Embed Size (px)
Citation preview

17
Lagascalia 13(1): 17-38 (1984)
ESTUDIO CARIOLOGICO DE BORAGINACEASESPAÑOLAS. II. ECHIUM L. DE ESPAÑA PENINSULAR
E ISLAS BALEARES
T. LUQUE
Departamento de Botánica, Facultad de Biología, Sevilla
(Recibido el 15 de junio de 1983)
Resumen. Se estudian cariológicamente los tazones del género Echium L. de España Penin-sular e Islas Baleares, excepto E. cantabricum (Laínz) Fernández Casas & Laínz. Cariológicamen-te los taxones estudiados se pueden separar en cuatro grupos, cuyos límites no coindicen con laclasificación propuesta por COINCY. Los resultados obtenidos basados en especies mediterráneasapoyan la idea de considerar x = 8 como número básico más primitivo del que han debido derivarx = 7, (6) y 5.
Summary. AH of the taxa of Echium L. from Peninsular Spain and the Balearic Islands, withthe exception of E. cantabricum (Laínz) Fernández Casas & Laínz, have been studied caryologica-Ily. The caryological characters do not allow the separation of four groups proposed by COIN(
The data obtained allow to consider x = 8 as the most primitive base number from which x = 7(6)and 5 shoiild derive.
INTRODUCCION
Dentro de Boraginaceae, Echium y géneros relacionados fueron separa-dos por DE CANDOLLE (1846) como tribu Echieae, idea que es compartida porGORKE (1893), POPOV (1953) y BRAMWELL (1978), aunque dicha tribu se haconsiderado tradicionalmente integrada en Lithospermeae desde que fuepropuesta su inclusión por BENTHAM & HOOKER (1876).
Se trata de un género que comprende aproximadamente unas 50 especies,distribuidas por la Región Mediterránea, Islas Atlánticas y Africa. En EspañaPeninsular e Islas Baleares se encuentra representado por 19 taxones, tres deellos endémicos (E. albicans Lag. & Rodr., E. salmanticum Lag. y E. flavumDesf.).

18
Desde el punto de vista cariológico, fue STREY (1931) el primero que alparecer estudió taxones de este género. LITARDIÉRE (1940-41, 1943), BRITTON
(1951), GADELLA 8i al. (1966), DAHLGREN & al. (1971), FERNANDES & LEITÁO
(1972), CARDONA (1973), FRITSCH (1973), FERNÁNDEZ CASAS (1977a, 1977b,1978, 1979), FERNÁNDEZ CASAS & al. (1980), CARDONA & CONTANDRIOPOU-
LOS (1980) y LUQUE (1980), se han ocupado del estudio de algunos taxones re-presentados en la Península Ibérica.
En el presente trabajo se han estudiado cariológicamente todos los taxo-nes salvo E. cantabricum (Laínz) Fernández Casas & Laínz, del que no se hanpodido conseguir plantas vivas, pero que ha sido estudiado recientemente porFERNÁNDEZ CASAS (1979: 11). Siempre que ha sido posible se ha indicado elnúmero cromosómico, longitud aparente y morfología de los cromosomas, asícomo fórmula idiogramática y asimetría que corresponde a cada cariotipo.
MATERIALES Y METODOS
Se ha realizado tanto el estudio de meiosis como de mitosis, para lo cuálse han estudiado plantas de origen silvestre correspondientes a 91 poblaciones.Para el estudio de meiosis, se han utilizado botones florales fijados en elcampo en líquido de Farmer (LOvE & LCIVE, 1975: 141) durante 24 horas y sehan conservado en alcohol etílico a 70%. Para el estudio de mitosis, se hanutilizado meristemos radicales de plantas obtenidas a partir de semillas y plán-tulas cultivadas en el Jardín Experimental del Departamento de Botánica de laFacultad de Biología, de la Universidad de Sevilla, los cuales fueron tratadoscon 8-hidroxiquinoleína 0,002 M (Dio & LEVAN, 1950), a 4 ± 2°C durante 3 -3,30 horas, posteriormente se siguió el mismo procedimiento que con losbotones florales.
La tinción, en ambos casos, se efectuó con carmín-alcohólico-clorhídrico(SNow, 1963) manteniéndose en dicho colorante entre 24 y 48 horas los meris-ternos radicales y hasta 96 horas los botones florales. Seguidamente se proce-dió a su preparación, que se ha realizado por aplastamiento y como líquido demontaje se ha utilizado ácido acético al 45%, y en caso de material escasamen-te teñido, carmín-acético (LOvE & LCIVE, 1975: 150).
Para los estudios de mitosis se han estudiado una media de 6 plantas porpoblación, mientras que para meiosis una media de 10 flores por planta. Lasplantas utilizadas se conservan en el Herbario del Departamento de Botánica

19
de la Facultad de Biología de Sevilla (SEV) y se hace referencia a ellas por elnúmero de pliego con que se conservan en dicho herbario.
Se ha utilizado la terminología propuesta por STEBBINS (1938, 1971) paraexpresar el tamaño aparente de los cromosomas y el grado de asimetría de loscariotipos, y para indicar la morfología de los cromosomas se ha seguido aLEVAN & al. (1965).
RESULTADOS
E. albicans Lag. & Rodr., Anal. Ci. Nat. 5: 269 (1802).
Material estudiado.Cádiz. Grazalema, 13.VII.1978, Devesa, Rivera & Valdés (SEV 64937), n = 8 y 2n = 16.
Granada. Subida al Monte Trevenque, 20.V11.1978, Candau & Luque (SEV 64935), n = 8. Mála-ga. Atajate, 10.V11.1978, Candau, Díez & Luque (SEV 64938), n = 8 y 2n = 16; ídem, Ronda,subida a San Pedro de Alcántara, 16.VI.1979, Malato, Luque & al. (SEV 60903), n = 8; ídem,Ronda, 13.V.1979, García (SEV 64936), n = 8.
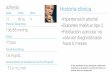
Se ha encontrado el número gamético n = 8 (Lám. I, fig. lb) y el zigótico2n = 16. Este último coincide con el indicado por FERNÁNDEZ CASAS (1977a:40) para plantas procedentes de Sierra de Cázulas y del Puerto del Lobo (Gra-nada). Se trata de un taxón diploide con número básico x = 8.
La longitud aparente de los cromosomas, de las plantas procedentes deAtajate (Málaga, SEV 64938) oscila entre 1,30 y 1,63 ,um, siendo por tanto pe-queños (Lám. I, fig. la). Dichos cromosomas pueden agruparse como sigue(Lám. I, fig. 1c): 6 metacéntricos (M), con centrómero en el punto medio (pares3, 5 y 6); 6 metacéntricos (m), con centrómero en la región media (pares 1, 4 y7); 2 submetacéntricos (sm), con centrómero en la región submedia (par 8) y 2subtelocéntricos (st) con centrómero en la región subterminal (par 2).
A este cariotipo corresponde la fórmula idogramática siguiente: 6M +6m + 2sm + 2st. Su asimetría cromosómica es de tipo 2A.
E. buinile Desf., FI. Atl. 1: 165 (1798).
Material estudiado.Alicante. Muchamiel, 6.V.1979, García, Luque & Valdés (SEV 60888), n = 8; ídem, entre
Alicante y Benidorm, cerca de Villajoyosa, 6.V.1979, García, Luque & Valdés (SEV 64953), n = 8;ídem, Villajoyosa, 6.V.1979, García, Luque & Valdés (SEV 60889), n = 8. Murcia. Aguilas,10.1V.1979, Devesa, Luque & Ubera (SEV 60887), n = 8.

