Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Estructura y producción primaria delEstructura y producción primaria delfitoplancton y perifiton en unfitoplancton y perifiton en un
humedal del bajo Paranáhumedal del bajo Paraná
Rodríguez, Patricia Laura
2008
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Rodríguez, Patricia Laura. (2008). Estructura y producción primaria del fitoplancton y perifitonen un humedal del bajo Paraná. Facultad de Ciencias Exactas y Naturales. Universidad deBuenos Aires.
Cita tipo Chicago:
Rodríguez, Patricia Laura. "Estructura y producción primaria del fitoplancton y perifiton en unhumedal del bajo Paraná". Facultad de Ciencias Exactas y Naturales. Universidad de BuenosAires. 2008.

UNIVERSIDAD DE BUENOS AIRES
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
DEPARTAMENTO DE ECOLOGÍA, GENÉTICA Y EVOLUCIÓN
ESTRUCTURA Y PRODUCCIÓN PRIMARIA DEL FITOPLANCTON Y
PERIFITON EN UN HUMEDAL DEL BAJO PARANÁ
Tesis presentada para optar por el título de Doctor de la Universidad de Buenos Aires en
el área Ciencias Biológicas
Patricia Laura Rodríguez
Director de tesis: Dra. Haydée Norma Pizarro
Consejero de Estudios: Dr. Héctor Guillermo Tell
Laboratorio de Limnología
Departamento de Ecología, Genética y Evolución
Facultad de Ciencias Exactas y Naturales
Universidad de Buenos Aires
Buenos Aires, 2008

ÍNDICE
CONTENIDO
PÁGINA
Resumen i
Summary ii
Agradecimientos iii
Dedicatoria v
Introducción 1
La producción primaria 5
Métodos de estimación de producción primaria 6
Curvas de Fotosíntesis-Irradiancia (P-I) 9
Producción integrada en la columna de agua (PA) 13
Objetivo General 15
Área de Estudio 17
La Reserva Natural Otamendi 18
Capítulo I
Estructura de la comunidad algal perifítica sobre sustratos naturales en el humedal de la RNO
24
Introducción 25
Materiales y Métodos 29
Perifiton sobre sustratos naturales 33
Resultados
Variables ambientales 35
Perifiton sobre sustratos naturales 42
Discusión 55
Capítulo II
Estructura y producción primaria del fitoplancton y perifiton en la Laguna Grande
61
Introducción 62
El fitoplancton 62

CONTENIDO PÁGINA
Antecedentes 64
La estructura del fitoplancton en la RNO 65
El perifiton 66
Antecedentes 69
La estructura del perifiton en la RNO 70
Hipótesis 71
Objetivos 71
Materiales y Métodos 72
Variables ambientales 72
Fitoplancton 74
Curva P-I 75
Curva P-P 78
Trabajo de laboratorio 79
Cálculo de las tasas de asimilación 79
Producción integrada en la columna de agua (PA) 81
Perifiton 82
Análisis de datos 86
Resultados 86
Variables ambientales 86
Fitoplancton 99
Perifiton sobre sustratos artificiales 106
Comparación entre fitoplancton y perifiton 112
Discusión 114
Capítulo III
Comparación de la estructura y producción primaria del fitoplancton y perifiton en dos ambientes lumínicos contrastados
131
Introducción 132
Materiales y Métodos 133
Fitoplancton
135

CONTENIDO PÁGINA
Perifiton sobre sustratos artificiales 135
Resultados 137
Discusión 147
Capítulo IV
Comparación de la producción primaria de dos fracciones de tamaño fitoplanctónicas en tres cuerpos de agua de la RNO
152
Introducción 153
Materiales y Métodos 157
Resultados
Laguna Grande 160
ROL y Laguna Grande 169
El Pescado, ROL y Laguna Grande 190
Discusión 201
Capítulo V
Efecto de la atenuación lumínica sobre la producción primaria del fitoplancton: Un estudio en mesocosmos
213
Introducción 214
Materiales y Métodos 217
Análisis de datos 221
Resultados 221
Discusión 233
Discusión Final 238
Bibliografía 244

i
ESTRUCTURA Y PRODUCCIÓN PRIMARIA DEL FITOPLANCTON Y PERIFITON EN UN HUMEDAL DEL BAJO PARANÁ
RESUMEN Se estudió la estructura y producción primaria del fitoplancton y del perifiton en cuerpos de agua húmicos de la Reserva Natural Otamendi, humedal de la llanura de inundación del bajo Paraná, Argentina. Inicialmente, se analizó estacionalmente la composición algal perifítica sobre sustratos naturales y se vio que la forma de vida del sustrato afectó la estructura algal. Los estudios de producción primaria, realizados a través de curvas P-I (método de 14C), tuvieron diferentes aproximaciones. En primer lugar se compararon los parámetros fotosintéticos y los patrones de productividad anuales del fitoplancton y perifiton sobre sustratos artificiales en la Laguna Grande. El fitoplancton contribuyó en un 96 % a la producción primaria microbiana y su dinámica anual estuvo fuertemente influenciada por la temperatura. Luego, se comparó la productividad de ambas comunidades en dos cuerpos de agua con ambientes lumínicos contrastantes. El perifiton en el ambiente limitado lumínicamente presentó valores más altos de producción primaria. Después, se estudió la variación de los parámetros fotosintéticos y el aporte de las fracciones de tamaño > 3 µm y picoplanctónica a la producción fitoplanctónica de tres cuerpos de agua. El picofitoplancton presentó en general los mayores valores de asimilación y eficiencia fotosintética y no se encontraron diferencias en la contribución de ambas fracciones de tamaño a la producción total. Con el aumento del estado trófico de los cuerpos de agua, la tendencia fue a la disminución del aporte del picoplancton a la producción y biomasa fotosintética fitoplanctónica. Finalmente, se estudió en mesocosmos el efecto de atenuación lumínica producida por las macrófitas flotantes en la producción fitoplanctónica de la Laguna Grande. Los valores de producción disminuyeron progresivamente en condiciones de 2 % de transmitancia de luz. En estos escenarios los organismos aclimatados a bajas intensidades lumínicas mostraron mayor eficiencia fotosintética. Palabras clave: perifiton, fitoplancton, humedal, producción primaria, parámetros fotosintéticos.

ii
STRUCTURE AND PRIMARY PRODUCTION OF PHYTOPLANKTON AND PERIPHYTON IN A WETLAND FROM THE LOWER PARANÁ RIVER
BASIN
SUMMARY The structure and primary production of phytoplankton and periphyton were studied in humic water bodies from the Natural Reserve Otamendi, wetland from the Lower Paraná River Basin, Argentina. First, the algal periphytic composition on natural substrata was analyzed and we observed that life form of the substrata affected the algal structure of the community. The different approximations of primary production studies were carried out trough P-I curves (14C method). Formerly, phytoplankton and periphyton annual photosynthetic parameters and productivity patterns were analyzed on artificial substrata in Laguna Grande shallow lake. Phytoplankton accounted the highest contribution to total microbial primary production (96 %) and temperature was the main environmental factor affecting their annual dynamics. Moreover, we compared the productivity of both communities in two water bodies with contrasting light environments. Periphyton in the more restricted light environment reached highest values of primary production. Next, dynamics of photosynthetic parameters and the contribution of phytoplankton size fractions > 3 µm and picoplankton to total production was analyzed in three shallow lakes. Overall, picophytoplankton showed highest assimilation and efficiency values, but no significant differences were found in terms of the input of both size fractions to total phytoplankton production. Notwithstanding a trend to a lowest contribution of picoplankton to total photosynthetic biomass and production with the increase in trophic status of the systems was observed. Finally, we studied in mesocosms the effects of light attenuation triggered by free floating plants on phytoplankton primary production in Laguna Grande. Primary production decreased under such scenarios where only 2 % of incident irradiance was transmitted. Moreover, low light acclimated organisms were the most light efficient. Key words: periphyton, phytoplankton, wetland, primary production, photosynthetic parameters.

iii
AGRADECIMIENTOS Cuando comenzamos los muestreos de esta tesis, con Haydée pensamos que iban a ser los agradecimientos más largos del mundo. Es que nos ayudaron tantas personas en los muestreos… Al principio venían siete por cada curva P-I, hasta que resolvimos el asunto. La cuestión es que durante estos años atravesamos tantas pero tantas situaciones que a esta altura cada una puede decir de la otra: Te conozco. Y eso está bueno. Gracias Haydée por llevarme a conocer el mundo, por cargar la parrilla conmigo, por transmitirme humildad, por aguantarme, por presentarme a carrera, por permitirme aprender todo lo que yo quisiera y por escucharme, escucharme siempre. Gracias por la confianza y por haber creído en mí aún cuando yo no lo hacía. Gracias por permitirme tener el mejor trabajo del mundo. Gracias por TODO. Muchas gracias Guillermo por compartirme el mundo de las algas y dejarme formar parte de tu grupo de investigación. Gracias también por tu alegría y tus sabios consejos, los de la biología y los de la vida. Irina, mi ángel guardián. Nunca me voy a olvidar que fuiste la primera persona con la que hablé en el laboratorio. Todavía no había cursado Limnología y me consideraste para hacer un seminario. Gracias por tu ejemplo de trabajo, de compromiso con lo que hacés y de lucha permanente. Por tu amor a la ciencia y el espíritu de equipo. Por compartir tu conocimiento y tu inmensa generosidad. Gracias.
Gabilingui, gracias por tanta confianza infundamentada. Gracias por estirarme tanto los cuentos y sacarle sangre a las rocas por mí. Por enseñarme tanto en la docencia y de mientras aconsejarme prudencia. Porque no hay idea que caiga al vacío cuando hablo con vos. Por cazarme siempre con cuidado, con dedales y esperanza, con sonrisas y jabón...
Gracias Inés por acompañarme a tantos muestreos, por la paciencia y por los mates. Por ayudarme siempre. Gracias Alicia por estar siempre dispuesta a darme una mano y algún consejo muy precavido. Gracias Luz por abrirme el camino y por largar todo lo que estás haciendo para prestarme atención. Al dúo de rubias, Laurita y Romi. A la primera su extrema sensibilidad y por tratar de cuidarme y ayudarme siempre, por preocuparse por mí y después negarlo (porque es chúcara). A Romi por su buen humor y por tratar que no nos peleemos con Laurita. Ah!!! y porque desde que entró Romi al laboratorio dejé de preparar mate porque a ella le salen mejor! A las dos por hacer más divertidas las tardes.
Como Rubén Lombardo comparte su número Kármico conmigo, yo me pienso que cualquier cosa que le diga la va a compartir y aprobar. Ha sido un placer para mí compartir nuestros diálogos estadísticos y esotéricos contigo. También mis monólogos, porque de esos hubo muchos. Por tu atención constante hacia los demás. A Paula y Rodri. A Paula por su acompañamiento durante toda la carrera del doctorado. Fue una suerte y un placer que lo hayamos podido compartir. Por bancarme durante todo este tiempo. Por sus cuentos de lugares remotos y por los regalitos! Y a Rodri, que después de todo nos hicimos amigos. Porque siempre me da opiniones acertadas que nunca quiero escuchar y luego me arrepiento. A los dos porque sin ellos estos años no hubieran sido lo mismo.
A Solange por su ayuda fundamental tanto en los muestreos como en el laboratorio de radioisótopos (donde hacía mucho calor!!!). A mis demás compañeros del laboratorio, Sole, Eugenia, Griselda y Gabita por aguantarme y por la buena onda. A mis amigos de la facultad y del oeste. A María Llames, que ya no se si es mi amiga de la facultad o del oeste. A Luciano. Compañeros de Ecología General y alumnos que me

iv
cedieron fotos de la Reserva Otamendi. A mi familia. A Luisa. Gracias por ayudarme a crecer, a ver dentro de mí, y encontrar mi camino.
Las personas del grupo de los siete y que son más de siete. Los que ayudaron en las curvas P-I y en los demás muestreos: María Llames, Iri, Inés, Rodri, Rubén, Paulita, Adrián, Lili, Valeria, Matías, Julieta, Daniela, Solange, Laurita y todos aquellos de los que no me acuerdo a propósito y los que no recuerdo sin querer…
Al Dr. Horacio Zagarese y Lic. Elizabeth Chornomaz por introducirnos en la metodología del carbono 14. Al Dr. Walter Helbling por contestar muy amablemente todas mis dudas y al personal de la Reserva Natural Otamendi (APN) por su buena predisposición. A la Dra. Nora Maidana por su ayuda en la determinación taxonómica de las diatomeas y a la Dra. María Dos Santos Afonso por el asesoramiento en cuestiones químicas. Al Dr. Paul del Giorgio por la determinación del COD de las muestras del capítulo I y al personal del INTA Delta por cedernos amablemente los datos de altura del río Paraná de Las Palmas y precipitaciones del área de estudio. Al Dr. Gonzalo Pérez por tomar los datos de Kd espectral del capítulo II y a personal del servicio de Hidrografía Naval por facilitarnos los horarios de salida y puesta de sol en las fechas de muestreo. Hay personas que para mí, durante estos años fueron sinónimo de alegría. Una de ellas es mi hermano Ricardo. Muchas gracias por venirme a visitar cuando me aislaba a escribir la tesis y por compartir la nada y el todo. Por ayudarme a reparar mi vida, por acompañarme siempre a donde sea que me lleve la biología y la vida. Por ser mi enfermero y mi amigo. Por estar y acompañarme SIEMPRE. No sé qué haría sin vos. Mi tío Nené me trajo alegría durante muchos almuerzos en la etapa de redacción y en la niñez también, ya que me traía chocolates. Gracias por enseñarme a amar y disfrutar la vida, por ser un ejemplo de dignidad ante todo y por el apoyo. Ojalá mi acompañamiento en sus vidas esté a la altura de lo que significan ellos para mí.
Gracias mamita querida, sin tu ayuda no sería nada. Gracias por apoyarme incluso cuando no sabías qué era lo que apoyabas. Por despertarme a la mañana para que no llegue tarde a ningún lado, por enviarme víveres y por tus consejos de mami. Gracias Martín por nuestra conexión especial. Porque hay cosas que no podría hablar con nadie más que con vos. Por introducirme en la mitología griega y luego compartirla conmigo. Por escucharme y por participarme de tu mundo mágico. A Facundo y Coca por recibirme siempre en mi hogar del oeste.
Don Benito, que fue el primer ecólogo que conocí. Me enseñó que los grillos no se matan y que los sapos comen las moscas. Que hay lugar para todos en el reino del Señor y que cada ser vivo cumple un rol. Me dijo que yo era cabeza dura y que el mundo era para los audaces. Me explicó que yo podía ver bajo el agua y por eso quise estudiar Limnología. Quise aprovechar mi virtud, si es que la tenía. Me diste ejemplo de tu vida y en 20 años que te conocí me diste consejos para siempre. Ya nos volveremos a ver. Las investigaciones de esta tesis fueron financiadas por los siguientes proyectos: CONICET PIP 5355, ANPCyT PICT 536 y 12332, y UBACYT X815 y X195.

v
-Pá, quiero estudiar Biología
-¿Qué?!!! ¿Acaso querés estudiar el comportamiento del bicho Moro?
A veces visita la sandía en la Gaona y a veces no…
A Benito

INTRODUCCIÓN

2
Existen actualmente muchas definiciones de humedales que se basan en diferentes
aspectos de los mismos y, si bien no existe consenso absoluto acerca de cuáles son los
elementos que conforman un humedal, en el transcurso de esta tesis adoptaremos la
definición de humedal propuesta por la Convención Ramsar (Secretaría de la Convención
de Ramsar 2006). Allí se los define como extensiones de marismas, pantanos, turberas o aguas de
régimen natural o artificial, permanente o temporal, estancada o corriente, dulce, salobre o salada,
incluyendo las extensiones de aguas marinas cuya profundidad en marea baja no exceda de seis metros.
Considerando lo abarcativo de esta definición, los humedales constituyen entonces
paisajes inundados ó inundables que, en nuestro planeta, cubren 8,6 millones de km2, lo
que representa el 6,4 % de la superficie de la tierra (Mitsch & Goselink 1993). La
presencia de macrófitas acuáticas y los suelos anaeróbicos son características que
distinguen a los humedales de otros tipos de cuerpos de agua. Los suelos saturados de
agua se tornan anaeróbicos debido a que el oxígeno disuelto presente es rápidamente
consumido por los organismos (van der Valk 2006). Las macrófitas, por otro lado, juegan
un rol fundamental en estos sistemas e influyen en su metabolismo (Engelhardt & Ritchie
2001).
Los humedales exhiben una enorme diversidad de ambientes de acuerdo a su
génesis, ubicación geográfica, régimen hídrico y químico, plantas dominantes y
características del suelo o sedimento (Maltby 1991). Se los considera ambientes valiosos ya
que, además que son importantes reservorios de agua dulce y que muchos de ellos son
ecosistemas altamente productivos, proveen de una serie de servicios tales como refugio
de biodiversidad, prevención de inundaciones y sequías, purificación del agua por la
retención de contaminantes y sedimentos y permiten la realización de actividades
recreativas. Más recientemente se ha comenzado a estudiar su rol a escala global en el
ciclo del carbono, azufre y nitrógeno a través de la biosfera (Mitsch & Wu 1995). En
particular, interesa su dinámica en cuanto al metabolismo del carbono en el contexto de
los estudios de cambio global. Los humedales son actualmente considerados como
pequeños sumideros de dióxido carbono y grandes fuentes de metano. El metano es un
importante gas de efecto invernadero ya que posee un potencial de calentamiento 21
veces mayor que el dióxido de carbono (Rohem 2005).
Las llanuras de inundación de algunos ríos presentan áreas periódicamente inundables que
alternan fases terrestres y fases acuáticas. Contienen un abanico de diferentes ambientes

3
formando humedales en los que los cambios físico-químicos son muy marcados. Estos
cambios tan frecuentes causan la respuesta de la biota a través de adaptaciones
morfológicas, fisiológicas, anatómicas, fenológicas y etológicas (Junk et al. 1989). Neiff
(1999) propuso la sigueinte definición para los grandes humedales sudamericanos: Sistemas
de cobertura sub-regional en los que la presencia temporal de una capa de agua de espesor variable
(espacial y temporalmente) condiciona flujos biogeoquímicos propios, suelos con acentuado hidromorfismo y
una biota peculiar por procesos de selección, que tiene patrones propios en su estructura y dinámica.
Pueden considerarse como macrosistemas cuya complejidad crece con la variabilidad hidrosedimentológica y
la extensión geográfica ocupada. En Sudamérica la mayoría de los grandes ríos están rodeados
por extensas llanuras aluviales, que pueden superar los 300000 km2 e incluir un gran
número de lagos poco profundos de diferentes características limnológicas (Welcomme
1985).
La cuenca del río Paraná es la segunda en importancia en Sudamérica, luego de la
del río Amazonas. Posee una longitud cercana a los 3780 km y una extensa llanura aluvial
de aproximadamente 40000 km2, contribuyendo anualmente con el 77 % de la descarga
del Río de la Plata (500 106 m3 año-1) (Carignan et al. 1994, Drago 1990). A diferencia de
los ríos tropicales Amazonas y Orinoco, que se caracterizan por poseer un régimen de
inundación unimodal, predecible y de gran intensidad, el río Paraná suele tener períodos
de inundación menos predecibles, más irregulares y de menor intensidad. En el río
Amazonas la diferencia entre aguas altas y bajas puede llegar a ser de 15 m, mientras que
en el río Paraná esta oscilación suele ser de 1 a 3 m (Carignan et al. 1994). Además el ciclo
hidrológico del río Paraná se encuentra superpuesto al ciclo estacional, lo que no ocurre
en ambientes tropicales.
Algunos cuerpos de agua de los humedales poseen grandes cantidades de materia
orgánica proveniente de las macrófitas y el plancton presentes en ellos y de la cuenca
circundante. La acumulación de grandes cantidades de materia orgánica en
descomposición les confiere a estos sistemas características húmicas, que imparten un
color amarillo al agua, con importantes consecuencias en la absorción de luz,
particularmente en la zona azul del espectro. Las sustancias húmicas son polímeros
orgánicos que contienen anillos aromáticos unidos a largas cadenas alquilo; y pueden ser
desde compuestos solubles de bajo peso molecular a agregados de macromoléculas
insolubles (Kirk 1994). La presencia de sustancias húmicas puede influenciar la estructura
de las redes tróficas planctónicas y el metabolismo general de todo el cuerpo de agua

4
(Williamsom et al. 1999, Jones 1992). En primer lugar, alteran el ambiente físico y
químico, modificando así la productividad primaria y toda la trama trófica que depende de
ella. Las sustancias húmicas absorben radiación de longitud de onda corta, modificando
de esta manera el clima lumínico del cuerpo de agua (Kirk 1994). La capa eufótica suele
estar reducida en sitios con altas concentraciones de sustancias húmicas; en este sentido
tendrían un efecto negativo sobre la productividad primaria (Jones 1992). Las sustancias
húmicas pueden acomplejarse con el fósforo y el hierro, reduciendo de esta manera su
biodisponibilidad para el fitoplancton, hecho que también contribuye a la disminución de
la productividad primaria (Jackson & Hecky 1980). Por otro lado, las sustancias húmicas
pueden actuar como fuente de carbono en las tramas tróficas y promover el crecimiento
del bacterioplancton (Tranvik 1988, Jones 1992), que pasa a competir con el fitoplancton
por los nutrientes inorgánicos (Drakare et al. 2002).
Gran parte del área de la Reserva Natural Otamendi (RNO) constituye un humedal
emplazado sobre la llanura de inundación del río Paraná en su tramo inferior. Los cuerpos
de agua de la RNO poseen características húmicas y presentan gran desarrollo de
macrofitia flotante y palustre. La macrofitia puede ser muy importante en su aporte a la
producción primaria en los humedales (van der Valk 2006) y en particular, las macrófitas
flotantes influencian la productividad de las comunidades que viven por debajo (Nahlik &
Mitsch 2006). Además de las macrófitas, las algas juntamente con las cianobacterias
contribuyen en gran medida a la productividad de estos ecosistemas (van der Valk 2006).
Las comunidades algales en los humedales comprenden una serie de ensambles
estructuralmente superpuestos e íntimamente relacionados, y a fines prácticos se pueden
reconocer las siguientes categorías de grupos de organismos (Goldsborough & Robinson
1996):
- Perifiton: son las algas adheridas a un sustrato natural o artificial, vivo o
muerto. Adquieren distintas denominaciones de acuerdo al tipo de sustrato
donde se encuentre asentada la comunidad. En los humedales se pueden
reconocer más comúnmente al epifiton y epipelon.
Epipelon: incluye las algas móviles que habitan en los sedimentos blandos.
Un tipo de comunidad relacionada es el plocon, que incluye grupos de
algas no migratorias que se forman en la superficie de los sedimentos,

5
compuesto principalmente por cianobacterias y diatomeas. Estos grupos se
desprenden ocasionalmente debido a la formación de burbujas de gas y
flotan cerca de la superficie del cuerpo de agua.
Epifiton: compuesto de algas postradas, erectas y filamentos heterotricos
que crecen tanto en las superficies de plantas sumergidas como raíces de las
plantas flotantes.
- Metafiton: son matas de algas flotantes o subsuperficiales, principalmente
compuestas de algas verdes filamentosas. Generalmente las matas se originan
como epifiton que es desprendido por la turbulencia del agua y pueden flotar
debido a la presencia de burbujas gaseosas dentro de la mata.
- Fitoplancton: incluye las algas que viven en suspensión en el seno de la
columna de agua. Pueden o no ser móviles, y en ocasiones pueden provenir del
desprendimiento desde el epipelon o epifiton.
El aporte de la fracción algal microbiana a la productividad de los humedales
puede ser importante e incluso en ocasiones puede exceder a la de las macrófitas
(Goldsborough et al. 2005). La contribución relativa de los distintos grupos de algas a la
productividad de un humedal dependerá, entre otros factores, de la presión de pastoreo,
el contenido de nutrientes, altura hidrométrica, estabilidad de la columna de agua,
presencia de sustrato colonizable, penetración de la luz en la columna de agua, presencia o
no de macrófitas, etc.
La producción primaria
Los términos tasas de fotosíntesis, producción y fijación de carbono suelen ser utilizados
en general como sinónimos aunque no tengan estrictamente el mismo significado. La
fotosíntesis es la conversión biológica de energía lumínica en energía de enlace químico
que es almacenada en la forma de compuestos de carbono. Provee la fuente primaria de
materia orgánica para el crecimiento y las demandas metabólicas de los demás organismos
del ecosistema. De esta manera, la tasa de fotosíntesis pone un límite superior sobre la
biomasa total y productividad de los ecosistemas y restringe el flujo de energía sobre la
superficie de este planeta (Falkowski & Raven 2007). El término producción bruta se
refiere a la tasa de fijación de carbono sin considerar las pérdidas respiratorias, en tanto

6
que sí se tienen en cuenta cuando se habla de producción neta. El término producción
primaria se refiere a la tasa con la cual la energía es fijada, por medio del proceso
fotosintético, en la forma de sustancias orgánicas que pasan a formar parte de la biomasa
de los autótrofos. Estas sustancias orgánicas luego pueden ser utilizadas como alimento
por otros organismos. Se utiliza a veces el término productividad primaria, que se refiere
en particular al rendimiento fotosintético (ej. producción por unidad de clorofila a). Por
último, la fijación de carbono se refiere a cuánto carbono es asimilado por unidad de
tiempo y generalmente se estima mediante la incorporación de 14C durante el proceso de
fotosíntesis pero sin tener en cuenta su destino (Kalff 2003). En este sentido, el carbono
fijado puede pasar a formar parte de los organismos, ser excretado o secretado al
ambiente en forma de carbono orgánico disuelto ó CO2 respiratorio. La tasa de fijación de
carbono en general es menor que la tasa de producción de oxígeno y la relación entre los
moles de O2 producidos por moles de CO2 incorporados se denomina cociente
fotosintético (CF) (Sakshaug et al. 1997). El CF suele ser mayor que 1 debido a que no
toda la energía capturada por los fotosistemas se utiliza en la fijación de carbono, una
fracción se utiliza por las células para reducir nitrato y en menor grado sulfato (Falkowski
& Raven 2007).
Métodos de estimación de producción primaria
Dada la ecuación general de la fotosíntesis oxigénica (el agua es el donante de electrones y
se libera oxígeno):
6CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O
las técnicas para estimar las tasas de fotosíntesis en comunidades acuáticas están basadas
en la estequiometría de esta ecuación, ya sea por la medición de las tasas de producción de
oxígeno, consumo de CO2, o por cambios en la concentración de materia orgánica
(Wetzel & Likens 2000).
La forma más antigua y directa de estimar la producción primaria es a través de la
medición del oxígeno disuelto en botellas claras y oscuras. La misma fue desarrollada para
el fitoplancton y se basa en la incubación de botellas claras y oscuras por cada
profundidad seleccionada en el perfil de la columna de agua. En general se utilizan,
dependiendo de la profundidad, tres o cuatro pares de botellas por encima de la

7
profundidad de compensación, uno alrededor de ella y uno o dos por debajo. Las botellas
se incuban a la profundidad de donde fue extraída el agua con la que se las llenó, durante
un período de tiempo relativamente largo que depende del estado trófico del cuerpo de
agua (el tiempo de incubación es mayor a medida que disminuye el estado trófico). Se
debe realizar una medición de la concentración del oxígeno disuelto inicial de cada sitio de
toma de muestra, a partir de la cual se realizarán las comparaciones posteriores de la
evolución del gas. En el caso de emplearse el método de Winkler para la determinación
del oxígeno disuelto (que implica la fijación in situ del oxígeno mediante la adición de
ioduro de potasio en medio alcalino y sulfato manganoso, APHA 2005), se debe realizar
una fijación del oxígeno de la muestra de una botella por cada profundidad a tiempo cero
(botella testigo). Luego de pasado el período de incubación, se retiran las botellas del
cuerpo de agua y se procede a la determinación de la concentración de O2 disuelto en las
botellas provenientes de cada profundidad. La botella clara contiene el O2 inicial más el
O2 producido por fotosíntesis menos el O2 consumido en la respiración. La botella oscura
contiene la diferencia entre el O2 inicial y el O2 respirado en tanto que la botella testigo
contiene el O2 inicial. Por diferencia entre las concentraciones de oxígeno
correspondientes a las distintas botellas puede obtenerse una estimación de las tasas de
fotosíntesis bruta (O2 botella clara – O2 botella oscura) y neta (O2 botella clara – O2
botella testigo) (Dokulil 1984).
El método del intercambio de oxígeno tiene la principal ventaja de permitir estimar
la producción primaria bruta y neta y la respiración de manera directa. Por otra parte, un
inconveniente que tiene esta técnica es que la respiración que se mide en la botella oscura
no es sólo de las algas sino de todos los componentes del plancton y asume además que la
respiración en la botella clara es equivalente a la que ocurre en la botella oscura. Este
supuesto no siempre es válido ya que en la oscuridad puede existir proliferación
bacteriana cuya respiración afectaría una estimación precisa de la respiración algal (Wetzel
& Likens 2000). Dado que el oxígeno no posee un radioisótopo de vida media larga, la
determinación de la concentración de este elemento se realiza por métodos químicos o
con electrodos. El problema en general suele ser que los cambios en las concentraciones
de O2 son pequeños y la concentración previa suele ser grande, de modo que se requieren
largos períodos de incubación; además es muy difícil obtener mediciones precisas sobre
todo si la biomasa de los organismos no es muy alta. Sin embargo, cuanto más sensible
sea el método que se utiliza para medir los cambios en la concentración de O2, mejores

8
estimaciones se pueden obtener con esta técnica (Holm-Hansen & Helbling 1995). Una
alternativa para la determinación de las tasas de intercambio de O2 es el empleo de 18O2,
isótopo estable del oxígeno. El 18O2 puede ser inyectado en las botellas o cámaras de
incubación y su abundancia puede ser determinada con un espectrómetro de masa. Si bien
con este método se obtienen buenas estimaciones, su empleo es tedioso y costoso, por lo
que no ha sido ampliamente utilizado para determinaciones de fotosíntesis en la
naturaleza (Falkowski & Raven 2007).
El método del oxígeno también se utiliza en el caso de las comunidades sésiles
como el perifiton, con algunas modificaciones metodológicas dependiendo si se trata de
comunidades sobre sustratos naturales o artificiales. En todos estos casos, suelen
emplearse clausuras que minimizan las fuentes extra de O2 como la relacionada a la
mezcla física del agua en la capa límite de la comunidad (Falkowski & Raven 2007).
La técnica de incorporación de 14C es más sensible que la de intercambio de oxígeno y es
por lejos la más utilizada para la estimación de las tasas fotosintéticas en comunidades
naturales (Falkowski & Raven 2007). Steeman Nielsen (1952) introdujo el uso de 14C para
estimar producción orgánica fitoplanctónica en el mar y desde entonces, la técnica ha sido
examinada y perfeccionada en repetidas oportunidades (Legendre et al. 1983, Peterson
1980, Lean & Burnison 1979) y adaptada para la estimación de la productividad de otras
comunidades microbianas, como por ejemplo el epipelon y el epifiton (Vadeboncoeur &
Lodge 1998, Wetzel 1965). Desde la introducción de la técnica, pocos temas limnológicos
han recibido tanta atención y esfuerzos empíricos y desarrollo de modelos matemáticos
como el tema de la producción primaria del fitoplancton (Eilers & Peeters 1988, Fee
1984, Jassby & Platt 1976, entre otros).
El método asume que la fijación y reducción del 14CO2 puede igualarse a la tasa de
utilización de 12CO2 excepto por un factor de discriminación isotópica. El factor asume
que el 14C, que es más pesado que el 12C, es incorporado por las células un 5 o 6 % más
lentamente que el 12C (Falkowski & Raven 2007, Holm-Hansen & Helbling 1995). Por
otra parte, dado que el período de semidesintegración del 14C es de 5200 años, el
decaimiento del isótopo durante las incubaciones no es motivo de preocupación y su
actividad puede determinarse fácilmente en un contador de centelleo líquido.
Una de las desventajas de este método, además de tener en cuenta las medidas de
precaución que involucra el trabajo con sustancias radiactivas, es que la botella oscura no

9
representa la respiración sino la incorporación de carbono marcado por procesos no
fotosintéticos (Holm-Hansen & Helbling 1995). Por este motivo, aún existe debate acerca
de la estimación que proporciona este método en cuanto a si se trata de producción
primaria neta o bruta (Wetzel & Likens 2000). Dependiendo de la extensión del período
de incubación y la tasa de crecimiento de las células, el método brinda una estimación
intermedia entre la fotosíntesis bruta y neta. Si las incubaciones se llevan a cabo durante
períodos cortos de tiempo (2-4 hs), el método da una buena aproximación a la tasa de
fotosíntesis bruta. A medida que se prolongan los tiempos de incubación, una fracción
mayor del carbono marcado es respirado y la tasa de incorporación del 14C se aproxima a
la fotosíntesis neta (Falkowski & Raven 2007).
Existen otros métodos que permiten para estimar producción primaria de
comunidades acuáticas mediante el empleo de sensores remotos, fotos satelitales,
fluorescencia natural, etc. También se han desarrollado instrumentos ópticos que detectan
la fluorescencia emitida por el fotosistema II luego de haber sido estimulado con luz
artificial (ej. fluorómetros PAM, FRRF y PPF). Estos fluorómetros permiten estimar el
máximo rendimiento fotosintético (Sakshaug et al. 1997) y tienen la ventaja que no
requieren incubación ni filtración de las muestras ya que no son manipulativos.
Independientemente del método que se emplee para la determinación de la tasa
fotosintética de las comunidades microbianas acuáticas, se utilizan en general dos formas
principales de incubar las muestras. Una de ellas se basa en la incubación de las muestras a
la misma profundidad o en el mismo sitio de donde fueron extraídas, es decir in situ. La
otra forma es mediante la utilización de baños de agua con control de temperatura,
utilizando mallas de atenuación neutra para simular distintas atenuaciones de la radiación
incidente (Helbling & Holm-Hansen 1995). Si bien con la incubación in situ se obtienen
datos más realistas, no siempre es logísticamente posible su realización.
Curvas de Fotosíntesis-Irradiancia (P-I)
La relación no lineal existente entre la tasa de fotosíntesis y la irradiancia suele ser
representada a través de curvas P-I (del inglés Photosynthesis-Irradiance). Éstas pueden
ser utilizadas para evaluar la respuesta ecofisiológica de los organismos ante la irradiancia
y también para predecir las tasas de fotosíntesis in situ (Hill 1996). Metodológicamente las
curvas se pueden realizar siguiendo tanto la evolución del oxígeno como la asimilación de

10
carbono. En las curvas P-I se pueden diferenciar tres zonas: una región de limitación
lumínica, otra de saturación y una tercera de fotoinhibición (Fig. 1) (Falkowski & Raven
2007).
En la zona de limitación lumínica, la fotosíntesis aumenta linealmente con la
irradiancia a una tasa que es igual al parámetro プ o eficiencia fotosintética, donde la
absorción lumínica determina la tasa de tranporte de electrones desde el H2O al CO2. El
parámetro プ suele llevar un superíndice B cuando está relativizado a la biomasa
fotosintética (プB). Idealmente no lleva unidades de tiempo dado que no es una tasa, sino
que está relacionada con el máximo rendimiento lumínico de la fotosíntesis a través del
proceso de absorción, y depende entonces de la composición espectral de la irradiancia
(Falkowski & Raven 2007). A lo largo de esta tesis se emplearán las unidades: µg C m2 s
(µg Chl a h µmol fotones)-1 para el parámetro プ, por ser las utilizadas más frecuentemente
en la literatura, en lugar de µg C m2 (µg Chl a µmol fotones)-1.
En la zona de saturación lumínica, con mayores incrementos en la intensidad
lumínica, la tasa fotosintética comienza a tener una respuesta no lineal y aproximarse a un
nivel de saturación llamado Pmáx, número de asimilación ó tasa fotosintética
máxima. En esta región la tasa de absorción de fotones excede la tasa de transporte
electrónico desde el H2O al CO2. Pmáx incorpora unidades de tiempo en sus
dimensiones (ej. µg C (µg Chl a h)-1) y es independiente de la absorción óptica o espectral
del aparato fotosintético (Falkowski & Raven 2007). La intersección entre Pmáx y プ
define un punto en el eje de las irradiancias llamado irradiancia correspondiente al
inicio de la saturación fotosintética, Ik, cuyas unidades son, por ejemplo µmol fotones
m-2 s-1. Puede calcularse como Ik = Pmáx / プ (Talling 1957), de modo que depende de la
longitud de onda de la radiación absorbida y es independiente de las unidades en que se
expresen Pmáx y プ, siempre y cuando ambos parámetros estén normalizados por la
misma variable (ej. clorofila a) (Falkowski & Raven op. cit.). El parámetro Ik informa sobre
el estado de fotoaclimatación de la comunidad, ya que bajos valores de Ik indican un uso
ineficiente de altas irradiancias y viceversa (Henley 1993).
Mayores aumentos en los niveles de irradiancia pueden llevar a una reducción en la
tasa fotosintética. Esta reducción, que depende tanto de la intensidad de la irradiancia y
del tiempo de la exposición, se llama fotoinhibición y se suele representar mediante el
parámetro ベ, que caracteriza la fotoinhibición por altas irradiancias y cuyas unidades
son las mismas que las correspondientes al parámetro プ (Falkowski & Raven 2007). En el
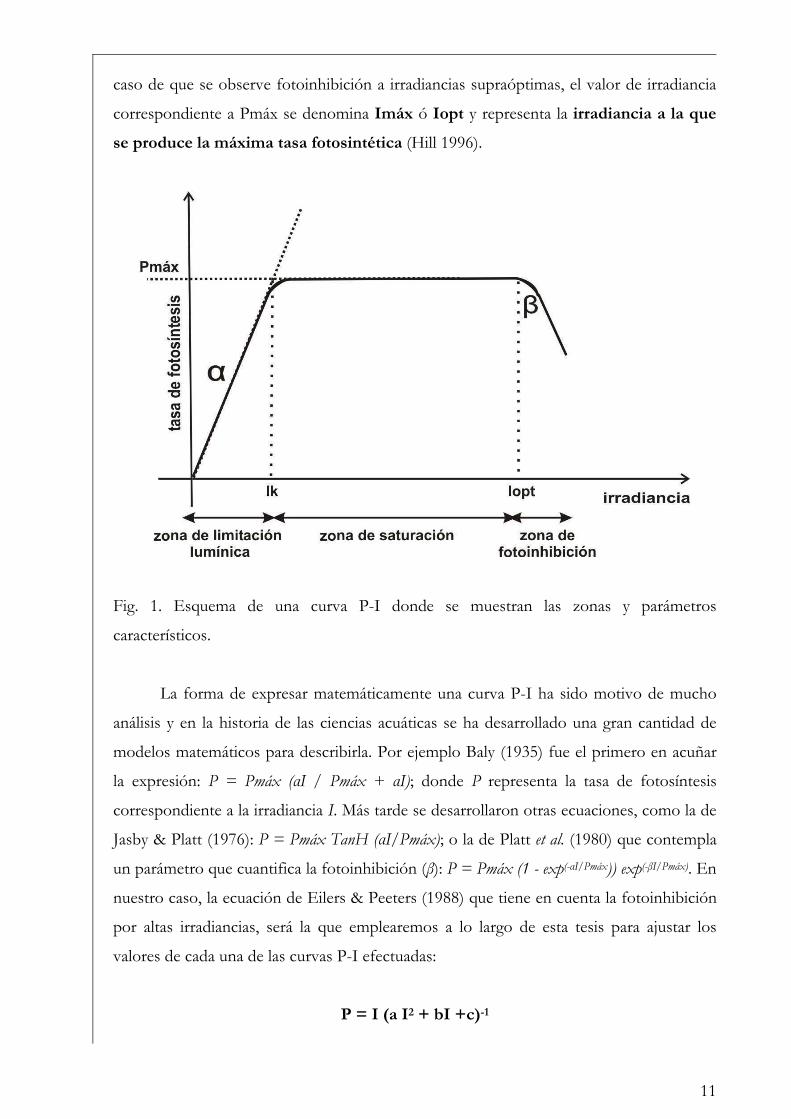
11
caso de que se observe fotoinhibición a irradiancias supraóptimas, el valor de irradiancia
correspondiente a Pmáx se denomina Imáx ó Iopt y representa la irradiancia a la que
se produce la máxima tasa fotosintética (Hill 1996).
Fig. 1. Esquema de una curva P-I donde se muestran las zonas y parámetros
característicos.
La forma de expresar matemáticamente una curva P-I ha sido motivo de mucho
análisis y en la historia de las ciencias acuáticas se ha desarrollado una gran cantidad de
modelos matemáticos para describirla. Por ejemplo Baly (1935) fue el primero en acuñar
la expresión: P = Pmáx (プI / Pmáx + プI); donde P representa la tasa de fotosíntesis
correspondiente a la irradiancia I. Más tarde se desarrollaron otras ecuaciones, como la de
Jasby & Platt (1976): P = Pmáx TanH (プI/Pmáx); o la de Platt et al. (1980) que contempla
un parámetro que cuantifica la fotoinhibición (ベ): P = Pmáx (1 - exp(-プI/Pmáx)) exp(-ベI/Pmáx). En
nuestro caso, la ecuación de Eilers & Peeters (1988) que tiene en cuenta la fotoinhibición
por altas irradiancias, será la que emplearemos a lo largo de esta tesis para ajustar los
valores de cada una de las curvas P-I efectuadas:
P = I (a I2 + bI +c)-1
12
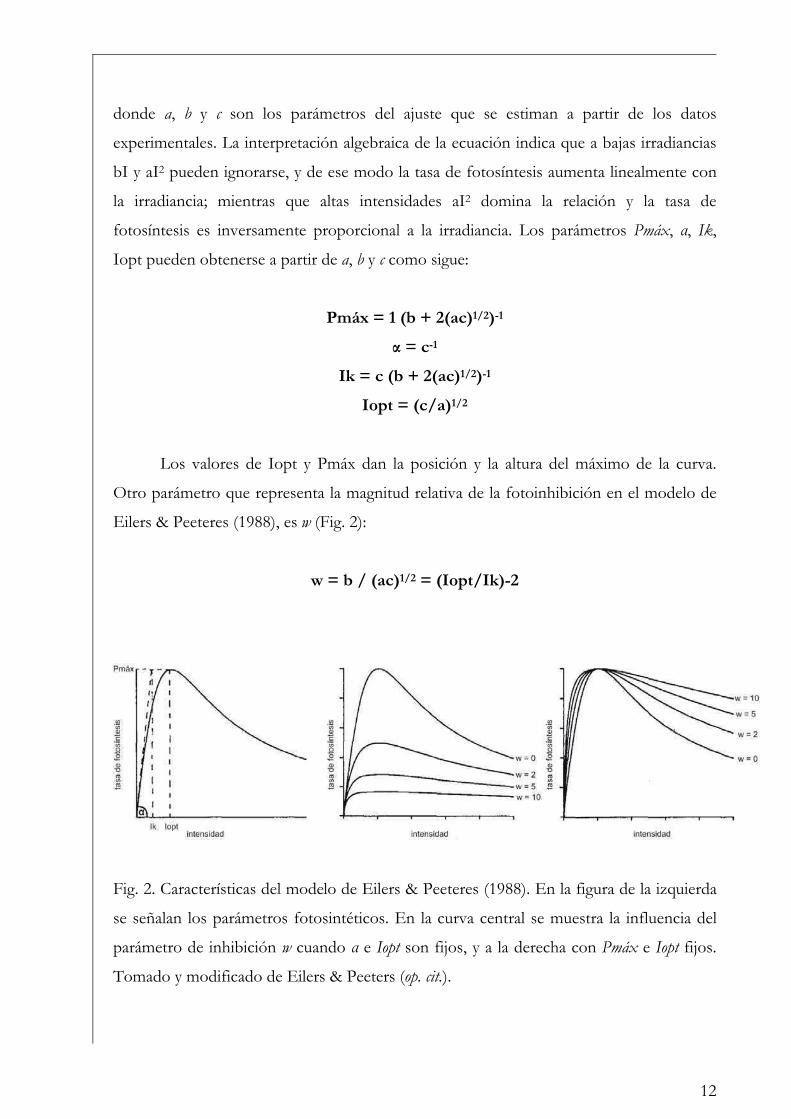
donde a, b y c son los parámetros del ajuste que se estiman a partir de los datos
experimentales. La interpretación algebraica de la ecuación indica que a bajas irradiancias
bI y aI2 pueden ignorarse, y de ese modo la tasa de fotosíntesis aumenta linealmente con
la irradiancia; mientras que altas intensidades aI2 domina la relación y la tasa de
fotosíntesis es inversamente proporcional a la irradiancia. Los parámetros Pmáx, プ, Ik,
Iopt pueden obtenerse a partir de a, b y c como sigue:
Pmáx = 1 (b + 2(ac)1/2)-1
プ = c-1
Ik = c (b + 2(ac)1/2)-1
Iopt = (c/a)1/2
Los valores de Iopt y Pmáx dan la posición y la altura del máximo de la curva.
Otro parámetro que representa la magnitud relativa de la fotoinhibición en el modelo de
Eilers & Peeteres (1988), es w (Fig. 2):
w = b / (ac)1/2 = (Iopt/Ik)-2
Fig. 2. Características del modelo de Eilers & Peeteres (1988). En la figura de la izquierda
se señalan los parámetros fotosintéticos. En la curva central se muestra la influencia del
parámetro de inhibición w cuando プ e Iopt son fijos, y a la derecha con Pmáx e Iopt fijos.
Tomado y modificado de Eilers & Peeters (op. cit.).

13
Producción integrada en la columna de agua (PA)
Cuando se desea comparar la productividad fitoplanctónica de distintos cuerpos de agua
se tiene en cuenta una magnitud muy útil que es la tasa de fotosíntesis por unidad de
área, PA, ó tasa integrada de fotosíntesis por unidad de superficie. Se puede estimar
determinando in situ las tasas de fotosíntesis por unidad de volumen a distintas
profundidades y luego integrando estas producciones por planimetría (Wetzel & Likens
2000, Kirk 1994).
Por otro lado, existe gran cantidad de modelos que se emplean para calcular la
producción integrada del fitoplancton en la columna de agua, muchos de los cuales
brindan resultados equivalentes (Falkowski & Raven 2007). La mayoría se basa en la
estimación de la PA a partir de los parámetros de la curva P-I (principalmente Pmáx e Ik),
clorofila a del fitoplancton, datos de irradiancia y del coeficiente de atenuación vertical
(Kd) del agua (Kirk 1994, Fee 1984). Por ejemplo, Talling (1957) demostró la
dependencia de PA con la concentración de clorofila a, el Kd, la irradiancia incidente (I0) y
la irradiancia al inicio de la saturación fotosintética (Ik), que expresó en la siguiente
ecuación:
PA = Pmáx vol (ln (2I0/Ik)/Kd) (1)
donde Pmáx vol, es el número de asimilación (Pmáx) multiplicado por la concentración
de clorofila a, que representa la productividad máxima por unidad de volumen. En la Fig.
3 se encuentran esquematizadas la PA y Pmáx vol en una columna de agua imaginaria.
Mientras Pmáx vol representa la producción por unidad de volumen a una sola
profundidad, PA representa la producción de toda la columna de agua integrando un área
de 1 m2 de superficie proyectada en profundidad (Falkowski & Raven 2007).
La ecuación (1) propuesta por Talling (1957) así como todos los modelos
matemáticos disponibles, presenta limitaciones ya que tiene implícitos una serie de
supuestos: la columna de agua es isotérmica, el fitoplancton está constituido por una sola
especie homogéneamente distribuida y no se tiene en cuenta la fotoinhibición. Sin
embargo, este modelo ha sido empleado en un amplio rango de ecosistemas de agua dulce
(Kalff 2003, Krause-Jensen & Sand-Jensen 1998).

14
Suele ser muy útil integrar los valores de PA en el tiempo para poder estimar tasas
diarias y anuales de producción, que permiten la comparación entre distintos cuerpos de
agua (Wetzel & Likens 2000).
Fig. 3. Representación esquemática de la Pmáx vol a una sola profundidad, donde ocurre
la máxima producción (cilindro rojo) y la PA, integrada en la columna de agua
(paralelepípedo negro).

15
OBJETIVO GENERAL
El objetivo general de este trabajo de tesis fue estudiar la producción primaria del
fitoplancton y perifiton presentes en cuerpos de agua húmicos del humedal de la Reserva
Natural de Otamendi (RNO). Se analizaron los parámetros fotosintéticos en función de
las principales variables ambientales en investigaciones comparativas entre las
comunidades, en distintos sitios y con esquemas temporales de estudio.
Para lograr el objetivo general se encararon distintos estudios que se presentan a
continuación en sucesivos capítulos. En el capítulo I, y debido a la carencia de estudios
estructurales de la comunidad perifítica, fundamentales para el posterior estudio de su
producción primaria, se describió la composición taxonómica algal del perifiton sobre
sustratos naturales en los principales cuerpos de agua de la RNO. Se analizó la fracción
algal presente en los sustratos más abundantes y representativos del sistema,
estacionalmente y en función del nivel hidrométrico. Además, se exploraron los patrones
de diversidad a distintas escalas espaciales. Una vez descrita esta comunidad, en el capítulo
II, se procedió a analizar la fluctuación temporal de la estructura y productividad del
fitoplancton y del perifiton sobre sustratos artificiales en la Laguna Grande, principal
cuerpo de agua permanente de la RNO, y las posibles variables ambientales reguladoras.
Se compararon los parámetros fotosintéticos de ambas comunidades derivados de curvas
P-I y se analizó el posible aporte de estas comunidades a la productividad primaria
microbiana del cuerpo de agua. Con el objeto de extender el análisis a otros ecosistemas
acuáticos de humedal, pero de características limnológicas contrastadas, en el capítulo III
se comparó la productividad y fracciones de masa del perifiton en dos sistemas de la
RNO (Laguna Grande y ROL), que presentan en particular ambientes lumínicos muy
distintos. También se compararon los parámetros fotosintéticos del perifiton con los
obtenidos para el fitoplancton en los dos cuerpos de agua.
Considerando que la Laguna Grande es el cuerpo de agua permanente más grande
y representativo del humedal donde el fitoplancton fue la comunidad microbiana
dominante durante el período de estudio, se procedió a profundizar su estudio analizando
el aporte de las distintas fracciones de tamaño fitoplanctónicas a la productividad
primaria. En el capítulo IV, por consiguiente, se estudió el aporte de las fracciones de
tamaño planctónicas > 3 µm (nano y microplancton) y la comprendida entre 0,2 y 3 µm
(picoplancton) a la productividad primaria del fitoplancton durante un año. A fines

16
comparativos, este análisis se extendió a otros dos cuerpos de agua de la RNO con un
cronograma de muestreo estacional. Por último, teniendo en cuenta que la presencia de
macrófitas flotantes es una característica distintiva en los sistemas acuáticos del humedal,
nos propusimos analizar su influencia en la productividad del fitoplancton. Para ello y en
el capítulo V, figuran los resultados de una experiencia manipulativa de campo en donde
estudiamos el efecto de la atenuación de la luz producida por una cobertura completa de
macrófitas flotantes en la producción primaria del fitoplancton de la Laguna Grande.

17
ÁREA DE ESTUDIO

18
La Reserva Natural Otamendi
Todas las investigaciones desarrolladas en esta tesis se llevaron a cabo en cuerpos de agua
ubicados en la Reserva Natural Otamendi (RNO, Administración de Parques Nacionales),
de aproximadamente 3000 ha. Los terrenos de la RNO están ubicados al sur de la ciudad
de Campana, provincia de Buenos Aires (34° 10´ a 34° 17´ S y 58 ° 48´ a 58° 53´ O) (Fig.
4). Entre los objetivos de la creación de la RNO en 1990 se destacaron la protección de
los ambientes naturales cercanos a la Ciudad de Buenos Aires tales como selvas ribereñas
del Paraná de las Palmas, bajos asociados al delta del Paraná, barrancas naturales con
bosques de tala y zonas altas con pastizales pampeanos. Se sumó además la necesidad de
proteger al amenazado ciervo de los pantanos. Por otro lado, dada la gran cantidad de
especies de aves que frecuenta o habita la reserva (alrededor de 240) fue declarada Área
Importante para la Conservación de las Aves (AICA) (Haene et al. 2003). En 2008 la
reserva fue declarada sitio de interés internacional por la Convención Ramsar sobre los
humedales. La misma es un tratado intergubernamental que sirve de marco para la acción
nacional y la cooperación internacional en pro de la conservación y uso racional de los
humedales y sus recursos (http://www.ramsar.org/indexsp.htm).
Fig. 4. Mapa de la zona de estudio en la RNO. Se señalan con una flecha los sitios
mayormente estudiados en el marco de esta tesis.

19
La RNO se encuentra ubicada sobre la margen derecha del Río Paraná de Las
Palmas. El ciclo hidrológico presenta un patrón irregular debido a la influencia de las
mareas del Río de la Plata y las sudestadas. La reserva se ubica dentro de dos grandes
unidades geomorfológicas, correspondiendo al "Bajío Rivereño" al norte y a la depresión
del río Luján al sur (Bonfils 1962). El sector norte abarca tanto una zona de barranca
como una llanura baja que la separa del río Paraná de las Palmas debido a la presencia de
albardones costeros. En el sector sur existe una extensa llanura de relieve cóncavo con
albardones a lo largo del río Paraná de las Palmas y Luján. Exceptuando la parte de la
barranca, ambas zonas son anegables, conformando un humedal, y es allí donde además
se localizan numerosos bañados temporarios o semipermanentes, meandros abandonados
y dos lagunas: "Laguna Grande" y "El pescado" (Chichizola 1993) (Fig. 4).
El clima de la zona es de tipo templado-húmedo debido al efecto moderador del
Río de la Plata. Predominan los vientos del sudoeste, que alternan con los provenientes
del sudeste que se presentan con frecuencia e intensidad variables (Chichizola 1993).
Según los datos climatológicos suministrados por la Estación Experimental Agropecuaria
INTA Delta del Paraná, la temperatura media anual durante el período de estudio (2004-
2007) fue de alrededor de 17,3 °C. Enero fue el mes más cálido, con una temperatura
máxima de 37 °C y media de aproximadamente 29,3 °C. Julio fue el mes más frío, con
temperaturas mínimas de aproximadamente -3 °C y media de 6,5 °C. La precipitación
media para los tres años fue de 1200 mm.
Los suelos de la llanura aluvial tienen drenaje limitado y en algunas partes se
observa salinización y alcalinización de los suelos, influenciada por la poca profundidad a
la que se encuentran los sedimentos marinos querandinos (Fig. 5). Allí puede observarse
también la presencia de vegetación halófila: espartillar de Spartina densiflora (esparto),
pastizal de pelo de chancho (Distichlis spicata), pastizal de Sporobolus pyramidatus y abrojo
(Xanthium cavanillesii), cortaderal (Cortaderia selloana) y pajonal de serruchetas (Eryngium sp.)
(Chichizola 1993, Virdó 1993).
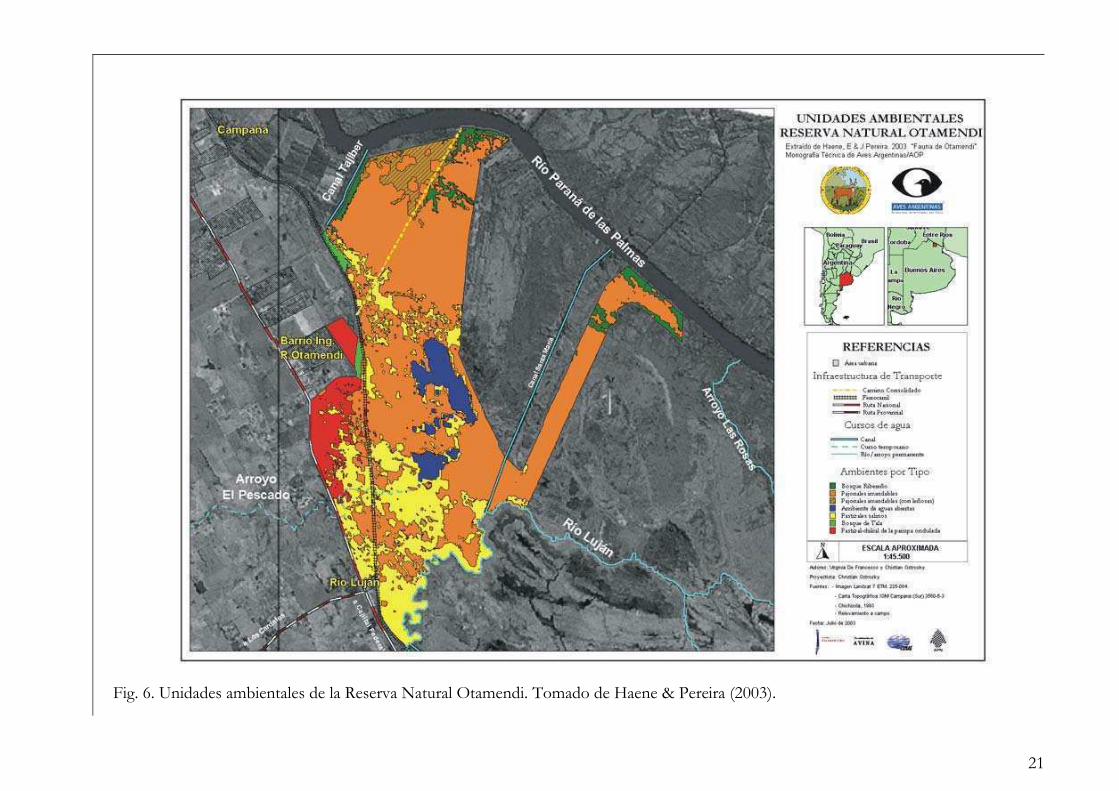
En cuanto a las comunidades vegetales, Haene et al. (2003) las agruparon en seis
unidades ambientales con distinto porcentaje de cobertura en la reserva: bosque ribereño
(3,3 %), pajonales inundables (63 %), ambientes de aguas abiertas (4,6 %), pastizales
salinos (22,3 %), bosque de tala (0,8 %) y pastizal-chilcal de la pampa ondulada (6 %) (Fig.
6).

20
Fig. 5. Vista de los afloramientos salinos (flecha) en la Reserva Natural Otamendi.
21
Fig. 6. Unidades ambientales de la Reserva Natural Otamendi. Tomado de Haene & Pereira (2003).

22
Se distingue en este humedal una gran cantidad de ambientes acuáticos de
diferentes características morfométricas y limnológicas (Izaguirre et al. 2001 b). La mayoría
de los mismos presenta un gran desarrollo de macrófitas acuáticas y palustres que se
presentan en diferentes asociaciones y coberturas. La macrófita más conspicua es
Schoenoplectus californicus (junco) junto con algunas especies asociadas como Typha latifolia
(totora), Sagitaria montividensis (saeta), Bacopa monnieri, Cleome titubans, Scirpus giganteus (paja
brava) y Juncus acutus entre otras. Además, tanto los meandros abandonados como la zona
litoral de la laguna presentan una cobertura fluctuante de macrófitas flotantes de diferente
porte tales como Lemma minima (lenteja de agua) y otras lemnáceas, Ricciocarpus natans,
Azolla filiculoides y en menor proporción Pistia stratiotes (repollito de agua), entre las más
abundantes.
Los estudios de esta tesis fueron llevados a cabo en varios cuerpos de agua de la
RNO: en dos meandros abandonados que llamaremos ROLs, (del inglés relictual oxbow
lakes; el mayor de ellos, ROL 1, 17 ha), en la laguna El Pescado (39 ha), el arroyo El
Pescado, y mayormente en la Laguna Grande (156 ha) (Fig. 4). Los ROLs tienden a ser
de forma alargada, poseen un contorno muy irregular y sus aguas son de color caramelo.
El mayor de ellos suele cubrirse con macrófitas flotantes gran parte del año, mientras que
el menor (ROL 2) posee un profuso desarrollo de vegetación emergente del tipo totora y
junco (Fig. 7). La laguna El Pescado está conectada a la Laguna Grande por medio del
arroyo El Pescado, de mayor profundidad que la laguna del mismo nombre (máximo ~60
cm). Las lagunas comparten algunas características, entre ellas la escasa profundidad
(máximo en El Pescado ~40 cm) y la coloración de las aguas, que es menos marcada que
en los ROLs. Las dos lagunas poseen un contorno bastante irregular (Fig. 7), que depende
en gran parte de la altura hidrométrica del sistema. La Laguna Grande posee una longitud
máxima de aproximadamente 3000 m, un ancho máximo de 1 km y una profundidad
máxima variable que raramente excede 1 m.
En el humedal el drenaje es limitado debido a la napa freática alta y fluctuante. Por
lo tanto, el área se mantiene periódicamente anegada por el agua de lluvia y esto provoca
condiciones edáficas reductoras (Chichizola 1993). Por otro lado, no existe habitualmente
conexión directa entre los cuerpos de agua permanentes del humedal, salvo en períodos
de fuertes lluvias. Tampoco existe un efecto evidente provocado por las crecidas o
desborde de los ríos Paraná de las Palmas y Luján, por lo que el principal origen del agua
de los sistemas es la precipitación pluvial.

23
En los cuerpos de agua permanentes (Laguna Grande y El Pescado) o
semipermanentes (ROLs) se encuentra gran cantidad de materia orgánica proveniente
principalmente del tejido macrofítico en descomposición, que les confiere características
húmicas (Kirk 1994). La posesión de altos niveles de sustancias húmicas y fósforo total,
permitirían clasificar estos cuerpos de agua como mixotróficos (Williamsom et al. 1999).
Fig. 7. Imágenes del área de estudio tomadas de Google Earth. En la imagen de la
izquierda se visualizan los ROLs mientras que en la imagen a la derecha éstos se
encontraban cubiertos de macrófitas flotantes (ROL 1) o emergentes (ROL 2).

24
CAPÍTULO I
ESTRUCTURA DE LA COMUNIDAD ALGAL PERIFÍTICA
SOBRE SUSTRATOS NATURALES EN EL HUMEDAL DE LA RNO

25
INTRODUCCIÓN
La interfase sólido-líquido de casi cualquier superficie, luego de ser sumergida,
seguramente será colonizada por bacterias y diatomeas de la columna de agua. Estos
organismos modificarán la superficie mediante la secreción de sustancias mucilaginosas,
creando una matriz donde se fijarán partículas orgánicas e inorgánicas y convivirán
bacterias, algas y otros organismos acuáticos (Liboriussen 2003). La terminología que
define a la comunidad microbiana adherida a un sustrato es amplia y ha cambiado a lo
largo del tiempo. Términos tales como biofilms, aufwuchs (del alemán: "que crece
sobre"), perifiton y bentos se han utilizado como sinónimos y en ocasiones han recibido
definiciones más precisas. El término que se presenta más citado en la literatura
actualmente parece ser el de perifiton. Etimológicamente hablando, la palabra perifiton
proviene del griego ┇ポ┈ί, que significa cerca, alrededor y ┍┌┋ό, cuyo significado es planta.
En el sentido biológico Wetzel (1983) define al perifiton como a "la compleja comunidad
microscópica de biota (algas, bacterias, hongos, animales y detritos orgánicos e
inorgánicos) adheridos a un sustrato sumergido natural o artificial, tanto vivo como
muerto".
En relación al perifiton existe además un amplio espectro de terminologías con las
que se especifica el tipo de sustrato adonde se adhiere la comunidad. Por ejemplo,
epifiton se refiere a la comunidad cuyo sustrato es una planta, epixylon se denomina a la
comunidad adherida a la madera, epiliton se utiliza para sustrato rocoso, episammon si es
arena, epipelon cuando se trata de sedimento, epizoon para animales, etc. (Wetzel 1983).
Por otro lado, puede ocurrir que en particular las algas del perifiton, se desprendan del
sustrato, ya sea porque una parte del ciclo vital comprende estadios sésiles y otras no, o
porque fue desprendida mecánicamente por efecto de la turbulencia. Cuando esto ocurre,
se les denomina ticoplancton o meroplancton. Éste último término se emplea en
general para denominar a las algas pertenecientes al plancton, pero que pasan una parte de
sus ciclos vitales adheridas a un sustrato (Kalff 2003).
Wetzel (2005) reconoce tres importantes características de la comunidad perifítica
dentro de los ecosistemas acuáticos. En primer lugar, la comunidad perifítica de la mayor
parte de los cuerpos de agua superficiales está compuesta de ensambles de algas
autotróficas, cianobacterias, bacterias heterotróficas, hongos, virus y protistas. En
segundo lugar, el intercambio de gases y nutrientes de la comunidad perifítica con el

26
medio ocurren a velocidades más lentas de las que ocurren en el agua circundante. De esta
manera, el metabolismo, crecimiento y productividad de la comunidad dependen en gran
parte del reciclado interno y de la conservación de los recursos capturados externamente,
resultando así en una alta eficiencia de utilización y retención de los mismos. Por último,
la mayor parte de la materia orgánica producida por los productores primarios es reciclada
dentro de la misma comunidad. El destino de la materia orgánica viva o muerta que no es
utilizada o respirada es complejo. En este sentido, Wetzel (2005) señala que la proporción
de la energía química y de los nutrientes contenidos en la comunidad perifítica que pasa a
los niveles tróficos superiores es materia de análisis y depende en gran parte del sustrato
de soporte.
En particular, la dependencia del epifiton con respecto al sustrato macrofítico debe
ser evaluada teniendo en cuenta las condiciones del hábitat, como por ejemplo la edad del
sustrato, el momento del año en que el sustrato fue colonizado, el tiempo que ha estado
disponible para la colonización y su posición física en el cuerpo de agua (Vermaat 2005).
La interacción con el sustrato parece ser más importante cuando la disponibilidad de
nutrientes en la columna de agua es menor (Eminson & Moss 1980), ya que algunos
sustratos pueden proveer de nutrientes a las algas epífitas (Cattaneo & Kalff 1979). A su
vez, la edad de la planta hospedadora condiciona su capacidad de liberar nutrientes,
disminuyendo ésta en tejidos más jóvenes (Burkholder 1996). Algunos nutrientes como
amonio, fosfato y azúcares simples son liberados por los tejidos vegetales durante su
senescencia en las primeras etapas de la descomposición (Wetzel 1981). El proceso de
liberación de nutrientes por parte de la planta puede ser impulsado por la ruptura de la
cutícula de la macrófita por la acción de bacterias celulíticas o bien de invertebrados que
dañan el tejido mientras consumen las algas adheridas a él (Burkholder op. cit.).
La presencia de epífitos puede tener efectos negativos en el crecimiento de la
planta hospedadora, tales como la reducción en la disponibilidad de luz, pueden interferir
en la captación de nutrientes y causar el desprendimiento de las hojas de la planta debido
a su peso (Vermaat 2005). Por otra parte, la presencia de epífitos podría tener también
efectos positivos sobre las macrófitas, como por ejemplo la atenuación de la radiación
ultravioleta antes que llegue a la superficie de la planta, actuando de esta manera como un
filtro (Brandt & Koch 2003). Las plantas a su vez, pueden liberar sustancias alelopáticas
(alcaloides, fenoles, taninos) que inhiben el crecimiento algal del perifiton (Anthoni et al.
1980). Finalmente, el resultado de la interacción planta-epífito dependerá de la estación

27
del año, de la disponibilidad de nutrientes en la columna de agua, de las condiciones
físicas y edad de la planta soporte, entre otros factores (Burkholder 1996).
Existen pocos trabajos en los que se describe la composición algal adherida sobre
sustratos macrofíticos en Argentina, entre los cuales pueden mencionarse por ejemplo los
trabajos de Tell (1979, 1977, 1973). El primero de ellos (Tell 1979) se focalizó en la flora
de algas clorofíceas de ambientes de Tierra del Fuego. En ese trabajo se propuso un
índice para estimar la superficie de fijación de las algas epifíticas al sustrato. Los otros dos
trabajos se llevaron a cabo en lagunas de la provincia de Buenos Aires. En Tell (1977) se
describió la comunidad algal epifítica sobre Ricciocarpus natans de la laguna Los Padres y en
Tell (1973) se estudió la composición de cianobacterias sobre macrófitas sumergidas de
las lagunas Chascomús, El Burro, Yalca y Vitel. Más tarde, Pozzobon & Tell (1995)
estudiaron la estructura y dinámica temporal de la comunidad perifítica sobre la hepática
Ricciocarpus natans en la laguna Los Padres. Tesolín & Tell (1996) describieron la
comunidad algal perifítica sobre distintas macrófitas flotantes de una laguna del valle
aluvial del río Paraná en su tramo inferior. Por su parte, Gómez et al. (2003) analizaron la
estructura y arquitectura de las algas epífitas sobre junco (Schoenoplectus californucus) en el
Río de la Plata. Bauer et al. (2007) evaluaron la utilidad de la comunidad perifítica que
crece sobre junco (S. californucus) como indicador de la calidad del agua en el Río de la
Plata. Más recientemente, Cano et al. (2008) estudiaron el aporte de la comunidad epifítica
creciendo sobre junco (S. californucus), epipelon y fitoplancton a la biomasa fotosintética
algal de una laguna de la provincia de Buenos Aires (Lacombe). Analizaron el patrón
temporal de dominancia de cada comunidad en el marco conceptual de un modelo
propuesto por Goldsborough & Robinson (1996) para los posibles estados de un
humedal.
El estudio de los patrones de diversidad permite evaluar la estructura de las comunidades
en el espacio y el tiempo, posibilitando la comparación entre distintos ecosistemas y de un
mismo ecosistema a lo largo de un período (ej. estacional, evolutivo). Existen distintos
índices y medidas de diversidad cuyo empleo depende del nivel de análisis y tipo de
comparación deseados. La プ diversidad es una propiedad de la unidad espacial; algunos de
sus estimadores otorgan mayor peso a las especies raras (ej. riqueza específica, índice de
Shannon-Wiener), mientras que otros a las más abundantes (ej. índice de Simpson). Por

28
otro lado, la ベ diversidad es esencialmente una medida de cuán diferentes o similares en
términos de especies son un grupo de comunidades a lo largo de un gradiente o entre
hábitats. La misma aproximación puede emplearse para examinar cambios de diversidad
en el tiempo (Magurran 2006). Cuanto menos especies compartan las diferentes
comunidades que se comparan, mayor será la diversidad ベ. La ペ diversidad tiene en cuenta
la totalidad de las especies encontradas en una región, mientras que la diversidad ポ se
refiere a una provincia biogeográfica Por último, la ボ diversidad representa el cambio en
composición específica (y abundancia) que ocurre entre unidades de ペ diversidad en un
sitio de ポ diversidad (Magurran op. cit.).
Se han llevado a cabo numerosos estudios en cuerpos de agua de la RNO, la mayor parte
ellos focalizados en distintos aspectos de la ecología de las comunidades planctónicas (de
Tezanos Pinto et al. 2007, O´Farrell et al. 2007, 2003; Rodríguez & Pizarro 2007, Sinistro et
al. 2007, 2006; Izaguirre et al. 2004, 2001 b), mientras que el perifiton no ha sido estudiado
aún en el humedal. En este capítulo se describirá la estructura de la comunidad perifítica
algal desarrollada sobre los sustratos naturales más conspicuos en cinco cuerpos de agua
del humedal de la RNO. También se relacionará la composición algal perifítica con las
variables ambientales a través del análisis multivariados y se analizarán los patrones de
diversidad en los diferentes sustratos a distintas escalas espaciales.
Se pondrán a prueba las siguientes hipótesis en relación a la comunidad perifítica
algal desarrollada sobre sustratos naturales (plantas flotantes y emergentes) en la RNO:
- La riqueza algal en los distintos sustratos será baja dadas las características ambientales
restrictivas del humedal (principalmente bajas irradiancias);
- La composición de la comunidad perifítica algal desarrollada sobre plantas palustres
(junco) mostrará predominio de algas resistentes a la desecación como resultado de su
adaptación a las fluctuaciones hidrométricas del sistema;
- Las algas presentes en los distintos tipos de sustrato mostrarán gran proporción de algas
planctónicas y metafíticas considerando que los cuerpos de agua tienen escasa
profundidad y las comunidades biológicas se encuentran estrechamente relacionadas.
29
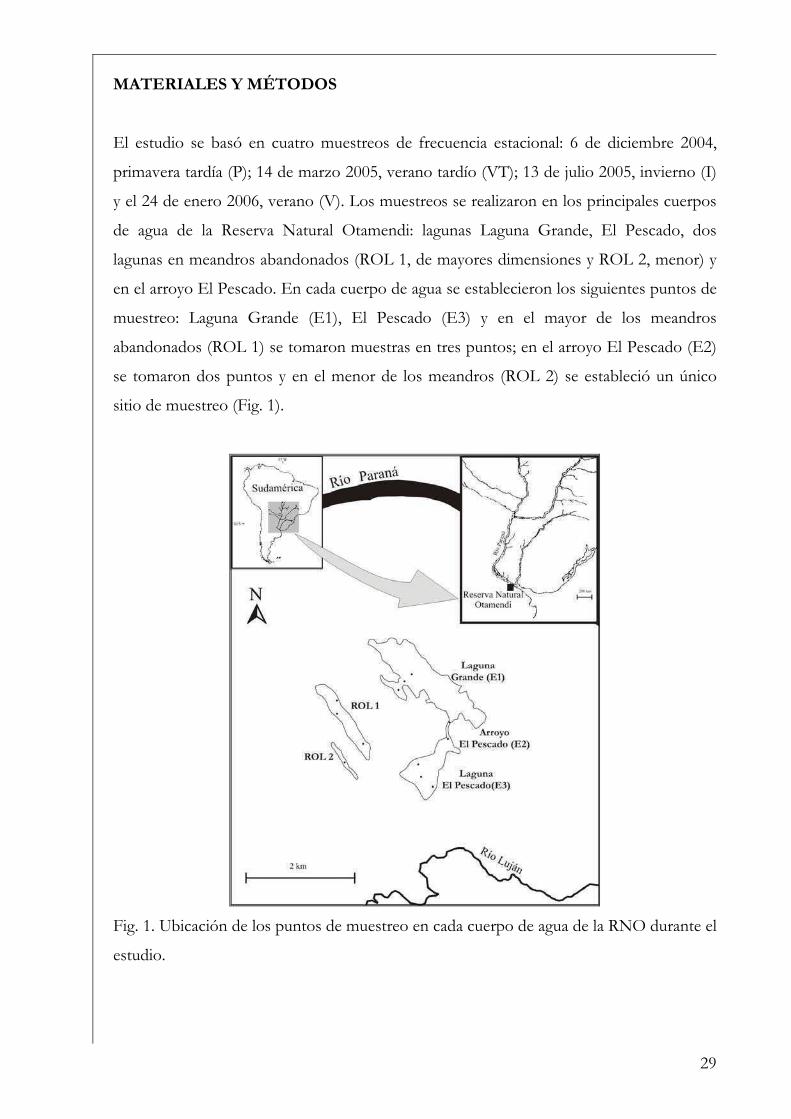
MATERIALES Y MÉTODOS
El estudio se basó en cuatro muestreos de frecuencia estacional: 6 de diciembre 2004,
primavera tardía (P); 14 de marzo 2005, verano tardío (VT); 13 de julio 2005, invierno (I)
y el 24 de enero 2006, verano (V). Los muestreos se realizaron en los principales cuerpos
de agua de la Reserva Natural Otamendi: lagunas Laguna Grande, El Pescado, dos
lagunas en meandros abandonados (ROL 1, de mayores dimensiones y ROL 2, menor) y
en el arroyo El Pescado. En cada cuerpo de agua se establecieron los siguientes puntos de
muestreo: Laguna Grande (E1), El Pescado (E3) y en el mayor de los meandros
abandonados (ROL 1) se tomaron muestras en tres puntos; en el arroyo El Pescado (E2)
se tomaron dos puntos y en el menor de los meandros (ROL 2) se estableció un único
sitio de muestreo (Fig. 1).
Fig. 1. Ubicación de los puntos de muestreo en cada cuerpo de agua de la RNO durante el
estudio.

30
Se registró la profundidad del agua en cada sitio de muestreo con una vara
graduada al milímetro y en un punto fijo de la Laguna Grande a partir de uno de los
pilotes de madera del muelle, que se utilizó como referencia para analizar las fluctuaciones
en el nivel hídrico del humedal. Se midieron in situ la temperatura, pH y conductividad
con sensor de campo HANNA HI 991301, y el oxígeno disuelto con sensor portátil
HANNA HI 9143. Los datos de precipitación pluvial fueron cedidos por la Estación
Experimental Agropecuaria Delta del Paraná (INTA). Se midió la radiación
fotosintéticamente activa subsuperficial e incidente en superficie con un radiómetro Li-
COR equipado con un sensor esférico Li-192 SA. Se determinó el porcentaje de
atenuación de luz incidente a través del cociente entre la radiación subacuática superficial
(I0) y la atmosférica, multiplicado por 100.
El contenido de ácidos húmicos se estimó a partir de la absorbancia a 254 nm de
agua de cada punto de muestreo previamente filtrada por filtros de fibra de vidrio
Millipore® de 0,7 µm de tamaño de poro (Kronberg 1999). La concentración de sólidos
en suspensión se determinó a partir de la filtración de un volumen conocido de agua a
través de filtros de fibra de vidrio Millipore® de 1 µm de tamaño de poro previamente
secados en estufa (60-70 °C). Los filtros con el material retenido fueron llevados
nuevamente a estufa hasta peso constante (APHA 2005).
Se determinaron las fracciones disueltas de los nutrientes luego de la filtración de la
muestra de agua a través de filtros de fibra de vidrio Millipore® de 0,7 µm de tamaño de
poro. Para el N-amonio se empleó el método del indofenol, para el fósforo disuelto (P-
disuelto) el del cloruro estañoso, para el N-nitrato + nitrito la reducción con cadmio y
para el sílice el método del molibdosilicato (APHA 2005). La concentración de nitrógeno
inorgánico disuelto (NID) se obtuvo como la suma de las fracciones de N-amonio + N-
nitrato + N-nitrito.
Para la determinación de la concentración de carbono orgánico disuelto se filtró
agua de cada sitio de muestreo a través de filtros de fibra de vidrio Millipore® de 0,7 µm
de tamaño de poro previamente calcinados a 500 °C. Las muestras se colocaron en
frascos oscuros luego de ser llevadas a pH 2 y se conservaron a 4 °C durante el traslado
para su posterior determinación en la Universidad de Québec (Montreal). Allí las muestras
fueron oxidadas con persulfato en caliente y luego determinadas en un analizador OI
1010 TOC.

31
La concentración de clorofila a fitoplanctónica se estimó a través de la filtración en
oscuridad de un volumen conocido de agua a través de filtros de fibra de vidrio Millipore®
de 0,7 µm de poro. El filtro se colocó envuelto en papel aluminio a -20 °C durante por lo
menos 24 hs. Luego se procedió a la extracción del pigmento, para lo cual se cortó el
filtro en trozos grandes que se colocaron en frascos opacos y se le agregó 8 mL de etanol
entre 60 y 70 °C. Los frascos se colocaron a 4 °C durante una noche, transcurrida la cual
se procedió a la lectura espectrofotométrica del sobrenadante a 665 nm y 750 nm, antes y
después de acidificar con HCl 0,1 N. Para el cálculo de las concentraciones se utilizó la
fórmula de Lorenzen (1967).
Con las variables ambientales obtenidas en los puntos de muestreo dentro de cada cuerpo
de agua se realizó un análisis de agrupamiento de los sitios en función de las variables
ambientales. A partir de los dendrogramas obtenidos (Fig. 2) se observó en general un
alto grado de similitud entre los puntos de un mismo cuerpo de agua. Este resultado nos
facilitó trabajar posteriormente con los promedios de las variables ambientales de cada
cuerpo de agua.
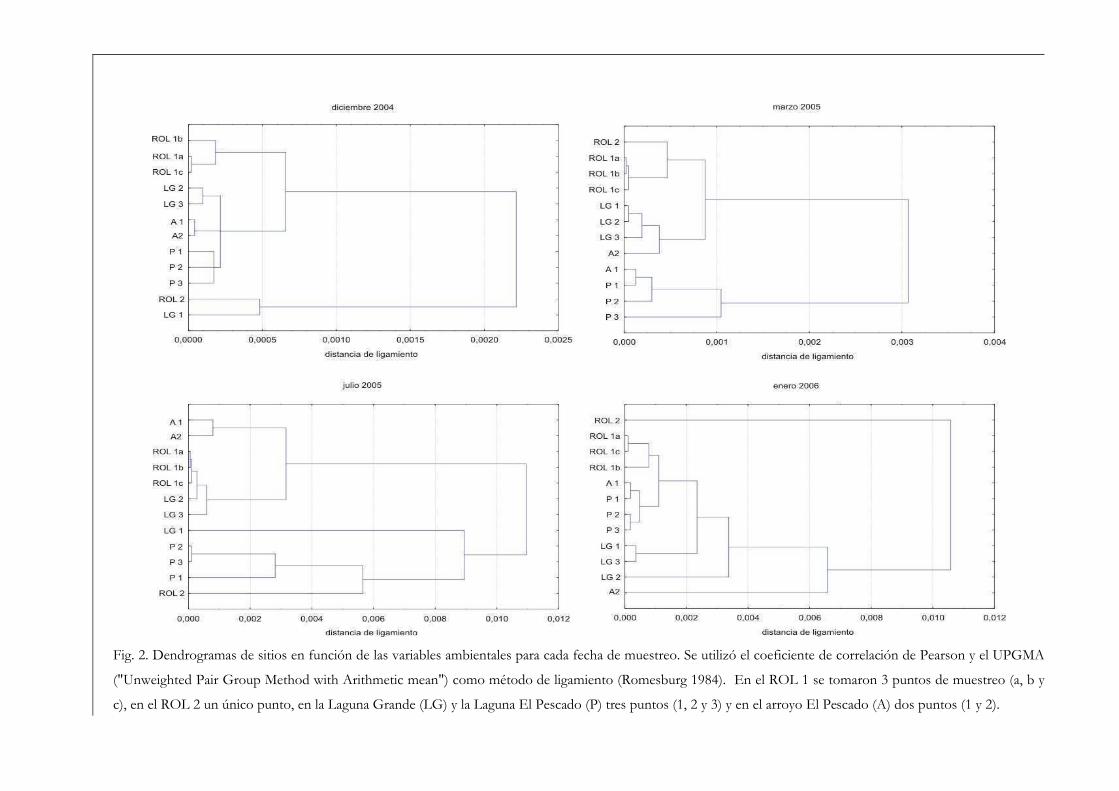
Fig. 2. Dendrogramas de sitios en función de las variables ambientales para cada fecha de muestreo. Se utilizó el coeficiente de correlación de Pearson y el UPGMA
("Unweighted Pair Group Method with Arithmetic mean") como método de ligamiento (Romesburg 1984). En el ROL 1 se tomaron 3 puntos de muestreo (a, b y
c), en el ROL 2 un único punto, en la Laguna Grande (LG) y la Laguna El Pescado (P) tres puntos (1, 2 y 3) y en el arroyo El Pescado (A) dos puntos (1 y 2).

33
Perifiton sobre sustratos naturales
Se tomaron muestras de las macrófitas presentes en los sitios señalados en la Fig. 1. Las
macrófitas más conspicuas en todos los sitios de muestreo fueron Schoenoplectus californicus
(Junco), Ricciocarpus natans y Pistia stratiotes (repollito de agua). En cada sitio de muestreo se
cortaron 5 trozos de junco de características fisonómicas similares de aproximadamente 10
cm de longitud (sólo se empleó para su observación la porción sumergida) y se recolectaron
por lo menos 10 individuos de Ricciocarpus natans y 3 de Pistia stratiotes en caso de estar
presentes. Las epífitas adheridas a la superficie de los trozos de junco y a las escamas (R.
natans) y raíces (P. stratiotes) de las plantas se observaron directamente bajo microscopio
óptico binocular Zeiss y los especímenes se dibujaron con cámara clara. En el caso de junco y
P. stratiotes el material fue raspado de su superficie (de las raíces en el caso de P. stratiotes) por
medio de un bisturí, y en el caso de R. natans se colocaron las escamas entre porta y
cubreobjetos para su observación.
Para la determinación taxonómica y del tipo de hábito de los especimenes
encontrados se siguió la siguiente bibliografía: para géneros Bourrelly (1970, 1972 y 1981),
para Cyanobacteria se utilizó Geitler (1932), Komárek & Anagnostidis (2005, 1999); para
Bacillariophyceae Lange-Bertalot (2001), Kramer & Lange-Bertalot (1986, 1988, 1991 a y
1991 b); para Chlorophyta Printz (1964), Ettl (1983) y Komárek & Fott (1983); para
Euglenophyta Huber-Pestalozzi (1955) y Tell & Conforti (1986) y para Xanthophyceae Ettl
(1977) y Pizarro (1995). Para la clasificación de entidades taxonómicas superiores se siguió el
esquema propuesto en van den Hoek et al. (1995). Los hábitos de los organismos se los
consideró de la siguiente manera: se consideró perifiton a todos aquéllos organismos
adheridos, independientemente del tipo de sustrato y de su ubicación en el cuerpo de agua;
fitoplancton a los organismos que viven en el seno del agua; ticoplancton a los organismos
accidentales en el plancton y metafiton a las algas principalmente filamentosas que pueden
desprenderse y formar matas macroscópicas.
Con el objeto de abarcar una mayor variabilidad dentro de un mismo cuerpo de agua
se unificaron los listados taxonómicos pertenecientes a los puntos de muestreo de cada lugar.
De esta manera, se construyó una matriz de presencia/ausencia con una sola columna por
ambiente.

34
Para estudiar la composición específica de los sitios de muestreo en relación a las variables
ambientales se realizó un análisis de correspondencia canónico (ACC) (ter Braak 1994, ter
Braak 1990, ter Braak 1987, ter Braak & Verdonschot 1995) a partir de las matrices de
presencia/ausencia de especies y variables ambientales. Para este análisis se seleccionaron
aquellos taxa con una constancia mayor a 10 en la matriz de todos los puntos de muestreo y
sustratos. Se realizó un ordenamiento para el sustrato Schoenoplectus californicus y otro para
Ricciocarpus natans, que mostraron la mayor frecuencia de aparición en los sitios de muestreo a
lo largo del período de estudio. Este análisis se realizó por separado ya que consideramos que
las variables ambientales deben afectar de forma diferente a los organismos presentes en
ambos tipos de sustrato por proveer cada uno de ellos microhábitats distintos. Se empleó una
matriz con las mismas 20 entidades taxonómicas en ambos casos, y los organismos que se
incluyeron en este análisis se encuentran identificados en la Tabla 3, donde también se señala
la abreviatura de cada especie empleada en el diagrama de ordenamiento. Antes de realizar el
análisis propiamente dicho se analizó la variable "inflation factor", los coeficientes de
correlación de las variables ambientales y se empleó una selección "forward" para detectar y
eliminar del análisis aquellas variables ambientales altamente correlacionadas entre sí. En
ambos casos se tuvieron en cuenta las siguientes variables ambientales: pH, conductividad,
temperatura, transmitancia lumínica, oxígeno disuelto, abs 254 nm, NID, P-disuelto, sólidos
en suspensión y concentración de clorofila a fitoplanctónica. Se evaluó la significancia del
primer eje canónico a través del test de Monte Carlo con 499 permutaciones (p < 0,05).
No se utilizó directamente la variable profundidad en el ACC para no emplear
promedios de las profundidades entre sitios de muestreo de un mismo cuerpo de agua. Esta
decisión se tomó teniendo en cuenta que la profundidad entre puntos de muestreo en el
mismo cuerpo de agua es muy variable debido a la irregularidad del fondo provocada por
elevadas cantidades de materia orgánica y restos de material vegetal. La variable
conductividad se empleó en el ACC como indicadora del nivel hidrométrico del sistema. La
conductividad estuvo relacionada de forma negativa con la profundidad mediante la
expresión: Log10conductividad = -5,12 10-3 profundidad +3,6 (R = 0,36; p = 0,013). Para
realizar esta regresión se tuvieron en cuenta todos los sitios de muestreo por separado (N =
46).

