Embed Size (px)
Citation preview

“RESPUESTA FISIOLÓGICA DEL FRIJOL CAUPÍ (Vigna unguiculata (L.) Walp) A LA COINOCULACIÓN DE BACTERIAS DIAZOTRÓFICAS DE LOS
GÉNEROS Azotobacter Y Rhizobium EN SUELOS DEL DEPARTAMENTO DEL CESAR”
DOLLY MELISSA OBANDO CASTELLANOS
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE AGRONOMÍA
MAESTRÍA EN CIENCIAS AGRARIAS-ÉNFASIS FISIOLOGÍA DE CULTIVOS BOGOTÁ
2012

2
“RESPUESTA FISIOLÓGICA DEL FRIJOL CAUPÍ (Vigna unguiculata (L.) Walp) A LA COINOCULACIÓN DE BACTERIAS DIAZOTRÓFICAS DE LOS
GÉNEROS Azotobacter Y Rhizobium EN SUELOS DEL DEPARTAMENTO DEL CESAR”
DOLLY MELISSA OBANDO CASTELLANOS
Tesis presentada como requisito parcial para optar al título de:
Magister en Ciencias Agrarias: Línea de Investigación en fisiología de cultivos
Director (a):
Ph.D. RUTH REBECA BONILLA BUITRAGO
Codirector (a):
Ph.D. VÍCTOR JULIO FLOREZ RONCANCIO
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE AGRONOMÍA
MAESTRÍA EN CIENCIAS AGRARIAS-ÉNFASIS FISIOLOGÍA DE CULTIVOS BOGOTÁ
2012

3
A Dios por darme todas las bendiciones y por
ayudarme a cumplir mis sueños…
A mi mami y a mi hermanita por ser la razón
de mi vida, quiero dedicarles este logro por todos
estos años que no hemos estado juntas, porque este
lindo paso acorte el tiempo para que se cumpla
nuestro sueño de vivir juntas otra vez….

4
Agradecimientos
A mi Madre por ser lo mas lindo de mi vida, por enseñarme todos los días a ser mejor ser
humano y por ser mi amiga incondicional que nunca me falla..
A mi Padre por ser mi ejemplo de lucha y perseverancia, por dejarme una profunda lección
de vida y por demostrarme que todo se puede lograr…
A Juan María Torres por ser mi segundo padre, con su ejemplo y su gran ayuda me
conduce cada día por los caminos del saber…
A Mi Diego por ser mi amor bonito, por ser mi mano derecha en todo momento y por ser mi
alegría cuando más lo necesito.
A la Dra. Ruth Bonilla por su Dirección, su apoyo y por haberme ayudado
incondicionalmente para que pudiera alcanzar este paso.
Al profesor Víctor Flórez por creer en mí y apoyarme cuando más lo necesité, por darme la
tranquilidad para superar las dificultades del proyecto.
A Dani, por su asesoría científica y por haberse preocupado desinteresadamente porque
todo resultara siempre bien.
Al Doctor Belisario Roncallo, Dubys, Dolly, Edelberto y a todas las personas de la E.E
Motilonia por haber contribuido enormemente en la segunda fase de del proyecto y porque
desde el pregrado siempre encontré el apoyo y la disposición de todos ustedes……
Gracias!!!!!
A mi “Patiñito” , Mabelita y Gus por alegrarme en momentos difíciles y por compartir su
amistad conmigo.
A Inesita, Lady y Mao por su asesoría técnica y por su entrega incondicional. Sin ustedes
no hubiera sido posible avanzar. ¡Los quiero mucho!

5
Contenido
.............................................................................. Pág.
Lista de figuras ................................................................................................................................... 8
Lista de tablas .................................................................................................................................... 9
Lista de símbolos y abreviaturas .................................................................................................... 10
Introducción ............................................................................................................................ ….....11
Objetivos ........................................................................................................................................ 10
Objetivo General ...................................................................................................... 14
Objetivos específicos ............................................................................................... 14
Capítulo I. Marco teórico .............................................................................................................. 15
1.1 El Nitrógeno ....................................................................................................... 15
1.1.1 Ciclo del Nitrógeno .......................................................................................... 15
1.2 Fijación biológica de nitrógeno (FBN) ................................................................. 16
1.2.1 Microorganismos diazotróficos asimbióticos .................................................... 17
1.2.1.1 Género Azotobacter ..................................................................................... 18
1.2.2 Microorganismos diazotróficos simbióticos ...................................................... 19
1.2.2.1 Genero Rhizobium ........................................................................................ 19
1.3 Importancia de la fijación biológica de nitrógeno (FBN) en leguminosas ............ 20
1.3.1 Fríjol caupí (Vigna unguiculata (L.) Walp) ........................................................ 21
1.3.2 Simbiosis Rizobio - leguminosas ..................................................................... 22
1.4 Coinoculaciones ................................................................................................. 23
Capítulo II. Caracterización y potencial de Azotobacter sp. en leguminosas forrajeras. . 25
Resumen .................................................................................................................. 25
2.1 Introducción ......................................................................................................... 26

6
2.2 Materiales y Métodos .......................................................................................... 27
2.2.1 Material vegetal y localización .......................................................................... 27
2.2.2 Selección y caracterización de Azotobacter sp. .................................................... 28
2.2.2.1 Toma de Muestra y Aislamiento………………………. ................................... 28
2.2.2.2 Caracterización bioquímica y promoción de crecimiento vegetal .................. 28
2.2.3 Ensayo en invernadero .................................................................................... 29
2.2.4 Identificación Molecular ................................................................................... 30
2.2.5 Análisis Estadístico .......................................................................................... 30
2.3 Resultados ......................................................................................................... 31
2.3.1 Selección y descripción de las capacidades de promoción de crecimiento
de las cepas ............................................................................................................. 31
2.3.2 Evaluación de las capacidades de promoción de crecimiento vegetal en
condiciones de invernadero ...................................................................................... 32
2.3.3 Identificación mediante determinación y secuenciación del grupo específico
nifH..... ...................................................................................................................... 34
2.4 Discusión ............................................................................................................ 34
2.4.1 Selección y descripción de las capacidades de promoción de crecimiento
de las cepas ............................................................................................................. 34
2.4.2 Evaluación de las capacidades de promoción de crecimiento vegetal en
condiciones de invernadero ...................................................................................... 36
2.4.3 Identificación mediante determinación y secuenciación del grupo específico
nifH ..... ..................................................................................................................... 38
Conclusiones ............................................................................................................ 38
Capítulo III Influencia de la coinoculación Rhizobium - Azotobacter sobre la
nodulación, crecimiento, y calidad nutricional del fríjol caupí (Vigna unguiculata (L.)
Walp) en condiciones de invernadero. ...................................................................................... 40
Resumen .................................................................................................................. 40
3.1 Introducción ......................................................................................................... 41
3.2 Materiales y Métodos .......................................................................................... 43

7
3.2.1 Identificación molecular de las cepas G56 y G58 mediante la amplificación del
16S rDNA .................................................................................................................. 43
3.2.2 Ensayo en invernadero. ..................................................................................... 45
3.2.2.1 Condiciones del experimento…………………………. ................................... 45
3.2.2.2 Parámetros Evaluados ................................................................................. 45
3.2.3 Diseño experimental y Análisis Estadístico ...................................................... 46
3.2.4 Microscopía electrónica de Barrido (MEB) ....................................................... 46
3.3 Resultados ......................................................................................................... 47
3.3.1 Identificación molecular de las cepas G56 y G58 mediante la amplificación
del 16S rDNA ........................................................................................................... 47
3.3.2 Evaluación en invernadero .............................................................................. 48
3.3.3 Determinación de colonización rizoférica mediante Microscopía electrónica
de Barrido (MEB)..... ................................................................................................. 52
3.4 Discusión ............................................................................................................ 53
3.4.1. Caracterización molecular de las cepas G56 y G58 ....................................... 53
3.4.2 Evaluación en invernadero .............................................................................. 54
3.4.3 Determinación de la colonización rizoférica del mejor coinóculo mediante
Microscopía electrónica de Barrido (MEB) ..... .......................................................... 57
Conclusiones ............................................................................................................ 58
Conclusiones generales de la investigación ............................................................. 59
Recomendaciones .......................................................................................................................... 60
Bibliografía ....................................................................................................................................... 61

8
Lista de figuras
Figura 1. Análisis de componentes principales de los tratamientos con las cepas (A)
Diagrama de dispersión del análisis del experimento en invernadero con las componentes
PC1 y PC2; (B) Diagrama de dispersión del análisis del experimento en invernadero con
las componentes PC1 y PC3; y (C) Diagrama de dispersión del análisis del experimento en
invernadero con las componentes PC2 y PC3. Tratamientos y relación de símbolos: testigo
absoluto ( ), testigo biológico (۞), cepa GV22 (Δ), cepa GV2 (),cepa GV6 (), cepa GV33
(), cepa GV23 (♦), cepa GV11 (◊),cepa GV1 (), cepa GV12 (+),cepa GV39 (), cepa
GV5 (▼), cepa GV28 (∇), cepa GV34 (✡), cepa GV17 (x), cepa GV 18 (★), cepa GV43
(✩). Cada símbolo es el promedio de tres mediciones ................................................................ 33
Figura 2. Determinación del grupo específico nifH para las cepas GV18, GV23 y GV34. ........ 34
Figura 3. Amplificación del 16S rDNA de las cepas G56 y G58 mediante electroforesis en
gel de agarosa al 2%. MP: Marcador de peso molecular 1000 kb plus....................................... 47
Figura 4. Árbol filogenético realizado mediante la amplificación del 16S rDNA. La filogenia
fue inferida por el método de Neighbor-Joining (Saitou y Nei, 1987) utilizando el software
Mega 5 (Tamura et al., 2011). Las distancias evolutivas se calcularon utilizando el
parámetro de Kimura 2-método (Kimura, 1980). Los números en los nodos representan el
porcentaje de bootstraping con 10000 repeticiones. La barra de escala = 0,01 sustituciones
por sitio... .......................................................................................................................................... 48
Figura 5. ± Desviación estándar - Letras diferentes representan diferencias
estadísticamente significativas en relación a cada variable evaluada de acuerdo al test de
Tukey HSD (p < 0.05). Cada gráficas representa variables de peso y longitud en función del
tiempo (15, 30 y 45 d.d.s). 5A: Longitud Parte aérea; 5B: Longitud de raíz; 5C: Peso seco
aérea; 5D: Peso seco raíz... ........................................................................................................... 49
Figura 6. Análisis de componentes principales de los tratamientos con las
inoculaciones de Azotobacter sp. y Rhizobium sp. (A) Diagrama de dispersión del
análisis del experimento en invernadero con las componentes PC1 y PC2; (B)
Diagrama de dispersión del análisis del experimento en invernadero con las
componentes PC1 y PC3; y (C) Diagrama de dispersión del análisis del experimento
en invernadero con las componentes PC2 y PC3. Tratamientos y relación de
símbolos: T1 (), T2 (Δ), T3 (), T4 (),T5 (),T6 (), T7 (x), T8 (∇), T9 (▼), Cada
símbolo es el promedio de tres mediciones....…………………..... ……………………51

9
Lista de tablas
Tabla 1 Descripción de las capacidades de promoción de crecimiento vegetal in vitro de
las cepas seleccionadas de Azotobacter sp……………………………. .......…...............31
Tabla 2. Descripción del porcentaje de variación acumulada del experimento en el
análisis de componentes principales y la contribución de la carga factorial para cada
parámetro evaluado..……………………………………………………….......…...............44
Tabla 3. Iniciadores utilizados para la secuenciación del gen 16S rDNA de las cepas G56 y
G58…………………………………………………………………………….......…...............44
Tabla 4. Descripción del porcentaje de variación acumulada del experimento en el
análisis de componentes principales y la contribución de la carga factorial para cada
parámetro evaluado..……………………………………………………….......…...............50

10
Lista de símbolos y abreviaturas
DNA Ácido desoxirribonucleico
RNA Ácido ribonucleico
PCR Reacción en cadena de la polimerasa
NCBI Centro nacional de información biotecnológica
FBN Fijación biológica de nitrógeno
UFC Unidades formadoras de colonia
AIA Ácido indol acético
ARA Actividad de reducción de acetileno
DO Densidad óptica
M Molar
L Litro
μg Microgramo
ml Mililitro
min Minuto
ng Nanogramo
nm Nanomol
rpm Revoluciones por minuto
mm Milimetro
mL Mililítro
cm Centímetro

11
Introducción
El sector agropecuario ha sido uno de los principales motores del desarrollo
económico del país (Proexport, 2011), con una superficie total agropecuaria estimada
en 50'900.000 hectáreas, ocupando el renglón pecuario 39'100.000 hectáreas en su
mayoría dedicadas a la ganadería, contribuyendo con 51% del PIB pecuario
(Fedegan, 2010). En el contexto nacional, la región Caribe reviste un papel importante
en el sector, con un inventario de 6,9 millones de cabezas, que generan el 38,7% de
la leche y el 38% de la carne que produce el país, cifras que demuestran la amplia
participación de la región y su notable contribución a la economía nacional (Fedegan -
SENA, 2010).
No obstante, la explotación ganadera ha sido manejada en sistemas de producción
extensivos con pastoreo de bovinos en monocultivos de gramíneas, lo cual afecta de forma
negativa la respuesta productiva de los animales y la calidad de las praderas, por la
reducción de la capacidad de rebrote y el vigor de crecimiento de las especies forrajeras,
en razón del agotamiento de las reservas de nutrientes. En consecuencia, se favorece la
invasión de las praderas por malezas, la compactación del suelo y exposición del suelo a la
erosión (Corpoica, 2004). Generalmente no se cuenta con planes de fertilización en su
etapa productiva y, en algunos casos, sólo se aplican fuentes de nitrógeno, siendo el
factor determinante que potencializa los rendimientos de la planta y su calidad
nutritiva (Cuesta-Muñoz, 2005).
Por el bajo costo y la abundancia de las especies forrajeras, es cada día más
relevante su uso en los sistemas ganaderos de esta región (Cuesta y Mila, 2003). Sin
embargo, debido a las limitaciones edafoclimáticas que caracterizan la microrregión del
Valle del Cesar la producción de forraje es escasa e irregular durante el año, con
reducciones en la producción y calidad de forraje durante la época seca comprometiendo
la producción de carne debido a la pérdida de peso de los animales.
El fríjol caupí (Vigna unguiculata (L.) Walp) es una de las principales leguminosas
forrajeras con potencial cultivable en la Región Caribe (DNP, 2010). Este cultivo se
constituye como una excelente provisión de forraje para los animales en temporadas
donde normalmente no existe ninguna otra fuente de alimento; a su vez, es una
fuente complementaria de la dieta, debido a que puede crecer sola o en asociación con
otras gramíneas ó leguminosas (Calegari, 1995).

12
Esta leguminosa fija el nitrógeno atmosférico en simbiosis mutualista con Rhizobium sp.
mediante la Fijación Biológica del Nitrógeno (FBN). El cultivo se nutre del nitrógeno
aportado por la FBN y del disponible en el suelo. La FBN puede aportar entre 25 y 90% del
nitrógeno necesario para el desarrollo del cultivo, pero esto sólo puede concretarse cuando
los factores ambientales no son limitantes (González et al., 1997; Perticari et al., 2003;
Perticari, 2005). Las condiciones edafoclimáticas que caracterizan la Región de Valle del
Cesar enfrentan a esta leguminosa a diversos tipos de estrés, entre los que se destaca la
sequía, factor que determina la capacidad simbiótica de los rizobios. Se sabe que la
nodulación y la fijación del nitrógeno son procesos muy sensibles a factores
medioambientales (Ravindar y Chandra, 2008).
De esta forma, las estrategias que favorezcan el establecimiento de esta leguminosa
forrajera con capacidad de nodular vigorosamente y de fijar el nitrógeno eficientemente,
combinado con estrategias agronómicas, permitirán obtener altos rendimientos y contribuir
con las reservas nitrogenadas del suelo (Perticari et al., 2003).
Debido a sus implicaciones agronómicas, las interacciones ecológicas entre los rizobios y
otras bacterias del suelo han sido de especial interés en los últimos años. Debido a una
interacción positiva entre las especies de Rhizobium y bacterias diazotróficas del suelo,
como Azotobacter y Azospririllum, del Gallo y Fabbri (1991), Dardanelli et al. (2008) y
Benintende et al. (2010), entre otros autores, han reportado aumento de la nodulación y
crecimiento óptimo de una amplia variedad de forrajes.
Azotobacter sp. (especialmente A. chroococcum) ha sido ampliamente reportado por
promover el crecimiento vegetal en condiciones edáficas de estrés por sequía. Especies de
este género se encuentran ampliamente distribuidos en suelos destinados a la agricultura,
predominante en regiones tropicales, y se pueden encontrar fácilmente en asociación con
cereales y en la rizosfera de leguminosas (Kole et al., 1988; Martínez-Toledo et al., 1991;
Gharib et al., 2009).
Por su capacidad para formar estructuras de resistencia en condiciones de sequía y de
mejorar significativamente el desarrollo vegetal, en Colombia Azotobacter sp. ha sido
ampliamente estudiada, entre otros cultivos, en algodón, sorgo, maíz, fríjol, tomate, ají
y lechuga, disminuyendo costos y aplicación de fertilizantes de síntesis química hasta
en un 50% (Bonilla et al., 2000; 2002; Bonilla y Morales 2007). Adicionalmente,
Bonilla y Morales (2007) evaluaron la respuesta a la inoculación de este
microorganismo en especies leguminosas arbóreas, donde se obtuvo una

13
disminución del tiempo de germinación y aumento de altura y producción de masa
seca.
Los efectos positivos de la inoculación combinada con Rhizobium - Azotobacter y otras
rizobacterias estimuladoras del crecimiento han demostrado que aumentan
significativamente el número y peso de nódulos, mayor fijación de nitrógeno, aumento del
contenido de macronutrientes y micronutrientes, en comparación con la inoculación
individual de Rhizobium sp. (Yahalom et al., 1987; Rodelas et al., 2000). El agente
potenciador de estos beneficios ha sido correlacionado con la influencia positiva de la
bacterias promotoras de crecimiento no simbióticas, ya que se verifican aumentos
significativos en factores de rendimiento como la acumulación de masa seca y el contenido
de nitrógeno en grano (Burns et al., 1981; Sarig et al., 1986; Rodelas et al., 2000).
Por lo anterior, la evaluación de las coinoculaciones con bacterias nativas de
naturaleza simbiótica y asimbiótica a partir de leguminosas forrajeras permitirá
determinar su potencial de promoción ante el efecto de la marcada estacionalidad en
las precipitaciones de la zona; permitiendo de esta forma la utilización de recursos
biológicos poco explorados y contribuyendo con tecnologías limpias que favorezcan y
mejoren las condiciones de especies forrajeras de interés en sistemas ganaderos de
la región.
El objetivo de la presente investigación fue contribuir con el mejoramiento de la
calidad del fríjol Caupí (Vigna unguiculata (L.) Walp) mediante la coinoculación de
bacterias diazotróficas nativas de los géneros Rhizobium y Azotobacter en el
departamento del Cesar.

14
Objetivos
Objetivo General
Contribuir con el mejoramiento de la calidad del fríjol caupí (Vigna unguiculata (L.)
Walp) mediante la coinoculación de bacterias diazotróficas de los géneros
Azotobacter y Rhizobium en el departamento del Cesar.
Objetivos específicos
1. Caracterizar y seleccionar microorganismos diazótrofos simbióticos y asimbióticos
nativos asociados al fríjol caupí (Vigna unguiculata (L.) Walp) en suelos del
departamento del Cesar.
2. Caracterizar y cuantificar la capacidad de promoción de crecimiento vegetal in vitro de los
microorganismos diazótrofos seleccionados.
3. Evaluar la respuesta del fríjol caupí (Vigna unguiculata (L.) Walp) en condiciones de
invernadero a la coinoculación de bacterias diazotróficas de los géneros Azotobacter y
Rhizobium eficientes in vitro.