20
El número cromosómico encontrado, n = 8, coincide con el indicado porDJERDJOUR & GUITTONNEAU (1977: 269) para plantas procedentes deArgelia. Se trata, al parecer, del primer estudio efectuado con material espa-ñol.
Todas las meiosis estudiadas han sido normales, con formación de 8 biva-lentes y emigración regular en anafase I (Lám. I, fig. 2).
No ha sido posible estudiar metafases somáticas, por lo que se desconoceel tamaño y morfología de los cromosomas, así como la fórmula idiogramáticay grado de asimetría que corresponde al cariotipo de este taxón.
E. asperrimum Lam., Tabi. Encycl. Méth. Bot. 1: 412 (1792).
Material estudiado.Alicante. Pilar de la Horadada, 5.V.1979, García, Luque & Valdés (SEV 64940), n = 7 y
2n = 14. Baleares. IBIZA. Santa Eulalia, 6.VI.1981, Deve,sa, Luque & Romero (SEV 64939), n = 7.Ciudad Real. Villanueva de la Fuente, 30.V.1980, Luque, Ubera & Valdés (SEV 64942), n = 7.Jaén. Castril, 20.V1.1979, Díez, Ubera & Valdés (SEV 60960), n = 7. Murcia. San Javier,5.V.1979, García, Luque & Valdés (SEV 64944), n = 7 y 2n = 14. Navarra. Yesa, Monasterio deLeyre, 17.VII.1980, Devesa, Luque & al. (SEV 64943), n = 7. Valencia. Fontanares, 2.VI.1978,Devesa, Pastor & Valdés (SEV 60958), n = 7. Zaragoza. Zuera, Estación de Ontiflar, 13.V11.1979,Cabezudo, Luque & (Ibera (SEV 64941), n = 7.
En las plantas estudiadas se ha encontrado n = 7 y 2n = 14. El númerodiploide encontrado coincide con el indicado por LITARD1ÉRE (1940-41: 320;1943: 157, sub E. hispidissimum R. Lit.) para plantas procedentes de Francia,Túnez y Argelia, por DELAY (1969: 8, sub E. pyrenaicum Desf.) para plantasfrancesas y por VAN LOON & SNELDERS (1979: 633) para plantas procedentesde Grecia. LITARDIÉRE (1.c.) encontró además, en las poblaciones proceden-tes de Túnez y Argelia, individuos con raíces mixoploides que presentaban a lavez células diploides (2n = 14) y tetraploides (2n = 28), lo que no ha sido con-firmado por ningún otro autor. Se trata, en cualquier caso, ele un taxóndiploide con número básico x = 7.
En todas las plantas estudiadas en meiosis, se ha comprobado que de los7 bivalentes formados tanto en profase como en metafase I, uno de ellos esnotablemente más grande que los demás (Lám. I, fig. 4b). Esta diferencia detamaño y grosor de los cromosomas, se mantiene en las metafases somáticasestudiadas, ya que de los 14 cromosomas, 12 presentaban un tamaño inferiora la mitad de los dos más grandes (Lám. I, fig. 4a), como fué ya observadopor LITARDIÉRE (1.c.) y DELAY (1.C.)..

21
La longitud aparente de los cromosomas de las plantas procedentes deAlicante (SEV 64940), oscila entre 1,10 y 2,95 ,um, siendo, por tanto, peque-ños y medianamente pequeños, perteneciendo a este último grupo solamentelos dos cromosomas más grandes anteriormente comentados. Dichoscromosomas se pueden agrupar de la siguiente forma (Lám. I, fig. 4c): 2 meta-céntricos (M), con centrómero en el punto medio (par 4); 6 metacéntricos (m),con centrómero en la región media (pares 3, 5 y 7); 4 submetacéntricos (sm),con centrómero en la región submedia (pares 2 y 6) y 2 subtelocéntricos (st),con centrómero en la región subterminal (par 1).
A este cariotipo corresponde la fórmula idiogramática: 2M + 6m + 4sm+ 2st, y su asimetría cromosómica es de tipo 2B.
LITARDIÉRE (1943: 164-165) indicó la presencia de un par de cromosomassatelizados en las células diploides y de dos pares en las tetraploides, que corres-pondían a los más grandes del cariotipo, lo que no ha sido observado en elpresente estudio.
E. italicum L., Sp. Pl. 139 (1753).
Material estudiado.
Baleares. Mallorca. Entre Soller y Pollensa, 8,.VI.1981, Devesa, Luque & Romero (SEV63293), n = 8; ídem, Mallorca, Pollensa, 8.VI.1981, Devesa Luque & Romero (SEV 63297), n = 8.
El número encontrado n = 8, coincide con el indicado por ARYAVAND(1977: 443) para plantas, procedentes de Irán. Está de acuerdo igualmente conel somático 2n = 16, indicado por LITARDIÉRE (1943: 157) para materialcultivado y procedente de Rumania, Alpes Marítimos y Córcega, porPODLECH & DIETERLE (1969: 220) para plantas de Afganistán, por HINDAKO-vÁ (1970: 52) para plantas procedentes de Checoslovaquia, por MARKOWA &IVANOVA (1971a: 124) para plantas recolectadas en Bulgaria, por STRID(1971: 491) para plantas procedentes de Albania, por CARDONA (1973: 10)para plantas españolas recolectadas en la Montaña del Toro (Menorca, Balea-res), por FRITSCH (1973: 451) y por KLIPHUIS (1977: 268) para plantas deHungría, por UHRIKovA & MAJOVSKY (1978) para plantas deChecoslovaquia, por KLIPHUIS & WIEFFERING (1979: 399) para plantas pro-cedentes de Rusia y por STRID & FRANZEN (1981: 838) para plantas recolecta-das en Grecia.
GADELLA & al. (1966: 485) indicaron 2n = 32 para plantas españolasrecolectadas entre Lanjarón y Orjiva (Granada) y cerca de Sagunto (Valen-cia). CARDONA (1. c.) indica que debe tratarse de una mala determinación por

22
E. vulgare var. argentae. Al menos las plantas de la primera localidad no pue-den pertenecer a E. italicum, ya que el área de distribución de este taxón nollega hasta Granada.
Como en E. asperrimum, de los ocho bivalentes que regularmente apare-cen en profase, uno de ellos presenta un tamaño muy superior a los demás,como puede apreciarse en la diacinesis de la Lám. I, fig. 3.
MARKOWA & IVÁNOVÁ (1971a: 125-126) dedujeron el cariotipo de E.italicum. Por los datos de estos autores se ha podido obtener la longitud apa-rente de sus cromosomas, que oscila entre 1,5 y 3,7m, por lo que se conside-ran entre pequeños y medianamente pequeños, siendo de este último grupo so-lamente dos. Estas medidas coinciden con las obtenidas a partir del idiogramarepresentado por FRITSCH (1973: 453), del que se ha deducido la siguientefórmula idiogramática: 4M + 8m + 4sm. La asimetría es de tipo 2B.
Estos autores, así COMO LITARDIÉRE (1943: 164) y CARDONA (1973: 10),indican que en el cariotipo de este taxón se encuentran dos cromosomas detamaño muy superior a los restantes, lo que confirma las observaciones demeiosis anteriormente expuestas. Este par de cromosomas más grandes estásatelizado. De acuerdo con LITARDIÉRE (1.c.) y FRITSCH (1.c.) uno de los pa-
res de cromosomas más pequeños se encuentra i gualmente satelizado. En lascélulas tetraploides de los individuos mixoploides observados por LITARDIÉRE(1. c.) y como cabría esperar, se encuentran cuatro cromosomas satelizadosde mayor tamaño.
E. italicum y E. asperrimum han sido confundidos frecuentemente, con-fusión que no es posible en base al número cromosómico 2n = 16 en el prime-ro y 2n -= 14 en el segundo. La presencia de un par de cromosomas de tamañomuy superior a los demás, caracteriza, sin embargo, a ambos taxones, y losdistingue claramente de los demás.
E. lusitanicum L., Sp. Pl. 140 (1753).
Material estudiado.Orense. Sierra del Invernadero, 20.V1.1978, Devesa & Pastor (SEV 60912), n = 8.
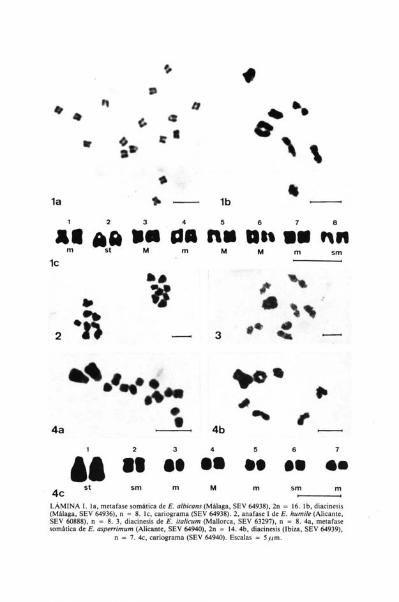
La población estudiada presenta n = 8, número que coincide con el indi-cado por BRITTON (1951: 241) para material cultivado y por LUQUE (1980:668) para plantas recolectadas entre Porto-Alegre y Castelo de Vide (AltoAlentejo, Portugal). Concuerda con el número somático 2n = 16 indicadopor STREY (1931: 718), LITARDIÉRE (1943: 157) y BRITTON (I.C.) para mate-rial cultivado y por FERNANDES & LEITÁO (1972: 392) para plantas de Portu-
4ae
4b
•5 6 7
11/10 e lie
SM m M m sm
2
n1113
ae111
•abe
`It
*s.
e II
•la - lb
24 6 73 5 8
Ami al Ois PA no ah eln
m m m m smlc
23 .* #
gia
LÁMINA I. la, metafase somática de E. albicans (Málaga, SEV 64938), 2n = 16. lb, diacinesis(Málaga, SEV 64936), n = 8. le, cariograma (SEV 64938). 2, anafase I de E. humile (Alicante,SEV 60888), n = 8. 3, diacinesis de E. italicum (Mallorca, SEV 63297), n = 8. 4a, metafasesomática de E. asperrimum (Alicante, SEV 64940), 2n = 14. 46, diacinesis SEV 64939),
n = 7. 4c, cariograma (SEV 64940). Escalas = 5 um.
•• 4: •,*re lo• •
2
#
N4c mO
3 4 5
.. • ."ti 4. .
• *
a3 •o • a
tle •1 "/ Zeia / ab w 1,
3a 111 3b
3c 12g U s. st s.
4a 4b
Lám. II. 1, anafase 1 de E. lusitanicum (Orense, SEV 60912), n = 8. 2, diacinesis de E. salmanri-cum (Cáceres, SEV 60913), n = 8. 3a, metafase somática de E. 17avum (Avda, SEV 64952). 2n =16. 3b, anafase 1 (Jaén, SEV 64951), n = 8. 3c, cariograma (SEV 64952). 4a, metafase somáticade E. boissieri (Málaga, SEV 64945). 2n = 10. 4b, metafase 1 (Jaén, SEV 64947), n = 5. 4c,
cariograma (SEV 64945). Escalas 5 m.

