Embed Size (px)
Citation preview

Tlamati Sabiduría, Volumen 7 Número Especial 2 (2016)
1
4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 21 y 23 de septiembre 2016
Memorias
Evaluación de mutaciones de Lphn3 asociadas con cáncer en procesos de
adhesión celular mediado por sus ligandos endógenos.
Isamar Piza Sosa (Becaria)
Universidad Autónoma de Guerrero
Unidad Académica de Ciencias Naturales
Programa de Verano UAGro
Área en la que participa: III Medicina y Ciencias de la Salud
Dr. Antony Boucard Jr. (Asesor-Investigador)
Profesor-Investigador del Centro de Investigación y de Estudios Avanzados del Instituto
Politécnico Nacional (CINVESTAV-IPN).
Resumen.
Las Latrofilinas (Lphn) son moléculas de adhesión celular que son parte de la familia de GPCR de adhesión
y se expresan tres isoformas diferentes en diversos tejidos. El papel de Lphn en procesos de adhesión se
ejemplificó principalmente en la estabilidad de las redes neuronales a través de la sinapsis. Sin embargo, la
función de Lphn en otros tejidos sigue siendo una incógnita. En cuanto a la función de Latrofilinas ha venido
de la identificación de ligandos endógenos con la que Lphn puede formar complejos intercelulares. Se cree
que estas interacciones tienen lugar entre Lphn y sus ligandos a través de sus respectivos dominios
extracelulares. Las mutaciones en el dominio GAIN de Lphn3 se han asociado con diversos tejidos que
presentan un fenotipo canceroso. Estas mutaciones son K561N, A760G, D798H, S810L y E811Q. Dado
que los cambios en la adhesión celular es distintivo de la carcinogénesis, hemos tratado de investigar los
efectos de estas mutaciones en la expresión de Lphn3, sobre el perfil de adherencia con respecto a su ligando
engógeno Teneurina 2. El perfil de expresión de las diferentes proteínas de Lphn3 se evaluó con la
transfección de células de mamífero con el ADNc de mutantes de Lphn3 e inmunodetección en ensayos de
Western Blot. Estos ensayos dieron como resultado la observación de que todos los mutantes que se
expresan en diferentes niveles, que la Lphn3 silvestre. Por otra parte, mostraron un patrón diferente de la
proteólisis en el sitio de GPS. Por último, el perfil de adhesión se verificó mediante la realización de ensayos
de agregación celular en los cuales no se observó ningún defecto para cualquiera de los mutantes en
comparación con la LPHN3 silvestre. Estos datos sugirieron que aunque las mutaciones de cáncer afectan
los niveles de expresión de Lphn3 estos cambios no interfieren con las funciones de adhesión.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
2
Palabras Clave: Receptores acoplado a proteinas G, Latrofilinas, Cancer, Adhesion celular, Neuronas,
Sinapsis.
Introducción
En el cerebro, las neuronas forman redes para comunicarse entre sí a través de la formación
de sinapsis, estas son uniones intracelulares que permiten la transferencia de información
transneuronal (1,2). La expresión de moléculas de adhesión dentro de estos compartimentos
asegura la correcta yuxtaposición y proporciona estabilidad a las conexiones neuronales. Las
latrofilinas son moléculas de adhesión que fueron identificadas como receptores de α-latrotoxina,
una neurotóxina del veneno de la araña viuda negra, y también como partes integrales de las
sinapsis neuronales (5,6). Existen tres isoformas de latrofilina (Lphn1, Lphn2, y Lphn3), Lphn1 y
Lphn3 se expresan mayoritariamente en el cerebro, y la expresión de Lphn2 es de manera ubicua
en otros tejidos (7). También Lphn3 se expresa en una proporción menor en tejidos como el
corazón, placenta, páncreas, riñones y testículos (8,9).Las Lphns forman parte de la familia de
receptores de adhesión acoplados a proteínas (aGPCRs), y están constituidos por un extremo
carboxilo intracelular, siete pases transmembranales y un compartimento N-terminal extracelular
integrado por el dominio olfactomedina, un dominio rico en serina/treonina, un dominio de unión
a hormona, y un dominio GAIN (GPCR-Autoproteolysis Inducing) (3,4).
El dominio GAIN comprende 320 residuos, y en él se encuentra un sitio de incisión conocido como
GPS, el cual constituye el punto de proteólisis del receptor. Esta secuencia se encuentra en todos
los miembros de la familia de los aGPCR, y curiosamente, gran parte de las mutaciones en este
dominio se expresan en enfermedades como el cáncer, por lo que el descubrimiento de ligandos
endógenos para el dominio GAIN constituirá un avance importante en el campo de la investigación
del cáncer relacionado con esta familia de moléculas de adhesión (10). Se han descrito tres ligandos
endógenos para Lphn: neurexinas (NRXNs) (17), teneurinas (TEN) (11) y FLRTs (12). Las TEN
son glucoproteínas transmembranales de tipo II, que contienen una secuencia corta N-terminal, una
región transmembranal única y una secuencia larga extracelular que cuenta con múltiples
repeticiones similares a EGF, una región de cisteínas conservadas y repeticiones únicas YD (13).
