcas también contribuyeron a acrecentar la
supuesta ferocidad de Architeuthis, que, para la imaginación
de los novelistas, podría entablar colosales luchas con los
cachalotes, y salir victorioso en algunas ocasiones. Esa
potencial batalla en las profundidades, e incluso en la
superficie del mar, donde los tentáculos del calamar gigante
actuarían a modo de grandes serpientes que se enroscarían
en el cuerpo de los cachalotes, también ha desatado la
imaginación de algunos científicos, como es el caso del
biólogo marino Victor Scheffer, quien, después de relatar
muchos aspectos bien documentados científicamente sobre
los grandes cetáceos en su libro The Year of the Whale
(1996), desata sus dotes creativas y se imagina una gran
lucha entre ambos colosos, que seguramente poco debe
tener que ver con la realidad.
El encuentro de un Architeuthis con un cachalote
aparece también, entre otras, en la novela Soundings escrita
por Henk Searls y publicada en 1982. Por otra parte, además
de narraciones, esta batalla entre gigantes ha sido tema de
numerosas e ingeniosas ilustraciones, todas ellas imaginadas
porque nadie todavía ha podido presenciar lo que acontece
en el encuentro entre un cachalote, que se sumerge a varios
cientos de metros de profundidad, y su presa el Architeuthis.
El término criptozoología fue acuñado por el zoólogo
belga Bernard Heuvelmans a mediados del siglo XX. La

310
criptozoología es, etimológicamente, el tratado de los
animales ocultos. Es el estudio objetivo de los animales que
únicamente son conocidos por indicios considerados
insuficientes por la mayoría (testimonios, fotografías, rastros
o piezas anatómicas), y que por consiguiente no han sido catalogados oficialmente. En el libro de Heuvelmans Dans le sillage
des monstres marins: le Kraken et le Poulpe Colossal publicado en
francés en 1958, y traducido al español en 1975 como Tras la
estela de los monstruos marinos, se encuentra la historia completa
del descubrimiento de los calamares gigantes Architeuthis,
que resulta muy ilustrativa desde el punto de vista histórico y
legendario, y que se trató en el capítulo segundo de este
libro.
Las obras de Heuvelmans, así como su intenso trabajo
realizado desde el centro de criptozoología que fundó en
1975 y cuya sede esta cerca de Le Bugue en el sur de
Francia, han influido notablemente en todos los estudiosos de
esta materia, la mayoría enmarcados dentro de la Sociedad
Internacional de Criptozoología. Estos relatos han influido
también notablemente a un amplio espectro del gran público,
cada vez más receptivo a este tipo de misterios, que llenan
de alguna manera la insatisfacción que produce en el espíritu
humano la crudeza y racionalidad de la ciencia experimental.
Sin lugar a dudas, el libro más completo sobre
Architeuthis escrito hasta la fecha es el del explorador,
artista y biólogo marino Richard Ellis, titulado The Search for
the Giant Squid, que se publicó en 1998. En este libro, con
un estilo divulgativo pero a la vez preciso, se proporcionan
un texto y unas ilustraciones que constituyen una síntesis de
la historia de las primeras investigaciones sobre los
calamares gigantes y el impacto que estas criaturas marinas
han tenido en la cultura y la historia.
Películas, documentales y otros
Los calamares y pulpos gigantes han aparecido en
infinidad de ocasiones en la historia del séptimo arte. Aparte
de las películas basadas en novelas dónde ya aparecían estos
fabulosos animales, como las ya mencionadas Veinte mil

311
leguas de viaje submarino de Julio Verne y Esfera de Michael
Crichton, se recurrió a estos “monstruos” de muchas formas.
Es curioso observar como la aparición de los calamares y
pulpos gigantes en el cine presenta un desarrollo
directamente relacionado con la evolución de la ciencia
ficción y los temores de la sociedad en relación con los
avances tecnológicos. En las películas de la primera mitad del
siglo XX se asumía la existencia de estos monstruos
gigantescos en las profundidades aún inexploradas del
océano, dónde eran perturbados por uno u otro
acontecimiento, ya fuera natural o causado por los avances
científicos. Luego, durante la segunda mitad del siglo XX, se
empezaron a buscar “explicaciones” a su aparición dirigidas
hacia mutaciones causadas por la radiación, la contaminación
o cambios genéticos.
En The Mysterious Island, producida en 1929, los
tripulantes de dos submarinos enemigos se persiguen a
través del océano hasta llegar a un extraño lugar lleno de
dragones y calamares gigantes. Acercándose a la mitad del
siglo XX, en la película Wake of the Red Witch, producida por
Cecil B. De Mille en 1942, un terrible calamar gigante da
muerte al personaje interpretado por John Wayne, siendo
esta una de las pocas películas en que este actor “muere”.
Algunos ejemplos de películas de mediados del siglo
XX en las que los “calamares gigantes” participaron son:
Killers of the Sea donde un guardia de Florida se dedica a
eliminar a criaturas marinas dañinas para el negocio de los
pescadores del lugar, entre ellos un enorme pulpo. En
Monster from the Ocean Floor un muy joven Robert Wagner
interpreta a un buzo contratado por un biólogo marino para
capturar a una especie de pulpo gigante.
Incluso en los dibujos animados, como la serie de
Beany y Cecil, dónde un niño vive aventuras junto a una
serpiente marina, conocen a un calamar en el episodio Beany
and Cecil meet Billy the Squid de 1959.
La radiación es la principal causa de la aparición de los
calamares y pulpos gigantes en las películas y series de la
segunda mitad del siglo XX. Para ello sólo un par de
ejemplos: En It Came from Beneath the Sea aparece un

312
pulpo gigante producido por la radiación. En Tentacles, John
Huston y Henry Fonda tienen que luchar también contra un
pulpo gigante producido por la contaminación. También por
esta época aparecen películas de serie B, como Octaman,
dónde un pulpo gigante muy parecido a un hombre con
tentáculos protege a otros cefalópodos sometidos a
experimentos por “malvados científicos”.
Hay que mencionar películas como Viaje al fondo del
mar de 1961, en la que aparecen un par de calamares
durante el viaje del submarino. Además, en la serie de
televisión que generó dicha película, producida durante los
años 60, más de un calamar, pulpo gigante o ser parecido
figuraron en capítulos de la serie.
El cine y las series de televisión japonesas han
aportado generosamente argumentos, calamares gigantes,
Fig. 79. Carteles de dos películas de mediados del siglo XX donde intervienen cefalópodos gigantes: Reap the Wild Wind con John
Wayne y Monster from the Ocean Floor con Robert Wagner.

313
pulpos gigantes y “tentáculos” a nuestra vida. El mítico
Godzilla, un devastador monstruo generado por las
radiaciones de desastres como los de Hiroshima y Nagasaki,
luchará con una serie de monstruos como Mosura (Motha),
Gamera, Polillas, galápagos gigantes y dinosaurios
antediluvianos también generados por los ensayos atómicos.
Entre ellos, Godzilla se enfrentará a Viras, un calamar del
espacio de 96 metros de largo y 120 toneladas y a Daidako,
un poderoso pulpo gigante. El mismo Viras se enfrentará en
otras películas con Gamera, otro ser de la “mitología”
monstruosa del Japón.
Y, junto a Godzilla, nacieron una gran cantidad de
series japonesas de ciencia ficción, como Ultraman y Ultra
Seven, donde abundaban los monstruos llenos de tentáculos
de calamar que luchaban contra el superhéroe de turno, ya
sea un extraterrestre bueno que protege la tierra, un súper
robot, un equipo de humanos provisto de la última tecnología
o la combinación de éstos. Un ejemplo reciente de este tipo
de series, llevado a la televisión norteamericana, son los
Power Rangers.
Una especial mención merece el cine y las series de
Anime, un tipo de dibujo animado generado en Japón y
heredero directo de series como Godzilla. Muchas series y
películas de animación tienen seres y monstruos con
tentáculos, e incluso calamares, contra los cuales los
protagonistas deben luchar y donde la destrucción, ya sea
universal o no, está a la vuelta de la esquina.
Tampoco se puede olvidar al mejor de todos los espías
del mundo, James Bond el agente 007, que en el Doctor No
(1962) tiene que escapar, al igual que en la novela, de la
fiereza de un calamar gigante. Otro encuentro, esta vez con
los pulpos, tiene el agente 007 en Octopusi (1983), aunque
esta vez recibe una pequeña ayuda de ellos.
En The Abyss (1989) dirigida por James Cameron, un
grupo de buzos prospectores de petróleo, que participan en
una misión de rescate de un submarino nuclear accidentado
cerca de una profunda fosa submarina, entablan contacto
con alienígenas que tienen un ligero parecido a calamares y

314
usan tentáculos de agua para explorar la base submarina de
los humanos.
En 1996 la película para televisión The Beast, basada
en una novela de Peter Benchley, muestra la lucha de un
pueblo contra un calamar gigante que asesina a varios de sus
habitantes y arruina el turismo. Tampoco podemos olvidar
Men in Black (1997), donde podemos ver extraterrestres con
tentáculos tipo calamar.
En el clásico de la ciencia ficción Dark City (1998)
podemos ver a un grupo de humanos que viven en una
ciudad donde siempre es de noche, la cual realmente es una
nave espacial controlada por extraterrestres parecidos a
calamares con poderes mentales. Incidentalmente, esta
película se considera precursora de Matrix (1999), donde los
robots que persiguen a los humanos son muy parecidos a
calamares gigantes, aunque hay que indicar que los
argumentos de ambas películas son diferentes.
En el comienzo del siglo XXI varias películas,
conocidas o no, tienen entre sus personajes a los calamares
gigantes. En primer lugar, los calamares y pulpos gigantes
están presentes, aunque brevemente, en El Señor de los
Anillos: La comunidad del Anillo (2001), basada en la novela
de R.R. Tolkien, donde un monstruo con tentáculos y
mandíbula típicos de calamar gigante ataca a los personajes
justo antes de entrar a las minas de Morya. Sin embargo,
hay otras películas en las que los calamares y pulpos
gigantes son realmente los protagonistas. Una de ellas es la
película de terror, dirigida por John Eyres en el 2000,
Octopus, donde la tripulación de un submarino nuclear que
investiga las desapariciones de barcos en el triangulo de las
Bermudas en busca de actos terroristas, se encuentra con un
terrorífico pulpo gigante.
Incluso en España se pueden encontrar películas
donde figuran los calamares gigantes. Un ejemplo reciente es
Kárate a Muerte en Torremolinos (2001) de Pedro Temboury.
Se trata de una delirante sátira de las series tipo B en la que
el malévolo doctor Malvedades llega a Torremolinos con la
intención despertar al monstruo Jocántaro, un ser mitad
centollo mitad calamar gigante.

315
Pero los calamares gigantes también son utilizados en
películas de animación como The Hungry Squid (2001),
producida en Canadá, que trata los problemas de una
adolescente con unos padres que apenas están con ella, y
cuyo pueblo es invadido por una serie de criaturas gigantes,
incluido un gigantesco calamar hambriento. Otras películas
de animación donde aparecen los calamares de una u otra
forma, son La Sirenita (1989) donde la malvada bruja tiene
tentáculos de pulpo en vez de piernas, y Simbad: La leyenda
de los siete mares (2003) en la cual los protagonistas son
perseguidos por un enorme calamar gigante.
En la actualidad, podemos encontrar dos películas
donde figuran los cefalópodos de una u otra forma. En la
película japonesa The Calamari Wrestler (2004), que
recuerda mucho a las series de ciencia ficción como
Ultraman, los Power Rangers u otras series juveniles de este
tipo, un “calamar gigante” se convierte en un campeón de
lucha enfrentándose a varios contrincantes. Regresando al
cine americano, aunque con origen en los comics de
mediados del siglo XX, en la recién estrenada Spiderman 2
(2004), se cuenta el enfrentamiento del superhéroe con uno
de sus más conocidos archienemigos, el Dr. Octopus.
Así, ya sea considerándolos en toda su realidad, es
decir, como criaturas de las profundidades, o como
monstruos que amenazan con destruir al mundo, o bien
aportando alguna de sus características, sobre todo
tentáculos, a diversas criaturas y robots, los calamares
gigantes están muy presentes en el cine y la televisión
fantástica y de ciencia ficción, aunque también se han
utilizado en los géneros de terror y comedia. Sin embargo,
los cefalópodos no se pueden considerar como simples
recursos a utilizar por el cine para sus películas ya que por si
mismos son organismos muy interesantes y difíciles de filmar
en los océanos. Y es en este sentido que desde hace tiempo
estos maravillosos pobladores de los mares son objeto, cada
vez mayor, de la realización de documentales.
Existe un gran número de documentales sobre los
organismos marinos en los cuales podemos encontrar
cefalópodos, como pueden ser la serie de 2002 Planeta Azul

316
producida por la BBC. También en la serie El mundo
submarino producida por Jacques Cousteau entre1970 y
1980 con los capítulos La Noche del Calamar y El Gran Pulpo,
o el documental de la BBC Aliens From Inner Space de 1997,
donde se aprecia la belleza y las características de estos
organismos. Sin embargo, en estos documentales, así como
en otros que aquí no se mencionan, no aparecen calamares
gigantes.
En los años recientes la idea de filmar “vivo” un
calamar gigante ha cobrado cada vez más fuerza entre los
productores de documentales, deseosos de ofrecer
novedades. Y, esta es la razón por la que han nacido los
documentales sobre calamares gigantes. El primero de ellos
fue Sea Monsters: Search for the Giant Squid producido por
la National Geographic en 1998, donde los expertos tratan de
filmar un Architeuthis utilizando cámaras submarinas sujetas
al lomo de cachalotes. El documental muestra filmaciones
anteriores de ejemplares varados y unas extraordinarias
imágenes de las profundidades realizadas por las cámaras
mientras los cachalotes bajan a comer, pero ninguna del
calamar gigante.
En 2001 Discovery Channel produjo el documental
Quest for the Giant Squid, en el cual la búsqueda del calamar
gigante se centró en las profundidades del mar de Cortez en
el golfo de México, y donde se utilizan excelente imágenes
del calamar de la corriente de Humboldt, Dosidicus gigas.
Aunque el documental no llega a presentar filmaciones de un
Architeuthis, ya que de hecho no consiguen siquiera
acercarse a uno, los científicos del Instituto Oceanográfico de
Woods Hole muestran muy buenas escenas submarinas de
otras especies. Siguiendo con la idea de los calamares
gigantes, pero esta vez centrados en Dosidicus gigas,
National Geographic produjo el documental Giant Squids
(2003), donde se muestra espectaculares imágenes de estos
cefalópodos en las profundas aguas azules del mar de
Cortez. Estas filmaciones no están exentas de peligro ya que
estos Dosidicus gigas es un depredador muy voraz y
agresivo, los pescadores locales llaman diablo rojo. Estos
animales se congregan en cardúmenes que pueden llegar a

317
contar con cientos de individuos siendo más temidos que los
propios tiburones.
A primeros de 2004 apareció el primer documental
sobre calamares gigantes producido íntegramente en
España: Proyecto Kraken: en busca del Calamar Gigante. En
esta película, de 53 minutos de duración, se relatan los
intentos realizados por ECOBIOMAR, en asociación con la
productora de documentales Transglobe Films, por filmar
vivo un Architeuthis en las profundidades de Carrandi, frente
a las costas de Asturias. Aunque la expedición no logró filmar
un calamar gigante, el documental tiene gran valor por ser el
primero producido en nuestro país sobre este tema en
directa competencia con Discovery Channel y National
Geographic. Instituciones que, a pesar de haber invertido
mucho más en recursos y fondos que la producción española,
aún no han obtenido resultados.
En agosto de 2004 salió el último documental
producido sobre los calamares gigantes, Chasing Giants: On
the Trail of the Giant Squid producido por el Discovery
Channel. En este documental se muestran las investigaciones
Fig. 80. El primer documental sobre los calamares gigantes
producido en España: Proyecto Kraken: en busca del Calamar Gigante. A la izquierda, algunos de los miembros que participaron
en el documental y, a la derecha portada del documental en su versión en Inglés.

318
de Steve O’Shea sobre los calamares gigantes en el cañón de
Kaikoura en Nueva Zelanda y su búsqueda para filmar uno
de ellos en su hábitat natural. Un gran aporte de este
documental es la observación y filmación, por primera vez,
de lo que el investigador identificó como paralarvas vivas de
Architeuthis.
A finales del mes de septiembre de 2005 saltó a los
medios de comunicación la noticia de que unos
investigadores japoneses habían conseguido, por primera
vez, fotografías de un calamar gigante vivo en su medio
natural. Efectivamente así ha sido. Después de tres años de
intentos (2002-2004), a las 09.15 del 30 de septiembre de
2004, Tsunemi Kubodera y Kyoichi Mori consiguieron
fotografiar un Architeuthis atrapado por uno de sus
tentáculos en una potera cebada, ubicada a 900 m de
profundidad en las proximidades de las islas japonesas de
Ogasawara en el Pacífico norte (26º57.3’ N, 142º 16.8’E), en
un área donde los registros de cachalotes son bastante
frecuentes. En su publicación (2005), estos autores explican
que se obtuvieron 550 imágenes con una cámara digital,
durante las cuatro horas que el animal estuvo enganchado en
la potera. Después de lo cual, el tentáculo se rompió siendo
recuperado por los investigadores. La morfología de las
estructuras del tentáculo permitió identificar sin ninguna
duda al calamar, y su longitud (5,5 m) estimar un tamaño
total del animal algo superior a los 8 metros, presuponiendo
que el tentáculo se corto por su base.
Sin restar mérito a estos investigadores, que han sido
los primeros en fotografiar un calamar gigante vivo y en su
medio natural, cabe, no obstante, realizar algunas
observaciones, que restan importancia a los resultados, no
sólo desde un punto de vista científico sino, sobre todo,
filmográfico. En primer lugar, las imágenes obtenidas son de
un animal apresado por un arte de pesca y no totalmente
libre. Ello significa que el comportamiento observado es el de
una situación de máximo estrés. En segundo término, la
ubicación de la cámara, por encima de la potera y de los
cebos, así como el ángulo de visón del objetivo, únicamente
permitieron obtener imágenes de la corona braquial y la

319
cabeza del calamar. En tercer lugar, que se trata de
fotografías, cuando, tanto para estudios etológicos como
para fines documentales, hubiese sido muy preferible
grabación en video. Y por último, que, a juicio de varios
expertos en fotografía submarina, el color del agua que
muestran las fotos publicadas cuestiona que fuesen
obtenidas a 900 m de profundidad, aún considerando el uso
de dispositivos luminosos. Quizá estas, y otras
consideraciones que aquí se nos escapan, sean el motivo por
el que los autores han tardado un año, a nuestro juicio
demasiado tiempo, en dar a conocer su hallazgo, y , además,
no lo hayan hecho en alguna de las dos revistas científicas
multidisciplinares de mayor impacto.
Por lo tanto, todavía está pendiente el filmar un
Architeuthis vivo y libre en su medio natural.
Para finalizar, y ya como curiosidad, los calamares
gigantes no sólo están presentes en las películas y en los
documentales, sino que también en la música y el la filatelia.
En 2003 el compositor japonés de música experimental Koji
Asano presentó una colección de 4 CD con 14 piezas de
música experimental, compuesta por él mismo, a la que puso
como título The Giant Squid.
El calamar gigante ha resultado ser un tema atractivo
para la filatelia, existiendo numerosos países que han editado
sellos de correos dedicados a este cefalópodo desde hace
más de 30 años.