23
gal. Se trata de un taxón diploide con número básico x = 8 del que al pareceres la primera vez que se estudia material español.
Se ha observado gran regularidad en todas las fases de la meiosis (Lám.II, fig. 1). No habiendo sido posible estudiar metafases somáticas. FERNÁNDES& LEITA0 (1972: 392) indicaron que en el cariotipo de esta especie se encuentrandos pares de cromosomas satelizados.
E. salmanticum Lag., Gen. Sp. Nov. 10 (1816).
Material estudiado.Avila. El Barco de Avila, Los Llanos, 16.VI.1981, Talavera & Valdés (SEV 63317), n = 8.
Cáceres. Entre Jerte y Tornavacas, 27.V11. 1978, Devesa, Pastor & Talavera (SEV 60913), n = 8.
En las plantas de las dos poblaciones estudiadas, se ha encontrado el nú-mero haploide n = 8. Al parecer, es la primera vez que se estudia cariológica-mente dicha especie, endémica del CW de España. Se trata de un taxóndiploide, con número básico x = 8. La meiosis es normal, habiéndose obser-vado la presencia de 8 bivalentes en diacinesis (Lám. I, fig. 2).
No se ha podido estudiar mitosis somáticas, por lo que se desconoce tan-to el tamaño como la morfología de sus cromosomas y, por tanto, la fórmulaidiogramática y asimetría de su cariotipo.
E. flavum Desf., Fi. Atl. 1: 165 (1798).
Material estudiado.Albacete. Laderas del río Mundo. Fábricas de Ri6par, 9.V.1979, García, Luque & Valdes
(SEV 60954), n = 8. Avila. Puerto de Menga, 28.VII.1978, Devesa, Pastor & Talavera (SEV64952), 2n = 16. Granada. Sierra Nevada, Peñones de San Francisco, 21.VII.1978, Candau & Lu-
que (SEV 60956), n = 8. Jaén. Sierra de Cazorla, Pico Cabañas, 1850 m.s.m., 24.VI.1980, Deve-
sa, Luque & al. (SEV 64951), n = 8. Málaga. Subida a Benahavís, 28.IV.1978, Luque & Valdés
(SEV 64950), n = 8.
El número haploide encontrado, n = 8, coincide con el indicado porFERNÁNDEZ CASAS (1978: 47) para plantas procedentes de La Uña (Cuenca),y el número somático (2n = 16) con el indicado por LITARDIÉRE (1943: 157)para plantas recolectadas en Argelia.
Este último autor encontró algunos individuos con raíces mixoploidescon células diploides (2n = 16) y tetraploides (2n = 32).
El número cromosómico encontrado difiere, sin embargo, del diploide2n = 10 indicado por FERNÁNDEZ CASAS (1977b: 341) para plantas españolasprocedentes del Pico Padrón de Bienservida en la Sierra de Alcaraz

24
(Albacete). Por los datos expuestos, E. flavum es un taxón diploide connúmero básico x = 8.
Se ha comprobado que en meiosis se forman 8 bivalentes en profase y me-tafase I, siendo igualmente regular la emigración de los cromosomas en anafa-se I (Lám. II, fig. 3b).
Las plantas procedentes de Avila (SEV 64952) presentan cromosomascon una longitud aparente que oscila entre 2,15 y 3,44 ,um (Lám. II, fig. 3a),siendo, por tanto, medianamente pequeños. Estos cromosomas pueden agru-parse de la siguiente manera (Lám. II, fig. 3c): 2 metacéntricos satelizados (M)con centrómero en el punto medio (par 6); 2 metacéntricos (m), con centróme-ro en la región media (par 3); 10 submetacéntricos (sm), con centrómero en laregión submedia (pares 1, 2, 4, 5 y 8) y 2 subtelocéntricos satelizados (st), concentrómero en la región subterminal (par 7).
A este cariotipo corresponde la fórmula idiogramática siguiente: 2Msat
2m + 10 sm + 2 Stsat . Su asimetría cromosómica es de tipo 3A.Es interesante resaltar que los cromosomas mitóticos de E. flavum pre-
sentan abundantes zonas heterocromáticas (Lám. II, figs. 3a, 3c).
E. boissieri Steudel, Nomencl. Bot., ed. 2, 1: 540 (1840).
Material estudiado.Cádiz. Entre Ubrique y el Puerto de Gáliz, 14.V11.1978, Devesa, Rivera & Valdés (SEV
64946), n = 5 y 2n = 10; ídem, Grazalema, 16.V1.1979, Luque, Malato & al. (SEV 61571), n = 5.Ciudad Real. Entre Villanueva de la Fuente y Povedilla, 30.V.1980, Luque, Ubera & Valdés (SEV64948), n = 5. Jaén. Puente de Genave, 3.VI.1978, Devesa, Pastor & Valdés (SEV 64947), n = 5.Málaga. Montecorto, 13.V11.1978, Devesa, Rivera & Valdés (SEV 64945), n = 5 y 2n = 10;ídem, Ronda, hacia San Pedro de Alcántara, 16.V1. 1979, Luque, Malato & al. (SEV 64949), n = 5.Sevilla. Entre Carmona y Marchena, 26.1V.1978, Candau & Soler (SEV 61567), n = 5. Teruel.Castelserás, 7.VI11.1978, Devesa, Pastor & Talavera (SEV 61568), 2n = 10.
Se ha encontrado el número gamético n = 5 y el somático 2n = 10, coin-cidiendo este último con el indicado por FERNÁNDEZ CASAS (1977a: 40;1977b: 341) para plantas españolas recolectadas en Granada y Cuenca. Se tra-ta, por tanto, de un taxón diploide con número básico x = 5.
En meiosis, los cromosomas se comportan regularmente con formaciónde 5 bivalentes, ,como se aprecia en metafase I de la Lám. II, fig. 4b, destacán-dose dos bivalentes por su tamaño respecto al resto, lo que se refleja en meta-fase somática con la presencia de 4 cromosomas algo más grandes.
La longitud de los cromosomas de las plantas procedentes de Málaga(SEV 64945) oscila entre 1,50 y 2,36 1um, (Lám. II, fig. 4a), siendo, por tanto,