Los vertebrados poseen cuatro diferentes genes (TEN1, 2, 3 y 4) (14). FLRTs también son una
familia de moléculas de la superficie celular codificadas por tres genes (FLRT1, FLRT2, y
FLRT3). FLRTs se expresan ampliamente en todos los tejidos (15,16), y se componen de 10
repeticiones ricas en leucina extracelular, un dominio de fibronectina de tipo III, un TMR, y una
cola citoplásmica. FLRTs estaban implicados en la señalización de FGF (16), y la adhesión celular
durante el desarrollo (17). Las NRXNs son proteínas transmembranales tipo I pre-sinápticas. En
humanos, existen tres genes que dan lugar a dos isoformas principales: NRXNα y NRXNβ (18).
La interacción de NRXNs con neuroliginas, moléculas de adhesión postsinapticas, es esencial para
el funcionamiento y la especificidad de la sinapsis (19).
Puesto que Lphn3 es un aGPCR, el objetivo de este proyecto fue evaluar el efecto de las
mutaciones K561N, A760G, D798H, S810L y E811Q reportadas previamente en pacientes con
cáncer, en la adhesión celular, utilizando como ligando para Lphn3 a TEN2.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Materiales y Métodos
Bacterias competentes.
Incubamos una colonia de bacterias DH5α en 5 ml de medio SOB (10 mM MgSO4) a 37°C.
Después de 16 horas adicionamos 0.5 ml de cultivo en 50 ml de medio SOB (10 mM MgSO4) el
cual fue incubado a 37°C en agitación constante hasta alcanzar una densidad óptica de 550. El
cultivo fue enfriado en hielo-agua y centrifugado a 4000 rpm durante 12 minutos a 4°C. La pastilla
bacteriana obtenida fue resuspendida en 16 ml de Buffer RF1 frío e incubada por 15 minutos en
hielo-agua. Se realizó una segunda centrifugación respetando los parámetros antes descritos y se
decantó el sobrenadante. Las bacterias fueron resuspendidas en 4 ml de Buffer RF2 frío e incubadas
durante 15 minutos en baño de hielo-agua. Finalmente se realizaron alícuotas de 200 µl en tubos
eppendorf y fueron almacenadas a 70°C.
Transformación de bacterias competentes.
200 µl de bacterias competentes DH5α fueron mezcladas con 10 ng de DNA plasmídico.
La mezcla fue incubada por 30 minutos en hielo y sucesivamente por 90 segundos a 42°C en baño
maría. Se volvió a incubar en hielo durante 2 minutos y se agregó 800 µl de medio SOB. La
suspensión final fue incubada a 37°C durante 45 minutos en agitación constante. Del volumen final
de la suspensión bacteriana, se sembraron 200 µl en cajas de medio LB con ampicilina.
Extracción de Ácidos nucleicos (Midipreparción)
Bacterias DH5α transformadas fueron cultivadas en 50 ml de medio LB por 16 horas a
37°C. El cultivo fue centrifugado a 7000 rpm por 20 minutos. La pastilla bacteriana fue
resuspendida en 2 ml de solución I fría, a la cual también se le agregó 300 µl de solución de
lisozima (10 ng/ ml en Tris Cl 10 mM pH 8.0). Consecutivamente se adicionó 4 ml de solución II
y se incubó durante 10 minutos a Tamb. 4 ml de CH3CL3 y 2 ml de solución III fría fueron agregados
posteriormente, incubando la mezcla total en hielo por 10 minutos. La mezcla fue centrifugada a
4000 rpm por 15 minutos a 4°C. Se recuperó la fase acuosa (contiene el DNA plasmídico) y se le
adicionó 0.6 veces de isopropanol respecto el volumen obtenido. Incubamos durante 10 minutos a
Tamb, y centrifugamos a 5000 rpm por 15 minutos a 4°C. La pastilla de ácidos nucleicos obtenida
fue lavada con etanol al 70% y disuelta en 300 µl de Buffer TE.
Purificación de DNA.
