Embed Size (px)
Citation preview

1. Introducción
Uno de los principales grupos de plagas para la agricultura chilena lo constituyen los
chanchitos blancos, dentro de los cuales destacan, Pseudococcus calceolariae,
Pseudococcus longispinus, Pseudococcus viburni y Planococcus citri.
El chanchito blanco de los cítricos (Planococcus citri (Risso) es una de las especies más
dañinas para la citricultura y afecta principalmente a los huertos que se encuentran entre
la IV y la VI Región (Ripa y Rodríguez, 1999). La importancia de esta especie recae en
que es una de las plagas más dañinas para los cítricos, el cual actualmente es el grupo de
frutales de mayor producción a nivel mundial (Agustí, 2004).
El chanchito blanco de los cítricos es controlado por varios enemigos naturales,
depredadores como Cryptolaemus montrouzieri Muls y Sympherobius maculipennis
Kimm; y parasitoides como Leptomastidea abnormis Girault y Coccidoxenoides peregrina
Timberlake.
La elección de criar Leptomastidea abnormis versus otros enemigos naturales, se
fundamenta, primero, en su forma de acción. Al igual que todos los parasitoides, es
especialista, es decir, sólo puede completar su ciclo de desarrollo en un único hospedero,
siendo en este caso P. citri. Esta característica es vital si se compara con la acción de los
depredadores como Cryptolaemus montrouzieri, los cuales son generalistas, es decir, se
alimentan de diferentes especies, en este caso de chanchito blanco.
El utilizar un controlador biológico generalista puede significar, en algunos casos, la
diseminación de éste a huertos vecinos en búsqueda de otros focos de plagas, ya sea por
preferencia o por un mayor grado de infestación.

Otra característica positiva es que parasita al P. citri en estadíos juveniles, principalmente
en primer y segundo estadío ninfal, lo que dificulta su diseminación, ya que durante esta
fase del desarrollo es cuando las ninfas migran a la zona de la roseta o al ombligo de los
cítricos. Es de gran importancia el control durante esta etapa, pues cuando las ninfas se
sitúan en esas zonas, es muy difícil lograr controlarlas con depredadores o con productos
químicos. Cabe destacar que la presencia de chanchito blanco en cítricos es causal de
rechazo para las exportaciones.
Con respecto a la crianza de uno u otro parasitoide, se seleccionó L. abnormis por su
facilidad de crianza en laboratorio, sobretodo si se compara con Coccidoxenoides
peregrina, la cual por su pequeño tamaño presenta la capacidad de escaparse fácilmente
de las baterías de crianza y contaminar otras. Por su parte L. abnormis también presenta
una buena adaptación a las condiciones climáticas de la zona central de Chile, sector
donde se concentran las principales plantaciones de cítricos; lo que facilita su posterior
establecimiento a largo plazo.
1.1 Objetivos
1.1.1 Objetivo general
- Establecer un pie de cría de Leptomastidea abnormis para uso comercial.
1.1.2 Objetivos específicos
- Establecer una crianza de laboratorio para Planococcus citri
- Recuperar en terreno el parasitoide Leptomastidea abnormis
- Producir en laboratorio Leptomastidea abnormis

2. Revisión bibliográfica
2.1 Consideraciones generales sobre control biológico
El manejo integrado de plagas (MIP) nace como una alternativa al manejo tradicional, el
cual está enfocado principalmente al control de las plagas mediante el uso de control
químico con pesticidas de amplio espectro, y generalmente, de largo efecto residual, el
que además afecta frecuentemente a los enemigos naturales de estos, evitando su acción
(Inia, 2006).
El MIP utiliza racionalmente los métodos químicos, biológicos y culturales para enfrentar
un problema de plagas (Inia, 2006).
El MIP se basa en la manipulación del agroecosistema con el fin de evitar que las plagas
alcancen niveles de densidad que sean ecológicamente dañinos. Esta manipulación
requiere: identificar la plagas y sus enemigos naturales, el daño y en que nivel estas
plagas lo causan; conocer el efecto del control químico sobre la plaga y sobre los
enemigos naturales, como también utilizar conjuntamente el control cultural (Ripa y
Rodríguez, 1999).
El control biológico en Chile se inicia a partir del año 1903 con la introducción de
enemigos naturales a los campos, sin conocer a fondo su biología y las posibles
interacciones con otras poblaciones de insectos, causando muchas veces graves errores
de internación (Rojas, 2005).
Como primera regla, debe conocerse muy bien la plaga, su morfología, ciclo de vida,
donde vive y sus hábitos, así como también a sus controladores biológicos y el daño real

que causa a un cultivo en específico, para luego determinar como ejercer un control sobre
ella (Ripa y Rodríguez, 1999).
Según Apablaza (2000) las plagas agrícolas son insectos, ácaros y otros
microorganismos que dañan plantas cultivadas o sus productos.
Se debe diferenciar entre plagas primarias y secundarias. Las plagas primarias son
aquellas que producen daños directos e indirectos de significación económica para el
cultivo, aún a niveles bajos de densidad. En cambio, plagas secundarias son aquellas que
están o no permanentemente asociadas a un cultivo, y que generalmente producen daños
subeconómicos (González, 1989).
El control cultural se refiere principalmente a las siguientes técnicas: manejos cero
labranza, eliminación de malezas hospederas de insectos plagas, rotación de cultivos, uso
de abonos orgánicos y uso de barreras entre cultivos entre otros, que tengan como
objetivo disminuir la población de la plaga (Apablaza, 2000).
Según González y Rojas (1966) el control biológico consiste en la regulación y supresión
del potencial reproductor de organismos a través de la acción de parasitoides, predatores
(depredadores) y patógenos. Desde que el hombre interviene en esta interacción se le ha
llamado control biológico, en cambio el control natural se refiere al que ocurre sin
intervención del hombre.
Para Capdeville (1945) el control biológico trata de la simbiosis entre los animales. Entre
los carnívoros se encuentran algunas interacciones como el predatismo y el
parasitoidismo, los cuales se explican a continuación.

El predatismo, es aquella forma de simbiosis en la cual uno de los simbiontes ataca a un
individuo o varios individuos de una o más especies, con el propósito de alimentarse de
él. El individuo así vive del otro y se le llama predador.
El predador es un insecto entomófago que durante su vida se alimenta de varios
individuos huéspedes y que completa su evolución fuera de cualquiera de ellos.
Parasitoidismo es aquella forma de simbiosis en la que un simbionte vive dentro o fuera
del cuerpo del organismo huésped, alimentándose a su expensa durante todo su período
de desarrollo. El individuo que así vive se le llama parasitoide.
Un parasitoide es un insecto entomófago que se alimenta exclusivamente de un individuo
huésped y termina su evolución por lo general dentro de dicho huésped.
Ambas categorías, predatismo y parasitoidismo, poseen una similitud general como un
parámetro en la dinámica de las poblaciones, porque son letales y tienden a ser factores
de mortalidad (De Bach, 1968).
2.2 Características generales del chanchito blanco de los cítricos
El chanchito blanco de los cítricos es uno de los fitófagos más agresivos de los cítricos, y
de otras especies como el chirimoyo (Llorens, 1990).
Se diferencia a grandes rasgos de otras especies por presentar secreciones algodonosas
de una longitud más o menos igual. Cabe destacar que para una determinación

taxonómica exacta se debe recurrir a un examen con microscopio (Ripa y Rodríguez,
1999).
Este insecto manifiesta principalmente dos tipos de daños, uno directo al insertar su
estilete y succionar savia, lo que en ataques serios debilita la fruta causando su caída
(Capdeville, 1945), e indirecto porque segrega melaza (sustancia azucarada). Esta
melaza es expulsada por el ano (el chanchito levanta la parte posterior de su cuerpo y
abre los dos últimos filamentos y así proyecta la gota de melaza a cierta distancia de
donde se encuentra) y sirve de sustrato al hongo fumagina. Se termina por depreciar al
fruto tanto por su presencia como por la del chanchito blanco que causa rechazo en las
exportaciones al ser detectado. Además, la fumagina produce una cubierta negra que
interfiere con los procesos fisiológicos normales de la planta (Capdeville, 1945). También
atraídas por la melaza aparecen las hormigas (Llorens, 1990).
Según Capdeville (1945) la acción en masa de los chanchitos blancos frecuentemente
provoca la defoliación de los árboles debido a los disturbios fisiológicos que causa.
El chanchito blanco de los cítricos se incluye en el Orden Hemíptera, el cual aporta un
elevado porcentaje de plagas a la fruticultura. La clasificación taxonómica para esta plaga
es la siguiente:
Orden : Hemíptera
Suborden : Coccinea
Superfamilia : Coccoidea
Familia : Pseudococcidae
Genero : Planococcus
Especie : citri

Los chanchitos blancos o cochinillas tienen cuerpo blando, desnudo, segmentado y
recubierto de secreciones céreas en forma de polvillo blanco o algodonoso. Son móviles
en todos los estados de desarrollo. Los estados larvarios son muy similares a las formas
adultas, sólo se diferencian en su tamaño y convexión. Presentan patas y antenas bien
visibles (Llorens, 1990).
Una vez que la hembra es fecundada esta se fija y empieza a producir una tenue
secreción algodonosa que originará el ovisaco algodonoso; como se trata de una especie
ovípara (Capdeville, 1945) los huevos se depositan en sacos sueltos u ovisacos. Según
Rojas (2005) cada hembra ovipone entre 400 a 500 huevos, pudiendo alcanzar hasta casi
mil unidades.
Las hembras adultas tienen forma oval, abombada dorsalmente. Estas producen largos
filamentos céreos que protegen la puesta en la parte posterior de su cuerpo. (Ripa y
Rodríguez, 1999). Generalmente se mantienen fijas, sin perder su capacidad de caminar
ya que conservan los tres pares de patas funcionales.
Por su parte los machos, forman un pupario alargado de textura filamentosa y color
blanco debajo de las hembras donde sufren dos mudas, para, al final del proceso emerger
como adulto alado (Llorens, 1990).
Esta especie pasa el invierno en todos los estados, excepto como macho adulto. En el
año se producen entre 2 a 4 generaciones dependiendo del clima (Ripa y Rodríguez,
1999).
Estos individuos son gregarios, es decir, forman colonias de individuos agrupados entre
ellos. Generalmente forman sus colonias sobre frutos, hojas y ramillas que se encuentran

