Embed Size (px)
Citation preview

Volumen 2 / N
o. 1 / Enero-Junio 2008Revista C
olombiana de C
iencias Hortícolas
Editorial 5
Seccióndefrutales
Efectodelatemperatura,inmersiónenaguayconcentracióndefitorreguladoressobrelagerminacióndesemillasdepapaya(Carica apayaL.)Effect of temperature, immersion in water and concentration of plant growth regulators on germination of papaya (Carica papaya L.) seedsArletteIvonneGilC.,DiegoMirandaL. 9
Característicasfísico-químicasdefrutosdefeijoainfluenciadasporlaposiciónenelcanopiPhysico-chemical characteristics of pineapple guava fruits as influenced by canopi positionRonaldR.Martínez-Vega,GerhardFischer,AníbalHerrera,BernardoChaves,OmarCamilo
Quintero 21
Influenciadelácidogiberélicoydelnitratodecalciosobreladuraciónposcosechadefrutosdefresa(Fragariasp.)Influence of gibberellin and calcium nitrate on post-harvest conservation of strawberry (Fragaria sp.) fruits
FánorCasierra-PosadayRicardoA.SalamancaG. 33
Respuestasfisiológicasymorfológicasdeplantasdemora(Rubussp.)sometidasaestrésporvientoinducidoPhysiological and morphological responses of blackberry (Rubus sp.) plants to wind-induced stress
FánorCasierra-Posada,OscarE.Aguilar-Avendaño 43
Seccióndehortalizas
Efectodediferentesláminasderiegoysustratosenlapropagacióndetomate(Solanum lycopersicumL.)Effect of different water levels and substrates on tomato (Solanum lycopersicum L.) propagation
YuliDeaquiz-Oyola,JavierÁlvarez-Herrera,AnaFraile 54
Comportamientodelosnutrientesentejidofoliarenbrócoli(Brassica oleraceavar.italica)‘Coronado’yrepollo(Brassica oleracea)hibrido‘Delus’cultivadosenlaSabanadeBogotáNutrient levels in foliar tissue of broccoli (Brassica oleracea var. italica) ‘Coronado’ and cabbage (Brassica oleracea) hybrid ‘Delus’ cultivated on the Bogota PlateauCarlosCarranza,OctavioLanchero,DiegoMiranda 66
Antagonismoin vitrodeTrichodermaspp.sobreaislamientosdeSclerotiniaspp.yRhizoctoniaspp.Antagonism of Trichoderma spp. against isolates of Sclerotinia spp. and Rhizoctonia spp. in vitro
LillianaHoyos-Carvajal,GloriaDuque,SergioOrduzP. 76
AbundanciaydiversidaddeBacillusthuringiensisdediferenteshábitatentresmunicipiosdeBoyacá,ColombiaAbundance and diversity of Bacillus thuringiensis in different habitats within three municipalities of Boyaca, ColombiaJeimyM.Poveda,JohnW.Martínez 87
Seccióndeornamentales
Aproximacióndeladinámicapoblacionaldelosmicroorganismosendiferentessustratosempleadosenelcultivoderosa(Rosaspp.var.Charlotte)enlaSabanadeBogotáOverview of the population dynamics of microorganisms in different substrates used in rose (Rosa spp. var. Charlotte) cultivation on the Bogota PlateauMilenaAlfonso,AlejandraCoca,WilliamRamírez,LilianaHoyos-Carvajal 98
Seccióndeotrasespecies
Estudiosdeadaptaciónymanejointegradodeestevia(Stevia rebaudianaBert.):nuevaalternativaagroindustrialdelCaribecolombiano.UnarevisiónEnvironmental suitability and integrated management studies in stevia (Stevia rebaudiana Bert.): a new agro-industrial alternative for the Colombian Caribbean. A review
AlfredoJarma-Orozco 109
Políticaeditorialeinstruccionesparalosautores 121


PRESIDENTEDiego Miranda
COMITÉ EDITORIAL
COMITÉ CIENTÍFICO
VICEPRESIDENTEMaría Soledad Hernández
EDITORFanor Casierra-Posada
COEDITORGerhard Fischer
Marco CabezasBernardo Chaves
Miguel EspitiaRebecca Lee
Nelson RodríguezEdison Valencia Kris Wyckhuys
SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS - SCCHBOGOTÁ - COLOMBIA
Miguel A. AltieriUniversity of California, Berkeley (Estados Unidos)
Galdino Andrade FilhoUniversidade Estadual de Londrina (Brasil)
Raúl CabreraTexas A+M University (Estados Unidos)
Ana Maria CastagninoUniversidad Nacional del Centro de la Provincia de Buenos Aires (Argentina)
Daniel H. DíazLaboratorios Agroenzimas (México)
Georg EbertKali + Salz Gruppe (Alemania)
José Miguel GuzmánUniversidad de Almería (España)
Miguel JordánUniversidad Mayor, Santiago (Chile)
Marcelo Francisco PompelliUniversidade Federal de Pernambuco (Brasil)
Jairo Antonio OsorioCorporación Colombiana de Investigación Agropecuaria (Colombia)
Philip A. StanslyUniversity of Florida (USA)
Ricardo RamírezUniversity of Guelph (Canadá)

© 2008 Revista Colombiana de Ciencias HortícolasSociedad Colombiana de Ciencias HortícolasISSN: 2011-2173
INFORMACIÓN, CORRESPONDENCIA Y CANJE
Sociedad Colombiana de Ciencias HortícolasFacultad de Agronomía Universidad Nacional de ColombiaA.A. 14490, Bogotá-ColombiaTeléfono: 571 - 316 5000 Ext. 19041/19043Tel-fax: 571 - 316 [email protected], [email protected]
CORRECTOR DE ESTILO EN INGLÉS
Phil Stansly
DISEÑO GRÁFICO Y ARMADA ELECTRÓNICAMauricio González, Carmen Lucía González [email protected]
IMPRESIÓNEditorial Gente Nueva
Bogotá, ColombiaJunio de 2008
La presente publicación es de carácter científico (Artículo 4, resolución 1508, Ministerio de Cultura, Octubre 3 de 2000).
A nivel internacional, la Revista Colombiana de Ciencias Hortícolas está integrada en las bases de datos de CAB Abstracts y AGRIS (FAO).

JUNTA DIRECTIVA DE LA SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS (SCCH)
Gerhard FischerUniversidad Nacional de Colombia, Bogotá
Wilson de Jesús Piedrahita Universidad Nacional de Colombia, Bogotá
VOCALES
Diego MirandaUniversidad Nacional de Colombia, Bogotá
María Soledad HernándezInstituto Sinchi, Bogotá
Marlon Hans Rodríguez Sociedad Colombiana de Ciencias Hortícolas
Alfredo JarmaUniversidad de Córdoba, Monterí[email protected]
Hernán PinzónAsesor Hortícola, Bogotá [email protected]
Stanislav MagnitskiyUniversidad Nacional de Colombia, Bogotá
PRESIDENTE VICEPRESIDENTE SECRETARIO EJECUTIVO TESORERO
Luís Enrique FlórezUniversidad de Bogotá
Jorge Tadeo [email protected]
FISCAL
Fánor Casierra-PosadaUniversidad Pedagógica y Tecnológica
de Colombia, [email protected]
EDITOR REVISTA


EDITORIALFÁNOR CASIERRA-POSADADirector Comité Editorial
Revista Colombiana de Ciencias Hortícolas
Vol. 2 - No. 1 - 2008
La Revista Colombiana de Ciencias Hortícolas se ha posicionado entre los miembros de la Sociedad Colombiana de Ciencias Hortícolas (SCCH) y en la comunidad cien-tífica nacional e internacional como una importante fuente de consulta, lo cual se ha podido verificar mediante las solicitudes de personas interesadas en la adquisición de los textos publicados. Así también, se espera que este interés se incremente con la ubicación de la revista en la plataforma de la International Society for Horticultural Sience, por medio de la cual los científicos e interesados en la horticultura tropical podrán tener acceso a los artículos allí publicados. De esta forma se pretende cumplir con el objetivo de dar respuesta a los múltiples interrogantes generados en el sector hortícola tropical. Consecuente con lo anterior, este medio divulgativo establece una estrategia de comunicación permanente entre los investigadores con el entorno regio-nal y con la comunidad técnico-científica nacional e internacional.
La presente entrega está dividida en tres partes. La sección de frutales, en la cual se publican artículos relacionados con germinación de semillas de papaya; características fisicoquímicas de frutos de feijoa; influencia del ácido giberélico en frutos de fresa y respuestas fisiológicas y morfológicas de plantas de mora expuestas al viento. En lo referente a las hortalizas, en esta sección se divulgan aspectos relacionados con la pro-pagación de tomate; el comportamiento de los nutrientes en brócoli; el antagonismo in vitro de Trichoderma spp. y la diversidad de Bacillus thuringiensis en diferentes regiones de Boyacá. Continuando con la estructura, se encuentra el componente de especies ornamentales, en la cual se divulga la dinámica poblacional de los microor-ganismos en diferentes sustratos en el cultivo de rosa. Finalmente, este órgano di-vulgativo hace alusión a la oferta tecnológica de Stevia rebaudiana como alternativa agroindustrial para el Caribe colombiano.
Por otro lado, en el marco de las actividades de extensión de la SCCH, hemos conti-nuado con la realización de eventos científicos, congresos, simposios y seminarios, en los que se han dado cita investigadores de carácter nacional e internacional, los cuales se han constituido en elementos de intercambio de opiniones y de posibilidades de co-operación internacional e interinstitucional. Una información más detallada de estos eventos puede consultarse en la página web de la SCCH (www.soccolhort.com).
De esta manera, se espera que esta revista siga siendo un instrumento que recoja opiniones, experiencias y soluciones a los problemas hortícolas en el trópico y, desde luego, pueda ayudar a generar instancias de debate científico en esta área, con miras a reorientar el desarrollo de la horticultura colombiana.


CONTENIDOVolumen 2 - No. 1 - 2008
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS
PÁG.
Editorial 5
Sección de frutales
Efecto de la temperatura, inmersión en agua y concentración de fitorreguladores sobre la germinación de semillas de papaya (Carica papaya L.)Effect of temperature, immersion in water and concentration of plant growth regulators on germination of papaya (Carica papaya L.) seeds
Arlette Ivonne Gil C., Diego Miranda L. ....................................................................................................................................................... 9
Características físico-químicas de frutos de feijoa influenciadas por la posición en el canopiPhysico-chemical characteristics of pineapple guava fruits as influenced by canopi position
Ronald R. Martínez-Vega, Gerhard Fischer, Aníbal Herrera, Bernardo Chaves, Omar Camilo Quintero ....................................................21
Influencia del ácido giberélico y del nitrato de calcio sobre la duración poscosecha de frutos de fresa (Fragaria sp.)Influence of gibberellin and calcium nitrate on post-harvest conservation of strawberry (Fragaria sp.) fruits
Fánor Casierra-Posada, Ricardo A. Salamanca G. ...................................................................................................................................33
Respuestas fisiológicas y morfológicas de plantas de mora (Rubus sp.) sometidas a estrés por viento inducidoPhysiological and morphological responses of blackberry (Rubus sp.) plants to wind-induced stress
Fánor Casierra-Posada, Oscar E. Aguilar-Avendaño ...............................................................................................................................43
Sección de hortalizas
Efecto de diferentes láminas de riego y sustratos en la propagación de tomate (Solanum lycopersicum L.) Effect of different water levels and substrates on tomato (Solanum lycopersicum L.) propagation
Yuli Deaquiz-Oyola, Javier Álvarez-Herrera, Ana Fraile ...........................................................................................................................54

REV. COLOMB. CIENC. HORTÍC.
�
Comportamiento de los nutrientes en tejido foliar en brócoli (Brassica oleracea var. italica) ‘Coronado’ y repollo (Brassica oleracea) hibrido ‘Delus’ cultivados en la Sabana de Bogotá Nutrient levels in foliar tissue of broccoli (Brassica oleracea var. italica) ‘Coronado’ and cabbage (Brassica oleracea) hybrid ‘Delus’ cultivated on the Bogota Plateau
Carlos Carranza, Octavio Lanchero, Diego Miranda ..................................................................................................................................66
Antagonismo in vitro de Trichoderma spp. sobre aislamientos de Sclerotinia spp. y Rhizoctonia spp.Antagonism of Trichoderma spp. against isolates of Sclerotinia spp. and Rhizoctonia spp. in vitro
Lilliana Hoyos-Carvajal, Gloria Duque, Sergio Orduz P. .............................................................................................................................76
Abundancia y diversidad de Bacillus thuringiensis de diferentes hábitat en tres municipios de Boyacá, ColombiaAbundance and diversity of Bacillus thuringiensis in different habitats within three municipalities of Boyaca, Colombia
Jeimy M. Poveda, John W. Martínez .......................................................................................................................................................87
Sección de ornamentales
Aproximación de la dinámica poblacional de los microorganismos en diferentes sustratos empleados en el cultivo de rosa (Rosa spp. var. Charlotte) en la Sabana de BogotáOverview of the population dynamics of microorganisms in different substrates used in rose (Rosa spp. var. Charlotte) cultivation on the Bogota Plateau
Milena Alfonso, Alejandro Coca, William Ramírez, Liliana Hoyos-Carvajal ..............................................................................................98
Sección de otras especies
Estudios de adaptación y manejo integrado de estevia (Stevia rebaudiana Bert.): nueva alternativa agroindustrial del Caribe colombiano. Una revisiónEnvironmental suitability and integrated management studies in stevia (Stevia rebaudiana Bert.): a new agro-industrial alternative for the Colombian Caribbean. A review
Alfredo Jarma-Orozco ......................................................................................................................................................................... 109
Política editorial e instrucciones para los autores 121

Arlette Ivonne GIl C.1
DIeGo MIrAnDA l.2, 3
reSUMen
En el cultivo de papaya, la mayoría de la semilla es importada, costosa y su calidad se asume de acuerdo con la información del marbete. Para esta fase del cultivo, no existen recomendaciones específicas en cuanto a temperaturas óptimas, tratamientos para las semillas o influencia de reguladores de crecimiento sobre la germinación, con lo que se corre el riesgo de tener pérdidas económicas. Esta investigación se realizó como un aporte al conocimiento sobre la calidad de la semilla de papaya utilizada para la propagación en cultivos com-erciales, debido a que los productores no disponen de métodos eficientes para su propagación. Con el fin de evaluar el efecto de la temperatura (20, 25, 30 y 35°C), el tiempo de inmersión en agua (0, 24, 48 y 72 horas) y la concentración de los fitorreguladores ácido giberélico, AG3, y ácido naftalenacético, ANA, (0, 100, 200 y 300 mg L-1) sobre la germinación de semillas de papaya se estableció un experimento en condiciones de laboratorio (humedad relativa 80-90% y temperaturas variables según tratamiento), en el cual se utilizaron dos materiales comerciales importados ampliamente utilizados en el país: la variedad Maradol y el híbrido Tainung-1. Una temperatura de 30°C, junto con 48 horas de inmersión de la semilla en agua, fueron los tratamientos con mayor respuesta en el porcentaje de germinación. Los tratamientos con AG3 también fueron de alta respuesta por parte de las semillas, sin embargo, las diferencias con los tratamientos de inmersión en agua no fueron significativas. Finalmente, no se recomienda utilizar el regulador ANA, debido a que provoca malformaciones en las radículas de las semillas, e inhibe su posterior desarrollo.
1 Facultad de Ciencias Agropecuarias, Universidad de Cundinamarca, Fusagasugá (Colombia).2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).3 Autor para correspondencia. [email protected]
Efecto de la temperatura, inmersión en agua y concentración de fitorreguladores sobre la germinación de semillas de papaya (Carica papaya L.)
Effect of temperature, immersion in water and concentration of plant growth regulators on germination of papaya (Carica papaya L.) seeds
Palabras clave adicionales: imbibición, variedad Maradol, híbrido Tainung-1, ácido giberélico, ácido naftalenacético.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 9-20, 2008
Fruto maduro de papaya con semillas.Foto: D. Miranda L.

REV. COLOMB. CIENC. HORTIC.
10
IntroDUCCIÓn
Additional keywords: imbibition, Maradol variety, Tainung-1 hybrid, gibberellic acid, naphthalene acetic acid.
Fecha de recepción: 16-01-2008 Aprobado para publicación: 03-06-2008
ABStrACt
Specific recommendations are not available to for temperatures, treatments, or growth regulators to optimize germination of papaya seed, with the consequent, risk of increasing of economic losses. An experiment was established in Colombia under laboratory conditions to evaluate the effect of temperature (20, 25, 30, or 35°C), immersion time in water (0, 24, 48, or 72 hours) and the concentration of the growth regulators gibberellic acid, GA3, and naphthalene acetic acid, NAA (0, 100, 200, or 300 ppm) on germination of papaya seeds at, 80-90% relative humidity. Two commercial varieties were tested: Maradol and the hybrid Tainung-1. A temperature of 30°C together with 48 hour seed immersion in water were the treatments resulting in the highest percentage of germination. The seeds had a strong response to the treatments with GA3; nevertheless, differences compared to immersion treatments were not significant. Finally, the growth regulator NAA is not recommended for seed treatments because it causes malformation of the seed radicles and inhibits their later development.
GIL/MIRANDA
La papaya (Carica papaya L.) es originaria de América tropical y, según algunos autores, espe-cíficamente de Centroamérica (entre México y Costa Rica), el cultivo se expandió por el resto de América del Sur y se ha desarrollado amplia-mente en las zonas tropicales y subtropicales en todo el mundo. Los tres principales productores de papaya para 2007 fueron Brasil, México y Ni-geria con 1.811.540, 919.225 y 765.000 t respec-tivamente, lo que representa el 25% para Brasil, el 12% para México y 10% para Nigeria de la producción mundial; Colombia ocupó el octavo lugar con 207.000 t, seguido por Guatemala con 184.000 t (Faostat, 2007).
Según Semicol (2002), la papaya es en la actua-lidad uno de los frutos de mayor consumo en el interior del país, gracias a sus poderes digestivos. En los últimos años, se está imponiendo la varie-dad Maradol roja mejorada, desarrollada en Cuba y los híbridos, como el Tainung-1 y Tainung-2 de origen taiwanés, no sólo por su exquisito sabor, sino también por sus características de duración
en poscosecha y su resistencia al transporte en largas distancias.
Sin embargo, los métodos de propagación de la papaya son sexuales y asexuales. Los métodos asexuales, como el uso de esquejes, estacas y aco-dos, aunque en algunos países se realizan a muy pequeña escala, rara vez se utilizan en planta-ciones comerciales debido a que el costo no se justifica por la vida económicamente corta de la plantación; sólo se recomienda la propagación por semillas, preferiblemente de semillas preger-minadas (Pereira de Araujo Filho, 1987).
Por definición, la germinación sensu stricto in-corpora aquellos eventos que comienzan con la toma de agua por la semilla seca quiescente y termina con la protrusión de la radícula, la elon-gación del eje embrionario, posterior crecimiento y desarrollo del embrión (Fosket, 1994; Bewley, 1997; Welbaum et al., 1998). Con la absorción de agua, el embrión y el endospermo (só lo en semi-llas endospérmicas) se expanden y la consiguien-

Vol. 2 - No. 1 - 2008
11EFECTOS SOBRE LA GERMINACIóN DE SEMILLAS DE pApAyA
te ruptura de las envolturas más externas permi-te el libre acceso de oxígeno para la respiración (Thomson, 1979). Se considera que la germina-ción está completa cuando todas las reservas en la semilla se han consumido y la plátula es capaz de vivir de forma independiente (Besnier, 1988; Bradbeer, 1988).
Las condiciones ambientales favorables son ne-cesarias para la germinación de las semillas. La humedad, el oxígeno, la temperatura y la luz afectan la germinación, pero con diferencias en-tre especies que poseen distintos requerimientos óptimos. Si las condiciones internas de la semilla y las condiciones ambientales son adecuadas, la germinación tendrá lugar (Lee y Heimsch, 1962).
Las semillas de papaya han sido clasificadas como intermedias por tolerancia a la desecación, entre las ortodoxas y las recalcitrantes, mostran-do signos de estrés por desecación con contenidos de humedad menores de 8% (Ellis et al., 1991); la desecación induce quiescencia metabólica o dormancia (Word et al., 2000), lo que ocasiona que durante la siembra la germinación sea lenta y se obtengan porcentajes de emergencia bajos. Para incrementar el porcentaje de germinación y emergencia, así como su homogeneidad, se han sugerido algunos métodos: una técnica utilizada es la pregerminación, en la cual las semillas se ponen a remojar en una cubeta ó recipiente con agua limpia de pH neutro, la cual se debe cam-biar cada 8-12 h por 2 ó 3 d. Después de 48 h de remojo, se observa si existe flotación de las semi-llas, éstas se llevan a otra cubeta para seguir el procedimiento de remojo. Si en 24 h no se han hundido, se deben de eliminar. Otro método con-siste en el remojo de las semillas en ácido giberé-lico a razón de 200 mg L-1 (Navarrete, 1996) ó 600 mg L-1 (Nagao y Furatani 1986; Furatani y Nagao 1987), agua (Lima et al., 1985; Salvador-Fi-gueroa et al., 2005), y 1,0 M de nitrato de potasio. La aplicación de los tratamientos anteriores tiene el propósito de eliminar (Lima et al., 1985) o con-trarrestar el efecto de las sustancias lipoprotéicas que retardan o inhiben la germinación.
El efecto de reguladores de crecimiento u otros factores sobre la germinación de semillas de pa-paya es bien conocido. El uso de reguladores de crecimiento en fase de germinación mejora el desempeño de las plántulas, acelerando la velo-cidad de emergencia o incrementando el poten-cial de las semillas de varias especies (Bevilaqua et al., 1993). El uso de compuestos químicos biológicamente activos, como los reguladores de crecimiento, puede disminuir el efecto de los factores adversos en la calidad y desempeño de las semillas. Por otro lado, las giberelinas se consideran las promotoras de la iniciación enzi-mática en el proceso de germinación y partici-pan en diferentes concentraciones dependiendo de los estadíos de las semillas. Para Hartmann et al. (2005) y Hedden y Proebsting (1999), las giberelinas desempeñan un papel importante en dos etapas de la germinación de las semillas en general. En la primera etapa, las giberelinas actúan en la fase inicial de inducción de enzi-mas y, posteriormente, activan las enzimas que intervienen en la movilización del sistema de alimentos de reserva. Pueden tener efecto sobre la elongación del tejido embrionario, quizás al-terando la extensibilidad de la pared celular, de este modo se facilita la toma de agua (Hilhorst, 1995).
El ANA (ácido α-naftalenacético) es una auxina sintética con fuerte actividad auxínica, la cual se relaciona con la presencia de una carga neta positiva en el anillo, situada a una distancia de 0,05 nm de la carga negativa del grupo COOH y promueve respuestas reguladas por las auxi-nas, como producción de etileno y elongación del hipocótilo en tomate; también previene la brotación de las yemas de tubérculos de patata almacenados (Mito y Bennett, 1995).
Leonel y Rodrigues (1996), trabajando con semillas de limón Cravo, obtuvieron mayor germinación con el uso de 50 mg L-1 de AG3 y Bevilaqua et al. (1993) verificaron que semillas de arroz, tratadas com 100 mg L-1 de AG3, presentaron mayor vigor en relación con aquellas que no habían sido tratadas.