15
Capítulo I. Marco Teórico
1.1 Nitrógeno
El nitrógeno molecular es el principal constituyente de la atmósfera. La concentración
de nitrógeno es resultado del balance entre la fijación del nitrógeno atmosférico por
acción bacteriana, eléctrica y química, y su liberación se realiza a través de la
descomposición de materias orgánicas por bacterias o por combustión. Es un
constituyente esencial de moléculas fundamentales de todos los seres vivos:
aminoácidos, proteínas, ácidos nucleicos, vitaminas, entre otros. En estado
combinado, el nitrógeno se presenta en diversas formas. Para que el nitrógeno pueda
ser asimilado, es necesario que sea reducido, los únicos seres vivos capaces de
realizar esta reacción son las Eubacteria y Archaea, por el proceso denominado
fijación biológica de nitrógeno (FBN) (Baca et al., 2000).
1.1.1 Ciclo del nitrógeno
La atmósfera es el principal reservorio de nitrógeno donde constituye hasta un 78%
de los gases. Sin embargo, como la mayoría de los seres vivos no pueden utilizar el
nitrógeno atmosférico para elaborar aminoácidos y otros compuestos nitrogenados,
dependen del nitrógeno presente en los minerales del suelo. Por lo tanto, a pesar de
la gran cantidad de nitrógeno en la atmósfera, la escasez de nitrógeno en el suelo
constituye un factor limitante para el crecimiento de los vegetales.
El ciclo del nitrógeno consta de las siguientes etapas:
Proteólisis: El nitrógeno no está disponible debido a su forma no asimilable por la
plantas; razón por la que los microorganismos desempeñan un rol determinante como
degradadores del nitrógeno orgánico para que pueda quedar libre y pueda ser
reutilizado. Este proceso se debe a que los microorganismos poseen la capacidad de
sintetizar proteinasas extracelulares que transforman las proteínas en unidades más
simples (péptidos), los cuales son degradados obteniendo como producto final la
liberación de aminoácidos individuales. El nitrógeno de las proteínas (y de los ácidos
nucleicos) se debe considerar como el final de la línea de síntesis de compuestos
nitrogenados (Pelczar et al., 1993; Baca et al., 2000).
Amonificación: Los productos finales de la proteólisis son los aminoácidos. Estos
compuestos suelen ser degradados a compuestos simples por los organismos que

16
viven en el suelo (bacterias y hongos), los cuales utilizan las proteínas y aminoácidos
para formar las proteínas que necesitan y liberar el exceso de nitrógeno como
amoníaco (NH3) o amonio (NH+4). Este proceso se denomina amonificación (Pelczar
et al., 1993; Baca et al., 2000).
Nitrificación: Según Pelczar et al. (1993), la oxidación del amoníaco a nitritos se llama
nitrificación. Una de las actividades de mayor importancia de algunas bacterias
autótrofas son las reacciones de oxidación que proporcionan a estos
microorganismos la energía requerida para procesos celulares. La nitrificación se da
en los siguientes pasos: Un grupo de bacterias (Nitrosomonas sp. y Nitrococcus sp.)
oxidan el amoníaco a nitrito (NO2-).
2 NH3 + 3 O2 2 NO2 - + 2 H+ + 2 H2O
Otro grupo de bacterias (Nitrobacter sp.) transforman el nitrito en nitrato, por este
motivo no se encuentra nitrito en el suelo, que además es tóxico para las plantas.
Desde el punto de vista de la fertilidad del suelo, el producto de estas reacciones
(nitratos) es la forma más accesible para las plantas.
2 NO2 - + O2 2 NO3
–
Desnitrificación: Algunos microorganismos son capaces de transformar nitratos a
nitrógeno gaseoso u óxidos nitrosos. Este proceso se denomina desnitrificación y
conlleva a pérdidas de nitrógeno en el suelo. Algunos de los microorganismos
relacionados en la reacción son Thiobacillus denitrificans (autótrofo), Micrococus
denitrificans (heterótrofo) y algunas especies de heterótrofos más comunes
perteneciente a los géneros Serratia, Pseudomonas, y Achromobacter. La
desnitrificación se disminuye en suelos bastante aireados, que tienen cantidades
moderadas de materia orgánica y nitratos; y sucede abundantemente en suelos
saturados de agua (anaerobios) y ricos en sustancias orgánicas (Pelczar et al., 1993
Baca et al., 2000).
1.2 Fijación biológica de nitrógeno (FBN)
La FBN es el proceso que transforma el nitrógeno atmosférico, que es inerte, en una
forma biológicamente útil siendo la principal forma de entrada en ecosistemas
desérticos (Zahran, 2001). Químicamente, la fijación de nitrógeno es similar al
proceso Haber-Bosch (N2 + 2H2 → 2NH3, 500ºC y 350 atm), método comercial de la
producción de amoniaco para fertilizantes nitrogenados, que consume una gran

17
cantidad de energía. Se estima que se necesitan 1,5 kg de combustible por cada kg
de nitrógeno fijado (Mahecha, 2002).
El proceso de FBN es utilizado en la naturaleza por diferentes géneros bacterianos.
Las plantas se benefician de este proceso cuando las bacterias mueren y liberan el
nitrógeno al suelo o cuando las bacterias viven en estrecha asociación con las plantas
(Willems, 2003). Si bien existe una amplia gama de organismos y asociaciones
vegetales que son capaces de fijar nitrógeno de la atmósfera, la relación simbiótica
entre rizobios y leguminosas es responsable de contribuir con la mayor cantidad de
nitrógeno fijado en especies agrícolas (Peoples et al., 1995; Chianu et al., 2011).
La fijación de nitrógeno en la biosfera se estima en unos 275 millones de toneladas
anuales, de los cuales 30 corresponden a causas naturales, 70 a fijación industrial y
175 al proceso de fijación biológica (Burns y Hardy, 1975), por parte microorganismos
diazotróficos asimbióticos (de vida libre) y simbióticos (mutualistas obligados) (Marín
et al., 1999). Las entradas en los ecosistemas terrestres de FBN a partir de la relación
simbiótica entre leguminosas y sus rizobios es de 70.000.000 toneladas de N2/ha/año
(Brockwell et al., 1995; Baca et al., 2000). Esta cantidad puede aumentarse en la
medida que la población mundial se incremente y las fuentes naturales que suplen los
fertilizantes nitrogenados disminuyan.
1.2.1 Microorganismos diazotróficos asimbióticos
Las bacterias diazótrofas asimbióticas son aquellas que poseen capacidad de fijar
nitrógeno atmosférico sin la necesidad de formar una simbiosis con plantas, debido a
que estas disponen de diferentes mecanismos de protección del complejo
nitrogenasa. Además de fijar el nitrógeno atmosférico, estas bacterias favorecen el
desarrollo del sistema radical de la planta con la cual conviven, a través de la
producción de reguladores de crecimiento u hormonas (Cassan et al., 2001),
favoreciendo una mayor absorción de nutrientes por parte de la planta. Como
resultado de esta asociación se han obtenido incrementos del orden de 5% a 30% en
los rendimientos de caña de azúcar, maíz, arroz, trigo y gramíneas forrajeras. Estas
bacterias se encuentran prácticamente distribuidas en todos los hábitats. Entre los
principales géneros que se encuentran en vida libre o endófitos asociados a la
rizósfera se destacan: Azotobacter, Azotococcus, Azospirillum, Beijerinckia,
Azotomonas, Bacillus, Citrobacter, Clostridium, Chromatium, Chlorobium,

18
Desulfovibrio, Desulfomonas, Gluconacetobacter, Herbaspirillum y Klebsiella
(Rodríguez et al., 2003).
1.2.1.1 Género Azotobacter
Los microorganismos de este género presentan forma bacilar, Gram negativos y en
cultivos viejos como Gram variables, las células son ovoides y miden alrededor de 2 a
4μm de diámetro, siendo las de mayor tamaño las de A. chroococcum que llegan a
medir hasta 6μm. Poseen estructuras de resistencia conocidas como quistes, son
aerobios pero algunos pueden vivir en tensiones bajas de oxígeno y móviles por
flagelos perítricos (Saribay, 2003). En medio libre de nitrógeno con glucosa como
única fuente de carbono, las células jóvenes de diferentes especies muestran una
forma bacilar con extremos redondeados. Las células en cultivos viejos tienden a ser
elipsoidales y en ciertos casos es común observar gránulos sudanofílicos y
metacromáticos (Tejera et al., 2005; Segura y Espín, 1998).
Las colonias jóvenes de estos microorganismos son generalmente lisas, opacas poco
convexas y viscosas. Bioquímicamente son catalasa y oxidasa positivas, reducen el
nitrato, producen sulfuro de hidrógeno e hidrolizan almidón, y producen hormonas
(giberelinas, auxinas y citoquininas); solubilizan fosfatos, realizan procesos de
biodegradación de plaguicidas, tales como Endosulfán (Castillo et al., 2005). Son
fijadores de nitrógeno en vida libre, fijan alrededor de 10 mg de N2 por gramo de
carbohidrato consumido (Holt, 2000). Para fijar nitrógeno requieren molibdeno, que
puede ser parcialmente sustituido por vanadio.
Azotobacter sp. es quimioheterótrofo, utiliza como fuente de carbono y energía una
gran variedad de ácidos orgánicos, azúcares o sus derivados alcohólicos como el
manitol, siendo el sustrato ideal para aislarlos y cultivarlos. Respecto a la fuente de
nitrógeno utilizan nitrato, sales de amonio y aminoácidos; adicionalmente, pueden
incorporar el nitrógeno del ambiente (Garzón et al., 2001). Para realizar la fijación de
nitrógeno necesitan magnesio, potasio, calcio, hierro y fósforo. Su crecimiento es
óptimo a pH de 7,0 a 7,5, temperatura de 30ºC, con susceptibilidad a pH ácido
(Balandreau, 1986), altas concentraciones de NaCl y temperaturas mayores a 35ºC
(Saribay, 2003).

19
1.2.2 Microorganismos diazotróficos simbióticos
Las bacterias diazotróficas simbióticas son aquellas que son capaces de fijar
nitrógeno atmosférico en simbiosis con las raíces de plantas leguminosas. Estas
raíces presentan nódulos en cuyas células se alojan las bacterias fijadoras de
nitrógeno; toman ácidos orgánicos de las plantas y contribuyen con compuestos
nitrogenados como los ureidos y las amidas de reserva. Este tipo de fijación de
nitrógeno es muy eficiente al recibir las plantas los compuestos nitrogenados
directamente de los microorganismos nitro-fijadores que viven en simbiosis (Espín,
2002). Se han descrito 40 especies en nueve géneros de bacterias que forman
nódulos en leguminosas, no obstante, existen bacterias simbióticas que pueden
colonizar otras plantas; por ejemplo, Rhizobium leguminosarum y cepas fotosintéticas
de Bradyrhizobium sp. encontradas en raíces de arroz (Yanni et al., 2001); Rhizobium
ettli en raíces de maíz (Gutierrez y Martinez, 2001).
1.2.2.1 Género Rhizobium
La taxonomía de los rizobios ha cambiado considerablemente en los últimos 20 años; el
género Rhizobium, un miembro de α-Proteobacteria, ahora se divide en varios géneros.
Mediante el estudio de plantas promiscuas hospederas dispersas geográficamente se ha
constatado que son fuente de muchas especies nuevas. Recientemente, una serie de
aislamientos se han registrado en los nódulos de las leguminosas, con capacidad de
fijación de nitrógeno, pero filogenéticamente ubicados fuera de los grupos tradicionales de
rizobios en α-proteobacterias. Entre las nuevas líneas géneros simbiontes que presentan la
fijación de nitrógeno en leguminosas se incluyen Blastobacter, Methylobacterium, Devosia,
Ochrobactrum y Phyllobacterium en α-proteobacterias Burkholderia, Herbaspirrillum,
Ralstonia y Cupriavidus en β--proteobacteria (Willems, 2003; Lindström y Martínez-
Romero, 2007).
De la filogenia del 16S rDNA está claro que Rhizobium sp. y Agrobacterium sp. están
fuertemente relacionadas y sus especies están entrelazadas (Young et al., 2003). Está
planteada la transferencia de todos estos taxones de Rhizobium sp. y unir a A. radiobacter
y A. tumefaciens en R. radiobacter; mientras que A. rhizogenes en R. rhizogenes (Young et
al., 2001). Por otra parte, A. rubi y A. vitis se transfieren a Rhizobium, como especies
distintas y A. larrymoorei como R. larrymoorei (Young, 2004; Young et al., 2006).
Desde la década de los 80, con la introducción de características genéticas (ADN-ADN y
las hibridaciones ADN-rARN, catálogos rARN y secuenciación del rADN) fue descubierta

20
una mayor diversidad entre los rizobios y se hizo evidente sus relaciones con otros grupos
de bacterias. Esto condujo a un aumento gradual en el número de géneros. En paralelo,
también ha aumentado significativamente en el número de especies, con 48 rizobios
reconocidos (Willems, 2003).
Los rizobios son bacilos de 0,5 a 1,0 x 1,2 a 3,0 µm, Gram negativos y móviles por flagelos
perítricos. Las colonias son usualmente blancas o beige, circulares convexas,
semitraslúcidas u opacas, a veces mucoides, usualmente de 2 a 4 mm de diámetro dentro
de 3 a 5 días de incubación en medio YMA (Vincent, 1970). El crecimiento en un medio de
carbohidratos es usualmente acompañado por abundante cantidad de exopolisacáridos
extracelulares. Desarrollan una pronunciada turbidez de 2 a 3 días en caldo aireado o
agitado (Kuykendall et al., 2005). Son aerobios, su temperatura óptima de crecimiento se
encuentra entre 25 y 30 ºC; algunas especies pueden crecer a temperaturas de 40 ºC. El
pH óptimo de crecimiento está entre 6,0 y 7,0; Sin embargo, pueden predominar entre
valores de 4,0 a 10,0. El tiempo de generación de las cepas de Rhizobium sp. está entre
1,5 a 5,0 horas (Kuykendall et al., 2005).
Es quimiorganoheterótrofo, utiliza un amplio rango de carbohidratos y sales de ácidos
orgánicos como única fuente de carbono, sin formación de gas. No son capaces de
metabolizar celulosa, almidón, caseína, quitina y agar; mientras que la peptona es
pobremente utilizada. Las sales de amonio, de nitrato y la mayoría de los aminoácidos
pueden servir como fuente de nitrógeno. Algunas cepas requieren factores de crecimiento
como biotina, pantotenato o ácido nicotínico (Kuykendall et al., 2005).
1.3 Importancia de la fijación biológica de nitrógeno (FBN) en leguminosas
El proceso de FBN es utilizado en la naturaleza por diferentes géneros bacterianos. Las
plantas se benefician de este proceso cuando las bacterias mueren y liberan el nitrógeno al
suelo o cuando las bacterias viven en estrecha asociación con las plantas (Willems, 2006).
Esta asociación simbiótica se presenta en leguminosas con microorganismos denominados
rizobios, que viven en los nódulos de las plantas fijando el nitrógeno en forma de amonio, el
cual es absorbido por las plantas (Herridge et al., 2008; Angelini et al., 2011).
Si bien existe una amplia gama de organismos y asociaciones vegetales que son capaces
de fijar nitrógeno de la atmósfera, la relación simbiótica entre rizobios y leguminosas es
responsable de contribuir con la mayor cantidad de nitrógeno fijado en especies agrícolas
(Peoples et al., 1995; Chianu et al., 2011). Aproximadamente el 80% del nitrógeno fijado
biológicamente en la agricultura proviene de la simbiosis entre las leguminosas y las

21
especies de Rhizobium, Bradyrhizobium, Sinorhizobium, Azorhizobium, Mesorhizobium y
Allorhizobium (Vance, 1998).
El empleo elevado e indiscriminado de fertilizantes nitrogenados representa una carga
medioambiental, que incluye contaminación del aire, lluvia ácida, contaminación del agua
por nitrato, eutrofización y reducción de la biodiversidad (Socolow, 1999). Esto indica que la
fijación biológica del nitrógeno de tipo simbiótico representa una alternativa económica y
ecológicamente viable, que contribuye a mitigar esta problemática de forma sustentable
(Sylvia et al., 2005).
Teniendo en cuenta que aproximadamente 11g de nitrógeno se consumen por persona
cada día (Frink et al., 1999), que las leguminosas aportan hasta el 80% de las necesidades
dietéticas en la mayor parte de los trópicos y subtrópicos (Singh et al., 2003) y que para el
año 2025 se espera que la población mundial haya crecido hasta un 40% (Mannion, 1998),
se necesitaría para entonces un incremento desmesurado en la producción agrícola
mundial con el fin de disminuir la brecha del abastecimiento de proteína para la población
humana (Siddhuraju et al., 1996; Rangel et al., 2004).
1.3.1 Fríjol Caupí (Vigna unguiculata (L.) Walp)
La familia Leguminosae es vital en la base nutricional actual, debido a que alimenta la
mayoría de la población mundial (Lewis et al., 2005; Singh et al., 2003). El fríjol caupí es
una leguminosa originaria de África, cultivado predominantemente en regiones tropicales y
subtropicales (Steele et al., 1985; Singh et al., 2003). Es uno de los cultivos más adaptados,
versátiles y nutritivos con un alto contenido proteico, importante en la dieta alimenticia para
la población de países en vía de desarrollo (Ramakrishnan et al., 2005).
Se adapta a una amplia gama de suelos arenosos y arcillosos, bien drenados, con una
preferencia por suelos ligeros que permiten un buen enraizamiento; crece también en
suelos de textura pesada fuertemente alcalinos. No tolera las inundaciones extendidas o la
salinidad y crece desde el nivel del mar hasta los 1500 msnm, dependiendo de la latitud
(Carvalho et al., 1998; Davis et al., 1991; Ehlers y Hall, 1997).
Esta especie tiene un alto potencial como abono verde, puede ser incorporado en el suelo
de 8 a 10 semanas después de la siembra proporcionando alrededor de 80 kg/ha de
nitrógeno para un cultivo posterior. Así mismo, es un excelente ensilaje para la utilización
en cultivos mixtos de sorgo, fríjol y forraje o mijo (Carvalho et al., 1998).

22
Para mejorar la calidad y el rendimiento del fríjol caupí, la inoculación de rizobios
seleccionados con alta capacidad de nodulación y fijación de nitrógeno es una estrategia
importante (Kremer y Peterson, 1983; Zilli et al., 2004). En comparación con otras especies,
el fríjol caupí presenta un amplio rango de hospederos de especies de rizobios. El cultivo es
tolerante a la baja fertilidad, presentando altas tasas de fijación de nitrógeno (Elowad y Hall,
1987).
1.3.2 Simbiosis rizobio - leguminosa
La simbiosis rizobio – leguminosa se inicia por los exudados de las raíces de plantas que
permiten la interacción con rizobios y la expresión de los genes de nodulación. Los
flavonoides como luteolina, narigenina y genisteína son los principales exudados que
interactúan con proteínas bacterianas “NodD” activando la expresión de los genes (nod)
(D'Haeze y Holsters, 2002). Estos genes están constituidos por una estructura básica que
consiste en oligómeros de N-acetilglucosamida, unidos por un enlace β 1-4, que conducen
a la síntesis de un grupo de moléculas de señalización que inducen la morfogénesis de los
nódulos (Oldroyd et al., 2005).
En la simbiosis, la planta huésped tiene la información genética para la infección simbiótica
y la nodulación. El papel de la bacteria es desencadenar este proceso que centra su
importancia en dos aspectos fundamentales de la relación rizobio - planta: la preinfección o
atracción quimiotáctica de la bacteria por la planta, seguida de la inducción de cambios
estructurales en los pelos radicales, y la infección donde la bacteria entra en el pelo y forma
canales, recubiertos por nuevo material de pared celular (Olivares y Barea, 1995).
El rizobio entra a la planta a través de los pelos radicales, formando una estructura llamada
hilo de infección (Masson-Boivin et al., 2009), que penetra las células corticales de la raíz.
Una vez en la célula, las bacterias se diferencian en bacteroides y comienza el proceso de
fijación de nitrógeno (Giles et al., 2008).
Cuando se da la unión de los rizobios a los pelos radicales, se induce un cambio en la
dirección de crecimiento apical, generándose una deformación y curvatura de los pelos
radicales o “curling” donde quedan atrapadas las bacterias. Se forma un canal de infección
por donde entran los rizobios al citoplasma de las células vegetales por un mecanismo
similar a la endocitosis, dando lugar a la infección y formación del nódulo propiamente
dicho (Carlson et al., 1994).

23
Dentro de los nódulos, los bacteroides están rodeados por una membrana derivada del
huésped llamada membrana peribacteroidal (MPB), que controla el intercambio molecular
entre el bacteroide y las leguminosas (Panter et al., 2000). A estos grupos de bacteroides
rodeados por la MPB se les llama simbiosomas (Brewin, 2003).
La relación entre el nitrógeno absorbido desde el suelo y el proveniente de la simbiosis
varía con la especie de leguminosa, las condiciones ambientales y las características
genéticas del microsimbionte (Shabaev et al., 1996). En primera instancia, mientras se
genera el sistema nodular, la planta utiliza nitrógeno del suelo, pero, si hay amplia
disponibilidad de nitrógeno en el ambiente de la raíz, el número de nódulos formados será
menor (González, 2002). Las plantas absorben el nitrógeno del suelo en forma de nitrato o
amonio, donde el exceso de nitrógeno en el suelo en forma de nitratos tiene un efecto
inhibitorio sobre la simbiosis en todos los pasos: infección, formación del nódulo y la fijación
de nitrógeno (Fernández, 2003). Así, el aporte global por la FBN para las leguminosas es
menor en suelos bien provistos de nitrógeno que en aquellos donde el nutriente es limitante
(González, 2002; Pietrarelli et al., 2008).
La simbiosis rizobio - leguminosa tiene importancia ecológica y agronómica significativa y
constituye una fuente importante de nitrógeno; por lo tanto, desempeña un papel esencial
en la estructura de los ecosistemas y la agricultura sostenible (Lindström et al., 2010).
1.4 Coinoculaciones
Las inoculaciones mixtas de bacterias promotoras de crecimiento vegetal han sido
reportadas desde hace más de 30 años. Sin embargo, en Colombia no existen evidencias
científicas sobre investigaciones en este campo, razón por la cual es de gran relevancia
evaluar herramientas biológicas de fácil acceso, con soportes científicos reproducibles en
nuestro país.
Jauhri et al. (1979) observaron que la respuesta de Vigna radiata, Glycine max y Pisum
sativum con una inoculación doble de Azotobacter sp. y Rhizobium sp. presentaron los
mejores rendimientos en comparación con la inoculación individual de Rhizobium sp.
Rodelas et al. (1999) estudiaron la inoculación mixta de Vicia faba L. con cuatro diferentes
combinaciones de Rhizobium - Azospirillum y Rhizobium - Azotobacter. Los resultados
evidenciaron que las coinoculaciones dieron lugar a cambios en el contenido total,
concentración y distribución de K, P, Ca, Mg, Fe, B, Mn, Zn y Cu, en comparación con las
plantas inoculadas sólo con Rhizobium.

24
Molla et al. (2001) evaluaron el potencial de crecimiento de las raíces y la nodulación en
soya con la aplicación de Azospirillum brasilense y A. lipoferum coinoculados con dos
cepas de Bradyrhizobium japonicum. Observaron estimulación significativa del crecimiento
de la raíz y mejora en la nodulación en los tratamientos con coinoculación con
Bradyrhizobium. El número total de raíces y su longitud, la masa seca de la raíz, el
desarrollo de pelos radicales y el número y peso fresco de nódulos aumentaron
significativamente en los tratamientos en mezclas. Destacándose el tratamiento
Azospirillum brasilense - Bradyrhizobium japonicum con respecto a los resultados
obtenidos con la mezcla con A. lipoferum en la mayoría de los parámetros agronómicos
evaluados.
Dardanelli et al. (2008) evidenciaron el efecto promotor de Azospirillum brasilense - Cd y
Rhizobium sp. en el crecimiento de plantas de fríjol, la nodulación, la producción de
flavonoides y de lipoquitooligosacaridos en un sistema de cultivo en hidroponía. Esta
inoculación mixta además promovió la elongación de la raíz, la ramificación de las mismas
y el aumento de la secreción de genes Nod. Los resultados también demostraron que la
asociación de bacterias asimbióticas con Rhizobium sp. permiten una exudación más
persistente de flavonoides.
Cassán et al. (2009) encontraron que la coinoculación con Az39 (Azospirillum sp.) y E109
(Bradyrhizobium sp.) en el cultivo de soja incrementó el número de nódulos por plantas y el
porcentaje de plantas noduladas comparadas con la simple inoculación con E109.
Gharib et al. (2009) estudiaron durante los veranos de 2007 y 2008 el efecto de la
inoculación mixta en fríjol. Se evaluó una cepa de Rhizobium leguminosarum, dos cepas de
Azotobacter chroococcum (AZ1) y una cepa de Bacillus cereus var megaterium
phosphaticium (BM3). Los resultados indicaron que la inoculación mixta tuvo un
efecto estadísticamente significativo sobre el crecimiento rápido, nodulación, fijación
de nitrógeno y actividad de reducción de acetileno.