320
Fig. 81. Sellos relativos a los calamares gigantes emitidos en Australia, Canadá, Francia, Nueva Zelanda y Togo. (http://www.pibburns.com/cryptost/kraken.htm).
En resumen, los calamares y los pulpos gigantes han
pasado de ser leyendas mitológicas de siglos pasados a
monstruo protagonista en muchos libros, películas y series de
televisión. En su aventura cinematográfica han sido desde
misteriosos seres de las profundidades a mutantes causados,
primero por las radiaciones atómicas y luego por alteraciones
genéticas, para terminar incluso como calamares espaciales.
Por otra parte, han prestado sus tentáculos a muchas otras
criaturas en infinidad de películas, y ha sido sólo durante la
segunda mitad del siglo XX, cuando los calamares gigantes
han pasado del cine de ciencia ficción, fantástico o de terror
a los documentales, dónde han reasumido su real posición en
el medio marino, transformándose en uno de los principales
objetivos de las actuales expediciones teutológicas y grandes
productoras de documentales.
Expediciones
El ecosistema existente en el cañón submarino de
Kaikoura en Nueva Zelanda es uno de los pocos lugares del
mundo donde aparece frecuentemente el calamar gigante.
Con objeto de observarlo en su medio natural, lo que nadie
ha logrado todavía, la ya mencionada Smithsonian Institution
y la mundialmente conocida Nacional Geographic Society, en
colaboración con científicos y técnicos de todo el mundo,
organizaron la expedición Kaikoura. Después de una
operación preparatoria en las Azores que tuvo lugar en julio

321
de 1996, donde se probaron algunos equipos y metodologías,
se desarrolló la expedición Kaikoura 1997.
Con un presupuesto de cinco millones de dólares, esa
expedición internacional se puso en marcha con Clyde F. E.
Roper como jefe de los científicos, tarea en la que estuvo
apoyado, entre otros, por su mujer Ingrid, por el
investigador inglés Malcolm R. Clarke, por el ingeniero James
Bellingham del Massachusetts Institute of Technology, y
Emory Kristof, un veterano fotógrafo de la Nacional
Geographic Society. Bellingham diseñó un vehículo
submarino autónomo no tripulado, llamado Odyssey, con el
apoyo de la Marina de los Estados Unidos de América. Este
aparato fue el encargado de sumergirse en las profundidades
para obtener las imágenes del Kraken. Kristof había
desarrollado una cámara de video digital preparada para
filmar a profundidades de hasta 1.830 metros llamada
Ropecam, que fue muy valiosa durante la expedición. La
filosofía básica de campaña era utilizar a los cachalotes,
principales depredadores de Architeuthis y muy abundantes
en Kaikoura, para filmar al evasivo calamar gigante. Dos
equipos embarcados a bordo de dos barcos estuvieron
trabajando en la zona entre el 25 de febrero y el 13 de
marzo de 1997. Desde el Taneka, un pesquero neozelandés
de 14 metros de eslora, Roper coordinó las operaciones que
se desarrollaban en esa embarcación, así como las que lo
hicieron desde el Mystique, un yate de 25 metros de eslora,
que estaba comandado por Kristof. El Odyssey y la cámara
Ropecam hicieron sus tareas filmando numerosos organismos
de profundidad atraídos por los cebos químicos preparados a
bordo. Además se utilizó una cámara de video llamada
Crittercam, cuya principal peculiaridad es llevar una ventosa
en su parte inferior, gracias a la cual puede adherirse a la
cabeza de un cachalote. Esa difícil operación se logró con
éxito, pero la cámara, después de filmar interesantes
escenas de una manada de esos enormes cetáceos durante
unos minutos, se soltó y ya no hubo forma de poder volver a
ser utilizada. Esas imagines submarinas, junto con las que la
Crittercam había obtenido en las Azores, fueron utilizadas por
National Geographic Television para producir un documental

322
sobre los cachalotes y su relación con Architeuthis, que, pese
a todos los esfuerzos, no había podido filmarse.
Search of the giant squid New Zealand 1999, fue la
segunda expedición en busca de poder filmar a los calamares
gigantes en su ambiente natural. Esta expedición se
desarrolló entre el 19 de febrero y el 26 de marzo de 1999,
también en el cañón submarino de Kaikoura. En esta ocasión
Roper contó con el apoyo del investigador neozelandés
Steven O’Shea, entonces en Auckland, buen conocedor de la
zona y también experto en calamares gigantes.

Fig. 82. Localización y perfil orográfico del cañón de Kaikoura en aguas de Nueva Zelanda.

324
Esta expedición se llevó a cabo a bordo del Kaharoa,
buque de investigación del National Institute of Water and
Atmospheric Research (NIWA), utlizándose el Deep Rover, un
vehículo submarino autónomo tripulado, capaz de sumergirse
hasta 900 metros de profundidad y desarrollado con
tecnología canadiense. El esfuerzo multinacional comprendía
científicos, tecnólogos, cámaras y tripulantes del submarino y
del barco de investigación pertenecientes a cuatro países: los
Estados Unidos de Norteamérica, Nueva Zelanda, Gran
Bretaña y Canadá. El proyecto fue patrocinado por diez
instituciones encabezadas por la Smithsonian Institution. La
expedición estuvo afectada negativamente por las adversas
condiciones atmosféricas reinantes, así como por varios
problemas técnicos, y no se consiguió el principal objetivo,
aunque se obtuvieron interesantes imágenes de la fauna
abisal del cañón así como de los cachalotes y otros
mamíferos marinos abundantes en Kaikoura, que se han
utilizado en numerosos proyectos culturales y exposiciones
realizados en los diversos países participantes. Roper, en la
carta que dirigió a todos los miembros de la expedición,
mantiene el optimismo de que en próximas expediciones se
pueda lograr filmar al huidizo Kraken.
Desgraciadamente, la salud le jugó una mala pasada a
Clyde Roper y tuvo que ser intervenido quirúrgicamente para
corregir graves lesiones cardiacas. Actualmente, ya retirado
y recuperado, conserva intacta todavía su ilusión por el
proyecto, pero el relevo parece haberlo tomado Steve
O’Shea, quien a principios del 2003 comunicó en su página
web la captura de paralarvas de Architeuthis, de menos de 1
cm cada una, mediante una red RMT50, y el fallido intento de
mantenerlas vivas en condiciones experimentales. Este fue el
principal resultado de una expedición en la que también
colaboraron C.C. Lu y M.R. Clarke, realizada bajo el
patrocinio de Dicovery Channel. O’Shea también ha
comunicado la organización de dos nuevas expediciones para
2004, una basada en pescas de zooplancton para tratar de
conseguir nuevas paralarvas de estas criaturas e intentar de
mantenerlas vivas, y otra cuyo objetivo será tratar de
filmarlo.

325
En las antípodas de Nueva Zelanda, concretamente
frente a las costas de Asturias, y desde hace muchos años,
los ejemplares de Architeuthis son tema de conversación
para las gentes del mar. Ellos los llaman peludines, debido al
aspecto pelado que muestran cuando son izados a bordo
después de un arrastre de varias horas, avistados flotando
sobre la superficie del mar o varados en la costa. Poca
atención habían recibido estos gigantes por parte de los
científicos asturianos, y españoles, hasta que Luis Laria,
presidente de la Coordinadora para la Protección de las
Especies Marinas (CEPESMA) llamó la atención sobre ellos,
contactando con el grupo de investigación de Ecología y
Biodiversidad (ECOBIOMAR) del Instituto de Investigaciones
Marinas que el Consejo Superior de Investigaciones
Científicas tiene en Vigo. Allí, el jefe del equipo, Ángel
Guerra, y sus colaboradores Ángel F. González, Francisco
Rocha y Jaime Otero, llevaban años estudiando los
cefalópodos y ya habían tenido en sus manos varios
ejemplares de calamar gigante, procedentes de Namibia y
Sudáfrica. En el invierno de 1999 se efectuó en Luarca la
disecación del primer ejemplar, una hembra inmadura de
104 Kg de peso, y desde entonces las operaciones de esta
índole se han sucedido ininterrumpidamente, con lo que se
han podido examinar 17 ejemplares hasta la fecha. La
frecuencia de aparición de los calamares gigantes en los
caladeros de Carrandi, próximos a los cañones submarinos
de Llanes, Lastre y Avilés, la proximidad de esta zona, que se
localiza a menos de 30 millas al norte de la costa, y la
existencia de un puerto base (Gijón) con todas las facilidades
para operaciones logísticas de gran envergadura, llevó a los
miembros de este grupo a sopesar la posibilidad de organizar
expediciones científicas para tratar de filmar, por primera
vez, esas extraordinarias y huidizas criaturas. Así nació el
Proyecto KRAKEN, que se concretó gracias a la colaboración
con la productora de documentales TRANSGLOBE FILMS, con
objeto de tener éxito donde las expediciones internacionales
que se habían realizado en Kaikoura habían fracasado.
Ese proyecto se concibió estructurado en cinco fases:
en primer lugar, una precampaña en 2001 para conocer el

326
fondo la batimetría, topografía y el ecosistema donde moran
esas criaturas en Asturias; en segundo término, varias
expediciones, la segunda realizada en el verano de 2002; en
tercer lugar, la producción de un documental que fue emitido
por primera vez en 2004; en cuarto, la presentación de los
resultados en exposiciones realizadas por toda España, que
Fig. 83. Parte de la campaña del proyecto KRAKEN, tal como fue
presentada en un cuadro explicativo para la prensa (modificado de infografía de EL MUNDO).

327
comenzaron en 2003 y aún siguen; y por último, en la
edición de un libro que resumiera, entre otras cosas, todas
las operaciones, resultados y descubrimientos del proyecto.
La expedición preliminar o precampaña zarpó del
puerto de Gijón el 5 de octubre de 2001 a bordo de los
buques Científico e Investigador, de 52 y 44 metros de
eslora, respectivamente. Veintidós miembros de la
expedición, entre científicos y técnicos, bajo la dirección de
Ángel Guerra, y dieciocho tripulantes se involucraron en el
proyecto durante una intensa semana. La enorme
profundidad en la que viven estos animales, requirió el
empleo de material técnico muy sofisticado, capaz de
soportar presiones de hasta 100 atmósferas a temperaturas
cercanas a los 2 grados centígrados. Los aparatos empleados
para filmar en completa oscuridad al Kraken fueron tres
sistemas de cámaras sujetas a boyas en superficie, dentro de
las cuales había un sofisticado sistema de grabación para
más de quince días de trabajo ininterrumpido, cuyo diseño
original fue obra de Joaquín Gracia, una cámara móvil, y un
vehículo submarino operado por control remoto (ROV) capaz
de descender hasta 300 m de profundidad, y así mismo
equipado con otra cámara.
Fig. 84. Durante el proyecto KRAKEN se tuvieron que realizar ajustes y arreglos en las boyas de las cámaras.

328
Todos estos sistemas eran controlados y estaban
comunicados con la cabina de recepción - Calipso - mediante
cable de fibra óptica o radio señal. Parte importante en esta
expedición fueron los sistemas de atracción utilizados,
compuestos por cebos acústicos, luminosos y orgánicos. Tras
levantar un plano topográfico detallado del fondo mediante
ecosondas, se delimitó la zona de trabajo, consistente en un
triángulo equilátero de dos millas de lado, en cuyos vértices
se logró colocar una de las tres boyas mencionadas, con su
cámara fija. Durante esta campaña se planeó la realización
de perfiles de temperatura, salinidad y profundidad mediante
un CTD, así como pescas de plancton mediante redes de
Bongo. Todo ello con objeto de precisar las condiciones
hidrográficas del hábitat donde viven los calamares gigantes
en el caladero de Carrandi, y tratar de capturar larvas y
juveniles de los mismos, que son desconocidas. Pese al mal
tiempo atmosférico, siempre presente, y a que no todos los
aparatos y planes funcionaron correctamente, en esa
precampaña se puso a punto la metodología de trabajo, se
conjuntaron los equipos humanos, a la par que se
adquirieron importantes datos que proporcionaron una visión
bastante precisa, y novedosa, de las características del
ecosistema donde moran estos gigantes marinos.
La segunda campaña se desarrolló del 10 al 27 de
septiembre de 2002. Está vez la meteorología acompañó y se
pudieron situar con facilidad las tres cámaras fijas en los
vértices del triángulo, que filmarían a 550, 660 y 780 metros
de profundidad, grabando durante 24 horas seguidas los
quince días de la campaña.
Desafortunadamente, y por causas desconocidas, la
primera boya desapareció al sexto día llevándose con ella
toda la grabación efectuada. Esa boya estaba ubicada a poco
más de doscientos metros de donde una pareja de
arrastreros capturó un Architeuthis macho de 43 Kg, durante
la realización de la expedición. El ejemplar fue trasladado a
bordo del Investigador y examinado por los sorprendidos
científicos. Se trataba del primer macho capturado al sur del
paralelo 55ºN, estaba funcionalmente maduro, y mostraba
esperma en varios de sus brazos.

329
Fig. 85. Ejemplar de Architeuthis macho capturado por una pareja de arrastreros junto a la zona donde se instaló una de las cámaras
de la segunda campaña.
Durante esta campaña también se trabajó con la
cámara móvil y con el vehículo submarino autónomo,
obteniéndose cientos de horas de filmación que guardaban
imágenes originales y novedosas de organismos marinos de
diferentes ecosistemas, pero ninguna del calamar gigante. En
la primavera de 2003 se inauguró en el Museo Nacional de
Ciencias Naturales, en Madrid, una exposición sobre el
proyecto donde se mostraron los materiales utilizados.
Durante seis meses, los visitantes pudieron contemplar el
interior de las boyas donde se instaló el sofisticado sistema
de grabación en discos duros, las cámaras y los cebos
utilizados, el buque Investigador y los sistemas de
posicionamiento con los que se instalaron las cámaras, las
imágenes del abismo y las criaturas que lo pueblan. En
definitiva, toda la información de lo que ha sido la campaña
y de lo que es el abismo cantábrico donde se llevaron a cabo
los rodajes. Por supuesto, también pudieron conocer de
primera mano todo lo que se sabe hasta la fecha sobre los
calamares gigantes y, en concreto, sobre las dos especies
“estrella” del caladero de Carrandi: el Architeuthis y el

330
Taningia danae con dos espectaculares maquetas a tamaño
real, con las que el público, que acudió en gran número al
Museo, pudo sentir la sensación de estar cara a cara frente a
un auténtico Kraken. El éxito de esa exposición ha llevado a
que el Museo del Mar de Galicia, en Vigo, se interesara por
ella, allí se inaugurará en el transcurso de 2005. Por otra
parte, el 1 de marzo de 2004, la segunda cadena de
Televisión Española, emitió el documental de 53 minutos El
proyecto Kraken: en busca del calamar gigante. Dirigido por
Fernando González Sitges y producido por Javier Linares, ese
documental de Transglobe Films muestra en imágenes las
vivencias de este proyecto, así como la biología del calamar
gigante, su historia y la aventura que supone intentar
capturarlo en imágenes. Ya se ha traducido a varios idiomas
y será proyectado en otras cadenas de televisión de todo el
mundo.
Fig. 86. Una escena durante el rodaje del documental: El Proyecto
Kraken, en busca del calamar gigante.

331
La ilusión de ese equipo por ser los primeros en
obtener las imágenes de un Kraken vivo frente a las costas
de Asturias está intacta, y ya se planea realizar una nueva
expedición en septiembre de 2005, con técnicas y
metodologías de última generación. Aunque no se puede
olvidar que quizás sea en Nueva Zelanda donde caiga la
suerte, y se defina así la incertidumbre abierta por la
competición deportiva que existe entre los investigadores
que indagan en el cañón submarino de Kaikoura y los que lo
hacen en las fosas de Asturias oriental en busca del
enigmático Kraken.
Los Peludines de Asturias
Muy posiblemente sea la costa Asturiana la única zona
de Europa en la que los calamares gigantes del género
Architeuthis tienen un nombre común, ya que son conocidos
desde tiempos remotos por los lugareños con el nombre de
peludines. Esta denominación vernácula también se da a
otros cefalópodos de tamaño considerable como a la pota
saltadora (Ommastrephes bartramii), especie que puede
considerarse más habitual en estas aguas, y cuyos
especimenes pueden encontrarse con relativa frecuencia
varados en pedreros o arenales, sobre todo en los meses de
otoño. Sin embargo, a estos animales se les puede distinguir
fácilmente por mantener una epidermis mucho más
consistente que los calamares gigantes.
Este nombre común tiene su origen en el estado en el
que se encuentran los ejemplares que aparecen ya sea
varados o son capturados por los arrastreros. En ambos
casos, debido al contacto del cuerpo con el sustrato o con la
red, la epidermis queda fragmentada y pelada en su mayor
parte, eliminando su característico color vinoso y dejando al
descubierto la coloración más pálida y marfil de la
musculatura. Curiosamente, sin embargo, al calamar gigante
se le llama Peludín sólo en la zona central y occidental de la
costa asturiana, cuando la mayoría de los registros más
importantes de su presencia se producen en la oriental.