REV. COLOMB. CIENC. HORTIC.
12
El objetivo de esta investigación fue evaluar el efecto de diferentes temperaturas y tiempos de inmersión de la semilla en agua y en concentra-ciones de AG3 y ANA sobre los porcentajes de germinación y los cambios morfológicos de se-millas de papaya de la variedad Maradol y el hí-brido Tainung-1.
MAterIAleS Y MÉtoDoS
Semillas de papaya (Carica papaya L.), de la va-riedad Maradol roja y el híbrido Tainung-1, ad-quiridas en casas comercializadoras de semillas. La semilla de Maradol, procedente de la casa Ca-risem (Cuba) con la presentación en lata de 50 g (lote 0225-6) y la del híbrido Tainung-1, en lata de 100 g (lote AG00807).
Para el remojo de semillas en agua, éstas se colo-caron en 30 mL de agua destilada durante 0 (tes-tigo), 24, 48 y 72 h, para su imbibición. Pasado este tiempo, se colocaron 50 semillas en cajas de Petri con papel toalla marca Scott, humedecida con 10 mL de agua destilada, con tres repeticio-nes por tratamiento. Cuatro temperaturas 20, 25, 30 y 35ºC fueron utilizadas en la evaluación. Se utilizó un germinador Seedburo Equipment Company® Des Plaines (IL, USA). La humedad relativa en la cámara de germinación varió entre 80% y 90%. La evaluación de la germinación de las semillas fue llevada hasta los 40 d, ya que, de acuerdo con trabajos realizados en papaya por Yahiro (1979), la germinación con tratamientos de pregerminación a temperaturas de 30ºC de-mora entre 20-30 días. Con los resultados fina-les, se obtuvo el porcentaje de germinación acu-mulado al final del tiempo.
Para evaluar el efecto de los fitorreguladores, se hizo inmersión de las semillas en 30 mL de solución con AG3 y ANA durante 48 h, en con-centraciones de 100, 200 y 300 mg L-1, además del testigo. No se hizo prueba de imbibición por cuanto ésta correspondió a otro experimento, y no se constituyó en objetivo de este experimen-
to. Para preparar la solución del fitorregulador, se pesaron 0,56 g de NaOH y se agregaron 20 mL de agua destilada. Posteriormente, se pesaron el AG3 y el ANA (marca MERCK®), para lograr las concentraciones requeridas se disolvieron en 5 mL de la solución de NaOH. Esta solución se completó con 25 mL de agua destilada. A las 24 h se cambiaba la solución de regulador, con el fin de reducir el efecto de posibles sustancias lipoproteicas sobre la germinación. Después del tiempo de inmersión, se colocaron lotes de 50 se-millas en cada caja de Petri con papel toalla Scott humedecido con 10 mL de agua destilada. Tres repeticiones, con 50 semillas por repetición, se llevaron al germinador. Similar a la prueba ante-rior, se obtuvo el porcentaje de germinación final en el tiempo. El criterio utilizado para definir la germinación de las semillas, para todas las prue-bas, fue la protrusión radicular, con una longitud de 5 mm a través de la cubierta seminal, según lo anotado Wood et al. (2000).
Sobre los resultados experimentales, se aplicó el análisis de varianza y se realizaron pruebas de hipótesis sobre las medias de los tratamientos y comparaciones ortogonales de grupos de trata-mientos. De esta forma, se pretendió determinar si los tratamientos fueron efectivos para aumen-tar el porcentaje de germinación.
Los tratamientos consistieron en una combina-ción factorial 4*2*11, con cuatro temperaturas, dos materiales de papaya, y 11 formas de mane-jo de la semilla, respectivamente, presentadas de la siguiente manera: 1. semilla sin inmersión en agua (0 h), 2. semilla con 24 h de inmersión en agua, 3. semilla con 48 h de inmersión en agua, 4. semilla con 72 h de inmersión en agua, 5. tes-tigo sin fitorregulador, 6. tratamiento con AG3, a una concentración de 100 mg L-1, 7. tratamiento con AG3, a una concentración de 200 mg L-1, 8. tratamiento con AG3, a una concentración de 300 ppm, 9. tratamiento con ANA, a una concentra-ción de 100 mg L-1, 10. tratamiento con ANA, a una concentración de 200 mg L-1 y 11. tratamien-to con ANA, a una concentración de 300 mg L-1.
GIL/MIRANDA

Vol. 2 - No. 1 - 2008
13
Para la estadística, se realizó un análisis de va-rianza, que se aplicó por separado para cada temperatura. Las combinaciones de material de papaya y manejo de la semilla se evaluaron me-diante contrastes ortogonales.
Adicionalmente, se incluyó la prueba de compara-ción múltiple de Tukey (P≤0,05). Para el análisis estadístico de todos los resultados obtenidos, se utilizó el programa SAS (Statistical Análisis Sys-tem) versión 8.1, con los procesos Anova para los análisis de varianza y GLM (General Linear Mo-del) para las pruebas de contrastes ortogonales.
reSUltADoS Y DISCUSIÓn
De acuerdo con el análisis realizado, el porcen-taje de germinación difirió entre los materiales evaluados, en particular bajo las temperaturas de 20, 25 y 35ºC. La semilla de papaya es muy sensible a los cambios de temperatura y de hu-medad, dichos cambios causan una disminución progresiva de la viabilidad y el porcentaje de ger-minación de la misma, por lo que se debe conser-var el menor tiempo posible bajo las condiciones del medio ambiente reinante. A las temperaturas de 20 y 25ºC, el porcentaje de germinación fue mayor con Maradol que con Tainung-1. A 30ºC, no hubo diferencias significativas entre los dos, pero a 35ºC, Tainung-1 superó a Maradol, esta última con un comportamiento muy deficiente, expresado en su bajo porcentaje de germinación. Adicionalmente, se presentaron diferencias sig-nificativas entre los tratamientos con fitorregu-ladores para las cuatro temperaturas evaluadas (tabla 1). Varias investigaciones independientes muestran que el ácido giberélico estimula la alfa amilasa y otras enzimas proteolíticas, promo-viendo la hidrólisis de material de reserva. La gi-berélina promueve el crecimiento por el aumento de la plasticidad de la pared celular, seguida por la hidrólisis de almidón en azúcar, que reduce el potencial hídrico de la célula. Esto resulta en la entrada de agua a su interior y promueve su alar-gamiento. Los pasos básicos posiblemente invo-lucrados en ese mecanismo se resumen en que
el AG3, producido en el embrión, es transferido a la capa de aleurona de las células, donde la α-amilasa es producida vía síntesis “de novo”. Ésta promueve la conversión de almidón en azúcar, que es usado en su totalidad para el crecimiento de la plántula (Arteca, 1996). A partir de 100 mg L-1 de AG3 hubo una demora en la germinación en el primer conteo de la prueba de germinación. Se verificó también que las semillas tratadas con concentraciones superiores a 100 mg L-1 presen-taron un aumento acentuado en el número de plántulas anormales, indicando un posible efec-to fitotóxico de la hormona. Este efecto también se observó por el número de semillas muertas.
Rosseto et al. (2000) constataron que semillas de maracuyá escarificadas y tratadas con 300 mg
EFECTOS SOBRE LA GERMINACIóN DE SEMILLAS DE pApAyA
Comparación de tratamientoscon contrastes ortogonales
Temperatura, °C20 25 30 35
Variedad, V * * *
Agua u Hormona, A|H * * *
V x A|H, Variedad x Agua u Hormona * * *
Hormona (H), sí o no * *
Variedad x Hormona, V x H
AG3 o ANA * * * *
Variedad x AG3 o ANA, V x GA|A * *
Horas inmersión lineal, HL *
V x HL
Horas inmersión cuadrática, HC * * * *
V x HC *
Horas inmersión cúbica, HCC * *
V x HCC
AG3 lineal, GL * * *
V x GL *
AG3 cuadrático, GC
V x GC
ANA lineal, AL * * *
V x AL *
ANA cuadrática, AC
V x AC
tabla 1. Interacciones entre inmersión, variedad y hormonas a cada temperatura.
* Indica diferencia significativa según la prueba de Tukey (P≤0,05).

REV. COLOMB. CIENC. HORTIC.
14
L-1 de AG3 tuvieron un incremento significativo en la germinación. Por otro lado el número de plantas anormales también aumentó, confir-mando el efecto fitotóxico inducido por el áci-do giberélico en concentraciones elevadas. Sólo el 5% de la germinación fue contabilizada en la unidad experimental, sin AG3, casi el último día de la investigación. Por el contrario, los porcen-tajes de germinación de las parcelas tratadas con AG3 fueron del 39 y 60% en 500 y 1.000 mg L-1, respectivamente. Se notó una considerable pro-moción de la germinación. Este incremento de la germinación en 500 y 1.000 mg L-1 continuó durante 14 d posteriores al tratamiento y 22 días, en el caso de 1.000 mg L-1, y sus porcentajes de germinación fueron superiores a los demás trata-mientos (Yahiro y Oryoji, 1980).
La prueba de comparación múltiple de Tukey, a temperatura de 20°C (tabla 2), muestra que para la variedad Maradol no se presentaron diferencias significativas en el porcentaje de germinación en-tre las semillas tratadas con AG3 y la inmersión en agua (24 y 48 h), lo que lleva a sugerir que no es necesario realizar una alta inversión en el fitorregulador, cuando se puede conseguir un porcentaje de germinación aceptable únicamen-te con la inmersión de las semillas en agua. Para Tainung-1, a esta temperatura se presentaron di-ferencias significativas entre la aplicación de AG3 y la inmersión de las semillas en agua durante 48 h, que obtuvo el promedio más alto entre los tratamientos de agua. Sin embargo, cabe destacar que los porcentajes promedios de germinación de la semilla fueron bajos (menores del 45%), debido
tabla 2. efecto del GA3, AnA y la inmersión en agua sobre el porcentaje de germinación de semillas de papaya variedad Maradol y el híbrido tainung-1 bajo cuatro temperaturas.
Material TratamientoGerminación (%)
20ºC 25ºC 30ºC 35ºCMaradol Agua 0 h 16,00 fghij 78,66 b 88,00 ab 2,66 h
Maradol Agua 24 h 30,00 abcde 81,33 ab 88,66 ab 4,00 h
Maradol Agua 48 h 33,33 abcd 84,00 ab 92,00 a 5,33 h
Maradol Agua 72 h 27,33 bcdefg 80,00 b 86,66 ab 0,00 h
Maradol Testigo hormonal 28,00 bcdef 86,66 ab 89,33 ab 1,33 h
Maradol AG3 100 mg L-1 39,33 abc 82,00 ab 86,66 ab 0,00 h
Maradol AG3 200 mg L-1 40,00 ab 84,66 ab 88,00 ab 0,00 h
Maradol AG3 300 mg L-1 42,66 a 94,00 a 95,33 a 0,00 h
Maradol ANA 100 mg L-1 11,33 ij 30,00 cd 36,66 cd 0,00 h
Maradol ANA 200 mg L-1 11,33 ij 20,66 de 25,33 ef 0,00 h
Maradol ANA 300 mg L-1 10,66 ij 13,33 e 17,33 f 0,00 h
Tainung-1 Agua 0 h 6,00 j 84,00 ab 86,66 ab 23,33 efg
Tainung-1 Agua 24 h 8,00 j 85,33 ab 87,33 ab 27,33 cdef
Tainung-1 Agua 48 h 12,00 ij 90,00 ab 92,00 a 30,66 cd
Tainung-1 Agua 72 h 10,66 ij 82,00 ab 78,66 b 25,33defg
Tainung-1 Testigo hormonal 14,66 ghij 88,66 ab 90,66 a 29,33 cde
Tainung-1 AG3 100 mg L-1 22,66 defghi 86,66 ab 91,33 a 33,33 bc
Tainung-1 AG3 200 mg L-1 25,33 defgh 90,66 ab 93,33 a 37,33 ab
Tainung-1 AG3 300 mg L-1 26,66 cdefg 94,00 a 97,33 a 40,00 a
Tainung-1 ANA 100 mg L-1 18,66 efdhij 36,00 c 42,66 c 27,33 cdef
Tainung-1 ANA 200 mg L-1 13,33 hij 23,33 cde 28,66 de 22,66 fg
Tainung-1 ANA 300 mg L-1 10,00 ij 21,33 de 24,00 ef 19,33 gPromedios con letras distintas indican diferencia significativa según la prueba de Tukey (P≤0,05).
GIL/MIRANDA

Vol. 2 - No. 1 - 2008
15
a que esta temperatura no es la óptima para la adecuación de un semillero de papaya.
Al analizar la prueba de Tukey para 25°C (tabla 2), se observó que no se presentaron diferencias significativas entre los tratamientos con AG3 y la inmersión de las semillas en agua por 24 y 48 h para los dos materiales evaluados, aunque entre estos dos tiempos de inmersión el tratamiento por 48 h fue el de mayor respuesta.
De acuerdo con la respuesta, se puede deducir que esta temperatura y una inmersión por 48 h en agua, son suficientes para obtener un por-centaje de germinación alto (entre el 85% y el 90%), sin tener que incurrir en altos costos por la adquisición de giberelina. Adicionalmente, se observó que los tratamientos con ANA fueron los menos adecuados, debido a que ANA no es promotor de la germinación, sino más bien un inhibidor.
De acuerdo con los resultados obtenidos en la prueba de comparación múltiple de Tukey para 30°C (tabla 2), no se presentaron diferencias sig-nificativas entre los tratamientos de giberelina y agua para ambos materiales evaluados, con lo que se puede concluir que no es necesario aplicar giberelina para obtener altos porcentajes de ger-minación (mayores al 90%), ya que las semillas inmersas 48 h en agua ofrecen prácticamente los mismos resultados. Además, se encontró que las semillas puestas a germinar en esta temperatura obtuvieron los más altos porcentajes de germi-nación. De este modo, no es necesario invertir en giberelina para obtener altos porcentajes de ger-minación, ya que las concentraciones endógenas presentes en la semilla son suficientes para cum-plir con este objetivo. Los tratamientos menos adecuados fueron con ANA, y a mayores con-centraciones del regulador, se presentó el menor porcentaje de germinación, lo que demuestra su efecto inhibitorio.
Los resultados obtenidos en la prueba de Tukey para 35°C (tabla 2) mostraron a Tainung-1 con
los mayores porcentajes de germinación cuando se aplicó giberelina, y hubo diferencias significa-tivas con 48 h de inmersión en agua. La variedad Maradol fue más susceptible a esta temperatura, siendo el mejor tratamiento las 48 h de inmer-sión en agua. Para esta temperatura, los porcen-tajes de germinación mostrados por las semillas fueron bajos (menores al 40%), lo que indica que es excesiva y no se recomienda su utilización.
Cabe anotar un aspecto relevante que fue ob-servado en todos los tratamientos evaluados: las semillas sin inmersión (0 h) obtuvieron bajos porcentajes de germinación, lo cual podría ser ex-plicado debido a que en la semilla no se reactiva-ron la expansión de las paredes celulares, así como tampoco se produjo la expansión del embrión y del endospermo, como consecuencia de la falta de agua, de acuerdo con Thomson (1979). Este com-portamiento fue explicado por Ha et al. (1997) quienes mencionan que las células deshidratadas de la pared celular se comportan como sólidos rígidos. Las cadenas de xiloglucano, sustancia de reserva bastante semejante a un polisacárido estructural presentando (D-glucosa; D-xilosa y D-galactosa en proporción 4:3:1) comprende el 20-25% de las paredes primarias de dicotiledó-neas (Buckeridge et al., 1992) y está íntimamen-te ligado a la celulosa. Según Lima y Buckeridge (2001), en frutos maduros la composición del fru-to es 30,2% pectinas, 10,1% hemicelulosas y el 0,2% de celulosas. Estas hemicelulosas presentan una composición compatible con la presencia de xiloglucano o glucomanano, siendo este último un polímero degradado durante el desarrollo, de-jando la pared celular con proporciones de mo-nosacáridos típicos de xiloglucanos, los cuales pueden unirse al hidrógeno de la celulosa, de este modo, ellas pueden unir las microfibrillas adya-centes y así restringir la expansión celular (Fry et al., 1993). Además, se restringió el acceso del oxígeno para la respiración de la semilla y es po-sible que las proteínas LEA (late embryogenesis abundant), proteínas implicadas en la tolerancia a la desecación, se hayan degradado lentamente, según lo anotado por Bewley (1997). Cuando se
EFECTOS SOBRE LA GERMINACIóN DE SEMILLAS DE pApAyA
REV. COLOMB. CIENC. HORTIC.
16
incrementa la temperatura, se varía la permeabi-lidad de las membranas celulares, aunque la velo-cidad de imbibición aumenta ligeramente (Q10= 1,5-1,8) por la disminución de la viscosidad del agua y el aumento de la energía cinética de sus moléculas, de acuerdo con Montaldi (1995). Este ligero aumento del ritmo de imbibición y la can-tidad de agua total absorbida es independiente de la temperatura, debido a que la imbibición es un fenómeno físico y el agua ingresa a las semillas por difusión.
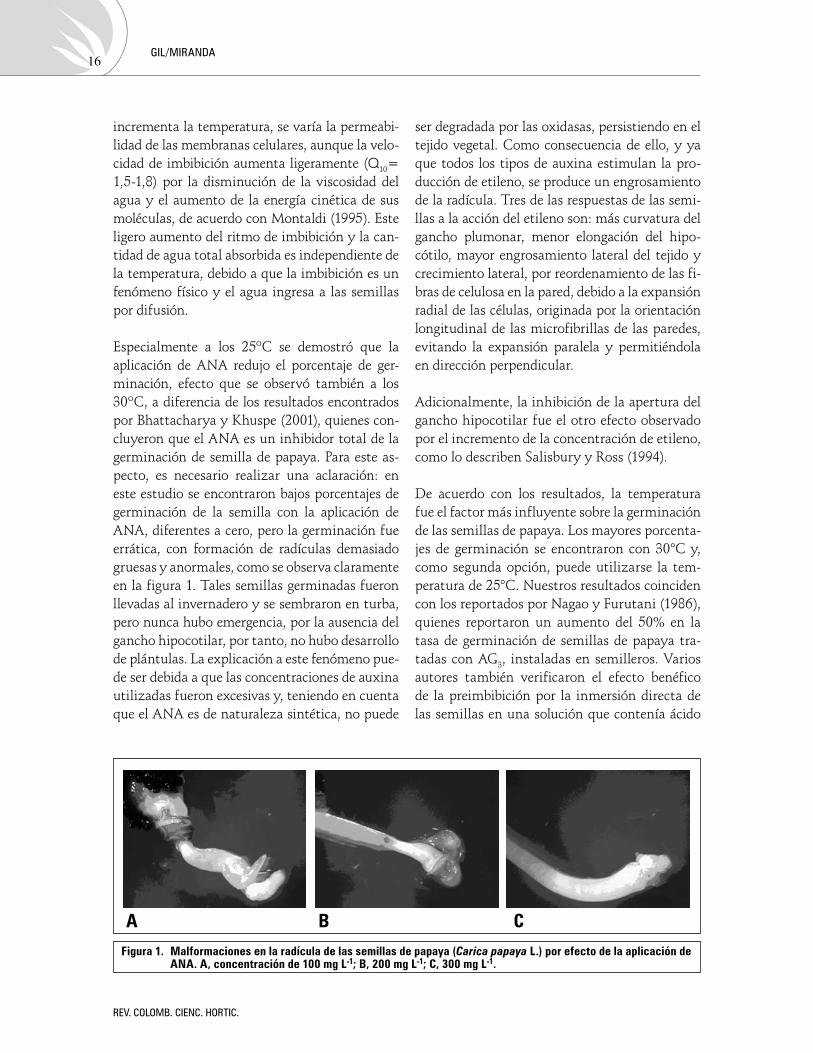
Especialmente a los 25ºC se demostró que la aplicación de ANA redujo el porcentaje de ger-minación, efecto que se observó también a los 30ºC, a diferencia de los resultados encontrados por Bhattacharya y Khuspe (2001), quienes con-cluyeron que el ANA es un inhibidor total de la germinación de semilla de papaya. Para este as-pecto, es necesario realizar una aclaración: en este estudio se encontraron bajos porcentajes de germinación de la semilla con la aplicación de ANA, diferentes a cero, pero la germinación fue errática, con formación de radículas demasiado gruesas y anormales, como se observa claramente en la figura 1. Tales semillas germinadas fueron llevadas al invernadero y se sembraron en turba, pero nunca hubo emergencia, por la ausencia del gancho hipocotilar, por tanto, no hubo desarrollo de plántulas. La explicación a este fenómeno pue-de ser debida a que las concentraciones de auxina utilizadas fueron excesivas y, teniendo en cuenta que el ANA es de naturaleza sintética, no puede
ser degradada por las oxidasas, persistiendo en el tejido vegetal. Como consecuencia de ello, y ya que todos los tipos de auxina estimulan la pro-ducción de etileno, se produce un engrosamiento de la radícula. Tres de las respuestas de las semi-llas a la acción del etileno son: más curvatura del gancho plumonar, menor elongación del hipo-cótilo, mayor engrosamiento lateral del tejido y crecimiento lateral, por reordenamiento de las fi-bras de celulosa en la pared, debido a la expansión radial de las células, originada por la orientación longitudinal de las microfibrillas de las paredes, evitando la expansión paralela y permitiéndola en dirección perpendicular.
Adicionalmente, la inhibición de la apertura del gancho hipocotilar fue el otro efecto observado por el incremento de la concentración de etileno, como lo describen Salisbury y Ross (1994).
De acuerdo con los resultados, la temperatura fue el factor más influyente sobre la germinación de las semillas de papaya. Los mayores porcenta-jes de germinación se encontraron con 30°C y, como segunda opción, puede utilizarse la tem-peratura de 25°C. Nuestros resultados coinciden con los reportados por Nagao y Furutani (1986), quienes reportaron un aumento del 50% en la tasa de germinación de semillas de papaya tra-tadas con AG3, instaladas en semilleros. Varios autores también verificaron el efecto benéfico de la preimbibición por la inmersión directa de las semillas en una solución que contenía ácido
GIL/MIRANDA
Figura 1. Malformaciones en la radícula de las semillas de papaya (Carica papaya l.) por efecto de la aplicación de AnA. A, concentración de 100 mg l-1; B, 200 mg l-1; C, 300 mg l-1.
A B C

Vol. 2 - No. 1 - 2008
17
giberélico (Rosseto et al., 2000). Estos resultados están de acuerdo con los encontrados por Bhatta-charya y Khuspe (2001), quienes evaluaron 10 cultivares de papaya, obteniendo los más bajos porcentajes de germinación (32%) a una tempe-ratura de 20ºC e incrementándose a un máximo de 80% con 30ºC. A temperaturas superiores de 30ºC, el porcentaje de germinación disminuyó cerca del 20%.
La explicación a estos resultados residiría en que, a una temperatura de 20°C, los complejos enzi-máticos no actúan eficientemente, debido a que no están activados por completo, a causa de la baja temperatura para esta especie. Para el ran-go de temperatura entre 25-30°C, los sistemas enzimáticos se activan y las enzimas actúan movilizando las reservas acumuladas durante el desarrollo del eje embrionario, ya que se ven fa-vorecidas por estas temperaturas. El ácido gibe-rélico, considerado activador enzimático endóge-no, promueve la germinación (Levitt, 1974). Por otra parte, las giberelinas están involucradas en el control de la hidrólisis de las reservas y en la síntesis de novo de alfa-amilasa, enzima respon-sable de la hidrólisis del almidón y su aplicación exógena influencia el metabolismo proteico, pu-diendo aumentar la tasa de síntesis de proteínas de las semillas (McDonald y Khan, 1983).
A los 35°C, la respuesta de las semillas empieza a disminuir, debido a que las enzimas comienzan su proceso de desnaturalización, provocado por la alta temperatura.
En cuanto a los tratamientos con agua y fitorre-guladores, se recomienda sumergir las semillas en agua durante 48 h, ya que se obtuvo el ma-yor porcentaje de germinación. Aunque los tra-tamientos con AG3 también fueron de alta res-puesta por parte de las semillas, las diferencias con los tratamientos de inmersión en agua no fueron significativas, por tanto se recomienda, no realizar una costosa inversión para la adqui-sición de la hormona, cuando se pueden reducir costos únicamente tratando a las semillas con
agua y teniendo en cuenta las temperaturas re-comendadas. Este resultado difiere del encontra-do por Bhattacharya y Khuspe (2001), quienes obtuvieron los mayores porcentajes de germina-ción con 200 mg L-1 de AG3.
De acuerdo con las evaluaciones, en los materia-les evaluados las concentraciones endógenas de fitohormonas en la semilla, posiblemente, son suficientes para llevar a cabo el proceso de ger-minación de manera exitosa. De acuerdo con Fa-rrant et al. (1993), durante la histodiferenciación, los contenidos de giberelinas, AIA y citoquininas son elevados, y posteriormente se observa un des-censo. Hay dos mecanismos principales que han sido documentados sobre el modo de acción de las giberelinas: el primero, podría ser el de inducir al endospermo y a la cubierta seminal a debilitar-se. Este proceso es requerido para la germinación de muchas especies, debido a que dichos tejidos confieren parte de la resistencia mecánica a la protrusión radicular. En el segundo mecanismo, las giberelinas podrían estar involucradas en la re-anudación de la actividad del ciclo celular durante la germinación, como se ha documentado en se-millas de tomate (Gallardo et al., 2002). Además, las giberelinas están implicadas en la moviliza-ción de las reservas acumuladas, para su posterior utilización durante la germinación. El hecho de que el ácido giberélico producido endógenamente pueda estar involucrado en la digestión del en-dospermo antes de la germinación fue indicado en estudios con mutantes de semillas de tomate (Groot y Karssen, 1987). En investigaciones en semillas del tomate mutante AG3-deficiente, no se produjo la endo-ß-mananasa necesaria para la digestión del endospermo, esto implica que la enzima fue inducida por ácido giberélico para la degradación y germinación. Se ha reportado que el ácido giberélico endógeno puede controlar la germinación de semillas de pimentón, tomate como también papaya (Andreoli y Khan, 1993). Al iniciar la germinación de la semilla, estos tres reguladores de crecimiento incrementan su con-centración y están implicados en los procesos de reanudación del metabolismo. La movilización
EFECTOS SOBRE LA GERMINACIóN DE SEMILLAS DE pApAyA

REV. COLOMB. CIENC. HORTIC.
18
reFerenCIAS BIBlIoGrÁFICAS
de las reservas sostiene el crecimiento de la plán-tula recién germinada, hasta que ésta es activa fotosintéticamente.
Se observaron diferencias en la respuesta de las se-millas de los dos materiales evaluados en cuanto a las temperaturas, siendo el híbrido Tainung-1 el que soporta las mayores temperaturas, respues-ta posiblemente relacionada con el vigor híbrido. No se observó fitotoxicidad en las estructuras de las semillas germinadas en ninguno de los dos materiales con las concentraciones mayores de ácido giberélico. De acuerdo con la respuesta las concentraciones endógenas de ácido giberélico en las semillas de los dos materiales evaluados son suficientes para promover la germinación.
ConClUSIoneS
Según los resultados obtenidos, la temperatura fue el factor más influyente sobre la germinación
de las semillas de papaya, siendo los 30°C el pun-to en el que se obtuvieron los mayores porcenta-jes de germinación, seguido de 25°C. Temperatu-ras superiores (35°C) o inferiores (20°C) fueron detrimentales para ambos materiales.
No se presentaron diferencias significativas para porcentaje de germinación entre los tratamientos de 48 h de inmersión de la semilla en agua y las concentraciones de ácido giberélico aplicados a las semillas. Este resultado lleva a recomendar que no se justifique económicamente una alta inversión en la adquisición de este fitorregulador, cuando se puede implementar un sistema fácil y económico, como lo es la inmersión de la semilla en agua por 48 h y temperaturas de 30°C y 25°C.
El ácido naftalenacético aplicado a las semillas de papaya, provocó malformaciones en las radí-culas e inhibió la formación del gancho hipoco-tilar, dando como resultado el impedimento del crecimiento de las plántulas.
GIL/MIRANDA
Andrade-Rodríguez, M.; J.J. Ayala-Hernández; I. Alia-Te-jacal; H. Rodríguez-Mendoza; C.M. Acosta-Durán y V. López-Martínez. 2008. Efecto de promotores de la germinación y sustratos en el desarrollo de plántu-las de papayo. Rev. Fac. Agron. LUZ 25(1), 617-635.
Andreoli, C. y A.A. Khan. 1993. Improving papaya seed-ling emergence by matriconditioning and gibberel-lin treatment. HortScience 28(7), 708-709.
Aragao, C.A.; B.F. Dantas; E. Alves; A.C. Cataneo; C. Ca-variani y J. Nakagawa. 2003. Atividade amilolítica e qualidade fisiológica de sementes armazenadas de Milho super doce tratadas com ácido giberélico. Rev. Bras. Sementes 25(1), 43-48.
Arteca, R.D. 1996. Plant growth substances: principles and applications. Chapman y Hall, New York, NY.
Besnier, F. 1988. Semillas: biología y tecnología. Edicio-nes Mundi-Prensa, Madrid.
Bevilaqua, G.A.P.; S.T. Peske, B.G. Santos-Filho y L. Bau-det. 1993. Desempenho de sementes de arroz irriga-do tratadas com regulador de crescimento. I. Efeito na emergência a campo. Rev. Bras. Sementes 15(1), 75-80.
Bewley, J.D. 1997. Seed germination and dormancy. Plant Cell 9, 1055-1066.
Bhattacharya, J. y S.S. Khuspe. 2001. In vitro and in vivo germination of papaya (Carica papaya L.) seeds. Scientia Hort. 91, 39-49.
Bradbeer, J.W. 1988. Seed dormancy and germination. Blackie Academic and Professional, Glasgow, UK.