25
pequeños y medianamente pequeños, pudiéndose agrupar como sigue (Lám.II, fig. 4c): 4 metacéntricos (m), con centrómero en la región media (pares 1 y5) y 6 submetacéntricos (sm) con centrómero en la región submedia (pares 2, 3y 4).
A este cariotipo corresponde la fórmula idiogramática siguiente: 4m +6sm. La asimetría cromosómica es de tipo 2A.
Como en la especie anterior, se observa que en los cromosomas de esta es-pecie se presenta abundantes zonas heterocromáticas, aunque en una propor-ción inferior.
E. vulgare L., Sp. Pl. 139 (1753).
Material estudiado.
Alicante. Cocentaina, Sierra de Mariola, subida al Montcabré, 7.V.1979, García, Luque &Valdés (SEV 60908), n = 16. Cuenca. Entre Salinas y Paredes, 6.VII.1979, Cabezudo, Luque &Ubera (SEV 64967), n = 16. Guadalajara. Torremocha del Campo, 30.V.1981, Devesa, Luque &Romero (SEV 64968), n = 16. Huesca. Gavín, ermita de San Bartolomé, 23.VIII.1979, Valdés(SEV 64962), 2n = 32. Lérida. Ardes, 12.VII.1979, Cabezudo, Luque & Ubera (SEV 64961), n =16; ídem, Espot, subida al Lago San Mauricio, 12.VII.1979, Cabezudo, Luque & Ubera (SEV60907), n = 16. Oviedo. Playa de Rodiles, 29.VII.1978, Devesa, Pastor & Talavera (SEV 64963),2n = 32. Palencia. Entre Cervera de Pisuerga y Potes, 13.VII.1980, Devesa, Luque & al. (SEV64964), 2n = 32. Santander. Picos de Europa, 1700-2000 m. s. m., 14.VII.1980, Devesa, Luque &al. (SEV 64965), 2n = 32. ídem, Entre Buelles y San Pedro de Baena, 14.VII.1980, Devesa,Luque & al. (SEV 64966), n = 16. Valencia. Entre Játiva y Mogente, 8.V.1979, García, Luque &Valdés (SEV 68197), n = 16.
Se han encontrado los números cromosómicos n = 16 y 2n = 32, el pri-mero coincide con el indicado por STREY (1931: 718) para material cultivado,por TISCHLER (1935: 13) para plantas recolectadas en Alemania, por BRITTON
(1951: 241-242) para material cultivado, por DELAY (1969: 7) para plantasfrancesas y por FERNÁNDEZ CASAS & al. (1980: 200) para plantas españolasprocedentes de Palencia.
El número somático encontrado, 2n = 32, coincide con el indicado porBRITTON (1. c.), por MULLIGAN (1957: 779) para plantas procedentes deCanadá, por LOvE & LOVE (1956: 209) para plantas de Islandia, por GADE-
LLA & KLIPHUIS (1966: 542) para plantas recolectadas en Holanda, por PM:Kovik (1970: 10) para plantas de Checoslovaquia, por MARKOWA & IVÁNOVÁ
(1971a: 124) para plantas procedentes de diversas localidades de Bulgaria, porFERNANDES & LEITÁO (1972: 392) para plantas recolectadas en BraganÇa(Tras-os-Montes e Alto Douro, Portugal), por FRITSCH (1973: 452) para ma-terial cultivado, por NATARAJAN (1978: 530) para plantas de Francia, por

26
VAN LOON & JONG (1978: 59) para plantas procedentes de Yugoslavia, porJAVÓRKOVÁ (1979: 401) para plantas de Checoslovaquia, por VAN DEN
BRAND & al. (1979: 396) para plantas procedentes de Holanda, por VAN LOON
& KIEFT (1980: 540) para plantas recolectadas en Yugoslavia y por STRID
(1980: 709) en plantas procedentes de Grecia.DELAY (1969: 7) encontró n = 8 en plantas francesas, y STRID (1971: 491)
2n = 16 en plantas procedentes de Albania, recuentos que difieren de todoslos demás indicados.
Se trata de un taxón con número básico x = 8 y dos niveles de ploidía, di-ploide al que corresponden los dos últimos recuentos comentados y tetraploi-des, al que corresponden todos los demás.
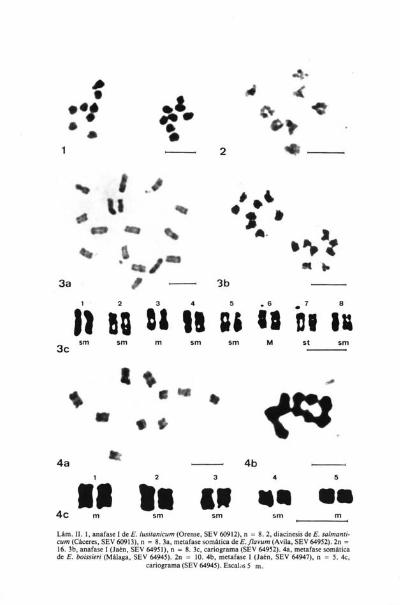
En meiosis, se ha comprobado que se forman frecuentemente tetravalen-tes en diacinesis, y sólo excepcionalmente se han encontrado 16 bivalentes(Lám. III, fig. lb).
La longitud aparente de los cromosomas de las plantas procedentes deOviedo (SEV 64963) oscila entre 1,29 y 2,21 film, siendo por tanto, pequeños ymedianamente pequeños (Lám. III, fig. la ), tamaño que se aproxima alindicado por FRITSCH (1973: 452) y por MARKOWA & IVÁNOVÁ (1971a: 126).
FRITSCH (1973: 452) indicó la presencia de ocho cromosomas isobraquia-les y 24 heterobraquiales, así como la presencia de dos pares satelizados, quehan sido igualmente indicados por FERNANDES & LEITÁO (1972: 392) y obser-vados en el presente estudio, habiéndose deducido, además que, la asimetríacromosómica es de tipo A.
E. plantagineum L., Mantissa Alt. 202 (1771).E. lycopsis L. ex Grufberg, Fi. AngL 12 (1754) p.p.
Material estudiado.Albacete. Entre Reolid y Villapalacios, 30.V.1980, Luque, Cibera & Valdés (SEV 64993), n = 8;
ídem, Villapalacios, 9.V.1980, García, Luque & Valdés (SEV 64989), n = 8. Avila. El Barco de
Avila, 5.VI.1979, Díez, Pastor & Silvestre (SEV 64990), n = 8. Cádiz. Entre Algodonales yGrazalema, Los Batanes, 16.VI.1979, Luque & al. (SEV 64991), 2n = 16; ídem, entre El Puertode Santa María y Puerto Real, 13.1V. 1978, Luque & Valdés (SEV 64987), n = 8. Huelva. Santa
°lana, 18.V.1978, Candau, Díez & Pérez (SEV 64986), n = 8. La Coruña. Santiago de Compos-
tela, 8.VI.1979, Díez, Pastor & Silvestre (SEV 60768), n = 8. Lugo. Quiroga, 7.VI.1979, Díez,
Pastor & Silvestre (SEV 64985), n = 8. Madrid. Casa de Campo, 3.VI.1980, Pardo (SEV 64988),
2n = 16. Málaga. Archidona, 29.VI.1978, Devesa, Pastor & Valdés (SEV 60762), 2n = 16. Mur-
cia. Entre Los Alcazares y Las Arrutias, 5.V.1979, García, Luque & Valdés (SEV 64992), n = 8 y
2n = 16. Sevilla. Sevilla, Tablada, 7.111.1978, Luque (SEV 64954), n = 8; ídem, alrededores deFacultad de Biología, 8.111.1978, Luque (SEV 64955), n = 8; ídem, entre Marchena y La Lante-juela, 24.1V. 1979, Candau, Fernández & Luque (SEV 64984), 2n = 16.

27
El número haploide encontrado (n = 8) coincide con el indicado porSTREY (1931: 718), y BRITTON (1951: 241) para material cultivado. El nú-mero somático (2n = 16) coincide con el indicado por L1TARDIÉRE (1943: 158)para plantas procedentes de Córcega, Túnez y Argelia y para material cultiva-do, por BRITTON (1. c.) para material cultivado, por BORGEN (1970: 148) paraplantas españolas procedentes de Gran Canarias, por STRID (1971: 491) paraplantas recolectadas en Albania, por FERNANDES & LEITÁO (1972: 392) paraplantas de diversas localidades de Portugal y por FRITSCH (1973: 452) paramaterial silvestre procedente del Jardín Botánico de Coimbra (Portugal). Setrata de un taxón diploide con número básico x = 8.
Las fases observadas de la meiosis han resultado ser regulares, formándo-se 8 bivalentes y emigrando los cromosomas en anafase I, (Lám. III, fig. 3).
La longitud aparente de los cromosomas de las plantas de Málaga (SEV60762) oscila entre 0,98 y 1,40 film, por lo que se consideran pequeños, igual-mente fueron considerados por FRITSCH (1973: 452), que indicó además lapresencia de un par de cromosomas satelizados, así como LITARDIÉRE (1943:161), mientras que FERNANDES & LEITÁO (1972: 392) indican dos pares decromosomas satelizados. En el presente estudio se ha confirmado la presenciade un sólo par satelizado.
El grado de asimetría del cariotipo es de tipo A.
E. sabulicola Pomel, Nouv. Mal. Fi. Ah. 90 (1874).
Material estudiado.Alicante. Santa Pola, 5.V.1979, García, Luque & Valdés (SEV 64994), n = 8. Almería.
Serón, 30.V.1978, Devesa, Pastor & Valdés (SEV 64997), n = 8 y 2n = 16. Baleares. Mallorca.Puerto de Sóller, 8.VI.1981, Devesa, Luque & Romero (SEV 63318), n = 8. Granada. Béznar,8.IV.1979, Devesa, Luque & Ubera (SEV 61566), n = 8. Murcia. Cartagena, 10.IV.1979, Deve-sa, Luque & Ubera (SEV 64996), 2n = 16. ídem, Fuente Alamo de Murcia, 4.V.1979, García,Luque & Valdés (SEV 64998), n = 8 y 2n = 16; ídem, Los Nietos, 5.V.1979, García, Luque &Valdés (SEV 64995), n = 8 y 2n = 16.
En las plantas estudiadas, se ha encontrado n = 8 (Lám. III, fig. 2) y 2n = 16,este número somático coincide con el indicado por LITARDIÉRE (1943:158, sub E. modestum Ball var. decipiens (Pomel) Maire) para plantas de Ar-gelia y por CARDONA (1973: 10) para plantas españolas procedentes deCapifort (Menorca). Se trata, por tanto, de un taxón diploide con númerobásico x = 8.
En las plantas procedentes de Almería (SEV 64997) el tamaño aparentede los cromosomas oscila entre 0,84 y 1,26 am, siendo por tanto pequeños,por lo que su asimetría cromosómica sería de tipo A.