25
Capítulo II. Caracterización y potencial de Azotobacter sp. en leguminosas
forrajeras.
Resumen
Las leguminosas forrajeras del trópico representan una alternativa para la
alimentación del ganado en épocas de sequía. En este sentido, los rizobios
desempeñan un papel importante para el proceso de fijación biológica de nitrógeno
(FBN) mediante la simbiosis microrganismo - planta. No obstante, las condiciones
edafoclimáticas limitan los procesos de FBN por diversos factores como el estrés
hídrico. El objetivo de esta investigación fue determinar la presencia y la capacidad de
promoción de crecimiento vegetal de cepas nativas de Azotobacter sp. asociadas al
fríjol caupí (Vigna unguiculata (L.) Walp) como coadyuvante de los procesos
biológicos que desempeña la FBN bajo las condiciones del Caribe seco Colombiano.
Para esto, se realizó el aislamiento y selección de cepas presuntivas de Azotobacter
sp. a partir de suelo rizosférico del fríjol caupí. Se obtuvieron 15 cepas que fueron
caracterizadas por sus mecanismos de promoción de crecimiento vegetal in vitro
(fijación de nitrógeno, producción de compuestos indólicos, solubilización de fosfatos
y producción de sideróforos). Se realizó un ensayo a nivel de invernadero inoculando
las semillas del fríjol caupí y realizando la germinación en turba estéril, se
trasplantaron a bolsas plásticas con una mezcla de arena y vermiculita estéril (2:1)
durante 50 días. Posteriormente se evaluó el contenido de biomasa y longitud en
planta, contenido de clorofila; y proteínas y carbohidratos totales. Las cepas que
favorecieron significativamente el crecimiento y desarrollo vegetal fueron identificadas
molecularmente mediante detección y secuenciación del primer del gen nifH. De
acuerdo al análisis multivariado de componentes principales, los tratamientos
inoculados con las cepas GV23 y GV18 presentaron diferencias significativas (p <
0.05) en comparación con los demás tratamientos en cuanto a la promoción del
crecimiento vegetal del fríjol caupí expresado en el contenido de biomasa, proteínas y
longitud radical, lo cual evidencia que las dos cepas podrían representar una
importante herramienta para complementar y estimular el desempeño de los procesos
de FBN simbiótico bajo condiciones de estrés de sequía en el Caribe seco
Colombiano.
Palabras clave: Fríjol caupí, Promoción de crecimiento vegetal, Azotobacter sp.

26
Abstract
The tropical legumes represent an alternative for feeding livestock in times of drought.
In this sense, the rhizobia play an important role in the process of biological nitrogen
fixation (BNF). However, the soil and climate processes FBN limited by several factors
such as water stress. The objective of this research was to determine the presence
and capacity to promote plant growth of native strains of Azotobacter sp. associated
with cowpea (Vigna unguiculata (L.) Walp) as an adjunct of the biological processes
that BNF plays under dry conditions Colombian Caribbean. For this, we performed the
isolation and selection of presumptive strains of Azotobacter sp. from rhizosphere soil
of cowpea beans. Fifteen strains were obtained which were characterized by their
mechanisms of plant growth promoting in vitro (nitrogen fixation, production of indole
compounds, phosphates and solubilization of siderophore production). A trial was
conducted at the greenhouse by inoculating the seeds of cowpea beans and making
sterile peat germination, were transplanted to plastic bags with a mixture of sterile
sand and vermiculite (2:1) for 50 days. Subsequently, we evaluated the content and
length in plant biomass, chlorophyll content, and total protein and carbohydrates.
Strains significantly favored plant growth and development were identified by
molecular detection and sequencing of nifH gene first. According to multivariate
principal component analysis, the treatments inoculated with strains GV23 and GV18
significant differences (p <0.05) in terms of promoting plant growth of cowpea beans
expressed in the contents of biomass, protein and root length, the which shows that
the two strains may represent an important tool to complement and stimulate the
performance of symbiotic BNF process under drought stress conditions in the
Colombian Caribbean dry.
Keywords: Cowpea Bean, Plant growth promotion, Azotobacter sp.
2.1 Introducción
El fríjol caupí (Vigna unguiculata (L.) Walp) es una leguminosa de ciclo corto que se
utiliza principalmente como abono orgánico y en rotación de cultivos, su forraje es de
elevado valor nutritivo, constituyendo una opción importante en sistemas de
integración agrícolas - ganaderos (Calegari, 1995). El agente potenciador de estos
beneficios es dependiente de la capacidad de fijación biológica de nitrógeno (FBN),
en la cual los rizobios representan la estrategia efectiva al desempeñar procesos
simbióticos que favorecen la nutrición vegetal (Prinyawiwatkul et al., 1996). No

27
obstante, en la microrregión del Valle del Cesar, la producción de forrajes es escasa e
irregular durante el año, debido a limitaciones edafoclimáticas que caracterizan a los
agroecosistemas del trópico bajo. Durante la época seca la producción de forraje se
reduce considerablemente y su calidad sufre un notable deterioro, comprometiendo la
producción de carne debido a la pérdida de peso de los animales (Bonilla et al.,
2000).
Durante las últimas décadas el uso de biofertilizantes mixtos que incluyen bacterias
simbióticas y asimbióticas se ha convertido en una manera de generar procesos de
origen biológico, consumo de poca energía no renovable y disminución en la
contaminación del medioambiente.
Por lo anterior, la evaluación de las respuestas de leguminosas forrajeras a la
inoculación con bacterias asimbióticas de vida libre permitirá determinar su potencial
de promoción ante el efecto de la marcada estacionalidad en las precipitaciones de la
zona, permitiendo la utilización de recursos biológicos poco explorados y
contribuyendo con tecnologías limpias que favorezcan y mejoren las condiciones de
especies forrajeras de interés en sistemas de ganadería de la región (Sarig et al.,
1986; Rodelas et al., 1996). El objetivo de esta investigación fue aislar y evaluar la
capacidad de Azotobacter sp. para promover el crecimiento del fríjol caupí (Vigna
unguiculata (L.) Walp) en ausencia de bacterias nodulantes tipo Rhizobium sp. en
condiciones semicontroladas de invernadero.
2.2 MATERIALES Y MÉTODOS
2.2.1 Material Vegetal y Localización
El fríjol caupí (Vigna unguiculata (L.) Walp) fue el material vegetal utilizado para el
ensayo realizado en invernadero en el Centro de Biotecnología y Bioindustria (CBB)
de Corpoica, ubicado en el Municipio de Mosquera (Cundinamarca) (4º 42' Latitud
Norte y 74º 12' Longitud Oeste) Temperatura media: 13.1°C; clima frío seco ; 2516
m.s.n.m).
2.2.2 Selección y caracterización de Azotobacter sp.
2.2.2.1 Toma de Muestra y Aislamiento
El muestreo se llevó a cabo a 20 cm de profundidad tomando 200 g por submuestra
de suelo rizosférico del fríjol caupí. Posteriormente, las submuestras se

28
homogenizaron y se transportaron refrigeradas al Laboratorio de Microbiología de
Suelos del CBB de Corpoica.
El aislamiento de Azotobacter sp. se realizó a partir de suelo rizosférico, mediante la
técnica de gránulos de suelo en placas de petri con medio Ashby, las cuales se
incubaron durante siete días a 30 ± 2°C. Para la caracterización se evaluaron las
características morfológicas y de crecimiento de las colonias en medio de cultivo
sólido semiespecífico Ashby y Ashby-benzoato (producción de polímeros y pigmentos
difusibles) de acuerdo a los protocolos descritos por Döbereiner et al., 1995 y
Fenglerowa, 1965; y la forma bacteriana, el tamaño y el tipo de agregación se
determinaron por medio de la coloración de Gram (Döbereiner et al., 1995).
2.2.2.2 Caracterización bioquímica y promoción de crecimiento vegetal
Las pruebas bioquímicas de catalasa y oxidasa se llevaron a cabo de acuerdo con el
protoclo descrito por Holt y Krieg (1994) y Holt et al. (2000). Para la prueba de
Catalasa, con asa de siembra se recogió el centro de una colonia pura de 24 a 48 h
de incubación y se extendió sobre un portaobjetos, luego se agregó una gota de H2O2
al 30%. La formación de burbujas indica la liberación de O2 característico de un
resultado positivo. La prueba de Oxidasa se realizó de acuerdo con el protocolo del kit
de prueba bioquímico para la detección de citocromo-oxidasa en microorganismos
Bactident-Merck®.
Los mecanismos de promoción del crecimiento vegetal como la fijación biológica de
nitrógeno, la solubilización de fosfatos, la producción de hormonas indólicas y la
producción de sideróforos, se evaluaron como sigue:
La actividad nitrogenasa indicadora de FBN se estimó por medio de la técnica de
reducción de acetileno, para lo cual el microorganismo se inoculó en 5 mL de medio
semisólido Ashby y se incubó a 30ºC por 48 h. La FBN fue cuantificada mediante
cromatografía de gases (Perkin Elmer, USA) con detector de ionización de llama y
una columna Poropak N 200/300 mesh de 6 pies y 3 mm de diámetro (Eckert et al.,
2001).
El método cuantitativo de fosfomolibdato fue empleado para determinar la
solubilización de fosfatos. Para esto, cada cepa fue cultivada en medio SRS (Sundara
y Sinha, 1963) sin indicador e incubada por 120 h a 150 rpm; posteriormente, se

29
tomaron alíquotas de 1 mL y se centrifugó a 10.000 rpm; empleando para el análisis
500 μL del sobrenadante (Fiske y Subbarow, 1925).
Producción de Hormonas Indólicas. A partir de la fermentación líquida de cada cepa
se tomaron 4 mL de cultivo y se centrifugaron a 10.000 rpm por 10 min; luego, se
tomó 1mL del sobrenadante y se llevó a reacción con 1mL del reactivo de Salkowsky
modificado; la reacción se incubó por 30 min en oscuridad y por triplicado.
Posteriormente se determinó la absorbancia en un espectrofotómetro a 540 nm y se
reemplazaron los datos en la ecuación de la recta de la curva patrón de Salkowsky
para determinar la producción de índoles totales (Carreño et al., 2000).
La producción de sideróforos se determinó mediante el ensayo de cromo azurol
sulfonato en agar sólido (CAS, Sigma–Aldrich, USA), conforme a lo descrito en
Schwyn y Neilands (1987).
2.2.3 Ensayo en invernadero
Para este ensayo semillas de fríjol caupí, suministradas por el banco de semillas de
Corpoica, fueron mantenidas en cámara de crecimiento a 28°C, con humedad relativa
del 40% y en oscuridad hasta germinación; posteriormente, se inocularon 40 mL de
cada cepa bacteriana a una concentración de 108 UFC/mL.
Longitud y biomasa. Se determinó a los 45 d.d.s: Longitud de la parte aérea (LPA),
longitud de raíz (LR), peso seco de la parte aérea (PSA), peso seco raíz (PSR),
diámetro del tallo (DT), número de hojas (NH) y área foliar (AF).
Determinación de carbohidratos totales y de proteínas. Estas determinaciones se
realizaron en tejidos de parte aérea y raíz.
La determinación de carbohidratos (CH) se realizó de acuerdo con la metodología
descrita por Dubois (1985). A 500 μL de muestra se adicionaron 250 μL de fenol al
5% y 1250 μL de ácido sulfúrico concentrado; esta mezcla se llevó al vortex y se dejó
reposar por 10 min, luego se dispuso en baño termostatado a 30°C por 15 min. La
absorbancia se leyó a 492 nm en espectrofotómetro y se ajustó la información en la
ecuación de la recta de la curva de calibración.
La determinación de proteínas se realizó de acuerdo con la metodología descrita por
Bradford (1987). A 100 μL de la muestra se adicionaron 100 μL de NaOH 1M,
disponiéndola en baño termostatado 5min a 90°C y se dejó enfriar. Posteriormente,
se agregaron 200 μL de HCl 0,5M y 800 μL del reactivo de Bradford. La absorbancia

30
se leyó a 595 nm en espectrofotómetro y se ajustó la información en la ecuación de la
recta de la curva de calibración
Determinación de Clorofila total. El contenido de clorofilas a y b del tejido foliar del
fríjol caupí se estimó por el método de Hiscox e Israelstam (1979), empleando las
ecuaciones descritas por Wellburn (1994); se utilizó como solvente el Dimetilsulfóxido
(DMSO).
2.2.4 Identificación Molecular
El gDNA fue extraído empleando el DNeasy Blood & Tissue Kit (Qiagen, Germany),
siguiendo las instrucciones fabricante, y cuantificado en Nanodrop (Thermo
Corporation, USA). El gDNA fue diluido en agua Milli-Q hasta alcanzar una
concentración de aproximadamente 50 ng/μL antes de realizar la PCR, realizada
empleando los prímers universales siguiendo el protocolo sugerido por Bürgmann et
al. (2004). El tamaño del amplicón (370 pb) fue confirmado usando electroforesis en
gel de 1.0% agarosa en buffer TAE conteniendo 1 μg/mL de bromuro de etidio.
2.2.5 Análisis Estadístico
Para el análisis y procesamiento de la información de las capacidades de promoción
de crecimiento vegetal in vitro, se utilizó el análisis de varianza univariante
(UNIANOVA) y la comparación de medias se realizó por el Test de Tukey al 5% de
probabilidad utilizando el software SAS 9.0 para Windows. La evaluación de la
capacidad de promoción de crecimiento vegetal se realizó por 50 días mediante un
diseño completamente al azar con 16 tratamientos (15 cepas presuntivas de
Azotobacter sp. y el testigo absoluto) y 5 repeticiones. Para el análisis de la
información de la respuesta agronómica en invernadero se realizó el análisis
multivariado mediante el análisis de componentes principales (ACP), utilizando el
software XLSTAT 2011.
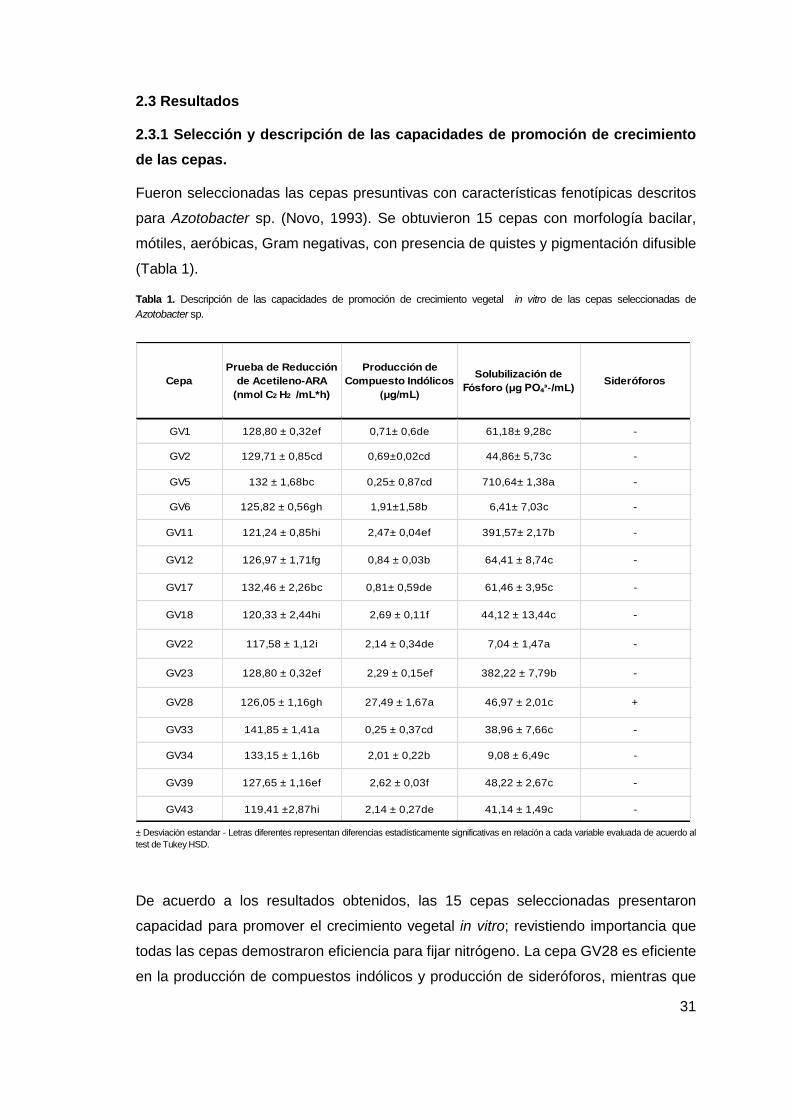
31
2.3 Resultados
2.3.1 Selección y descripción de las capacidades de promoción de crecimiento
de las cepas.
Fueron seleccionadas las cepas presuntivas con características fenotípicas descritos
para Azotobacter sp. (Novo, 1993). Se obtuvieron 15 cepas con morfología bacilar,
mótiles, aeróbicas, Gram negativas, con presencia de quistes y pigmentación difusible
(Tabla 1).
Tabla 1. Descripción de las capacidades de promoción de crecimiento vegetal in vitro de las cepas seleccionadas de
Azotobacter sp.
± Desviación estandar - Letras diferentes representan diferencias estadísticamente significativas en relación a cada variable evaluada de acuerdo al
test de Tukey HSD.
De acuerdo a los resultados obtenidos, las 15 cepas seleccionadas presentaron
capacidad para promover el crecimiento vegetal in vitro; revistiendo importancia que
todas las cepas demostraron eficiencia para fijar nitrógeno. La cepa GV28 es eficiente
en la producción de compuestos indólicos y producción de sideróforos, mientras que
GV1 128,80 ± 0,32ef 0,71± 0,6de 61,18± 9,28c -
GV2 129,71 ± 0,85cd 0,69±0,02cd 44,86± 5,73c -
GV5 132 ± 1,68bc 0,25± 0,87cd 710,64± 1,38a -
GV6 125,82 ± 0,56gh 1,91±1,58b 6,41± 7,03c -
GV11 121,24 ± 0,85hi 2,47± 0,04ef 391,57± 2,17b -
GV12 126,97 ± 1,71fg 0,84 ± 0,03b 64,41 ± 8,74c -
GV17 132,46 ± 2,26bc 0,81± 0,59de 61,46 ± 3,95c -
GV18 120,33 ± 2,44hi 2,69 ± 0,11f 44,12 ± 13,44c -
GV22 117,58 ± 1,12i 2,14 ± 0,34de 7,04 ± 1,47a -
GV23 128,80 ± 0,32ef 2,29 ± 0,15ef 382,22 ± 7,79b -
GV28 126,05 ± 1,16gh 27,49 ± 1,67a 46,97 ± 2,01c +
GV33 141,85 ± 1,41a 0,25 ± 0,37cd 38,96 ± 7,66c -
GV34 133,15 ± 1,16b 2,01 ± 0,22b 9,08 ± 6,49c -
GV39 127,65 ± 1,16ef 2,62 ± 0,03f 48,22 ± 2,67c -
GV43 119,41 ±2,87hi 2,14 ± 0,27de 41,14 ± 1,49c -
Cepa Sideróforos
Prueba de Reducción
de Acetileno-ARA
(nmol C2 H2 /mL*h)
Producción de
Compuesto Indólicos
(μg/mL)
Solubilización de
Fósforo (μg PO₄ᶟ-/mL)
32
GV5, GV11 y GV23 mostraron la capacidad de solubilizar eficientemente fosfatos. De
acuerdo a lo anterior, los resultados evidenciaron que las cepas son eficientes por
diversos mecanismos bajo condiciones controladas. No obstante, fue necesario
establecer las capacidades en planta, bajo condiciones semicontroladas en
invernadero.
2.3.2 Evaluación de las capacidades de promoción de crecimiento vegetal en
condiciones de invernadero.
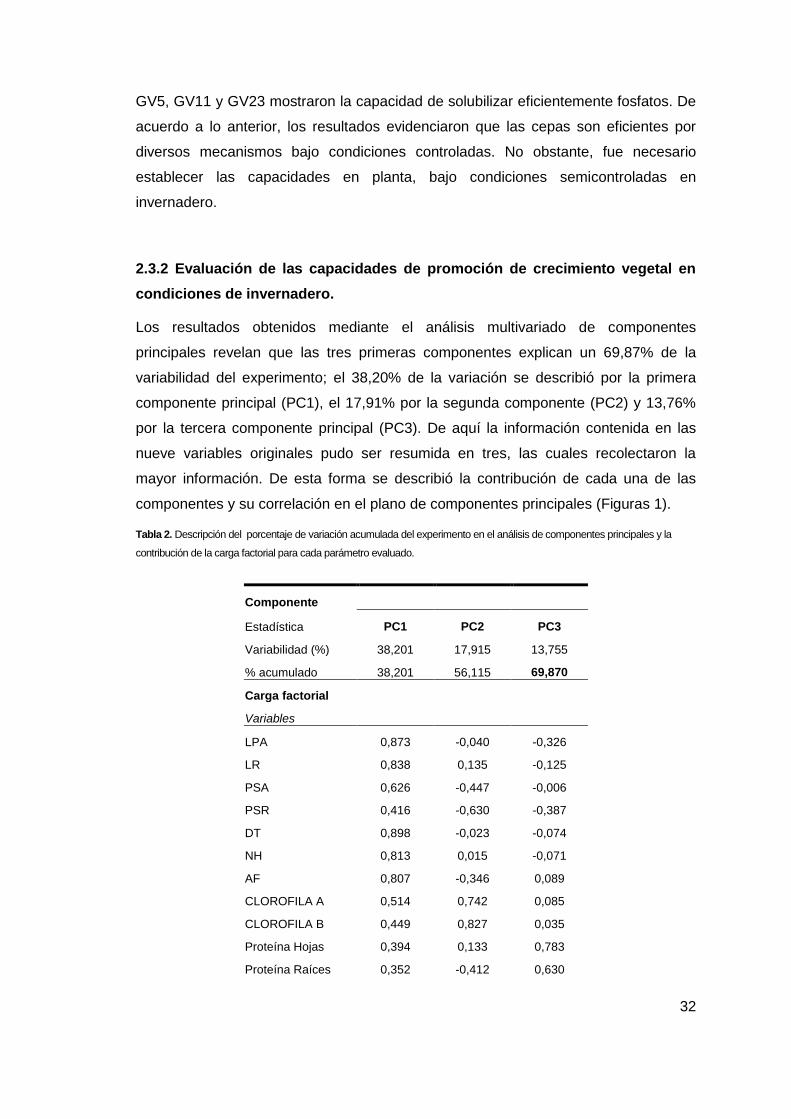
Los resultados obtenidos mediante el análisis multivariado de componentes
principales revelan que las tres primeras componentes explican un 69,87% de la
variabilidad del experimento; el 38,20% de la variación se describió por la primera
componente principal (PC1), el 17,91% por la segunda componente (PC2) y 13,76%
por la tercera componente principal (PC3). De aquí la información contenida en las
nueve variables originales pudo ser resumida en tres, las cuales recolectaron la
mayor información. De esta forma se describió la contribución de cada una de las
componentes y su correlación en el plano de componentes principales (Figuras 1).
Tabla 2. Descripción del porcentaje de variación acumulada del experimento en el análisis de componentes principales y la
contribución de la carga factorial para cada parámetro evaluado.
Componente
Estadística PC1 PC2 PC3
Variabilidad (%) 38,201 17,915 13,755
% acumulado 38,201 56,115 69,870
Carga factorial
Variables
LPA 0,873 -0,040 -0,326
LR 0,838 0,135 -0,125
PSA 0,626 -0,447 -0,006
PSR 0,416 -0,630 -0,387
DT 0,898 -0,023 -0,074
NH 0,813 0,015 -0,071
AF 0,807 -0,346 0,089
CLOROFILA A 0,514 0,742 0,085
CLOROFILA B 0,449 0,827 0,035
Proteína Hojas 0,394 0,133 0,783
Proteína Raíces 0,352 -0,412 0,630