Vol. 2 - No. 1 - 2008
19
Buckeridge, M.S.; D.C. Rocha; J.S.G Reid y S.M.C. Die-trich. 1992. Xylocglucan structure ans postgermi-native metabolism in seeds of Copaifera langsdor-fii from savanna and forest populations. Physiol. Plant. 86, 145-151.
Ellis, R.H.; T.D. Hong y E.H. Roberts. 1991. Effect of storage temperature and moisture on the germina-tion of papaya seeds. Seed Sci. Res. 1, 69-72.
Faostat. 2007. FAO statistics division. En: http://faostat .fao.org/site/567/DesktopDefault .aspx?PageID=567#ancor; consulta: enero de 2008.
Farrant, J.M.; N.W. Pammenter y P. Berjak. 1993. Seed development in relation to desiccation tolerance: a comparison between desiccation-sensitive (recal-citrant) seeds of Avicennia marina and desiccation-tolerant types. Seed Sci. Res. 3, 1-13.
Fosket, D.E. 1994. Plant growth and development: a mo-lecular approach. Academic Press, San Diego, CA.
Fry, S.C.; S. Aldington; P.R. Hetherington y J. Aitken. 1993. Oligosaccharides as signals and substrates in the plant cell wall. Plant Physiol. 103, 1-5.
Furutani, S.C. y M.A. Nagao. 1987. Influence of temper-ature, KNO3, GA3 and seed drying on emergence of papaya seedlings. Scientia Hort. 32(1-2), 67-72.
Gallardo, K.; C. Job; S.P.C. Groot; M. Puype; H. Demol; J. Vandekerckhove y D. Job. 2002. Proteomics of Arabidopsis seed germination: a comparative study of wild-type and gibberellin-deficient seeds. Plant Physiol. 129, 823-837.
Groot, S.P.C. y C.M. Karssen. 1987. Gibberellins regulate seed germination in tomato by endosperm weak-ening: a study with gibberellin-deficient mutants. Planta 171, 525-531.
Ha, M.A.; D.C. Apperley y M.C. Jarvis. 1997. Molecular rigidity in dry and hydrated onion cell walls. Plant Physiol. 115, 593-598.
Hartmann, H.T., D.E. Kester, F.T. Davies y R.L. Geneve. 2005. Plant propagation: principles and practices. 7th ed. Prentice Hall, Upper Saddle River, NY.
Hedden, P. y W.M. Proebsting. 1999. Genetic analysis of gibberellin biosynthesis. Plant Physiol. 119, 365-370.
Hilhorst, H.W.M. 1995. A critical update on seed dorman-cy. I. primary dormancy. Seed Sci. Res. 5, 61-73.
Hore, J.K. y S.K. Sen. 1993. Viability of papaya (Carica papaya L.) seeds under different pre-storage treat-ments. Environ. Ecol. 11(2), 273-75.
Lee, A.E. y C. Heimsch. 1962. Development and struc-ture of plants: a photographic study. Holt Rinehart and Winston, New York, NY.
Leonel, S. y J.D. Rodrigues. 1996. Germinação de semen-tes de limoeiro cravo. p. 167. En: Anais Congreso Brasileiro de Fruticultura 14, Sociedade Brasileira de Fruticultura. Curitiba, Brasil.
Levitt, J. 1974. Introduction to plant physiology. CV Mosby Company, Saint Louis, MO.
Lima, D.S., D.I. Lima, G.R. Velenzuela y P. Macias. 1985. Estudio de la viabilidad de la semilla de Carica pa-paya L. (variedad Maradol Roja). Centro Agrícola 12, 119-130.
Lima, D.U., de, y M.S. Buckeridge. 2001. Interaction between cellulose and storage xyloglucans: the in-fluence of degree of galactosylation. Car. Pol. 46, 157-163.
McDonald, M.B., Jr. and A.A. Khan. 1983. Acid scarifi-cation and protein synthesis during seed germina-tion. Agron. J. 75, 111-114.
Nishina, M.S.; M.A. Nagao y S.C. Furutani. 2004. Op-timizing germination of papaya seeds, fruits and nuts. Cooperative Extension Service, College of Tropical Agriculture and Human Resources, Uni-versity of Hawaii, Manova.
Mito, N. y A.B. Bennett. 1995. The diageotropica muta-tion and synthetic auxins differentially affect the expression of auxin-regulated genes in tomato. Plant Physiol. 109, 293-297.
Montaldi, E. 1995. Principios de fisiología vegetal. Edi-ciones Sur, La Plata, Argentina.
Nagao, M.A. y S.C. Furutani. 1986. Improving germi-nation of papaya seed by density separation, po-tassium nitrate, and gibberellic acid. HortScience 21(6), 1439-1440.
Navarrete, R.J.A. 1996. Producción del papayo (Carica papaya L.) variedad Maradol en Yucatán. Instituto Tecnológico Agropecuario. Centro de Investigación y Graduados Agropecuarios. Mérida, Yucatán, México.
Pereira de Araújo Filho, J. 1987. Cultivo de la papaya. pp. 117-122. En: Producción, manejo y exportación de frutas tropicales. Federacafé, Manizales, Colombia.
Rosseto, C.A.V.; R.C.C. Coneglian; J. Nakagawa; M.K. Shimizu y V.A. Marin. 2000. Germinação de se-mentes de maracujá-doce (Passiflora alata Dryand) em função de tratamento pré-germinativo. Rev. Brás. Sementes 22(1), 247-252.
EFECTOS SOBRE LA GERMINACIóN DE SEMILLAS DE pApAyA

REV. COLOMB. CIENC. HORTIC.
20
Salisbury, F.B. y C.W. Ross. 1994. Fisiología vegetal. Gru-po Editorial Iberoamérica, México D.F.
Salvador-Figueroa, M., M. de L. Adriano-Anaya y C. Becerra-Ortíz. 2005. Efecto del remojo en agua sobre la germinación de semillas de papaya var. Maradol. Revista Chapingo Serie Horticultura 11, 27-30.
Semicol. 2002. Catalogo de productos. En: www.semi-col.com.co; consulta: enero de 2005.
Thomson, J.R. 1979. Introducción a la tecnología de las semillas. Acribia, Zaragoza, España.
Weaver, R.J. 1987. Reguladores del crecimiento de las plantas en la agricultura. 5a. ed. Trillas, Mexico D.F.
Welbaum, G.E.; K.J. Bradford; Y. Kyu-Ock.; D.T. Booth y M.O. Oluoch. 1998. Biophysical, physiological and biochemical processes regulating seed germination. Seed Sci. Res. 8, 161-172.
Wood, C.B.; H.W. Pritchard y D. Amritphale. 2000. Des-iccation-induced dormancy in papaya (Carica pa-paya L.) seeds is alleviated by heat shock. Seed Sci. Res. 10, 135-145.
Yahiro, M. 1979. Effects of seed-pretreatments on the promotion of germination in papaya, Carica papa-ya L. Mem. Fac. Agr. Kagoshima Univ. 15, 49-54.
Yahiro, M. y Y. Oryoji. 1980. Effects of gibberellin and cytokinin treatments on the promotion of germina-tion in papaya, Carica papaya L., seeds. Mem. Fac. Agr. Kagoshima Univ. 16, 45-51.
GIL/MIRANDA

ronAlD r. MArtÍneZ-veGA1
GerHArD FISCHer1, 3
AnÍBAl HerrerA1
BernArDo CHAveS1
oMAr CAMIlo QUIntero2
Feijoas en fructificación. Foto: O.C. Quintero
Características físico-químicas de frutos de feijoa influenciadas por la posición en el canopi
Physico-chemical characteristics of pineapple guava fruits as influenced by canopi position
reSUMen
Con el fin de evaluar el efecto de la posición del fruto en el árbol de la feijoa cv. Quimba, se cosecharon frutos fisiológicamente maduros en cinco cuadrantes diferentes de la copa: (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior. El estudio se llevó a cabo en un huerto en las inmediaciones del municipio de la Vega (Cundinamarca, Colombia) a 2.350 msnm. Se observó que bajo un día nublado las diferencias en la intensidad lumínica, que se filtra a lo largo del dosel, son menos marcadas que en un día soleado. Las características evaluadas en los frutos fueron peso fresco, porcentaje de masa seca, pH, acidez total titulable (ATT), sólidos solubles totales (SST), relación de madurez y color. Los análisis mostraron que el peso fresco del fruto fue mayor en el cuadrante medio externo al igual que lo fue la relación de madurez en el superior; mientras el color de las feijoas fue más verde intenso en la base y en la parte interna de la copa. El porcentaje de masa seca, el pH, la concentración de SST y la ATT no se vieron mayormente afectadas por la posición del fruto. Se observó que los frutos del cuadrante medio externo presentaron buenas características en general. Los frutos del cuadrante superior mostraron mayor madurez pero menor peso.
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).2 Gerencia Disfruta las Feijoas, Bogotá (Colombia). 3 Autor para correspondecia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 21-32, 2008

REV. COLOMB. CIENC. HORTIC.
22
Palabras clave adicionales: intensidad lumínica, peso fruto, pH, grados Brix, acidez titulable, color.
ABStrACt
The influence of canopy position on fruit quality of pineapple guava ‘Quimba’ was evaluated in an orchard located near the municipality of La Vega (Cundinamarca, Colombia) at 2,350 m altitude above sea level. Physiologically mature fruits were harvested from five different canopy quadrants: 1) internal base, 2) external base, 3) mid-height internal, 4) mid-height external, and 5) top. Fruit characteristics evaluated were fresh weight (FW), dry matter (DM) content, pH, total titratable acidity (TTA), total soluble solids (TSS), maturity index and color. Greatest FW was observed from the external mid-height quadrant. The maturity index was enhanced in the superior quadrant, and color was intensified at the base and internal parts of the canopy. Percentage DM, pH, TSS and TTA were not highly affected by fruit position. It was observed that fruits on the external medium quadrant presented appropriate conditions in general but that that differences in the sunlight intensity filtering through the canopy were less marked on cloudy days than on sunny days. Fruits on the upper quadrant exhibited greater maturity but lower weight.
Additional keywords: light intensity, fruit weight, fruit Brix, fruit acidity, fruit color.
Fecha de recepción: 18-12-2007 Aprobado para publicación: 03-06-2008
La feijoa (Acca sellowiana [O. Berg] Burret, fami-lia Myrtaceae) es un arbusto subtropical con-siderado por mucho tiempo como una planta ornamental, es originaria del sur de América y se distribuye desde la zona meridional de Brasil y Paraguay hasta la zona de Chile, Argentina y Uruguay. Se caracteriza por su amplia capacidad de adaptación, encontrándose en Colombia en cultivos comerciales en zonas con altitudes sobre 1.800 y 2.650 msnm, temperaturas entre 13°C y 21°C (promedio 16°C), precipitación anual de 700 a 1.200 mm, una exposición a la radiación de 1.500 horas brillo solar/año y humedad relativa promedio del 70% (Fischer, 2003).
La variabilidad en la calidad de los frutos es principalmente el resultado de la competencia,
IntroDUCCIÓn
el tipo de rama que los sostiene, la posición del fruto en la rama (Farina et al., 2006), la posición en el dosel del árbol y el ambiente (Ferguson et al., 1999); adicionalmente, la calidad del fruto es fijada genéticamente (Mengel et al., 2001). La composición de los frutos es afectada por la re-lación hojas-fruto (Montanaro et al., 2006), la madurez y los niveles de luz bajo los que el fruto se desarrolla, los cuales afectan el tiempo de co-secha e influyen sobre la vida en almacenamien-to (Tombesi et al., 1993).
La luz desempeña un papel crítico en la interacción entre la posición del fruto en el árbol y su calidad (Senger, 2000) y es uno de los factores ambientales de mayor importancia, cuyo efecto sobre la cali-dad de los mismos no se ha evaluado. Según Rom
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

Vol. 2 - No. 1 - 2008
23
(1996), el proceso de la diferenciación floral no se presenta a niveles lumínicos por debajo del 30% dentro del árbol, en comparación con luz plena.
Larcher (2003) afirma que, para las plantas, la luz no es solamente una fuente de energía (efec-to fotoenergético), sino también funciona como un estímulo para procesos de desarrollo (efectos fotocibernéticos) y ocasionalmente puede ser un factor de estrés (efecto fotoinhibidor o fotodes-tructivo). Su efecto sobre el crecimiento de las plantas cultivables (Zhang y Xu, 2002), la den-sidad floral y el rendimiento en especies frutales (Farina et al., 2006) están relacionados casi direc-tamente con la cantidad de radiación intercep-tada, la cual es significativamente mayor que la radiación absorbida (Wang, 2001).
El dosel de la planta se caracteriza por un gra-diente vertical de luz, particularmente cuando la densidad es alta (Pons et al., 2001), producto del sombrío dentro del árbol y entre los árboles (Zhang y Xu, 2002), mostrando una disminución en la distribución de la irradiancia a través de los estratos de la planta como consecuencia de su ar-quitectura (Galvis et al., 1999). En la mayoría de los casos, dentro de un árbol hay más sombrío que entre diferentes árboles y, por tanto, las condicio-nes lumínicas que afectan una planta individual son en mayor grado determinadas por la estruc-tura propia de la misma, la forma de la copa y la elevación solar, principalmente en árboles ralos y jóvenes (Zhang y Xu, 2002). Así, por ejemplo, en la parte basal central de un árbol de duraznero se encontraron condiciones de sombra y los valores más bajos de radiación fotosintéticamente activa (RFA) (9-26%), mientras que la porción media presentó 19- 33% (Luchsinger et al., 2002).
Gariglio et al. (2007) afirman que en el mismo ci-clo de crecimiento es necesario un umbral mínimo de intensidad de luz en el interior del canopi para lograr el establecimiento de los frutos, el adecuado tamaño de los mismos al momento de la cosecha, una alta concentración de los sólidos solubles y una coloración para las exigencias comerciales.
Se han observado diferencias en varias espe-cies frutales (tamaño, color, contenido de azú-cares, acidez) entre frutos obtenidos de partes sombreadas y de partes expuestas de una mis-ma planta (Crisosto et al., 1997; Fischer, 2000; Senger, 2000; Snelgar et al., 1998). Murray et al. (2005) encontraron en ciruelas expuestas com-pletamente a la radiación solar un color rojo intenso, mayores periodos de maduración y al-macenamiento, menor porcentaje de materia fresca, menor firmeza, mayor concentración de sólidos solubles y menor acidez titulable en com-paración con frutos sombreados.
Para el caso de manzanas, los datos son con-tradictorios y dependientes de la variedad. Así, varios cultivares de manzana roja únicamen-te desarrollan el color rojo en el lado del fruto que está expuesto a la radiación solar (Nilsson y Gustavsson, 2007), siendo la luz indispensa-ble para la formación de los pigmentos antocia-ninos (Westwood, 1993; Stoll, 1997). Durante la maduración de las manzanas ‘Aroma’ en la periferia del dosel, ellas desarrollaron su color rojo típico, mientras las ubicadas en el interior permanecieron verdes. Además, los frutos de la periferia de la copa tuvieron mayor contenido de materia seca, concentración de sólidos solu-bles y azúcares solubles, y un nivel de la acidez titulable ligeramente más bajo en comparación con las manzanas del interior (Nilsson y Gus-tavsson, 2007). De igual forma, Durán (1982) encontró que a mayor radiación solar en las seis a ocho semanas precedentes a la recolección de la manzana, mayor es la cantidad de azúcares, la dureza de la pulpa y menor incidencia del par-deamiento interno.
Teniendo en cuenta las diferencias entre los fru-tos por ubicación en la copa, el objetivo de este trabajo fue evaluar el efecto de la posición del fruto en el árbol de feijoa sobre sus caracterís-ticas físico-químicas. Estos conocimientos per-mitirán establecer patrones de manejo de la copa del árbol para dirigir la producción hacía estratos con mayor rendimiento y calidad.
CARACTER ÍST ICAS F ÍS ICO-QUÍMICAS DE FRUTOS DE FE IjOA

REV. COLOMB. CIENC. HORTIC.
24
MAterIAleS Y MÉtoDoS
Los frutos de feijoa usados en este ensayo cre-cieron en la finca “El Cortijo”, localizada en in-mediaciones del municipio de la Vega (Cundina-marca), ubicada a 4º39’ N y 71º57’ W, con una altitud de 2.350 msnm, precipitación promedio anual de 1.423 mm, temperatura promedio anual de 17ºC, humedad relativa promedio de 85% y un brillo solar anual de 1.377 h.
Para el estudio se utilizaron 10 árboles del ‘Clon 41’ (‘Quimba’), de 6 años de edad, sembrados a una distancia de 4,0 x 2,5 m en un lote el cual presenta una pendiente de 25%. Las ramas de los tres pisos estuvieron agobiadas horizontalmente, obteniéndose una forma piramidal de la corona. Los frutos se cosecharon en la madurez fisioló-gica, la cual correspondió al punto de mayor de-sarrollo en tamaño (Rodríguez et al., 2006) y de fácil separación (fuerza de retención en el árbol equivalente a 2.000 kg-f).
Las determinaciones físico-químicas se llevaron a cabo en el laboratorio de análisis físico-quími-cos del Instituto Colombiano de Tecnología de Alimentos (ICTA) y en el laboratorio de Fisio-logía de Cultivos de la Facultad de Agronomía de la Universidad Nacional de Colombia, sede Bogotá.
El peso fresco y seco se midió con una balan-za Ohaus, AdventurerTM, con una precisión de 0,001 g; para la obtención del porcentaje de ma-teria seca se secaron los frutos en una estufa de secado WTB, Binder a 70ºC hasta alcanzar peso constante.
Para la determinación de los grados Brix se usó un refractómetro portátil Carl Zeiss; el pH con un potenciómetro Orion, modelo 420A; la aci-dez titulable se determinó mediante titulación con NaOH 0,1 N hasta pH 8,2 y se expresó como porcentaje de ácido cítrico. El color se midió con un colorímetro Minolta, modelo CR-300, utili-zando los parámetros del sistema de medición de
color Hunter (L, a, b) (Purvis y Barmore, 1981; Alonso y Granell, 1995) donde:
L: Correspondiente a brillo: valores de claridad o luminosidad que van desde negro (L=0) hasta blanco (L=100).
a: Correspondiente a la escala de colores verde a rojo, en la cual los valores verdes son negativos (-) y los valores rojos son positivos (+).
b: Correspondiente a la escala de colores azul a amarillo, en la cual los valores azules son negati-vos (-) y los valores amarillos son positivos (+) y se expresó mediante el índice de color 1000a/Lb (Jiménez-Cuesta et al., 1981).
La radiación incidente se midió con una barra cuántica Li-Cor, modelo Line Quantum, jun-to con un (Radiómetro/Quantum/Fotómetro) Li-Cor, modelo LI-189 en un día soleado y uno nublado en cinco árboles de la feijoa en los cua-drantes estudiados (tabla 1).
El diseño que se utilizó en este ensayo fue uno completamente al azar. Las variables de respues-ta se analizaron mediante el programa SAS (Sta-tistical Analysis System), realizando un análisis de varianza (Anova) y la prueba de comparación de medias de Tukey, además se diseñaron con-trastes ortogonales.
reSUltADoS Y DISCUSIÓn
Radiación incidente en la copa
La radiación incidente en el árbol se ve disminui-da a medida que se profundiza en el dosel del ár-bol, como consecuencia de la sombra que produ-cen las hojas y las ramas. Los datos de radiación incidente medidos en la feijoa van de acuerdo a lo reportado por Galvis et al. (1999), quienes di-vidieron el árbol de feijoa en tres estratos y re-portan menor irradiancia en el estrato bajo, se-guida por el estrato medio y el alto, donde hubo
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

Vol. 2 - No. 1 - 2008
25
la mayor incidencia lumínica. De igual forma, se observó que en un día nublado la diferencia de radiación incidente en cada uno de cuadrantes es menor que la encontrada para un día soleado, lo que concuerda con las mediciones de Awad et al. (2001). También Stoll (1997) afirma que la luz difusa tiene la capacidad de penetrar más al inte-rior de la copa que la directa.
Según los porcentajes de la radiación incidente (ta-bla 1) se pueden clasificar las copas de las feijoas del estudio como “coronas de sol” porque las hojas de la base interna recibieron 35 a 45% de la luz externa en un día soleado y nublado, respectiva-
mente. Larcher (2003) caracteriza como “coronas de sol” a aquellos árboles árboles en los cuales las hojas más internas de la copa reciben en promedio 10% a 20% de la luz externa. La mayor incidencia de la luz en este estudio se puede explicar debido a la formación piramidal de los árboles con tres pisos horizontales (agobio de las ramas laterales) y una constante eliminación de los chupones.
peso fresco del fruto
Los frutos que presentaron el mayor peso fresco fueron los tomados del cuadrante medio exter-no, con 52,14 g (figura 2), que percibió la segunda mayor radiación incidente (tabla 1).
Figura 1. Distribución de los cuadrantes en el árbol: (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior.
Base interna (Cuadr.1)
Base externa(Cuadr. 2)
Medio interno (Cuadr. 3)
Medio externo (Cuadr. 4)
Superior (Cuadr. 5)
plena exposición (encima de la copa)
Día soleado 35 54 70 81 90 100
Día nublado 45 48 60 65 85 100
tabla 1. Porcentaje de radiación incidente en cada uno de los cuadrantes dentro de la copa del árbol de feijoa, para un día soleado (en promedio 1.920 µmol m-2 s-1) y nublado (en promedio 348 µmol m-2 s-1).
CARACTER ÍST ICAS F ÍS ICO-QUÍMICAS DE FRUTOS DE FE IjOA

REV. COLOMB. CIENC. HORTIC.
26
El follaje ubicado en la parte media de la copa provee mejores condiciones para alcanzar tasas máximas de fotosíntesis, favoreciendo el llena-do de los frutos, comparado con la parte basal del árbol (Palmer, 1986). Por otro lado, el peso fresco del fruto fue la variable con más diferen-cias entre los cuadrantes de la copa, por lo cual se supone que el estado hídrico de la planta ha jugado un papel importante en este resultado. De acuerdo con Murray et al. (2005), el elevado peso fresco de estos frutos se podría explicar por la mayor tasa de transpiración relacionada con la irradiancia que se presentó en esta parte del ár-bol, lo que significaría una prolongada afluencia de agua y nutrientes al fruto. Esto sugiere que mayor disponibilidad lumínica incrementa y pro-longa el transporte de la corriente xilemática a estos órganos.
En el campo, los efectos de la luz y temperatura no son fáciles de separar porque, comúnmente, la intensidad lumínica incrementa la temperatu-ra (Westwood, 1993). Por tanto, se supone que el menor peso fresco de los frutos en la parte superior del árbol (figura 2) se debe también a efectos térmicos. Las temperaturas superiores a un umbral térmico suelen disminuir la trans-cripción, traducción y actividad de muchas en-zimas relacionadas con la maduración del fruto (Gariglio et al., 2007). El calentamiento de los frutos con la radiación se debe principalmente a su baja tasa transpiratoria, como midieron Gal-vis et al. (1999) en el estrato superior de árboles de feijoa y por la menor superficie específica del fruto respecto a las hojas para disipar calor (Ga-riglio et al., 2007). También Lakso (1986) afir-ma que superficies poco transpirantes, como el fruto, aumentan la temperatura. Por otro lado, el supuesto menor contenido de clorofila en los frutos del estrato superior (figura 5) conllevó a una reducida fotosíntesis y crecimiento de estos órganos, como lo reporta Proeitti (1990) en otro fruto verde como es el olivo.
Los pesos de los frutos en los cuadrantes de la base externa y del medio interno no presentaron
diferencias significativas, sin embargo la desvia-ción estándar fue alta en los frutos de todos los cuadrantes, principalmente del cuadrante 2 (base interna).
Los frutos de la base interna presentaron los valo-res más bajos de peso fresco (21,12 g), que puede explicarse por la baja incidencia de la luz en este estrato, siendo el factor lumínico fundamental para una adecuada fotosíntesis y producción de fotoasimilados para el llenado del fruto. Por otro lado, la menor carga de frutos en este estrato po-dría haber reducido la fotosíntesis foliar, debido a que las hojas adyacentes a los frutos exhiben una capacidad fotosintética más alta que las otras ho-jas del canopi (Urban et al., 2003), y así la pro-ducción y traslocación de carbohidratos a estos vertederos (Lakso, 1986; Lakso y Flore, 2001).
Murray et al. (2005) encontraron que duraznos sombreados por 6 semanas hasta cosecha tuvie-ron mayor reducción en el tamaño del fruto en comparación con frutos sombreados por 3 sema-nas antes de cosecha, concluyendo que la luz es más importante durante la segunda mitad del estado III de desarrollo frutal.
porcentaje de materia seca del fruto
El porcentaje de materia seca (MS) en los fru-tos osciló entre 12,69 y 13,23 y según el análisis de varianza no se presentaron diferencias sig-nificativas en esta variable entre los diferentes cuadrantes del dosel del árbol y del análisis de contrastes, como lo explican Montanaro et al. (2006), quienes encontraron en kiwi que el som-breado no afectó la acumulación total de MS, de-bido a que los asimilados pueden ser fácilmente traslocados por el floema y dirigirse hacía partes no sombreadas.
En otros casos, hubo una mayor acumulación de MS en frutos expuestos directamente a la luz, indicando la importancia del estado hídrico en estos órganos. Jifon y Syvertsen (2001), en na-ranja ‘Hamlin’ y Murray et al. (2005), en ciruelos
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