en zonas poco aireadas de los árboles. Los daños más llamativos se encuentran en
huertos con alta humedad, poca ventilación y poda deficiente (Llorens, 1990).
Según Prado (1991) se encuentra a Planococcus citri asociado a los siguientes cultivos:
caqui, chirimoyo, guayabo, limonero, mandarino, mango, naranjo y pomelo. Para lo
anterior se determina que Planococcus citri puede ser considerada una plaga primaria, es
decir que comúnmente necesita control durante la temporada; pero también puede ser
plaga secundaria para otros, que puede o no necesitar control.
A su vez, González (1989) clasifica a los hospedantes del chanchito blanco de los cítricos
en primarios y secundarios. En la primera clasificación se encuentra al chirimoyo, los
cítricos y el granado; en cambio en la segunda se encuentran el caqui, mango, y algunas
plantas ornamentales.
Las colonias de chanchito blanco se alojan principalmente en el pedicelo, en la unión
entre frutos y entre el fruto y la hoja. En los cítricos es posible encontrarlos en la base del
fruto (bajo la roseta), lo que en el caso de frutos para exportación origina problemas de
rechazo dada la dificultad para identificar los estados ninfales (Ripa y Rodríguez, 1999).
2.3 Control biológico de Planococcus citri
Planococcus citri presenta una amplia lista de enemigos naturales, tanto depredadores
como parasitoides los que se detallan en el Cuadro 1.
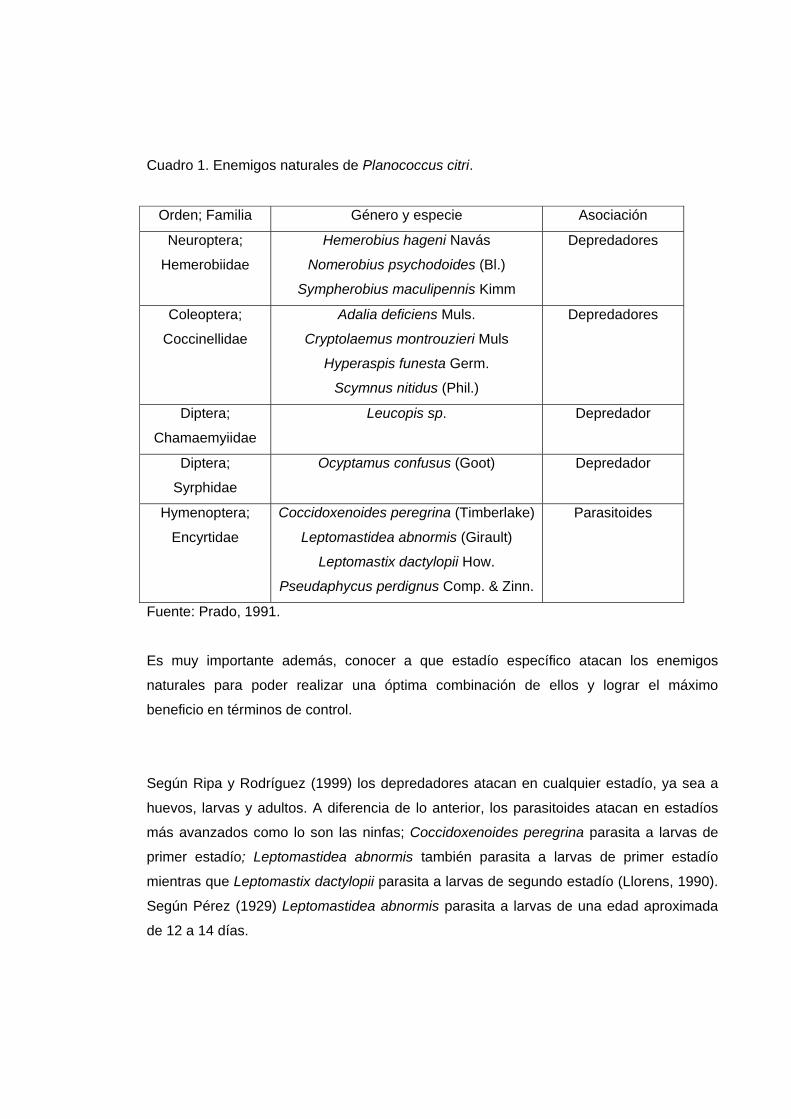
Cuadro 1. Enemigos naturales de Planococcus citri.
Orden; Familia Género y especie Asociación
Neuroptera;
Hemerobiidae
Hemerobius hageni Navás
Nomerobius psychodoides (Bl.)
Sympherobius maculipennis Kimm
Depredadores
Coleoptera;
Coccinellidae
Adalia deficiens Muls.
Cryptolaemus montrouzieri Muls
Hyperaspis funesta Germ.
Scymnus nitidus (Phil.)
Depredadores
Diptera;
Chamaemyiidae
Leucopis sp. Depredador
Diptera;
Syrphidae
Ocyptamus confusus (Goot) Depredador
Hymenoptera;
Encyrtidae
Coccidoxenoides peregrina (Timberlake)
Leptomastidea abnormis (Girault)
Leptomastix dactylopii How.
Pseudaphycus perdignus Comp. & Zinn.
Parasitoides
Fuente: Prado, 1991.
Es muy importante además, conocer a que estadío específico atacan los enemigos
naturales para poder realizar una óptima combinación de ellos y lograr el máximo
beneficio en términos de control.
Según Ripa y Rodríguez (1999) los depredadores atacan en cualquier estadío, ya sea a
huevos, larvas y adultos. A diferencia de lo anterior, los parasitoides atacan en estadíos
más avanzados como lo son las ninfas; Coccidoxenoides peregrina parasita a larvas de
primer estadío; Leptomastidea abnormis también parasita a larvas de primer estadío
mientras que Leptomastix dactylopii parasita a larvas de segundo estadío (Llorens, 1990).
Según Pérez (1929) Leptomastidea abnormis parasita a larvas de una edad aproximada
de 12 a 14 días.

2.4 Características generales de Leptomastidea abnormis
Leptomastidea abnormis se clasifica taxonómicamente (Prado, 1991) de la siguiente
manera:
Orden : Hymenoptera
Superfamilia : Chalcidoidea
Familia : Encyrtidae
Genero : Leptomastidea
Especie : abnormis
La super familia Chalcidoidea incluye varias familias de importantes parasitoides de
plagas, lo que es muy beneficioso ya que estos atacan a insectos de diversos órdenes
(Apablaza, 2000).
La familia Encyrtidae se caracteriza por ser un grupo amplio de insectos pequeños, que
comúnmente son parasitoides de homópteros, pulgones, escamas, mosquitas blancas y
en algunos casos de lepidópteros (Apablaza, 2000).
L. abnormis es originaria de Sicilia, Italia (González y Rojas, 1966) y fue introducida en
Chile el año 1931 desde EEUU por A. Graf, y posteriormente esta especie fue internada
en varias oportunidades (durante los años 1933, 1934 y 1939) hasta que se logró su
establecimiento, junto a otra especie más como de Coccidoxenoides peregrina (Rojas,
2005).

Leptomastidea abnormis es una avispita de color pardo; el cuerpo del adulto mide
alrededor de 0,75 mm con antenas largas. Las alas presentan bandas transversales de
color negruzco (Ripa y Rodríguez, 1999).
Se caracteriza también por presentar una cabeza bastante grande, uniformemente
coloreada de amarillo claro, a excepción de las mandíbulas y los ocelos, que son de color
rojo rubí, y los grandes ojos compuestos de aspecto amarillento, con tornasoles verdosos.
El tórax es amarillo testáceo, con numerosas cerdas gruesas casi blancas. El abdomen es
de color amarillo mas claro (Pérez, 1929).
L. abnormis es fácilmente distinguible de otros parásitos por su apariencia llamativa, que
deriva de su costumbre de posarse y colocar sus alas en alto. Un ala la coloca dando la
impresión de que esta estuviera fracturada en su base, y además presenta bandas
características que sirven para identificarla (Capdeville, 1945).
El apareo de machos y hembras se verifica con toda normalidad a una temperatura de 16º
a 18º C, generalmente hacia el medio día, durante unos 15 min. Acto seguido se dispone
la hembra a comenzar la puesta (Pérez, 1929).
Este encírtido es un parasitoide interno solitario (Capdeville, 1945) específico de
Planococcus citri. Esto significa que la hembra adulta ovipone en el chanchito y dentro de
él se desarrolla el parasitoide, lo que finalmente produce muerte del hospedero. Durante
este período ocurren algunos cambios en la forma del insecto parasitado, el cual pasa a
denominarse momia y contiene en su interior al parasitoide. La momia de L. abnormis es
de color anaranjado, de forma ovoidal y su tamaño no supera 1 mm de largo (Ripa y
Rodríguez, 1999).

La oviposición por parte del parasitoide se efectúa casi tan pronto el insecto ha terminado
su desarrollo. Las hembras adultas oviponen en los estadíos primarios del chanchito
blanco, siendo sus huevos depositados libres en la hemolinfa (Inia, 2006). En campo, la
regla es encontrar un huevo por cada chanchito blanco parasitado. El desarrollo desde
huevo a adulto demora entre 25 y 45 días dependiendo de la temperatura (Capdeville,
1945).
La temperatura a su vez influye en dos factores muy importantes en el desarrollo de la
dinámica poblacional, la descendencia y la longevidad de los adultos. Al aumentar
progresivamente la temperatura entre los 18º y 24º C, aumenta en relación directa la
descendencia, pero después de ese rango y hasta los 34º C casi no existe variación. La
longevidad es inversamente proporcional a la temperatura, es decir, disminuye la
longevidad al aumentar la temperatura. Sin embargo, en comparación con otros parásitos
del P. citri, como lo son Leptomastix dactylopii y Anagyrus pseudococci, Leptomastidea
abnormis es el que se adapta mejor a un rango mucho más amplio de temperaturas
(Tingle y Copland, 1989).
El huevecillo depositado en la especie víctima es sumamente pequeño. Es de color
blanco, con una membrana semi transparente, que debe de ser muy flexible para que
pueda pasar sin romperse por el oviscapto de la hembra (Pérez, 1929).
Los huevos fertilizados del parasitoide dan origen a individuos hembras y los no
fertilizados a machos. Al nacer la larva, ésta comienza a alimentarse de la hemolinfa de P.
citri y en sus últimos estadíos termina por consumir totalmente el contenido del cuerpo
ocasionando su muerte (Inia, 2006).
De la biología estudiada por Pérez (1929) se deduce que el ciclo evolutivo completo es de
unos 33 días en los meses de calor, y de unos 37 a 45 en los de más frío. Esto ha de
suponer que el insecto tuviera de cuatro a cinco generaciones por año.

