Embed Size (px)
Citation preview
TEMA: Gluconeogénesis
MARCO TEORICO:
Importancia Biológica
La glucosa es considerada como el combustible y bloque estructural casi universal de todos los organismos vivos, desde los microbios hasta el hombre.
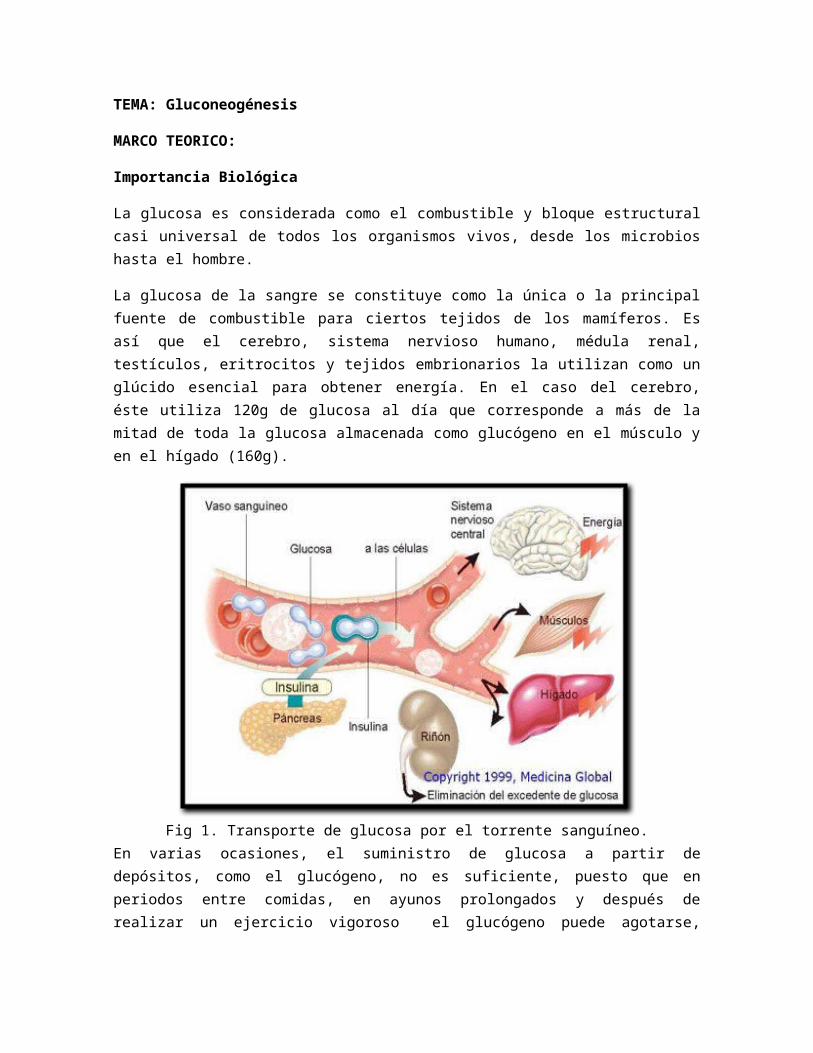
La glucosa de la sangre se constituye como la única o la principal fuente de combustible para ciertos tejidos de los mamíferos. Es así que el cerebro, sistema nervioso humano, médula renal, testículos, eritrocitos y tejidos embrionarios la utilizan como un glúcido esencial para obtener energía. En el caso del cerebro, éste utiliza 120g de glucosa al día que corresponde a más de la mitad de toda la glucosa almacenada como glucógeno en el músculo y en el hígado (160g).
Fig 1. Transporte de glucosa por el torrente sanguíneo.En varias ocasiones, el suministro de glucosa a partir de depósitos, como el glucógeno, no es suficiente, puesto que en periodos entre comidas, en ayunos prolongados y después de realizar un ejercicio vigoroso el glucógeno puede agotarse, entonces surge la necesidad por parte de los organismos de buscar una vía alternativa para sintetizar glucosa.
Frente a esta necesidad surge la gluconeogénesis que significa “formación de un azúcar nuevo”, esta vía se encarga de obtener glucosa a partir de precursores no glucosídicos así a partir del piruvato y compuestos relacionados de 3 y 4 carbonos se producirá glucosa.