Vol. 2 - No. 1 - 2008
27
japoneses, encontraron que frutos no sombrea-dos presentaron mayor tamaño y relación MS/agua, por cuanto la acumulación de materia seca, según los autores, depende del aporte que realiza el flujo del floema, mientras la acumulación de la materia fresca depende tanto del flujo del floema como del xilema y las tasas de transpiración.
pH del fruto
El pH de los frutos mostró poca variación (2,88-2,97) y según el análisis de varianza no hubo dife-rencias significativas entre los valores obtenidos. Sin embargo, el análisis de contrastes mostró di-ferencias significativas (P≤0,05) en el pH de los frutos tomados de la parte baja (2,96) con los cre-cidos en la parte media del dosel (2,90); también se presentaron diferencias significativas (P≤0,05) entre los frutos de la parte media de la copa (2,90) con los de la parte alta (2,97). La radiación solar directa parece tener un efecto sobre el grado de desarrollo del pH. Hunter y Archer (2002) afir-man que para la uva la exposición del follaje debe
ser alta, con el objeto de limitar la carga de pota-sio en la baya y, en consecuencia, la formación de sales del ácido tartárico y aumento del pH.
Sólidos solubles totales (SST) del fruto
Los valores de los SST medidos oscilaron entre 10,35 y 11,63 ºBrix, dependiendo del cuadrante del dosel. Los frutos tomados de la parte media interna (10,35 ºBrix, promedios de los cuadran-tes 3 y 4) tuvieron un valor significativamente menor (P≤0,01) que los que crecieron en otras partes de la copa (figura 3). En cuanto al análisis de contrastes, se observó que hubo diferencias altamente significativas (P≤0,01) entre los fru-tos de la parte interior (10,75 ºBrix, cuadrantes 1 y 3) y los de los cuadrantes exteriores (2, 4 y 5) que mostraron en promedio de 11,45 ºBrix. Por otro lado, también se observaron diferencias al-tamente significativas (P≤0,05) entre los frutos de la parte media del dosel (10,84 ºBrix) y los de la parte superior (11,63 ºBrix), presentando esta última el más alto valor de los SST en la copa.
Figura 2. Peso fresco de los frutos de feijoa dentro de cada cuadrante de la copa, (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior). Promedios con letras distintas son significativamente diferentes según la prueba de tukey (P≤0,05).
CARACTER ÍST ICAS F ÍS ICO-QUÍMICAS DE FRUTOS DE FE IjOA

REV. COLOMB. CIENC. HORTIC.
28
Una de las variables que se podría esperar tu-viera las mayores variaciones es el porcentaje de sólidos solubles, debido principalmente a la in-fluencia de la intensidad lumínica en la activi-dad fotosintética del fruto verde (como la feijoa), contribuyendo directamente en su composición química (Gross, 1987). También Murray et al. (2005) afirman que la mejora en la concentración de sólidos solubles se puede atribuir a una mayor fotosíntesis en hojas adyacentes y, por ende, una mayor disponibilidad de carbohidratos para los frutos en desarrollo, explicando las mayores con-centraciones de los SST en las partes exteriores y superiores de la copa de feijoa; sin embargo, la concentración de los SST en chirimoya no fue influenciada por la posición del fruto en el árbol (González y Cuevas, 2008). Posiblemente, tam-bién, aparte del estado de desarrollo del fruto (Stoll, 1997), existe un efecto genético sobre la formación de los grados Brix.
Acidez total titulable (ATT) del fruto
El análisis de varianza de la ATT no mostró di-ferencias significativas entre los frutos de cada
cuadrante; sin embargo, el análisis de contrastes arrojó diferencias significativas entre los frutos de los dos cuadrantes bajos (2,28%) con los que crecieron en la parte alta del dosel (2,07%), ade-más entre los frutos de la parte media (2,25%) y los frutos de la parte alta (2,07%). Posiblemente, en la parte sombreada del árbol los frutos demo-ran más tiempo en la degradación de los ácidos (Stoll, 1997), debido a que la sombra y la supues-ta menor temperatura en estos estratos originan una madurez fisiológica retardada que en aque-llos que crecen en las partes de la copa directa-mente expuestas a la luz
Por el contrario, González y Cuevas (2008) ob-servaron frutos de chirimoya, formados en las partes exteriores del dosel y posicionados hacia los ápices de las ramas, con una mayor acidez que los situados en el interior de la copa, coinci-diendo con lo reportado por Heinicke (1966) en manzano. Posiblemente, la formación de la ATT en el fruto no depende solamente de la posición en el árbol, sino también de factores como el es-tado de madurez, su color, la intensidad lumíni-ca y otros factores genéticos y varietales.
Figura 3. Concentración de sólidos solubles totales (grados Brix) de los frutos de feijoa dentro de cada cuadrante de la copa, (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior. Promedios con letras distintas son significativamente diferentes según la prueba de tukey (P≤0,05).
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

Vol. 2 - No. 1 - 2008
29
Relación de madurez (RM) del fruto
Como muestra la figura 4, la RM (SST/ATT) en frutos de la posición media interna (4,73) de la copa fue la más baja y presentó un valor signi-ficativamente menor (P≤0,01) que la RM de la parte superior del dosel (5,73). Con respecto a la prueba de contrastes, se observó que hubo dife-rencias significativas (P≤0,05) de los frutos de la parte interior del árbol (4,86, promedio de los cuadrantes 1 y 3) con respecto a los de la parte exterior (5,28, cuadrantes 2, 4 y 5). De igual ma-nera, se presentaron diferencias altamente sig-nificativas (P≤0,01) entre los frutos de la parte baja (5,01) y los del dosel medio (4,91) y entre estos con los frutos del dosel superior (5,73).
La relación de madurez sirve para medir la ca-lidad organoléptica de los frutos (Rodríguez et al., 2006). Menores porcentajes de ácido cítrico (2,07%) y mayores en SST (11,83 °Brix) del cua-drante superior en relación con los frutos de los demás cuadrantes, explica porqué estos frutos tienden a presentar los valores más altos de rela-
ción de madurez como frutos más dulces de esta posición de la copa. Por otro lado, en el medio interno de la copa se encontraron las RM más bajas, que se dieron como consecuencia de los bajos niveles de sólidos solubles en este lugar. La poca diferencia en la ATT y la RM entre los dife-rentes cuadrantes de la copa comprueba que los factores que controlan la acidez en el fruto son en mayor grado fijados genéticamente (Mengel et al., 2001).
Color del fruto
La prueba de Tukey mostró diferencias (P≤0,05) en el color de la cáscara de los frutos entre el cua-drante medio interno, y el superior (figura 5). Los frutos de la parte superior mostraron un índice de color menos negativo, lo que indica un verde más amarillento en comparación con los frutos del cua-drante medio interno que desarrollaron un verde más profundo. También en la prueba de contrastes se observaron diferencias significativas (P≤0,05) en el color entre los frutos de la parte media y los frutos del cuadrante superior. Igualmente, entre
Figura 4. relación de madurez (SSt/Att) de los frutos de feijoa dentro de cada cuadrante de la copa, (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior. Promedios con letras distintas son significativamente diferentes según la prueba de tukey (P≤0,05).
CARACTER ÍST ICAS F ÍS ICO-QUÍMICAS DE FRUTOS DE FE IjOA

REV. COLOMB. CIENC. HORTIC.
30
los frutos de la parte baja y los frutos del dosel alto hubo diferencias altamente significativas (P≤0,01) en esta variable. Este resultado confirma que una radiación muy intensa durante el periodo de ma-duración incrementa la formación de pigmentos amarillos y, al mismo tiempo, acelera la degrada-ción de la clorofila (Stoll, 1997), observada en la parte superior de la copa de feijoa.
El color verde más intenso de los frutos sombrea-dos indica que sus cloroplastos tuvieron un may-or contenido de clorofila y una menor cantidad de compuestos asociados con la fotosíntesis com-parado con los frutos más iluminados de la copa (Pons et al., 2001). Estos autores afirman que la clorofila se incrementa en órganos fotosintéticos sombreados, lo que indica aclimatación de los cloroplastos a condiciones de baja luminosidad. La gran mayoría de trabajos de investigación muestra que los frutos de las partes internas del árbol permanecen con color más verde debido a la mayor síntesis de pigmentos diferentes a la
clorofila en presencia de alta luminosidad (Noè y Eccher, 1996; Awad et al., 2001; Murray et al., 2005).
Como complemento a estos estudios, Kasper-bauer (1971), Noè y Eccher (1996) y Awad et al. (2001) encontraron que diferentes longitudes de onda se filtran a lo largo del dosel, siendo posi-blemente este diferencial de calidad en la luz el responsable de las variaciones que se presentan en los frutos dentro del árbol.
Otras causas de las variaciones en los frutos dentro del árbol se deben a la señalización por medio de hormonas, que se relocalizan como consecuencia de las variaciones en el microam-biente dentro del dosel, lo cual fue verificado por Pons et al. (2001), quienes lograron contrar-restar los reducidos niveles de nitrógeno foliar y la capacidad fotosintética de hojas sometidas a sombrío por medio de aplicaciones de bencila-minopurina.
Figura 5. Índice de color de los frutos de feijoa dentro de cada cuadrante de la copa, (1) base interna, (2) base externa, (3) medio interno, (4) medio externo y (5) superior. Promedios con letras distintas son significativamente diferentes según la prueba de tukey (P≤0,05).
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

Vol. 2 - No. 1 - 2008
31
ConClUSIoneS
• Debido a que la radiación solar incidente ilumi-na menos la parte interior de la copa, algunas de las variables productivas y cualitativas del fruto de feijoa son afectadas, como se encontró para el peso fresco y los grados Brix, mientras la acidez total titulable se incrementó ligera-mente en las partes menos iluminadas.
• Posiblemente, la exposición directa a la luz in-cidente en la parte superior del dosel contribu-
ye a un mayor contenido de sólidos solubles en estos frutos, conllevando a una elevada rela-ción de madurez, pero, a su vez, disminuyendo la intensidad del color verde y el peso fresco de estos órganos en dicha posición.
• En futuros estudios sobre el tema se recomien-da complementarlos con mediciones de foto-síntesis y transpiración, además con evalua-ciones en la fase posrecolección para encontrar posibles relaciones entre la posición del fruto y su calidad y longevidad en poscosecha.
reFerenCIAS BIBlIoGrÁFICAS
Alonso, J.M. y A. Granell. 1995. A putative vacuolar pro-cessing protease is regulated by ethylene and also during fruit ripening in citrus fruit. Plant Physiol. 109, 541-547.
Awad, M.A.; P.S. Wagenmakers y A. Jager. 2001. Effects of light on flavonoid and chlorogenic acid levels in the skin of ‘Jonagold’ apples. Scientia Hort. 88, 289-298.
Crisosto, C.H.; J.R. Scott; T. DeJong y K.R. Day. 1997. Orchard factors affecting postharvest stone fruit quality. HortScience 32(5), 820-823.
Duran, S. 1982. Frigoconservación de la fruta. Editorial Aedos, Barcelona, España.
Farina, V.; R. Lo Bianco y P. Inglese. 2006. Shoot growth, crop load, and fruit quality within vase-shaped canopies of ‘Fairtime’ peach trees. Europ. J. Hort. Sci. 71(5), 227-230.
Ferguson, I.; R. Volz y A. Woolf. 1999. Preharvest factors affecting physiological disorders of fruit. Postharv. Biol. Technol. 15, 255-262.
Fischer, G. 2003. Ecofisiología, crecimiento y desarrollo de la feijoa. pp. 9-26. En: Fischer, G., D. Miranda, G. Cayón y M. Mazorra (eds.). Cultivo, poscosecha y exportación de la feijoa (Acca sellowiana Berg). Produmedios, Bogotá.
Fischer, G. 2000. Efectos de las condiciones en precose-cha sobre la calidad poscosecha de los frutos. Rev. Comalfi 27(1-2), 39-50.
Galvis, A.; M.S. Hernández y G. Fischer. 1999. Transpi-ración de la feijoa (Acca sellowiana Burret) en la Sa-bana de Bogotá. Rev. Comalfi 26(1-2), 56-61.
Gariglio, N.F.; R.A. Pilatti y M. Agustí. 2007. Re-querimientos ecofisiológicos de los árboles frutales. pp. 41-103. En: Sozzi, G.O. (ed.). Árboles frutales: ecofisiología, cultivo y aprovechamiento. Editorial Facultad de Agronomía, Universidad de Buenos Ai-res, Buenos Aires.
González, M. y J. Cuevas. 2008. Optimal crop load and positioning of fruit in cherimoya (Annona cherimola Mill.) trees. Scientia Hort. 115(2), 129-134.
Gross, J. 1987. Pigments in fruits. Academic Press, London.
Heinicke, D.R. 1966. Characteristics of ’McIntosch’ and ’Red Delicious’ apples as influenced by exposure to sunlight during the growing season. Proc. Amer. Soc. Hort. Sci. 89, 10-13.
Hunter, J.J. y E. Archer. 2002. Papel actual y perspectivas fu-turas de la gestión del follaje. En: ACE Revista de Eno-logía, http://www.acenologia.com/ciencia59_2.htm; consulta: 8 de abril de 2008.
Jifon, J.L. y J.P. Syvertsen. 2001. Effects of moderate shade on citrus leaf gas exchange, fruit yields, and quality. Proc. Fla. State Hort. Sci. 114, 177-181.
Jiménez-Cuesta, M.; J. Cuquerella and J.M. Martínez-Jávega. 1981. Determination of a color index for cit-rus degreening. Proc. Intl. Soc. Citricult. 2, 750-753.
CARACTER ÍST ICAS F ÍS ICO-QUÍMICAS DE FRUTOS DE FE IjOA

REV. COLOMB. CIENC. HORTIC.
32
Kasperbauer, M.J. 1971. Spectral distribution of light in a tobacco canopy and effects of end-of-day light quality on growth and development. Plant Physiol. 47, 775-778.
Lakso, A.N. 1986. Photosynthesis in fruit trees in rela-tion to environmental factors. pp. 6-13. En: Lakso, A.N. y F. Lenz (eds.). The regulation of photosyn-thesis in fruit trees. N.Y. Agr. Exp. Sta. Spec. Publ., Cornell University, Geneva, NY.
Lakso, A.N. y J.A. Flore. 2001. Carbohydrate partitioning and plant growth. pp. 21-30. En: Baughter, T.A. y S. Singha (eds.). Concise encyclopedia of temperate tree fruit. Food Products Press, New York, NY.
Larcher, W. 2003. Physiological plant ecology. 4th ed. Springer, Berlín.
Luchsinger, L.; P. Ortin; G. Reginato y R. Infante. 2002. Influence of canopy fruit on the maturity and qual-ity of ‘Angelus’ peaches. Acta Hort. 592, 515-521.
Mengel., K.; E.A. Kirkby; H. Kosegarten y T. Appel. 2001. Principles of plant nutrition. Kluwer Aca-demic Publ., Dordrecht, The Netherlands.
Montanaro, G.; B. Dichio; C. Xiloyannis y G. Celano. 2006. Light influences transpiration and calcium accumulation in fruit of kiwifruit plants (Actinidia deliciosa var. deliciosa). Plant Sci. 170, 520-527.
Murray, X.J.; D.M. Holcroft; N.C. Cook y S.J.E. Wand. 2005. Postharvest quality of ‘Laetitia’ and ‘Son-gold’ (Prunus salicina Lindell) plums as affected by preharvest shading treatments. Postharv. Biol. Technol. 37, 81-92.
Nilsson, T. y K. Gustavsson. 2007. Postharvest physiol-ogy of ‘Aroma’ apples in relation to position on the tree. Postharv. Biol. Technol. 43, 36-46.
Noè, N. y T. Eccher. 1996. ‘Golden Delicious’ apple fruit shape and russeting are affected by light condi-tions. Scientia Hort. 65, 209-213.
Palmer, J.W. 1986. Seasonal variation of light saturated photosynthesic rate of Golden Delicious apple leaves as influenced by leaf type and crop load. pp. 30-33. En: Lakso, A.N. y F. Lenz (eds.). The regula-tion of photosynthesis in fruit trees. N.Y. Agr. Exp. Sta. Spec. Publ.,Cornell University, Geneva, NY.
Pons, T.L.; W. Jordiy y D. Kuiper. 2001. Acclimation of plants to light gradients in leaf canopies: evidence for
a possible role for cytokinins transported in the tran-spiration stream. J. Exp. Bot. 52(360), 1563-1574.
Proeitti, P. 1990. Photosynthesis and respiration in olive fruit. Acta Hort. 286, 211-214.
Purvis, A.C. y C.R. Barmore. 1981. Involvement of eth-ylene in chlorophyll degradation in peel of citrus fruits. Plant Physiol. 68, 854-856.
Rodríguez, M.; H.E. Arjona y J.A. Galvis. 2006. Madura-ción del fruto de feijoa (Acca sellowiana Berg) en los clones 41 (Quimba) y 8-4 a temperatura ambiente en condiciones de Bogotá. Agron. Colomb. 42(1), 68-76.
Rom, C.R. 1996. Environmental factors regulating growth; light, temperature, water and nutrition. pp. 11-30. En: Maib, K.M., P.K. Andrews, G.A. Lang y K. Mullinix (eds.). Tree fruit physiology; growth and development. Washington State Fruit Commis-sion (publ. by Good Fruit Grower), Washington.
Senger, K.A. 2000. Effects of light availability and cano-py position on peach fruit quality. Master Thesis. Virginia Polytechnic Institute and State University, Blacksburg, VA.
Snelgar, W.P.; G. Hopkirk; R.J. Seelye; P.J. Martin y P.J. Manson. 1998. Relationship between canopy den-sity and fruit quality of kiwifruit. New Zealand J. Crop Hort. Sci. 26, 223-232.
Stoll, K. 1997. Der Apfel. Enrico Negri, Zürich, Suiza.
Tombesi, A.; E. Antognozzi y A. Pallioti. 1993. Influence of light exposure on characteristics and storage life of kiwifruit. N.Z. J. Crop Hort. Sci. 21, 87-92.
Urban, L.; X. Le Roux; H. Sinoqut; S. Jaffuel y M. Jan-noyer. 2003. A biochemical model of photosynthe-sis for mango leaves: evidence for the effect of fruit on photosynthetic capacity of nearby leaves. Tree Physiol. 23(5), 289-300.
Wang, Y.P. 2001. Modelling radiation absorption by plant canopies. pp. 30-32. En: Kirschbaum, M.U.F. y R. Mueller (eds.). Net ecosystem exchange. Co-operative Research Centre for Greenhouse Ac-counting, Canberra.
Westwood, M.N. 1993. Temperate-zone pomology: Phys-iology and culture. Timber Press, Portland, OR.
Zhang, X-Q. y D. Xu. 2002. Modeling radiation transfer within the canopy of a Chinese fir plantation. For. Ecol. Manage. 170, 107-116.
MARTÍNEZ-VEGA/FISCHER/HERRERA/CHAVES/QUINTERO

FÁnor CASIerrA-PoSADA1, 2
rICArDo A. SAlAMAnCA G.1
Plantación nueva de fresa en Arcabuco, Boyacá (Colombia).Foto: F. Casierra-posada
Influencia del ácido giberélico y del nitrato de calcio sobre la duración poscosecha de frutos de fresa (Fragaria sp.)
Influence of gibberellin and calcium nitrate on post-harvest conservation of strawberry (Fragaria sp.) fruits
1 Grupo de Investigación Ecofisiología Vegetal, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja (Colombia).
2 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 33-42, 2008
reSUMenLos factores precosecha, como el material genético, las condiciones ambientales durante el cultivo, la nutri-ción y el manejo de cultivo tienen una marcada influencia sobre el comportamiento poscosecha de frutas, especialmente debido a su carácter perecedero. Este trabajo se realizó en un cultivo a campo abierto en Tuta, Colombia, con plantas de fresa de los cultivares Chandler y Sweet Charlie. Se asperjaron 15 días luego de la antesis y al momento de la cosecha con diferentes concentraciones de ácido giberélico y de nitrato de calcio, con miras a determinar su efecto sobre la pérdida de peso y la vida en poscosecha de los frutos. Los tratamien-tos comprendieron un testigo sin aplicación y 500, 1.500 y 3.000 mg L-1 de ácido giberélico ó 50, 150 y 200 mg L-1 de nitrato de calcio aplicados a las hojas y frutos de las plantas. Al momento de la cosecha se realizaron pruebas de calidad en la determinación de sólidos solubles totales, acidez titulable y peso del fruto. En otros frutos se determinó la pérdida diaria, como la pérdida máxima de peso. Los resultados no mostraron ningún efecto de los productos aplicados o de sus concentraciones sobre la calidad del fruto. Los frutos de ‘Chandler’ se conservaron por mayor tiempo en poscosecha que los de ‘Sweet Charlie’. Los frutos tratados con nitrato de calcio duraron 16,4% más tiempo, mientras que los aplicados con ácido giberélico duraron 11,6% más que los testigos. La pérdida de peso fue proporcional a la duración en poscosecha. Comparando los resultados, se observó una respuesta de los frutos a los tratamientos, fuertemente dependiente del cultivar.

REV. COLOMB. CIENC. HORTIC.
34
Palabras clave adicionales: pérdida de peso, calidad de fruta, Chandler, Sweet Charlie.
ABStrACtPre-harvest factors, such as genetic background, environmental conditions during cultivation, nutrition and crop handling, noticeably influence the post-harvest condition of perishable fruits. This study was carried out in an open field crop of strawberries in Tuta, Colombia to evaluate effects of gibberellin and calcium nitrate on both weight loss and fruit shelf life. Leaves and fruits of ‘Chandler’ and ‘Sweet Charlie’ plants were sprayed with 500, 1,500 and 3,000 ppm gibberellin and 50, 150 and 200 ppm calcium nitrate 15 days after anthesis and at harvest. Total fruit soluble solids, titratable acidity and fruit fresh weight were evaluated post-harvest and daily weight loss and maximal post-harvest life were assessed for each treatment. The results showed no effect of the sprayed products on fruit quality. However, the fruits of cv. Chandler lasted longer during post-harvest than those of Sweet Charlie. Also, fruits treated with calcium nitrate lasted 16,4% longer, while gibberellin solutions prolonged their post-harvest life only 11,6% in comparison to control fruits. Post-harvest weight loss was directly proportional to post-harvest time. The response of fruit to treatments was strongly dependent on the cultivar.
Additional keywords: weight loss, fruit quality, Chandler, Sweet Charlie.
Fecha de recepción: 25-03-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
A diferencia de los países de la zona templada, donde la producción se limita al verano, en Co-lombia es posible la producción de fresa (Fragaria sp.) durante todo el año, con oferta alta de no-viembre a febrero y junio a septiembre y oferta media en octubre y de marzo a mayo (Corabas-tos, 2004). En 2006, se cultivaron en Colombia 703 ha de fresa en los departamentos más pro-ductores, Cundinamarca, Antioquia y Cauca, con un rendimiento promedio de 24 t ha-1 a nivel nacional (Agronet, 2008).
La fresa representa un cultivo de alto valor con una vida poscosecha bastante corta, que requiere de tratamientos de poscosecha para prolongar su vida útil (Esmel et al., 2004). El color, el tama-ño, la firmeza, la acidez y el sabor dulce hacen de la fresa una de las frutas más populares durante la primavera y el verano en países de zona tem-plada. A pesar de esto, algunos consumidores se sienten insatisfechos con su sabor y con la calidad de algunos tipos de fresa (Azodanlou et al., 2003). Por este motivo, se han realizado ensayos, en los
CASIERRA-pOSADA/SALAMANCA

Vol. 2 - No. 1 - 2008
35
cuales, mediante la aplicación en precosecha de fuentes de calcio y reguladores del crecimiento, se busca mejorar la calidad de los frutos. Se han utilizado muchos reguladores de crecimiento con el fin de incrementar el tamaño de la fruta (Guar-diola y García-Luis, 2000; Stern et al., 2007). La eficiencia de estos productos es fácilmente eva-luable con respecto al tamaño de la fruta, pero es necesario conocer su influencia en las caracterís-ticas bioquímicas y de calidad en la misma.
El papel de las auxinas sobre el desarrollo de frutos de fresa está ampliamente reportado. Las auxinas son las responsables del crecimiento del receptáculo floral y, por tanto, del incremento del tamaño de los frutos. En fresa, Nitsch (1950) demostró que los compuestos hormonales pro-ducidos durante el desarrollo de las semillas esta-ban asociados con el crecimiento del fruto.
Se reportan algunos efectos positivos de las apli-caciones de ácido giberélico sobre la producción de fruta, el acortamiento del periodo entre la siembra y la primera fructificación, el número de frutos y la duración del periodo de cosecha; sin embargo, esta hormona también puede reducir el peso de la fruta (Singh et al., 1960; Tehrani-far y Battey, 1997). Choma y Himelrick (1984) observaron que las aplicaciones de ácido gibe-rélico a cultivares con diferente sensibilidad al fotoperiodo incrementaban el peso y número de frutos. A pesar de esto, se puede esperar una res-puesta contraria de las plantas cuando se usan dosis muy altas de la hormona (Dennis y Ben-nett, 1969; Weidman y Stang, 1983; Tehranifar y Battey, 1997).
El calcio se aplica comúnmente en varios cultivos frutícolas con miras a mantener o incrementar la calidad de los frutos. Se ha aplicado a fresas del cultivar Sweet Charlie tiosulfato de calcio para lograr un incremento en la vida poscosecha de los frutos (Esmel et al., 2004). El calcio ha sido reportado ampliamente como un elemento esen-cial que desempeña un papel importante en el mantenimiento de la calidad poscosecha en cul-
tivos hortofrutícolas. El efecto del calcio sobre el comportamiento poscosecha de productos agrícolas se atribuye, en particular, a su función como estabilizador de la membrana celular (Kir-kby y Pilbeam, 1984).
El objetivo del presente trabajo fue evaluar el efecto de la aplicación de diferentes dosis de áci-do giberélico y de nitrato de calcio, realizadas en la mitad del periodo de desarrollo de los frutos y al momento de la cosecha, sobre la vida poscose-cha de frutos de dos cultivares de fresa.
MAterIAleS Y MÉtoDoS
Este ensayo se realizó en Tuta, Colombia, pobla-ción ampliamente reconocida por ser productora de fresa, ubicada a 05º 41’ 36” N, 73º 13’ 51” W y 2.600 m de altura sobre el nivel del mar. La temperatura media es de 14ºC y la precipitación anual de 935 mm. Como material vegetal se uti-lizaron plantas de fresa (Fragaria sp.) de 18 me-ses de los cultivares Chandler y Sweet Charlie, dado que son cultivares comúnmente sembrados en la zona. Las plantas fueron seleccionadas, rotuladas y se les eliminó la fructificación exis-tente al iniciar el ensayo. Al inicio del trabajo se marcaron 150 flores en diferentes plantas y los tratamientos respectivos se realizaron a los 15 días después de la antesis y al momento de la co-secha. Estas fechas se determinaron basadas en que en la región los frutos duran 28-30 días desde la apertura de la flor hasta el punto de cosecha. Los tratamientos consistieron en la aplicación foliar de las soluciones acuáticas del ácido (Pro-gibb®, Bayer CropScience) en concentraciones de 0, 500, 1.500 ó 3.000 mg L-1 y nitrato de calcio (Ca(NO3)2 4H2O, Merck) en concentraciones de 0, 50, 150 ó 200 mg L-1.
Al momento de la cosecha se realizaron las prue-bas de calidad en 50 frutos con madurez de cose-cha (fruto con el 80% de color rojo), que consis-tieron en determinar los sólidos solubles totales (ºBrix), la acidez total titulable y el peso fresco
INFLUENCIA DEL áC IDO G IBERéL ICO y DEL N ITRATO DE CALC IO