33
CH HOJAS 0,197 0,072 0,518
CH RAÍCES 0,190 0,406 -0,458
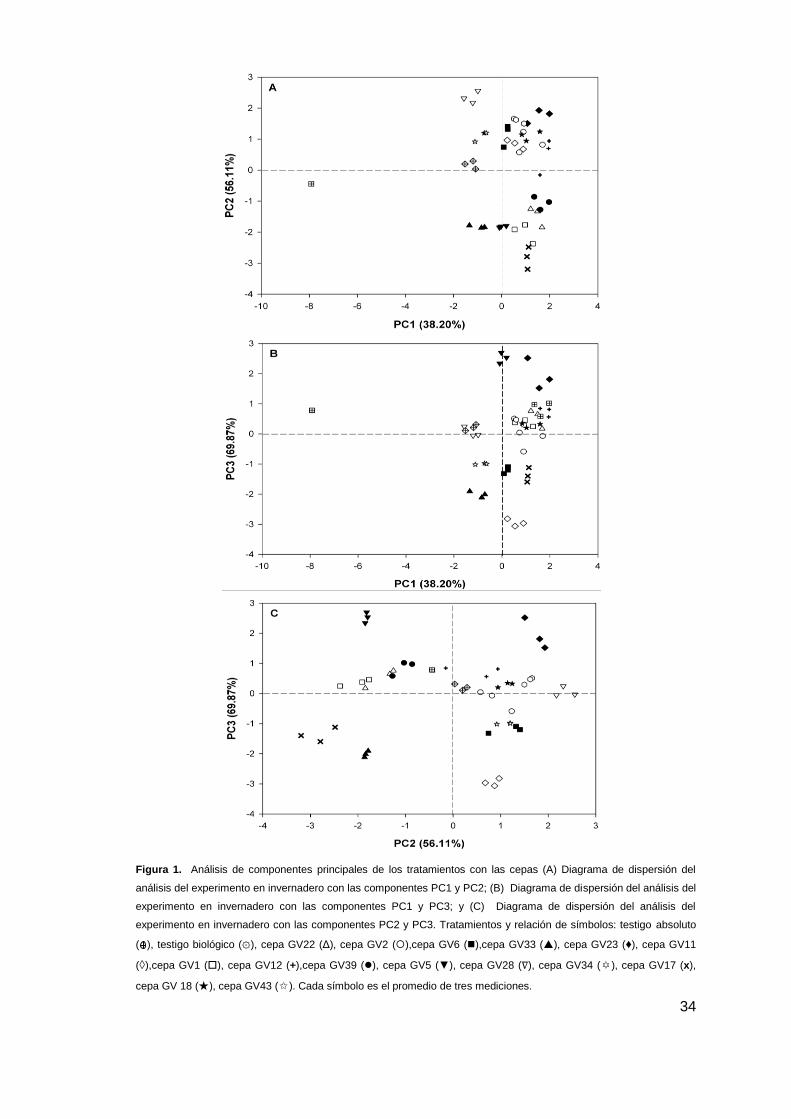
Los tratamientos con inoculación de las cepas GV23, GV18 y GV34 presentaron las
mejores respuestas en la promoción de crecimiento vegetal del fríjol caupí en los 3
componentes principales analizados; Se observa que cuando la componente principal 1
(PC1) se encuentra al lado positivo del eje indica un aumento en el tamaño de la planta
(parte aérea y raíz) cuando se realizó inoculación con las cepas GV23, GV18 y GV34. En el
caso de (PC2) cuando los tratamientos se encuentran al lado positivo del eje indican un
aumento en el contenido de clorofila total en las hojas siendo inversamente proporcional al
peso seco radical, es decir, el resultado más representativo lo representa el tratamiento
inoculado con la cepa GV23 . Con respecto a (PC3), cuando está en el lado positivo implica
que en la planta hay una mayor translocación de asimilados hacia la parte aérea desde las
raíces, en donde el tratamiento GV23 presenta diferencias significativas en esta
componente en comparación con los demás tratamientos evaluados.
34
Figura 1. Análisis de componentes principales de los tratamientos con las cepas (A) Diagrama de dispersión del
análisis del experimento en invernadero con las componentes PC1 y PC2; (B) Diagrama de dispersión del análisis del
experimento en invernadero con las componentes PC1 y PC3; y (C) Diagrama de dispersión del análisis del
experimento en invernadero con las componentes PC2 y PC3. Tratamientos y relación de símbolos: testigo absoluto
( ), testigo biológico (۞), cepa GV22 (Δ), cepa GV2 (),cepa GV6 (),cepa GV33 (), cepa GV23 (♦), cepa GV11
(◊),cepa GV1 (), cepa GV12 (+),cepa GV39 (), cepa GV5 (▼), cepa GV28 (∇), cepa GV34 (✡), cepa GV17 (x),
cepa GV 18 (★), cepa GV43 (✩). Cada símbolo es el promedio de tres mediciones.

35
2.3.3 Identificación mediante determinación y secuenciación del grupo
específico nifH
De acuerdo al análisis multivariado fueron seleccionadas por su capacidad eficiente
para mejorar la respuesta fisiológica del fríjol caupí en condiciones de invernadero,
las cepas GV18, GV23 y GV34. El análisis comparativo de las secuencias del
fragmento de nifH obtenidas con las registradas en la base de datos del NCBI mostró
que las 3 cepas se acercaron en un porcentaje del 98% de identidad con la secuencia
de Azotobacter sp. (Figura 2).
Figura 2. Determinación del grupo específico nifH para las cepas GV18, GV23 y GV34
2.4 Discusión
2.4.1 Selección y descripción de las capacidades de promoción de crecimiento de
las cepas.
Los resultados evidenciaron que las cepas son eficientes por diversos mecanismos
bajo condiciones controladas. La capacidad del Azotobacter sp. de mantener una
eficiencia en la fijación de nitrógeno depende de las condiciones de crecimiento y
disponibilidad de nutrientes. En Azotobacter sp. se dispone al menos de cuatro
mecanismos para proteger la nitrogenasa, considerándose como los más importantes
el mecanismo de autoprotección y la protección conformacional (Soto y Baca, 2001).
En la prueba de fijación de nitrógeno, todas las cepas presentaron eficiencia, con una
producción entre 117,58 - 141,85 nmol C2H2 mL-1*h. Estos resultados coinciden con

36
los reportados por Lu et al. (2009) quienes obtuvieron cepas de Azotobacter sp. que
alcanzaron una máxima producción de 139,79 C2H2 mL-1*h. Adicionalmente, estos
valores son superiores a los registrados por múltiples autores con respecto a la
producción de otras cepas diazotróficas de vida libre como A. beijerinckii (Ravikumar
et al., 2004), A. lipoferum (Mascarua-Esparza et al.,1988), A. lipoferum (Mehnaz y
Lazarovits, 2006), entre otros.
En cuanto a la producción de reguladores de crecimiento vegetal Azotobacter sp. junto
con Azospirillum sp. y Pseudomonas sp. han sido reportados como sintetizadores de este
tipo de compuestos (Patten y Glick 1996, González et al., 1986). La cepa GV28 presentó
diferencias significativas (p < 0.05) con respecto a las 15 cepas en evaluación en cuanto a
la producción de compuestos indólicos (27,49 μg/mL). Ravikumar et al. (2004) reportaron
cepas aisladas de Azotobacter chroococcum con una producción de ácido
indolacético de 18,5 μg/mL, seguido por A. beijerinckii con 16,5 μg mL y A. vinelandii
13,8 μg/mL; Torres et al. (2000), realizaron el asilamiento de 18 cepas del género
Azotobacter sp. en 20 cultivos de arroz en el Tolima (Colombia) y encontraron
importantes producciones de hormonas indólicas tipo ácido indolacético (AIA),
incluyendo a A. vinelandii con 32,22 μg/mL y A. chroococcum con 30,07 μg/mL de
AIA.
La solubilización de los fosfatos insolubles en la rizosfera es uno de los mecanismos
más comunes de la acción de las bacterias promotoras del crecimiento vegetal
(BPCV), lo cual permite mejorar la disponibilidad de nutrientes para las plantas
(Richardson et al., 2001; Rodríguez et al., 2006). En esta prueba, GV5 (710,64 μg
PO43-/mL), GV11 (391,57 μg PO4
3-/mL) y GV23 (382,22 μg PO43-/mL) mostraron la
capacidad de solubilizar eficientemente fosfatos.
Yi et al. (2007) reportaron resultados similares por cepas de Enterobacter sp. – EnHy-
401 (632,6 μg PO43-/mL), Artrhobacter sp. – ArHy-505 (428,90 μg PO4
3-/mL) y
Azotobacter sp. - AzHy – 510 (229,03 μg PO43-/mL), lo cual permite inferir que las
cepas con valores similares en este estudio tienen gran habilidad de solubilizar
fosfato tricálcico; Chaiharn et al. (2011) reportaron valores de producción de 216
cepas, destacándose Acinetobacter cepa CR 1,8, la cual tuvo el mayor valor de
solubilización (334 ± 0,4 μg PO43-/mL). Así mismo, Anandham et al. (2007) reportaron
la eficiencia en solubilicación de fosfato tricálcico en medio líquido Pikovskaya por
Burkholderia sp. – CBPB-HOD (359,27 ug PO43-/mL) a las 48 horas de incubación.

37
No obstante, la producción de sideróforos por Azotobacter sp. en esta investigación
fue positiva sólo por la cepa GV28. Con respecto a esta capacidad, numerosos
resultados demuestran que la producción de sideróforos está reportado para los
géneros promotores de crecimiento vegetal como Pseudomonas, Rhizobium, Bacillus
y Azotobacter (Bachhawaat, 1987; Cornelis y Matthisj, 2007; Crowley 2006; Glick et
al., 1999; Shah et al., 1992; Tortora et al., 2011).
2.4.2 Evaluación de las capacidades de promoción de crecimiento vegetal en
condiciones de invernadero.
El aumento en el tamaño de la planta (parte aérea y raíz) cuando se realizó inoculación con
las cepas GV23, GV18 y GV34, evidencia el efecto positivo que ejercen sobre el fríjol caupí
(PC1). Lo que permite inferir que la inoculación con estos microrganismos estimula la
eficiencia de las plantas en la acumulación de reservas y transporte efectivo de asimilados
hacia los vertederos en posteriores estadíos como floración y llenado de grano (Radford,
1967; Hunt, 1990; Borrego et al., 2000).
En el caso de PC2 cuando los tratamientos se encuentran al lado positivo del eje indican un
aumento en el contenido de clorofila total, siendo inversamente proporcional al peso seco
radical, es decir, el resultado más representativo está reflejado en el comportamiento del
tratamiento inoculado con GV23. De otra parte, la disminución de la biomasa seca en
raíces probablemente está relacionada a las condiciones particulares en las que se realizó
este estudio.
Estudios de inoculaciones de bacterias no simbióticas asociadas al fríjol caupí en
condiciones de invernadero han sido muy limitados; esta respuesta permite inferir que de
alguna forma la planta entró en condiciones de estrés por la búsqueda de mecanismos
para la fijación eficiente del nitrógeno en ausencia de bacterias nodulantes o rizobios.
Como lo mencionan Freiberg et al. (1997), quienes afirman que la deficiencia de
nitrógeno mineral frecuentemente limitan la planta en relación al crecimiento y toma
recursos de la interacción simbiótica que han desarrollado entre las plantas y una variedad
de organismos fijadores de nitrógeno. De acuerdo a lo anterior se puede inferir que la
limitación de nitrógeno (mineral o de naturaleza simbiótica) indujo a la planta a tomar
nutrientes en mayor escala por vía fotosintética para la obtención de fotoasimilados.
Adicionalmente, se ha descrito que bajo condiciones de estrés la planta invierte sus
recursos de forma diferente entre los órganos en función de captación de luz y asimilación
de dióxido de carbono y fotosintatos (hojas), captación de agua y nutrientes minerales

38
(raíces) (Poorter y Nagel, 2000). Existe por tanto, un compromiso para la planta en una
mayor asignación de biomasa a las hojas para garantizar una eficiente captación de
asimilados (Castro-Diez, 2002; Poorter y Nagel, 2000).
Con respecto a (PC3), cuando está en el lado positivo implica que en la planta hay una
mayor translocación de asimilados hacia la parte aérea desde las raíces. Esta asignación y
translocación de fotoasimilados desde la raíz puede explicar los bajos gradientes entre los
fotoasimilados de la raíz y su aumento en la parte aérea (Cruz et al., 2005).
Correlacionando las respuestas observadas en (PC2) y (PC3), se puede inferir que la
planta, en ausencia de población rizobiana y sin formación de nódulos maximizó el trabajo
fotosintético y la acumulación de reservas, debido a que el carbono requerido para el
llenado de las semillas deriva de reservas que dependen de la disponibilidad de la fuente
fotosintética durante este proceso (Peoples et al., 1995).
Por esta razón el aumento de la clorofila total junto con la inoculación de Azotobacter sp.
podrían deducir una eficiencia en la captación de asimilados, como medida alternativa a la
fijación simbiótica de nitrógeno simbiótica. Aunque en este estudio se infieren condiciones
de estrés por las respuestas observadas en PC2 y PC3 también se observó que todos los
tratamientos con inoculación superaron al testigo en todos los componentes en estudio, lo
cual evidencia que Azotobacter sp. es un factor benéfico que favorece y mejora el
desarrollo del fríjol caupí por diferentes mecanismos. El efecto promotor de crecimiento
vegetal de Azotobacter sp. no sólo se ha relacionado con fijación de nitrógeno como
recursos disponibles proporcionado a la planta, también se ha demostrado que están
involucrados en el crecimiento vegetal por la producción de reguladores de
crecimiento, tales como fitohormonas y vitaminas, siendo ampliamente reportadas
(González et al., 1986;Martínez - Toledo et al., 1991; Arshad y Frankenberger, 1991;
Obando et al., 2010).
Además, el efecto potenciador de la absorción de nutrientes y el crecimiento vegetal
se ha correlacionado con otras capacidades como solubilización de minerales y
producción de sideróforos (Crowley et al., 1991; 2006), incremento en el volumen de
la raíz (Bowen y Rovira, 1999), inducción de resistencia sistémica a patógenos (Van
Peer et al., 1991), inhibición del crecimiento de organismos antagónicos e interacción
sinérgica con otros microorganismos del suelo (Bashan et al., 1996); siendo estos
factores activadores de procesos metabólicos de compuestos orgánicos en las
plantas que a su vez estimulan el crecimiento vegetal, ejerciendo un efecto directo en
factores de nutrición y rendimiento (El-Seifi et al.,2004).

39
2.4.3 Identificación mediante determinación y secuenciación del grupo
específico nifH
La caracterización molecular de bacterias diazotróficas puede determinarse a partir
de varios genes presentes en estos microorganismos que codifiquen para funciones
específicas, como los nif para la nitrogenasa (Roesch et al., 2006). El análisis
comparativo de las secuencias del fragmento de nifH determinó que las cepas GV18,
GV23 y GV34 se acercaron en un porcentaje del 98% de identidad con la secuencia
de Azotobacter sp.
Los miembros de este género son bacterias de vida libre, aeróbicos diazótrofos
heterótrofos que se encuentran en ambientes de suelos y de agua dulce. Son
especialmente interesantes desde el punto de vista de la fijación de Nitrógeno ya que
dependiendo de la especie, poseen dos o tres sistemas de nitrogenasa genéticamente
distintas, cuya expresión está determinada por la disponibilidad de metales como el Hierro y
Molibdeno (Bishop y Joerger, 1990). nifH junto con nifD y nifK codifican en su orden para
el complejo Fe-proteína y las subunidades alfa y beta codifican para el complejo MoFe-
proteína; que en su conjunto forman el complejo de enzima nitrogenasa (Evans et al.,
1988).
De acuerdo a esto, la identificación de este fragmento sumado a la caracterización
fenotípica y bioquímica, reconfirman la presencia de tres cepas estrechamente
relacionadas con Azotobacter sp. con capacidades de fijación de nitrógeno y descrita
ampliamente por su capacidad de promoción de crecimiento vegetal en diversas
especies de interés agrícola en el mundo (Mehnaz y Lazarovits, 2006).
Conclusiones
Bajo las condiciones en las cuales se realizó el presente estudio se concluye que las
cepas GV18, GV23 y GV34 promovieron capacidades de crecimiento vegetal del fríjol
caupí (Vigna unguiculata (L.) Walp), incrementando el aumento en el tamaño de la
planta (parte aérea y raíz), contenido de clorofila y aumento en la translocación de
asimilados hacia las estructuras foliares presentes en el proceso del llenado posterior
del fruto (grano). De acuerdo a lo anterior, Azotobacter sp. es eficiente en la
promoción del crecimiento de esta especie en ausencia de bacterias nodulantes tipo
Rhizobium sp. en condiciones semicontroladas de invernadero, lo cual permite inferir
que podría utilizarse en coinóculos mixtos sin afectar parámetros de rendimiento
agronómico.

40
Capítulo III. Efectos de la coinoculación Rhizobium - Azotobacter sobre la
nodulación, el crecimiento, y el valor nutricional del fríjol caupí (Vigna
unguiculata (L.) Walp) en condiciones de invernadero.
Resumen
En esta investigación se estudió la respuesta fisiológica del fríjol caupí (Vigna
unguiculata (L.) Walp) a la coinoculación de Rhizobium sp. y Azotobacter sp. en
condiciones de invernadero en la estación experimental Motilonia en el municipio de
Agustín Codazzi (Cesar). El trabajo se realizó con las cepas nativas GV23 y GV18 de
Azotobacter sp. previamente evaluadas y caracterizadas, y las cepas nativas G56 y
G58 de Rhizobium sp., las cuales mediante la aplicación de la técnica de
caracterización molecular 16S rDNA reveló en un 99% de similitud en su identidad. Las
semillas del fríjol caupí se coinocularon y sembraron en suelo estéril de la zona de
estudio; las observaciones se realizaron hasta la floración. Se determinó el porcentaje
de germinación y parámetros agronómicos de crecimiento a los 15, 30 y 45 días
después de la siembra (d.d.s). El tratamiento que presentó los mejores resultados en
las plantas fue replicado en condiciones gnobióticas in vitro para determinar la
colonización rizosférica de los microorganismos evaluados mediante microscopía
electrónica de barrido-SEM. El análisis de varianza mostró diferencias
estadísticamente significativas (p < 0.05) del tratamiento T6, en las variables de
longitud y acumulación de biomasa en las diferentes etapas de evaluación, en
comparación con los demás tratamientos; mediante el análisis multivariado de
componentes principales, los tratamientos T6 y T9 presentaron diferencias
significativas con respecto a los demás tratamientos en cuanto a la promoción del
crecimiento vegetal del fríjol caupí expresado en el contenido de biomasa, número y
peso de nódulos y longitud radical. Se infiere que la colonización de los
microorganismos del tratamiento T6 en los nódulos y sistema radical del fríjol caupí
está asociado al mejoramiento de los indicadores de crecimiento y rendimiento de la
planta con el coinóculo empleado.
Palabras clave: Fríjol caupí, Promoción de crecimiento vegetal, Coinoculaciones,
Azotobacter sp.

41
Abstract
In this research we studied the physiological response of cowpea (Vigna unguiculata
(L.) Walp) to co-inoculation of Rhizobium sp. and Azotobacter sp. in the greenhouse at
the E.E Motilonia, Agustin Codazzi (Cesar). The work was performed with the native
strains GV23 and GV18 of Azotobacter sp. previously tested and characterized, and
the native strains G56 and G58 of Rhizobium sp., which by applying the technique of
16S rDNA molecular characterization revealed 99% similarity in their identity. The
seeds of bean and cowpea planted co - inoculated in sterile soil in the area of study,
observations were made until flowering. We determined the percentage of germination
and agronomic parameters of growth at 15, 30 and 45 days after sowing. The
treatment presented the best results in the plants was replicated in vitro gnobiotics
able to determine rhizosphere colonization of microorganisms assessed by scanning
electron microscopy - SEM. Analysis of variance showed statistically significant
differences (p <0.05) T6 treatment, the variables of length and biomass accumulation
in different stages of evaluation, compared with other treatments, by multivariate
principal component analysis, treatments T6 and T9 showed significant differences
compared to other treatments in promoting plant growth expressed in cowpea
biomass content, number and weight of nodules and root length. It is inferred that the
colonization of microorganisms in the lymph T6 treatment and cowpea bean root
system is associated with improved indicators of growth and yield of the plant with the
coinoculated.
Keywords: Cowpea bean, Plant growth promotion, Mixed culture, Azotobacter sp.
3.1 Introducción
El fríjol caupí (Vigna unguiculata (L.) Walp) es una de las principales leguminosas
forrajeras con potencial cultivable en la Región Caribe Colombiana, que ocupa un
área sembrada de 4.600 hectáreas (DNP, 2010). Este cultivo se constituye como una
excelente provisión de forraje para los animales en temporadas donde normalmente
no existe ninguna otra fuente de alimento; a su vez es un recurso complementario de
la dieta, debido a que puede crecer sola o en asociación con otras gramíneas o
leguminosas (Calegari, 1995). Sin embargo, su oferta es muy baja en proporción a los
requerimientos para consumo animal y humano, por lo que actualmente se promueve
la siembra de esta especie en regiones tropicales (DNP, 2010).