3. Materiales y métodos
La crianza de Leptomastidea abnormis se realizó en el laboratorio de Control Biológico
ubicado en la Facultad de Agronomía de la Pontificia Universidad Católica de Valparaíso
(La Palma, Quillota) durante el año 2006.
En el laboratorio, la sala de crianza presentó las siguientes condiciones: humedad relativa
al menos de 60%, y una temperatura promedio de 23º a 24º C. Para mantener esta
temperatura se utilizó una estufa y paralelamente funcionó un extractor para remover y
extraer el aire interior (Anexo 1).
3.1 Crianza en laboratorio de Planococcus citri:
El trabajo comenzó con la captura en terreno de colonias de chanchito blanco de la
especie Planococcus citri, durante los meses de marzo y abril. Se recolectó material en
huertos de cítricos y chirimoyos en la zona de Quillota, los que presentaban antecedentes
de ataques de chanchito blanco.
Se colectó colonias tanto de ninfas como de adultas, de éstas últimas se identificó
claramente las que estaban comenzando la postura de huevos a través de la
característica lanosidad blanca que las recubre.
La identificación de campo se verificó en el laboratorio con la ayuda de una lupa
estereoscópica. Con el uso de una aguja, se separaron las hembras adultas de la colonia.
Éstas se colocaron en cajas de acrílico con tapa con muselina fina para facilitar la
respiración del insecto y mantenerlas separadas de posibles depredadores.

Se comenzó la crianza con hembras adultas ya que a partir de ellas se obtienen nuevas
colonias puras. Si se utilizaran masas de huevos, estos podrían contener entre ellos
huevos de depredadores que luego contaminarían la crianza. Por otra parte si se
utilizaran ninfas, estás podrían estar parasitadas de campo lo que también contaminaría la
crianza.
En las cajas de acrílico se mantuvo a las hembras adultas por un período de 7 a 10 días
y se esperó la postura de masas de huevos por parte de las hembras. Para facilitar la
postura se mantuvieron las cajas a una temperatura estable de 22º a 23º C.
Cuando las hembras adultas presentaron una masiva producción de masas de huevos,
comenzó la inoculación del chanchito blanco sobre el sustrato.
Las baterías de crianza tipo Flanders son cajas de madera con una tapa de vidrio, esta
permite la fácil observación a través del vidrio y el manipuleo durante la crianza. En el
fondo de la batería se colocó papel de diario seco, para absorver la humedad que se
genera dentro de la batería y evitar el contacto directo entre la mielecilla que produce el
chanchito blanco y la madera. Luego, dentro de la batería se colocó el sustrato.
Como sustrato se utilizó zapallo tipo cacho y papas brotadas, ambos aportados por la
empresa Xilema S.A. El uso de uno u el otro depende principalmente de la fecha en que
se deba realizar la crianza ya que ésta determina la disponibilidad del sustrato; es así
como en invierno se tuvo disponible zapallo y en primavera papas brotadas.
En el caso de utilizar zapallo tipo cacho, este debe colocarse dentro de la sala de crianza
aproximadamente 1 h antes de colocarlo dentro de la batería para que se acondicione a la
temperatura de la sala (ya que se almacenan en un lugar de menor temperatura). En el
caso de no tomar esta precaución, la superficie del zapallo podría condensarse y ser

causante de una posterior pudrición de los mismos. Transcurrido este tiempo, se
colocaron tres atriles de madera y sobre cada uno de ellos los zapallos. Los atriles se
utilizaron para mantener fija la posición del zapallo y evitar que se movieran y aplastaran
parte de la colonia.
Es importante revisar los zapallos con cierta frecuencia, ya que el chanchito blanco, en
especial P. citri, tiene la capacidad de generar mucha melaza lo que favorece el
humedecimiento de los zapallos, y por lo tanto los hace más sensibles a ataques de
hongos. Los zapallos que se contaminaron con hongos se eliminaron de inmediato, pero
antes, se rescataron las colonias de chanchito blanco con el uso de agujas y se colocaron
sobre nuevos zapallos para evitar perder ese material.
Cuando se utilizó papas brotadas como sustrato la inoculación se llevó a cabo en una sala
de crianza aparte. Las papas se deben lavar con agua y cloro utilizando una esponja
suave. Se debe tener la precaución de no dañar ningún ojo (brote) de la papa para evitar
herirla. Luego de la limpieza, se deben secar al sol por unas 5 h. Luego las papas se
colocaron en bandejas plásticas. Cerca de ellas se colocaron zapallos que presentaban
grandes colonias de chanchito blanco. Sobre los brotes de las papas se colocaron masas
de huevos y ninfas para que comenzara la fijación de la colonia, pero también hubo
inoculación por la simple migración de las ninfas presentes en los zapallos. Cuando los
brotes estaban cubiertos por la colonia, se trasladaron las bandejas a las baterías. Se
colocaron dos bandejas por cada batería tipo Flanders.
En el momento en que la población de chanchito blanco aumentó significativamente, es
decir, cuando se observó gran producción de masas de huevos y fijación de las ninfas, se
decidió duplicar las baterías.
Para duplicar las baterías, se colocaron también tres zapallos dentro de la batería, pero
sólo los de los costados presentaban colonias sobre ellos, en cambio, el zapallo del medio

era nuevo. Las ninfas de las colonias migran fácilmente de un zapallo al otro. Mediante
este procedimiento se multiplican las colonias y se obtiene la segunda generación de
chanchito blanco ya criado en laboratorio. Este proceso se llevó a cabo varias veces
durante el invierno para mantener en todo momento nuevas colonias de chanchito blanco.
A finales de septiembre se había conseguido un total de seis baterías.
3.2 Recuperación de Leptomastidea abnormis en terreno:
El material colectado en campo que se utilizó en el método anterior, es decir, las muestras
de frutos de cítricos y chirimoyos, ramillas y hojas infestadas con chanchito blanco se
colocaron dentro de una batería tipo Flanders.
En este caso fue importante limpiar meticulosamente el vidrio que cubre la batería puesto
que se requiere mantener la mejor visibilidad para observar fácilmente si hay nacimiento
de parasitoides.
La observación de las baterías se realizó diariamente, ya que además se debió revisar si
había nacimiento de otros enemigos naturales depredadores como ocurrió con el
coccinélido Cryptolaemus montrouzieri. En este caso se deben retirar las larvas y los
adultos de éste para evitar la depredación de la colonia.
Al cabo de unos días, fue posible observar nacimiento de adultos de L. abnormis. A
medida que nacen los parasitoides, deben examinarse para determinar que efectivamente
correspondan a la especie y no a otro parasitoide que también pudiera encontrarse en el
campo.

Al confirmar que los ejemplares que habían nacido eran efectivamente L. abnormis estos
se trasladaron a las baterías en las cuales estaba establecida la colonia del hospedero.
Gran parte de los individuos de la población correspondían a ninfas de primer y segundo
estadío.
3.3 Producción en laboratorio de Leptomastidea abnormis:
Cuando comenzó el nacimiento del parasitoide se empezó a formar el pie de cría. Los
adultos capturados se trasladaron a las baterías de crianza del chanchito blanco para que
comenzara su reproducción y así lograran parasitar a las ninfas de la colonia de
chanchitos.
Con el incremento de la población de L. abnormis se pudo seguir extrayendo adultos de
las baterías y trasladarse a otras para aumentar su crianza. La extracción fue llevada a
cabo con un aspirador especial. Este esta formado por un tubo de vidrio con un tapón de
goma en la parte superior, perforado en dos partes del cual se conecta una de estas
perforaciones con una manguera plástica desde la cual se realiza la aspiración.
Durante esta etapa es esencial mantener un stock de baterías de crianza con colonias de
chanchito blanco en activo crecimiento. Esto para utilizar los hospederos en el momento
oportuno en que nacen los parasitoides.
Al finalizar esta etapa se dio por concluida la formación del pie de cría de L. abnormis, lo
que después podría generar un sistema de crianza masiva de este parasitoide.

4. Resultados y discusión
4.1 Desarrollo del método de crianza
Etapa 1. Obtención del sustrato
Las papas pueden conseguirse todo el año, teniendo en cuenta que siempre se debe
comprar papas maduras, es decir, que estén completamente suberizadas (lo que significa
que tengan por lo menos unos cuatro meses de producción en la tierra), ya que las papas
pelonas (las que aparecen en el mercado a partir de septiembre) son papas nuevas, es
decir, poco suberizadas, lo que las hace muy sensible al roce, por lo tanto muy delicadas
de manipular. Se compran en partidas de 50 kg, ya que facilita el traslado, la manipulación
y la guarda. A medida que se requiera más sustrato se deberá comprar.
Los zapallos cacho se compran durante el verano. Luego se almacenan en una bodega
con buena ventilación. Es importante que los zapallos no estén en contacto uno con el
otro para evitar daños.
Etapa 2. Tratamiento del sustrato
El primer paso fue lavar las papas y luego secarlas. Además se puede utilizar algún
fitorregulador para acelerar el proceso de brotación. Este procedimiento se realiza
principalmente en la temporada de otoño invierno, debido a que durante estas fechas la
brotación de los tubérculos tarda mucho más que durante la primavera y el verano. La
hormona que se utiliza es el ácido giberélico. Un producto comercial en base a este
ingrediente es Giberplus, distribuido por la empresa Anasac. Este producto viene
formulado en tabletas. La dosis recomendada por Anasac para acelerar la brotación de
los tubérculos de papas es de 5 ppm. Esto quiere decir que se debe disolver 5 g de
ingrediente activo formulado en 100 l de agua. Luego de preparar la solución deben

sumergirse los tubérculos en ella. El tiempo de inmersión es de 5 min. Este procedimiento
puede disminuir el tiempo de brotación durante la época invernal a la mitad. Alrededor de
la tercera semana se obtienen los resultados (Anasac, 2006).
Etapa 3. Brotación de las papas
Para comenzar la brotación de los tubérculos, estos se colocaron en cajones de madera
en una sala oscura. La oscuridad es un factor imprescindible para generar brotes
etiolados, es decir, brotes sin clorofila. Ellos se caracterizan por poseer un tejido mucho
más blando que los brotados a plena luz. Esta característica es esencial para facilitar la
fijación de las ninfas de chanchito blanco mas adelante.
Se utilizaron cajones de madera de 30 X 40 cm. Dentro de cada cajón se colocaron entre
25 y 30 papas.
Cuando aparecen los primeros brotes, se debieron recortar promoviendo la brotación
lateral del mismo, aumentando así considerablemente la superficie donde pudieronn
fijarse las ninfas de chanchito blanco.
Se consideró que el sustrato estaba en condiciones para continuar la próxima etapa
cuando se consiguieron brotes con las siguientes características:
- brotes de no menos de 2 mm de diámetro y
- brotes de por lo menos 2 cm de largo
La duración de esta etapa varía de acuerdo a las condiciones ambientales. En los meses
de invierno puede tardar 8 a 12 semanas, y en primavera - verano desde dos semanas a
un mes.