Se adicionó 300 µl de LiCl 5M a los ácidos nucleicos extraídos. La mezcla fue centrifugada
a 10000 rpm por 10 min a 4 °C. Al sobrenadante recuperado se le agregó un volumen equivalente

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
4
de isopropanol. Se centrifugo nuevamente a 10000 rpm por 10 min a Tamb y se decantó el
sobrenadante. La pastilla obtenida fue lavada con etanol al 70% a Tamb y disuelta en 500 µl de
buffer TE pH 8 + RNasa pancreática. Se incubó por 30 min a Tamb y después se agregaron 500 µl
de NaCl 1.6M + PEG 8000 al 13%. La mezcla fue centrifugada a 13000 rpm por 5 min a 4 °C y la
pastilla obtenida fue disuelta en 400 µl de buffer TE pH 8. Consecutivamente se adicionó 400 µl
de fenol y se centrifugó a 13000 rpm por 5 min. La fase acuosa fue separada de la orgánica para
agregarle 200 µl de fenol y 200 µl de cloroformo. Se realizó una segunda centrifugación, se separó
la fase acuosa y se le adicionó 400 µl de cloroformo. Una tercera centrifugación fue realizada para
recuperar la fase acuosa a la cual se le adicionó 100 µl de acetato de amonio 10 M. Posteriormente
se agregó 1 ml de etanol absoluto, se incubó por 10 min a Tamb y centrifugó a 13000 rpm por 5 min
a 4 °C. El ADN obtenido fue lavado con 200 µl de etanol frio al 70% y disuelto en 100 µl de buffer
TE pH 8. Finalmente se cuantificó en el equipo nanodrop 2000 de Thermo SCIENTIFIC.
Transfección con Polietilenimina (PEI).
Realizamos transfecciones transitorias en placas de seis pozos utilizando PEI como medio
para la introducción de plásmidos.En un tubo eppendorf se agregaron 420 µl de medio DMEM no
suplementado, posteriormente 7 µg de ADN plasmídico y finalmente 35 µl de PEI, esto constituye
el complejo ADN-PEI. Este complejo se mantuvo en una relación de 5 μl de PEI [1mg/ml] y 6 μl
de DMEM no suplementado por cada μg de ADN. Una vez formado el complejo se incubó a
temperatura ambiente por 30 min. Pasado este tiempo, se agregó nuevamente medio DMEM no
suplementado hasta completar 1 ml. Se removió el medio de las células a transfectar y el complejo
fue agregado gota a gota en cada pozo. Las células con el complejo fueron incubadas por 30 min
en un ambiente de CO2 al 5 % a 37 °C. Finalmente, se agregó gota a gota 1 ml de medio DMEM
suplementado al 20 % con SFB. Después de 24 horas se removió el medio de las células y se
agregaron 2 ml de medio DMEM suplementado al 10% con SFB.
Extracción de proteínas totales y de membrana
Los cultivos celulares fueron lavados con PBS y almacenados a -70°C por 30 minutos,
después fueron incubados por 30 segundos a 37°C en baño de agua y su posterior tratamiento se
llevó a cabo en agua-hielo. Para la extracción de proteínas totales, se adicionó a cada pozo 200 µl
de buffer de carga 1X y las células fueron removidas con un escraper. Para la extracción de
proteínas de membrana se adicionó 1 ml de PBS frío con BSA+ inhibidor de proteasas, las células
fueron removidas con un escraper y centrifugadas por 5 minutos a 5000 rpm a 4 ˚C. La pastilla
que contiene la fracción membranal, fue solubilizada en Buffer IP frío (1% tritón X-100, 2mM
CaCl2, 2Mm MgCl2, 150mM NaCl, 0.1Mm EDTA, inhibidores de proteasas1X), e incubada por
2 horas a 4°C en rotación suave. Finalmente se centrifugó por 30 minutos a 15000 rpm a 4°C,
recuperando el sobrenadante y almacenándolo a -70°C. Las proteínas extraídas fueron calentadas
a 37˚C por 30 minutos para su análisis por electroforesis.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Western Blot
Las muestras fueron analizadas en geles de poliacrilamida al 8%. Después de la
electroforesis fueron transferidas a membranas de nitrocelulosa (BIO-RAD). La membrana fue
bloqueada con leche al 5% en TBST 1X durante 2 horas. Después de cada incubación fueron
lavadas 3 veces con TBST al 1X por 10 minutos cada una. Para la detección de las proteínas
utilizamos el anticuerpo primario Anti-HA de ratón a una dilución 1:1000 en TBST 1X+BSA al
3%, y al anticuerpo secundario IRDye® 800CW de burro de LI-COR a una dilución de 1:15000 en
TBST 1X+BSA al 3%. Para el revelado de las membranas se utilizó el sistema de imagen
Odyssey® Fc de LI-COR.