35
Para analizar los patrones de diversidad se calcularon la riqueza específica total del perifiton
de todos los sitios y fechas de muestreo (ペ diversidad) y la específica de cada cuerpo de agua
por estación del año (プ diversidad) a partir de la matriz de presencia-ausencia de especies. La
ベ diversidad (entre sitios) se estimó a partir del índice de Whittaker (ベW) (Magurran 2006):
ベW = S / プ
donde S es el número total de especies registradas en el sistema (ペ diversidad) y "プ" es el
número de especies de la muestra.
Con el objeto de comparar la composición específica entre pares de sustratos y entre
pares de sitios se empleó el índice de similitud de Jaccard (J) (Magurran 2006):
J = a (a + b +c)-1
donde a representa el número de especies presentes en ambos tipos de sustrato o sitios; b el
número de especies presentes sólo en uno de ellos y c el número de especies presentes sólo
en el otro sustrato o sitio. Este índice adquiere el valor 1 cuando todas las especies son
compartidas entre las comunidades que se están comparando y vale 0 cuando no se comparte
ninguna entidad.
RESULTADOS
Variables ambientales
Durante el período estudiado no se observó una cobertura completa de macrófitas flotantes
en los sitios de muestreo sino que ésta fue dispersa en el ROL 1 y principalmente restringida
a la zona litoral en las lagunas Grande y El Pescado. El ROL 2 se encontró cubierto en gran
parte de su superficie por vegetación emergente del tipo totora y junco. Las precipitaciones
acumuladas durante el mes previo al muestreo fueron de 66 mm en diciembre 2004, 45 mm
en marzo 2005, 108 mm en julio 2005 y 255 mm en enero 2006. El nivel del agua tomado en
un punto fijo del muelle de la Laguna Grande osciló entre 23 cm (diciembre 2004, aguas
bajas) y 49 cm (enero 2006, aguas altas), tomando valores intermedios de 35 y 34 cm en los
muestreos de marzo 2005 y julio 2005.
En la Tabla 1 se muestran los rangos de variación de las variables ambientales
registradas durante el estudio así como su promedio, desvío estándar y coeficiente de
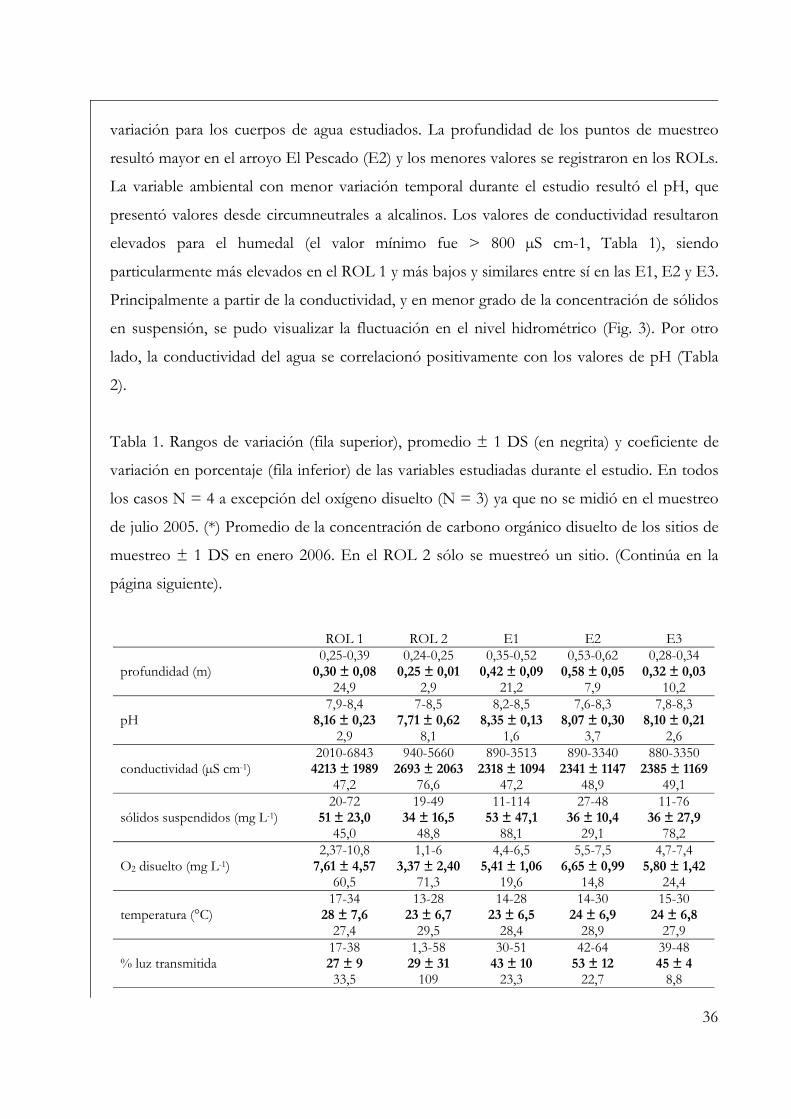
36
variación para los cuerpos de agua estudiados. La profundidad de los puntos de muestreo
resultó mayor en el arroyo El Pescado (E2) y los menores valores se registraron en los ROLs.
La variable ambiental con menor variación temporal durante el estudio resultó el pH, que
presentó valores desde circumneutrales a alcalinos. Los valores de conductividad resultaron
elevados para el humedal (el valor mínimo fue > 800 µS cm-1, Tabla 1), siendo
particularmente más elevados en el ROL 1 y más bajos y similares entre sí en las E1, E2 y E3.
Principalmente a partir de la conductividad, y en menor grado de la concentración de sólidos
en suspensión, se pudo visualizar la fluctuación en el nivel hidrométrico (Fig. 3). Por otro
lado, la conductividad del agua se correlacionó positivamente con los valores de pH (Tabla
2).
Tabla 1. Rangos de variación (fila superior), promedio ± 1 DS (en negrita) y coeficiente de
variación en porcentaje (fila inferior) de las variables estudiadas durante el estudio. En todos
los casos N = 4 a excepción del oxígeno disuelto (N = 3) ya que no se midió en el muestreo
de julio 2005. (*) Promedio de la concentración de carbono orgánico disuelto de los sitios de
muestreo ± 1 DS en enero 2006. En el ROL 2 sólo se muestreó un sitio. (Continúa en la
página siguiente).
ROL 1 ROL 2 E1 E2 E3
profundidad (m) 0,25-0,39
0,30 ± 0,08 24,9
0,24-0,25 0,25 ± 0,01
2,9
0,35-0,52 0,42 ± 0,09
21,2
0,53-0,62 0,58 ± 0,05
7,9
0,28-0,34 0,32 ± 0,03
10,2
pH 7,9-8,4
8,16 ± 0,23 2,9
7-8,5 7,71 ± 0,62
8,1
8,2-8,5 8,35 ± 0,13
1,6
7,6-8,3 8,07 ± 0,30
3,7
7,8-8,3 8,10 ± 0,21
2,6
conductividad (µS cm-1) 2010-6843
4213 ± 1989 47,2
940-5660 2693 ± 2063
76,6
890-3513 2318 ± 1094
47,2
890-3340 2341 ± 1147
48,9
880-3350 2385 ± 1169
49,1
sólidos suspendidos (mg L-1) 20-72
51 ± 23,0 45,0
19-49 34 ± 16,5
48,8
11-114 53 ± 47,1
88,1
27-48 36 ± 10,4
29,1
11-76 36 ± 27,9
78,2
O2 disuelto (mg L-1) 2,37-10,8
7,61 ± 4,57 60,5
1,1-6 3,37 ± 2,40
71,3
4,4-6,5 5,41 ± 1,06
19,6
5,5-7,5 6,65 ± 0,99
14,8
4,7-7,4 5,80 ± 1,42
24,4
temperatura (°C) 17-34
28 ± 7,6 27,4
13-28 23 ± 6,7
29,5
14-28 23 ± 6,5
28,4
14-30 24 ± 6,9
28,9
15-30 24 ± 6,8
27,9
% luz transmitida 17-38 27 ± 9 33,5
1,3-58 29 ± 31
109
30-51 43 ± 10
23,3
42-64 53 ± 12
22,7
39-48 45 ± 4
8,8
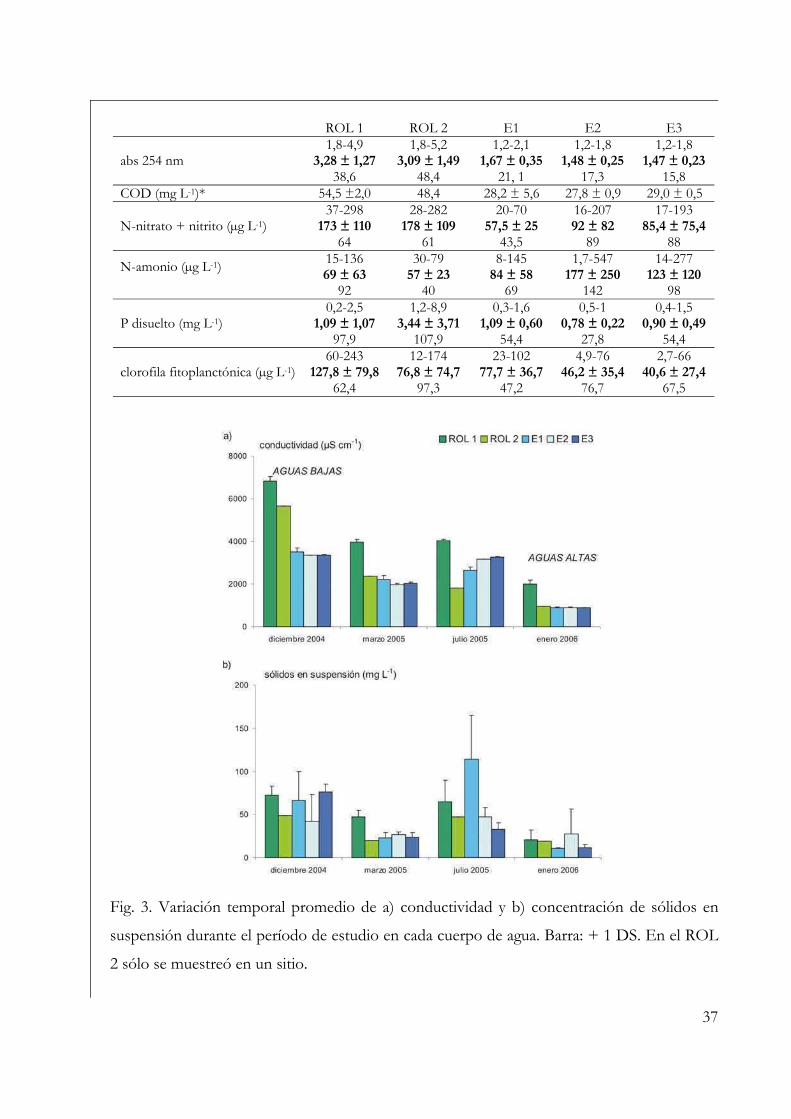
37
ROL 1 ROL 2 E1 E2 E3
abs 254 nm 1,8-4,9
3,28 ± 1,27 38,6
1,8-5,2 3,09 ± 1,49
48,4
1,2-2,1 1,67 ± 0,35
21, 1
1,2-1,8 1,48 ± 0,25
17,3
1,2-1,8 1,47 ± 0,23
15,8 COD (mg L-1)* 54,5 ±2,0 48,4 28,2 ± 5,6 27,8 ± 0,9 29,0 ± 0,5
N-nitrato + nitrito (µg L-1) 37-298
173 ± 110 64
28-282 178 ± 109
61
20-70 57,5 ± 25
43,5
16-207 92 ± 82
89
17-193 85,4 ± 75,4
88
N-amonio (µg L-1)
15-136 69 ± 63
92
30-79 57 ± 23
40
8-145 84 ± 58
69
1,7-547 177 ± 250
142
14-277 123 ± 120
98
P disuelto (mg L-1) 0,2-2,5
1,09 ± 1,07 97,9
1,2-8,9 3,44 ± 3,71
107,9
0,3-1,6 1,09 ± 0,60
54,4
0,5-1 0,78 ± 0,22
27,8
0,4-1,5 0,90 ± 0,49
54,4
clorofila fitoplanctónica (µg L-1) 60-243
127,8 ± 79,862,4
12-174 76,8 ± 74,7
97,3
23-102 77,7 ± 36,7
47,2
4,9-76 46,2 ± 35,4
76,7
2,7-66 40,6 ± 27,4
67,5
Fig. 3. Variación temporal promedio de a) conductividad y b) concentración de sólidos en
suspensión durante el período de estudio en cada cuerpo de agua. Barra: + 1 DS. En el ROL
2 sólo se muestreó en un sitio.
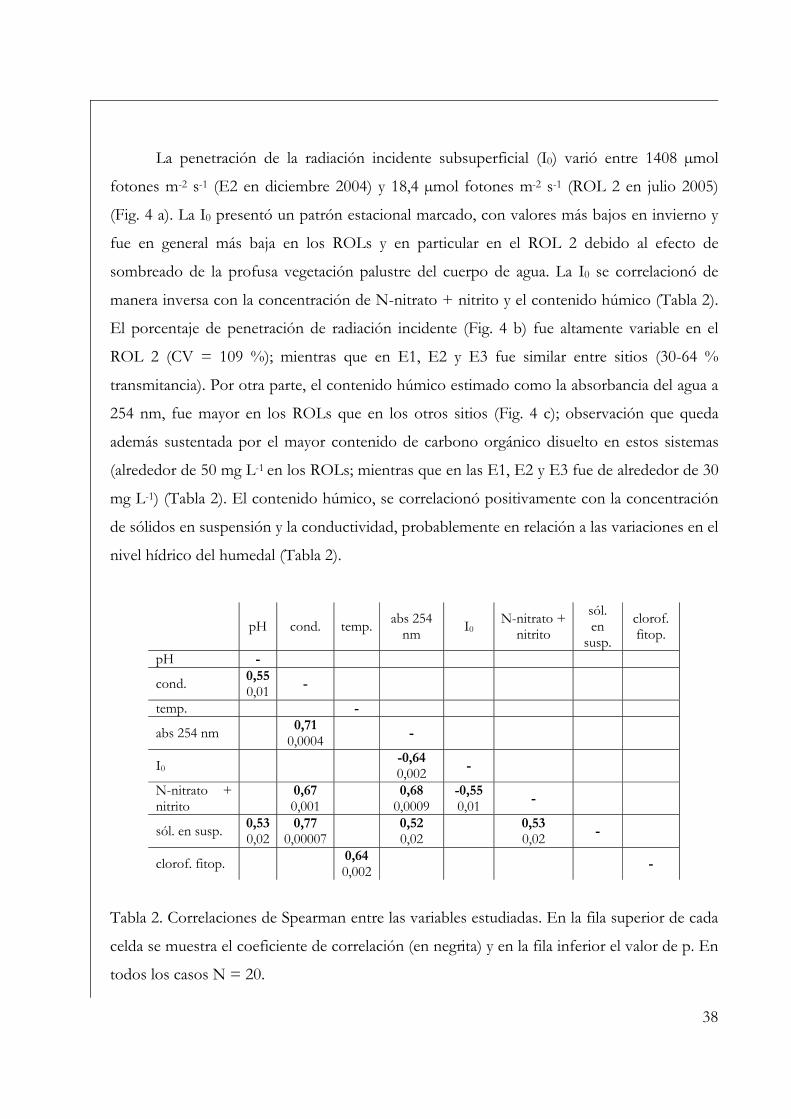
38
La penetración de la radiación incidente subsuperficial (I0) varió entre 1408 µmol
fotones m-2 s-1 (E2 en diciembre 2004) y 18,4 µmol fotones m-2 s-1 (ROL 2 en julio 2005)
(Fig. 4 a). La I0 presentó un patrón estacional marcado, con valores más bajos en invierno y
fue en general más baja en los ROLs y en particular en el ROL 2 debido al efecto de
sombreado de la profusa vegetación palustre del cuerpo de agua. La I0 se correlacionó de
manera inversa con la concentración de N-nitrato + nitrito y el contenido húmico (Tabla 2).
El porcentaje de penetración de radiación incidente (Fig. 4 b) fue altamente variable en el
ROL 2 (CV = 109 %); mientras que en E1, E2 y E3 fue similar entre sitios (30-64 %
transmitancia). Por otra parte, el contenido húmico estimado como la absorbancia del agua a
254 nm, fue mayor en los ROLs que en los otros sitios (Fig. 4 c); observación que queda
además sustentada por el mayor contenido de carbono orgánico disuelto en estos sistemas
(alrededor de 50 mg L-1 en los ROLs; mientras que en las E1, E2 y E3 fue de alrededor de 30
mg L-1) (Tabla 2). El contenido húmico, se correlacionó positivamente con la concentración
de sólidos en suspensión y la conductividad, probablemente en relación a las variaciones en el
nivel hídrico del humedal (Tabla 2).
pH cond. temp.abs 254
nm I0
N-nitrato + nitrito
sól. en
susp.
clorof. fitop.
pH -
cond. 0,55 0,01
-
temp. -
abs 254 nm 0,71
0,0004 -
I0 -0,64 0,002
-
N-nitrato + nitrito
0,67 0,001
0,68
0,0009 -0,55 0,01
-
sól. en susp. 0,53 0,02
0,77 0,00007
0,52 0,02
0,53 0,02
-
clorof. fitop. 0,64 0,002
-
Tabla 2. Correlaciones de Spearman entre las variables estudiadas. En la fila superior de cada
celda se muestra el coeficiente de correlación (en negrita) y en la fila inferior el valor de p. En
todos los casos N = 20.
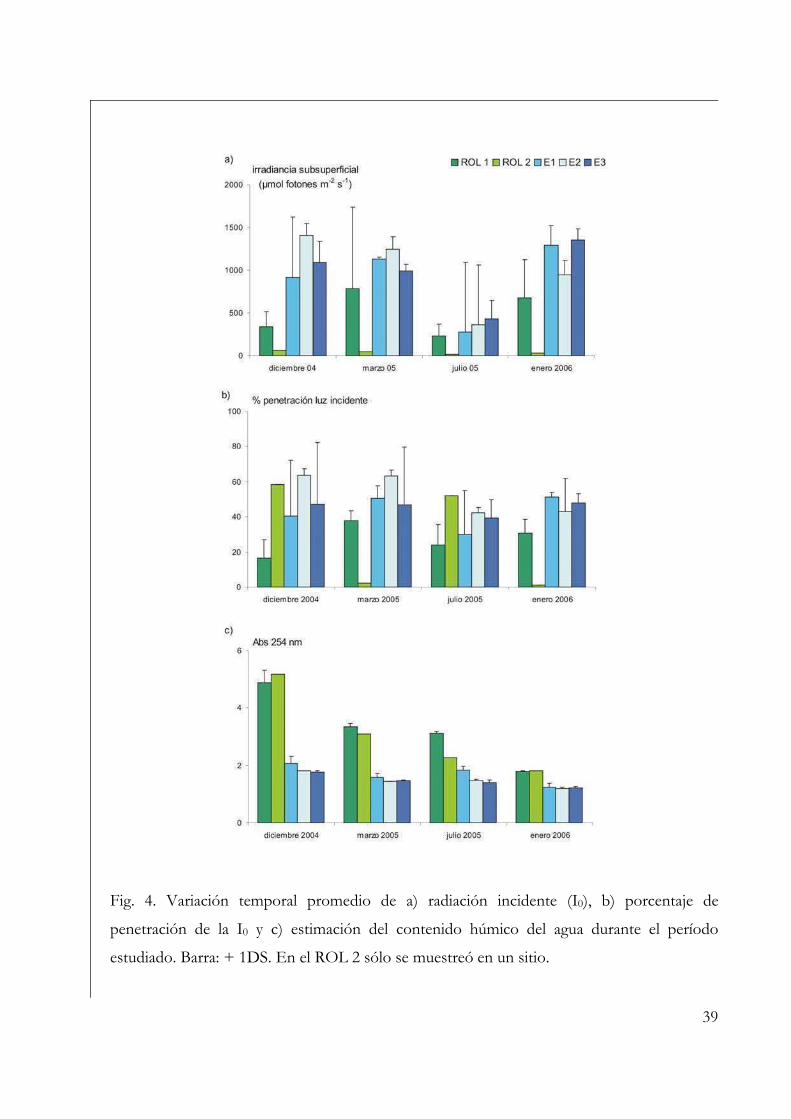
39
Fig. 4. Variación temporal promedio de a) radiación incidente (I0), b) porcentaje de
penetración de la I0 y c) estimación del contenido húmico del agua durante el período
estudiado. Barra: + 1DS. En el ROL 2 sólo se muestreó en un sitio.

40
La concentración de N-nitrato + nitrito tendió a ser más alta en los ROLs, alcanzando
un máximo de 300 µg L-1 en el ROL 1 en diciembre 2004 (aguas bajas) (Fig. 5 a). En enero
2006 (aguas altas) las concentraciones fueron más bajas y tendieron a homogeneizarse entre
los cuerpos de agua, siendo el valor mínimo observado de 16 µg L-1 en la E2. La
concentración de N-nitrato se correlacionó de manera positiva con la conductividad, la
absorbancia a 254 nm y la concentración de sólidos en suspensión (Tabla 2). El patrón de
fluctuación de las concentraciones de N-amonio fue más variable, observándose el valor
máximo y mínimo en la E2, siendo el primero de 550 µg L-1 en julio 2005 y el valor más bajo
de 1,7 µg L-1 en diciembre 2004 (Fig. 5 b). La concentración de NID para los cuerpos de
agua E1, E2 y E3 presentó mayores valores en invierno, mientras que en los ROLs en
diciembre 2004 se detectaron los valores más altos. En los ROLs, en general la concentración
de NID se encontró por encima de los valores considerados como limitantes para el máximo
crecimiento algal (100 µg L-1 sensu Reynolds 2006), mientras que en los demás cuerpos de
agua se presentaron valores por debajo de este límite en algunas oportunidades (Fig. 5 c).
En general, la concentración de P-disuelto tendió a ser más alta en el ROL 2 (valor
máximo = 9 mg L-1 en diciembre 2004) (Fig. 6 a). Los valores más bajos para este nutriente
se detectaron en invierno, a excepción del ROL 1, cuyo valor más bajo ocurrió en diciembre
2004 (0,2 mg L-1). En todas las oportunidades y sitios de muestreo las concentraciones de P-
disuelto se encontraron por encima de los valores considerados limitantes para el crecimiento
algal (5-10 µg L-1 sensu Reynolds 1984). Por último, la concentración de clorofila a
fitoplanctónica fue similar entre cuerpos de agua durante diciembre 2004 (aguas altas) (Fig. 6
b). EL valor más alto se encontró en el ROL 1 en marzo 2005 (243 µg L-1), mientras que las
menores concentraciones fueron en invierno, tal como sustenta la correlación encontrada
entre esta variable y la temperatura del agua (Tabla 2).
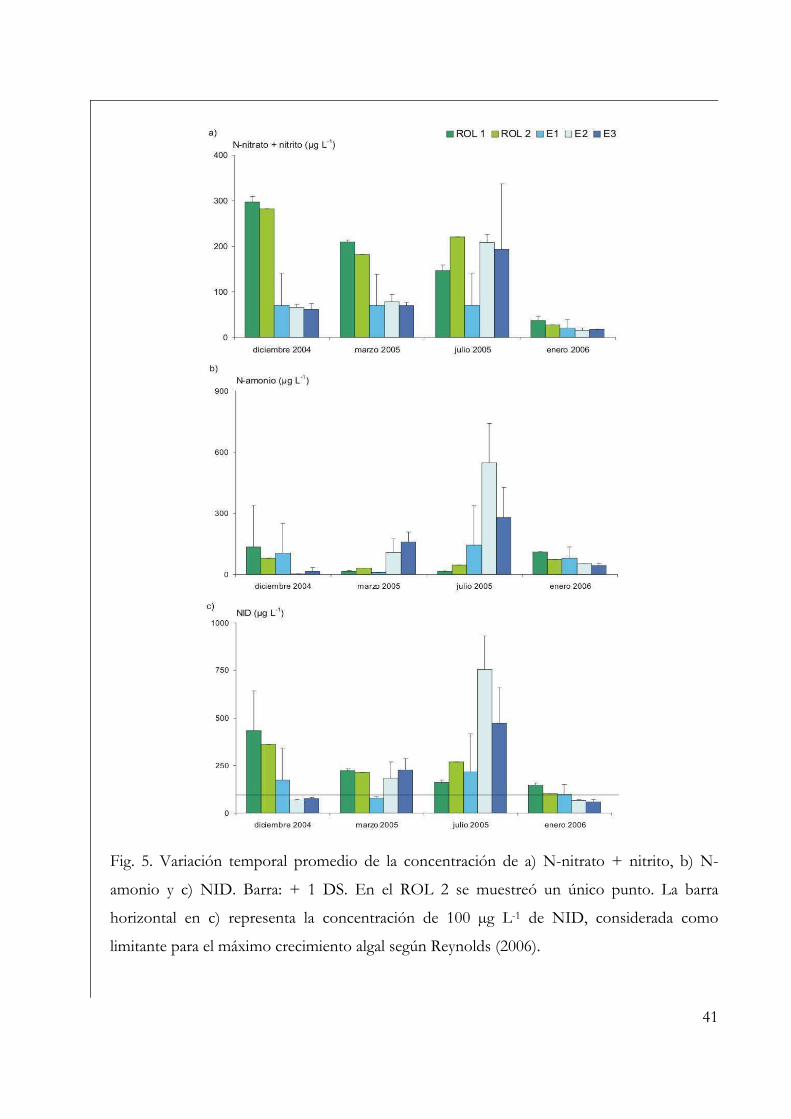
41
Fig. 5. Variación temporal promedio de la concentración de a) N-nitrato + nitrito, b) N-
amonio y c) NID. Barra: + 1 DS. En el ROL 2 se muestreó un único punto. La barra
horizontal en c) representa la concentración de 100 µg L-1 de NID, considerada como
limitante para el máximo crecimiento algal según Reynolds (2006).
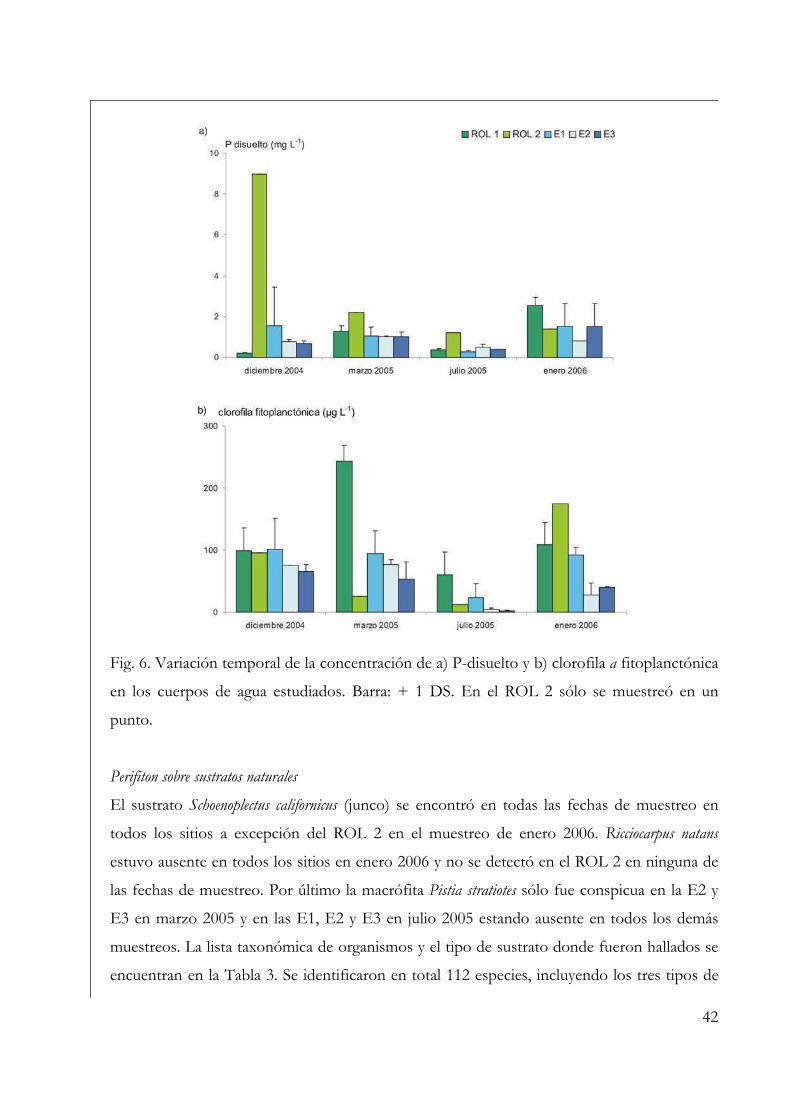
42
Fig. 6. Variación temporal de la concentración de a) P-disuelto y b) clorofila a fitoplanctónica
en los cuerpos de agua estudiados. Barra: + 1 DS. En el ROL 2 sólo se muestreó en un
punto.
Perifiton sobre sustratos naturales
El sustrato Schoenoplectus californicus (junco) se encontró en todas las fechas de muestreo en
todos los sitios a excepción del ROL 2 en el muestreo de enero 2006. Ricciocarpus natans
estuvo ausente en todos los sitios en enero 2006 y no se detectó en el ROL 2 en ninguna de
las fechas de muestreo. Por último la macrófita Pistia stratiotes sólo fue conspicua en la E2 y
E3 en marzo 2005 y en las E1, E2 y E3 en julio 2005 estando ausente en todos los demás
muestreos. La lista taxonómica de organismos y el tipo de sustrato donde fueron hallados se
encuentran en la Tabla 3. Se identificaron en total 112 especies, incluyendo los tres tipos de

43
sustratos analizados. De las especies registradas, 51 pertenecían a la clase Bacillariophyceae
(45,5 %), 21 a la división Cyanobacteria (18,8 %), 26 a la división Chlorophyta (23,2 %), 7 a
Euglenophyta (6,3 %), 5 a la clase Xanthophyceae (4,4 %), 1 a Synurophyceae (0,9 %) y 1 a la
división Cryptophyta (0,9 %). En la Fig. 7 se muestra la variación temporal de la composición
de la comunidad en grupos algales. Las diatomeas dominaron la mayor parte del año, salvo
en enero 2006 cuando codominaron junto con cianobacterias y clorofitas. En cuanto al tipo
de hábito, la mayor parte de los organismos encontrados fueron perifíticos (67 %), mientras
que los restantes fueron planctónicos (26,8 %), ticoplanctónicos (2,7 %) y metafíticos (3,5
%). Cabe destacar que dentro del hábito perifítico se incluyeron tanto las especies epifíticas
sensu estricto (formas adnatas, pedunculadas y erectas asociadas al sustrato) como las algas
bentónicas (aquéllas que se encuentran en el fondo del cuerpo de agua, generalmente
asociadas al sedimento). El porcentaje de algas perifíticas halladas en los distintos tipos de
sustrato en todos los sitios de muestreo fue mayor en diciembre 2004 (92 %, aguas bajas),
mientras que fue menor (64 %) en el muestreo de aguas altas (enero 2006), cuando las algas
planctónicas constituyeron el 31 % de la comunidad (Fig. 8).
La riqueza específica de algas epífitas sobre los sustratos analizados fue mayor en
Schoenoplectus californicus (102) que en Ricciocarpus natans (74) ó Pistia stratiotes (59). De las 112
especies halladas, 26 fueron exclusivas de junco (23 %), mientras que sólo 4 (3,6 %) fueron
exclusivas de R. natans y 3 (2,7 %) de P. stratiotes. El resto de las especies (71 %) fueron
compartidas entre dos o entre los tres tipos de sustrato (Tabla 3). En cuanto a la similitud en
la composición algal entre los distintos sustratos durante todo el estudio ésta fue mayor entre
S. californicus y R. natans (J = 0,61) que entre cualquiera de estos dos sustratos y P. stratiotes
(para S. californicus-P. stratiotes J = 0,49; para R. natans-P. stratiotes J = 0,56).
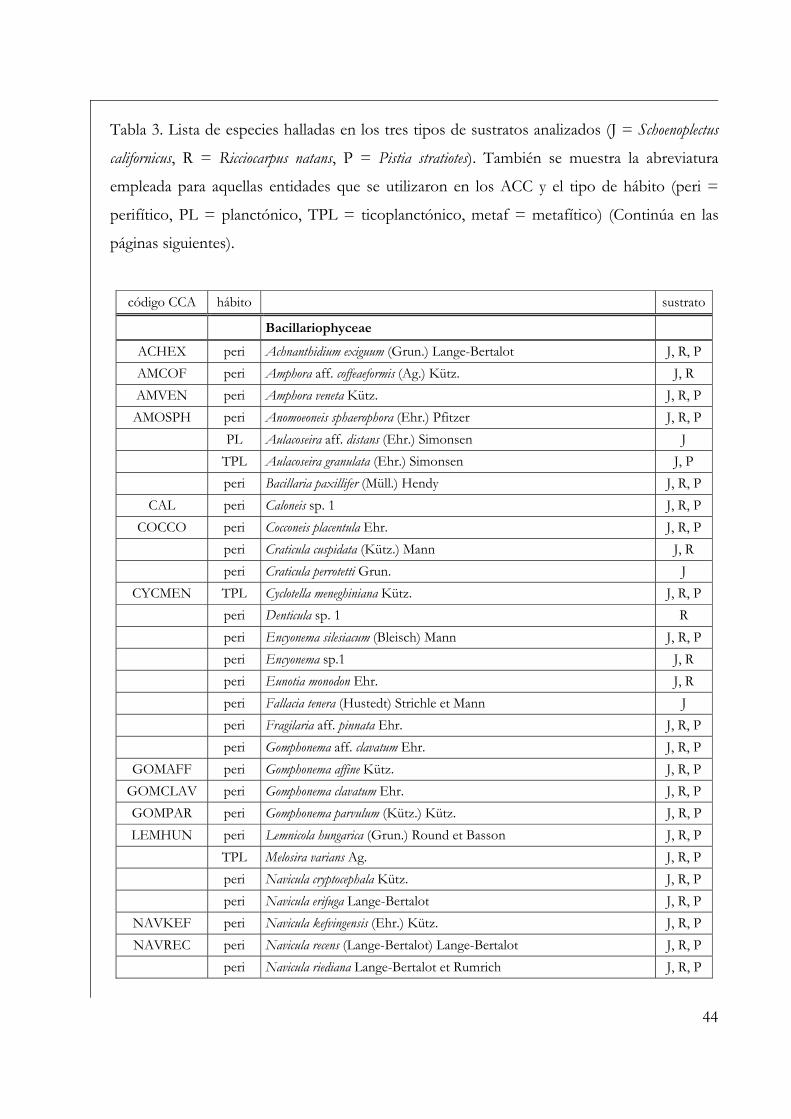
44
Tabla 3. Lista de especies halladas en los tres tipos de sustratos analizados (J = Schoenoplectus
californicus, R = Ricciocarpus natans, P = Pistia stratiotes). También se muestra la abreviatura
empleada para aquellas entidades que se utilizaron en los ACC y el tipo de hábito (peri =
perifítico, PL = planctónico, TPL = ticoplanctónico, metaf = metafítico) (Continúa en las
páginas siguientes).
código CCA hábito sustrato
Bacillariophyceae
ACHEX peri Achnanthidium exiguum (Grun.) Lange-Bertalot J, R, P
AMCOF peri Amphora aff. coffeaeformis (Ag.) Kütz. J, R
AMVEN peri Amphora veneta Kütz. J, R, P
AMOSPH peri Anomoeoneis sphaerophora (Ehr.) Pfitzer J, R, P
PL Aulacoseira aff. distans (Ehr.) Simonsen J
TPL Aulacoseira granulata (Ehr.) Simonsen J, P
peri Bacillaria paxillifer (Müll.) Hendy J, R, P
CAL peri Caloneis sp. 1 J, R, P
COCCO peri Cocconeis placentula Ehr. J, R, P
peri Craticula cuspidata (Kütz.) Mann J, R
peri Craticula perrotetti Grun. J
CYCMEN TPL Cyclotella meneghiniana Kütz. J, R, P
peri Denticula sp. 1 R
peri Encyonema silesiacum (Bleisch) Mann J, R, P
peri Encyonema sp.1 J, R
peri Eunotia monodon Ehr. J, R
peri Fallacia tenera (Hustedt) Strichle et Mann J
peri Fragilaria aff. pinnata Ehr. J, R, P
peri Gomphonema aff. clavatum Ehr. J, R, P
GOMAFF peri Gomphonema affine Kütz. J, R, P
GOMCLAV peri Gomphonema clavatum Ehr. J, R, P
GOMPAR peri Gomphonema parvulum (Kütz.) Kütz. J, R, P
LEMHUN peri Lemnicola hungarica (Grun.) Round et Basson J, R, P
TPL Melosira varians Ag. J, R, P
peri Navicula cryptocephala Kütz. J, R, P
peri Navicula erifuga Lange-Bertalot J, R, P
NAVKEF peri Navicula kefvingensis (Ehr.) Kütz. J, R, P
NAVREC peri Navicula recens (Lange-Bertalot) Lange-Bertalot J, R, P
peri Navicula riediana Lange-Bertalot et Rumrich J, R, P
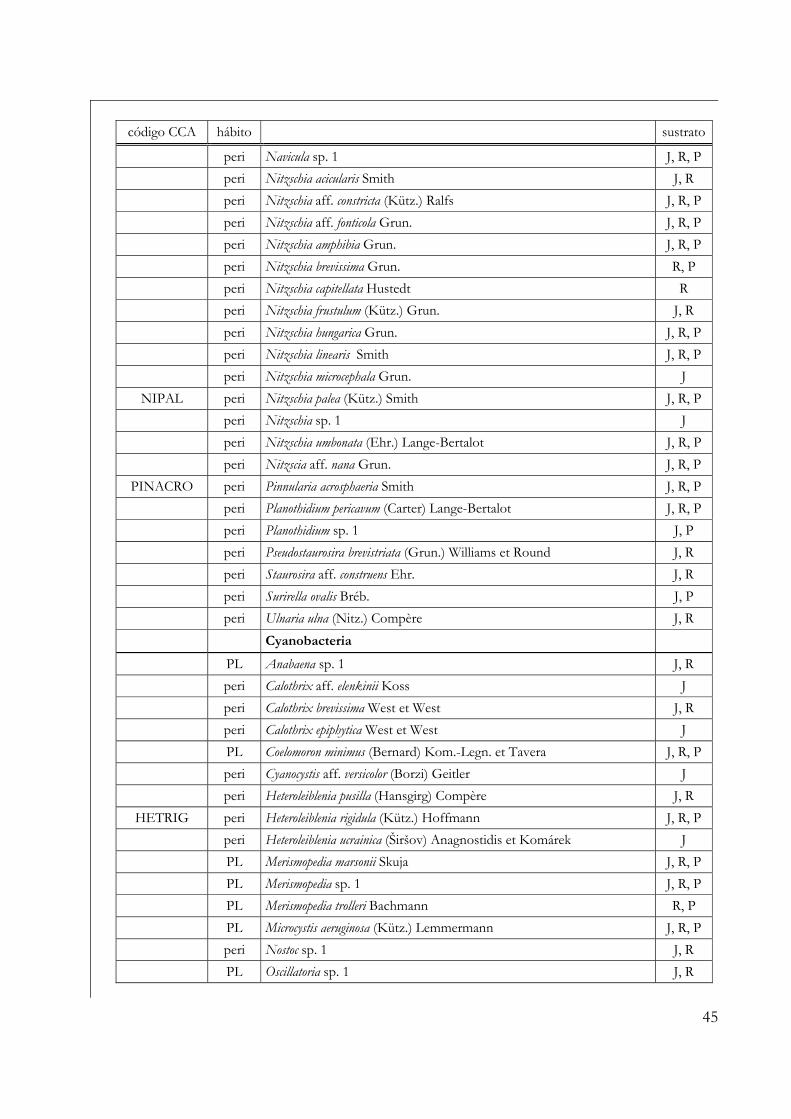
45
código CCA hábito sustrato
peri Navicula sp. 1 J, R, P
peri Nitzschia acicularis Smith J, R
peri Nitzschia aff. constricta (Kütz.) Ralfs J, R, P
peri Nitzschia aff. fonticola Grun. J, R, P
peri Nitzschia amphibia Grun. J, R, P
peri Nitzschia brevissima Grun. R, P
peri Nitzschia capitellata Hustedt R
peri Nitzschia frustulum (Kütz.) Grun. J, R
peri Nitzschia hungarica Grun. J, R, P
peri Nitzschia linearis Smith J, R, P
peri Nitzschia microcephala Grun. J
NIPAL peri Nitzschia palea (Kütz.) Smith J, R, P
peri Nitzschia sp. 1 J
peri Nitzschia umbonata (Ehr.) Lange-Bertalot J, R, P
peri Nitzscia aff. nana Grun. J, R, P
PINACRO peri Pinnularia acrosphaeria Smith J, R, P
peri Planothidium pericavum (Carter) Lange-Bertalot J, R, P
peri Planothidium sp. 1 J, P
peri Pseudostaurosira brevistriata (Grun.) Williams et Round J, R
peri Staurosira aff. construens Ehr. J, R
peri Surirella ovalis Bréb. J, P
peri Ulnaria ulna (Nitz.) Compère J, R
Cyanobacteria
PL Anabaena sp. 1 J, R
peri Calothrix aff. elenkinii Koss J
peri Calothrix brevissima West et West J, R
peri Calothrix epiphytica West et West J
PL Coelomoron minimus (Bernard) Kom.-Legn. et Tavera J, R, P
peri Cyanocystis aff. versicolor (Borzi) Geitler J
peri Heteroleiblenia pusilla (Hansgirg) Compère J, R
HETRIG peri Heteroleiblenia rigidula (Kütz.) Hoffmann J, R, P
peri Heteroleiblenia ucrainica (Širšov) Anagnostidis et Komárek J
PL Merismopedia marsonii Skuja J, R, P
PL Merismopedia sp. 1 J, R, P
PL Merismopedia trolleri Bachmann R, P
PL Microcystis aeruginosa (Kütz.) Lemmermann J, R, P
peri Nostoc sp. 1 J, R
PL Oscillatoria sp. 1 J, R

46
código CCA hábito sustrato
peri Phormidium animale (Ag. ex Gom.) Anagn.et Komárek J
peri Phormidium autumnale (Ag.) Gom. J
PL Planktothrix agardhii (Gom.) Anagn.et Komárek J, R
PL Pseudoanabaena catenata Lauterb. J, R, P
PL Pseudoanabaena sp. 1 J, R, P
PL Raphidiopsis mediterranea Skuja J, R, P
peri Characium sp. 1 J, R
peri Characium ensiforme Hermann J
Chlorophyta
peri Aphanochaete repens Braun R
PL Closterium sp. 1 P
PL Coelastrum sp. 1 J
PL Didymocystis planctonica Korshikov P
PL Dimorphococcus sp. 1 R
EPIB peri Epibolium sp. 1 J, R, P
metaf Gloeocystis sp. 1 J
peri Gloeotila sp. 1 J, R, P
PL Hydrodiction sp. 1 J
peri Klebsormidium klebsii (Smith) Mattox et Blackwell J
peri Microspora sp. 1 J
PL Monoraphidium arcuatum (Kors.) Hind. J, P
PL Monoraphidium circinale (Nyg.) Nyg. P
metaf Mougeotia sp. 1 J, P
OEDO peri Oedogonium sp. 1 J, R, P
PL Pediastrum sp. 1 J
peri Protoderma viride Kütz. J
PL Scenedesmus sp. 1 J, P
peri Schizomeris sp. 1 J, R
metaf Spirogyra sp. 1 J, R, P
STIGEO peri Stigeoclonium sp. 1 J, R, P
peri Tetraspora gelatinosa (Vauch.) Desv. J, R
peri Ulothrix sp. 1 J, R, P
UROCON peri Uronema confervicolum Lagerheim J, R, P
Euglenophyta
PL Euglena acus Ehr. J
PL Euglena archaeoplastidiata Chadefaud J, R
PL Euglena cingula Gojdics J, R
PL Euglena sp. 1 J, R
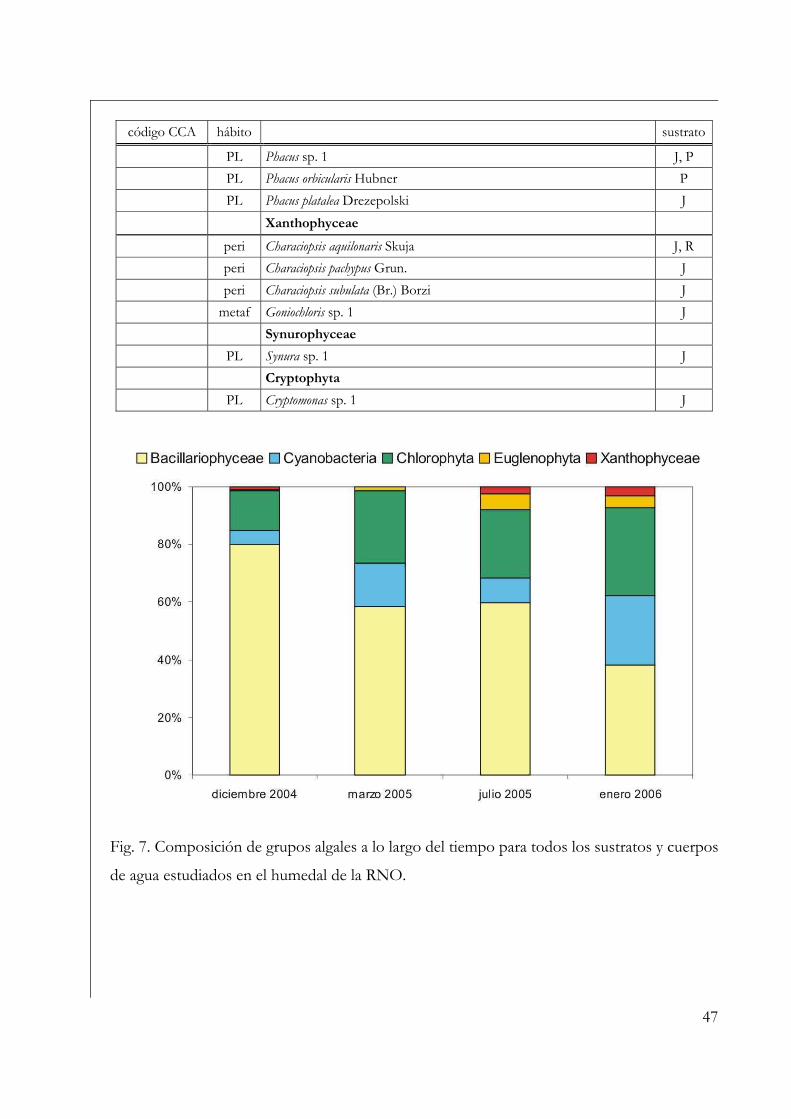
47
código CCA hábito sustrato
PL Phacus sp. 1 J, P
PL Phacus orbicularis Hubner P
PL Phacus platalea Drezepolski J
Xanthophyceae
peri Characiopsis aquilonaris Skuja J, R
peri Characiopsis pachypus Grun. J
peri Characiopsis subulata (Br.) Borzi J
metaf Goniochloris sp. 1 J
Synurophyceae
PL Synura sp. 1 J
Cryptophyta
PL Cryptomonas sp. 1 J
Fig. 7. Composición de grupos algales a lo largo del tiempo para todos los sustratos y cuerpos
de agua estudiados en el humedal de la RNO.
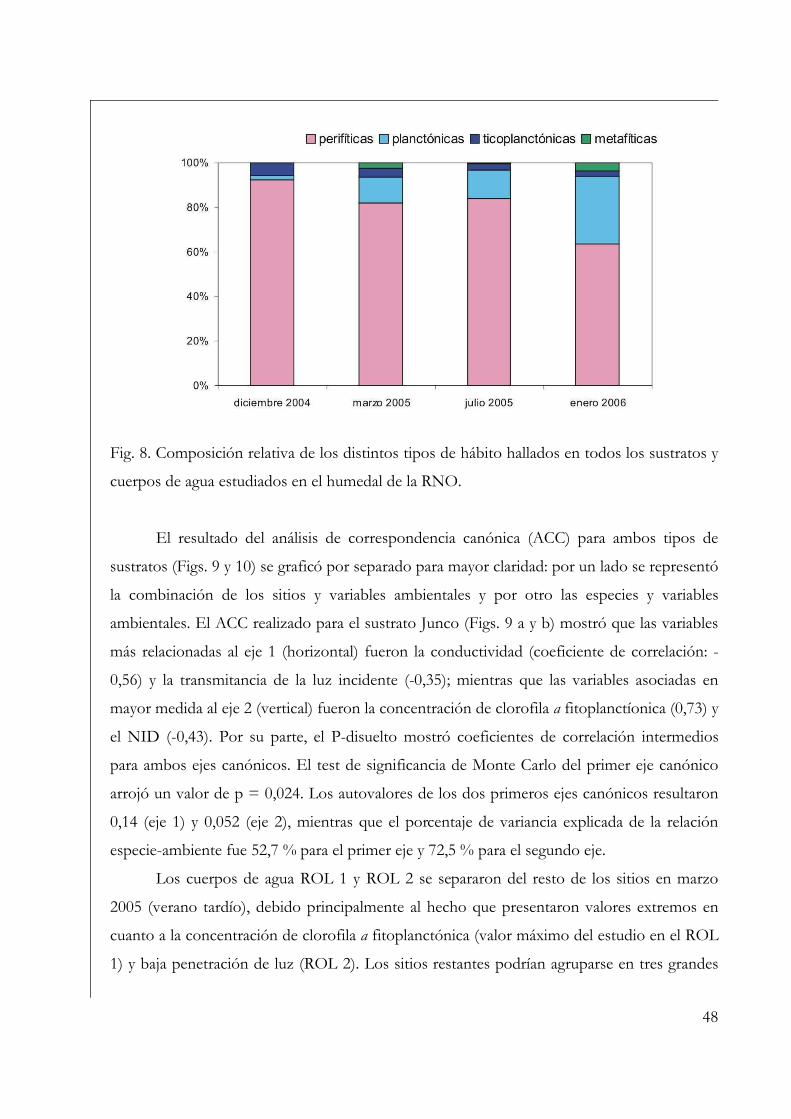
48
Fig. 8. Composición relativa de los distintos tipos de hábito hallados en todos los sustratos y
cuerpos de agua estudiados en el humedal de la RNO.
El resultado del análisis de correspondencia canónica (ACC) para ambos tipos de
sustratos (Figs. 9 y 10) se graficó por separado para mayor claridad: por un lado se representó
la combinación de los sitios y variables ambientales y por otro las especies y variables
ambientales. El ACC realizado para el sustrato Junco (Figs. 9 a y b) mostró que las variables
más relacionadas al eje 1 (horizontal) fueron la conductividad (coeficiente de correlación: -
0,56) y la transmitancia de la luz incidente (-0,35); mientras que las variables asociadas en
mayor medida al eje 2 (vertical) fueron la concentración de clorofila a fitoplanctíonica (0,73) y
el NID (-0,43). Por su parte, el P-disuelto mostró coeficientes de correlación intermedios
para ambos ejes canónicos. El test de significancia de Monte Carlo del primer eje canónico
arrojó un valor de p = 0,024. Los autovalores de los dos primeros ejes canónicos resultaron
0,14 (eje 1) y 0,052 (eje 2), mientras que el porcentaje de variancia explicada de la relación
especie-ambiente fue 52,7 % para el primer eje y 72,5 % para el segundo eje.
Los cuerpos de agua ROL 1 y ROL 2 se separaron del resto de los sitios en marzo
2005 (verano tardío), debido principalmente al hecho que presentaron valores extremos en
cuanto a la concentración de clorofila a fitoplanctónica (valor máximo del estudio en el ROL
1) y baja penetración de luz (ROL 2). Los sitios restantes podrían agruparse en tres grandes

49
grupos. Según nuestro análisis, uno de ellos estuvo caracterizado por altas conductividades y
tenores de P-disuelto, alta penetración de luz, bajo NID y baja concentración de clorofila a
fitoplanctónica. Este grupo estuvo integrado por el arroyo El Pescado (E2) en diciembre
2004 (P) y marzo 2005 (VT) y el ROL 1 y la laguna El Pescado en diciembre 2004. Otro
grupo, integrado por los sitios E2, E3 y ROL 2 en invierno y ROL 2 en diciembre 2004
estuvo caracterizado por altos tenores de NID. Por último, el resto de los sitios fue
determinado por bajas conductividades, transmitancia de luz, NID y concentraciones de P
disuelto. El eje 1 (horizontal) separó los sitios de acuerdo al nivel hidrométrico del humedal,
dado que los muestreos de aguas altas (V = enero 2006) se encuentran a la derecha del eje
mientras que los realizados en períodos de aguas bajas (P = diciembre 2004) se encuentran a
la izquierda del cuadrante y están caracterizados por altos valores de conductividad.
Finalmente, los muestreos realizados en estados hidrológicos intermedios (VT = marzo 2005
e I = julio 2005) se encuentran en la zona media de la figura (Fig. 9 a).
En cuanto al ordenamiento de las especies en función de las variables ambientales
(Fig. 9 b) se pueden distinguir cuatro grandes grupos y una especie que se separa de ellos
(Cocconeis placentula), caracterizada por altos tenores de NID y P disuelto, alta conductividad,
transmitancia de luz y baja concentración de clorofila a fitoplanctónica. Luego, se definen dos
grupos de tres especies cada uno, que se separan principalmente por su tolerancia a la
disponibilidad de luz, por la conductividad y estado trófico (estimado a través de la
concentración de clorofila a fitoplanctónica), siendo similares en cuanto a los niveles de NID
y P disuelto. Otro grupo, compuesto por 5 especies, se encuentra en el cuadrante superior
derecho y está caracterizado por bajos niveles de nutrientes, luz y conductividad y altas
concentraciones de clorofila a fitoplanctónica. Por último, la mayor parte de las especies se
encontraron en condiciones de bajas concentraciones de clorofila a fitoplanctónica, altas
niveles de nutrientes, conductividad y luz.

50
Fig. 9. Ordenamiento del sustrato Schoenoplectus californicus para a) sitios y variables ambientales
y b) especies y variables ambientales. P = primavera (diciembre 2004), VT = verano tardío
(marzo 2005), I = invierno (julio 2005) y V = verano (enero 2006). Los códigos empleados
para las abreviaturas de especies se encuentran señalados en la Tabla 3.

51
En el análisis de correspondencia canónica (ACC) realizado para el sustrato Ricciocarpus
natans (Figs. 10 a y b) se observó que las variables más relacionadas al eje 1 (horizontal)
fueron la concentración de clorofila a fitoplanctónica (coeficiente de correlación: 0,37) y la
conductividad (-0,36); mientras que las variables asociadas en mayor grado al eje 2 (vertical)
fueron la transmitancia de luz incidente (0,61) y la concentración de NID (-0,61). El
coeficiente de correlación de la variable P-disuelto adquirió valores intermedios para ambos
ejes. El test de significancia de Monte Carlo del primer eje canónico arrojó un valor de p =
0,004. Los autovalores de los dos primeros ejes canónicos resultaron 0,197 (eje 1) y 0,111 (eje
2), mientras que el porcentaje de variancia explicada de la relación especie-ambiente fue 44,1
% para el primer eje y 69,1 % para el segundo eje.
En el caso del análisis de ordenamiento para R. natans, la estacionalidad permitió
separar tres grupos de sitios a excepción del ROL 1 en diciembre 2004 (VT), que se mostró
caracterizado por altas concentraciones de clorofila a fitoplanctónica y penetración de luz
incidente (Fig 10 a). El diagrama de especies y variables ambientales se halla representado en
la Fig. 10 b. Se distingue allí un gran grupo de especies en el centro del gradiente ambiental y
cuatro especies no se incluyeron en este grupo por estar en los extremos del gradiente. Las
especies Achnanthidium exiguum, Navicula recens y Stigeoclonium sp. y Uronema confervicolum
mostraron cierto grado de preferencia por valores altos de penetración de irradiancia, P
disuelto y clorofila a fitoplanctónica en el primer caso; elevados valores de NID en cuanto a
N. recens y principalmente a bajas conductividades en el caso de las clorofitas.

52
Fig. 10. Ordenamiento del sustrato Ricciocarpus natans para a) sitios y variables ambientales y
b) especies y variables ambientales. P = primavera (diciembre 2004), VT = verano tardío
(marzo 2005), I = invierno (julio 2005) y V = verano (enero 2006). Los códigos empleados
para las abreviaturas de especies se encuentran señalados en la Tabla 3.

53
Los patrones temporales de プ diversidad de los sitios se encuentran representados en
la Fig. 11 a. El mayor valor de riqueza específica observado ocurrió en la E3 en marzo 2005
(65), mientras que el más bajo se registró en el ROL 2 durante el mismo muestreo (1). El
ROL 2 fue, a excepción del muestreo de diciembre 2004, el sitio menos rico en especies. En
el muestreo de diciembre 2004 se observó la menor diferencia entre valores de riqueza del
sitio más y menos rico en especies (15), mientras que en el muestreo de marzo la diferencia
fue máxima (64). En cuanto al patrón temporal de ベ diversidad, se observó en general que el
sitio con mayores valores de ベW fue el ROL 2, salvo en el muestreo de diciembre 2004,
cuando el mayor valor se observó en el arroyo El Pescado (5,9) (Fig. 11 b). EL valor más
bajo de ベW fue 1,7 y se detectó en la E3 en marzo 2005, mientras que el más alto fue 112 y de
detectó en el ROL 2 en la misma fecha de muestreo. El patrón general que se observa a partir
del análisis de la Fig. 11 b es que los sitios más ricos en especies (ej. E3 en marzo 2005)
fueron los que compartieron más especies con el resto de los sitios de muestreo (menor ベ
diversidad), mientras que aquéllos con menor riqueza específica (ej. ROL 2) fueron más
diferentes al resto de los sitios (mayor ベ diversidad).
Cuando se analizan los patrones de diversidad para todo el estudio, se observa la
misma tendencia general anteriormente descrita. El sitio más rico en especies (laguna El
Pescado) fue también el que tuvo la menor ベ diversidad; mientras que el ROL 2, el sitio con
menor riqueza, mostró el mayor valor de ベ diversidad (Tabla 4).
En cuanto a la similitud entre pares de sitios, en la Tabla 5 se muestra el resultado del
cálculo del índice de Jaccard para todas las combinaciones posibles de cuerpos de agua. Se
obtuvo que el ROL 2 fue el que más se diferenció del resto de los sitios estudiados (cuando
estuvo presente el ROL 2 en la comparación se obtuvieron los valores más bajos del índice),
mientras que el resto de los sitios mostró valores relativamente altos y similares.
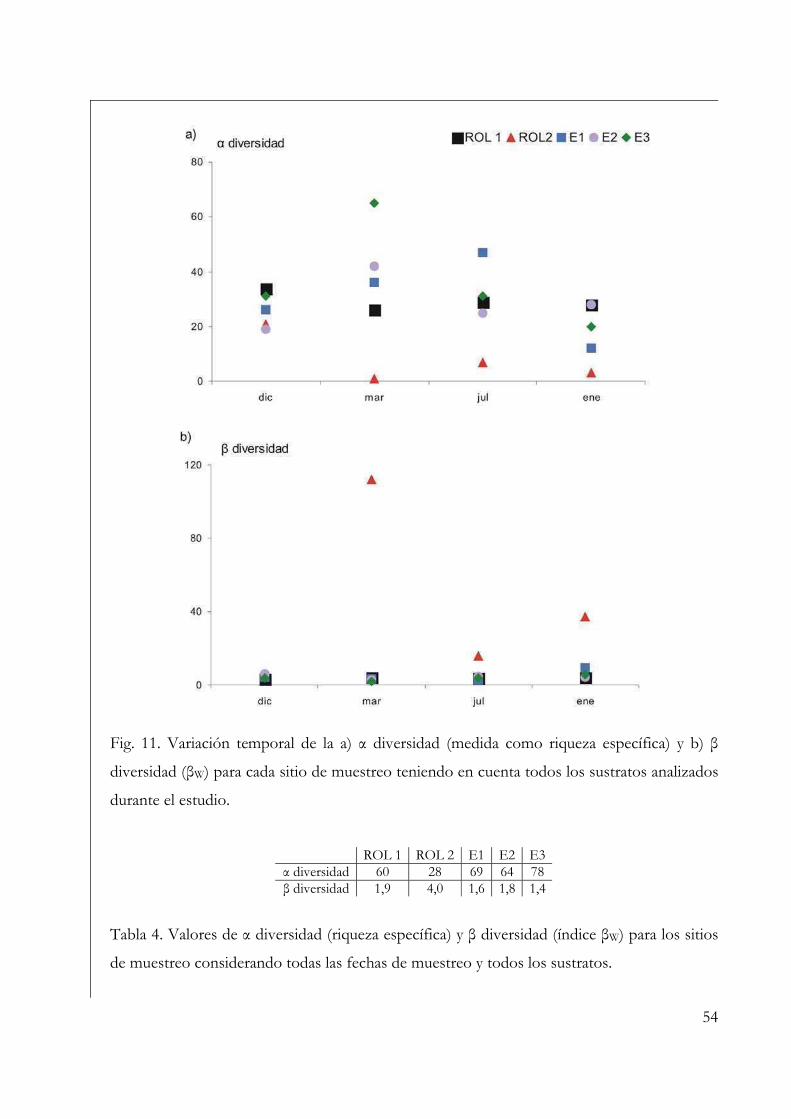
54
Fig. 11. Variación temporal de la a) プ diversidad (medida como riqueza específica) y b) ベ
diversidad (ベW) para cada sitio de muestreo teniendo en cuenta todos los sustratos analizados
durante el estudio.
ROL 1 ROL 2 E1 E2 E3プ diversidad 60 28 69 64 78ベ diversidad 1,9 4,0 1,6 1,8 1,4
Tabla 4. Valores de プ diversidad (riqueza específica) y ベ diversidad (índice ベW) para los sitios
de muestreo considerando todas las fechas de muestreo y todos los sustratos.

55
ROL 1 ROL 2 E1 E2 E3ROL 1 - ROL 2 0,33 -
E1 0,56 0,35 - E2 0,53 0,37 0,56 - E3 0,52 0,33 0,53 0,58 -
Tabla 5. Valores del índice de Jaccard entre pares de cuerpos de agua teniendo en cuenta
todas las fechas de muestreo y todos los sustratos analizados.
DISCUSIÓN
En los valles aluviales de los grandes ríos, el pulso de inundación suele ser el principal
modelador tanto de la biota como de las características físicas y químicas del ecosistema
(Junk et al. 1989). En la Reserva Natural Otamendi, el efecto de una napa freática alta y
fluctuante y las precipitaciones juegan un rol fundamental en las variaciones del nivel
hidrométrico de los cuerpos de agua del humedal (Chichizola 1993). En este estudio, el
contraste entre los períodos de aguas altas y aguas bajas se visualizó principalmente a partir
de los valores de conductividad, sólidos en suspensión y absorbancia del agua a 254 nm. La
homogeneización de algunas variables abióticas (conductividad, sólidos en suspensión,
contenido húmico, NID, P-disuelto) y del fitoplancton (ej. composición, densidad y
biovolumen algal según señaló de Tezanos Pinto 2008) en el verano de aguas altas evidenció
la posibilidad de la existencia de procesos de conectividad entre los cuerpos de agua del
humedal, a pesar de que no se registraron inundaciones por el desborde de los ríos aledaños.
Las lagunas Grande y El Pescado y el arroyo del mismo nombre presentaron
características limnológicas más similares entre sí que con los ROLs teniendo en cuenta
principalmente el contenido húmico y otras variables abióticas. Por su parte, los ROLs
fueron más ricos en carbono orgánico disuelto y nutrientes en general, mientras que las
lagunas y el arroyo presentaron ocasionalmente bajos valores de NID, particularmente en los
meses más cálidos. Sin embargo, esto no significa necesariamente que el NID haya sido
limitante, sino que probablemente sea reciclado más rápidamente en los meses de mayor

56
crecimiento algal, cuando sería más plausible encontrarlo incorporado a la biomasa de los
organismos.
En el caso del perifiton, es muy difícil establecer un rango de nutrientes (ej. NID o P)
o relación entre concentración de nutrientes (N:P), para el cual el crecimiento algal se vería
limitado. Este nivel de nutrientes depende entre otros factores de la estructura de la
comunidad en cuestión (Borchardt 1996) y del tipo de sustrato en el que se encuentra
asentada, ya que algunos de ellos pueden ser una fuente extra de nutrientes para las algas
(Kalff 2003). Las algas perifíticas pueden tomar los nutrientes de diversas fuentes, entre ellas,
de la columna de agua, del reciclado en la propia matriz perifítica y del sustrato mismo
(Wetzel 2005). Además, depende del estado trófico del cuerpo de agua; en este sentido
Eminson & Moss (1980), sostuvieron que el aporte potencial de nutrientes del sustrato al
epífito decrecería con el aumento de la eutrofia ya que tomarían nutrientes de la columna de
agua cuando sean abundantes en ésta, y del sustrato cuando la disponibilidad de nutrientes en
el medio fuera baja. De esta manera, es muy difícil establecer un nivel de nutrientes en la
columna de agua que restrinja el crecimiento del perifiton. Sin embargo, entre los valores
que figuran en la bibliografía, podemos mencionar que Bothwell (1988) determinó que
concentraciones de P-PO43- de entre 0,3-0,6 µg L-1 saturaban el crecimiento de diatomeas
bentónicas en un río templado del hemisferio norte. Por otro lado, otras comunidades algales
como las integradas mayoritariamente por clorofitas necesitarían mayores concentraciones de
P (25-50 µg L-1) para alcanzar un crecimiento máximo (Borchardt 1996). En el caso de la
concentración de NID se han reportado valores similares a los considerados limitantes para
el máximo crecimiento fitoplanctónico (55-100 µg L-1, Borchardt op. cit. y las referencias que
allí se citan).
La composición en grupos algales mayoritarios hallada por nosotros en la RNO
(diatomeas, clorofilas y cianobacterias) es la citada en la bibliografía como comúnmente
encontrada en humedales (Goldsborough et al. 2005). También Putz (1997) encontró en
sistemas lénticos de aguas húmicas del valle de inundación del río Amazonas que la
comunidad perifítica estaba principalmente compuesta por diatomeas la mayor parte del año.
Gómez et al. (2003) observaron en epífitas de junco en el Río de La Plata un patrón
sucesional similar al hallado en este trabajo, con dominancia de diatomeas gran parte del año
a excepción del verano, cuando dominaron las algas verdes. En nuestro estudio, las

57
diatomeas dominaron gran parte del año, a excepción del período de aguas altas, cuando se
detectó además una gran proporción de algas planctónicas en la composición de la
comunidad. Este hecho puede explicarse teniendo en cuenta que en sistemas someros las
distintas comunidades algales interactúan íntimamente y sus hábitats suelen solaparse
(Goldsborough & Robinson 1996) debido a los procesos de mezcla y resuspensión de los
sedimentos (Schallenberg & Burns 2004). En este sentido, Gómez et al. (2003) observaron
que en junco (Schoenoplectus californicus) la comunidad estaba dominada por una gran
proporción de algas planctónicas y bentónicas. En nuestro estudio encontramos que el 60%
de las diatomeas encontradas fueron bentónicas (relacionadas con los sedimentos del fondo),
coincidentemente con lo discutido por las autoras.
Por otro lado en el muestreo de aguas altas, la cobertura de macrófitas flotantes estaba
reducida a su mínima expresión en los cuerpos de agua del humedal, hecho que trajo
aparejado que no se registre perifiton en esta oportunidad. Goldsborough & Robinson (1996)
desarrollaron un modelo conceptual en el que definen cuatro estados posibles en la
dominancia de los grupos algales mayoritarios en los humedales de acuerdo principalmente al
nivel hidrométrico, presencia de macrófitas, herbivoría y cantidad de luz. En este sentido, los
autores predicen que en condiciones de ausencia de macrófitas y altos niveles hidrométricos
el fitoplancton es la comunidad dominante en el humedal y denominaron a estas
circunstancias "estado de lago".
De acuerdo a los valores del índice de Jaccard en relación a la comparación de la
comunidad algal entre sustratos, obtuvimos que las macrófitas más parecidas entre sí en
relación a su composición perifítica algal fueron Schoenoplectus californicus y Ricciocarpus natans.
Hay mucha bibliografía donde se discute la posible existencia de especificidad alga-sustrato y,
en este sentido, Tell (1977) afirma que si bien se conocen algunos casos de especificidad de
algas epífitas por un sustrato determinado, en general éste les resulta indiferente. Por este
motivo, y en concordancia a lo discutido por el autor, era esperable que los sustratos tuvieran
una gran proporción de especies en común. Gran parte de los organismos encontrados por
nosotros en ambos tipos de sustrato ya han sido registrados en otros trabajos en distintos
tipos de ambientes en sustratos similares (Bauer et al. 2007, Gómez et al. 2003, Tesolín & Tell
1996, Pozzobon & Tell 1995, Ács & Buczó 1994, Tell 1977). Por este motivo podemos
afirmar que los especímenes encontrados en este trabajo poseen una gran afinidad por el

58
hábito epifítico. Por ejemplo, en Tell (1977) se describe la presencia de Lemnicola hungarica en
los vértices de las células de las escamas de Ricciocarpus natans, tal como fuera observado por
nosotros en este estudio, además de la presencia del alga verde Epibolium sp. en este sustrato.
Las especies de diatomeas halladas en este estudio presentan, de acuerdo a la
bibliografía (van Dam et al. 1994), rangos de tolerancia similares a determinadas variables
ambientales, presentando la mayoría características que les permiten prosperar en este tipo de
ambientes. La mayoría está adaptada a vivir en cuerpos de agua con pH entre neutrales y
alcalinos, de salinidad intermedia a elevada; la mayoría de ellas son autótrofas en cuanto al
metabolismo del N, tolerando bajas a moderadas concentraciones de O2, ocurren en sitios de
estado trófico elevado y en general toleran cierto grado de desecación. Por este motivo, no es
sorprendente que algunas especies ocupen distintos lugares en los diagramas de
ordenamiento para los dos tipos de sustrato dada la estrechez del gradiente ambiental
analizado; es decir, los ambientes presentaron siempre conductividades elevadas, condiciones
eutróficas semejantes, altas concentraciones de P-disuelto, baja penetración de luz, y en
general bajas concentraciones de NID. Por consiguiente, si bien pueden estar ocupando
posiciones dispares en los diagramas, estas diferencias representan situaciones muy
semejantes en la naturaleza.
Otro factor que cobra relevancia a la hora de analizar los esquemas de ordenamiento
de las especies son los distintos tipos de hábitats que proveen los sustratos y la interacción
con el ambiente abiótico que los organismos tienen con él. La concentración de clorofila a
fitoplanctónica que es indicadora del estado trófico, junto con la concentración de NID
fueron las dos variables que afectaron más a la estructura del perifiton tanto en Schoenoplectus
californicus como en Ricciocarpus natans. Entonces, la abundancia del fitoplancton se vio
reflejada en este análisis como así también el nutriente cuya dinámica estaría relacionada con
su consumo mayoritario, y que podría llegar a ser materia de competencia entre las
comunidades. Por otro lado, la conductividad influyó más en el ordenamiento de S. californicus
que en el de R. natans en tanto que el porcentaje de transmitancia de luz incidente, afectó más
al de R. natans que al de junco. Considerando que la conductividad representa aquí además a
la altura hidrométrica, queda en evidencia el efecto de sus fluctuaciones en la comunidad
asentada en el junco. En R. natans, la intensidad lumínica es la variable que modula al

59
perifiton considerando que la restricción en la cantidad de luz se hace más fuerte cuando el
organismo vive debajo de una planta flotante debido al sombreado provocado por ésta.
La comparación de la ペ diversidad del perifiton sobre sustratos naturales con la de otros
sistemas de características similares es dificultosa, ya que para nuestro conocimiento no se
registran muchos trabajos en este sentido. Podemos especular que la ペ diversidad de la
comunidad algal fue baja porque fue similar a la que registraron Tesolín & Tell (1996) sobre
macrófitas flotantes (Eichhornia crassipes, Salvinia rotundifolia, Lemna gibba y Ricciocarpus natans) en
una sola laguna somera de la llanura de inundación del río Paraná (136 taxa). También fue
menor a la registrada por Putz & Junk (1997) en 9 arroyos cercanos a Manaos en Brasil (329)
y a la detectada por Rodrigues & Bicudo (2001) en un gradiente de ambientes lénticos y
lóticos del valle aluvial del río Paraná Superior (228). Una posible explicación para la baja ペ
diversidad del perifiton sobre sustratos naturales puede ser que las altas concentraciones de
fitoplancton (ej. concentración de clorofila a) restrinjan su desarrollo, compitiendo por la luz
y los nutrientes (Borchardt 1996, Hill 1996).
Tanto el índice ベW como el de Jaccard (J) se emplearon en este trabajo como
estimadores de la ベ diversidad. Ambos señalaron la diferencia en composición específica del
ROL 2 con respecto a los demás ambientes acuáticos. Ésta se debió a la poca cantidad de
especies registradas en este sitio, debido muy probablemente a que el profuso desarrollo de
plantas emergentes provocó impedimentos físicos en la colonización de los microorganismos
además de la baja penetración de la irradiancia incidente. La alta ベW del ROL 2 en marzo
2005 (verano tardío) quedó también manifestada en el ACC para junco (Fig. 9 a), donde este
sitio quedó alejado del conjunto de sitios en el diagrama de ordenamiento.
En este capítulo hemos descrito la comunidad perifítica algal sobre sustratos naturales en los
principales ambientes acuáticos de la RNO. Hemos observado que las especies encontradas
son ubicuas y tienen rangos de tolerancia similares que les permiten prosperar en este tipo de
ambientes. Se encontró que la mayor parte de las especies encontradas en los sustratos
mayoritarios (junco y Ricciocarpus natans) eran compartidas entre ellos sin manifestarse
especificidad clara. Hemos advertido que en períodos de aguas altas la comunidad presentaba
un mayor número de representantes del plancton, mientras que durante el resto del año la

60
comunidad estaba compuesta principalmente por algas epífitas o bentónicas, debido a la
escasa profundidad de los cuerpos de agua estudiados y a los procesos de remoción
provocados por el viento. A partir del análisis de ordenamiento para los sitios y las especies
consideramos que la fluctuación en el nivel hidrométrico afectó más a la comunidad perifítica
sobre Schoenoplectus californicus en tanto que las condiciones de irradiancia afectaron más a la
adherida en R. natans. Estaríamos además observando el posible efecto modulador del
fitoplancton sobre el perifiton en el humedal. El cuerpo de agua más alejado y con menor
probabilidad de conectarse de manera directa con el resto (ROL 2) fue el que más se
diferenció de los demás en cuanto a sus características limnológicas y patrones de diversidad.
La posesión de pocas especies en el ROL 2 fue el determinante de su mayor ベ diversidad y
bajos valores del índice de Jaccard, provocados por el desarrollo de vegetación emergente
que pudo influenciar en la colonización física del perifiton y en la penetración de irradiancia.
Por último, hemos aportado además nueva información sobre la diversidad de la ficoflora
microbiana del humedal de Otamendi, incorporando 112 especies perifíticas a las ya
registradas 289 especies fitoplanctónicas (de Tezanos Pinto 2008). El humedal de la RNO en
un sistema rico en especies y su conocimiento desde lo estructural constituye un punto de
sumo interés en la actualidad, a la luz de los fenómenos de cambio global que están afectando
al planeta.

61
CAPÍTULO II
ESTRUCTURA Y PRODUCCIÓN PRIMARIA
DEL FITOPLANCTON Y PERIFITON EN LA LAGUNA GRANDE

62
INTRODUCCIÓN
El Fitoplancton
El fitoplancton es el conjunto de microorganismos fotosintéticos adaptados a pasar parte o
toda su vida en suspensión en el seno del agua (Reynolds, 2006). Cuando estos organismos
pasan una parte de sus ciclos vitales adheridos a un sustrato, se les denomina meroplancton,
para distinguirlos del fitoplancton verdadero también llamado holoplancton o euplancton.
Constituyen por lejos, el grupo de organismos más estudiado en las aguas continentales y los
océanos. Se han escrito innumerables artículos acerca de su fisiología y ecología en estudios
tanto de campo como de laboratorio. La estructura y la productividad de esta comunidad en
relación a las variables ambientales y sus interacciones biológicas han recibido mucha
atención durante décadas por parte de los limnólogos (Kalff 2003). Existen numerosas
clasificaciones para estos organismos, tanto desde puntos de vista estructurales como
funcionales. Sieburth et al. (1978) los agrupó de acuerdo a su tamaño celular; Margalef (1978)
por su parte, relacionó las estrategias de vida y la morfología de estos organismos con el
ambiente. Reynolds (1980) propuso un modelo que más tarde retoma y revisa (Reynolds et al.
2002), donde clasifica al fitoplancton en grupos funcionales de acuerdo a sus rangos de
tolerancia a variables ambientales tales como los niveles de nutrientes, pH, régimen de
mezcla, irradiancia, color del agua, etc.
En general, el fitoplancton de agua dulce está compuesto principalmente por
organismos pertenecientes a las divisiones y/o clases algales Chlorophyta, Euglenophyta,
Bacillariophyceae, Chrysophyceae, Synurophyceae, Cryptophyta y Pyrrophyta (van den
Hoeck 1995). Las bacterias fotosintéticas pertenecientes a la división Cyanobacteria, a pesar
de ser procariotas, suelen ser agrupadas con el fitoplancton más que con el bacterioplancton
por sus similitudes con el resto de las algas (Kalff 2003). Es decir que la definición de
fitoplancton agrupa a los organismos en mayor medida desde un punto de vista funcional y
de caracteres adaptativos compartidos, que desde el punto de vista filogenético.
Es posible observar dentro del fitoplancton, una gran variedad de formas de vida y
hábitos, desde organismos unicelulares cocoides o flagelados hasta organismos pluricelulares
cenobiales, filamentosos o coloniales. Existe además una amplia gama de adaptaciones a la

63
vida pelágica, desde la presencia de flagelos, mucílago, espinas y acumulación de lípidos, a la
formación de cadenas, colonias o la posesión de vesículas de gas, tal como ocurre en algunas
cianobacterias, a quienes permiten permanecer en suspensión en la columna de agua y regular
su flotabilidad (Kirk 1994, Reynolds 2006).
El espectro de estrategias de nutrición del fitoplancton es muy variado (Jones 2000).
Los fotoautótrofos estrictos son capaces de fotosintetizar liberando oxígeno y de crecer con
carbono inorgánico (CO2) como única fuente de carbono, además de utilizar otros nutrientes
en su forma inorgánica. Los fotoautótrofos obligados pueden tomar y metabolizar algunos
compuestos orgánicos de carbono, pero sólo pueden crecer si disponen de luz y CO2
(Falkowski & Raven 2007). Por otro lado, existen fotoautótrofos facultativos, también
llamados mixotróficos, que además de fotosintetizar son capaces en ciertas circunstancias
ambientales, de utilizar carbono orgánico como suplemento en reemplazo de luz y CO2 para
el crecimiento. La osmotrofia y la fagotrofia son dos variantes del metabolismo mixotrófico
(Jones 1994). En el primer caso, los organismos absorben compuestos orgánicos en forma
soluble a través de la membrana plasmática o por pinocitosis. Por ejemplo Chlamydomonas
spp., pueden utilizar compuestos orgánicos como el acetato para suplementar su nutrición
fotosintética (Falkowski & Raven 2007). Los organismos fagotróficos son aquéllos capaces
de ingerir partículas en vacuolas alimenticias para su subsecuente digestión. La fagotrofia en
fotoautótrofos facultativos ocurre por ejemplo en crisofíceas, criptofitas, haptofíceas y
dinoflagelados (Falkowski & Raven 2007).
Todos los organismos fitoplanctónicos comparten la posesión de clorofila a como
pigmento fotosintético y además, muchos de ellos poseen pigmentos accesorios,
característica que les permite ampliar el rango de longitudes de onda disponibles para la
fotosíntesis (Kirk 1994). Esta propiedad tiene gran importancia ecológica ya que la radiación
penetra diferencialmente en el agua, puesto que ciertas longitudes de onda son absorbidas
y/o dispersadas por los elementos disueltos y en suspensión que contiene el medio líquido.
Por ejemplo, en los lagos húmicos penetran bajas intensidades lumínicas y predominan
longitudes de onda larga ya que los ácidos húmicos absorben en particular la radiación de
onda corta del espectro (Arvola et al. 1999). De esta manera, en cuerpos de agua húmicos se
ven favorecidos los grupos poseedores de ficobilinas tales como las cianobacterias y

64
criptofíceas, que poseen ficoeritrina y ficocianina, con picos de absorción en el verde (≈ 565
nm) y rojo (≈ 620 nm) respectivamente (Kirk 1994).
Antecedentes
La comunidad fitoplanctónica se está estudiando en Argentina desde hace décadas,
enfocándose su análisis desde numerosos puntos de vista. En particular en la llanura de
inundación del río Paraná se han llevado a cabo numerosas investigaciones como por
ejemplo, estudios realizados tanto en lagunas y ríos del valle aluvial del río Paraná como en el
propio curso principal, tanto en el tramo superior (Train & Rodrigues 1997), como en el
tramo medio (García de Emiliani 1993, 1997; Zalocar de Domitrovic 1990, 1992) y el inferior
(Mercado & Gómez 1999, Unrein 2002). Por su parte, Izaguirre et al. (2001 a) estudiaron la
variación de la composición específica del fitoplancton a lo largo de una transecta que se
extendía desde el río Paraná en su tramo inferior hasta una laguna del valle de inundación,
conectada al río a través de un arroyo. Otros trabajos hacen referencia a aspectos tales como
la dinámica de los nutrientes en el río Paraná. En este sentido, Bonetto et al. (1994) trabajaron
en el delta del río en su tramo inferior, en tanto que Carignan & Planas (1994) estudiaron la
limitación lumínica y de nutrientes del fitoplancton en lagunas de la planicie de inundación
del río Paraná en su tramo medio. Unrein & Tell (1994) analizaron la limitación por
nutrientes en el propio río en su tramo inferior. Por su parte, Bonetto el al. (1983, 1979)
estudiaron la composición del fitoplancton y su producción primaria en el tramo superior del
río en tanto que Izaguirre & O'Farrell (1999) estudiaron la dinámica de la producción
primaria de la comunidad fitoplanctónica en una laguna del valle de inundación del río
Paraná por el método de intercambio de oxígeno. Algunos estudios más recientes llevados a
cabo en el área se centraron en el análisis de las interacciones tróficas entre las distintas
comunidades acuáticas, como por ejemplo los trabajos de Sinistro et al. (2006, 2007).
También se ha publicado un trabajo acerca de la productividad primaria del fitoplancton en la
Laguna Grande de la Reserva Natural Otamendi (Rodríguez & Pizarro 2007) con resultados
preliminares de este capítulo de tesis.

65
La Estructura del Fitoplancton en la RNO
La estructura del fitoplancton en distintos cuerpos de agua de la Reserva Natural de
Otamendi ha sido estudiada y descrita detalladamente, principalmente para la Laguna Grande
y lagunas en meandros abandonados (ROLs, del inglés "relictual oxbow lakes") por Izaguirre
et al. (2004, 2001 b) y O´Farrell et al. (2007, 2003). En el trabajo de Izaguirre et al. (2004) se
describe la composición específica del fitoplancton y su dinámica anual en la Laguna Grande
y los ROLs. Los autores encontraron que en la laguna, las cianobacterias constituían en
general el grupo dominante en la mayor parte del período de estudio. Entre diciembre y
enero dominaban las clorofíceas del orden Chlorococcales, acompañadas por varias especies
de Cryptomonas, constituyendo ambos grupos más del 65 % de la densidad total. Por otro
lado, las diatomeas fueron cuantitativamente menos importantes en esa época del año, pero
contribuyeron sustancialmente a la densidad total de organismos y a la riqueza específica
entre enero y abril. El resto del año codominaron las diatomeas, criptofíceas y clorofíceas.
Los autores destacaron en su trabajo que en la Laguna Grande las cianobacterias y
euglenoideos estaban bien representados y en menor proporción se encontraban las
crisofíceas, tribofíceas y dinofíceas. Los muestreos fueron realizados en dos puntos de la
laguna, uno ubicado en la zona litoral y el otro en la zona pelágica, y observaron densidades
máximas de fitoplancton más altas en la zona litoral (≈ 30000 ind mL-1) que en la zona
pelágica (≈ 25000 ind mL-1). El patrón anual de densidades resultó estacional, con valores
más bajos en invierno y más altos en primavera y verano. En O´Farrell et al. (2003) señalaron
que la densidad total de fitoplancton en la zona litoral de la Laguna Grande osciló entre ≈
700 y ≈ 30000 ind mL-1 bajo la cobertura de plantas flotantes. Las algas verdes dominaron
llegando a tener densidades de hasta aproximadamente 25000 ind mL-1 y los principales
grupos acompañantes fueron las criptofíceas y los euglenoideos.
Recientemente, O´Farrell et al. (2007) estudiaron en estos mismos ambientes, la
relación entre caracteres morfológicos externos de las algas y las distintas condiciones de
iluminación del sitio donde fueron encontrados lo especímenes. Por ejemplo, observaron que
en los ambientes menos iluminados había organismos de menor longitud con alta relación
superficie/volumen (S/V). En los sitios más iluminados los individuos eran más largos y
poseían una menor relación S/V, y también advirtieron la presencia de flagelados en estos

66
sitios. Por otro lado, relacionaron la presencia/ausencia de vacuolas en las cianobacterias con
la intensidad lumínica. Las vacuolas de gas le confieren flotabilidad a las cianobacterias,
permitiéndoles posicionarse en la columna de agua en zonas con distintas intensidades
lumínicas y contenido de nutrientes (Kirk 1994). Los autores, observaron a partir de las
muestras de un experimento realizado en la Laguna Grande (de Tezanos Pinto et al. 2007), la
disminución del contenido vacuolar de algunas cianobacterias luego de la remoción de la
cobertura de macrófitas en mesocosmos. Además, registraron una mayor abundancia de
cianobacterias con vacuolas en los lugares menos iluminados. Por último, con respecto al
picoplancton autotrófico, encontraron densidades más bajas en sitios con alto contenido
húmico. Esta observación la relacionaron con la presión de predación ejercida por parte de
los flagelados heterotróficos y ciliados sobre el picoplancton autotrófico en los ROLs, con
mayor contenido húmico.
En resumen, el fitoplancton de la Laguna Grande se caracteriza por estar compuesto
por algas adaptadas a restricciones lumínicas, impuesta en primer lugar por el contenido
húmico del agua y luego por una cobertura fluctuante de macrófitas flotantes. Por otro lado,
es destacable la presencia de grupos que poseen biliproteínas como pigmentos accesorios
(cianobacterias y criptofíceas) que les confieren un mayor rango de absorción lumínica en las
longitudes de onda del rojo (ficocianina) y verde (ficoeritrina) predominantes en los cuerpos
de agua húmicos (Eloranta 1999, Kirk 1994). Además, la presencia de organismos
mixotróficos ha sido comprobada experimentalmente en la Laguna Grande por Sinistro et al.
(2006). Esta estrategia nutricional es ventajosa en ambientes con pobre iluminación o
deficientes en nutrientes (Jones 2000, Jansson et al. 1996). Las condiciones de baja irradiancia
son características de los cuerpos de agua de la RNO en tanto que la limitación de nutrientes
fue analizada experimentalmente por Unrein (2001), quien demostró que el fósforo no sería
el factor limitante para el fitoplancton en este humedal, pero que bajo ciertas condiciones sí
puede serlo el nitrógeno.
El Perifiton
Si bien se han llevado a cabo numerosos estudios acerca de distintos aspectos tanto
estructurales como funcionales de la comunidad perifítica, se han realizado muchas menos

67
investigaciones acerca de esta comunidad en comparación con el fitoplancton
(Vadeboncoeur et al. 2002, Lodge et al. 1998). Este sesgo en cuanto al número de artículos no
está dado por una menor importancia en términos ecológicos, ya que por el contrario, las
algas perifíticas pueden aportar la mayor parte de la producción primaria en cuerpos de agua
bien iluminados tales como arroyos, lagos someros, humedales y ambientes costeros
(Vadeboncoeur et al. 2001, Dodds et al. 1999). Según Sand-Jensen & Borum (1991), es
probable que esta desigualdad radique, por un lado, en una larga tradición histórica de
estudios de ecología del fitoplancton y por otro, en las dificultades metodológicas que
involucra el estudio del perifiton. Señalan los mismos autores, que la comunidad adherida
está generalmente distribuida de manera muy heterogénea y es más difícil de cuantificar en
términos absolutos cuando se trabaja con sustratos naturales.
El desarrollo de estudios acerca de la comunidad perifítica adherida a sustratos
artificiales estratégicamente colocados por el investigador es un intento de minimizar los
inconvenientes en la toma de muestras y cuantificación de los atributos de la comunidad
(Cattaneo & Kalff 1979). Por otro lado, tanto el tipo de sustrato (rocas, macrófitas,
sedimentos) como sus propiedades físicas y químicas afectan las propiedades del perifiton.
Las rocas por ejemplo, dependiendo de su origen, pueden o no ser una fuente extra de
nutrientes para el perifiton como lo sugirieron Pizarro et al. (2004), pero son relativamente
más estables como sustrato desde un punto de vista físico que el sedimento o las macrófitas
(Vadeboncoeur & Lodge 2000). Las algas epifíticas pueden tomar nutrientes liberados por la
planta hospedadora o los nutrientes de la columna de agua en competencia directa con el
fitoplancton. Por otro lado, el epipelon, puede tomar el carbono inorgánico disuelto y los
nutrientes por difusión desde los sedimentos, donde las concentraciones intersticiales de
nutrientes son usualmente varios órdenes de magnitud mayores que en la columna de agua
(Blumenshine et al. 1997). La utilización de sustratos artificiales facilita el diseño experimental
ya que permite conocer con exactitud el tiempo de colonización de la comunidad. Además
facilita su cuantificación con el consiguiente beneficio en los análisis numéricos y estadísticos
posteriores, entre otras ventajas. Por otro lado, posibilita el análisis de la interacción exclusiva
de los organismos adheridos con la columna de agua, sacrificando a veces el realismo por
simplicidad y reproducibilidad (Kalff 2003).

68
La morfometría de los cuerpos de agua influye sobre la productividad primaria del perifiton
ya que es el principal determinante de la cantidad de hábitats disponibles para la colonización
de la comunidad. Por ejemplo, influye en el tipo y desarrollo de macrófitas que colonizarán el
cuerpo de agua y que servirán de sustrato al epifiton; en la disponibilidad de sustratos
bénticos, ya que solamente constituyen hábitats disponibles si reciben suficiente luz como
para permitir la fotosíntesis de la fracción algal. La importancia de la profundidad del cuerpo
de agua es altamente variable ya que los lagos someros pueden ser claros y estar dominados
por perifiton, pero por otro lado, pueden estar dominados por fitoplancton, sólidos en
suspensión o materia orgánica disuelta y por lo tanto, poseer una penetración de luz limitada
para el alcance del perifiton (Vadeboncoeur & Steinman 2002).
La disponibilidad de luz, temperatura, el contenido de nutrientes, la predación y las
variaciones en el nivel hidrométrico son factores que interactúan entre sí y afectan el
desarrollo y producción primaria de la comunidad perifítica (Rosemond et al. 2000, Cronk &
Mitsch 1994). En particular, la luz es un requisito fundamental para una existencia fototrófica
y cualquier variación en la cantidad y calidad de luz en el cuerpo de agua influye de manera
crucial en la fisiología, crecimiento y estructura de la comunidad perifítica (Hill 1996). El rol
de la luz puede estudiarse en diferentes escalas espaciales cuando se estudia la comunidad
perifítica. En la escala de paisaje, las características de la cuenca afectan la carga de nutrientes
y la cantidad de materia orgánica que ingresa al cuerpo de agua. Por ejemplo, un exceso de
nutrientes provenientes de la cuenca (por ejemplo como consecuencia de la actividad
agrícola) puede provocar floraciones algales del fitoplancton que alterarán, entre otros
factores, la cantidad y la calidad de luz que recibe el perifiton; del mismo modo, la materia
orgánica proveniente de la cuenca circundante puede afectar el color del agua (Vadeboncoeur
& Steinman 2002).
A una escala menor, la del cuerpo de agua, el fitoplancton posee una ventaja en
cuanto a la competencia por la luz con el perifiton, dado que está suspendido en el agua y de
esta manera puede sombrear a las comunidades fotótrofas que crecen debajo (Sand-Jensen &
Borum 1991). Más aún, dada la naturaleza sésil del perifiton, esta comunidad experimenta un
rango limitado de intensidades lumínicas dependiendo de la profundidad a la que se
encuentren los sustratos donde habita (Vadeboncoeur & Lodge 2000). De esta manera,
sufren los efectos de la variación en la cantidad y calidad de luz directamente. Algunos

69
organismos como las diatomeas con rafe, poseen movilidad dentro del sustrato. Esta
característica les permite regular su ambiente lumínico mediante algún mecanismo de
fototaxis, que les posibilita desplazarse de los extremos de un gradiente lumínico si la escala
de éste es de milímetros (Hill 1996). Por último, una vez que la luz llega a la comunidad
perifítica es atenuada aún más por las propias algas y partículas orgánicas e inorgánicas
presentes en el sustrato. Estos componentes de la matriz del perifiton absorben y dispersan la
luz, atenuándola aún más en una escala más pequeña y la magnitud de esta nueva reducción
depende del espesor del biofilm (Vadeboncoeur & Steinman 2002, Dodds et al. 1999).
Al igual que en el caso del fitoplancton, la respuesta fotosintética de la comunidad
perifítica algal es una función no lineal de la intensidad lumínica, de modo que la realización
de curvas P-I resulta de suma utilidad para conocer la respuesta ecofisiológica de la
comunidad ante las distintas irradiancias y condiciones ambientales (Hill 1996).
Antecedentes
La literatura acerca de estudios que involucran al perifiton es escasa cuando se la compara
con la concerniente al fitoplancton en humedales templados sudamericanos. En cuanto al
valle aluvial del río Paraná, entre los estudios que analizan aspectos estructurales del perifiton,
Tesolín & Tell (1996) publicaron la estructura del perifiton sobre sustratos naturales en una
laguna del tramo inferior del río. Por su parte, Pizarro (1999) analizó la variación espacial y
temporal de la biomasa del epifiton sobre Echinochloa polystachya en una laguna turbia del
Paraná inferior. Rodrigues & Bicudo (2001) estudiaron la composición específica del
perifiton sobre sustratos naturales y artificiales en distintos ambientes de Paraná superior.
Más recientemente, Leandrini & Rodrigues (2008) examinaron la variación temporal de la
biomasa perifítica sobre sustratos naturales en ambientes semilóticos del Paraná superior.
En cuanto a los estudios funcionales del perifiton, se vienen llevando a cabo estudios
de la productividad de la comunidad perifítica en lagunas de llanura de inundación desde
hace más de dos décadas. Apesteguía et al. (1978) analizaron la productividad primaria de
perifiton sobre sustratos artificiales en ambientes acuáticos del Paraná medio mediante la
determinación del carbono orgánico. Rai & Hill (1984, en Putz & Junk 1997) publicaron el
primer estudio cuantitativo de producción primaria de esta comunidad en un lago de la

70
llanura de inundación del río Amazonas. Desde entonces se ha realizado un gran número de
trabajos acerca de la funcionalidad del perifiton en distintos sustratos en este tipo de sistemas
(Moschini-Carlos et al. 2000, Putz 1997, Engle & Melack 1993, entre otros).
La Estructura del Perifiton en la RNO
La comunidad algal perifítica sobre sustratos naturales en la RNO que se estudió en el
capítulo I se caracterizó por mostrar dominancia de diatomeas epifíticas y/o bentónicas
adaptadas a un rango de condiciones ambientales similares (ej. alto pH, conductividad
elevada, tolerantes a cierto grado de desecación y elevado estado trófico). También se
encontraron bien representadas las clorofitas Stigeocloniom sp. y Oedogonium sp. y algunos
agregados de cianobacterias y formas cocoides y filamentosas de esta división. Las algas
presentes en el sustrato más conspicuo del humedal (Schoenoplectus californicus, junco) se
mostraron más afectadas por las variaciones en el nivel hidrométrico que aquellas adheridas a
las escamas de Ricciocarpus natans, para quienes la fluctuación de la intensidad de la irradiancia
fue una de las variables más importantes. En general, la composición específica de la fracción
algal del perifiton fue homogénea entre los ambientes de la RNO a excepción del ROL 2,
donde se encontraron pocos ejemplares en relación al profuso desarrollo de vegetación
emergente.
Las determinaciones de productividad y variables de masa en este capítulo se llevaron
a cabo enteramente en comunidades desarrolladas sobre sustratos artificiales debido a que
éstos permiten, entre otras ventajas, homogeneizar los tiempos de colonización y estandarizar
a una superficie similar y conocida las variables de medición. Por otro lado, el sustrato
artificial, al ser inerte en cuanto a su captación de carbono nos asegura que las tasas de
asimilación estimadas correspondan al perifiton. Finalmente, y dado que la composición algal
en los sustratos naturales fue similar a aquella desarrollada sobre los sustratos artificiales,
consideramos que nuestros resultados son extrapolables a la comunidad natural.
El trabajo realizado y que se describe en este capítulo se llevó a cabo enteramente en un área
de la Laguna Grande, representativa de la transición entre las zonas pelágica y litoral. La
laguna se encontró durante todo el período de estudio en un estado de aguas turbias

71
(Scheffer et al. 1993), caracterizado por altas densidades de fitoplancton. En trabajos previos,
se describe que la laguna se encontraba cubierta en gran parte de su superficie por macrófitas
flotantes y presentaba densidades más bajas de fitoplancton (Izaguirre et al. 2004, O´Farrell et
al. 2003). En base a estos antecedentes, nos interesaba saber cómo variaba la productividad
del fitoplancton y perifiton a lo largo del año, en este sistema sujeto a fuertes cambios
hidrométricos y de estacionalidad marcada. También se analizaron los patrones observados
en la productividad de ambas comunidades en relación a su estructura y a las variables
ambientales. Por último, realizamos estimaciones comparativas de la contribución relativa de
estas comunidades a la productividad primaria microbiana del sistema.
Hipótesis
I) La estacionalidad y la altura hidrométrica afectan la dinámica anual de la producción
primaria de ambas comunidades;
II) Las variaciones observadas en los patrones de productividad del fitoplancton y del
perifiton estarán acompañados de cambios en la estructura de las comunidades;
III) La productividad del perifiton tendrá un patrón anual inverso al del fitoplancton;
IV) La contribución del perifiton a la producción primaria microbiana de la laguna será
menor que la del fitoplancton por encontrarse la laguna en un estado de aguas turbias
dominado por fitoplancton.
Objetivos
- Analizar la estructura del fitoplancton y del perifiton;
- Estudiar la fluctuación anual de los parámetros fotosintéticos del fitoplancton y perifiton
sobre sustratos artificiales;

72
- Estudiar la variación temporal de la producción fitoplanctónica por unidad de área
integrada en la columna de agua y del perifiton por unidad de área de sustrato;
- Comparar los parámetros fotosintéticos de ambas comunidades en relación a la
estacionalidad y el nivel hidrométrico;
-Analizar las variables mencionadas en los objetivos previos en relación a la estructura de la
comunidad y las variables ambientales;
- Estimar el aporte de la productividad de ambas comunidades a la producción primaria
microbiana de la Laguna Grande.
MATERIALES Y MÉTODOS
El trabajo se llevó a cabo en la Laguna Grande de la Reserva Natural Otamendi durante los
años 2005, 2006 y 2007 con distinta frecuencia de toma de muestras. Las muestras de
fitoplancton que se tomaron en una primera etapa (año 2005) tuvieron frecuencia estacional
(4 muestreos) en tanto que en una etapa posterior, se llevaron a cabo muestreos mensuales
desde abril 2006 hasta marzo 2007. Las muestras de fitoplancton fueron tomadas con un
balde integrando los primeros 40-50 cm de profundidad. El perifiton sobre sustratos
artificiales, tuvo un esquema de muestreos que involucró un ensayo piloto en invierno de
2005 y muestreos mensuales desde febrero 2006 hasta marzo 2007. Se delimitó un área de 25
m2 como zona de muestreo en un sector representativo de la transición entre la zona pelágica
y litoral de la laguna en donde se colocaron los dispositivos para la colonización del perifiton
y se tomaron las muestras de fitoplancton y de agua para análisis químicos y la determinación
de parámetros abióticos in situ.
Variables ambientales
Se midieron en el campo: el pH, la conductividad y la temperatura con un sensor de campo
Hanna HI 991301 y el oxígeno disuelto con un sensor portátil Hanna HI 9143. El porcentaje

73
de saturación de oxígeno con respecto a la temperatura se obtuvo a partir del nomograma de
Mortimer (Wetzel 1981). La profundidad del sitio de muestreo se registró con una vara
graduada en uno de los pilotes del muelle de la laguna ubicado a no más de 2 m del punto de
muestreo. Los datos de precipitación y altura hidrométrica del río Paraná de las Palmas a la
altura de Las Rosas (Fig. 6, Área de Estudio, página 21) fueron cedidos por la Estación
Experimental Agropecuaria Delta del Paraná (INTA). Se registró la intensidad de la radiación
subacuática comprendida entre 400 y 700 nm (RFA, radiación fotosintéticamente activa) con
un radiómetro Li-Cor (Lincoln, Nebraska, USA) equipado con un sensor esférico modelo Li-
193SA. La intensidad de RFA se registró a intervalos de profundidad de 5 cm con el objeto
de calcular el coeficiente de extinción vertical, Kd (Kirk 1994). Se calcularon las
profundidades ópticas (⦆) de acuerdo a Kirk (1994) como ζ = Kd z (z es la profundidad). La
profundidad óptica correspondiente al 1 % de la irradiancia incidente en superficie es igual a
4,6; por lo tanto en toda zona eufótica habrá como máximo 4,6 profundidades ópticas. Una ⦆
menor a 4,6 implica que toda la columna de agua corresponde a capa eufótica en tanto que
una mayor, implica que la profundidad de la capa eufótica es menor que la profundidad total
del sitio (Falkowski & Raven 2007). Se estimó la irradiancia media integrada en la columna de
agua como:
Imedia = I0 (1 - e-Kd z) (Kd z)-1
donde I0 es la irradiancia subsuperficial, z la profundidad del sitio y Kd el coeficiente de
atenuación vertical (Helbling et al. 1994, Neale et al. 1991). Con el objeto de obtener un perfil
Irradiancia vs. profundidad discriminado por longitudes de onda y calcular los Kd por │, en
el muestreo correspondiente al mes de mayo se tomaron datos espectrales en la laguna con
un espectroradiómetro marca Oceanics. Las mediciones fueron efectuadas in situ por el Dr.
Gonzalo Pérez del Instituto Tecnológico de Chascomús (INTECH).
Los análisis de nutrientes, contenido húmico, sólidos en suspensión y clorofila a del
fitoplancton se realizaron a partir muestras de agua integradas y se colocaron en botellas
plásticas previamente enjuagadas con HCl 2 %, que fueron transportadas al laboratorio de
Limnología (FCEyN, UBA) en frío y oscuridad. Durante el año 2005 se empleó la
absorbancia del agua a 254 nm como estimador del contenido húmico del agua (Kronberg

74
1999) y la filtración de la muestra se realizó a través de filtros de fibra de vidrio de 0,7 µm de
tamaño de poro. El contenido de ácidos húmicos se estimó en 2006 y 2007 a través del
coeficiente de absorción a 440 nm (g440) (Kirk 1994). Para ello, se midió la absorbancia a
440 nm de agua de cada sitio previamente filtrada por un filtro de policarbonato de 0,2 µm.
También se determinó la absorbancia del agua filtrada a 320 nm y se calculó el coeficiente de
absorción a esa longitud de onda (a320, Williamsom et al. 1999).
Las fracciones disueltas de los nutrientes se determinaron luego de la filtración de la
muestra a través de un filtro de fibra de vidrio Millipore® de 0,7 µm de tamaño de poro. Para
la determinación del fósforo disuelto (P-disuelto) se utilizó el método del cloruro estañoso,
para el N-nitrato+nitrito la reducción con cadmio, para el sílice el método del molibdosilicato
(APHA 2005) y para la determinación del N-amonio se empleó el método del salicilato. Estas
determinaciones se realizaron empleando un espectrofotómetro Hach® DR/2010 y los
respectivos reactivos Hach®. Las fracciones totales de nitrógeno y fósforo se estimaron con
la metodología empleada para el N-nitrato+nitrito y P-disuelto previa digestión conjunta de
los compuestos de N y P en la muestra (Grasshoff et al. 1983). La concentración de sólidos
en suspensión se determinó de forma análoga a la descrita en el capítulo I (página 30). La
alcalinidad se determinó a través de la titulación de la muestra sin filtrar con HCl 0,1 N el
mismo día de la salida de campo (Mackereth et al. 1978). La concentración de carbono
inorgánico disuelto (CID) se estimó a partir de la alcalinidad, pH y temperatura (Stumm &
Morgan 1996).
Fitoplancton
Para el reconocimiento de los especímenes se empleó la bibliografía taxonómica detallada en
la página 33 del capítulo I. Los análisis cuantitativos se realizaron a partir de muestras
obtenidas en cada sitio de muestreo en botellas plásticas que se fijaron con solución de lugol
acético a una concentración final de 1 %. Los recuentos se realizaron en microscopio
invertido por el método de Utermöhl (1958) con un error del 20 % (Venrick 1978). A partir
de agosto de 2006 se estimó la concentración de ficocianina y ficoeritrina en el agua con un
fluorómetro marca Cyclops (Turner designs) provisto de los sensores correspondientes. La
concentración de clorofila a se estimó a través de la filtración en oscuridad de un volumen

75
conocido de muestra a través de filtros de fibra de vidrio Millipore® de 0,7 µm de poro y se
procedió como figura en la página 31.
Las estimaciones de productividad primaria se llevaron a cabo empleando la metodología de
asimilación de 14C descrita en Holm-Hansen & Helbling (1995). Durante 2005 se llevaron a
cabo curvas de fotosíntesis-irradiancia (P-I) y curvas de fotosíntesis en función de la
profundidad (P-P). Las estimaciones de producción integrada en la columna (PA) realizadas a
partir de los dos tipos de procedimientos no mostraron diferencias significativas (test de t, p
= 0,52). Considerando que durante la instalación de los dispositivos para realizar la curva P-P
se provocaba la resuspensión de los sedimentos, difícil de controlar ya que el fondo de la
laguna es muy poco compacto y sumado esto al escaso perfil de la columna de agua que no
permitía tomar muchos de puntos en profundidad, decidimos luego estimar la PA de agua a
partir de la curva P-I.
Para efectuar la estimación de PA a partir de la curva P-I consideramos el Kd y la
irradiancia para obtener la profundidad correspondiente a cada irradiancia de la curva P-I y
luego integrar el área bajo la curva de P vs z.
Curva P-I
Las muestras de fitoplancton se colocaron en tubos de acrílico transparente (corte neto a 400
nm) de 67,5 mL. Se emplearon además dos tubos oscuros para estimar la incorporación de
14C por procesos no fotosintéticos y cuyo valor de asimilación fue restado al de los tubos
transparentes. A cada tubo se le agregó 1 µCi de NaH14CO3 (Perkin Elmer Life Sciences,
Inc., USA) en la mayor oscuridad posible. Los tubos se colocaron a incubar en un
contenedor plástico de 1,55 m por 1,20 m y 0,3 m de profundidad. En el interior del
contenedor se colocó una estructura con grampas plásticas para el sostén de los tubos. El
incubador se llenó con agua corriente y se controló la temperatura del dispositivo a intervalos
regulares de tiempo. Se utilizaron mallas de atenuación neutra de distinto espesor para lograr
distintas atenuaciones entre el 0 % y el 100 % de la radiación solar incidente en superficie
(Fig. 1). Se expuso un par de tubos a cada atenuación lumínica. En cada curva P-I se
utilizaron como mínimo entre 6 a 7 atenuaciones. Las incubaciones se realizaron por dos
horas alrededor del mediodía en días soleados y de poca cobertura nubosa. Se registró la luz

76
incidente en superficie durante el período de incubación. Se utilizó el sensor esférico Li-Cor
apoyado sobre una superficie negra para evitar la reflexión y sobreestimación de la radiación
incidente. Finalizada la incubación se tomó el promedio de la radiación durante el período de
incubación.
Fig. 1. Dispositivo empleado para la incubación de las muestras para la realización de las
curvas P-I. Se señalan las mallas neutras (atenuaciones) y la ubicación de los tubos en el
incubador.
La toma de muestras en la Laguna Grande para todas las curvas P-I se efectuó en la
proximidad del muelle (Fig. 2), con la excepción del muestreo correspondiente a diciembre
2005. En esta oportunidad, una sequía extrema nos obligó a tomar el agua de un sitio más
interno en la laguna, aproximadamente a 30 metros del área original (Fig. 3). Sin embargo,
consideramos este punto representativo del sitio de muestreo original ya que los sitios
ubicados en la transición de las zonas pelágica y litoral en la Laguna Grande poseen
características físicas y químicas muy similares entre sí ocurriendo lo mismo con el
fitoplancton (Izaguirre et al. enviado, de Tezanos Pinto 2008).