Cuando terminó el proceso de brotación, las papas se colocaron en bandejas plásticas de
30 x 25 x 6 cm. Se colocaron entre 11 a 18 papas por bandeja, con un promedio de 15. La
cantidad dependerá del tamaño de las mismas. Cada bandeja tiene un peso promedio de
1.9 kg, lo que equivale a que cada papa tenga un peso promedio de 125 g.
Etapa 4. Inoculación con Planococcus citri
Previo a la inoculación se recortaron los brotes para dejarlos de un largo similar y evitar
que los brotes demasiado largos se rompieran al manipular las bandejas.
Para la inoculación se pueden colocar directamente masas de huevos sobre las papas, o
si ya se ha empezado una crianza de Planococcus citri en otro sustrato como zapallo,
estos se pueden colocar al lado de las papas e inocularse simplemente por el movimiento
de las ninfas, las cuales colonizarán el nuevo sustrato.
En el caso de realizar la inoculación de las papas a partir de otro sustrato como el zapallo
tipo cacho, se debe evitar el contacto de ambos con otros enemigos naturales como
ocurre con el Cryptolaemus montrouzieri ya que si hay postura de huevos, estos son casi
imperceptibles y difíciles de diferenciar de las masas de huevos de los propios chanchitos
blancos. Si ocurre, es muy probable que las larvas de C. montrouzieri se alimenten de las
ninfas de chanchito blanco y por lo tanto es de esperar que devoren las ninfas que ya han
sido parasitadas por Leptomastidea abnormis y por consiguiente también se alimente del
parasitoide en cuestión.
Se estimó que la inoculación con Planococcus citri había finalizado cuando se observó a
simple vista los brotes de un color amarillento que indicaba la presencia de ninfas.
Idealmente se requieren ninfas de primer y segundo estadío. Todos los brotes deben
estar colonizados aparentemente igual. Cuando esto se logró, se colocaron las bandejas
plásticas dentro de las baterías tipo Flanders. Se colocaron dos bandejas por batería.

Etapa 5. Inoculación con Leptomastidea abnormis
En el momento que comenzó el nacimiento de adultos de Leptomastidea abnormis, y a su
vez se tuvo colonias de ninfas de P. citri en los brotes de papas, estas últimas se
inocularon con el parasitoide para continuar su multiplicación. Se liberó un cierto número
de individuos por batería (dosis).
Etapa 6. Cosecha de Leptomastidea abnormis
Transcurrido el tiempo de desarrollo de L. abnormis, emergieron los adultos del
parasitoide. Estos individuos debieron ser recolectados y contabilizados para inocular
nuevas baterías y así continuar con la crianza. Para colectarlos se utilizó un aspirador
sencillo, con el cual se succiona el aire que lo rodea y por lo tanto al mismo parasitoide en
sí. Para facilitar la captura del parasitoide se trabajó en la sala de crianza en oscuridad a
excepción de una lámpara que se colocó muy cerca del vidrio de la batería. Este efecto
estimula a los parasitoides a acercarse a la fuente de luz, por lo que se posan sobre el
vidrio lo que facilita su recolección.
4.2 Formación del pie de cría de P. citri
A partir de la recolección de colonias de chanchito blanco de campo en el mes de abril, se
obtuvo hacia finales de septiembre un total de seis baterías con colonias de P. citri,
utilizando zapallo cacho como sustrato. Se comenzó con dos baterías, y al transcurrir
cuatro meses éstas se habían triplicado.
Utilizando papas como sustrato sólo se formaron tres baterías durante el mes de
septiembre. Esta vez, el proceso fue más acelerado puesto que se contaba con más
material para llevar a cabo la inoculación del sustrato.

4.3 Caracterización de los sustratos
Los zapallos que se utilizan son de la especie Cucurbita moschata. Debe elegirse los
zapallos que presenten las siguientes características: base redondeada, de tamaño
mediano, cuello corto, con presencia de un trozo de pedúnculo y que no presente ningún
tipo de herida ni infecciones causadas por hongos u otros tipos de microorganismos. El
peso ideal es de 1.5 kg.
Las papas (Solanum tuberosum L.) que se utilizan como sustrato son de la variedad
Desiree y Yagana, aunque se debiera preferir la primera si hay disponibilidad de ambas,
ya que ésta genera brotes de mayor tamaño.
Cuadro 2. Peso promedio por papa antes de la brotación
Número papas /kg Peso/papa (gramos) Peso promedio por papa
7 115, 120, 200, 150, 115,
120, 180
140
6 180, 195, 170, 180, 180 ,
120
170
6 200, 100, 160, 180, 180,
190
160
7 100, 180, 170, 120, 100,
110, 170
135
5 200, 210, 200, 130, 220 192
Mediante los datos expuestos en el Cuadro 2, se calcula el promedio de papas por kg,
antes de la brotación es de 6,2 papas con un peso promedio de 159,4 g cada una.

4.4 Brotación de las papas
Las papas se colocaron en cajones de madera porque por sus poros se permite la
ventilación, en cambio las cajas de acrílico (plástico) no la permiten, lo que aumentaría la
humedad en la sala y requeriría reposición de ellas periódicamente. Debajo de las papas
se colocó papel de diario para que absorviera la humedad que se genera.
La sala de brotación debe presentar entre 10º y 12º C, y una humedad entre 65 y 75%.
Este proceso demora aproximadamente tres meses, pero se puede acelerar con los
reguladores de crecimiento anteriormente citados.
Las papas que se deben utilizar para la crianza de chanchito blanco deben presentar
ciertas características que se describen en el Cuadro 3. Además luego de la brotación las
papas sufren ciertos cambios como la pérdida de peso (Cuadro 4).
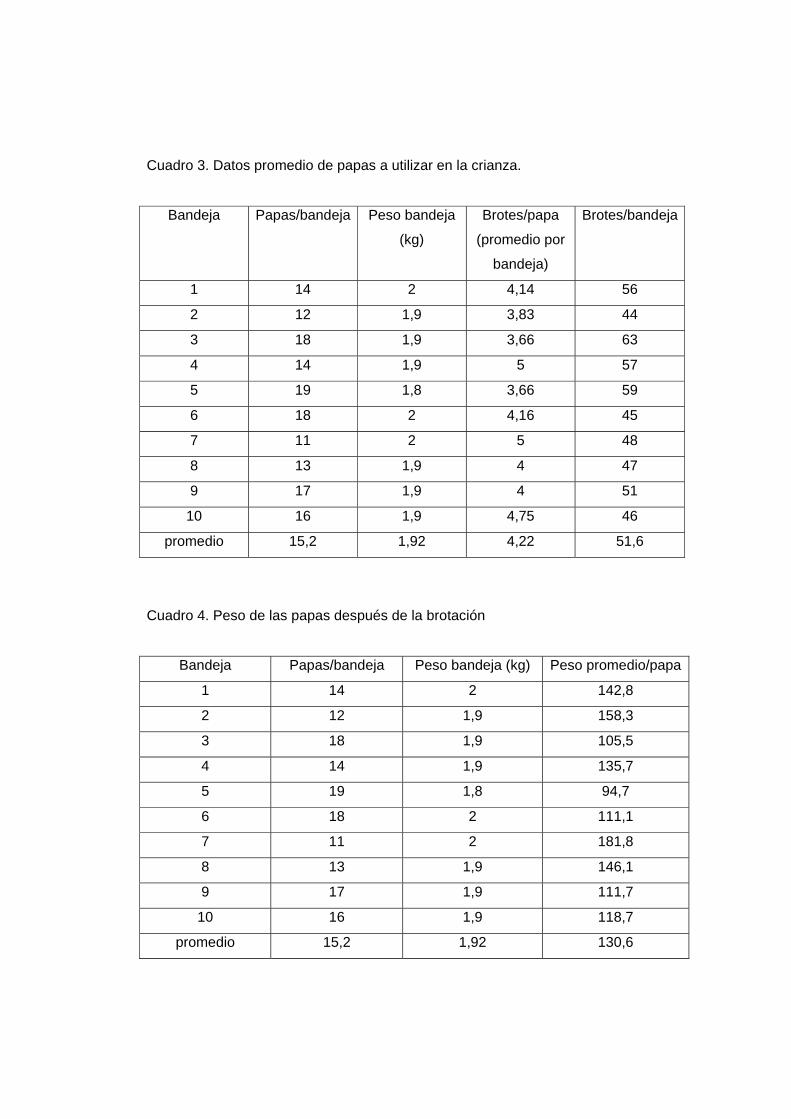
Cuadro 3. Datos promedio de papas a utilizar en la crianza.
Bandeja Papas/bandeja Peso bandeja
(kg)
Brotes/papa
(promedio por
bandeja)
Brotes/bandeja
1 14 2 4,14 56
2 12 1,9 3,83 44
3 18 1,9 3,66 63
4 14 1,9 5 57
5 19 1,8 3,66 59
6 18 2 4,16 45
7 11 2 5 48
8 13 1,9 4 47
9 17 1,9 4 51
10 16 1,9 4,75 46
promedio 15,2 1,92 4,22 51,6
Cuadro 4. Peso de las papas después de la brotación
Bandeja Papas/bandeja Peso bandeja (kg) Peso promedio/papa
1 14 2 142,8
2 12 1,9 158,3
3 18 1,9 105,5
4 14 1,9 135,7
5 19 1,8 94,7
6 18 2 111,1
7 11 2 181,8
8 13 1,9 146,1
9 17 1,9 111,7
10 16 1,9 118,7
promedio 15,2 1,92 130,6