Ensayos de agregación celular
Células HEK293T fueron transfectadas por el método de PEI con los plásmidos que
expresaban a Lphn3 y sus diferentes mutantes y también con teneurina 2. Las células fueron lavadas
con PBS y removidas con 500 µl de solución PBS+EGTA+ DNasa. Las resuspensiones se
realizaron utilizando pipetas pasteur siliconizadas. En un tubo eppendorf se agregaron 100 µl de
Lphn3 o alguna de las mutantes (receptor) y 100 µl de teneurina 2 (ligando). Posterior a esto se
agregó 300 µl de solución incubadora (DMEM que contiene 50 mM de HEPES-NaOH (pH 7.4), 10 %
de SFB, 10 mM de CaCl2 y 10 mM de MgCl2) obteniendo un volumen final de 500 µl. la mezcla final
se incubo por 2 horas a Tamb en rotación suave. Después de este tiempo las muestras fueron
analizadas por microscopia de epifluorescencia y los agregados fueron analizados con el programa
imageJ.
Resultados
Identificación de plásmidos por endonucleasas
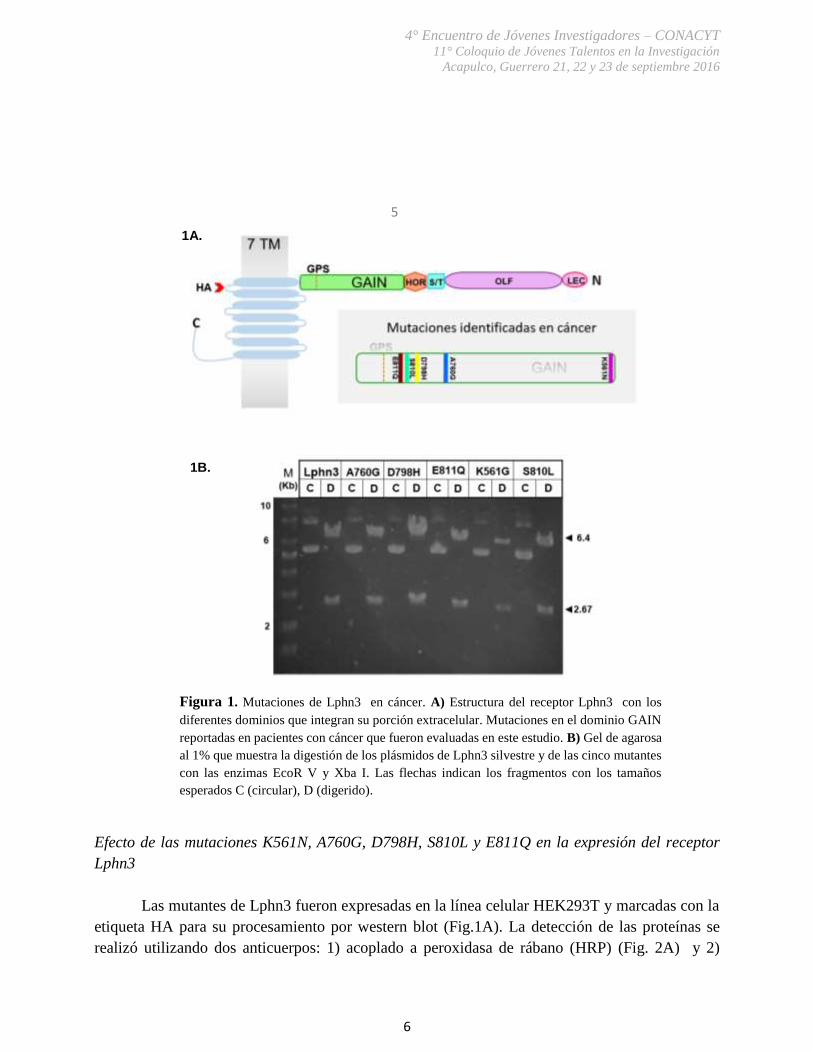
Las construcciones plasmídicas que contienen las mutantes K561N, A760G, D798H,
S810L y E811Q en la secuencia del dominio GAIN de Lphn3 fueron generadas previamente en el
laboratorio (Fig. 1A). Evaluamos por endonucleasas el tamaño de cada uno de los plásmidos
utilizando enzimas que recocieran un sitio de restricción dentro de la secuencia de Lphn3. Las
enzimas utilizadas fueron EcoR V y Xba I. El patrón de digestión generado fueron dos fragmentos,
uno de 2,679 bp y otro de 6,406 bp para todos los plásmidos. La suma de ambos corresponde al
tamaño total del plásmido (9,085 pb) (Fig. 1B). El vector de expresión utilizado para todas las
construcciones fue pCMV.