332
Desde el año 1962, un 80% de los registros de
Peludines en Asturias (Tabla 11) pueden considerarse
provenientes de capturas accidentales, con excepción de 5
ejemplares en el año 2001 y otros 4 en el 2003, que
aparecieron coincidiendo con la presencia de barcos que
realizaban trabajos de investigaciones geofísicas, muy
probablemente relacionadas con la búsqueda de bolsas de
gas y de petróleo en la región meridional del Golfo de
Vizcaya. Como ya se ha expuesto, las actividades de dichos
barcos fueron coincidentes en espacio y tiempo con los
varamientos, y puede que influyeran decisivamente en sus
muertes.
Según las informaciones recabadas a los patrones de
barcos que faenan con artes de arrastre en la zona era
habitual tirar por la borda ejemplares de calamar gigante
capturados por la red para evitar trabajo innecesario,
aunque, esporádicamente, alguno de ellos era descargado en
lonja como las de Gijón y Avilés. Aunque la aparición de esos
ejemplares tenía algún interés publicitario que era
aprovechado por alguna pescadería local que compraba el
ejemplar, éste terminaba posteriormente en la basura.
Descartados por carecer de valor económico, y,
desafortunadamente, desatendidos por los investigadores,
quienes no supieron ver su interés científico, no debe llamar
la atención que esos eventos fueran tan poco conocidos o,
tan siquiera registrados correctamente.
En varias ocasiones hemos escuchado con sorpresa
comentar que varios de los Peludines mantenían todavía
signos vitales al ser encontrados flotando o varados,
moviendo sus brazos con pequeñas contracciones o incluso
realizando expulsiones de agua a través del sifón. En ese
sentido, Laria recuerda perfectamente el relato de un turista
granadino que a las siete de la mañana recorría en solitario
un pedrero de la playa de la Griega en el municipio de
Colunga, precisamente en la zona donde ignitas y fósiles de
dinosaurios son la atención de múltiples visitantes. A esa
temprana hora, mientras saltaba de piedra en piedra,
escuchó un chapoteo entre unas grandes rocas a unos

Tabla 11. Registros de Architeuthis dux en las costas asturianas desde 1960
AÑO FECHA LUGAR OBSERVACIONES
1960 ¿Enero? Lonja de Aviles Ejemplar de 100 Kg. devuelto al mar. 1962 ---- Las Tiberas Dos tentáculos de 6 m. 1968 ---- Puerto de Vega Calamar de más de 7 m. Informa la embarcación Matías Bengoechea. Recogen el pico 1969 ---- Las Tiberas Tentáculo de más de 6m. Capturado por la embarcación Saturno Juan 1969 10-Julio Luanco (Gozón) Calamar gigante recogido por unos pescadores deportivos de Oviedo. Peso 250 Kg. Es el
más grande de cuantos se registraron en Asturias. 1969 ---- Puerto de Figueras Calamar de 7,90 m. y 120 Kg. recogido muerto por José Manuel Méndez, Bote Carducha 1971 Marzo Carrandi Un ejemplar de 80 Kg. recogido por el bardo Divino San Antonio. 1972 Febrero Puerto de El Musel Flotando y en descomposición (Martino Andres). Unos tres metros. 1980 ---- Vertical de Cudillero Se intenta meter a bordo un tentáculo de más de 100 Kg., pero se descarta al observar su
mal estado. Embarcación Oñaegea 1981 -------- Calamar de 10m. Se expone en una pescadería de Gijón y es vendido en la lonja de Avilés. 1981 Navidades Playa de Luarca Ejemplar varado en mal estado. Recogido por Secundino González y Manuel Pico. 1983 ---- Las Tiberas Calamar muerto de 150Kg. Recogido por “El Dioni” de Avilés 1984 ---- Cal. Sierra d Oviedo Tentáculo seccionado encontrado en el estomago de un tiburón (más de 10cm. de
perímetro) 1984 Abril 3 millas de Tazones Ejemplar capturado en arrastre por Antonio Coro (3-4 metros con brazos) 1992 ¿? Playa de Xagó Restos de manto y cabeza, enredados en un paño de volanta. Guardia Civil 1992 El Musel Calamar de 5 m. 1995 27- Sept. Playa de los Pozos Calamar de 160 Kg. Embarcaciones Nuevo Juanito y Hermanos de Avilés (pareja) con arte
de modalidad bou. 1996 Febrero Perlora Ejemplar seccionado por el primer tercio del manto y sin brazos ni tentáculos 1999 Noviemb. Carrandi Calamar muerto de 107 Kg. Recogido por el arrastrero Minchos VI 1999 Diciemb. Carrandi Calamar de 147 Kg. y unos 13 m. de largo capturado vivo por la pareja de arrastre Minchos
V y Minchos VI con base en Cillero (Lugo). Subastado en la lonja de Avilés y adquirido por Pescaderías Jiménez de Avilés. En la actualidad expuesto en el Aula del Mar (CEPESMA)
2000 2-Abril ------ Tentáculo de 2 m en el estomago de un marrajo. Embarcación Teresa José (Luarca)

Tabla 11. Continuación
AÑO FECHA LUGAR OBSERVACIONES 2001 12-Enero Agudo de Tierra Hembra de 9,67 m. de longitud y 92 Kg. 2001 Mayo Caladero Lastres Ejemplar 9 m. y más de 90 Kg. 2001 2-Julio Oeste de Carrandi Ejemplar de 60 Kg. y 7,10 m. Capturado por la pareja Elena María y Bautista Pino (Cillero) 2001 12-Sept. Playa de Rodiles (Villaviciosa) Ejemplar de 60 Kg. y longitud sin determinar debido a que le faltan los tentáculos. Apareció
varado 2001 20-Sept. Playa de Torimbia (Llanes) Ejemplar varado de 104 Kg. y 10 m. Expuesto en el Museo del Mar de Biarritz 2001 23-Sept. Caladero de El Corbiro Hembra inmadura de 73,5 Kg. capturada en arte de arrastre 2001 23-Oct. Playa de S. Cosme de
Barreiros (Lugo) Ejemplar de 90 Kg. y 9,5 m.
2002 11-Marzo Playa del Gayo (Luanco, Gozón)
Hembra inmadura de 70 Kg. que apareció varada
2002 13-Sept. Carrandi Ejemplar macho de 6 m. y 50 Kg. (Proyecto Kraken) 2002 10-Oct. El Agudo Ejemplar de 87 Kg. Embarcaciones Luscinda y Peña la Deva 2003 13-Sept. La Griega (Colunga) Ejemplar de 11 m. y 140 Kg. 2003 15-Sept. La Isla (Colunga) Ejemplar hembra de 10 m. y 80 Kg. 2003 16-Sept. Gozón Ejemplar agonizante (Hespérides) 2003 23-Sept. Bañugues (Gozón) Ejemplar macho en maduración de 60 Kg. Varado. 2003 10-Oct. Pozos- Carrandi Ejemplar de 10 m. y 67,50 Kg. Embarcaciones Travesía y Valdés Vega de Avilés 2003 18-Oct. La Griega (Colunga) Ejemplar hembra de 70 Kg. Tentáculos rotos AÑO FECHA LUGAR OBSERVACIONES
*Existe constancia de unos diez registros más en este periodo que están todavía por confirmar.

cuantos metros de distancia. Despacio se fue
acercando hacia la zona pensando que se trataba
de algún pez que habría quedado atrapado al
bajar la marea. Sin embargo, cuando se aproximó
lo suficiente el susto fue mayúsculo al observar
que un ser realmente desconocido para él se
movía, emitiendo lo que parecían respiraciones
forzadas. Sobresaltado por el acontecimiento se
alejo unos metros para, de nuevo y más sereno,
volver a observarlo, dándose cuenta entonces de
que aquello no era fruto de su imaginación, sino
que se trataba de algo real y que, a pesar de la
leyenda, cerca, muy cerca de él se encontraba un
calamar gigante. Corriendo, él dice que no sabe
como, por el abrupto pedrero llegó hasta el
camping donde sus compañeros de viaje
descansaban, a quienes explicó el sorprendente
encuentro.
En nuestro caso, sin mayores pruebas que
relatos exaltados, nos parecía poco probable que
los calamares gigantes pudiesen sobrevivir lo
suficiente como para ser observados moribundos.
Sin embargo, los hechos nos han probado lo
contrario gracias a uno de los ejemplares recogido
por Laria con la colaboración de la Guardia Civil
del Mar en las cercanías del Cabo de Peñas, el
cual nos demostró que, en muchos casos, las
personas que dicen haberlos observado todavía
con vida estaban en lo cierto y no era fruto de los
nervios que en ese momento podrían estar
sufriendo. Laria comenta que él también siempre
fue bastante escéptico hasta que pudo observar
directamente signos de vida en el ejemplar
recuperado por la Benemérita.
El peludin del Canea
Las siete menos cuarto de la mañana. Nubes
oscuras se ciernen en lontananza y la mar, como
a ellos les gusta llamarla, pica por el arrecio de la
brisa fresca del nordeste.
De pronto, tres relámpagos iluminan la
penumbra del amanecer seguidos de los truenos
que hacen retemblar el viejo cascaron de la
Siempre Carmela, aquella agotada lancha de popa
de gallo en la que tantos años el Canea veía
amanecer.
Mario, el Canea, estremecido por aquel
estruendo tasca los ojos dormitantes y alerta a los
demás. Todavía en camiseta de tirantes abre el
pestillo de la caseta intentando sujetar el
esquileiro y los rixones que, ante el zarandeo de
la marejada golpean los trastes de cubierta. De
repente, otro relámpago ilumina como nunca la
cubierta y la inquieta mar, dejando ver una
sombra en la superficie cercana a la borda de
estribor. Después, entre las luces del alba, ve

como eso se acerca más… cada vez más. El Canea
se frota la cara intentando quitarse el agua del
fuerte chubasco para cerciorarse de lo que estaba
presintiendo. No puede ser, no puede ser, piensa,
pero casi simultáneamente grita con fuerza:
- ¡Mon, Mon, coged los ganchos, coged los
ganchos!
A tumbos y soñoliento, Mon sale del traste
y, mientras avanza por la pequeña cubierta hasta
el pincho de proa para coger un gancho, pregunta
a voces:
- ¿Que pasa Canea?
- ¡Ven con el gancho, rápido!, contesta
Canea.
Que duda cabe que aquello extraño, aquello
inusual que flota en la penumbra, nunca nos
debería pasar desapercibido, más si no tiene, o no
se le otorga valor, puede caer en la más sombría
inexistencia. De alguna manera, este ser que ha
nacido leyenda, recorrió la historia
contemporánea sin apenas mérito, interesando
poco a esta sociedad ávida de monstruos a los
que aplacar o ensalzar míticamente para
desahogar nuestros temores y sueños. Cuantas
veces los marinos han contado historias a la luz
del candil después de la caída de la tarde en
aquellas largas noches de invierno con el lar
chispeando y, colgada sobre él, una sopa de ajo
caldeando. Un cálido ágape servido con esmero a
los vecinos que, después de escudriñar el
horizonte, auspiciaban el descanso forzado por la
rugidora bajamar. Mario el Canea sabía como
poner los ojos como platos, no solo a sus
pequeños, sino también a los no tanto. Los unos y
los otros pagarían después consecuencias del
insomnio que años, bastantes años después,
siguen recordando.
- Yo no le doy gancho, está vivo- decía Mon
a voces mientras se separaba como un resorte del
carel de la lancha.
Efectivamente, ante ellos y a escasos dos metros
en la superficie del agua, un enorme peludín
agitaba sus gruesos brazos, algunos de los cuales
se aferraban fuertemente a las maderas del
costado, a la vez que emitía resoplidos roncos que
salpicaban con agua la propia cubierta.
Los doce metros de eslora de la Siempre
Carmela se quedaban ridículos ante tan enorme
calamar. Ninguno de sus tripulantes habría jamás
imaginado tal tamaño aunque conocían bien esos
animales, ya que era fácil que en los meses de
primavera y otoño, en las playas de Otur, La
Segunda o la Tercera de Luarca, vieran
descomponerse alguno de ellos pasto de la pulga
y con un característico olor a amoniaco.
El Canea, por el simple motivo de que no le
tomaran por loco al llegar a tierra cuando contase

lo que habían visto, ávido y de forma certera,
logró hincar el gancho en el manto de tan
descomunal animal, que, al sentirse clavado, con
solo dos quites arrebató de aquellas férreas
manos el utensilio, como si de un mondadientes
se tratase alejándose luego del barco.
El Canea y Mon, mirándose mutuamente con
asombro, no daban crédito a lo sucedido y
acordaron no contarlo hasta bastante tiempo
después para evitar las seguras guasas en los
corrillos del puerto.
Y el mudo habló
En el mes de septiembre de 1969, desde el
pequeño puerto de Figueras en la orilla Asturiana
de la ría del Eo, José Manuel Méndez, en su
pequeño bote La Carducha, de apenas cinco
metros, salió como siempre bien entrada la tarde
en busca de calamares.
Lo que nunca podría haber imaginado el
bueno de Pepe el mudo es que aquella tarde no
regresaría como de costumbre con alguno de los
preciados calamares comunes, sino con uno muy
especial de nada menos que 120 kilos.
En la desembocadura de la propia ría
encontró flotando un peludín y, a toda prisa y
después de lograr atarlo para remolcarlo, lo
acercó al puerto. Dicen las muchas personas que
recuerdan verlo llegar lleno de júbilo y
extremadamente nervioso que aquel día “el mudo
habló”. Pensándolo bien, no seria para menos ya
que parece ser que el gran calamar todavía se
encontraba vivo en el momento que logro pasarle
una soga por el cuerpo.
Los peludines y los niños asturianos
CEPESMA se creó en 1996 y en 1998 abrió
en Luarca el “Aula del Mar”, museo que cuenta
con una amplia exposición de animales marinos
con sus correspondientes paneles explicativos:
más de 400 ejemplares de especies distintas,
entre los que se puede observar ocho de los
calamares gigantes más grandes y mejor
conservados del mundo. A través de esta
exposición, así como de documentales y
conferencias, se pretende dar a conocer el
ecosistema marino y sensibilizar a la población de
la necesidad de conservarlo, mostrando las
agresiones más habituales que conducen a su
paulatino deterioro. Uno de los objetivos
fundamentales de CEPESMA es concienciar a la
población, especialmente a los niños, de la gran
importancia que tiene el preservar los
ecosistemas marinos. Durante los pasados años,

más de 10.000 niños y adolescentes participaron
en las actividades organizadas en el Aula.
Entre las numerosas actividades formativas
permanentes que se organizan en el Aula del Mar,
está la convocatoria anual de un concurso de
redacción entre los escolares de Luarca. Es
increíble el derroche de entusiasmo y de
imaginación que se detecta a partir de los
trabajos de los niños. Sería muy largo, aunque
muy instructivo recoger aquí los escritos
archivados en los ficheros de esta institución. No
obstante, como muestra se expone íntegramente
a continuación la redacción que consiguió el
primer premio en el certamen celebrado en
Noviembre de 2002 bajo el título “Calamares
Gigantes en Asturias”. Su autora es la niña de
nueve años Merce López Martínez del Colegio
Público Padre Galo de Luarca.
“El calamar que nun se queria ir del colexu”
Soi una neña de 9 años qu’e estudia 5º de
Primaria, nun colexu que se llama Padre Galo, al
que voi dende que yera pequeña y al que tengo
muchísimo cariño.
El colexu tien dos plantes, na d’abaxo tan
los neños de preescolar hasta 2º; dende 3º hasta
6º tamos nel piso d’arriba, neste piso del colexu
tá l’Aula del Mar. Polos pasillos del colexu hái
vitrinas de cristal que tienen muchas especies
marinas y tamién munchos utensilios del mar,
colos que aprendemos muchísimas cousas y en
una parte di esi pasillu, tá metiu nuna vitrina, un
calamar xigante del que voi a cuntar la hestoria
d’esti cuento...
Tolos dies, cuando voi al colexu y subo al
pisu d’arriba quédome mirando il calamar que tá
nel pasillu, al lláu delau mio clas. Siempre me
prestó muncho mirarlo ¡veslu tan quietu! como si
nun sénterase de nada y ensin embargo velo
too...
Estos dies tan recoyendo munchas de las
cousas que tán polos pasillos porque l’Aula vase
pá otru llugar y a mi dame muncha pena, porque
l’colexu quedase como un poco triste..., tamién
sin toas esas cousas que nos rodeaban y noté que
il calamar tamien taba mui triste...
Taba mirando fixamente cuando escuché
que me decian: Y nel cole qué tal ¿bien? Miré a un
lláu y otru, pero nun había nadie, nadie, ... ensin
embargo alguien falaba...
Di un respigo...! nun pue ser, pensé, pero el
calamar repitióme: neña ¿pasate dalguna cousa?
Taba asustada, nun sabia que decir, las
palabras nun me salian... y era como si me
quedase muda...!
Tovia asustada, contesté: ¡taba mirando como
llevan algunas cousas del colexu!

Ay neniña, si supieres que triste toi
pensando si me llevan a mi tamen desti colexu...
Tolos dias pola mañana escuchó la sirena
sonar y veo como subis alborotaos y felices
pa’clas, veu pasar los mayestros ¡fixate si los
conozco a toos!
Pa sa ‘l d’ inglés y digo: ¡Hellow! Dicese
asina ¿verdá?
Faigo una flexión colos tentáculos cuando
pasa il mayestro de gimnasia y quédome quietu
cuando pasa a “direutora” del colexu ¡por si
acasu...!
Tovía tengo deprender munchísimas cousas
más: tocar a flauta, dibuxar, y sobre tolo ir a clas
de matemáticas, conocimiento, llengua...
¡Tovia toi mui floxo nestas asignaturas! Por
eso prestariame quedarme nel colexu.
Mais tarde, baxando les escaleres, de cuatro
en cuatro, con mucha por baxar al patiu a xugar
¡como me presta veros nel recréu! xugando tan
contentos: las neñas a la comba, los neños
cambiando cromos...
¡Como me prestaría tar xugando con
vosotros ...!
Y hasta los pequeños, que veo poco, porque
tan nel pisu d’abaxo, como me presta velos subir
cuando van al video o a música con esos
mandiloninos de cuadrinos marielos, azules y
hasta esti añu son de colorines, como me presta
oílos cantar cuando entran del recréu:
Yo conozco un tren
que se llama Cilipin
y va por la ría
haciendo pi, pi ...
¡Cuánto vos voi a echar de menos ...!
Emocionada y con ganas de llorar, polo que
me cuntara del colexu dixe-y que nun se
preocupara, que la so nueva casa yera mos
guapa, toa de madera y nosotros siempre vamos
a querete aunque te vayas del colexu.
Además ¡tengo una idea! pá que t’allegres
un poco voi ponete un nome, porque los
calamares teneis nomes moi raros, a ver ...
Deixame pensar ... ya tá: ¡el calamar
Graduado! porque esti colexu cuando mio pa’ yera
pequeñu, llamaban “las Escuelas Graduadas”,
pero ¡meyor tovia! de Graduado ponemus “Gradu”
y de Padre Galo – “Galo” ¡ya tá!
¡El calamar GraduGalo! - ¡Que te paez?
Suena mui bien, préstame il nome, neniña.
¡Ya toi más contento! Yeres una neña muy
simpática y amable. ¡Gracias neniña por
escuitame!
Queriu calamar: quiero que sepas los neños
del colexu Padre Galo, siempre te querremus,
porque como diz mio ma’ ¡lo importante ye querer
a la xente! O nesti casu a los animales.