42
Debido a las limitaciones edafoclimáticas que caracterizan los agroecosistemas del
trópico bajo, en la microrregión del Valle del Cesar la producción de forraje es escasa
e irregular durante el año. Durante la época seca, la producción de forraje se reduce
considerablemente y su calidad se deteriora comprometiendo la producción de carne
debido a la pérdida de peso de los animales. Estas limitaciones pueden atribuirse a la
disminución en la activación del proceso simbiótico, la formación de nódulos y por
ende en la poca efectividad en la fijación biológica de nitrógeno, los cuales son
sensibles estrés ambiental (Hungría et al., 2000).
De otra parte, debido a sus implicaciones agronómicas, las interacciones ecológicas entre
los rizobios y otras bacterias del suelo han sido de especial interés en los últimos años. Se
han registrado evidencias de interacciones positivas entre las especies de Rhizobium y
bacterias diazotróficas del suelo, como Azotobacter y Azospririllum, del Gallo y Fabbri
(1991) Dardanelli et al. (2008) y Benintende et al. (2010), entre otros autores, han reportado
aumento de la nodulación y crecimiento óptimo de una amplia variedad de forrajes.
Azotobacter sp. (especialmente A. chroococcum) ha sido ampliamente reportado por
promover el crecimiento vegetal en condiciones edáficas de estrés por sequía. Este género
se encuentra ampliamente distribuido en suelos destinados a la agricultura, predominante
en regiones tropicales y se pueden encontrar fácilmente en asociación con cereales y en la
rizosfera de leguminosas (Kole et al., 1988; Martínez-Toledo et al., 1991; Gharib et al.,
2009).
Los efectos positivos de la inoculación combinada con Rhizobium - Azotobacter y otras
rizobacterias estimuladoras del crecimiento han demostrado que aumentan
significativamente el número y peso de nódulos, mayor fijación de nitrógeno, aumento del
contenido de macronutrientes y micronutrientes, en comparación con la inoculación
individual de Rhizobium sp. (Yahalom et al., 1987; Rodelas et al., 2000). El agente
potenciador de estos beneficios ha sido corelacionado con la influencia positiva de la
bacterias promotoras de crecimiento no simbióticas, ya que se verifican aumentos
significativos en factores de rendimiento como la acumulación de masa seca y el contenido
de nitrógeno en grano (Burns et al., 1981; Sarig et al., 1986; Rodelas et al., 2000).
En Colombia no ha sido explorado el estudio de coinoculaciones con rizobios y bacterias de
vida libre en leguminosas; su evaluación permitirá determinar su potencial de promoción
del crecimiento de las plantas ante el efecto de la marcada estacionalidad en las
precipitaciones del Caribe seco, permitiendo de esta forma la utilización de recursos
biológicos poco explorados y contribuyendo con tecnologías limpias que favorecen y

43
mejoran las condiciones de especies forrajeras de interés en sistemas de ganadería
de la región.
La presente investigación tuvo como objetivo de esta investigación fue estudiar bajo
condiciones semicontroladas la influencia de la coinoculación con Rhizobium y
Azotobacter sobre la nodulación, crecimiento y calidad nutricional del fríjol caupí
(Vigna unguiculata (L.) Walp) en el Municipio de Agustín Codazzi (Cesar).
3.2 Materiales y Métodos
3.2.1 Identificación molecular de las cepas G56 y G58 mediante la amplificación
del fragmento 16S rDNA
Extracción de ADN genómico. La extracción de ADN genómico (gADN) se realizó
utilizando el Wizard kit (Promega Corporation, Madison, USA). El gADN extraído fue
analizado de acuerdo a Sambrook y Russell, (2001) y cuantificado mediante el uso de
espectrofotometría (NanoDrop, Thermo Corporation).
Amplificación de la secuencia 16S rDNA. Para la amplificación del 16S rDNA se utilizó
la técnica de reacción en cadena de la polimerasa (PCR) utilizando los cebadores
27F y 1492R descritos por Osborne et al. (2005). Cada reacción de amplificación tuvo
un volumen final de 50 μL. Las reacciones de amplificación se realizaron en un
Termociclador PTC 100 (MJ Research, USA). Los ciclos de amplificación consistieron
en una desnaturalización inicial a 94 ºC durante 3 min seguido por 30 ciclos de
desnaturalización a 94 ºC durante 1 min, una etapa de anillamiento a 58 ºC durante 1
min y una etapa de extensión a 72 ºC durante 2 min. Finalmente, una última etapa de
extensión a 72 ºC durante 5 minutos.
Purificación del producto de amplificación. La purificación de los fragmentos
amplificados 32 μL de producto de PCR se mezclaron con 0,1 volúmenes de acetato
de sodio 3M (pH 5,5) y 2,5 volúmenes de etanol frío al 99,8%. La mezcla se incubó a
20 ºC durante la noche. Posteriormente la mezcla fue centrifugada durante 30
minutos a 16.000 g a 4 ºC, donde el sobrenadante fue descartado. Se añadieron 300
µL de etanol frío al 70% y la muestra se centrifugó durante 10 min a 16.000g y 4 ºC, y
nuevamente se descartó el sobrenadante. El precipitado se mantuvo a temperatura
ambiente hasta el secado completo y luego se resuspendió en 20 μL de H2O para
PCR (Sambrook et al., 1989).

44
Secuenciación de los fragmentos. La secuenciación de fragmento 16S rDNA de la
cepa se llevó a cabo en el Laboratorio de Genoma de Embrapa Agrobiología-Brasil
empleando los cebadores descritos en la Tabla 2. Para la reacción de secuenciación
se utilizaron 200 ng de los productos purificados de PCR, 1 µL de cada cebador 5,0
mM, 4,0 µL del kit "TE Dynamics Kit" (GE Healthcare) y agua ultrapura
(ultraPURETM, Invitrogen Co) hasta un volumen final de 10 µL. Las condiciones para
la reacción de secuenciación fueron: 25 ciclos de desnaturalización anillamiento y
extensión a 95 ºC durante 25 seg, 58 ºC durante 15 segundos, 60 ºC durante 1
minuto, respectivamente. Después de marcar la reacción se precipitaron las muestras
por adición de 1,0 µL de acetato de amonio 7,5 M y 27,5 µL de etanol y se incubaron
a 4 ºC durante la noche, después de la incubación se centrifugó a 3000 g por 45
minutos a 4 ºC, el sobrenadante fue descartado y el pellet se lavó con 100 µL de
etanol 70% y nuevamente centrifugado a 3000 g durante 10 min a 4 ºC. Las muestras
fueron secadas al aire y se resuspendieron en 7,5 µL de buffer de secuenciación
Mega BACE ("loading") y analizadas en un secuenciador automático Mega
BACE1000 (GE Healthcare). Las secuencias contiguas se reunieron a partir de las
secuencias directas y reversas con los programas PHRED/PHRAP (Ewing et al.,
1998; Ewing y Green, 1998) o CAP3/CONSED (Gordon et al., 1998; Huang y Madan,
1999) en un entorno Linux para el sector de bioinformática, Embrapa Agrobiología.
Tabla 3. Iniciadores utilizados para la secuenciación del fragmento 16S rDNA de las cepas G56 y
G58.
Las secuencias obtenidas fueron comparadas con otras en la base de datos de
BLAST (http://www.ncbi.nlm.nih.gov/BLAST/) del Centro Nacional de Información
Biotecnológica NCBI. Para la construcción del árbol filogenético se utilizó la
metodología Neighbor-Joining (Saitou y Nei, 1987), las secuencias de interés fueron
Iniciador Secuencia (5’ 3’) Referencia
27f AGA GTT TGA TCC TGG CTC AG
Osborne et al., 2005
1492R GGY TAC CTT GTT ACGA CTT Osborne et al., 2005
362F CTC CTA CGG GAG GCA GCA GTT GGG Menna et al., 2006
786F CGA AAG CGT GGG GAG CAA ACA GG Menna et al., 2006
RU6 ATG GCT GTC GTC AGC TCG Wang et al., 1996
U3 GWA TTA CCG CGG CKG CTG Wang et al., 1996
U2 TGC TGC CTC CCG TAG GAG Wang et al., 1996
1110R TTA GCA AGT AAG GAT ATG GGT Cho et al., 2006

45
extraídas de la base de datos de BLAST y se alinearon con la función ClustalW del
programa informático MEGA 5 (Tamura et al., 2011).
3.2.2 Ensayo en invernadero
3.2.2.1 Condiciones del experimento
Soporte y material vegetal. Las semillas del fríjol caupí (Vigna unguiculata (L.) Walp)
fueron suministradas por el banco de semillas de Corpoica E.E. Codazzi, ubicada en
el Municipio de Agustín Codazzi (Cesar) (10° 12” Latitud Norte – 73° 13” Long. Oeste;
Temperatura media: 28.7°C; clima semiárido; 180 m.s.n.m). Las semillas fueron
seleccionadas y desinfectadas con etanol al 70% por 5 min y luego con solución de
hipoclorito de sodio 3% durante 5 min. El soporte fue suelo de la misma zona de
estudio previamente esterilizado y se dispuso en bolsas de plástico con capacidad de
3 kg con 2,5 kg de soporte estéril.
Microorganismos y condiciones de crecimiento. Los microorganismos nativos
previamente caracterizados se preservaron en el Banco de Trabajo del Laboratorio de
Microbiología de Suelos del CBB - CORPOICA. Para este ensayo se utilizaron las
cepas G56 y G58 de Rhizobium sp. y las cepas GV23 y GV18 de Azotobacter sp.; las
cepas se incubaron durante 24 h en agitación a 120 rpm a 30 °C en medio de cultivo
YMA (Vincent, 1970) y LB (Bertani, 1952; Luria y Burrous, 1957) respectivamente. Los
controles de calidad presentaron 100% de pureza y la concentración celular de 108
UFC/mL.
3.2.2.2 Parámetros Evaluados
Las variables evaluadas a los 15, 30 y 45 días después de la siembra (d.d.s) fueron:
Longitud parte aérea (LPA), longitud de raíz (LR), peso seco parte aérea (PSA) y
peso seco raíz (PSR); para los 45 d.d.s se determinó además el número de los
nódulos (NN), peso fresco de los nódulos (PFN) y peso seco de nódulos (PSN).
Determinación de carbohidratos totales y de proteínas. Estas determinaciones se
realizaron en tejidos de parte aérea y raíz.
La determinación de carbohidratos (CH) se realizó de acuerdo con la metodología
descrita por Dubois (1985). A 500 μL de muestra se adicionaron 250 μL de fenol al
5% y 1250 μL de ácido sulfúrico concentrado; esta mezcla se llevó al vortex y se dejó
reposar por 10 min, luego se dispuso en baño termostatado a 30°C por 15 min. La

46
absorbancia se leyó a 492 nm en espectrofotómetro y se ajustó la información en la
ecuación de la recta de la curva de calibración.
La determinación de proteínas se realizó de acuerdo con la metodología descrita por
Bradford (1987). A 100 μL de la muestra se adicionaron 100 μL de NaOH 1M,
disponiéndola en baño termostatado 5min a 90°C y se dejó enfriar. Posteriormente,
se agregaron 200 μL de HCl 0,5M y 800 μL del reactivo de Bradford. La absorbancia
se leyó a 595 nm en espectrofotómetro y se ajustó la información en la ecuación de la
recta de la curva de calibración
3.2.3 Diseño experimental y Análisis Estadístico
La evaluación se realizó por 45 días aplicando un diseño experimental
completamente al azar con 9 tratamientos y 10 repeticiones; el control químico
correspondió a fertilización con 50 kg/ha de Urea y los controles biológicos fueron
AC1 (Azotobacter chroococcum) y L5 (Rhizobium sp.) provisionadas por el banco de
germoplasma del laboratorio de Microbiología de Suelos del CBB – Corpoica.
Los tratamientos evaluados para determinar la respuesta del fríjol caupí a la
inoculación mixta fueron los siguientes: T1: Testigo Absoluto; T2: Testigo Químico (50
kg/ha de urea); T3: Testigo Biológico 1 (Inoculación 100% Azotobacter – AC1); T4:
Testigo Biológico 2 (inoculación 100% Rhizobium – L5); T5: Testigo inoculación mixta
AC1 + L5; T6: inoculación mixta GV18 + G56; T7: inoculación mixta GV18 + G58; T8:
inoculación mixta GV23 + G56; T9: inoculación mixta GV23 + G58.
Para el análisis y procesamiento de la información de longitud y aumento de biomasa
a los 15, 30 y 45 días después de la siembra, se utilizó el análisis de varianza
univariante (UNIANOVA) y la comparación de medias se realizó por el Test de Tukey
al 5% de probabilidad utilizando el software SAS 9.0 para Windows. Para el análisis
de la información de la respuesta agronómica en el estadio de madurez fisiológica (45
d.d.s) se realizó el análisis multivariado mediante el análisis de componentes
principales (ACP), utilizando el software XLSTAT 2011.
3.2.4 Microscopía electrónica de Barrido (MEB)
A partir de los resultados obtenidos en invernadero se realizó la determinación de la
colonización rizosférica del tratamiento que presentó la mejor respuesta del fríjol
caupí a partir de cortes longitudinales de nódulo (presencia de rizobios) y raíz

47
(presencia de Azotobacter sp.). La visualización se realizó mediante Microscopía
Electrónica de Barrido (MEB) (FEI QUANTA 200); el procesamiento de la muestra se
realizó mediante la metalización en un Sputter SDC-050 de Marca (Balzers), en
condiciones de prevacío (<10-1 torr) con argón como gas de ataque (plasma) sobre
una placa (cátodo) de oro-paladio (8:2). La película se depositó sobre las muestras
(ánodo) a corriente de descarga de +/- 50mA y el espesor típico fue de +/- 200 nm. La
muestra ya procesada fue visualizada en diferentes campos mediante el microscopio
de barrido permitiendo destacar la morfología del material y demás propiedades de la
misma.
3.3 Resultados
3.3.1 Identificación molecular de las cepas G56 y G58 mediante la amplificación
del fragmento 16S rDNA
Previamente se realizó una reactivación y caracterización confirmativa de las cepas,
incubadas 72 h en medio de cultivo YMA, en la cual se observó colonias de
aproximadamente 5.0 mm de diámetro, con borde circular, liso y convexo, y con formación
de moco blanco opaco. La reacción de Gram fue negativa; por lo cual se evidenciaron
características representativas del género Rhizobium de acuerdo a las descripciones
realizadas por Kuykendall et al. (2005) y Carlson et al. (1994).
Para la identidad molecular los resultados mostraron que las cepas G56 y G58 amplificaron
la banda del tamaño esperado, aproximadamente 1500 pb (Figura 3).
Figura 3. Amplificación del fragmento 16S rDNA de las cepas G56 y G58 mediante electroforesis en gel de agarosa al 2%. MP:
Marcador de peso molecular 1000 kb plus.
GG5566 GG5588 MMPP
1650 pb
1000 pb

48
La secuenciación del fragmento mostró que las cepas G56 y G58 se encuentran asociadas
con el género Rhizobium con un 99% de identidad; consecuentemente el análisis de las
secuencias obtenidas fue representado mediante la construcción de un árbol filogenético
con 18 secuencias de referencias que fueron utilizadas a partir de la base de datos del
NCBI, de acuerdo a los porcentajes de similitud obtenidos en el análisis realizado en
BLAST (Figura 4).
Figura 4. Árbol filogenético realizado mediante la amplificación del 16S rDNA. La filogenia fue inferida por el método de
Neighbor-Joining (Saitou y Nei, 1987) utilizando el software Mega 5 (Tamura et al., 2011). Las distancias evolutivas se calcularon
utilizando el parámetro de Kimura 2-método (Kimura, 1980). Los números en los nodos representan el porcentaje de
bootstraping con 10000 repeticiones. La barra de escala = 0,01 sustituciones por sitio.
3.3.2 Evaluación en invernadero
Longitud de la parte aérea. Los resultados obtenidos presentaron diferencias
significativas (p < 0.05) de los tratamientos T2 y T8 con respecto a los demás
tratamientos evaluados a los 15 d.d.s. En el muestreo realizado a los 30 d.d.s la
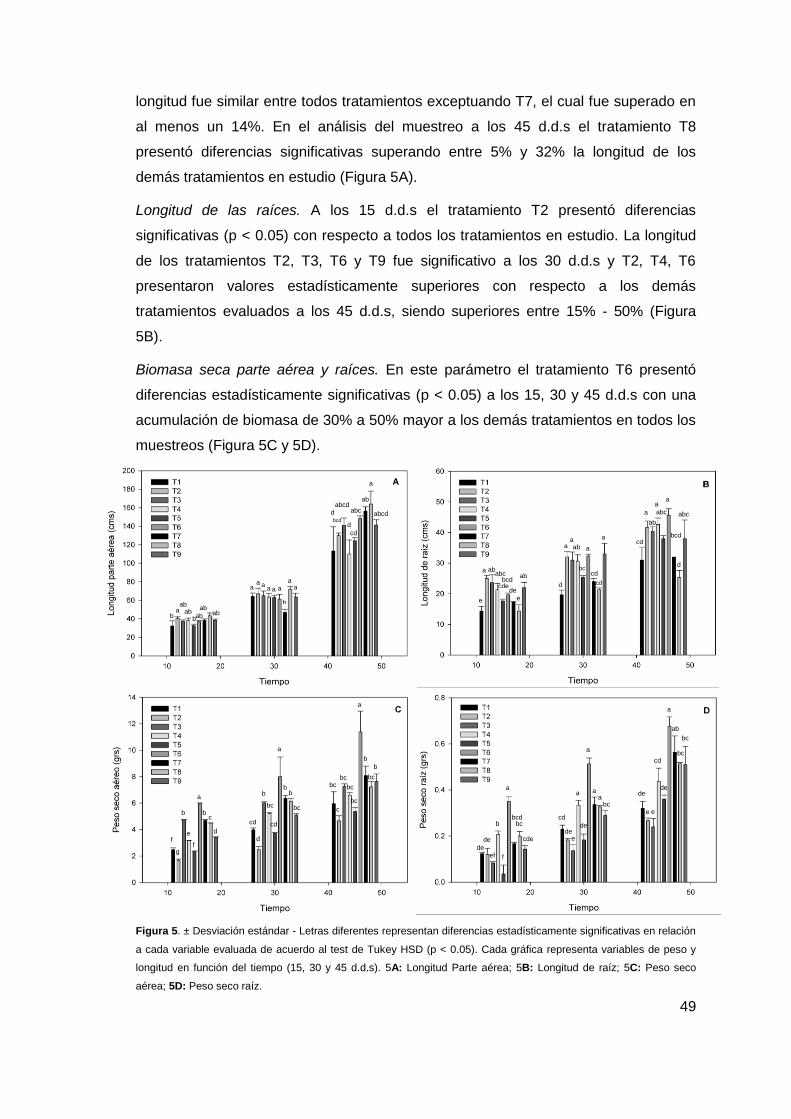
49
longitud fue similar entre todos tratamientos exceptuando T7, el cual fue superado en
al menos un 14%. En el análisis del muestreo a los 45 d.d.s el tratamiento T8
presentó diferencias significativas superando entre 5% y 32% la longitud de los
demás tratamientos en estudio (Figura 5A).
Longitud de las raíces. A los 15 d.d.s el tratamiento T2 presentó diferencias
significativas (p < 0.05) con respecto a todos los tratamientos en estudio. La longitud
de los tratamientos T2, T3, T6 y T9 fue significativo a los 30 d.d.s y T2, T4, T6
presentaron valores estadísticamente superiores con respecto a los demás
tratamientos evaluados a los 45 d.d.s, siendo superiores entre 15% - 50% (Figura
5B).
Biomasa seca parte aérea y raíces. En este parámetro el tratamiento T6 presentó
diferencias estadísticamente significativas (p < 0.05) a los 15, 30 y 45 d.d.s con una
acumulación de biomasa de 30% a 50% mayor a los demás tratamientos en todos los
muestreos (Figura 5C y 5D).
Figura 5. ± Desviación estándar - Letras diferentes representan diferencias estadísticamente significativas en relación
a cada variable evaluada de acuerdo al test de Tukey HSD (p < 0.05). Cada gráfica representa variables de peso y
longitud en función del tiempo (15, 30 y 45 d.d.s). 5A: Longitud Parte aérea; 5B: Longitud de raíz; 5C: Peso seco
aérea; 5D: Peso seco raíz.
a
b b a ab
ab ab
ab ab
b
a a a
a a a a a
a
ab
abcd abc
abcd
bcd
cd
d
d
a
e e
cde bcd
de
ab ab
abc bc
a a
cd
cd
a a ab
d
a
d
a
a
cd
ab
abc abc
bcd
bc
b b
f
d
f
c
a
bc
a
bc
b bc
cd
b b
e
g
bc bc
d
cd
bc
c
b
b
a
a
de
de
ef f
a
b bc bcd
cde
a a
a
bc
bc
ab
de
cd e e
e de
bc
de de
cd
a
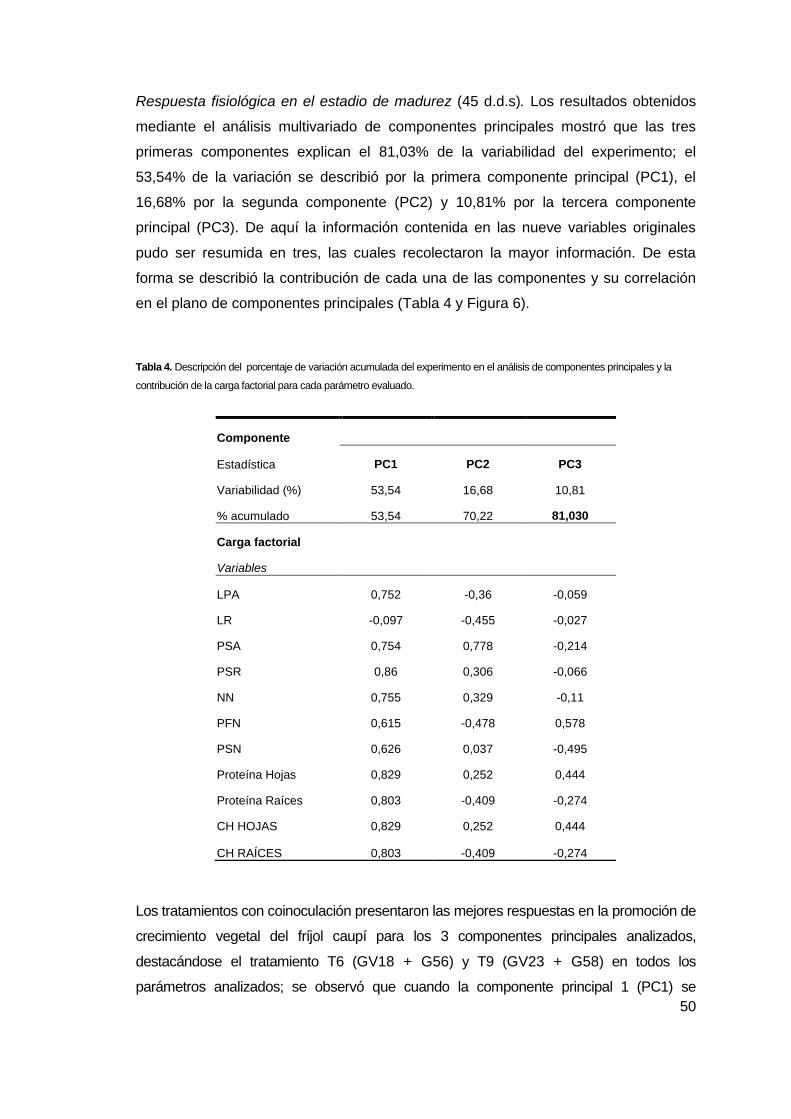
50
Respuesta fisiológica en el estadio de madurez (45 d.d.s). Los resultados obtenidos
mediante el análisis multivariado de componentes principales mostró que las tres
primeras componentes explican el 81,03% de la variabilidad del experimento; el
53,54% de la variación se describió por la primera componente principal (PC1), el
16,68% por la segunda componente (PC2) y 10,81% por la tercera componente
principal (PC3). De aquí la información contenida en las nueve variables originales
pudo ser resumida en tres, las cuales recolectaron la mayor información. De esta
forma se describió la contribución de cada una de las componentes y su correlación
en el plano de componentes principales (Tabla 4 y Figura 6).
Tabla 4. Descripción del porcentaje de variación acumulada del experimento en el análisis de componentes principales y la
contribución de la carga factorial para cada parámetro evaluado.
Componente
Estadística PC1 PC2 PC3
Variabilidad (%) 53,54 16,68 10,81
% acumulado 53,54 70,22 81,030
Carga factorial
Variables
LPA 0,752 -0,36 -0,059
LR -0,097 -0,455 -0,027
PSA 0,754 0,778 -0,214
PSR 0,86 0,306 -0,066
NN 0,755 0,329 -0,11
PFN 0,615 -0,478 0,578
PSN 0,626 0,037 -0,495
Proteína Hojas 0,829 0,252 0,444
Proteína Raíces 0,803 -0,409 -0,274
CH HOJAS 0,829 0,252 0,444
CH RAÍCES 0,803 -0,409 -0,274
Los tratamientos con coinoculación presentaron las mejores respuestas en la promoción de
crecimiento vegetal del fríjol caupí para los 3 componentes principales analizados,
destacándose el tratamiento T6 (GV18 + G56) y T9 (GV23 + G58) en todos los
parámetros analizados; se observó que cuando la componente principal 1 (PC1) se