4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
6
Efecto de las mutaciones K561N, A760G, D798H, S810L y E811Q en la expresión del receptor
Lphn3
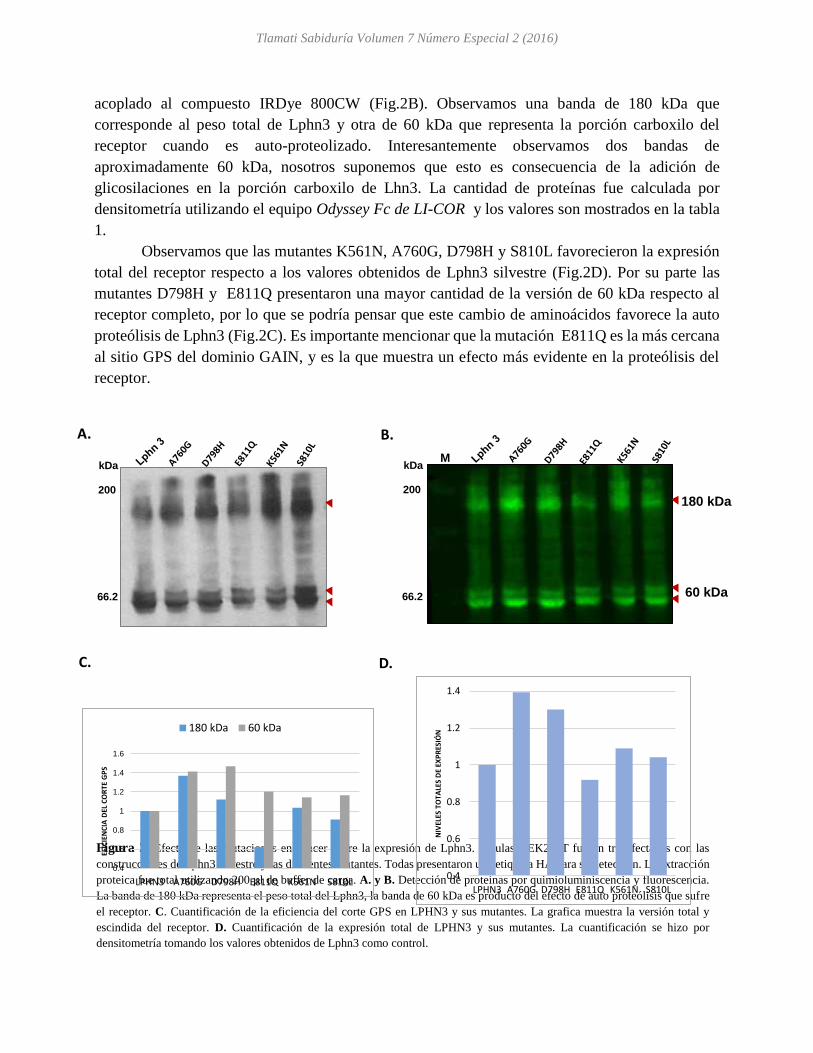
Las mutantes de Lphn3 fueron expresadas en la línea celular HEK293T y marcadas con la
etiqueta HA para su procesamiento por western blot (Fig.1A). La detección de las proteínas se
realizó utilizando dos anticuerpos: 1) acoplado a peroxidasa de rábano (HRP) (Fig. 2A) y 2)
Figura 1. Mutaciones de Lphn3 en cáncer. A) Estructura del receptor Lphn3 con los
diferentes dominios que integran su porción extracelular. Mutaciones en el dominio GAIN
reportadas en pacientes con cáncer que fueron evaluadas en este estudio. B) Gel de agarosa
al 1% que muestra la digestión de los plásmidos de Lphn3 silvestre y de las cinco mutantes
con las enzimas EcoR V y Xba I. Las flechas indican los fragmentos con los tamaños
esperados C (circular), D (digerido).
1A.
1B.
5
Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
acoplado al compuesto IRDye 800CW (Fig.2B). Observamos una banda de 180 kDa que
corresponde al peso total de Lphn3 y otra de 60 kDa que representa la porción carboxilo del
receptor cuando es auto-proteolizado. Interesantemente observamos dos bandas de
aproximadamente 60 kDa, nosotros suponemos que esto es consecuencia de la adición de
glicosilaciones en la porción carboxilo de Lhn3. La cantidad de proteínas fue calculada por
densitometría utilizando el equipo Odyssey Fc de LI-COR y los valores son mostrados en la tabla
1.
Observamos que las mutantes K561N, A760G, D798H y S810L favorecieron la expresión
total del receptor respecto a los valores obtenidos de Lphn3 silvestre (Fig.2D). Por su parte las
mutantes D798H y E811Q presentaron una mayor cantidad de la versión de 60 kDa respecto al
receptor completo, por lo que se podría pensar que este cambio de aminoácidos favorece la auto
proteólisis de Lphn3 (Fig.2C). Es importante mencionar que la mutación E811Q es la más cercana
al sitio GPS del dominio GAIN, y es la que muestra un efecto más evidente en la proteólisis del
receptor.
Figura 2. Efecto de las mutaciones en cáncer sobre la expresión de Lphn3. Células HEK293T fueron transfectadas con las
construcciones de Lphn3 silvestre y las diferentes mutantes. Todas presentaron una etiqueta HA para su detección. La extracción
proteica fue total utilizando 200 µl de buffer de carga. A. y B. Detección de proteínas por quimioluminiscencia y fluorescencia.