En base a los pesos promedios entre papas no brotadas (159,4 g) y papas brotadas
(130,6 g) se calcula la primera pérdida que ocurre por pérdida de peso. La pérdida de
peso promedio (causado por pérdida de agua) fue de 16%. Este resultado se deberá tener
en cuenta en el momento de realizar los cálculos para determinar la cantidad de sustrato
a comprar.
Considerando lo evaluado anteriormente, al comprar 50 kg de papas, con un peso
promedio de 125 g cada una, se obtendrían alrededor de 400 papas, y si éstas se colocan
en bandejas de 15 papas se lograría tener 26 bandejas lo que equivale a 13 baterías
Flanders.
4.5 Inoculación con Leptomastidea abnormis
La primera inoculación se realizó con L. abnormis proveniente del campo. Los datos del
Cuadro 5 corresponden al total de adultos inoculados en las baterías.
Cuadro 5. Individuos provenientes de campo inoculados por batería.
Batería Individuos inoculados
1 29
2 26
3 165
4 75
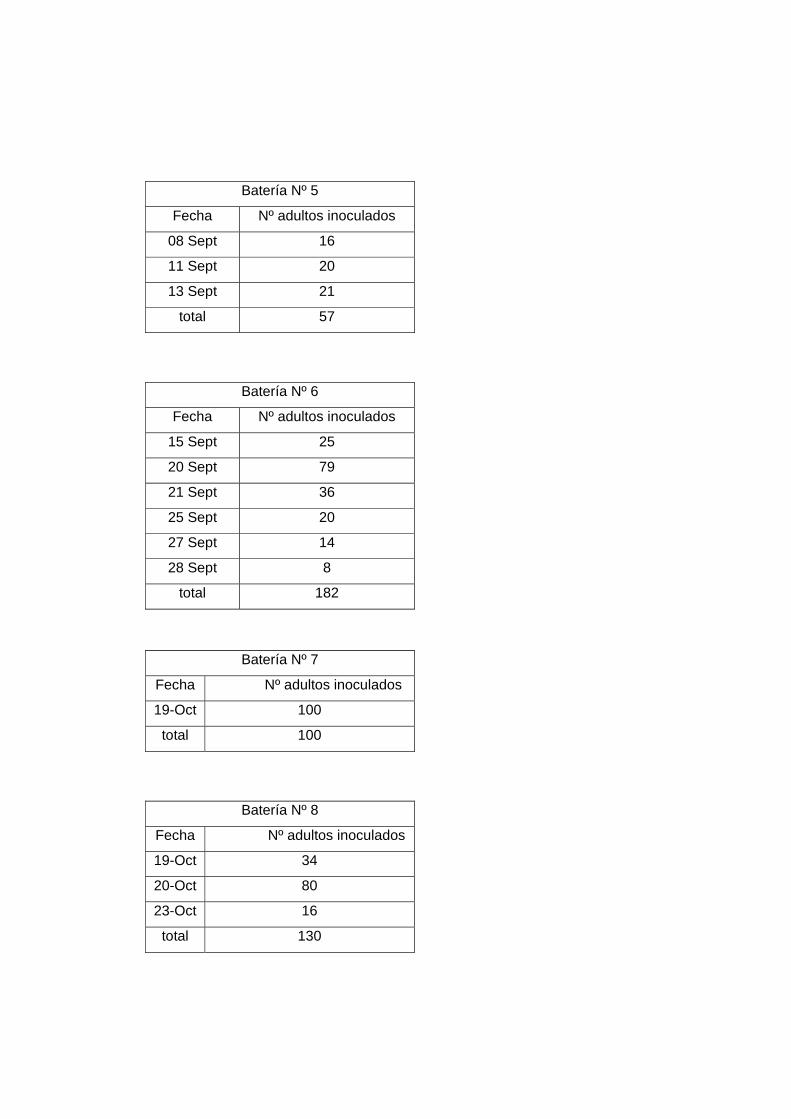
Nota: El desglose de individuos inoculados por batería y las fechas en que se realizaron
están descritos en el Anexo 2.

Durante esta primera fase, se inocularon cuatro baterías con un total de 295 individuos.
Sólo hubo nacimiento del parasitoide en la batería Nº 4.
En las primeras tres baterías no hubo nacimiento del parasitoide porque hubo pudrición
de los zapallos cacho. Una de las principales causas de estas pudriciones es la alta
humedad relativa (70%) y mediana a alta temperatura (24º C) que se mantiene dentro de
la sala de crianza, la cual no cuenta con sistemas automatizados que mantengan la sala
en las condiciones óptimas. Estas condiciones son óptimas para el desarrollo de hongos.
Otra de las causales puede ser problemas en la manipulación de los zapallos, durante la
cosecha, el transporte y el almacenaje de éstos, siendo una de las principales causas
deterioro por golpes que pueda predisponer la entrada de microorganismos a través de la
piel del zapallo, y que luego al estar sometidos a alta temperatura y alta humedad relativa
(medio óptimo para multiplicación de microorganismos) el proceso de pudrición se haya
acelerado notablemente.
De la batería Nº 4 hubo nacimiento de 239 individuos, los cuales se inocularon
nuevamente en otras dos baterías (Cuadro 6). Estos individuos corresponden a la primera
generación de L. abnormis en el laboratorio.
Cuadro 6. Inoculación de adultos correspondientes a la primera generación obtenida en
laboratorio.
Batería Individuos inoculados
5 57
6 182

Nuevamente sólo hubo nacimientos en una de las baterías, en este caso en la batería Nº
6. Los zapallos de la batería Nº 5 se pudrieron.
En la batería Nº 6 nacieron 430 individuos (segunda generación obtenida en el
laboratorio) los que se inocularon en otras tres baterías (Cuadro 7). La diferencia con las
primeras crianzas, es el cambio del sustrato, esta vez la crianza se realizó sobre papas.
Este cambio se efectuó porque ya no había disponibilidad de zapallo cacho en el
mercado, y también porque se esperaba que las papas fueran más resistentes a
problemas como las pudriciones que atacaron a los zapallos.
Cuadro 7. Inoculación con adultos correspondientes a la segunda generación obtenida en
laboratorio.
Batería Individuos inoculados
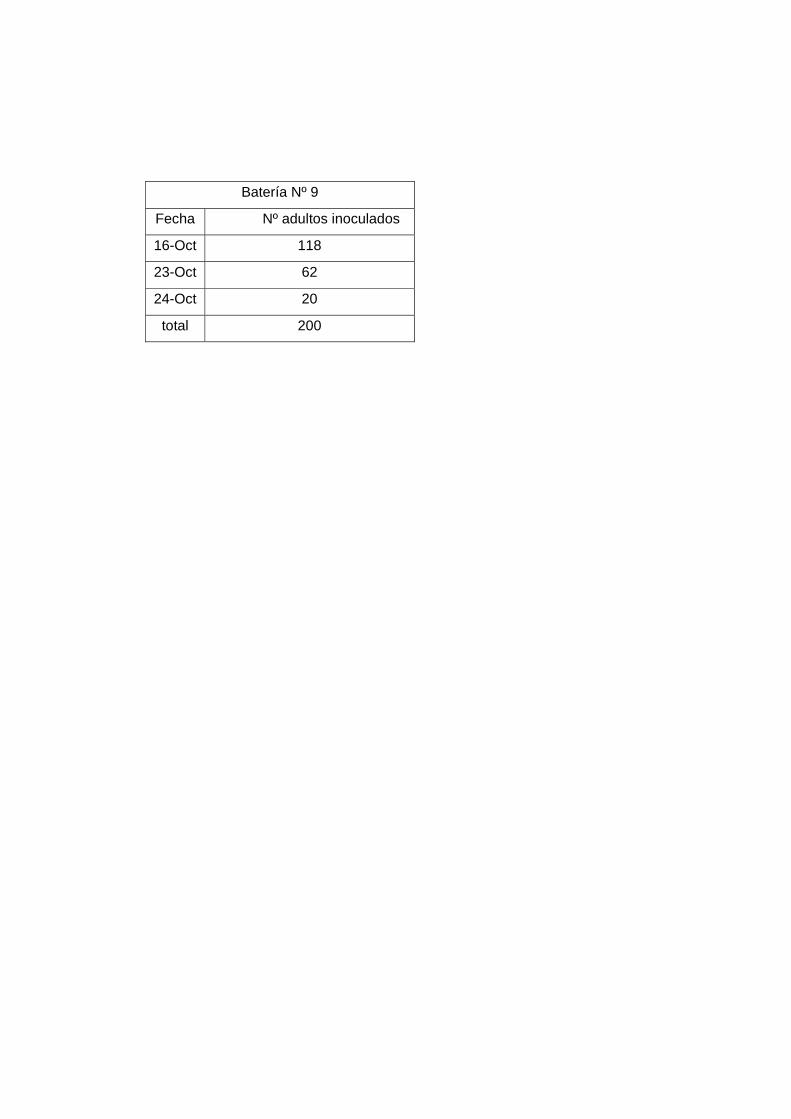
7 100
8 130
9 200
De estas baterías (Nº 7, 8, 9) sólo hubo nacimiento esporádico de algunos individuos y en
fechas en que no se esperaban (al menos dos meses después de la inoculación).
Una de las principales causas de este problema podría ser el aumento de las
temperaturas lo que afecta diversos procesos que finalmente determinan el éxito o no de
la crianza.
Primero, la temperatura afecta el apareamiento. Durante esta fase (mes de octubre) la
temperatura de la sala se mantuvo al menos en unos 22º C como promedio, lo que supera