77
Fig. 2. Vista de la Laguna Grande desde la orilla. La flecha indica aproximadamente la zona
de muestreo.
Fig. 3. Vista de la zona de muestreo (diciembre 2005) donde se observa el bajo nivel
hidrométrico de la Laguna Grande.

78
Curva P-P
Este tipo de análisis, que se realizó durante 2005, implicó la instalación de un dispositivo que
permite el sostén de los tubos en sentido horizontal en el mismo cuerpo de agua (Fig. 4). Se
emplearon los mismos tubos que en el caso de la curva P-I. Se tomaron muestras a tantas
profundidades como lo permitía el nivel del agua; cada 2,5 cm aproximadamente. Se llenaron
los tubos con agua proveniente de cada profundidad y se les inoculó el radiactivo como se
describió para el caso de la curva P-I. Los tubos, dos por profundidad, se incubaron durante
dos horas en la laguna. Se incubaron dos tubos oscuros cuyo valor de asimilación fue restado
al de los transparentes. Durante el período de incubación se tomaron mediciones de luz
incidente como se describió más arriba.
Finalizado el período de incubación los tubos provenientes de los dos tipos de experiencias
(curva P-I y P-P), fueron transportados en frío y oscuridad al laboratorio de 14C y 3H del
Departamento de Química Biológica de la FCEyN donde fueron procesados dentro de las 2
hs posteriores al final del período de incubación. En diciembre de 2005, fecha de extrema
sequía en el humedal, estaba programada una curva P-P que no pudo efectuarse por
cuestiones logísticas.
Fig. 4. Dispositivo con tubos (flecha) utilizado para incubar las muestras en la profundidad
de donde fueron extraídas para la realización de las curvas P-P.

79
Trabajo de Laboratorio
Los tubos provenientes de ambos tipos de experiencias fueron procesados de igual manera.
En primer lugar se procedió a extraer 1 mL de cada muestra, que fue colocado en viales de
centelleo líquido marca Polistor® a los que previamente se les había adicionado 3 gotas de
NaOH 0,1 N para evitar posibles pérdidas de carbono inorgánico marcado por difusión a la
atmósfera. Este vial fue utilizado para la determinación de la actividad de carbono 14
específica en cada muestra. Luego se le adicionó el líquido de centelleo (Opti-phase Hi Safe
3, Perkin Elmer, Life Sciences, Inc., USA) y se determinó la actividad de los viales en un
contador de centelleo líquido Beckman LS-6500 de la FCEyN.
Un volumen conocido de cada tubo (40 mL) fue filtrado en condiciones de luz tenue
a través de filtros de fibra de vidrio Millipore® de 0,7 µm de tamaño de poro. Los filtros
fueron colocados en viales de centelleo líquido que se colocaron en una atmósfera clorhídrica
durante una noche. Luego fueron ventilados bajo campana y se dejaron secar. Una vez secos,
se les adicionó el líquido de centelleo, se mezclaron bien y se dejaron reposar una hora antes
de ser contados en el contador de centelleo líquido durante 1 minuto por muestra.
Cálculo de las Tasas de Asimilación
Las tasas de asimilación se obtuvieron a partir de la siguiente fórmula (Tilzer 1984, Pregnall
1991, Holm-Hansen & Helbling 1995, Wetzel & Likens 2000):
µg C (µg Chl a h)-1 = DPMinc CID 1,06 (DPMad Tinc Chl a)-1 (1),
donde en el numerador, DPMinc es la radiactidad incorporada por cada muestra, CID es la
concentración de carbono inorgánico disuelto (en µg C L-1) y 1,06 es el factor de
discriminación isotópica que considera la velocidad de los procesos enzimáticos involucrados
en la fotosíntesis, ya que el 14C es incorporado aproximadamente un 6 % menos rápido que
el 12C. En el denominador, DPMad es la radiactividad del carbono adicionado, Tinc la
cantidad de horas de incubación y Chl a es la concentración de clorofila a (µg L-1). Tanto a
DPMinc como a DPMad se les efectuaron las correcciones volumétricas correspondientes:

80
DPMad = DPMmL V (X)-1 (2)
DPMinc = DPMfiltro V (W)-1 (3)
donde V es el volumen de la muestra (67,5 mL), X es el volumen en el que se determinó la
actividad adicionada a la muestra (1 mL) y W son los mL filtrados (Tilzer 1984).
Los valores de asimilación así obtenidos fueron posteriormente ajustados a la
ecuación de Eilers & Peeters (1988):
P = I (aI2 + bI + c)-1
donde P es la productividad primaria (µg C (µg Chl a h)-1), I es la irradiancia (µmol fotones m-
2 s-1) y a, b y c son parámetros del ajuste. Los parámetros fueron obtenidos por iteraciones
sucesivas mediante la técnica de cuadrados mínimos, empleando el algoritmo de Levenberg-
Marquardt.
La pendiente inicial de la curva P-I, "プ" (µg C (µg Chl a h)-1 µmol fotones m2 s), la tasa
máxima de asimilación, Pmáx (µg C (µg Chl a h)-1), la irradiancia al inicio de la saturación, Ik
(µmol fotones m-2 s-1) y la irradiancia óptima, Iopt (µmol fotones m-2 s-1) pueden expresarse,
diferenciando la ecuación de Eilers & Peeters (1988), como sigue:
プ = 1 / c
Pmáx = 1 / (b + 2 (ac)1/2)
Ik = c / (b + 2 (ac)1/2)
Iopt = (c / a)1/2
El parámetro Ik también puede obtenerse como Ik = Pmáx / プ (Talling 1957). Para estimar el
estado de limitación lumínica de la comunidad, se compararon los valores de Ik de cada
comunidad con la correspondiente Imedia del día de muestreo alrededor del mediodía. Si los
valores de Ik de la comunidad eran mayores a la Imedia del cuerpo de agua alrededor del
mediodía (que se consideró como momento de máxima irradiancia de ese día), la comunidad
de consideró limitada lumínicamente (Torremorell et al. 2008, Knoll et al. 2003). De este

81
modo, calculando la relación Imedia/Ik, en caso de obtener valores menores a 1, se puede
decir que la comunidad estaba limitada por luz, ya que estaba recibiendo en el momento del
día de mayor irradiancia, valores de radiación por debajo de la intensidad con la que la
fotosíntesis deja de estar limitada por luz.
Se calculó para el fitoplancton la producción máxima por unidad de volumen (Pmáx
vol) como el producto del número de asimilación, Pmáx por la concentración de clorofila a
(Krause-Jensen & Sand-Jensen 1998). Esta variable permite estimar la máxima producción
del fitoplancton por unidad de volumen a una sola profundidad, a diferencia de la
producción integrada en la columna de agua (PA), que involucra toda la capa productiva.
Producción integrada en la columna de agua (PA)
Para la estimación de las tasa de PA (mg C m-2 h-1) se calculó el área bajo la curva de
producción primaria (en mg m-3 h-1) en función de la profundidad (m) (Wetzel & Likens
2000). Los cálculos integrales se efectuaron con un programa computacional. La profundidad
se obtuvo despejando a partir de la irradiancia correspondiente en la curva P-I y la ecuación
de penetración de la irradiancia en la columna de agua (Kirk 1994):
Iz = I0 e –Kd z (4)
donde Iz es la irradiancia a la profundidad z, I0 es la irradiancia subsuperficial, Kd el
coeficiente de atenuación vertical y z la profundidad (Kirk 1994).
Se llevaron las tasas de PA por hora a valores diarios a partir de la irradiancia máxima
medida el día de muestro alrededor del mediodía y los datos de salida y puesta de sol
provistos por el Servicio de Hidrografía Naval. Se empleó la siguiente fórmula para
aproximar la irradiancia cada 15 minutos entre la salida y puesta de sol el día de muestreo
(McBride 1992):
It = Imáx seno (π (t – tsalida)/ Δt)
donde It es la irradiancia a la hora t, Imáx es la irradiancia máxima medida alrededor del
mediodía, tsalida corresponde a la hora de salida del sol y Δt es la diferencia entre la hora de

82
puesta y salida del sol para el día de muestro. Si bien esta curva supone que la irradiancia se
distribuye en forma de sinusoide perfecta a lo largo del día es probable que haya variaciones
en la mayor parte de las situaciones prácticas (Kirk 1994). Sin embargo, constituye una buena
aproximación en nuestro caso para poder hacer una estimación de la PA diaria ya que
contamos con la irradiancia máxima de cada día de muestreo en la RNO.
Se estimó la irradiancia cada 15 minutos para el día de muestreo, luego se promedió
para un intervalo de tiempo de dos horas y se calculó la producción primaria a través de la
ecuación de Eilers & Peeters (1988) cada 5 cm de profundidad. La irradiancia en el agua se
obtuvo a partir de la relación entre la irradiancia subsuperficial y la del aire para el día del
muestreo. Luego, aplicando la ecuación de penetración de la luz en profundidad (4) se
obtuvo la irradiancia a cada z (Kirk 1994). Se sumaron las producciones para los intervalos de
dos horas a cada profundidad en cada día de muestreo. La PA se obtuvo integrando P vs z
como se describió más arriba. Los valores de producción diarios se multiplicaron luego por el
número de días de cada mes y se sumaron, obteniendo de esta manera una estimación anual
de la producción fitoplanctónica por unidad de área para la zona de muestreo.
Perifiton
Las determinaciones de clorofila a, fracciones de masa, estructura y productividad de la
comunidad perifítica se llevaron a cabo sobre sustratos artificiales. Los sustratos consistieron
en rectángulos de policarbonato de 7,3 cm de largo por 1,63 cm de ancho y 1 mm de espesor.
En la Fig. 5 se muestra el dispositivo de acrílico transparente utilizado para el sostén de los
sustratos artificiales y la instalación del sistema de flotación (sujeto a la fluctuación
hidrométrica de la laguna) en la Laguna Grande. Los sustratos se incubaron a una
profundidad de 10 cm en cada oportunidad.

83
Fig. 5. Izquierda: dispositivo para el sostén de los sustratos artificiales (perifitómetro).
Derecha: instalación del dispositivo con su sistema de flotación (flecha) en la Laguna Grande.
Para la estimación del tiempo en que la comunidad alcanza su máximo valor de
biomasa fotosintética, se llevó a cabo una experiencia piloto en septiembre de 2004 en la que
se colocaron a colonizar sustratos y se les determinó a tres de ellos la concentración de
clorofila a una vez por semana durante 45 días. Se estimó que se necesitan por lo menos 20
días para tener una comunidad relativamente madura en primavera. Por lo tanto, se estimó
que un mes sería suficiente para tener una comunidad madura en cualquier momento del año.
Durante el período de estudio el tiempo de colonización varió entre 24 y 35 días.
Para la determinación taxonómica de las algas se empleó la bibliografía citada en la
página 33 del capítulo I. En análisis cualitativo se llevó a cabo a partir del raspado del material
proveniente de tres sustratos y su observación en el microscopio óptico. Para el recuento de
los organismos se tomaron tres sustratos, que fueron raspados y llevados cada uno a un
volumen conocido de agua. Luego se les agregó lugol acético a una concentración final de 1
%. Los recuentos se realizaron en microscopio invertido por el método de Utermöhl (1958)
con un error del 20 % (Venrick 1978), llevándose los valores de densidad a cm2 de sustrato
colonizado. La concentración de clorofila a de la comunidad perifítica se obtuvo empleando
la misma metodología descrita para el fitoplancton previo raspado del material adherido al
sustrato con un elemento filoso. Dada la variabilidad de la comunidad perifítica, en cada
oportunidad se tomaron tres sustratos artificiales (réplicas).

84
Se estimaron por triplicado las fracciones de masa peso seco, cenizas y peso seco libre
de cenizas a partir de otros sustratos artificiales. Para ello, se colocaron filtros de fibra de
vidrio Millipore® de 0,7 µm de tamaño de poro en la mufla a 500 °C durante 3 horas y luego
se les determinó el peso. Estos filtros se usaron para filtrar el material proveniente de los
sustratos el mismo día de la toma de muestras. Los filtros con el material perifítico retenido
se llevaron a estufa (60-70 °C), se les determinó su peso seco (PS) y luego se los llevó
nuevamente a la mufla (500 °C, 3 hs) y se volvieron a pesar para obtener el peso de las
cenizas. Por diferencia entre el PS y las cenizas (C) se obtuvo el peso seco libre de cenizas
(PSLC) (APHA 2005). Con estos datos se calculó el índice autotrófico (IA) como el cociente
entre el PSLC y la concentración de clorofila a perifítica. Este cociente informa sobre si la
composición de la comunidad es más autotrófica o heterotrófica: valores superiores a 200
indican una alta proporción de organismos heterotróficos, no clorofílicos y detritus orgánico
(APHA 2005, Lowe & Pan 1996). Además, la comunidad se clasificó de acuerdo al sistema
de Lakatos (1989) en referencia a su peso seco, contenido de cenizas y concentración de
clorofila a siguiendo el esquema que figura en la Tabla 1.
TIPO PESO SECO (PS) (mg cm-2)I perifiton de alta biomasa > 4 II perifiton de biomasa media 2-4 III perifiton de baja biomasa < 2
CENIZAS (C) (%) I perifiton inorgánico > 75 II perifiton inorgánico-orgánico 50-75 III perifiton orgánico-inorgánico 25-50 IV perifiton orgánico < 25
CLOROFILA a (%) I perifiton autotrófico > 0,6 II perifiton auto-heterotrófico 0,25-0,6 III perifiton hetero-autotrófico 0,1-0,25 IV perifiton heterotrófico < 0,10
Tabla 1. Clasificación de la comunidad perifítica propuesta por Lakatos (1989).
Para la realización de las curvas P-I de perifiton se utilizaron los mismos dispositivos
experimentales de incubación que en el caso del fitoplancton y la metodología seguida fue
similar utilizando los sustratos artificiales colonizados. Cada sustrato que fue colocado en un
tubo para su posterior incubación tenía identificada una de sus caras con una marca inocua.

85
Esa marca permitió reconocer la cara que sería expuesta directamente a la radiación mientras
que a la otra se le eliminó la mayor cantidad de material posible con un elemento filoso en el
campo. Posteriormente los sustratos se introdujeron en los tubos de incubación con agua
proveniente del sitio de muestreo previamente filtrada con red de 15 µm de poro. A cada
tubo se le adicionaron 3 µCi de NaH14CO3. Luego de la incubación (2 hs) las muestras
fueron trasladadas al laboratorio en frío y oscuridad para su posterior procesamiento en el
laboratorio de 14C y 3H del Departamento de Química Biológica de la FCEyN. El
procesamiento posterior del material fue el mismo que se describió en el caso del
fitoplancton previo raspado y filtrado del material de la cara del sustrato expuesta a la luz.
Para obtener las tasas de asimilación se empleó la fórmula (1) descripta para el
fitoplancton, pero utilizando en (3) la superficie colonizada raspada en lugar del volumen
filtrado:
DPMinc = DPMfiltro V (superficie colonizada raspada)-1
También se ajustaron los datos de productividad de perifiton a la ecuación de Eilers &
Peeters (1988). Para analizar el estado de limitación lumínica de la comunidad, se calculó la
relación entre la irradiancia correspondiente a la profundidad donde fueron colonizados los
sustratos (I10 cm) alrededor del mediodía el día de muestreo y el Ik de la comunidad. Valores
del cociente I10 cm/Ik menores a 1 indican limitación lumínica de manera análoga al
fitoplancton.
Para comparar las tasas de producción por unidad de área de sustrato entre las
distintas fechas se estimó la producción por unidad de área de sustrato a los 10 cm de
profundidad (PA 10 cm) a partir de la ecuación de Eilers & Peeters (1988), y la ecuación de
penetración de la irradiancia en profundidad (4) (Kirk 1994). Estos valores fueron llevados
luego a tasa diaria y anual de manera análoga al caso del fitoplancton.
Dado que la Laguna Grande es un sistema de características lumínicas particulares, los
valores de irradiancia óptima (Iopt) de ambas comunidades en general se encontraron por
encima de la irradiancia recibida por cada una de ellas (Imedia en el caso del fitoplancton e I10
cm en el caso del perifiton). Las consideraciones acerca del estado de fotoaclimatación y de

86
una posible limitación lumínica de los organismos se efectuaron entonces considerando el
valor de Ik en cada oportunidad. Del mismo modo, no se analizó la fotoinhibición por altas
irradiancias en las curvas P-I ya que las irradiancias encontradas en el ambiente fueron en
general menores a las correspondientes a la zona de fotoinhibición (APÉNDICE I).
Análisis de datos
Se calcularon los coeficientes de variación de las variables analizadas y se llevaron a cabo
correlaciones no paramétricas (Spearman) entre todas las variables. Se efectuaron también
regresiones múltiples entre las variables biológicas de ambas comunidades y las variables
ambientales. Para este análisis se utilizó el método de selección de variables de a pasos
sucesivos ("forward stepwise"). En el caso del fitoplancton (N = 14) se tuvieron en cuenta las
siguientes variables como independientes: profundidad, pH, temperatura, Imedia, NID y P
disuelto. Para el caso del perifiton (N = 12) se empleó la conductividad, temperatura,
Irradiancia a los 10 cm, abs 254 nm, NID, P disuelto, Si y clorofila a fitoplanctónica. En los
casos que fue necesario, se transformó la variable dependiente para cumplir el supuesto de
normalidad empleando Log10. Se efectuó una regresión curvilínea entre Pmáx vol y la
concentración de clorofila a del fitoplancton. En las regresiones se empleó el Modelo II de
regresión (Sokal & Rohlf 1979). También se efectuaron ANOVA no paramétricos: test de
Kruskal-Wallis (K-W) para comparar más de dos muestras independientes y Mann-Whitney
(M-W) para comparar dos muestras independientes. En todos los casos se consideró un valor
de p < 0,05 como significativo (Zar 1996).
RESULTADOS
Variables ambientales
En la Tabla 2 se muestran los promedios y rangos de variación de las variables ambientales
estudiadas. Durante el período de estudio la laguna presentó algunos manchones aislados y
fluctuantes de macrófitas flotantes en la zona litoral, sin alcanzar una cobertura importante.
Se observó también vegetación palustre del tipo junco cercana al área de muestreo. El nivel
del agua fue bajo en la mayor parte del período estudiado (Fig. 6 a), y en pocos casos superó
los 0,5 m de profundidad: febrero y fines de agosto de 2005, marzo, septiembre y noviembre

87
de 2006 y marzo de 2007. Los valores de profundidad no difirieron estacionalmente (K-W, p
= 0,22). Las precipitaciones acumuladas durante el mes previo al muestreo fueron máximas
en el mes de marzo 2006 (353 mm), mientras que fueron mínimas en mayo del mismo año (7
mm) (Fig. 6 a). La profundidad se correlacionó de manera positiva con las precipitaciones
acumuladas durante el mes previo al muestreo (Tabla 3) pero no se correlacionó con la altura
hidrométrica del río Paraná a la altura del arroyo Las Rosas (Tabla 4), lo que sugiere que las
precipitaciones pluviales regularon la altura hidrométrica de la laguna. El coeficiente de
variación (CV) de la profundidad fue de 39 % y el mismo valor se obtuvo para la
temperatura, que siguió un patrón estacional marcado, siendo la máxima registrada de 29,8
°C en diciembre 2005 y la mínima de 5,8 °C en julio 2006 (Fig. 6 b). Los valores de esta
variable difirieron significativamente entre estaciones del año (K-W, p = 0,001). Por su parte,
los valores máximos de oxígeno disuelto los constituyen los picos observados en febrero
2005 y noviembre 2006 (14,2 y 14,3 mg L-1 de O2, respectivamente). En estas oportunidades
se observó sobresaturación de oxígeno en el agua y lo mismo ocurrió en diciembre 2005.
Cabe destacar que en estas tres oportunidades se observaron floraciones de cianobacterias
filamentosas en la laguna. Los valores más bajos fueron medidos en febrero de 2006 (3,43 mg
L-1 de O2) y marzo de 2007 (3,57 mg L-1 de O2) (Fig. 6 c). La concentración de oxígeno
disuelto mostró durante 2006 una tendencia general a presentar valores más altos en los
meses más fríos, sin embargo no se observó una correlación significativa entre esta variable y
la temperatura ni diferencias en los valores entre estaciones del año considerando todos los
datos. Por otro lado, si se tienen en cuenta sólo las mediciones efectuadas en 2006 y 2007, la
concentración de oxígeno disuelto se mostró correlacionada de manera negativa (R = -0,71;
N = 12) con la temperatura y presentó valores mayores en otoño e invierno (K-W, p =
0,009).
Los valores de pH del agua fueron altos y se mantuvieron alrededor de 8 durante el
estudio. Esta variable fue la que presentó el menor CV en el estudio (6 %). Los máximos
fueron 9,4 y 9,5 en febrero de 2005 y noviembre de 2006; mientras que los mínimos hallados
fueron 7,9 y 7,8 en marzo y abril de 2006 respectivamente (Tabla 2). El pH se correlacionó
de manera positiva con la concentración de clorofila a fitoplanctónica y el oxígeno disuelto,
lo que podría estar indicando que las variaciones en el pH podrían ser debidas en gran parte a
la fotosíntesis de las algas (Tabla 3). La alcalinidad fue elevada siendo en promedio mayor en

88
2005 (9 ± 4 mEq L-1) que en 2006 y 2007 (valor medio: 5,6 ± 1,5 mEq L-1). En el último
período se mantuvo relativamente constante, sin mostrar patrones temporales definidos. Su
valor máximo coincidió con la sequía de la laguna en diciembre de 2005 (15 mEq L-1, Tabla
2). Los valores de conductividad en general fueron elevados. El valor mínimo fue 1000 µS
cm-1 y se registró en abril de 2006. El valor más alto se midió en diciembre 2005 (4500 µS
cm-1), producido por un posible efecto de concentración debido al bajo nivel hidrométrico.
Cabe destacar la gran variación que se registró en esta variable en el mes de agosto 2005. El
15 de agosto de 2005 se midió un valor de conductividad de 3000 µS cm-1 y el 30 de agosto el
valor disminuyó a 1440 µS cm-1 debido a una fuerte lluvia ocurrida en los días previos al
muestreo de fines de agosto. Esta disminución en la conductividad fue acompañada por un
incremento de 20 cm en la profundidad. En el año 2006 los valores de conductividad
tendieron a ser más altos en la primavera (Fig. 6 d).
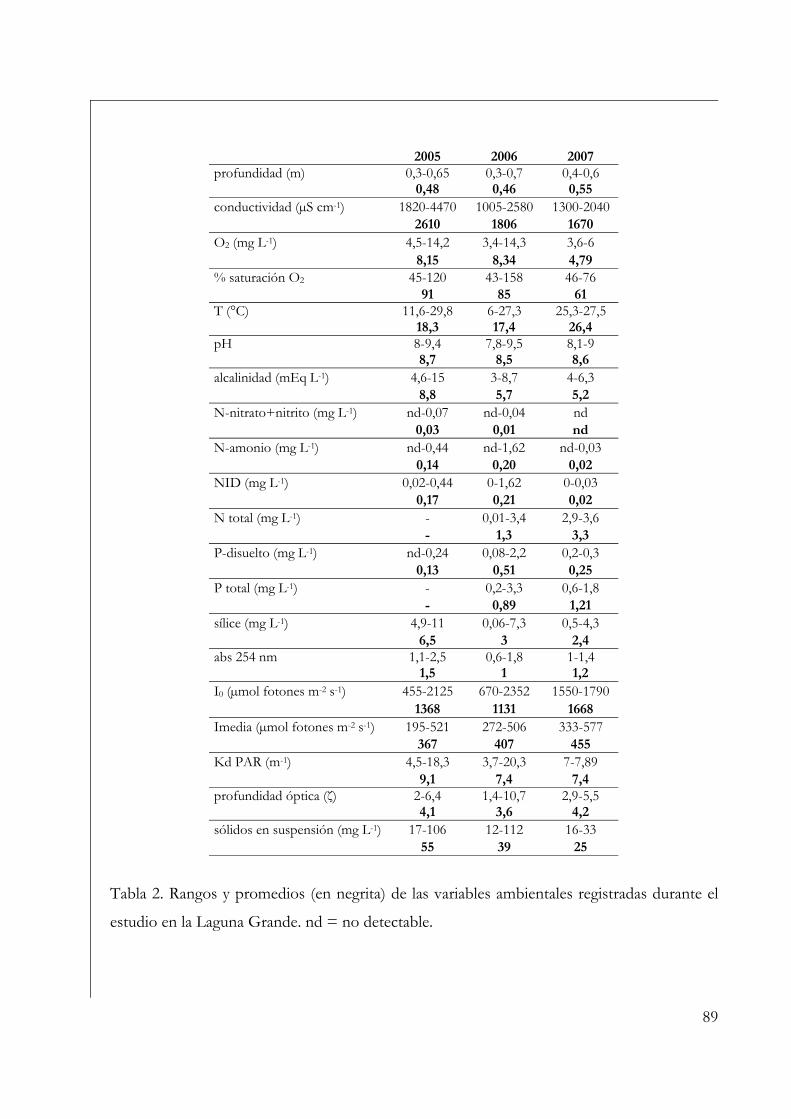
89
2005 2006 2007 profundidad (m) 0,3-0,65 0,3-0,7 0,4-0,6 0,48 0,46 0,55 conductividad (µS cm-1) 1820-4470 1005-2580 1300-2040 2610 1806 1670 O2 (mg L-1) 4,5-14,2 3,4-14,3 3,6-6 8,15 8,34 4,79 % saturación O2 45-120 43-158 46-76 91 85 61 T (°C) 11,6-29,8 6-27,3 25,3-27,5 18,3 17,4 26,4 pH 8-9,4 7,8-9,5 8,1-9 8,7 8,5 8,6 alcalinidad (mEq L-1) 4,6-15 3-8,7 4-6,3 8,8 5,7 5,2 N-nitrato+nitrito (mg L-1) nd-0,07 nd-0,04 nd 0,03 0,01 nd N-amonio (mg L-1) nd-0,44 nd-1,62 nd-0,03 0,14 0,20 0,02 NID (mg L-1) 0,02-0,44 0-1,62 0-0,03 0,17 0,21 0,02 N total (mg L-1) - 0,01-3,4 2,9-3,6 - 1,3 3,3 P-disuelto (mg L-1) nd-0,24 0,08-2,2 0,2-0,3 0,13 0,51 0,25 P total (mg L-1) - 0,2-3,3 0,6-1,8 - 0,89 1,21 sílice (mg L-1) 4,9-11 0,06-7,3 0,5-4,3 6,5 3 2,4 abs 254 nm 1,1-2,5 0,6-1,8 1-1,4 1,5 1 1,2 I0 (µmol fotones m-2 s-1) 455-2125 670-2352 1550-1790 1368 1131 1668 Imedia (µmol fotones m-2 s-1) 195-521 272-506 333-577 367 407 455 Kd PAR (m-1) 4,5-18,3 3,7-20,3 7-7,89 9,1 7,4 7,4 profundidad óptica (⦆) 2-6,4 1,4-10,7 2,9-5,5 4,1 3,6 4,2 sólidos en suspensión (mg L-1) 17-106 12-112 16-33 55 39 25
Tabla 2. Rangos y promedios (en negrita) de las variables ambientales registradas durante el
estudio en la Laguna Grande. nd = no detectable.
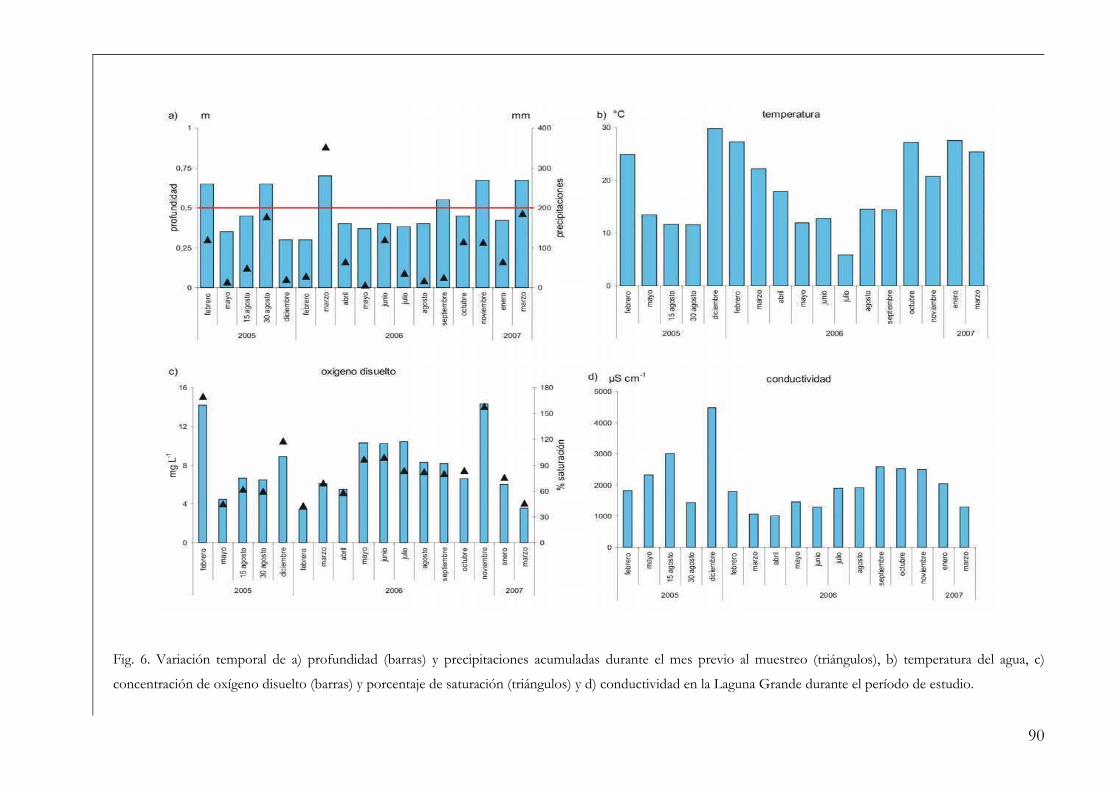
90
Fig. 6. Variación temporal de a) profundidad (barras) y precipitaciones acumuladas durante el mes previo al muestreo (triángulos), b) temperatura del agua, c)
concentración de oxígeno disuelto (barras) y porcentaje de saturación (triángulos) y d) conductividad en la Laguna Grande durante el período de estudio.
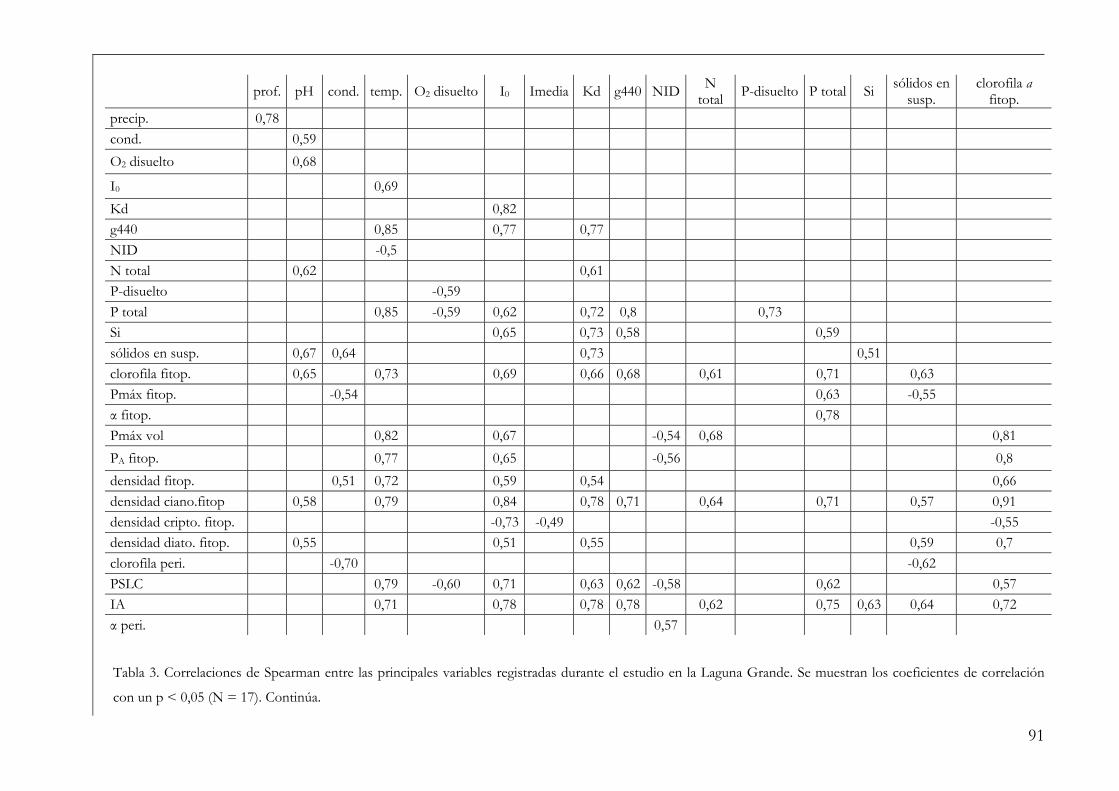
91
prof. pH cond. temp. O2 disuelto I0 Imedia Kd g440 NIDN
total P-disuelto P total Si
sólidos en susp.
clorofila a fitop.
precip. 0,78
cond. 0,59
O2 disuelto 0,68
I0 0,69
Kd 0,82
g440 0,85 0,77 0,77
NID -0,5
N total 0,62 0,61
P-disuelto -0,59
P total 0,85 -0,59 0,62 0,72 0,8 0,73
Si 0,65 0,73 0,58 0,59
sólidos en susp. 0,67 0,64 0,73 0,51
clorofila fitop. 0,65 0,73 0,69 0,66 0,68 0,61 0,71 0,63
Pmáx fitop. -0,54 0,63 -0,55
プ fitop. 0,78
Pmáx vol 0,82 0,67 -0,54 0,68 0,81
PA fitop. 0,77 0,65 -0,56 0,8
densidad fitop. 0,51 0,72 0,59 0,54 0,66
densidad ciano.fitop 0,58 0,79 0,84 0,78 0,71 0,64 0,71 0,57 0,91
densidad cripto. fitop. -0,73 -0,49 -0,55
densidad diato. fitop. 0,55 0,51 0,55 0,59 0,7
clorofila peri. -0,70 -0,62
PSLC 0,79 -0,60 0,71 0,63 0,62 -0,58 0,62 0,57
IA 0,71 0,78 0,78 0,78 0,62 0,75 0,63 0,64 0,72
プ peri. 0,57
Tabla 3. Correlaciones de Spearman entre las principales variables registradas durante el estudio en la Laguna Grande. Se muestran los coeficientes de correlación
con un p < 0,05 (N = 17). Continúa.
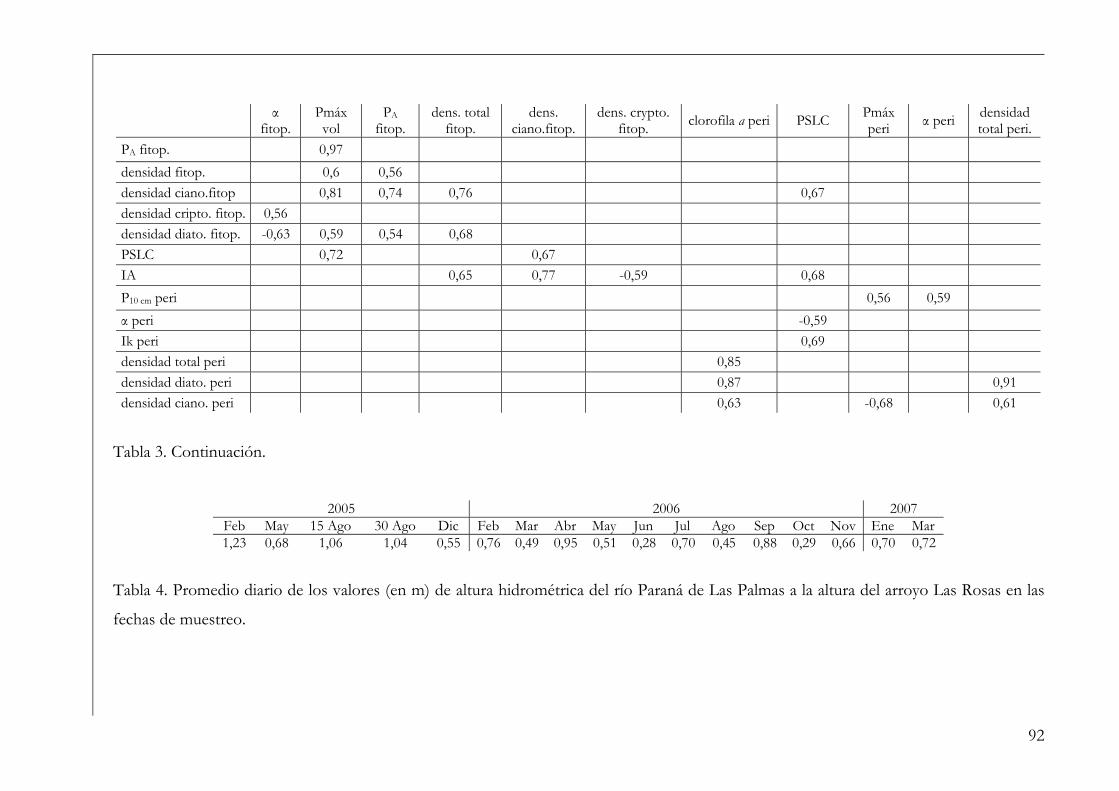
92
プ
fitop. Pmáx vol
PA fitop.
dens. total fitop.
dens. ciano.fitop.
dens. crypto. fitop.
clorofila a peri PSLC Pmáx peri
プ peri densidad total peri.
PA fitop. 0,97
densidad fitop. 0,6 0,56
densidad ciano.fitop 0,81 0,74 0,76 0,67
densidad cripto. fitop. 0,56
densidad diato. fitop. -0,63 0,59 0,54 0,68
PSLC 0,72 0,67
IA 0,65 0,77 -0,59 0,68
P10 cm peri 0,56 0,59
プ peri -0,59
Ik peri 0,69
densidad total peri 0,85
densidad diato. peri 0,87 0,91
densidad ciano. peri 0,63 -0,68 0,61
Tabla 3. Continuación.
2005 2006 2007 Feb May 15 Ago 30 Ago Dic Feb Mar Abr May Jun Jul Ago Sep Oct Nov Ene Mar1,23 0,68 1,06 1,04 0,55 0,76 0,49 0,95 0,51 0,28 0,70 0,45 0,88 0,29 0,66 0,70 0,72
Tabla 4. Promedio diario de los valores (en m) de altura hidrométrica del río Paraná de Las Palmas a la altura del arroyo Las Rosas en las
fechas de muestreo.

93
La irradiancia subacuática superficial (I0) fluctuó entre 2352 µmol fotones m-2 s-1
en noviembre de 2006 y 455 µmol fotones m-2 s-1 a mitad de agosto de 2005. Esta variable
mostró un patrón estacional, con valores más elevados en los meses más cálidos,
tendencia sustentada además por la correlación positiva encontrada entre esta variable y la
temperatura (Tabla 3). La irradiancia integrada en la columna de agua (Imedia) no siguió
un patrón estacional y no mostró diferencias significativas entre estaciones del año (K-W,
p > 0,05). Su valor más alto fue 580 µmol fotones m-2 s-1 en enero 2007 y el más bajo 195
µmol fotones m-2 s-1 a mediados de agosto 2005 (Fig. 7). El coeficiente de atenuación
vertical (Kd) siguió un patrón estacional, presentándose los valores más bajos en invierno.
El valor más alto fue 20,3 m-1 (octubre de 2006) y el más bajo fue 3,75 m-1 (julio de 2006)
(Fig. 8 a). El Kd se mostró correlacionado de manera positiva con la concentración de
clorofila a del fitoplancton, la densidad total de fitoplancton, los sólidos en suspensión, la
temperatura, el contenido húmico, P y N total y sílice. Por otro lado, los valores de Kd
espectral fueron menores para las longitudes de onda del rojo, indicando que esta │ era la
que más penetraba en la laguna en esta oportunidad (Kd rojo = 3,8 m-1; Kd verde = 5,8
m-1 y Kd azul = 5,4 m-1).
En general, las profundidades ópticas (⦆) fueron menores al valor 4,6; indicando
que la irradiancia llegaba al fondo del cuerpo de agua y sólo en cinco oportunidades, fin
agosto y diciembre 2005, octubre y noviembre 2006 y marzo 2007, las ⦆ fueron mayores al
mencionado valor. A excepción del muestreo de fines de agosto, se observó en estos
casos que la comunidad estaba dominada por cianobacterias filamentosas, mientras que
en agosto el alto valor de ⦆ podría estar relacionado con la resuspensión de los sedimentos
debida a las fuertes lluvias recientes. Las mediciones de absorbancia efectuadas a las
distintas longitudes de onda para estimar el contenido húmico arrojaron resultados
similares, presentando un patrón estacional marcado, con valores más bajos en invierno
(Fig. 8 b); además el contenido húmico se mostró positivamente correlacionado con la
temperatura (Tabla 3).
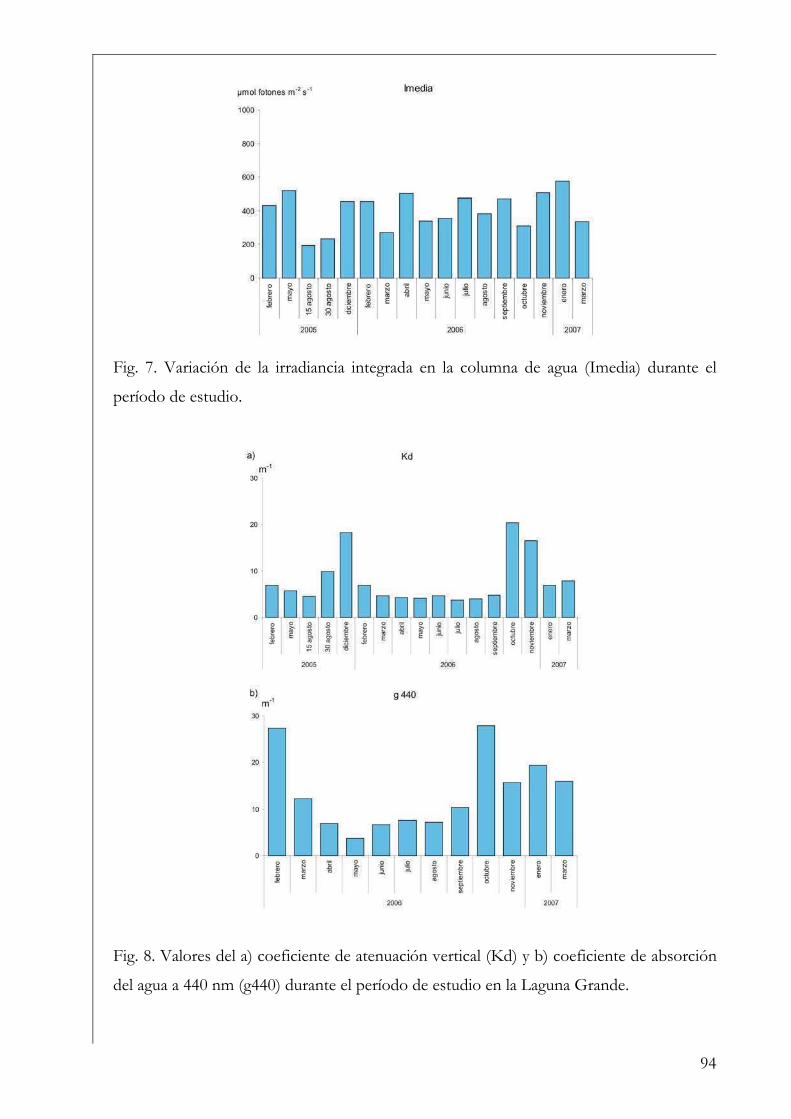
94
Fig. 7. Variación de la irradiancia integrada en la columna de agua (Imedia) durante el
período de estudio.
Fig. 8. Valores del a) coeficiente de atenuación vertical (Kd) y b) coeficiente de absorción
del agua a 440 nm (g440) durante el período de estudio en la Laguna Grande.

95
La variación en las concentraciones de P-disuelto se muestra en la Fig. 9 a. El
menor valor hallado fue una concentración por debajo del límite de detección en febrero
de 2005, coincidente con un crecimiento masivo de cianobacterias filamentosas, por lo
que el bajo valor podría explicarse por el consumo del nutriente por parte de estos
organismos. El resto de las concentraciones se encontraron por encima de las
consideradas limitantes para el máximo crecimiento algal (alrededor de 5-10 µg L-1 sensu
Reynolds 1984). Los máximos observados fueron de 2,2 mg L-1 y 1 mg L-1 en febrero y
marzo de 2006, respectivamente. Luego, los valores se mantuvieron por debajo de 0,62
mg L-1. No se observó un patrón temporal definido pero sí una correlación inversa con el
contenido de oxígeno disuelto (Tabla 3). En cambio, las concentraciones de P total
mostraron un patrón estacional marcado, con valores más altos hacia los meses más
cálidos (Fig. 9 b), como confirma la correlación positiva encontrada entre esta variable y la
temperatura (Tabla 3). La concentración de P total también se mostró asociada de manera
negativa con el oxígeno disuelto, mientras que de manera positiva con la I0, Kd,
concentración de clorofila a del fitoplancton y densidad de cianobacterias
fitoplanctónicas, ente otras variables (Tabla 3). Entre el 12 % (noviembre 2006) y el 90 %
(marzo 2006) del P total se encontró como P-disuelto, sin observarse un patrón estacional
definido en esta relación.
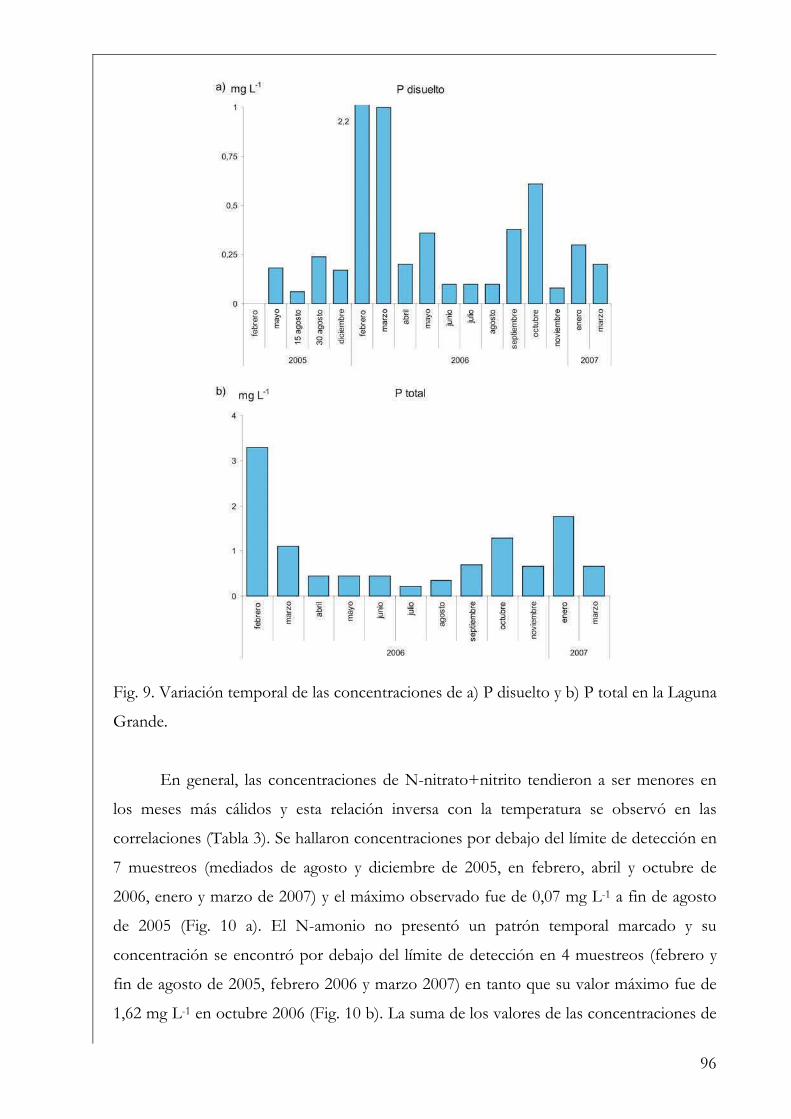
96
Fig. 9. Variación temporal de las concentraciones de a) P disuelto y b) P total en la Laguna
Grande.
En general, las concentraciones de N-nitrato+nitrito tendieron a ser menores en
los meses más cálidos y esta relación inversa con la temperatura se observó en las
correlaciones (Tabla 3). Se hallaron concentraciones por debajo del límite de detección en
7 muestreos (mediados de agosto y diciembre de 2005, en febrero, abril y octubre de
2006, enero y marzo de 2007) y el máximo observado fue de 0,07 mg L-1 a fin de agosto
de 2005 (Fig. 10 a). El N-amonio no presentó un patrón temporal marcado y su
concentración se encontró por debajo del límite de detección en 4 muestreos (febrero y
fin de agosto de 2005, febrero 2006 y marzo 2007) en tanto que su valor máximo fue de
1,62 mg L-1 en octubre 2006 (Fig. 10 b). La suma de los valores de las concentraciones de
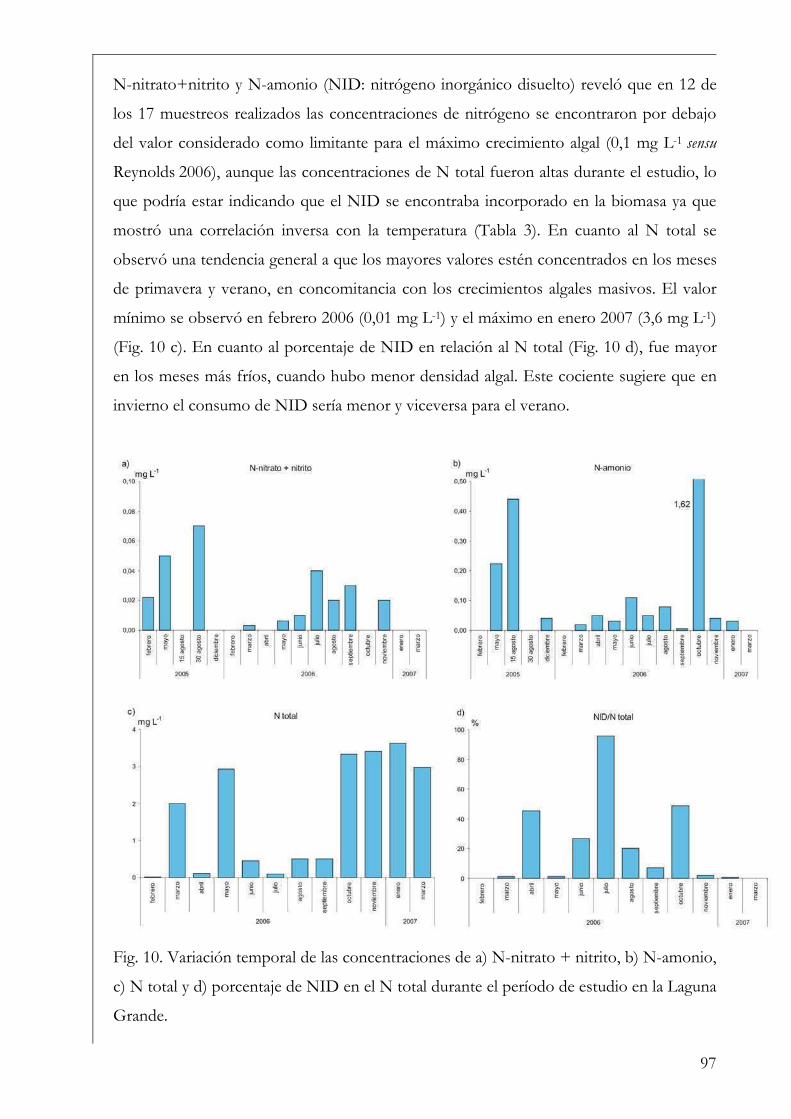
97
N-nitrato+nitrito y N-amonio (NID: nitrógeno inorgánico disuelto) reveló que en 12 de
los 17 muestreos realizados las concentraciones de nitrógeno se encontraron por debajo
del valor considerado como limitante para el máximo crecimiento algal (0,1 mg L-1 sensu
Reynolds 2006), aunque las concentraciones de N total fueron altas durante el estudio, lo
que podría estar indicando que el NID se encontraba incorporado en la biomasa ya que
mostró una correlación inversa con la temperatura (Tabla 3). En cuanto al N total se
observó una tendencia general a que los mayores valores estén concentrados en los meses
de primavera y verano, en concomitancia con los crecimientos algales masivos. El valor
mínimo se observó en febrero 2006 (0,01 mg L-1) y el máximo en enero 2007 (3,6 mg L-1)
(Fig. 10 c). En cuanto al porcentaje de NID en relación al N total (Fig. 10 d), fue mayor
en los meses más fríos, cuando hubo menor densidad algal. Este cociente sugiere que en
invierno el consumo de NID sería menor y viceversa para el verano.
Fig. 10. Variación temporal de las concentraciones de a) N-nitrato + nitrito, b) N-amonio,
c) N total y d) porcentaje de NID en el N total durante el período de estudio en la Laguna
Grande.
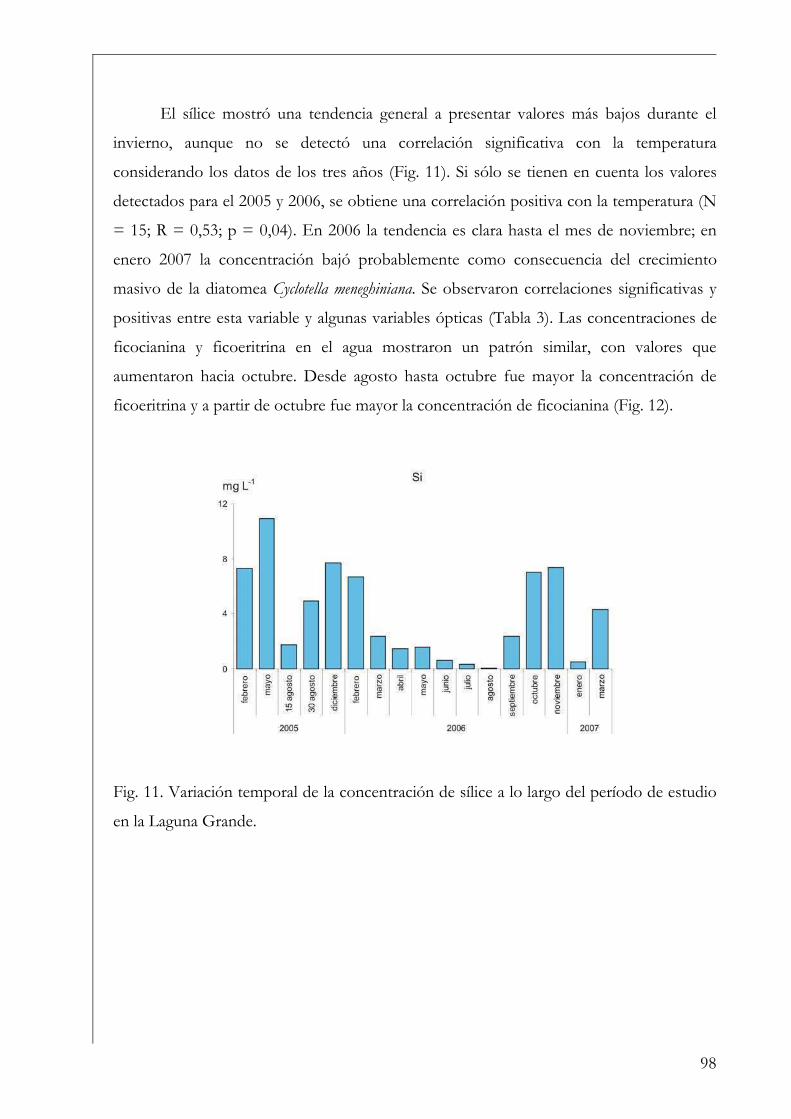
98
El sílice mostró una tendencia general a presentar valores más bajos durante el
invierno, aunque no se detectó una correlación significativa con la temperatura
considerando los datos de los tres años (Fig. 11). Si sólo se tienen en cuenta los valores
detectados para el 2005 y 2006, se obtiene una correlación positiva con la temperatura (N
= 15; R = 0,53; p = 0,04). En 2006 la tendencia es clara hasta el mes de noviembre; en
enero 2007 la concentración bajó probablemente como consecuencia del crecimiento
masivo de la diatomea Cyclotella meneghiniana. Se observaron correlaciones significativas y
positivas entre esta variable y algunas variables ópticas (Tabla 3). Las concentraciones de
ficocianina y ficoeritrina en el agua mostraron un patrón similar, con valores que
aumentaron hacia octubre. Desde agosto hasta octubre fue mayor la concentración de
ficoeritrina y a partir de octubre fue mayor la concentración de ficocianina (Fig. 12).
Fig. 11. Variación temporal de la concentración de sílice a lo largo del período de estudio
en la Laguna Grande.
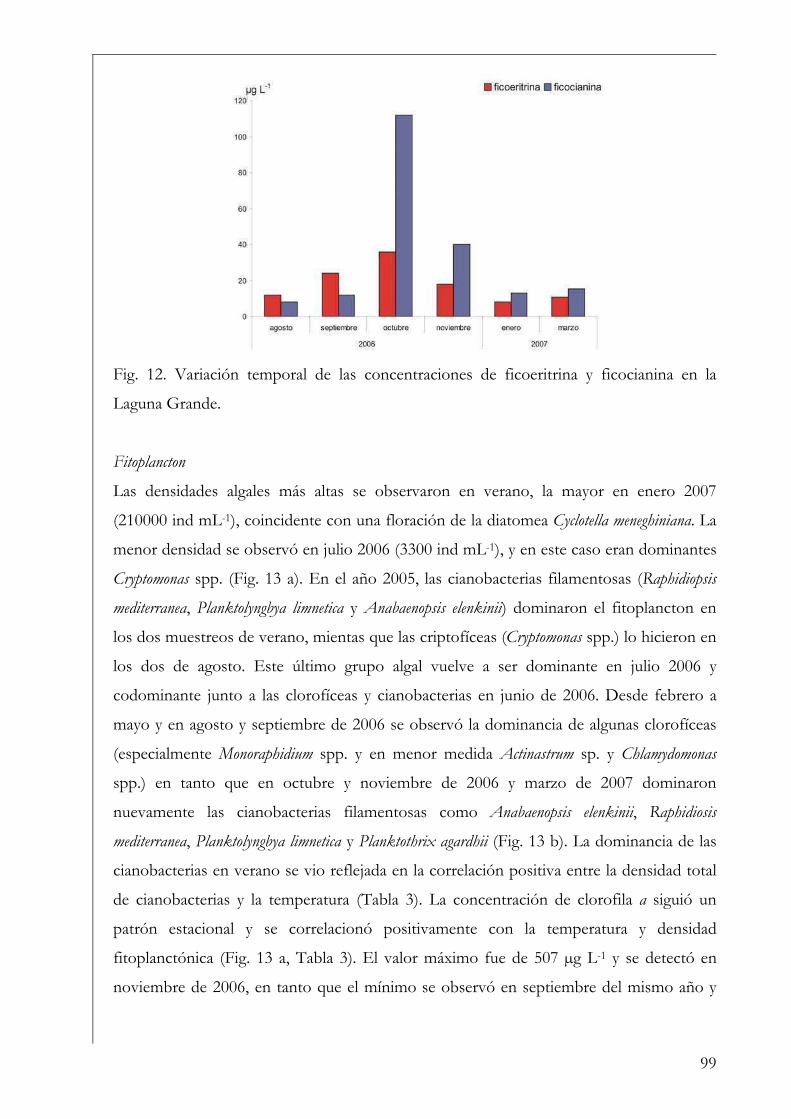
99
Fig. 12. Variación temporal de las concentraciones de ficoeritrina y ficocianina en la
Laguna Grande.
Fitoplancton
Las densidades algales más altas se observaron en verano, la mayor en enero 2007
(210000 ind mL-1), coincidente con una floración de la diatomea Cyclotella meneghiniana. La
menor densidad se observó en julio 2006 (3300 ind mL-1), y en este caso eran dominantes
Cryptomonas spp. (Fig. 13 a). En el año 2005, las cianobacterias filamentosas (Raphidiopsis
mediterranea, Planktolyngbya limnetica y Anabaenopsis elenkinii) dominaron el fitoplancton en
los dos muestreos de verano, mientas que las criptofíceas (Cryptomonas spp.) lo hicieron en
los dos de agosto. Este último grupo algal vuelve a ser dominante en julio 2006 y
codominante junto a las clorofíceas y cianobacterias en junio de 2006. Desde febrero a
mayo y en agosto y septiembre de 2006 se observó la dominancia de algunas clorofíceas
(especialmente Monoraphidium spp. y en menor medida Actinastrum sp. y Chlamydomonas
spp.) en tanto que en octubre y noviembre de 2006 y marzo de 2007 dominaron
nuevamente las cianobacterias filamentosas como Anabaenopsis elenkinii, Raphidiosis
mediterranea, Planktolyngbya limnetica y Planktothrix agardhii (Fig. 13 b). La dominancia de las
cianobacterias en verano se vio reflejada en la correlación positiva entre la densidad total
de cianobacterias y la temperatura (Tabla 3). La concentración de clorofila a siguió un
patrón estacional y se correlacionó positivamente con la temperatura y densidad
fitoplanctónica (Fig. 13 a, Tabla 3). El valor máximo fue de 507 µg L-1 y se detectó en
noviembre de 2006, en tanto que el mínimo se observó en septiembre del mismo año y
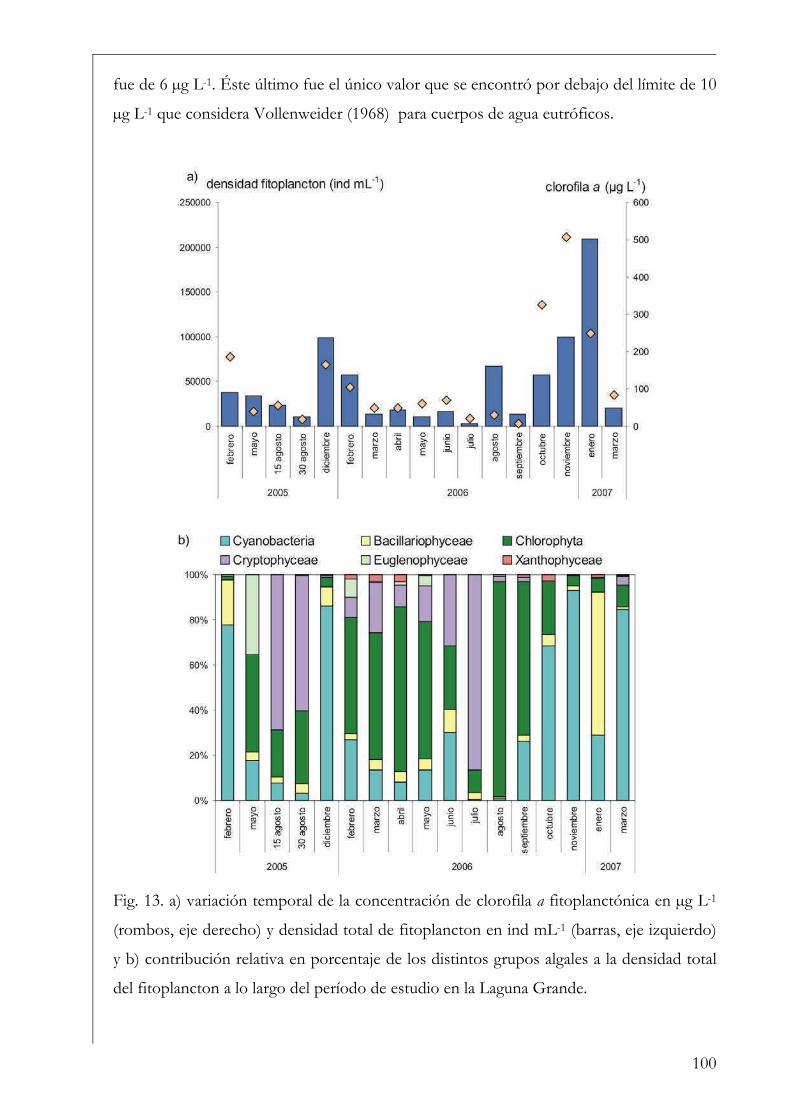
100
fue de 6 µg L-1. Éste último fue el único valor que se encontró por debajo del límite de 10
µg L-1 que considera Vollenweider (1968) para cuerpos de agua eutróficos.
Fig. 13. a) variación temporal de la concentración de clorofila a fitoplanctónica en µg L-1
(rombos, eje derecho) y densidad total de fitoplancton en ind mL-1 (barras, eje izquierdo)
y b) contribución relativa en porcentaje de los distintos grupos algales a la densidad total
del fitoplancton a lo largo del período de estudio en la Laguna Grande.

101
El rango de variación del número de asimilación (Pmáx) fue de 1,3 µg C (µg Chl a h)-1 en
enero 2007 y 12 µg C (µg Chl a h)-1 en marzo de 2007 (Fig. 14 a). No se observó una
tendencia estacional clara ni correlación con la temperatura. La eficiencia fotosintética (プ)
varió entre 0,005 µg C m2 s (µg Chl a h µmol fotones)-1 y 0,07 µg C m2 s (µg Chl a h)-1 en
enero 2007 y fin de agosto 2005 (Fig. 14 b), coincidiendo con la floración de la diatomea
Cyclotella meneghiniana en el primer caso y la dominancia de Cryptomonas spp. en el segundo.
Esta observación se advierte también en las correlaciones, ya que la densidad de
diatomeas se correlacionó de forma negativa con プ y la densidad de las criptofíceas lo hizo
de manera positiva (Tabla 3).
La irradiancia al inicio de la saturación fotosintética (Ik) no mostró una dinámica
estacional y presentó su valor más alto, 500 µmol fototes m-2 s-1, en el muestreo de agosto
2006; el valor más bajo se detectó a fines de agosto 2005 y fue de 34 µmol fototes m-2 s-1
(Fig. 14 c), en coincidencia con la mayor eficiencia fotosintética y la dominancia de
Cryptomonas spp. El fitoplancton estuvo limitado por luz en dos oportunidades, como
puede apreciarse en la Fig. 14 c, donde se señala con una flecha los momentos en los que
la Imedia fue menor que el parámetro Ik. La magnitud de la limitación lumínica,
representada como el cociente Imedia/Ik, varió entre 0,76 (la más limitada, agosto 2006) y
0,78 (la menos limitada, mayo 2006).
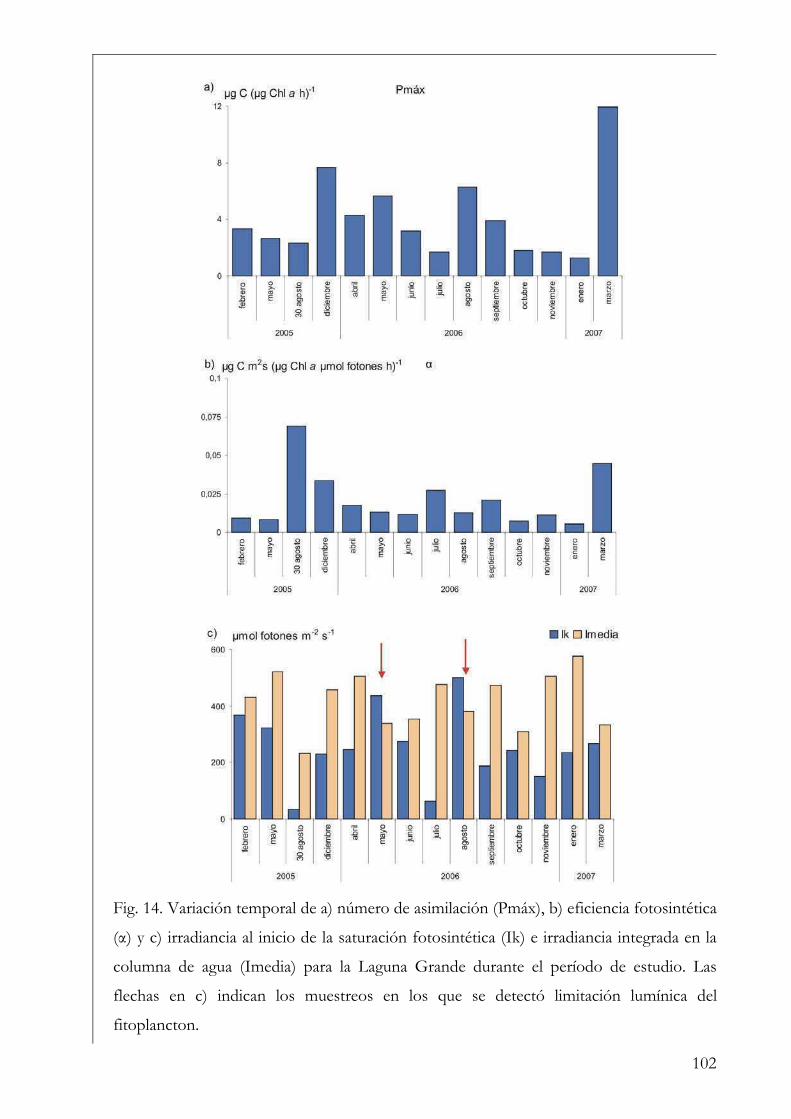
102
Fig. 14. Variación temporal de a) número de asimilación (Pmáx), b) eficiencia fotosintética
(プ) y c) irradiancia al inicio de la saturación fotosintética (Ik) e irradiancia integrada en la
columna de agua (Imedia) para la Laguna Grande durante el período de estudio. Las
flechas en c) indican los muestreos en los que se detectó limitación lumínica del
fitoplancton.
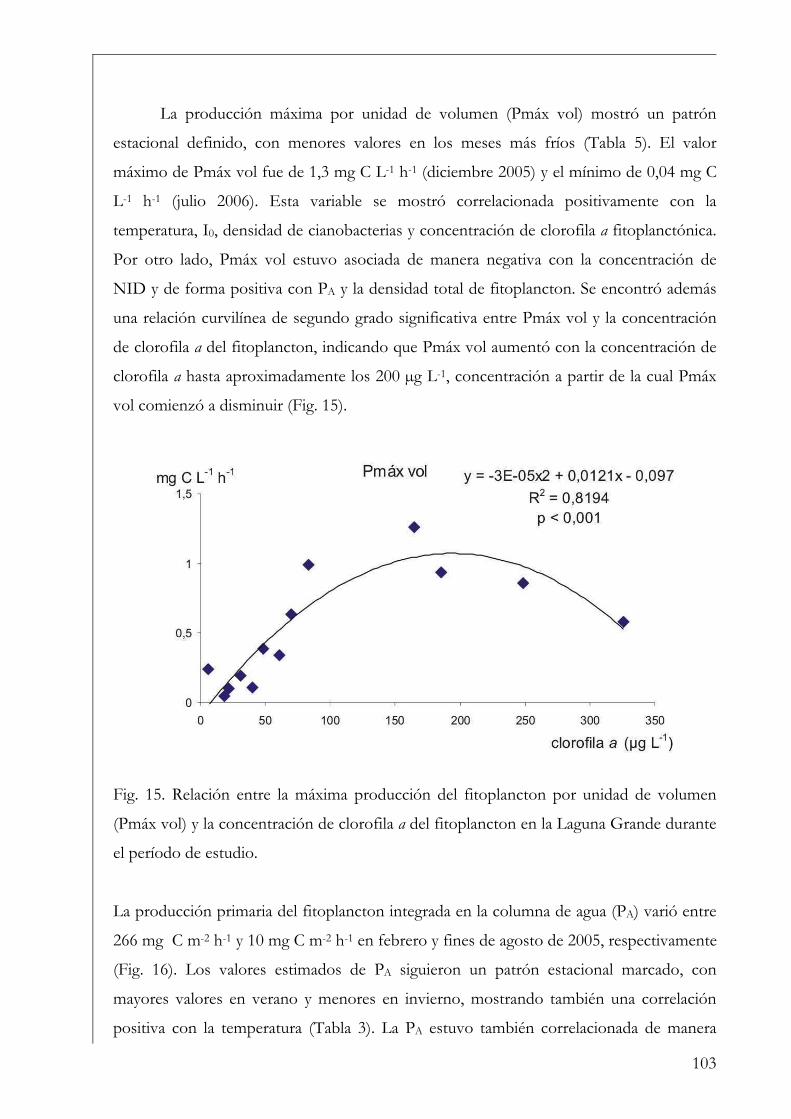
103
La producción máxima por unidad de volumen (Pmáx vol) mostró un patrón
estacional definido, con menores valores en los meses más fríos (Tabla 5). El valor
máximo de Pmáx vol fue de 1,3 mg C L-1 h-1 (diciembre 2005) y el mínimo de 0,04 mg C
L-1 h-1 (julio 2006). Esta variable se mostró correlacionada positivamente con la
temperatura, I0, densidad de cianobacterias y concentración de clorofila a fitoplanctónica.
Por otro lado, Pmáx vol estuvo asociada de manera negativa con la concentración de
NID y de forma positiva con PA y la densidad total de fitoplancton. Se encontró además
una relación curvilínea de segundo grado significativa entre Pmáx vol y la concentración
de clorofila a del fitoplancton, indicando que Pmáx vol aumentó con la concentración de
clorofila a hasta aproximadamente los 200 µg L-1, concentración a partir de la cual Pmáx
vol comienzó a disminuir (Fig. 15).
Fig. 15. Relación entre la máxima producción del fitoplancton por unidad de volumen
(Pmáx vol) y la concentración de clorofila a del fitoplancton en la Laguna Grande durante
el período de estudio.
La producción primaria del fitoplancton integrada en la columna de agua (PA) varió entre
266 mg C m-2 h-1 y 10 mg C m-2 h-1 en febrero y fines de agosto de 2005, respectivamente
(Fig. 16). Los valores estimados de PA siguieron un patrón estacional marcado, con
mayores valores en verano y menores en invierno, mostrando también una correlación
positiva con la temperatura (Tabla 3). La PA estuvo también correlacionada de manera
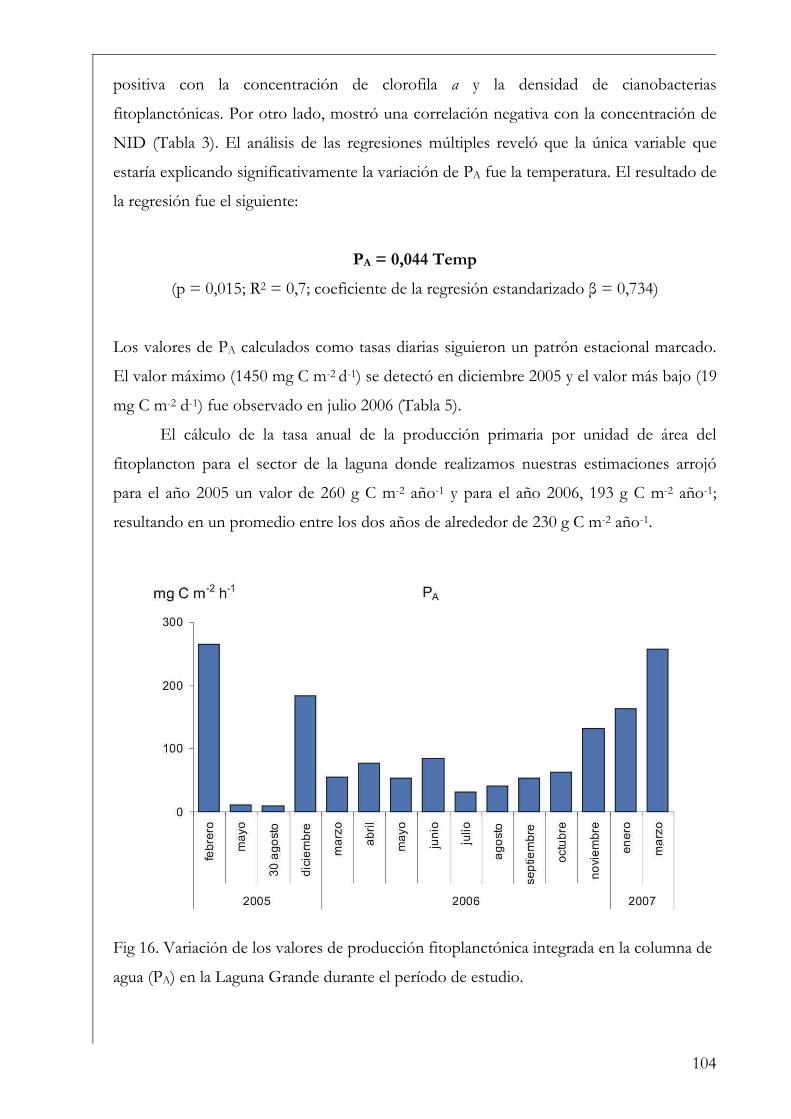
104
positiva con la concentración de clorofila a y la densidad de cianobacterias
fitoplanctónicas. Por otro lado, mostró una correlación negativa con la concentración de
NID (Tabla 3). El análisis de las regresiones múltiples reveló que la única variable que
estaría explicando significativamente la variación de PA fue la temperatura. El resultado de
la regresión fue el siguiente:
PA = 0,044 Temp
(p = 0,015; R2 = 0,7; coeficiente de la regresión estandarizado ベ = 0,734)
Los valores de PA calculados como tasas diarias siguieron un patrón estacional marcado.
El valor máximo (1450 mg C m-2 d-1) se detectó en diciembre 2005 y el valor más bajo (19
mg C m-2 d-1) fue observado en julio 2006 (Tabla 5).
El cálculo de la tasa anual de la producción primaria por unidad de área del
fitoplancton para el sector de la laguna donde realizamos nuestras estimaciones arrojó
para el año 2005 un valor de 260 g C m-2 año-1 y para el año 2006, 193 g C m-2 año-1;
resultando en un promedio entre los dos años de alrededor de 230 g C m-2 año-1.
Fig 16. Variación de los valores de producción fitoplanctónica integrada en la columna de
agua (PA) en la Laguna Grande durante el período de estudio.
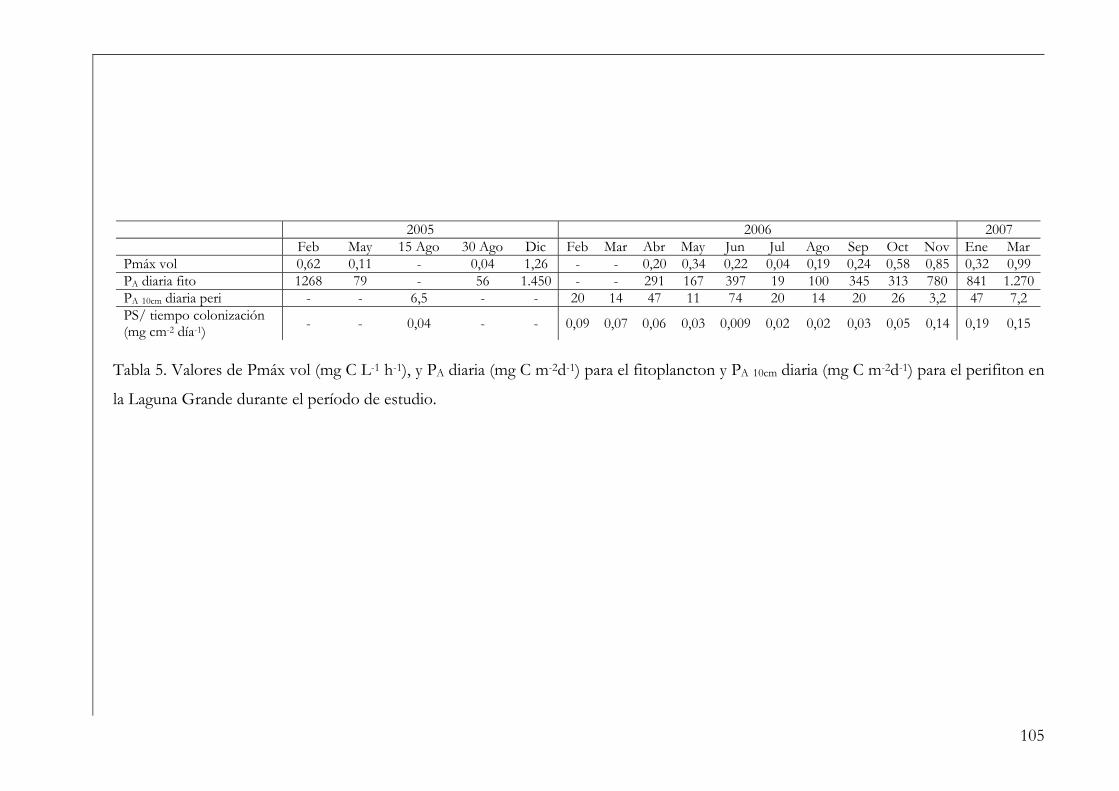
105
2005 2006 2007 Feb May 15 Ago 30 Ago Dic Feb Mar Abr May Jun Jul Ago Sep Oct Nov Ene Mar Pmáx vol 0,62 0,11 - 0,04 1,26 - - 0,20 0,34 0,22 0,04 0,19 0,24 0,58 0,85 0,32 0,99 PA diaria fito 1268 79 - 56 1.450 - - 291 167 397 19 100 345 313 780 841 1.270 PA 10cm diaria peri - - 6,5 - - 20 14 47 11 74 20 14 20 26 3,2 47 7,2 PS/ tiempo colonización (mg cm-2 día-1)
- - 0,04 - - 0,09 0,07 0,06 0,03 0,009 0,02 0,02 0,03 0,05 0,14 0,19 0,15
Tabla 5. Valores de Pmáx vol (mg C L-1 h-1), y PA diaria (mg C m-2d-1) para el fitoplancton y PA 10cm diaria (mg C m-2d-1) para el perifiton en
la Laguna Grande durante el período de estudio.

106
Perifiton sobre sustratos artificiales
En cuanto a las densidades algales se observó que tendieron a aumentar con la
disminución de la temperatura hasta noviembre 2006, aunque no se detectó una
correlación entre esas variables (Fig. 17 a). Por otro lado, en enero y marzo 2007 la
composición algal estaba constituida netamente por algas planctónicas. El máximo
hallado fue de 112000 ind cm-2 en marzo 2007 y el mínimo de 5750 ind cm-2 en octubre
2006. La clase Bacillariophyceae fue en general el grupo dominante la mayor parte del
estudio (Fig. 17 b). Entre las diatomeas dominantes, Navicula spp., Lemnicola hungarica y
Amphora veneta fueron las pennadas más importantes. En marzo 2006, cuando dominaron
las clorofíceas, Monoraphidium spp. eran las más abundantes. En noviembre 2006 la
comunidad estaba dominada por cianobacterias, siendo algunas especies de Heteroleiblenia
el organismo más conspicuo. En el muestreo de enero 2007 la especie dominante, al igual
que en el caso del fitoplancton, fue Cyclotella meneghiniana, alcanzando una de las
densidades más altas del estudio (alrededor de 98000 ind cm-2, representando el 92 % de
la densidad total de la comunidad). Finalmente, en marzo 2007 Merismopedia sp. fue la
cianobacteria dominante, alcanzando cerca del 50 % de la densidad total en esa fecha.
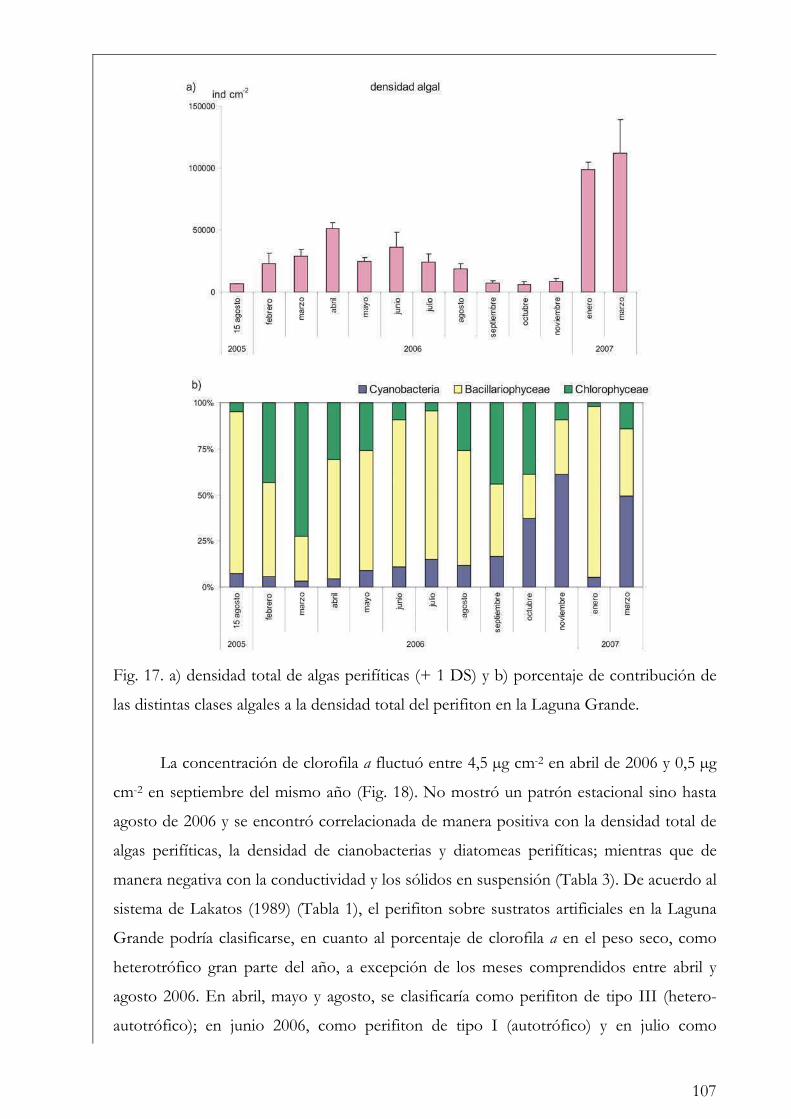
107
Fig. 17. a) densidad total de algas perifíticas (+ 1 DS) y b) porcentaje de contribución de
las distintas clases algales a la densidad total del perifiton en la Laguna Grande.
La concentración de clorofila a fluctuó entre 4,5 µg cm-2 en abril de 2006 y 0,5 µg
cm-2 en septiembre del mismo año (Fig. 18). No mostró un patrón estacional sino hasta
agosto de 2006 y se encontró correlacionada de manera positiva con la densidad total de
algas perifíticas, la densidad de cianobacterias y diatomeas perifíticas; mientras que de
manera negativa con la conductividad y los sólidos en suspensión (Tabla 3). De acuerdo al
sistema de Lakatos (1989) (Tabla 1), el perifiton sobre sustratos artificiales en la Laguna
Grande podría clasificarse, en cuanto al porcentaje de clorofila a en el peso seco, como
heterotrófico gran parte del año, a excepción de los meses comprendidos entre abril y
agosto 2006. En abril, mayo y agosto, se clasificaría como perifiton de tipo III (hetero-
autotrófico); en junio 2006, como perifiton de tipo I (autotrófico) y en julio como

108
perifiton de tipo II (auto-heterotrófico). Es decir, que la comunidad fue en proporción
altamente heterotrófica gran parte del año, a excepción de los meses de menores
temperaturas, cuando la fracción autotótrofica de la comunidad cobró mayor relevancia.
En cuanto al peso seco, las cenizas y el peso seco libre cenizas siguieron el mismo patrón
temporal estacional (Fig. 19 a-c). El peso seco varió entre 6,9 mg cm-2 en enero 2007 y
0,25 mg cm-2 en junio 2006, mientras que las cenizas entre 1,7 y 0,06 mg cm-2 en enero
2007 y junio 2006, respectivamente. De acuerdo a la clasificación de Lakatos (1989)
(Tabla 1) en relación a los valores de peso seco, el perifiton sobre sustratos artificiales fue
en general tipo III (perifiton de baja biomasa) la mayor parte del año, salvo los meses de
febrero a abril 2006 (perifiton tipo II, de biomasa intermedia) y noviembre 2006 y enero y
marzo 2007 (perifiton tipo I, alta biomasa). La velocidad de colonización estimada como
el valor de PS relativizado al número de días de colonización adquirió sus valores más
altos en los meses de mayor temperatura, siendo máxima en enero (0,19 mg cm-2 día-1) y
mínima en junio (0,009 mg cm-2 día-1) (Tabla 5). El porcentaje de cenizas con respecto al
peso seco osciló entre el 14 y 37 % (agosto y mayo 2006 respectivamente), pudiendo
considerarse como perifiton predominantemente orgánico (Tabla 1). El peso seco libre de
cenizas (PSLC) mostró un patrón estacional bien marcado (Fig. 19 c), siendo su valor
máximo 5,2 mg cm-2 y el mínimo 0,2 mg cm-2 en enero 2007 y junio 2006,
respectivamente. La variable que mejor explicó su variación fue la temperatura mediante
la expresión:
PSLC = 0,045 Temp – 0,77
(p = 0,005; R2 = 0,6; ベ = 0,77)
El índice autotrófico (IA) mostró un patrón estacional, con menores valores en los
meses más fríos (Fig. 19 d), como indica también la correlación positiva de esta variable
con la temperatura (Tabla 3). Es decir, que la comunidad era en proporción más
autotrófica en los meses más fríos. También se encontró una correlación positiva
significativa entre el IA y el coeficiente de atenuación vertical (Kd), que estaría indicando
que cuando la atenuación de la luz es mayor, el IA también, siendo en ese caso la
comunidad perifítica en proporción más heterotrófica.
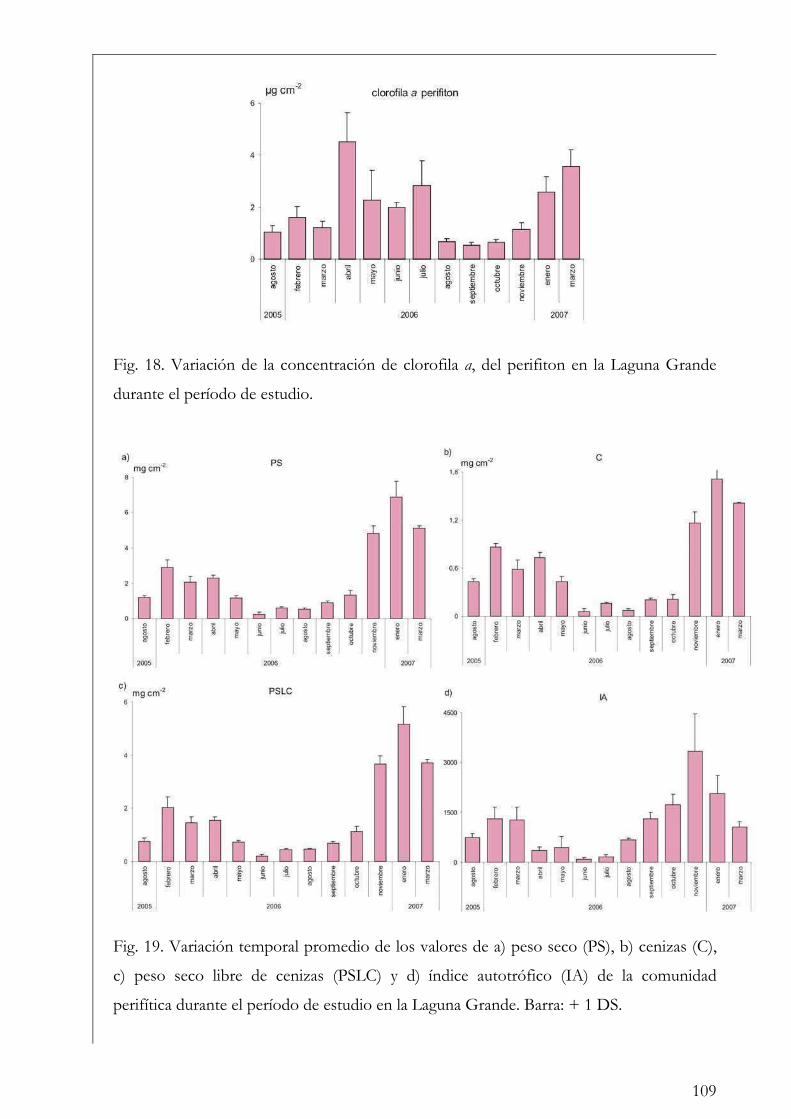
109
Fig. 18. Variación de la concentración de clorofila a, del perifiton en la Laguna Grande
durante el período de estudio.
Fig. 19. Variación temporal promedio de los valores de a) peso seco (PS), b) cenizas (C),
c) peso seco libre de cenizas (PSLC) y d) índice autotrófico (IA) de la comunidad
perifítica durante el período de estudio en la Laguna Grande. Barra: + 1 DS.