La banda de 180 kDa representa el peso total del Lphn3, la banda de 60 kDa es producto del efecto de auto proteólisis que sufre
el receptor. C. Cuantificación de la eficiencia del corte GPS en LPHN3 y sus mutantes. La grafica muestra la versión total y
escindida del receptor. D. Cuantificación de la expresión total de LPHN3 y sus mutantes. La cuantificación se hizo por
densitometría tomando los valores obtenidos de Lphn3 como control.
180 kDa
60 kDa
M
200
66.2
kDa
200
66.2
kDa
0.4
0.6
0.8
1
1.2
1.4
LPHN3 A760G D798H E811Q K561N S810L
NIV
ELES
TO
TALE
S D
E EX
PR
ESIÓ
NA. B.
C. D.
0.4
0.6
0.8
1
1.2
1.4
1.6
LPHN3 A760G D798H E811Q K561N S810L
EFIC
IEN
CIA
DEL
CO
RTE
GP
S
180 kDa 60 kDa
7

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
8
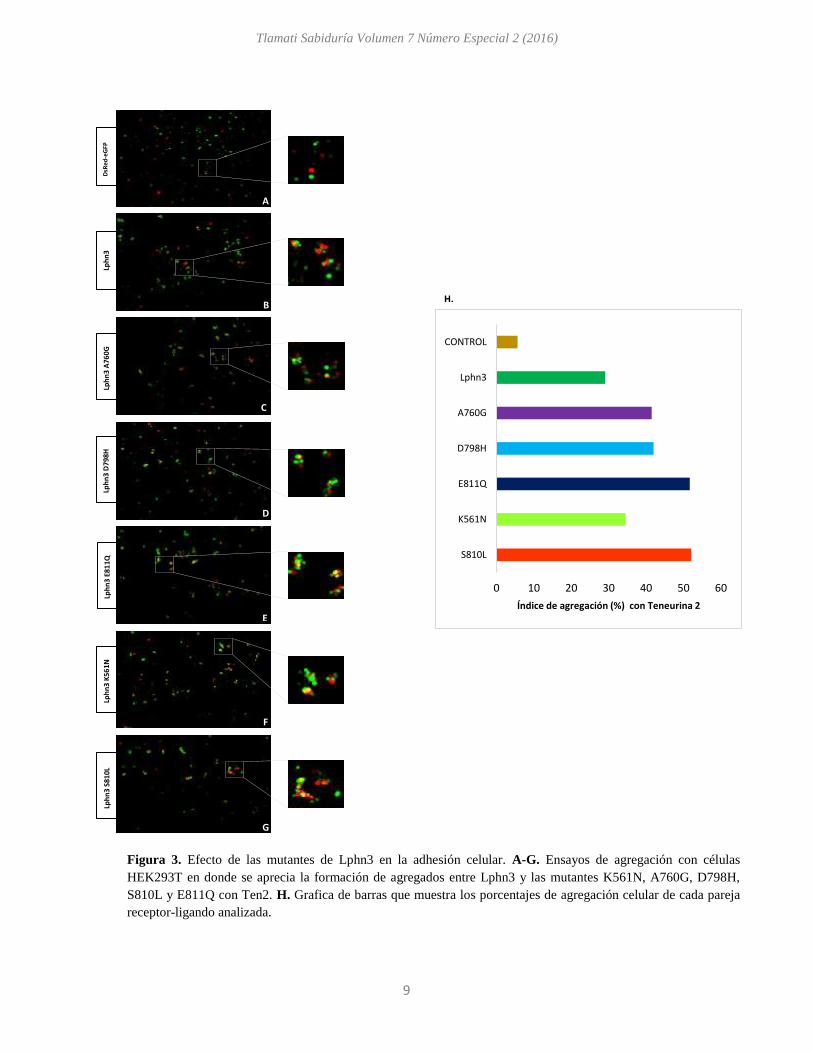
Ensayos de agregación de LPHN3 y sus mutantes en procesos de adhesión celular.
Lphn3 es una molécula de adhesión que ha sido relacionada con procesos de estabilidad y
conectividad neuronal. Estos procesos requieren de la formación de interacciones proteína-proteína
altamente estable. Uno de los ligandos reportados para Lphn3 es la molécula de adhesión teneurina
2. En el 2012 Boucard y colaboradores reportaron que las interacciones Lphn3- ten 2 son altamente
estables que permiten la formación de agregados celulares. Tomando en cuenta lo anterior,
mediante ensayos de agregación celular evaluamos el posible efecto de las mutaciones puntuales
reportadas en cáncer de Lphn3 en la adhesión célula - célula. Para ello co-transfectamos a Lphn3,
K561N, A760G, D798H, S810L y E811Q con eGFP, y a teneurina 2 con DsRed. Realizamos
combinaciones de células que expresaran al receptor (Lphn3) y a su ligando (Ten2), para que la
interacción receptor–ligando promoviera la formación de agregados celulares. Observamos la
formación de agregados celulares con todas las mutantes de Lphn3 al estar en contacto con células
que expresaban a Ten2 (Fig. 3A-G). Las mutantes A760G, D798H, S810L y E811Q presentaron
un aumento en los índices de agregación respecto al receptor silvestre de Lphn3 (Fig. 3H).