28
E. confusum Coincy, Morot Journ. Bot. 14: 298 (1900).
Material estudiado.Baleares. Ibiza, alrededores de la Catedral, 6.VI.1981, Devesa, Luque & Romero (SEV
63172),n = 8 y 2n = 16.
En lag plantas estudiadas se ha encontrado n = 8, y en células somáticasde tejido conectivo, 2n = 16. Se trata de un taxón diploide con número básicox = 8. Según la bibliografía consultada, es la primera vez que se estudia cario-lógicamente este taxón.
El comportamiento de los cromosomas en meiosis es normal, y el tamañoaparente de los cromosomas en metafase mitótica es inferior a 2 ,um, siendo,por tanto, pequeños.
E. creticum L., Sp. Pl. 139 (1753).
subsp. creticum.
Material estudiado.Gerona. Entre Rosas y Cadaqués, 2.V1.1981, Devesa, Luque & Romero (SEV 63296), n = 8;
ídem, Puerto de la Selva, 2.V1.1981, Devesa, Luque & Romero (SEV 63295), n = 8.
El número encontrado, n = 8, concuerda con el somático 2n = 16 indi-cado por LITARD1ÉRE (1943: 156, sub E. australe Lam.) para plantas cultiva-das y procedentes de Argelia. Se trata, de un taxón diploide con númerobásico x = 8, siendo al parecer la primera vez que se estudia material español.
En meiosis, los cromosomas se comportan de una manera normal, conformación de ocho bivalentes (Lám. III, fig. 4).
subsp. coincyanum (Lacaita) R. Fernandes, Bol. Soc. Brot., ser. 2,43: 153(1969).
Material estudiado.Alicante. Cocentaina, Base de la Sierra de Mariola, 7.V.1979, García, Luque & Valdés (SEV
64971), n = 16; ídem, Santa Pola, 5.V.1979, García, Luque & Valdés (SEV 64975), n = 16. Alme-
ría. Velez Rubio, 3.V.1979, García, Luque & Valdés (SEV 60894), n = 16. Cádiz. Subida a Gra-
zalema, 16.V1.1979, Luque & al. (SEV 64977), 2n = 16. Granada. Entre Diezma y Guadix,
2.V.I979, García, Luque & Valdés (SEV 64972), n = 8. Jaén. Sierra de la Cabrilla, 1500-1800
m.s.m., 20.V1.1979, Díez, Ubera & Valdés (SEV 60896), n = 8. Málaga. Antequera, Base del
Torcal, 29.V.1978, Devesa, Pastor & Valdés (SEV 64973), n = 16. Murcia. Entre Aguilas y Ma-
zarrón. Sierra del Cantar, 31.V.1978, Devesa, Pastor & Valdés (SEV 60893), 2n = 16; ídem, Sie-
rra del Cantar, 10.IV.1979, Devesa, Luque &libera (SEV 64974), n = 16. Sevilla. Alcalá de Gua-
daira, 10.111.1978, Luque (SEV 64970), n = 8.

29
Se ha encontrado en las plantas de tres poblaciones, n = 8, en cinco n = 16y en las dos restantes el número somático 2n = 16. Este último coincide con elindicado por FRITSCH (1973: 453, sub E. spinescens Med.) para material culti-vado. Se trata de un taxón con número básico x = 8 y dos niveles de ploidía,ya que cinco de las poblaciones estudiadas son diploides con 2n = 16 y cincotetraploides con 2n = 32.
Se ha observado en meiosis, frecuente formación de tetravalentes en lasplantas tetraploides (Lám. III, fig. 6), presentándose fases normales en los di-ploides (Lám. III, fig. 5).
La longitud aparente de los cromosomas de las plantas procedentes deMurcia (SEV 60893) oscila entre 0,84 y 1,40 film, siendo por tanto, pequeños.En cuanto al cariotipo, puede solamente indicarse que en los diploides se pre-senta un par de cromosomas satelizados, y que la asimetría es de tipo A.
E. tuberculatum Hoffmanns. & Link, Fi. Port. 1: 183 (1810).
Material estudiado.Ciudad Real. Valdepeñas, 27.V.1980, Luque, Ubera & Valdés (SEV 64957), n = 16. Huelva.
Entre Higuera de la Sierra y Valdezufre, 22.X.1979, Rivera (SEV 64960), 2n = 32. Sevilla. EntreEl Pedroso y Cazalla de la Sierra, 18.V.1979, Pastor & Valdés (SEV 60914), n = 16.
Se ha encontrado n = 16 y 2n = 32, siendo por tanto, plantas tetraploi-des con número básico x = 8. Dicho número cromosómico difiere del diploide(2n = 16) indicado por LITARDIÉRE (1943: 157, sub E. hoffmannseggii R. Li-tard.) y por GARDÉ & MALHEIROS-GARDÉ (1953: 123) para material cultivado,y por FERNANDES & LEITÁO (1972: 392) para plantas portuguesas de origensilvestre. FRITSCH (1973: 453, sub E. hoffmannseggii R. Litard.) indicó el mis-mo número, pero encontró en un placa metafásica 2n = 32.
Se trata, por tanto, de un taxón con dos niveles de ploidía, siendo diploi-des las poblaciones portuguesas y tetraploides las españolas estudiadas.
El porcentaje de tetravalentes en profase I es muy elevado (Lam. III, fig. 7),siendo excepcional la formación de solamente bivalentes (Lám. 111, fig. 8). El
tamaño aparente de los cromosomas de los cariotipos estudiados es inferior a2pm, considerándose por tanto, pequeños.
E. gaditanum Boiss., Voy. Bot. Midi Esp. 2: 422 (1841).
Material estudiado.Cádiz. Entre Rota y Chipiona, 21.V.1978, Pérez Vega (SEV 64981), n = 16; ídem, entre
Jimena de la Frontera y San Roque, 10.VII.1978, Candau, Díez & Luque (SEV 64982), n = -16.