51
encuentra al lado positivo del eje indica un incremento en casi todos los parámetros de
crecimiento y nutrición vegetal cuando se realizó inoculación mixta con los tratamientos T6
y T9. Para el caso de (PC2) cuando los tratamientos se encuentran al lado positivo del eje
indican un aumento en el contenido de biomasa en la parte aérea, siendo el resultado más
representativo de este parámetro el mostrado por T6 y T9. Con respecto a (PC3), cuando
está en el lado positivo implica que en la planta hay una mayor translocación y acumulación
de asimilados en hojas a partir del área radical, en donde el tratamiento T4 presenta
diferencias significativas en esta componente en comparación con los demás tratamientos
evaluados.
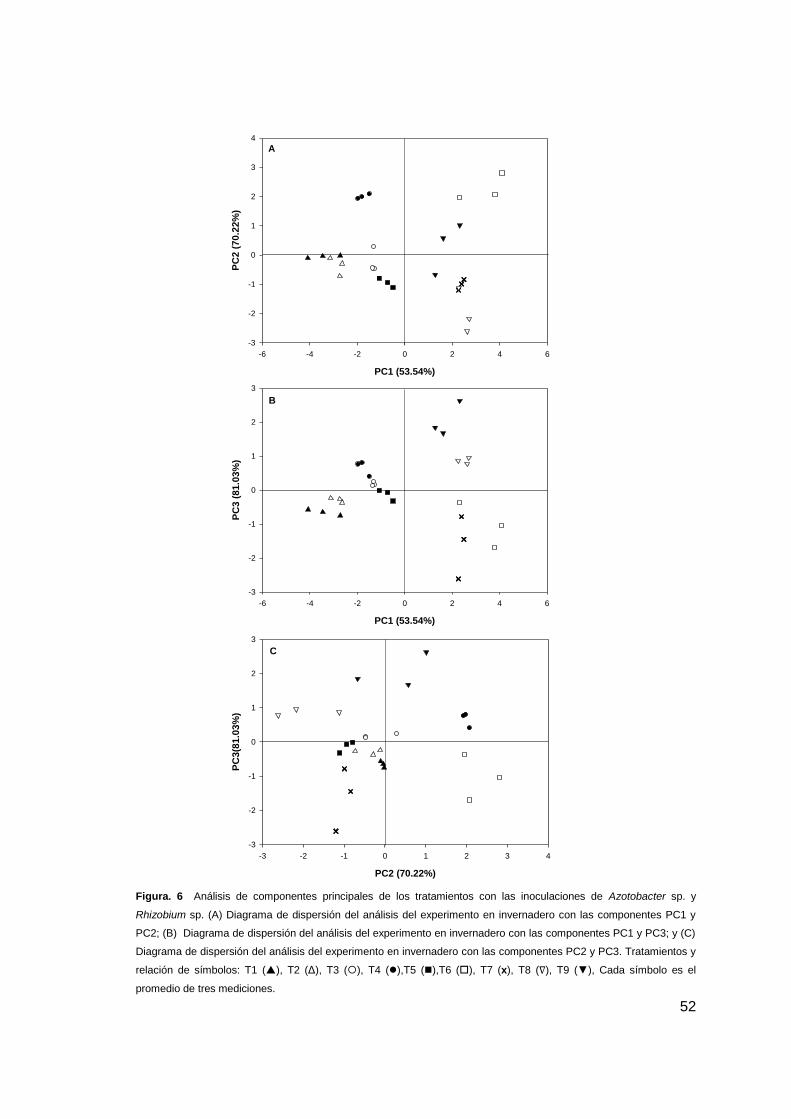
52
Figura. 6 Análisis de componentes principales de los tratamientos con las inoculaciones de Azotobacter sp. y
Rhizobium sp. (A) Diagrama de dispersión del análisis del experimento en invernadero con las componentes PC1 y
PC2; (B) Diagrama de dispersión del análisis del experimento en invernadero con las componentes PC1 y PC3; y (C)
Diagrama de dispersión del análisis del experimento en invernadero con las componentes PC2 y PC3. Tratamientos y
relación de símbolos: T1 (), T2 (Δ), T3 (), T4 (),T5 (),T6 (), T7 (x), T8 (∇), T9 (▼), Cada símbolo es el
promedio de tres mediciones.
PC1 (53.54%)
-6 -4 -2 0 2 4 6
PC
2 (
70.2
2%
)
-3
-2
-1
0
1
2
3
4
A
PC2 (70.22%)
-3 -2 -1 0 1 2 3 4
PC
3(8
1.0
3%
)
-3
-2
-1
0
1
2
3
C
PC1 (53.54%)
-6 -4 -2 0 2 4 6
PC
3 (
81
.03
%)
-3
-2
-1
0
1
2
3
B
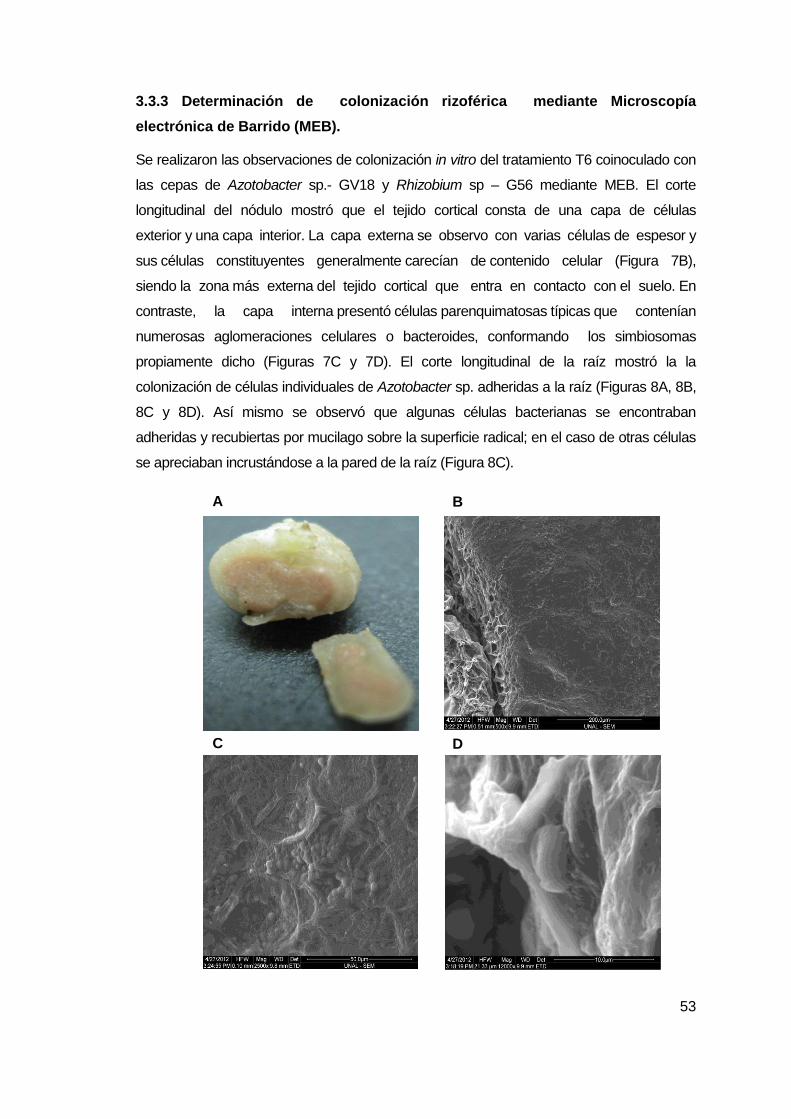
53
3.3.3 Determinación de colonización rizoférica mediante Microscopía
electrónica de Barrido (MEB).
Se realizaron las observaciones de colonización in vitro del tratamiento T6 coinoculado con
las cepas de Azotobacter sp.- GV18 y Rhizobium sp – G56 mediante MEB. El corte
longitudinal del nódulo mostró que el tejido cortical consta de una capa de células
exterior y una capa interior. La capa externa se observo con varias células de espesor y
sus células constituyentes generalmente carecían de contenido celular (Figura 7B),
siendo la zona más externa del tejido cortical que entra en contacto con el suelo. En
contraste, la capa interna presentó células parenquimatosas típicas que contenían
numerosas aglomeraciones celulares o bacteroides, conformando los simbiosomas
propiamente dicho (Figuras 7C y 7D). El corte longitudinal de la raíz mostró la la
colonización de células individuales de Azotobacter sp. adheridas a la raíz (Figuras 8A, 8B,
8C y 8D). Así mismo se observó que algunas células bacterianas se encontraban
adheridas y recubiertas por mucilago sobre la superficie radical; en el caso de otras células
se apreciaban incrustándose a la pared de la raíz (Figura 8C).
A
D C
B
54
Figura. 7. Colonización de las raíces del fríjol caupí por Rhizobium sp. A) Nódulo efectivo utilizado para MEB B)
Ampliación mediante MEB del corte longitudinal del nódulo que muestra su estructura general; observando el tejido ó
“simbiosomas” (derecha) y el tejido de la corteza con células vacías ó membrana peribacteroidal - MPB (izquierda). C)
y D) Acercamiento de la figura B donde se observa el simbiosoma propiamente dicho y la formación de bacteroides.
Figura. 8. Colonización de las raíces del fríjol caupí por Azotobacter sp. A) Raíz con formación de nódulos a la cual se
realizó el corte longitudinal para observación de Azotobacter sp. B) Observación de pelos radicales y con adhesión de
células de Azotobacter C) y D) Acercamiento que muestra las adhesión de las células de Azotobacter a la superficie
de la raíz.
3.4 Discusión
3.4.1. Caracterización molecular de las cepas G56 y G58.
La secuenciación del fragmento mostró que las cepas G56 y G58 se encuentran asociadas
con el género Rhizobium con un 99% de similitud en su identidad. La inferencia filogenética
A
C D
B

55
a partir de las secuencias del fragmento de 16S rDNA confirmó la cercanía con este
género. El análisis filogenético indica que el subgrupo de G56 y G58 presentan similitud
con Rhizobium sp. y Agrobacterium tumefaciens (reclasificada actualmente a R.
radiobacter); revistiendo importancia que de la filogenia del 16S rDNA está claro que
Rhizobium y Agrobacterium están fuertemente relacionadas y sus especies están
entrelazadas (Young et al., 2003).
Sin embargo, para la identificación a nivel de especie de la cepa en estudio Gaunt et al.
(2001) reportaron la necesidad de utilizar diferentes cebadores como recA y atpD que
permitan acercarse aun más a la especie de Rhizobium sp. Así mismo Laguerre et al.
(2001) compararon mediante análisis filogenético la correlación existente entre los genes
nod, nif y la técnica del 16s rDNA para una mejor clasificación de los rizobios encontrando
una mayor resolución para la determinación de especies en este género.
3.4.2 Evaluación en invernadero
Los resultados evidenciaron que los parámetros de longitud y acumulación de
biomasa seca fueron en su mayoría significativamente superiores a los tratamientos
con inoculación mixta e individual (exceptuando la longitud de la planta a los 15
d.d.s). Se destacó el incremento de biomasa seca total en donde T6 (GV18 + G56)
presentó valores superiores entre 30% y 50% mayor a los demás tratamientos en
todas las etapas de crecimiento vegetal. Similares resultados reportaron Groppa et al.
(1998) quienes trabajaron con experimentos en macetas y encontraron incrementos
de 23% en la producción de materia seca debido a la coinoculación en relación con la
inoculación simple, aunque este incremento no fue estadísticamente significativo.
Así mismo, el incremento de la biomasa influenciada en el tratamiento 6 en la etapa
de plántula puede atribuirse en parte a la estimulación del desarrollo diferencial
embrión inducido por reguladores de crecimiento bacteriano que penetran en la
cubierta de la semilla junto con el agua, lo cual acelera el crecimiento de la radícula
con mejoras en la toma de agua y minerales (Cassán et al., 2009). Este concepto es
concordante con lo planteado por Bashan et al. (2004) quienes enfatizan en la
relevancia de la FBN en el para el incremento de la biomasa en las etapas posteriores
del desarrollo una vez que un número significativo de bacterias se han establecido en
los tejidos vegetales. De acuerdo a esta hipótesis, el papel de las bacterias en la
promoción del crecimiento vegetal sería determinante desde las primeras etapas de
desarrollo tales como la germinación y la formación de plántulas.

56
La comunidad bacteriana en el suelo y la rizosfera pueden afectar la nutrición mineral
vegetal, cambios en la absorción de raíces características debido a una modificación
de la morfología de la raíz o alteración de los mecanismos de captación y la tasa de
crecimiento relativo (Tinker, 1984). No obstante, estos beneficios están directamente
relacionados con la adaptación bacteriana y su riqueza en el suelo, es decir, la
concentración celular inicial es crucial en los efectos posteriores.
En el presente estudio se puede atribuir el éxito de las coinoculaciones a su
aplicación temprana, la eficiencia en los mecanismos de promoción de las cepas y la
concentración del inóculo inicial (108 UFC/mL), como lo sugiere Rodelas et al. (1996),
quienes registraron mejoras en los parámetros agronómicos de R. leguminosarum bv.
viceae y V. faba por la inoculación mixta de Rhizobium – Azospirillum brasilense y
efectos benéficos sobre la nodulación y parámetros de crecimiento de esta
leguminosa, los cuales aumentaron en función de la concentración adecuada en el
momento de la inoculación, obteniendo que concentraciones iguales o superiores a
108 UFC/mL garantizan un mayor efecto promotor de crecimiento. Resultados
similares fueron encontrados en estudios realizados con cepas de Azotobacter
chroococcum (cepa H23) y A. vinelandii (cepas ATCC 12837 y Dv42) coinoculados
con R. leguminosarum bv. viceae cepa Z25, en condiciones controladas del cultivo.
Los tratamientos con coinoculación presentaron las mejores respuestas en la promoción de
crecimiento vegetal del fríjol caupí para los 3 componentes principales analizados,
destacándose el tratamiento T6 (GV18 + G56) y T9 (GV23 + G58). En estos tratamientos
se favoreció el incremento en todos los parámetros de crecimiento y nutrición vegetal
[(PC1) y (PC2)], es decir, estos tratamientos influyeron positivamente en desarrollo óptimo
del fríjol caupí en comparación con todos los tratamientos evaluados. Para el caso de
(PC3) el aumento en la acumulación de asimilados en hojas a partir del área radical del
tratamiento con inoculación individual T4 (Inoculación 100% Rhizobium sp.) evidencian que
las inoculaciones incrementan la acumulación de nutrientes para su posterior inversión de
nutrientes en llenado del grano, lo cual representa el estímulo de Rhizobium sp. en función
de mejorar la eficiencia de las plantas en la acumulación de reservas y transporte efectivo
de asimilados hacia los vertederos en posteriores estadios (Borrego et al., 2000).
Sin embargo, la mejora observada por T6 y T9 representan un beneficio integral de la
planta que evidencia que la inoculación mixta confiere al fríjol caupí atributos de
crecimiento, nutrición y rendimiento con respecto a la inoculación individual de Rhizobium
sp.

57
Estos resultados coinciden con los reportados por Rodelas et al. (1996), quienes
encontraron que la inoculación Azotobacter - R. leguminosarum bv. viceae promovieron el
crecimiento de Vicia faba con aumentos significativos de masa seca total y número de
nódulos, obteniendo una rápida tasa de crecimiento y acumulación de nitrógeno total.
Gharib et al. (2009) indicaron que la inoculación mixta Rhizobium - Azotobacter - Bacillus
megaterium var. phosphaticium mejoraron todos los parámetros de crecimiento y
acumulación de nutrientes de naturaleza nitrogenada en comparación con la inoculación de
Rhizobium sp.
El incremento en el peso seco y número de nódulos en los tratamientos T6 y T9 influyeron
positivamente en el contenido de nutrientes (carbohidratos y proteínas totales) y
acumulación de biomasa seca total de acuerdo al análisis multivariado. Varios estudios han
sugerido que hormonas como las auxinas y citoquinas desempeñan funciones esenciales
en el desarrollo de nódulos y su efecto benéfico en la nutrición de leguminosas (Mathesius
et al., 1998). Estudios realizados por Schmidt et al. (1998) demostraron que la
coinoculación de semillas de Medicago sativa con una cepa de R. melilotti ineficaz en la
síntesis de AIA y una cepa eficiente de A. brasilense incrementó significativamente el
número de nódulos en la raíz principal, así como el peso y la cantidad de nitrógeno en
comparación con las plantas inoculadas sólo con Rhizobium sp.
El género Azotobacter ha sido reconocido por su eficiencia en la producción de
fitohormonas y sus efectos se asocian con el crecimiento vegetal, especialmente en la
estimulación de la elongación y aumento de la zona radical. Martínez-Toledo et al. (1991) y
Rodelas et al. (1993) evidenciaron que las bacterias pertenecientes a este género
(específicamente Azotobacter vinelandii ATCC 12837 y Azotobacter chroococcum la cepa
H23) son productores activos de fitohormonas y vitaminas. La síntesis y excreción de los
reguladores de crecimiento en la rizosfera debe explicar la capacidad de estas cepas para
promover el crecimiento y la nodulación de leguminosas, así como los efectos sobre la
nutrición de las plantas evidenciado en otras investigaciones (Okon y Itzigsohn, 1995;
Ravikumar et al., 2004; Yi et al., 2007).
El hecho de que en el presente estudio se trabajó con cepas nativas de Azotobacter sp. y
Rhizobium sp. reviste gran importancia para futuras evaluaciones, ya que se puede
suponer una ventaja en la capacidad de adaptación al suelo, además no presentó acción
antagónica ante Rhizobium sp. ni es factor inhibitorio de la formación de los nódulos, siendo
factores cruciales para colonización exitosa de las raíces y su persistencia en ellas
(Rodelas et al., 2000); lo cual favorecería el crecimiento y rendimiento del fríjol caupí bajo
las condiciones climáticas de la región.

58
3.4.3 Determinación de la colonización rizoférica del mejor coinóculo mediante
Microscopía electrónica de Barrido (MEB).
Los resultados obtenidos mediante microscopía electrónica de barrido permitieron
establecer la presencia de los microorganismos utilizados en la coinoculación del
tratamiento T6. Este estudio confirma las estructuras características del nódulo y la
presencia de rizobios; La morfología y anatomía nodular ha sido previamente descrita
(Cordero y Beltrati, 1989; Sprent y Parsons, 2000; Sprent, 2001).
El corte longitudinal del nódulo mostró la presencia de tejidos corticales sin células
bacterianas y tejidos activos con presencia de simbiosomas, siendo este término
utilizado para el compartimento celular que se forma durante las interacciones
endosimbiontes en el que se toma el microorganismo (rizobio) por la célula huésped y
se convierte completamente envuelto por una membrana derivada del hospedero
(Zahran, 2001).
Así mismo el acercamiento de la figura 8C presentó la formación de estructuras
fibrilares que recubrían los bacteroides inmersos en el sistema peribacteroidal. Se ha
descrito que la formación de microfibrillas extracelulares de celulosa juegan un papel
en la fijación bacteriana a las células vegetales (Matthyssee et al., 1981; Parniske,
2000). La producción de microfibrillas extracelulares por Rizobium radiobacter
conduce a una firme adhesión célula - célula después de su unión inicial a su planta
huésped (Matthyssee et al., 1981). Estas estructuras de celulosa también son
producidas por ciertas Cepas de Rhizobium en cultivo puro (Deinema y Zevenhuizen,
1971; Parniske, 2000) y se han detectado mediante microscopía electrónica
microfibrillas extracelulares asociadas con R. trifolii, las cuales se observaron unidas a
pelos radicales del trébol (Zahran, 1999).
De igual forma, el corte longitudinal de las raíces del fríjol caupí evidenció la
presencia de algunas bacterias individuales de Azotobacter sp. adheridas a la
superficie de las raíces y envueltas en microfibrillas (Figura 8C). Varios estudios han
sido reportados hacia la utilización de microscopias de alta gama en microorganismos
diazotróficos (Berg et al., 1979; Kabir et al., 1995; Bianucci et al., 2011) para la visualización
de estructuras representativas y colonización de la planta. Del mismo modo autores como
Puente et al. (1999) han empleado microscopía electrónica de barrido con el fin de
identificar Azospirillum halopraeferans y Azospirillum brasilense en la superficies de las
raíces de plantas y determinar su contribución al proceso de colonización de las raíces
mediante la formación de microfibrillas.

59
Se ha demostrado que las bacterias promotoras de crecimiento vegetal producen material
fibrilar y no es producido por las células vegetales como se planteó inicialmente. Este
hecho se soporta en por observaciones realizadas mediante MEB de colonias puras de
Azospirillum, que presentaban similares estructuras fibrilares en su superficie (Vanbleu y
Vanderleyden, 2003). La red de microfibrillas permite a las bacterias anclar a las raíces
para un mejor acceso a los exudados de plantas, y recíprocamente permite a la planta
llegar a las sustancias excretadas por las bacterias antes de su consumo por otras
bacterias. Además, proporciona resistencia contra fuerzas externas físicas aplicadas a la
raíz, como el lavado y la agitación (Bashán et al., 1991).
Las características descritas demostraron el establecimiento de los microorganismos
acorde con su adaptación a la planta hospedera (Formación de nódulos y/o adhesión a la
raíz), de acuerdo diversos factores como la disponibilidad y competencia de nutrientes
proporcionados por la planta huésped, interacción y adecuada señalización
microorganismo – planta, entre otros (Bashan et al., 2004); siendo componentes que
juegan un papel importante en el establecimiento y asociación de Rhizobium sp. y
Azotobacter sp. con su anfitrión.
Conclusiones
A partir de los resultados encontrados en esta investigación se permite concluir que
existe un efecto de estimulación del crecimiento del fríjol caupí (Vigna unguiculata (L.)
Walp) por la aplicación de cepas nativas Azotobacter sp. junto con Rhizobium sp.
(inoculación mixta GV18 + G56 y GV23 + G58), promoviendo incremento en peso,
biomasa seca, aumentos de la concentración de proteínas y carbohidratos, mayor
peso de nódulos y número de nódulos; así como la eficiente translocación de
asimilados hacia las estructuras foliares presentes en el proceso del llenado posterior
del fruto.