¡Si te vas diremos a verte de vez en cuando
a la to nova casa prometido!
Termino diciendote:
¡Adiós calamar queriu, nunca te
olvidaremos, onque te hayas ido!
Merce
Es una experiencia pedagógica contrastada
que los niños desarrollan más fácilmente su
sensibilidad con las clases y cursos de pintura y,
por lo tanto, tienen una mayor capacidad para
poder expresar y recrear sus vivencias, sus
sentimientos y sus anhelos. Desde este punto de
vista, el dibujo es un material muy considerado
por los pedagogos ya que permite conocer mejor
al niño y lo que piensa. La incorporación del gusto
por la estética en los niños debe hacerse como
toda iniciación, a edad temprana, ya que así les
resulta más fácil asimilar los conocimientos
iniciales que constituirán la base cultural que
podrán aumentar y consolidar a lo largo de su
vida.
Son numerosas las ciudades que tienen
muestras artísticas como un refuerzo visual capaz
de generar un sentimiento de aprecio a la belleza
por parte de la infancia y que anima a los niños a
la creatividad. También hay en las ciudades y los
pueblos, por pequeños que sean, edificios
susceptibles de ser contemplados bajo un prisma
artístico, y su contemplación y análisis pueden ser
un punto de partida magnífico para la información
y la formación artística desde muy tempranas
edades. En Luarca, además de nobles y bellos
edificios, hay un museo con numerosos objetos y
seres relacionados con el mar, que también
pueden ser útiles instrumentos para la iniciación
artística de los niños. Así se entendió desde un
principio y son ya numerosos los certámenes de
dibujo basados en sus colecciones, donde uno de
los elementos más llamativos y sugerentes del
museo para la imaginación infantil son los
peludines.
Figura 87. Cuatro dibujos del calamar gigante. Niños
de tres años del colegio Padre Galo.

La ingenuidad creativa que destila la
redacción reproducida unas líneas más arriba sólo
es comparable a la de la colección de dibujos
sobre los peludines realizados por los niños de
Luarca, de la que se expone una muestra
representativa de los niños de tres años.

Capítulo 8
ASPIRACIONES Y PERSPECTIVAS DE
FUTURO
Aumentar el conocimiento sobre las
profundidades submarinas
Todo lo que se ha dicho sobre los calamares
gigantes hasta ahora nos conduce a la conclusión
de que se conoce muy poco sobre su historia
natural. Pero la cuestión es todavía más honda:
apenas conocemos las profundidades de los
mares y océanos que nos circundan, ello en un
planeta que debería haberse llamado “AGUA” en
vez de “TIERRA”, porque más de sus dos terceras
partes están cubiertas por el líquido elemento.
En su conjunto, los dominios bati y
abisopelágico de los océanos representan más del
90% de la biosfera localizada sobre la corteza
terrestre. Por otra parte, los fondos marinos de
las zonas batial, abisal y hadal constituyen un
dominio profundo que se extiende desde la base
del talud continental hasta las mayores
profundidades abarcando cerca de 307 millones
de kilómetros cuadrados, lo que representa el
80% de la superficie de los fondos marinos y
cerca de 65% de la superficie de la Tierra.
Tanto los dominios pelágicos como
bentónicos profundos constituyen el ambiente
menos favorable para el desarrollo de la vida,
debido a las extremas condiciones que lo
caracterizan: elevadas presiones hidrostáticas;
temperatura constante e inferior a 4ºC; oscuridad
casi total o total, impidiendo todo tipo de
producción fotosintética; y aporte limitado de
nutrientes. Tales condiciones deberían limitar, a
priori, considerablemente el desarrollo de los
seres vivos, sin embargo, las evaluaciones más
recientes han mostrado que la biodiversidad en
los sedimentos oceánicos profundos es una de las
más elevadas del planeta. Por otra parte, el
paradigma de “la uniformidad en las condiciones
de la vida en la Tierra” ha variado
considerablemente después del descubrimiento de
nuevos ecosistemas basados en la quimiosíntesis,
asociados a las surgencias hidrotermales ubicadas
en los ejes de las grandes dorsales oceánicas, y a
las fuentes de fluidos fríos - ricos en metano - en
las zonas marginales activas y pasivas de las
placas continentales.
Entre los seres vivos que moran a grandes
profundidades, los cefalópodos son uno de los

grupos que están proporcionando mayores
sorpresas y novedades. Un ejemplo de ello lo
proporciona el reciente hallazgo de un cefalópodo
abisal cosmopolita aún no identificado, que fue
publicado en 2001 en la prestigiosa revista
Science.
En septiembre de 1988, el submarino
tripulado francés Nautile filmó por primera vez un
extraño calamar en aguas del Atlántico occidental,
frente a las costas de Brasil y a una profundidad
de 4.735 m. En julio de 1992 un nuevo ejemplar
fue filmado desde ese mismo submarino en la
región oriental del Atlántico, cerca de las costas
de África, entre 2.950 y 3.010 m. En noviembre
de 1998, el submarino japonés Shinkai 6500
encontró otro en el Índico a 2.340 m. En enero de
2000 fue filmado nuevamente por un ROV
comercial a 2.195 m en el golfo de México,
estimándose su longitud total en 1,5 m. En mayo
de 2000, otra vez el Nautile lo filmó en el océano
Índico a 2.576 m de profundidad. En octubre de
2000 lo hizo el submarino tripulado
norteamericano Alvin, a 1.940 m, en el golfo de
México. Por último, en mayo de 2001 lo filmó el
submarino Tiburón, a 3.380 m, en el Pacífico
central, estimándose su longitud total entre 4 y 5
metros.
En la mayoría de los casos este cefalópodo,
que se identificó como un tipo de calamar
(Teuthida), fue encontrado cerca del fondo, con
sus enormes aletas terminales ondulando
lentamente y con los diez apéndices alrededor de
la boca- sin diferencia entre brazos y tentáculos-
extendidos a lo largo del eje corporal y doblados
abruptamente en su parte anterior, formando un
codo del que partían filamentos muy largos. Estos
larguísimos filamentos (hasta 20 veces la longitud
del manto del animal) parecen estar desprovistos
de ventosas, son contráctiles y son los más largos
conocidos hasta ahora en cualquier otro
cefalópodo.

Fig. 88. Foto de un cefalópodo abisal desconocido
que ha sido fotografiado en varios océanos del
mundo.
La morfología de estos ejemplares es muy
diferente a la de cualquier otra familia conocida.
No obstante, podría tratarse de adultos de la
familia Magnapinnidae, descrita en 1998 por
Vecchione y Young a partir de dos juveniles y una
paralarva, capturados en aguas epipelágicas del
Pacífico central y oriental.
Probablemente, la inferencia más
importante que se pueda extraer a partir de estas
ocho observaciones sobre una especie
desconocida de cefalópodo; todas realizadas con
pocos años de diferencia y a profundidades
similares, que al mismo tiempo fueron realizadas
en sitios tan lejanos como el golfo de México, y
los océanos Atlántico, Índico y Pacífico, es que se
trata de un animal batipelágico relativamente
frecuente y de distribución cosmopolita. Gracias a
estas observaciones se puede decir que esta
especie es un cefalópodo de tamaño conspicuo
que puede considerarse común en el mayor
ecosistema existente en el mundo, el océano
abisal, pero que todavía no ha sido capturado, y
que nunca antes había sido observado. Esto pone
en evidencia lo poco que conocemos aún del
ecosistema más vasto de la tierra y de la vida en
las grandes profundidades, y ¿por qué
conformarnos con conocer mejor Marte que las
profundidades submarinas?
Equipar a España con instrumentos
modernos de exploración submarina
A partir del final de la Segunda Guerra
Mundial se ha producido un creciente interés por
el estudio de la biodiversidad y el funcionamiento
de los ecosistemas profundos de los océanos. Este
estudio es necesario actualmente, entre otras
razones, porque muchos de esos ecosistemas son
lugares de actividad industrial, donde se produce
un impacto antropogénico; también porque en
ellos la actividad biológica es responsable de la
transformación y de los intercambios
biogeoquímicos en la interfase agua-sedimento,
que juegan un importante papel en el equilibrio
químico del océano. Este intercambio no puede
ser estimado de forma global sin considerar la
variación espacial y temporal de las comunidades
bentónicas y de los diferentes procesos biológicos

que reinan en ellas. Por último, pero no menos
importante, en muchos de esos ecosistemas se
han producido adaptaciones muy peculiares en los
organismos, que nos pueden ayudar a
comprender los procesos evolutivos. En este
sentido, algunos ecosistemas profundos, sobre
todo los de las fuentes hidrotermales, son
verdaderos laboratorios naturales protegidos de la
agresiva radiación solar ultravioleta, donde poder
estudiar la vida en las condiciones de un océano
primitivo.
Si estos ambientes contienen los
ecosistemas más vastos todavía inexplorados de
nuestro planeta su estudio debiera ser prioritario.
Sin embargo esto no es así, ¿por qué existe tanto
retraso en su estudio? El principal motivo de esta
demora reside en el lento desarrollo que han
tenido los equipos técnicos indispensables para
trabajar en las extremas condiciones reinantes en
dichos lugares. Un aspecto trascendental que
explica esta lentitud es, a diferencia de lo ocurrido
en la investigación espacial o militar, por ejemplo,
la escasez de presupuestos destinados al
desarrollo de nuevas tecnologías para el estudio
del ecosistema marino. En la actualidad, los
científicos disponen de medios para realizar
muestreos bastante precisos, como los corer de
gravedad con testificador de caja de gran
superficie, o los corer multitubulares para
muestreo del sedimento superficial con un mínimo
de perturbación. Pero, sólo recientemente se ha
podido abordar el estudio directo de los
organismos marinos en su medio gracias al
desarrollo y puesta en funcionamiento de
vehículos manejados por control remoto (ROVs) y
de submarinos tripulados. Ambos tipos de
vehículos submarinos están provistos de sensores
para medir diferentes parámetros ambientales,
analizadores químicos, cámaras de
experimentación, cámaras de fotos y videos, así
como pinzas y trampas para la obtención de
muestras. De esta forma, y por primera vez, los
científicos han poseído herramientas eficaces para
abordar estudios de biodiversidad,
comportamiento y funcionamiento in situ de
importantes procesos biológicos.
Desgraciadamente, ese tipo de investigación es
tan cara que únicamente se la pueden permitir de
forma rutinaria los Estado Unidos de América y
Francia, y, en menor medida, Japón, Canadá y
Reino Unido. La antigua Unión Soviética realizaba
importantes estudios con estos vehículos pero
ahora, ya como Rusia, y debido a la falta de
medios, ha dejado de contar en el concierto
mundial. En cuanto a España este tipo de
investigación es prácticamente inexistente,
careciéndose en la práctica de los medios propios

necesarios para siquiera intentar realizar este tipo
de estudios.
Un buen ejemplo de un planteamiento
eficiente para la investigación de los abismos
submarinos puede ser el del Acuario de
Monterrey. El cañón submarino de Monterrey, en
California, es el más profundo que hay en la costa
del Pacífico de los Estados Unidos de América del
Norte. Su profundidad sobrepasa los 4.000
metros, pero además está muy cerca de la costa;
de tal manera, que los investigadores del Acuario
de Monterrey pueden salir del puerto a las siete
de la mañana y estar sobre 2.000 metros de
profundidad una hora y media más tarde, con un
barco que navegue a diez u once nudos. Ese
cañón representa un auténtico laboratorio
submarino donde estudiar los ecosistemas
profundos. El Acuario, cuyo objetivo es inspirar e
impulsar la conservación de los océanos, cuenta
con un vehículo submarino operado por control
remoto (ROV) capaz de descender hasta 4.000
metros de profundidad. Su barco nodriza es el
Western Flyers. Este instrumento de
investigación, exploración y enseñanza submarina
ha dado ya magníficos resultados en esos tres
campos. El Tiburón, como se llama el ROV, es un
vehículo no tripulado que manejan operarios
desde la cubierta del barco, cuya operatividad es
de seis días. Se trata de un vehículo submarino
no tripulado, de estructura abierta con sistema de
flotabilidad y estabilización vertical automática,
muy maniobrable y versátil, que recibe las
órdenes y ofrece la información desde y a una
cabina de control, recepción y almacenamiento de
datos.
El Tiburón está conectado al barco nodriza
por un cable con doble armadura exterior de
acero inoxidable, dentro de la cual discurren los
cables eléctricos y de fibra óptica. Su propulsión
es eléctrica y lleva un navegador acústico. Este
ROV lleva también un brazo pinza manipulador de
4 ejes y está provisto de una cámara de fotos
digital con objetivo gran angular y posibilidad de
macro, así como de una cámara de video, y
sistemas de iluminación. Diseñado para que se
puedan instalar en él sistemas modulares
fácilmente acoplables con diferentes tipos de
sensores, carruseles para recogida de muestras
biológicas, sedimentos, agua, sonares de distinto
tipo, hidrófonos, etc., este vehículo submarino
permite, además de la observación y la obtención
de muestras de muy diferente naturaleza, la
realización de trabajos de investigación en muy
diferentes campos científicos como la
oceanografía, la ecología y la geología.
Disponer de instrumentos como el descrito
en varios puntos de la geografía española, donde

Asturias sería un candidato ideal por tener cerca
cañones submarinos parecidos a los de
Monterrey, no sólo permitiría elevar las
posibilidades para observar y filmar a los
calamares gigantes en su medio ambiente natural
sino que, además, ampliaría la precisión de toda
la gama de sensores que se utilizan
habitualmente en oceanografía, biogeoquímica,
biología marina, contaminación, impacto
medioambiental, etc. Gracias a estos equipos
submarinos España podría aumentar su capacidad
para estudiar los ambientes abisales, así como se
posibilitaría la fotografía y filmación de fenómenos
indetectables con los equipos habituales. Además,
este tipo de tecnología se podría utilizar para el
posicionamiento y recuperación de equipos fijos,
así como para realizar reparaciones submarinas,
tal y como se hace ya en otros países.
Recientemente, aquí en España y con
ocasión del desastre del petrolero Prestige, hemos
tenido la posibilidad de comprobar lo útiles que
son los submarinos tripulados tipo Nautile, y los
ROVs. Quizá este desgraciado acontecimiento
haga que en España se abra una puerta a la
investigación en aguas profundas. Es de esperar
que todos tengamos ahora más claro que
nuestras conexiones con las profundidades
marinas son muy estrechas, puesto que
dependemos de los fondos como fuentes de
recursos de muy diferente tipo, incluso
alimenticios. Por otra parte, lo que ocurre en las
aguas profundas influye en aguas más someras
donde los intereses humanos son, por ahora,
mayores. Además, los científicos están usando
animales de aguas profundas como fuente de
nuevas medicinas, y quedan todavía numerosas
especies por descubrir. Teniendo en cuenta los
posibles beneficios, tanto materiales como
intelectuales, que podamos conseguir en el
futuro, parece evidente hoy en día que
necesitamos conocer mejor, proteger y conservar
esas aguas profundas. Y para ello necesitamos los
instrumentos científicos adecuados.
Los calamares gigantes: organismos
emblemáticos para inspirar la conservación
de los océanos
Resulta tópico afirmar que “una imagen vale
más que mil palabras”, pero no por ello deja de
ser bastante cierto. En cuantas reuniones
internacionales, publicaciones, conferencias y
otros foros se ha hablado ya, bien y
convincentemente, sobre la necesidad de cuidar el
ambiente, y que esa protección y conservación
debe extenderse a los océanos.
Desafortunadamente, la realidad muestra que esa

idea no ha calado lo suficiente en la sociedad y
todavía se maltrata de muy diferentes maneras
nuestra hidrosfera. Quizá ello se deba a que gran
parte de los océanos son propiedad de nadie, o
también a que los efectos nocivos de la
contaminación se perciben menos en el mar que
en la tierra. Sin embargo, un delfín muerto al ser
atrapado por una red, o un solitario alejado de su
manada que se acerca a nuestras costas,
despierta y seguirá despertando, sentimientos
misericordes, a través de los cuales se hace más
cercana, plástica y comprensible la necesidad de
conservar los océanos. De esta manera, se puede
considerar a los delfines como animales
emblemáticos para inspirar la conservación de los
océanos.
Salvando las distancias, ya que no en vano
aquellos son mamíferos y éstos son fríos
invertebrados, los calamares gigantes han
demostrado tener una notable capacidad de
atracción para el gran público. A través de
aquellos ejemplares que vararon hace un par de
años en Asturias coincidiendo con exploraciones
acústicas realizadas durante prospecciones
geofísicas en el golfo de Vizcaya se ha levantado
una marea popular de protesta. Desde numerosas
instancias se pide más información sobre las
posibles repercusiones de ese tipo de
exploraciones en los organismos y ecosistemas
profundos marinos. Los calamares gigantes, se
dijo entonces, pueden ser la punta de un iceberg
que oculta graves lesiones a los ecosistemas y
animales marinos, cuyas repercusiones
ambientales, económicas y culturales son
impredecibles. Si esto es así, hemos hallado un
nuevo animal emblemático a través del cual,
Asturias, o cualquier otro lugar, puede inspirar la
conservación de los océanos de la que tan
necesitados estamos.
Bibliografía
Akimushkin, I.I. 1963. Cephalopods of the Seas of the
U.S.S.R. Academy of Sciences of the U.S.S.R.,
Institute of Oceanology, Moscow, 235 pages, 60
figures (Translation, Israel Program for Scientific
Translations, Jerusalem, 223 pages)
Aldrich, F.A. 1968. The distribution of giant squids
(Cephalopoda, Architeuthidae) in the north
Atlantic and particularly about the shores of
Newfoundland. Sarsia, 34: 393-398.
Aldrich, F.A. 1991. Some aspects of the systematic and
biology of squid of the genus Architeuthis based
on a study of specimens from Newfoundland

waters. Bulletin of Marine Science, 49(1-2): 457-
481.
Aldrich, F.A. y M.M. Aldrich. 1968. On regeneration of
the tentacular arm of the giant squid Architeuthis
dux Steenstrup (Decapoda, Architeuthidae).
Canadian Journal of Zoology, 46(5): 845-847.
Aldrich, F.A. y E.L. Brown. 1967. The giant squid in
Newfoundland. Newfoundland Quarterly, 65(3): 4-
8.
Alexeyev, D.O. 1994. New data on the distribution and
biology of squids from the southern Pacific.
Ruthenica, 4(2):151-166.
Allan, J.K.1948. A rare giant squid. Australian Museum
Magazine, 9(9): 306-308.
Anonymous. 1978. Big squid is largest on record. Curry
Coastal Pilot, July 6, 1978: 11.
Anonymous. 1996. Out of its depth. Age, Melbourne
Australia, 12 June 1996.
Anonymous. 1999. Giant squid found dead in Bahamian
waters. Soundings, University of Miami Rosenstiel
School of Marine & Atmospheric Science, May 15,
1999.
Antonelis, G.A., M.S. Lowry, C.H. Fiscus, B.S. Stewart y
R.L. DeLong. 1994. Diet of the northern elephant
seal. En: B.J. LeBoeuf and R.M. Laws (Eds.)
Elephant Seals: Population Ecology, Behavior, and
Physiology: 211-223, University of California
Press.
Arfelli, C.A., A.F. de Amorim y A.R.G. Tomas. 1991.
First record of a giant squid Architeuthis sp.
Steenstrup, 1857 (Cephalopoda, Architeuthidae)
in Brazilian waters. Boletín do Instituto da Pesca,
18: 83-88.
Aronson, R.B. 1991. Ecology, paleobiology and
evolutionary constraint in the Octopus. Bulletin of
Marine Science, 49(1-2): 245-255.
Atkinson D. y R.M. Sibly. 1997. Why are organisms
usually bigger in colder environments? Making
sense of a life history puzzle. Trends Ecology and
Evolution, 12: 235-239.
Bandel, K. y S.v. Boletzky. 1988. Features of
development and functional morphology required
in the reconstructuion of early coleoid
cephalopods. En: Wiedmann, J. and J. Kullmann
(Eds.) Cephalopod –Present ad past: 229-246.
Schweizerbart’sche Werlagsbuchhandlung.
Stuttgart.
Bardarson, G. 1920. Om den marine molluskfauna ved
vestkysten af Island. Kongelige Danske
Videnskabernes Selskab Biologiske Meddelelser,
2(3): 1-139.
Belloc, G. 1950. Catalogue des types de Céphalopodes
du Musee Océanographique de Mónaco. Bulletin
de l'Institut Océanographique, 970: 1-10.
Belyayev, G.M. 1964. Rostra of cephalopods in oceanic
bottom sediments. Deep-Sea Research, 11(1):
113-126.