30
Huelva. Almonte, Matalascañas, 20.V111.1978, Valdés (SEV 64979), 2n = 32; ídem, Almonte,Matalascañas, 19.VIII.1979, Valdés (SEV 64980), 2n = 32; ídem, Almonte, Matalascañas,1.IV.1980, Díez (SEV 60890), n = 16.
El número somático encontrado (2n = 32) concuerda con el indicado porFRITSCH (1973: 453) para material cultivado y por FERNÁNDEZ CASAS (1977a:40) para plantas de Cádiz. Se trata, por tanto, de un taxón tetraploide con nú-mero básico x = 8.
FRITSCH (1973: 456) indica que tanto el número n = 16 atribuído porBRITTON (1951) a E. rosulatum Lange, como el número somático 2n = 32indicado por STREY (1931) y LITARDIÉRE (1943) para este mismo taxón, co-rresponden en realidad a E. gaditanum. No ha podido comprobarse el mate-rial testigo utilizado por estos autores, pero dado que utilizaron para sus estu-dios plantas procedentes de diversos jardines botánicos, es fácil que la deter-minación del material fuera incorrecta.
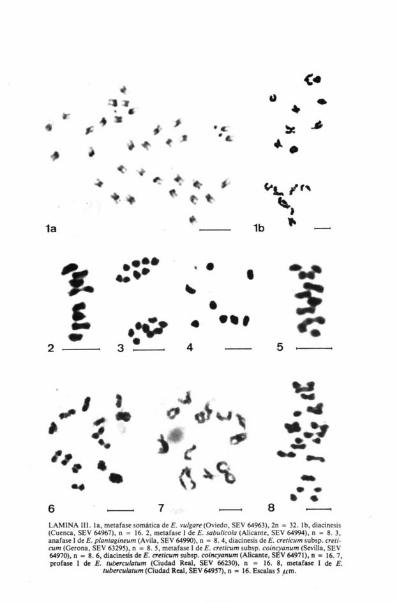
En meiosis, se ha observado que en la mayoría de las placas, los cromoso-mas forman tetravalentes y sólo excepcionalmente bivalentes (Lam. IV, fig. lb).
Las plantas procedentes de Almonte (Huelva, SEV 64979) poseen cromo-somas con longitud aparente comprendida entre 0,98 y 1,96 juin, considerán-dose pequeños (Lám. IV, fig. la). Dichos cromosomas se pueden agrupar dela siguiente manera (Lám. IV, fig. 1c): 4 metacéntricos (M), con centrómeroen el punto medio (pares 13 y 16); 6 metacéntricos (m), con centrómero en laregión media (pares, 2, 3 y 7), y 22 submetacéntricos (sm), con centrómero enla región submedia (pares 1, 4, 5, 6, 8, 9, 10, 11, 12, 14 y 15).
A este cariotipo corresponde la fórmula idiogramática: 4M + 6m +22sm, y asimetría cromosómica de tipo 3B.
E. rosulatum Lange, Ind. Sem. Horto Haun. 1857: 22 (1857).
Material estudiado.La Coruña. El Ferrol, Playa de Doniños, 9.VI.1979, Díez, Pastor & Silvestre (SEV 60900),
n = 16.
El número encontrado (n = 16) coincide con el somático 2n = 32 indica-do por FERNANDES & LEITÁO (1972: 392) para plantas de diversas localidadesde Portugal. Se trata de un taxón tetraploide con número básico x = 8, siendoal parecer, la primera vez que se estudia material español. Como se ha indica-do anteriormente, los recuentos atribuídos a este taxón por STREY (1931),LITARDIÉRE (1943) y BRITTON (1951) corresponden a E. gaditanum, segúnFRITSCH (1973: 456).
dt.
t. 4 fi *
IIla
5•3
1,411
#37
e',Ir
44.:* 11• o
• a • «
lb I.• *,
•
•
4
be
•
6 7 8
LAMINA III. la , metafase somática de E. vulgare (Oviedo, SEV 64963), 2n = 32. lb, diacinesis(Cuenca, SEV 64967), n = 16. 2, metafase I de E. sabulicola (Alicante, SEV 64994), n = 8. 3,anafase I de E. plantagineum (Avila, SEV 64990), n = 8. 4, diacinesis de E. creticum subsp. creti-curn (Gerona, SEV 63295), n = 8. 5, metafase I de E. creticum subsp. coincyanum (Sevilla, SEV64970), n = 8. 6, diacincsis de E. creticum subsp. coincyanum (Alicante. SEV 64971), n = 16. 7,profase I de E. tuberculatum (Ciudad Real, SEV 66230), n = 16. 8, metafase I de E.
tuberculatum (Ciudad Real, SEV 64957), n = 16. Escalas 5 pm.

4
<ti *
SM SM
14 15 16
111111
la lb
4 5 6
pi OI21 NI U anM SM SM SM
9 10 12
3174 01811 ea aa11
aaSM SM SM
2LAMINA IV. la , metafase somática de E. gadiumum (Huelva, SEV 64979), 2n = 32. lb, diaci-nesis (Huelva. SEV 64980). n = 16. I c. carioarama (Huelva. SEV 64979). 2. diacinesis de E. rosa-latum (La Coruña, SEV 60900), n = 16. 3, metafase I de E. arenarium (Ibiza, SEV 63291), n = 8.
Escalas = 5 p.m.

31
Se ha comprobado que ocasionalmente se forman bivalentes (Lám. IV.fig. 2), mientras que la formación de tetravalentes resulta muy frecuente.
E. parviflorum Moench, Meth. 423 (1794).
Material estudiado.Baleares. Menorca, Cales Piques, Cintadella, 18.111.1980, Cardona (SEV 64983), 2n = 16.
Se ha encontrado en células de tejido conectivo, el número cromosómico2n = 16, que concuerda con el indicado por DAHLGREN & al. (1971: 252) paraplantas españolas procedentes de Mallorca y por CARDONA & CONTANDRIO-
POULOS (1980: 275) para plantas de Cabrera (España). Difiere, en cambio, delnúmero somático 2n = 12 aportado por KLIPHUIS & BARKOUDAH (1977: 241)para material procedente de Siria.
Del cariotipo representado por DAHLGREN & al. (1971: 253), se hadeducido que la longitud aparente de los cromosomas varía entre 0,77 y 1,29
iam, por lo que se consideran pequeños, siendo la asimetría de este cariotipadetipo A.
E. arenarium Guss., Ind. Sem. Horto Boccad. 5 (1826).
Material estudiado.Baleares. Ibiza, Alrededores de la Catedral, 6.VI.1981, Devesa, Luque & Romero (SEV
63291), n = 8.
El número encontrado (n = 8) coincide con el somático 2n = 16 indicadopor KAMARI & PAPATSOU (1973: 267) para plantas de Grecia. Se trata, portanto, de un taxón diploide con número básico x = 8, siendo ésta, al parecer,la primera vez que se estudia con material español.
Se ha comprobado que en diacinesis y metafase I se forman constante-mente bivalentes (Lám. IV, fig. 3).
Del cariotipo representado por KAMARI & PAPATSOU (1. c.) puedededucirse que el tamaño de los cromosomas es inferior a 2 ,um, por lo que seconsideran pequeños, y que su asimetría debe ser de tipo A.
DISCUSION
Del estudio de los cariotipos de los taacones del género Echium representa-dos en España peninsular y Baleares, se deduce que la longitud aparente de los

32
cromosomas oscila entre 0,84 y 3,44pm, por lo que se pueden considerar co-mo pequeños y medianamente pequeños, siendo E. sabulicola y E. creticumsubsp. coincyanum los que los presentan de tamaño más pequeño, con 0,84 -1,26,um, y 0,84- 1,40 1um, respectivamente y E. flavum, con 2,15 - 3,44 1um, elque los tiene más grandes.
Los taxones estudiados se pueden separar claramente en cuatro grupos,en cuanto al tamaño de sus cromosomas. El primero está constituído por E.sabulicola, E. creticum y E. plantagineum, con cromosomas pequeños. El se-gundo, por E. italicum y E. asperrimum, con cromosomas pequeños y media-namente pequeños y con cariotipos muy característicos, ya que presentan unpar de cromosomas marcadamente más grandes que los demás. A un tercergrupo pertenece E. flavum que presenta los cromosomas más grandes de lasespecies estudiadas. El cuarto grupo, al que pertenecen los restantes taxonesestudiados, presentan un tamaño cromosómico intermedio entre los delprimero y segundo grupo, aunque solapado con ellos, de los que se separa porel grado de asimetría cromosómica.
Por el número cromosómico, los 19 taxones estudiados pueden igualmen-te formar cuatro grupos, que no coinciden exactamente con los formados deacuerdo con el tamaño de los cromosomas. Estos grupos se han formado porlos taxones con 2n = 10, 2n = 14, 2n = 16 y 2n = 32 cromosomas. Los tresprimeros grupos están constituidos por taxones diploides, y el último por te-traploides. En los tres primeros se mantiene casi constante la proporción entrecromosomas metacéntricos (isobraquiales) y submetacéntricos y subte-locéntricos (heterobraquiales), mientras que el cuarto grupo formado portaxones tetraploides, la proporción varía, ya que el porcentaje de cromo-somas metacéntricos es bastante inferior al que se presenta en las especiesdiploides, como se puede apreciar en el cuadro 1. Los cariotipos más asi-métricos corresponden a los taxones tetraploides, con asimetría de tipo 3A y3B. El tipo más primitivo en cuanto a la asimetría se refiere, corresponde a E.boissieri con 2n = 10, seguido de E. asperrimum con 2n = 14, en que laasimetría es de tipo 2B. En cuanto a la especies con 2n = 16 presentan cario-tipos primitivos y avanzados, ya que varían entre 2A y 3A, aunque parecenpredominar los de 2A. E. italicum, perteneciente a este grupo, tiene asimetría2B, que es la misma que presenta E. asperrimum, con 2n = 14, y E. fiavum conasimetría de tipo 3A, que a pesar de presentar 2n = 16, parece ser queconstituye una línea evolutivamente diferente.
COINCY (sec. LITARDIÉRE, 1943, y SAUVAGE & VINDT, 1954), dividió aEchium, excluyendo Lobostemon, en tres secciones. Los taxones macaronési-