60
CONCLUSIONES GENERALES DE LA INVESTIGACIÓN
Las cepas GV18 y GV23 de Azotobacter sp. promovieron capacidades de crecimiento
vegetal del fríjol caupí (Vigna unguiculata (L.) Walp), incrementando el aumento en el
tamaño de la planta (parte aérea y raíz), contenido de clorofila y aumento en la
translocación de asimilados hacia las estructuras foliares presentes en el proceso del
llenado posterior del fruto (grano).
Azotobacter sp. es eficiente en la promoción del crecimiento de esta especie en ausencia
de bacterias nodulantes del género Rhizobium en condiciones semicontroladas de
invernadero, lo cual permite inferir que podría utilizarse en coinóculos mixtos sin afectar
parámetros de rendimiento agronómico.
Se evidencia un efecto de estimulación del crecimiento del fríjol caupí (Vigna unguiculata
(L.) Walp) por la aplicación de Azotobacter sp. junto con Rhizobium sp. (inoculación mixta
GV18 + G56 (T6) y GV23 + G58 (T9)), promoviendo incremento en peso, biomasa seca,
aumentos de la concentración de proteínas y carbohidratos, mayor peso de nódulos y
número de nódulos; así como la eficiente translocación de asimilados hacia las estructuras
foliares presentes en el proceso del llenado posterior del fruto.
La coinoculación de cepas nativas de Azotobacter sp. - Rhizobium sp. se constituye como
un estudio inicial que permite avisorar la implementación de tecnologías limpias que
contribuyan con el mejoramiento de la calidad del fríjol caupí (Vigna unguiculata (L.) Walp)
en la microregión del Valle del Cesar.

61
RECOMENDACIONES
Determinar molecularmente a nivel de especie de los microorganismos utilizados en el
presente estudio.
Realizar ensayos con las mejores coinoculaciones en condiciones de campo.
Evaluar las coinoculaciones en condiciones de estrés por sequía, el cual predomina en el
área de estudio.
Evaluar futuros prospectos de inoculantes en otras zonas del país con el propósito de
evaluar el potencial de los coinóculos y su adaptación en diferentes condiciones
edafoclimáticas.

62
BIBLIOGRAFÍA
Anandham, R., Indiragandhi, P., Madhaiyan, M., Kim, K., Yim, W., Saravanan, V. S., Hung,
J., Sa. T. (2007). Thiosulfate oxidation and mixotrophic growth of ethylobacterium oryzae.
Canadian Journal of Microbiology. 53:869–876. CrossRef.
Angelini, J., Ibáñez, F., Taurian, T., Tonelli, M.L., Valetti, L., Fabra, A. (2011). A Study on the
Prevalence of Bacteria that Occupy Nodules within Single Peanut Plants. Curr Microbiol
62:1752–1759.
Arshad, M., Frankenberger, W.T. (1991). Microbial production of plant hormones. Plant and
Soil 133:1-8.
Baca, B., Soto, L., Pardo, M. (2000). Fijación biológica de nitrógeno. Elementos. 38:
39-49.
Bachhawaat, G. (1987). Iron transport in Azospirillum brasilense: role of the siderophore
spirilobactin. J Gen Microbiol 133:1759–1765. DOI:10.1099/00221287-133-7-1759.
Balandreau, J. (1986). Ecological factors and adaptive processes in N2-fixing bacterial
populations of the plant environment. Plant and Soil. Vol 90. Pp 73.
Bashan, Y., Holguín, G., Ferrera-Cerrato, R. (1996). Interacciones entre plantas y
microorganismos benéficos. II. Bacterias asociativas de la rizosfera. Terra. 14: 195-210.
Bashan, Y., Mitiku, G., Whitmoyer, R.E., Levanony, H. (1991). Evidence that fibrillar
anchoring is essential for A. brasilense Cd attachment to sand. Plant Soil. 132, 73–83.
Bashan., Y. Holguin., G. de Bashan., L. (2004). Azospirillum–plant relationships:
physiological, molecular, agricultural, and environmental advances, Can. J. Microbiol. 50:
521–577.
Benintende, S., Uhrich, M., Herrera, F., Gangge, M., Sterren M., Benintende, M. (2010).
Comparación entre coinoculación con Bradyrhizobium japonicum y Azospirillum brasilense
e inoculación simple con Bradyrhizobium japonicum en la nodulación, crecimiento y
acumulación de N en el cultivo de soja. Agriscientia, 27 (2): 71-77.
Berg, R.H., Vasil, V. and Vasil, I.K. (1979). The biology of Azospirillum-sugarcane
association. II. Ultrastructure. Protoplasma 101, 143-163.
Bertani, G. (1951). Studies on lysogenesis. I. The mode of phage liberation by lysogenic
Escherichia coli. J. Bacteriol. 62:293-300.

63
Bianucci, E., Fabra, A., Castro, S. (2011). Involvement of glutathione and enzymatic
defense system against cadmium toxicity in Bradyrhizobium sp. Strains (peanut
symbionts). Biometals. DOI 10.1007/s10534-011-9480-z.
Bishop, P., Joerger, R. (1990). Genetics and molecular biology of alternative nitrogenase
systems. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41:109-125.
Bonilla, R., Novo, R., Vanegas, N., Galvis, A., Martínez, M., Parra, D., Vanegas, O.
(2000). Generación de tecnologías para la utilización de la fijación no simbiótica de
nitrógeno como alternativa de fertilización.
Bonilla, R., Murillo, J., Gnecco, M., Valero, L., Aaron, M., Orozco G. (2002).
Mejoramiento y conservación de suelos algodoneros: Técnicas de labranza e
Incorporación de abonos verdes. En: Colombia Innovación y Cambio Tecnológico -
Corpoica ISSN: 1657-0901 ed: Productos Editoriales y Audiovisuales-Produmedios
v.2 fasc. p.53 – 59.
Bonilla, R., Morales, G. (2007). Monibac biofertilizante con base en cepas nativas de
Azobacter sp. para incrementar la productividad y sostenibilidad del algodonero
Colombia, Innovación y Cambio Tecnológico – Corpoica. ISSN: 1657-0901, vol:4 fasc:
págs: 30 – 34.
Borrego, F., FernandeZ, A., Lopez, V., Parga, M., Carvajal, A. (2000). Análisis de
crecimiento en siete variedades de papa (Solanum tuberosum L.). Agronomía
Mesoamericana 11(1), 145-149.
Bowen, G.D., Rovira, A.D. (1999). The Rhizosphere and its management to improve
plant growth. Adv. Agron. 66: 1-102.
Bradford, M. (1976). Anal. Biochem. 12,248-254.
Brewin, N.J. (2004). Plant cell wall remodelling in the rhizobium legume symbiosis. Crit
Rev Plant Sci. 23: 293–316.
Brockwell, J. (1982). Inoculation methods for field experimenters and farmers. In
Nitrogen Fixation in Legumes (J. M. Vincent, Ed.), pp. 211-227. Academic Press,
Sydney.
Bürgmann, H., Widmer, F., Von Sigler, W., Zeyer, J. (2004). New molecular screening
tools for analysis of free-living diazotrophs in soil. Appl Environ Microbiol 70: 240–247.
Burns, R. C., Hardy, R. W. F. (1975). Nitrogen Fixation in Bacteria and Higher
Plants. New York: Springer Verlag.

64
Burns, T., Bishop, P. Israel, D. (1981). Enhanced nodulation of leguminous plant roots
by mixed cultures of Azotobacter vinelandii and Rhizobium. Plant and Soil. 62, 399-
412.
Calegari, A. 1995. Leguminosa para Adubacâo verde De verâo no Paraná. Circular
No. 80 MAIO/95 IAPAR. Instituto Agronômico Do Paraná-Londrina-PR.
Carlson, R., Price, N., Stacey, G. (1994). The biosynthesis of Rhizobial lipo-
oligosaccharide nodulation signal molecules. Mol Plant Microbe Interact. Vol. 7 6: 684-
695.
Carreño, R., Campos-Reales, N., Elmerich, C., BACA, B. (2000). Physiological
evidence for differently regulated tryptophan-dependent pathways for indole-3- acetic
acid synthesis in Azospirillum brasilense. Mol Gen Genet., 264 (4): 521–530.
Carvalho, M., Sgarbieri, V. (1998). Relative importance of phytohemagglutinin (lectin)
and trypsin-chymotrypsin inhibitor on bean (Phaseolus vulgaris L) protein absorption
and utilization by the rat. J. Nutr. Sci. Vitaminol. 44(5): 685-696.
Cassán, F., Bottini, R., Schneider, G., Piccoli, P. (2001). Azospirillum brasilense and
Azospirirllum lipoferum hidrolize conjugates of GA20 and metabolize the resultant
aglycones to GA1. in seedlings of rice dwarf mutants. Plant Physiol. 125: 2053-2058.
Cassán, F., Perrig, D., Sgroy, V., Masciarelli, O., Penna, C., Luna, V. (2009).
Azospirillum brasilense Az39 and Bradyrhizobium japonicum E109, incoulated singly
or in combination, promote seed germination and early seedling growth in corn (Zea
mays L.) and soybean (Glycine max L.). European Journal of soil Biology 45: 28-35.
Castillo, J., Casas, J., Martinez, M. (2005). Evaluación de la degradación de
endosulfan por Azotobacter chroococcum y determinación del efecto plaguicida sobre
la fijación biológica de nitrógeno y sobre la producción de auxinas. Tesis de
Microbiología Agrícola y Veterinaria. Pontificia Universidad Javeriana. pp 13, 14.
Castro-Diez, P. (2002). Factores que limitan el crecimiento de la vegetación leñosa
mediterránea. Respuestas de las plantas: de organo a comunidad. pp 47-85 en: J.
Charco (editor). La regeneración natural del bosque mediterráneo en la península
ibérica. ARBA-Ministerio de Medio Ambiente, Madrid, España.
Chaiharn, M., Chunhaleuchanon, S., Lumyong, S. (2009). Screening siderophore
producing bacteria as potential biological control agent for fungal rice pathogens in
Thailand. World J Microbiol Biotechnol 25(11):1919–1928. doi:10.1007/s11274-009-
0090-7.

65
Chianu, J.N., Nkonya, E.M., Mairura, F.S., Chianu, J.N., Akinnifesi, F.K. (2011).
Biological nitrogen fixation and socioeconomic factors for legume production in sub-
Saharan Africa: a review. Agron. Sustain. Dev. 31:139–154.
Corporación Colombiana de Investigación Agropecuaria. (2004). Primer informe del
Convenio Colciencias –Fedegan-Corpoica. Proyecto “Estrategias de innovación
tecnológica para mejorar la competitividad y sostenibilidad de los sistemas de
producción de carne y leche bovina en el Valle del Sinú y Sabanas de Bolívar,
Córdoba y Sucre”. 17 de marzo 2004. 26 pp.
Cho, J., Lee, D. Jeon, J. Kim, J., Han. H. (2006). Microbial population dynamics of
kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 257, 262-267.
Cordero, L., Beltrati., C.M. (1989). Estrutura e desenvolvimento de nódulos
radiculares de Anadenanthera falcata. Speg. Revista Brasileira de Botânica.12:61-70.
Cornelis, P., Matthisj, S. (2007). Pseudomonas siderophores and their biological
significance. In: Ajit V, Chincholkar SB (eds) Microbial siderophores (Soil Biology).
Heidelberg, Berlin, Germany, pp 193–200.
Crowley, D.E., Wang, Y.C., Reid C.P.P., Szaniszlo, P.J. (1991). Mechanisms of iron
acquisition from siderophores by microorganisms and plants. Plant Soil 130: 179-198.
Crowley, D. (2006). Microbial siderophores in the plant rhizospheric. In: Barton LL,
Abadía J (eds) Iron nutrition in plants and rhizospheric microorganisms. Springer,
Netherlands, pp 169–198.
Cruz, N., Ortiz, F., Sanchez, Y., Mendoza, M. (2005). Biomasa e índices fisiológicos
en chile morrón cultivado en altas densidades. Rev. Fitotec. Mex. 28(3), 287-293.
Cuesta, M., Mila, P. (2003). Manejo y productividad de praderas renovadas en el
Trópico Alto. Fundamentos teóricos y metodología de evaluación. En: Renovación y
manejo de praderas y utilización de ensilajes en el Trópico Alto. Resultados finales
Guachucal y Buesaco. 17 y 19 de julio de 2003.Plan de Modernización Tecnológica
de la Ganadería Bovina Colombiana. pp 29-35.
Cuesta-Muñoz, P. (2005). Fundamentos de manejo de praderas para mejorar la
productividad de la ganadería del trópico colombiano. En: Colombia Revista Corpoica
ISSN: 0122-8706 ed: Produmedios v.6 fasc.2 pp.5-13.
Dardanelli, M., Rodríguez, D., Megías, M., Okon, Y. (2008). Influencia de la
coinoculación Azospirillum-rizobios sobre el crecimiento y la fijación de nitrógeno de
leguminosas de interés agronómico. In: Cassán, F., García Salamone, I. (Eds.),

66
Azospirillum sp.: Cell Physiology, Plant Interactions and Agronomic Research in
Argentina. Asociación Argentina de Microbiología, Buenos Aires, Argentina, pp. 141–
151.
Davis, D.W., Oelke, E.A., Oplinger, E.S., Doll, J.D., Hanson, C.V., Putnam, D.H.
(1991). Alternative Field Crops Manual. University of Wisconsin, pp. 429.
Deinema, M., Zevenhuizen, L. P. (1971). Formation of cellulose fibrils by Gram
negative bacteria and their role in bacterial flocculation. Arch. Mikrobiol. 78:42-57.
Del Gallo, M., Fabbr,. P. (1990). Inoculation of Azospirillum brasilense Cd on chick
pea (Cicer arietinum). Symbiosis 9: 283-287.
Departamento Nacional de Planeación (DNP). República de Colombia. (2010).
Anuario Estadístico del Sector Agropecuario-Min Agricultura. url:
http://www.dnp.gov.co/ PortalWeb/Portals/0/ archivos / documentos/ddrs/
Indicadores/Tabla%201.pdf, Agosto 2011.
D'Haeze, W., Holsters, M. (2002). Nod factor structures, responses, and perception
during initiation of nodule development. Glycobiology. 12: 79R–105R.
Döbereiner, J., Baldani, V., Baldani J. (1995). Como isolar e identificar bactérias
diazotróficas de plantas ñao leguminosas. Brazil, EMBRAPA-SPI.
Dubois, M., Gilles, K., Hamilton, J., Rebers, P., Smith, F. (1951). A colorimetric
method for the determination of sugars, Nature 168-167.
Eckert, B., Baller, O., Kirchhof, G., Halbritter, A., Stoffels, M., Hartmann, A. (2001).
Azospirillum doebereinerae sp. nov., a nitrogen-fixing bacterium associated with the
C4-grass Miscanthus. International Journal of Systematic and Evolutionary
Mcrobiology, 51: 17-26.
Ehlers, J.D., Hall, A.E. (1997). Cowpea (Vigna unguiculata L. Walp.) Field Crops
Research 53:187-204.
El-Seifi, S.K., Sarg, S.M.H., Abdel-Fattah, A.L., Mohamed, M.A. (2004). Effect of
biofertilizers and nitrogen levels on the productivity and quality of Chinese garlic under
sandy soil conditions. Zagazig J. Agric. Res. 31(3).
Elowad, H.O.A., Hall, A.E. (1987). Influences of early and late nitrogen fertilization on
yield and nitrogen fixation of cowpea under well-watered and dry field conditions. Field
Crops Res., 15: 229-244.

67
Ewing, B., Green, P. (1998). BaseCalling of Automated Sequencer Traces Using
Phred. II. Error Probabilities. Genome Research 8: 186-194.
Ewing, B., Hillier, L., Wendl, M.C. and Green, P. (1998). BaseCalling of Automated
Sequencer Traces Using Phred. I. Accuracy Assessment. Genome Research 8: 175-
185.
Espin, G. (2002). Biología de Azotobacter vinelandii. Instituto de Biotecnología,
Universidad Nacional Autónoma de México. En:
www.microbiologia.org.mx/microbiosenlinea/CAPITULO_09/Capitulo09.pdf Consulta:
3 de Febrero de 2012.
Evans, D., Jones, R., Woodley, P., Robson, R. (1988). Further analysis of nitrogen
fixation genes in Azotobacter chroococcum: identification and expression in Klebsiella
pneumoniae of nifS, nifV, nifM, and nifB genes and localization of nifEIN, nifU-, nifA-
and fixABC-like genes. J. Gen. Microbiol. 134:931-942.
Fedegan-Sena. (2010). Parcelas demostrativas con materiales forrajeros importados
de Brasil: Región Caribe. ISBN: 978-958-8498-11-9 pp. 1-36 Consultado el 26 de
Marzo de 2012 en:
http://portal.fedegan.org.co/pls/portal/docs/PAGE/FNG_PORTLETS/PUBLICACIONE
S/LIBROS_CARTILLAS/CARTILLAS/CARIBE_SINCUB.PDF.
Fedegan. (2010). La Ganadería Colombiana y las Cadenas Láctea y Cárnica. Plan
Estratégico de la Ganadería Colombiana PEGA 2019. Consultado el 24 de Marzo de
2012 en:
http://www.campoestrategico.com/Media/Default/BlogPost/blog/CIFRASDEREFEREN
CIASECTORGANADEROENCOLOMBIA.pdf.
Fenglerowa, W. (1965). Simple method for counting Azotobacter in soil samples. Acta
Microbiologica Polonica, 14 (2): 203–206.
Fernández C, M. 2003. Manual de nodulación. Editorial Hemisferio Sur, Buenos Aires,
Argentina. 53 p. www.nitragin.com.ar
Fiske, C., Subbarow, Y. (1925) The colorimetric determination of phosphorus. J Biol
Chem 66:26.
Freiberg, C., Fellay, R., Bairoch, A., Broughton, W.J., Rosenthal, A. Perret. X. (1997).
Molecular basis of symbiosis between Rhizobium and legumes. Nature 387:394–401.
Frink, C.R., Waggoner, P.E. y Ausubel, J.H. (1999). Nitrogen fertilizer: retrospect and
prospect. Proc. Natl. Acad. Sci. USA 96: 1175-1180.

68
Garzón, S., Lamprea, S., Martinez, M. (2001). Desarrollo de una preparación liquida
de Azotobacter utilizando como medio de cultivo los subproductos de la industria
sucroquímica. Tesis de Microbiología Industrial. Pontificia Universidad Javeriana. Pp
25.
Gaunt M.W., Turner, S.L., Rigottier-Gois, L., Lloyd-Macgilps, S.A. and Young, J.P.W.
(2001). Phylogenies of atpD and recA support the small subunit rRNA-based
classification of rhizobia. International Journal of Systematic and Evolutionary
Microbiology, 51, 2037–2048.
Gharib, A., Shahen., M. Ragab, A. (2009). Influence of Rhizobium inoculation
combined with Azotobacter chrococcum and Bacillus megaterium var phosphaticum
on growth, nodulation, yield and quality of two snap been (Phasealus vulgaris L.)
Cultivars. 4 th Conference on Recent Technologies in Agriculture. pp1-15.
Giles, E., Oldroy, G., Downie, A. (2008). Coordinating nodule morphogenesis with
rhizobial infection in legumes. The Annual Review of Plant Biology, 59: 519-546.
Glick, B., Pattern, C., Holguin, G., Penrose, D. (1999). Biochemical and genetic
mechanisms used by plant growth promoting bacteria. Imperial College Press,
London.
González, J., Salmerón, V., Martínez, M., Ballesteros, F., RAMOS, A. (1986).
Production of auxins gibberelins and citokinins by Azotobacter vinelandii ATCC 12837
in chemically defined media and dialysed soil media. Soli Biol. Bioch. 18. p. 119-120.
González, N., Perticari, A., Stegman, B., Rodríguez, E. (1997). Nutrición nitrogenada.
En: Giorda L.M.y Baigorri, H.E.J. (Eds.). El cultivo de la soja en Argentina. INTA,
Centro Regional Córdoba. EEA Marcos Juárez- EEA. p. 188- 198.
González, N. (2002). Algunos elementos de juicio para interpretar el fenómeno de la
nodulación en soja. Publicación de las Jornadas de Cosecha Gruesa. INTA CIAM,
Mar del Plata. 4 p.
Gordon, D., Abajian, C., Green, P. (1998). Consed: a graphical tool forsequence
finishing. Genome Research 8: 195-202.
Gutierrez, A., Martinez, E. (2001). Natural endophytic association between Rhizobium
ettli and maize (Zea mays L.). Journal of Biotechnology. 91: 117-126.

69
Groppa, M. D., Zawoznik, M. S., Tomaro, M.L. (1998). Effect of Co-inoculation with
Bradyrhizobium japonicum and Azospirillum brasilense on soybean plants. Soil Biol.
34(2): 75-80.
Herridge, D.F., Peoples, M.B., Boddey, R.M. (2008). Global inputs of biological
nitrogen fixation in agricultural systems. Plant Soil 311, 1-18.
Hiscox, J.D., Israelstam, G.F.(1979). A method for the extraction of chlorophyll from
leaf tissue without maceration. Can. J. Bot. 57, 1332–1334.
Hoagland, D.R., Arnon, D.I. (1950). A water culture method for growing plants without
soil. California Agricultural Experimental Station Circular 347.
Holt, J., Krieg, N., (1994). Enrichment and isolation. In: Methods for General and
Molecular Biology . P. Gerhardt, R.G.E. Murray, W.A. Wood y N.R. Krieg (eds.).
American Society for Microbiology. Washington, DC. p. 179-215.
Holt, J., Krieg, N., Sneath, P., Staley, J., Staley, W. (2000). Manual of determinative
Bacteriology. Ninth edition. Lippincott Williams and Wilkins. Baltimore. p. 787.
Huang, X., Madan, A. (1999). Cap 3: a DNA sequence assembly program. Genome
Res 9: 868-877.
Hungría, M., Andrade, D.S., Chueirea, L.M., Probanza, A., Gutierrez-Mañero, F.J.,
Megías, M., 2000. Isolation and characterization of new efficient and competitive bean
(Phaseolus vulgaris L.) rhizobia from Brazil. Soil Biology & Biochemistry.32: 1515–
1528.
Hunt, R. (1990). Basic growth análisis. Academic Press,London.
Jauhri, K.S., Bhatnagar, R.S., Iswaran, V. (1979). Associative effect of inoculation of
different strains of Azotobacter and homologous Rhizobium on the yield of moong
(Vigna radiata), soybean (Glycine max) and pea (Pisum sativum). Plant and soil. 53
(1),pp105-108.
Kabir, M.M., Faure, D., Haurat, J., Normand, P., Jacoud, C., Wadoux, P., Bally, R.
(1995). Oligonucleotide probes based on 16S rRNA sequences for the identification of
four Azospirillum species. Can. J. Microbiol. 41, 1081-1087.
Kimura, M. (1980). A simple method for estimating evolutionary rates of base
substitutions through comparative studies of nucleotide sequences. J Mol Evol.
16(2):111-20.