Berry, S.S. 1912. Note on the occurrence of a giant
squid off the California coast. Nautilus, 25(10):
117-118.
Berry, S.S. 1914. Another giant squid in Monterey Bay.
Nautilus, 28(2): 22-23.
Berzin, A.A. 1971. The sperm whale. Pacific Scientific
Research Institute of Fisheries and Oceanography,
Moscow, 1971. [English translation from Russian
by E. Hoz and Z. Blake, Israel Program for
Scientific Translations, 1972, 393 pp.]
Bettencourt, V. y A. Guerra. 2000. Growth increments
and biomineralization process in cephalopod
statoliths. Journal of Experimental Marine Biology
and Ecology, 248: 191-205.
Bizikov, V.A. 1991. A new method of squid age
determination using the gladius. N.T.R. - I.T.P.P.
Special Publication, 1: 39-51.
Blake, J.H. 1909. A giant squid. Nautilus, 23(3): 43-44.
Boletzky, S.v. 1989. Recent studies on spawning,
embryonic development, and hatching in the
Cephalopoda. Advances in Marine Biology, 25: 85-
115.
Bolstad, K.S. y S. O’Shea. 2004. Gut contents of the
giant squid Architeuthis dux (Cephalopoda:
Oegopsida) from New Zealand waters. New
Zealand Journal of Zoology, 31:15-21.
Botas, J.A., A. Bode, E. Fernández, y R. Anadón. 1988.
Descripción de una intrusión de agua de elevada
salinidad en el Cantábrico central: distribución de
los nutrientes inorgánicos y su relación con el
fitoplancton. Investigación Pesquera, 52: 561-
574.
Bower, J.R. y Y. Sakurai. 1996. Laboratory observations
on Todarodes pacificus (Cephalopoda:
Ommastrephidae) egg masses. American
Malacological Bulletin, 13(1/2): 65-71.
Boyle, P.R. (Ed.). 1983. Cephalopod Life Cycles. Vol.1.
Species Account. Academic Press. London, 475
pp.
Boyle, P.R. 1984. Giant squid stranded at Aberdeen.
Porcupine Newsletter, 3(1): 12.
Boyle, P.R. 1986. Report on a specimen of Architeuthis
stranded near Aberdeen, Scotland. Journal of
Molluscan Studies, 52(1): 81-82.
Boyle, P.R. (Ed.). 1987. Cephalopod Life Cycles. vol. 2.
Comparative Reviews. Academic Press. London,
441 pp.
Brinkmann, A. 1916. Kjaempeblaekspruten
(Architeuthis dux Stp.) i Bergens Museum.
Naturen. Illustrert Maanedsskrift for Populaer
Naturvidenskap, Bergen, 40(6): 175-182.
Brix, O. 1983. Giant squids may die when exposed to
warm water currents. Nature, 303(5916): 422-
423.
Brix, O., A. Bardgard, A. Cau, A. Colosimo, S. G. Condo
y B. Giardina. 1989. Oxygen-binding properties of
cephalopod blood with special reference to

environmental temperatures and ecological
distribution. Journal of Experimental Zoology,
252(1): 34-42.
Brunetti, N.E., B. Elena, G.R. Rossi, M. Sakai, S.E.
Pineda y M.L. Ivanovic, 1998. Description of an
Architeuthis from Argentine waters. South African
Journal of Marine Science, 20: 355-361.
Brunetti, N.E., M.L. Ivanovic, M. Sakai y L.N. Pascual.
2005. Two new records of giant squid
(Architeuthis sp) from Patagonian region. Revista
de Investigación y Desarrollo Pesquero (en
prensa).
Budelman, B.U., R. Shipp y S.v. Boletzky. 1997.
Cephalopoda. En: Microscopic Anatomy of
Invertebrates. Vol. 6A, Mollusca II (Harrison, F.W.
and A.J. Kohn, Eds.): 119-414. Wiley-Liss, New
York.
Bustamante, P., K. Das, A. Guerra, V. Lahaye, F.
Rocha, A.F. González and L. Laria. 2006. The
giant squid (Architeuthis): a very high level
predator? Cephalopod Life Cycles, Cephalopod
International Advisory Council Symposium,
Abstract book: 72. Hobart, Tasmania.
Cadenat, J. 1935. Note sur la première capture dans le
Golfe de Gascogne du céphalopode géant,
Architeuthis nawaji. Compte-rendu de
l'Association Française pour l'Avancement des
Sciences, 59: 513.
Cadenat, J. 1936. Note sur une céphalopode géant
(Architeuthis harveyi Verrill) capturé dans le golfe
de Gascogne. Bulletin de Muséum d'Histoire
Naturelle, Paris, (2)8(3): 277-285.
Calow, P. 1987. Fact and theory -an overview. En:
Boyle, P.R. (Ed.). Cephalopod Life Cycles. vol. 2,
Comparative Reviews: 351-365. Academic Press.
London.
Cherel, Y. 2003. New records of the giant squid
Architeuthis dux in the Southern Indian ocean.
Journal of the Marine Biological Association of the
United Kingdom, 83: 1295-1296.
Cherel, Y and Hobson, K. A. 2005. Stable isotopes,
beaks and predators: a new tool to study the
trophic ecology of cephalopods, including giant
and colossal squids. Proceedings of the Royal
Society B, 272: 1601-1607.
Clarke, M.R. 1962. Stomach contents of a sperm whale
caught off Madeira in 1959. Norsk Hvalfangsttid,
51(5): 173-191.
Clarke, M.R. 1966. A review of the systematics and
ecology of oceanic squids. Advances in Marine
Biology, 4: 91-300.
Clarke, M.R. 1967. A deep-sea squid, Taningia danae
Joubin, 1931. Symposium Zoological Society,
London 19: 127-143.
Clarke, M.R. 1980. Cephalopoda in the diet of sperm
whales of the southern hemisphere and their

bearing on sperm whale biology. Discovery
Report, 37: 1-324.
Clarke, M.R. 1986. A Handbook for the Identification of
Cephalopod Beaks. Clarendon Press, Oxford, 273
pp.
Clarke, M.R. 1988. Evolution of buoyancy and
locomotion in recent cephalopods. En: Wilbur,
K.M. (Ed.) The Mollusca, vol. 12. Clarke, M.R. y
E.R. Trueman (Eds.) Paleontology and Neontology
of Cephalopods: 203-213. Academic Press Inc.
Clarke, M.R. (Ed.). 1996. The role of cephalopods on
the world’s oceans. Philosophical Transactions of
the Royal Society of London, Series B, 351
(1343): 977-1112.
Clarke, M.R. 1998. The value of statolith shape for
systematics, taxonomy, and identification. En:
Voss, N.A., M. Vecchione, R.B. Toll & M.J.
Sweeney (Eds.) Systematics and Biogeography of
Cephalopods, Vol. I. Smithsonian Contributions to
Zoology, 586: 69-76.
Clarke, M.R. y N. MacLeod 1974. Cephalopod remains
from a sperm whale caught off Vigo, Spain.
Journal of the Marine Biological Association of the
United Kingdom, 54: 959-968.
Clarke, M.R. y N. MacLeod 1976. Cephalopod remains
from sperm whales caught off Iceland. Journal of
the Marine Biological Association of the United
Kingdom, 56: 733-749.
Clarke, M.R. y N. MacLeod 1982. Cephalopod remains
from the stomachs of sperm whales caught in the
Tasman Sea. Memoirs of the National Museum
Victoria, 43: 25-42.
Clarke, M.R., H.R. Martins y P. Pascoe. 1993. The diet
of sperm whales (Physeter macrocephalus
Linnaeus, 1758) off the Azores. Philosophical
Transactions of the Royal Society of London, ser.
B, 339: 67-82.
Clarke, M.R. y N. Merrett 1972. The significance of
squid, whale and other remains from the
stomachs of bottom-living deep-sea fish. Journal
of the Marine Biological Association of the United
Kingdom, 52: 599-603.
Clarke, M.R. y E.R. Trueman (Editores). 1988. The
Mollusca. Volume 12. Paleontology and
Neontology of Cephalopods. Academic Press. San
Diego, 355 pp.
Clarke, M.R. y P.L. Pascoe 1997. Cephalopod species in
the diet of a sperm whale (Physeter catodon)
stranded at Penzance, Cornwall. Journal of the
Marine Biological Association of the United
Kingdom, 77: 1255-1258.
Clarke, M.R. y R.E. Young 1998. Description and
analysis of cephalopod beaks from stomachs of
six species of odontocete cetaceans stranded on
Hawaiian shores. Journal of the Marine Biological
Association of the United Kingdom, 78: 623-641.

Clarke, R. 1955. A giant squid swallowed by a sperm
whale. Norwegian Whaling Gazette, 1955(10):
589-593.
Clarke, R. 1956. Sperm whales of the Azores. Discovery
Report, 28: 237-298.
Clarke, R. y O. Paliza. 2000. The Humboldt Current
squid Dosidicus gigas (Orbigny, 1835). Revista de
Biología Marina y Oceanografía, 35(1): 1-39.
Clarke, W.J. y G.C. Robson 1929. Notes on the
stranding of giant squids on the North-East coast
of England. Proceedings of the Malacological
Society, 18(4): 154-158.
Coelho, M.L. 1985. Review of the influence of
oceanographic factors on cephalopod distribution
and life cycles. Northwest Atlantic Fisheries
Organisation (NAFO) Scientific Council Studies, 9:
47-57.
Collet, R. 1911-1912. Norges Pattedyr. H. Aschehoug &
Co., Kristiania, 744 pp.
Collins, M.A.1998. A female giant squid (Architeuthis)
stranded on the Aberdeenshire coast. Journal of
Molluscan Studies, 64(4): 489-492.
Collins, M.A., C. Lordan, K. Flannery, D.T.G. Quigley y
F.G. Howard 1997. New records of cephalopods
caught in Irish and Scottish waters. Journal of the
Marine Biological Association of the United
Kingdom, 77: 561-564.
Crosse, H. y P. Fischer. 1862. Nouveau documents sur
les céphalopodes gigantesques. Journal de
Conchyliologie, 10: 124-140.
Dawe E.G. y J.K.T. Brodziak. 1998. Trophic relations,
ecosystem variability and recritment. In:
Rodhouse P.G., E.G. Dawe, R. O’Dor, (Eds.).
Squid recruitment dynamics. FAO Fisheries
Technical Papers, 376: 125-150.
Dell, R.K. 1952. The Recent Cephalopoda of New
Zealand. Dominion Museum Bulletin Wellington,
16: 1-157.
Dell, R.K. 1970. A specimen of the giant squid
Architeuthis from New Zealand. Records of the
Dominion Museum, 7(4): 25-36.
Dunning, M.C., M.R. Clarke y C.C. Lu. 1993.
Cephalopods in the diet of oceanic sharks caught
off eastern Australia, pages 119-131. En: T.
Okutani, R.K. O'Dor y T. Kubodera (eds.) Recent
Advances in Fisheries Biology, Tokai University
Press, Tokyo, 752 pp.
Ehrhardt, N.M., P. Jacquemin, F. García, G. González,
J.M. López, J. Ortiz. y A. Solís. 1983. On the
fishery and biology of the giant squid Dosidicus
gigas in the Gulf of California, Mexico. FAO
Fisheries Technical Paper, 231: 306-340.
Ellis, R. 1998. The search for the giant squid. Penguin
Books, 322 pp.
Engeser, Th. y K. Bandel. 1988. Phylogenetic
classification of coleoid cephalopods. En:
Wiedmann, J. and J. Kullmann (Eds.) Cephalopods

– Present and Past: 105-115. Schweizerbart’sche
Verlagsbuchhandlung (Nägele u. Obermiller)
Stuttgart.
Fernández, E., J. Cabal, J.L. Acuña, A. Bode, A. Botas y
C. García-Soto. 1993. Plankton distribution across
a slope current-induced front in the southern Bay
of Biscay. Journal of Plankton Research. 15: 619-
641.
Fernández-Núñez, M.M. y C.L. Hernández-González.
1995. Report of a specimen of Architeuthis
Steenstrup caught in Canarian waters. En:
Guerra, A. (Ed.) Abstracts of the 12th
International Malacological Congress, Vigo, Spain:
343.
Fiscus, C.W. y D.W. Rice. 1974. Giant squids,
Architeuthis sp., from stomachs of sperm whales
captured off California. California Fish and Game,
60(2): 91-93.
Fiscus, C.W., D.W. Rice y A.A. Wolman. 1989.
Cephalopods from the stomachs of sperm whales
taken off California. NOAA Technical Report, NMFS
83: 1-12.
Fisher, W. y J.C. Hureau (eds.). 1985. FAO species
identification sheets for fishery purposes.
Southern Ocean (Fishing areas 48, 58 y 88)
(CCAMLR Convention Area). Vol. 1. Rome, FAO,
232 pp.
Förch, E.C. 1983. New Plymouth's giant squid. Catch
'83. Ministry of Agriculture and Fisheries,
Wellington, 10(10): 6-7.
Förch, E.C. 1984. Giant squid still a mystery. Catch '84.
Ministry of Agriculture and Fisheries, Wellington,
11(11): 19.
Förch, E.C. 1998. The marine fauna of New Zealand:
Cephalopoda; Oegopsida; Architeuthidae (Giant
squid). NIWA Biodiversity Memoir, 110: 113 pp.
Frost, N. 1934. Notes on a giant squid (Architeuthis
sp.) captured at Dildo, Newfoundland, in
December, 1933. Annual Report of the
Newfoundland Fishery Research Commission for
1933, 2(2): 100-113.
Frost, N. 1936. A further species of giant squid
(Architeuthis sp.) from Newfoundland waters.
Annual Report of the Newfoundland Fishery
Research Commission for 1935, 2(5): 89-95.
Gannon, E. 1996. Monster catch from the depths.
Herald-Sun Newspaper, Melbourne Australia, 11
June 1996.
Gaskin, D.E. y M.W. Cawthorn. 1967. Diet and feeding
habits of the sperm whale (Physeter catodon L.)
in the Cook Strait region of New Zealand. New
Zealand Journal of Marine and Freshwater
Research, 1(2): 156-179.
Gauldie, R.W., I.F. West y E.C. Förch. 1994. Statocyst,
statolith, and age estimation of the giant squid,
Architeuthis kirki. Veliger, 37(1): 93-109.