110
Los valores correspondientes al número de asimilación (Pmáx) fueron bajos en general,
siendo el máximo 2 µg C (µg Chl a h)-1 (octubre 2006) y el mínimo 0,05 µg C (µg Chl a h)-
1 (noviembre 2006). El patrón general de variación de Pmáx no estuvo asociado a la
temperatura, como puede observarse en la Fig. 20 a. La eficiencia fotosintética (プ)
tampoco presentó un patrón temporal definido (Fig. 20 b); el valor máximo registrado
para プ fue 0,008 µg C m2 s (µg Chl a h µmol fotones)-1 en octubre de 2006 y el mínimo,
de 0,0001 µg C m2 s (µg Chl a h µmol fotones)-1 en noviembre del mismo año. Se
encontró una correlación positiva entre プ y el NID y negativa entre プ y el PSLC (Tabla 3).
La irradiancia al inicio de la saturación fotosintética (Ik) no mostró un patrón
estacional marcado, aunque los valores más bajos se observaron en general en los meses
más fríos. El valor más alto se detectó en febrero 2006 y fue 492 µmol fotones m-2 s-1; en
tanto que el menor valor fue de 86 µmol fotones m-2 s-1 y se encontró en agosto 2006.
Esta variable se correlacionó positivamente con el contenido húmico y el PSLC (Tabla 3).
En la Fig. 20 c se puede observar que la comunidad se encontró limitada lumínicamente
en el muestreo de octubre 2006, siendo la magnitud de la limitación igual a 0,83.
La producción por unidad de área de sustrato a los 10 cm de profundidad
(profundidad a la cual fueron colonizados los sustratos, PA 10 cm) adquirió su valor más alto
en el muestreo de junio de 2006 (255 µg C m-2), siendo éste un orden de magnitud mayor
que la mayoría de los valores estimados durante el estudio y el valor más bajo fue
detectado en noviembre de 2006 (6 µg C m-2) (Fig. 21). Esta variable no se mostró
correlacionada de manera significativa con ninguna variable ambiental estudiada en este
trabajo. Las estimaciones de PA 10cm extrapoladas a valores diarios se muestran en la Tabla
5. Los valores diarios no difirieron significativamente entre estaciones del año (K-W, p >
0,05). Se estimó, en base a los valores de producción para cada muestreo, una tasa anual
de 10 g C m-2 año-1 por unidad de área de sustrato.
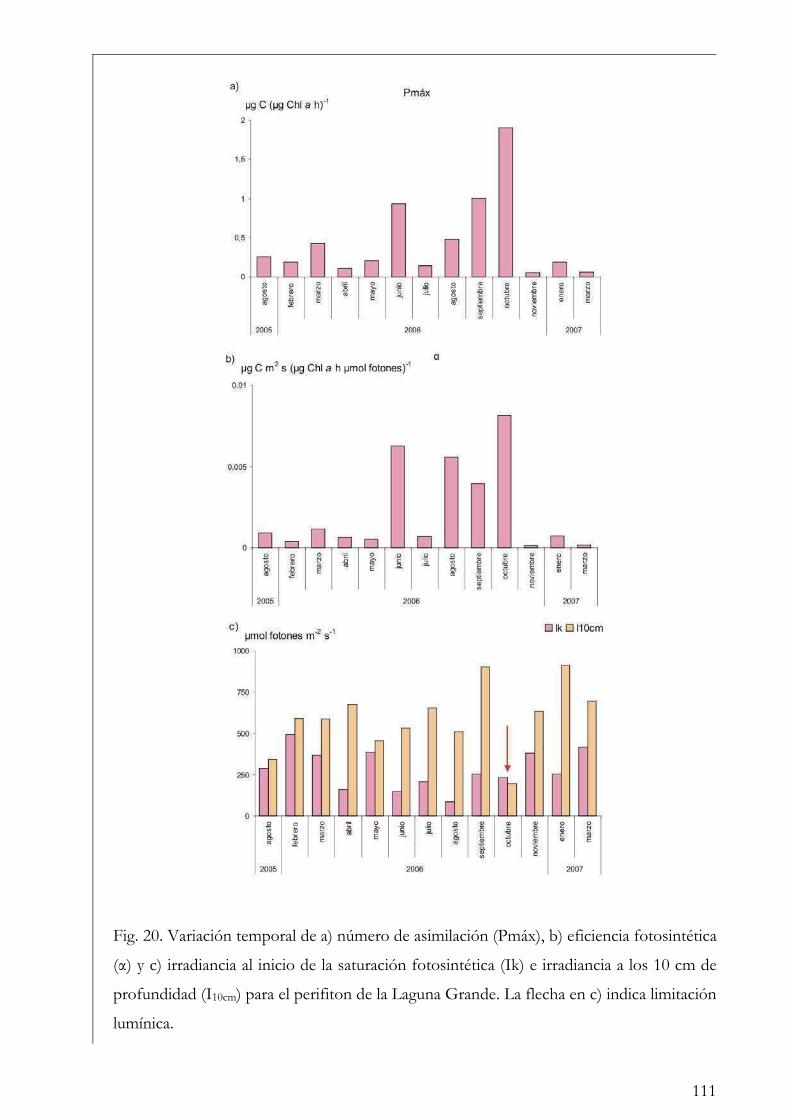
111
Fig. 20. Variación temporal de a) número de asimilación (Pmáx), b) eficiencia fotosintética
(プ) y c) irradiancia al inicio de la saturación fotosintética (Ik) e irradiancia a los 10 cm de
profundidad (I10cm) para el perifiton de la Laguna Grande. La flecha en c) indica limitación
lumínica.
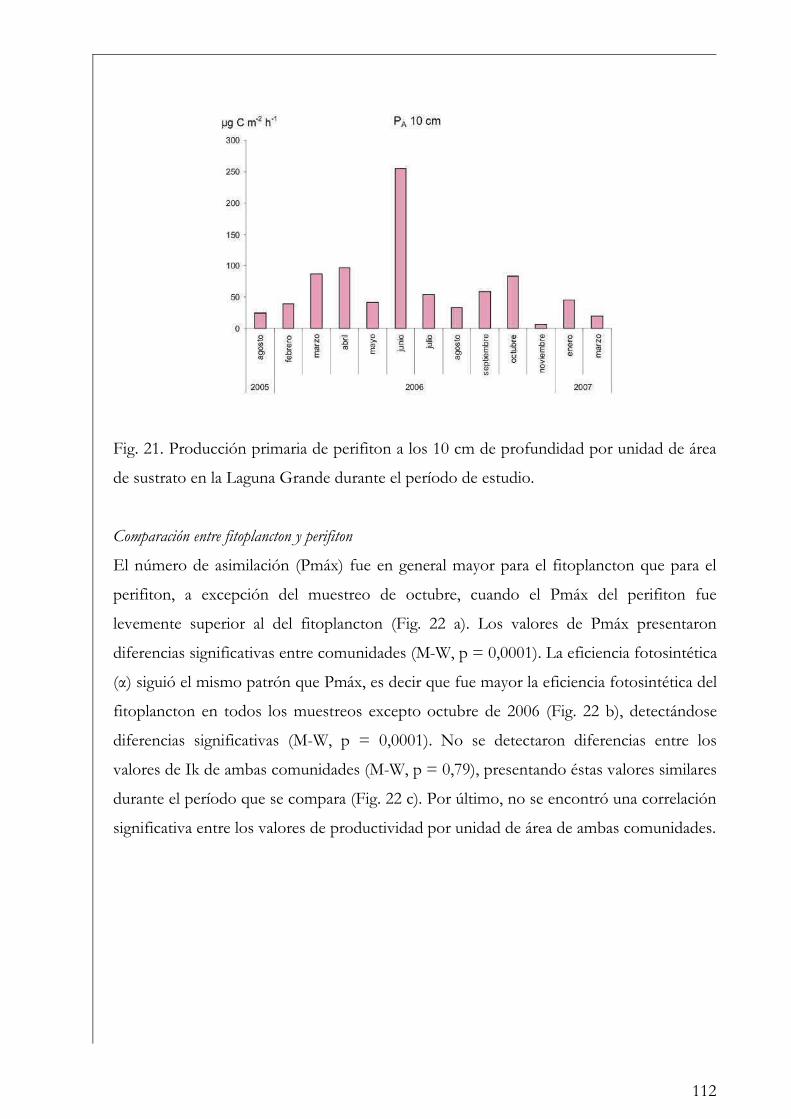
112
Fig. 21. Producción primaria de perifiton a los 10 cm de profundidad por unidad de área
de sustrato en la Laguna Grande durante el período de estudio.
Comparación entre fitoplancton y perifiton
El número de asimilación (Pmáx) fue en general mayor para el fitoplancton que para el
perifiton, a excepción del muestreo de octubre, cuando el Pmáx del perifiton fue
levemente superior al del fitoplancton (Fig. 22 a). Los valores de Pmáx presentaron
diferencias significativas entre comunidades (M-W, p = 0,0001). La eficiencia fotosintética
(プ) siguió el mismo patrón que Pmáx, es decir que fue mayor la eficiencia fotosintética del
fitoplancton en todos los muestreos excepto octubre de 2006 (Fig. 22 b), detectándose
diferencias significativas (M-W, p = 0,0001). No se detectaron diferencias entre los
valores de Ik de ambas comunidades (M-W, p = 0,79), presentando éstas valores similares
durante el período que se compara (Fig. 22 c). Por último, no se encontró una correlación
significativa entre los valores de productividad por unidad de área de ambas comunidades.
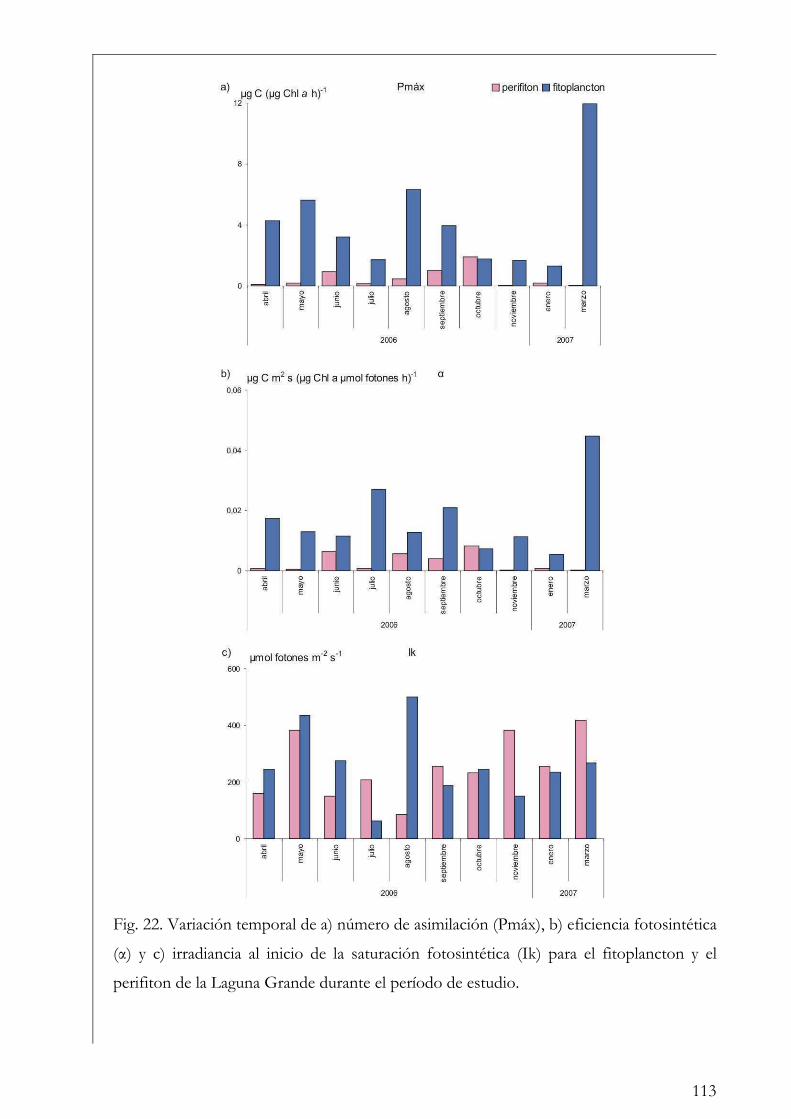
113
Fig. 22. Variación temporal de a) número de asimilación (Pmáx), b) eficiencia fotosintética
(プ) y c) irradiancia al inicio de la saturación fotosintética (Ik) para el fitoplancton y el
perifiton de la Laguna Grande durante el período de estudio.

114
DISCUSIÓN
La Laguna Grande es un cuerpo de agua somero, donde el viento ejerce un importante
papel en la mezcla constante de sus aguas. Su ambiente lumínico es restringido en cuanto
a la cantidad y calidad de luz que penetra, debido principalmente a las altas
concentraciones de ácidos húmicos. Sumado a esto, la cobertura fluctuante de macrófitas
flotantes realza aún más esta restricción, atenuando la irradiancia en sí y proveyendo una
importante fuente autóctona de ácidos húmicos debido a la descomposición del material
vegetal (Kirk 1994). Estas restricciones sin embargo no minimizaron el desarrollo
fitoplanctónico y por el contrario, la laguna se encontró en un estado de aguas turbias
(Scheffer et al. 1993) durante el estudio, dominado por esta comunidad. La Laguna
Grande posee características mixotróficas de acuerdo a la clasificación de Williamsom et
al. (1999) dadas sus altas concentraciones de P total y contenido húmico. Además de la
restricción lumínica, posee en ocasiones bajos tenores de nitrógeno inorgánico, y ambas
características podrían hacer de este cuerpo de agua un ambiente difícil para la vida de los
organismos estrictamente autotróficos. Por estas razones, no es sorprendente encontrar
en la laguna organismos mixotróficos tal como corroboró Sinistro et al. (2006) y
cianobacterias adaptadas a bajos contenidos de nutrientes (Oliver & Ganf 2000) y a un
amplio rango de disponibilidad de luz (Kirk 1994), grupos algales comunes en cuerpos de
agua húmicos en general (Lepistö et al. 2004, Lepistö & Holopainen 2003, Arvola et al.
1999, Ahtiainen et al. 1985).
Los niveles hidrométricos registrados en la laguna durante este estudio estuvieron
dentro de los observados previamente para este cuerpo de agua por otros autores
(Izaguirre et al. 2004, O´Farrell et al. 2003). Se observó durante el estudio una fuerte
influencia de las precipitaciones en las variaciones del nivel hídrico de la laguna. Si bien en
promedio las oscilaciones entre aguas altas y bajas fueron de aproximadamente 40 cm, en
ocasiones se puede llegar a secar parte de la laguna, como ocurrió en diciembre 2005, y en
ese caso las variaciones son mayores (≈ 70 cm). Para sistemas tan someros, una oscilación
de tan sólo unos pocos centímetros puede llegar a ser importante. Sin embargo, no
encontramos correlación de las variables ambientales (a excepción de las precipitaciones
acumuladas durante el mes previo al muestreo) y biológicas con la profundidad, siendo
ésta una laguna de un valle de inundación, donde las fluctuaciones en el nivel

115
hidrométrico son las modeladoras de la biota (Junk et al. 1989). Esto puede deberse a que
la estacionalidad, tan marcada, enmascaró el efecto de la variación hidrométrica. Por lo
visto por nosotros, la temperatura fue el factor que influenció en mayor medida los
patrones de variación de los parámetros limnológicos en general y del fitoplancton en
particular, pudiendo ser en este estudio la variable modeladora más evidente. La
estacionalidad influenció la dinámica de toda la masa de agua; influyó en la disponibilidad
de radiación y en la atenuación de la luz en la columna de agua y también estuvo
relacionada con la disponibilidad de los principales nutrientes.
Los valores obtenidos para el coeficiente de atenuación vertical (Kd) son altos
cuando se los compara con los de otros cuerpos de agua (Kirk 1994). En la Laguna
Grande su fluctuación a lo largo del estudio estuvo relacionada con la temperatura, y con
variables que se correlacionaron con ella fuertemente. El Kd estuvo influenciado no sólo
por el color del agua sino que el fitoplancton y su dinámica estacional lo condicionaron
fuertemente, como se puede deducir a partir de las correlaciones positivas con estas
variables. En cuanto al Kd espectral, si bien se observó en mayo 2006 que el Kd
correspondiente a las longitudes de onda del rojo tuvo un valor más bajo (mayor
penetración), no se puede decir que la longitud de onda del rojo era la única que
penetraba en la columna de agua. Esto es debido a que la profundidad óptica para los tres
Kd era menor a 4,6; por lo tanto en el somero perfil de 37 cm de profundidad no se llegó
a extinguir completamente ninguna longitud de onda. Sin embargo, esto es lo que podría
ocurrir en la laguna cuando la profundidad óptica es mayor, como por ejemplo en
diciembre de 2005, octubre y noviembre de 2006 y marzo de 2007, cuando el crecimiento
masivo de cianobacterias filamentosas limitó la penetración de la luz en la columna de
agua.
Con respecto al contenido húmico del agua, los valores de los estimadores
empleados en este trabajo son altos cuando se los compara por ejemplo con la Laguna
Iberá (Argentina), donde Loiselle et al. (2005) reportaron valores máximos de 1,9 y 2,3 m-1
para g440. También son altos con respecto a los que recopila Kirk (1994) para un amplio
rango de ambientes de agua dulce y los obtenidos por Gallegos et al. (2008) para dos lagos
húmicos de Nueva Zelanda. Es decir, que teniendo en cuenta el contenido húmico del
agua, la Laguna Grande podría considerarse un sitio muy restringido lumínicamente.
Durante el año 2006 y principios del 2007 las concentraciones de oxígeno disuelto
mostraron un patrón de variación temporal que se explica en relación a la solubilidad de

116
este gas con la temperatura (Wetzel 1981). Sin embargo esta tendencia se desdibujó
durante los crecimientos masivos de cianobacterias filamentosas, ante cuya presencia los
valores de oxígeno detectados fueron mayores que los que cabría esperar para la
solubilidad del gas con la temperatura del agua. Por este motivo es que no detectamos una
correlación entre la concentración de oxígeno disuelto y la temperatura considerando
todos los valores del estudio. Por otro lado, en situaciones de anoxia en el fondo del
cuerpo de agua, la liberación de fósforo por parte de los sedimentos anóxicos es un
proceso muy importante (Nürnberg 1984) y es altamente probable que este fenómeno sea
muy frecuente en la Laguna Grande, lo que podría explicar la correlación registrada entre
O2 y P-disuelto. Por su parte, O´Farrell et al. (2009) detectaron en esta laguna la presencia
de una oxiclina en la interfase de los sedimentos donde las concentraciones de oxígeno
disuelto disminuían abruptamente hasta hacerse nulas.
Los valores de pH fueron altos durante el estudio, incluso más elevados que los
reportados previamente para la laguna por O´Farrell et al. (2003) e Izaguirre et al. (2004).
Podría esperarse que, dado el elevado contenido húmico de sus aguas, el pH fuera más
bajo (Kortelainen 1999). Sin embargo en el humedal se observa salinización y
alcalinización del suelo que, según Chichizola (1993), estarían influenciados por la escasa
profundidad a la que se encuentran los sedimentos querandinos, y sería en estos sectores
donde se encuentra vegetación halófila (por ejemplo, Spartina desinflora, Distichlis spicata,
entre otras). Esta particularidad de la cuenca generaría que las aguas tengan una alta
reserva alcalina como puede desprenderse a partir de los valores de estimados medidos
por nosotros, que fueron elevados cuando se los compara con los obtenidos por
Carvahho et al. (2001) para lagos y lagunas las principales llanuras aluviales de Sudamérica
(Amazonas, Paraná Superior, Paraná Medio y Orinoco, entre otras). Además, la
correlación positiva encontrada entre pH y los elevados valores de conductividad
sustentaría esta explicación. Por otra parte, cuando O´Farrell et al. (2003) e Izaguirre et al.
(2004) realizaron sus investigaciones, la laguna se encontraba cubierta en gran parte por
macrófitas flotantes, hecho que limitaría el desarrollo del fitoplancton, a diferencia de los
resultados obtenidos en estudio, donde mayores tasas de fotosíntesis fitoplanctónica
podrían además contribuir a los altos valores de pH observados.
Por otro lado, Unrein (2001) comprobó experimentalmente en la Laguna Grande
que el fósforo no es limitante en la laguna y que sí puede serlo el nitrógeno. A veces, el
nitrógeno puede limitar la producción del fitoplancton en lagos eutróficos de latitudes

117
medias, en especial donde las concentraciones de fósforo son relativamente altas y la
relación N/P inorgánicos es baja (Reynolds 1984), como ocurrió en general en la laguna.
En lagos con alto contenido húmico es común la limitación por nitrógeno (Jansson et al.
2001, 1996) y por eso es también común encontrar organismos mixotróficos (Pålsson &
Granéli 2004). Se observó también que en general una pequeña fracción del N está en la
forma de NID, siendo las formas orgánicas y aquellas incorporadas a la biomasa las más
abundantes. En invierno, cuando el consumo por parte del fitoplancton sería menor, se
detectó la mayor relación entre NID/N total. Es muy probable entonces que el NID sea
consumido y reciclado muy rápidamente en este cuerpo de agua durante los períodos de
mayor crecimiento algal.
La concentración de Si se halló dentro de los valores observados por Reynolds
(1984) tanto para ríos de llanura (alrededor de 18 mg L-1) y lagos templados (1,2-12 mg L-
1). El mismo autor comenta que, en general en los lagos, las concentraciones de Si están
sujetas a marcados cambios estacionales, y regulan potencialmente el crecimiento de las
diatomeas en aguas naturales, y como consecuencia, la composición de especies del
fitoplancton. En la Laguna Grande, si bien no se obtuvo una correlación significativa
entre Si y temperatura se puede advertir un patrón estacional considerando el año 2006,
con valores más bajos en invierno. Es interesante destacar que este patrón se desdibuja
principalmente en enero 2007, cuando se registró la floración de Cyclotella meneghiniana. El
consumo masivo de parte de esta población de diatomeas puede haber hecho descender
los niveles de Si por debajo del valor que se esperaría para esa época del año.
El fitoplancton mostró un claro patrón estacional cuando se analizan las distintas
variables a través de las cuales estimamos su dinámica temporal. La temperatura en este
sentido se destacó como un importante factor modelador de las comunidades
fitoplanctónicas en otros trabajos realizados en llanuras de inundación en latitudes medias
(Huszar & Reynolds 1997, Train & Rodrigues 1997, García de Emiliani 1993). Las
densidades de fitoplancton registradas en este estudio son más elevadas que las registradas
por Izaguirre et al. (2004) y O´Farrell et al. (2003) para la misma laguna, diferencia que
puede radicar en la diferencia de cobertura vegetal, como se discutió más arriba en
relación al pH. A partir del 2004, la cobertura de plantas flotantes en la laguna fue
disminuyendo hasta quedar prácticamente restringida a la zona litoral, como durante
nuestro estudio (obs. pers.). A pesar de esta diferencia en densidad, la composición

118
específica fitoplanctónica encontrada por nosotros es similar a la registrada por O´Farrell
et al. (2007, 2003) e Izaguirre et al. (2004). En este sentido, es muy probable que las
elevadas concentraciones de ácidos húmicos, que generan condiciones lumínicas
particulares en la columna de agua, sean el factor determinante de la composición
florística de la comunidad fitoplanctónica. Es interesante entonces destacar que Unrein
(2001) registró un pico de densidad de Cyclotella meneghiniana en la misma época del año
que nosotros en la laguna. El autor comprobó experimentalmente que una de las
consecuencias del aumento de pH en la Laguna Grande era la dominancia de esta especie.
Esta podría ser la explicación para la correlación positiva observada entre el pH y la
densidad de diatomeas. Por su parte, van Dam et al. (1994) ubicó a esta especie entre las
alcalinófilas (ocurren a pH > 7) y como heterótrofa facultativa para el metabolismo del
nitrógeno, lo que podría estar explicando su dominancia en enero, cuando la
concentración de NID era baja (30 µg L-1 NID).
En nuestro caso la ocurrencia de la dominancia de las cianobacterias en verano
explicaría la correlación positiva encontrada entre la densidad de estos organismos y la
temperatura. Por otro lado, la posesión de ficobilinas como pigmentos accesorios por
parte de las cianobacterias sustentaría la mayor concentración de ficocianina detectada en
el muestreo de octubre 2006 (pico de absorción a ≈ 620 nm, Kirk 1994). Las ficobilinas
constituyen una ventaja adaptativa que permite a los organismos ampliar el rango de
captación lumínica a mayores longitudes de onda, que son importantes en cuanto a su
penetración en este tipo de aguas (Eloranta 1999).
En referencia a la dominancia de clorofíceas cocoides el resto del año, Steinberg
(2006) observó que las sustancias húmicas afectaban el crecimiento de algunas especies
del género Monoraphidium, estimulándolo en algunos casos. Por su parte, Sinistro et al.
(2007) observaron que estas especies eran predadas por el mesozooplancton de la laguna.
Este hecho provocaría que su presencia y dominancia en algunos casos esté controlada en
parte por la presión de predación. Sinistro (2007) observó entre los años 2005 y 2006 un
patrón estacional de fluctuación de las densidades de zooplancton en el mismo sector de
la laguna donde realizamos este estudio. Detectó densidades de zooplancton más bajas en
invierno y mayores en verano. Extrapolando las observaciones de Sinistro (2007) a
nuestro estudio, ya que se trata de un período cercano (agosto 2005, enero, marzo y
septiembre 2006), observamos que en los momentos en los que las densidades de
zooplancton serían más elevadas son aquéllos en los que dominaron las cianobacterias

119
filamentosas, que son predadas en menor grado que las clorofíceas cocoides (Sinistro et al.
2007). Por otro lado en épocas del año en las que figuran los valores más bajos de
densidades registrados por Sinistro (2007), nosotros encontramos dominancia de
clorofitas, más palatables, así como también criptofíceas. Es decir que además que estas
algas están adaptadas a crecer en sitios con alto contenido húmico de acuerdo a las
observaciones de Steinberg (2006), es muy probable que la presión de predación regule
sus patrones de abundancia durante gran parte del año.
Los valores de producción primaria fitoplanctónica integrada en la columna de
agua, PA, registrados por nosotros (10-266 mg C m-2 h-1) están en general dentro del rango
de los obtenidos por Conzonno & Claverie (1987) para la Laguna de Chascomús (24-195
mg C m-2 h-1) y dentro de la mayor parte de los valores estimados por Petrucio & Barbosa
(2004) para lagos tropicales del SE de Brasil. Nos referimos a la mayor parte de los
valores ya que estos últimos autores obtuvieron dos valores por encima de 1000 mg C m-2
h-1 en el mencionado trabajo durante la estación seca. Los valores más altos de PA
ocurrieron en verano cuando la temperatura del agua era más alta. La dependencia de la
producción integrada en la columna de agua con la temperatura fue ya descrita por Talling
(1957) y estaría sustentando que la temperatura haya resultado la variable explicativa de la
variación de PA en este estudio a partir de las regresiones múltiples. Durante las altas
temperaturas estivales se produjeron grandes desarrollos de cianobacterias y de diatomeas
y fue en estas condiciones que se alcanzaron los valores más altos de PA. La activa
fotosíntesis durante esas floraciones se puede corroborar a través de la correlación
positiva encontrada entre el pH y la densidad total de fitoplancton, la densidad de
cianobacterias y la concentración de O2 disuelto. Por otro lado, la correlación negativa
entre PA y NID estaría sugiriendo la existencia de alto consumo del NID por los
organismos en los meses de mayor crecimiento algal y producción. Dentro de las
cianobacterias filamentosas que se desarrollaron masivamente en verano encontramos a
Anabaenopsis elenkinii, fijadora de nitrógeno gaseoso, que podría estar aportando nitrógeno
en verano a través de este proceso fisiológico.
Las tasas diarias de variación de PA (19-1450 mg C m-2 d-1) estuvieron
aproximadamente dentro del rango observado por Stenuite et al. (2007) en el lago
Tanganyica (110-1410 mg C m-2 d-1). Sin embargo, las tasas anuales estimadas por estos
autores (123-205 g C m-2 año-1) son algo menores que las obtenidas en este trabajo
(promedio de los dos años: 230 g C m-2 año-1). Por otro lado, nuestras estimaciones

120
diarias de PA estuvieron por debajo de las reportadas por Oduor & Schagerl (2007) para
lagos hipereutróficos africanos y la PA anual estimada aquí fue aproximadamente el doble
que la obtenida por Conzonno & Claverie (1988) para la laguna de Chascomús (121 g C
m-2año-1). Recientemente, para la misma laguna se ha estimado una tasa anual de
aproximadamente 3 kg C m-2 año-1(Torremorell et al. 2008), cifra sustancialmente más alta
que la obtenida en nuestro estudio. Por su parte Putz & Junk (1997) recopilaron valores
de estimaciones de productividad primaria de fitoplancton y perifiton para lagunas del
valle de inundación del río Amazonas. Para sistemas de aguas húmicas encontraron que el
valor promedio era de 2,3 kg C m-2año-1, un orden de magnitud mayor que el estimado
por nosotros en sistemas de características limnológicas similares pero en una latitud
donde la estacionalidad está dada por las fluctuaciones entre períodos húmedos lluviosos
y secos, lo que provoca que el principal factor modelador de esos sistemas sea el nivel
hidrométrico. Finalmente, los valores diarios de PA fitoplanctónica se encuentran dentro
del rango de cuerpos de agua de productividad intermedia que recopila Kalff (2003) y son
similares a los obtenidos por Carignan & Planas (1994) para lagunas del valle aluvial del
Paraná medio en verano.
Los valores de Pmáx vol estimados por nosotros (37-1256 mg de C m-3 h-1) fueron
en general mayores que los reportados por Carignan & Planas (1994) y también a los
obtenidos por Pertrucio et al. (2005) en el SE de Brasil. Por otro lado, en este estudio
Pmáx vol presentó una relación de segundo orden con la concentración de clorofila a, y
esto no ocurrió con los valores de PA, que mostraron relación lineal aunque no
significativa (p = 0,07) con la concentración declorofila a. En el caso de Pmáx vol, la
relación encontrada implicaría que a concentraciones de clorofila a mayores a
aproximadamente 200 µg L-1 el efecto de sombreado producido por el propio
fitoplancton limitaría la producción primaria por unidad de volumen. Estos resultados
estarían sugiriendo que en la profundidad donde se daría la máxima producción, la Pmáx
vol se ve afectada por altas concentraciones de clorofila, mientras que si se integra en la
columna de agua este efecto no se hace evidente. Esto puede deberse a que cuando se
suman las contribuciones de cada una de las profundidades para calcular PA, queda
enmascarada la disminución de la producción por efecto del sombreado del fitoplancton.
Existe un efecto de dilución al integrar profundidades con menor aporte a la PA, como
ocurre en los sistemas húmicos en general, donde la zona eufótica está restringida a las
capas superiores de la columna de agua (Arvola et al. 1999).

121
El rango de variación del número de asimilación del fitoplancton, Pmáx (1,3-12 µg
C (µg chl a h)-1), se encontró dentro del rango de los reportados por Macedo et al. (2001,
2002) para una laguna costera de Portugal, por Torremorell et al. (2008) para la Laguna
Chascomús y por Carignan & Planas (1994) para una laguna del valle aluvial del río Paraná
en su tramo medio. Por su parte, Stenuite et al. (2007) reportaron valores más bajos (2,8-
5,9 µg C (µg chl a h)-1 para el lago Tanganyica. En cuanto a la eficiencia fotosintética (プ),
Reynolds (1984) revisó la bibliografía y observó que los valores para el fitoplancton en
general se encontraban entre 2-37 mg C (mg chl a)-1 E-1 m2, con la mayor parte en el
rango 6-18 mg C (mg chl a)-1 E-1 m2. Si expresamos nuestros valores en las mismas
unidades, nuestros valores de プ estarían comprendidos entre 1,6 y 19 mg C (mg chl a)-1
mol fotones-1 m2, es decir aproximadamente dentro del rango de valores reportados por
este autor. Se observó en este estudio que la comunidad menos eficiente (enero 2007) fue
la dominada por las diatomeas céntricas (Cyclotella meneghiniana) mientras que la más
eficiente lo estuvo por las criptofíceas. Los valores de eficiencia a bajas intensidades
lumínicas dependen de varios factores, entre ellos el clima lumínico, el contenido de
nutrientes, el tamaño celular, la capacidad que poseen las células de capturar la irradiancia
incidente y la composición de pigmentos (Côté & Platt 1983, Cleveland & Perry 1987,
Sakshaug et al. 1997). Entonces, podríamos sugerir que la baja concentración de nitrógeno
podría ser una causa para la baja eficiencia observada durante la floración de Cyclotella
meneghiniana. Por el contrario, la mayor eficiencia fotosintética fue registrada en momentos
de baja Imedia y temperatura. Esto estaría indicando una alta eficiencia fotosintética de la
comunidad dominada por Crypyomonas spp.
Los mayores valores del parámetro Ik registrados para el fitoplancton (435 y 500
µmol fototes m2 s-1 en mayo y agosto 2006) resultaron en general elevados cuando se los
compara con los obtenidos por Carignan & Planas (1994) (217-287 µmol fotones m-2 s-1),
Macedo et al. (2001) (5-335 µmol fotones m-2 s-1), Torremorell et al. (2008) (71-158 µmol
fotones m-2 s-1) y los que recopiló Kirk (1994) para agua dulce (16-323 µmol fotones m-2 s-
1). Se han registrados valores elevados (1230 µmol fototes m-2 s-1) por ejemplo, en
poblaciones de la cianobacteria Microcystis creciendo a altas irradiancias (Kirk 1994). El
parámetro Ik brinda información sobre el estado de fotoaclimatación de la comunidad y
según Henley (1993), un valor alto de Ik estaría indicando un uso eficiente de altas
irradiancias y un valor bajo, un uso eficiente de bajas irradiancias. En los muestreos
durante los cuales se observaron los dos valores más altos de este parámetro, la

122
comunidad estaba limitada lumínicamente y probablemente la posibilidad de desarrollarse
en un medio deficiente en luz se deba a los procesos de mezcla en la laguna. Dada la
escasa profundidad del cuerpo de agua, la mezcla es muy frecuente e importante
permitiendo que las algas accedan a capas superiores donde las irradiancias son mayores.
En los lagos con altos coeficientes de atenuación (Kd) los procesos de mezcla producen
gran variabilidad en el clima óptico y en pocos centímetros la variación de la penetración
de la luz es muy grande (Helbling et al. 2001 a). Otros factores que influencian la
capacidad de las células de fotoaclimatarse son la disponibilidad de nutrientes (Falkowski
& LaRoche 1991), los cambios de temperatura repentinos (Sakshaug et al 1997, Prézelin et
al. 1991), la composición específica y la historia lumínica previa (Côté & Platt 1983).
Inversamente, en invierno (julio 2005 y agosto 2006) se observaron los valores más
bajos de Ik indicando que se encontraba adaptada a fotosintetizar a bajas irradiancias
cuando la comunidad estaba dominada por Cryptomonas spp. Una vez más, destacamos la
gran adaptación de los organismos de este grupo taxonómico a ambientes con bajas
intensidades lumínicas pudiéndose visualizar también a través de este parámetro
fotosintético.
La composición específica de las algas del perifiton fue bastante homogénea durante el
estudio y es común encontrar la dominancia de diatomeas, clorofitas y cianobacterias
(Kalff 2003). Entre las diatomeas, que fue el grupo preponderante durante la mayor parte
del estudio, las especies más conspicuas Lemnicola hungarica y Amphora veneta son
clasificadas por van Dam et al. (1994) como autótrofas para el metabolismo del N pero
que toleran altas concentraciones de N orgánico en el medio. En cuanto a la alcalinidad,
los mismos autores consideran a A. veneta como una especie alcalinobionte, es decir que
ocurre exclusivamente a pH mayor que 7. En cuanto al nivel de saturación de oxígeno,
pueden soportar entre moderadas y bajas concentraciones de este gas. Además, como ya
se discutió más arriba, la laguna posee pH elevados y altas concentraciones de N
orgánicos provenientes en parte de la descomposición de las macrófitas, y del plancton.
Estas condiciones les confieren a estos organismos una ventaja especial en este sistema y
no es raro entonces encontrarlos dominando el perifiton. Por su parte, Putz (1997)
encontró en sistemas húmicos de la llanura de inundación del río Amazonas, la
dominancia de diatomeas perifíticas sobre sustratos artificiales (95 %) sin detectar
cambios en la dominancia de los distintos grupos algales debidos a la estacionalidad. En

123
nuestro estudio encontramos frecuentemente especies compartidas con el fitoplancton
como en marzo 2006, cuando las clorofíceas del género Monoraphidium dominaban tanto
en el fitoplancton como en el perifiton. Lo mismo ocurrió en enero 2007 con Cyclotella
meneghiniana, y en marzo 2007 con Merismopedia sp. que sólo dominó en el perifiton. Por
otro lado, la densidad de la fracción algal del perifiton fue máxima cuando la comunidad
no estuvo dominada por algas perifíticas, como en enero y marzo 2007. Si no se tienen en
cuenta estos muestreos al analizar la variación estacional de las densidades algales
perifíticas, se podría observar que la tendencia es hacia su aumento en los meses de
invierno, inversamente a lo que ocurre en el caso del fitoplancton, observándose entonces
un patrón especular. La dominancia de algas planctónicas en el perifiton en tres
oportunidades está relacionada con el solapamiento de los hábitats en los humedales
(Goldsborough & Robinson 1996), más aún si se considera la escasa profundidad y la
mezcla producida por el viento en la Laguna Grande.
Los valores de clorofila a del perifiton estimados por nosotros sobre sustratos
artificiales resultaron bajos cuando se los compara con los hallados por Kaggwa et al.
(2006) para sustratos naturales en humedales tropicales en Uganda y por Leandrini &
Rodrigues (2008) para ambientes del Paraná superior para perifiton sobre sustratos
naturales. Sin embargo, lo están dentro de los reportados por Moschini-Carlos et al. (2000)
para sustratos artificiales de vidrio en la represa Jurumirim en Brasil, por Laugaste &
Lessok (2004) para epifiton sobre junco y por Putz (1997) para perifiton sobre acetato en
sistemas húmicos de la llanura aluvial del río Amazonas. También se encuentran dentro
de los citados por Goldsborough & Robinson (1996) para epifiton de humedales de
distintas latitudes y tipo de sustrato. La comunidad perifítica fue en proporción altamente
heterotrófica gran parte del año, a excepción de los meses de menores temperaturas,
cuando la fracción autotótrofica de la comunidad fue más importante y la PA 10 cm fue
mayor, indicando que los procesos de producción superarían el metabolismo
heterotrófico. En los meses más cálidos el desarrollo de bacterias y animales provocaría
que los procesos de respiración dominen en la matriz perifítica.
Los valores de peso seco (PS) obtenidos en este trabajo (0,25-7 mg cm-2) están
dentro del orden de los encontrados en la bibliografía para epifiton en humedales de
distintas partes del mundo (Goldsborough & Robinson 1996). Sin embargo, son
relativamente bajos cuando se los compara con trabajos de Brasil, como por ejemplo el
realizado por Doyle (1991, en Putz & Junk 1997) (2-29 mg cm-2) y Engle & Melack (1993)

124
(32 mg cm-2). Además, de acuerdo al sistema de Lakatos (1989) la biomasa del perifiton
fue en general baja a excepción de los meses de mayor temperatura, donde el componente
heterotrófico de la comunidad cobró importancia. Por otro lado, no es sorprendente que
las cenizas hayan seguido un patrón de variación estacional en la Laguna Grande si
consideramos que una buena parte de este valor debe estar constituido por los frústulos
silíceos de las diatomeas y que las concentraciones de sílice siguieron un patrón estacional
durante gran parte del estudio.
En cuanto a los valores de peso seco libre de cenizas (PSLC) están dentro del
rango de los obtenidos por Putz (1997) sobre sustratos artificiales para sistemas de la
llanura de inundación del río Amazonas ricos en ácidos húmicos y por Leandrini &
Rodrigues (2008) para perifiton sobre sustratos naturales en ambientes semilóticos del
valle aluvial del río Paraná en su tramo superior. Por otro lado, son bajos cuando se los
compara con los obtenidos por Liboriussen & Jeppesen (2005) para sustratos artificiales
en sistemas no húmicos y por Kaggwa et al. (2006) en sustratos naturales en África. Dado
que el PSLC mostró un patrón estacional marcado, siendo la temperatura su principal
factor abiótico modelador, y que la concentración de clorofila a perifítica no siguió un
patrón estacional directo, es probable que la asociación entre temperatura y PSLC esté
dada fundamentalmente por los cambios en la biomasa de heterótrofos. Era común
observar en los sustratos de verano larvas de Chironomidae de gran tamaño, que bien
podrían estar contribuyendo al aumento en la proporción heterotrófica y explicando por
lo tanto este patrón. Este crecimiento estacional de la fracción heterótrofa del perifiton
podría estar explicando también la correlación negativa entre el PSLC y el NID y entre el
O2 y el PSLC; cuando hubo menos NID disponible habría menos algas pero más
heterótrofos que consumen O2.
El índice autotrófico (IA) corrobora el carácter predominantemente heterotrófico
de la comunidad perifítica en la Laguna Grande durante la mayor parte del año. Se
considera que valores mayores a 200 caracterizan a una comunidad heterotrófica (APHA
2005), y en este estudio en solamente dos oportunidades se registraron valores por debajo
de este límite (junio y julio 2006). Es interesante advertir que al aumentar el valor del
coeficiente de atenuación vertical (Kd), la comunidad era más heterotrófica. Esto estaría
indicando que la luz es un recurso que afecta el metabolismo de la comunidad. Por otro
lado, el alto contenido en fósforo del sitio estaría generando gran proliferación bacteriana
en el perifiton.

125
La productividad primaria del perifiton sobre sustratos artificiales por unidad de
superficie a los 10 cm de profundidad (PA 10 cm) fue baja cuando se la compara con otros
trabajos de distintas latitudes y distinto tipo de sustrato (O´Reilly 2006, Vadeboncoeur et
al. 2003, Putz 1997, Kairesalo 1980). Si bien P10 cm mostró un patrón temporal irregular,
su valor máximo coincide con unos de los valores más bajos del índice autotrófico (junio
2006). Esta observación corrobora con mediciones de producción primaria el carácter
autotrófico de la comunidad que había sido definido a partir del IA. La comunidad
autotrófica estaría restringida en su desarrollo debido a la limitación lumínica causada por
el desarrollo del fitoplancton, las altas concentraciones de ácidos húmicos y el
componente heterotrófico del propio perifiton. El fitoplancton, al mostrar un patrón
estacional bien marcado, mostró valores de densidad, clorofila a y producción primaria
(PA) menores en invierno, cuando el perifiton podría entonces desarrollarse, dado que
también habría más NID disponible y de esta manera podría alcanzar valores de
producción primaria más altos. Hansson (1992) por su parte, encontró mayores valores de
biomasa perifítica relacionados con la disminución de la temperatura.
El perifiton, y en particular el epipelon, suele ser menos sensible que el
fitoplancton a la disponibilidad de nutrientes en la columna de agua. Se sabe que puede
tomar carbono inorgánico (Vadeboncoeur & Lodge 1998) y otros nutrientes de los
espacios intersticiales del sustrato, donde las concentraciones suelen ser varios órdenes de
magnitud mayor que en la columna de agua. Por otro lado, el epifiton es más dependiente
de las concentraciones de nutrientes de la columna de agua (Kalff 2003), aunque puede
tomar nutrientes del sustrato vegetal (Cattaneo & Kalff 1979). En este estudio se empleó
un sustrato inerte en cuanto a la liberación de nutrientes y colocado a 10 cm de
profundidad. De esta forma, el perifiton, lejos del fondo y de potenciales fuentes de
nutrientes extra, se vio posiblemente limitado por la baja disponibilidad de NID de la
columna de agua. Sumado a esto, el fitoplancton le aventajaba en la competencia por la
luz y estaría contribuyendo a la atenuación de la irradiancia incidente sobre el perifiton.
Por su parte, Rai & Hill (1984, en Putz & Junk 1997) estimaron la producción primaria de
perifiton sobre sustratos de acetato de celulosa en una laguna húmica de Brasil y
obtuvieron tasas diarias de entre 0,01-4,6 g C m-2d-1, con una media de 1,5 g C m-2d-1. En
nuestro caso, la media diaria para todo el estudio fue de 0,023 g C m-2d-1, con un valor
máximo de 0,074 g C m-2d-1 en junio de 2006. Los valores de Rai & Hill (op. cit.) son entre
uno y dos órdenes de magnitud mayores que los estimados en este trabajo, lo que nos

126
permitiría inferir acerca de la baja productividad del perifiton estudiado. Por otro lado,
Doyle (1991, en Putz & Junk 1997) estimó la productividad de perifiton adherido a raíces
de macrófitas acuáticas y calculó una tasa de 310 g C m-2 año-1, un orden de magnitud
mayor que la estimada por nosotros (10 g C m-2año-1), salvando las diferencias de sustrato.
Por su parte, Cronk & Mitsch (1994) estimaron una tasa anual de 2-85 g C m-2 año-1 para
epifiton en humedales de Norteamérica. Los autores observaron los menores valores en
condiciones de aguas bajas. Este amplio rango de valores encontrados en la bibliografía da
una idea de la gran variabilidad del perifiton según las características del cuerpo de agua y
el tipo de sustrato analizado.
El número de asimilación del perifiton (Pmáx) obtenido a partir de las curvas P-I
se encontró entre los citados en la bibliografía para algas bentónicas, (Krause-Jensen &
Sand-Jensen 1998), epifiton de humedales (Goldsborough & Robinson 1996) y lagos
africanos (O´Reilly 2006). En cuanto al parámetro プ, también se encontró entre los
valores encontrados en otros trabajos (Liboriussen & Jepessen 2005, Goldsborough &
Robinson 1996). En general, los valores de irradiancia al inicio de la saturación
fotosintética (Ik, 86-492 µmol fotones m-2 s-1) se encontraron entre los que considera Hill
(1996) como comunes en el perifiton (100-400 µmol fotones m-2 s-1). Pueden observarse
valores más altos del parámetro Ik cuando se estudian comunidades aclimatadas a
irradiancias mayores, como puede observarse en el trabajo de Dodds et al. (1999) y no es
éste precisamente el caso de la Laguna Grande. Sin embargo al colonizarse la comunidad
a 10 cm de profundidad es probable que haya percibido la variabilidad en las condiciones
lumínicas causadas por la mezcla y de este modo haya recibido mayores irradiancias en
algún momento a lo largo del período de colonización.
Los parámetros Pmax y プ fueron mayores para el fitoplancton que para el
perifiton, indicando que la comunidad de vida libre asimila más carbono y de forma más
eficiente. Esto ocurrió así a excepción del muestreo correspondiente al mes de octubre
2006. En esta oportunidad, cuando todo el NID se encontraba en la forma de amonio, se
observaron altos valores de contenido húmico, Kd (20 m-1), P-disuelto, ficocianina,
clorofila a fitoplanctónica y sólidos en suspensión. La profundidad óptica en este caso fue
alta para el sistema (9,1), indicando que a los 23 cm de profundidad (profundidad máxima:
45 cm) llegaba el 1 % de la luz incidente en superficie. Es probable que estas
circunstancias hayan sido provocadas por altas tasas de descomposición de materia
orgánica. Sería de esperar que bajo estas condiciones las cianobacterias dominantes en el

127
plancton (Anabaenopsis elenkinii) se encontraran restringidas a los primeros centímetros de
profundidad. Por otro lado, ninguna de las comunidades estudiadas debió verse limitada
por nitrógeno dados los altos valores estimados (1620 µg L-1 N-amonio, relación NID/P
disuelto 2,7). Podemos suponer entonces que las altas concentraciones de NID
promovieron una alta eficiencia fotosintética por parte de la fracción algal del perifiton a
pesar de estar limitada lumínicamente. Es decir que, probablemente, el nitrógeno podría
ser un factor limitante para el perifiton en este sistema. Esta explicación sustenta además
la correlación positiva encontrada entre la eficiencia (プ) fotosintética del perifiton y el
NID.
Mientras que la dinámica de la producción fitoplanctónica estuvo explicada por la
temperatura, en el caso del perifiton el análisis de regresión múltiple no arrojó ninguna
variable ambiental significativa que explique los patrones de variación de la producción, si
bien el NID pareció marcar en parte su dinámica. Por otro lado, es probable que la
predación, que no fue analizada en este estudio, sea quien ejerza la fuerza modeladora de
la dinámica de esta comunidad. Esta posibilidad sería una buena hipótesis para poner a
prueba si consideramos los altos valores del índice autotrófico y que los mayores valores
de producción primaria y densidad algal perifítica (sin incluir los muestreos de 2007
cuando la flora estaba dominada por fitoplancton) se observaron cuando el IA era más
bajo (en invierno), y por tanto había menor componente heterótrofo en la comunidad.
Las variables estudiadas por nosotros nos permiten hacer inferencias en el control desde
los recursos, mientras que quedaría como interrogante investigar qué pasa con el control
ejercido por los predadores, tanto para el perifiton como para el fitoplancton.
En resumen de lo planteado hasta aquí en cuanto a la productividad de ambas
comunidades, hemos visto que en un escenario libre de macrófitas flotantes la dinámica
de la productividad del fitoplancton integrada en la columna de agua se presenta con una
estacionalidad muy marcada en tanto que la producción del perifiton por unidad de área
de sustrato no presenta un patrón definido. Se observa de todos modos que la producción
primaria del perifiton tiende a ser mayor en los momentos que es menor el desarrollo del
fitoplancton, que afectaría la disponibilidad de luz y nutrientes que recibe el perifiton . Los
parámetros fotosintéticos del perifiton, y en particular la eficiencia, parecerían responder a
la disponibilidad de NID de manera positiva. En general, los parámetros fotosintéticos
Pmáx y プ fueron mayores para el fitoplancton, lo que podría estar vinculado con la

128
capacidad del fitoplancton de desplazarse en la columna de agua (presencia de flagelos,
vesículas de gas en cianobacterias) y así obtener mejores condiciones de iluminación y de
nutrientes. Por último, ambas comunidades experimentaron en algún momento del año
limitación lumínica relacionada a altos valores de Ik provocados probablemente por el
efecto de la variabilidad lumínica dada por la mezcla.
Finalmente y en relación a las hipótesis planteadas al principio de este capítulo, resta
analizar la contribución relativa de la producción primaria de ambas comunidades a la
productividad microbiana del sistema. En el caso del fitoplancton, para poder realizar esta
estimación debemos suponer que la comunidad es homogénea y de igual composición
específica en toda la laguna y que la productividad estimada por nosotros es representativa
de toda la laguna. Basamos esta suposición en que muestreamos en un área de transición
entre la zona pelágica y litoral donde la producción podría ser menor y mayor
respectivamente que las estimadas en este trabajo. Si suponemos el valor de 230 g C m-2
año-1 estimado por nosotros como un aporte promedio, obtendríamos que el fitoplancton
produciría en las 156 ha de la laguna, 360 toneladas de C por año.
Por otro lado, haciendo una suposición de máxima productividad de perifiton en la
que toda la laguna está cubierta de juncos (de 2 cm de diámetro) y que todos los juncos
son sustrato colonizable; que la productividad del perifiton es similar a la que estimamos
sobre sustratos artificiales a los 10 cm de profundidad (10 g C m-2 año-1) y que así se
mantiene en toda la estructura vertical de junco (extendiéndose en la capa eufótica),
obtendríamos una tasa máxima de 16 toneladas de C por año. Es decir que el máximo
aporte que podría realizar el perifiton a la productividad del sistema es un orden de
magnitud menor que el aporte del fitoplancton. Analizando estos números en porcentaje,
obtenemos que el fitoplancton en estas condiciones constituiría el 96 % de la
productividad microbiana mientras que el perifiton el 4 %. Esto significa que en
condiciones reales, en caso que la productividad estimada por nosotros fuera
representativa de otras áreas de la laguna, el aporte del perifiton sería aún mucho menor,
ya que nuestra estimación se basó en suponer que toda la laguna está cubierta de juncos.
En períodos de aguas turbias, con gran desarrollo de fitoplancton, el aporte de la
comunidad perifítica a la productividad primaria total microbiana sería mínimo.
Podríamos hipotetizar entonces, que el perifiton estaría ocupando el rol funcional de las
macrófitas sumergidas en la hipótesis de equilibrios alternativos (Scheffer et al. 1993). Se

129
han observado en la bibliografía (Liboriussen & Jeppesen 2003, Vadeboncoeur &
Steinman 2002, Meulemans 1988) contribuciones similares para el perifiton en cuerpos de
agua oligotróficos o eutróficos dominados por fitoplancton, como es el caso de la Laguna
Grande.
Una limitación que tiene esta extrapolación y que nos podría estar llevando a
subestimar la productividad del perifiton en la laguna es que en los sustratos naturales,
como se vio en el capítulo I de esta tesis, fue común observar un gran desarrollo de algas
verdes filamentosas del tipo Stigeoclonium sp., y en los sustratos artificiales, su
representación fue menor. La menor presencia de algas verdes de gran porte en sustratos
artificiales ya ha sido reportada para sistemas de características similares por Putz (1997)
en lagunas del valle de inundación del río Amazonas. Esta diferencia de composición
específica entre ambos tipos de sustratos podría estar relacionada también con el tiempo
de colonización de los sustratos artificiales. Para que los filamentos de Stigeoclonium sp.
puedan desarrollarse seguramente se necesita de un período mayor de tiempo ya que es un
alga que cubre gran parte de la superficie del sustrato con su porción basal, además de
poseer filamentos erectos. Nosotros observamos estos individuos en juncos más viejos, lo
que podría estar sugiriendo que es una especie tardía en la sucesión del perifiton en los
sustratos naturales. Los juncos más jóvenes poseían una composición algal similar a la de
los sustratos artificiales (obs. pers.). Entonces, la estimación anterior debe ser hecha con
cautela y considerando la edad de los juncos, es decir que sería aplicable para juncos
jóvenes y no se ha considerado completamente el aporte de Stigeoclonuim sp. a la
productividad, aporte que podría llegar a ser importante. Por otro lado, la suposición de
que todos los juncos que cubren la laguna estén colonizados es una sobreestimación de la
realidad, ya que dentro de las densas matrices de juncos las condiciones para el
crecimiento del perifiton se deterioran notablemente (Putz 1997) como también se
discutió en el capítulo I en relación a la poca cantidad de especies encontradas en el ROL
2, donde había un profuso desarrollo de plantas palustres.
Haciendo estas salvedades, podemos inferir que el fitoplancton supera al perifiton
en cuanto a su aporte a la productividad microbiana de la laguna. Esta observación estaría
indicando que en este tipo de sistema, con gran desarrollo de fitoplancton y contenido
húmico, el perifiton, que no puede posicionarse en la columna de agua en busca de
irradiancias óptimas, ve limitado su desarrollo y productividad primaria. Sin embargo,
creemos que esta situación ocurre en condiciones donde el desarrollo de la macrofitia

130
flotante es limitado debido probablemente a las bajas concentraciones de NID, ya que un
desarrollo mayor de plantas provee de sustrato al perifiton (Sand-Jensen & Borum 1991) y
también limitaría el desarrollo del fitoplancton (Scheffer & van Nes 2007).

131
CAPÍTULO III
COMPARACIÓN DE LA PRODUCCIÓN PRIMARIA DEL FITOPLANCTON
Y PERIFITON EN DOS AMBIENTES LUMÍNICOS CONTRASTADOS

132
INTRODUCCIÓN
La luz juega un rol esencial en la interacción competitiva entre fitoplancton y perifiton ya
que el fitoplancton, absorbe y dispersa la luz antes que ésta alcance la comunidad
perifítica, modificando así su calidad y cantidad (Sand Jensen & Borum 1991). La
disminución de la productividad y biomasa perifítica como resultado de esta interacción
ha sido observada en lagos eutróficos. Recientemente, Vadeboncoeur et al. (2008)
analizaron el rol de la producción primaria perifítica en función de las variables
morfométricas, contenido de nutrientes y luz de distintos cuerpos de agua empleando una
base de datos bibliográfica. Los autores señalaron que la contribución del perifiton a la
productividad del cuerpo de agua era menor que la del fitoplancton en lagos someros
eutróficos. Obtuvieron que la importancia relativa del perifiton era mayor en lagos
someros oligotróficos y en lagos de altas latitudes. En sistemas oligotróficos, el
fitoplancton suele estar limitado por nutrientes, lo que disminuye su desarrollo y permite
así una mejor penetración de luz con el consecuente beneficio para el desarrollo del
perifiton. Por otro lado, en sistemas oligotróficos, el epipelon puede explotar una fuente
extra de nutrientes, ya que puede tomarlos de los espacios intersticiales de los sedimentos,
lo que favorece su productividad por sobre la del fitoplancton (Liboriussen & Jepessen
2003). Las algas perifíticas pueden entonces ser responsables de una gran parte de la
producción primaria en los cuerpos de agua someros bien iluminados (Dodds et al 1999).
La mayor parte de los estudios donde se analiza la comparación de la
productividad de las dos comunidades hacen referencia al estado trófico del cuerpo de
agua (Vadeboncoeur et al. 2003, Liboriussen & Jepessen 2003, Hansson 1988). Por su
parte, Robinson et al (1997 a y b) estudiaron el efecto de la fluctuación en el nivel
hidrométrico sobre la biomasa fotosintética y productividad de las comunidades algales en
un humedal de Estados Unidos. Encontraron que el metafiton era la comunidad que
contribuía en mayor medida a la biomasa y productividad algal, y que el aumento en el
nivel hídrico favorecía al fitoplancton sobre las comunidades sésiles (epifiton y epipelon).
Para nuestro conocimiento, no hay muchos trabajos donde se compare la productividad
de fitoplancton y perifiton en sistemas húmicos. Kairesalo (1980) encontró que en el lago
húmico Pääjärvi, al sudeste de Finlandia, el fitoplancton poseía el mayor aporte a la
productividad primaria del lago, y que sólo en la zona litoral las algas epífitas y epipélicas
excedían la productividad del fitoplancton.

133
El objetivo general del trabajo que se presenta en este capítulo fue realizar una
aproximación al estudio de la producción primaria del fitoplancton y del perifiton sobre
sustratos artificiales en dos ambientes someros de la RNO (ROL 1, de aquí en adelante
ROL, y Laguna Grande) con distintas características limnológicas y lumínicas provocadas
principalmente por el contenido húmico y la cobertura de plantas flotantes. Se plantearon
las siguientes hipótesis:
- La producción integrada en la columna de agua para el fitoplancton y la producción del
perifiton a los 10 cm de profundidad por unidad de área de sustrato son mayores en la
Laguna Grande que en el ROL ya que éste presenta mayores restricciones lumínicas,
impuestas por el mayor contenido húmico y cobertura de plantas flotantes;
- El fitoplancton posee valores mayores de Pmáx y プ que el perifiton en los dos ambientes
probablemente debido a su capacidad de movilizarse en la columna y posicionarse en
condiciones de mejores irradiancias, a diferencia del perifiton que es sésil.
Para poner a prueba las hipótesis planteadas se llevaron a cabo los siguientes objetivos
específicos:
- Comparación espacial y temporal de la producción areal y los parámetros fotosintéticos
del fitoplancton y del perifiton obtenidos a partir de curvas P-I en ambos sitios, en
verano y en invierno;
- Evaluación de variables de estructura (composición a nivel de clases algales y
concentración de clorofila a del fitoplancton y perifiton además de las restantes variables
de masa del perifiton) en ambos sitios, en verano y en invierno.
MATERIALES Y MÉTODOS
Este trabajo se realizó en dos cuerpos de agua someros (profundidad máxima < 1 m) de
la Reserva Natural Otamendi. Uno de ellos fue la Laguna Grande (156 ha) y el otro el
mayor de los meandros abandonados (ROL, 17 ha). Se eligieron comparar dos períodos

134
estacionales opuestos, razón por la cual se llevó a cabo un muestreo en el invierno de
2006 y el otro en el verano 2007. Si bien durante todo ese año la cobertura de macrófitas
flotantes en la laguna Grande fue fluctuante, el sitio de muestreo se encontró libre de
cobertura durante las dos fechas de muestreo. Por su parte el ROL presentó una cubierta
del 80 % de su espejo de agua en invierno y del 100 % en el verano. Las macrófitas
flotantes dominantes fueron de pequeño porte tales como Ricciocarpus natans, Azolla
filiculoides, y lemnáceas (Wolffiella oblonga y Lemna minima entre otras) (Fig. 1). Ambos
sistemas presentaron características húmicas aunque el ROL presentó mayor contenido
húmico que la Laguna Grande: en la Laguna Grande la concentración de carbono
orgánico disuelto fue de alrededor de 30 mg L-1 mientras que en el ROL fue de 50 mg L-1
(datos presentados en el capítulo I de esta tesis).
Fig. 1. Detalle de la cobertura de macrófitas flotantes presentes en el ROL durante el
estudio.
En este estudio se analizaron las siguientes variables ambientales: profundidad, pH,
temperatura del agua, conductividad, oxígeno disuelto, Kd, g440, N-amonio, N-nitrato +
nitrito, N total, P-disuelto, P total y sólidos en suspensión. Los sensores utilizados así
como la toma de muestras y la metodología seguida para el análisis de estas variables se

135
explicitan en la páginas 72 a 74 del capítulo II. En el ROL, las mediciones de RFA
(radiación fotosintéticamente activa) se realizaron cada 5 cm en tantos puntos como fuera
posible dada la escasa profundidad del cuerpo de agua. Con estos datos se calculó el
coeficiente de atenuación vertical, Kd (Kirk 1994).
Fitoplancton
Se tomaron muestras de agua de cada sitio, que se fijaron con solución de lugol acético a
una concentración final de 1 %. Se realizó el análisis cuantitativo a nivel de clase algal para
el nano y microplancton bajo microscopio invertido con un error del 20 % (Ütermohl
1958, Venrick 1978) utilizándose para el reconocimiento de los individuos la bibliografía
ya mencionada oportunamente en esta tesis. Se estimó la concentración de clorofila a y se
llevaron a cabo las correspondientes curvas P-I con agua tomada de una zona de
transición entre la zona pelágica y litoral de la Laguna Grande y de la parte central en el
ROL. En el caso de las muestras de fitoplancton del ROL se inocularon 2 µCi de
NaH14CO3. Los parámetros de las curvas P-I fueron obtenidos por iteración utilizando el
modelo de Eilers & Peeters (1988). También se calculó la producción integrada en la
columna de agua. La metodología de cada variable fue descripta en el capítulo II (páginas
75 a 86).
Perifiton sobre sustratos artificiales
La comunidad perifítica se estudió mediante el método de los sustratos artificiales
permitiendo así la cuantificación de las variables de masa, densidad y producción primaria
del perifiton. El tipo de sustrato artificial empleado, así como sus dimensiones, el marco
para su sostén (perifitómetro) y su respectivo dispositivo de flotación (Fig. 2) fueron
similares a los que describieron en el capítulo II (páginas 82 a 84) para el perifiton de la
Laguna Grande. Al igual que en el capítulo anterior, a cada sustrato se le realizó una
marca neutra para la identificación de sus caras (Fig. 3). Los sustratos se dejaron colonizar
durante un mes antes de realizar las experiencias.
Se determinó la concentración de clorofila a del perifiton, el peso seco (PS), las
cenizas (C) y el peso seco libre de cenizas (PSLC) en tres sustratos de cada sitio, y se
calculó el índice autotrófico (IA) empleando la metodología descrita en el capítulo II
(páginas 82 a 84). El análisis estructural de la fracción algal del perifiton se realizó

136
mediante el análisis cuantitativo en muestras previamente raspadas del sustrato, llevadas a
un volumen conocido y fijadas con solución de lugol acético a una concentración final de
1 %. Los recuentos se efectuaron en microscopio invertido por el método de Ütermohl
(1958) con un 20 % de error (Venrick 1978). También se clasificó al perifiton en función
del sistema de Lakatos (1989), cuyas categorías se encuentran resumidas en la Tabla 1 del
capítulo II (página 84).
Fig. 2. Sistema de flotación (marco rectangular) empleado para la sujeción del
perifitómetro en el ROL.
Fig. 3. Sustrato artificial colonizado donde se muestran las marcas neutras (flechas)
empleadas para la identificación de las caras.

137
Las curvas P-I del perifiton de cada sitio se llevaron a cabo simultáneamente el
mismo día que las del fitoplancton y en el mismo dispositivo experimental (Fig. 1,
capítulo II, página 86). Se filtró el agua que se colocó en cada tubo por una red de 15 µm
de poro y se colocó un sustrato colonizado con la cara marcada hacia la luz; previamente
se eliminó con un bisturí la mayor parte posible del material adherido en la otra cara del
sustrato. En el caso del perifton del ROL se inocularon 3 µCi de NaH14CO3 en las
muestras. Considerando que los sustratos se colonizaron a 10 cm de profundidad, se
calculó entonces la producción por unidad de área a esa profundidad (PA 10 cm). El
procedimiento analítico y el ajuste de los datos para la obtención del carbono asimilado y
la PA 10 cm fueron los mismos empleados para el perifiton de la Laguna Grande y que se
describieron en el capítulo II.
Se emplearon test no paramétricos (Mann-Whitney) con el objetivo de explorar si
había diferencias entre las variables perifíticas entre sitios y épocas del año y entre los
parámetros fotosintéticos (Pmáx, プ, Ik) entre sitios y comunidades (Zar 1996).
RESULTADOS
En la Tabla 1 se muestran los valores obtenidos para las variables ambientales en cada
sitio de muestreo durante el estudio. Los valores de profundidad fueron en ambos sitios
mayores en verano (aguas altas) que en invierno (aguas bajas), patrón seguido por los
valores de conductividad, que fueron más altos en invierno. Tanto en el ROL como en la
Laguna Grande la concentración de oxígeno disuelto fue mayor en invierno que en
verano, llegando a registrarse anoxia debajo de la cobertura macrofítica del ROL en
verano. Tanto en el ROL como en la Laguna Grande la temperatura y el contenido
húmico registraron valores más altos en verano.
Las aguas del ROL tuvieron valores de pH superiores a 8 en tanto que en la
laguna, los valores fueron circumneutrales y en ambos sitios la alcalinidad fue elevada. En
el ROL, la concentración de sólidos en suspensión fue dos órdenes de magnitud mayor en
verano con respecto al invierno en tanto que en la laguna, los valores obtenidos fueron
muy similares entre invierno y verano. La irradiancia subsuperficial (I0) y en menor
medida la irradiancia a los 10 cm de profundidad (I10 cm) presentaron valores más altos en

138
verano en la Laguna Grande, mientras que en el ROL ocurrió lo inverso, debido a la
mayor cobertura de macrofitia presente en este sitio en verano. La irradiancia integrada en
la columna de agua (Imedia) fue mayor en la Laguna Grande que en el ROL en los dos
muestreos mientras que el coeficiente de atenuación vertical (Kd) fue siempre mayor en el
ROL (Tabla 1).
En la Laguna Grande en invierno, el 90 % del N total se encontraba en la forma de
NID mientras que en el ROL el 85 % del N total era NID. En verano sucedió lo
contrario, en la Laguna Grande todo el nitrógeno se encontraba en forma orgánica o
particulada, mientras que en el ROL, el NID era sólo el 1 % del N total. La misma
tendencia se observó en cuanto a las concentraciones de P-disuelto y P total. En la laguna
en invierno el 50 % del P total se hallaba en la forma de P-disuelto mientras que en
verano este porcentaje fue del 30 %. En el ROL en invierno todo el fósforo se encontraba
en la forma disuelta y en verano sólo el 12%. Por último, el ROL presentó
concentraciones más elevadas de nutrientes, con la excepción del P-disuelto en invierno
(Tabla 1).
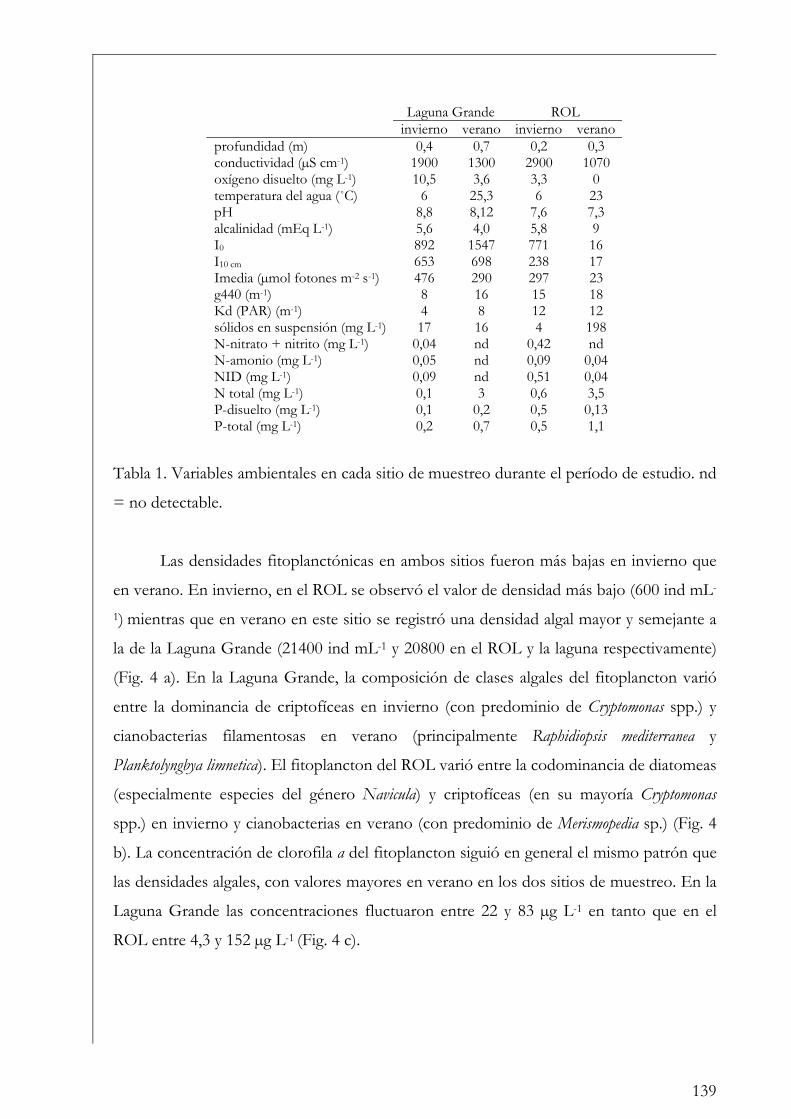
139
Laguna Grande ROL invierno verano invierno verano profundidad (m) 0,4 0,7 0,2 0,3 conductividad (µS cm-1) 1900 1300 2900 1070 oxígeno disuelto (mg L-1) 10,5 3,6 3,3 0 temperatura del agua (˚C) 6 25,3 6 23 pH 8,8 8,12 7,6 7,3 alcalinidad (mEq L-1) 5,6 4,0 5,8 9 I0 892 1547 771 16 I10 cm 653 698 238 17 Imedia (µmol fotones m-2 s-1) 476 290 297 23 g440 (m-1) 8 16 15 18 Kd (PAR) (m-1) 4 8 12 12 sólidos en suspensión (mg L-1) 17 16 4 198 N-nitrato + nitrito (mg L-1) 0,04 nd 0,42 nd N-amonio (mg L-1) 0,05 nd 0,09 0,04 NID (mg L-1) 0,09 nd 0,51 0,04 N total (mg L-1) 0,1 3 0,6 3,5 P-disuelto (mg L-1) 0,1 0,2 0,5 0,13 P-total (mg L-1) 0,2 0,7 0,5 1,1
Tabla 1. Variables ambientales en cada sitio de muestreo durante el período de estudio. nd
= no detectable.
Las densidades fitoplanctónicas en ambos sitios fueron más bajas en invierno que
en verano. En invierno, en el ROL se observó el valor de densidad más bajo (600 ind mL-
1) mientras que en verano en este sitio se registró una densidad algal mayor y semejante a
la de la Laguna Grande (21400 ind mL-1 y 20800 en el ROL y la laguna respectivamente)
(Fig. 4 a). En la Laguna Grande, la composición de clases algales del fitoplancton varió
entre la dominancia de criptofíceas en invierno (con predominio de Cryptomonas spp.) y
cianobacterias filamentosas en verano (principalmente Raphidiopsis mediterranea y
Planktolyngbya limnetica). El fitoplancton del ROL varió entre la codominancia de diatomeas
(especialmente especies del género Navicula) y criptofíceas (en su mayoría Cryptomonas
spp.) en invierno y cianobacterias en verano (con predominio de Merismopedia sp.) (Fig. 4
b). La concentración de clorofila a del fitoplancton siguió en general el mismo patrón que
las densidades algales, con valores mayores en verano en los dos sitios de muestreo. En la
Laguna Grande las concentraciones fluctuaron entre 22 y 83 µg L-1 en tanto que en el
ROL entre 4,3 y 152 µg L-1 (Fig. 4 c).
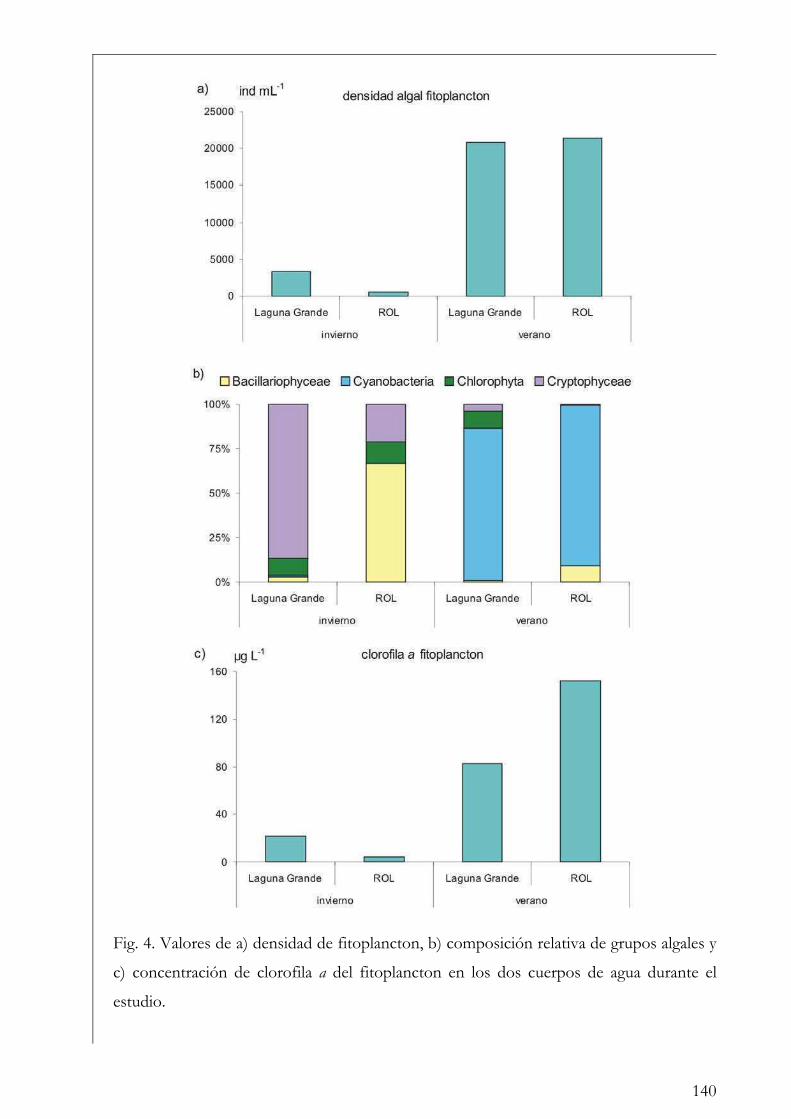
140
Fig. 4. Valores de a) densidad de fitoplancton, b) composición relativa de grupos algales y
c) concentración de clorofila a del fitoplancton en los dos cuerpos de agua durante el
estudio.
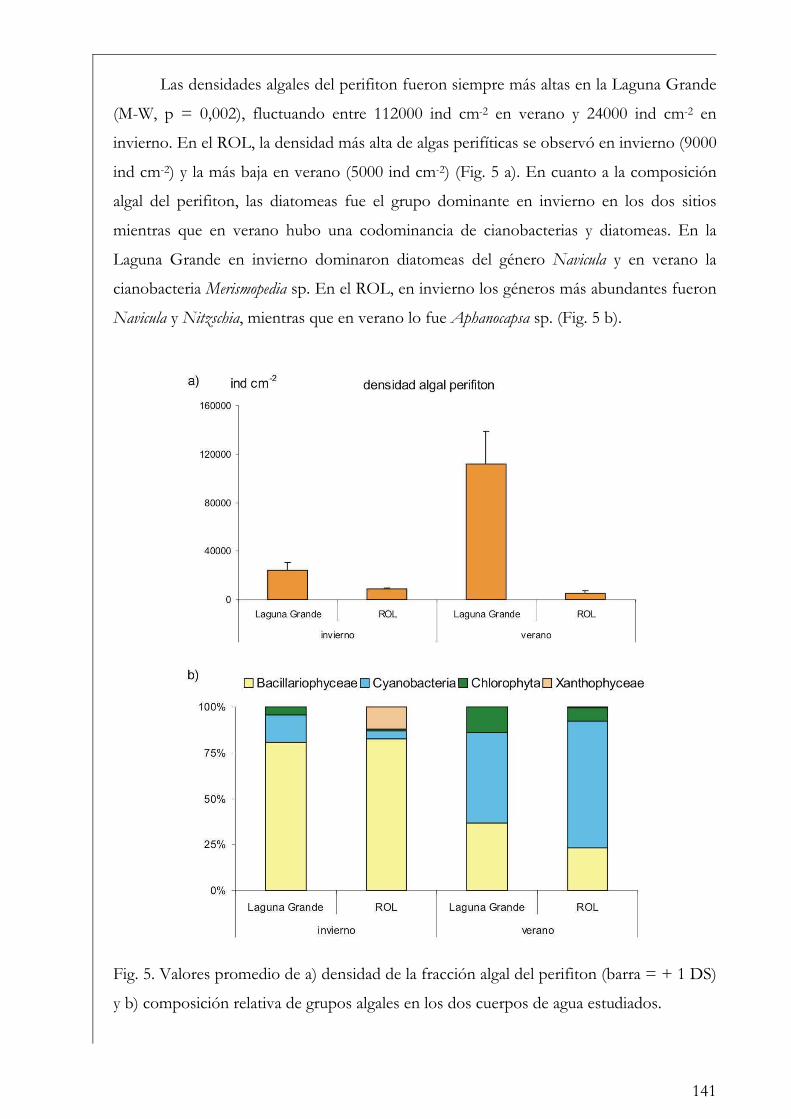
141
Las densidades algales del perifiton fueron siempre más altas en la Laguna Grande
(M-W, p = 0,002), fluctuando entre 112000 ind cm-2 en verano y 24000 ind cm-2 en
invierno. En el ROL, la densidad más alta de algas perifíticas se observó en invierno (9000
ind cm-2) y la más baja en verano (5000 ind cm-2) (Fig. 5 a). En cuanto a la composición
algal del perifiton, las diatomeas fue el grupo dominante en invierno en los dos sitios
mientras que en verano hubo una codominancia de cianobacterias y diatomeas. En la
Laguna Grande en invierno dominaron diatomeas del género Navicula y en verano la
cianobacteria Merismopedia sp. En el ROL, en invierno los géneros más abundantes fueron
Navicula y Nitzschia, mientras que en verano lo fue Aphanocapsa sp. (Fig. 5 b).
Fig. 5. Valores promedio de a) densidad de la fracción algal del perifiton (barra = + 1 DS)
y b) composición relativa de grupos algales en los dos cuerpos de agua estudiados.

142
La concentración de clorofila a del perifiton fue siempre mayor en la laguna (M-
W, p = 0,002) y en los dos sitios los valores más altos se detectaron en verano (Fig. 6 a).
La menor concentración estimada fue de 0,31 µg cm-2 en el ROL en invierno, y la más alta
fue 3,55 µg cm-2 en la laguna en verano. El peso seco (PS), las cenizas (C) y el peso seco
libre de cenizas (PSLC) siguieron el mismo patrón general con valores más altos en
verano. Los valores obtenidos para el PS, C y el porcentaje de cenizas se encuentran en la
Tabla 2. Los valores de PS fueron significativamente más altos en la Laguna Grande que
en el ROL (M-W, p = 0,002). La concentración de cenizas fue significativamente mayor
en la Laguna Grande (M-W, p = 0,002). El rango de variación de los promedios del PSLC
fue de 0,094 mg cm-2 para el ROL en invierno y 3,71 mg cm-2 para la laguna en verano,
siendo significativamente más altos en la Laguna Grande (M-W, p = 0,002) (Fig. 6 b). El
índice autotrófico siguió un patrón estacional inverso cuando se comparan los dos sitios.
En invierno fue mayor en el ROL y en verano en la Laguna Grande. Es decir, que la
laguna fue en composición perifítica más autotrófica en invierno mientras que el ROL lo
fue en verano (Tabla 2). No se detectaron diferencias significativas entre los valores del
índice autotrófico entre los dos sitios de muestreo. En cuanto a la clasificación propuesta
por Lakatos (1989) en relación a los valores de peso seco, porcentaje de cenizas y
porcentaje de clorofila a en el peso seco, los resultados se muestran en la Tabla 2.
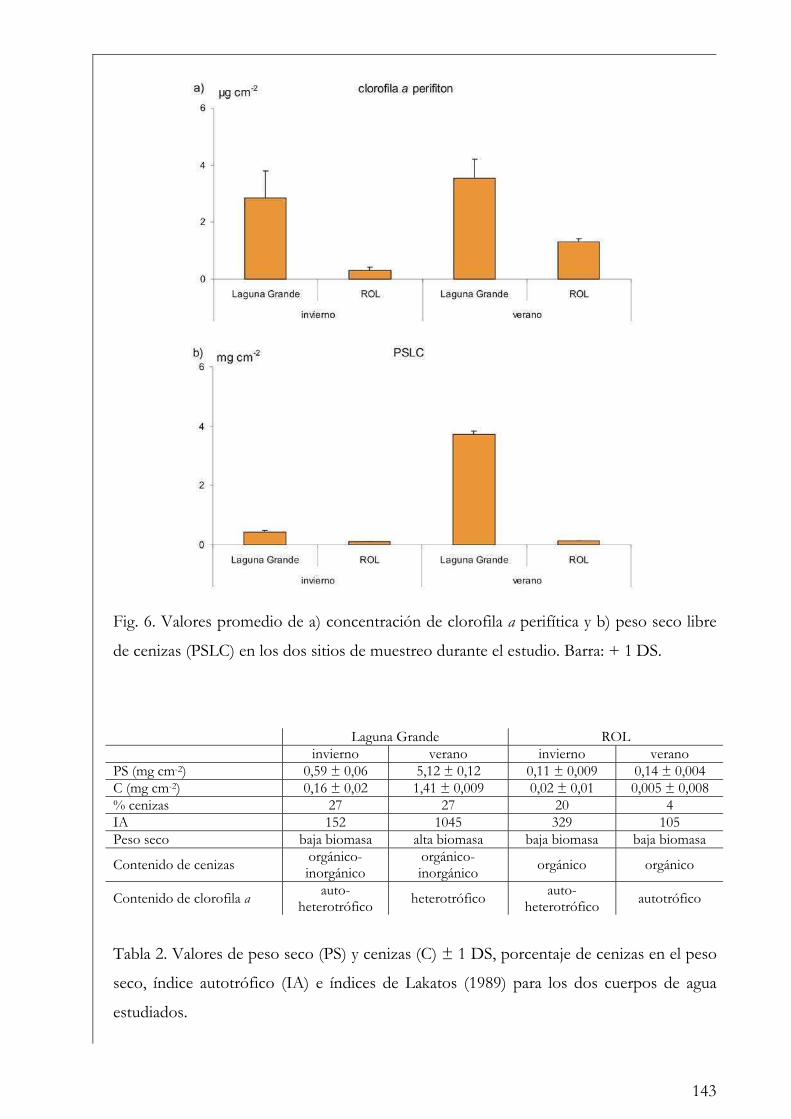
143
Fig. 6. Valores promedio de a) concentración de clorofila a perifítica y b) peso seco libre
de cenizas (PSLC) en los dos sitios de muestreo durante el estudio. Barra: + 1 DS.
Laguna Grande ROL invierno verano invierno verano PS (mg cm-2) 0,59 ± 0,06 5,12 ± 0,12 0,11 ± 0,009 0,14 ± 0,004 C (mg cm-2) 0,16 ± 0,02 1,41 ± 0,009 0,02 ± 0,01 0,005 ± 0,008 % cenizas 27 27 20 4 IA 152 1045 329 105 Peso seco baja biomasa alta biomasa baja biomasa baja biomasa
Contenido de cenizas orgánico-inorgánico
orgánico-inorgánico
orgánico orgánico
Contenido de clorofila a auto-
heterotrófico heterotrófico
auto-heterotrófico
autotrófico
Tabla 2. Valores de peso seco (PS) y cenizas (C) ± 1 DS, porcentaje de cenizas en el peso
seco, índice autotrófico (IA) e índices de Lakatos (1989) para los dos cuerpos de agua
estudiados.