70
Kole, M., Page, J., Altosaar, I. (1988). Distribution of Azotobacter in Eastern Canadian
soils and in association with plant rhizospheres. Can. J. Microbiol. 34, 815-817.
Kremer, R.J., Peterson, H.L. (1983). Effects of carrier and temperature on survival of
Rhizobium spp. in legume inocula: development of an improved type of inoculant.
Applied and Environmental Microbiology. 45:1790-1794.
Kuykendall, D., Young, J., Martínez, E., Kerr, A., Sawada, H. (2005). Rhizobium.
Frank, 1889, 338. Bergey`s Manual os Systematic bacteriology. Editorial Springer
US. p. 325-340.
Laguerre, G., Nour, S.M., Macheret, V., Sanjuan, J., Drouin, P. and Amarger, N.
(2001). Classification of rhizobia based on nodC and nifH gene analysis reveals a
close phylogenetic relationship among Phaseolus vulgarissymbionts. Microbiology
147, 981–993.
Lewis, B., Schrire, B., Mackinder, B., Lock, M. (2005). Legumes of the world. Kew
Publishing, Surrey.
Lindström, K., Martínez-Romero, E. (2007). International committee on systematics of
prokaryotes subcommittee on the taxonomy of Agrobacterium and Rhizobium:
minutes of the meeting, 26 July 2006, Toulouse, France. Int J of Systematic and
Evolutionary Microbiology. Vol 57: 1365-1366. DOI 10.1099/ijs.0.63744-0.
Lu Bing-Lin, L., Wan, W., Li, J., Guo T.(2009). Nitrogen fixation ability of Azotobacter and its
effect on growth of spring wheat. Chinese Journal of Eco-Agriculture. CNKI: SUN:
ZGTN.0.2009-05-018. S512.1; S144.
Luria, S. E., Burrous, J. W. (1957). Hybridization between Escherichia coli and Shigella. J.
Bacteriol. 74:461-476.
Mahecha, L. (2002). El silvopastoreo: Una alternative de producción que disminuye el
impacto ambiental de la ganadería bovina. Rev. Col. Cienc. Pec. 15(2). 226-231.
Mannion, A.M. (1998). Future trends in agriculture: the role of biotechnology. Outlook
on Agriculture 27, 219-224.
Marín, V.A., Baldani, V.L., Dos Santos, K.R. y Baldani, J.I. (1999). Fixação biológica
de nitrogênio: bactérias fixadoras de nitrogênio de importância para a agricultura
tropical. En: www.cnpab.embrapa.br
Martínez-Toledo, M., González-López, J., de la Rubia, T., Moreno, J., Ramos-
Cormenzana, A. (1988). Effect of inoculation with Azotobacter chroococcum on

71
nitrogenase activity of Zea mays roots grown in agricultural soils under aseptic and
non-sterile conditions. Biology and Fertility of Soils 6, 170-176.
Mascarua-Esparza, M., Villa-González, R., Caballero-Mellado, J. (1988). Acetylene
reduction and indoleacetic acid production by Azospirillum isolates from Cactaceous
plants. Pant and Soil, 106: 91-95.
Masson-Boivin, C., Giraud, E., Perret, X., Batut, J. (2009). Establishing nitrogen-fixing
symbiosis with legumes: how many rhizobium recipes? .Trends Microbiol 17:458–466.
Mathesius, U., Schlaman, H., Spaink, H., Sautter, C., Rolfe, B., Djordjevic, M. (1998).
Auxin transport inhibition precedes root nodule formation in white clover roots and is
regulated by flavonoids and derivatives of chitin oligosaccharides, Plant J.14: 23–34.
Matthyssee, A.G., Holmes, K. V., Gurlitz, R. H. (1981). Elaboration of cellulose fibrils
by Agrobacterium tumefaciens during attachment to carrot cells. J. Bacteriol. 145:583-
595.
Mehnaz, S., Lazarovits, G. (2006). Inoculation effects of Pseudomonas putida,
Gluconacetobacter azotocaptans, and Azospirillum lipoferum on corn plant growth
under greenhouse conditions. Microbial Ecology, 51: 326-335.
Menna, P., Hungria, M., Barcellos, F.G., Bangel. E.V., Hess. P.N., Martines-Romero,
E. (2006). Molecular phylogeny base don the 16S rRNA gene of eleite rhizobial
starins used in Brazilian comercial inoculants. Systematic and Applied Microbiology,
v.29, p. 315-322.
Molla, A., Shamsuddin, Z., Halimi, M., Morziah, M., Puteh, A.( 2001). Potential for
enhancement of root growth and nodulation of soybean coinoculated with Azospirillum and
Bradyrhizobium in laboratory systems, Soil. Biol. Biochem.33: 45-463.
Novo, R. (1993). Microbiología del suelo y biofertilización. En: Memorias de la
Fundación de Asesorías para el Sector Rural (Fundases). Santa Fe de Bogotá. p.
101.
Obando, M., Burgos, L., Rivera, D., Garrido, M., Baldani, V.L., Bonilla, R. (2010).
Caracterización de bacterias diazotróficas asimbióticas asociadas al Eucalipto
(Eucalyptus sp.) en Codazzi, Cesar. Acta Biológica Colombiana. 15(3):105-120.
Okon, Y., Itzigsohn, R. (1995).The development of Azospirillum as a commercial
inoculant for improving crop yields. Biotechnology Advances. (13): 365-374.

72
Oldroyd, G., Harrison, M., Udvardi, M. (2005). Peace talks and trade deals. Keys to
long-term harmony in legume-microbe symbioses. Plant Physiology, 137: 1205-1210.
Osborne, C.A., Galic, M., Sangwan, P., Janssen, P.H. (2005). PCR-generated artefact
from 16S rRNA gene-specific primers. FEMS Microbiology Letters, v.248, p.183–187.
Olivares, J., Barea, M. (1995). Microorganismos del suelo en nutrición vegetal:
simbiosis rhizobium-leguminosa y micorrizas VA. Organización de las Naciones
Unidas para la Educación, la Ciencia y la Cultura. Biblioteca digital de la Universidad
de
Chile.http:/mazinger.sisib.uchile.cl/repositorio/lb/ciencias_quimicas_y_farmaceuticas/l
achicam01/frameizq.html.
Parniske, M. (2000). Intracellular accommodation of microbes by plants: a common
developmental program for symbiosis and disease?. Current Opinion in Plant Biology.
3:320–328
Panter, S., Thomson, R., de Bruxelles, G. Laver, D., Trevaskis, B., Udvardi, M. (2000).
Identification with proteomics of novel proteins associated with the peribacteroid
membrane of soybean root nodules. Mol Plant Microbe Interact 13:325–333.
Patten, C., Glick B. (1996). Bacterial biosynthesis of indole-3-acetic acid. Canadian
Journal of Microbiology 42: 207-220.
Pelczar, M., Reid, R., Chan, P. (1993). Microbiología. 4 edición. México: McGraw–Hill.
p. 8-36.
Peoples, M., Herridge, F., Lahda, K. (1995). Biological nitrogen fixation: an efficient
source of nitrogen for sustainable agricultural production? Plant Soil.174:3–28.
Perticari, A., Arias, N., Baigorri, H., De Battista, J., Montecchia, M., Pacheco J.,
Simonella, A., Toresani, S., Ventimiglia, L., Vicente, R. (2003). Inoculación y fijación
biológica de nitrógeno en el cultivo de soja. En: El libro de la soja. Buenos Aires.
Servicios y Marketing Agropecuario, p.69-76.
Perticari, A. (2005). Inoculación de calidad para un máximo aprovechamiento de la
FBN. Congreso Mundo Soja. Buenos Aires. p. 121-126.
Pietrarelli, L., Zamar, J., Leguía, H., Alessandria, E., Sánchez, J., Arborno, M., Luque,
S. (2008). Effects of different management practices on nodulation and on the yield of
soybean crops. Agriscientia, 25(2):81-87.

73
Proexport. (2011). Sector Agroindustrial Colombiano. Volumen I No. I. Enero de 2011.
Consultado el 24 de Marzo de 2012 en: http://www.botschaft-
kolumbien.de/descargas_proexport/berlin_2011/espanol/inversion/agroindustria/perfil_
agroindustria.pdf
Poorter, H., Nagel, O. (2000). The role of biomass allocation in the growth response of
plants to different levels of light, CO2, nutrients and water: a quantitative review.
Australian Journal of Plant Physiology 27: 1191-1191.
Prinyawiwatkul, W., Watters, M., Beuchat, L., Phillips, R. (1996). Cowpea flour: a
potential ingredient in food products. Critical Reviews in Food Science and Nutrition
36:414 -436.
Puente, M.E., Holguin, G., Glick, B.G., Bashan, Y. (1999). Root-surface colonization of
black mangrove seedlings by Azospirillum halopraeferens and Azospirillum brasilense
in seawater. FEMS Microbiology Ecology 29: 283-292.
Radford, P.J. (1967). Growth analysis formulae. Their use and abuse. Crop Sci. 7,
171-175.
Rangel, A., Saraiva, K., Schwengber, P., Narciso, M.S., Domont, G.B., Ferreira, S.T.,
Pedrosa, C. (2004). Biological evaluation of a protein isolate from cowpea (Vigna
unguiculata) seeds. Food Chem ,87: 491-499.
Ramakrishnan, K., Gnanam R., Sivakumar, P. Manickam, A. (2005). In vitro somatic
embryogenesis from cell suspension cultures of cowpea (Vigna unguiculata (L.)
Walp). Plant Cell Rep. 24: 449–461.
Ravikumar, S., Kathiresan, K., Thadedus, M., Lgnatiammal, S., Babu, M., Shanthy, S.
(2004). Nitrogen-fixing Azotobacter from mangrove habitat and their utility as marine
biofertilizers.J.Exp.Mar.Biol.Ecol.312:5-17.
Ravindar, K., Chandra, R. (2008). Influence of PGPR and PSB on Rhizobium
Leguminosarum Bv. Viciae strain competition and symbiotic performance in Lentil.
World Journal of Agricultural Sciences 4 (3): 297-301, 2008.
Richardson, A.E., Hadobas, P.A., Hayes, J.E., O’Hara, C.P., Simpson, R.J. (2001).
Utilization of phosphorus by pasture plants supplied with myo-inositol hexaphosphate
is enhanced by the presence of soil micro-organisms. Plant Soil 229:47-56.
Rodelas, B., González-López, J., Salmerón, V., Pozo, C., Martínez-Toledo, M.V.
(1996). Enhancement of nodulation, N2-fixation and growth of faba bean (Vicia faba L.)

74
by combined inoculation with Rhizobium leguminosarum bv. viceae and
Azospirillumbrasilense. Symbiosis 21:175–186.
Rodelas, B.; Salmerón, V.; Martínez – Toledo, M.V.; Pozo, C. Y González – López, J.
(1999). Interacción Rhizobium/Azospirillum y Rhizobium/Azotobacter. Efecto sobre el
crecimiento, nodulación y fijación simbiótica de N2 en Vicia faba (L.). Grupo de
Microbiología Ambiental, Departamento de Microbiología (Facultad de Farmacia) e
Instituto del Agua, Universidad de Granada. 18071 Granada, España. Abstract
Volume 29 Isue 2 pp 165 – 169.
Rodelas, B., González, J., Pozo, C., Martínez-Toledo, M., Salmerón, V. (2000).
Interacción Rhizobium/ Azospirillum y Rhizobium / Azotobacter. Efecto sobre el
crecimiento, nodulación y fijación simbiótica de N2 en Vicia faba (L.). Grupo de
Microbiología Ambiental Departamento de Microbiología (Facultad de Farmacia) e
Instituto del Agua, Universidad de Granada. 18071 Granada, España.
Rodríguez, A. J., Rojas, M. M., Trujillo, I. D., Manzano, J., Heydrich, M. (2003).
Caracterización de la comunidad microbiana endófita de la caña de azúcar. En:
Memorias. V Taller Internacional sobre Recursos Fitogenéticos FITOGEN 2003.
Instituto de Investigaciones de Pastos y Forrajes, La Habana, Cuba.
Rodriguez, H., Fraga, R., Gonzalez, T., Bashan, Y. (2006). Genetics of phosphate
solubilization and its potencial applications for improving plant growth-promoting
bacteria. Plant Soil 287:15–21. doi:10.1007/s11104-006-9056-9.
Roesch, L., Olivaeres, F., Pereira, L., Selbach, P., Saccol de Sa, E., Oliveira, F.
(2006). Characterization of diazotrophic bacteria associated with maize: effect on
plant genotype, ontogeny and nitrogen-supply. World and J. of Microb and Biotech.
Vol 22. Pp.3818-3829.
Roth, LE., Jeon, K.W., Stacey, G. (1988). Homology in endosymbiotic systems: the
term ‘symbiosome’. In Molecular Genetics of Plant–Microbe Interactions. Edited by
Palacios R, Verma DPS. St. Paul, Minnesota; APS Press; pp:220-225.
Saribay, G. (2003). Growth and nitrogen fixation dynamics of Azotobacter
chroococcum nitrogen-free and own containing medium. Tesis de Maestría en
Ciencias Aplicadas. Departamento de ingeniería de alimentos. The Middle East
Technical University. Turkia. Pp 1-45.
Saitou, N., Nei, M. (1987). The neighbor-joining method: A new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution 4:406-425.

75
Sambrook, J., Fritschi, E.F., Maniatis, T. (1989). Molecular cloning: a
laboratorymanual, Cold Spring Harbor Laboratory Press, New York.
Sambrook, J., Russell, D.W. (2001). Molecular Cloning: A laboratory manual. 3rd ed.
Cold Spring Harbor Laboratory Press, New York, USA, 99p.
Sarig, S., Kapulnik, Y., Okon, Y. (1986). Effect of Azospirillum inoculation on nitrogen
fixation and growth of several winter legumes. Plant Soil. 90 :335–342.
Schwyn, B., Neilands, J.B. (1987). Universal chemical assay for the detection and
determination of siderophores. Anal Biochem. 160(1):47–56. doi:10.1016/0003-
2697(87)90612-9.
Segura, D., spín, G. (1998). Mutational Inactivation of a Gene Homologous to
Escherichia coli ptsP Affects Poly-(R)-Hydroxybutyrate Accumulation and Nitrogen
Fixation in Azotobacter vinelandii. Journal of Bacteriology. 180(18):4790-4798.
Schmidt, W., Martin, P., Omay, H., Bangerth, F. (1988). Influence of Azospirillum
brasilense on nodulation of legumes, in: W. Klingmuller (Ed.), Azospirillum IV
Genetics, Physiology, Ecology, Springer-Verlag, Heidelberg, Berlin, 1988, pp.92–100.
Shabaev, V., Smolin, V., Provorov, N., Simarov, B. (1996). The influence of various
Rhizobium meliloti strains on lucerne mass and nitrogen accumulation on the mineral
nitrogen background. Biology Bulletin, 23:291-296.
Shah, S., Karkhanis, V., Desai, A. (1992). Isolation and characterization of
siderophore, with antimicrobial activity from Azospirillum lipoferum. M. Curr Microbiol
25(6):34–35. doi:10.1007/ bf01577233.
Siddhuraju, P., Vijayakumari, K. y Janardhanan, K. (1996). Chemical composition and
nutritional evaluation of an underexploited legume, Acacia nilotica (L.) Del. Food
Chemistry, 57(3), 385–391.
Singh, B.B., Ajeigbe, H.A., Tarawali, S.A., Fernandez-Rivera, S., Abubakar, M. (2003).
Improving the production and utilization of cowpea as food and fodder. Field Crops
Res. 84, 169–177.
Sprent, J.I., Parsons, R. (2000). Nitrogen fixation in legumes and non legume trees.
Field crop research. 65: 183-196.
Sprent, J.I. (2001). Nodulation in legumes. Kew: Royal Botanical Gardens.
Sylvia, D., Hartel, P., Fuhrmann, J., Zuberer, D. (2005). Principles and applications of
soil Microbiology. Segunda edición. Editorial Prentice Hall. New Jersey, EE.UU. 644 p.

76
Socolow, R.H. (1999). Nitrogen management and the future of food: lessons from the
management of energy and carbon. Proc. Natl. Acad. Sci. USA 96: 6001-6008.
Soto Urzua, L., Baca, B.E. (2001). Mecanismos de proteccion de la nitrogenasa a la
inactivacion por oxigeno. Rev Latinoam Microbiol. Jan-Mar; 43(1):37-49.
Steele, W.M., Allen, D.J., Summerfield, R.J. (1985). Cowpea (Vigna unguiculata (L.)
Walp.). In: Summerfield, R.J., Roberts, E.H. (Eds.), Grain Legume Crops. Collins,
London, UK, pp. 520–583.
Sundara, R., Shinha, M. (1963).Organis phosphate solubilizers in soil. Soil Science
and Plant Nutrition. 9 (2):45-49.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011).
MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood,
Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and
Evolution 28 (10): 2731-2739.
Tinker, P. 1984. The role of microorganisms in mediating and facilitating the uptake of
plant nutrients from soil. Plant Soil. 76: 77-91.
Tejera, N., Lluch, C., Martınez-Toledo M.V., Gonzalez-Lopez, J. (2005). Isolation and
characterization of Azotobacter and Azospirillum strains from the sugarcane
rhizosphere. Plant Soil., 270: 223-232.
Torres, M., Valencia, S., Bernal J., Martinez P. (2000). Isolation of Enterobacteria,
Azotobacter sp. and Pseudomonas sp., Producers of Indole-3- Acetic Acid and
Siderophores, from Colombian Rice Rhizosphere. Revista Latinoamericana de
Microbiología. Vol 42. Pp 171-176.
Tortora, M., Díaz-Ricci, J., Pedraza, R.(2011). Azospirillum brasilense siderophores
with antifungal activity against Colletotrichum acutatum. Arch Microbiol 193:275–286
DOI 10.1007/s00203-010-0672-7.
Van Peer, R., Niemann, G.J., Schippers, B. (1991). Induced resistance and
phytoalexin accumulation in biological control of Fusarium wilt of carnation by
Pseudomonas sp. Strain WCS417r. Phytopathology. 81: 728-734.
Vanbleu, E., Vanderleyden, J. (2003). Molecular genetics of rhizosphere and plant
root colonization. In: Elmerich, C., Newton, W.E. (Eds.), Associative and Endophytic
Nitrogen-fixing Bacteria and Cyanobacterial Associations. Kluwer Academic
Publishers, Amsterdam, pp. 85–105.

77
Vance, C.P. (1998). Legume symbiotic nitrogen fixation: agronomic aspects: In:
Spaink, H. P., et al. (Eds.). The Rhizobiaceae. Kluwer Academic Publishers,
Dordrecht, pp. 509-530.
Vincent, J.M. (1970). A manual for the practical study of root-nodule bacteria/J.M.
Wang, R.F., Cao, W.W. and Cerniglia, C.E. (1996). Phylogenetic analysis of
Fusobactenum prausnitzii based upon 16s rRNA gene sequence and PCR
confirmation. Int. J. Syst. Bacteriol. 46341-343.
Wellburn, A. (1994). The spectral determination of chlorophylls a and b, as well as
total carotenoids, using various solvents with spectrophotometers of different
resolution. J. Plant Physiol. 144, 307–313.
Willems, A., Fernández-López, M., Muñoz-Adelantado, E., Goris, J., De Vos, P.,
Martínez-Romero, E., Toro, N., Gillis, M. (2003). Description of New Ensifer strains
from nodules and proposal to transfer Ensifer adhaerens Cassida 1982 to
Sinorhizobium as Sinorhizobium adhaerens comb. nov. Request for an Opinion. Int. J.
Syst. Evol. Microbiol. 53, 1207–1217.
Yahalom, E., Okon, Y., Dovrat, A. (1987). Azospirillum effects on susceptibility to
Rhizobium nodulation and on nitrogen fixation of several forage legumes. Can. J.
Microbiol. 33, 510±514.
Yanni, Y., Rizk, R., Fattah, F.K., Squartine, A. (2001). The beneficial plant growth-
promoting association of Rhizobium leguminosarum bv. trifolii with rice root. Australian
Journal of Plant Physiology. 28: 845-870.
Yi, Y., Huang, W., Ge, Y. (2008). Exopolysaccharide: a novel important factor in the
microbial dissolution of tricalcium phosphate. World J Microbiol Biotechnol. 24:1059–
1065. DOI 10.1007/s11274-007-9575-4.
Young, J.M., Kuykendall, L.D., Martínez-Romero, E., Kerr, A., and Sawada, H. (2001).
A revision of Rhizobium Frank 1889, with an emended description of the genus, and
the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola
de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R.
rubi, R. undicola and R. vitis. International Journal of Systematic and Evolutionary
Microbiology, 51, 89–103.
Young, J.M. (2003). The genus Ensifer (Casida, 1982) takes priority over
Sinorhizobium (Chen et al., 1988) and Sinorhizobium morelense (Wang et al., 2002) is
a later synonym of Ensifer adhaerens (Casida, 1982). Is the combination

78
Sinorhizobium adhaerens (Casida, 1982) Willems et al., 2003 legitimate? Request for
an opinion. Int. J. Syst. Evol. Microbiol., 53, 2107-2110.
Young, J.M. (2004). Renaming of Agrobacterium larrymoorei Bouzar and Jones 2001
as Rhizobium larrymoorei (Bouzar and Jones 2001) comb. nov. Int. J. Syst. Evol.
Microbiol. 54, 149.
Young, J.M., Pennycook, S.R. and Watson, D.R.W. (2006). Proposal that
Agrobacterium radiobacter has priority over Agrobacterium tumefaciens. Request for
an Opinion. Int. J. Syst. Evol. Microbiol. 56, 491–493.
Zahran, H. (2001). Rhizobia from wild legumes: diversity, taxonomy, ecology, nitrogen
fixation and biotechnology. J. Biotech, 91, 143-153 .
Zilli, J.E., Valisheski, R.R., Freire Filho, F.R., Prata Neves, M.C., Rumjanek, N.G.,
(2004). Assessment of cowpea Rhizobium diversity in Cerrado areas of northeastern
Brazil. Brazil. J. Microb. 35, 281–287.





