Gervais, P. 1875. Remarques au sujet des grands
céphalopodes decabrachides constituant le genre
Architeuthis. Journal de Zoologie, 4(2): 88-97.
Girard, A. A. 1892. Les céphalopodes des Iles Acores et
de l'Ile de Madère. Jornal de Ciencias
Matemáticas, Physicas e Naturaes, Lisboa, (ser.
2), 2(7): 210-220.
González, A.F. y A. Guerra. 1993. Composición química,
aspectos nutritivos y ecotoxicología de los
cefalópodos. En: Simal X. y Xose A. Fidalgo
(Eds.). Contaminación Ambiental e Alimentaria.
Cadernos do Laboratorio Ourensán de
Antropoloxía Social. Nº 5, Serie de Cursos e
Congresos, 3 (Bromatoloxía): 121-139.
González, A.F. y A. Guerra. 1996. Reproductive biology
of the short-finned squid Illex coindetii
(Cephalopoda, Ommastrephidae) of the
northeastern Atlantic. Sarsia 81: 107-118.
González, A.F., A. Guerra, S. Pascual y P. Briand. 1998.
Vulcanoctopus hydrothermalis gen. et sp. nov.
(Mollusca, Cephalopoda): an octopod from a
deep-sea hydrothermal vent site. Cahiers de
Biologie Marine, 39: 169-184.
González, A.F., A. Guerra, F. Rocha y J. Gracia. 2002.
Recent findings of the giant squid Architeuthis in
the northern Spanish waters. Journal of the
Marine Biological Association of the United
Kingdom, 82: 859-861.
González, A.F., A. Guerra y F. Rocha. 2003. New data
on the life history of the hooked squid Taningia
danae. Sarsia, 88: 297-301.
González A.F., P.N. Trathan, C. Yau, P.G. Rodhouse.
1997. Interactions between oceanography,
ecology and fishery biology of the ommastrephid
squid Martialia hyadesi in the south Atlantic.
Marine Ecology Progress Series, 152: 205-215.
González, M., M. Fernández-Casado, M.P. Rodriguez, A.
Segura y J.J. Martín. 2000. First record of the
giand squid Architeuthis sp (Architeuthidae) in the
Mediterranean Sea. Journal of the Marine
Biological Association of the United Kingdom, 80:
745-746.
Grieg, J.A. 1933. Cephalopods from the west coast of
Norway. Bergens Museum Aarbog,
Naturvidenskapelig rekke, 1933(4): 1-25.
Gronningsaeter, A. 1946. Sjormen-blekksprutten.
Naturen, 70(12): 379-380.
Guerra, A. 1992. Mollusca, Cephalopoda. En: Fauna
Ibérica, vol. 1. Ramos, M.A. et al., (Eds.). Museo
Nacional de Ciencias Naturales. CSIC. Madrid. 327
pp.
Guerra, A. 1996. Algunas consideraciones sobre el
envejecimiento de los cefalópodos. Iberus, 14 (2):
125-130.
Guerra, A. 2003. Los cefalópodos. Tomo III, cap.10,
part.VI: 366-374. Serie Naturaleza. Proyecto

Galicia. Hércules ediciones S.A. Santiago de
Compostela.
Guerra, A. y P. Sánchez. 1985. Comparación de las
pesquerías de cefalópodos de dos áreas de
afloramiento de la costa occidental africana.
Simposio. Internacional sobre el. Aforamiento del.
Oeste de África. Instituto de Investigaciones
Pespesqueras, Barcelona, vol II: 749-760.
Guerra, A., A.F. González, E.G. Dawe, y F. Rocha.
2004. Records of giant squid in the north-eastern
Atlantic, and two records of male Architeuthis sp.
off the Iberian Peninsula. Journal of Marine
Biological Association of the United Kingdom, 84:
427-431.
Guerra, A., A.F. González, F. Rocha, J. Gracia y M.
Vecchione. 2004. Calamares gigantes varados.
Víctimas de exploraciones acústicas. Investigación
y Ciencia, nº 334: 35-37.
Hall, H. 1990. Mugged by a squid. Ocean Realm, 6: 8.
Hamabe, M. 1957. A giant squid collected at Kuniga,
Oki Islands. Nihonkai Suisan Shiken Kenyu
Rebrajy Btysym, 82:3. [En japonés]
Hamilton, J.E. 1915. Belmullet Whaling Station. Report
to the Committee. Report of the British
Association for the Advancement of Science,
1914: 125-161.
Hanlon, R.T. y J.B. Messenger. 1996. Cephalopod
Behaviour. Cambridge University Press, 232 pp.
Harting, P. 1860. Description de quelques fragments de
deux céphalopodes gigantesques. Verhandelingen
der Koninklijke Akademie van Wetenschappen,
Amsterdam, 9(1)[1861]: 16 pages, 3 plates.
[Published separately, 1860; Publiées par
l'Académie Royale des Sciences a Amsterdam]
Harvey, M. 1874. Gigantic cuttlefishes of
Newfoundland. Annals and Magazine of Natural
History, (ser. 4)13(73): 67-69.
Hatanaka, H., S. Kawahara, Y. Uozumi y S. Kasahara.
1985. Comparison of life cycles of five
ommastrephid squid fished by Japan. Todarodes
pacificus, Illex illecebrosus, Illex argentinus,
Nototodarus sloani sloani and Nototodarus gouldi.
Northwest Atlantic Fisheries Organisation (NAFO)
Scientific Council Studies, 9: 59-68.
Heppell, D. 1978. Giant squid stranded at North
Berwick. Conchologists' Newsletter, 65: 89.
Heppell, D. y S.M. Smith. 1983. Recent Cephalopoda in
the collections of the Royal Scottish Museum,
Edinburgh. Royal Scottish Museum Information
Series, Natural History, 10: 1-81.
Herring, P.J., P.N. Dilly y C. Cope. 1992. Different types
of photophore in the oceanic squids Octopoteuthis
and Taningia (Cephalopoda: Octopoteuthidae).
Journal of Zoology, 227: 479-491.
Hess, S.C. y R.B. Toll. 1981. Methodology for specific
diagnosis of cephalopod remains in stomach
contents of predators with reference to the

broadbill swordfish, Xiphiasgladius. Journal of
Shellfish Research, 1(2): 161-170.
Heuvelmans, B. 1997. The kraken and the colosal
octopus. Kegan Paul.
Higa, L.1981. Recent finds. Hawaiian Shell News,
29(6): 9.
Hilgendorf, F.1880. Einen riesigen Dintenfisch aus
Japan, Megateuthis martensii g.n., sp.n. Sitzungs-
Bericht der Gesellschaft naturforschender Freude
zu Berlin, 4: 65-67.
Hochberg, F.G. y W.G. Fields. 1980. Cephalopoda: The
squids and octopuses: 429-444. En: R.H. Morris
et al. Intertidal Invertebrates of California.
Stanford University Press, 690 pp.
Holthe, T.1975. A new record of a giant squid from
Trondheimsfjorden Fauna, 28(3): 174.
Honma, Y., T. Kitami y R. Mizusawa. 1983. Record of
Cephalopoda in the waters adjacent to Niigata
and Sado Island in the Japan Sea, based partially
on the pelagic squids stranded ashore. Bulletin of
the Biogeographical Society of Japan, 38(3): 23-
29.
Hoving, H.-J.T., M.A.C. Roeleveld, M.R. Lipinski y Y.
Melo. 2004. Reproductive system of the giant
squid Architeuthis in Southern African waters.
Journal of Zoology, London, 264: 153-169.
Iwai, E.1956. Descriptions on unidentified species of
dibranchiate cephalopods. I. An oegopsiden squid
belonging to the genus Architeuthis. Scientific
Reports of the Whales Research Institute, Tokyo,
11: 139-151.
Jackson, G.D., C.C. Lu y M. Dunning 1991. Growth
rings within the statolith microstructure of the
giant squid Architeuthis. Veliger, 34(3): 331-334.
Jeffreys, J.G. 1869. British Conchology, volume 5. John
van Voorst, London.
Jepson, P.D. et alia. 2003. Gass-bubble lesions in
stranded cetaceans. Nature, 425 : 575-576.
Joubin, L. 1895. Contribution a l'étude des
Céphalopodes de l'Atlantique Nord. Résultats des
campagnes scientifiques accomplies sur yacht par
Albert I Prince souverain de Monaco, 9: 1-63.
Joubin, L.1900. Céphalopodes provenant des
campagnes de la Princesse Alice (1891-1897).
Résultats des campagnes scientifiques accomplies
sur son yacht par Albert I Prince souverain de
Monaco, 17: 1-135.
Joubin L. 1931. Notes preliminaries sur les
céphalopodes de croisiéres du Dana (1921-1922)
3e Partie. Institut Oceanographique. Monaco, 10:
169-211.
Kamita, T.1962. Visits by unusual aquatic animals to
the sea coasts of San-in District, Japan Sea. San-
in Bunka Kenkyu Kiyo, 2: 1-35. [En japonés]
Keil, A.1963. Riesentintenfische aus dem Pottwal-
Magen. Natur und Museum. Frankfurt, 93(8):
319-323.

Kent, W.S. 1874. Note on a gigantic cephalopod from
Conception Bay, Newfoundland. Proceedings of
the Scientific Meetings of the Zoological Society of
London, 1874(12): 178-182.
Kent, W.S. 1875. A further communication upon certain
gigantic cephalopods recently encountered off the
coast of Newfoundland. Proceedings of the
Scientific Meetings of the Zoological Society of
London, 1874(32): 489-494.
Kirk, T.W. 1880. On the occurrence of giant cuttlefish
on the New Zealand coast. Transactions and
Proceedings of the New Zealand Institute 1879,
12(2): 310-313.
Kirk, T.W. 1882. Descriptions of new Cephalopods.
Transactions and Proceedings of the New Zealand
Institute, 14(2): 283-286, 1 plate.
Kirk, T.W. 1888. Brief description of a new species of
large decapod (Architeuthis longimanus).
Transactions and Proceedings of the New Zealand
Institute 1887, 20(1): 34-39.
Kjennerud, J. 1958. Description of a giant squid,
Architeuthis, stranded on the West Coast of
Norway. Universitetet I Bergen. Årbok 1958,
Naturvitenkapelig Rekke, 9: 1-14.
Knudsen, J.1957. Some observations on a mature male
specimen of Architeuthis from Danish waters.
Proceedings of the Malacological Society of
London, 32(5): 189-198.
Kodolov, L.S.1972. Squids of the Bering Sea: 157-160.
En: P.A. Moiseev (ed.), Soviet fisheries
investigations in the northeastern Pacific, Part V.
Proceedings of All-Union Scientific Research
Institute of Marine Fisheries and Oceanography,
70. [Translated from Russian by N. Kaner, Israel
Program for Scientific Translations, 1972, 462
pp].
Kristensen, T.K. y J. Knudsen. 1983. A catalogue of the
type specimens of Cephalopoda (Mollusca) in the
Zoological Museum, University of Copenhagen.
Steenstrupia, 9(10): 217-227.
Kubodera, T. 2003. Architeuthis in Japanese waters.
Book of Abstracts of the CIAC meeting, Phuket,
Thailand: 50.
Kubodera, T. y K. Mori. 2005. First-ever observations of
a live giant squid in the wild. Proceedings of the
Royal Society B, 272: 2583-2586.
Kuroiwa, M. 1998. Exploration of the Jumbo Squid,
Dosidicus gigas, resources in the Southeastern
Pacific Ocean with notes on the history of jigging
surveys by the Japan Marine Fisheries Research
Center. En: Contributed Papers to International
Symposium on Large Pelagic Squids (T. Okutani,
ed.). Japan Marine Fishery Resources Research
Center, Tokyo, pp. 89-105.
Ladman, N.H., J.K. Cochram, C. Cerrato, J. Mark, C.F.E.
Roper y C.C. Lu. 2004. Habitat and age of the

giant squid (Architeuthis sanctipauli) inferred from
isotopic analyses. Marine Biology, 144: 685-691.
Leviton, A.E. y F.H. Gibbs Jr. 1988. Standards in
herpetology and ichthyology. Standard symbolic
codes for institutional resource collections in
herpetology and ichthyology. Supplement No. 1:
Additions and corrections. Copeia, 1988 (1): 28-
228.
Leviton, A.E., F.H. Gibbs Jr., E. Heal, y C.D.
Dawson.1985. Standards in herpetology and
ichthyology: Part I. Standard symbolic codes for
institutional resource collections in herpetology
and ichthyology. Copeia, 1985(3): 802-832.
Lipinski, M.R. 1993. The deposition of statoliths: a
working hypothesis, pages 241-262. In: T.
Okutani, R.K. O'Dor and T. Kubodera (eds.)
Recent Advances in Fisheries Biology, Tokai
University Press, Tokyo, 752 pp.
Lipinski, M.R. 1997. Morphology of giant squid
Architeuthis statoliths. South African Journal of
Marine Science, 18: 299-303.
Loates, G. 1989. A portfolio of drawings of Architeuthis
dux. Journal of Cephalopod Biology, 1(1): 86-94.
Lonnberg, E. 1891. Ofversigt ofver Sveriges
Cephalopoder. Kongliga Svenska Vetenskaps-
Akademiens Handlingar, 17(6): 1-41.
Lordan, C., M.A. Collins y C. Perales-Raya. 1998.
Observations on morphology, age and diet of
three Architeuthis caught off the west coast of
Ireland in 1995. Journal of the Marine Biological
Association of the United Kingdom, 78: 903-917.
Lu, C.C. 1986. Smallest of the largest - first record of
giant squid larval specimen. Australian Shell
News, 53: 9.
Lu, C.C., R. Boucher-Rodoni y A. Tillier. 1995.
Catalogue of types of recent Cephalopoda in the
Museum National d'Histoire Naturelle (France).
Bulletin du Muséum national d'Histoire naturelle,
Paris, (series 4) 17(3-4): 307-343.
Lu C.C. y M.R. Clarke. 1975. Vertical distribution of
cephalopods at 40oN, 53 oN and 60 oN at 20 oW
in the north Atlantic. Journal of the Marine
Biological Association of the United Kigdom, 55:
143-163.
MacAlaster, E. 1977. Cephalopods in the Nova Scotia
Museum collection. Curatorial Report, Nova Scotia
Museum, 33: 1-26.
MacPherson, J.H. y C.J. Gabriel. 1962. Marine Molluscs
of Victoria. Melbourne University Press, 475 pp.
Maddock, L. y J.Z. Young. 1987. Quantitative
differences among the brains of cephalopods.
Journal of Zoology. London., 212: 739-767.
Mangold, K. 1989. Céphalopodes. En: Traité de
Zoologie (Grassé, P.P.) Tome V, fascicule 4.
Masson, Paris, 804 pp.
Marshall, B.A. 1996. Molluscan name-bearing types in
the Museum of New Zealand Te Papa Tongarewa.

Tuhinga: Records of the Museum of New Zealand,
9: 1-85.
Martin, A.R. y M.R. Clarke. 1986. The diet of sperm
whales (Physeter macrocephalus) captured
between Iceland and Greenland. Journal of the
Marine Biological Association of the United
Kingdom, 66: 779-790.
Massy, A.L. 1909. The Cephalopoda Dibranchiata of the
coasts of Ireland. Fisheries, Ireland, Scientific
Investigations 1907, 1: 1-39.
Massy, A.L. 1913. Further records of the Cephalopoda
Dibranchiata of the coasts of Ireland. Fisheries,
Ireland, Scientific Investigations 1912, 5: 1-12.
Massy, A.L. 1928. The Cephalopoda of the Irish Coast.
Proceedings of the Royal Irish Academy, 38(B2):
25-37.
Masuda, S., K. Yokawa, A. Yatsu y S. Kawahara. 1998.
Growth and Population Structure of Dosidicus
gigas in the Southeastern Pacific Ocean. En:
Contributed Papers to International Symposium
on Large Pelagic Squids (T. Okutani, Ed.): 107-
118. Japan Marine Fishery Resources Research
Center, Tokyo.
McSweeny, E.S. 1970. Description of the juvenile form
of the Antarctic squid Mesonychoteuthis hamiltoni
Robson. Malacologia, 10: 323-332.
Mitsukari, K. y S. Ikeda. 1895. Notes on a gigantic
cephalopod. Zoological Magazine, Tokyo, 7(77):
39-50.
More, A.G. 1875a. Notice of a gigantic cephalopod
(Dinoteuthis proboscideus) which was stranded at
Dingle, in Kerry, two hundred years ago. The
Zoologist, London, (series 2)118: 4526-4532.
More, A.G. 1875b. Gigantic squid on the west coast of
Ireland. Annals and Magazine of Natural History,
(ser. 4)16(92): 123-124.
Murray, A. 1874a. Letter to the Secretary....
Proceedings of the Boston Society of Natural
History, 16: 161-163.
Murray, A. 1874b. Capture of a gigantic squid at
Newfoundland. American Naturalist, 8(2): 120-
123. [See also, Correction..., American Naturalist,
8(4): 226-227.]
Muus, B.J. 1959. Skallus, Sotaender, Blaeksprutter.
G.E.C. Gads Forlag, Kobenhaven, 239 pp.
Myklebust, B. 1946. Et nytt funn av
kjempebleblekksprut i Romsdal. Naturen, 70(12):
377-379.
Nesis, K.N. 1974. Giant squids. Priroda, 6: 55-60.
Nesis, K.N. 1982/87. Abridged key to the cephalopod
mollusks of the world's ocean. 385+ii pp. Light
and Food Industry Publishing House, Moscow. (In
Russian.). Translated into English by B. S.
Levitov, ed. by L. A. Burgess (1987), Cephalopods
of the world. T. F. H. Publications, Neptune City,
NJ, 351pp.

Nesis, K.N. 1983. Dosidicus gigas. En: Cephalopod Life
Cycles, Vol. 1. Species Accounts (P.R. Boyle, Ed.):
216-231. Academic Press, London.
Nesis, K.N. 1985. Oceanic cephalopods: Distribution,
life forms and evolution. Nauka, Moskow pp 1-
287.
Nesis, K.N. 1987. Cephalopods of the world; squids,
cuttlefishes and allies (English Translation). TFH
Publications, Neptune New Jersey pp 1-351.
Nesis, K.N. 1996. Mating, spawning and death in
oceanic cephalopods: a review. Ruthenica, 6: 23-
64.
Nesis, K.N. 2003. Distribution of recent Cephalopoda
and implications for Plio-Pleistocene events.
Berliner Palaobiologische Abhandlungen, 3: 199-
224.
Nesis, K.N., A.M. Amelekhina, A.R. Boltachev y G.A.
Shevtsov. 1985. Records of giant squids of the
genus Architeuthis in the north Pacific and south
Atlantic. Zoologicheskii Zhurnal, 64(4): 518-528.
Nigmatullin, C.M. 1976. Discovery of the giant squid
Architeuthis in equatorial waters of the Atlantic
Ocean. Biologiya Morya, Vladivostok, 2(4): 29-31.
Nigmatullin, Ch.M., K.N. Nesis y A.I. Arkhipkin. 2001. A
review of the biology of the jumbo squid Dosidicus
gigas (Cephalopoda: Ommastrephidae). Fisheries
Research, 54: 9-19.
Nishimura, S. 1960. Visits by unusual aquatic animals
to the sea coasts of Niigata prefecture, Japan Sea,
in the winter of 1959-1960. Saishu to Shiiku, 22:
213-216. [En japonés]
Nishimura, S. 1968. A preliminary list of the pelagic
Cephalopoda from the Japan Sea. Publications of
the Seto Marine Biological Laboratory, 16(1): 71-
83.
Nixon, M. 1984. Stranding of giant squid on Scottish
coast. Cephalopod Newsletter, (8): 4.
Nixon, M. y J.Z. Young. 2003. Brains and lives of
cephalopods. Oxford Academic Press, 406 pp.
Nordgard, O. 1923. The Cephalopoda Dibranchiata
observed outside and in the Trondhjemsfjord.
Meddelelse Fra Trondhjems Biologiske Station,
16: 1-14.
Nordgard, O. 1928. Faunistic notes on marine
evertebrates III. Det Kongelige Norske
Videnskabers Selskab. Forhandlinger, 1(26): 70-
72.
Norman, M. 2000. Cephalopods. A world guide.
ConchBooks. Germany, 320 pp.
Norman, M.D. y C.C. Lu. 1997. Sex in giant squid.
Nature, 389: 683-684.
O'Dor, R.K. y N. Balch. 1985. Properties of Illex
illecebrosus egg masses potentially influencing
larval oceanographic distribution. Northwest
Atlantic Fisheries Organisation Scientific Council
Studies, 9: 69-76.