la temperatura ideal para el apareamiento que rodea los 16º y 18º C que señala Pérez
(1929) en su estudio sobre esta especie.
La temperatura también afecta la tasa de oviposición. Como señalan Tingle y Copland
(1989) el parasitoidismo de esta especie alcanza valores máximos a temperaturas
moderadas, lo que no siempre ocurrió en el mes de octubre fecha en la que ocurrió la
oviposición, ya que ciertos días la sala podía alcanzar temperaturas cercanas a los 27º C
y más. Además, el grado de parasitoidismo desciende a medida que transcurren los días
(Tingle y Copland, 1989), lo que afectaría la tasa de oviposición si los individuos fueron
inoculados en la nueva batería luego de unos días de haber nacido.
Otra causa podría ser que la temperatura se mantuvo sobre el umbral de desarrollo del
hospedero, causando muerte de los estados juveniles del P. citri, esto ocurre porque las
ninfas son el estadío de desarrollo más susceptibles a las temperaturas debido a que no
poseen protección, como si ocurre en otros estadíos. Si en la colonia, la cantidad de
ninfas de segundo y tercer estadío es pequeña, la posibilidad de obtener altos
rendimientos en términos de nacimiento de parasitoides es muy baja puesto que como
señala Capdeville (1945) la regla es esperar nacimiento de un parasitoide por cada
individuo hospedero.
En el Cuadro 8 se presentan los datos de las inoculaciones realizadas.
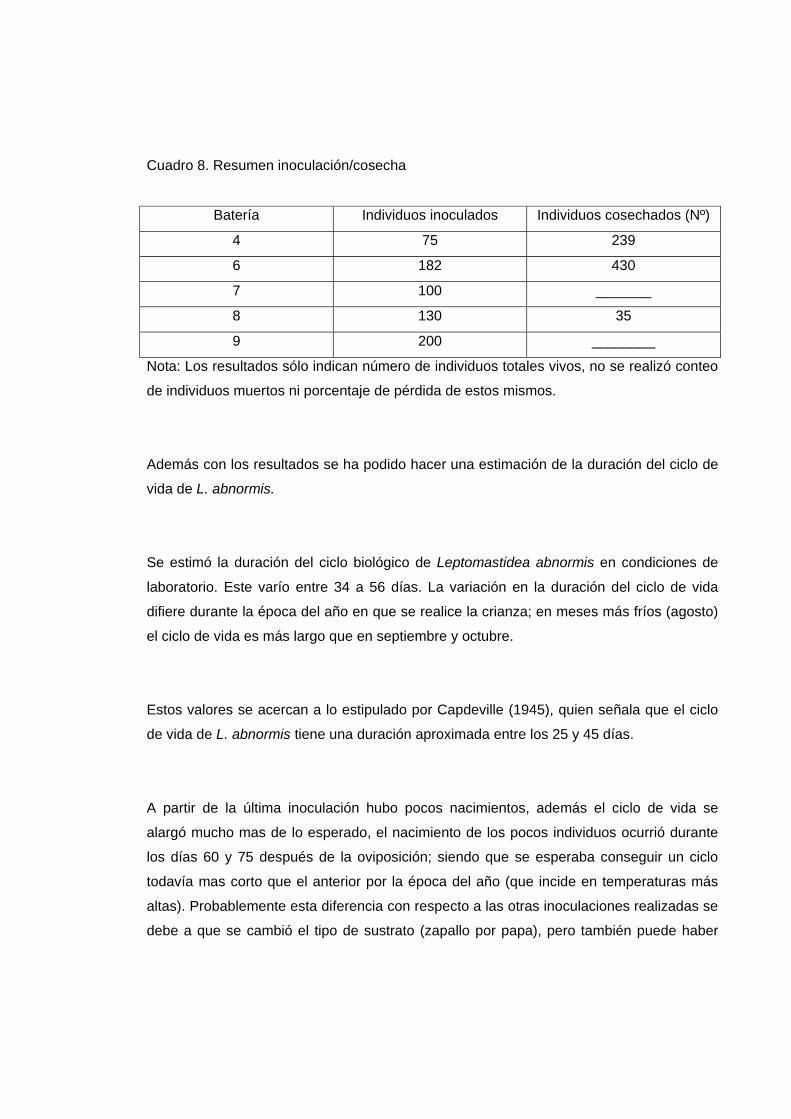
Cuadro 8. Resumen inoculación/cosecha
Batería Individuos inoculados Individuos cosechados (Nº)
4 75 239
6 182 430
7 100 _______
8 130 35
9 200 ________
Nota: Los resultados sólo indican número de individuos totales vivos, no se realizó conteo
de individuos muertos ni porcentaje de pérdida de estos mismos.
Además con los resultados se ha podido hacer una estimación de la duración del ciclo de
vida de L. abnormis.
Se estimó la duración del ciclo biológico de Leptomastidea abnormis en condiciones de
laboratorio. Este varío entre 34 a 56 días. La variación en la duración del ciclo de vida
difiere durante la época del año en que se realice la crianza; en meses más fríos (agosto)
el ciclo de vida es más largo que en septiembre y octubre.
Estos valores se acercan a lo estipulado por Capdeville (1945), quien señala que el ciclo
de vida de L. abnormis tiene una duración aproximada entre los 25 y 45 días.
A partir de la última inoculación hubo pocos nacimientos, además el ciclo de vida se
alargó mucho mas de lo esperado, el nacimiento de los pocos individuos ocurrió durante
los días 60 y 75 después de la oviposición; siendo que se esperaba conseguir un ciclo
todavía mas corto que el anterior por la época del año (que incide en temperaturas más
altas). Probablemente esta diferencia con respecto a las otras inoculaciones realizadas se
debe a que se cambió el tipo de sustrato (zapallo por papa), pero también puede haber

sido causado por problemas externos a la misma batería como son cambios en las
condiciones de la sala, probablemente muy altas temperaturas en conjunto con una
menor humedad relativa y una mala ventilación de la sala que pudo haber creado un
ambiente inhóspito para el nacimiento de L. abnormis.

5. Conclusiones
Se logró establecer un método de crianza para el parasitoide Leptomastidea abnormis.
Este método varia según la época del año, ya que esta influye en que sustrato se
encuentra disponible. Aún cuando los resultados no fueron exitosos para considerar un
crianza comercial, la metodología es la apropiada para lograrlo si se cuenta con mayor
tecnología, sobretodo para mantener las condiciones ambientales óptimas en la sala de
crianza, manteniendo un estricto control sobre la temperatura y la humedad relativa.
Obtener la plaga a parasitar desde el campo es un procedimiento simple pero que
requiere de tiempo y programación. La plaga debe colectarse durante los meses de marzo
y abril, donde se encuentran hembras adultas en el campo. Para lograr reproducirla y
obtener una abundante población de individuos en el estadío de desarrollo necesario al
momento de inocular con el parasitoide.
Recuperar el parasitoide en terreno para comenzar a establecer una crianza es un
método engorroso, pues se debe realizar en el momento del año en que los parasitoides
se encuentran en actividad de oviposición. Si este procedimiento no se realiza en las
fechas adecuadas (otoño o primavera) se entorpece y atrasa todo el proceso de crianza.

6. Literatura citada
Agustí, M. 2004. Fruticultura. 493 p. Mundi-Prensa, Madrid, España.
Anasac. 2006. Giberplus. Disponible en http://www.anasac.cl/. Leído el 20 Octubre 2006.
Apablaza, J. 2000. Introducción a la entomología general y agrícola. 339 p. Universidad
Católica de Chile, Santiago, Chile.
Capdeville, C. 1945. Plagas de la agricultura en Chile. 355 p. Imprenta Pacífico, Quillota,
Chile.
De Bach, P. 1968. Control biológico de las plagas de insectos y malas hierbas. 949 p.
C.E.C.S.A, México DF., México
González, R.H. y S. Rojas. 1966. Estudio analítico del control biológico de las plagas
agrícolas en Chile. Agricultura Técnica 26 (4): 133-147.
González, R. 1989. Insectos y ácaros de importancia agrícola y cuarentenaria en Chile.
310 p. Universidad de Chile, Santiago, Chile.
Inia. 2006. Manejo integrado de plagas en cítricos. Disponible en http://www.mipcitricos.cl.
Leído el 8 Junio 2006.
Llorens, J.M. 1990. Homóptera I: cochinillas de los cítricos y su control biológico. 260 p.
Pisa Ediciones, Valencia, España.
Pérez, M. 1929. Biología del chalcidid Leptomastidea abnormis (Girault), parásito de
Pseudococcus citri Risso. Rec. Roy. Sp. Soc. Natl. Hist. 15: 73-80.

Prado, E. 1991. Artrópodos y sus enemigos naturales asociados a plantas cultivadas en
Chile. 204 p. Serie Boletín Técnico Nº 169. Instituto de Investigaciones Agropecuarias,
Santiago, Chile.
Ripa, R. y F. Rodríguez. 1999. Plagas de cítricos, sus enemigos naturales y manejo. 151
p. Instituto de Investigaciones Agropecuarias, La Cruz, Chile.
Rojas, S. 2005. Control biológico de plagas en Chile: historia y avances. 123 p. Instituto
de Investigaciones Agropecuarias, La Cruz, Chile.
Tingle, C.C.D. and M.J.W. Copland. 1989. Progeny production and adult longevity of the
mealybug parasitoids Anagyrus pseudococci, Leptomastix dactylopii and Leptomastidea
abnormis (Hym. Encyrtidae) in relation to temperature. Entomophaga 34 (2): 111-120.