144
El número de asimilación (Pmáx) en general alcanzó valores más altos para el
fitoplancton en ambos sitios con la excepción del verano en el ROL, cuando el perifiton
alcanzó un valor mayor (0,97 µg C (µg hl a h)-1) (Fig. 7 a). El valor más alto de Pmáx fue
observado para el fitoplancton de la Laguna Grande en verano (12 µg C (µg hl a h)-1) y el
más bajo para el perifiton de la Laguna Grande en verano (0,14 µg C (µg hl a h)-1). Los
valores de Pmáx para el fitoplancton de la Laguna Grande y el perifiton del ROL fueron
mayores en verano, mientras que en invierno el fitoplancton alcanzó mayores valores en
ambos sitios. La eficiencia fotosintética (プ) siguió el mismo patrón espacial y temporal que
Pmáx (Fig. 7 b), siendo el máximo registrado para el fitoplancton de la laguna en verano
(0,04 µg C m2 s (µg hl a h µmol fotones)-1) y el más bajo para el perifiton de la laguna en
verano (0,0002 µg C m2 s (µg hl a h µmol fotones)-1). No se detectaron diferencias
significativas entre los valores de Pmáx y プ de ambas comunidades (uniendo los datos de
ROL y laguna) ni entre los valores de ROL y laguna (uniendo ambas comunidades).
En la Laguna Grande, los valores de irradiancia al inicio de la saturación
fotosintética (Ik) fueron más altos para el perifiton (Fig. 8). Los mayores valores de Ik se
observaron para ambas comunidades en la Laguna Grande en verano, siendo el valor
máximo el correspondiente al perifiton (418 µmol fotones m-2 s-1). El valor más bajo se
observó para el fitoplancton de la laguna en invierno (63 µmol fotones m-2 s-1). En el caso
de la comunidad perifítica, los valores más elevados se observaron en la laguna, mientras
que para el fitoplancton el Ik fue mayor en el ROL en invierno y mayor en la laguna en
verano. Se observó que ambas comunidades estuvieron limitadas lumínicamente en el
ROL en verano. Finalmente, no se detectaron diferencias significativas entre los valores
de Ik de ambas comunidades (unificando los datos de ROL y laguna) ni entre los valores
de ROL y laguna (unificando ambas comunidades).
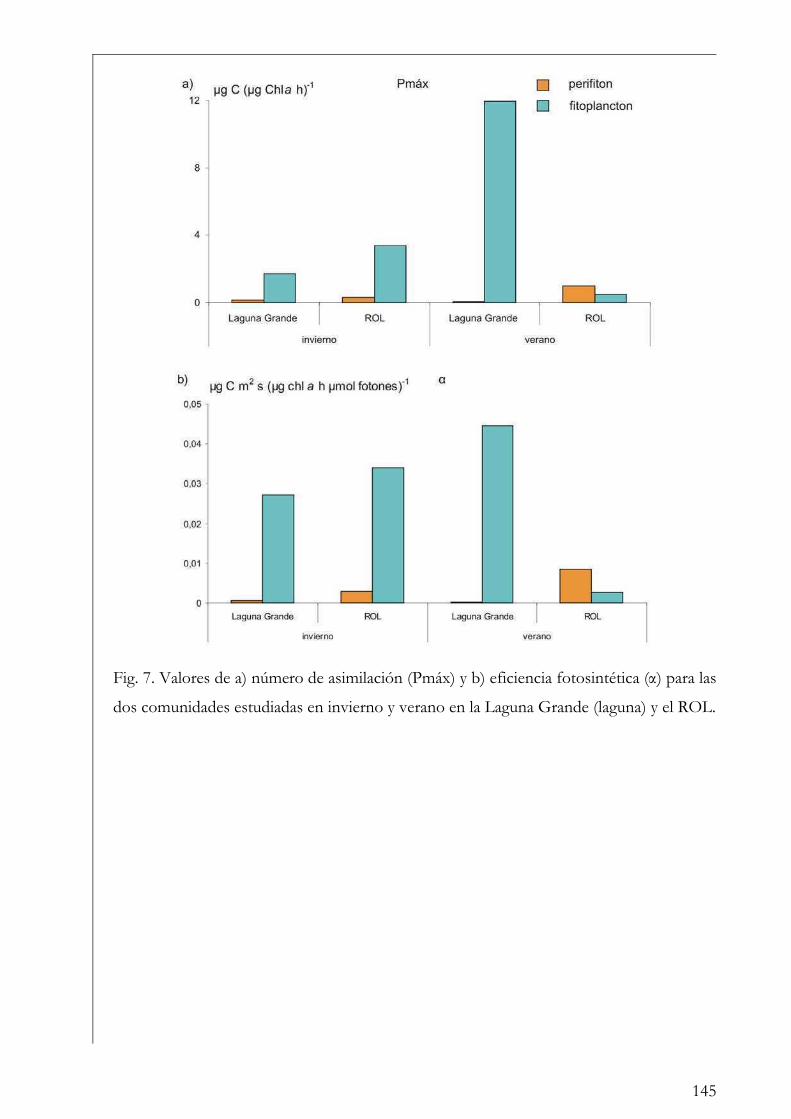
145
Fig. 7. Valores de a) número de asimilación (Pmáx) y b) eficiencia fotosintética (プ) para las
dos comunidades estudiadas en invierno y verano en la Laguna Grande (laguna) y el ROL.
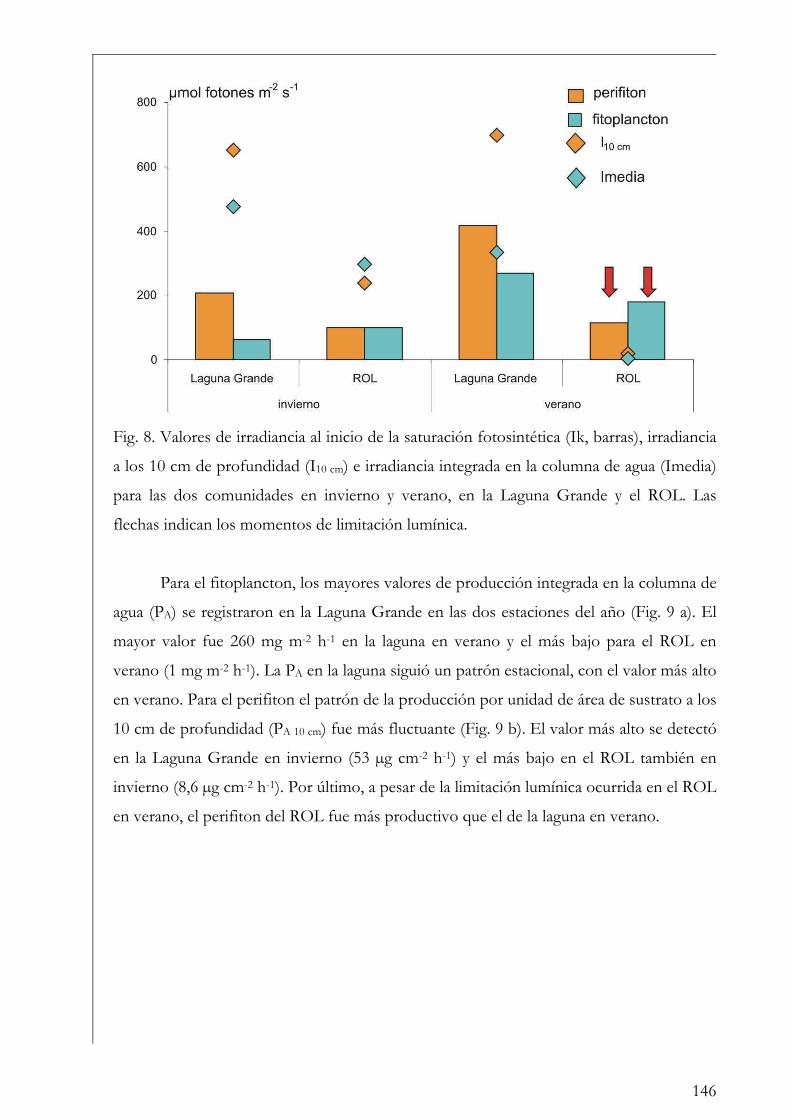
146
Fig. 8. Valores de irradiancia al inicio de la saturación fotosintética (Ik, barras), irradiancia
a los 10 cm de profundidad (I10 cm) e irradiancia integrada en la columna de agua (Imedia)
para las dos comunidades en invierno y verano, en la Laguna Grande y el ROL. Las
flechas indican los momentos de limitación lumínica.
Para el fitoplancton, los mayores valores de producción integrada en la columna de
agua (PA) se registraron en la Laguna Grande en las dos estaciones del año (Fig. 9 a). El
mayor valor fue 260 mg m-2 h-1 en la laguna en verano y el más bajo para el ROL en
verano (1 mg m-2 h-1). La PA en la laguna siguió un patrón estacional, con el valor más alto
en verano. Para el perifiton el patrón de la producción por unidad de área de sustrato a los
10 cm de profundidad (PA 10 cm) fue más fluctuante (Fig. 9 b). El valor más alto se detectó
en la Laguna Grande en invierno (53 µg cm-2 h-1) y el más bajo en el ROL también en
invierno (8,6 µg cm-2 h-1). Por último, a pesar de la limitación lumínica ocurrida en el ROL
en verano, el perifiton del ROL fue más productivo que el de la laguna en verano.
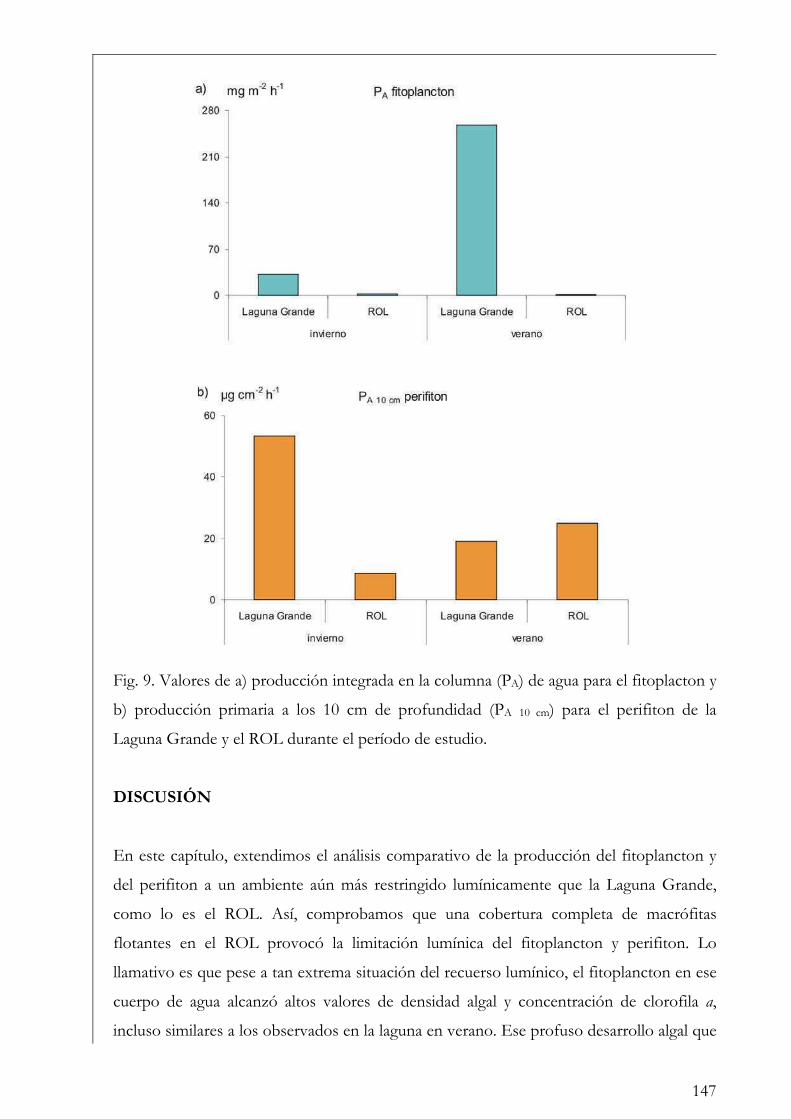
147
Fig. 9. Valores de a) producción integrada en la columna (PA) de agua para el fitoplacton y
b) producción primaria a los 10 cm de profundidad (PA 10 cm) para el perifiton de la
Laguna Grande y el ROL durante el período de estudio.
DISCUSIÓN
En este capítulo, extendimos el análisis comparativo de la producción del fitoplancton y
del perifiton a un ambiente aún más restringido lumínicamente que la Laguna Grande,
como lo es el ROL. Así, comprobamos que una cobertura completa de macrófitas
flotantes en el ROL provocó la limitación lumínica del fitoplancton y perifiton. Lo
llamativo es que pese a tan extrema situación del recuerso lumínico, el fitoplancton en ese
cuerpo de agua alcanzó altos valores de densidad algal y concentración de clorofila a,
incluso similares a los observados en la laguna en verano. Ese profuso desarrollo algal que

148
pudo ser la respuesta a la penetración de luz por los espacios entre las plantas flotantes
(Fig. 1) que posibilitó el crecimiento, no se visualizó en los valores de producción. La
comunidad fitoplanctónica en el ROL fue menos productiva que en la laguna y los bajos
valores de PA obtenidos en el ROL en verano podrían estar relacionados entonces a una
particular composición algal. El fitoplancton del ROL estuvo compuesto en ese verano
principalmente por agregados de cianobacterias a diferencia de las cianobacterias
filamentosas que se desarrollaron en la Laguna Grande y que fueron más productivas. La
comunidad de la laguna también resultó tener mayor eficiencia fotosintética optimizando
la luz recibida por unidad de biomasa productiva. Por otra parte, la mayor representación
de formas más pequeñas de cianobacterias en el ROL probablemente esté relacionada con
la ventaja adaptativa que presentan los organismos de menor tamaño en relación a la
captación de luz (Raven 1998). El efecto paquete es menor en células más pequeñas, y
ésto tiene que ver con la pérdida de eficiencia de los pigmentos fotosintéticos en la
absorción lumínica cuando se encuentran empaquetados en unidades estructurales (Kirk
1994).
El perifiton también manifestó particularidades en el ROL ya que, aunque estuvo
un orden de magnitud menos desarrollado y también limitado lumínicamente en verano,
alcanzó valores de Pmáx, プ y PA 10 cm ligeramente mayores a los observados para el
perifiton de la laguna que no estaba limitado lumínicamente. Los parámetros
fotosintéticos del perifiton del ROL en verano fueron también mayores a los del
fitoplancton del mismo sitio. Estas características del perifiton del ROL probablemente
guarden relación con el carácter autotrófico que tuvo la comunidad en ese verano, que
implica que los procesos de producción superaban los de pérdida dentro de la misma
matriz perifítica. El perifiton de la laguna si bien estaba más desarrollado, alcanzando el
mayor valor de peso seco del estudio, tuvo un gran desarrollo del componente
heterotrófico (bacterias, protozoos, animales, etc.) que se visualizó en el valor de índice
autotrófico, corroborado también con la clasificación de tipo heterotrófico de Lakatos
(1989). Es decir, que por unidad de superficie colonizada, el "poco" perifiton desarrollado
en el ROL produjo más y de manera más eficiente.
Si bien ambos cuerpos de agua son húmicos, el clima lumínico del ROL fue más
restringido aún que el de la Laguna Grande; porque además de presentar una importante
cobertura de macrófitas flotantes durante los dos muestreos, el contenido en ácidos

149
húmicos y Kd (en invierno) fueron mayores. De acuerdo a estas características, la flora del
ROL debe presentar una gama de adaptaciones que le permita prosperar en estas
condiciones. En este sentido, Izaguirre et al. (2001 b) estudiaron el fitoplancton de los
ROLs y encontraron que las especies que habitaban en estos sitios eran reconocidas como
tolerantes a bajos tenores de oxígeno disuelto y bajas irradiancias. Observaron, como lo
hicimos nosotros en este estudio, que los organismos que constituían el fitoplancton del
ROL pertenecían principalmente a las clases Cyanobacteria (órdenes Chroococcales y
Oscilatoriales) (alrededor del 80 % de la densidad total de organismos) y Bacillariophyceae
(en particular, diatomeas pennadas).
El hecho de que los valores de los parámetros fotosintéticos Pmáx y プ del
fitoplancton del ROL tendieran a ser mayores en invierno en comparación al verano
podría estar relacionado con la mayor disponibilidad de nutrientes inorgánicos en esa
época del año (Falkowski 1981) o bien con la mayor disponibilidad de luz debido a una
cobertura menos tupida de macrófitas flotantes, ya que en verano el fitoplancton del ROL
se encontró limitado lumínicamente por las bajas irradiancias.
Para poder prosperar en ambientes húmicos y por ende de irradiancia limitada en
cuanto a la intensidad y calidad (Eloranta 1999, Jones 1992), las algas deben estar
adaptadas mediante distintos tipos de estrategias a la explotación del ambiente abiótico en
estas circunstancias. En el fitoplancton es común observar adaptaciones para el
desplazamiento en la columna de agua para obtener mayores irradiancias. Ejemplos de
este tipo de estrategia lo constituyen la posesión de flagelo o vesículas gaseosas que
permiten regular la flotabilidad a algunas cianobacterias (Kirk 1994). En el fitoplancton,
también pueden encontrarse distintas estrategias nutricionales, como por ejemplo la
mixotrofia, que representa una ventaja en ambientes como la Laguna Grande, donde los
tenores de NID son bajos (Sinistro et al. 2006, Jansson et al. 1996). Las adaptaciones en el
perifiton son principalmente de tipo fisiológicas, regulando sus mecanismos fotosintéticos
y si bien su hábito sésil les impide desplazarse en la columna de agua sí lo pueden hacer en
la propia matriz perifítica siendo ésta una estrategia para optimizar la obtención del
recurso (Hill 1996).
La concentración de clorofila a perifítica fue mayor en la Laguna Grande en los
dos muestreos, patrón que siguió también la densidad algal. Los valores de biomasa
fotosintética perifítica fueron bajos cuando se los compara con los obtenidos por Pizarro

150
(1999) para epifiton sobre Echinochloa polystachya en una laguna del valle de inundación del
Bajo Paraná, por Leandrini & Rodrigues (2008) para epifiton de ambientes semilóticos
del Paraná Superior y por Putz (1997) para perifiton sobre sustratos artificiales en el valle
aluvial del río Amazonas; pero dentro de los valores obtenidos por Scott et al. (2005) en
un humedal con limitación de nutrientes (N y P). Por otro lado, Jones et al. (2002)
obtuvieron valores entre 1,2 y 3,9 µg cm-2 para comunidades perifíticas sujetas a una
intensa presión de pastoreo. Es decir que los valores obtenidos por nosotros se asemejan
a los obtenidos en estudios donde la comunidad perifítica estaba sujeta a grandes pérdidas
debidas a la predación o la limitación de nutrientes. En nuestro caso, probablemente los
bajos valores de biomasa fotosintética en el ROL estén relacionados con las bajas
irradiancias, aunque sería necesario explorar el efecto del pastoreo.
Las estimaciones realizadas del número de asimilación (Pmáx) y la PA 10 cm para el
perifiton en la Laguna Grande y el ROL fueron bajas cuando se las compara con las
obtenidas por Putz (1997) para sistemas húmicos de la llanura aluvial del río Amazonas.
Arvola et al. (1999) encontraron una relación inversa entre la asimilación del fitoplancton y
el contenido húmico en lagos finlandeses y tal vez sucede lo mismo con el perifiton, si
bien no encontramos referencias que sustenten esta hipótesis y se requieren más estudios
en este sentido para comprobarlo.
Los valores de irradiancia al inicio de la saturación fotosintética (Ik) estuvieron,
para la comunidad algal perifítica, aproximadamente en el rango 100-400 µmol fotones m-
2 s-1, que es donde se encuentra la mayor parte de los valores referidos en la bibliografía
(Hill 1996). La misma observación se aplica para los valores de Ik del fitoplancton (Kirk
1994). Por otra parte, el hecho que los valores de Ik del perifiton en la Laguna Grande
hayan tendido a ser mayores a los del fitoplancton podría tener relación con que el
perifiton estudiado se desarrolló a los 10 cm de profundidad, ubicación que pudo haberle
permitido obtener mayores irradiancias que el fitoplancton, que en condiciones
supraóptimas de iluminación puede desplazarse en la columna de agua. Si bien no se
detectaron diferencias significativas entre ambientes, se observó en general una tendencia
a valores más bajos de Ik en el ROL para ambas comunidades, lo que también indicaría
mayor fotoaclimatación a bajas intensidades lumínicas (Henley 1993).

151
En la Laguna Grande, el principal factor modelador de la productividad primaria y
dinámica fitoplanctónica fue la temperatura, como se comprobó en el capítulo II de esta
tesis, mientras que en el ROL el factor principal pareció ser la disponibilidad de luz, de
acuerdo a lo que se puede observar de los resultados de este trabajo. Quedó de manifiesto
que en ambientes naturales no siempre una cobertura completa de macrófitas flotantes
genera comunidades microbianas poco desarrolladas como advertimos en el ROL.

152
CAPÍTULO IV
COMPARACIÓN DE LA PRODUCCIÓN PRIMARIA DE DOS FRACCIONES
DE TAMAÑO FITOPLANCTÓNICAS EN TRES CUERPOS DE AGUA DE LA
RNO

153
INTRODUCCIÓN
La primera subdivisión del fitoplancton en categorías de tamaño data de 1892 (Schüt
1892, en Callieri 2008) aunque la distinción entre microplancton, nanoplancton y
ultraplancton no fue introducida sino hasta los años cincuenta del siglo siguiente, con el
advenimiento de mallas y filtros de distintos tamaños de poro con los que se logró el
fraccionamiento de los organismos. Si bien Sieburth et al. (1978) acuñaron por primera
vez los términos picoplancton y femtoplancton para el componente heterotrófico del
plancton, esta terminología se extendió luego al fitoplancton. La nomenclatura empleada
actualmente para clasificar a los organismos planctónicos de acuerdo a su tamaño celular
puede resumirse en la siguiente tabla (Callieri 2008):
tamaño (µm)
Microplancton 20-200
Nanoplancton 2-20
Picoplancton 0,2-2
Femtoplancton 0,02-0,2
La clasificación del fitoplancton en fracciones de tamaño permite profundizar en
su estudio y de esta manera apreciar las diferencias fisiológicas y ecológicas entre los
grupos de organismos de distinto tamaño celular. Dentro de la fracción autotrófica del
picoplancton, denominada picofitoplancton, se encuentran fotótrofos como las
picocianobacterias y picoeucariotas, desde células solitarias hasta microcolonias de hasta
50 células que se encuentran en casi todos los tipos de cuerpos de agua de distinto estado
trófico (Stockner et al. 2000). El picofitoplancton consiste de varios grupos de algas pero
está principalmente representado por cianobacterias cocoides unicelulares de los géneros
Synechococcus y Synechocystis (Stockner & Antia 1986).
La adquisición de recursos de parte del fitoplancton depende del tamaño de los
organismos, que puede tener grandes efectos en la fisiología y tasas de crecimiento celular,
alterando la abundancia y estructura de tamaños de la comunidad. Se ha demostrado
experimentalmente que los organismos más pequeños poseen tasas más altas de
incorporación de nutrientes por unidad de biomasa y constantes de saturación media más
bajas que los organismos más grandes debido a la mayor relación superficie a volumen

154
(S/V). Las células más pequeñas poseen requerimientos metabólicos celulares mínimos
más bajos que las células más grandes, lo que les permite prosperar y sobrevivir cuando la
disponibilidad de recursos es baja (Irwin et al. 2006). Además, la capa límite difusiva es
menor alrededor de las células más pequeñas, lo que incrementa la disponibilidad de los
nutrientes en mayor medida cuando éstos se encuentran en bajas concentraciones (Raven
1998).
El resultado de la interacción de los organismos con la irradiancia también
depende del tamaño celular. Los diámetros de las células picoplanctónicas (600-2000 nm)
se encuentran en el orden de las correspondientes a las longitudes de onda visible (400-
700 nm), de modo que el efecto paquete es mínimo, lo que les permite una mayor
absorción por unidad de pigmento en comparación con células de mayor tamaño (Raven
1998). El efecto paquete se refiere a la disminución en la efectividad de absorción
lumínica por parte de los pigmentos fotosintéticos cuando éstos se encuentran dentro de
unidades discretas (células, cloroplastos) en comparación a cuando se encuentran
uniformemente distribuidos en solución (Kirk 1994). Por otro lado, el menor efecto
paquete en las células de menor tamaño también implica un daño mayor por parte de la
radiación visible y ultravioleta (Raven 1998). En este sentido, Helbling et al. (2001 a y b)
comprobaron que en general las células más pequeñas son más resistentes a la inhibición
de la fotosíntesis, pero son más vulnerables al daño en el DNA por parte de la radiación
ultravioleta. En cambio, el microplancton es más sensible a la inhibición fotosintética pero
más resistente al daño en el DNA. También Helbling et al. (2001 a) mostraron que el
tamaño celular es muy importante en la determinación del estado de aclimatación de las
células a nuevas condiciones de irradiancia, demostrando que las células nano y
picoplanctónicas se aclimatan mucho más rápido que las del microplancton. Además,
dada la mayor relación S/V de las células más pequeñas, éstas estarían mejor adaptadas a
la utilización de la radiación en ambientes con gran atenuación de la radiación solar. En
cuerpos de agua con restricciones lumínicas como los que se encuentran en el humedal de
la RNO, los procesos de mezcla producen mayor variabilidad en las condiciones de
irradiancia que en cuerpos de agua relativamente claros. Los procesos metabólicos de las
células deben ajustarse rápidamente a esas fluctuaciones de irradiancia para de esta manera
poder prosperar en estos sitios tan cambiantes (Helbling et al. 2001 a).
La temperatura es otro factor importante que influencia la estructura en fracciones
de tamaño del fitoplancton (Jasser & Arvola 2003). Por ejemplo, Jasser & Arvola (2002)

155
estudiaron la dinámica estacional de abundancias del picofitoplancton y del fitoplancton
de mayor tamaño en lagos boreales con distinto contenido húmico en Finlandia. Los
autores detectaron la máxima densidad del picoplancton en verano y observaron también
un pico primaveral desfasado con respecto al fitoplancton de mayor tamaño. Sugirieron
que este defasaje podría deberse a la sensibilidad del picoplancton autotrófico a las bajas
temperaturas. En este sentido, y según los estudios realizados en latitudes templadas, el
picoplancton autotrófico suele presentan un patrón estacional marcado, con dos picos de
abundancia anuales: uno en primavera o a principios del verano y otro en otoño (Callieri
& Stockner 2002).
La contribución relativa del picoplancton, nano y microplancton a la biomasa,
densidad y producción primaria fitoplanctónica de los cuerpos de agua parece ser una
función del estado trófico de los mismos. En este sentido, la abundancia y biomasa
picoplanctónica aumenta a medida que lo hace el estado trófico de los cuerpos de agua
dulce y en el mar. Sin embargo, su contribución relativa a la biomasa total de organismos
y productividad primaria total disminuye (Bell & Kalff 2001, Agawin et al. 2000, Jasser
1997). En este sentido, su aporte a la productividad suele ser más importante en los
sistemas pobres en nutrientes (Raven 1998). Vörös et al. (1991) encontraron que el
picoplancton autotrófico aportaba hasta el 50 % de la producción primaria en algunos
lagos mesotróficos e hipereutróficos de Hungría. Greisberger et al. (2008) analizaron el
aporte de tres fracciones de tamaño fitoplanctónicas (< 0,2 µm, 2-11 µm y > 11 µm) a la
concentración de clorofila a y productividad fitoplanctónicas en un lago alpino
mesotrófico de Austria. Encontraron que el aporte del picoplancton (< 2 µm) dependía
de la estacionalidad, contribuyendo con hasta un 58 % de la productividad primaria en
otoño. Steitz & Velimirov (1999) señalaron que el picoplancton llegó a contribuir con el
74 % (con una media del 35 %) de la fijación de carbono total en un lago somero
oligotrófico de Austria durante la estación de crecimiento.
El picofitoplancton en los cuerpos de agua de la Reserva Natural Otamendi está
constituido mayoritariamente por picocianobacterias (Izaguirre et al. enviado, Sinistro et al.
2007, 2006). Sinistro et al. (2006) observaron que en la Laguna Grande el picoplancton
autotrófico era predado principalmente por algas mixotróficas, nanoflagelados
heterotróficos y ciliados. También vieron que los picoeucariotas constituían una pequeña
fracción del picoplancton, principalmente representados por células tipo Chlorella sp. Por
su parte, O´Farrell et al. (2007) encontraron que la abundancia de picofitoplancton en los

156
sitios con mayor contenido húmico (principalmente los ROLs) tendía a ser más baja. Por
último, bajo condiciones de anoxia en el ROL y en mesocosmos en la Laguna Grande se
ha advertido la presencia de bacterias fotosintéticas anaeróbicas (Izaguirre et al. enviado).
En este capítulo se analizaron los parámetros fotosintéticos y la contribución relativa de
las fracciones de tamaño comprendidas entre 0,2 y 3 µm (picoplancton) y la > a 3 µm
(nano y micoplancton) a la productividad fitoplanctónica. Los muestreos se llevaron a
cabo básicamente en la Laguna Grande y se realizaron comparaciones puntuales con un
ROL y la laguna El Pescado, tres cuerpos de agua de la RNO con características
limnológicas contrastadas. Se pusieron a prueba las siguientes hipótesis:
- la contribución del picoplancton a la producción fitoplanctónica en la Laguna Grande es
menor que la del nano y micoplancton dado su elevado estado trófico;
- la producción primaria del picofitoplancton y de la fracción > 3 µm presentará, en
términos de producción integrada en la columna de agua (PA) un patrón de fluctuación
estacional en la Laguna Grande;
- los parámetros fotosintéticos プ y Pmáx serán mayores para el picofitoplancton, mientras
que el parámetro Ik será menor para estos organismos en los tres cuerpos de agua por
estar mejor adaptados bajas irradiancias;
- la producción total integrada en la columna de agua será más baja en el ROL que en los
otros dos cuerpos de agua debido a las altas concentraciones de ácidos húmicos y
cobertura de plantas la mayor parte del año;
- en la Laguna Grande y El Pescado la dinámica de la productividad será similar por
compartir estas lagunas características limnológicas similares.
Para poner a prueba las hipótesis planteadas, se llevaron a cabo los siguientes objetivos
específicos:
- determinación de las variables ambientales en los tres cuerpos de agua

157
- estudio de la variación anual de los parámetros fotosintéticos a través de la realización de
curvas P-I, estimación de la concentración de clorofila a y las tasas de producción
primaria integrada en la columna de agua (PA) y volumétrica (Pmáx vol) de las fracciones
de tamaño > 3 µm y la comprendida entre 0,2 y 3 µm en la Laguna Grande, analizando su
fluctuación en relación a las variables ambientales;
- comparación estacional de los parámetros fotosintéticos, la PA y Pmáx vol del nano y
microplancton y del picoplancton en tres ambientes acuáticos de la Reserva Natural
Otamendi, lagunas Grande y El Pescado y la mayor de las lagunas en meandros
abandonados (ROL), contrastados en cuanto a su contenido húmico y cobertura de
macrófitas flotantes.
MATERIALES Y MÉTODOS
Este estudio se realizó en 3 cuerpos de agua del la RNO: Laguna Grande, el mayor de los
meandros abandonados (ROL) y en la laguna El Pescado. En la Fig. 4 de la sección Área
de estudio (página 18) se señala la ubicación de los puntos de muestreo en cada cuerpo de
agua. En la Laguna Grande la estación de muestreo se estableció en un zona
representativa de la transición entre la zona litoral y pelágica; en El Pescado, las muestras
fueron obtenidas del centro de la laguna y en el ROL las muestras se tomaron en el centro
del meandro. En todas las oportunidades, se obtuvieron muestras integradas de la
profundidad que nos permitía la columna de agua sin remoción del fondo. Las muestras
en El Pescado fueron tomadas por personal de la RNO, dada la imposibilidad de acceder
hasta el lugar por nuestros propios medios.
La toma de muestras en los 3 cuerpos de agua se realizó con el siguiente esquema
temporal: en la Laguna Grande la frecuencia fue mensual, desde febrero del 2006 hasta
marzo del 2007; en la laguna El Pescado se realizó un muestreo en agosto de 2006 y otro
en enero de 2007. En el ROL se efectuaron seis muestreos con frecuencia
aproximadamente bimensual: marzo, mayo, julio, agosto y noviembre de 2006 y enero de
2007. La diferencia en la frecuencia de toma de muestras en los tres cuerpos de agua
radica en que nos interesaba el seguimiento minucioso de los patrones de productividad
en la Laguna Grande ya que es el mayor cuerpo de agua de la RNO, con un importante

158
aporte a la productividad microbiana del humedal. Con el objeto de comparar con otro
tipo de cuerpo de agua de características contrastantes, realizamos estimaciones en el
ROL más espaciadas en el tiempo. Por último, en la laguna El Pescado, con características
similares a la Laguna Grande (por su conexión con ésta a través del arroyo El Pescado) y
de difícil acceso, con el objeto de extender la comparación en el humedal, se realizaron
dos relevamientos estacionales puesto que consideramos que temperatura es un factor
estructurador importante del fitoplancton en este sistema.
En este estudio se tuvieron en cuenta las siguientes variables ambientales:
profundidad, pH, temperatura del agua, conductividad, oxígeno disuelto, RFA (radiación
fotosintéticamente activa), Kd, g440, N-amonio, N-nitrato + nitrito, N total, P-disuelto, P
total y sólidos en suspensión. Los sensores utilizados así como la toma de muestras y la
metodología seguida para el análisis de estas variables se explicitan en la páginas 72 a 74
del capítulo II. La concentración de nitrógeno inorgánico disuelto (NID) se obtuvo como
la suma de las concentraciones de N-amonio y N-nitrato + nitrito. La profundidad
correspondiente al 1 % de la penetración de la RFA (Z1%) se obtuvo a partir de la
ecuación 4 de la página 81 del capítulo II. La profundidad óptica (⦆) se obtuvo como ⦆ =
Kd z (Kirk 1994). Para la laguna El Pescado, las determinaciones de pH, temperatura y
conductividad se realizaron a partir de la muestra de agua tomada por personal de la RNO
en tanto que los valores de irradiancia subsuperficial (I0), Kd e Imedia fueron estimados
ya que no se pudieron efectuar mediciones in situ. Las estimaciones se realizaron mediante
la suposición que las relaciones entre el Kd, I0 e Imedia se mantienen proporcionalmente
constantes entre la Laguna Grande y El Pescado. Considerando entonces los valores de
estas variables de un muestreo anterior efectuado en enero de 2006 (capítulo I) y
asumiendo constante el factor de proporcionalidad entre esas variables, se estimaron los
nuevos valores correspondientes a El Pescado a partir de conocer los de la Laguna
Grande.
Fitoplancton
Se tomaron muestras de agua de cada sitio que se fijaron con solución de lugol acético a
una concentración final de 1 %. Se realizó el análisis cuantitativo a nivel de clase algal para
el nano y microplancton bajo microscopio invertido con un error del 20 % (Ütermohl
1958, Venrick 1978). Para el reconocimiento de los organismos se empleó la bibliografía
citada en el capítulo I (página 33).

159
Para el análisis de la concentración de clorofila a, se tomaron las muestras de agua
en frascos plásticos que se mantuvieron en frío y oscuridad hasta su posterior
procesamiento en laboratorio. Con el objeto de analizar comparativamente las diferencias
entre las técnicas de análisis se realizaron dos determinaciones de clorofila a, uno para el
fitoplancton > 0,7 µm y el análisis fraccionado de la fracción de tamaño > 3 µm y la
comprendida entre 0,2 y 3 µm. Para la estimación de la fracción > 0,7 µm se siguió la
metodología que figura en la página 31 del capítulo I. La concentración de clorofila a de
las fracciones de tamaño > 3 µm y 0,2-3 µm se estimó a partir de la filtración secuencial
de un volumen conocido de la muestra de agua, en primer lugar a partir de un filtro
Millipore® de policarbonato de 3 µm de tamaño de poro y luego, el agua filtrada se hizo
pasar a través de otro filtro de iguales características de 0,2 µm de poro. Las filtraciones
fueron efectuadas en oscuridad y con baja presión de vacío. La extracción de los
pigmentos se realizó de manera análoga a la ya descrita para el fitoplancton > 0,7 µm.
El trabajo de campo para la realización de las curvas P-I, que incluye la toma de
muestras, la incubación y la medición de la irradiancia incidente se efectuó de igual
manera que la descrita en el capítulo II (páginas 75 a 76) con la salvedad que a las
muestras del ROL se le adicionaron 2 µCi de NaH14CO3 para las incubaciones. Tal como
para la estimación de clorofila, se realizó un análisis para el fitoplancton > 0,7 µm y otro
para las fracciones > 3 µm y 0,2-3 µm. La relacionada con la fracción > 0,7 µm
corresponde a lo señalado en el capítulo II. Para analizar la producción de cada fracción
de tamaño en el laboratorio se procedió al fraccionamiento del fitoplancton. Se siguió la
metodología sugerida en Callieri & Stockner (2002). Las filtraciones se efectuaron en
condiciones de luz tenue y baja presión de vacío. Se filtraron 25 mL de agua proveniente
de cada cuerpo de agua, en primer lugar a través de filtros de policarbonato de 3 µm
(Millipore®) de tamaño de poro. El agua que pasó a través de este filtro se filtró a su vez
por un segundo filtro de policarbonato de 0,2 µm (Millipore®) de poro. Los filtros con el
material retenido se colocaron en viales de centelleo líquido y se procedió de igual manera
a la ya descrita en el capítulo II.
Cabe señalar que se empleó un filtro de 3 µm en lugar de uno de 2 µm para
asegurarnos una separación más eficiente en fracciones de tamaño ya que los filtros de 2
µm retienen mayor cantidad de picoplancton que los de 3 µm (Izaguirre com. pers.). Es
por este motivo que en este capítulo nos referiremos indistintamente a la fracción > 3 µm
como tal y como fracción nano y microplanctónica; mientras que a la comprendida entre

160
0,2 y 3 µm la llamaremos de esa manera o picofitoplancton, de manera análoga a Vörös et
al. (1991). La metodología para la obtención de las tasas de asimilación, los parámetros
fotosintéticos, Pmáx vol, tasas diarias y anuales de producción primaria integradas en la
columna de agua fue la misma que se detalló oportunamente en el capítulo II.
Se realizaron correlaciones no paramétricas (Spearman) entre todas las variables
estudiadas tanto para la Laguna Grande como para el ROL. En el caso de la laguna, al
contar con un mayor número de observaciones (N = 12), se efectuaron regresiones
múltiples entre PA y Pmáx vol para las fracciones de tamaño > 3 µm y picofitoplancton y
las variables ambientales. Para este análisis se utilizó el método de selección de variables
de a pasos sucesivos (forward stepwise). Se tuvieron en cuenta como variables
independientes: temperatura, pH, NID, P disuelto y la Imedia. La variable dependiente
fue trasformada mediante la aplicación de Log10 para cumplir el supuesto de normalidad
cuando correspondiese. También se efectuaron comparaciones entre los valores de
producción y clorofila a de las distintas fracciones de tamaño y para una misma fracción
de tamaño entre cuerpos de agua a través del test no paramétrico de Mann-Whitney (M-
W) (Zar 1996).
RESULTADOS
Laguna Grande
La profundidad de la Laguna Grande fue en general baja durante el período de estudio,
registrándose sólo en cuatro oportunidades valores por encima de los 50 cm, lo que para
este humedal podría considerarse como períodos de aguas altas. La altura hidrométrica
estuvo asociada principalmente con el nivel de precipitaciones del mes anterior al
muestreo. En las Tablas 2 y 3 y Figs. 6-13 del capítulo II de esta tesis (páginas 89 a 100) se
encuentran los valores y figuras de los parámetros físicos y químicos así como la
concentración de clorofila a de la fracción > 0,7 µm y densidad del nano y microplancton
de la Laguna Grande y que corresponden al mismo período de muestreo. La
concentración de O2 disuelto mostró valores más altos en los meses más fríos, a
excepción de los meses de primavera y verano, cuando se registraron floraciones de
cianobacterias filamentosas. La Imedia osciló alrededor de 400 µmol fotones m-2 s-1
durante los años 2006 y principio de 2007. El NID estuvo disponible en mayor medida en
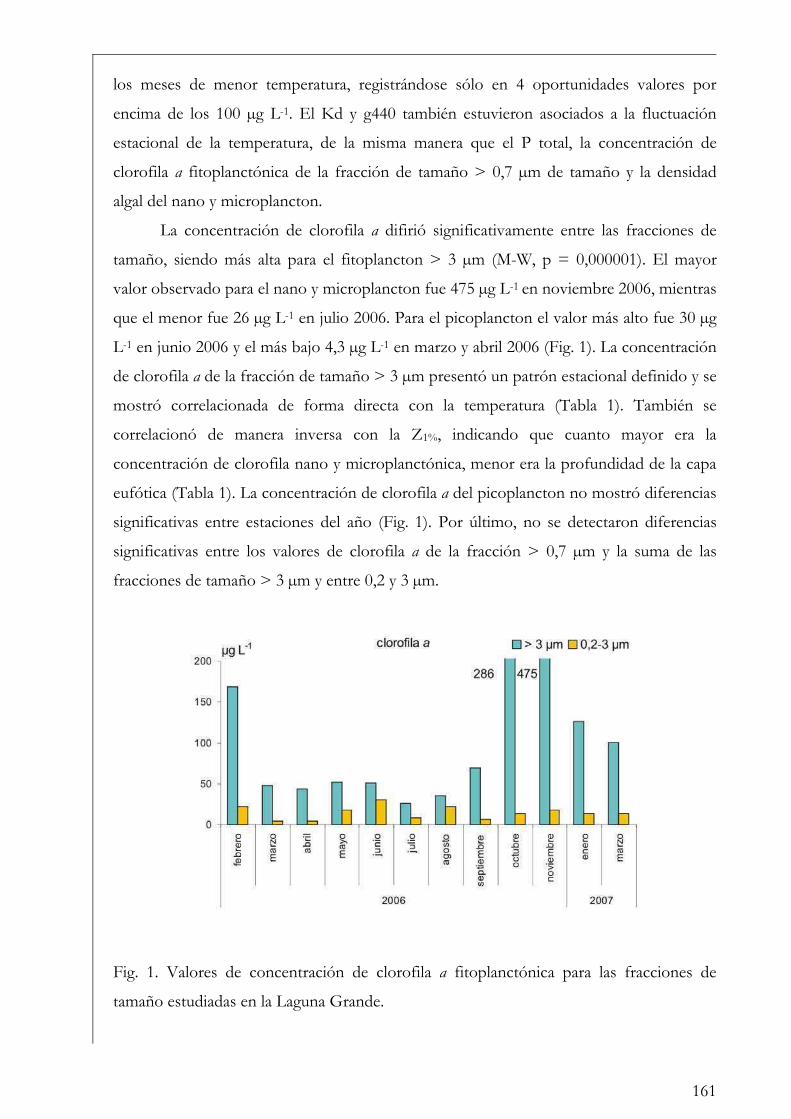
161
los meses de menor temperatura, registrándose sólo en 4 oportunidades valores por
encima de los 100 µg L-1. El Kd y g440 también estuvieron asociados a la fluctuación
estacional de la temperatura, de la misma manera que el P total, la concentración de
clorofila a fitoplanctónica de la fracción de tamaño > 0,7 µm de tamaño y la densidad
algal del nano y microplancton.
La concentración de clorofila a difirió significativamente entre las fracciones de
tamaño, siendo más alta para el fitoplancton > 3 µm (M-W, p = 0,000001). El mayor
valor observado para el nano y microplancton fue 475 µg L-1 en noviembre 2006, mientras
que el menor fue 26 µg L-1 en julio 2006. Para el picoplancton el valor más alto fue 30 µg
L-1 en junio 2006 y el más bajo 4,3 µg L-1 en marzo y abril 2006 (Fig. 1). La concentración
de clorofila a de la fracción de tamaño > 3 µm presentó un patrón estacional definido y se
mostró correlacionada de forma directa con la temperatura (Tabla 1). También se
correlacionó de manera inversa con la Z1%, indicando que cuanto mayor era la
concentración de clorofila nano y microplanctónica, menor era la profundidad de la capa
eufótica (Tabla 1). La concentración de clorofila a del picoplancton no mostró diferencias
significativas entre estaciones del año (Fig. 1). Por último, no se detectaron diferencias
significativas entre los valores de clorofila a de la fracción > 0,7 µm y la suma de las
fracciones de tamaño > 3 µm y entre 0,2 y 3 µm.
Fig. 1. Valores de concentración de clorofila a fitoplanctónica para las fracciones de
tamaño estudiadas en la Laguna Grande.
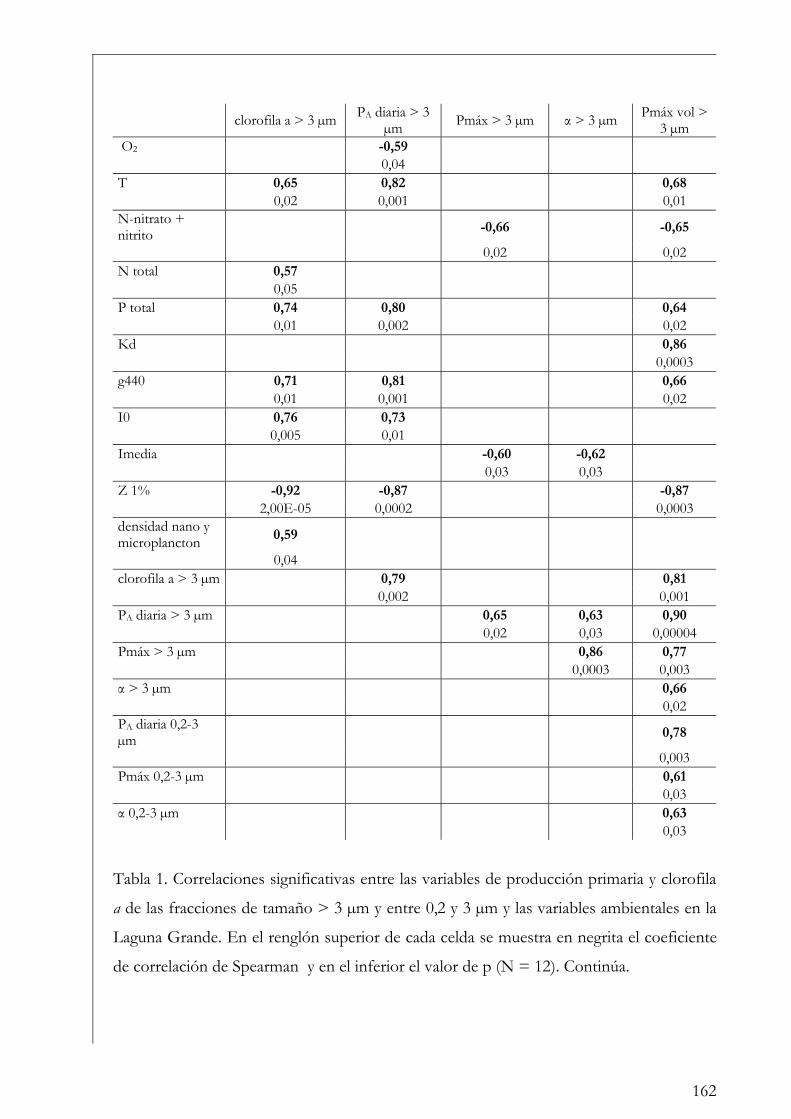
162
clorofila a > 3 µmPA diaria > 3
µm Pmáx > 3 µm プ > 3 µm
Pmáx vol > 3 µm
O2 -0,59 0,04 T 0,65 0,82 0,68 0,02 0,001 0,01 N-nitrato + nitrito
-0,66 -0,65
0,02 0,02 N total 0,57 0,05 P total 0,74 0,80 0,64 0,01 0,002 0,02 Kd 0,86 0,0003 g440 0,71 0,81 0,66 0,01 0,001 0,02 I0 0,76 0,73 0,005 0,01 Imedia -0,60 -0,62 0,03 0,03 Z 1% -0,92 -0,87 -0,87 2,00E-05 0,0002 0,0003 densidad nano y microplancton
0,59
0,04 clorofila a > 3 µm 0,79 0,81 0,002 0,001 PA diaria > 3 µm 0,65 0,63 0,90 0,02 0,03 0,00004 Pmáx > 3 µm 0,86 0,77 0,0003 0,003 プ > 3 µm 0,66 0,02 PA diaria 0,2-3 µm
0,78
0,003 Pmáx 0,2-3 µm 0,61 0,03 プ 0,2-3 µm 0,63 0,03
Tabla 1. Correlaciones significativas entre las variables de producción primaria y clorofila
a de las fracciones de tamaño > 3 µm y entre 0,2 y 3 µm y las variables ambientales en la
Laguna Grande. En el renglón superior de cada celda se muestra en negrita el coeficiente
de correlación de Spearman y en el inferior el valor de p (N = 12). Continúa.
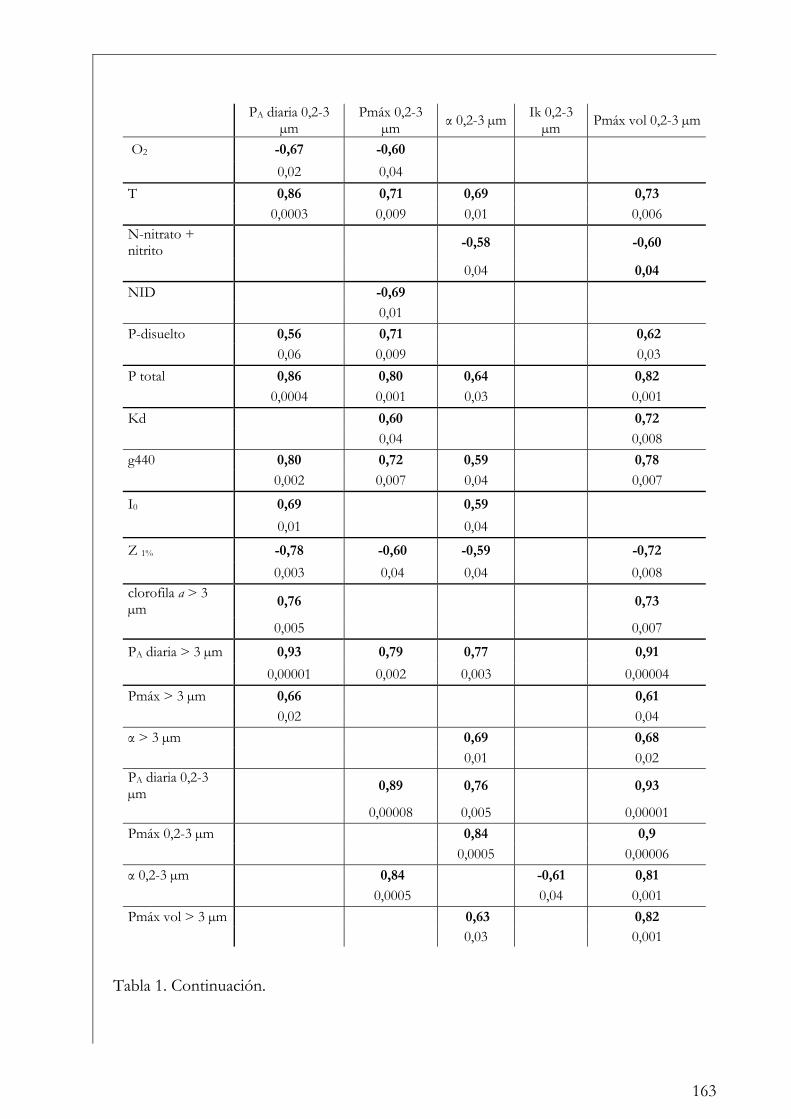
163
PA diaria 0,2-3
µm Pmáx 0,2-3
µm プ 0,2-3 µm
Ik 0,2-3 µm
Pmáx vol 0,2-3 µm
O2 -0,67 -0,60
0,02 0,04
T 0,86 0,71 0,69 0,73
0,0003 0,009 0,01 0,006 N-nitrato + nitrito
-0,58 -0,60
0,04
0,04
NID -0,69
0,01
P-disuelto 0,56 0,71 0,62
0,06 0,009 0,03
P total 0,86 0,80 0,64 0,82
0,0004 0,001 0,03 0,001
Kd 0,60 0,72
0,04 0,008
g440 0,80 0,72 0,59 0,78
0,002 0,007 0,04 0,007
I0 0,69 0,59
0,01 0,04
Z 1% -0,78 -0,60 -0,59 -0,72
0,003 0,04 0,04 0,008 clorofila a > 3 µm
0,76 0,73
0,005 0,007
PA diaria > 3 µm 0,93 0,79 0,77 0,91
0,00001 0,002 0,003 0,00004
Pmáx > 3 µm 0,66 0,61
0,02 0,04
プ > 3 µm 0,69 0,68
0,01 0,02 PA diaria 0,2-3 µm 0,89 0,76 0,93
0,00008 0,005 0,00001
Pmáx 0,2-3 µm 0,84 0,9
0,0005 0,00006
プ 0,2-3 µm 0,84 -0,61 0,81
0,0005 0,04 0,001
Pmáx vol > 3 µm 0,63 0,82
0,03 0,001
Tabla 1. Continuación.

164
El número de asimilación (Pmáx) resultó mayor para los organismos de menor
tamaño (M-W, p = 0,01), con la excepción de los meses de junio y agosto de 2006 (Fig. 2
a). Para la fracción fitoplanctónica > 3 µm el valor de Pmáx más alto fue 11 µg C (µg Chl
a h)-1 en marzo de 2007 y el más bajo de 0,6 µg C (µg Chl a h)-1 en septiembre 2006. Si
bien puede advertirse que los valores más bajos de Pmáx para el nano y microplancton
estuvieron concentrados en los meses de invierno, no se observó un patrón estacional
conspicuo y no se observaron diferencias significativas entre estaciones del año o
correlación con la temperatura. Para el picoplancton autotrófico, el valor máximo de
Pmáx fue de 94 µg C (µg Chl a h)-1 y se detectó en marzo de 2006; mientras que el más
bajo fue de 0,54 µg C (µg Chl a h)-1 en agosto del mismo año (Fig. 2 a). Para esta fracción
de tamaño se observó un patrón estacional claro, detectándose una correlación directa
con la temperatura (Tabla 1).
La eficiencia fotosintética (プ) siguió el mismo patrón temporal que el número de
asimilación y los valores correspondientes a la fracción de tamaño comprendida entre 0,2
y 3 µm fueron significativamente más elevados que los de la fracción > 3 µm (M-W, p =
0,01) (Fig. 2 b). El valor más alto para el fitoplancton > 3 µm fue de 0,04 µg C m2 s (µg
Chl a h µmol fotones)-1 en octubre de 2006 y el más bajo de 0,002 µg C m2 s (µg Chl a h
µmol fotones)-1 en agosto del mismo año, sin observarse ningún patrón estacional
definido. Los valores máximos y mínimos del parámetro プ correspondientes a la fracción
de tamaño comprendida entre 0,2 y 3 µm coincidieron cronológicamente con los del
fitoplancton > 3 µm. El máximo fue de 2,7 µg C m2 s (µg Chl a h µmol fotones)-1 y el
mínimo fue 0,002 µg C m2 s (µg Chl a h µmol fotones)-1. El patrón de fluctuación
estacional para la fracción más pequeña fue análogo al observado para el número de
asimilación y también se observó correlación positiva con la temperatura (Tabla 1).
Los valores de producción máxima por unidad de volumen (Pmáx vol) para la
fracción > 3 µm variaron entre 1,3 mg C L-1 h-1 en octubre 2006 y 0,02 mg C L-1 h-1 en
julio 2006. Para el picofitoplancton, oscilaron entre 0,94 mg C L-1 h-1 en febrero 2006 y
0,01 mg C L-1 h-1 en agosto 2006 (Fig. 3). No se hallaron diferencias significativas en los
valores de Pmáx vol entre fracciones de tamaño pero en los dos casos se observó un
patrón estacional marcado, mostrando ambas fracciones de tamaño una correlación
positiva con la temperatura, Kd, g440, P total; e inversa con la concentración de N-nitrato
+ nitrito y Z1% (Tabla 1). Las regresiones múltiples efectuadas para el Pmáx vol de cada
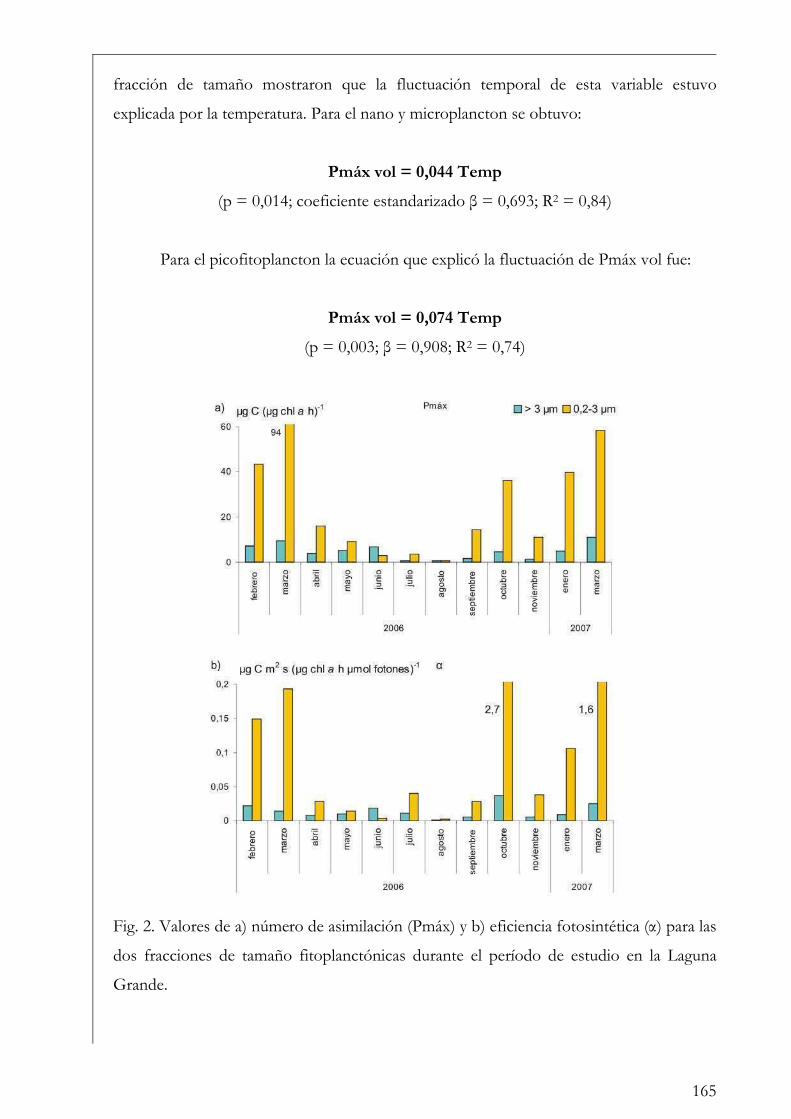
165
fracción de tamaño mostraron que la fluctuación temporal de esta variable estuvo
explicada por la temperatura. Para el nano y microplancton se obtuvo:
Pmáx vol = 0,044 Temp
(p = 0,014; coeficiente estandarizado ベ = 0,693; R2 = 0,84)
Para el picofitoplancton la ecuación que explicó la fluctuación de Pmáx vol fue:
Pmáx vol = 0,074 Temp
(p = 0,003; ベ = 0,908; R2 = 0,74)
Fig. 2. Valores de a) número de asimilación (Pmáx) y b) eficiencia fotosintética (プ) para las
dos fracciones de tamaño fitoplanctónicas durante el período de estudio en la Laguna
Grande.
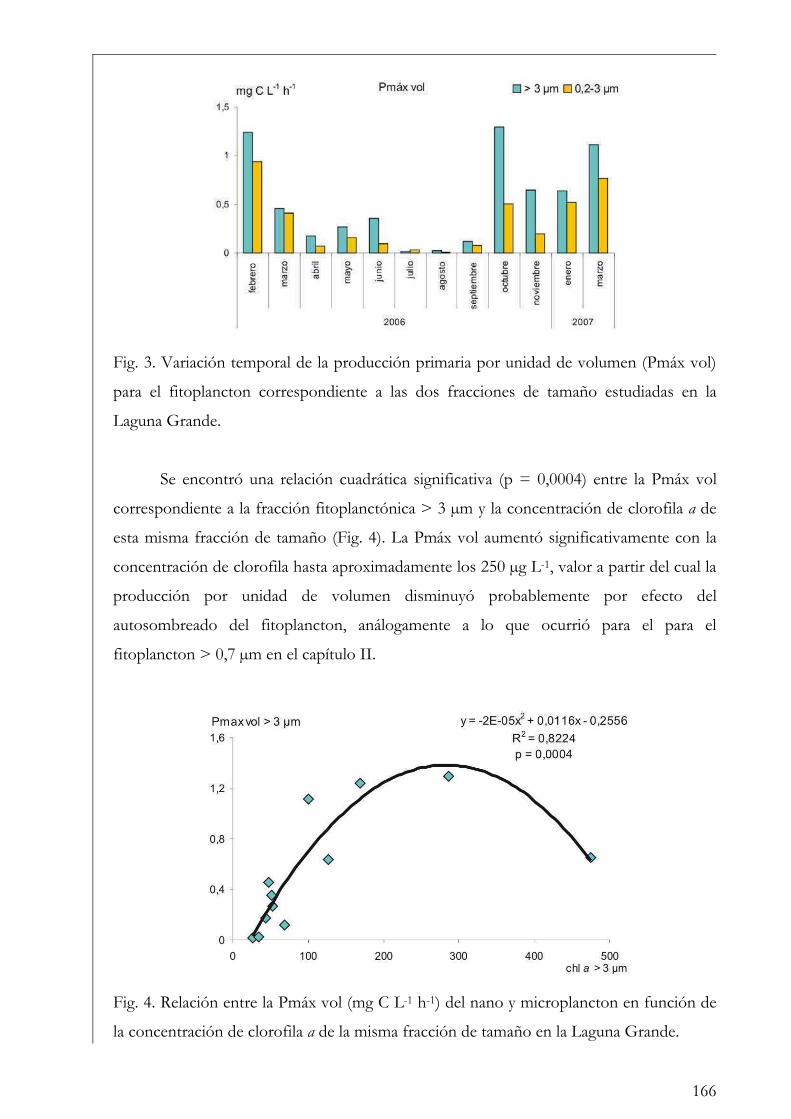
166
Fig. 3. Variación temporal de la producción primaria por unidad de volumen (Pmáx vol)
para el fitoplancton correspondiente a las dos fracciones de tamaño estudiadas en la
Laguna Grande.
Se encontró una relación cuadrática significativa (p = 0,0004) entre la Pmáx vol
correspondiente a la fracción fitoplanctónica > 3 µm y la concentración de clorofila a de
esta misma fracción de tamaño (Fig. 4). La Pmáx vol aumentó significativamente con la
concentración de clorofila hasta aproximadamente los 250 µg L-1, valor a partir del cual la
producción por unidad de volumen disminuyó probablemente por efecto del
autosombreado del fitoplancton, análogamente a lo que ocurrió para el para el
fitoplancton > 0,7 µm en el capítulo II.
Fig. 4. Relación entre la Pmáx vol (mg C L-1 h-1) del nano y microplancton en función de
la concentración de clorofila a de la misma fracción de tamaño en la Laguna Grande.
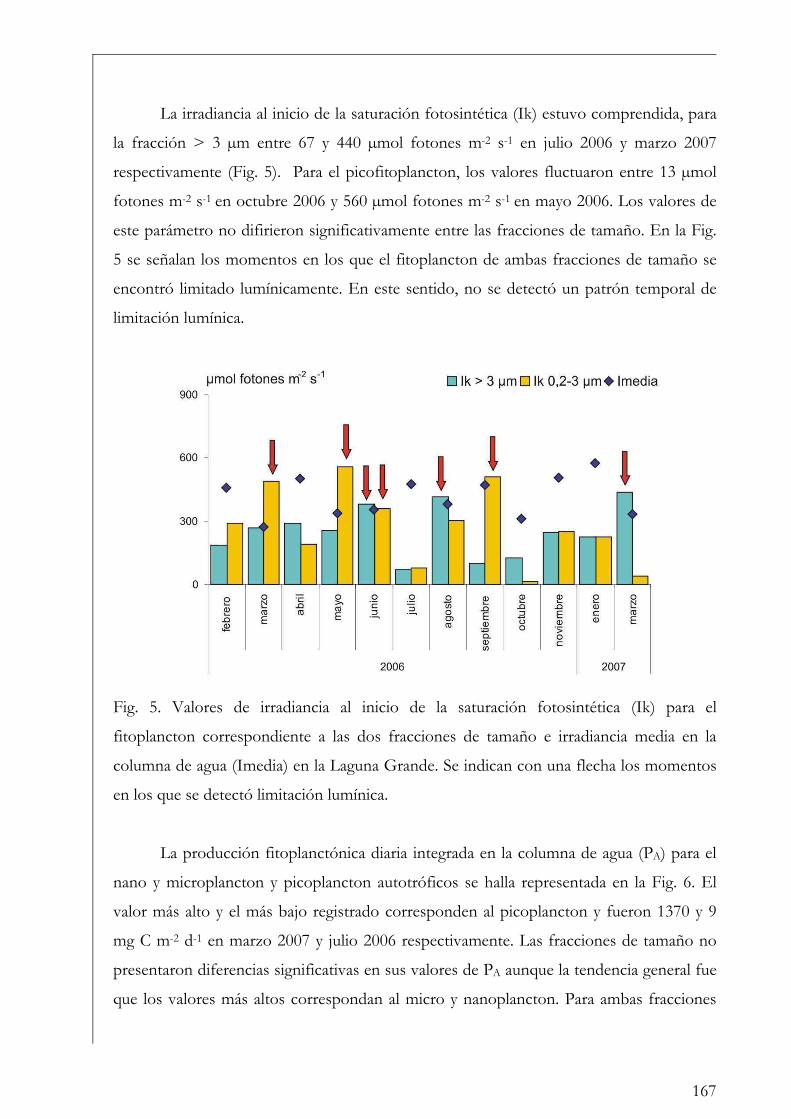
167
La irradiancia al inicio de la saturación fotosintética (Ik) estuvo comprendida, para
la fracción > 3 µm entre 67 y 440 µmol fotones m-2 s-1 en julio 2006 y marzo 2007
respectivamente (Fig. 5). Para el picofitoplancton, los valores fluctuaron entre 13 µmol
fotones m-2 s-1 en octubre 2006 y 560 µmol fotones m-2 s-1 en mayo 2006. Los valores de
este parámetro no difirieron significativamente entre las fracciones de tamaño. En la Fig.
5 se señalan los momentos en los que el fitoplancton de ambas fracciones de tamaño se
encontró limitado lumínicamente. En este sentido, no se detectó un patrón temporal de
limitación lumínica.
Fig. 5. Valores de irradiancia al inicio de la saturación fotosintética (Ik) para el
fitoplancton correspondiente a las dos fracciones de tamaño e irradiancia media en la
columna de agua (Imedia) en la Laguna Grande. Se indican con una flecha los momentos
en los que se detectó limitación lumínica.
La producción fitoplanctónica diaria integrada en la columna de agua (PA) para el
nano y microplancton y picoplancton autotróficos se halla representada en la Fig. 6. El
valor más alto y el más bajo registrado corresponden al picoplancton y fueron 1370 y 9
mg C m-2 d-1 en marzo 2007 y julio 2006 respectivamente. Las fracciones de tamaño no
presentaron diferencias significativas en sus valores de PA aunque la tendencia general fue
que los valores más altos correspondan al micro y nanoplancton. Para ambas fracciones
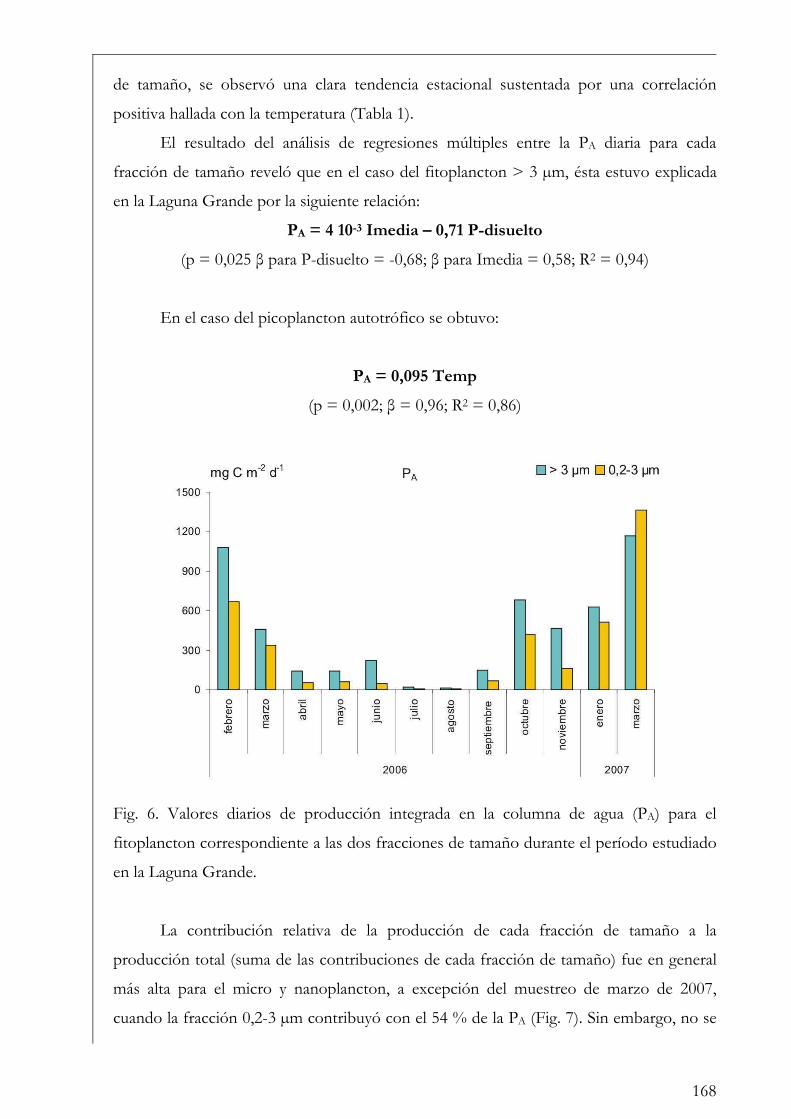
168
de tamaño, se observó una clara tendencia estacional sustentada por una correlación
positiva hallada con la temperatura (Tabla 1).
El resultado del análisis de regresiones múltiples entre la PA diaria para cada
fracción de tamaño reveló que en el caso del fitoplancton > 3 µm, ésta estuvo explicada
en la Laguna Grande por la siguiente relación:
PA = 4 10-3 Imedia – 0,71 P-disuelto
(p = 0,025 ベ para P-disuelto = -0,68; ベ para Imedia = 0,58; R2 = 0,94)
En el caso del picoplancton autotrófico se obtuvo:
PA = 0,095 Temp
(p = 0,002; ベ = 0,96; R2 = 0,86)
Fig. 6. Valores diarios de producción integrada en la columna de agua (PA) para el
fitoplancton correspondiente a las dos fracciones de tamaño durante el período estudiado
en la Laguna Grande.
La contribución relativa de la producción de cada fracción de tamaño a la
producción total (suma de las contribuciones de cada fracción de tamaño) fue en general
más alta para el micro y nanoplancton, a excepción del muestreo de marzo de 2007,
cuando la fracción 0,2-3 µm contribuyó con el 54 % de la PA (Fig. 7). Sin embargo, no se
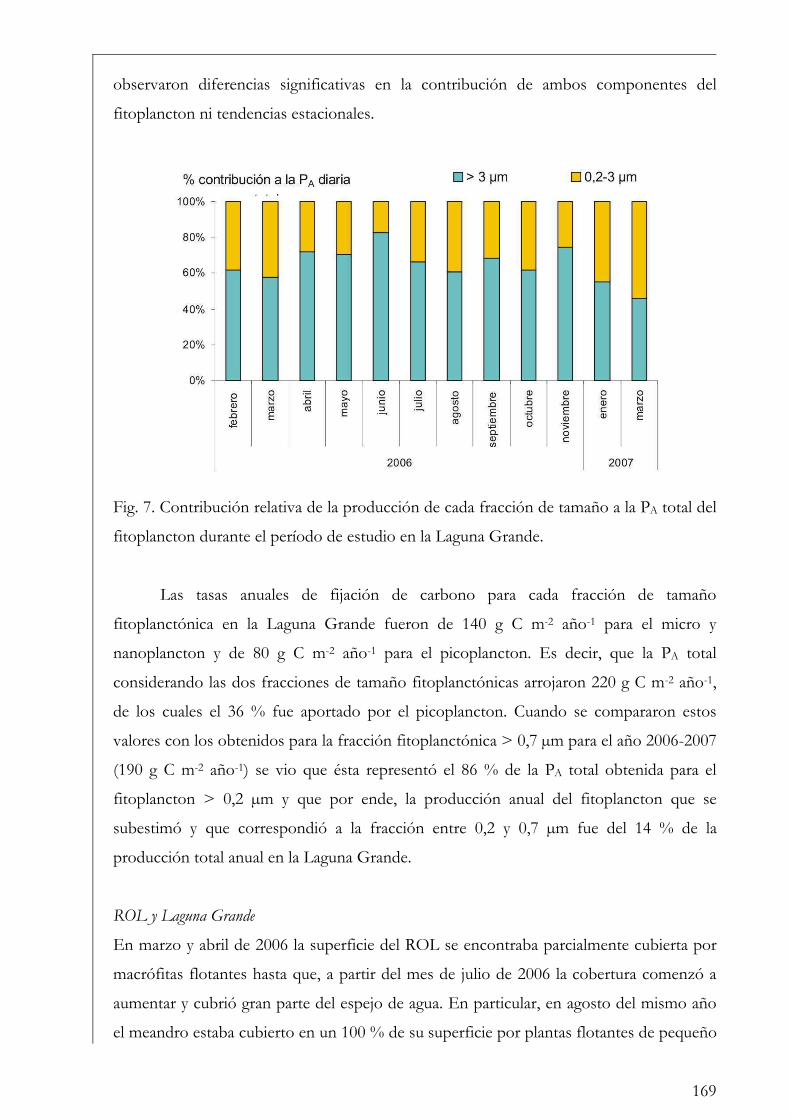
169
observaron diferencias significativas en la contribución de ambos componentes del
fitoplancton ni tendencias estacionales.
Fig. 7. Contribución relativa de la producción de cada fracción de tamaño a la PA total del
fitoplancton durante el período de estudio en la Laguna Grande.
Las tasas anuales de fijación de carbono para cada fracción de tamaño
fitoplanctónica en la Laguna Grande fueron de 140 g C m-2 año-1 para el micro y
nanoplancton y de 80 g C m-2 año-1 para el picoplancton. Es decir, que la PA total
considerando las dos fracciones de tamaño fitoplanctónicas arrojaron 220 g C m-2 año-1,
de los cuales el 36 % fue aportado por el picoplancton. Cuando se compararon estos
valores con los obtenidos para la fracción fitoplanctónica > 0,7 µm para el año 2006-2007
(190 g C m-2 año-1) se vio que ésta representó el 86 % de la PA total obtenida para el
fitoplancton > 0,2 µm y que por ende, la producción anual del fitoplancton que se
subestimó y que correspondió a la fracción entre 0,2 y 0,7 µm fue del 14 % de la
producción total anual en la Laguna Grande.
ROL y Laguna Grande
En marzo y abril de 2006 la superficie del ROL se encontraba parcialmente cubierta por
macrófitas flotantes hasta que, a partir del mes de julio de 2006 la cobertura comenzó a
aumentar y cubrió gran parte del espejo de agua. En particular, en agosto del mismo año
el meandro estaba cubierto en un 100 % de su superficie por plantas flotantes de pequeño

170
porte. Esta cobertura se mantuvo hasta el final del estudio observándose el ROL como se
aprecia en la Fig. 8. En la Tabla 3 se muestra el promedio y los rangos de variación de las
variables ambientales registradas para el ROL y la Laguna Grande durante el período de
estudio.
Fig. 8. Meandro abandonado (ROL) cubierto en un 100 % de su superficie por macrófitas
flotantes de pequeño porte ubicadas entre manchones de vegetación palustre.
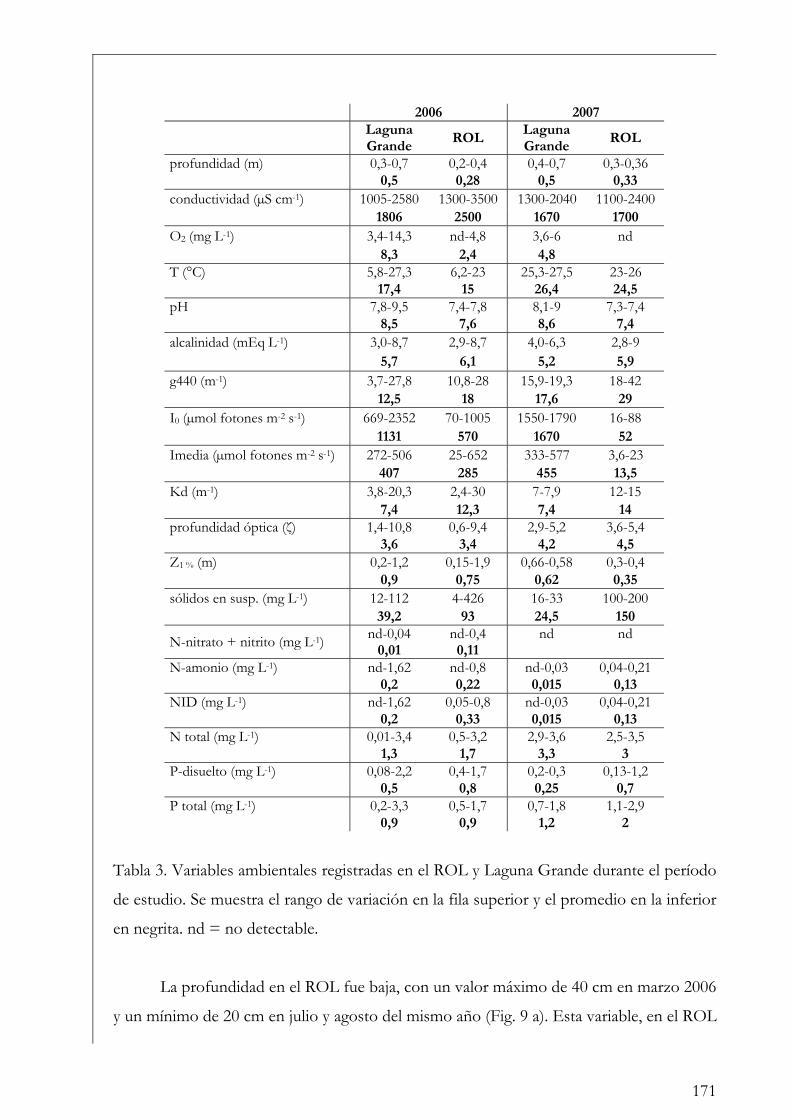
171
2006 2007
Laguna Grande
ROL Laguna Grande
ROL
profundidad (m) 0,3-0,7 0,2-0,4 0,4-0,7 0,3-0,36 0,5 0,28 0,5 0,33 conductividad (µS cm-1) 1005-2580 1300-3500 1300-2040 1100-2400 1806 2500 1670 1700 O2 (mg L-1) 3,4-14,3 nd-4,8 3,6-6 nd 8,3 2,4 4,8 T (°C) 5,8-27,3 6,2-23 25,3-27,5 23-26 17,4 15 26,4 24,5 pH 7,8-9,5 7,4-7,8 8,1-9 7,3-7,4 8,5 7,6 8,6 7,4 alcalinidad (mEq L-1) 3,0-8,7 2,9-8,7 4,0-6,3 2,8-9 5,7 6,1 5,2 5,9 g440 (m-1) 3,7-27,8 10,8-28 15,9-19,3 18-42 12,5 18 17,6 29 I0 (µmol fotones m-2 s-1) 669-2352 70-1005 1550-1790 16-88 1131 570 1670 52 Imedia (µmol fotones m-2 s-1) 272-506 25-652 333-577 3,6-23 407 285 455 13,5 Kd (m-1) 3,8-20,3 2,4-30 7-7,9 12-15 7,4 12,3 7,4 14 profundidad óptica (⦆) 1,4-10,8 0,6-9,4 2,9-5,2 3,6-5,4 3,6 3,4 4,2 4,5 Z1 % (m) 0,2-1,2 0,15-1,9 0,66-0,58 0,3-0,4 0,9 0,75 0,62 0,35 sólidos en susp. (mg L-1) 12-112 4-426 16-33 100-200 39,2 93 24,5 150
N-nitrato + nitrito (mg L-1) nd-0,04
0,01 nd-0,4 0,11
nd
nd
N-amonio (mg L-1) nd-1,62 nd-0,8 nd-0,03 0,04-0,21 0,2 0,22 0,015 0,13 NID (mg L-1) nd-1,62 0,05-0,8 nd-0,03 0,04-0,21 0,2 0,33 0,015 0,13 N total (mg L-1) 0,01-3,4 0,5-3,2 2,9-3,6 2,5-3,5 1,3 1,7 3,3 3 P-disuelto (mg L-1) 0,08-2,2 0,4-1,7 0,2-0,3 0,13-1,2 0,5 0,8 0,25 0,7 P total (mg L-1) 0,2-3,3 0,5-1,7 0,7-1,8 1,1-2,9 0,9 0,9 1,2 2
Tabla 3. Variables ambientales registradas en el ROL y Laguna Grande durante el período
de estudio. Se muestra el rango de variación en la fila superior y el promedio en la inferior
en negrita. nd = no detectable.
La profundidad en el ROL fue baja, con un valor máximo de 40 cm en marzo 2006
y un mínimo de 20 cm en julio y agosto del mismo año (Fig. 9 a). Esta variable, en el ROL
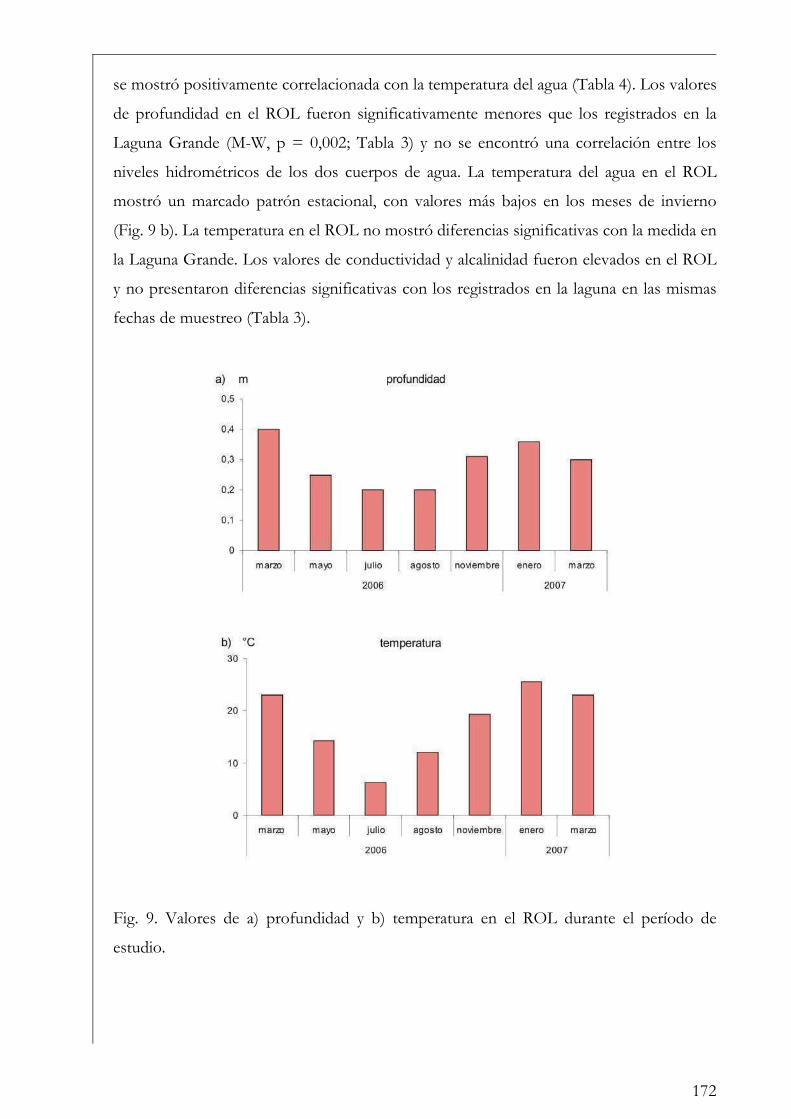
172
se mostró positivamente correlacionada con la temperatura del agua (Tabla 4). Los valores
de profundidad en el ROL fueron significativamente menores que los registrados en la
Laguna Grande (M-W, p = 0,002; Tabla 3) y no se encontró una correlación entre los
niveles hidrométricos de los dos cuerpos de agua. La temperatura del agua en el ROL
mostró un marcado patrón estacional, con valores más bajos en los meses de invierno
(Fig. 9 b). La temperatura en el ROL no mostró diferencias significativas con la medida en
la Laguna Grande. Los valores de conductividad y alcalinidad fueron elevados en el ROL
y no presentaron diferencias significativas con los registrados en la laguna en las mismas
fechas de muestreo (Tabla 3).
Fig. 9. Valores de a) profundidad y b) temperatura en el ROL durante el período de
estudio.
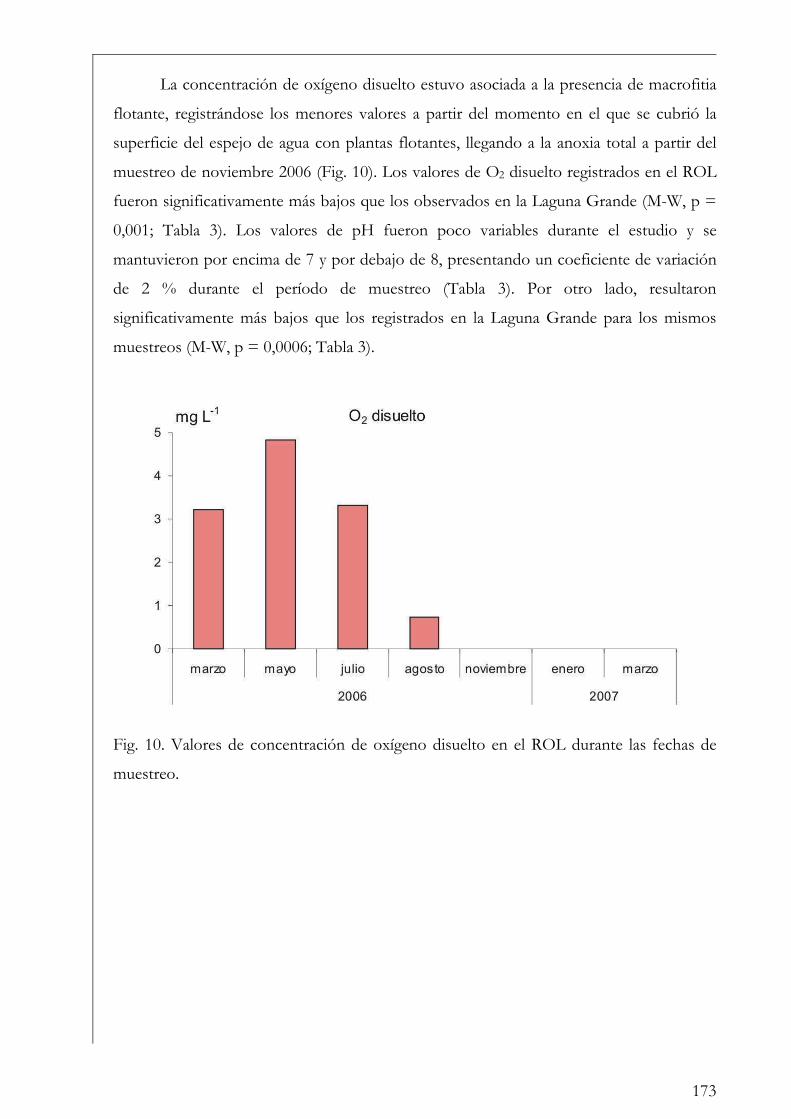
173
La concentración de oxígeno disuelto estuvo asociada a la presencia de macrofitia
flotante, registrándose los menores valores a partir del momento en el que se cubrió la
superficie del espejo de agua con plantas flotantes, llegando a la anoxia total a partir del
muestreo de noviembre 2006 (Fig. 10). Los valores de O2 disuelto registrados en el ROL
fueron significativamente más bajos que los observados en la Laguna Grande (M-W, p =
0,001; Tabla 3). Los valores de pH fueron poco variables durante el estudio y se
mantuvieron por encima de 7 y por debajo de 8, presentando un coeficiente de variación
de 2 % durante el período de muestreo (Tabla 3). Por otro lado, resultaron
significativamente más bajos que los registrados en la Laguna Grande para los mismos
muestreos (M-W, p = 0,0006; Tabla 3).
Fig. 10. Valores de concentración de oxígeno disuelto en el ROL durante las fechas de
muestreo.
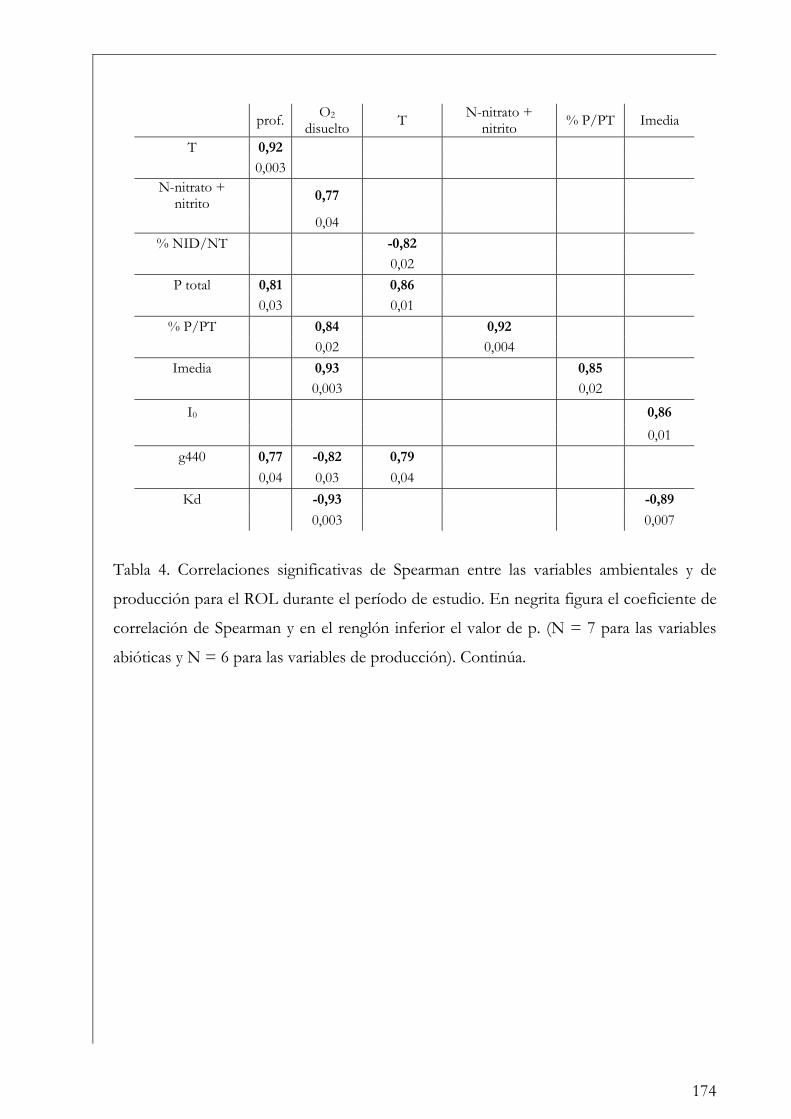
174
prof. O2
disuelto T
N-nitrato + nitrito
% P/PT Imedia
T 0,92
0,003 N-nitrato +
nitrito 0,77
0,04
% NID/NT -0,82
0,02
P total 0,81 0,86 0,03 0,01
% P/PT 0,84 0,92 0,02 0,004
Imedia 0,93 0,85
0,003 0,02
I0 0,86
0,01
g440 0,77 -0,82 0,79
0,04 0,03 0,04
Kd -0,93 -0,89
0,003 0,007
Tabla 4. Correlaciones significativas de Spearman entre las variables ambientales y de
producción para el ROL durante el período de estudio. En negrita figura el coeficiente de
correlación de Spearman y en el renglón inferior el valor de p. (N = 7 para las variables
abióticas y N = 6 para las variables de producción). Continúa.
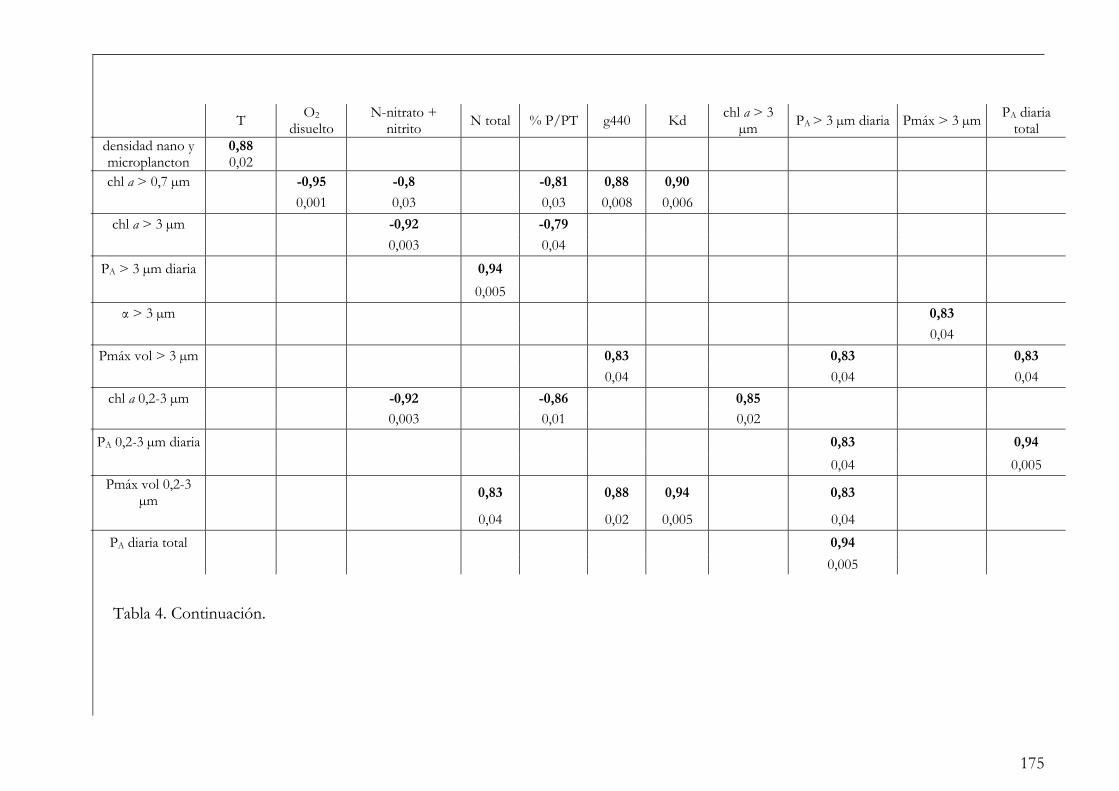
175
T O2
disuelto N-nitrato +
nitrito N total % P/PT g440 Kd
chl a > 3 µm
PA > 3 µm diaria Pmáx > 3 µm PA diaria
total densidad nano y microplancton
0,88 0,02
chl a > 0,7 µm -0,95 -0,8 -0,81 0,88 0,90
0,001 0,03 0,03 0,008 0,006
chl a > 3 µm -0,92 -0,79
0,003 0,04
PA > 3 µm diaria 0,94
0,005
プ > 3 µm 0,83
0,04
Pmáx vol > 3 µm 0,83 0,83 0,83
0,04 0,04 0,04
chl a 0,2-3 µm -0,92 -0,86 0,85 0,003 0,01 0,02
PA 0,2-3 µm diaria 0,83 0,94
0,04 0,005 Pmáx vol 0,2-3
µm 0,83 0,88 0,94 0,83
0,04 0,02 0,005 0,04
PA diaria total 0,94
0,005
Tabla 4. Continuación.