O’Dor, R.K y D.M. Webber. 1986. The constraints on
cephalopods: why squid aren’t fish. Canadian
Journal of Zoology, 64: 1591-1605.
Ohsumi, S. y Y. Satake. 1976. Provisional report on
investigations of sperm whales off the coast of
Japan under a special permit. International
Whaling Commission Meeting (La Jolla, 1976),
Document IWC/SC SP76/16.
Okiyama, M. 1965. Some information on the eggs and
larvae of the common squid, Todarodes pacificus
Steenstrup. Bulletin of the Japanese Regional
Fisheries Research Laboratory, 15: 39-53.
Okiyama, M. 1993. Kinds, abundance and distribution
of the oceanic squids in the Sea of Japan. En: T.
Okutani, R.K. O'Dor y T. Kubodera (Eds.): 403-
415. Recent Advances in Fisheries Biology, Tokai
University Press, Tokyo.
Okiyama, M. y S. Kasahara. 1975. Identification of the
so-called "common squid eggs" collected in the
Japan Sea and adjacent waters. Bulletin of the
Japanese Regional Fisheries Research Laboratory,
26: 35-40.
Okutani T. 1974. Epipelagic decapod cephalopods
collected by micronekton tows during the
EASTROPCA Expeditions, 1967-1968. Bulletin
Tokai Regional Fisheries Research Laboratory, 80:
29-118.
Okutani, T. (ed.). 1998. Contributed papers to
International Symposium on Large Pelagic Squids.
Japan Marine Fishery Resources Research Center.
Tokyo, Japan. 269 pp.
Okutani T. y M.R. Clarke. 1985. Identification key and
species description for Antarctic squids. BIOMASS
Handbook, 21: 1-57.
Okutani, T., Y. Satake, S. Ohsumi y T. Kawakami.
1976. Squids eaten by sperm whales caught off
Joban District, Japan, during January-February,
1976. Bulletin of Tokai Regional Fisheries
Research Laboratory, 87: 67-113.
Okutani T. y Y. Satake. 1978. Squids in the diet of 38
sperm whales caught in the Pacific off northern
Honshu, Japan, February 1977. Bulletin Tokai
Regional Fisheries Research Laboratory, 93: 13-
27.
Ortea, J.A. y G. Flor. 1986. Asturias desde el mar.
Ayalga. Salinas. Asturias, 218 pp.
O’Shea, S. 2004. The giant octopus Haliphron atlanticus
(Mollusca: octopoda) in New Zealand waters. New
Zealand Journal of Zoology, 31: 7-13.
Owen, R. 1881. Descriptions of some new and rare
Cephalopoda (Part II). Transactions of the
Zoological Society of London, 11(5): 131-170.
Packard, A. 1972. Cephalopod and fish: The limits of
convergence. Biological Reviews of the Cambridge
Philosophical Society, 47: 241-307.
Packard, A. 1988. The skin of Cephalopods (Coleoids):
General and special adaptations. En: Wilbur,

K.M.(Ed.), vol. 11. Form and Function. Trueman,
E.R. y M.R. Clarke (Eds.): 37-67. Academic Press
Inc.
Packard, A.S. Jr. 1873. Colossal cuttlefishes. American
Naturalist, 7(2): 87-94.
Pereira, J., R. Rosa, A. Moreno, M. Henriques, J.
Sendao y T.C. Borges. 2005. First recorded
specimen of the giant squid Architeuthis sp in
Portugal. Journal of the Marine Biological
Association of the United Kingdom, 85: 175-176.
Pérez-Gándaras, G. y A. Guerra. 1978. Nueva cita de
Architeuthis (Cephalopoda: Teuthoidea):
descripción y alimentación. Investigación
Pesquera, 42(2): 401-414.
Pérez-Gandaras, G. y A. Guerra 1989. Architeuthis de
Sudáfrica: nuevas citas y consideraciones
biológicas. Scientia Marina, 53(1): 113-116.
Pfeffer, G. 1912. Die Cephalopoden der Plankton-
Expedition. Zugleich eine monographische
ubersicht der Oegopsiden Cephalopoden.
Ergebnisse der Plankton-Expedition der Humboldt-
stiftung, 2: 1-815.
Pinkas, L., M.S. Oliphant y I.L.K. Iverson. 1971. Food
habits of albacore, bluefin tuna, and bonito in
California waters. California Department of Fish
and Game, Fish Bulletin, 152: 1-105.
Pippy, J. y F.A. Aldrich. 1969. Hapatoxylon trichiuri
(Holden, 1802) (Cestoda, Trypanorhyncha) from
the giant squid Architeuthis dux Steenstrup, 1857
in Newfoundland. Canadian Journal of Zoology,
47: 263-264.
Pontoppidan, E. 1752. Det forste forsog paa Norges
naturlige Historie. Ludolph Henrich Lillie, 1752-
1753, Kobenhavn.
Pörtner, H.O., R.K. O' Dor y D.L. Macmillan. 1994.
Physiology of Cephalopod Molluscs: Lifestyle and
Peformance Adaptations. Gordon and Breach
Publishers, 214 pp.
Posselt, H.J. 1890. Cephalopoda, volume 2: 135-145.
En: C.G.J. Petersen (ed.) Det videnskabelige
Udbytte af Kanonbaaden "Hauchs" Togter i de
Danske Have indenfor skagen i Aarene 1883-
1886, 5 volumes and atlas, Kjobenhavn, 1889-
1893.
Posselt, H.J. 1898. Gronlands Brachiopoder og Bloddyr.
[Conspectus Faunae Groenlandicae. Brachiopoda
et Mollusca.] Meddelelser om Gronland, udgivne
af Commissionen for Ledelsen af de geologiske og
geographiske Undersogelser i Gronland, 23(1): 1-
298.
Puente, M. 1996. Testing waters to learn more about
giant squid. USA Today Newspaper, 1 February
1996.
Rae, B.B. 1950. Description of a giant squid stranded
near Aberdeen. Proceedings of the Malacological
Society of London, 28: 163-167.

Rasero, M., A.F. González, B.G. Castro y A. Guerra.
1996. Predatory relationships of two symaptric
squid, Todaropsis eblanae and Illex coindetii
(Cephalopoda: Ommastrephidae) in Galician
waters. Journal of the marine biological
Association of the United Kigdom, 76: 73-87.
Rathjen, W.F. 1973. Northwest Atlantic squids. Marine
Fisheries Review, 35(12): 20-26.
Ré, M.E., P.J. Barón, J.C. Berón, A.E. Gosztonyi, L.
Kuba, M.A. Monsalve y N.H. Sardella. 1998. A
giant squid Architeuthis sp. (Mollusca,
Cephalopoda) stranded on the Patagonian shore
of Argentina. South African Journal of Marine
Biology, 20: 109-122.
Rees, W.J. y G.E. Maul. 1956. The Cephalopoda of
Madeira, records and distribution. Bulletin of the
British Museum (Natural History), Zoology, 3(6):
259-281.
Rey, I., O. Soriano, B.A. Dorda, A.F. González, F. Rocha
and A. Guerra. 2006. First Cytochrome C Oxidase
I and 16S RNA genes partial sequences and
phylogenetic relationships in the hooked giant
squid Taningia danae. Cephalopod Life Cycles,
Cephalopod International Advisory Council
Symposium. Abstract book: 95. Hobart,
Tasmania.
Ríos, A.F., F. Fraga, F.G. Figueiras, R. Prego y F.F.
Pérez. 1987. Campañas oceanográficas "Asturias
I, II, III y IV". Datos Informativos del Instituto
Investigaciones Marinas, Vigo 22: 1-140
Ritchie, J. 1918. Occurrence of a giant squid
(Architeuthis) on the Scottish coast. Scottish
Naturalist, 73: 133-139.
Ritchie, J. 1920. Giant squid cast ashore on N. Uist,
Outer Hebrides. Scottish Naturalist, 99: 57.
Ritchie, J. 1922. Giant squids on the Scottish coast.
Report of the British Association for the
Advancement of Science, 1921: 423.
Robison, B.H. 1989. Depth of occurrence and partial
chemical composition of a giant squid,
Architeuthis, off Southern California. Veliger,
32(1): 39-42.
Robson, C.W. 1887. On a new species of giant cuttlefish
stranded at Cape Campbell, June 30th 1886
(Architeuthis kirkii). Transactions and Proceedings
of the New Zealand Institute, 19(1): 155-157.
Robson, G.C. 1933. On Architeuthis clarkei, a new
species of giant squid, with observations on the
genus. Proceedings of the Zoological Society of
London, 1933(3): 681-697.
Rocha, F., A. Guerra y A.F. González. 2001. A review of
the reproductive strategies in cephalopods.
Biological Reviews, 76: 291-304.
Rodhouse, P. 1991. Population structure of Martialia
hyadesi (Cephalopoda: Ommastrephidae) at the
Antarctic Polar Front and the Patagonian shelf,

South Atlantic. Bulletin of Marine Science, 49(1-
2): 404-418.
Rodhouse P.G. y M.R. Clarke. 1985. Growth and
distribution of young Mesonychoteuthis hamiltoni
Robson (Mollusca: Cephalopoda): an Antarctic
squid. Vie Milieu,34: 223-230.
Rodhouse P.G. y U. Piatkowski. 1995. Fine-scale
distribution of juvenile cephalopods in the Scotia
Sea and adaptive allometry of the brachial crown.
Marine Biology, 124 : 111-117.
Rodríguez-Navarro, A., A. Guerra, Ch. S. Romanek, F.
Rocha and A. F. González. 2006. Life history of
the giant squid Architeuthis as revealed from
stable isotope and trace elements signatures
recorded in its beak. Cephalopod Life Cycles,
Cephalopod International Advisory Council
Symposium, Abstract book: 97. Hobart,
Tasmania.
Roe, H.S.J. 1969. The food and feeding habits of the
sperm whales (Physeter catodon L.) Taken off the
west coast of Iceland. Journal du Conseil, 33(1):
93-102.
Roeleveld, M.A.C. 2000. Giant squid beaks: implications
for systematics. Journal of the Marine Biological
Association of the United Kingdom, 80(1): 185-
187.
Roeleveld, M.A.C. 2002. Tentacle morphology of the
giant squid Architeuthis from the North Atlantic
and Pacific oceans. Bulletin of Marine Science, 71:
725-737.
Roeleveld, M.A. y J. Knudsen. 1980. Japetus
Steenstrup: On the Merman (called the Sea Monk)
caugth in the Øresund in the time of King
Christian III. Steenstrupia, 6(17): 293-332.
Roeleveld, M.A. y M.R. Lipinski. 1991. The giant squid
Architeuthis in southern African waters. Journal of
Zoology, London, 224 (3): 431-477.
Roper, C.F. y K.J. Boss. 1982. The giant squid.
Scientific American, 246(4): 82-89.
Roper, C.F.E. y R.E. Young. 1972. First record of
juvenile giant squid, Architeuthis (Cephalopoda:
Oegopsida). Proceedings of the Biological Society
of Washington, 85(16): 205-222.
Roper, C.F.E. y G.L. Voss. 1983. Guidelines for
taxonomic descriptions of cephalopod species.
Memoirs National Museum Victoria, 44: 49-63.
Roper, C.F.E., M.J. Sweeney y C. Nauen. 1984. FAO
species catalogue. Cephalopods of the world. An
annotated and illustrate catalogue of species of
interest to fisheries FAO Fisheries Synopsis, 3
(125), 277 pp.
Roper, C.F.E., y M. Vecchione. 1993. A geographic and
taxonomic review of Taningia danae Joubin, 1931
(Cephalopoda: Octopoteuthidae), with new
records and observations on bioluminescence. En:
Okutani, T., R.K. O’Dor y T. Kubodera (Eds.).

Recent Advances in Fisheries Biology: 441-456
Tokai University Press, Tokyo.
Ruiz, A. y R. Fondacaro. 1996. Un calamar gigante en
aguas de al plataforma patagónica. Jornadas
patagónicas de Medio Ambiente, Universidad
Nacional de la Patagonia San Juan Bosco, 9-11
October 1996, Esquel – Chubut. Patagonia
Argentina. Abstracts: 95.
Santos, M.B., G.J. Pierce, A.F. González, F. Santos,
M.A. Vázquez, y M.A. Collins. 2001. First records
of Taningia danae (Cephalopoda, Octopodidae) in
Galician waters (NW Spain) and in Scottish waters
(United Kingdom). Journal of the Marine Biological
Association of the United Kigdom, 81: 355-356.
Sasaki, M. 1929. A monograph of the dibranchiate
cephalopods of the Japanese and adjacent waters.
Journal of the College of Agriculture, Hokkaido
Imperial University, 20(supplement): 1-357, 159.
Satomi, N. 1979. Hokurikuno-shizenshi, Umi-hen
[Natural History of Kokuriku district -Marine].
Kougen-sha Press, Toyama Japan, 159 pp.
Saunders, W.B. y N.H. Landman. 1987. Nautilus. The
biology and paleobiology of a living fossil. Plenum
Press: 632 pp.
Sivertsen, E. 1955. Blekksprut. Kongelige Norske
Videnskabers Selskab (Museet) Arbok, 1954: 5-
15.
Sleptsov, M.M. 1955. Biology of cephalopod mollusks
from the Far Eastern Seas and the northwestern
Pacific. Trudy Instituta Okeanologii Im P.P.
Shirshova, Akademiya Nauk SSSR, 18: 69-77. [En
ruso]
Staub, F. 1993. Requin bleu, calmar géant et cachalot.
Proceedings of the Royal Society of Arts and
Sciences of Mauritius, 5(3): 141-145.
Steenstrup, J. 1849. Meddelelse om tvende
kiaemlpestore blaeksprutter, opdrevne 1639 og
1790 ved Islands Kyst, og om nogle andre
nordiske dyr. Forhandlinger ved de Skandinaviske
Naturforskeres Syvende Mode 5, (1849):950-957.
[Traducción inglesa: Note on two giant
cephalopods drifted ashore in 1639 and 1790 on
the coast of Iceland, and on some other northern
animals. En: Volsoe, A., J. Kundsen and W. Rees,
The cephalopod papers of Japetus Steenstrup.
Danish Sciende Press Ltd. Copenhagen, 1962: 9-
13].
Steenstrup, J. 1855. Kjaeber af en kolossal
blaeksprutte. Oversigt over det Kongelige Danske
Videnskabernes Selskabs Forhandlinger,
1855(5/6):199-200.Traducción inglesa: On the
jaws of a giant cuttle-fish. En Volsoe, et al., 1962:
14-15.
Steenstrup, J. 1857. Oplysninger om Atlanterhavets
colossale blaeksprutter. Forhandlinger ved de
Skandinaviske Naturforskeres Syvende Mode 7,
(1856):182-185. Traducción inglesa: Prof.

Steenstrup presented several particulars about
the giant cuttle-fishes of the Atlantic Ocean. En
Volsoe, et al., 1962: 17-19.
Steenstrup, J. 1882. Notae teuthologicae, 1-4. Oversigt
over det Kongelige Danske Videnskabernes
Selskabs Forhandlinger, 1882:143-168. En:
Volsoe, et al., 1962: 145-162.
Steenstrup, J. 1898. Sepolia atlantica. Kolossale
Blaeksprutter fra det nordlige Atlanterhav. Danske
Videnskabernes Selskabs Skrifter, 5 Raekke,
Naturvidenskabelig og Mathematisk, 4(3)
(1898):409-456, figures A-G, 4 plates. En Volsoe,
et al., 1962: 258-308.
Stephen, A. 1937. Notes on Scottish cephalopods.
Scottish Naturalist, 227: 131-132.
Stephen, A. 1944. The Cephalopoda of Scottish and
adjacent waters. Transactions of the Royal Society
of Edinburgh, 61(1): 247-270.
Stephen, A. 1950. Giant squid, Architeuthis, in
Shetland. Scottish Naturalist, 62(1): 52-53.
Stephen, A. 1953. Rare invertebrates recently found in
the Scottish area. Scottish Naturalist, 65(2): 120-
121.
Stephen, A. 1962. The species of Architeuthis inhabiting
the North Atlantic. Proceedings of the Royal
Society of Edinburgh, 68(2): 147-161.
Stephen, A., B.B. Rae y E. Wilson. 1957. Rare marine
invertebrates recently found in the Scottish area.
Scottish Naturalist, 69(3): 178-181.
Stevenson, J.A. 1935. The cephalopods of the Yorkshire
coast. The Journal of Conchology, 20(4): 102-
116, 7 plates.
Stommel, H. 1975. La circulación abisal. Selecciones de
Scientific American. Oceanografía: 75-80.
Storm, V. 1897. Om 2 udenfor Trondhjemsfjorden
fundne kjaempeblaekspruter. Naturen. Illustreret
Maanedskrift for Populaer Naturvidenskab,
Bergen, 21(4): 97-102.
Sweeney, M.J. 2001. Records of Architeuthis specimens
from published reports. National Museum of
Natural History. Smithsonian Institution, 132 pp.
Tanaka, I. 1950. Glory of rare fishes. Hagi Bunka
Sosho, no. 2. [En japonés]
Tarasevich, M.N. 1968. Food composition of sperm
whales in the northern Pacific. Zoologicheskii
Zhurnal, 47(4): 595-601. [En ruso]
Toll, R.B. y S.C. Hess. 1981a. Cephalopods in the diet
of the swordfish, Xiphias gladius, from the Florida
Straits. Fishery Bulletin, U.S., 79(4): 765-774.
Toll, R.B. y S.C. Hess. 1981b. A small, mature, male
Architeuthis (Cephalopoda: Oegopsida) with
remarks on maturation in the family. Proceedings
of the Biological Society of Washington, 94(3):
753-760.
Tsuchiya, K. y K. Mori. 1998. A note on an early
juvenile specimen of Architeuthis sp. collected
from the south off Central Honshu, Japan

(Cephalopoda: Architeuthidae). Venus, 57(3):
225-230.
Van Heukelem, W.F. 1983. Octopus maya. En: Boyle,
P.R. editor. Cephalopod life cycles: species
accounts. Vol.1. Academic Press, London, pp.
311-324.
Vecchione, M. y R.E. Young. 1998. The Magnapinnidae,
a newly discovered family of oceanic squid
(Cephalopoda: Oegopsida). En: Payne, A.I.L.,
M.R. Lipinski, M.R. Clarke y M.A.C. Roeleveld
(Eds.). Cephalopod Biodiversity, Ecology and
Evolution. South African Journal of Marine
Science, 20: 429-437.
Vecchione, M., R.E. Young, A. Guerra, D.J. Lindsay,
D.A. Clague, J.M. Bernhard, W.W. Sager, A.F.
González, F.J. Rocha, y M. Segonzac. 2001.
Worlwide observations of remarkable deep-sea
squids. Science, 294(5551): 2505-2506.
Vélain, C. 1875. Observations effectuées a l'Ile Saint-
Paul. Comptes Rendus Hebdomadaires des
Séances de l'Académie des Sciences, 80(15):
998-1003.
Vélain, C. 1877. Passage de Venus sur le Soleil (9
December 1874). Expédition Française aux Iles
Saint-Paul et Amsterdam. Zoologie. Observations
générales sur la faune des deux îles suivies d'une
description des Mollusques. Archives de Zoologie
Expérimentale et Générale, 6: 1-144.
Vélain, C. 1878. Passage de Venus sur le Soleil (9
December 1874) Expédition Française aux Iles
Saint-Paul et Amsterdam sous le commandement
de M. Mouchez, Capitaine de Vaisseau.
Remarques générales au sujet de la faune des
Iles Saint-Paul et Amsterdam suivies d'une
description de la faune malacologique des deux
îles. Thèse présentée a la Faculté des Sciences de
Paris, A. Hennuyer, Paris, 143 pages, 9 figures, 5
plates.
Verrill, A.E. 1874a. Occurrence of gigantic cuttle-fishes
on the coast of Newfoundland. American Journal
of Arts and Sciences, 7(38): 158-161.
Verrill, A.E. 1874b. The giant cuttle-fishes of
Newfoundland and the common squids of the New
England coast. American Naturalist, 8(3): 167-
174.
Verrill, A.E. 1875a. The colossal cephalopods of the
North Atlantic. American Naturalist, 9(1): 21-36.
Verrill, A.E. 1875b. The colossal cephalopods of the
North Atlantic, II. American Naturalist, 9(2): 78-
86.
Verrill, A.E. 1875c. Brief contributions to zoology from
the Museum of Yale College. No. XXXV. Notice of
the occurrence of another gigantic cephalopod
(Architeuthis) on the coast of Newfoundland, in
December, 1874. American Journal of Science and
Arts, (ser. 3), 10(57): 213-214.