Anexos
Anexo 1. Condiciones de la sala
MARZO
Fecha Tº actual Tº máxima Tº mínima HR% 24 22,6 31,6 14,9 29 21,9 30,0 14,6 30 22,0 22,8 21,6 31 21,0 22,3 20,3
ABRIL
Fecha Tº actual Tº máxima Tº mínima HR% 3 22,3 23,5 21,0 4 21,0 22,6 20,6 5 21,5 23,7 21,0 6 20,8 25,2 17,8 7 22,3 23,1 20,7 10 22,1 26,7 20,6 11 22,2 22,5 21,0 12 21,3 22,5 20,8 68 13 21,3 23,2 20,8 66 17 20,6 24,6 19,6 60 18 21,0 24,6 19,6 63 19 21,1 22,5 20,6 60 20 21,1 24,1 18,3 68 21 21,9 24,3 20,8 68 24 21,9 27,8 20,8 69 25 21,1 22,6 20,6 63 26 21,9 25,3 21,0 63 27 21,3 24,0 21,0 66 28 21,6 24,0 20,0 66
Nota: Los datos de humedad relativa (HR) del mes de marzo y parte del mes de abril no se encuentran disponibles porque todavía no se contaba con un higrometro.
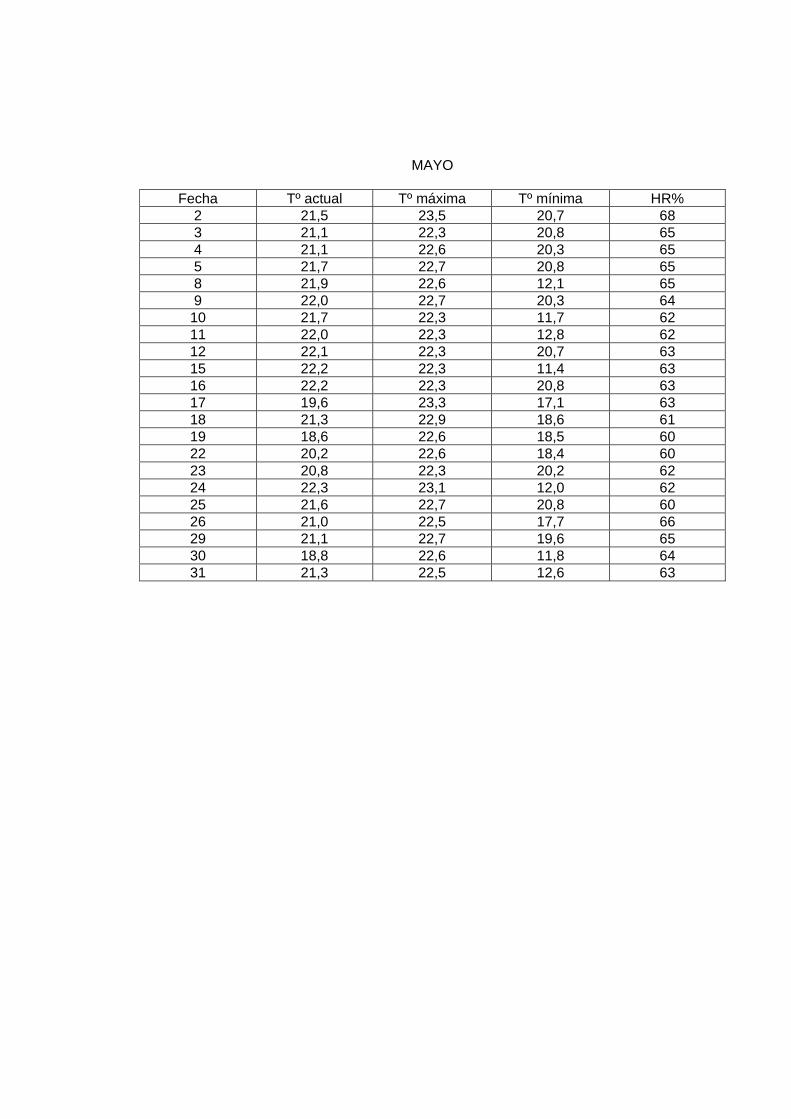
MAYO
Fecha Tº actual Tº máxima Tº mínima HR% 2 21,5 23,5 20,7 68 3 21,1 22,3 20,8 65 4 21,1 22,6 20,3 65 5 21,7 22,7 20,8 65 8 21,9 22,6 12,1 65 9 22,0 22,7 20,3 64 10 21,7 22,3 11,7 62 11 22,0 22,3 12,8 62 12 22,1 22,3 20,7 63 15 22,2 22,3 11,4 63 16 22,2 22,3 20,8 63 17 19,6 23,3 17,1 63 18 21,3 22,9 18,6 61 19 18,6 22,6 18,5 60 22 20,2 22,6 18,4 60 23 20,8 22,3 20,2 62 24 22,3 23,1 12,0 62 25 21,6 22,7 20,8 60 26 21,0 22,5 17,7 66 29 21,1 22,7 19,6 65 30 18,8 22,6 11,8 64 31 21,3 22,5 12,6 63
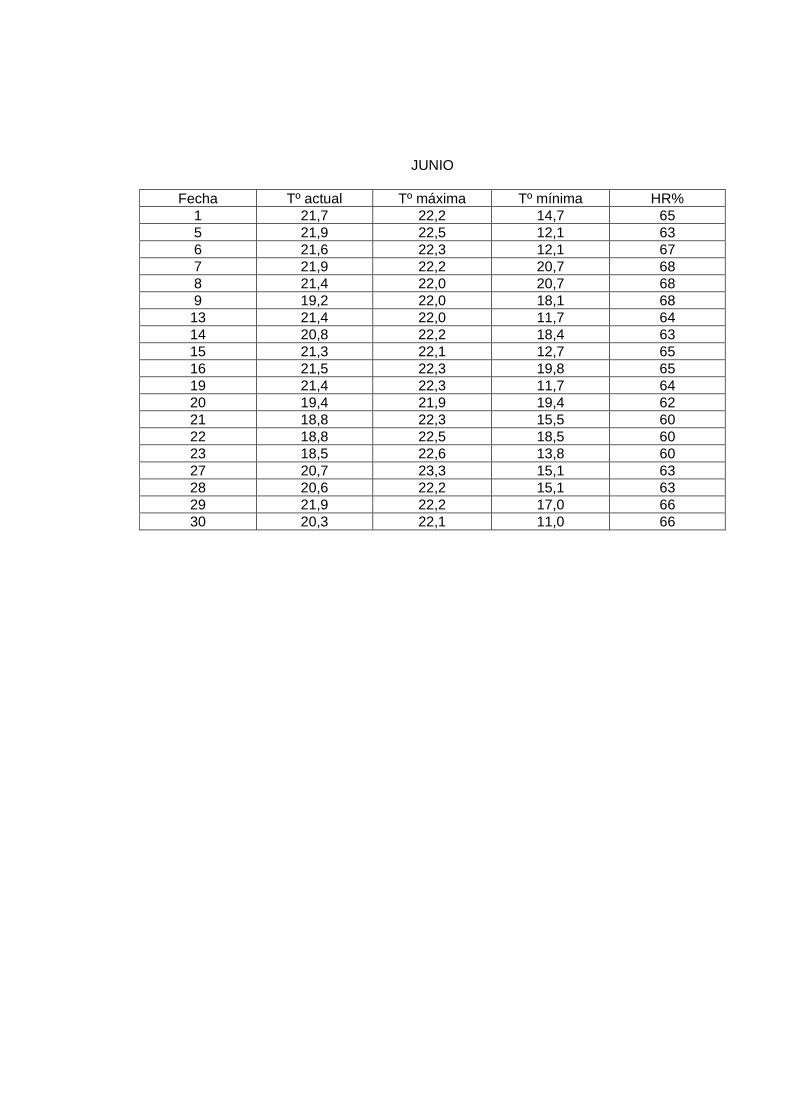
JUNIO
Fecha Tº actual Tº máxima Tº mínima HR% 1 21,7 22,2 14,7 65 5 21,9 22,5 12,1 63 6 21,6 22,3 12,1 67 7 21,9 22,2 20,7 68 8 21,4 22,0 20,7 68 9 19,2 22,0 18,1 68 13 21,4 22,0 11,7 64 14 20,8 22,2 18,4 63 15 21,3 22,1 12,7 65 16 21,5 22,3 19,8 65 19 21,4 22,3 11,7 64 20 19,4 21,9 19,4 62 21 18,8 22,3 15,5 60 22 18,8 22,5 18,5 60 23 18,5 22,6 13,8 60 27 20,7 23,3 15,1 63 28 20,6 22,2 15,1 63 29 21,9 22,2 17,0 66 30 20,3 22,1 11,0 66
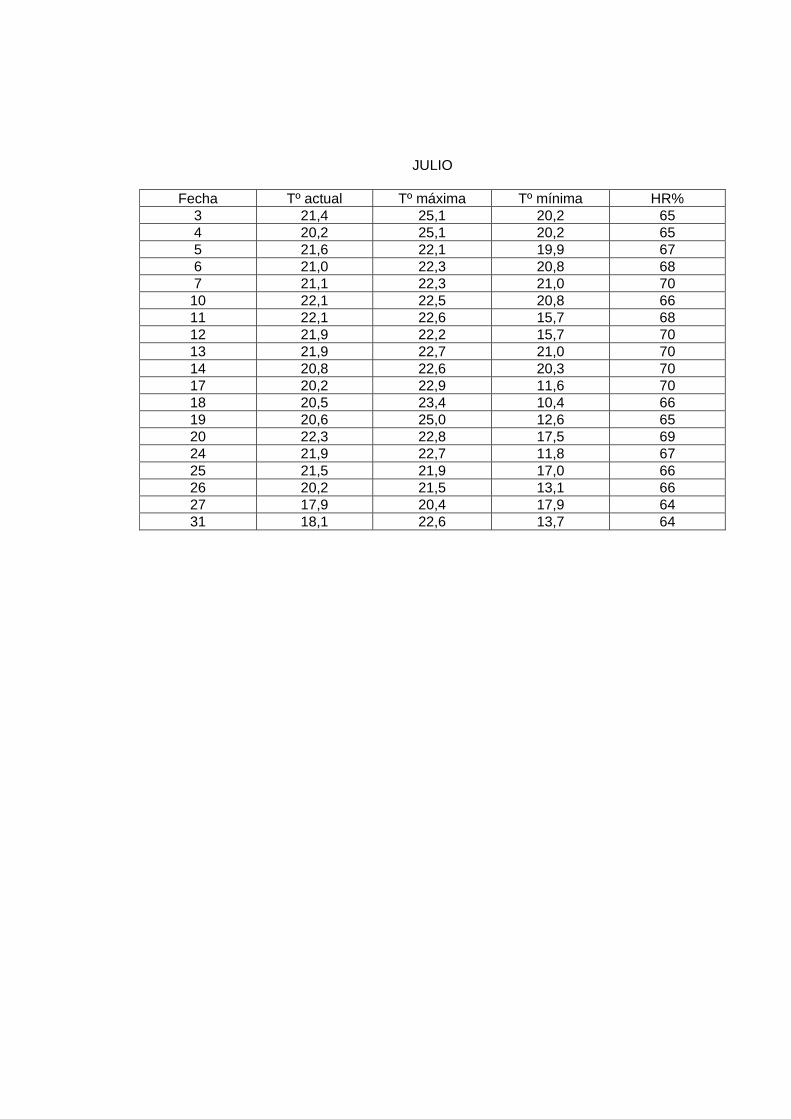
JULIO
Fecha Tº actual Tº máxima Tº mínima HR% 3 21,4 25,1 20,2 65 4 20,2 25,1 20,2 65 5 21,6 22,1 19,9 67 6 21,0 22,3 20,8 68 7 21,1 22,3 21,0 70 10 22,1 22,5 20,8 66 11 22,1 22,6 15,7 68 12 21,9 22,2 15,7 70 13 21,9 22,7 21,0 70 14 20,8 22,6 20,3 70 17 20,2 22,9 11,6 70 18 20,5 23,4 10,4 66 19 20,6 25,0 12,6 65 20 22,3 22,8 17,5 69 24 21,9 22,7 11,8 67 25 21,5 21,9 17,0 66 26 20,2 21,5 13,1 66 27 17,9 20,4 17,9 64 31 18,1 22,6 13,7 64
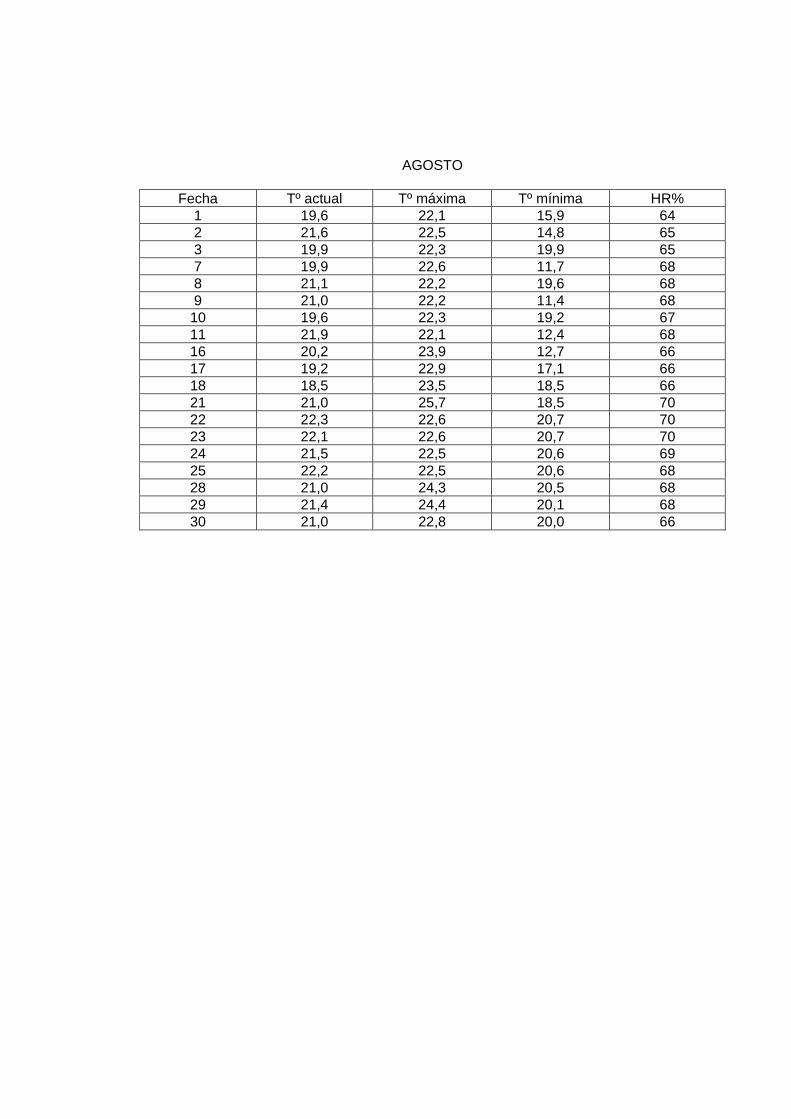
AGOSTO
Fecha Tº actual Tº máxima Tº mínima HR% 1 19,6 22,1 15,9 64 2 21,6 22,5 14,8 65 3 19,9 22,3 19,9 65 7 19,9 22,6 11,7 68 8 21,1 22,2 19,6 68 9 21,0 22,2 11,4 68 10 19,6 22,3 19,2 67 11 21,9 22,1 12,4 68 16 20,2 23,9 12,7 66 17 19,2 22,9 17,1 66 18 18,5 23,5 18,5 66 21 21,0 25,7 18,5 70 22 22,3 22,6 20,7 70 23 22,1 22,6 20,7 70 24 21,5 22,5 20,6 69 25 22,2 22,5 20,6 68 28 21,0 24,3 20,5 68 29 21,4 24,4 20,1 68 30 21,0 22,8 20,0 66