176
La irradiancia incidente subsuperficial (I0) al igual que la irradiancia integrada en la
columna de agua (Imedia) disminuyeron a partir de la presencia permanente de las
macrófitas flotantes en agosto de 2006 (Fig. 11). La Imedia se mostró positivamente
correlacionada con la concentración de oxígeno disuelto y el porcentaje de P-disuelto en
el P total (Tabla 4). Los valores de I0 difirieron de manera significativa cuando se comparó
la Laguna Grande con el ROL (M-W, p = 0,04; Tabla 3), siendo más bajos en este último
sitio. El contenido húmico, estimado con el coeficiente de absorción a 440 nm (g440) y,
por otro lado, el coeficiente de atenuación vertical (Kd) del ROL fueron más altos a partir
de agosto 2006, cuando el meandro se cubrió completamente de macrófitas flotantes (Fig.
12 a y b). Ambas variables mostraron además una correlación negativa con concentración
de oxígeno disuelto (Tabla 4). La misma tendencia al aumento con la mayor cobertura de
macrófitas, se observó en cuanto a la concentración de sólidos en suspensión (Fig. 12 c).
La profundidad óptica fue menor al valor 4,6 en todos los muestreos a excepción de
noviembre 2006 y enero 2007 (Fig. 13). En estas dos oportunidades el 1 % de la
irradiancia incidente en superficie no llegaba al fondo del sitio de muestreo, sino hasta los
15 y 30 cm, siendo la profundidad total de 31 y 36 cm respectivamente en noviembre
2006 y enero 2007. Este hecho probablemente esté relacionado con el mayor contenido
húmico y mayor Kd en estos muestreos, asociados con el mayor desarrollo de
fitoplancton, como indica la correlación positiva entre la concentración de clorofila a de la
fracción fitoplanctónica > 0,7 µm y el Kd y g440 (Tabla 4).
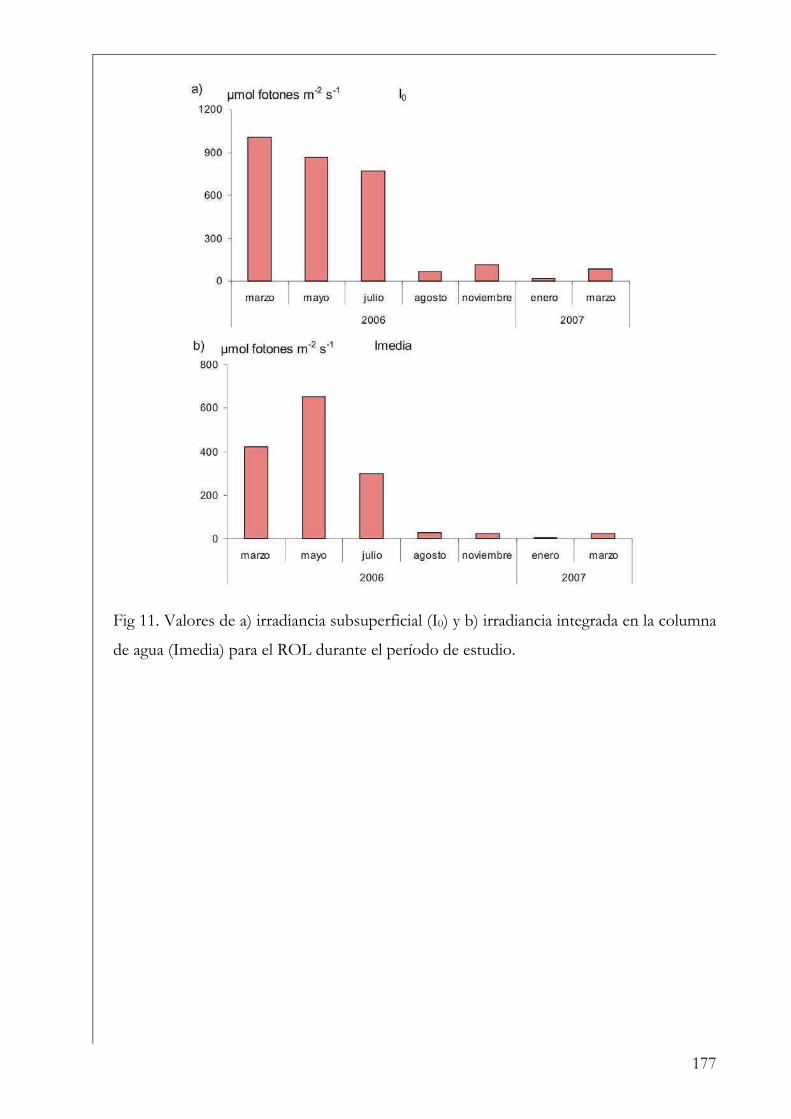
177
Fig 11. Valores de a) irradiancia subsuperficial (I0) y b) irradiancia integrada en la columna
de agua (Imedia) para el ROL durante el período de estudio.
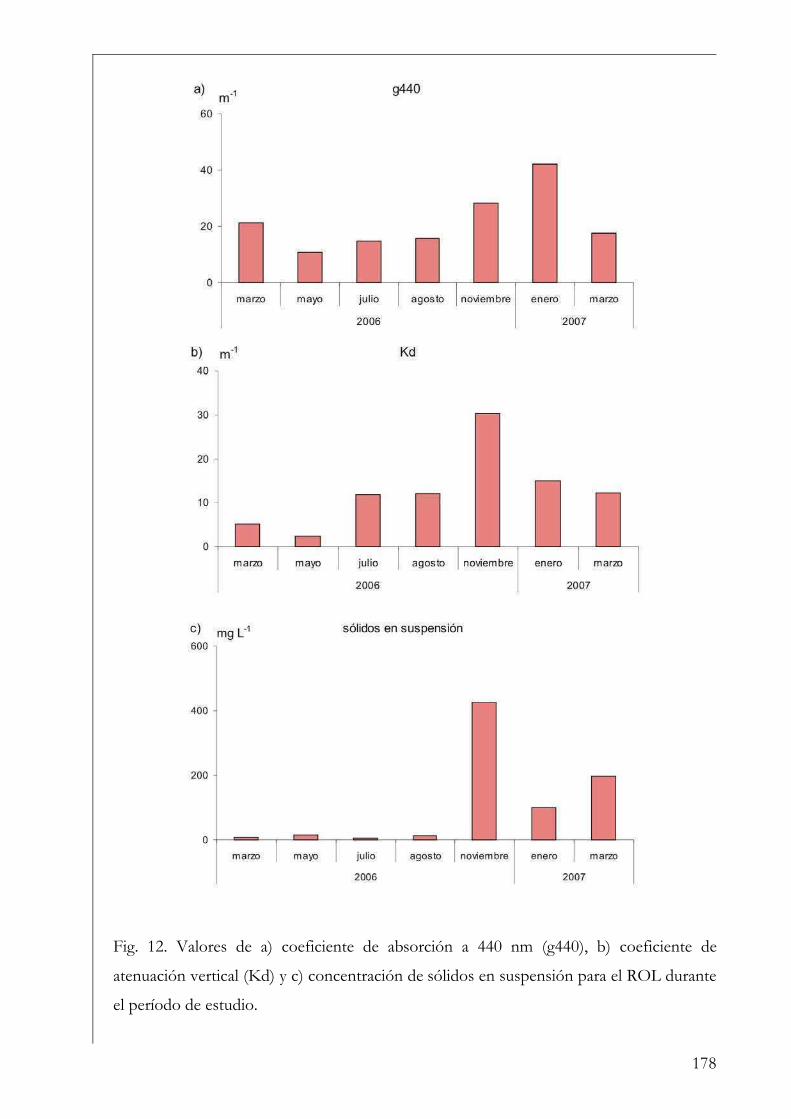
178
Fig. 12. Valores de a) coeficiente de absorción a 440 nm (g440), b) coeficiente de
atenuación vertical (Kd) y c) concentración de sólidos en suspensión para el ROL durante
el período de estudio.
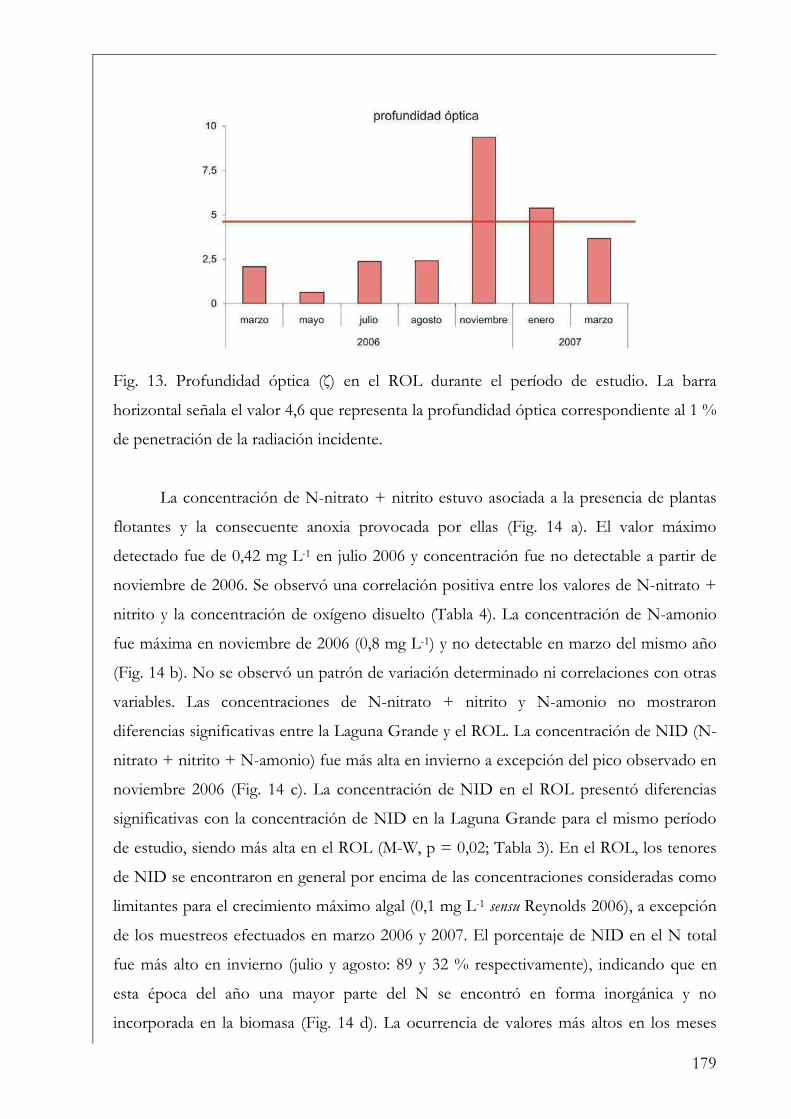
179
Fig. 13. Profundidad óptica (⦆) en el ROL durante el período de estudio. La barra
horizontal señala el valor 4,6 que representa la profundidad óptica correspondiente al 1 %
de penetración de la radiación incidente.
La concentración de N-nitrato + nitrito estuvo asociada a la presencia de plantas
flotantes y la consecuente anoxia provocada por ellas (Fig. 14 a). El valor máximo
detectado fue de 0,42 mg L-1 en julio 2006 y concentración fue no detectable a partir de
noviembre de 2006. Se observó una correlación positiva entre los valores de N-nitrato +
nitrito y la concentración de oxígeno disuelto (Tabla 4). La concentración de N-amonio
fue máxima en noviembre de 2006 (0,8 mg L-1) y no detectable en marzo del mismo año
(Fig. 14 b). No se observó un patrón de variación determinado ni correlaciones con otras
variables. Las concentraciones de N-nitrato + nitrito y N-amonio no mostraron
diferencias significativas entre la Laguna Grande y el ROL. La concentración de NID (N-
nitrato + nitrito + N-amonio) fue más alta en invierno a excepción del pico observado en
noviembre 2006 (Fig. 14 c). La concentración de NID en el ROL presentó diferencias
significativas con la concentración de NID en la Laguna Grande para el mismo período
de estudio, siendo más alta en el ROL (M-W, p = 0,02; Tabla 3). En el ROL, los tenores
de NID se encontraron en general por encima de las concentraciones consideradas como
limitantes para el crecimiento máximo algal (0,1 mg L-1 sensu Reynolds 2006), a excepción
de los muestreos efectuados en marzo 2006 y 2007. El porcentaje de NID en el N total
fue más alto en invierno (julio y agosto: 89 y 32 % respectivamente), indicando que en
esta época del año una mayor parte del N se encontró en forma inorgánica y no
incorporada en la biomasa (Fig. 14 d). La ocurrencia de valores más altos en los meses
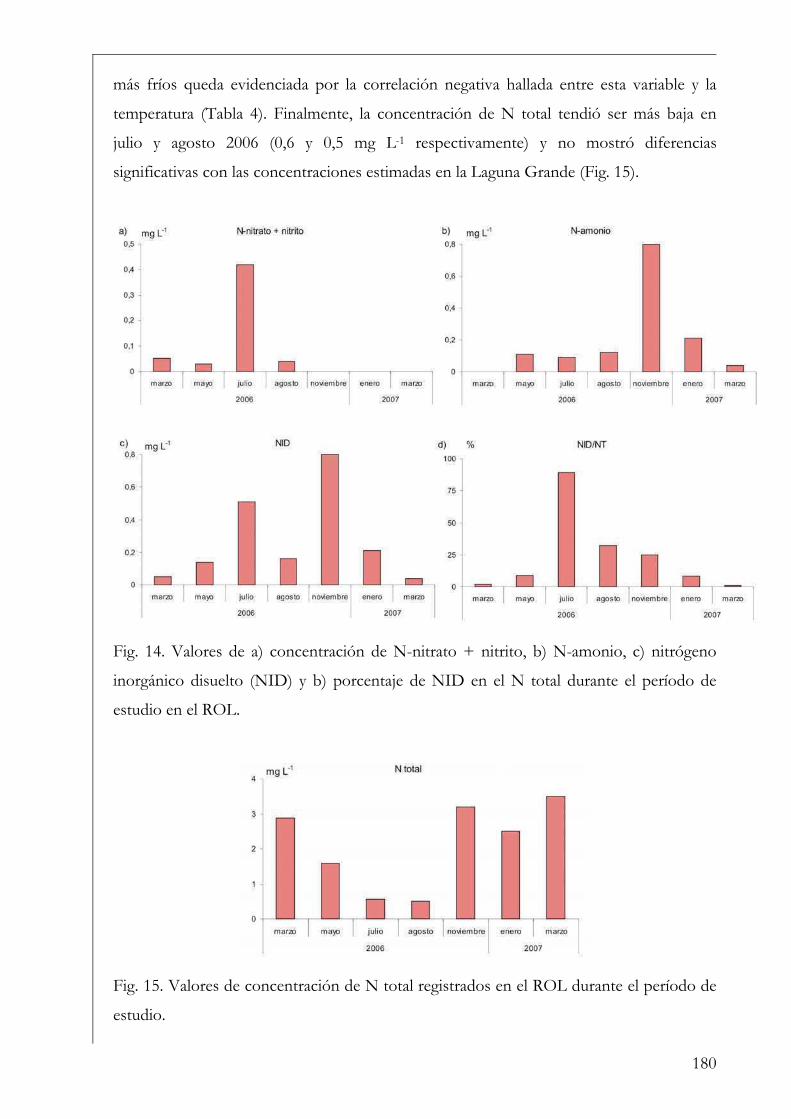
180
más fríos queda evidenciada por la correlación negativa hallada entre esta variable y la
temperatura (Tabla 4). Finalmente, la concentración de N total tendió ser más baja en
julio y agosto 2006 (0,6 y 0,5 mg L-1 respectivamente) y no mostró diferencias
significativas con las concentraciones estimadas en la Laguna Grande (Fig. 15).
Fig. 14. Valores de a) concentración de N-nitrato + nitrito, b) N-amonio, c) nitrógeno
inorgánico disuelto (NID) y b) porcentaje de NID en el N total durante el período de
estudio en el ROL.
Fig. 15. Valores de concentración de N total registrados en el ROL durante el período de
estudio.

181
La concentración de P-disuelto en el ROL osciló entre 0,1 mg L-1 (marzo 2007) y
1,7 mg L-1 (marzo 2006) (Fig. 16 a) y mostró una tendencia a disminuir con la cobertura
de plantas flotantes a excepción de un pico observado en el mes de enero 2007 (1,2 mg L-
1). Los valores registrados en este sitio fueron más altos que los estimados en la Laguna
Grande para el mismo período de estudio (M-W, p = 0,03; Tabla 3). La concentración de
P-disuelto en el ROL se encontró siempre por encima de los valores que considera
Reynolds (1984) limitantes para el máximo crecimiento algal. La concentración de P total
fluctuó entre 0,5 y 2,9 mg L-1 (en julio 2006 y enero 2007 respectivamente) y siguió el
mismo patrón temporal que el P disuelto. El P total se correlacionó positivamente con la
temperatura del agua (Fig. 16 b, Tabla 4). Por otro lado, durante el año 2006, más del 50
% de P se encontraba en la forma disuelta. La relación P-disuelto/P total tendió a
disminuir conforme aumentaba la cobertura de macrófitas flotantes, llegando a ser del 12
% en marzo 2007 (Fig. 16 c).
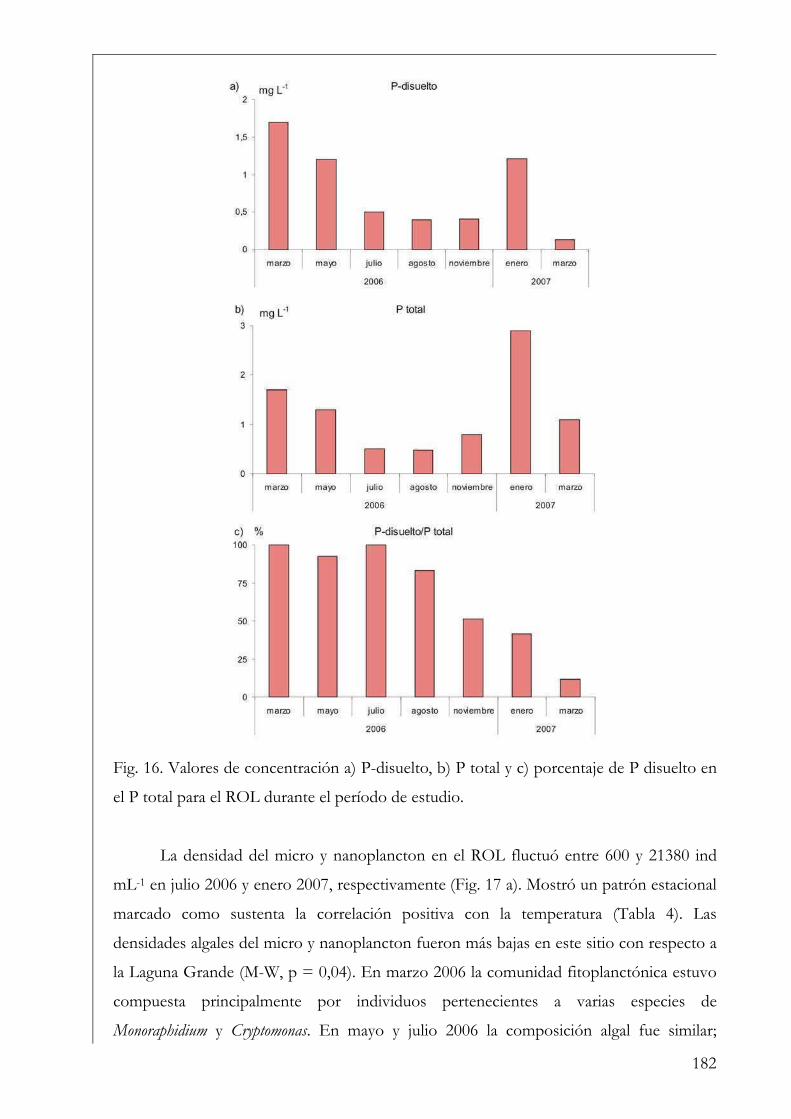
182
Fig. 16. Valores de concentración a) P-disuelto, b) P total y c) porcentaje de P disuelto en
el P total para el ROL durante el período de estudio.
La densidad del micro y nanoplancton en el ROL fluctuó entre 600 y 21380 ind
mL-1 en julio 2006 y enero 2007, respectivamente (Fig. 17 a). Mostró un patrón estacional
marcado como sustenta la correlación positiva con la temperatura (Tabla 4). Las
densidades algales del micro y nanoplancton fueron más bajas en este sitio con respecto a
la Laguna Grande (M-W, p = 0,04). En marzo 2006 la comunidad fitoplanctónica estuvo
compuesta principalmente por individuos pertenecientes a varias especies de
Monoraphidium y Cryptomonas. En mayo y julio 2006 la composición algal fue similar;
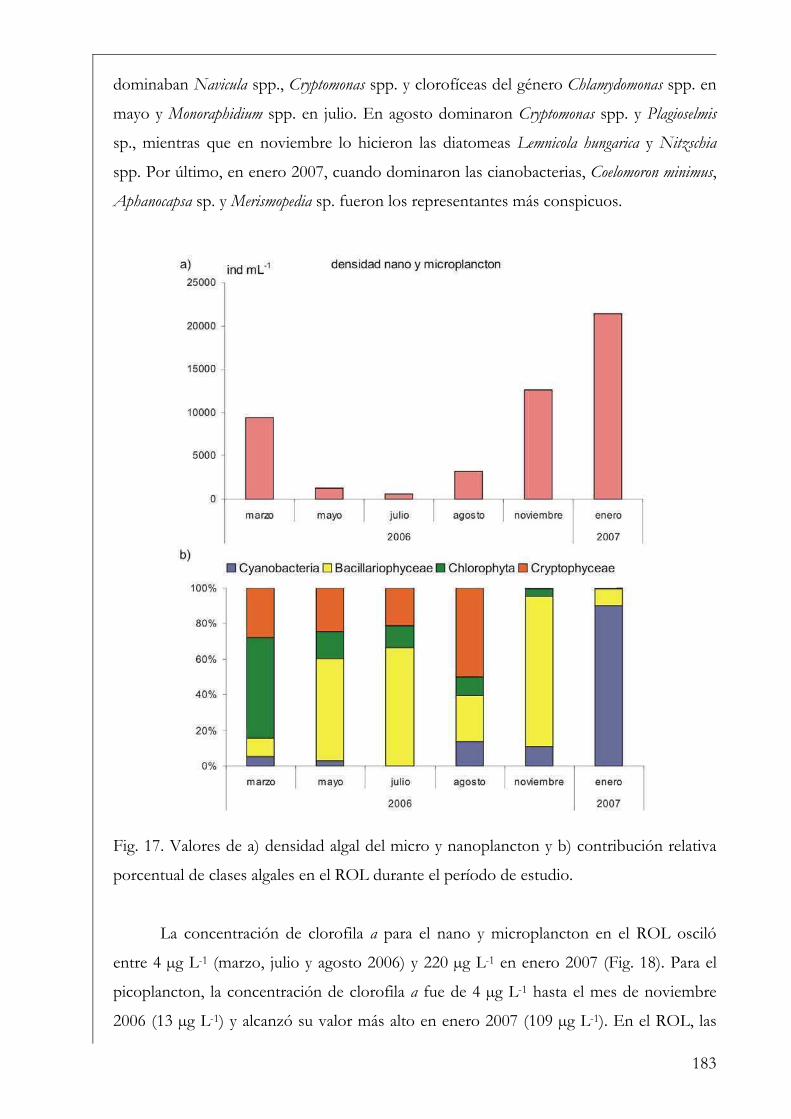
183
dominaban Navicula spp., Cryptomonas spp. y clorofíceas del género Chlamydomonas spp. en
mayo y Monoraphidium spp. en julio. En agosto dominaron Cryptomonas spp. y Plagioselmis
sp., mientras que en noviembre lo hicieron las diatomeas Lemnicola hungarica y Nitzschia
spp. Por último, en enero 2007, cuando dominaron las cianobacterias, Coelomoron minimus,
Aphanocapsa sp. y Merismopedia sp. fueron los representantes más conspicuos.
Fig. 17. Valores de a) densidad algal del micro y nanoplancton y b) contribución relativa
porcentual de clases algales en el ROL durante el período de estudio.
La concentración de clorofila a para el nano y microplancton en el ROL osciló
entre 4 µg L-1 (marzo, julio y agosto 2006) y 220 µg L-1 en enero 2007 (Fig. 18). Para el
picoplancton, la concentración de clorofila a fue de 4 µg L-1 hasta el mes de noviembre
2006 (13 µg L-1) y alcanzó su valor más alto en enero 2007 (109 µg L-1). En el ROL, las
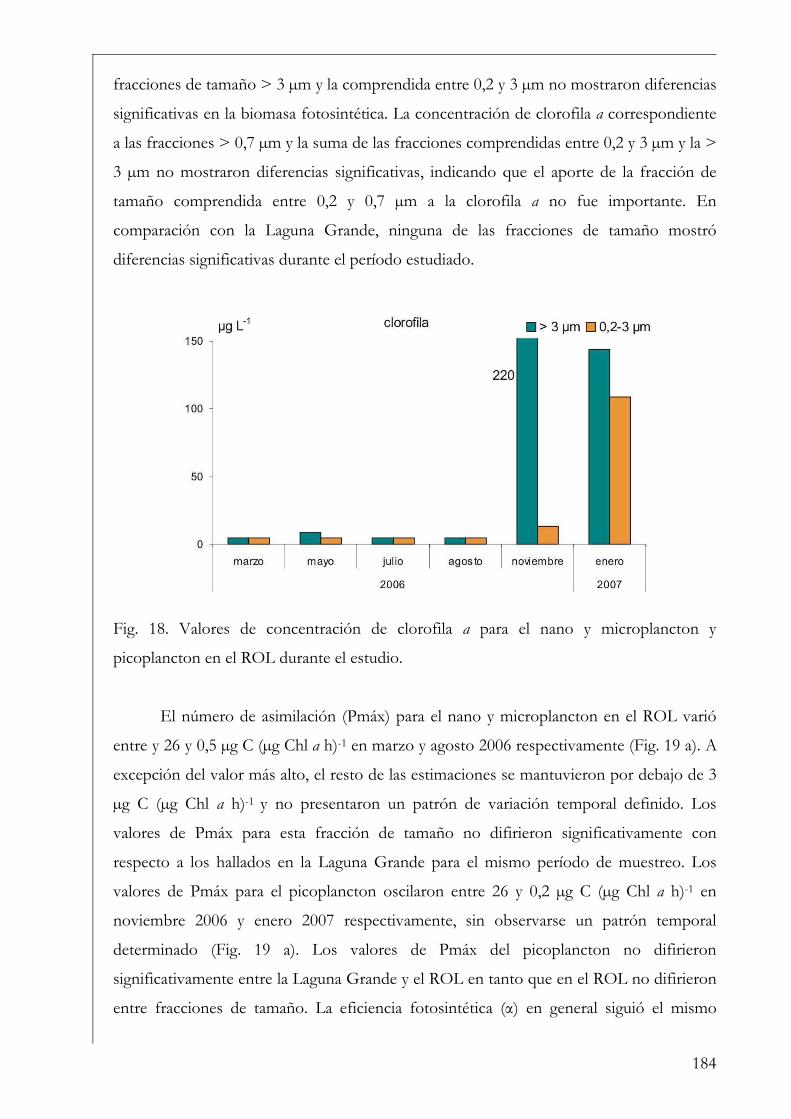
184
fracciones de tamaño > 3 µm y la comprendida entre 0,2 y 3 µm no mostraron diferencias
significativas en la biomasa fotosintética. La concentración de clorofila a correspondiente
a las fracciones > 0,7 µm y la suma de las fracciones comprendidas entre 0,2 y 3 µm y la >
3 µm no mostraron diferencias significativas, indicando que el aporte de la fracción de
tamaño comprendida entre 0,2 y 0,7 µm a la clorofila a no fue importante. En
comparación con la Laguna Grande, ninguna de las fracciones de tamaño mostró
diferencias significativas durante el período estudiado.
Fig. 18. Valores de concentración de clorofila a para el nano y microplancton y
picoplancton en el ROL durante el estudio.
El número de asimilación (Pmáx) para el nano y microplancton en el ROL varió
entre y 26 y 0,5 µg C (µg Chl a h)-1 en marzo y agosto 2006 respectivamente (Fig. 19 a). A
excepción del valor más alto, el resto de las estimaciones se mantuvieron por debajo de 3
µg C (µg Chl a h)-1 y no presentaron un patrón de variación temporal definido. Los
valores de Pmáx para esta fracción de tamaño no difirieron significativamente con
respecto a los hallados en la Laguna Grande para el mismo período de muestreo. Los
valores de Pmáx para el picoplancton oscilaron entre 26 y 0,2 µg C (µg Chl a h)-1 en
noviembre 2006 y enero 2007 respectivamente, sin observarse un patrón temporal
determinado (Fig. 19 a). Los valores de Pmáx del picoplancton no difirieron
significativamente entre la Laguna Grande y el ROL en tanto que en el ROL no difirieron
entre fracciones de tamaño. La eficiencia fotosintética (プ) en general siguió el mismo
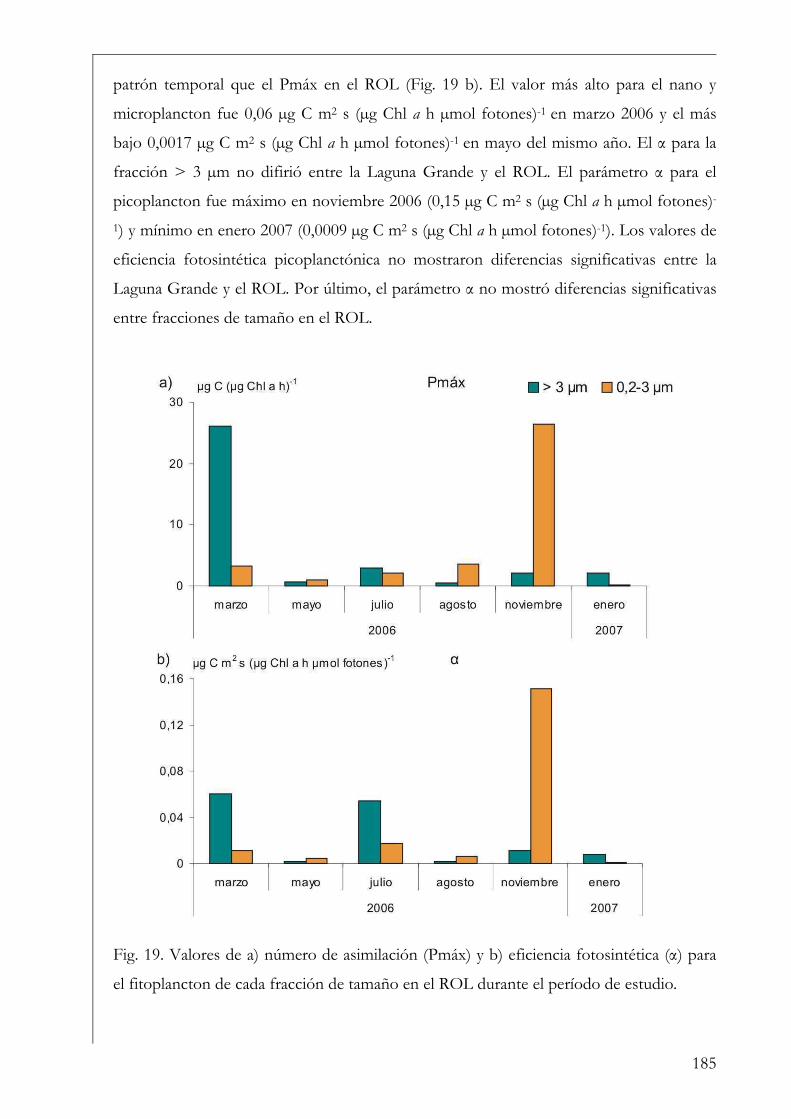
185
patrón temporal que el Pmáx en el ROL (Fig. 19 b). El valor más alto para el nano y
microplancton fue 0,06 µg C m2 s (µg Chl a h µmol fotones)-1 en marzo 2006 y el más
bajo 0,0017 µg C m2 s (µg Chl a h µmol fotones)-1 en mayo del mismo año. El プ para la
fracción > 3 µm no difirió entre la Laguna Grande y el ROL. El parámetro プ para el
picoplancton fue máximo en noviembre 2006 (0,15 µg C m2 s (µg Chl a h µmol fotones)-
1) y mínimo en enero 2007 (0,0009 µg C m2 s (µg Chl a h µmol fotones)-1). Los valores de
eficiencia fotosintética picoplanctónica no mostraron diferencias significativas entre la
Laguna Grande y el ROL. Por último, el parámetro プ no mostró diferencias significativas
entre fracciones de tamaño en el ROL.
Fig. 19. Valores de a) número de asimilación (Pmáx) y b) eficiencia fotosintética (プ) para
el fitoplancton de cada fracción de tamaño en el ROL durante el período de estudio.
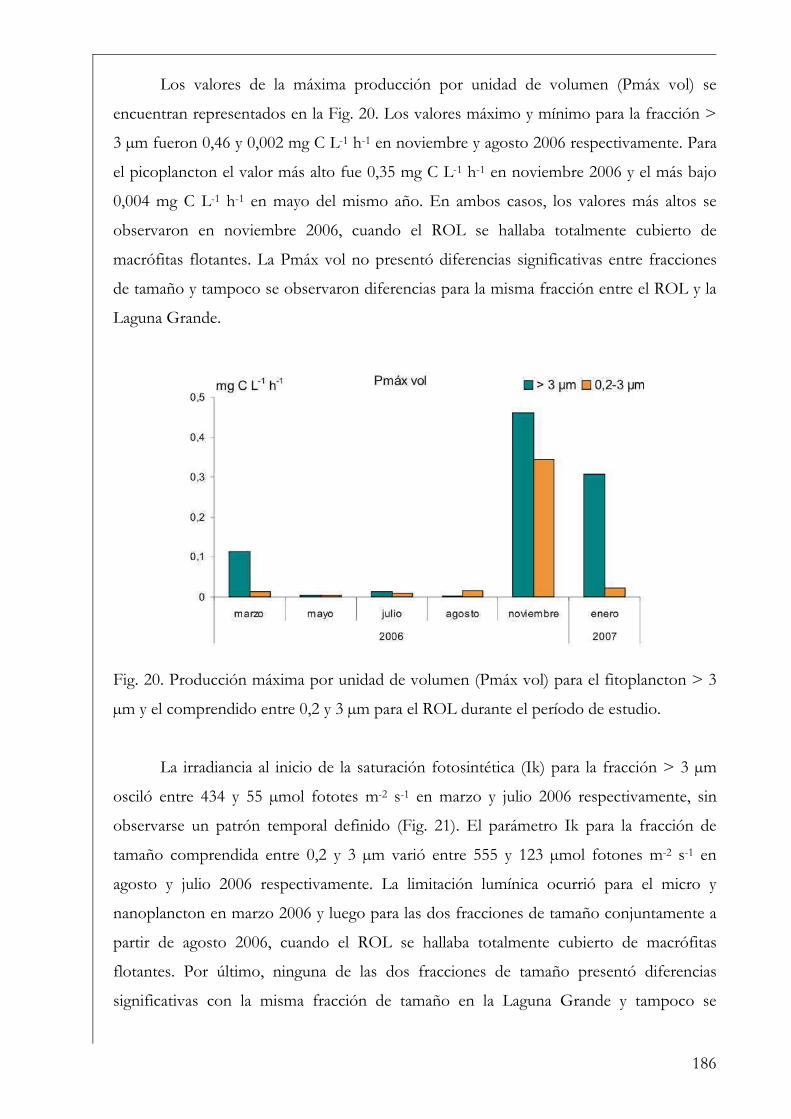
186
Los valores de la máxima producción por unidad de volumen (Pmáx vol) se
encuentran representados en la Fig. 20. Los valores máximo y mínimo para la fracción >
3 µm fueron 0,46 y 0,002 mg C L-1 h-1 en noviembre y agosto 2006 respectivamente. Para
el picoplancton el valor más alto fue 0,35 mg C L-1 h-1 en noviembre 2006 y el más bajo
0,004 mg C L-1 h-1 en mayo del mismo año. En ambos casos, los valores más altos se
observaron en noviembre 2006, cuando el ROL se hallaba totalmente cubierto de
macrófitas flotantes. La Pmáx vol no presentó diferencias significativas entre fracciones
de tamaño y tampoco se observaron diferencias para la misma fracción entre el ROL y la
Laguna Grande.
Fig. 20. Producción máxima por unidad de volumen (Pmáx vol) para el fitoplancton > 3
µm y el comprendido entre 0,2 y 3 µm para el ROL durante el período de estudio.
La irradiancia al inicio de la saturación fotosintética (Ik) para la fracción > 3 µm
osciló entre 434 y 55 µmol fototes m-2 s-1 en marzo y julio 2006 respectivamente, sin
observarse un patrón temporal definido (Fig. 21). El parámetro Ik para la fracción de
tamaño comprendida entre 0,2 y 3 µm varió entre 555 y 123 µmol fotones m-2 s-1 en
agosto y julio 2006 respectivamente. La limitación lumínica ocurrió para el micro y
nanoplancton en marzo 2006 y luego para las dos fracciones de tamaño conjuntamente a
partir de agosto 2006, cuando el ROL se hallaba totalmente cubierto de macrófitas
flotantes. Por último, ninguna de las dos fracciones de tamaño presentó diferencias
significativas con la misma fracción de tamaño en la Laguna Grande y tampoco se
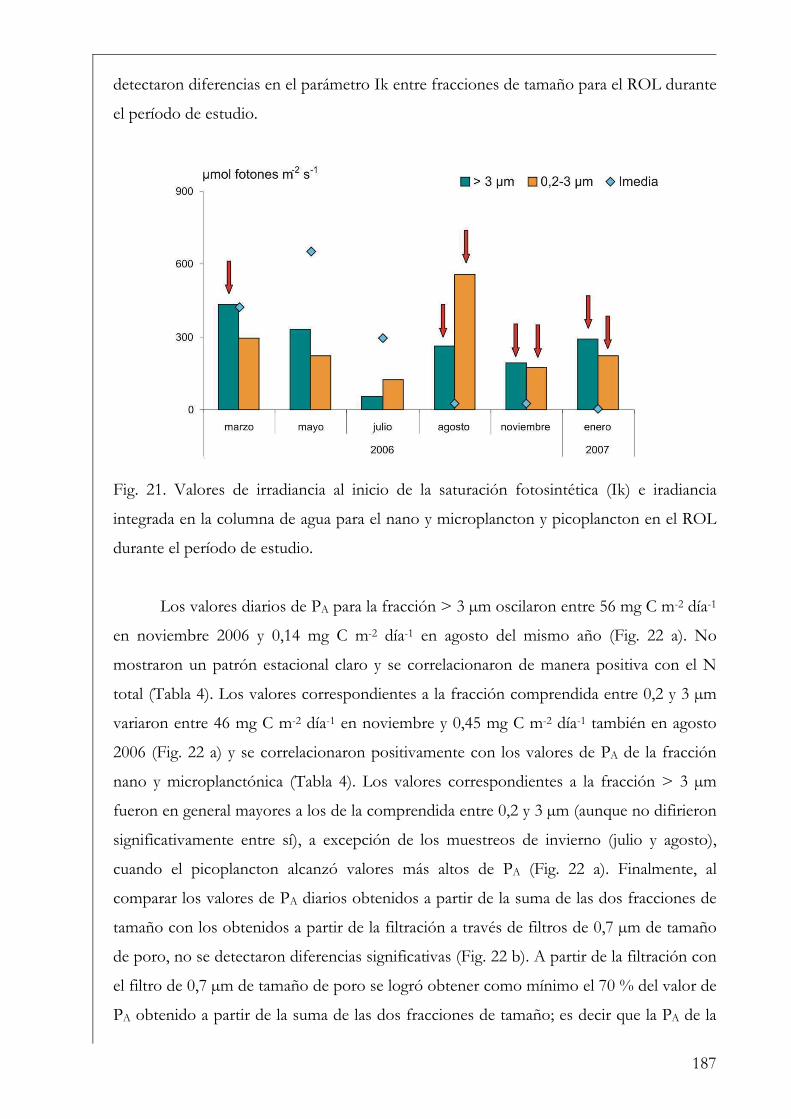
187
detectaron diferencias en el parámetro Ik entre fracciones de tamaño para el ROL durante
el período de estudio.
Fig. 21. Valores de irradiancia al inicio de la saturación fotosintética (Ik) e iradiancia
integrada en la columna de agua para el nano y microplancton y picoplancton en el ROL
durante el período de estudio.
Los valores diarios de PA para la fracción > 3 µm oscilaron entre 56 mg C m-2 día-1
en noviembre 2006 y 0,14 mg C m-2 día-1 en agosto del mismo año (Fig. 22 a). No
mostraron un patrón estacional claro y se correlacionaron de manera positiva con el N
total (Tabla 4). Los valores correspondientes a la fracción comprendida entre 0,2 y 3 µm
variaron entre 46 mg C m-2 día-1 en noviembre y 0,45 mg C m-2 día-1 también en agosto
2006 (Fig. 22 a) y se correlacionaron positivamente con los valores de PA de la fracción
nano y microplanctónica (Tabla 4). Los valores correspondientes a la fracción > 3 µm
fueron en general mayores a los de la comprendida entre 0,2 y 3 µm (aunque no difirieron
significativamente entre sí), a excepción de los muestreos de invierno (julio y agosto),
cuando el picoplancton alcanzó valores más altos de PA (Fig. 22 a). Finalmente, al
comparar los valores de PA diarios obtenidos a partir de la suma de las dos fracciones de
tamaño con los obtenidos a partir de la filtración a través de filtros de 0,7 µm de tamaño
de poro, no se detectaron diferencias significativas (Fig. 22 b). A partir de la filtración con
el filtro de 0,7 µm de tamaño de poro se logró obtener como mínimo el 70 % del valor de
PA obtenido a partir de la suma de las dos fracciones de tamaño; es decir que la PA de la
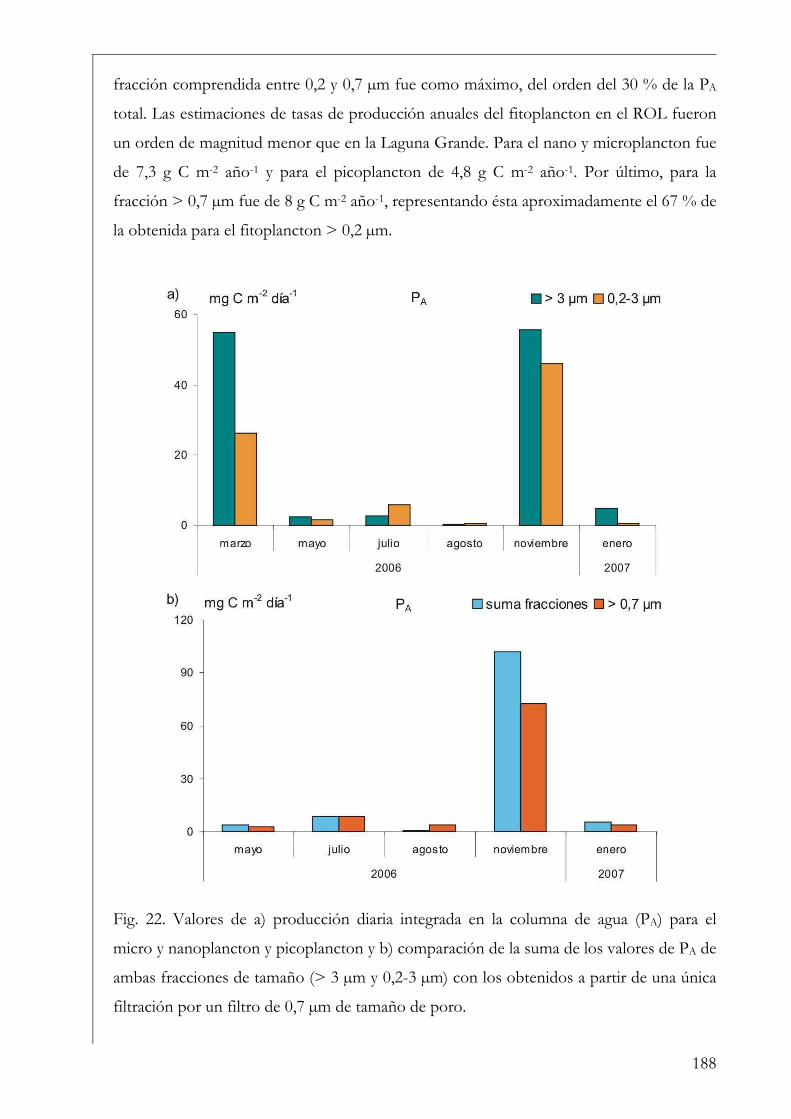
188
fracción comprendida entre 0,2 y 0,7 µm fue como máximo, del orden del 30 % de la PA
total. Las estimaciones de tasas de producción anuales del fitoplancton en el ROL fueron
un orden de magnitud menor que en la Laguna Grande. Para el nano y microplancton fue
de 7,3 g C m-2 año-1 y para el picoplancton de 4,8 g C m-2 año-1. Por último, para la
fracción > 0,7 µm fue de 8 g C m-2 año-1, representando ésta aproximadamente el 67 % de
la obtenida para el fitoplancton > 0,2 µm.
Fig. 22. Valores de a) producción diaria integrada en la columna de agua (PA) para el
micro y nanoplancton y picoplancton y b) comparación de la suma de los valores de PA de
ambas fracciones de tamaño (> 3 µm y 0,2-3 µm) con los obtenidos a partir de una única
filtración por un filtro de 0,7 µm de tamaño de poro.
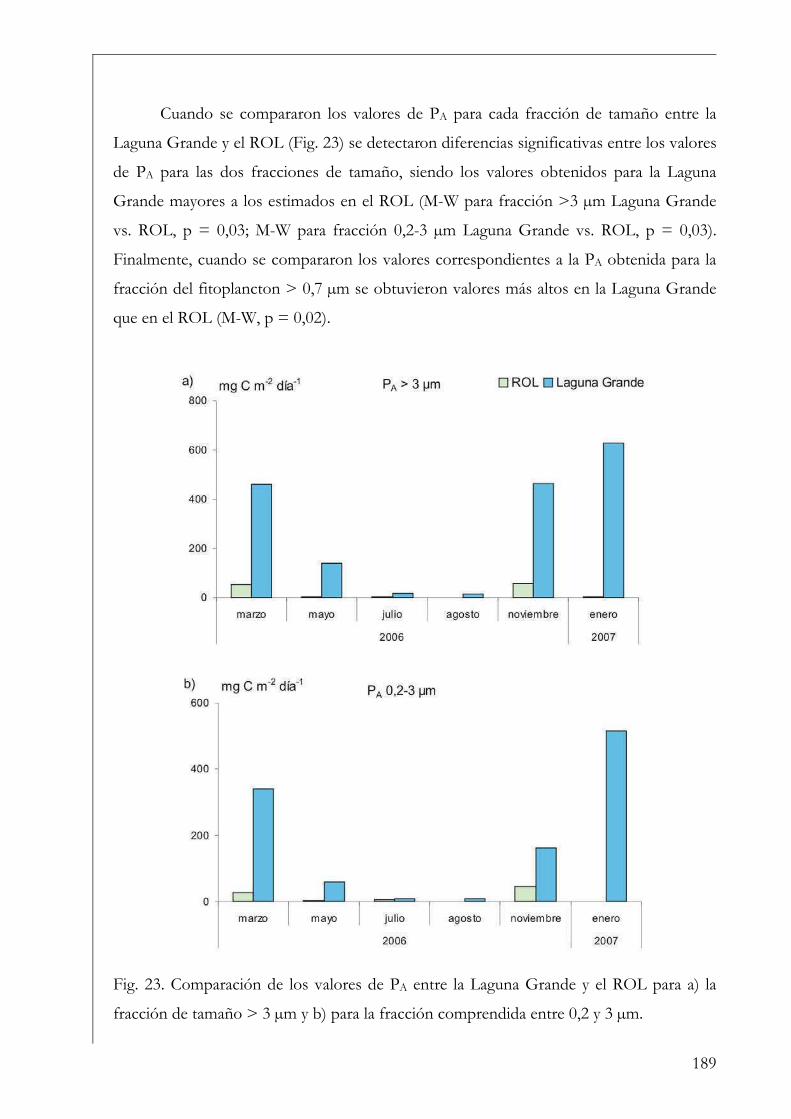
189
Cuando se compararon los valores de PA para cada fracción de tamaño entre la
Laguna Grande y el ROL (Fig. 23) se detectaron diferencias significativas entre los valores
de PA para las dos fracciones de tamaño, siendo los valores obtenidos para la Laguna
Grande mayores a los estimados en el ROL (M-W para fracción >3 µm Laguna Grande
vs. ROL, p = 0,03; M-W para fracción 0,2-3 µm Laguna Grande vs. ROL, p = 0,03).
Finalmente, cuando se compararon los valores correspondientes a la PA obtenida para la
fracción del fitoplancton > 0,7 µm se obtuvieron valores más altos en la Laguna Grande
que en el ROL (M-W, p = 0,02).
Fig. 23. Comparación de los valores de PA entre la Laguna Grande y el ROL para a) la
fracción de tamaño > 3 µm y b) para la fracción comprendida entre 0,2 y 3 µm.
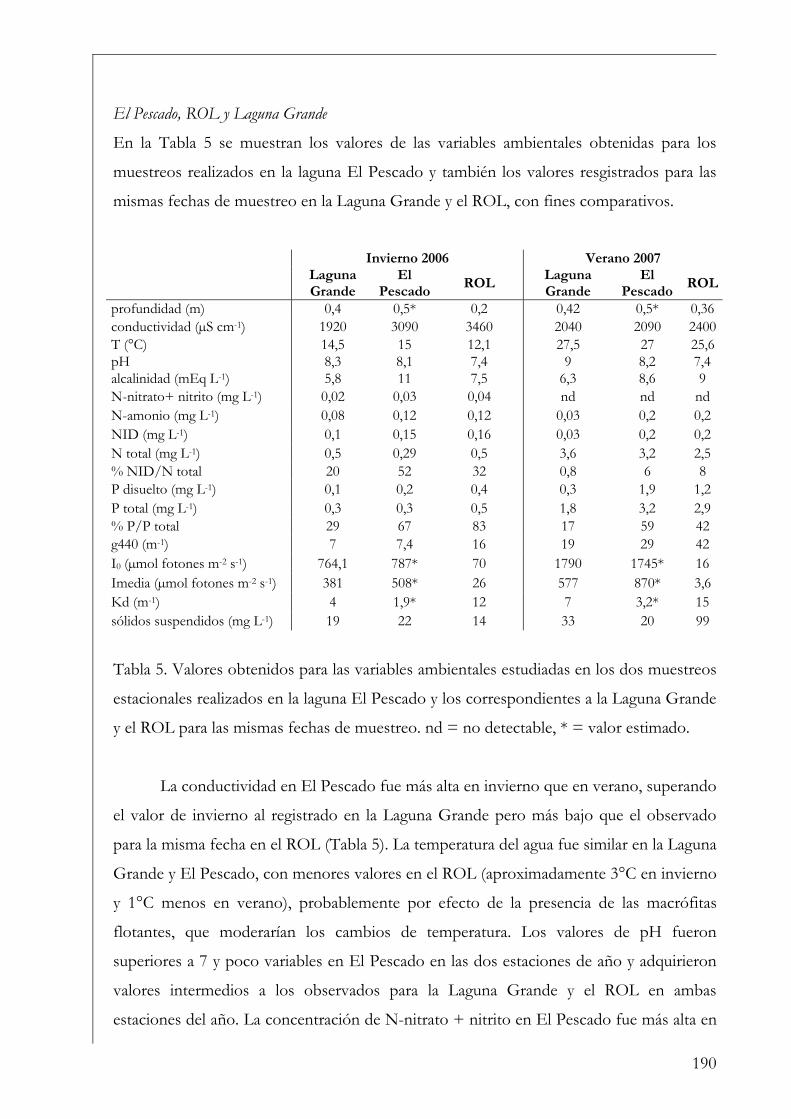
190
El Pescado, ROL y Laguna Grande
En la Tabla 5 se muestran los valores de las variables ambientales obtenidas para los
muestreos realizados en la laguna El Pescado y también los valores resgistrados para las
mismas fechas de muestreo en la Laguna Grande y el ROL, con fines comparativos.
Invierno 2006 Verano 2007
Laguna Grande
El Pescado
ROL Laguna Grande
El Pescado
ROL
profundidad (m) 0,4 0,5* 0,2 0,42 0,5* 0,36conductividad (µS cm-1) 1920 3090 3460 2040 2090 2400T (°C) 14,5 15 12,1 27,5 27 25,6pH 8,3 8,1 7,4 9 8,2 7,4 alcalinidad (mEq L-1) 5,8 11 7,5 6,3 8,6 9 N-nitrato+ nitrito (mg L-1) 0,02 0,03 0,04 nd nd nd N-amonio (mg L-1) 0,08 0,12 0,12 0,03 0,2 0,2 NID (mg L-1) 0,1 0,15 0,16 0,03 0,2 0,2 N total (mg L-1) 0,5 0,29 0,5 3,6 3,2 2,5 % NID/N total 20 52 32 0,8 6 8 P disuelto (mg L-1) 0,1 0,2 0,4 0,3 1,9 1,2 P total (mg L-1) 0,3 0,3 0,5 1,8 3,2 2,9 % P/P total 29 67 83 17 59 42 g440 (m-1) 7 7,4 16 19 29 42 I0 (µmol fotones m-2 s-1) 764,1 787* 70 1790 1745* 16 Imedia (µmol fotones m-2 s-1) 381 508* 26 577 870* 3,6 Kd (m-1) 4 1,9* 12 7 3,2* 15 sólidos suspendidos (mg L-1) 19 22 14 33 20 99
Tabla 5. Valores obtenidos para las variables ambientales estudiadas en los dos muestreos
estacionales realizados en la laguna El Pescado y los correspondientes a la Laguna Grande
y el ROL para las mismas fechas de muestreo. nd = no detectable, * = valor estimado.
La conductividad en El Pescado fue más alta en invierno que en verano, superando
el valor de invierno al registrado en la Laguna Grande pero más bajo que el observado
para la misma fecha en el ROL (Tabla 5). La temperatura del agua fue similar en la Laguna
Grande y El Pescado, con menores valores en el ROL (aproximadamente 3°C en invierno
y 1°C menos en verano), probablemente por efecto de la presencia de las macrófitas
flotantes, que moderarían los cambios de temperatura. Los valores de pH fueron
superiores a 7 y poco variables en El Pescado en las dos estaciones de año y adquirieron
valores intermedios a los observados para la Laguna Grande y el ROL en ambas
estaciones del año. La concentración de N-nitrato + nitrito en El Pescado fue más alta en

191
invierno (0,03 mg L-1) que en verano, cuando resultó no detectable en los tres cuerpos de
agua. Los valores de N-amonio en El Pescado fueron más similares a los registrados en el
ROL en los dos muestreos (0,12 mg L-1 en invierno y 0,2 mg L-1 en verano). La
concentración de NID en El Pescado se encontró por encima de los valores considerados
como limitantes para el máximo crecimiento algal (sensu Reynolds 2006). Como ya se ha
mencionado previamente tanto para la Laguna Grande como para el ROL, en El Pescado
también se puede advertir que fue en invierno cuando la mayor parte del N se encontraba
en la forma inorgánica. El porcentaje entre el NID y el N total fue mayor en El Pescado
en invierno con respecto a los otros cuerpos de agua estudiados (52 %). En El Pescado,
tanto la concentración de P disuelto como P total fue mayor en verano. Los valores de P
disuelto (0,2 mg L-1 en invierno y 1,9 mg L-1 en verano) se encontraron por encima de los
considerados limitantes para el máximo crecimiento algal (sensu Reynolds 1984). El
contenido húmico en El Pescado fue cuatro veces mayor en verano que en invierno; fue
similar al de la Laguna Grande en invierno y más alto en verano. Por último, los valores
estimados de Kd muestran que en El Pescado se detectaron los valores más bajos tanto
en invierno como en verano, siendo aproximadamente la mitad de los obtenidos para la
Laguna Grande; registrándose así los mayores valores de Imedia en la laguna El Pescado
(Tabla 5).
Las densidades algales del micro y nanoplancton en El Pescado alcanzaron valores
intermedios entre la Laguna Grande y el ROL (Fig. 24 a). La densidad más alta en este
cuerpo de agua se observó en verano (66000 ind mL-1) y la más baja en invierno (4000 ind
mL-1). La composición relativa de las clases algales se halla representada en la Fig. 24 b.
Las diatomeas Cyclotella meneghiniana y Nitzschia spp. fueron dominantes en invierno,
acompañadas de las clorofíceas Monoraphidium spp. y Chlamydomonas spp.; también se
registraron en este muestro algunas crisofíceas. En el verano dominaron las
cianobacterias, siendo Planktolyngbya limnetica la especie dominante. También se detectaron
clorofíceas (Monoraphidium circinale), otras cianobacterias (Arthospira sp. y Merismopedia
tenuisima) y crisofíceas.
La concentración de clorofila a para el nano y microplancton de la laguna El
Pescado fue de 4 y 24 µg L-1 en invierno y verano respectivamente, mostrando una
tendencia estacional; mientras que para el picoplancton, la concentración de clorofila a en
los dos muestreos fue de 4 µg L-1 (Fig. 25). Por otro lado, cuando se compararon los
valores de concentración de clorofila a de la fracción de tamaño > 0,7 µm con los
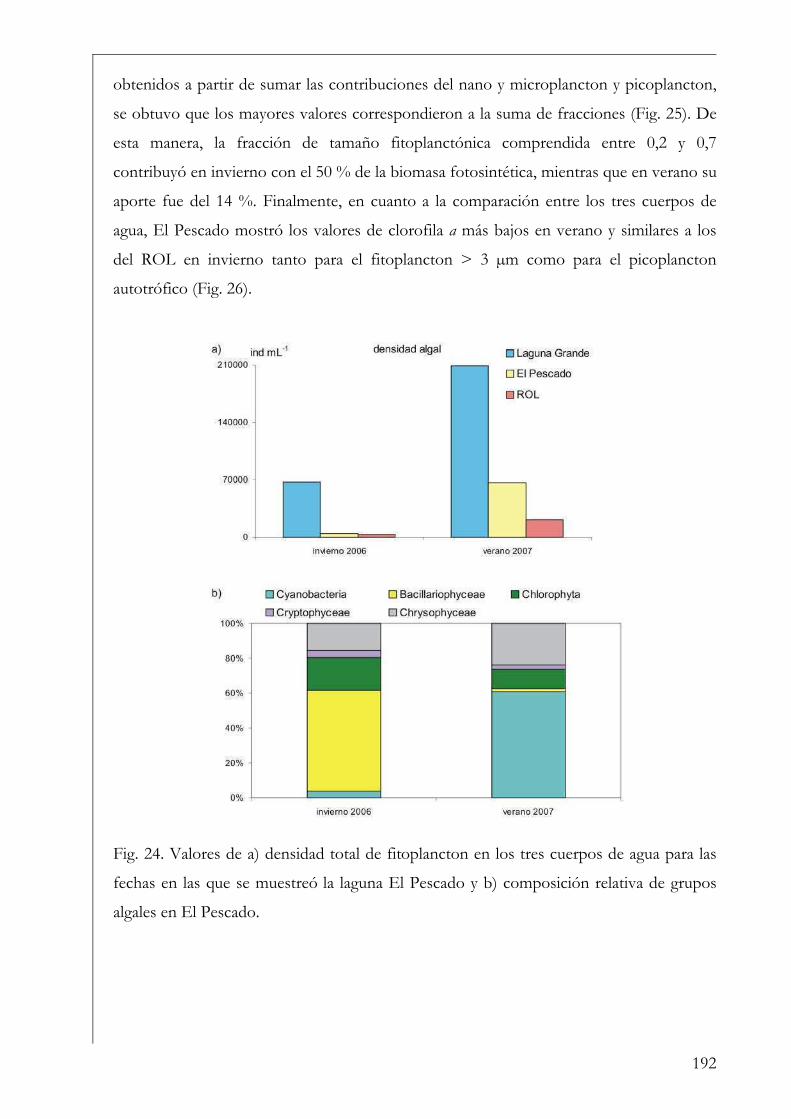
192
obtenidos a partir de sumar las contribuciones del nano y microplancton y picoplancton,
se obtuvo que los mayores valores correspondieron a la suma de fracciones (Fig. 25). De
esta manera, la fracción de tamaño fitoplanctónica comprendida entre 0,2 y 0,7
contribuyó en invierno con el 50 % de la biomasa fotosintética, mientras que en verano su
aporte fue del 14 %. Finalmente, en cuanto a la comparación entre los tres cuerpos de
agua, El Pescado mostró los valores de clorofila a más bajos en verano y similares a los
del ROL en invierno tanto para el fitoplancton > 3 µm como para el picoplancton
autotrófico (Fig. 26).
Fig. 24. Valores de a) densidad total de fitoplancton en los tres cuerpos de agua para las
fechas en las que se muestreó la laguna El Pescado y b) composición relativa de grupos
algales en El Pescado.
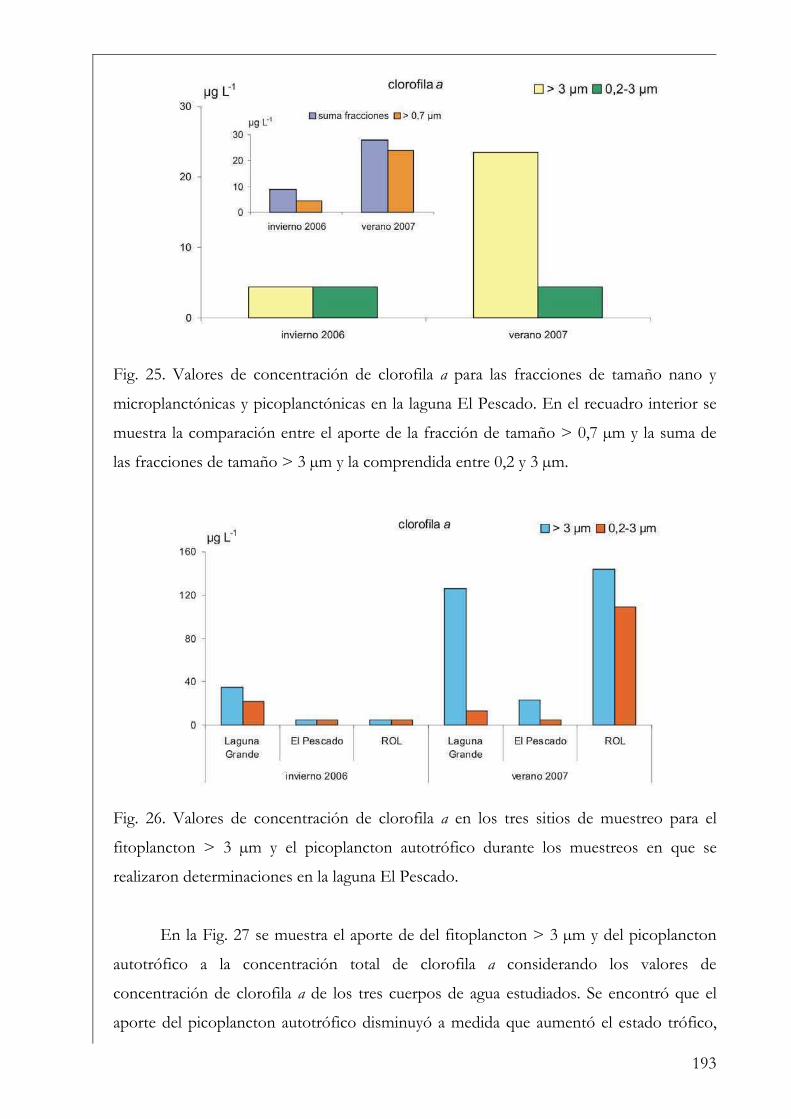
193
Fig. 25. Valores de concentración de clorofila a para las fracciones de tamaño nano y
microplanctónicas y picoplanctónicas en la laguna El Pescado. En el recuadro interior se
muestra la comparación entre el aporte de la fracción de tamaño > 0,7 µm y la suma de
las fracciones de tamaño > 3 µm y la comprendida entre 0,2 y 3 µm.
Fig. 26. Valores de concentración de clorofila a en los tres sitios de muestreo para el
fitoplancton > 3 µm y el picoplancton autotrófico durante los muestreos en que se
realizaron determinaciones en la laguna El Pescado.
En la Fig. 27 se muestra el aporte de del fitoplancton > 3 µm y del picoplancton
autotrófico a la concentración total de clorofila a considerando los valores de
concentración de clorofila a de los tres cuerpos de agua estudiados. Se encontró que el
aporte del picoplancton autotrófico disminuyó a medida que aumentó el estado trófico,
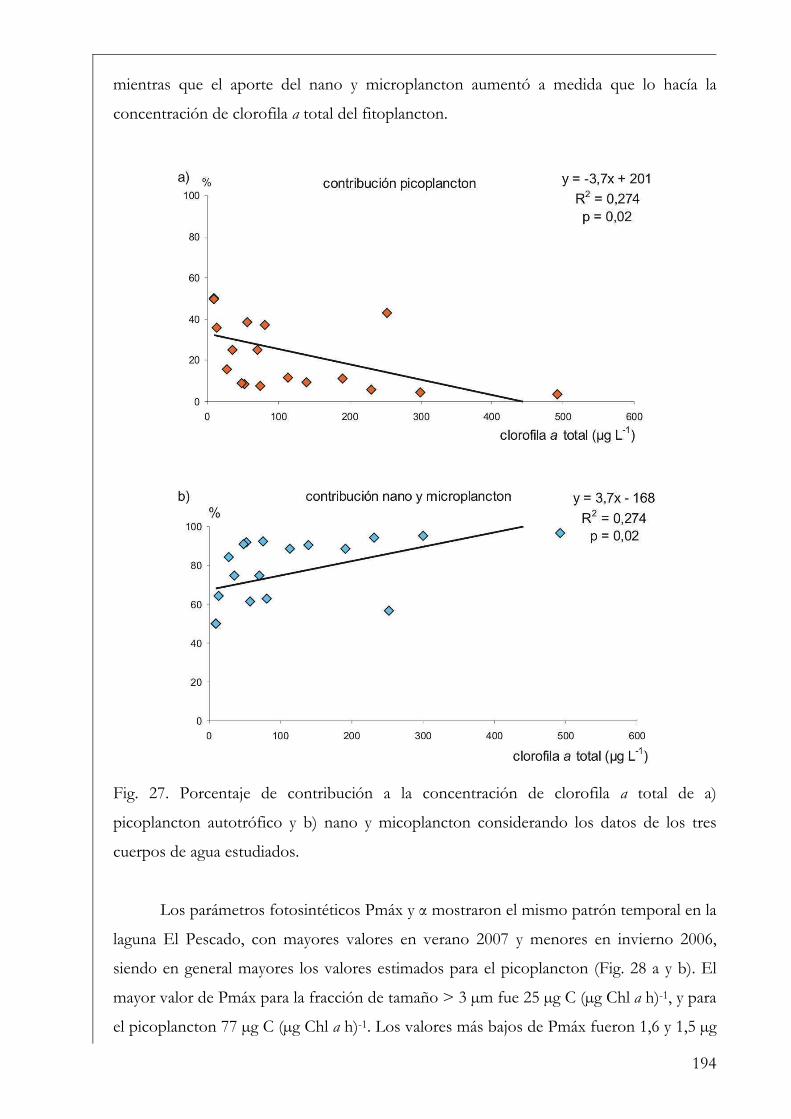
194
mientras que el aporte del nano y microplancton aumentó a medida que lo hacía la
concentración de clorofila a total del fitoplancton.
Fig. 27. Porcentaje de contribución a la concentración de clorofila a total de a)
picoplancton autotrófico y b) nano y micoplancton considerando los datos de los tres
cuerpos de agua estudiados.
Los parámetros fotosintéticos Pmáx y プ mostraron el mismo patrón temporal en la
laguna El Pescado, con mayores valores en verano 2007 y menores en invierno 2006,
siendo en general mayores los valores estimados para el picoplancton (Fig. 28 a y b). El
mayor valor de Pmáx para la fracción de tamaño > 3 µm fue 25 µg C (µg Chl a h)-1, y para
el picoplancton 77 µg C (µg Chl a h)-1. Los valores más bajos de Pmáx fueron 1,6 y 1,5 µg
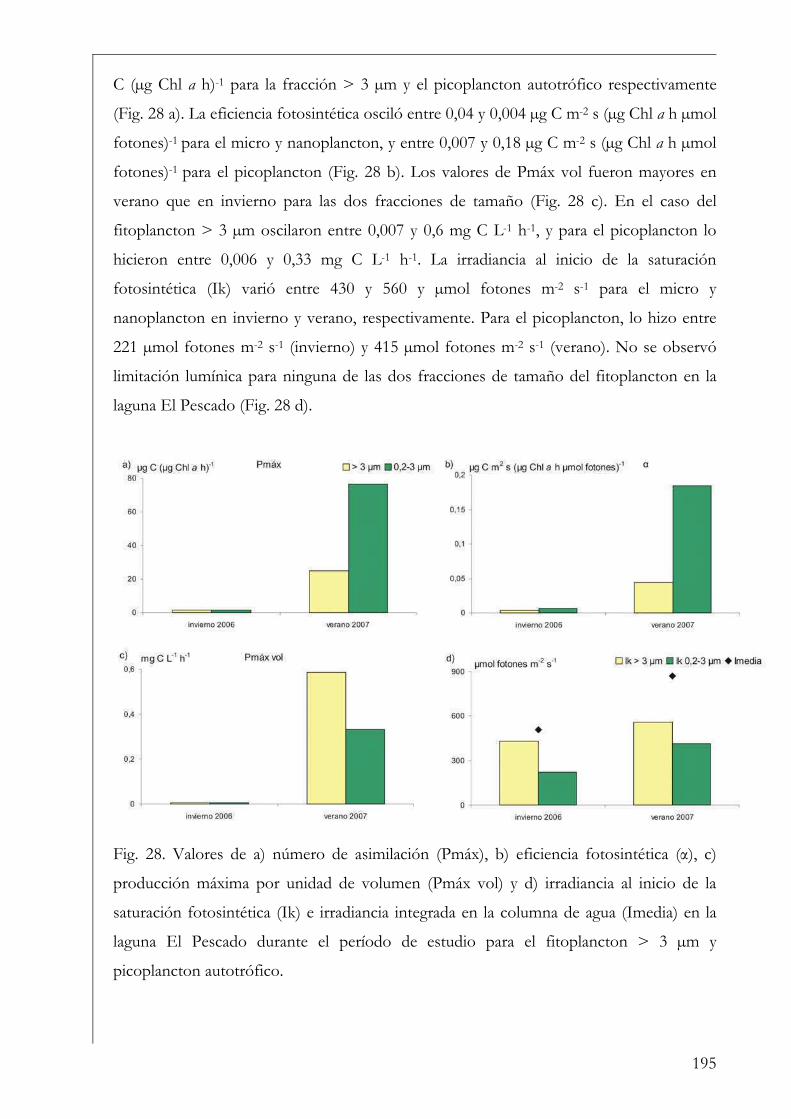
195
C (µg Chl a h)-1 para la fracción > 3 µm y el picoplancton autotrófico respectivamente
(Fig. 28 a). La eficiencia fotosintética osciló entre 0,04 y 0,004 µg C m-2 s (µg Chl a h µmol
fotones)-1 para el micro y nanoplancton, y entre 0,007 y 0,18 µg C m-2 s (µg Chl a h µmol
fotones)-1 para el picoplancton (Fig. 28 b). Los valores de Pmáx vol fueron mayores en
verano que en invierno para las dos fracciones de tamaño (Fig. 28 c). En el caso del
fitoplancton > 3 µm oscilaron entre 0,007 y 0,6 mg C L-1 h-1, y para el picoplancton lo
hicieron entre 0,006 y 0,33 mg C L-1 h-1. La irradiancia al inicio de la saturación
fotosintética (Ik) varió entre 430 y 560 y µmol fotones m-2 s-1 para el micro y
nanoplancton en invierno y verano, respectivamente. Para el picoplancton, lo hizo entre
221 µmol fotones m-2 s-1 (invierno) y 415 µmol fotones m-2 s-1 (verano). No se observó
limitación lumínica para ninguna de las dos fracciones de tamaño del fitoplancton en la
laguna El Pescado (Fig. 28 d).
Fig. 28. Valores de a) número de asimilación (Pmáx), b) eficiencia fotosintética (プ), c)
producción máxima por unidad de volumen (Pmáx vol) y d) irradiancia al inicio de la
saturación fotosintética (Ik) e irradiancia integrada en la columna de agua (Imedia) en la
laguna El Pescado durante el período de estudio para el fitoplancton > 3 µm y
picoplancton autotrófico.
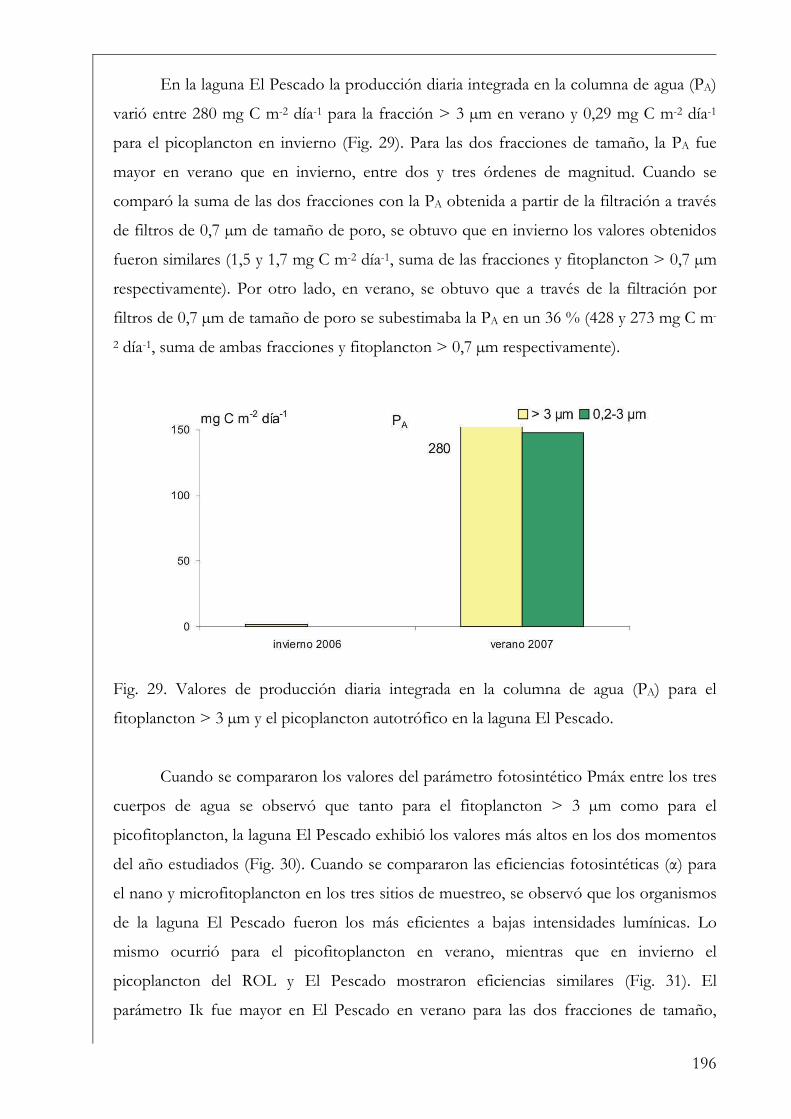
196
En la laguna El Pescado la producción diaria integrada en la columna de agua (PA)
varió entre 280 mg C m-2 día-1 para la fracción > 3 µm en verano y 0,29 mg C m-2 día-1
para el picoplancton en invierno (Fig. 29). Para las dos fracciones de tamaño, la PA fue
mayor en verano que en invierno, entre dos y tres órdenes de magnitud. Cuando se
comparó la suma de las dos fracciones con la PA obtenida a partir de la filtración a través
de filtros de 0,7 µm de tamaño de poro, se obtuvo que en invierno los valores obtenidos
fueron similares (1,5 y 1,7 mg C m-2 día-1, suma de las fracciones y fitoplancton > 0,7 µm
respectivamente). Por otro lado, en verano, se obtuvo que a través de la filtración por
filtros de 0,7 µm de tamaño de poro se subestimaba la PA en un 36 % (428 y 273 mg C m-
2 día-1, suma de ambas fracciones y fitoplancton > 0,7 µm respectivamente).
Fig. 29. Valores de producción diaria integrada en la columna de agua (PA) para el
fitoplancton > 3 µm y el picoplancton autotrófico en la laguna El Pescado.
Cuando se compararon los valores del parámetro fotosintético Pmáx entre los tres
cuerpos de agua se observó que tanto para el fitoplancton > 3 µm como para el
picofitoplancton, la laguna El Pescado exhibió los valores más altos en los dos momentos
del año estudiados (Fig. 30). Cuando se compararon las eficiencias fotosintéticas (プ) para
el nano y microfitoplancton en los tres sitios de muestreo, se observó que los organismos
de la laguna El Pescado fueron los más eficientes a bajas intensidades lumínicas. Lo
mismo ocurrió para el picofitoplancton en verano, mientras que en invierno el
picoplancton del ROL y El Pescado mostraron eficiencias similares (Fig. 31). El
parámetro Ik fue mayor en El Pescado en verano para las dos fracciones de tamaño,
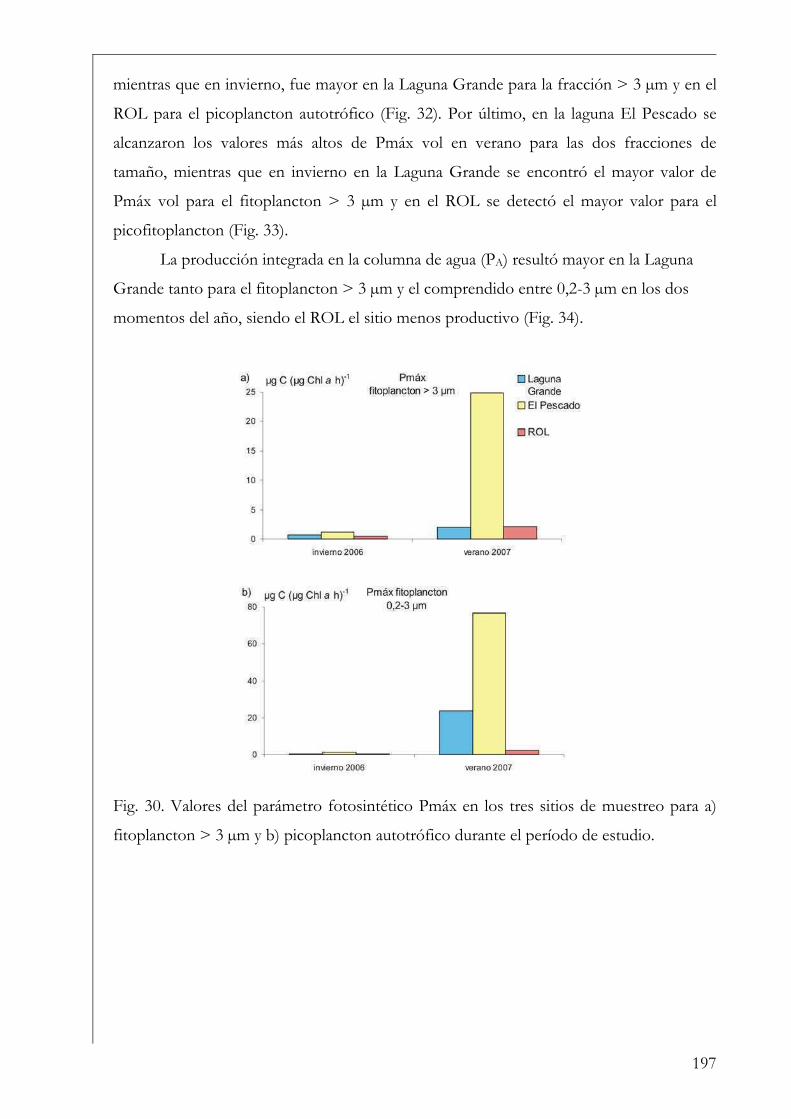
197
mientras que en invierno, fue mayor en la Laguna Grande para la fracción > 3 µm y en el
ROL para el picoplancton autotrófico (Fig. 32). Por último, en la laguna El Pescado se
alcanzaron los valores más altos de Pmáx vol en verano para las dos fracciones de
tamaño, mientras que en invierno en la Laguna Grande se encontró el mayor valor de
Pmáx vol para el fitoplancton > 3 µm y en el ROL se detectó el mayor valor para el
picofitoplancton (Fig. 33).
La producción integrada en la columna de agua (PA) resultó mayor en la Laguna
Grande tanto para el fitoplancton > 3 µm y el comprendido entre 0,2-3 µm en los dos
momentos del año, siendo el ROL el sitio menos productivo (Fig. 34).
Fig. 30. Valores del parámetro fotosintético Pmáx en los tres sitios de muestreo para a)
fitoplancton > 3 µm y b) picoplancton autotrófico durante el período de estudio.
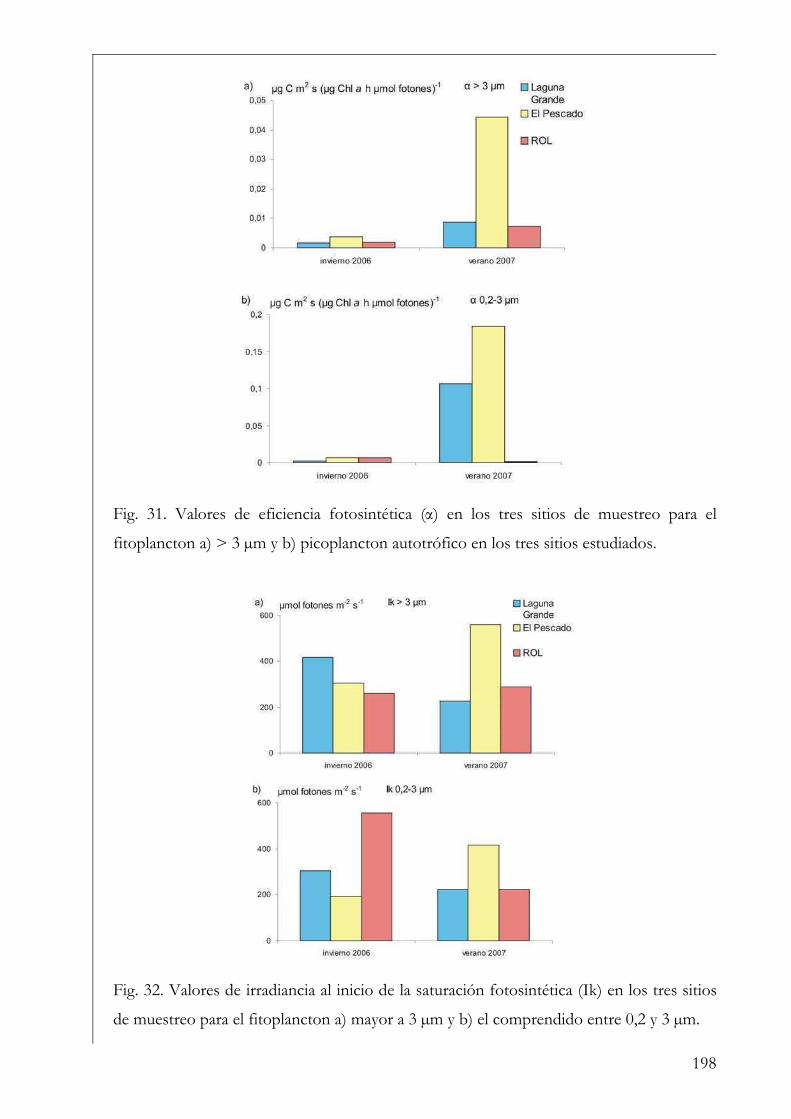
198
Fig. 31. Valores de eficiencia fotosintética (プ) en los tres sitios de muestreo para el
fitoplancton a) > 3 µm y b) picoplancton autotrófico en los tres sitios estudiados.
Fig. 32. Valores de irradiancia al inicio de la saturación fotosintética (Ik) en los tres sitios
de muestreo para el fitoplancton a) mayor a 3 µm y b) el comprendido entre 0,2 y 3 µm.
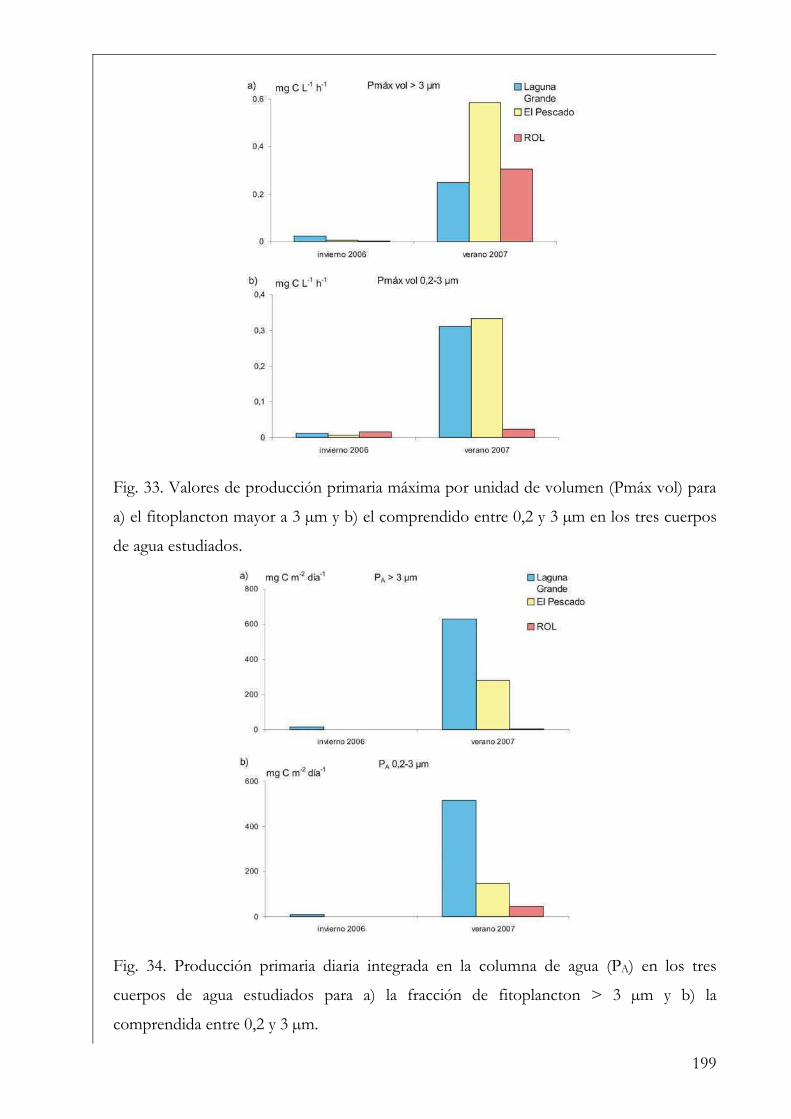
199
Fig. 33. Valores de producción primaria máxima por unidad de volumen (Pmáx vol) para
a) el fitoplancton mayor a 3 µm y b) el comprendido entre 0,2 y 3 µm en los tres cuerpos
de agua estudiados.
Fig. 34. Producción primaria diaria integrada en la columna de agua (PA) en los tres
cuerpos de agua estudiados para a) la fracción de fitoplancton > 3 µm y b) la
comprendida entre 0,2 y 3 µm.
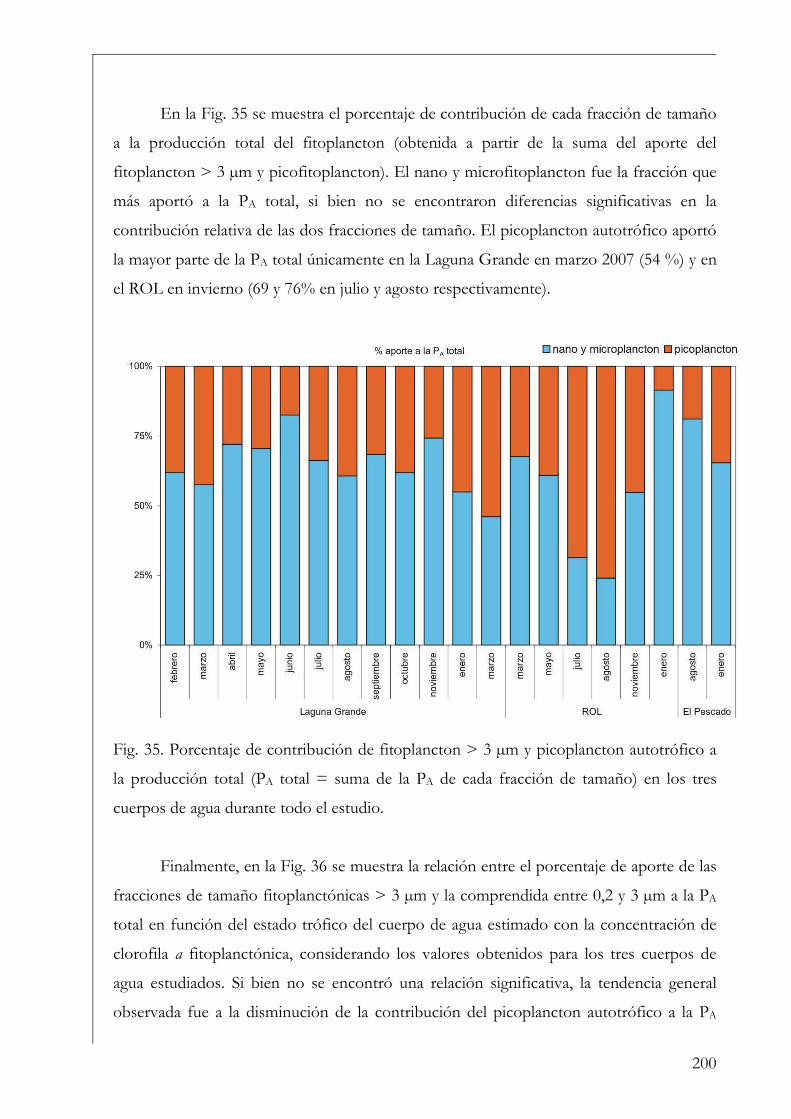
200
En la Fig. 35 se muestra el porcentaje de contribución de cada fracción de tamaño
a la producción total del fitoplancton (obtenida a partir de la suma del aporte del
fitoplancton > 3 µm y picofitoplancton). El nano y microfitoplancton fue la fracción que
más aportó a la PA total, si bien no se encontraron diferencias significativas en la
contribución relativa de las dos fracciones de tamaño. El picoplancton autotrófico aportó
la mayor parte de la PA total únicamente en la Laguna Grande en marzo 2007 (54 %) y en
el ROL en invierno (69 y 76% en julio y agosto respectivamente).
Fig. 35. Porcentaje de contribución de fitoplancton > 3 µm y picoplancton autotrófico a
la producción total (PA total = suma de la PA de cada fracción de tamaño) en los tres
cuerpos de agua durante todo el estudio.
Finalmente, en la Fig. 36 se muestra la relación entre el porcentaje de aporte de las
fracciones de tamaño fitoplanctónicas > 3 µm y la comprendida entre 0,2 y 3 µm a la PA
total en función del estado trófico del cuerpo de agua estimado con la concentración de
clorofila a fitoplanctónica, considerando los valores obtenidos para los tres cuerpos de
agua estudiados. Si bien no se encontró una relación significativa, la tendencia general
observada fue a la disminución de la contribución del picoplancton autotrófico a la PA
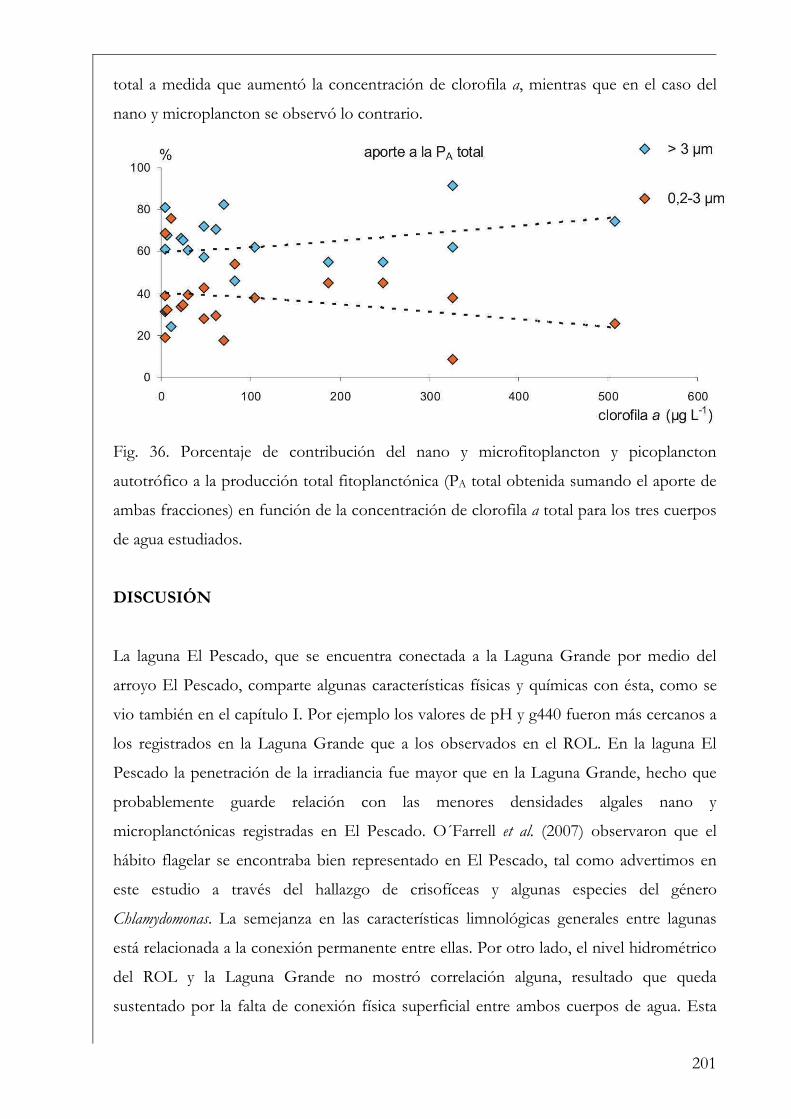
201
total a medida que aumentó la concentración de clorofila a, mientras que en el caso del
nano y microplancton se observó lo contrario.
Fig. 36. Porcentaje de contribución del nano y microfitoplancton y picoplancton
autotrófico a la producción total fitoplanctónica (PA total obtenida sumando el aporte de
ambas fracciones) en función de la concentración de clorofila a total para los tres cuerpos
de agua estudiados.
DISCUSIÓN
La laguna El Pescado, que se encuentra conectada a la Laguna Grande por medio del
arroyo El Pescado, comparte algunas características físicas y químicas con ésta, como se
vio también en el capítulo I. Por ejemplo los valores de pH y g440 fueron más cercanos a
los registrados en la Laguna Grande que a los observados en el ROL. En la laguna El
Pescado la penetración de la irradiancia fue mayor que en la Laguna Grande, hecho que
probablemente guarde relación con las menores densidades algales nano y
microplanctónicas registradas en El Pescado. O´Farrell et al. (2007) observaron que el
hábito flagelar se encontraba bien representado en El Pescado, tal como advertimos en
este estudio a través del hallazgo de crisofíceas y algunas especies del género
Chlamydomonas. La semejanza en las características limnológicas generales entre lagunas
está relacionada a la conexión permanente entre ellas. Por otro lado, el nivel hidrométrico
del ROL y la Laguna Grande no mostró correlación alguna, resultado que queda
sustentado por la falta de conexión física superficial entre ambos cuerpos de agua. Esta

202
falta de conexión superficial durante la mayor parte del año, permitiría que se desarrollen
en el ROL características distintivas con respecto a las lagunas. Sólo en períodos de nivel
hidrométrico extremadamente altos hemos observado conexión superficial entre los
cuerpos de agua del humedal, como por ejemplo en febrero 2005 (obs. pers.).
Las macrófitas flotantes de pequeño porte, como Ricciocarpus natans y lemnáceas,
fueron un elemento dominante en el ROL durante gran parte del estudio. La cobertura
total del espejo de agua a partir del invierno de 2006 trajo aparejado, entre otros efectos
asociados a la disminución de la penetración de la luz, la anoxia en la columna de agua, el
aumento de la profundidad óptica, el aumento del coeficiente de atenuación vertical, de
los sólidos en suspensión y del contenido húmico. También la disminución en la
concentración de N-nitrato + nitrito y un aumento en la del N-amonio. A diferencia de lo
que ocurría en la Laguna Grande, donde la concentración de sólidos en suspensión seguía
el mismo patrón que la concentración de clorofila a, en el ROL la fluctuación temporal de
la concentración de sólidos suspendidos estuvo asociada con la presencia de macrofitia
flotante, tal como toda la dinámica de este cuerpo de agua.
El ROL es un cuerpo de agua muy somero y de menores dimensiones que las
lagunas Grande y El Pescado. El pH del agua resultó ser más bajo en el ROL que en la
Laguna Grande y en El Pescado probablemente como consecuencia de las menores
densidades y producción primaria fitoplanctónicas. La penetración de la luz fue menor en
el ROL que en la Laguna Grande y en general, las concentraciones de nutrientes (en
particular N y P) fueron más altas en el ROL. Las formas inorgánicas del N y P fueron
más abundantes en invierno, de manera análoga a lo ocurrido en la Laguna Grande y El
Pescado. Las concentraciones de P disuelto mostraron una tendencia a su disminución a
partir del momento en que el cuerpo de agua se cubrió de macrófitas flotantes,
probablemente causado por el mayor consumo del bacterioplancton, cuya respiración
contribuiría a la generación de condiciones anóxicas.
La composición algal micro y nanoplanctónica del ROL fue similar a la reportada
en estudios previos por Izaguirre et al. (2004) y O´Farrell et al. (2003) para este mismo
cuerpo de agua. Las densidades fitoplanctónicas halladas fueron en general bajas y
menores a las registradas en la Laguna Grande. Izaguirre et al. (2004, 2001 b) registraron
para el ROL densidades máximas de alrededor de 10000 ind mL-1, mientras que en este
estudio en una sola oportunidad (enero 2007) se superó sustancialmente ese valor (21380
ind mL-1) coincidentemente con la mayor densidad de esa fracción encontrada también en

203
la Laguna Grande. Es interesante mencionar que no se observó el predominio de
cianobacterias filamentosas en el ROL como sí se hizo en la laguna El Pescado en verano.
En el ROL, donde la penetración de la irradiancia fue menor, se vieron aventajadas las
formas cocoides y agregados de cianobacterias con mayor relación superficie/volumen,
característica que permite maximizar la captación de luz. También en relación al ROL,
advertimos que gran parte de la comunidad estaba constituida por diatomeas perifíticas
provenientes de la comunidad desarrollada sobre sustratos naturales. Este hecho es muy
común en cuerpos de agua someros donde las comunidades algales suelen solapar sus
hábitats (Goldsborough & Robinson 1996).
Cuando se analizaron los valores de la concentración de clorofila a fitoplanctónica para el
micro y nanoplancton de la Laguna Grande puede observarse que presentaron un patrón
estacional durante el estudio, análogo al encontrado en el capítulo II para el fitoplancton
> 0,7 µm de tamaño. Para el picoplancton, los valores de clorofila a estimados fueron
altos cuando se los compara con algunos valores de la bibliografía, si bien no
encontramos precisamente datos provenientes de sistemas húmicos de características
similares (Drakare et al. 2003, Steitz & Velimirov 1999, Stockner 1988). Los altos valores
hallados por nosotros pueden deberse en parte a que en este estudio consideramos como
picoplancton a la fracción comprendida entre 0,2 y 3 µm, es decir, que también
consideramos la concentración de clorofila a de una fracción del nanoplancton además de
la del picoplancton. Sin embargo, de manera parecida a lo hallado para picocianobacterias
por Vörös et al. (1998), cuando registramos valores de concentración de clorofila a total
que superaban los 100 µg L-1, la contribución del picoplancton raramente excedió el 10 %
teniendo en cuenta los valores de los tres cuerpos de agua del humedal. Considerando que
en el humedal el picoplancton está representado mayoritariamente por picocianobacterias
tal como señalaran Sinistro et al. (2007, 2006) e Izaguirre et al. (enviado) nuestros
resultados son comparables a los de Vörös et al. (op cit.).
La disminución del aporte del picoplancton a la concentración total de clorofila a a
medida que aumenta la biomasa fotosintética total es un hecho ya registrado en la
bibliografía. Agawin et al. (2000) encontraron que en el mar en aguas poco productivas, el
picoplancton tenía mayor biomasa fotosintética que el micro y nanoplancton, mientras
que en regiones más productivas del océano, ocurría lo inverso. Bell & Kalff (2001),
recopilando datos de la bibliografía para el mar y el agua dulce, mostraron que la

204
concentración de clorofila a picoplanctónica aumentaba con el estado trófico del cuerpo
de agua, es decir que los sistemas eutróficos contienen más picoplancton que sus
contrapartes oligotróficos marinos o de agua dulce. Sin embargo, encontraron que la
contribución relativa a la biomasa fotosintética total fitoplanctónica por parte del
picoplancton disminuía, análogamente a lo encontrado por nosotros. En nuestro caso,
esta relación la encontramos en un rango de concentraciones de clorofila a total altas (9 a
500 µg L-1), es decir, vimos esta diferencia siempre dentro de un marco eutrófico.
En el ROL, resulta llamativo el aumento de la concentración de clorofila a a partir
del mes de noviembre, cuando el cuerpo de agua estaba totalmente cubierto de macrófitas
flotantes. Este aumento en la concentración de clorofila a coincidió con un aumento en
las densidades algales. El incremento de la biomasa fitoplanctónica en condiciones de
restricción lumínica impuesta por las macrófitas flotantes podría estar dado por efecto del
viento, que permitiría la existencia de áreas libres de macrófitas en algunos momentos
posibilitando el mayor ingreso de luz a la columna de agua, permitiendo así el desarrollo
planctónico. Además la dominancia de algas perifíticas en el plancton durante el muestreo
de noviembre 2006 (Nitzschia spp. y Lemnicola hungarica) podría estar indicando el
desprendimiento de diatomeas del fondo o del epifiton de las macrófitas, lo que podría
estar influenciando las estimaciones de clorofila a. Este hecho ha sido observado por
Carrick et al. (1993) en un lago somero de Florida. Los autores atribuyeron las altas
concentraciones de clorofila fitoplanctónica (media 105 µg L-1) a la resuspensión inducida
por el viento de algas meroplanctónicas del fondo afótico del lago. También Schallenberg
& Burns (2004) observaron aumentos en la producción primaria del fitoplancton por
efecto de la resuspensión de los sedimentos en un lago somero de Nueva Zelanda. En
noviembre 2006 en particular la biomasa fotosintética en el ROL estaba dominada por el
nano y microplancton. Es posible que el picoplancton en el ROL estuviera sujeto a una
fuerte presión de competencia con el bacterioplancton, lo que permitiría el mayor
desarrollo de las fracción de mayor tamaño, al disminuir la competencia con el
picoplancton autotrófico (Drakare et al. 2003).
La falta de diferencias significativas entre las estimaciones de biomasa fotosintética
realizadas por los dos tipos de filtraciones (fraccionamiento y filtros de 0,7 µm de tamaño
de poro) estaría indicando que el aporte de la fracción comprendida entre 0,2 y 0,7 µm a
la biomasa fotosintética total es pequeño en la Laguna Grande y en el ROL. Sin embargo,
no se puede decir lo mismo cuando se analizan los resultados de El Pescado, donde el

205
aporte de la fracción comprendida entre 0,2 y 0,7 µm fue importante especialmente en
invierno, cuando contribuyó con el 50 % de la concentración de clorofila a
fitoplanctónica.
Según Falkowski (1981) los números de asimilación tienden a ser mayores al disminuir el
tamaño celular tal como encontramos nosotros especialmente en la Laguna Grande, y
explicó este hecho por las mayores tasas metabólicas de las células más pequeñas. La
mayor parte de los valores de Pmáx estimados por nosotros para el caso del picoplancton
resultaron elevados cuando se los comparó con los que recopiló Stockner (1988) para el
agua dulce y los estimados para lagos subalpinos de Austria por Callieri et al. (2005) y
Greisberger et al. (2007); por Malinsky-Rushansky et al. (1997) para el lago Kinneret y por
Schweizer & Heusel (1992) para un lago eutrófico de Holanda, entre otros trabajos. Para
el microplancton, Happey-Wood (1993) encontró valores de hasta 51 mg C (mg Chl a h)-1
en un lago de montaña en Inglaterra, aunque en el mencionado trabajo el picoplancton
poseía valores más bajos de Pmáx que los encontrados por nosotros.
En el ROL se registraron en verano los valores de Pmáx más bajos de ambas
fracciones de tamaño, probablemente como resultado del mayor contenido húmico del
ROL. Si bien no se encontraron diferencias significativas, el contenido de ácidos húmicos
del ROL fue siempre superior al de la Laguna Grande. Arvola et al. (1999) encontraron
una tendencia negativa entre el color del agua y la asimilación fotosintética en diez lagos
húmicos de Finlandia. Los autores argumentaron entonces que la iluminación restringida
debida al contenido húmico era la causante de esta tendencia inversa. En la Laguna
Grande, el número de asimilación Pmáx correspondiente a la fracción picoplanctónica
mostró un patrón estacional muy marcado, indicando que la temperatura fue un factor
muy importante para este parámetro en este cuerpo de agua. Kirk (1994) atribuye la
disminución de los valores del parámetro Pmáx a temperaturas más bajas a la velocidad de
las reacciones enzimáticas oscuras de carboxilación, que son más lentas a menor
temperatura. En El Pescado, si bien realizamos dos muestreos estacionales, se advirtió
que en verano el Pmáx fue más alto que en invierno. Es probable que el patrón de
variación de Pmáx en este sitio sea también estacional como en la Laguna Grande.
En el ROL, tanto Pmáx como la eficiencia fotosintética (プ), Pmáx vol y PA fueron
máximas para el picoplancton en noviembre 2006 bajo la cobertura total de macrofitia,
cuando su contribución a la concentración de clorofila a fue menor. Una posible

206
explicación y que requiere para su sustento el desarrollo de estudios posteriores, podría
estar relacionada con la incorporación de 14C por parte de bacterias fotosintéticas
(Prakash et al. 1991) que incorporarían carbono sin poseer clorofila a. Esta hipótesis está
sustentada en el hecho que se ha registrado la presencia de bacterias fotosintéticas
anoxigénicas en el ROL (Izaguirre et al. enviado). Estas bacterias necesitan amonio como
sustrato para la fotosíntesis sin liberar oxígeno como producto de la reacción (Jost et al.
2008) y en este sentido, nosotros estimamos un pico de amonio en el muestreo de
noviembre, que podría estar favoreciendo este tipo de metabolismo bacteriano. Otro
factor a considerar es la dependencia lumínica de la fijación en oscuridad por parte de
estos organismos en los ecosistemas acuáticos ya que Casamayor et al. (2008) observaron
que las bacterias fotosintéticas son capaces de fijar carbono en oscuridad después de ser
expuestas a la luz.
En cuanto al número de asimilación de la fracción > 3 µm en la Laguna Grande, el
valor máximo hallado fue coincidente y similar al valor máximo observado para la
fracción > 0,7 µm en marzo 2007. De manera tal que los valores encontrados en el
presente análisis para el micro y nanoplancton, también se hallan entre los valores
comúnmente observados en la bibliografía para estudios de comunidades naturales,
algunos de los cuales ya fueron citados en el capítulo II. Si bien no se observó para la
fracción > 3 µm un patrón estacional tan marcado como en el caso del picofitoplancton,
los valores más bajos de Pmáx coinciden con lo meses más fríos del año (julio y agosto).
En la laguna El Pescado los valores de Pmáx para el micro y nanoplancton fueron más
elevados que en la Laguna Grande, en particular en verano. Este hecho podría estar
relacionado con el mayor contenido de nutrientes en este cuerpo de agua, dado que el
Pmáx es influenciado por la disponibilidad de los mismos (Falkowski 1981). Este
parámetro no depende directamente de la composición espectral de la radiación (Sakshaug
et al. 1997) y en cambio sí puede depender, entre otros factores de la composición
específica de la comunidad (Côté & Platt 1983). Considerando que en la Laguna Grande
dominaron las diatomeas y en El Pescado las cianobacterias filamentosas acompañadas de
crisofíceas, la diferente composición específica podría entonces explicar también los
mayores Pmáx en El Pescado, además de la mayor penetración de irradiancia que se
estimó en esta laguna.
En el ROL los valores del número de asimilación del picoplancton, a excepción
del muestreo de marzo 2006, estuvieron comprendidos entre 0,9 y 3,6 µg C (µg Chl a h)-1

207
y no se observó un cambio que pudiera atribuirse a la cobertura de macrófitas. El mayor
valor observado, que fue alto (26 µg C (µg Chl a h)-1 en marzo 2006), estuvo ligeramente
por encima del máximo teórico de 25 µg C (µg Chl a h)-1 señalado por Falkowski (1981).
En esta fecha, las altas temperaturas y mayores irradiancias debido a la falta de cobertura
vegetal además de las altas concentraciones de P-disuelto pudieron posibilitar mayores
tasas de asimilación fotosintética. Además, la dominancia de clorofíceas pudo también
posibilitar una mayor asimilación fotosintética en el ROL.
En la Laguna Grande las correlaciones negativas halladas entre los valores de
Pmáx y Pmáx vol de ambas fracciones de tamaño y la eficiencia fotosintética
picoplanctónica (プ) con el NID o N-nitrato + nitrito probablemente guarden una estrecha
relación con la variación estacional del NID como consecuencia de su incorporación a la
biomasa. Estas variables (Pmáx, Pmáx vol y プ del picoplancton) resultaron en general
mayores en los meses de verano, cuando había muy poco NID disponible debido muy
probablemente a su consumo.
Los valores de producción máxima por unidad de volumen (Pmáx vol) registrados
por nosotros para ambas fracciones de tamaño en los cuerpos de agua estudiados fueron
elevados y estuvieron dentro del orden de los registrados por Harding (1997) para el lago
sudafricano hipereutrófico Zeekoevlei. Este lago estaba, cuando fue estudiado, sujeto a
una mezcla permanente por acción del viento y la Pmáx vol estuvo relacionada con la
temperatura, análogamente a lo ocurrido en la Laguna Grande para el fitoplancton de las
dos fracciones de tamaño (> 3 µm y picoplancton). No se detectaron diferencias
significativas entre Pmáx vol de la laguna y el ROL debido a la gran variabilidad estacional
en las mediciones. En el caso del picoplancton, las estimaciones realizadas por nosotros
fueron en general mayores a los obtenidos por Rahaingomanana et al. (2002) para sistemas
húmicos del valle de inundación del río Doce en Brasil en verano, por Drakare et al.
(2002, 2003) lagos húmicos de Suecia y por Schweizer & Heusel (1992) para un lago
eutrófico de Holanda.
Las correlaciones positivas encontrada entre el Kd y g440 con la Pmáx vol de las
dos fracciones de tamaño en la Laguna Grande y el ROL vol guardarían relación con la
influencia del fitoplancton sobre las propiedades ópticas del agua (Kirk 1994). En este
mismo sentido, la profundidad de la capa eufótica disminuyó a medida que aumentó la
Pmáx vol en los dos ambientes como resultado de la disminución de la penetración de la
luz por la presencia de altas concentraciones de fitoplancton. De forma similar a lo que

208
ocurrió con la concentración de clorofila a, los valores de Pmáx vol aumentaron debajo de
la cobertura de macrófitas flotantes en el ROL, en particular en noviembre 2006 y enero
2007. Si bien nosotros no encontramos una relación estadística entre esta variable y la
concentración de clorofila a en el ROL, Krause-Jensen & Sand-Jensen (1998) encontraron
una relación positiva entre la Pmáx vol y la biomasa fotosintética a partir de datos de la
literatura, de modo que muy probablemente la estadística no esté reflejando en este caso
lo que ocurre en la naturaleza.
En la Laguna Grande, concentraciones de clorofila a de nano y microplancton
mayores a aproximadamente 250 µg L-1 provocaron la disminución de la Pmáx vol.
Concentraciones tan altas de clorofila se observaron en el caso de floraciones de
cianobacterias. En estas condiciones la presencia de los propios organismos provocó aún
más la atenuación de la radiación incidente y la disminución de las tasas de crecimiento
algales. Este hecho ya ha sido observado por otros autores en otro tipo de sistemas (ej.
Shigesada & Okubo 1981). Esta observación queda además sustentada por la correlación
negativa encontrada entre la profundidad correspondiente al 1 % de la radiación incidente
y la concentración de clorofila del nano y microplancton, que estaría indicando el
adelgazamiento de la capa eufótica en presencia de una alta biomasa de organismos.
La mayor eficiencia fotosintética (プ) del picoplancton tanto en la Laguna Grande
como en El Pescado podría estar relacionado con el menor efecto paquete de los
organismos de menor tamaño y su uso más eficiente de la irradiancia (Helbling et al. 2001
a, Raven 1998). En el ROL el patrón temporal fue más irregular, ya que en tres
oportunidades la fracción de organismos > 3 µm alcanzó una eficiencia fotosintética
mayor, de modo que no se detectaron diferencias en el parámetro プ entre fracciones de
tamaño. Los valores obtenidos fueron muy variables y en general se hallan entre los
reportados en la bibliografía tanto para el fitoplancton > 3 µm como para el picoplancton
(Callieri et al. 2005, Reynolds 1984). Los altos números de asimilación y eficiencia
fotosintética que se registraron en algunas oportunidades para el picoplancton podrían
explicarse por las bajas irradiancias que se encuentran en los cuerpos de agua del humedal
debido a la presencia de ácidos húmicos y de macrófitas flotantes en el ROL. En este
sentido, Callieri et al. (2007) encontraron que la eficiencia fotosintética del picoplancton
era mayor que la de los organismos de mayor tamaño a menores irradiancias y contenido
de nutrientes.