REV. COLOMB. CIENC. HORTIC.
36
del fruto. Los frutos restantes se llevaron al La-boratorio de Fisiología Vegetal de la Universidad Pedagógica y Tecnológica de Colombia (Tunja) y se mantuvieron en un ambiente a temperatu-ra promedio del 15±2°C y humedad relativa de 71%±5% hasta que los frutos mostraran sínto-mas de senectud y perdieran por completo su calidad. Para este estudio se tomaron 100 frutos por cultivar y por tratamiento; a diario se pesó cada fruto de manera individual, para determi-nar su pérdida de peso fresco. El trabajo consis-tió en un ensayo factorial de 2*2*3, cuyo primer factor fue el cultivar (Chandler o Sweet Char-lie), el segundo factor, los dos productos (ácido giberélico o nitrato de calcio) y el tercer factor las tres concentraciones (baja, media, alta) de los productos. Dada la naturaleza diferente de los dos productos utilizados, se dosificó de acuerdo con cada uno de estos; por tanto, para facilitar el acceso de los datos al programa estadístico, se utilizó el concepto de dosis baja, media, alta, las cuales correspondían a 0, 500, 1.500 y 3.000 mg L-1 de ácido giberélico y 50, 150 y 200 mg L-1 de nitrato de calcio, respectivamente. Los re-sultados se analizaron mediante un análisis de varianza (Anova) y la prueba de separación de promedios de Tukey (P≤0,05). El análisis esta-dístico se realizó mediante la aplicación de SPSS, versión 11.5.1.
reSUltADoS Y DISCUSIÓn
El efecto de los tratamientos implementados so-bre la calidad del fruto se evaluó en relación con el contenido de sólidos solubles totales y con la acidez total titulable, en la cual, según el análisis de varianza, no se encontraron diferencias signi-ficativas para ninguna de las variables evaluadas. A pesar de esto, vale la pena destacar que para ‘Chandler’ y ‘Sweet Charlie’ se encontraron valo-res para los sólidos solubles totales correspondien-tes a 8,02 y 8,92 ºBrix, respectivamente; mientras que en el caso de la acidez total titulable, Chan-dler presentó 0,09% y Sweet Charlie1,10%. De igual manera, Roussos et al. (2008) no encontra-
ron un impacto significativo de la aplicación de reguladores del crecimiento sobre estos mismos parámetros en fresa, así como tampoco hubo cambios importantes en el contenido de carbo-hidratos, ni en el color de la fruta, a pesar del incremento en la concentración de antocianinas como consecuencia de los tratamientos. Adicio-nalmente, los mismos autores reportaron que la actividad antioxidante del jugo de los frutos fue ligeramente mayor en los frutos provenientes de las plantas control, lo que podría ser el resultado de un mayor contenido de fenoles, o-difenoles, flavonoides y flavanoles. Estos resultados tam-bién han sido reportados por Ozguven y Yilmaz (2002), quienes no encontraron cambios en la acidez titulable en frutos de fresa luego de una aplicación de ácido giberélico. De hecho, el efecto del ácido giberélico sobre las características fisi-coquímicas de los frutos de fresa son altamente dependientes del cultivar, de la dosis aplicada y de la sensibilidad de las plantas al fotoperiodo, como reporta Kirschbaum (1998), quien presenta un listado de características de las plantas y fru-tos de fresa que fueron afectados por aplicaciones de ácido giberélico en precosecha.
Por otro lado, en diferentes reportes se menciona que la aplicación de reguladoras del crecimiento a las plantas tiene un efecto directo sobre el ta-maño de fruto sin afectar el contenido de sólidos solubles ni la acidez titulable, tanto en fresa (Roh-loff et al., 2002) como en otras especies (Amoros et al., 2004; Stern et al., 2007). Sin embargo, este resultado no se evidenció en el presente traba-jo porque no hubo respuesta de los frutos a las aplicaciones, en cuanto al incremento en tama-ño. Sólo se encontró que los frutos del cultivar Chandler fueron 40,7% más pesados que los de Sweet Charlie, pero este es un parámetro resul-tante del material vegetal utilizado y no por de-fecto de los productos aplicados.
Se presentó un efecto positivo de los tratamientos implementados sobre la prolongación de la vida en poscosecha de los frutos. Al respecto, hubo dife-rencias altamente significativas (P≤0,01) del culti-
CASIERRA-pOSADA/SALAMANCA

Vol. 2 - No. 1 - 2008
37
var. El cultivar Chandler mostró una duración de 18,4% más tiempo en poscosecha que el cultivar Sweet Charlie. La interacción producto y con-centración fue significativa (P≤0,05). Los frutos tratados con ácido giberélico duraron 11,6% más tiempo en referencia con los frutos de las plantas control, mientras aquellos tratados con nitrato de calcio tuvieron 16,4% más tiempo en poscosecha, en comparación con los controles. En cuanto al AG3, en concentración ascendente, los frutos du-raron 14,8; 10,3 y 17,0% más tiempo que los fru-tos de las plantas control. En lo relacionado con la significancia de la interacción de los tres factores, sólo se presentó diferencia altamente significativa para la interacción cultivar * producto (P≤0,01).
El calcio contribuye a unir las sustancias pécticas en las paredes celulares (Demarty et al., 1984). Las implicaciones poscosecha de un buen sumi-nistro de calcio radican en que altos contenidos de calcio en los frutos causan una reducción en la
tasa de maduración, respiración, producción de etileno y ablandamiento de los frutos (Ferguson, 1984). El calcio se absorbe principalmente a tra-vés de los tejidos jóvenes de los ápices radicales y se transloca a través del torrente de transpira-ción (Mengel et al., 2001; por tanto, aplicaciones precosecha con productos al suelo que conten-gan calcio tienen como consecuencia un mejo-ramiento en el comportamiento poscosecha y en la calidad de frutos en fresa. Sin embargo, en el presente ensayo, las aplicaciones se realizaron vía foliar, con lo cual fue también posible me-jorar parcialmente su comportamiento en pos-cosecha, a pesar de la condición de inmovilidad del elemento cuando es aplicado a los tejidos aé-reos, como lo corroboraron Eaves y Leefe (1962). Igualmente, Cheour et al. (1990; 1991) encontra-ron que aplicaciones de cloruro de calcio redu-cen la maduración y el desarrollo de hongos en poscosecha e incrementan la firmeza del fruto durante la cosecha y la poscosecha.
Figura 1: efecto de la aplicación de ácido giberélico y de nitrato de calcio sobre la duración de frutos de fresa (Fragaria sp.) en poscosecha. las barras sobre las columnas indican la desviación estándar.
INFLUENCIA DEL áC IDO G IBERéL ICO y DEL N ITRATO DE CALC IO

REV. COLOMB. CIENC. HORTIC.
38
Por otro lado, se encontró un efecto favorable de aplicaciones foliares de calcio sobre la calidad de la fruta, pero no hubo efecto significativo so-bre la producción de fruta o sobre la reducción de frutos deformados (Wojcik y Lewandouski, 2003). En contraste, Erincik et al. (1998), en en-sayos realizados tanto en campo como en inver-nadero, reportaron que no hubo diferencias sig-nificativas de las aplicaciones foliares de cloruro de calcio sobre el contenido de Ca en los frutos de fresa, ni tampoco sobre la producción de fru-tos, firmeza, sólidos solubles, acidez y color ex-terno de los frutos. Makus y Morris (1989; 1998) sustentan la conclusión que aplicaciones tanto foliares como edáficas de calcio no incrementan la calidad de frutos de fresa ni la distribución del elemento dentro de los frutos. Por tanto, se planteó la importancia de la fuente de calcio y fue así, como Esmel et al. (2004) utilizaron tio-sulfato de calcio por medio de fertirrigación en plantas de fresa, sin encontrar significancia en la
calidad de frutos. Todas estas apreciaciones con-ducen a concluir que hay un componente gené-tico involucrado en los resultados, dado que los investigadores mencionados realizaron sus ensa-yos utilizando diferentes cultivares de fresa. Este aspecto se evidenció de igual manera en el pre-sente trabajo, como se discutirá posteriormente.
Los resultados del ensayo mostraron diferencias altamente significativas de los factores cultivar, producto y concentración sobre la pérdida de peso en poscosecha, pero no se presentó diferencia para las interacciones entre los factores (P≤0,01). Los frutos del cultivar Chandler perdieron 5,4% más peso que los de Sweet Charlie. A su vez, los frutos de plantas tratadas con nitrato de calcio en precosecha perdieron en promedio 23,64% más peso que aquellos tratados con ácido giberélico, en los que el promedio de pérdida fue de 3,8%, en relación con el control sin aplicación. Adicio-nalmente, se pudo constatar una pérdida de peso
Figura 2: efecto de la aplicación de ácido giberélico y de nitrato de calcio sobre pérdida de peso en frutos de fresa (Fragaria sp.) en poscosecha. las barras sobre las columnas indican la desviación estándar.
CASIERRA-pOSADA/SALAMANCA

Vol. 2 - No. 1 - 2008
39
de frutos de manera proporcional al incremento en la concentración del producto aplicado en pre-cosecha. Las dosis inferior, media y alta indujeron 5,9; 8,0 y 14,5% de pérdida de peso en la posco-secha de frutos en relación con los frutos de las plantas control.
De hecho, la mayor pérdida de peso encontrada en los frutos, como consecuencia de la aplicación de nitrato de calcio, fue resultado de que los fru-tos tratados con este compuesto duraron más tiempo en poscosecha; por tanto, la exposición de sus epidermis a las condiciones ambientales que les inducían la perdida de peso fue más prolon-gada. Las pérdidas de peso de los frutos de fresa durante la poscosecha son la consecuencia de la respiración de los frutos (California Strawberry Commission, 2007) y también de las pérdidas de agua (Keutgen y Pawelzik, 2008). A pesar de estas modificaciones encontradas en poscosecha, estos últimos autores no encontraron cambios en el contenido relativo de materia seca. La pérdida de peso estaba acompañada por una apariencia y cambios de color poco atractivos de los frutos hacia el final de la vida en poscosecha, lo cual se evidenció claramente en el presente ensayo, en el cual los frutos adquirían una falta de brillo y se tornaban de una coloración más oscura en la medida en que perdían humedad, resultados que también fueron reportados por Nunes et al. (1995) y Knee y Aggrawal (2000).
Los resultados del presente ensayo estuvieron influenciados por el componente genético. Fue así, como los dos cultivares utilizados respon-dieron de manera diferente a las aplicaciones del ácido giberélico y de nitrato de calcio. La figura 3 muestra que en conjunto los frutos del culti-var Chandler duraron un día más en poscosecha que los de Sweet Charlie, independientemente del producto aplicado o de su concentración, lo cual pone de manifiesto que la respuesta de los frutos de fresa a tratamientos orientados al me-joramiento de la calidad o a la prolongación de la vida poscosecha es fuertemente dependiente del
material vegetal utilizado en los ensayos. Esto fue corroborado por diferentes investigadores citados anteriormente quienes encontraron res-puestas contradictorias o inconsistentes de di-versos cultivares a las aplicaciones de diferentes fuentes de calcio, en relación con la producción y calidad del fruto.
Adicionalmente, se reporta que los frutos de fre-sa pierden su posibilidad de mercadeo con una pérdida de humedad del orden de 6% (Nunes et al., 1995), lo cual fue alcanzado en todos los tratamientos, casi desde el primer día de alma-cenamiento. Evaluaciones similares en cuanto al comportamiento de frutos en poscosecha, en respuesta a tratamientos previos, arrojaron resul-tados similares. Es así como Keutgen y Pawelzik (2008) evaluaron el comportamiento de frutos de fresa expuestos a ozono y encontraron que en todos los tratamientos, los frutos alcanzaron el 6% de pérdida de humedad al quinto día luego de la cosecha, adicionalmente, que los tratamientos con ozono, durante el crecimiento, reducen las pérdidas de peso fresco en 1%, lo cual es un valor pequeño que no incide de manera considerable en las posibilidades de mercado de los frutos, debido a los cambios en la apariencia de los mismos.
Debe hacerse énfasis en que uno de los objetivos del presente estudio era la evaluación de los tra-tamientos en cuanto a la calidad y duración de los frutos en poscosecha, sin condiciones espe-ciales de almacenamiento que pudieran interve-nir con la pérdida de humedad, de esta manera, los frutos expuestos a condiciones extremas de almacenamiento, tendrían la posibilidad de ex-presar su durabilidad afectada por los productos aplicados en precosecha. En concordancia con los resultados del presente estudio, se reporta que la vida poscosecha de los frutos se ve ampliamente afectada por factores como la variedad, el sitio cultivo, el comportamiento climático durante la temporada, las prácticas precosecha y las con-diciones ambientales en poscosecha (California Strawberry Commission, 2007).
INFLUENCIA DEL áC IDO G IBERéL ICO y DEL N ITRATO DE CALC IO
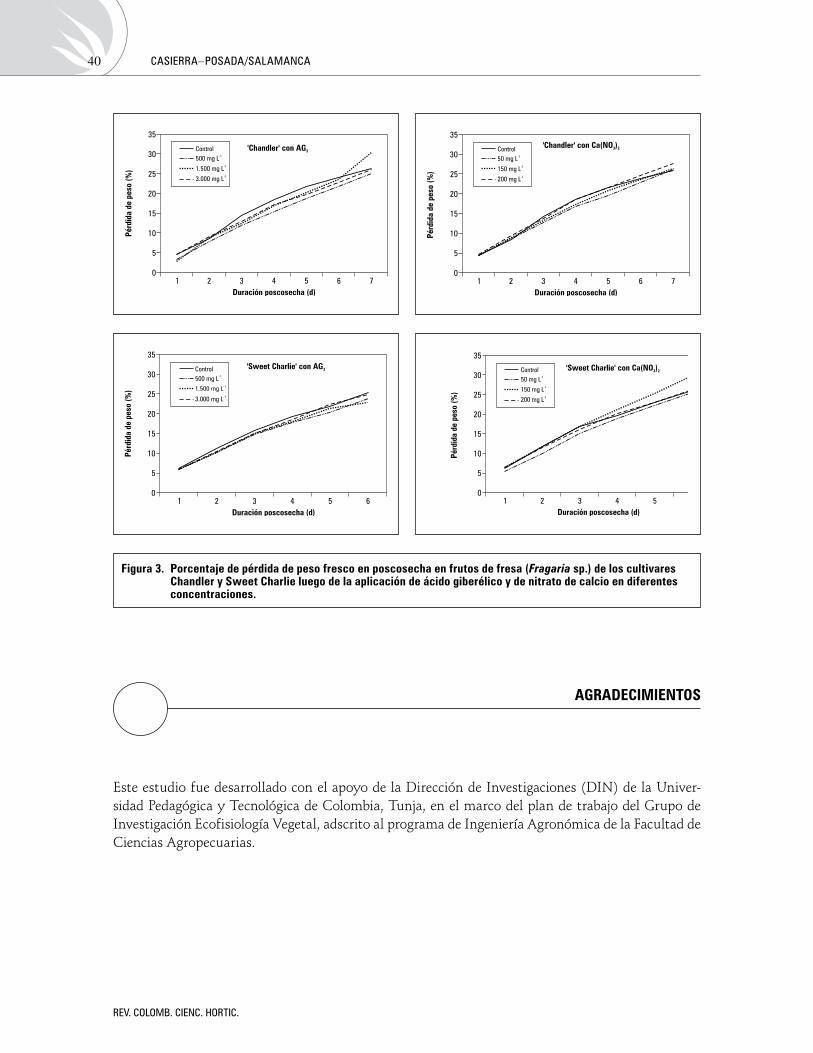
REV. COLOMB. CIENC. HORTIC.
40
Figura 3. Porcentaje de pérdida de peso fresco en poscosecha en frutos de fresa (Fragaria sp.) de los cultivares Chandler y Sweet Charlie luego de la aplicación de ácido giberélico y de nitrato de calcio en diferentes concentraciones.
AGrADeCIMIentoS
Este estudio fue desarrollado con el apoyo de la Dirección de Investigaciones (DIN) de la Univer-sidad Pedagógica y Tecnológica de Colombia, Tunja, en el marco del plan de trabajo del Grupo de Investigación Ecofisiología Vegetal, adscrito al programa de Ingeniería Agronómica de la Facultad de Ciencias Agropecuarias.
CASIERRA-pOSADA/SALAMANCA

Vol. 2 - No. 1 - 2008
41
Agronet. 2008. Área cosechada, producción y rendimien-to de fresa 1992-2006. En: http://www.agronet.gov.co/; consulta: 21 de mayo de 2008.
Amoros, A.; P. Zapata; M.T. Pretel; M.A. Botella; M.S. Almansa y M. Serrano. 2004. Role of naphthalene acetic acid and phenothiol treatments on increas-ing fruit size and advancing fruit maturity in lo-quat. Scientia Hort. 101, 387-398.
Azodanlou, R.; C. Darbellay; J.L. Luisier; J.C. Villettaz y R. Amado. 2003. Quality assessment of strawberries (Fragaria species). J. Agric. Food Chem. 51, 715-721.
California Strawberry Commission. 2007. Best handling practices for fresh strawberries. En: http://www.calstrawberry.com/fileData/docs/Best_Handling_Practice_for_Fresh_Strawberries_-_Final_03_12_07.pdf; consulta: 5 de marzo de 2008.
Cheour, F.; C. Willemot; J. Arul; Y. Desjardins; J. Makhlout; P.M. Charest y A. Gosselin. 1990. Foliar application of calcium chloride delays post-harvest ripening of strawberry. J. Amer. Soc. Hort. Sci. 115, 789-792.
Cheour, F.; C. Willemot; J. Arul; J. Makhlout y Y. Des-jardins. 1991. Post-harvest response of two straw-berry cultivars to foliar applications of calcium chloride. HortScience 26, 1186-1188.
Choma, M.E. y D.G. Himelrick. 1984. Responses of day-neutral, June-bearing and everbearing strawberry cultivars to gibberellic acid and phthalamide treat-ments. Scientia Hort. 22, 257-264.
Corabastos. 2004. Fresa. Portal de Frutas y Hortalizas de Colombia para el Mundo. En: http://frutasyhor-talizas.com.co/portal/includej/product_view.php; consulta: 9 de mayo de 2008.
Demarty, M.; C. Morvan y M. Thellier. 1984. Calcium and the cell wall. Plant Cell Environ. 7, 441-448.
Dennis, F.G. y H.O. Bennett. 1969. Effect of gibberel-lic acid and deflowering upon runner and inflores-cence development in an everbearing strawberry. J. Amer. Soc. Hort. Sci. 94, 534-537.
Eaves, C.A. y J.S. Leefe. 1962. Note on the influence of calcium on the firmness of strawberries. Can. J. Plant Sci. 42, 746-747.
reFerenCIAS BIBlIoGrÁFICAS
Erincik, O.; L.V. Madden; J.C. Scheerens y M.A. Ellis. 1998. Evaluation of foliar applications of calcium chloride for control of Botrytis fruit rot on straw-berry and effects on strawberry fruit quality. Adv. Strawberry Res. 17, 7-13.
Esmel, C.E.; J.R. Duval y A.A. Steven. 2004. The influ-ence of calcium thiosulfate on yieldand posthar-vest quality of ‘Sweet Charlie’ strawberry. Proc. Fla. State Hort. Soc. 117, 48-51.
Ferguson, I. 1984. Calcium in plant senescence and fruit ripening. Plant Cell Environ. 7, 477-489.
Guardiola, J.L. y A. Garcia-Luis. 2000. Increasing fruit size in citrus. Thinning and stimulation of fruit growth. Plant Growth Regul. 31, 121-132.
Keutgen, A. y E. Pawelzik. 2008. Influence of pre-har-vest ozone exposure on quality of strawberry fruit under simulated retail conditions. Postharv. Biol. Technol. 49, 10-18.
Kirkby, E.A. y D.J. Pilbeam. 1984. Calcium as a plant nutrient. Plant Cell Environ. 7, 397-405.
Kirschbaum, D.S. 1998. Temperature and growth regu-lator effects on growth and development of straw-berry (Fragaria x ananassa Duch.). Master Sc. the-sis. University of Florida, Gainesville, FL.
Knee, M. y D. Aggrawal. 2000. Evaluation of vacuum containers for consumer storage of fruits and veg-etables. Postharv. Biol. Technol. 19, 55-60.
Makus, D.J. y J.R. Morris. 1989. Influence of soil and foliar applied calcium on strawberry fruit nutrient and postharvest quality. Acta Hort. 265, 443-446.
Makus, D.J. y J.R. Morris. 1998. Preharvest calcium ap-plications have little effect on mineral distribution in ripe strawberry fruit. HortScience 33, 64-66.
Mengel, K., E.A. Kirkby, H. Kosegarten y T. Appel. 2001. Principles of plant nutrition. 5th ed. Kluwer Acade-mic Publishers, Dordrecht, The Netherlands.
Nitsch, J.P. 1950. Growth and morphogenesis of the strawberry as related to auxin. Amer. J. Bot. 37, 211-215.
INFLUENCIA DEL áC IDO G IBERéL ICO y DEL N ITRATO DE CALC IO

REV. COLOMB. CIENC. HORTIC.
42
Nunes, M.C.N.; J.K. Brecht; A.M.M.B Morais y S.A. Sargent. 1995. Physical and chemical quality char-acteristics of strawberries after storage are reduced by a short delay to cooling. Postharv. Biol. Technol. 6, 17-28.
Ozguven, A.I. y C. Yilmaz. 2002. The effect of gibber-ellic acid treatments on the yield and quality of strawberry (Fragaria x ananassa) cv. Camarosa. Acta Hort. 567, 277-280.
Rohloff, J.; S. Fiskaa H. y T. Iversen. 2002. The effect of plant biochemical regulators on strawberry pro-duction in field trials under drip irrigation man-agement at 5 locations in Norway. Acta Hort. 567, 463-466.
Roussos P.A.; N.-K. Denaxa y T. Damvakaris. 2008. Strawberry fruit quality attributes after applica-tion of plant growth stimulating compounds. Sci-entia Hort. 119(2), 138-146.
Singh, J.P.; G.S. Randhawa y N.L. Jain. 1960. Response of strawberry to gibberellic acid. Indian J. Hort. 17, 21-30.
Stern, R.A.; M.A. Flaishman y R. Ben-Arie. 2007. Effect of synthetic auxins on fruit size of five cultivars of Japanese plum (Prunus salicina Lindl.). Scientia Hort. 112, 304-309.
Tehranifar, A. y N.H. Battey. 1997. Comparison of the effects of GA3 and chilling on vegetative vigour and fruit set in strawberry. Acta Hort. 439, 627-631.
Weidman, R.W. y J. Stang. 1983. Effects of gibberellins (GA4+7), 6-benzyladenine (6-BA) and promalin (GA4+7 + 6-BA) plant growth regulators on plant growth, branch crown and flower development in ‘Scott’ and ‘Raritan’ strawberries. Adv. Strawberry Prod. 2, 15-17.
Wojcik, P. y M. Lewandowski. 2003. Effect of calcium and boron sprays on yield and quality of ‘Elsanta’ strawberry. J. Plant Nutr. 26, 671-682.
CASIERRA-pOSADA/SALAMANCA

FÁnor CASIerrA-PoSADA1, 2
oSCAr e. AGUIlAr-AvenDAño1
Fruto de mora cultivada cerca de Fusagasugá, Colombia. Foto: G. Fischer
Respuestas fisiológicas y morfológicas de plantas de mora (Rubus sp.) sometidas a estrés por viento inducido
Physiological and morphological responses of blackberry (Rubus sp.) plants to wind-induced stress
reSUMen
Cuando las plantas se exponen a agentes ambientales causales de estrés, como el viento, van a producir ciertas alteraciones: de orden físico (rompimiento de sus estructuras, desecación del follaje, etc.); de orden morfoló-gico, denominadas respuestas tigmomorfogénicas, y de orden fisiológico como modificaciones en el patrón de distribución de materia seca (MS) o en sus relaciones hídricas. Con el fin de determinar el grado de respuesta de plantas de mora a la estimulación por viento, se llevó a cabo un ensayo bajo condiciones controladas de laboratorio. Dos grupos de plantas fueron sometidos a ventilación artificial a velocidades 2 y 4 m s-1, adicional-mente se contó con un grupo de plantas testigo, las cuales no fueron afectadas por el estímulo. Se determinó el patrón de distribución de MS, la evapotranspiración, la eficiencia agronómica en el uso del agua (g MS/L H2O) y el contenido relativo de humedad. El viento modificó la forma en que las plantas distribuyeron su MS. Las plantas influenciadas por ventilación artificial asignaron a ramas y hojas más fotoasimilados que a los órganos reproductivos. Hubo una tendencia a incrementarse la evapotranspiración con el aumento del nivel de estrés. La eficiencia agronómica en el uso del agua y el contenido relativo de humedad de las plantas disminuyeron significativamente a medida que aumentaba la velocidad del viento. Las plantas de mora mostraron alta sensibilidad al viento particularmente en la alteración de sus relaciones hídricas. El hecho de que las plantas dirigieron más fotosintatos a órganos estructurales que reproductivos es una evidencia de tigmomorfogénesis en esta especie.
1 Grupo de Investigación Ecofisiología Vegetal, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja (Colombia).
2 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 43-53, 2008