SEPTIEMBRE
Fecha Tº actual Tº máxima Tº mínima HR% 1 20,3 22,5 20,0 66 4 20,0 23,9 19,4 65 5 20,2 24,6 19,4 65 6 21,4 23,8 20,2 66 7 20,5 22,1 20,5 65 8 20,6 22,7 20,0 65 11 21,9 25,6 20,5 67 12 21,7 22,5 20,8 68 13 21,4 24,1 20,8 68 14 21,6 23,2 20,8 65 15 20,7 23,4 19,6 62 20 22,2 25,2 20,3 65 21 21,9 25,3 21,3 65 22 21,5 22,5 21,0 68 25 21,9 29,9 21,0 68 26 21,4 24,1 21,4 69 27 21,5 25,3 19,7 65 28 22,1 25,5 21,0 65 29 22,3 24,9 21,3 67
OCTUBRE
Fecha Tº actual Tº máxima Tº mínima HR% 2 22,7 24,3 21,1 68 3 21,9 24,1 21,1 68 4 21,3 22,8 14,0 66 5 22,2 22,8 14,0 64 6 22,3 26,3 14,0 62 10 21,7 29,4 20,6 64 11 22,5 24,9 21,3 65 12 21,3 26,2 21,3 68 13 21,9 23,3 21,3 68 16 22,6 27,0 20,5 64 17 21,7 22,7 21,3 68 18 22,2 22,7 21,3 68 19 21,4 22,8 19,2 68 20 22,0 25,5 19,2 68 23 21,4 27,9 19,2 69 24 22,0 25,7 19,4 69 25 22,0 26,1 20,2 68 26 22,6 28,4 18,3 68 27 21,4 22,8 21,4 68 30 22,3 26,7 21,1 64 31 21,9 25,9 21,5 64
NOVIEMBRE
Fecha Tº actual Tº máxima Tº mínima HR%
2 21,4 26,5 12,6 64 3 21,6 24,9 18,5 65 6 21,6 26,8 19,1 65 7 21,6 29,4 14,1 65 8 22,2 26,1 14,4 62 9 21,6 24,6 13,5 63 10 22,3 27,1 12,7 64 13 21,5 25,9 12,6 66 20 21,7 29,9 14,2 65 21 22,5 23,5 19,1 65 22 22,1 29,6 12,1 66 23 21,9 26,9 20,8 67 24 21,7 25,7 21,4 67 27 22,0 29,1 17,7 65 28 21,7 26,1 18,3 65 29 22,0 29,9 21,5 64
DICIEMBRE
Fecha Tº actual Tº máxima Tº mínima HR% 1 22,3 30,7 21,5 64 4 21,6 28,6 18,3 64 5 21,7 27,3 19,0 64 6 22,9 29,1 21,6 63 7 22,0 30,7 21,5 62 11 21.6 27.7 18.1 64 12 21.9 28.8 21.5 65 13 21.6 30.9 21.5 65 14 21.6 29.2 21.5 66 15 21.6 29.6 21.5 66 18 22.3 29,0 13.7 65 20 22.3 31.5 13.1 64 21 22,0 28.3 14.9 62 22 22,0 29.9 18,0 62 26 22.2 30,0 12.9 62 27 22.5 31,0 16.3 60 28 22.5 31.6 21.6 60 29 22.8 31.4 21.5 60
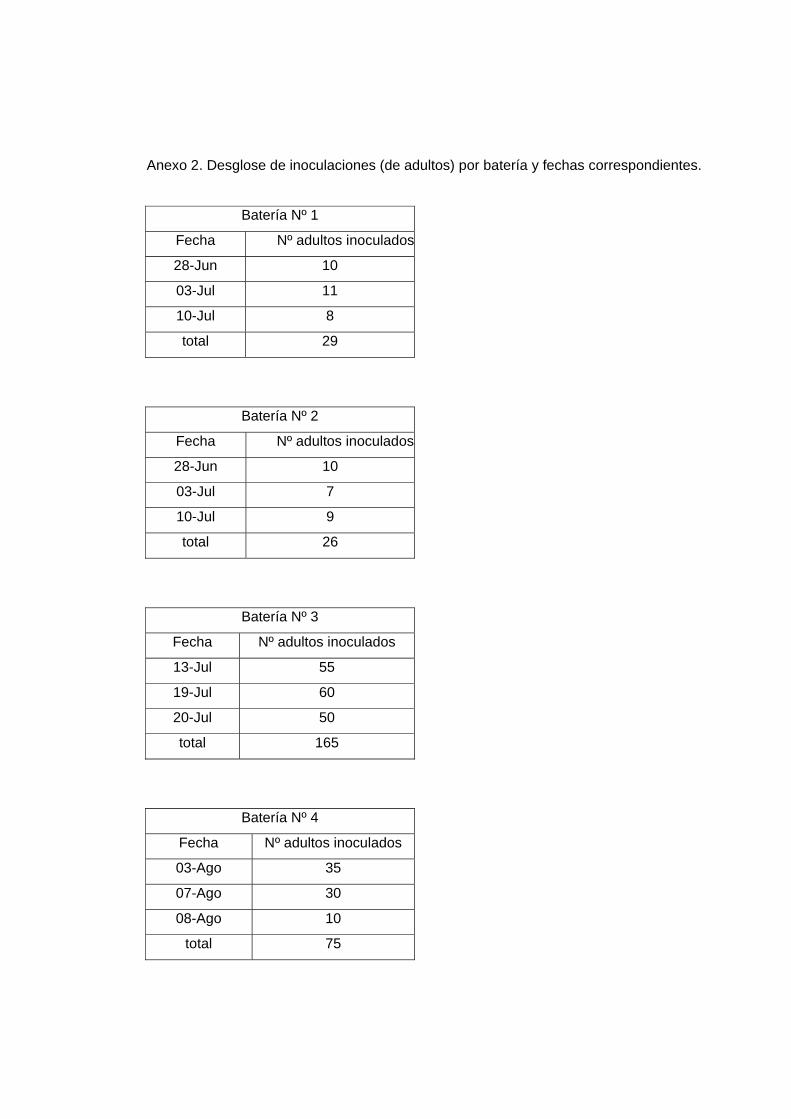
Anexo 2. Desglose de inoculaciones (de adultos) por batería y fechas correspondientes.
Batería Nº 1
Fecha Nº adultos inoculados
28-Jun 10
03-Jul 11
10-Jul 8
total 29
Batería Nº 2
Fecha Nº adultos inoculados
28-Jun 10
03-Jul 7
10-Jul 9
total 26
Batería Nº 3
Fecha Nº adultos inoculados
13-Jul 55
19-Jul 60
20-Jul 50
total 165
Batería Nº 4
Fecha Nº adultos inoculados
03-Ago 35
07-Ago 30
08-Ago 10
total 75
Batería Nº 5
Fecha Nº adultos inoculados
08 Sept 16
11 Sept 20
13 Sept 21
total 57
Batería Nº 6
Fecha Nº adultos inoculados
15 Sept 25
20 Sept 79
21 Sept 36
25 Sept 20
27 Sept 14
28 Sept 8
total 182
Batería Nº 7
Fecha Nº adultos inoculados
19-Oct 100
total 100
Batería Nº 8
Fecha Nº adultos inoculados
19-Oct 34
20-Oct 80
23-Oct 16
total 130
Batería Nº 9
Fecha Nº adultos inoculados
16-Oct 118
23-Oct 62
24-Oct 20
total 200






