209
Si bien Ik puede utilizarse como un indicador del estado de fotoaclimatación del
fitoplancton (Henley 1993), la irradiancia incidente en el campo fluctúa constantemente y
la aclimatación toma tiempo. De este modo, dado que el parámetro Ik varía
constantemente, en principio nunca se correspondería totalmente con la irradiancia
instantánea ya que depende de la historia lumínica previa (Sakshaug et al. 1997, Prézelin
1991). En este sentido, la única tendencia temporal clara que se observó en los tres
cuerpos de agua estudiados fue la disminución de los valores de Ik de ambas fracciones de
tamaño en invierno (en particular el muestreo de julio 2006 en Laguna Grande y ROL)
probablemente como consecuencia de las menores irradiancias incidentes acumuladas
durante esa estación del año. Según Sakshaug et al. (1997) esta variabilidad en los valores
de Ik sería particularmente importante para el fitoplancton de cuerpos de agua con una
fuerte mezcla. Tal es el caso, por ejemplo de la Laguna Grande, donde los valores de Ik
resultaron muy variables, en particular para los organismos más pequeños (coeficiente de
variación de los valores de Ik para el picoplancton: 66 %; mientras que para la fracción >
3 µm: 48 %). Esta mayor variabilidad de los valores de Ik picoplanctónicos estaría
relacionada con una mayor capacidad de aclimatación ya que en general las células más
pequeñas se aclimatan más rápido que las más grandes a nuevas condiciones de irradiancia
(Helbling et al. 2001). Por otro lado si bien no se observó una disminución conspicua de
los valores del parámetro Ik como consecuencia de la cobertura total de macrófitas
flotantes en el ROL, sí se observó que ambas fracciones de tamaño del fitoplancton se
encontraron limitadas lumínicamente a partir de agosto. Es destacable entonces que en el
ROL, aún bajo condiciones de limitación lumínica la Pmáx vol de ambas fracciones de
tamaño haya sido mayor que en otros muestreos, lo que estaría indicando la alta eficiencia
que tienen los organismos fotosintéticos de este sitio del humedal.
La producción primaria integrada en la columna de agua (PA) fue mayor en la Laguna
Grande para las dos fracciones de tamaño, cuando se comparan los tres sistemas. A su
vez, el micro y nanoplancton presentó una tendencia a ser la fracción más productiva por
unidad de área en esta laguna. En lagos someros susceptibles a la resuspensión por acción
del viento, la contribución del picoplancton a la biomasa total es pequeña, y estaría
afectada por la contribución alta y variable del microplancton (Bell & Kalff 2001). La
dinámica de las dos fracciones de tamaño estuvo fuertemente marcada por la temperatura,
si bien en el caso del micro y nanoplacton la variación conjunta de la Imedia y el fósforo

210
fueron las variables que mejor explicaron su dinámica. Para el picoplancton la temperatura
resultó ser la única variable que explicó su dinámica anual. La estacionalidad del
picoplancton, si bien en abundancia y no en producción, ha sido reportada para el lago
Baikal por Belykh et al. (2006). También se encontró que la temperatura era un factor
ambiental muy importante en la dinámica estacional del crecimiento picoplanctónico en el
lago Kinneret (Malinsky-Rushansky et al. 2002). Por su parte, Mózes et al. (2006)
observaron que en sectores mesotróficos y eutróficos del lago Balaton (Hungría) la
dinámica del picoplancton era estacional.
Los valores diarios totales de PA registrados para los tres cuerpos de agua fueron
bajos cuando se los compara con el lago hipereutrófico de Sudáfrica Zeekoevlei (Harding
1997) y con un estanque mesotrófico de Bangladesh (Khondker & Kabir 1995). Si bien
los valores de Pmáx vol registrados en este estudio fueron altos cuando se los comparó
con los encontrados en la bibliografía, los valores de producción integrada en la columna
de agua fueron bajos, y más aún los de producción anual para la Laguna Grande. Esto
tiene que ver con que en los sistemas de características húmicas, la disminución del
volumen productivo genera que la relación entre la producción por unidad de volumen
(correspondiente a la obtenida a la profundidad donde ocurre la mayor producción) y la
producción areal (integrada en la columna de agua) es mayor a la que cabría esperar en un
cuerpo de agua claro con una mayor capa eufótica (Arvola et al. 1999).
El aporte del picoplancton a la PA diaria fue generalmente bajo para los cuerpos de
agua estudiados considerando que Vörös et al. (1991) observaron que en el hipereutrófico
lago Balaton de Hungría, el aporte del picoplancton a la producción fitoplanctónica era
del 57 % en primavera. Por otro lado, Nagata et al. (1994) encontraron un aporte mayor,
estimando que el 80 % de la producción primaria del lago Baikal era debida al
picoplancton autotrófico. La excepción que encontramos nosotros fue el alto aporte del
picoplancton del ROL a la PA total en invierno que pudo deberse en parte a la mayor
biodisponibilidad de nitrógeno. En este sentido, Vörös et al. (1991) observaron la
disminución del crecimiento del picoplancton por efecto de la carencia de nitrógeno. Por
otro lado, la mayor atenuación lumínica del ROL en invierno también pudo favorecer al
picoplancton, dado que se ha citado en la bibliografía el aumento del aporte del
picoplancton a la producción primaria con la profundidad (Callieri et al. 2007, Malinsky-
Rushansky 1997, Vörös et al. 1991), debido al uso eficiente de bajas intensidades lumínicas
(Fogg 1986, Stockner & Antia 1986). Sin embargo, considerando las bajas tasas anuales de

211
PA fitoplanctónica tanto en la Laguna Grande como en el ROL, sería importante en
estudios posteriores analizar los patrones de productividad considerando el aporte de la
fracción mayor a 0,2 µm, ya que se produjeron en ambos sitios subestimaciones del 14 y
33 % (laguna y ROL respectivamente) al emplear filtros de 0,7 µm de tamaño de poro.
En teoría, las agua húmicas coloreadas proveen un ambiente adecuado para el
picoplancton autotrófico. En estos ambientes en general los nutrientes presentan baja
biodisponibilidad, lo que podría favorecer al picoplancton (Callieri 2008). Por su parte,
Drakare et al. (2003) observaron que la mayor abundancia de picoplancton ocurre a
concentraciones intermedias de carbono orgánico disuelto (COD). En los cuerpos de
agua con altas concentraciones de COD, como los de la RNO, una interacción frecuente
es la competencia del picofitoplancton con el bacterioplancton por nutrientes, en especial
por fósforo (Callieri 2008, Drakare et al. 2002). De esta manera, sería de esperar que la
competencia del picoplancton con las bacterias por los nutrientes sea mayor en el ROL,
restringiendo así la productividad primaria de esta fracción. Sería entonces por este
motivo que, cuando hay más nitrógeno como en el invierno, el picoplancton aporta más a
la PA total que el micro y nanoplancton en este sitio. En este sentido, Drakare et al. (2002)
observaron que el picofitoplancton pudo desarrollarse en un lago húmico de Suecia luego
que disminuyó la abundancia de bacterioplancton. Los autores concluyeron que la ventaja
del menor tamaño celular del picofitoplancton no es suficiente como para garantizar una
competencia exitosa con el bacterioplancton, aunque sí sería una ventaja en la
competencia con el fitoplancton de mayor tamaño.
En este estudio, observamos una tendencia general a la disminución de la
contribución del picoplancton autotrófico a la productividad con el incremento del estado
trófico. Agawin et al. (2000) y Bell & Kalff (2001) encontraron que la contribución relativa
del picofitoplancton a la producción total del fitoplancton disminuye en aguas muy
productivas o ricas en nutrientes debido al incremento en producción del fitoplancton de
mayor tamaño. Por su parte, Jasser (1997) y Callieri et al. (2007) obtuvieron resultados
similares.
Finalmente, en relación a las hipótesis planteadas al inicio de este capítulo hemos
observado una tendencia al menor aporte a la producción del picoplancton autotrófico en
el estudio anual que realizamos en la Laguna Grande, de elevado estado trófico. Además,
comprobamos el patrón estacional en la dinámica anual de la producción primaria de las

212
dos fracciones de tamaño fitoplanctónicas en la Laguna Grande, e inferimos un
comportamiento similar en la laguna El Pescado. Para la Laguna Grande, comprobamos
que los parámetros fotosintéticos プ y Pmáx fueron mayores para el picoplancton
autotrófico aunque no observamos una tendencia clara en los valores de Ik en cuanto a las
fracciones de tamaño estudiadas. Observamos que la subestimación efectuada al analizar
la fracción de tamaño > 0,7 µm puede ser importante al tener en cuenta que estos cuerpos
de agua son poco productivos cuando se analiza la producción integrada en la columna de
agua, si bien por unidad de volumen pueden mostrar valores elevados de producción. Por
último, el ROL presentó menores valores de producción primaria integrada, muy
probablemente debido a las mayores restricciones lumínicas encontradas en este cuerpo
de agua. Sin embargo, en condiciones de limitación lumínica impuestas por una cobertura
completa de plantas flotantes, el desarrollo del fitoplancton no se vio limitado, indicando
una gran adaptación a las condiciones del lugar.

213
CAPÍTULO V
EFECTO DE LA ATENUACIÓN LUMÍNICA SOBRE LA PRODUCCIÓN
PRIMARIA DEL FITOPLANCTON:
UN ESTUDIO EN MESOCOSMOS

214
INTRODUCCIÓN
Las macrófitas acuáticas conforman una comunidad conspicua en la mayoría de los
humedales de agua dulce de latitudes templadas a tropicales y su presencia puede
influenciar profundamente el funcionamiento de los lagos someros (por ejemplo moderan
la temperatura del agua, alteran la química del agua, restringen la acción del viento, entre
otros efectos) (van der Valk 2006). Además, las macrófitas proveen refugio a los
pequeños animales contra la predación por parte de otros más grandes, alteran la
dinámica de los nutrientes del sistema y en particular las macrófitas sumergidas previenen
la resuspensión de los sedimentos. Los cuerpos de agua vegetados generalmente poseen
comunidades más ricas de invertebrados y peces y atraen mayor número de aves acuáticas
que los lagos no vegetados (Scheffer 1998). El papel funcional y estructural de las
macrófitas sumergidas en cuerpos de agua someros ha sido ampliamente discutido
habiéndose escrito innumerables trabajos de investigación así como también libros al
respecto (Scheffer 1998, Jeppesen et al. 1998).
Scheffer et al. (1993) propusieron una hipótesis en la que se contempla la existencia
de estados alternativos en cuerpos de agua someros: un estado de aguas claras, dominado
por macrófitas sumergidas, y otro estado turbio dominado por fitoplancton. Más tarde
Scheffer et al. (2003) incluyeron en el modelo un tercer estado de equilibrio dominado por
plantas flotantes libres, que revisan en Scheffer & van Nes (2007). Los autores sostienen
que un estado alternativo dominado por plantas flotantes sería plausible de ocurrir en
condiciones de no limitación por nutrientes. Si estos recursos son limitantes, existiría
dominancia del fitoplancton ya que las algas son más eficientes que las macrófitas en la
toma de nutrientes cuando éstos se encuentran en bajas concentraciones. Dada la
sensibilidad de las macrófitas a los cambios en las concentraciones de nutrientes y a que
su presencia o ausencia refleja el estado nutritivo de los cuerpos de agua, pueden ser
utilizadas como indicadores de condiciones ambientales (Melzer 1999).
En lagos templados someros con niveles altos de nutrientes, el fitoplancton es más
eficiente en su captación que las macrófitas sumergidas. En estas condiciones, el
fitoplancton reemplazaría a las macrófitas sumergidas y luego el fitoplancton sería
reemplazado por las macrófitas flotantes, que son mejores competidoras por la luz
(Scheffer & van Nes 2007). Por otro lado, Scheffer et al. (2003) sostienen que las plantas
flotantes no son dominantes en grandes lagos templados debido a que su distribución

215
generalmente está restringida a sitios de menores dimensiones con áreas bien protegidas
de la acción del viento. Estas hipótesis tienen implicancias fundamentales en el manejo de
los lagos someros y surgieron ante la necesidad de encontrar una manera de restaurarlos
como consecuencia de la eutroficación (Scheffer 1998). Su rango actual de aplicación es
más amplio ya que provee un marco teórico básico para el estudio de la dinámica de estos
sistemas.
Las plantas flotantes libres afectan la dinámica del cuerpo de agua ya que provocan
la disminución de la fotosíntesis por parte de los microorganismos acuáticos puesto que
una cobertura más o menos conspicua disminuye la intensidad lumínica que penetra en el
cuerpo de agua además de provocar condiciones de anoxia en situaciones de cobertura
completa. También reducen el intercambio gaseoso entre la atmósfera y la columna de
agua (Nahlik & Mitsch 2006). Por otro lado las plantas compiten con el fitoplancton por
los nutrientes ya que como se mencionó, las macrófitas flotantes los toman de la columna
de agua (Scheffer et al. 2003). Además, las plantas aportan material vegetal al agua que, al
descomponerse, se convierte en una fuente importante de ácidos húmicos. Los ácidos
húmicos influyen sobre la disponibilidad de los nutrientes (en particular del fósforo
debido a la formación de complejos con el hierro y las sustancias húmicas, Jackson &
Hecky (1980)) y sobre el clima lumínico del sistema puesto que afectan la penetración de
la radiación en el cuerpo de agua entre otros factores (Jones 1992, Kirk 1994).
Las plantas flotantes de gran porte son muy sensibles a las bajas temperaturas,
tolerando mejor temperaturas altas a medianas, de forma tal que a nivel mundial se
encuentran principalmente restringidas a latitudes tropicales y subtropicales (Sculthorpe
1967). En este contexto, constituyen un importante indicador en estudios de
calentamiento global ya que el aumento de temperatura podría promover la expansión
geográfica de las plantas flotantes libres a zonas donde actualmente no se encuentran
(Meerhoff & Mazzeo 2004).
Existen numerosos estudios acerca de la dinámica de la biomasa de las plantas
flotantes, en particular de Eichhornia crassipes y su fauna asociada (Neiff & Poi de Neiff
1984); algunos de ellos fueron realizados en lagunas de la llanura de inundación del río
Paraná (Sabattini & Lallana 2007, Poi de Neiff & Neiff 2006, Neiff et al. 2001, Poi de
Neiff & Carignan 1997). Por otro lado, Carignan et al. (1994) estudiaron la limitación de
Eichhornia crassipes por nitrógeno en seis lagunas del valle aluvial del río Paraná.
Recientemente, Henry-Silva et al. (2008) estudiaron el efecto de distintas concentraciones

216
de nutrientes sobre el crecimiento de tres macrófitas flotantes de diferente porte en Brasil
(Eichhornia crassipes, Pistia stratiotes y Salvinia molesta). Por su parte, Meerhoff et al. (2003)
analizaron experimentalmente el rol de las macrófitas sumergidas y flotantes sobre las
características físico-químicas del agua, la estructura del zooplancton y los peces en un
lago somero de Uruguay. En este estudio los autores observaron que la presencia de
plantas flotantes libres afectaba a la comunidad generando un efecto en cascada sobre el
fitoplancton.
Ciertos cuerpos de agua de latitudes tropicales y subtropicales poseen
características morfométricas y limnológicas que posibilitan en general la existencia de
plantas flotantes la mayor parte del año, y en algunos casos su cobertura puede llegar a
tapizar todo el espejo de agua (Neiff & Poi de Neiff 1978). El efecto de las macrófitas
flotantes sobre la estructura de la comunidad del fitoplancton ha sido analizado por
diferentes autores. En un estudio realizado en una laguna del valle de inundación del río
Paraná Medio, Zalocar de Domitrovic (1993) observó que la presencia de Eichhornia
crassipes constituye un importante factor estructurador de la comunidad fitoplanctónica.
Bicudo et al. (2007) estudiaron los efectos de la remoción de Eichhornia crassipes sobre las
variables limnológicas y del fitoplancton en una represa en Brasil. Los autores
documentaron que la remoción del jacinto de agua provocó un aceleramiento del proceso
de eutrofización, llevando al sistema hacia un estado de aguas turbias con dominancia de
fitoplancton. Sinistro et al. (2006) comprobaron experimentalmente en microcosmos que
la atenuación lumínica provocada por la cobertura de macrófitas flotantes provocaba
cambios en la estructura de la comunidad microbiana en la Laguna Grande de la Reserva
Otamendi (RNO). También, de Tezanos Pinto et al. (2007) demostraron
experimentalmente en mesocosmos instalados en la misma laguna, que la atenuación
lumínica provocada por una cobertura completa de macrófitas flotantes era la fuerza
modeladora de la estructura del fitoplancton en tanto que en escenarios de aguas abiertas,
lo era la limitación por nutrientes.
La cobertura de macrófitas flotantes de la Laguna Grande de la RNO estuvo
restringida principalmente a la zona litoral del cuerpo de agua durante el período de
estudio. Sin embargo, en años anteriores la laguna se encontraba cubierta en gran parte de
su superficie por plantas de mediano y pequeño porte (de Tezanos Pinto et al. 2007,
Izaguirre et al. 2004, O´Farrell et al. 2003). Entre las macrófitas flotantes representativas de
la Laguna Grande, se pueden mencionar a Ricciocarpus natans, Azolla filiculoides, lemnáceas

217
de pequeño porte y en menor proporción Pistia stratiotes. La primer planta flotante cuya
distribución se vio afectada fue la flotante de mayor porte, Pistia stratiotes (obs. pers.),
probablemente debido a que su tasa de crecimiento decrece a bajos tenores de nutrientes,
tal como lo demostraron Henry-Silva et al. (2008).
El objetivo del presente estudio fue analizar el efecto de la atenuación lumínica provocada
por una cobertura completa de plantas flotantes en la producción primaria del
fitoplancton en un sistema húmico. Se realizó un experimento manipulativo en
mesocosmos en la Laguna Grande de la RNO en donde se estudiaron la estructura,
producción primaria y la eficiencia fotosintética del fitoplancton además de otras variables
físicas y químicas del agua. Se comparó un escenario libre de macrófitas con otro que
simulaba una cobertura completa y permanente de plantas. La experiencia manipulativa
tuvo una duración de un mes, con una frecuencia de muestreo de 10 días.
Hipótesis planteadas
i) la atenuación lumínica provocada por una cobertura completa de macrófitas genera la
disminución de la producción primaria del fitoplancton en un sistema húmico;
ii) la menor intensidad de luz generada por la cobertura de macrófitas se traduce en un
aumento de la eficiencia fotosintética del fitoplancton probablemente como consecuencia
de la aclimatación de la comunidad a menores irradiancias.
MATERIALES Y MÉTODOS
Se realizó un experimento en la Laguna Grande de la Reserva Natural Otamendi entre los
días 8 de marzo de 2006 y 7 de abril de 2006. Se utilizaron mesocosmos que consistieron
en clausuras de policarbonato (90 cm de diámetro, 180 L, profundidad promedio 0,5 m),
que fueron enterrados en el sedimento en un área libre de macrófitas flotantes de la zona
litoral (Fig. 1). Durante la instalación de cada mesocosmos se les colocó una red en su
base para evitar el ingreso no controlado de peces. La red fue removida justo antes de
afirmar cada mesocosmos en el sedimento. Los mesocosmos fueron asegurados al
sustrato por medio de estacas. Luego, con el objetivo de evitar la hiperpredación del

218
fitoplancton por parte del zooplancton, se agregaron en cada mesocosmos 10 peces
autóctonos de tamaño similar (11 ind m-2, Jenynsia multidentata). Empleamos una densidad
de peces similar a la empleada por Iglesias et al. (2008) en una experiencia en mesocosmos
en Uruguay (9 ind m-2) ya que no se disponían de datos de biomasa o densidad de peces
para la Laguna Grande. Por otro lado, contamos con el asesoramiento del Dr. Erik
Jeppesen (National Environmental Research Institute, Department of Freshwater
Biology, Silkeborg, Dinamarca) quien nos sugirió esa densidad luego de analizar in situ los
dispositivos.
Se aplicaron 2 tratamientos (por triplicado) asignados al azar a los mesocosmos.
En tres de ellos se simuló el efecto de sombreado producido por la presencia de
macrófitas que se logró mediante la utilización de mallas de nylon tipo media sombra
(tratamiento cubierto) con 2 % de transmitancia de la luz incidente. El escenario libre de
macrófitas flotantes (tratamiento libre) se obtuvo dejando a los mesocosmos libres de
toda cobertura (transmitancia del 100 % de la luz incidente). El tratamiento que
representaba la cobertura total de macrófitas flotantes simulaba una intensidad lumínica
similar a la que nosotros medimos in situ por debajo de una carpeta completa de
macrófitas flotantes en la zona litoral de laguna. La utilización de mallas de atenuación
neutra en lugar de plantas flotantes verdaderas permite analizar el efecto de sombreado
sin considerar otros efectos que puedan provocar las plantas flotantes. En estudios
previos realizados en la misma laguna se observó que las macrófitas flotantes de pequeño
y mediano porte afectaban el crecimiento del fitoplancton debido a la limitación lumínica
que generaban, más que por efecto de competencia por nutrientes o secreción de
sustancias alelopáticas (de Tezanos Pinto et al. 2007).

219
Fig. 1. Mesocosmos instalados en un área libre de macrófitas flotantes de la zona litoral de
la Laguna Grande.
La experiencia comenzó a los 10 días de instalados los mesocosmos, tiempo que se
estimó necesario para la homogeneización de todas las unidades experimentales. Al
tiempo cero, se cubrieron 3 mesocosmos con la mencionada malla dejándose libres los 3
restantes. A los 10 días del tiempo cero, se realizó la primera estimación de productividad
primaria para luego realizar dos más, alrededor del mediodía, con una frecuencia de 10
días entre ellas.
En cada oportunidad y cada 5 cm de profundidad se realizaron mediciones de los
siguientes parámetros en los mecosmos: temperatura, pH, conductividad, oxígeno
disuelto y radiación fotosintéticamente activa (RFA) empleando los mismos equipos
señalados en el capítulo II de esta tesis (páginas 72-73). Con el objeto de analizar un
posible efecto dado por la clausura, los valores de temperatura y profundidad en los
mesocosmos fueron comparados con los del agua libre de la laguna. Con los datos de
irradiancia en función de la profundidad se calcularon el coeficiente de atenuación vertical
(Kd), la irradiancia media en la columna de agua (Imedia) (Helbling et al. 1994, Neale et al.
1991) y las profundidades ópticas (⦆) de acuerdo a Kirk (1994).
Se tomaron muestras integradas de toda la columna de agua con un muestreador
vertical de PVC que consistió en un tubo de 0,8 m de longitud y 0,05 m de diámetro, con
cierre mecánico (Fig. 2). Las muestras de agua integradas se utilizaron para estimar el
coeficiente de absorción del agua a 440 nm (g440), la concentración de nutrientes (N-
nitrato + nitrito, N-amonio, P-disuelto, N y P total) y clorofila a. Las filtraciones se

220
llevaron a cabo a través de filtros de fibra de vidrio Millipore® de 0,7 µm de tamaño de
poro. Las determinaciones se efectuaron empleando la misma metodología descrita en la
página 74 de esta tesis.
Se trabajó con datos de composición relativa de clases algales de la fracciones de
tamaño nano y microplanctónicas y densidad total del fitoplancton. Las muestras cuali y
cuantitativas fueron tomadas integrando en profundidad y fijando con solución de lugol
acético a una concentración final del 1 %. Los análisis taxonómicos se realizaron
siguiendo la bibliografía ficológica usual en tanto que los recuentos fueron hechos bajo
microscopio invertido de acuerdo al método de Utermöhl (1958) con un 20% de error
(Venrick 1978). Estos análisis fueron realizados y cedidos por la Lic. Paula de Tezanos
Pinto y forman parte de su trabajo de tesis doctoral (de Tezanos Pinto 2008).
Fig. 2. Dispositivo empleado para tomar las muestras integradas de agua en los
mesocosmos (flecha).
Las estimaciones de producción primaria se llevaron a cabo a partir de muestras
integradas en la columna de agua tomadas con el muestreador vertical. Se empleó el
mismo diseño, dispositivos y la misma metodología de la técnica de asimilación de 14C
descritos en los capítulos anteriores. Se expusieron las muestras de fitoplancton de cada
mesocosmos simultáneamente a 2 intensidades lumínicas: 100 % (luz natural) y 10 %

221
(logrado con malla de atenuación neutra) y un par de tubos oscuros por cada
mesocosmos. Los tubos (por duplicado para cada muestra) se incubaron 2 hs alrededor
del mediodía, luego de la adición de 1 ┃Ci NaH14CO3 (Perkin Elmer Life Sciences, Inc.,
USA).
Con los datos de producción primaria para cada intensidad lumínica se estimó la
producción integrada en la columna de agua (PA) como el área bajo la curva de
producción en función de la profundidad. Esta estimación la efectuamos considerando
que el perfil de agua tenía pocos centímetros de profundidad en cada mesocosmos y que
la muestra que utilizamos para incubar a cada atenuación lumínica era representativa de la
columna de agua. Luego, cada valor de PA se estandarizó con la concentración de clorofila
a y con la Imedia de cada mesocosmos. Los cocientes de PA/clorofila a y PA/I media se
consideraron como estimadores de eficiencia fotosintética de cada comunidad.
Se estimó el parámetro Ik para cada mesocosmos con el objeto de inferir si existía
una posible limitación lumínica de la comunidad en cada mesocosmos. Para ello se
ajustaron los valores de productividad correspondientes a cada irradiancia a la ecuación de
Eilers & Peeters (1988). Cabe aclarar que los valores de Ik así calculados representan
aproximaciones al verdadero valor de Ik puesto que para su obtención nos basamos en el
ajuste de tres puntos de la curva P-I. Por este motivo los valores de Ik así obtenidos sólo
se utilizaron para sugerir posibles patrones.
Análisis de datos
Se efectuaron ANOVA de medidas repetidas para detectar diferencias significativas de
variables entre tratamientos y en el tiempo. Las variables que no cumplían los supuestos
del método fueron transformadas mediante Log10. Cuando las interacciones resultaron
significativas se realizaron contrastes de Tukey (Sokal & Rohlf 1979). También se
efectuaron correlaciones no paramétricas (Spearman) entre las variables ambientales y de
producción (Zar 1996).
RESULTADOS
En la Tabla 1 se resumen los rangos y promedios de las principales variables ambientales
registradas para los dos tratamientos durante el estudio. La profundidad de la columna de
agua fue poco variable entre tratamientos, sin mostrar diferencias significativas, mientras

222
que disminuyó con el tiempo (Tabla 2). La disminución del nivel hidrométrico también se
observó en las aguas libres de la laguna, con una baja de 11 cm entre los días 10 y 30. La
temperatura del agua en los mesocosmos mostró una variación máxima de 4,2 °C entre el
día 10 y 30, en tanto que en la laguna el rango de variación fue de 3,5 °C y no mostró
diferencias significativas con la temperatura de las clausuras. Por otro lado, tanto en los
tratamientos como en la Laguna Grande, la temperatura aumentó con el tiempo (Fig. 3 a,
Tabla 2). De acuerdo a estos resultados, descartamos un posible efecto en la temperatura
del agua provocado por la clausura. Por otro lado, la concentración de oxígeno disuelto
fue menor en el tratamiento cubierto, donde también se observó una tendencia a la
disminución de su concentración conforme avanzaba el tiempo, aunque fue no
significativa (Fig. 3 b, Tabla 2). La concentración de oxígeno disuelto se correlacionó
positivamente con el pH, la Imedia, la concentración de N total y la densidad algal, y de
forma negativa con las fracciones de fósforo disuelto y total (Tabla 3). El pH del agua fue
en general más alto en el tratamiento libre (Tabla 1), aunque esta observación no quedó
sustentada estadísticamente. El incremento de la conductividad se correlacionó
significativamente con la disminución de la profundidad (Tablas 1, 2 y 3).

223
tratamiento cubierto libre profundidad (m) 0,4-0,6 0,4-0,6 0,5±0,06 0,5±0,05 temperatura (°C) 17,3-20,7 17,5-20,8 19±1,3 19±1,4 O2 disuelto (mg L-1) 0,1-2,2 1,1-6,5 0,9±0,7 3,7±1,7 pH 7,8-8 7,7-8,5 7,9±0,1 7,7±0,2 conductividad (µS cm-1) 1090-1420 1200-1670 1260±123 1410±150 Imedia (µmol fotones m-2s-1) 7-15 130-325 10±2,5 250±71 g440 (m-1) 11-20 9-26 17±3,2 17±6,3 Kd (m-1) 2-7,1 3,6-10,2 5±1,5 7±2 P-disuelto (mg L-1) 0,2-0,3 1,2-2,6 2,4±0,9 1,8±0,5 P total (mg L-1) 2,1-3,4 1,4-2,8 2,7±0,5 2,1±0,5 N-nitrato + nitrito (mg L-1) 0-0,05 0-0,08 0,03±0,02 0,04±0,03 N-amonio (mg L-1) 0,07-0,3 0-0,03 0,2±0,08 0,005±0,01 N total (mg L-1) 0,3-1,2 0,2-1,3 0,7±0,3 0,7±0,4
Tabla 1. Rango de variación (fila superior) y promedio ± 1 DS (fila inferior, en negrita) de
las variables ambientales registradas para los dos tratamientos.

224
tratamiento tiempo interacción
profundidad ns F2,8 = 127
p= 0,000001ns
temperatura ns F2,8 = 72
p= 0,000008ns
O2 disuelto F1,4 = 12 p= 0,03
ns ns
conductividad ns F2,8 = 383
p= 0 ns
Imedia F1,4 = 409
p= 0,00004ns ns
g440 ns F2,8 = 20,7 p= 0,0007
ns
P-disuelto F1,4 = 9,2 p = 0,04
F2,8 = 5,4 p= 0,03
ns
P total ns F2,8 = 20
p= 0,0008 ns
N-nitrato + nitrito ns F2,8 = 55
p= 0,00002 ns
N-amonio F1,4 = 197 p = 0,0001
F2,8 = 18 p= 0,001
F2,8 = 4,8 p= 0,04
N total ns F2,8 = 34
p= 0,0001 ns
densidad total fitoplanctonF1,4 = 27 p = 0,006
F2,8 = 6,4 p= 0,02
F2,8 = 26 p= 0,0003
clorofila a F1,4 = 115 p = 0,0004
ns ns
PA/Imedia F1,4 = 30,5 p = 0,005
ns ns
Tabla 2. Resultados de los ANOVA de medidas repetidas efectuados para las variables
estudiadas a lo largo de período de estudio. ns= no significativo.
225
Fig. 3. Variación promedio de: a) temperatura del agua y b) oxígeno disuelto para cada
tratamiento a lo largo del experimento. Barras = +1DS.
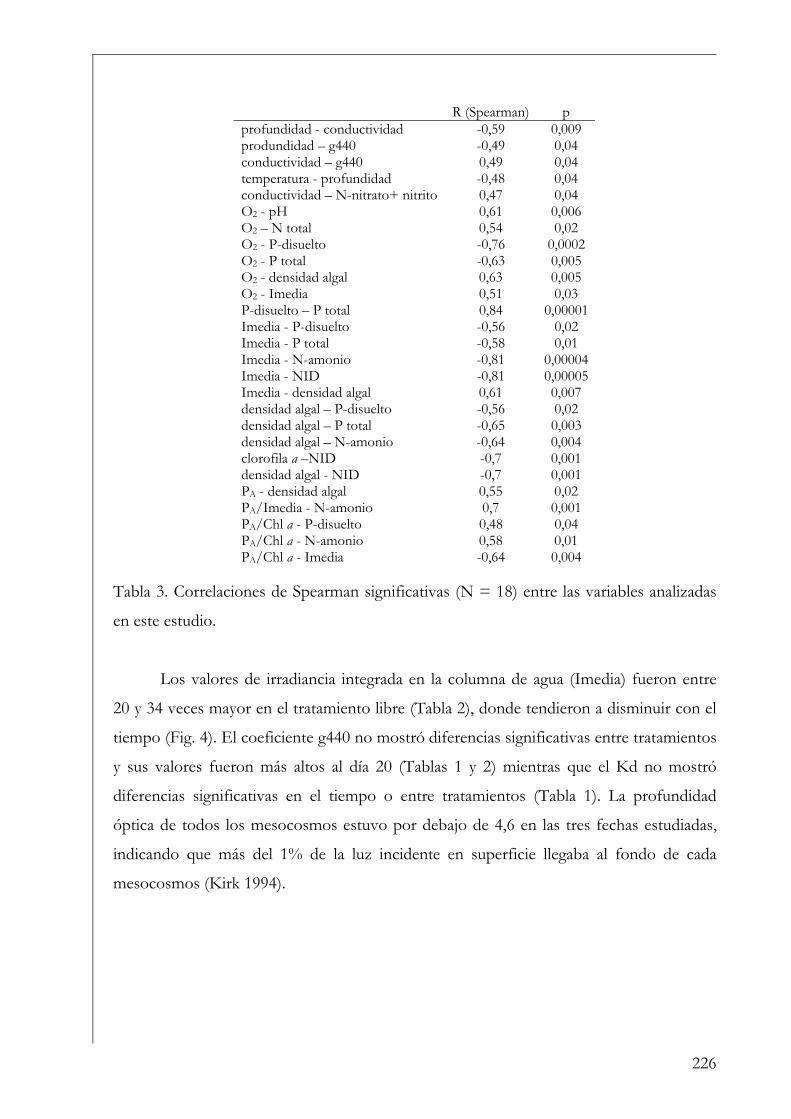
226
R (Spearman) p profundidad - conductividad -0,59 0,009 produndidad – g440 -0,49 0,04 conductividad – g440 0,49 0,04 temperatura - profundidad -0,48 0,04 conductividad – N-nitrato+ nitrito 0,47 0,04 O2 - pH 0,61 0,006 O2 – N total 0,54 0,02 O2 - P-disuelto -0,76 0,0002 O2 - P total -0,63 0,005 O2 - densidad algal 0,63 0,005 O2 - Imedia 0,51 0,03 P-disuelto – P total 0,84 0,00001 Imedia - P-disuelto -0,56 0,02 Imedia - P total -0,58 0,01 Imedia - N-amonio -0,81 0,00004 Imedia - NID -0,81 0,00005 Imedia - densidad algal 0,61 0,007 densidad algal – P-disuelto -0,56 0,02 densidad algal – P total -0,65 0,003 densidad algal – N-amonio -0,64 0,004 clorofila a –NID -0,7 0,001 densidad algal - NID -0,7 0,001 PA - densidad algal 0,55 0,02 PA/Imedia - N-amonio 0,7 0,001 PA/Chl a - P-disuelto 0,48 0,04 PA/Chl a - N-amonio 0,58 0,01 PA/Chl a - Imedia -0,64 0,004
Tabla 3. Correlaciones de Spearman significativas (N = 18) entre las variables analizadas
en este estudio.
Los valores de irradiancia integrada en la columna de agua (Imedia) fueron entre
20 y 34 veces mayor en el tratamiento libre (Tabla 2), donde tendieron a disminuir con el
tiempo (Fig. 4). El coeficiente g440 no mostró diferencias significativas entre tratamientos
y sus valores fueron más altos al día 20 (Tablas 1 y 2) mientras que el Kd no mostró
diferencias significativas en el tiempo o entre tratamientos (Tabla 1). La profundidad
óptica de todos los mesocosmos estuvo por debajo de 4,6 en las tres fechas estudiadas,
indicando que más del 1% de la luz incidente en superficie llegaba al fondo de cada
mesocosmos (Kirk 1994).
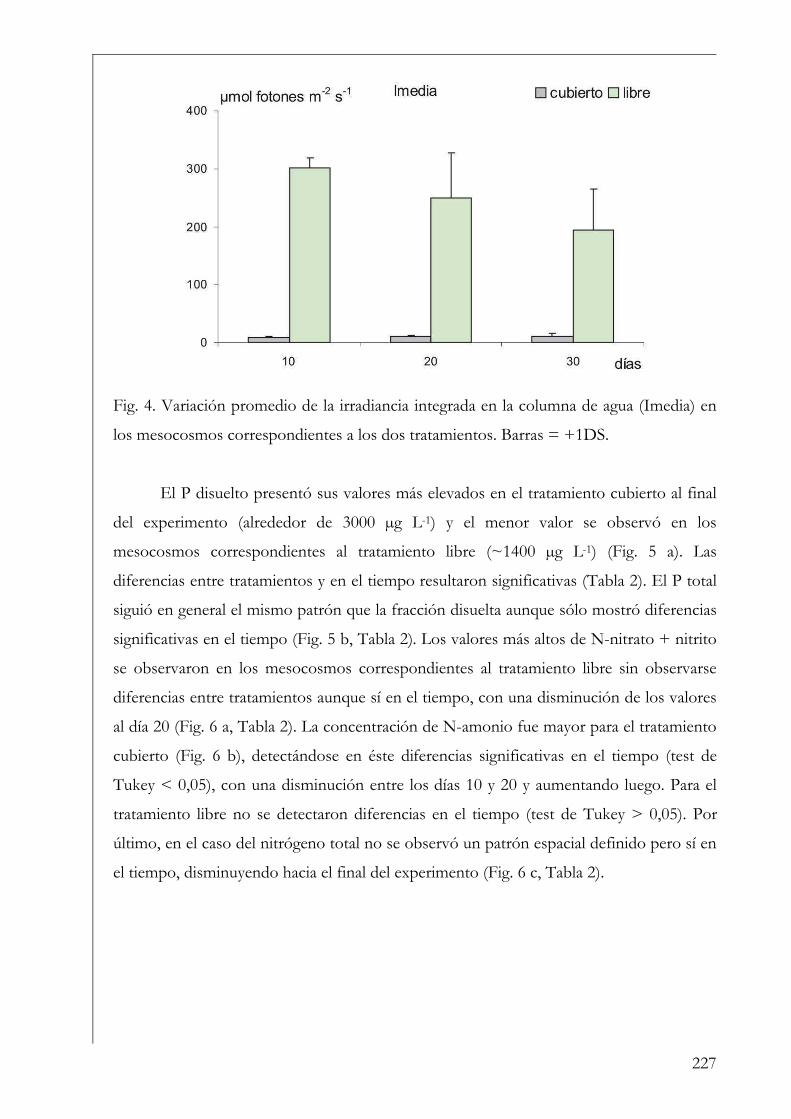
227
Fig. 4. Variación promedio de la irradiancia integrada en la columna de agua (Imedia) en
los mesocosmos correspondientes a los dos tratamientos. Barras = +1DS.
El P disuelto presentó sus valores más elevados en el tratamiento cubierto al final
del experimento (alrededor de 3000 µg L-1) y el menor valor se observó en los
mesocosmos correspondientes al tratamiento libre (~1400 µg L-1) (Fig. 5 a). Las
diferencias entre tratamientos y en el tiempo resultaron significativas (Tabla 2). El P total
siguió en general el mismo patrón que la fracción disuelta aunque sólo mostró diferencias
significativas en el tiempo (Fig. 5 b, Tabla 2). Los valores más altos de N-nitrato + nitrito
se observaron en los mesocosmos correspondientes al tratamiento libre sin observarse
diferencias entre tratamientos aunque sí en el tiempo, con una disminución de los valores
al día 20 (Fig. 6 a, Tabla 2). La concentración de N-amonio fue mayor para el tratamiento
cubierto (Fig. 6 b), detectándose en éste diferencias significativas en el tiempo (test de
Tukey < 0,05), con una disminución entre los días 10 y 20 y aumentando luego. Para el
tratamiento libre no se detectaron diferencias en el tiempo (test de Tukey > 0,05). Por
último, en el caso del nitrógeno total no se observó un patrón espacial definido pero sí en
el tiempo, disminuyendo hacia el final del experimento (Fig. 6 c, Tabla 2).
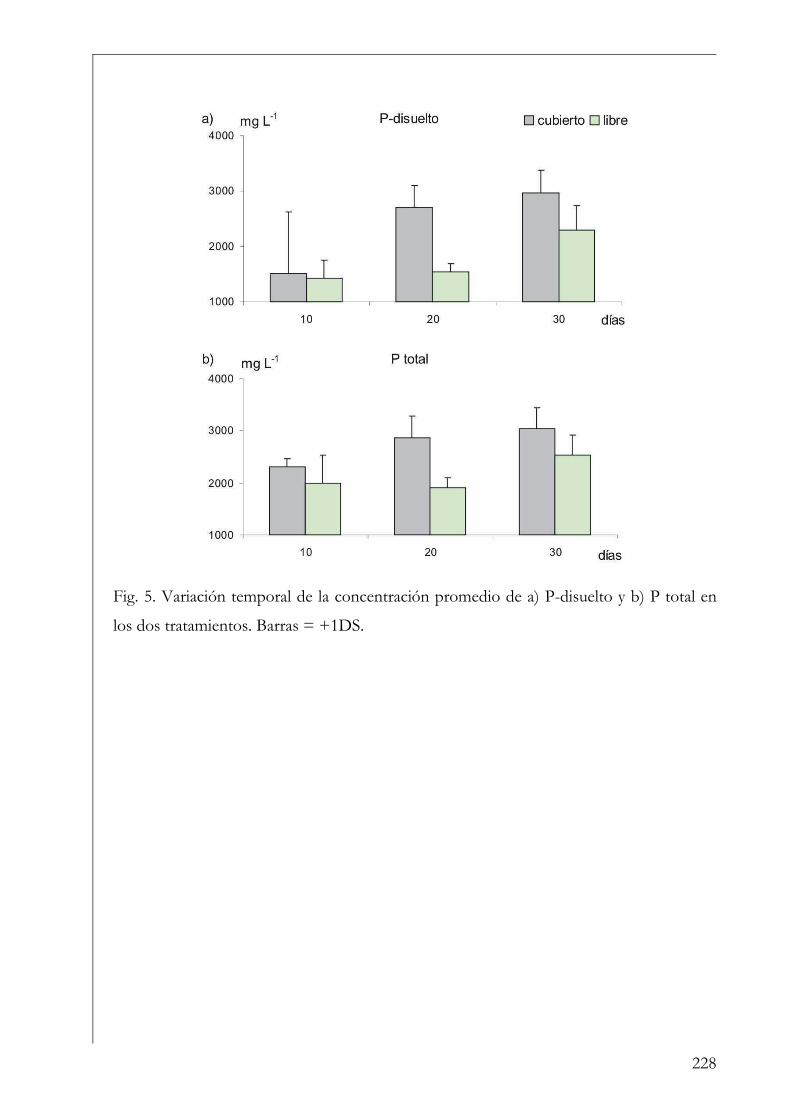
228
Fig. 5. Variación temporal de la concentración promedio de a) P-disuelto y b) P total en
los dos tratamientos. Barras = +1DS.
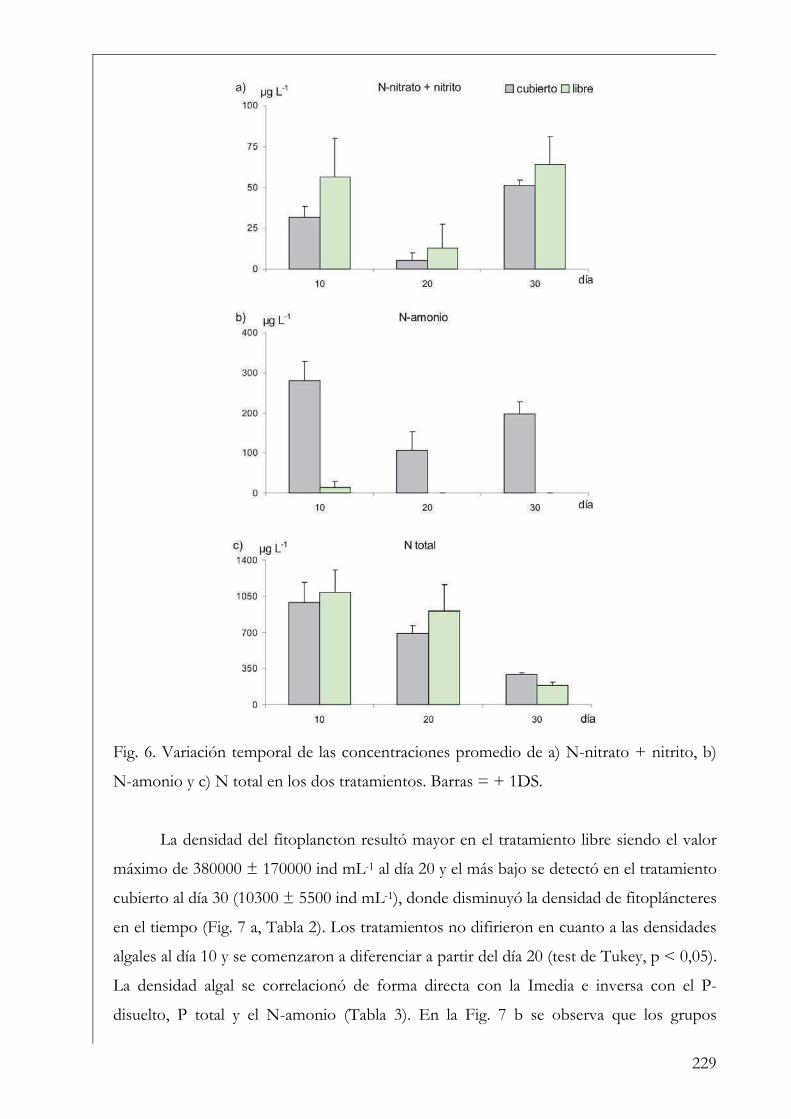
229
Fig. 6. Variación temporal de las concentraciones promedio de a) N-nitrato + nitrito, b)
N-amonio y c) N total en los dos tratamientos. Barras = + 1DS.
La densidad del fitoplancton resultó mayor en el tratamiento libre siendo el valor
máximo de 380000 ± 170000 ind mL-1 al día 20 y el más bajo se detectó en el tratamiento
cubierto al día 30 (10300 ± 5500 ind mL-1), donde disminuyó la densidad de fitopláncteres
en el tiempo (Fig. 7 a, Tabla 2). Los tratamientos no difirieron en cuanto a las densidades
algales al día 10 y se comenzaron a diferenciar a partir del día 20 (test de Tukey, p < 0,05).
La densidad algal se correlacionó de forma directa con la Imedia e inversa con el P-
disuelto, P total y el N-amonio (Tabla 3). En la Fig. 7 b se observa que los grupos
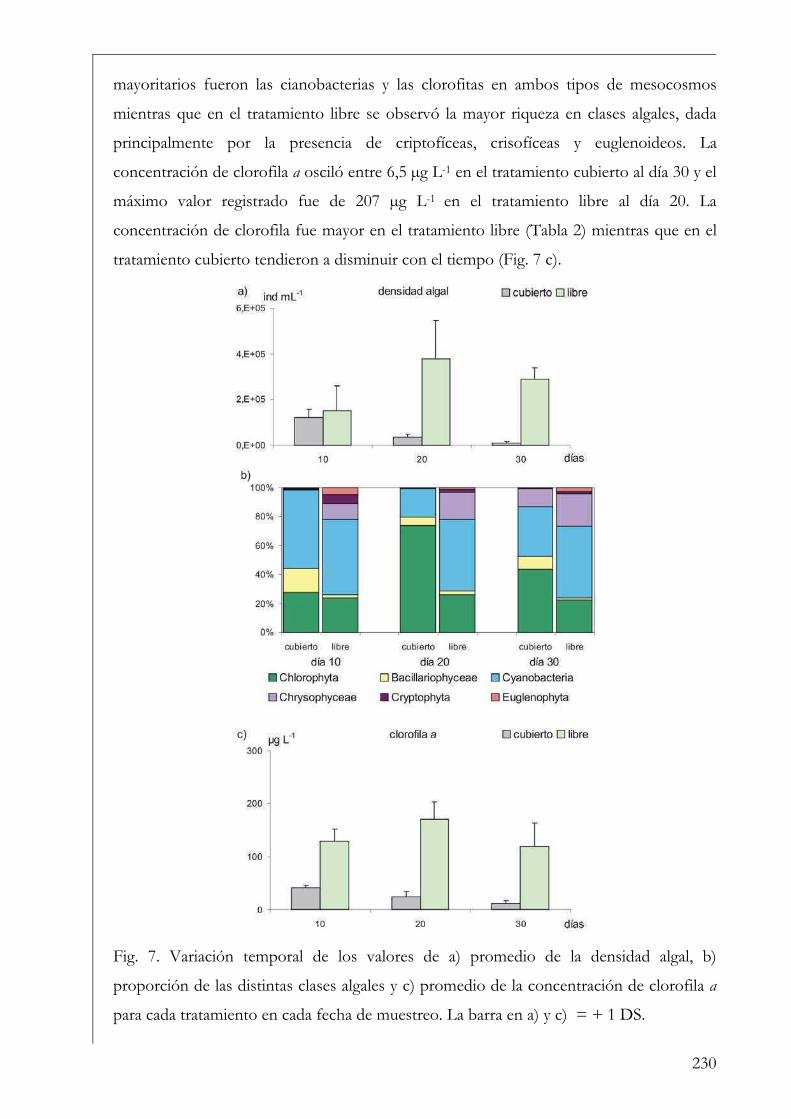
230
mayoritarios fueron las cianobacterias y las clorofitas en ambos tipos de mesocosmos
mientras que en el tratamiento libre se observó la mayor riqueza en clases algales, dada
principalmente por la presencia de criptofíceas, crisofíceas y euglenoideos. La
concentración de clorofila a osciló entre 6,5 µg L-1 en el tratamiento cubierto al día 30 y el
máximo valor registrado fue de 207 µg L-1 en el tratamiento libre al día 20. La
concentración de clorofila fue mayor en el tratamiento libre (Tabla 2) mientras que en el
tratamiento cubierto tendieron a disminuir con el tiempo (Fig. 7 c).
Fig. 7. Variación temporal de los valores de a) promedio de la densidad algal, b)
proporción de las distintas clases algales y c) promedio de la concentración de clorofila a
para cada tratamiento en cada fecha de muestreo. La barra en a) y c) = + 1 DS.

231
La producción primaria integrada en la columna de agua (PA) mostró sus valores
mínimo y máximo al día 30, siendo éstos 2,5 y 94 mg C m-2 h-1 en los tratamientos
cubierto y libre respectivamente (Fig. 8 a). Si bien no se hallaron diferencias significativas
en el tiempo o entre tratamientos, se observó una tendencia a la disminución progresiva
en el tiempo en el tratamiento cubierto en tanto que en el tratamiento libre, PA tendió a
aumentar en el mismo período. La PA se correlacionó de forma positiva con la densidad
algal (Tabla 3). La producción por metro cuadrado relativizada con la Imedia (PA/Imedia)
mostró su valor más bajo (0,002 mg C mmol fotones-1, tratamiento cubierto) y más alto
(1,8 mg C mmol fotones-1, tratamiento libre) a los 10 días de comenzado el experimento
(Fig. 8 b). Los valores del tratamiento cubierto fueron significativamente más altos que los
del libre (Tabla 2), y si bien no se detectaron diferencias significativas en el tiempo, se
pudo observar una tendencia similar a la detectada para los valores promedio de PA. La
producción primaria relativizada con la concentración de clorofila a (PA/Chl a) osciló
entre 0,04 y 2,6 µg C µg Chl a-1 h-1 a los 10 días en los tratamientos libre y cubierto
respectivamente (Fig. 8 c). No se detectaron diferencias significativas entre tratamientos
ni en el tiempo, aunque en el tratamiento libre se observó que los valores promedio
tendieron a aumentar en el tiempo. Esta variable se mostró correlacionada inversamente
con la Imedia y directamente con el N-amonio y P-disuelto (Tabla 3).
Los valores estimados de Ik mostraron que el fitoplancton del tratamiento cubierto
estaría limitado por luz en las tres fechas de muestreo ya que los Ik resultaron ser mayores
que la Imedia (Fig. 9). En el tratamiento libre, el fitoplancton en un sólo mesocosmos se
encontraría limitado lumínicamente a los 10 y 20 días, mientras que al día 30 ninguno de
ellos lo estaría. Por último, las estimaciones del parámetro Ik no difirieron entre
tratamientos o en el tiempo.
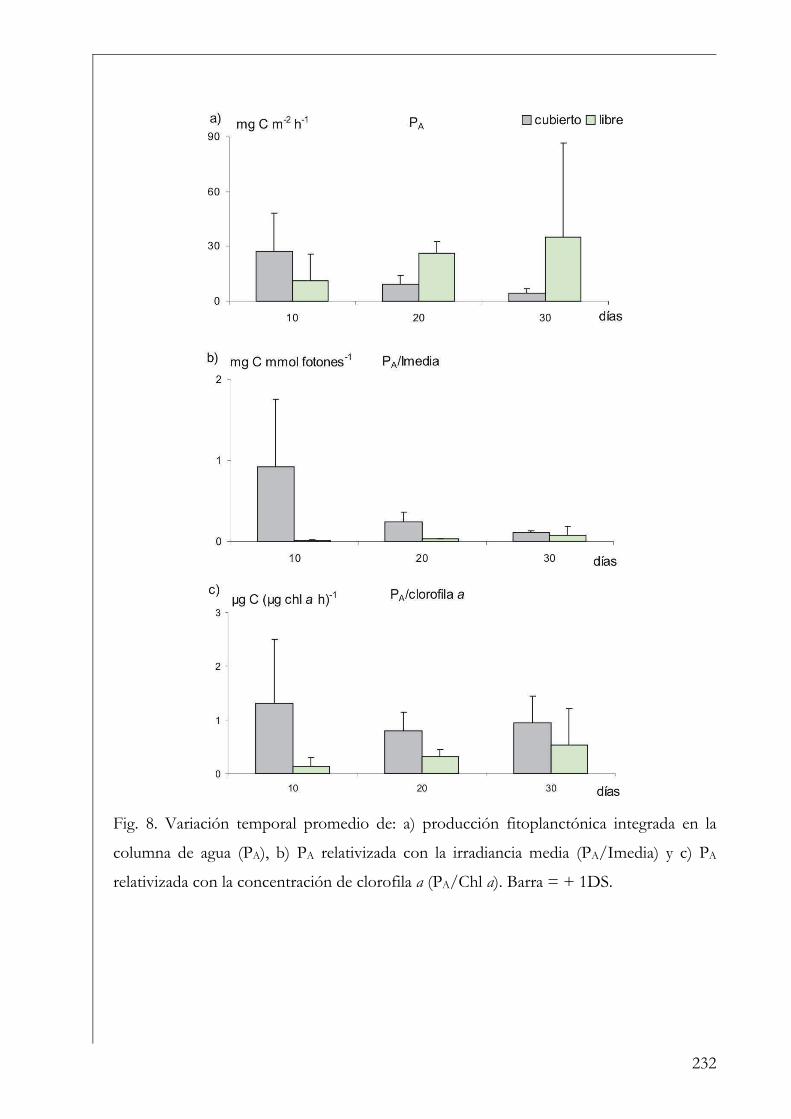
232
Fig. 8. Variación temporal promedio de: a) producción fitoplanctónica integrada en la
columna de agua (PA), b) PA relativizada con la irradiancia media (PA/Imedia) y c) PA
relativizada con la concentración de clorofila a (PA/Chl a). Barra = + 1DS.
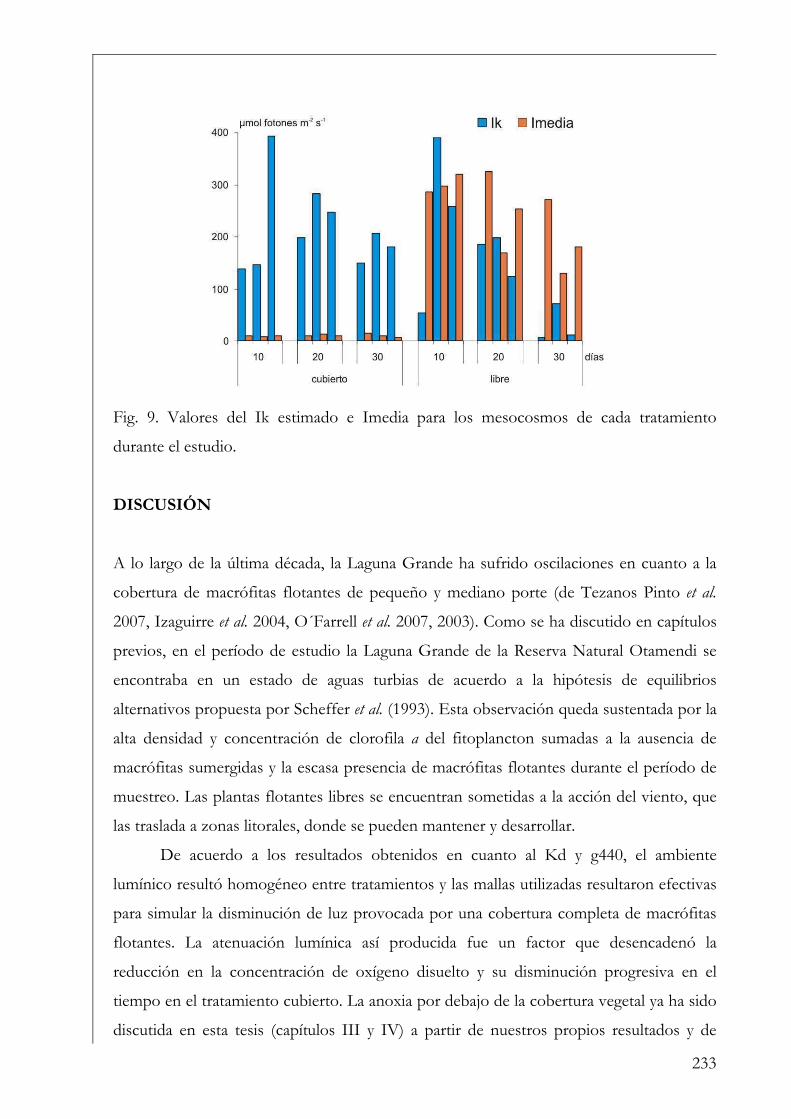
233
Fig. 9. Valores del Ik estimado e Imedia para los mesocosmos de cada tratamiento
durante el estudio.
DISCUSIÓN
A lo largo de la última década, la Laguna Grande ha sufrido oscilaciones en cuanto a la
cobertura de macrófitas flotantes de pequeño y mediano porte (de Tezanos Pinto et al.
2007, Izaguirre et al. 2004, O´Farrell et al. 2007, 2003). Como se ha discutido en capítulos
previos, en el período de estudio la Laguna Grande de la Reserva Natural Otamendi se
encontraba en un estado de aguas turbias de acuerdo a la hipótesis de equilibrios
alternativos propuesta por Scheffer et al. (1993). Esta observación queda sustentada por la
alta densidad y concentración de clorofila a del fitoplancton sumadas a la ausencia de
macrófitas sumergidas y la escasa presencia de macrófitas flotantes durante el período de
muestreo. Las plantas flotantes libres se encuentran sometidas a la acción del viento, que
las traslada a zonas litorales, donde se pueden mantener y desarrollar.
De acuerdo a los resultados obtenidos en cuanto al Kd y g440, el ambiente
lumínico resultó homogéneo entre tratamientos y las mallas utilizadas resultaron efectivas
para simular la disminución de luz provocada por una cobertura completa de macrófitas
flotantes. La atenuación lumínica así producida fue un factor que desencadenó la
reducción en la concentración de oxígeno disuelto y su disminución progresiva en el
tiempo en el tratamiento cubierto. La anoxia por debajo de la cobertura vegetal ya ha sido
discutida en esta tesis (capítulos III y IV) a partir de nuestros propios resultados y de

234
estudios previos en este humedal (Izaguirre et al. 2001 b). También hubo anoxia en el
fondo de todos los dispositivos experimentales, acompañada de un aumento en el tiempo
de la concentración de fósforo, probablemente en relación a la liberación de este nutriente
desde los sedimentos (Nürnberg 1994). La marcada disminución del nivel de agua en los
tratamientos y en la laguna (11 cm desde los 10 a los 30 días de comenzado el
experimento) trajo aparejado el aumento de la conductividad, del g440 y la reducción en la
⦆. En este sentido, la tendencia a la reducción de los valores de Imedia en el tratamiento
libre con el tiempo pudo ser consecuencia del aumento por concentración de las
sustancias húmicas y fitoplancton, mediada por la disminución del nivel hidrométrico con
la consecuente absorción de luz que aparejan.
En este estudio se observó que, debido a las condiciones reductoras en los
mesocosmos correspondientes al tratamiento cubierto, la forma predominante de
nitrógeno fue el amonio, mientras que en el tratamiento libre lo fue el nitrato, como
también observó Sinistro (2007) en la Laguna Grande. En general, las formas químicas
reducidas suelen ser el estado predominante en los cuerpos de agua bajo condiciones de
anoxia (Kalff 2003). La concentración de nitrógeno inorgánico disuelto (NID) en el
tratamiento libre, se mantuvo por debajo del valor de 100 µg L-1, considerado limitante
para el máximo crecimiento algal (Reynolds 2006). Este hecho estuvo relacionado con su
consumo por parte de las altas densidades de fitoplancton presentes en estos
mesocosmos, como indican las correlaciones negativas encontradas entre NID e Imedia,
clorofila a y densidad algal.
El fitoplancton siguió un patrón esperado ya que las menores densidades en el
tratamiento cubierto estarían dadas por la disminución en el crecimiento debido
principalmente a la limitación lumínica, en tanto que la composición algal se encaminó
hacia una comunidad menos variada. En O´Farrell et al. (2009) figuran los valores de
riqueza específica en ambos tratamientos durante el experimento. Pudo observarse que
partiendo de un número de especies semejante entre tratamientos (entre 70 y 80 especies),
el número de especies descendió significativamente a 24 en los mesocosmos cubiertos
mientras que en los libres la riqueza final fue de 60 especies. Es interesante destacar que la
composición de grupos algales en el tratamiento libre fue similar a la hallada en las aguas
libres de la laguna en la misma época del año, que figura en el capítulo II de esta tesis
(pág. 100).

235
Los valores de PA registrados en este estudio para los dos tratamientos están
dentro del rango de los estimados por Henry (1991) y Henry et al. (1993) en una represa
en Brasil y son bajos cuando se los compara con los estimados en la Laguna Grande en el
capítulo II de esta tesis para las aguas libres de la laguna en la misma época del año. Esta
diferencia entre los valores obtenidos en este experimento y aquéllos registrados a partir
de muestras tomadas directamente de la laguna podría estar relacionada con la reducción
de la mezcla en los mesocosmos dada por el viento en la columna de agua. En sistemas
someros con gran atenuación lumínica como la Laguna Grande, los procesos de mezcla
generan mayor variabilidad en el clima lumínico en comparación con cuerpos de agua
relativamente más claros (Helbling et al. 2001 a). Este proceso permite a los organismos
experimentar irradiancias mayores por algunos períodos de tiempo y pone en contacto a
los organismos con otras fuentes de nutrientes, hecho que estimula la fotosíntesis
(Schallenberg & Burns 2004).
La tendencia al aumento de la PA en el tiempo en el tratamiento libre y a su
disminución en el cubierto fue clara aunque estadísticamente no significativa. La
disminución de la producción por efecto de una cubierta de macrófitas fue registrada por
Stanley et al. (2003), quienes realizaron estimaciones de productividad algal y bacteriana en
varios ambientes de un humedal donde se presentaban distintas formas de vida de
macrófitas. Los autores encontraron que en presencia de macrófitas flotantes (Nymphaea
odorata) la producción fitoplanctónica era menor. En nuestro caso, son varios los factores
que pueden estar generando la falta de diferencias significativas entre los mesocosmos.
Uno de ellos fue la variabilidad entre réplicas, común en experimentos de campo a esta
escala. Por otro lado, un diseño que involucrara un tiempo de experimentación mayor tal
vez nos hubiera permitido detectar diferencias significativas, en cuyo caso se hubiera
requerido la utilización de mesocosmos de mayor volumen que evitaran el deterioro de
estos sistemas confinados. Otra posible explicación para la falta de diferencia entre los
valores de PA entre tratamientos está relacionada con las características lumínicas
naturalmente restringidas de la laguna, dadas por su naturaleza húmica. De esta manera, el
fitoplancton de este sitio está compuesto por organismos que poseen variadas estrategias
que les permiten prosperar bajo estas condiciones (mixotrofia, pigmentos accesorios)
(Sinistro et al. 2006; Kirk 1994). La presencia de este tipo de adaptaciones dificulta la
detección de cambios en el funcionamiento a nivel de toda la comunidad, ya que la

236
respuesta en términos de producción se compensaría con cambios tanto estructurales
como funcionales.
La PA relativizada con la Imedia (PA/Imedia) fue utilizada en este capítulo como
un estimador de la "eficiencia en el uso de la luz" por parte del fitoplancton. Observamos
que los mesocosmos con menor densidad de fitoplancton, es decir los correspondientes al
tratamiento cubierto, son los que presentaron mayores valores de eficiencia, aún en
condiciones de limitación lumínica (Ik estimado > Imedia). Es decir que los pocos
organismos presentes estarían utilizando más eficientemente la poca luz que tienen. El
cociente PA/Chl a nos permite estimar el número de asimilación Pmáx obtenido a partir
de una curva P-I y nos permite independizarnos de la biomasa algal. Los valores que
detectamos en este estudio estuvieron en general por debajo de los valores de Pmáx
obtenidos por nosotros para la laguna en el marco de esta tesis. Esta diferencia entre el
sistema natural y el experimental probablemente tenga que ver, también en este caso, con
la mezcla limitada en los mesocosmos. Al estratificar la columna de agua mediante la
instalación de los mesocosmos, se pierde este efecto y por lo tanto la productividad es
más baja. En cuanto a los valores obtenidos en estas condiciones, si bien las diferencias
entre tratamientos no fueron estadísticamente significativas, se puede observar una
tendencia a que los mayores valores de PA/Chl a se encuentren en los mesocosmos
cubiertos, observación que permitiría suponer mayor eficiencia con menos luz y que
quedaría apoyada por la correlación inversa encontrada entre PA/Chl a y la Imedia.
En nuestro estudio hemos podido observar que una cobertura completa de macrófitas
flotantes agudiza la restricción lumínica impuesta primeramente por las altas
concentraciones de ácidos húmicos. Pudimos demostrar que la atenuación severa de la
irradiancia que puede darse naturalmente cuando una carpeta flotante de plantas se
desplaza por acción del viento, generó respuestas en el fitoplancton, provocando cambios
estructurales con pérdida de densidad y riqueza algal así como una tendencia a la
disminución en la producción primaria. Sin embargo, la comunidad a pesar de una posible
limitación lumínica se tornó más eficiente, de modo de economizar la poca luz disponible.
Estos resultados difieren del marco conceptual derivado de ecosistemas terrestres que
afirma que la disminución en variabilidad (medida como riqueza específica) conduce a la
disminución en la eficiencia de captación de recursos y a su vez, a la menor producción de
biomasa (Cardinale et al. 2004). Nuestro estudio demuestra que, en lagunas donde

237
aparecen cubiertas de macrófitas flotantes, sus fluctuaciones en cobertura generan fuertes
cambios en el medio ambiente que afectan a la ecología del fitoplancton. El aumento en
las restricciones lumínicas provoca una respuesta en el ambiente y en el fitoplancton que
se reorganiza, con pérdida de especies, una disminución en la abundancia y en la
producción total, pero con aumento en la eficiencia de captación de luz y de producción
por unidad de biomasa. Por el contrario, la ausencia de macrófitas flotantes lleva a una
comunidad fitoplantónica más heterogénea y más productiva.

238
DISCUSIÓN FINAL

239
Los ambientes acuáticos de la Reserva Natural Otamendi (RNO) comparten la posesión
de altas concentraciones de carbono orgánico disuelto (COD) provenientes en gran parte
de la descomposición del sustrato macrofítico. Las características húmicas dadas por la
alta concentración de COD le confieren a las aguas un color caramelo característico
generando así la disminución de la penetración de la luz y un cambio en la calidad de la
misma, provocando la mayor extinción en las longitudes de onda corta del espectro
electromagnético. De este modo, las comunidades fotótrofas que adquieren prevalencia
en este tipo de ambientes poseen una gama de adaptaciones estructurales y funcionales
que les posibilita prosperar en estas condiciones.
La Laguna Grande se encontró dominada por fitoplancton durante las
investigaciones desarrolladas en el marco de esta tesis, y su dominancia posibilitó que éste
sea el cuerpo de agua más productivo de la RNO. El nivel hidrométrico de la laguna
estuvo dado principalmente por las precipitaciones y no mostró relación con la altura del
río Paraná de Las Palmas a la altura del arroyo Las Rosas. El meandro abandonado de
mayores dimensiones (ROL 1) se cubrió con macrófitas flotantes de pequeño porte
durante gran parte del período de estudio y la laguna El Pescado mostró características
limnológicas similares a la Laguna Grande dadas por su conexión con ésta a través del
arroyo El Pescado. Entre las similitudes se destacó el contenido de COD, la profundidad
y ausencia de macrófitas flotantes. Sin embargo en El Pescado se detectó en general
mayor contenido de nutrientes en su forma inorgánica, relacionado con el menor
desarrollo de fitoplancton; en este sentido, la penetración de la irradiancia fue mayor en
este cuerpo de agua. Estas características probablemente posibilitaron que en este sitio se
observen mayores valores de Pmáx y プ fitoplanctónicos.
La comunidad perifítica sobre sustratos naturales no mostró grandes diferencias de
composición específica en los 5 ambientes estudiados pero sí observamos que la
posibilidad de conexión directa entre los cuerpos de agua generaba características más
similares entre las comunicades que en caso de no poseerla. Dentro de los sistemas
analizados, las lagunas en meandros abandonados (ROLs) presentaron particularidades y
en especial, en el más alejado y con mayor desarrollo de plantas macrófitas emergentes, se
encontró la comunidad menos rica, diferenciándose así del resto. La vegetación palustre
provocó gran variabilidad en el ambiente óptico, además de proveer restricciones
espaciales para la colonización del perifiton. Los humedales de por sí, se caracterizan por
presentar fluctuaciones muy importantes del nivel hídrico, hecho que le confiere una

240
dinámica muy especial a todo el ecosistema. Las comunidades deben estar adaptadas a
períodos de aguas altas y bajas, lo que hemos visualizado en la particular composición de
la comunidad perifítica sobre sustratos naturales, encontrando adaptaciones a la sequedad
en mayor medida en las especies presentes en el junco (planta palustre que queda expuesta
al aire en períodos de aguas bajas). Por otro lado, en la comunidad adherida a macrófitas
flotantes, como el caso de Ricciocarpus natans, fue la luz la variable que modeló en mayor
medida a la comunidad adherida a sus escamas. Si bien la riqueza de especies encontrada
en el perifiton de los cuerpos de agua de la RNO no fue muy alta en comparación con
otros humedales, es importante el aporte a la biodiversidad microbiana del humedal.
En el estudio anual comparado de la producción del fitoplancton y del perifiton en
la Laguna Grande, se evidenció que este sistema se encuentra en un estado de aguas
turbias con gran desarrollo de fitoplancton. En este sentido, el fitoplancton fue la
comunidad más productiva contribuyendo con un 96 % a la producción anual total en
tanto que el perifiton lo hizo en un 4% de acuerdo a nuestras estimaciones. Si bien los
valores de producción anual evaluados por nosotros para el fitoplancton ubicaron a la
Laguna Grande entre cuerpos de agua de productividad intermedia (Kalff 2003), los
valores de Pmáx vol fueron elevados. La alta relación entre la producción por unidad de
volumen sobre la producción areal es característica de los sistemas húmicos. Debido a la
atenuación de la luz en la columna de agua, los organismos tienden a ubicarse en las capas
superiores (Arvola et al. 1999) de modo que cuando se integran los aportes individuales de
cada profundidad (PA), el valor resultante es bajo en relación a considerar el valor de
máxima producción a una sola profundidad (Fig. 1).

241
Fig. 1. Representación esquemática de la producción integrada en la columna de agua, PA
(paralelepípedo negro) y máxima producción por unidad de volumen (Pmáx vol, cilindro
rojo) en un lago húmico hipotético.
Pudimos visualizar que el fitoplancton es un componente importante en la
atenuación de la luz de la columna de agua y que probablemente esté afectando al
crecimiento y producción del perifiton sobre sustratos artificiales. Los tenores de NID
fueron en general bajos en la Laguna Grande, situación que probablemente afecte al
perifiton en mayor medida que al fitoplancton, ya que el perifiton alcanzó sus mayores
densidades y tasas de producción primaria en condiciones de mayor disponibilidad de
NID, menor abundancia de fitoplancton y menor componente heterotrófico del propio
perifiton. El perifiton estuvo estructurado en mayor proporción por organismos
heterotróficos cuya dinámica también estuvo marcada por las variaciones de temperatura.
Observamos que en verano el componente autotrófico del perifiton fue mayor en el
ROL, posibilitando así que en este sitio se alcancen los mayores valores de producción
perifítica, a pesar de la limitación lumínica que experimentaba la comunidad debido a la

242
cobertura completa de plantas flotantes. En esa oportunidad, el perifiton tendió a
presentar mayores valores de Pmáx y プ que el fitoplancton, sugiriendo una alta asimilación
y eficiencia fotosintética de la comunidad adherida, dominada por agregados de
cianobacterias planctónicas y metafíticas.
En el marco de los estudios abordados en esta tesis, hemos visto que la
temperatura fue la variable reguladora más evidente. Cabe destacar que el patrón de
fluctuación de la altura hidrométrica, correlacionado con las precipitaciones pluviales
locales, se vio enmascarado por el de la fluctuación de temperatura, lo que nos alerta a ser
cautelosos en la conclusión general sobre las variables modeladoras. Las altas
temperaturas promovieron la dominancia de cianobacterias filamentosas con altas tasas de
producción en el fitoplancton, que provocaron además el acortamiento de la capa eufótica
y disminución de la productividad por unidad de volumen (Pmáx vol) del propio
fitoplancton en la Laguna Grande. En el estudio fraccionado, registramos que el aporte
del picoplancton a la producción fitoplanctónica total anual resultó ser importante (36 %)
y presentó un patrón estacional marcado para las fluctuaciones de Pmáx y プ, siendo la
temperatura la única variable que explicó la dinámica de la producción areal
picoplanctónica. En ese estudio, el picoplancton resultó tener mayor tasa de asimilación y
eficiencia fotosintética que la fracción de mayor tamaño. También observamos una
tendencia a la disminución de aporte de la fracción picoplanctónica a la PA total y a la
biomasa fotosintética con el aumento del estado trófico cuando se consideraron los datos
de los todos los cuerpos de agua del humedal. Por otro lado, en el caso del micro y
nanoplancton, la dinámica anual de PA estuvo explicada por la combinación del P-disuelto
y la Imedia. El hecho que la temperatura haya resultado la variable modeladora de la
dinámica del picoplancton y del fitoplancton > 0,7 µm pone en evidencia que el
picofitoplacton fue la fracción de tamaño que marcó la tendencia estacional del
fitoplancton en la Laguna Grande.
En el ROL 1 las fracciones de tamaño no mostraron diferencias significativas en
sus parámetros Pmáx y プ. Esto probablemente tenga que ver con que la estrategia del
menor tamaño tal vez ya no sea una ventaja en un sitio tan restringido lumínicamente. Tal
vez sí lo sea una en la competencia por los recursos con el micro y nanplancton (Drakare
et al. 2002), pero no con las bacterias presentes en el ROL (Izaguirre et al. enviado). En el
ROL, el invierno fue el único momento en que el picoplancton tuvo una contribución
mayor que la fracción del nano y microplancton a la PA, probablemente relacionado con

243
la menor competencia del picoplancton con las bacterias dada por la mayor disponibilidad
de NID.
Además de la restricción lumínica generada por las características del agua, existe
otro factor que agudiza esta situación, que es el desarrollo de macrofitia flotante. Las
plantas acuáticas y palustres son comunes en humedales y en particular las flotantes,
pueden desarrollarse hasta cubrir por completo el espejo de los cuerpos de agua. En el
experimento manipulativo que presentamos en la parte final de la presente tesis hemos
puesto de manifiesto que la disminución de luz provocada por una cubierta completa de
macrófitas genera cambios estructurales y funcionales en el fitoplancton. Desde el punto
de vista estructural, provoca la disminución de la densidad total y la caída en la
variabilidad de clases algales. La comunidad menos desarrollada y menos variable produjo
a su vez a tiempo final un 87 % menos que la que permaneció libre de sombreado. Sin
embargo, la comunidad sometida a bajas intensidades lumínicas presentó mayor eficiencia
fotosintética, hecho que constituiría una estrategia adaptativa en ambientes restringidos
lumínicamente. Estos resultados se pueden extrapolar parcialmente a condiciones
naturales, ya que hemos visto que una cubierta continua de macrófitas en el ROL afectó
negativamente a la producción y a la eficiencia de una comunidad fitoplanctónica muy
bien desarrollada, con abundancias celulares semejantes a las de la Laguna Grande sin
cubierta de plantas. Queda en evidencia entonces que cada ambiente acuático del humedal
posee carcaterísticas propias que deben tenerse en cuenta a la hora de las generalizaciones.
El aporte de esta tesis consistió en describir los patrones de productividad de las
comunidades fotótrofas microbianas más importantes del humedal de la RNO,
representativo de ecosistemas de latitudes templadas, donde la fluctuación en el nivel
hídrico suele quedar enmascarada por la estacionalidad. Las características de estos
cuerpos de agua húmicos someros provocan que las comunidades microbianas que los
habitan extremen sus adaptaciones estructurales y funcionales para poder desarrollarse.
Este trabajo provee información de base para el desarrollo futuro de estudios en
relación a la dinámica del carbono, entre otros temas de importancia no sólo local sino
global, a la luz de los procesos actuales de cambio climático. El conocimiento de la
estructura y funcionalidad de los humedales resulta de gran importancia teniendo en
cuenta su preponderante rol en relación al hombre y los servicios que brindan.
Consideramos que es necesario conocerlos para poder conservarlos y disponer de planes
de manejo que permitan tanto su utilización como su resguardo.

244
BIBLIOGRAFÍA Ács, E. & K. Buczkó (1994) Daily changes of reed periphyton composition ina a shallow Hungarian lake (Lake Velence). En: Proceedings of the 13th International Diatom Symposium. D. Marino & M. Montresor (Eds). Biopress Limited, Bristol, pp. 1–10. Agawin, N. S., C. M. Duarte & S. Agustí (2000) Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnology and Oceanography, 45: 591–600. Ahtiainen, M., A. Holopainen & A. Hovi (1985) Phytoplankton, primary production and dark fixation in a polyhumic lake in eastern Finland . Aquatic Ecology, 15: 77-88. American Public Health Association (2005) Standard Methods for the Examination of Water and Wastewaters, 21st Edition. Centennial Edition. APHA, American Water Works Association, Water Environmental Federation, Washington, DC. Anthoni, U., C. Christophersen, J. Madsen, S. Wium-Andersen & N. Jacobsen (1980) Biologically active sulphur compounds from the green alga Chara globularis. Phytochemistry, 19: 1228-1229. Apesteguía, C., C. Marta & R. Tombolini (1978) Producción del perifiton en ambientes acuáticos del río Paraná Medio. I. Técnicas de análisis. Ecología, 3: 43-55. Arvola, L., P. Eloranta, M. Järvinen, J. Keskitalo & A. Holopainen (1999) Phytoplankton. En: Limnology of Humic Waters. J. Keskitalo & P. Eloranta (Eds.). Backuys Publishers, Leiden, pp. 137-171. Baly, E. C. (1935) The kinetics of photosynthesis. Proceedings of the Royal Society of London, Serie B, Biological Sciences, 117: 218-239. Bauer, D. E., N. Gómez & P. R. Hualde (2007) Biofilms coating Schoenoplectus californicus as indicators of water quality in the Río de la Plata Estuary (Argentina). Environmental Monitoring and Assessment, 133: 309-320. Bell, T. & J. Kalff (2001) The contribution of picophytoplankton in marine and freshwater systems of different trophic status and depth. Limnology and Oceanography, 46: 1243-1248. Belykh, O. I., G. Ekaterina, T. Sorokovikova, A. Saphonova & I. Tikhonova (2006) Autotrophic picoplankton of Lake Baikal: composition, abundance and structure. Hydrobiologia, 568: 9-17. Bicudo, D. C., B. Fonseca, L. Bini, L. O. Crossetti, C. E. de M. Bicudo & T. Araújo-Jesus (2007) Undesirable side-effects of water hyacinth control in a shallow tropical reservoir. Freshwater Biology, 52: 1120-1133.