REV. COLOMB. CIENC. HORTIC.
44
Palabras clave adicionales: viento-inducido, tigmomorfogénesis, contenido relativo de humedad, eficiencia agronómica en el uso del agua.
Additional keywords: wind-induced, thigmomorphogenesis, relative water content, agronomic water use efficiency.
Fecha de recepción: 04-02-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
ABStrACt
The effects on plants of wind-induced stress may be physical –breakage of structural organs, desiccation of foliage, etc.– morphological –thigmomorphogenic responses– and physiological –changes in the distribution pattern of dry matter, or modifications in plant water relations–. This research was carried out under laboratory conditions to evaluate the response of blackberry plants to wind stress. Three groups of plants were subjected to artificial ventilation at speeds of 2 and 4 m s-1 or left untreated. The distribution pattern of dry matter (DM), evapotranspiration, water use efficiency (g DM/L H2O), and relative water content were measured. Results showed that wind changed the way dry matter was distributed within the plants. The exposed to wind allocated more photoassimilates to branches and leaves than to reproductive organs compared to unexposed plants. Stress level shifts were observed to determine concomitant evaporation increases. Water use efficiency and relative water content of plants decreased significantly at increasing wind speed. In summary, blackberry plants showed high sensitivity to wind, especially through alteration of water relations. The fact that plants allocated more photoassimilates to structural than to reproductive organs is evidence of thigmomorphogenic response in this species.
El cultivo de mora (Rubus sp.) es uno de los cul-tivos frutícolas promisorios y más importantes con los que cuenta el sector agrícola colombia-no. Se estima que el área nacional destinada a este cultivo en 2007 fue de aproximadamente 10.229 ha, con un volumen de total producción de 97.936 t de fruta. Los rendimientos en el país, bajo condiciones óptimas, varían de 6 a 16 t con un promedio de 9,6 t ha-1 (Agronet, 2008).
Se sabe que en la naturaleza existen diversos factores que inducen condiciones de estrés en las plantas; el viento es posiblemente el principal agente ambiental, responsable de propiciar estrés de orden mecánico, el cual potencialmente indu-ce cambios morfológicos o fisiológicos, orienta-dos a la adaptación de las plantas sometidas a este factor de estrés. Estos cambios pueden aso-ciarse en la planta con ajustes fisiológicos, bien
CASIERRA-pOSADA/AGUILAR-AVENDAñO

Vol. 2 - No. 1 - 2008
45
sea una redistribución de los fotoasimilados, en orden de evitar fallas estructurales en la planta, o por otra parte, se pueden presentar alteraciones en el balance osmótico del vegetal para impedir su deshidratación (Schulze et al., 2005).
Los cambios morfológicos parecen estar relacio-nados casualmente a los cambios en la función de varios sistemas en la planta. Por ejemplo, la in-hibición del crecimiento debido a perturbaciones mecánicas puede deberse en parte a una dismi-nución en la producción de nuevos fotosintatos (Grace y Thompson, 1973) y a un incremento en la reducción de fotosintatos por el proceso respi-ratorio (Todd et al., 1972).
Los vientos fuertes pueden ocasionar daños seve-ros en plantas de importancia agrícola. Los efectos de corrientes de viento pueden resultar en una rá-pida desecación del follaje o del hipocótilo. Cuan-do esto ocurre en plántulas jóvenes, éstas pueden volcarse y morir. Si las condiciones del estrés son muy fuertes, pueden dar pie a lesiones físicas las cuales pueden resultar en daños al follaje o en el rompimiento de tallos o ramas (Fischer, 2000).
La transpiración en las plantas ocurre a través de los estomas o por difusión a través de otros te-jidos superficiales. Muchas de las superficies en las plantas se encuentran cubiertas con una ba-rrera cerosa que le permite a la planta mantener las pérdidas de agua al mínimo. Los daños a esta barrera pueden ser ocasionados por la fricción de las plantas unas con otras mediante la acción del viento. El daño inicial que pueda ocasionar la corriente de aire se ve limitado a la acción de disturbar la defensa cerosa, pero eventualmente las tasas de transpiración se irán a ver afectadas (Pitcairn et al., 1986).
En algunas localidades colombianas, el cultivo de mora se establece en terrenos pendientes, donde el flujo de corrientes fuertes de viento es alto, exponiendo a las plantas a este agente cau-sante de estrés.
Este trabajo está orientado a determinar el efec-to de corrientes fuertes de viento sobre posibles cambios morfológicos o fisiológicos, conocidos como respuesta tigmomorfogénicas en plantas de mora, mantenidas bajo condiciones de cáma-ra de crecimiento.
MetoDoloGÍA
El ensayo se llevó a cabo en una cámara de cre-cimiento (fitotrón) en la Facultad de Ciencias Agropecuarias de la Universidad Pedagógica y Tecnológica de Colombia en Tunja, Colombia, ubicada a 2.690 msnm. Se controlaron las condi-ciones ambientales calculadas para el desarrollo del ensayo: temperatura promedio 21,8°C, hu-medad relativa 81,2% y flujo fotónico fotosinté-tico equivalente a 49,7 μmol m-2 s-1, proveniente de luz natural a través de teja plástica.
Se evaluaron plantas de mora sin espinas (Rubus sp.), de una edad cercana a dos meses, obtenidas a partir de acodos aéreos. Las plantas permane-cieron en bolsas plásticas con capacidad de 4 kg. Con el fin de trabajar con un material uniforme, se eliminaron los brotes basales de las plantas, dejando un solo un brote principal, manejando una altura uniforme.
Una vez homogeneizado el material vegetal, se procedió a separarlo aleatoriamente en tres gru-pos de 10 plantas. Cada uno de los grupos de plantas fue expuesto individualmente a una ve-locidad de viento-inducido, proveniente de ven-tilación artificial. Mediante la implementación de un anemómetro, se establecieron las diferen-tes velocidades de viento.
Un primer grupo de plantas se expuso a una ve-locidad de viento de 2 m s-1, un segundo grupo a una velocidad de 4 m s-1 y, finalmente, se contó con un tercer grupo (plantas testigo), el cual no estuvo expuesto a la ventilación. Para los prime-ros dos grupos se determinó un promedio de 6
RESpUESTAS F IS IOLóGICAS y MORFOLóGICAS DE pLANTAS DE MORA

REV. COLOMB. CIENC. HORTIC.
46
h de exposición por día al estímulo, logrando de esta manera una estimulación considerable de las ramas y las hojas.
Como variables respuesta se determinaron: la asignación y distribución de materia seca, la eva-potranspiración, la eficiencia agronómica en el uso del agua y el contenido relativo de humedad.
El peso seco de los tejidos vegetales se determinó con la ayuda de una estufa calibrada a 70°C du-rante 24 h, hasta peso constante. A partir de los pesos secos de los diferentes órganos, fue posible establecer los patrones de distribución de materia seca para los diferentes tratamientos. La evapo-transpiración se determinó mediante la adición del volumen de agua faltante respecto a un peso de saturación, agregada en cada riego durante todo el ensayo, con base en la evapotranspiración fue posible calcular la cantidad necesaria en li-tros para producir un gramo de materia seca (efi-ciencia agronómica en el uso del agua) (Feddes, 1985). De acuerdo con la metodología propuesta por Barr y Weatherley (1962), un estimativo que permite conocer el estatus hídrico de la planta, en condiciones de un posible déficit hídrico en las cé-lulas. El método consistió en tomar muestras de tejidos foliares y rápidamente determinar el peso del trozo de hoja, posteriormente se hidrataron a plena saturación durante 24 h en cajas de Pe-tri con papel filtro saturado de agua desionizada. Luego de 4 h se halló el peso de las muestras fo-liares en plena saturación. Finalmente se halló el peso seco de la muestra. Con los datos obtenidos, se calculó el contenido relativo de humedad me-diante la fórmula RWC (%) = [(W-DW) / (TW-DW)] x 100; en la cual W – peso de la muestra en fresco, TW – peso de la muestra saturada, DW – peso seco de la muestra.
El arreglo factorial del experimento fue de un diseño estadístico completamente al azar con 10 replicaciones por tratamiento y una planta como unidad experimental. Los resultados obte-nidos fueron sometidos a un análisis de varian-cia (Anova), las pruebas de comparación múlti-
ple halladas mediante la prueba de Tukey. Los análisis estadísticos se realizaron con la versión 11.5.1 de SPSS® (Statistical Product and Service Solutions, Chicago, IL).
reSUltADoS Y DISCUSIÓn
Distribución de materia seca
Se encontró que la ganancia y distribución de carbono fueron afectadas por la velocidad del viento, la cual tuvo un efecto sobre la propor-ción en la cual se distribuyen los fotosintatos en los diferentes órganos de la planta (figura 1). La MS asignada a las raíces fue mayor (39,9%) en las plantas que no fueron influenciadas por la ventilación, en cambio, aquellas sometidas a las velocidades de viento 2 y 4 m s-1 asignaron gradualmente carbono en menor porcentaje 37,3 y 33,4%, respectivamente. Por otra parte, la dis-tribución de fotosintatos a las ramas se mostró favorecida por la acción de las corrientes de vien-to; en promedio la distribución de materia seca a estos órganos para las dos velocidades de viento fue de 26,76%, mientras que en las plantas que no estuvieron expuestas al estrés la distribución de MS se mostró en menor proporción (21,5%). De la misma forma que en las ramas, la distri-bución de fotoasimilados a las hojas incrementó gracias a la acción del viento. El máximo nivel de exposición al estrés aumentó el porcentaje de materia seca distribuido a las hojas (36,4%), en cambio en las plantas control y las sometidas a una velocidad de viento 2 m s-1 indicaron menor porcentaje de carbono asignado a hojas (32,0%). Si bien la distribución de materia seca en órganos aéreos de la planta como ramas y hojas incremen-tó gradualmente con la velocidad del viento, no ocurrió lo mismo con los órganos reproductivos (flores y frutos), por el contrario, aparentemente las plantas, al encontrarse sometidas a condicio-nes de estrés, asignaron una menor cantidad de fotosintatos a estos órganos. La MS acumulada en botones florales, flores y frutos se redujo a 3,8 y 2,4%, con el flujo de viento a velocidad 2 y 4 m
CASIERRA-pOSADA/AGUILAR-AVENDAñO
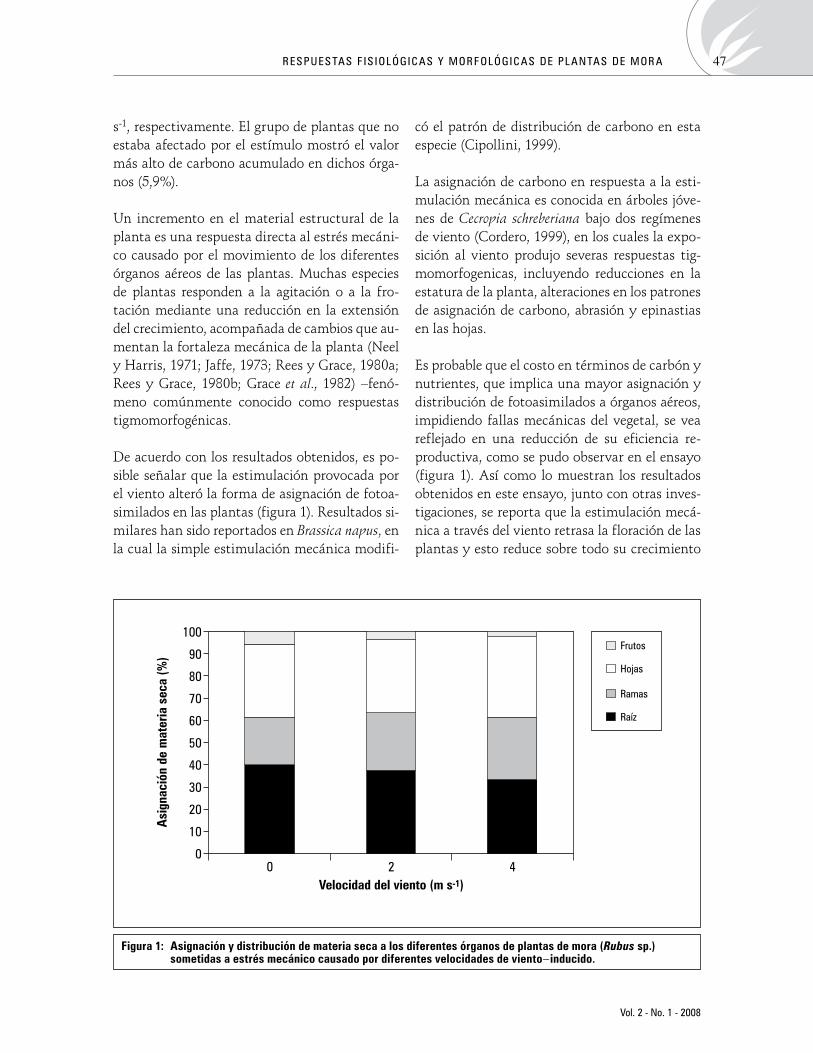
Vol. 2 - No. 1 - 2008
47
Figura 1: Asignación y distribución de materia seca a los diferentes órganos de plantas de mora (Rubus sp.) sometidas a estrés mecánico causado por diferentes velocidades de viento-inducido.
s-1, respectivamente. El grupo de plantas que no estaba afectado por el estímulo mostró el valor más alto de carbono acumulado en dichos órga-nos (5,9%).
Un incremento en el material estructural de la planta es una respuesta directa al estrés mecáni-co causado por el movimiento de los diferentes órganos aéreos de las plantas. Muchas especies de plantas responden a la agitación o a la fro-tación mediante una reducción en la extensión del crecimiento, acompañada de cambios que au-mentan la fortaleza mecánica de la planta (Neel y Harris, 1971; Jaffe, 1973; Rees y Grace, 1980a; Rees y Grace, 1980b; Grace et al., 1982) –fenó-meno comúnmente conocido como respuestas tigmomorfogénicas.
De acuerdo con los resultados obtenidos, es po-sible señalar que la estimulación provocada por el viento alteró la forma de asignación de fotoa-similados en las plantas (figura 1). Resultados si-milares han sido reportados en Brassica napus, en la cual la simple estimulación mecánica modifi-
có el patrón de distribución de carbono en esta especie (Cipollini, 1999).
La asignación de carbono en respuesta a la esti-mulación mecánica es conocida en árboles jóve-nes de Cecropia schreberiana bajo dos regímenes de viento (Cordero, 1999), en los cuales la expo-sición al viento produjo severas respuestas tig-momorfogenicas, incluyendo reducciones en la estatura de la planta, alteraciones en los patrones de asignación de carbono, abrasión y epinastias en las hojas.
Es probable que el costo en términos de carbón y nutrientes, que implica una mayor asignación y distribución de fotoasimilados a órganos aéreos, impidiendo fallas mecánicas del vegetal, se vea reflejado en una reducción de su eficiencia re-productiva, como se pudo observar en el ensayo (figura 1). Así como lo muestran los resultados obtenidos en este ensayo, junto con otras inves-tigaciones, se reporta que la estimulación mecá-nica a través del viento retrasa la floración de las plantas y esto reduce sobre todo su crecimiento
RESpUESTAS F IS IOLóGICAS y MORFOLóGICAS DE pLANTAS DE MORA

REV. COLOMB. CIENC. HORTIC.
48
reproductivo (Whitehead, 1962; Henry y Tho-mas, 2002; Anten et al., 2005).
Friedrich y Fischer (2000) reportaron que movi-mientos suaves de 1,7 m s-1 fueron óptimos para la producción de materia seca en frutales caduci-folios, puesto que un viento demasiado calmado puede conducir a un empobrecimiento del CO2 alrededor de la hoja, mientras que corrientes de viento alrededor del valor mencionado incremen-tan la transpiración y aumentan el exceso de ca-lor en las hojas.
Evapotranspiración
El consumo de agua por parte de las plantas, así como su pérdida por evaporación y por transpira-ción, no estuvo afectado significativamente por
el viento; sin embargo, fue posible observar un incremento promedio de cerca del 5% de la eva-potranspiración cuando las plantas fueron some-tidas a la ventilación artificial en comparación con el grupo de plantas que no fueron sometidas al estímulo (figura 2).
Un aumento en la evapotranspiración indica la tasa a la cual el flujo de vapor de agua de una hoja es afectada por la turbulencia del viento. Este aire turbulento transporta el agua lejos de las hojas hacia la atmósfera. De esta forma, es posible afirmar que el viento afecta la evapo-transpiración considerablemente.
El incremento de la pérdida de agua en las plan-tas afectadas por el estrés por viento se puede atribuir a tres factores: el aumento de la conduc-
Figura 2. evapotranspiración en plantas de mora (Rubus sp.) sometidas a estrés mecánico causado por diferentes velocidades de viento-inducido. Promedios con letras distintas indican diferencia significativa según la prueba de tukey (P≤0,01), las barras sobre las columnas indican la desviación estándar.
CASIERRA-pOSADA/AGUILAR-AVENDAñO

Vol. 2 - No. 1 - 2008
49
tancia estomática, la abrasión de las barreras de vapor y los efectos del viento sobre la transpira-ción. Sólo las dos primeras son una respuesta di-recta de las plantas a los efectos del viento y sólo la primera es el resultado de cambios morfológi-cos (Cleugh et al., 1998); sin embargo, corrientes suaves de viento, cerca de 1,7 m s-1, tendrían un efecto favorable para las plantas, mediante un incremento moderado en la tasa de transpira-ción y la reducción del sobrecalentamiento de las hojas, como lo describen Friedrich y Fischer (2000).
Aunque la transpiración de una hoja es predomi-nantemente una función de la cantidad de ener-gía recibida por la hoja, el viento puede influir en la manera en que la hoja pierde energía, y, por tanto, puede afectar significativamente la trans-piración. El viento influye en la transpiración mediante la remoción de la llamada capa de sa-turación del aire superficial de la hoja, así como por el cambio en la temperatura de la hoja (Woo-lley, 1960). Existen también otros mecanismos por los cuales el viento puede afectar la trans-piración, como: a) disminución de la presión del aire sobre el borde lateral de la hoja, causando el aumento en la evaporación de este lado; b) venti-lación de los espacios intracelulares; c) flexión de las hojas en el viento, provocando la compresión de los espacios intracelulares y el consiguiente bombeo de aire saturado fuera de los estomas (Woolley, 1960).
Eficiencia agronómica del uso del agua
Al cuantificar en gramos la MS que produjeron las plantas por un litro de agua consumida (efi-ciencia agronómica del uso del agua), se eviden-ció que existe un efecto significativo del viento sobre la eficiencia agronómica del uso del agua. Las plantas control se mostraron las más efi-cientes (3,58 g L-1) en comparación con aquellas sometidas a la ventilación. La producción de fi-tomasa se vio reducida en 29,44% en el grupo de
plantas sometidas a velocidad del viento 4 m s-1. Para la velocidad de viento 2 m s-1, la eficiencia agronómica en el uso del agua se redujo en un 11,47% respecto a la reportada en las plantas sin ventilación artificial (figura 3).
En general, las plantas expuestas al viento pre-sentan cambios anatómicos y morfológicos que se asemejan a aquellos encontrados en especies tolerantes al estrés por sequía, aparentemente diferentes, pero con la funcionalidad en térmi-nos de control del uso del agua. Las plantas bajo condiciones de estrés por sequía tienden a ser usuarios parsimoniosos del agua, mientras que las afectadas por el estrés por viento tienden a ser más derrochadoras. Como se reporta en la presente investigación, al inducir las plantas al viento, éstas van a presentar gradualmente al ni-vel de exposición al estrés, menor eficiencia en el uso del agua, comparadas con las plantas no estimuladas por el viento (figura 3).
La eficiencia en el uso del agua relaciona toda la cantidad de carbono adquirido por la planta (Bri-ggs y Shantz, 1913; Blum 2005). Sin embargo, es difícil medir de manera experimental el agua transpirada por las plantas. Por tal razón, la efi-ciencia agronómica del uso del agua constituye un estimativo más práctico, ya que relaciona la biomasa total con el agua evapotranspirada.
Diversos estudios han detallado el uso del agua que hacen las plantas bajo condiciones de vien-to-inducido. Barker et al. (1989) encontraron que plantas de algodón expuestas a corrientes de viento fueron más pequeñas, presentaron menor área foliar y, por tanto, fueron menos efi-cientes en el uso del agua que aquellas plantas que crecieron en condiciones tranquilas. Com-portamiento similar fue presentado (Finnell, 1928), en el cual se encontró que con velocida-des de viento de 7 m s-1 varias especies vegetales redujeron su crecimiento, pero asimismo incre-mentaron el uso del agua.
RESpUESTAS F IS IOLóGICAS y MORFOLóGICAS DE pLANTAS DE MORA

REV. COLOMB. CIENC. HORTIC.
50
Contenido relativo de humedad
Al determinar el estado hídrico de las plantas bajo condiciones de estrés, se encontraron diferencias altamente significativas (P≤0,01) en el contenido de humedad de los tejidos. Las plantas control, las cuales no estuvieron influenciadas por la ventila-ción artificial, revelaron un contenido de hume-dad de 82,89%, mientras que las plantas someti-das a las corrientes de aire presentaron niveles de desecación. Se observó un contenido de humedad de 53,92% para las plantas que se encontraban es-timuladas a una velocidad de viento de 2 m s-1. El mayor nivel de desecación se observó en el grupo de plantas que se encontraban bajo la acción del viento a mayor velocidad (4 m s-1), el contenido de humedad en este grupo de plantas fue menor (46,6%) y se mostró a la mitad del que revelaron las plantas control (figura 4).
Uno de los primeros síntomas de la deficiencia de agua en los tejidos de las plantas es la disminu-
ción del contenido relativo de agua. La reducción en el contenido relativo de humedad en plantas bajo condiciones de estrés puede estar asociada con la disminución en la energía de las plantas y se ha observado en muchas especies (Lopez et al., 2002; Halder y Burrage, 2003).
El contenido relativo de humedad representa un indicador útil del estado del balance hídrico de una planta, fundamentalmente debido a que ex-presa la cantidad absoluta de agua que requiere la planta para alcanzar su plena saturación arti-ficial (González y González-Vilar, 2001). La ca-pacidad de las plantas para sobrevivir a un grave déficit de agua depende de su capacidad para li-mitar la pérdida de agua a través de la epidermis de la hoja después que los estomas han alcanza-do una apertura mínima (El Jaafari, 2000).
El viento tiene una influencia directa en la dese-cación de los tejidos, como se pudo evidenciar en el presente ensayo (figura 4).
Figura 3. eficiencia agronómica en el uso del agua en plantas de mora (Rubus sp.) sometidas a estrés mecánico causado por diferentes velocidades de viento-inducido. Promedios con letras distintas indican diferencia significativa según la prueba de tukey (P≤0,01), las barras sobre las columnas indican la desviación estándar.
CASIERRA-pOSADA/AGUILAR-AVENDAñO

Vol. 2 - No. 1 - 2008
51
Los valores normales del contenido relativo de humedad oscilan entre el 98%, en hojas turgen-tes, y cerca del 40% en tejidos severamente di-secados y hojas muertas. En la mayoría de las especies cultivadas, el contenido de humedad típico de las plantas que se encuentran cerca a la marchitez es de 60% a 70%.
Bajo condiciones de estrés ambiental, las mem-branas de la planta están sujetas a cambios asociados a menudo con el incremento en la permeabilidad y la pérdida de la integridad (Blo-khina et al., 2003).
ConClUSIoneS
•Las plantas de mora presentaron un alto grado de respuesta fisiológica a la estimulación por viento. Las relaciones hídricas se vieron com-prometidas, particularmente aquellas relacio-nadas con la eficiencia en la producción de
biomasa, así como el contenido de humedad de los tejidos. Por su parte, existe la eviden-cia de respuestas tigmomorfogénicas en esta especie, dado que se alteraron los patrones de distribución de materia seca, en los cuales se evidenció un direccionamiento superior de fo-toasimilados a órganos estructurales (ramas y hojas), evitando de esta forma posibles fallas mecánicas del vegetal, mientras que la bioma-sa acumulada en órganos reproductivos y fru-tos se redujo gracias a la acción del viento.
•El establecimiento del cultivo de mora en zo-nas donde la acción de fuertes corrientes de viento probablemente limitará el rendimiento y la producción del cultivo. Se hace necesaria entonces la evaluación de medidas que permi-tan controlar el efecto de este estímulo sobre las plantas, como la instalación de rompevien-tos, o el manejo de las densidades de siembra, de modo que las plantas se escuden unas a otras, reduciendo la posibilidad de alteraciones morfológicas y fisiológicas.
Figura 4. Contenido de humedad en plantas de mora (Rubus sp.) sometidas a estrés mecánico causado por diferentes velocidades de viento-inducido. Promedios con letras distintas indican diferencia significativa según la prueba de tukey (P≤0,01), las barras sobre las columnas indican la desviación estándar.
RESpUESTAS F IS IOLóGICAS y MORFOLóGICAS DE pLANTAS DE MORA