33
CROMOSOMAS
Submetacéntricos
2n Metacéntricos Subtelocéntricos Asimetría
10 80% 20% 2A14 80% 20% 2B16 75% 25% 2A, 2B, 3A32 25% 75% 3A, 3B
Cuadro 1. Caracteres cariolágicos de los tazones de Echium con 2n = 10, 14, 16 y 32.
cos integran la Sect. Pachylepis Coincy, y han sido estudiados cariológica-mente en su mayor parte por BORGEN (1969, 1970, 1974, 1977), BRAMWELL &
al. (1971, 1972) y LARSEN (1960). Los mediterráneos quedan separados en dossecciones de acuerdo con los caracteres del anillo basal interno de la corola:Sect. Eleuterolepis Coincy ( = Sect. Echium), con anillo basal formado por 10escamas bien separadas, y Sect. Gamolepis Coincy, con el anillo basal forma-do por una membrana o engrosamiento continuo. De las especies estudiadas,E. sabulicola, E. confusum y E. arenarium pertenecen a la Sect. Gamolepis, ylas restantes a la Sect. Eleuterolepis. Los grupos formados por los caracterescariológicos no guardan por tanto ninguna relación con las secciones propues-tas por COINCY. Esta diferencia pone de manifiesto lo artificioso de la clasifi-cación de COINCY y la necesidad de clasificar el género de una manera más na-tural.
STEBBINS (1938) consideró x = 8 como número básico de este género.Los estudios posteriores de BRIrroN (1951), BORGEN (1969), DELAY (1969), yFERNANDES & LEITÁO (1972), añadieron x = 7, y los de BRAMWELL (1973),FRITSCH (1973) y KLIPHUIS & BARKOUDAH (1977: 241), x = 6.
LITARDIÉkE (1943) suponía para Echium la existencia de los númerosbásicos x = 4 y x = 7. Para él, salvo E. asperrimum con x = 7, los demás ta-xones serían tetraploides, hexaploides u octoploides con número básico x = 4.No existirían taxones diploides, ya que no se ha detectado ninguna especiecon 2n = 8. El número cromosómico indicado por FERNÁNDEZ CASAS & al.(1980: 199) para E. strictum L. fil., 2n = 8, puede considerarse raro, ya quetodos los recuentos efectuados por otros autores han coincidido en indicarn = 8. (UNDER & LAMBERT, 1965: 237; BRAMWELL & al. 1971: 377, para subsp.exasperatum (Webb.) Bramwell) ó 2n = 16 (LARsEN, 1960: 38; BORGEN,
1969: 90; BRAMWELL, 1. C.).

34
Los taxones incluidos en el presente estudio poseen x = 5, 7 y 8. Se puedeafirmar, por tanto, que en el género Echium se presenta una serie disploidecon números básicos x = 5, 6, 7 y 8, no considerándose válida la hipótesis deLITARDIÉRE (1943) de tomar como número básico fundamental x = 4.
De acuerdo en parte con las ideas de JOHNSTON (1953), hay que suponerque el género Echium tenga su origen en Africa del Sur, en que se ha originadouna línea evolutiva independiente, constituida por numerosos taxones prefe-rentemente leñosos con fascículos de pelos en la base de los estambres, y quepara muchos autores constituye un género independiente, Lobostemon. Deantepasados comunes, y por migración por la Región Atlántica hacia el hemis-ferio boreal, se han originado todos los taxones de Echium de Macaronesia,leñosos, y posteriormente los mediterráneos, preferentemente herbáceos, queevolucionaron intensamente en esta región, que hay que considerar como uncentro secundario de evolución del género.
Todas las especies de Africa del Sur presentan x = 7, y las de Macarone-sia x = 8, por los que hay que considerar que el número básico primitivo de estegénero ha de ser uno de estos dos.
BRAMWELL (1973) sugiere que el número básico primario en Echium esx = 8, y que de él han derivado por pérdida cromosómica x = 7 y x = 6.FRITSCH (1973) considera x = 8 como más primitivo porque la mayoría de lasespecies con este número básico presentan los cromosomas de sus cariotiposmuy similares en cuanto a diferencias de tamaño y morfología, lo que deacuerdo con STEBBINS (1971) es signo de primitivismo. De especies con estascaracterísticas, han derivado otras que como E. italicum presentan dos cromo-somas de tamaño superior a los demás, de las cuales, a su vez, pueden haber de-rivado otras con x = 7, como E. asperrimum, igualmente con dos cromoso-mas grandes. Considera, en cambio, que el número básico x = 6 no puede de-rivar directamente de x = 7.
Los resultados obtenidos en el presente estudio, basado en especies medi-terráneas, apoyan la idea de considerar x = 8 como el número básico másprimitivo para este grupo. Este es el número básico que corresponde igual-mente a las especies macaronésicas, y cabe suponer que es el número básicoancestral del género, que en su evolución ha desarrollado una línea con x = 7correspondientes a las especies de Africa del Sur, manteniéndose el númerobásico primitivo en los grupos macaronésicos y mediterráneos. De este núme-ro básico primitivo, han podido originarse los demás.
Como se indicó en la introducción, este género ha sido considerado poralgunos autores como tribu independiente de Lithospermeae.

35
El estudio cariológico pone de manifiesto que, por la asimetría de sus ca-riotipos (comprendida entre 2A y 3B) y por su reducido tamaño de los cromo-somas, se asemeja a los caracteres cariológicos descritos para Lithospermums. I. (LuQuE & VALDÉS, 1984), mientras que por sus números básicos bajos seaproximaría a otros géneros de Lithospermeae.
Morfologicamente, se diferencia marcadamente del resto de los géneros,sobre todo por su corola zigomorfa, así como por los caracteres anatómicosdel fruto (SEIBERT, 1978). Por todo ello, se considera que ha debjdo tener unorigen independiente del resto de Lithospermeae, aunque por las característi-cas de las núculas, se encuentra próximo a Neatostema (JOHNSTON, 1953).Esta interpretación difiere de la propuesta por LAWRENCE (1937) que en basea los caracteres anatómicos de los órganos florales, considera a Echium máspróximo a Boragineae (= Anchuseae) que a Lithospermeae.
BIBLIOGRAFIA
ARYAVAND, A. (1972) in A. LOVE (ed.) IOPB Chromosome number reports. LVII. Taxon 26:443-444.
BENTHAM, G. & J. D. HOOKER (1876) Genera Plantarum 2. London.
BORGEN, L. (1969) Chromosome numbers of vascular plants from the Canary Islands, with spe-cial reference to the occurrence of polyploidy. NyII. Mag. Bol. 16: 81-121.
(1970) Chromosorne numbers of Macaronesian flowering plants. Nytt. Mag. Bol. 17:145-161.
(1974) Chromosome numbers of Macaronesian flowering plants. II. Norw. Journ.Bol. 21: 195 -211.
(1977) Check-lis( of chromosome numbers counted in Macaronesian vascular plants.Oslo.
BRAtviwen, D. (1973) Studies in the genus Echium from Macaronesia. Monogr. Biol. Canariensis4: 71-82.
(1978) Boraginaceae in V. H. HEYwooD (ed.)Flowering Plants of the World. London.
, C. J. HUMPHIES, B. G. MURRAY & S. J. Oweris (1971) Chromosome numbers inplants from the Canary Islands. Bol. Not. (Lund) 124: 376-382.
, C. J. HUMPHIES, B. D. MURRAY & S. J. OwENs (1972) Chromosome studies in theflora of Macaronesia. Bol. Not. (Lund) 125: 139-152.
BárrroN, D. M. (1951) Cytogenetic studies on the Boraginaceae. Brittonia 7: 233-266.
CARDONA, M. A. (1973) Contribution á l'étude cytotaxonomique de la flore des Baleares. I. ActaPhytotax. Barcinonensia 14: 1 -20.
& J. CONTANDRIOPOULOS (1980) Números cromosómicos para la flora española. 162-182. Lagascalia 9: 272-284.