Los precursores más importantes en animales son compuestos de 3 carbonos tales como: lactato, piruvato, glicerol y ciertos aminoácidos entre los cuales figura principalmente la alanina.

Fig 2. Síntesis de glúcidos a partir de precursores sencillos.
La gluconeogénesis en mamíferos tiene lugar principalmente en el hígado y en menor extensión en la médula renal. La glucosa producida pasa a la sangre para abastecer a todos los tejidos de los organismos vivos.Es así que después de un ejercicio vigoroso, el lactato producido por la glucólisis anaerobia en el músculo esquelético vuelve al hígado y se convierte en glucosa, que vuelve de nuevo al músculo y es almacenado en forma de glucógeno, a este ciclo se lo conoce como ciclo de Cori.

Fig 3. Ciclo de Cori
Por otro lado, en las semillas germinadas de plantas, las grasas y proteínas almacenadas se convierten a través de rutas entre las cuales se encuentra la gluconeogénesis, en el disacárido sacarosa y en almidón para ser transportados por la planta en desarrollo y posteriormente ser almacenados. La glucosa y sus derivados son los precursores en la síntesis de los componentes de las paredes celulares de las plantas, nucleótidos y coenzimas de un gran número de metabolitos esenciales.
En microorganismos, la gluconeogénesis se produce a partir de compuestos orgánicos sencillos tales como: acetato, lactato y propionato los cuales están presentes en el medio de cultivo.
Las reacciones de la gluconeogénesis son las mismas en todos los organismos, el contexto metabólico y la regulación de la vía difieren de organismo a organismo y de tejido a tejido.
Glucólisis y Gluconeogénesis
La glucólisis y la gluconeogénesis son vías metabólicas que se dan en el citosol de la célula.
La finalidad de la glucólisis es obtener piruvato a partir de glucosa, en cambio la gluconeogénesis tiene como objetivo obtener glucosa a partir de piruvato. Éstas no son rutas idénticas que transcurren en direcciones opuestas, si bien es cierto, comparten la mayoría de los pasos, 7 de las 10 reacciones enzimáticas de la gluconeogénesis son la inversa de las reacciones glucolíticas. Sin embargo, existen 3 reacciones que no se pueden dar en forma inversa para la gluconeogénesis, puesto que existe una razón termodinámica para éstas, debido a que están muy desplazadas en el equilibrio son irreversibles in vivo, éstas reacciones de la glucólisis se muestran en la fig 4.

Fig 4. Reacciones de la glucosa que son irreversibles.
En las células, estas 3 reacciones se catalizan por una gran variación negativa de energía libre, mientras que otras reacciones glucolíticas tienen una energía libre próxima a cero. Esto se lo puede visualizar en la Fig 5.
Fig 5. Variaciones de energía libre en la glucólisis de los eritrocitos. ΔG’° hace referencia a las condiciones estándar. ΔG presenta los cálculos realizados con las concentraciones reales de los
intermediarios glucolíticos en condiciones fisiológicas de los eritrocitos a pH 7.
En la gluconeogénesis, los 3 pasos irreversibles se “rodean” mediante un conjunto diferente de enzimas que catalizan reacciones bastante exergónicas lo que las hace irreversibles.

Fig 6. Reacciones de la Gluconeogénesis.
A continuación se presenta el esquema general de la gluconeogénesis con los pasos comunes que tiene con la glucólisis.
Fig 7. Esquema general de Glucólisis y Gliconeogénesis.