245
Blumenshine, S. C., Y. Vadeboncoeur, D. M. Lodge, K. L. Cottingham & S. E. Knight (1997) Benthic-Pelagic Links: Responses of Benthos to Water-column enrichment. Journal of the North American Benthological Society, 16: 466-479. Bonetto, C., L. De Cabo, N. Gabellone, A. Vinocur, J. Donadelli & F. Unrein (1994) Nutrient dynamics in the deltaic floodplain of the Lower Parana River. Archiv für Hydrobiologie, 131: 277-295. Bonetto, C., Y. Zalocar de Domitrovic & E. R. Vallejos (1983) Fitoplancton y producción primaraia del Río Alto Paraná (Argentina). Physis, 101: 81-93. Bonetto, C., Y. Zalocar, P. M. Caro & E. R. Vallejos (1979) Producción primaria del fitoplancton del río Paraná en el área de su confluencia con el río Paraguay. Ecosur, 6: 207-227. Bonfils, C. G. (1962) Los suelos del Delta de río Paraná. Factores generadores, clasificación y uso. Revista de Investigaciones Agrícolas, 16: 257-370. Borchardt, M. A. (1996) Nutrients. En: Algal Ecology: Freshwater Benthic Ecosystems. R. J. Stevenson, M. L. Bothwell & R. L. Lowe (Eds.). Academic Press, Londres, pp. 183-227. Bothwell, M. L. (1988) Growth rate responses of lotic periphytic diatoms to experimental phosphorus enrichment: The influence of temperature and light. Canadian Journal of Fisheries and Aquatic Sciences, 45: 261-270. Bourrelly, P. (1981) Les alges d´eau douce. II: Les algues jaunes et bruns. N. Boubée, París, 517 p. Bourrelly, P. (1972) Les alges d´eau douce. I: Les algues vertes. N. Boubée, París, 572 p. Bourrelly, P. (1970) Les alges d´eau douce. III: Les algues bleues et rouges. N. Boubée, París, 512 p. Brandt, L. E. & E. W. Koch (2003) Periphyton as UV-B filter on seagrass leaves: a result of different transmittance in the UV-B and PAR. Aquatic Botany, 76: 317-327. Burkholder, J. A. (1996) Interactions of benthic algae with their substrata. En: Algal Ecology: Freshwater Benthic Ecosystems. R. J. Stevenson, M. L. Bothwell & R. L. Lowe (Eds.). Academic Press, Londres, pp. 253-297. Callieri, C. (2008) Picophytoplankton in freshwater ecosystems: the importance of small-sized phototrophs. Freshwater Reviews, 1: 1-28. Callieri C. & J. Stockner (2002) Freshwater autotrophic picoplankton: a review. Journal of Limnology, 61: 1-14. Callieri, C., B. Modenutti, C. Queimaliños, R. Bertoni & E. Balseiro (2007) Production and biomass of picophytoplankton and larger autotrophs in Andean ultraoligotrophic

246
lakes: differences in light harvesting efficiency in deep layers. Aquatic Ecology, 41: 511-523. Callieri, C., S. Moro, E. Caravati, N. D. Crosbie & T. Weisse (2005) Strain-specific photosynthetic response of freshwater picocyanobacteria. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 29: 777-782. Cano, M. G., M. A. Casco, L. C. Solari, M. E. Mac Donagh, N. A Gabellone & M. C. Clips (2008) Implications of rapid changes in chlorophyll a of plankton, epipelon and epiphyton in a Pampean shallow lake: an interpretation in terms of a conceptual model. Hydrobiologia, 614: 33-45. Cardinale, B. J., A. R. Ives & P. Inchausti (2004) Effects of species diversity on the primary productivity of ecosystems: extending our spatial and temporal scales of inference. Oikos, 104: 437-450. Carignan, R. & D. Planas (1994) Recognition of nutrient and light limitation in turbid mixed layers: Three approaches compared in the Paraná floodplain (Argentina). Limnology and Oceanography, 39: 580-596. Carignan, R., J. J. Neiff & D. Planas (1994) Limitation of water hyacinth by nitrogen in subtropical lakes of the Paraná floodplain (Argentina). Limnology and Oceanography, 39: 439-443. Carric, H. J., F. J. Alridge & C. L. Schelske (1993) Wind influences phytoplankton biomass and composition in a shallow, productive lake. Limnology and Oceanography, 38: 1179-1182. Carvalho, P., L. M. Bini, S. M. Thomaz, L. Gonçalves de Oliveira, B. Robertson, W. L. Gomes Tavechio & A. J. Darwisch (2001) Comparative limnology of South American floodplain lakes and lagoons. Acta Scienciatarum, 23: 265-273. Casamayor, E. O., J. García Canzaiano & C. Pedrós Alió (2008) Carbon dioxide fixation in the dark by photosynthetic bacteria in sulfide-rich stratified lakes with oxic-anoxic interfaces. Limnology and Oceanography, 53: 1193-1203. Cattaneo, A. & J. Kalff (1979) Primary production of algae growing on natural and artificial aquatic plants: A study of interactions between epiphytes and their substrate. Limnology and Oceanography, 14: 1031-1037. Chichizola, S. (1993) Las comunidades vegetales de la Reserva Natural Estricta Otamendi y sus relaciones con el ambiente. Parodiana, 8: 227-263. Cleveland, J. S. & M. J. Perry (1987) Quantum yield, relative specific absorption and fluorescence in nitrogen-limited Chaetoceros gracilis. Marine Biology, 94; 489-497. Conzonno, V. & E. F. Claverie (1987) Phytoplankton primary production in Chascomús Pond (Provincia de Buenos Aires, Argentina). Ecosur, 25: 7-16.

247
Côté, B. & T. Platt (1983) Day to day variations in the spring summer photosynthetic parameters of coastal marine phytoplankton. Limnology and Oceanography, 28: 320-44. Cronk, J. K. & W. J. Mitsch (1994) Periphyton productivity on artificial and natural surfaces in constructed freshwater wetlands under different hydrologic regimes. Aquatic Botany, 48: 325-341. de Tezanos Pinto, P. (2008) Influencia de las macrófitas flotantes en la comunidad fitoplanctónica en un humedal del Bajo Paraná (Reserva Natural de Otamendi, Argentina). Tesis Doctoral. Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 195 pp. de Tezanos pinto, P., L. Allende & I. O´Farrell (2007) Influence of free-floating plants on the structure of a natural phytoplankton assemblage: an experimental approach. Journal of Plankton Research, 29: 47-56. Dodds, W. K., B. J. Biggs & R. L. Lowe (1999) Photosynthesis-Irradiances patterns in benthic microalgae: variations as a function of assemblage thickness and community structure. Journal of Phycology, 35: 42-53. Dokulil, M. (1984) Metodología de medición de fotosíntesis en fitoplancton. En: Embalses, Fotosíntesis y Producción Primaria. N. Bahamonde & S. Cabrera (Eds.). Alfabeta, Santiago, Chile, pp. 73-84. Drago, E. C. (1990) Hydrological and geomorphological characteristics of the hydrosystem of the middle Paraná river. Acta Limnologica Brasiliensia, 3: 907-930. Drakare, S., P. Blomqvist, A. K. Bergström & M. Jansson (2003) Relationship between picophytoplankton and environmental variables in lakes along a gradient of water colour and nutrient content. Freshwater Biology, 48: 729-740. Drakare, S., P. Blomqvist, A. Bergström & M. Jansson (2002) Primary production and phytoplankton composition in relation to DOC input and bacterioplankton production in humic lake Örträsket. Freshwater Biology, 47: 41-52. Eilers, P. H. C. & J. C. H. Peeters (1988) A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecological Modelling, 42, 199-215. Eloranta, P. (1999) Humus and water physics. En: Limnology of Humic Waters. J. Keskitalo & P. Eloranta (Eds.). Backuys Publishers, Leiden, pp. 59-74. Eminson, D. & B. Moss (1980) The composition and ecology of periphyton communities in freshwaters. 1. The influence of host type and external environment on community composition. British Phycological Journal, 15: 429–446. Engelhardt, K. A. & M. E. Ritchie (2001) Effects of macrophyte species richness on wetland ecosystem functioning and services. Nature, 411: 687-689.

248
Engle, D. L & J. M. Melack (1993) Consequences of riverine flooding for seston and the periphyton of floating meadows in an Amazon floodplain lake. Limnolology and Oceanography, 38: 1500-1520. Ettl, H. (1983) Chlorophyta I. Phytomonadina. En: Süßwasserflora von Mitteleuropa, 9. A. Pascher (ed.). Gustav Fischer Verlag, Jena, 807 p. Ettl, H. (1977) Xanthophyceae. En: Süßwasserflora von Mitteleuropa, 3. A. Pascher (ed.). Gustav Fischer Verlag, Suttgart, Nueva York, 530 p. Falkowski, P. G. (1981) Light-shade adaptations and assimilation numbers. Journal of Plankton Research, 3: 203–16. Falkowski, P. G. & J. A. Raven (2007) Aquatic Photosynthesis, Second Edition. Princeton University Press, Oxford, 484 p. Falkowski, P. G. & J. LaRoche (1991) Acclimation to spectral irradiance in algae. Journal of Phycology, 27: 8-14. Fee, E. J. (1984) Un modelo para la determinación de la producción primaria. En: Embalses, Fotosíntesis y Producción Primaria. N. Bahamonde & S. Cabrera (Eds.). Alfabeta, Santiago, Chile, pp. 175-178. Fogg, G. E. (1986) Picoplankton. Proceedings of the Royal Society of London. Serie B, 228: 1-30. Gallegos, C. L., R. J. Davies-Colley & M. Gall (2008) Optical closure in lakes with contrasting reflectance. Limnology & Oceanography, 53: 2021-2034. García de Emiliani, M. O. (1997) Effects of water level fluctuations on phytoplankton in a river-floodplain lake system (Paraná River, Argentina). Hydrobiologia, 357: 1-15. García de Emiliani, M. O. (1993) Seasonal succession of phytoplankton in a lake of the Paraná river floodplain, Argentina. Hydrobiologia, 264: 101-114. Geitler, L. (1932) Cyanophyceae. Kryptogamen-Flora von Deutschland, Osterreich und der Schweiz, 14. Akademische Verlagsges, Leipzig, 1196 p. Goldsborough, L. G. & G. C. Robinson (1996) Patterns in wetlands. En: Algal Ecology: Freshwater Benthic Ecosystems. R. J. Stevenson, M. L. Bothwell & R. L. Lowe (Eds.). Academic Press, Londres, pp. 78-117. Goldsborough, L. G., R. L. McDougal & A. K. North (2005) Periphyton in freshwater lakes and wetlands. En: Periphyton: Ecology, Exploitation and Management. M. E. Azim, M. C. Verdegem, A. A. van Dam & M. C. M. Beveridge (Eds.). CABI Publishing, Estados Unidos, pp. 71-89.

249
Gómez, N., M. Licursi & R. R. Hualde (2003) Epiphytic algae on the bulrush (Scirpus californicus (Mey) Steud) in the Río de la Plata (Argentina): structure and architecture. Archiv für Hydrobiologie, 147: 231-247. Grasshoff, K., M. Ehrhardt & K. Kremling, 1983. Determination of nutrients. En: Methods of Seawater Analysis, 2nd edition. Verlang Chemie, Weinheim, pp. 125-188. Greisberger, S., M. T. Dokullil & K. Teubner (2008) A comparison of phytoplankton size-fractions in Mondsee, an alpine lake in Austria: distribution, pigment composition and primary production rates. Aquatic Ecology, 42: 379-389. Haene, E. & J. Pereira (2003) Fauna de Otamendi. Aves Argentinas, Asociación Ornitológica del Plata, 191 p. Haene, E., V. De Francesco, C. Ostrosky & A. Di Giacomo (2003) La Reserva Nartural Otamendi. Descripción general. En: Haene, E. & J. Pereira (eds.) Fauna de Otamendi. Aves Argentinas, Asociación Ornitológica del Plata, p. 5-16. Hansson, L. (1992) Factors regulating periphytic algal biomass. Limnology and Oceanography, 37: 322-328. Happey-Wood, C. M. (1993) Diurnal and seasonal variation in the contributions of autotrophic pico-, nano- and microplankton to the primary production of an upland. Journal of plankton Research, 15: 125-159. Harding, W. R. (1997) Phytoplankton primary productivity in a shallow, well- mixed, hyertrophic Soth African lake. Hydrobiologia, 344: 87-102. Helbling, E. W., V. E. Villafañe & E. S. Barbieri (2001 a) Sensitivity of winter phytoplankton communities from Andean lakes to artificial ultraviolet-B radiation. Revista Chilena de Historia Natural, 74: 273-282. Helbling, E. W., A. G. J. Buma, M. K. de Boer & V. E. Villafañe (2001 b) In situ impact of solar ultraviolet radiation on photosynthesis and DNA in temperate marine phytoplankton. Marine Ecology Progress Series, 211: 43-49. Helbling, E. W., V. E. Villafañe & O. Holm-Hansen (1994) Effects of ultraviolet radiation on Antarctic marine phytoplankton photosynthesis with particular attention to the influence of mixing. En: Ultraviolet Radiation in Antarctica: Measurements and Biological Effects. S. Weiler & P. Penhale (Eds.). American Geophysical Union, Washington, Estados Unidos, pp. 207-27. Henley W. J. (1993) Measurement and interpretation of photosinthetic light-response curves in algae in the context of photoinhibition and diel changes. Journal of Phycology, 29, 729-739. Henry, R. (1993) Primary production by phytoplankton and its controlling factors in Jurumirim reservoir (São Paulo, Brazil). Revista Brasileira de Biología, 53: 489-99.

250
Henry, R., M. A. Nunes, P. M. Mitsuka, N. Lima, & S. M. C. De e Casanova (1998) Variacão espacial e temporal da produtividade primária pelo fitoplâncton na represa de Jurumirim (Rio Paranapanema, Sp). Revista Brasileira de Biologia, 58: 571-590. Henry-Silva, G., A. F. M. Camargo & M. M. Pezzato (2008) Growth of free floating aquatic macrophytes in different concentrations of nutrients. Hydrobiologia, 610: 153-160. Hill, W. (1996) Effects of light. En: Algal Ecology: Freshwater Benthic Ecosystems. R. J. Stevenson, M. L. Bothwell & R. L. Lowe (Eds.). Academic Press, Londres, pp. 121-148. Holm-Hansen O. & E. W. Helbling (1995) Técnicas para la medición de la productividad primaria en el fitoplancton. En: Manual de métodos ficológicos. K. Alveal, M. E. Ferrario, E. C. Oliveira & E. Sar (Eds.). Universidad de Concepción, Concepción, Chile, p. 329-350. Huber-Pestalozzi, G. (1955) Euglenophyceen. En: Das Phytoplankton des Süßwassers. G. Huber-Pestalozzi (ed.). Schwerzerbart'sche Verlags-buchhandlung, Stuttgart. 4: 1-606. Huszar, V. L. & C. S. Reynolds (1997) Phytoplankton periodicity and sequences of dominance in an Amazonian floodplain lake (Lago Batata, Pará, Brasil): responses to gradual environmental change. Hydrobiologia, 346: 169-181. Iglesias, C., N. Mazzeo, G. Goyenola, C. Fosalba, F. Teixeira de Mello, S. García & E. Jeppesen (2008) Field and experimental evidence of the effect of Jenynsia multidentata, a small omnivorous–planktivorous fish, on the size distribution of zooplankton in subtropical lakes. Freshwater Biology, 53: 1797-1807. Irwin, A. J., Z. V. Finkel, O. M. Schofield & P. G. Flakowshi (2006) Scaling-up from nutrient physiology to the size-structure of phytoplankton communities. Journal of Plankton Research, 28: 459-471. Izaguirre, I. & I. O´Farrell (1999) Temporal fluctuations of phytoplankton chlorophyll a and primary production in a turbid floodplain lake of the River Paraná (Argentina). Gayana Botanica, 56: 39-51. Izaguirre, I., H. Pizarro, P. de Tezanos Pinto, P. Rodríguez, I. O´Farrell, F. Unrein & J. Gasol. Light as the primary structuring agent of photosynthetic picoplankton structure and productivity in wetlands. Enviado a Freshwater Biology. Izaguirre, I., I. O´Farrell, F. Unrein, R. Sinistro, M. dos Santos Afonso & G. Tell (2004) Algal assemblages across a wetland, from a shallow lake to relictual oxbow lakes (Lower Paraná River, South America). Hydrobiologia, 511, 25-36. Izaguirre, I., I. O´Farrell & G. Tell (2001 a) Variation in phytoplankton composition and limnological features in a water-water ecotone of the Lower Paraná Basin (Argentina). Freshwater Biology, 46, 63-74.

251
Izaguirre, I., R. Sinistro, I. O´Farrell, F. Unrein & G. Tell (2001 b) Algal assemblages in anoxic relictual oxbow lakes from the Lower Paraná floodplain (Argentina). Nova Hedwigia, 123: 95-106. Jackson, T. A. & R. E. Hecky (1980) Depression of primary productivity by humic matter in lake and reservoir waters of the boreal forest zone. Canadian Journal of Fisheries and Aquatic Sciences, 37: 2300-2317. Jansson, M., A. Bergström, P. Blomqvist & S. Drakare (2001) Allochtonous organic carbon and phytoplankton/bacterioplankton production relationships in lakes. Ecology, 81: 3250-3255. Jansson, M., P. Blomqvist, A. Jonsson & A. K. Bergström (1996) Nutrient limitation of bacterioplankton, autotrophic and mixotrophic phytoplankton, and heterotrophic nanoflagellates in Lake Östräsket. Limnology and Oceanography, 41: 1552-1559. Jassby, A. D. & T. Platt (1976) Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnology and Oceanography, 21: 540-547. Jasser, I. (1997) The dynamics and importance of picoplankton in shallow, dystrophic lake in comparison with surface waters of two deep lakes with contrasting trophic status. Hydrobiologia, 342/343: 87-93. Jasser, I. & L. Arvola (2003) Potential effects of abiotic factors on the abundance of autotrophic picoplankton in four boreal lakes. Jorunal of Plankton Research, 25: 873-883. Jasser, I. & L. Arvola (2002) Seasonal coupling of atotrophic picoplankton and phytoplankton in three boreal lakes. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 28: 190-193. Jeppesen, E., M. Søndergaard, M. Søndergaard & K. Christoffersen (1998) The structuring role of submerged macrophytes in Lakes. Ecological Studies 131, Springer-Verlag, New York, 423 p. Jones, J. I., J. O. Young, J. W. Eaton & B. Moss (2002) The influence of nutrient loading, dissolved inorganic carbon and higher trophic levels on the interaction between submerged plants and periphyton. Journal of Ecology, 90: 12-24. Jones, R. I. (2000) Mixotrophy in planktonic protists: an overview. Freshwater Biology, 45: 219-226. Jones, R. I. (1994) Mixotrophy in planktonic protists as a spectrum of nutritional strategies. Marine Microbial Food Webs, 8: 87-96. Jones, R. I. (1992) The Influence of Humic Substances on Lacustrine Planktonic Food Chains. Hydrobiologia, 229: 73-91.

252
Jost, G., M. Zubkov, E. Yakushev, M. Labrenz & K. Jürgens (2008) High abundance and dark CO2 fixation of chemolithoautotrophic prokaryotes in anoxic waters of the Baltic sea. Limnology and Oceanography, 53: 14-22. Junk, W. J., P. B. Bayley & R. Sparks (1989) The flood pulse concept in river-floodplain systems. En: Proceedings of the International Large River Sympsoium. J. P. Podge (ed.). Canadians Special Publication of Fisheries and Aquatic Sciences, Ohawa, Canada, pp. 110-127. Kaggwa, R. C., D. Kasule, A. A. van Dam & F. Kansiime (2006) An initial assessment of the use of wetlands plants as substrates for periphyton production in seasonal wetland fishponds in Uganda. International Jorunal of Ecology and Environmental Sciences, 32: 63-74. Kairesalo, T. (1980) Comparison of in situ photosynthetic activity of epiphytic, epipelic and planktonic algal communities in an oligotrophic lake, Southern Finland. Journal of Phycology, 16: 57-62. Kalff, J. (2003) Limnology, Inland Water Ecosystems. Prentice Hall, New Jersey, 592 pp. Khondker, M. & M. A. Kabir (1995) Phytoplankton primary production in a mesotrophic pond in sub-tropical Bangladesh. Hydrobiologia, 304: 39-47. Kirk, J. T. O. (1994) Light and photosynthesis in aquatic ecosystems. Second Edition. Cambridge University Press, Estados Unidos, 509 p. Knoll, L. B., M. J. Vanni & W. H. Renwick (2003) Phytoplankton primary production and photosynthetic parameters in reservoirs along a gradient of watershed land use. Limnology and Oceanography, 48: 608-617. Komárek, J. & K. Anagnostidis (2005) Cyanoprokariota. 2. Teil: Oscillatoriales. En: Süßwasserflora von Mitteleuropa. Büdel, B., G. Gärtner, L. Krienitz & M. Schagerl (eds.). Elsevier, Heidelberg, 759 p. Komárek, J. & K. Anagnostidis (1999) Cyanoprokariota. 1. Teil: Chroococcales. En: Süßwasserflora von Mitteleuropa. Ettl, H., G. Gärtner, H. Heynig & D. Mollenhauer (eds.). Gustav Fisher, Stuttgart, 548 p. Komárek, J. & B. Fott (1983) Chlorophyceae (Grünalgen) Ordnung Chlorococcales. En: Das Phytoplankton des Süベwassers. G. Huber Pestalozzi, H. Heynig & D. Mollenhauer (Eds.) Gustav Fischer, Jena, 548 p. Kortelainen, P. (1999) Acidity and buffer capacity. En: Limnology of humic waters. J. Keskitalo & P. Eloranta (Eds.). Backyus Publishers, Leiden, pp. 95-115. Krammer, K. & H. Lange-Bertalot (1991 a) Bacillariophyceae. 3. Centrales, Fragilariaceae, Eunotiaceae. En: Süßwasserflora von Mitteleuropa 2/3. H. Ettl, J. Gerloff, H. Heying & D. Mollenhauer (Eds.). Gustav Ficher Verlag, Jena, 576 p.

253
Krammer, K. & H. Lange-Bertalot (1991 b) Bacillariophyceae. 4. Achnanthaceae, Kritischie Ergänzungen zu Navicula (Lineolate) und Gomphonema. En: Süßwasserflora von Mitteleuropa 2/4. H. Ettl, J. Gerloff, H. Heying & D. Mollenhauer (Eds.). Gustav Ficher Verlag, Jena, 437 p. Krammer, K. & H. Lange-Bertalot (1988) Bacillariophyceae. 2. Bacillariaceae, Epithemiaceae, Surirellaceae. En: Süßwasserflora von Mitteleuropa 2/2. H. Ettl, J. Gerloff, H. Heying & D. Mollenhauer (Eds.), Gustav Ficher Verlag, Jena, 596 p. Krammer, K. & H. Lange-Bertalot (1986) Bacillariophyceae. 1. Naviculaceae. En: Süßwasserflora von Mitteleuropa 2/1. H. Ettl, J. Gerloff, H. Heying & D. Mollenhauer (Eds.). Gustav Ficher Verlag, Jena, 876 p. Krause-Jensen, D. & K. Sand-Jensen (1998) Light attenuation and photosynthesis of aquatic plant communities. Limnology and Oceanography, 43: 396-407. Kronberg, L. (1999) Content of humic substances in freshwater. En: Limnology of Humic Waters. J. Keskitalo & P. Eloranta (Eds.), Backyus Publishers, Leiden, pp. 9-10. Lakatos, G. (1989) Composition of reed periphyton (biotecton) in the Hungarian part of lake Fertö. Biol. Forschun. für Burg., 71: 125-134. Lange-Bertalot, H. (2001) Navicula sensu stricto. 10 Genera separated from Navicula sensu lato. Frustulia. En: Diatoms of Europe. Volume 2. H. Lange Bertalot (ed.), 526 p. Laugaste, R. & K. Lessok (2004) Planktonic algae and epiphyton of the littoral in Lake Peipsi, Estonia. Limnologica, 34: 90-97. Lean, D. R. & B. K. Burnison (1979) An evaluation of errors in the 14C method of primary production measurement. Limnology and Oceanography, 24: 917-928. Leandrini, J. A. & L. Rodrigues (2008) Temporal variation of periphyton biomass in semilotic environments of the upper Paraná River floodplain. Acta Limnologica Brasiliensia, 20: 21-28. Legendre, L., S. Demers, C. M. Yentsch & C. S. Yentsch (1983) The 14C method: Patterns of dark CO2 fixation and DCMU correction to replace the dark bottle. Limnology and Oceanography, 28: 996-1003. Lepistö, L. & A. Holopainen (2003) Occurrence of Cryptophyceae and katablepharids in boreal lakes. Hydrobiologia, 502: 307-314. Lepistö, L., A. Holopainen & H. Vuoristo (2004) Type-specific and indicator taxa of phytoplankton as a quality criterion for assessing the ecological status of Finish boreal lakes. Limnologica, 34: 236-248. Liboriussen, L. (2003) Productio, regutaion and ecophysiology of periphyton in shallow freshwater lakes. PhD Thesis. National Environmental Research Intsitute, ministry of the Environment, Denmark, 48 p.

254
Liboriussen, L. & E. Jeppesen (2005) Structure, biomass, production and depth distribution of periphyton on artificial substratum in shallow lakes with contrasting nutrient concentrations. Freshwater Biology, 51: 95-109. Liboriussen, L. & E. Jeppesen (2003) Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biology, 48: 418-431. Lodge, D. M., S. C. Blumenshine & Y. Vadeboncoeur (1998) Insights and Application of Large-Scale, Long-Term Ecological Observations and Experiments. En: Experimental Ecology: Issues and Perspectives. W. J. Resetarits Jr. & J. Bernardo (Eds.). Oxford, Lonres, pp. 203-235. Loiselle, S. A., L. Bracchini, A. Cózar, A. M. Dattilo & C. Rossi (2005) Extensive spatial analysis of the light environment in a subtropical shallow lake, Laguna Iberá, Argentina. Hydrobiologia, 534: 181-191. Lorenzen, C. J. (1967) Determination of chlorophyll and pheopigments: spectrophotometric equations. Limnology and Oceanography, 12: 343-346. Lowe R. L. & Y. Pan (1996) Benthic algal communities as biological monitors. En: Algal Ecology: Freshwater Benthic Ecosystems. R. J. Stevenson, M. L. Bothwell & R. L. Lowe (Eds.). Academic Press, Londres, pp. 705-739. Macedo, M. F., P. Duarte & J. G. Ferreira (2002) The influence of incubations periods on phytosynthesis-irradiance curves. Journal of Experimental Marine Biology and Ecology, 274: 101-120. Macedo, M. F., P. Duarte, P. Mendes, & J. G. Ferreira (2001) Annual variation of environmental variables, phytoplankton species composition and photosynthetic parameters in a coastal lagoon. Journal of Plankton Research, 23: 719-32. Mackereth, F. J. H., J. Heron & J. F. Talling (1978) Water Analysis: Some Revised Methods for Limnologists. Freshwater Biological Association, Windermere, UK, 120 pp. Magurran, A. E. (2006) Measuring Biological Diversity. Blackwell Science Ltd., Estados Unidos, 256 p. Malinsky-Rushansky, N., T. Berman & Z. Dubinsky (1997) Seasonal photosynthetic activity of autotrophic picoplankton in Lake Kinneret, Israel. Journal of Plankton Research, 19: 979-993. Maltby, E. (1991) Wetlands and their values. En: Wetlands. M. Finlayson & M. Moser (Eds.). International Waterfowl and Wetlands Research Bureau, Slimbridge, pp. 8-26. Margalef, R. (1978) Life forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta, 1: 493-509.

255
McBride, G. B. (1992) Simple calculation of daily photosynthesis by means of five photosynthesis-light equations. Limnology and Oceanography, 37: 1796-7808. Mercado, L. M. & N. Gómez (1999) Effects of a nuclear power plant on phytoplankton structure of the lower Parana River. Interciencia, 24: 36-41. Meerhoff, M. & N. Mazzeo (2004). Importancia de las plantas flotantes libres de gran porte en la conservación y rehabilitación de lagos someros de Sudamérica. Ecosistemas, 13: 12-22. Meerhoff, M., N. Mazzeo, B. Moss & L. Rodríguez-Gallego (2003) The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aquatic Ecology, 37: 377-391. Melzer, A. (1999) Aquatic macrophytes as tools for lake management. Hydrobiologia, 395/396: 181-190. Meulemans, J. T. (1988) Seasonal changes in biomass and production of periphyton growing upon reed in Lake Maarsseveen I. Archiv für Hydrobiologie, 112: 21-42. Mitsch, W. J. & X. Wu (1995) Wetlands and global change. En: Advances in Solil Science, Soil Management and Greenhouse Effect. R. Lal, J. Kimble & B. A. Stewart (Eds.). CRC Lewis Publishers, Florida, pp. 205-230. Mitsch, W. J. & J. G. Gosselink (1993) Wetlands, 2nd edition. John Wiley, Nueva York, 730 p. Moschini-carlos, V., R. Henry & M. R. Pompêo (2000) Seasonal variation of biomasa and productivity of the periphytic community on artificial substrata in the Jurumirim Reservori (São Paulo, Brazil). Hydrobiologia, 434: 35-40. Mózes, A., M. Présing & L. Vörös (2006) Seasonal dynamics of picocyanobacteria and picoeukaryotes in a large shallow lake (Lake Balaton, Hungary). International Review of Hydrobiology, 91: 38-50. Nagata, T., K. Takai, K. Kawanobe, D. Kim, R. Nakazato, N. Guselnikova, N. Bondarenko, O. Mologawaya, T. Kostrnova, V. Drucker, Y. Satoh & Y. Watanabe (1994) Autotrophic picoplankton in southern Lake Baikal: abundance, growth and grazing mortality during summer. Journal of Plankton Research, 16: 945-959. Nahlik, A. M. & W. J. Mitsch (2006) Tropical treatment wetlands dominated by free-floating macrophytes for water quality improvement in Costa Rica. Ecological Engineering, 28: 246-257. Neale, P., J. F. Talling, S. I. Heaney, C. S, Reynolds & J. W. Lund (1991) Long time series from the English Lake District: Irradiance-dependent phytoplankton dynamics during the spring maximum. Limnology and Oceanography, 36: 751-760.

256
Neiff, J. J. (1999) El régimen de pulsos en ríos y grandes humedales de Sudamérica. En: Tópicos sobre humedales subtropicales y templados de Sudamérica. A. I. Malvárez (ed.) Oficina Regional de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe (ORCYT), Montevideo, Uruguay, pp. 97-146. Neiff, J. J. & A. Poi de Neiff (1984) Cambios estacionales en la biomasa de Eichhornia crassipes (Mart.) Solms y su fauna en una laguna del Chaco (Argentina). Ecosur, 11: 51-60. Neiff, J. J. & A. Poi de Neiff (1978) Estudios sucesionales de los camalotales chaqueños y su fauna asociada. I. Etapa seral Pistia stratiotes – Eichornia crassipes. Physis, 37: 29-39. Neiff, J. J., A. Poi de Neiff & S. L. Casco (2001) The effect of prolongued floods on Eichornia crassipes growth in Paraná Rover floodplain lakes. Acta Limnologica Brasiliensis, 13: 51-60. Nürnberg, G. K. (1984) The prediction of internal phosphorus load in lakes with anoxic hypolimnia. Limnology and Oceanography, 29: 111-124. Oduor, S. O. & M Schagerl (2007) Phytoplankton primary productivity characteristics in response to photosynthetically active radiation in three Kenyan Rift Valley saline–alkaline lakes. Journal of plankton Research, 29: 1041-1050. O´Farrell, I., P. de Tezanos Pinto, P. Rodríguez, G. Chaparro & H. Pizarro (2009) Experimental evidence of the dynamic effect of free-floating plants on phytoplankton ecology. Freshwater Biology, 54: 363-375. O´Farrell, I., P. de Tezanos Pinto & I. Izaguirre (2007) Phytoplankton morphological response to the underwater Light conditions in a vegetated wetland. Hydrobiologia, 578: 65-77. O´Farrell. I., R. Sinistro, I. Izaguirre & F. Unrein (2003) Do steady state assemblages occur in shallow lentic environments from wetlands? Hydrobiologia, 502, 197-209. Oliver, R. L. & G. G. Ganf (2000) Freshwaters blooms. En: The Ecology of Cyanobacteria: Their Diversity in Time and Space. B. Whitton & M. Potts, (Eds.), Kluwer Academic Publishers, pp. 149-194. O´Reilly, C. (2006) Seasonal dynamics of periphyton in a large tropical lake. Hydrobiologia, 553: 293-301. Pålsson, C. & W. Granéli (2004) Nutrient limitation of autrotrophic and mixotrophic phytoplankton in a temperate and tropical humic lake gradient. Journal of Plankton Research, 26: 1005-1014. Peterson, B. J. (1980) Aquatic primary productivity and the 14C-CO2 method: A history of the productivity problem. Annual Review of Ecology and Systematics, 11: 359-385.

257
Petrucio, M. M. & F. A. Barbosa (2004) Diel variations of phytoplankton and bacterioplankton production rates in four tropical lakes in the middle Rio Doce basin (southeastern Brazil). Hydrobiologia, 513: 71-76. Petrucio, M. M., F. A. Barbosa & S. M. Thomaz (2005) Bacteria and phytoplankton production rates in eight river stretches of the middle Rio Doce hydrographic basin (southeast Brazil). Brazilian Archives of Biology and Technology, 48: 487-496. Pizarro, H. (1999) Periphyton biomass on Echinocloa polystachya (H. B. K.) Hitch. Of a lake of the Lower Paraná River floodplain, Argentina. Hydrobiologia, 397: 22-239. Pizarro, H. (1995) The genus Characiopsis Borzi (Mischococcales, Tribophyceae). Taxonomy, biogeography and ecology. J. Cramer, Stuttgart, 113 p. Pizarro, H., L. Allende & S. M. Bonaventura (2004) Littoral epilithon of lentic water bodies at Hope Bay, Antarctic Peninsula: biomass variables in relation to environmental conditions. Hydrobiologia, 529: 237-250. Platt, T., C. L. Gallegos & W. G. Harrison (1980) Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. Journal of Marine Research, 38: 687-701. Poi de Neiff, A. & J. J. Neiff (2006) Riqueza de especies y similaridad de los invertebrados que viven en plantas flotantes de la planicie de inundación del río Paraná (Argentina). Interciência, 31: 220-225. Poi de Neiff, A. & R. Carignan (1997) Macroinvertebrates on Eichhornia crassipes roots in two lakes of the Paraná River floodplain. Hydrobiologia, 345: 185-196. Pozzobon, M. V. & G. Tell (1995) Estructura y dinámica de la comunidad perifítica sobre Ricciocarpus natans (Hepaticae) de la Laguna de Los Padres (Buenos Aires, Argentina). Boletín de la Sociedad Argentina de Botánica, 30: 199-208. Prakash, A., R. W. Sheldon & W. H. Sutcliffe Jr. (1991) Geographic variation of oceanic 14C dark uptake. Limnology and Oceanography, 36: 30-39. Pregnall, A. M. (1991) Photosynthesis/Translocation: Aquatic. En: Carbon Isotope Techniques. D. C. Coleman & B. Fry (Eds.). Academic Press, San Diego, pp. 53-75. Prézelin, B. B., M. M. Tilzer, O. Schofield & C. Haese (1991) The control of the production process of phytoplankton by the physical structure of the aquatic environment with special reference to its optical properties. Aquatic Sciences, 52: 137-185. Printz, H. (1964) Die Chaetophoralen der Binnengewasser (eine systematische Ubersicht). Hydrobiologia, 24: 1-376. Putz, R. (1997) Periphyton communities in Amazonian black- and whitewater habitats: Community structure, biomass and productivity. Aquatic Sciences, 59: 74-93.

258
Putz, R. & W. J. Junk (1997) Phytoplankton and periphyton. En: The Central Amazon floodplain: Ecology of a Pulsing System. W. J. Junk (ed.). Ecological Studies, 126, Berlin, Springer-Verlag, pp. 207-222. Rahaingomanana, N., F. A. R. Barbosa & M. M. Petrucio (2002) Fractioned Primary Production of Phytoplankton in Lakes of the Rio Doce Valley (Southeastern Brazil). Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 28: 695-699. Raven, J. A. (1998) The twelfth Tansley Lecture. Small is beautiful: the picophytoplankton. Functional Ecology, 12: 503-513. Reynolds, C. S. (2006) The Ecology of Phytoplankton. Cambridge University Press, Cambridge, 535 p. Reynolds, C. S. (1984) The Ecology of Freshwater Phytoplankton. Cambridge University Press, Londres, 384 p. Reynolds, C. S. (1980) Phytoplankton assemblages and their periodicity in stratifying lake systems. Holartic Ecology, 3: 141-159. Reynolds, C. S., V. Huszar, C. Kruk, L. Naselli-Flores & S. Melo (2002) Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 24: 417-428. Robinson, G., S. E. Gurney, & L. G. Goldsborough (1997 a) Response of benthic and planktonic algal biomass to experimental water-level manipulation in a praire lakeshore wetland. Wetlands, 17: 167-181. Robinson, G., S. E. Gurney, & L. G. Goldsborough (1997 b) The primary productivity of benthic and planktonic algae in a prairie wetland under controlled water-level regimes. Wetlands, 17: 182-194. Rodrigues, L. & D. Bicudo (2001) Similarity among periphyton algal communities in a lentic-lotic gradient of the upper Paraná river floodplain, Brazil. Revista Brasileira de Botânica, 24: 235-248. Rodríguez, P. & H. Pizarro (2007) Phytoplankton productivity in a highly colored shallow lake of a South American floodplain. Wetlands, 24 (4): 1153-1160. Roehm, C. (2005) Respiration in wetland ecosystems. En: Respiration in Aquatic Ecosystems. P. del Giorgio & P. J. le B. Williams (Eds.). Oxford University Press, pp. 83-102. Romesburg, H. C. (1984) Cluster Analysis for Researches. Lifetime Learning Publications. Belmont, California, 334 p. Rosemond, A. D., P. J. Mullholland & S. H. Brawley (2000) Seasonality shifting limitation of stream periphyton: response of algal populations and assemblage biomass and

259
productivity to variation in light, nutrients and herbivores. Canadian Journal of Fisheries and Aquatic Sciences, 57: 66-75. Sabattini, R. A. & V. H. Lallana (2007) Aquatic Macrophytes. En: The Middle Paraná River: Limnology of a Subtropical Wetland. M. H. Iriondo, J. C. Paggi & M. J. Parma (Eds.). Springer-Verlag, Berlin, Heidelberg, pp. 205-226. Sakshaug, E., A. Bricaud, Y. Dandonneau, P. G. Falkowski, D. A. Kiefer, L. Legendre, A. Morel, J. Parslow & M. Takahashi (1997) Parameters of photosynthesis: definitions, theory and interpretation of results. Journal of Plankton Research, 19: 1637-1670. Sand-Jensen, K. & J. Borum (1991) Interactions among phytoplankton, periphyton and macrophytes in temperate freshwaters and estuaries. Aquatic Botany, 41: 137-175. Schallenberg, M. & C. Burns (2004) Effects of sediment resuspension on phytoplankton production: teasing apart the influences of light, nutrients and algal entrainment. Freshwater Biology, 49: 143-159. Scheffer, M. 1998. Ecology of Shallow Lakes. Chapman & Hall, London, 357 p. Scheffer, M. & E. H. van Nes (2007) Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia, 196, 455-466. Scheffer, M., S. Szabó, A. Gragnani, E. van Nes, S. Rinaldi, N. Kautsky, J. Norberg, R. M. Roijackers & R. J. Franken (2003) Floating Plant Dominance as a stable state. Proceedings of the National Academy of Sciences of the United States of America, 100: 4040-4045. Scheffer, M., S. H. Hosper, M. L. Meijer, B. Moss & E. Jeppesen (1993) Alternative equilibria in shallow lakes. Trends in Ecology and Evolution, 8: 275-279. Schweizer, A. & R. E. Heusel (1992) Picoplankton photosynthesis and diurnal variations in photosynthesis-irradiance relationship in a eutrophic ans meso-oligotrophic lake. Hydrobiologia, 238: 131-138. Scott, J. T., R. D. Doyle & C. T. Filstrup (2005) Periphyton nutrient limitations and nitrogen fixation potential along a wetland nutrient-depletion gradient. Wetlands, 25: 439-448. Sculthorpe, C. D. (1967) The biology of aquatic vascular plants. Arnold, Londres, 610 p. Secretaría de la Convención de Ramsar (2006) Manual de la Convención de Ramsar: Guía a la Convención sobre los Humedales (Ramsar, Irán, 1971), 4ta. edición. Secretaría de la Convención de Ramsar, Gland (Suiza). Shigesada, N. & A. Okubo (1981) Analysis of the self-shading effect on algal vertical distribution in natural waters. Journal of Mathematical Biology, 12: 311-326.

260
Sieburth, J. Mc N., V. Smetacek & J. Lenz (1978) Pelagic ecosystem structure: heterotrophic compartments of the plankton and their relationship to size fractions. Limnology and Oceanography, 23: 1256-1263. Sinistro, R. (2007) Estructura de las comunidades planctónicas y análisis de sus interacciones tróficas en un humedal de la cuenca del Paraná inferior (Reserva de Otamendi, Buenos Aires, Argentina). Tesis Doctoral. Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 167 p. Sinistro, R., M. L. Sánchez, M. C. Marinone & I. Izaguirre (2007) Experimental study of the zooplankton impact on the trophic structure of phytoplankton and the microbial assemblages in a temperate wetland (Argentina). Limnologica, 37: 88-99. Sinistro, R., I. Izaguirre & V. Asikian (2006) Experimental study on the microbial plankton community in a South American wetland (Lower Paraná River Basin) and the effect of the light deficiency due to floating macrophytes. Journal of Plankton Research, 28: 753-768. Sokal, R. R. & F. J. Rohlf (1979) Biometría: Principios y métodos estadísticos en la investigación biológica. Blume Ediciones, Madrid, 832 p. Stanley, E. H., M. D., Johnson & A. K. Ward (2003) Evaluating the influence of macrophytes on algal and bacterial production in multiple habitats of a freshwater wetland. Limnology and Oceanography, 48: 1101-1111. Steeman Nielsen, E. (1952) The use of radioactive carbon (14C) for measuring organic production in the sea. J. Cons. Int. Explor. Mer., 18: 117-140. Steinberg, C. E., S. Kamara, V. Y. Prokhotskaya, L. Manusadžianas, T. A. Karasyova, M. A. Timofeyev, Z. Jie, A. Paul, T. Meinelt, V. F. Farjalla, A. Y. Matsuo, B. K. Burnison & R. Menzel (2006) Dissolved humic substances – ecological driving forces from the individual to the ecosystem level?. Freshwater Biology, 51: 1189-1210. Steitz, A. & B. Velimirov (1999) Contribution of Picocyanobacteria to total primary production and community respiratory losses in a backwater system. Journal of Plankton Research, 21: 2341-2360. Stenuite, S., S. Pirlot, M. Hardy, H. Sarmento, H. Tarbe, B. Leporcq & J. P. Descy (2007) Phytoplankton production and growth rate in Lake Tanganyica: evidence of a decline in primary productivity in recent decades. Freshwater Biology, 52: 2226-2239. Stockner, J. (1988) Phototrophic picoplankton: an overview from marine and freshwater ecosystems. Limnology and Oceanography, 33: 765-775. Stockner, J. & N. Antia (1986) Algal picoplankton from marine and freshwater: a multidisciplinary perspective. Canadian Journal of Fisheries and Aquatic Sciences, 43: 2472-2503.

261
Stockner, J., C. Callieri, & G. Cronberg (2000) Picoplankton and other non-bloom forming cyanobacteria in lakes. En: The Ecology of Cyanobacteria: Their Diversity in Time and Space. B. Whitton & M. Potts, (Eds.), Kluwer Academic Publishers, pp. 195-238. Stumm, W. & J. Morgan (1996) Aquatic Chemistry. Chemical Equilibria and Rates in Natural Waters. John Wiley and sons, Inc., New York, 1022 p. Talling, J. F. (1957) The phytoplankton population as a compound photosynthetic system. New phytologist, 56: 133-149. Tell, G. (1979) Chlorophyceae epífitas y epizoicas de agua dulce de Tierra del Fuego. Ecosur, 6: 1-23. Tell, G. (1977) Estudios ecológicos sobre algas epífitas de Ricciocarpus natans. Ecosur, 4: 117-135. Tell, G. (1973) "Cyanophyta" epífitas de las lagunas Chascomús, El Burro, Yalca y Vitel (Prov. Buenos Aires, Argentina). Darwiniana, 18: 129-152. Tell, G. & V. Conforti (1986) Euglenophyta pigmentadas de la Argentina, Biblioteca Phycologica 75. J. Cramer, Berlin, Stuttgart, 301 p. ter Braak, C. J. F. (1994) Canonical community ordination. Part I: Basic theory and linear methods. Ecoscience, 1: 127-140. ter Braak, C. J. F. (1990) CANOCO-a Fortran program for canonical community ordination by partial, detrended, canonical correspondence analysis, principal component analysis and redundancy analysis (Version 3.11). Agricultural Mathematics Group. Wageningen, The Netherlands, 95 p. ter Braak, C. J. F. (1987) Ordination. En: Data analysis in community and lanscape ecology. R. H. G. Jongman, C. J. F. ter Braak & O. F. R. van Tongeren (Eds.). Pudoc Wageningen, pp. 91-173. ter Braak, C. J. F & P. F. M. Verdonschot (1995) Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquatic Sciences, 57: 255-289. Tesolín, G. & G. Tell (1996) The epiphytic algae on floating macrophytes of a Paraná river floodplain lake. Hydrobiologia, 333: 111-120. Tilzer, M. (1984) La medición de la producción primaria en el fitoplancton. En: Embalses, Fotosíntesis y Producción Primaria. N. Bahamonde & S. Cabrera (Eds.). Alfabeta, Santiago, Chile, pp. 85-91. Torremorell, A., M. Llames, G. Pérez, R. Escaray, J. Bustingorry & H. Zagarese (2008) Annual patterns of phytoplankton density and primary production in a large, shallow lake: the central role of light. Freshwater Biology, 54: 437-449.

262
Train, S. & L. C. Rodrigues (1997) Temporal fluctuations of the phytoplankton community of the Baía River, in the upper Paraná River floodplain, Mato Grosso do Sul, Brazil. Hydrobiologia, 361: 125-134. Tranvik, L. J. (1988) Availability of dissolved organic carbon for planktonic bateria in oligotrophic lakes of different humic content. Microbial Ecology, 16: 311-322. Unrein, F. (2002) Changes in phytoplankton community along a transversal section of the Lower Paraná floodplain, Argentina. Hydrobiologia, 468: 123-134. Unrein, F. (2001) Efectos de los nutrientes y el pH sobre el crecimiento y la estructura del fitoplancton en ambientes de la llanura aluvial del Paraná Inferior. Tesis Doctoral. Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 175 p. Unrein, F. & G. Tell (1994) Respuesta del fitoplancton al enriquecimiento con fósforo y nitrógeno en aguas de una laguna de inundación del río Paraná. Revista de Hydrobiologia Tropical, 27: 315-328. Utermöhl, M. (1958) Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitteilungen der Internationale Vereinigung Theoretische und Angewandte Limnologie, 9: 1-38. Vadeboncoeur, Y. & A. Steinman (2002) Periphyton function in lake ecosystems. The Scientific World, 2: 1449-1468. Vadeboncoeur, Y. & D. M. Lodge (2000) Periphyton production on wood and sediment: substratum-specific response to laboratory and whole-lake nutrient manipulation. Journal of the North American Benthological Society, 19 (1): 68-81. Vadeboncoeur, Y. & D. M. Lodge (1998) Dissolved inorganic carbon sources for epipelic algal production: sensitivity of primary productions estimates to spatial and temporal distribution of 14C. Limnology and Oceanography, 42: 1222-1226. Vadeboncoeur, Y., G. Peterson, M. J. Vander Zanden & J. Kalff (2008) Benthic algal production across lake size gradients: Interactions among morphometry, nutrients and Light. Ecology, 89: 2542-2552. Vadeboncoeur, Y., E. Jeppesen, M. J. Vander Zanden, H. H. Schierup, K. Christoffersen, & D. M. Lodge (2003) From Greenland to green lakes: Cultural eutrophication and the loss of benthic pathways in lakes. Limnology and Oceanography, 48: 1408-1418. Vadeboncoeur, Y., M. J. Vander Zanden & D. M. Lodge (2002) Putting the lake back together: Reintegrating benthic pathways into lake food webs models. BioScience, 52: 44-53. Vadeboncoeur, Y., D. M. Lodge & S. R. Carpenter (2001) Whole-lake fertilization effects on distribution of primary production between benthic and pelagic habitats. Ecology, 82: 1065-1077.

263
van Dam, H., A. Merters & J. Sinkeldam (1994) A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Netherlands Journal of Aquatic Ecology, 28: 117-133. van den Hoek, C., D G. Mann & H. M. Jahns (1995) Algae. An introduction to phycology. Cambridge University Press, 623 p. van der Valk, A. (2006) The Biology of Freshwater Wetlands. Oxford University Press, 173 p. Venrick, E. L. (1978) How many cells to count? En: Phytoplankton Manual. A. Sournia (ed.). UNESCO, Paris, pp. 167-180. Vermaat, J. E. (2005) Periphyton dynamics and influencing factors. En: Periphyton: Ecology, Exploitation and Management. M. E. Azim, M. C. Verdegem, A. A. Van Dam & M. C. M. Beveridge (Eds.). CABI Publishing, Estados Unidos, pp. 35-49. Virdó, A. N. (1993) Reserva Natural Estricta Ingeniero Rómulo Otamendi: Lineamientos básicos para la formulación de su plan de manejo y una propuesta de gestión. Tesis de Licenciatura, Departamento de Geografía, Facultad de Filosofía y Letras, Universidad de Buenos Aires. Vollenweider, R. A. (1968) Scientific fundamentals of the eutrophication of lakes and flowing waters with particular reference to Nitrogen and Phosphorous as factors in Eutrophication. Paris, OECD, 192 pp. Vörös, L., P. Gulyás & J. Németh (1991) Occurrence, Dynamics and Production of Picoplankton in Hungarian Shallow Lakes. International Revue Ges Hydrobiologie, 76: 617-629. Welcomme, R. L. (1985) River fisheries. Food and Agriculture Organisation of the United Nations. Technical Paper 262, 330 p. Wetzel, R. G. (2005) Periphyton in the aquatic ecosystem and food webs. En: Periphyton: Ecology, Exploitation and Management. M. E. Azim, M. C. Verdegem, A. A. Van Dam & M. C. Beveridge (Eds.). CABI Publishing, Estados Unidos, pp. 51-67. Wetzel, R. G. (1983) Opening remarks. En: Periphyton of Freshwater Ecosystems. R. G. Wetzel (ed.). Dr. W. Junk Publishers, The Hague, Netherlands, 346 p. Wetzel, R.G. (1981) Limnología. Omega, Barcelona, 679 p. Wetzel, R. G. (1965) Techniques and problems of primary productivity measurements in higher aquatic plants and periphyton. Memorie dell´Istituto Italiano de Idrobiologia, 18: 249-267. Wetzel, R. G. & G. E. Likens (2000) Limnological Analyses, Third Edition. Springer, USA, 429 pp.

264
Williamsom, C., D. P. Morris, M. L. Pace & O. G. Olson (1999) Dissolved organic carbon and nutrients as regulators of lake ecosystems: Resurrection of a more integrated paradigm. Limnology and Oceanography, 44: 795-803. Zalocar de Domitrovic, Y. (1993) Fitoplancton de una laguna vegetada por Eichhornia crassipes en el valle de inundación del río Paraná (Argentina). Ambiente Subtropical 3: 39-67. Zalocar de Domitrovic, Y. (1992) Fitoplancton de ambientes inundables del río Paraná (Argentina). Revue Hydrobiologie Tropicale, 25: 177-188. Zalocar de Domitrovic, Y. (1990) Efecto de las fluctuaciones del nivel hidrométrico sobre el fitoplancton en tres lagunas isleñas en el área de la confluencia de los ríos Paraná y Paraguay. Ecosur, 16: 13-29. Zar, J. (1996) Biostatical analysis. Third edition. Prentice-Hall, Englewood Cliffs, New Jersey, 660 p.

ANEXO I
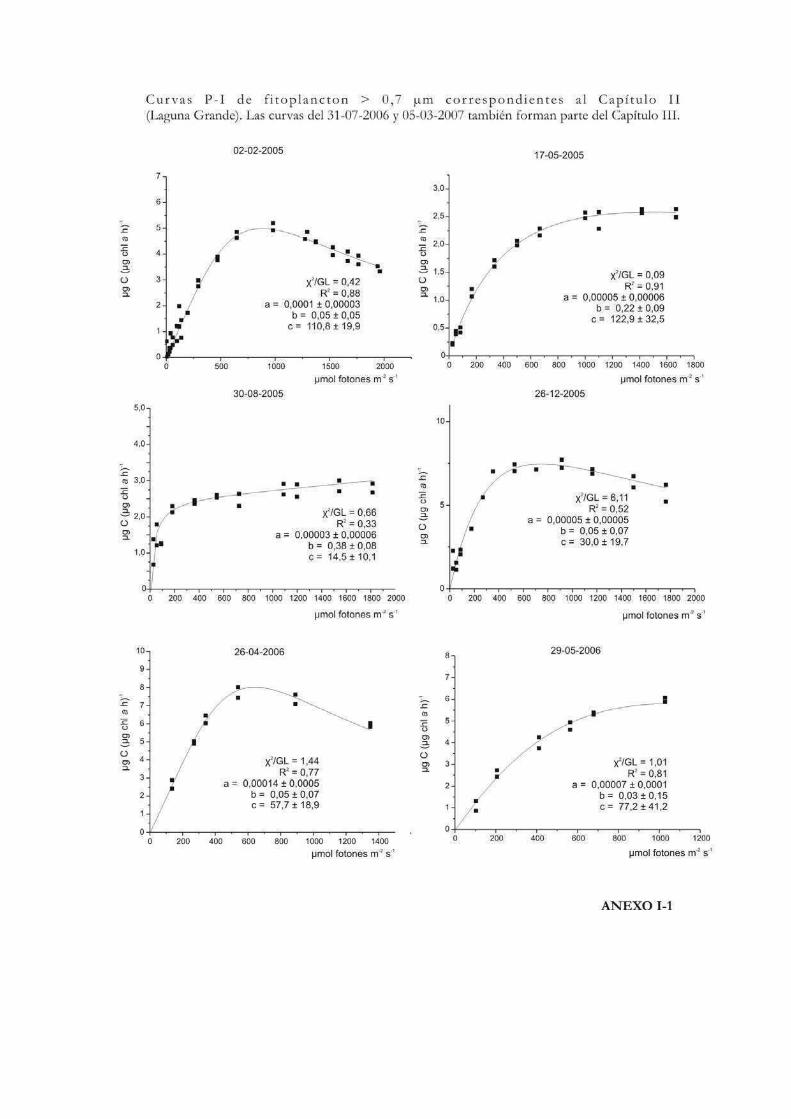
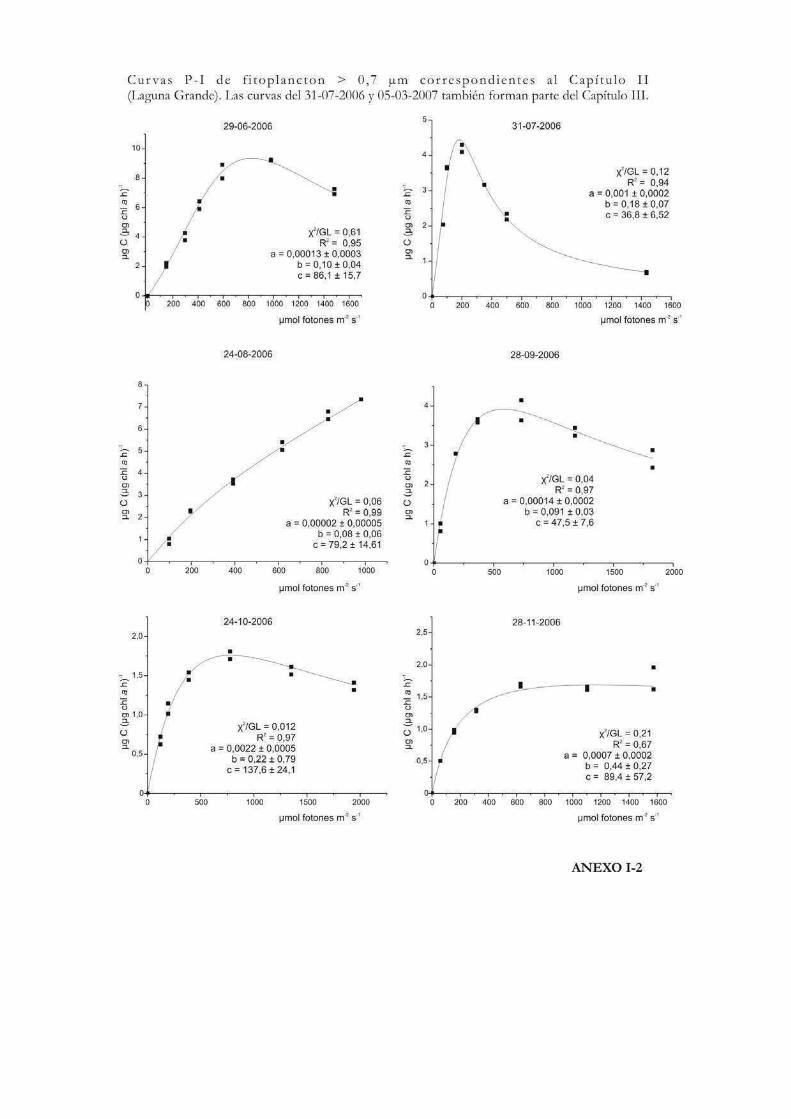
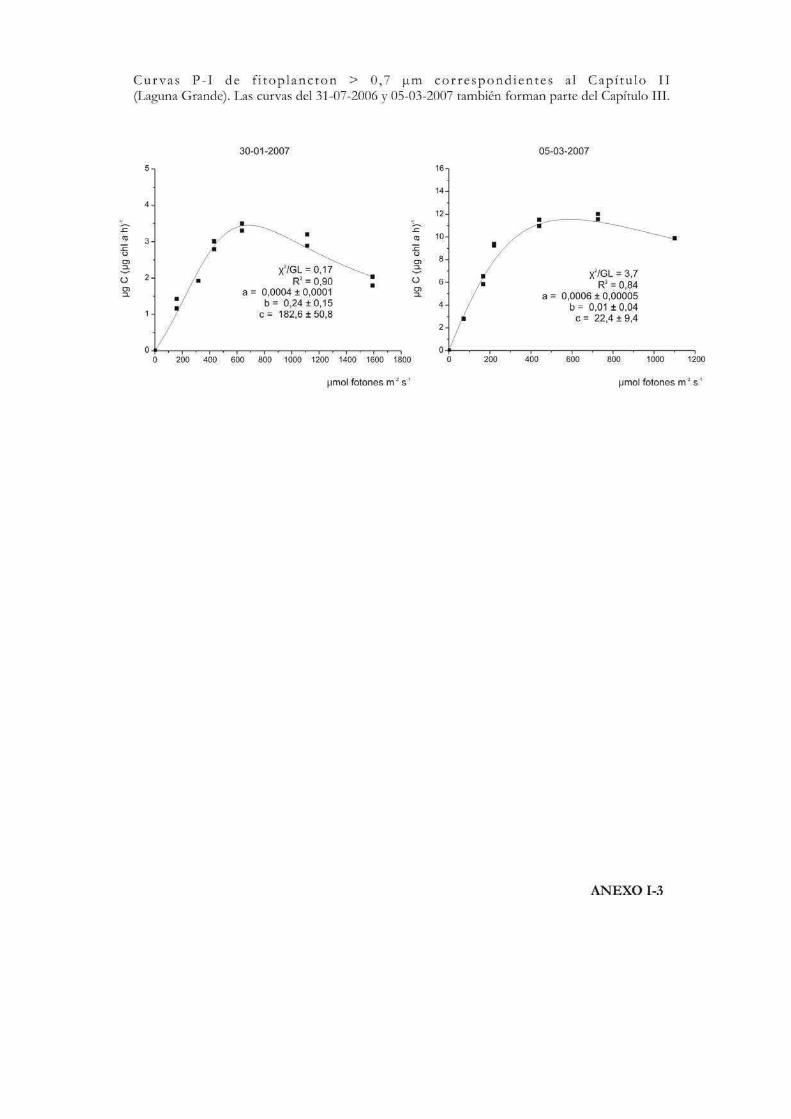
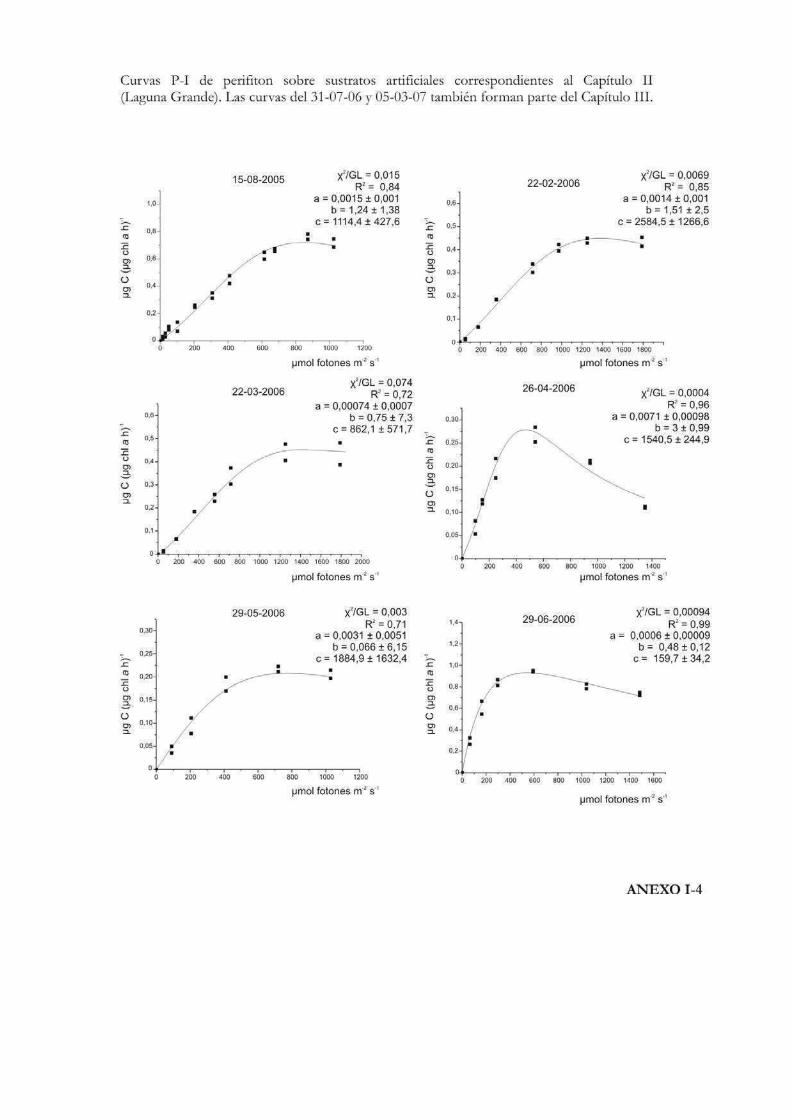
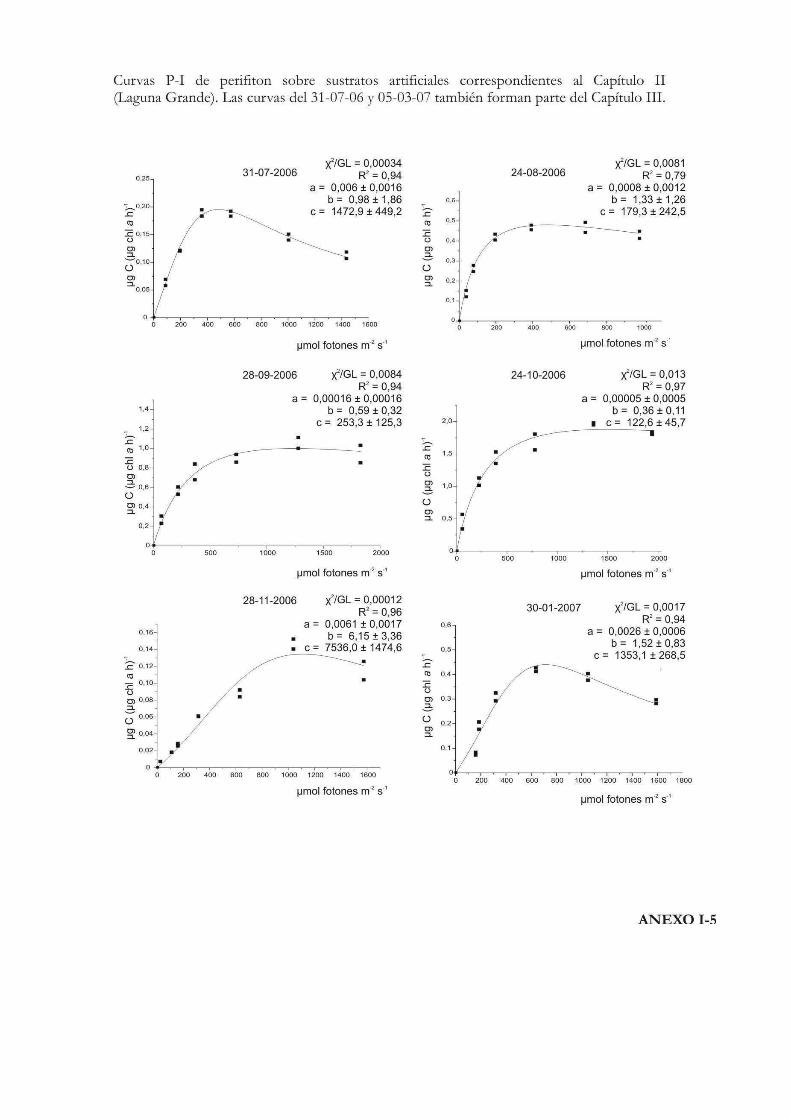
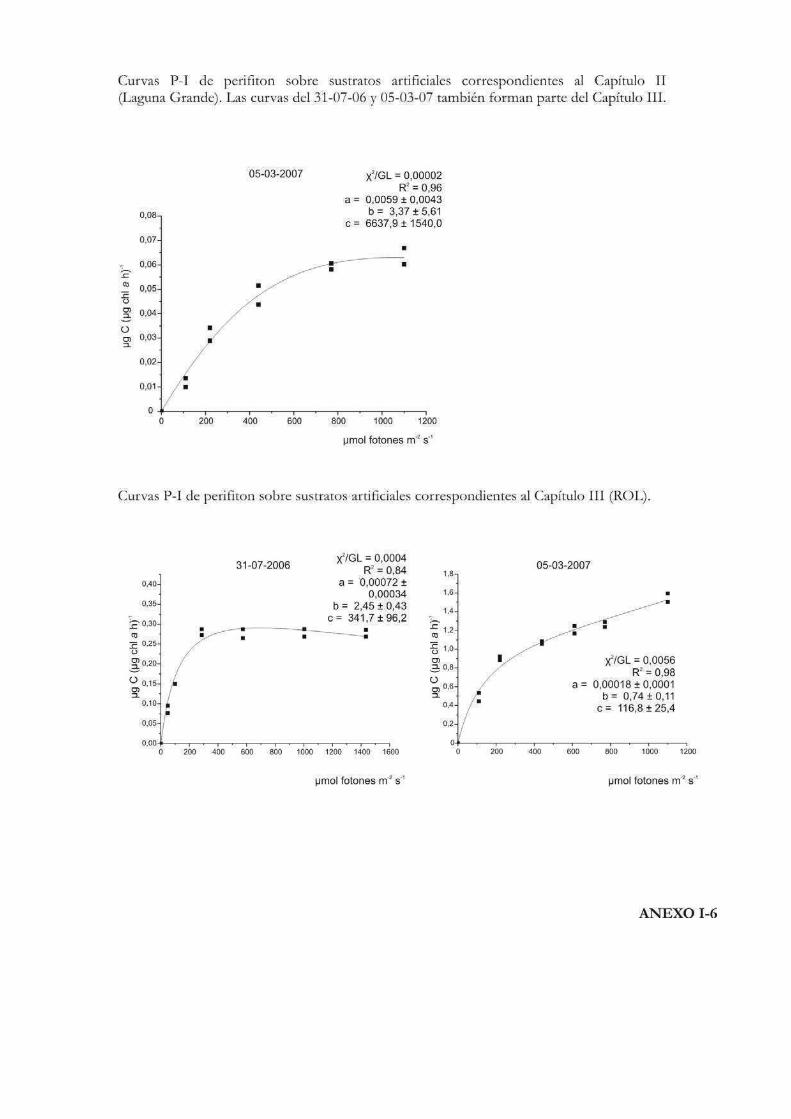
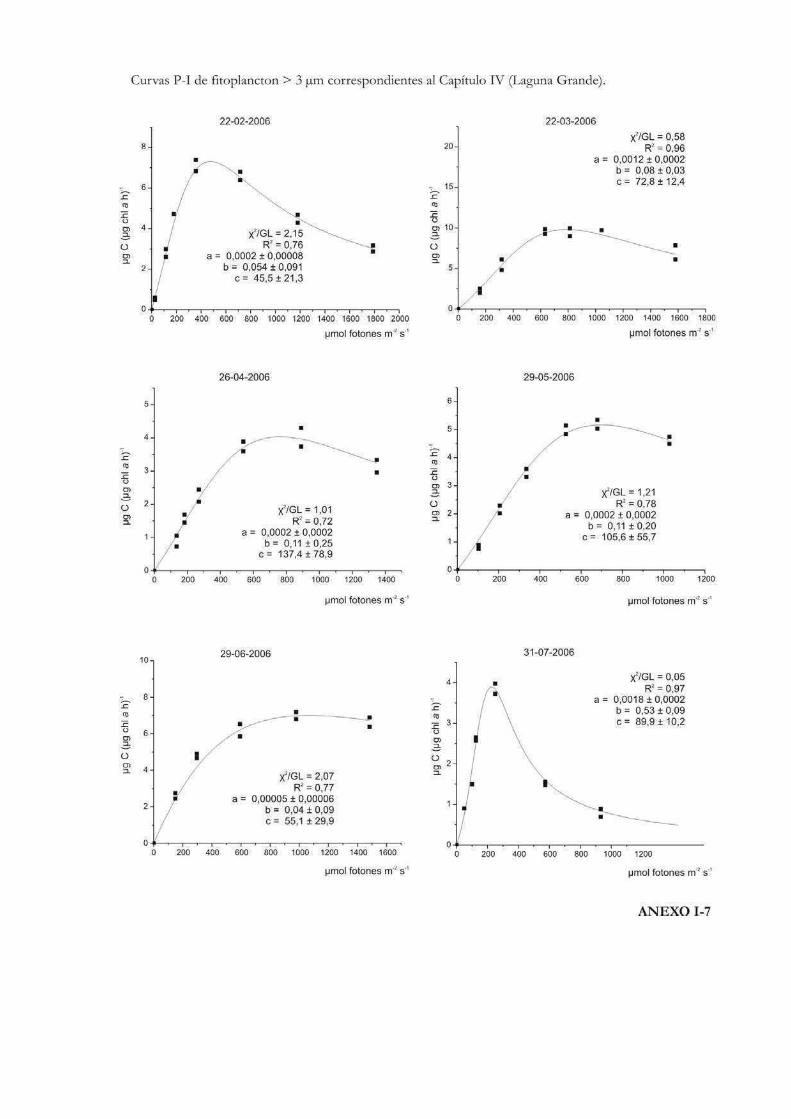
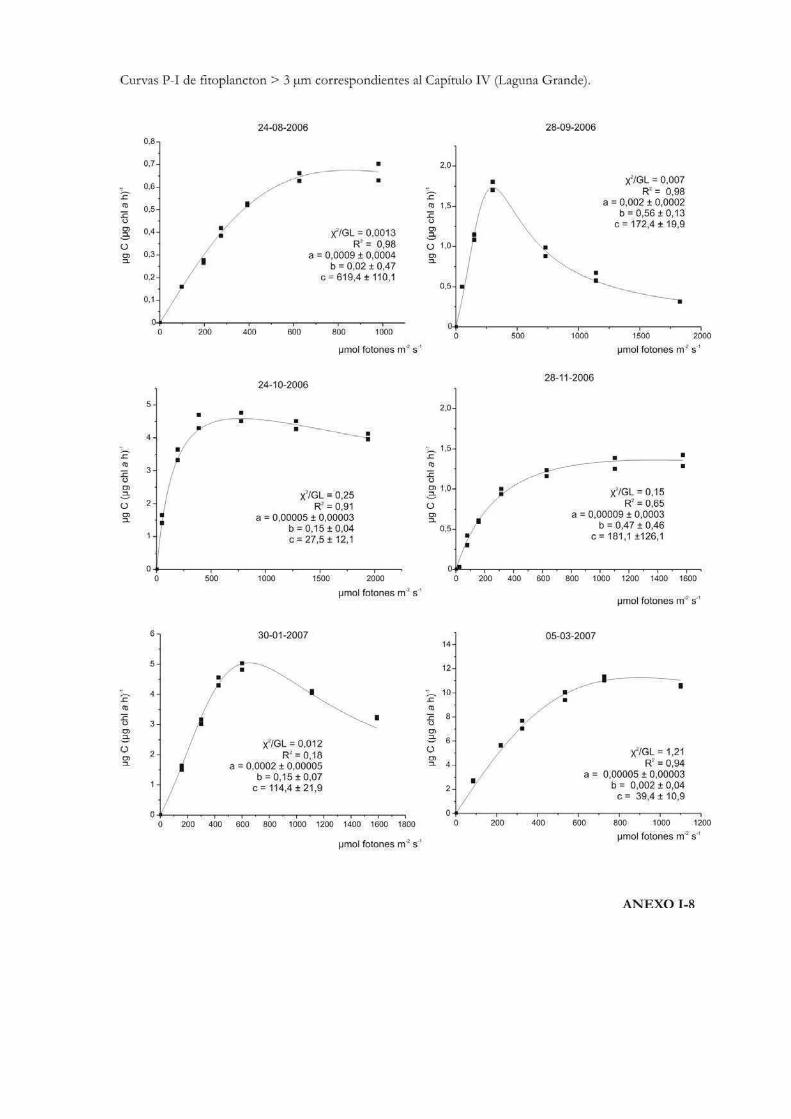
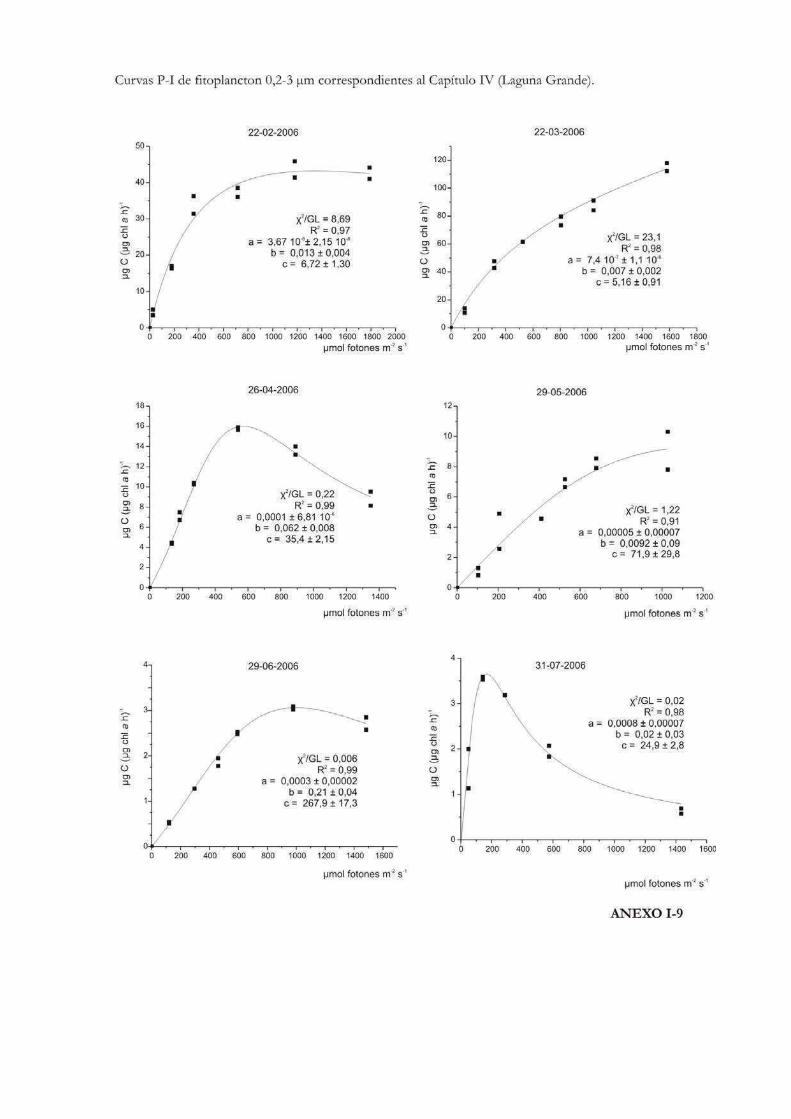
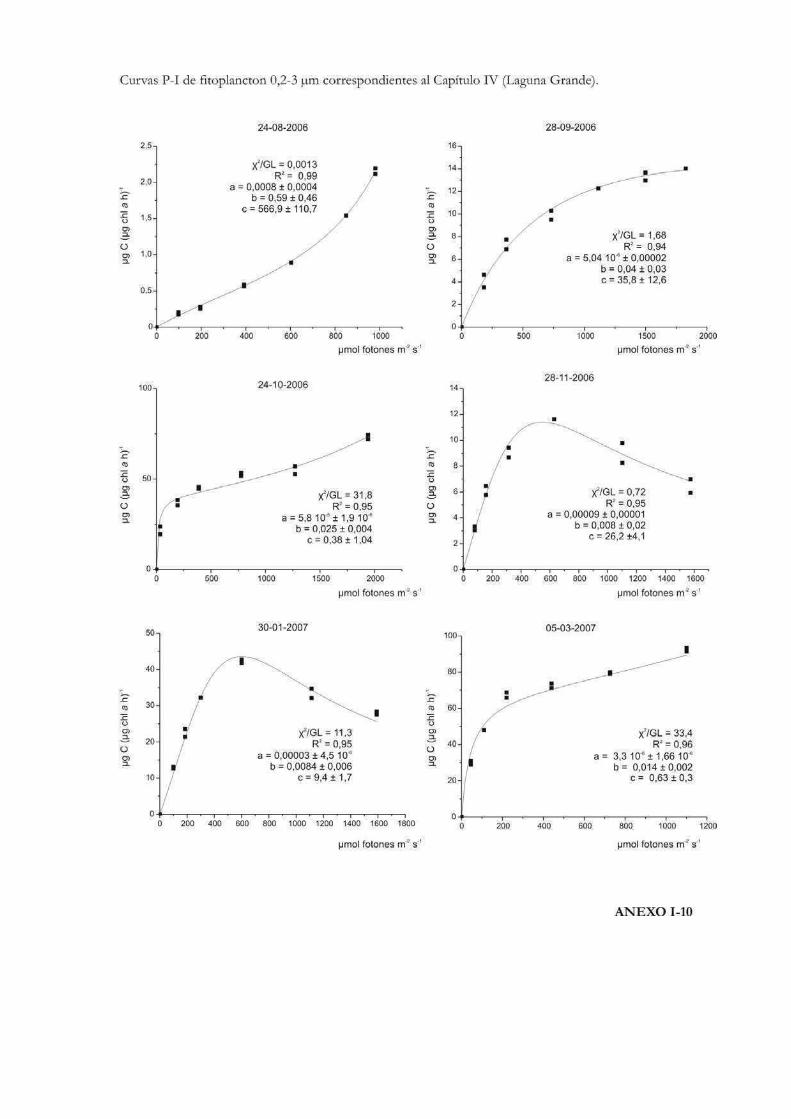
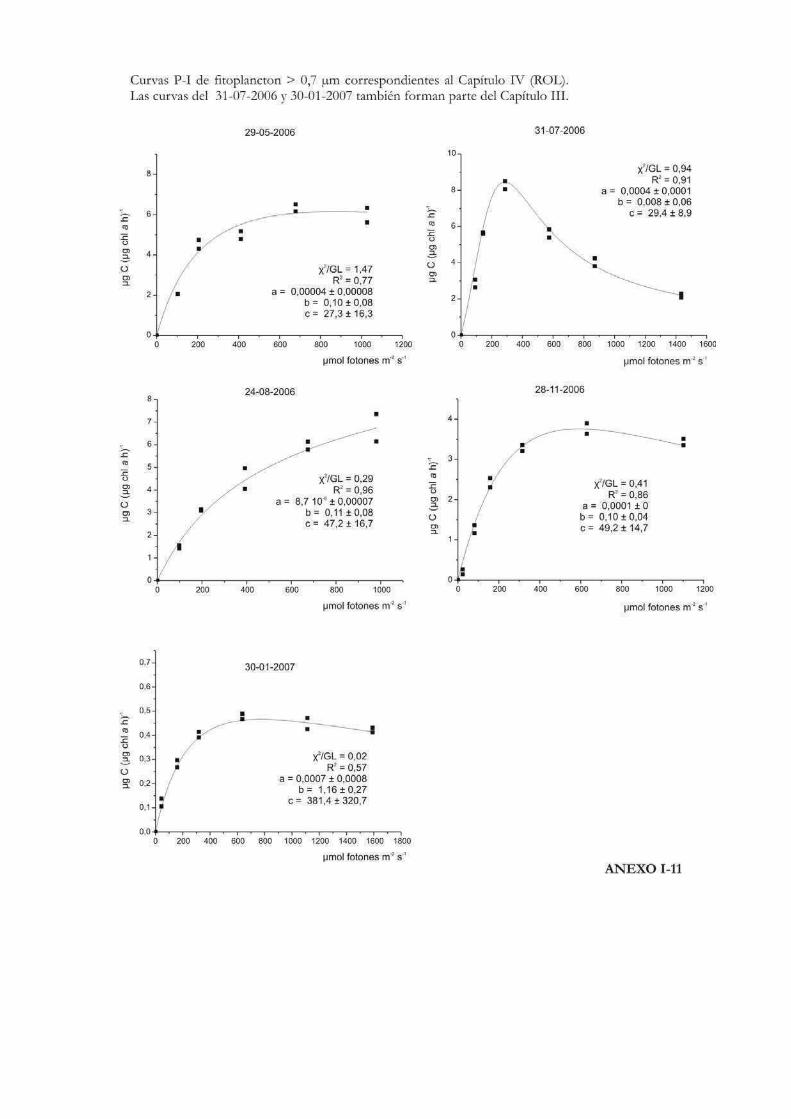
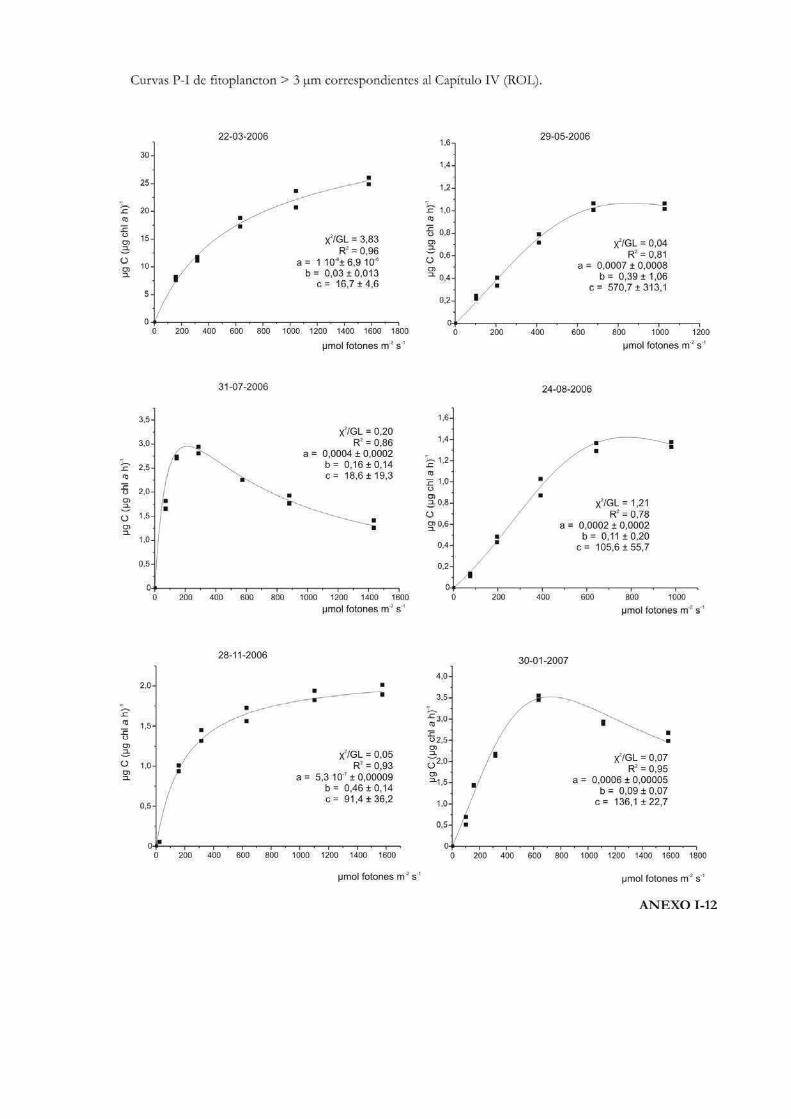
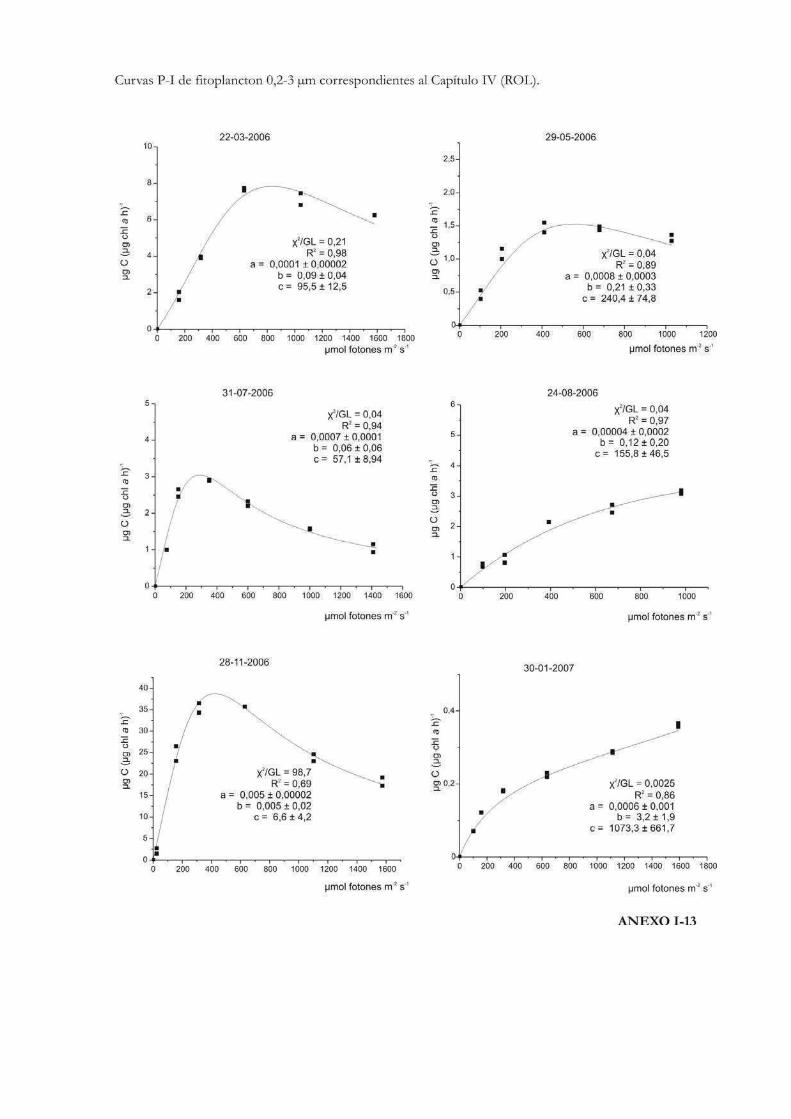
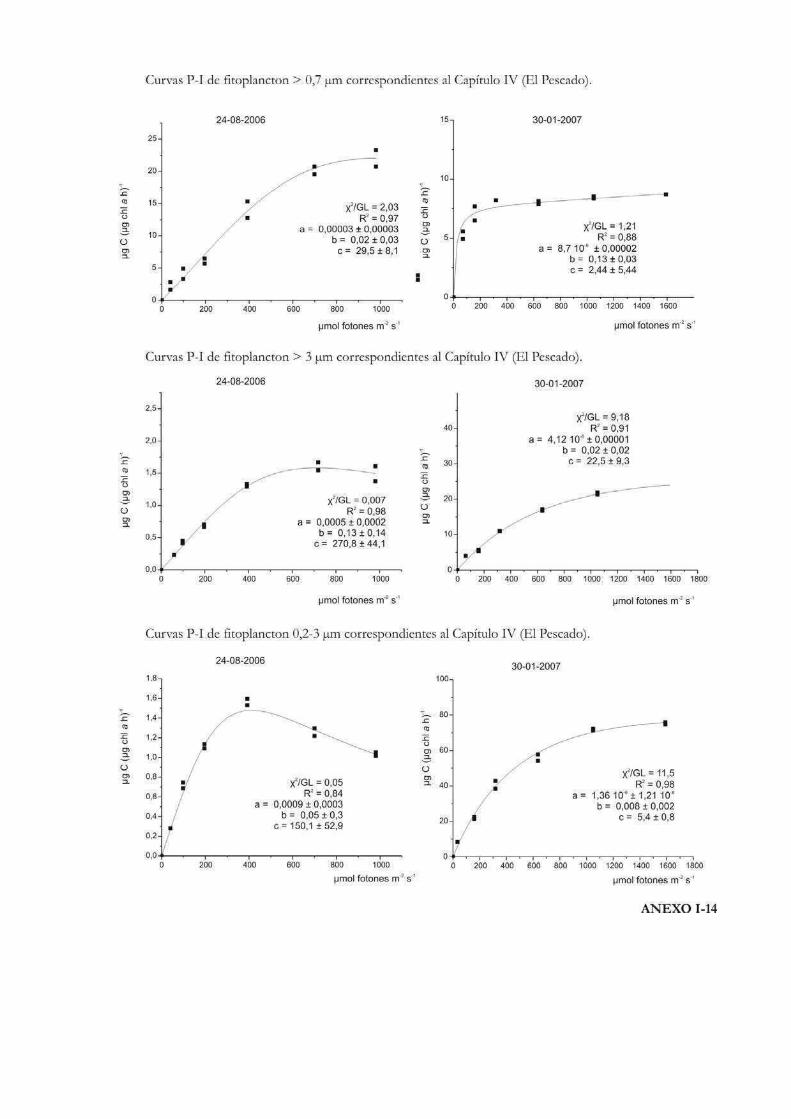

ANEXO II

Experimental evidence of the dynamic effect offree-floating plants on phytoplankton ecology
INES O’FARRELL* , †, PAULA DE TEZANOS PINTO* , †, PATRICIA L. RODRIGUEZ* , †,
GRISELDA CHAPARRO* AND HAYDEE N. PIZARRO* , †
*Departamento de Ecologıa, Genetica y Evolucion, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos
Aires, Argentina†Consejo Nacional de Investigaciones Cientıficas y Tecnicas (CONICET), Argentina
SUMMARY
1. There is an increasing concern to understand the role of free-floating plants (FFP) on the
functioning and structure of shallow lakes, particularly the mechanisms by which their
dominance is self-stabilizing and how they may outcompete phytoplankton.
2. In a field experiment with mesocosms, we simulated three commonly encountered
scenarios in warm temperate shallow lakes: FFP dominance, FFP fluctuation and FFP
absence. We explored the effects of several key processes, triggered by FFP dynamics, on
the composition, diversity and production of phytoplankton, and on the physicochemical
conditions.
3. The effects of persistent floating mats on light interception triggered a complex response
by the phytoplankton: species were lost and biomass was low, yet high diversity and
productivity potential were maintained. A contrasting phytoplankton response charac-
terized mesocosms lacking FFP, where light was sufficient but nitrogen was limiting.
Fluctuating FFP cover brought periodic shifts between these two limiting resources for the
phytoplankton, which most probably explain the maintenance of richness, diversity and
production in these shallow lakes.
4. These results support the recently proposed framework of alternative stable states,
providing experimental field evidence of the mechanisms resulting from the shifts
between floating plants and phytoplankton dominance.
Keywords: disturbance, free-floating plants, light regime, phytoplankton, shallow lake
Introduction
Many aspects of the functioning and structure of
shallow lakes depend on the presence of aquatic
plants, both in temperate and tropical zones (Moss,
1990; Jeppesen et al., 1997; Talling & Lemoalle, 1998;
Thomaz & Bini, 2003). The effects of submerged plants
on ecosystem functioning have been widely assessed
(Jeppesen et al., 1998; Scheffer, 1998), although surpris-
ingly the role of free-floating plants (FFP) has received
much less attention (Pompeo &Moschini-Carlos, 2003;
Meerhoff &Mazzeo, 2004; Bicudo et al., 2007; Meerhoff
et al., 2007a,b). However, there is increasing concern
about the effects of FFP because their geographical
range is likely to expand as a consequence of increasing
winter minimum air temperatures (Meerhoff, 2006)
and the accelerating eutrophication (Scheffer et al.,
2003) of freshwater ecosystems.
Lake ecosystems where FFP occur often fluctuate
between states of dense plant cover and their com-
plete absence; these states mainly depend on lake size
and morphometry, water level, nutrient concentration
Correspondence: Ines O’Farrell, Departamento de Ecologıa,
Genetica y Evolucion, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, C1428 EHA, Buenos Aires,
Argentina.
E-mail: [email protected]
Freshwater Biology (2009) 54, 363–375 doi:10.1111/j.1365-2427.2008.02117.x
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd 363

NOTE
PHYTOPLANKTON PRODUCTIVITY IN A HIGHLY COLORED SHALLOW LAKE
OF A SOUTH AMERICAN FLOODPLAIN
Patricia Rodrıguez and Haydee Pizarro
Laboratorio de Limnologıa Departamento de Ecologıa Genetica y Evolucion Facultad de
Ciencias Exactas y Naturales
Universidad de Buenos Aires
Piso 4 Pabellon 2, Ciudad Universitaria
C 1428 EHA Buenos Aires, Argentina
Consejo Nacional de Investigaciones Cientıficas y Tecnicas
E-mail: [email protected]
Abstract: We described phytoplankton productivity in a floodplain wetland of the Lower Parana River,
Argentina. Four samplings encompassing periods of high and low water levels were conducted in a highly
colored shallow lake. Photosynthesis-irradiance (P-E) curves and the areal photosynthetic rate (PA) were
estimated following the 14C assimilation technique. Likewise, physical and chemical variables and
phytoplankton composition, density, and chlorophyll a concentration were measured. Phytoplankton
assemblages shifted from cyanobacteria blooms in summer to cryptophycean dominance in winter, and
co-dominance of cryptophytes and chlorophytes in autumn. Assimilation number Pmax ranged from 3–
7.8 mg C (mg Chl a h)21 and peaked in early summer (low water level) when water color was highest,
suggesting that phytoplankton productivity was not depressed by the high content of colored humic
acids. Photosynthetic efficiency (a) ranged from 0.021–1 mg C (mg Chl a h)21mmol photons21 m2 s and
reached its maximum value during winter even when the assemblage, dominated by cryptophyceans, did
not achieve light saturation. In early and late summer and in autumn, optimal irradiance (Eopt) ranged
from 544–1,397 mmol photons m22 s21. The highest PA (207 mg C m22 h21) was registered in late
summer (high water level) when the lowest mean irradiance (Emean) was observed (341 mmol photons
m22 s21). The lowest PA (28 mg C m22 h21) occurred in winter when Emean was maximum (1,432 mmol
photons m22 s21). Our results indicate that productivity was similar to those recorded for other latitudes
and appeared not limited by the humic content of the water because phytoplankton was dominated by
algae well-adapted to low light conditions.
Key Words: areal photosynthetic rate, Argentina, humic wetland, P-E curves
INTRODUCTION
This study was conducted to describe some
functional aspects of a representative wetland of the
Lower Parana River Basin. The floodplain wetlands
of the Parana River are part of a large wetland
corridor, which originates in the swampland of Mato
Grosso, Pantanal. These wetland systems are impor-
tant reservoirs of fresh water and biodiversity for
large areas of South America. Studies are needed to
describe the structure and function of these wetlands
to enhance protection of these important systems.
South American floodplain wetlands are habitats
where water level fluctuations determine biogeo-
chemical fluxes, soils have accentuated hydromorph-
ism, and biota are well adapted to a wide range of
water availability (Neiff et al. 1994). Floodplains
along large rivers contain many shallow lakes with
different limnological properties including lagoons,
marshes, swamps, and relict oxbow lakes (Welcomme
1985). Some are vegetated with a well-developed
cover of free floating macrophytes. Strong variation
in water level of these wetlands occurs from seasonal
storm events (Junk et al. 1989).
In general, wetlands are rich in humic compounds
from autochthonous and allochthonous organic
matter. When plant tissue decomposes, most of the
organic matter is broken down by microbial action.
During the decomposition process, humic substances
are formed and impart a yellow-brownish color to
the water. Humic coloration has major consequences
for the absorption of light, particularly at the blue
end of the visible spectrum (Kirk 1994). Modifica-
tion of the light environment can impact algal
productivity. Since light permits a phototrophic
existence, availability and variation in the quantity
WETLANDS, Vol. 27, No. 4, December 2007, pp. 1153–1160’ 2007, The Society of Wetland Scientists
1153





