36
DAHLGREN, R., TH. KARLSSON & P. LASSEN (1971) Studies on the flora of the Balearic Islands. I.Chromosome numbers in Balearic angiosperms. Bol. Not. (Lund) 124: 249-269.
DE CANDOLLE, A. (1846) Prodromus Systematis Naturalis Regni Vegetabilis 10. Parisiis.
DELAY, J. (1969) Boraginacées. I. Inform. Annuelles Caryosyst. Cytogénét. 3: 7-8.DI FuLvio, T. E. (1965) Recuentos cromosómicos en Boraginaceae. Kurtziana 2: 131-132.
DIERDJOUR, B. & G.-G. GUITrONNEAU (1977) in A. LOVE (ed.) IOPB Chromosome number re-ports. LVI. Taxon. 26: 269.
FERNANDES, A. & M. T. LEITÁO (1972) Contribution a la connaissance cytotaxinomiques desSpermatophyta du Portugal. V. Boraginaceae. Bol. Soc. Brot., 2 1 ser., 46: 389-405.
& M. QUEIRÓS (1971) Sur la caryologie de quelques plantes récoltées pendant la IllémeRéunion de Botanique Péninsulaire. Mem. Soc. Brot. 21: 343-385.
FERNÁNDEZ CASAS, J. (1977a) Recuentos cromosómicos en plantas vasculares españolas.Saussurea 8: 40.
(1977b) Números cromosómicos de plantas españolas. IV. Anal. Inst. Bol. Cavanilles34: 335-349.
(1978) Recuentos cromosómicos en plantas vasculares españolas. 2. Saussurea 9: 45-50.
(1979) Echium cantabricum (Laínz) Fernández Casas & Laínz, stat. nov. Mém. Soc.Bol. Genéve 1: 111-114.
, A. PONS-SOROLLA & A. SUSANNA (1980) Números cromosomáticos de plantas occi-dentales (64-69). Anal. Jardín Bol. Madrid 37: 199-201.
FarrscH, B. (1973) Karyologische Untersuchungen in der Gattuna Echium L. Bol. Not. (Lund)126:450-458.
GADELI.A, TH W. J. & E. KLIPHUIS (1966) Chromosome numbers of flowering plants in the Nether-lands. II. Proc. Koninkl. Nederland. Akad. Wetensch. ser. C, 69: 541-556. , E. KLIPHUIS & E. A. MENNEGA (1966) Chromosome numbers of some flowering
plants of Spain and S France. Acta Bol. Neerland. 15: 484-489.GARDÉ, A. & N. MALHEIROS-GARDÉ (1953) ContribuiÇáo para o estudo cariologico de algunas es-
pecies de angiospermicas. I. Genét. Ibér. 5: 115-124.GüRKE, M. (1893) Borraginaceae (Asperifoliaceae) in A. ENGLER & K. PaArrn. (eds.)Die Natürli-
chen Pflanzenfamilien 4(3a): 71-131. Berlin.
JAvüaxovA, V. (1979) in A. LOve (ed.) 10PB Chromosome number reports. LXIV. Taxon 28:400-401.
JOHNSTON, 1. M. (1953) Studies in the Boraginaceae XXV. A revaluation of some genera of theLithospermeae. Journ. Arnold Arbor. 34: 258-299.
KAmmtí, G. & S. PAPATSOU (1973) Chromosome studies in some Mediterranean angiosperms.Bol. Not. (Lund) 26: 266-268.
KuPtruls, E. (1977) in A. LÓVE (ed.) 10PB Chromosome number reports. LVI. Taxon 26: 267-268.
& Y. I. BARKOUDAH (1977) Chromosome numbers in some Syrian angiosperms. ActaBol. Neerland 26: 239-249.
& J. H. WIEFFERING (1979) in A. LüVE (ed.) IOPB Chromosome number reports.LXIV. Taxon 28: 398-400.

37
LARSEN, K. (1960) Cytological and experimental studies on the flowering plants of the Canary Is-lands. Biol. Skr. Danske Vid. Selsk. 11: 1-60.
LAWRENCE, J. R. (1937) A correlation of the taxonomy and the floral anatomy of certain of theBoraginaceae. A mer. Journ. Bol. 24:433-444.
LEVAN A., K. FREDGA & A.A. SANDBERG (1965) Nomenclature for centromeric position of chro-mosomes. Hereditas 52:201-220.
UNDER, R. & A. M. LAMBERT (1965) Etude caryologique d'endemiques canarienses. Bull. Soc. Bol.
&once 112:234-238.LITARDIÉRE, R. DE (1940-41) Recherches caryologiques et caryotaxonimiques sur les Boragina-
cées. Bu!!. Soc. Hist. Nat. Afrique Nord 32: 315-330.
(1943) Recherches caryologiques et caryotaxonomiques sur les Boraginacées II. Nom-bres chromosomiques dans le genre Echium. Boissiera 7: 155-165.
LOvE, A. & D. LOvE (1956) Cytotaxonomical conspectus of the Icelandie flora. Acta Horti Coto-burg. 20: 65-291.
& D. LOvE (1975)Plant chromosome. Vaduz.
LuQuE, T. (1980) Números cromosómicos de algunas Boraginaceas de Portugal. Bol. Soc. Brot.,2 a ser., 53: 663-670.
& B. VALDÉS (1984) Karyological studies on Spanish Boraginaceae. Lithospermum L.Bol. Journ. Linn. Soc. 88: 335-350.
MARKOWA, M. L. & P. IvANovA (1971) Karyologische Untersuchung der Vertreter der Fam. Bo-raginaceae, Labiatae und Scrophulariaceae in Bulgarian. II. lzvest. Bot. Inst., Otd. Biol.Nauk. 21: 123-131.
MICHAELIS, G. (1964) Chromosomenzahlen einiger Kanarischer Endemismen. Planta (Berlin) 62:194.
MULLIGAN, G. A. (1957) Chromosome numbers of Canadian weeds. I. Canad. Journ. Bol. 35:779-789.
NATARAJAN, G. (1978) in A. LOvE (ed.) IOPB Chromosome number reports LXII. Taxon 27:526-531.
PAticovit, M. (1970) in MAtovsKV & al. (eds.) Index of chromosome numbers of Slovakian Flora(Part 1). Acta Fac. Rerum. Nat. Univ. Comenianae, Bol. 16: 10.
PODLECH, D. & A. DIETERLE (1969) Chromosomenstudien an afganischen Pflanzen. Candollea24: 185-243.
PoPov, M. G. (1953) Boraginaceae in B. K. SH1514KIN (ed.) Flora of the USSR 19. Moskva-Leningrad.
SAUVAGE, CH. & J. VINDT (1954) Flora du MarOC 2. Tanger.
SEIBERT, J. (1978) Fruchtanatomische Untersuchungen an Lithospermeae (Boraginaceae).Dissert. Bol. 44. Vaduz.
SNOW, R. (1963) Alcoholic hydrochloric acid-carmine as a stain for chromosome in squashpreparations. Stain Technol. 38: 9-13.
STEBBINS, G. L. (1938) Cytological characteristic associated with the different growth habits inthe dicotyledons. Amer. Journ. Bol. 25: 189-198.

38
(1971) Chromosomal Evolution in Higher Plants. London.
STREY, M. (1931) Karyologische Studien an Boraginoideae. Planta (Berlin) 14: 682-730.
STRID, A. (1971) Chromosome numbers in some Albanian angiosperms. Bot. No!. (Lund) 124:490-496.
(1980) in A. LOvE (ed.) Chromosome number reports LXIX. Taxon 29: 709-710.
& R. FRANZEN (1981) in A. LOVE (ed.) IOPB Chromosome number reports LXXIII.Taxon 30: 829-842.
TISCHLER, G. (1935) Die Bedeutung der Polyploidie für die Verbreitung der Angiospermen. Bot.Jahrb. 67: 1-36.
Tilo, J. H. & A. LEVAN (1950) The use of oxyquinoline in chromosome analysis. Anal. Es!.Exper. Aula Dei 2: 21-64.
UHRIKovd, A. & J. MAJovsKY (1978) in A. LOvE (ed.) 10PB Chromosome number reports.LXI. Taxon 27: 378-379.
VAN DEN BRAND, C., F. C. M. VAN MEEL & J. H. WIEFFERING (1979) in A. LOVE (ed.) IOPB Chro-mosome number reports. LXIV. Taxon 28: 395-397.
VAN LooN, J. CH. & H. DE JONG (1978) in A. LAVE (ed.) IOPB Chromosome number reports.LIX. Taxon 27: 56-60.
& B. IGErr (1980) in A. LOvE (ed.) IOPB Chromosome number reports. LXVIII.Taxon 29: 538-542.
& H. M. SNELDERS (1979) in A. LOvE (ed) IOPB Chromosome number reports. LXV.Taxon 28: 632-634.