Lphn3 A760G D798H E811Q K561N S810L
180 kDa 0.689 0.944 0.772 0.427 0.713 0.628
60 kDa 0.728 1.03 1.07 0.876 0.832 0.848
Tabla 1. Cuantificación proteica de Lphn3 y sus mutantes. La cuantificación se
realizó por densitometría utilizando el sistema de Odyssey de LI-COR. Se
seleccionaron las dos bandas de 60 kDa como única banda para su cuantificación.
Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
A
B
C
D
E
F
G
DsR
ed-e
GFP
Lp
hn
3
Lph
n3
A76
0G
Lph
n3
D79
8H
Lph
n3
E81
1Q
Lph
n3
K56
1N
Lph
n3
S810
L
0 10 20 30 40 50 60
S810L
K561N
E811Q
D798H
A760G
Lphn3
CONTROL
Índice de agregación (%) con Teneurina 2
H.
Figura 3. Efecto de las mutantes de Lphn3 en la adhesión celular. A-G. Ensayos de agregación con células
HEK293T en donde se aprecia la formación de agregados entre Lphn3 y las mutantes K561N, A760G, D798H,
S810L y E811Q con Ten2. H. Grafica de barras que muestra los porcentajes de agregación celular de cada pareja
receptor-ligando analizada.
9

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
10
Discusión y conclusiones
Este proyecto tuvo como propósito evaluar las mutaciones de LPHN3 que están asociadas con
cáncer mediante procesos de adhesión celular mediado por sus ligandos endógenos. Se pretendió
examinar por medio de ensayos de agregación si se presentaba adhesión celular entre el ligando
endógenos de LPHN3 que es Teneurina-2 con las mutantes de LPHN3.
De los resultados obtenidos en esta investigación, se puede deducir que la unión de latrofilina-3
con teneurina 2 puede mediar la unión célula-célula, en células que presentan estas proteínas en su
superficie estas se reconocer y así poder formar agregados. Los ensayos de agregación detectan el
establecimiento de interacción celular. Se llegó a la conclusión de que existe una alta afinidad de
latrofilina-3 y teneurina-2, que son capaces de poder formar uniones intracelulares.
Agradecimientos
Por medio de esta estancia, pude obtener mejores conocimiento y aprendí más técnicas de
laboratorio. Quiero agradecer al verano UAGro por permitirme participar como joven investigador
para el cuarto encuentro de jóvenes investigadores-CONACYT. Quiero agradecer al Dr. Antony
Boucard por aceptarme en el laboratorio de Biología Celular y poder llevar a cabo este proyecto de
investigación, también agradezco a Monserrat Ávila Zozoya que fungió como mi asesora para que
este proyecto se pudiera llevar acabo en tiempo y forma. El proyecto fue financiado con los
recursos de una convocatoria de Ciencia Basica de SEP-CONACyT #221568 y con recursos
federales del CINVESTAV-IPN (otorgados al Doctor Antony Boucard.
Referencias
Benson D. L.,Huntley G. W. (2012) Synapse adhesion: a dynamic equilibrium conferring stability and
flexibility. Curr.Opin. Neurobiol. 22, 397–404.
Missler M., Südhof T. C., Biederer T. (2012) Synaptic cell adhesion. Cold Spring Harbor Perspect.
Biol. 4, a005694.
Sugita S., Ichtchenko K., Khvotchev M., Südhof T. C. (1998) α-Latrotoxin receptor CIRL/latrophilin
1 (CL1) defines an unusual family of ubiquitous G-protein-linked receptors. G-protein coupling not
required for triggering exocytosis. J. Biol. Chem. 273, 32715–32724.
Araç D., Boucard A. A., Bolliger M. F., Nguyen J., Soltis S. M., Südhof T. C., Brunger A. T.
(2012) A novel evolutionarily conserved domain of cell-adhesion GPCRs mediates
autoproteolysis. EMBO J. 31, 1364–1378.
Krasnoperov V. G.,Bittner M. A., Beavis R., Kuang Y., Salnikow K. V., Chepurny O. G.,Little A.
R., Plotnikov A. N., Wu D., Holz R. W., Petrenko A. G. (1997) α-Latrotoxin stimulates exocytosis by
the interaction with a neuronal G protein-coupled receptor. Neuron 18,925–937.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Lelianova V. G., Davletov B. A., Sterling A., Rahman M. A., Grishin E. V., Totty N. F.,Ushkaryov Y.