Verrill, A.E. 1876. Note on gigantic cephalopods,-- a
correction. American Journal of Science and Arts,
(ser. 3), 12(69): 236-237.
Verrill, A.E. 1877. Occurrence of another gigantic
cephalopod on the coast of Newfoundland.
American Journal of Science and Arts, (ser. 3),
14(83): 425-426.
Verrill, A.E. 1878. Notice of recent additions to the
marine fauna of the eastern coast of North
America. American Journal of Science and Arts,
(series 3), 16(93)[Sep.]: 207-215.
Verrill, A.E. 1880a. The cephalopods of the north-
eastern coast of America. Part I. The gigantic
squids (Architeuthis) and their allies; with
observations on similar large species from foreign
localities. Transactions of the Connecticut
Academy of Sciences, 5(5): 177-257.
Verrill, A.E. 1880b. Synopsis of the Cephalopoda of the
North-eastern coast of America. American Journal
of Science, (ser. 3), 19(112): 284-295.
Verrill, A.E. 1881. The cephalopods of the north-eastern
coast of America. Part II. The smaller
cephalopods, including the "squids: and the
octopi, with other allied forms. Transactions of the
Connecticut Academy of Sciences, 5(5): 259-446.
Verrill, A.E. 1882a. Occurrence of an additional
specimen of Architeuthis at Newfoundland.
American Journal of Science, (ser. 3), 23(133):
71-72.
Verrill, A.E. 1882b. [Review of] Descriptions of some
new and rare Cephalopoda; by R. Owen.
American Journal of Science, (ser. 3), 23(133):
72-75.
Verrill, A.E. 1882c. [Review of] Description of new
Cephalopoda; by T.W. Kirk. American Journal of
Science, (ser. 3), 24(144): 477.
Villanueva, R. y P. Sánchez. 1993. Cephalopods of the
Benguela Current off Namibia: new additions and
considerations on the genus Lycoteuthis. Journal
of Natural History, 27: 15- 46.
Voss, G.L. 1956. A review of the cephalopods of the
Gulf of Mexico. Bulletin of Marine Science, 6(2):
85-178.
Voss, G.L. 1967. Squids; jet-powered torpedos of the
deep. National Geographic Magazine, 131(3):
386-411.
Voss, G.L. 1968. The Biology and bathymetric
distribution of deep sea cephalopods. Studies in
Tropical Oceanography, 5: 511-535.
Voss, N. A. 1980. A generic revision of the Cranchiidae
(Cephalopoda; Oegopsida). Bulletin of Marine
Science, 30: 365-412.
Voss, N.A., M. Vecchione, R.B. Toll y M. Sweeney
(Eds.). 1998. Systematics and Biogeography of
Cephalopods. Smithsonian Institution,
Smithsonian Contributions to Zoology, nº 586.
Washington D.C., vol.2, 599 pp.

Vovk, A.N., K.N. Nesis y B.G. Panfilov. 1975.
Distribution and deep sea cephalopods in the
South Atlantic and adjacent waters (mainly
analyses of sperm whales feeding), pp. 162-164.
(Abstract.) V th Meeting on the Investigation of
Molluscs, 11-14 February 1975, Leningrad.
Academy of Sciences USSR, Institute of Zoology.
[Traducción: Malacological Review, 11(1978):
131.].
Ward, P. y K. Bandel. 1987. Life history strategies in
fossil cephalopods. En: Boyle, P.R. (Ed.).
Cephalopod Life Cycles. vol. 2. Comparative
Reviews: 329-350. Academic Press. London.
Watt, J, G.J. Pierce y P.R. Boyle. 1997. A guide to the
premaxillae and vertebrae of North Sea fishes.
ICES (Co-operative Research Repport), no. 220:
1-231.
Wells, M.J. 1978. Octopus. Physiology and Behaviour of
an Advanced Invertebrate. Chapman and Hall,
London: 417 pp.
Wells, M.J. 1990. The dilemma of the jet set. New
Scientist, Febrero, 1990: 44-47.
Wilhelm, O. 1930. Las mortandades de jibias
(Omastrephes gigas) en la Bahía de Talcahuano.
Boletín de la Sociedad de Biología de Concepción
(Chile), 3: 23-37.
Wirz, K. 1959. Étude biométrique du système nerveux
des Céphalopodes. Bulletin Biologique de la
France et de la Belgique, 93: 78-117.
Wood, J.B. y R.K. O’Dor. 2000. Do larger cephalopods
live longer? Effects of temperature and phylogeny
on interspecific comparisons of age and size at
maturity. Marine Biology 136: 91-99.
Xampeny, J. y S. Filella. 1976. Datos sobre tres
cachalotes Physeter marcocephalus L. capturados
frente las costas atlánticas de Galicia, España
(Cetacea, Physeteridae). Miscellánea Zoológica, 3:
235-242.
Yamashiro, C., L. Mariátegui, J. Rubio, J. Arguelles, R.
Tafur, A. Taipe, y M. Rabí. 1998. Jumbo Flying
Squid fishery in Peru. En: Contributed Papers to
International Symposium on Large Pelagic Squids
(T. Okutani, Ed.): 119-125. Japan Marine Fishery
Resources Research Center, Tokyo.
Yatsu, A., K. Katto, F. Kakizoe, K. Yamanaka y K.
Mizuno. 1999. Distribution and biology of
Sthenoteuthis oualaniensis in the Indian Ocean -
Preliminary results from the research cruise of the
R/V Shoyo-Mary in 1995. En: Contributed Papers
to International Symposium on Large Pelagic
Squids (T. Okutani, Ed.): 145-153. Japan Marine
Fishery Resources Research Center, Tokyo.
Yatsu, A., R. Tafur y C. Maravi. 1999. Embryos and
Rhynchoteuthion paralarvae of the Jumbo Flying
Squid Dosidicus gigas (Cephalopoda) obtained
through artificial fertilization from Peruvian
Waters. Fisheries Science, 65(6): 904-908.

Yatsu, A., K. Yamanaka y C. Yamashiro. 1999. Tracking
experiments of the Jumbo Flying Squid, Dosidicus
gigas, with an ultrasonic telemetry system in the
Eastern Pacific Ocean. Bulletin of the National
Research Institute of far Seas Fisheries, 36: 55-
60.
Young, J.Z. 1964. A model of brain. Clarendon Press.
Oxford.
Young, J.Z. 1971. The anatomy of the nervous system
of Octopus vulgaris. Clarendon Press. Oxford.
Young, J.Z. 1977. Brain behaviour and evolution in
cephalopods. Symposium of the Zoological
Society of London, 38: 377-434. Academic Press.
London and New York.
Young, R.E. y R.F. Harman. 1988. "Larva", "Paralarva"
and "subadult" in cephalopod terminology.
Malacologia, 29 (1): 201-207.
Yukhov, V.L. 1974. The discovery of giant squids.
Priroda, Moscow, (6): 60-63, 3 figures. [En ruso]
Zeidler W. 1981. A giant deep-sea squid, Taningia sp.,
from South Australian waters. Transactions Royal
Society South Australia, 105: 218.
Zeidler, W. 1996. Forget the calamari! Australia Nature,
25(2): 7-8.
Zeidler, W. y K.L. Gowlett-Holmes. 1996. A specimen of
giant squid, Architeuthis sp., from South
Australian waters. Records of the South Australian
Museum, 29(1): 85-91.
Zuev, G.V. y K.N. Nesis. 1971. Volume II. Squid
(Biology and Fishing). En: Selected English
Translations of Publications on Cephalopods by Kir
N. Nesis (M.J. Sweeney, Ed.). Smithsonian
Institution Libraries (2003), 291 pp.

LOS AUTORES
Ángel Guerra Sierra (Madrid, España,
1947) se incorporó al Instituto de
Investigaciones Pesqueras en 1973. Doctor
en Biología por la Universidad de Barcelona
(1977). Desde 1976 trabaja en el Instituto
de Investigaciones Marinas de Vigo.
Profesor de Investigación del Consejo
Superior de Investigaciones Científicas
(CSIC), director del Departamento de Recursos
Marinos y Ecología Marina y jefe del grupo de
investigación Ecología y Biodiversidad Marina
(ECOBIOMAR). Ha dirigido veinticinco proyectos
de investigación y dirigido o participado en treinta
y ocho contratos de investigación con empresas y
administraciones. Hasta la fecha, ha publicado o
editado ocho libros, contribuyendo con capítulos
específicos en nueve volúmenes colectivos y es
autor de un centenar y medio de trabajos
científicos en revistas nacionales e
internacionales, así como de más de un centenar
de comunicaciones presentadas a congresos,
simposios y reuniones. Ha dirigido doce campañas
oceanográficas en el Atlántico, Pacífico y
Mediterráneo. Fue presidente del Centro Nacional
de Investigaciones Marinas (CSIC), Vicedirector
del Instituto de Investigaciones Marinas,
presidente de la sociedad internacional Unitas
Malacológica entre 1992 y 1995, periodo en el
cual organizó el XII Internacional Malacological
Congress. Fundador y miembro del primer y
quinto comité ejecutivo del Cephalopod
Internacional Advisory Council, es también
miembro de numerosas asociaciones nacionales e
internacionales de Biología Marina, perteneciendo
al consejo editorial de cuatro revistas científicas
de carácter internacional y al comité consultor de
veinte revistas de investigación, habiendo sido
editor jefe de las revistas científicas de la
Sociedad Española de Malacología. Es consultor
de varias comisiones de la Unión Europea y
asesor científico de varias comisiones nacionales e
internacionales. Su labor investigadora y docente,
realizada fundamentalmente a través diecisiete
tesis doctorales, cinco tesis de licenciatura y de
los cursos del ciclo de doctorado impartidos en las
tres universidades gallegas, son pioneras en
España en el campo de la Biología y Ecología de
los Cefalópodos. Ha impartido cursos y
conferencias en varias universidades europeas y
americanas. Su tarea de divulgación de la Ciencia
se concreta, además, en la publicación de una
treintena de artículos. Ha recibido varios premios
y distinciones por su labor científica.

Ángel F. González González (Vigo,
España, 1966) estudió Ciencias Biológicas
en la Universidad de Navarra,
incorporándose en 1991 al grupo de
investigación ECOBIOMAR del Instituto de
Investigaciones Marinas de Vigo (CSIC),
donde desarrolló su Tesis Doctoral
(Universidad de Vigo, 1994). En 1995
disfrutó de una beca post-doctoral en la Graduate
School of Oceanography (Universidad de Rhode
Island, USA), estudiando la dinámica trófica y
ordenación pesquera de los cefalópodos en
Georges Bank (Noreste USA). En 1996 realizó
trabajos de investigación en el British Antarctic
Survey, NERC (Cambridge, Reino Unido), siendo
investigador principal de un proyecto que
abordaba las relaciones entre la oceanografía y
las pesquerías de cefalópodos en el Atlántico Sur.
Posteriormente, fue el representante del Comité
para la Conservación de los Recursos Marinos
Antárticos (CCAMLR) y del NERC, y jefe de la
primera campaña de investigación realizada
durante el invierno austral en la Antártica, con
objeto del establecimiento de una nueva
pesquería en aguas de South Georgia y South
Sándwich (área 48.3 CCAMLR). Desde entonces
ha continuado su actividad investigadora en el
IIM-CSIC, realizando estancias cortas en Francia,
Alemania e Inglaterra. Ha participado en 17
proyectos de investigación, publicado sesenta
artículos en revistas científicas, varios capítulos
de libros, presentado más de medio centenar de
ponencias en congresos nacionales e
internacionales, es miembro del comité editorial
de la revista Fisheries Research y revisor de otras
doce revistas científicas. Ha dictado cursos de
doctorado en varias Universidades y es profesor
de dos Master para licenciados. Actualmente, es
Científico Titular de CSIC.
Francisco J. Rocha Valdés (Santiago, Chile,
1967) estudió Licenciatura en Ciencias con
Mención en Biología en la Universidad de Chile
(1986-1990), después de lo cual se radicó en
España, donde se doctoró en Biología por la
Universidad de Oviedo (1994). En Chile, su
actividad científica se desarrolló en la Universidad
de Chile (1988 a 1991) y el Instituto de Fomento
Pesquero (entre 1990 y 1991). Una vez en
España, el Dr. Rocha se incorporó en 1992 al
grupo de investigación ECOBIOMAR del Instituto
de Investigaciones Marinas de Vigo (CSIC).

Sus líneas de investigación se han orientado
hacia la biología, ecología y pesquerías de
organismos marinos, especialmente cefalópodos,
en Europa y Pacífico sureste. Como miembro de
ECOBIOMAR, ha participado en la disección y
estudio de los ejemplares de calamar gigante
encontrados en Asturias desde el inicio,
participando como investigador en el proyecto
Kraken y siendo coautor de varios trabajos
científicos sobre los resultados obtenidos. Como
investigador ha participado en 16 proyectos
internacionales y convenios de investigación. Sus
publicaciones suman un total de 44 artículos
científicos, 18 informes técnicos y tres volúmenes
colectivos, junto a 49 presentaciones a congresos.
Entre 1996 y 2000 fue editor adjunto de la
Revista científica Iberus, de la Sociedad Española
de Malacología, publicando un total de siete
volúmenes. En cuanto a su actividad docente, ha
dirigido tres tesis de Licenciatura y una de
doctorado. Finalmente, ha dictado clases en
diversos cursos, tanto de carrera, como
internacionales y de ciclo de doctorado, en las
Universidades de Chile (1988-1991 y 1999),
Pontificia Universidad Católica de Chile (1990-
1991), Nacional Andrés Bello (1991), Austral de
Chile (2003) y de Vigo (España, 1997-2005).
Joaquín Gracia Salvador (Valencia,
España, 1964) es experto en
programación y técnico especialista en
electrónica, además de ser buzo
profesional y diseñador de equipos
submarinos para investigación científica.
Ha trabajado como técnico y analista de
sistemas en diversos centros de todo el
mundo, como la central hidráulica y nuclear de
Cortes de Payas - Cofrentes (Valencia), y en
investigación y desarrollo de sistemas electrónicos
para Breakthrough Computers, PTY, LTD
(Towsville - Queensland (Australia). En 1998 fue
contratado dentro del grupo ECOBIOMAR del
Instituto de Investigaciones Marinas de Vigo
(CSIC) cómo especialista en informática y
desarrollo de sistemas electrónicos, donde
permaneció hasta 2001. Entre 2001 y 2004 ha
participado activamente en la realización del
proyecto Kraken, contratado por Transglobe Films
como Director de Producción. Joaquín Gracia ha
participado en los estudios del calamar gigante de
Asturias desde el inicio, primero como miembro
de ECOBIOMAR y luego desde Transglobe Films. A
él se debe la creación y desarrollo de la mayoría
de los equipos informáticos y submarinos
utilizados durante las campañas del proyecto

Kraken. Actualmente es director gerente de su
propia empresa de I+D: Marexi, Tecnología
Marina Ximo, S.L., destinada al diseño y
construcción de equipos submarinos de alta
tecnología. Ha impartido clases en la Universidad
James Cook y en el centro de formación de
Breakthrough Computers en Australia (1995-
1996). Tanto en Australia como en España a
participado en numerosos proyectos de
investigación y desarrollo relacionados con el
estudio de la fauna marina y los ecosistemas.
Gracias a ello tiene una amplia experiencia en
investigación marina y desarrollo de instrumental
científico y es coautor de varios trabajos de
investigación y ponencias a congresos.
Luis José Laria De La Maza (Arriendas,
España, 1959) es naturalista y buzo
profesional con amplia experiencia en
fotografía y filmación subacuática. A
realizado numerosos reportajes submarinos
en los mares de Cuba, Santo Domingo,
Bahamas, México, Panamá, Costa Rica,
California, Baja California, Egipto, Arabia
Saudita, Malasia, Borneo, Maldivas, Indonesia,
Tailandia, Sulawesi, Argentina, Antártica, etc.
Radicado en Luarca, su interés por la
conservación de la naturaleza le llevó a fundar, en
1996, la Coordinadora para el Estudio y
Protección de las Especies Marinas (CEPESMA) y,
posteriormente, el Museo Aula del Mar donde se
encuentra una de las mejores y mayores
colecciones de calamares gigantes del mundo.
Presidente de CEPESMA desde 1996, es
responsable de la Red de Varamientos de
Mamíferos Marinos del Principado de Asturias y
del Centro de Recuperación de Especies Marinas
en Luarca. Luis Laria ha publicado trabajos de
divulgación y notas de prensa relativas a la
conservación de especies y sus estudios sobre
mamíferos marinos y el calamar gigante. Al
mismo tiempo, ha colaborado con la Universidad
de Oviedo en cursos de extensión universitaria
sobre oceanografía y recursos marinos. Encargado
de las colecciones del Aula del Mar, no sólo ha
participado activamente en el estudio del calamar
gigante, sino que fue miembro del Proyecto
Kraken y ha ayudado en la organización y puesta
a punto de varias exposiciones museísticas sobre
organismos marinos, especialmente cefalópodos y
mamíferos marinos, en diversos puntos de España
y Francia.
View publication statsView publication stats





