GLUCONEOGÉNESIS
GLUCÓLISIS
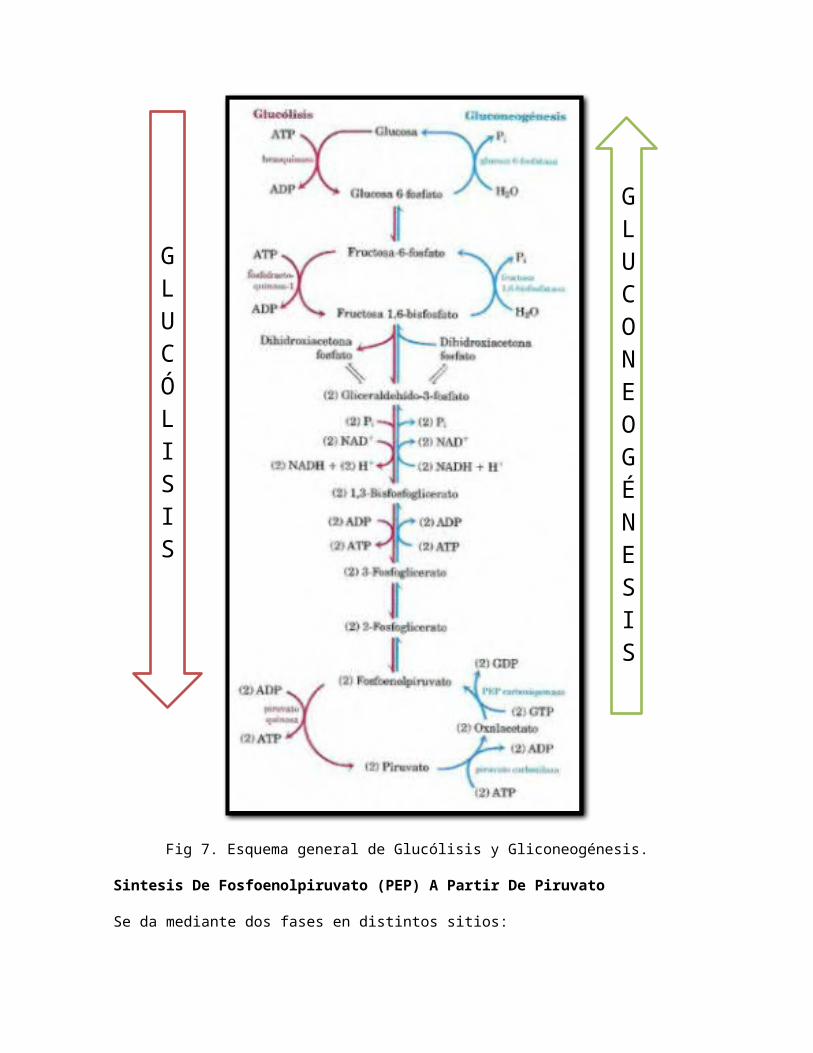
Sintesis De Fosfoenolpiruvato (PEP) A Partir De Piruvato
Se da mediante dos fases en distintos sitios:
a) Conversión de piruvato en oxalacetato en la mitocondria. La membrana mitocondrial no tiene un transportador para el oxalacetato por lo cual este debe reducirse a malato para ser transportado al citosol y asi generar NADH citosólico al continuar con el proceso de formación de de PEP.
b) En el citosol el malato transportado se transforma en oxalacetato al reoxidarse nuevamente:
Malato +NAD+ Oxalacetato + NADH + H+
Posteriormente el oxalacetato es convertido en PEP, gracias a que un compuesto de alta energía denominado GTP proporciona el grupo fosforilo. Esta reacción es reversible pues la formación de un compuesto fosfato de alta energía como es el PEP se compensa con la hidrólisis de otro como es el GTP.En este proceso se usaron 2 equivalentes fosfato de alta energía: uno del ATP y otro del GTP, los cuales proporcionan 50kJ/mol cada uno en el medio celular, a diferencia de en la glucólisis donde solo se obtenía ATP. Si bien la variación de energía libre estándar de la reacción en dos fases desde e l piruvato al PEP es de 0.9kJ/mol, en condiciones fisiológicas su valor real es de -25kJ/mol, esto debido al alto consumo de PEP por otras reacciones que mantienen este compuesto en concentraciones relativamente bajas haciendo de esta reacción un proceso irreversible.Podemos determinar y observar también que el grupo CO2 que se une al piruvato en la mitocondria para formar el oxalacetato es el mismo que se elimina en el CO2 para formar por descarboxilación el PEP. Esta secuencia de carboxilación-descarboxilación permite activar al piruvato para que la descarboxilación del oxalacetato facilite la formación de PEP.Como sabemos el NADH citosólico será usado posteriormente en la gluconeogénesis para producir el gliceraldehído 3-fosfato, por lo cual se comprende que la biosíntesis de la glucosa se da siempre y cuando de suministre este compuesto de reducción. El transporte de malato fuera de la mitocondria y su reconversión a oxalacetato representa una forma de transporte de este equivalente de reducción haciendo del proceso de formación de PEP importante para mantener el equilibrio entre el NADH producido y consumido.