A. (1997) α-Latrotoxin receptor, latrophilin, is a novel member of the secretin family of G protein-
coupled receptors. J. Biol. Chem. 272, 21504–21508.
Sugita S., Ichtchenko K., Khvotchev M., Südhof T. C. (1998) α-Latrotoxin receptor CIRL/latrophilin
1 (CL1) defines an unusual family of ubiquitous G-protein-linked receptors. G-protein coupling not
required for triggering exocytosis. J. Biol. Chem. 273, 32715–32724.
Ichtchenko K, Bittner MA, Krasnoperov V, Little AR, Chepurny O, Holz RW, Petrenko AG. A novel
ubiquitously expressed alphalatrotoxin receptor is a member of the CIRL family of G-proteincoupled
receptors. J Biol Chem 1999; 274: 5491–8.
Lang J, Ushkaryov Y, Grasso A, Wollheim CB. Ca2+-independent insulin exocytosis induced by
alpha-latrotoxin requires latrophilin, a G protein-coupled receptor. EMBO J 1998; 17: 648–57.
Kan Z, Jaiswal BS, Stinson J, Janakiraman V, Bhatt D, Stern HM, Yue P, Haverty PM, Bourgon R,
Zheng J, Moorhead M, Chaudhuri S, Tomsho LP, Peters BA, Pujara K, Cordes S, Davis DP, Carlton
VE, Yuan W, Li L, Wang W, Eigenbrot C, Kaminker JS, Eberhard DA, Waring P, Schuster SC,
Modrusan Z, Zhang Z, Stokoe D, de Sauvage FJ, Faham M, Seshagiri S. Diverse somatic mutation
patterns and pathway alterations in human cancers. Nature 2010; 466: 869–73.
Silva J. P., Lelianova V. G., Ermolyuk Y. S., Vysokov N., Hitchen P.
G., Berninghausen O.,Rahman M. A., Zangrandi A., Fidalgo S., Tonevitsky A.
G., Dell A., Volynski K. E., Ushkaryov Y. A.(2011) Latrophilin 1 and its endogenous ligand
Lasso/teneurin-2 form a high affinity transsynaptic receptor pair with signaling capabilities. Proc. Natl.
Acad. Sci. U.S.A. 108,12113–12118.
O'Sullivan M. L., de Wit J., Savas J. N., Comoletti D., Otto-Hitt S., Yates J. R. 3rd., Ghosh A.
(2012) FLRT proteins are endogenous latrophilin ligands and regulate excitatory synapse
development. Neuron 73, 903–910.
Oohashi T., Zhou X. H., Feng K., Richter B., Mörgelin M., Perez M. T., Su W. D.,Chiquet-
Ehrismann R., Rauch U., Fässler R. (1999) Mouse ten-m/Odz is a new family of dimeric type II
transmembrane proteins expressed in many tissues. J. Cell Biol. 145,563–577.
Boucard AA, Maxeiner S, Sudhof TC. Latrophilins function as heterophilic cell-adhesion molecules
by binding to teneurins: regulation by alternative splicing. J Biol Chem 2014; 289:387–402.
Lacy S. E., Bönnemann C. G.,Buzney E. A., Kunkel L. M. (1999) Identification of FLRT1, FLRT2,
and FLRT3: a novel family of transmembrane leucine-rich repeat proteins.Genomics 62, 417–426.
Böttcher R. T., Pollet N., Delius H., Niehrs C. (2004) the transmembrane protein XFLRT3 forms a
complex with FGF receptors and promotes FGF signalling. Nat. Cell Biol. 6, 38–44.
Maretto S.,Müller P. S., Aricescu A. R., Cho K. W., Bikoff E. K., Robertson E. J. (2008)Ventral
closure, headfold fusion and definitive endoderm migration defects in mouse embryos lacking the
fibronectin leucine-rich transmembrane protein FLRT3. Dev. Biol.318, 184–193.
Ushkaryov, Y. A., A. G. Petrenko, M. Geppert, and T. C. Südhof. 1992. “Neurexins: Synaptic Cell Surface
Proteins Related to the Alpha-Latrotoxin Receptor and Laminin.” Science (New York, N.Y.) 257 (5066):
50–56.
Tsetsenis, Theodoros, Antony A. Boucard, Demet Araç, Axel T. Brunger, and Thomas C. Südhof. 2014.
“Direct Visualization of Trans-Synaptic Neurexin-Neuroligin Interactions during Synapse Formation.” The
11

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
12
Journal of Neuroscience: The Official Journal of the Society for Neuroscience 34 (45): 15083–96.
doi:10.1523/JNEUROSCI.0348-14.2014.





