REV. COLOMB. CIENC. HORTIC.
52
•Es posible que en la implementación de ba-rreras rompevientos no sólo sus resultados se vean reflejados en la reducción de los daños directos del viento, sino que, a su vez, puedan
mejorar el rendimiento y la calidad de un cul-tivo, ya que potencialmente modificarían las condiciones, creando un microclima favorable para su desarrollo.
Agronet. 2008. Área cosechada, producción y rendimien-to de mora, 1992-2007. En: http://www.agronet.gov.co; consulta: 28 de mayo de 2008.
Anten, N.P.R.; R. Casado-Garcia y H. Nagashima. 2005. Effects of mechanical stress and plant density on mechanical characteristics, growth and lifetime reproduction of tobacco plants. Amer. Naturalist 166, 650-660.
Barker, G.L.; J.L. Hatfield y D.F. Wanjura. 1989. Influ-ence of wind on cotton growth and yield. Trans-act. ASAE 32(1), 97-104.
Barr, H.D. y P.E Weatherley. 1962. A re-examination of the relative turgidity technique for estimating wa-ter deficit in leaves. Aust. J. Biol. Sci. 15, 413-428.
Blokhina, O.; E. Virolainen y K.V. Fagerstedt. 2003. An-tioxidants, oxidative damage and oxygen depriva-tion stress. Ann. Bot. 91, 179-194.
Blum, A. 2005. Drought resistance, water-use efficiency, and yield potential - are they compatible, dissonant, or mutually exclusive. Aust. J. Agr. Res. 56, 1159-1168.
Briggs, L.J. y H.L. Shantz. 1913. The water requirements of plants. II. A review of the literature. USDA Bur. Plant Ind. Bull. 284, 1-96.
Cipollini, D.F. 1999. Costs to flowering of the produc-tion of a mechanically hardened phenotype in Brassica napus L. Intl. J. Plant Sci. 160, 735-741.
Cleugh, H.A.; J.M. Miller y M. Böhm. 1998. Direct me-chanical effects of wind on crops. Agrofor. Syst. 41, 85-112.
Cordero, R.A. 1999. Ecophysiology of Cecropia schreberi-ana saplings in two wind regimes in an elfin cloud forest: growth, gas exchange, architecture and stem biomechanics. Tree Physiol. 19, 153-163.
El Jaafari, S. 2000. Durum wheat breeding for abiotic stresses resistance: defining physiological traits
reFerenCIAS BIBlIoGrÁFICAS
and criteria. Options Mediterranéennes Serie A 40, 251-256.
Feddes, R.A. 1985. Cropwater use and dry matter pro-duction: state of the art. pp. 221-234. En: Les be-soins en eau des cultures. Conference Internatio-nale. September 11-14, 1984. París.
Finnell, H.H. 1928. Effect of wind on plant growth. J. Amer. Soc. Agron. 20(11), 1206-1210.
Fischer, G. 2000. Efecto de las condiciones en precosecha sobre la calidad poscosecha de los frutos. Rev. Co-malfi 27(1-2), 39-50.
Friedrich, G. y M. Fischer. 2000. Physiologische Grundlagen des Obstbaues. Ulmer Verlag, Stuttgart, Alemania.
González, L. y M. González-Vilar. 2001. Determination of relative water content. pp. 207-212. En: Reigosa, M.J. (ed). Handbook of plant ecophysiology tech-niques. Kluwer Academic Publishers, Dordrecht, The Netherlands..
Grace, J. y J.R. Thompson. 1973. The after-effect of wind on the photosynthesis and transpiration of Festuca arundinacea. Physiol. Plant. 28, 541-547.
Grace, J.; C.E.R. Pitcairn; G. Russell y M. Dixon. 1982. The effects of shaking on the growth and water relations of Festuca arundinacea Schreb. Ann. Bot. 49, 207-215.
Halder, K.P. y S.W. Burrage. 2003. Drought stress effects on water relations of rice grown in nutrient film technique. Pakistan J. Biol. Sci. 6, 441-444.
Henry, H.A.L. y S.C. Thomas. 2002. Interactive effects of lateral shade and wind on stem allometry, biomass allocation and mechanical stability in Abutilon theo-phrasti (Malvaceae). Amer. J. Bot. 89, 1609-1615.
Jaffe, M.J. 1973. Thigmomorphogenesis: the response of plant growth and development to mechanical stimulation. Planta 114, 143-157.
CASIERRA-pOSADA/AGUILAR-AVENDAñO

Vol. 2 - No. 1 - 2008
53
López, C.M.L.; H. Takahashi y S. Yamazaki. 2002. Plant water relations of kidney bean plants treated with NaCl and foliarly applied glycinebetaine. J. Agron. Crop Sci. 188, 73-80.
Neel, P.L. y R.W. Harris. 1971. Motion-induced inhibi-tion of elongation and induction of dormancy in liquidambar. Sci. 173, 58-59.
Pitcairn, C.E.R.; C.E. Jeffree y J. Grace. 1986. Influence of polishing and abrasion on the diffusive conduc-tance of leaf surface of Festuca arundinacea Schreb. Plant Cell Environ. 9, 191-196.
Rees, D.J. y J. Grace. 1980a. The effect of wind on the extension growth of Pinus contorta Douglas. Fo-restry 53, 145-153.
Rees, D.J. y J. Grace. 1980b. The effect of shaking on the extension growth of Pinus contorta Douglas. Forestry 53, 154-166.
Schulze, E.-D.; E. Beck y K. Müller-Hohenstein. 2005. Plant ecology. Springer Verlag, Heidelberg, Alemania.
Todd, G.W.; D.L. Chadwick y S.D. Tsai. 1972. Effect of wind on plant respiration. Physiol. Plant. 27, 342-346.
Whitehead, F.H. 1962. Experimental studies of the effect of wind on plant growth and anatomy II. Helian-thus annus. New Phytol. 61, 59-62.
Woolley, J.T. 1960. Mechanisms by which wind influ-ences transpiration. Plant Physiol. 36, 112-114.
RESpUESTAS F IS IOLóGICAS y MORFOLóGICAS DE pLANTAS DE MORA

YUlI DeAQUIZ-oYolA1
JAvIer ÁlvAreZ-HerrerA1, 2
AnA FrAIle1
Frutos del tomate ‘Calima’. Foto: G. Fischer
Efecto de diferentes láminas de riego y sustratos en la propagación de tomate (Solanum lycopersicum L.)
Effect of different water levels and substrates on tomato (Solanum lycopersicum L.) propagation
reSUMen
Uno de los principales problemas de la producción de tomate es el alto costo de la plantulación, así como el desconocimiento de láminas de riego adecuadas en germinación de semillas. Se evaluó el efecto de diferen-tes láminas de riego y sustratos en la germinación de semillas del híbrido larga vida Calima, utilizando 25 tratamientos compuestos por las láminas de riego 0,8; 1,0; 1,2; 1,4 y 1,6 y los sustratos turba rubia, mezcla de cascarilla de arroz quemado con suelo negro, mezcla de turba rubia con arena, mezcla de turba rubia con cascarilla de arroz quemado (1:1 v/v) y mezcla de turba rubia con cascarilla de arroz quemado para un total de 125 unidades experimentales. La combinación de la lámina de riego de 1,2 con la mezcla de turba rubia con cascarilla quemada presentó la mejor respuesta en cuanto a porcentaje, tiempo medio y velocidad media de la germinación, además en altura de la planta, longitud de la raíz, peso en fresco y en seco tanto para las hojas y el tallo. La lámina de riego de 1,6 y el sustrato mezcla de turba rubia con cascarilla quemada superó los demás tratamientos en cuanto área foliar, peso fresco y seco de raíz. El tratamiento óptimo económicamente fue el de la lámina de riego de 1,2 y la mezcla de turba rubia con cascarilla quemada.
1 Grupo de Investigaciones Agrícolas, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colom-bia (UPTC), Tunja (Colombia).
2 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 54-65, 2008

Vol. 2 - No. 1 - 2008
55
ABStrACt
Inappropriate substrates and irrigation levels often result in poor seed germination and consequently high priced tomato seedlings. The objective of this study was to assess the effect of different substrates and levels of irrigation on germination of ‘Calima´ tomato seeds, a long life hybrid. A randomized complete block design was used with 5 replicates and 25 treatments: five irrigation levels (0.8, 1.0, 1.2, 1.4 and 1.6 irrigation coefficient) and five substrates (blond peat, a mixture of burned rice husks with black soil, a mixture of blond peat with sand, a mixture of blond peat with rice husks (1:1 v/v) and a mixture of blond peat with burned rice husks). The combination of the 1.2 irrigation coefficient and the mixture of peat moss with burned rice husks attained the best response in terms of percentage, time and average rate of germination, seedling height, root length, and fresh and dry weight of leaves and stems. The 1.6 irrigation coefficient and blond peat substrate mixture with rice husks exceeded other treatments in leaf area and fresh and dry weight of the roots. The economically best treatment was that of the 1.2 irrigation coefficient and the substrate mixture of blond peat with burned rice husks
Palabras clave adicionales: germinación, híbrido, evaporación, plántula, mezcla.
Additional keywords: germination, hybrid, evaporation, seedling, mixture.
Fecha de recepción: 05-02-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
El tomate (Solanum lycopersicum L.) es una de las hortalizas de mayor importancia en el mundo (Gómez et al., 2000). En Colombia es la segunda hortaliza más importante y presenta un rendi-miento promedio de 25,78 t ha-1 en el país (FAO, 2007).
La disponibilidad de agua es una condición esen-cial para la germinación de las semillas. Ésta determina la imbibición y posterior activación de procesos metabólicos, como rehidratación, reparación de membranas y de ADN, elongación celular y aparición de la radícula (Dubreucq et al., 2000). En condiciones naturales, las plantas deben sincronizar sus ciclos de crecimiento y reproducción con un adecuado abastecimiento hídrico (Foley y Fennimore, 1998). La disponi-bilidad de agua durante el crecimiento de una planta madre afecta el desarrollo de sus semillas,
alterando su capacidad germinativa (Pallas et al., 1977; Benech et al., 1992; Gutterman, 2000).
Estudios fisiológicos del proceso de maduración de las semillas muestran que la tolerancia a la de-secación es diferente entre las especies y depende principalmente de dos factores: la velocidad a la cual se produce la pérdida de agua y el contenido final después del proceso de desecación (Hong y Ellis, 1992; Ellis y Hong, 1994; Wechsberg et al., 1994; Hay y Probert, 1995).
La aparición de enfermedades y el alto costo de las semillas de tomate mejoradas genéticamen-te hacen necesaria la utilización de sistemas de producción de plántulas en bandejas de germina-ción, en las que se utilizan sustratos enriquecidos con fertilizantes (Castilla, 1995). No obstante, el costo de los sustratos importados es alto, por lo
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE

REV. COLOMB. CIENC. HORTIC.
56
que surge la necesidad de disponer de materiales producidos localmente que brinden condiciones adecuadas de germinación y de fácil adquisición.
Quesada y Méndez (2005) encontraron que los sustratos que generaron los mejores resultados fueron los que incluían las mezclas de abono orgánico Juan Viñas + granza; lombricompost + abono orgánico Juan Viñas + granza y abo-no orgánico Juan Viñas, en almácigos de brócoli, pepino y tomate.
Los experimentos desarrollados para evaluar el efecto de diferentes tipos de vermicompost sobre la germinación, crecimiento, floración y fructifi-cación de varias especies hortícolas y ornamen-tales como pepino (Cucumis sativus L.), tomate (Solanum lycopersicum L.), petunias (Petunia gran-diflora L.), maravillas (Calendula officinalis L.), crisantemos (Chysanthemum sinense L.) demos-traron que las mejores respuestas de estas espe-cies se presentan cuando se sustituye del 10% al 20% del volumen total del medio de crecimiento comercial con los diferentes tipos de vermicom-post (Riggle, 1998; Subler et al., 1998).
Por lo anterior, el objetivo de este trabajo fue eva-luar el efecto de diferentes láminas y sustratos en germinación y obtención de plántulas de un híbrido de tomate larga vida, para asegurar una óptima calidad y eficiencia, reduciendo costos y optimizando el proceso de propagación.
MAterIAleS Y MÉtoDoS
El experimento se realizó en el invernadero de vidrio de la Facultad de Ciencias Agropecuarias de la Universidad Pedagógica y Tecnológica de Colombia en la ciudad de Tunja, 5º 32’ N, 73º 23’ W y 2.782 msnm. La temperatura promedio dentro del invernadero fue de 20ºC y la hume-dad relativa del 65%.
Como material vegetal, se empleó el híbrido de tomate (Solanum lycopersium L.) larga vida ‘Ca-lima’, el cual se seleccionó y se sembró en ban-dejas de polietileno de 72 alvéolos. La lámina de riego suministrada correspondió a cinco coefi-cientes de la evaporación (Ev), los sustratos em-pleados para la germinación fueron turba rubia canadiense (TR), mezcla de cascarilla de arroz quemado con suelo negro (CQSN), mezcla de turba rubia con arena (TRA), mezcla de turba rubia con cascarilla de arroz (TRCA) y mezcla de turba rubia con cascarilla de arroz quemado (TRCQ), todas las mezclas se hicieron en pro-porción 1:1 en volumen.
El diseño estadístico utilizado fue completamen-te al azar. Se evaluó la combinación de cinco di-ferentes sustratos y cinco láminas de riego (0,8; 1,0; 1,2; 1,4 y 1,6) para un total de 25 tratamien-tos (tabla 1), con cinco repeticiones, para un to-tal 125 unidades experimentales (UE).
La aplicación del primer riego se inició luego de la emergencia de las plántulas. Se instaló den-tro del invernadero un evaporímetro plástico a escala del tanque tipo A con dimensiones de 29 cm de diámetro y 6,1 cm de profundidad, reali-zando lecturas diarias de la evapotranspiración (Etp). Para el suministro de agua a la planta, se utilizó una jeringa de 10 mL con el fin de tener una mayor exactitud en las cantidades aplicadas, utilizando la siguiente fórmula:
En la cual, Etp = evapotranspiración en mm me-dida en el tanque evaporímetro; C = coeficiente multiplicador; A = área del alvéolo; = eficien-cia del riego (0,9).
La aplicación del primer riego se realizó luego de la emergencia de las plántulas. En la etapa de
DEAQUIZ-OyOLA/áLVAREZ-HERRERA/FRAILE

Vol. 2 - No. 1 - 2008
57
plántula, se midió área foliar, mediante el anali-zador Li-Cor® 3000A (Lincoln, NE); peso fresco y seco (70ºC durante 48 h), de raíz, tallo y hojas, longitud de raíz principal, altura final, eficiencia de tratamientos que se calcularon con base en la relación peso seco de plántula/consumo de agua y análisis económico que se calculó con base en el costo de tratamiento.
Se realizaron los análisis de varianza (Anova) con el fin de determinar la existencia de diferencias estadísticas entre sustratos y láminas de riego, asimismo mediante la prueba de comparación de Tukey con una confiabilidad del 95%, se estable-cieron los mejores tratamientos. Este análisis se realizó en el programa SAS v. 8.1e (Cary, NC).
reSUltADoS Y DISCUSIÓn
Germinación
El porcentaje de germinación presentó diferen-cias significativas entre tratamientos, la mejor respuesta se encontró con el sustrato TRCQ y un coeficiente de riego de 1,2 (figura 1). Entre sustra-tos, se observaron diferencias significativas, pero no entre láminas de riego; la mezcla TRCQ dio el mejor resultado. Debido a que el sustrato tuvo mayor retención de humedad y mayor espacio poroso, lo que permite una adecuada disponibi-lidad de oxigeno y mayor actividad respiratoria acorde con Handreck y Black (2002), quienes afirman que la porosidad total del sustrato afecta la capacidad de intercambio gaseoso del medio, disminuyendo el contenido de oxígeno que las semillas requieren para germinar.
Ensayos realizados por Maldonado et al. (2002) concluyeron que la capacidad de germinación de las semillas de Lycopersicon chilense estuvieron restringidas a condiciones de abastecimiento hí-drico favorable, provocando una reducción en la germinación, debido probablemente a que las enzimas hidrolíticas de los cotiledones no fueron activadas, lo que no desencadenó la activación metabólica de la semilla (Obroucheva y Antipo-va, 1997; Nonogaki y Morohashi, 1999).
La combinación entre la lámina de riego y los sustratos presentaron diferencias significativas que influyeron en la velocidad y uniformidad de germinación de las semillas, la mezcla TRCQ expresó la mayor respuesta, al igual que la lámi-na de 1,2. Esto debido a la influencia de la hume-
Tratamientos Sustratos Coeficiente de riego
T1 CQSN 1,6
T2 CQSN 1,0
T3 CQSN 0,8
T4 CQSN 1,2
T5 CQSN 1,4
T6 TRA 1,6
T7 TRA 1,0
T8 TRA 0,8
T9 TRA 1,2
T10 TRA 1,4
T11 TR 1,6
T12 TR 1,0
T13 TR 0,8
T14 TR 1,2
T15 TR 1,4
T16 TRCA 1,6
T17 TRCA 1,0
T18 TRCA 0,8
T19 TRCA 1,2
T20 TRCA 1,4
T21 TRCQ 1,6
T22 TRCQ 1,0
T23 TRCQ 0,8
T24 TRCQ 1,2
T25 TRCQ 1,4
tabla 1. tratamientos del estudio.
Abreviaturas
CQSN: mezcla de cascarilla de arroz quemado con suelo negro.
TRA: mezcla de turba rubia con arena.
TR: Turba rubia.
TRCA: mezcla de turba rubia con cascarilla de arroz.
TRCQ: mezcla de turba rubia con cascarilla de arroz quemado.
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE

REV. COLOMB. CIENC. HORTIC.
58
Figura 1: Germinación en plántulas de tomate bajo diferentes láminas de riego sembrados en diferentes sustratos.
tabla 2. Fórmulas empleadas para el cálculo de las variables de germinación.
dad y la disponibilidad de agua para la semilla en la germinación, que depende de la estructura porosa y la distribución del sustrato.
La velocidad media de germinación (VMG) au-menta en forma directa con la temperatura y la retención de humedad del sustrato, por lo que
para este caso se ve que los sustratos con mejor retención presentaron una VMG mayor, debido a que las semillas lograron absorber mayor agua, permitiendo el rompimiento de la cubierta, la emergencia de la radícula y el crecimiento de la plántula. Gul et al. (1995) compararon 10 sustra-tos, algunos con turba y arena en el cultivo de
Variable Ecuación Unidades
Velocidad media de germinación VMG = Semillas germinadas /día
Tiempo medio de germinación TMG = días
Porcentaje de germinación PG = *100 %
ni = Número de semillas germinadas en el i-ésimo día;
ti = Tiempo en días, para la germinación en el i-ésimo día.
N = Número de semillas germinadas.
NS = Número de semillas totales.
A1, A2,..., Ax: Número de semillas germinadas en el día 1, en el día 2, y en el día x.
T1, T2,..., Tx: Número de días entre la siembra y el primer día 1 de germinación, entre el día 2 y entre el día x.
ni
ti
N* (A1 + A2 + Ax)A1 *T1 + A2 *T2+ Ax *Tx)
NNs
DEAQUIZ-OyOLA/áLVAREZ-HERRERA/FRAILE

Vol. 2 - No. 1 - 2008
59
tomate cv. Carpy y obtuvieron plántulas una se-mana antes que en suelo.
En la variable tiempo medio de germinación, hubo diferencias significativas en el sustrato TRCQ y la lámina de 1,2 indujeron una rápida germinación en comparación a los demás tra-tamientos, lo que corrobora los resultados de VMG. Se asume que una vez la semilla tiene las condiciones adecuadas de humedad, temperatura y aireación el intercambio de gases entre sustrato y embrión acelera la emergencia.
El retraso de la germinación en los tratamien-tos con un coeficiente de riego de 1,0 y en los sustratos CQSN y TRA se dio por algún tipo de dormancia como mecanismo de defensa de las semillas (Bewley, 1997). Una vez que se rompió esta dormancia la germinación se activó. Se-gún Samperio (2001), la semilla absorbe agua, lo que produce un reblandecimiento en la capa protectora y se inicia el proceso enzimático que activa el crecimiento de la raíz y ésta empieza a alargarse, por tanto, un contenido adecuado de humedad en la semilla garantiza que el proceso germinativo se lleve a cabo en menor tiempo.
Abou Hadid et al. (1988) determinaron el tiempo de germinación de semillas de tomate cv. Alfredo en distintos sustratos, entre ellos uno constituido por arcilla: arena: aserrín (1:1:1) comparadas con semillas cultivadas en camas de suelo semillero, y encontraron que el tiempo de germinación se redu-jo entre tres a siete días, dependiendo del sustrato usado y, además, registraron un menor requeri-miento de agua para la germinación de las semi-llas, con respecto a las camas de suelo semillero.
Altura de las plántulas
En la altura se encontraron diferencias significa-tivas entre tratamientos. La mayor respuesta en altura se presentó en el sustrato TRCQ y una lámina de 1,2 de la evapotranspiración, con 8,14 cm, frente al sustrato CQSN y un coeficiente 0,8 que mostró plántulas de 2,58 cm (figura 2). La
homogeneidad en los sustratos y la VMG influyó en el crecimiento de la plántula.
El desarrollo de las plantas, en su etapa inicial, está directamente relacionado con la uniformi-dad en la germinación y ésta, a su vez, se puede atribuir exclusivamente a las características pro-pias del sustrato (Fernández-Bravo et al., 2005). Estas condiciones están relacionadas con factores físicos y químicos como el pH, contenido de nu-trientes, capacidad de intercambio gaseoso, agua disponible y temperatura, entre otros (Singh, 1998).
Quesada y Méndez (2005) encontraron que la mezcla de diferentes sustratos en la propagación de tomate influye en la altura de la plántula, ya que la deficiente preparación de los materiales se constituye en una limitante física.
Longitud de la raíz principal
Las semillas de tomate bajo el efecto de las dife-rentes láminas de riego y sustratos presentaron diferencias significativas en la longitud de raíz, el sustrato TRCQ y el coeficiente de riego de 1,2 generaron la mejor repuesta con una longitud de raíz principal de 15 cm, en relación al sustrato CQSN y un coeficiente de 0,8 que mostró 2,46 cm (figura 3). Esto reitera que el sustrato influye en el crecimiento vegetativo de la plántula y que está directamente relacionado con la retención de humedad, capacidad de aireación y la entrada de agua a la pared celular de la raíz, lo que per-mite el flujo masivo de solutos y el transporte de nutrientes.
Wilches y Álvarez (2007) encontraron que las plántulas de tomate sometidas a diferentes lá-minas de riego no presentaron diferencias sig-nificativas en la longitud radical; sin embargo, la lámina de coeficiente 2,2 mostró la mayor longitud de raíz principal. El hecho de que no se presentaran diferencias se debe al poco tiempo que tienen las plántulas para expresar el efecto de los tratamientos.
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE
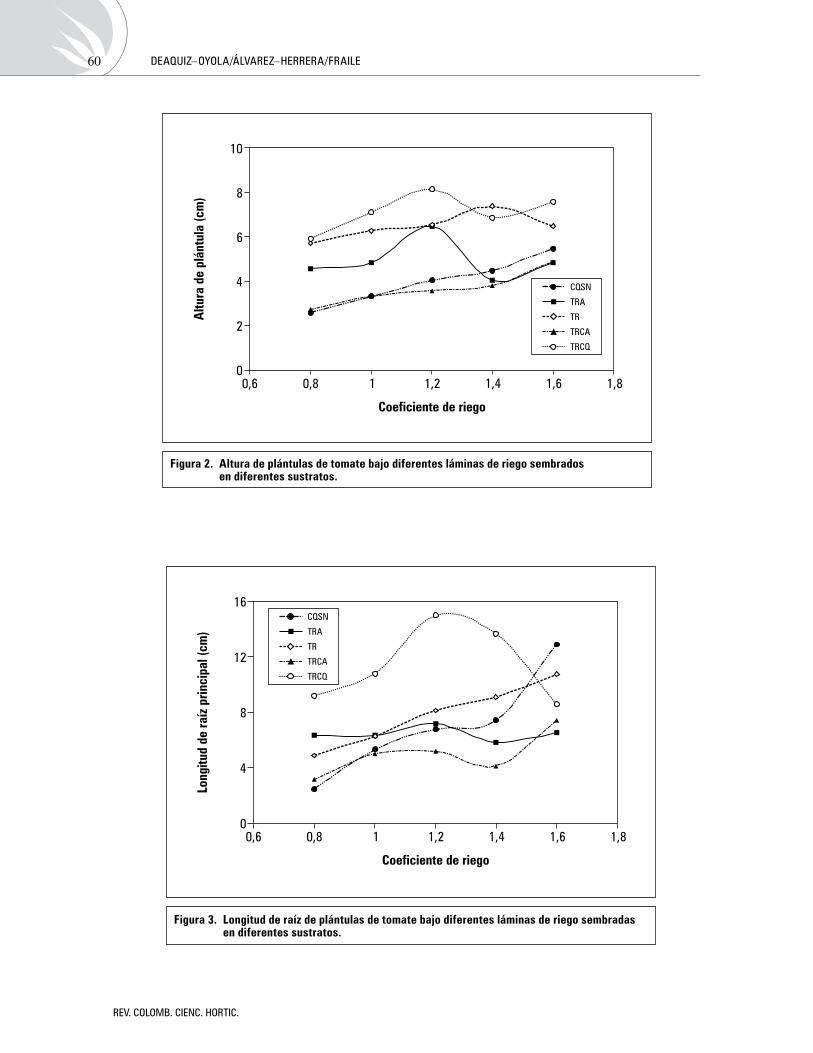
REV. COLOMB. CIENC. HORTIC.
60
Figura 2. Altura de plántulas de tomate bajo diferentes láminas de riego sembrados en diferentes sustratos.
Figura 3. longitud de raíz de plántulas de tomate bajo diferentes láminas de riego sembradas en diferentes sustratos.
DEAQUIZ-OyOLA/áLVAREZ-HERRERA/FRAILE
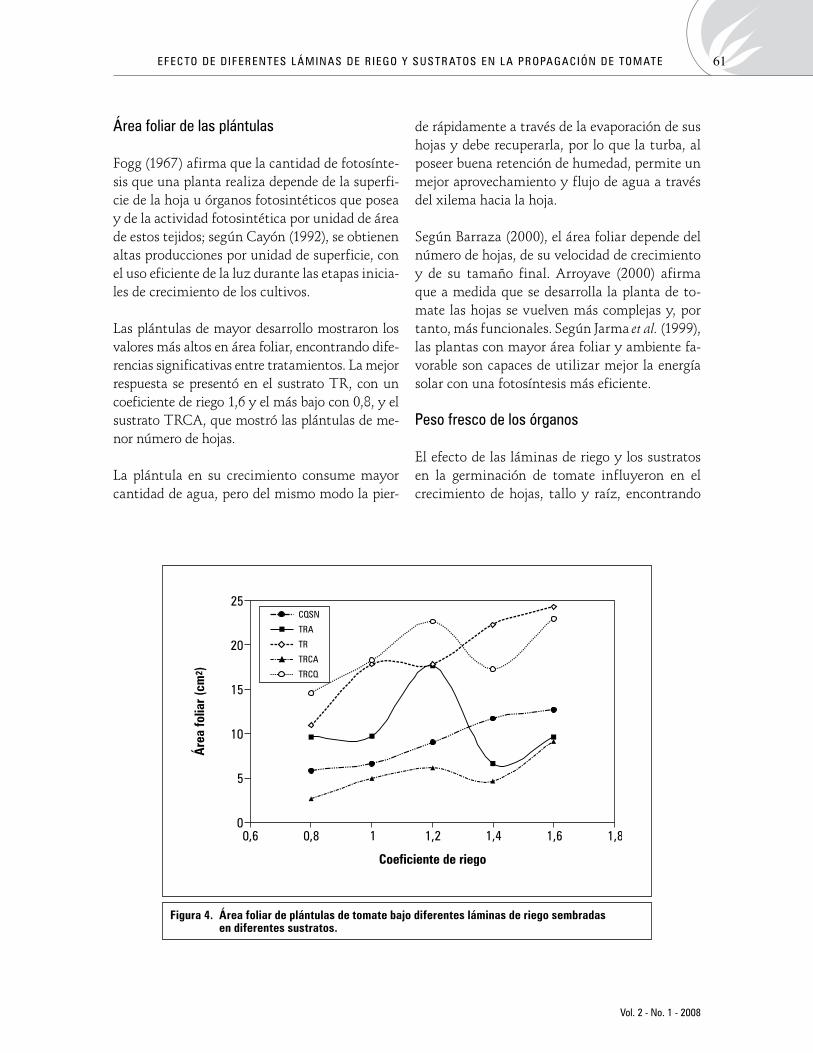
Vol. 2 - No. 1 - 2008
61
área foliar de las plántulas
Fogg (1967) afirma que la cantidad de fotosínte-sis que una planta realiza depende de la superfi-cie de la hoja u órganos fotosintéticos que posea y de la actividad fotosintética por unidad de área de estos tejidos; según Cayón (1992), se obtienen altas producciones por unidad de superficie, con el uso eficiente de la luz durante las etapas inicia-les de crecimiento de los cultivos.
Las plántulas de mayor desarrollo mostraron los valores más altos en área foliar, encontrando dife-rencias significativas entre tratamientos. La mejor respuesta se presentó en el sustrato TR, con un coeficiente de riego 1,6 y el más bajo con 0,8, y el sustrato TRCA, que mostró las plántulas de me-nor número de hojas.
La plántula en su crecimiento consume mayor cantidad de agua, pero del mismo modo la pier-
de rápidamente a través de la evaporación de sus hojas y debe recuperarla, por lo que la turba, al poseer buena retención de humedad, permite un mejor aprovechamiento y flujo de agua a través del xilema hacia la hoja.
Según Barraza (2000), el área foliar depende del número de hojas, de su velocidad de crecimiento y de su tamaño final. Arroyave (2000) afirma que a medida que se desarrolla la planta de to-mate las hojas se vuelven más complejas y, por tanto, más funcionales. Según Jarma et al. (1999), las plantas con mayor área foliar y ambiente fa-vorable son capaces de utilizar mejor la energía solar con una fotosíntesis más eficiente.
peso fresco de los órganos
El efecto de las láminas de riego y los sustratos en la germinación de tomate influyeron en el crecimiento de hojas, tallo y raíz, encontrando
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE
Figura 4. Área foliar de plántulas de tomate bajo diferentes láminas de riego sembradas en diferentes sustratos.