Biotina en Rx de la piruvato carboxilasa
La biotina es una vitamina que en este caso actúa como cofactor unido de forma covalente mediante un enlace amida al grupo alfa-amino de un residuo de lisina, formando la biotinil –enzima. Esta reacción ocurre en dos fases en sitios catalíticos distintos. En el sitio 1 el ión bicarbonato es convertido en CO2 a expensas del ATP, este CO2 se incorpora al biotinil-enzima formando carboxibiotinil-enzima, Luego un largo brazo formado por la biotina y la

cadena lateral de la lisina transportan el CO2 desde el sitio catalítico 1 al 2 en la superficie enzimática donde reacciona con el piruvato formando el oxalacetato y regenerando el biotinil-enzima.
Rutas alternativas desde el piruvato a fosfoenolpiruvato
Cabe especificar que la formación de PEP tiene dos rutas alternativas que dependen del precursor disponible y se ven influenciadas por la disponibilidad del NADH citosólico.Si el precursor es el piruvato la reacción ocurre como ya se la especificó anteriormente, pero el proceso cambia si como precursor se trabaja con el lactato producción por glucólisis en condiciones de hipoxia en eritrocitos o el músculo por ejemplo. Este lactato se convierte en piruvato en el citoplasma de hepatocitos por acción de la lactato deshidrogenasa produciendo NADH citosólico por lo que el transporte del malato fuera de la mitocondria ya no será necesario. El piruvato obtenido entra en la mitocondri y se transforma en oxalacetato de la misma forma que en la anterior ruta especificada y dentro de dicho orgnanelo este compuesto se transforma en PEP por acción de una isozima de la PEP carboxiquinasa, posteriormente el PEP saldrá de la mitocondri para continuar con el proceso de gluconeogénsis.
Si bien hemos hablado de la glucólisis y la gluconeogénesis como opuestas debemos recalcar que tres de sus reacciones en la gluconeogénesis no se realizan por simple reversión, sino que cumplen un rodeo mediante el cual se obtienen los mismos productos que en el caso de la glucólisis sería los reactantes pues la dirección es opuesta.
(Segundo Rodeo): Fru 1,6-bifosfato Fru 6-fosfato
El primer rodeo es la síntesis ya analizada del PEP, el segundo rodeo es la formación de la fructosa 6-fosfato a partir de la fructosa 1,6-bifosfato.En esta reacción la enzima usada es la fructosa 1,6-bifosfatasa dependiente de Mg2+, la cual reemplaza a la PFK-1 usada en la glucólisis. El proceso ocurre mediante la hidrólisis del fosfato en el carbono 1 de la fru 1,6-bifosfato y no por síntesis de ATP. Fructosa 1,6-bifosfato + H2O Fructosa 6-fosfato + PiG´°=-16,3kJ/mol
(Tercer Rodeo):Glucosa 6-fosfato Glucosa libre
El tercer rodeo de la gluconeogénesis es la reacción final para convertir la glucosa 6-fosfato en glucosa libre, catalizada por la glucosa 6-fosfatasa que reemplazaría a la hexoquinasa usada en la glucólisis. En esta reacción al igual que en la anterior se produce una hidrólisis en lugar de la fosforilación del ADP para formar ATP en el caso de que

iríamos en sentido inverso de la glucólisis, lo cual no sería factible, sería energéticamente desfavorable. La enzima usada en este rodeo no se encuentra ni en el cerebro, ni en los músculos por lo que la gluconeogénesis no se realiza en éstos órganos y la glucosa se obtiene por los alimentos ingeridos o por su producción en el hígado y el riñón.





