REV. COLOMB. CIENC. HORTIC.
62
diferencias significativas entre tratamientos. La mayor respuesta en hojas y tallo se obtuvo en el sustrato TRCQ, con un coeficiente de 1,2 frente al sustrato TRCA, con el coeficiente de 0,8 que mostró las plántulas de menor masa (figura 5).
La óptima captación de nutrientes y agua se ve reflejada en los mejores tratamientos, lo que con-llevó a una mayor división celular e incremento en masa fresca y a la absorción de oxigeno, que es esencial en el proceso de respiración y de acti-vidad metabólica.
Scagel (1987) menciona que una de las princi-pales funciones del tallo es formar y mantener las hojas y las estructuras de reproducción, con-ducir agua y nutrientes y almacenar sustancias. A medida que incrementa su tamaño garantiza
una mayor acumulación y transporte de solutos en la planta (Alóni, 1987), lo que manifiesta un mayor volumen de agua en sus tejidos, una ma-yor movilidad de los nutrientes y una actividad fisiológica eficiente.
peso seco de los órganos
Con la lámina de 1,2 y el sustrato TRCQ se pre-sentaron diferencias significativas en relación con los demás tratamientos. La lámina de 0,8 y el sustrato TRCA resultó en las plántulas de menor masa seca de hojas y tallo. En el caso de la raíz, el sustrato TRCQ, con un coeficiente de 1,6, presentó el valor más alto con 0,051 g, frente al sustrato TRCA y un coeficiente de 0,8 que ob-tuvo plántulas de 0,0013 g (figura 6). Esto se de-bió al pobre desarrollo alcanzado por la plántula,
Figura 5. Peso fresco de hojas, tallo y raíz en plántulas de tomate bajo diferentes láminas de riego sembradas en diferentes sustratos.
DEAQUIZ-OyOLA/áLVAREZ-HERRERA/FRAILE

Vol. 2 - No. 1 - 2008
63
debido a la influencia del sustrato, lo que causó una disminución en la actividad metabólica y el almacenamiento de reservas alimenticias y redu-jo el volumen celular.
Fernández-Bravo et al. (2005) encontraron que plántulas de tomate sembradas en turba obtu-vieron los mejores resultados de masa seca de tallo frente a otros sustratos, favoreciendo el cre-cimiento inicial de la plántula.
Según Tadeo (2000), existe una relación directa entre la masa fresca y la masa seca, en la que una mayor cantidad de agua influye directamente en la masa seca de la planta. De igual forma, una ma-yor masa seca implica que el agua dentro de los tejidos estará retenida en mayor cantidad y con mayor fuerza.
Análisis económico del sistema de propagación
Al evaluar los tratamientos económicamente, se encontraron diferencias significativas. El trata-miento más costoso fue el TR con un coeficiente de 1,6 con un valor de $66,88 pesos colombianos por plántula, frente al sustrato CQSN con un co-eficiente de 0,8, el cual tuvo el menor costo con un valor de $56,51. Al comparar los resultados de cada una de las variables con el costo por tratamiento, se ve que la mezcla entre turba y cascarilla quema-da es más rentable y económica, pues se obtienen plántulas vigorosas y óptimas para transplante.
El mejor tratamiento costó $63,69 y está cons-tituido por la combinación de la lámina de riego 1,2 y el sustrato TRCQ. De igual forma, fue el de
Figura 6. Peso seco de hojas, tallo y raíz en plántulas de tomate bajo diferentes láminas de riego sembradas en diferentes sustratos.
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE

REV. COLOMB. CIENC. HORTIC.
64
mejor eficiencia económica, lo que significa que generó mayor masa seca por unidad de masa de agua empleada.
ConClUSIoneS•La mezcla de turba rubia con cascarilla que-
mada y la lámina de 1,2 presentó los mejores
resultados en altura, longitud de raíz, peso fresco y seco de hojas y tallo.
•La lámina de 0,8 y el sustrato turba rubia ob-tuvo la mejor eficiencia en masa seca por agua consumida.
•El tratamiento óptimo económicamente fue el de la lámina de riego 1,2 y la mezcla de turba rubia con cascarilla quemada.
reFerenCIAS BIBlIoGrÁFICAS
Abou Hadid, A.; S. El Beltagy y H. El Saied. 1988. Soil mixture for tomato transplants. Egypt. J. Hort. 15, 23-28.
Alóni, B. y E. Pressman. 1987. The effects of salinity and gibberelic acid on blackheart disorder in celery. Agron. J. 79, 205-208.
Arroyave, H. 2000. Siembre los híbridos Petoseed. La compañía de las semillas híbridas. Bogotá. pp. 1-20.
Barraza, F.V. 2000. Crecimiento del Chile Manzano (Capsicum pubescens R. y P.) en cuatro soluciones nutritivas bajo invernadero. Tesis de Maestría, Ciencias en Horticultura, Universidad Autónoma de Chapingo, México.
Benech, A.; R.L. Fenne y P.J. Edwards. 1992. Changes in dormancy level in Sorghum halepense seeds induced by water stress during seed development. Funct. Ecol. 6, 596-605.
Bewley, J.D. 1997. Seed germination and dormancy. Plant Cell 9, 1055-1066.
Castilla, N. 1995. Manejo del cultivo intensivo del to-mate. pp. 189-226. En: Nuez, F. (ed.). El cultivo del tomate. Ediciones Mundi-Prensa, Madrid.
Cayón, G. 1992. Fotosíntesis y productividad de culti-vos. Rev. Comalfi 19(2), 23-31.
Dubreucq, B.; N. Berger; E. Vincent; M. Boisson; M. Caboche y L. Lepiniec. 2000. The Arabidopsis AtEPR1 extensin-like gene is specifically expressed in endosperm during seed germination. Plant J. 23, 643-652.
Ellis, R. y T.D. Hong. 1994. Desiccation tolerance and potential longevity of developing seeds of rice (Oryza sativa L.). Ann. Bot. 73, 501-506.
FAO. 2007. FAOStat estadística databases, agricultura, En: http://faostat.fao.org/site/340/default.aspx; con-sulta: 4 de febrero de 2008.
Fernández-Bravo, C.; N. Urdaneta; W. Silva; H. Poliszu y M. Marín. 2005. Germinación de semillas de to-mate (Lycopersicon esculentum Mill.) cv. Río Grande sembradas en bandejas plásticas, utilizando distin-tos sustratos. Rev. Fac. Agron. LUZ 23(2), 188-195.
Fogg, G.E. 1967. El crecimiento de las plantas. Editorial Universitaria de Buenos Aires, Argentina.
Foley, M.E. y S.A. Fennimore. 1998. Genetic basis for seed dormancy. Seed Sci. Res. 8, 173-182.
Gómez, O.; A. Casanova; H. Laterot e Y.G. Anais. 2000. Mejora genética y manejo del cultivo del tomate para la producción en el Caribe. Instituto de In-vestigaciones Hortícolas “Liliana Dimitrova”, La Habana, Cuba.
Gul, A.; A. Sevgican; F. Tognoni; G. Serra y A. Monteiro. 1995. Effect of growing media on glasshouse toma-to yield and quality. Acta Hort. 303, 145-150
Gutterman, Y. 2000. Maternal effects on seed during de-velopment. pp. 59-84. En: Fenner, M. (ed.). Seed: the ecology of regeneration in plant communities. CAB International, Wallingford, UK.
Handreck, K. y N. Black. 2002. Growing media for or-namental plants and turf. 3th ed. UNSW Press, Australia.
DEAQUIZ-OyOLA/áLVAREZ-HERRERA/FRAILE

Vol. 2 - No. 1 - 2008
65
Hay, F.R. y R.J. Probert. 1995. Seed maturity and the effects of different drying conditions on desicca-tion tolerance and seed longevity in foxglove (Digi-talis purpurea L.). Ann. Bot. 76, 639-647.
Hong, T.D. y R.H. Ellis. 1992. Development of desiccation tolerance in Norway maple (Acer platanoides L.) seeds during maturation drying. Seed Sci. Res. 2, 169-172.
Jarma, A.; C. Buitrago y S. Gutiérrez. 1999. Respuesta del crecimiento de la habichuela (Phaseolus vulgaris L. var. BlueLake) a tres niveles de radiación inci-dente. Rev. Comalfi 26(1-3), 62-73.
Maldonado, C.; E. Pujado y F.A. Squeo. 2002. El efecto de la disponibilidad de agua durante el crecimiento de Lycopersicon chilense sobre la capacidad de sus semillas para germinar a distintas temperaturas y concentraciones de manitol y NaCl. Rev. Chil. His-tor. Natur. 75, 651-660.
Nonogaki, H. y Y. Morohashi. 1999. Temporal and spa-tial pattern of the development of endo-ß-man-nanase activity in germinating and germinated lettuce seeds. J. Exp. Bot. 50, 1307-1313.
Obroucheva, N.V. y O.V. Antipova. 1997. Physiology of the initiation of seed germination. Russ. J. Plant Physiol. 44, 250-264.
Pallas, J.E.; J.R. Stansell y R.R. Bruce. 1977. Peanut seed germination as related to soil water regime during pod development. Agron. J. 69, 381-383.
Quesada, G. y C. Méndez. 2005. Análisis fisicoquímico de materias primas y sustratos de uso potencial en almácigos de hortalizas. Rev. Agr. Trop. 35, 1-13.
Riggle, D. 1998. Vermicomposting research and educa-tion. BioCycle 39, 54-56.
Samperio, G. 2001. Germinación de semillas: Manual de divulgación para uso en instituciones de educación. Toluca, Estado de México. En: http://www.rlc.fao.org/es/agricultura/aup/pdf/mexico.pdf; consulta: febrero de 2008.
Scagel, R.F. 1987 El reino vegetal. 2ª ed. Ediciones Ome-ga, Barcelona.
Singh, B.P. y U.M. Sainju. 1998. Soil physical and mor-phological properties and root growth. HortScience 33(6), 966-971.
Subler, S.; C.A. Edwards y J.D. Metzger. 1998. Compar-ing vermicomposts and composts. BioCycle 39, 63-66.
Tadeo, F. 2000. Fisiología de las plantas y el estrés. pp. 481-498. En: Azcon-Bieto, J. y M. Talón (eds.). Fundamentos de fisiología vegetal. Mc Graw–Hill/Interamericana de España, Barcelona.
Wechsberg, G.E.; C.M. Bray y R.J. Probert. 1994. Expres-sion of dehydrin-like proteins in orthodox seeds of Ranunculus scleratus during development and water stress. Seed Sci. Res. 4, 241-246.
Wilches, F. y J. Álvarez. 2007. Tamaños de alveolo y diferentes láminas de riego en obtención de plán-tulas de tomate (Lycopersicon esculentum Mill.). Tra-bajo de grado. Facultad de Ciencias Agropecuarias, UPTC, Tunja.
EFECTO DE D IFERENTES LáMINAS DE R IEGO y SUSTRATOS EN LA pROpAGACIóN DE TOMATE

CArloS CArrAnZA1
oCtAvIo lAnCHero1
DIeGo MIrAnDA1, 2
Plantulas de tomate. Foto: álvarez-Herrera.
Comportamiento de los nutrientes en tejido foliar en brócoli (Brassica oleracea var. italica) ‘Coronado’ y repollo (Brassica oleracea) híbrido ‘Delus’ cultivados en la Sabana de Bogotá
Nutrient levels in foliar tissue of broccoli (Brassica oleracea var. italica) ‘Coronado’ and cabbage (Brassica oleracea) hybrid ‘Delus’ cultivated on the Bogota Plateau
reSUMen
De las Brassicas más importantes que se cultivan en la Sábana de Bogotá son el brócoli y el repollo, pero no existe información de los nutrientes en estrés salino. En el centro agropecuario Marengo de la Universidad Nacional de Colombia, en tres parcelas de 300 m2 cada una, con 8,3 y 11,1 plantas/m2 para brócoli y repollo respectivamente, regadas con aguas del distrito La Ramada, se realizaron muestreos cada ocho días y se deter-minó el contenido total de nutrientes N, P, K, Ca, Mg y B en el tejido foliar. Los datos obtenidos se ajustaron a curvas cuadráticas. En brócoli, los contenidos de N, P, Ca y Mg estuvieron entre los rangos adecuados los primeros 51 días después del trasplante (ddt) y posteriormente fueron deficientes. En repollo el P, Ca y Mg se ubicaron en los rangos adecuados. El K fue deficiente en todo el ciclo del cultivo para brócoli y repollo. El contenido del B fue alto en brócoli a los 19 ddt y en repollo a los 47 ddt. De acuerdo con la respuesta de las plantas, la presencia de sales tanto en el suelo como en el agua de riego influyó en la absorción, transporte distribución de los elementos minerales en las especies de Brassica evaluadas.
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 2 Autor para correspondencia. [email protected]
Brócoli en el Centro Agropecuario Marengo, Mosquera, Bogotá.Foto: C. Carranza
Palabras clave adicionales: absorción, deficiencia, macronutrientes, boro, salinidad.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 66-75, 2008

Vol. 2 - No. 1 - 2008
67
ABStrACt
The objective of this study was to evaluate changes in plant nutrient levels over time in broccoli and cabbage crops grown in the open field on the Bogota Plateau. The study was conducted at Agricultural Center Marengo of the National University of Colombia located at km 12 route Bogota-Mosquera, 2,516 m altitude above sea level, and characterized by an average temperature of 13.7 ºC, precipitation 669.9 mm year-1 and 4.2 daily sunshine hours, in a moderately saline soil. Three plots of 300 m2 each, with respectively 8.3 and 11.1 plants per m2 of broccoli and cabbage were irrigated with water of La Ramada district. Samples were taken every 8 days and the content of the plant nutrients N, P, K, Ca, Mg and B in foliar tissue was determined. Data generally fit a quadratic curve over time. In broccoli, the contents of N, P, Ca and Mg remained within an adequate range during the first 51 days after the transplanting (dat) but later became deficient. In cabbage, the contents of P, Ca and Mg were found to be within suitable ranges. Potassium was deficient during the whole crop cycle in both broccoli and cabbage. Boron was high in broccoli until 19 dat and, in cabbage, until 47 dat. In accordance with plant response, the presence of salts both in the soil and irrigation water probably influenced the absorption, transport and distribution of mineral elements in evaluated species of Brassicaceae.
Additional keywords: absorption, deficiency, macronutrients, boron, salinity.
Fecha de recepción: 19-12-2007 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
La producción agrícola mundial está ampliamente afectada por un sinnúmero de peligros ambien-tales, entre los cuales la salinidad es uno de los más importantes y está asociado con la sequía. Las tierras salinas no solamente están distribuidas en zonas desérticas o semidesérticas, también ocurren frecuentemente en suelos planos, fértiles, valles y regiones costeras densamente pobladas y con sistemas de irrigación (Sen et al., 2002). De 230 millones de hectáreas irrigadas, 45 millones han sido afectadas por sales (19,5%) y de 1.500 millones de hectáreas bajo agricultura de “tierras secas”, 32 millones están afectadas por sales en diferentes grados (Munns, 2003).
En la clasificación americana de suelos USDA (2006) Soil Taxonomy, se estableció que, a partir del valor de 2 dS m-1, el suelo se considera salino y las propiedades morfológicas y fisicoquímicas del perfil quedan influenciadas por el carácter salino. Por otra parte, el laboratorio de salinidad de los Estados Unidos estableció como límite 4
dS m-1 para que la salinidad comience a ser tóxi-ca para las plantas (Greenway y Munns, 1980; Marschner, 2002). Existe también un espectro continuo de tolerancia de plantas a condiciones salinas clasificándolas como glicófitas, que son sensibles a la sal hasta las halófitas, las cuales so-breviven en concentraciones muy altas de sales (Volkmar et al., 1998).
El estrés por salinidad es un fenómeno complejo e involucra no sólo el efecto osmótico, sino tam-bién el efecto de iones tóxicos y el desbalance nutricional (Ashraf, 2004). El factor iónico de la salinidad radica en la toxicidad iónica. Los iones que más problemas inducen son el cloruro (Cl-) y el sodio (Na+), aunque otros, como el nitrato (NO-
3), el sulfato (SO42-) el amonio (NH4
+) o el borato (H2BO3
-2) también pueden ser tóxicos. Su acumulación en las hojas reduce la tasa foto-sintética, induce la pérdida de pigmentos y pro-voca la senescencia y la abscisión (Azcón-Bieto y Talón, 2008).
COMpORTAMIENTO DE LOS NUTRIENTES EN TEj IDO FOL IAR

REV. COLOMB. CIENC. HORTIC.
68
El brócoli (Brassica oleracea var. italica) y repollo (Brassica oleracea) pertenecen a la familia Brassi-caceae y según la Encuesta Nacional Agropecua-ria realizada por el Ministerio de Agricultura y Desarrollo Rural en 2006, en Colombia se sem-braron 2.186 ha, donde se cosecharon 2.150 ha para un volumen total de 50.767 t (CCI, 2006) y para 2007 en la Sabana de Bogotá, se sembra-ron 500 ha con una producción anual de 14.457 t (Asohofrucol, 2007). Otras zonas productoras incluyen sectores de Cundinamarca, Boyacá, An-tioquia, Nariño, Tolima y Caldas (Semillas Arro-yave, 2007). El híbrido Delus es semi-tardío, se cosecha entre 115 y 120 días después del trasplan-te, tiene buena tolerancia a bajas temperaturas y es medianamente resistente a la salinidad (Semi-llas Arroyave, 2007).
A pesar de que el brócoli y el repollo son de las Brassicas más importantes que se cultivan en la Sabana de Bogotá, no existe información acerca del comportamiento de los nutrientes cuando la planta está en condiciones de estrés salino. Por lo anterior, esta investigación busca determinar el comportamiento de los contenidos de nutrientes en el tejido foliar de brócoli y repollo cultivados en un suelo salino de la Sabana de Bogotá.
MAterIAleS Y MÉtoDoS
El ensayo se realizó en el Centro Agropecuario Marengo de la Universidad Nacional de Colom-bia, sede Bogotá, ubicado en el km 12 vía Bogo-tá-Mosquera, con coordenadas 4° 42’ N; 74° 12’ W, a 2.543 msnm, temperatura anual 12,6 ºC, precipitación 669,9 mm año-1 y brillo solar 4,2 h diarias. Para cada especie, el diseño experimental consistió en tres repeticiones, con parcelas de 300 m2 por repetición, con la siembra de plántulas de
brócoli variedad ‘Coronado’ a una densidad de 8,3 plantas/m2 y para repollo híbrido ‘Delus’ en una densidad de 11,1 plantas/m2.
La fertilización se efectuó a partir de los resultados del análisis de suelo (tabla 1) realizado en el Labo-ratorio de Suelos y Aguas, Facultad de Agronomía, Universidad Nacional de Colombia. Se emplearon los siguientes métodos: carbono orgánico (CO): método de Walkley-Black; N: estimado a partir del CO (factor empleado: 0,0862); Ca, K, Mg, Na intercambiables: extracción con acetato de amonio 1N pH 7, valoración por absorción atómica; CIC (Capacidad de Intercambio Catiónico): desplaza-miento del NH4 intercambiado con NaCl 1M, va-loración volumétrica; P aprovechable: método de Bray II, valoración colorimétrica; Cu, Fe, Mn, Zn: extracción con DTPA, valoración por absorción atómica; B: extracción con fosfato monobásico (Azometina-H) (ICA, 1989; IGAC, 1990), valo-ración colorimétrica; Arcilla (Ar), limo (L), arena (A): método de Bouyoucos, previa dispersión con hexametafosfato de sodio; textura: triángulo de clasificación textural del USDA. Los nutrientes en deficiencia se suplementaron con fertilizantes químicos edáficos de mayores N 15%, P 30% y K 10%, y menores N 8,0% (Amoniacal 1,0% y Urei-co 7,0%); P2O5 5,0%; CaO 18,0%; MgO) 6,0%; S 1,6%; B 1,0%; Cu 0,75%; Mo 0,005%; Zn 2,5. En mezcla 1:1 con una dosis de 10 g/planta-1, los pri-meros 30 días, y posteriormente dos aplicaciones 45 y 70 días después del trasplante con N 15%, P 15% y K 15% en dosis de 20 g/planta. El suelo pre-sentó una conductividad eléctrica de 2,16 dS m-1 y el agua de riego 0,95 dS m-1. El manejo agronómi-co de malezas, plagas y enfermedades se efectuó oportunamente, dependiendo de las poblaciones, niveles y síntomas; finalmente, no se presentaron problemas relevantes que afectaran el desarrollo de las plantas.
pHCO N Ca K Mg Na CIC p Cu Fe Mn Zn B Ar L A
T% meq 100 g-1 mg kg-1 %
5,59 4,08 0,35 10,5 0,46 3,51 4,94 27,8 48,1 1,04 426 1,49 21,8 0,94 17 47 36 Fr
tabla 1. Características fisicoquímicas del suelo, Centro Agropecuario Marengo, Universidad nacional de Colombia.
CARRANZA/LANCHERO/MIRANDA

Vol. 2 - No. 1 - 2008
69
Los muestreos se realizaron cada ocho días en tres plantas por parcela y por repetición, para un total de siete muestreos en brócoli y ocho en repollo. En el Laboratorio de Agua y Suelos de la Facultad de Agronomía, Universidad Nacional de Colombia, se determinó el contenido total de nutrientes en el tejido foliar de acuerdo con las metodologías del ICA (1989) e IGAC (1990). N (nitrógeno total): método de micro-Kjeldahl, va-loración volumétrica; P: calcinación de la mues-tra, valoración calorimétrica con vanadato y mo-libdato de amonio; Ca, K, Mg: calcinación de la muestra, valoración por espectrofotometría de absorción atómica; B: calcinación de la muestra, valoración colorimétrica con Azometina-H. Los datos obtenidos fueron ajustados a una regresión cuadrática de la forma:
Y = ax2 + bx + c,
En la cual:
Y: la concentración del elemento
x : días después del trasplante
a, b y c: coeficientes del modelo
reSUltADoS Y DISCUSIÓn
Los cultivos pueden ser adversamente afectados por la salinidad, induciendo desórdenes nutricio-nales. Estos desórdenes pueden resultar de los efectos de la salinidad sobre la disponibilidad de nutrientes, respuesta competitiva, transporte o partición dentro de la planta (Grattan y Grieve, 1999). El contenido nutricional de las Brassicas es variable y depende de las condiciones ambienta-les en las que se desarrolla la planta, la edad de la misma y manejo del cultivo. En nuestro estudio, la concentración de nitrógeno total en el tejido foliar de brócoli presentó un mayor porcenta-je (5,78%) al inicio del cultivo, encontrándose en los rangos óptimos (4,0-6,5%) (Castellanos, 1998), y luego disminuyó a través del ciclo pro-
ductivo hasta el momento de la cosecha (2,78%), llegando al rango de deficiencia en las hojas (fi-gura 1A). En repollo, los rangos óptimos para el N en tejido foliar varían entre 3% y 4%, conside-rados niveles adecuados para el tejido foliar (Ma-rambio, 2004). En nuestro caso, la concentración obtenida fue alta (5,97%) en los muestreos iní-ciales y disminuyó progresivamente hasta los 68 días después del trasplante (ddt) (2,71%) (figura 2A). Por tanto, en los primeros 53 ddt el conteni-do de N estuvo en el rango óptimo y de ahí hasta el final del ciclo productivo estuvo en el rango de deficiencia. Este comportamiento fue inves-tigado por Greenway y Munns (1980), quienes aseguraron que la salinidad en muchos suelos incrementa el contenido de Na y, consecuente-mente, afecta la absorción de otros elementos minerales. Es posible que las disminuciones en el contenido de N en hojas también estén asociadas al alto consumo del N en las etapas intermedias del desarrollo del cultivo.
Las relaciones salinidad-nitrógeno en las espe-cies hortícolas son muy complejas. Sin embargo, la mayoría de estudios reportan que la toma y acumulación de N en el tallo de las plantas pue-de ser reducido bajo condiciones de salinidad, aunque otros estudios han encontrado respues-tas contrarias, o simplemente no han encontra-do efecto, como lo menciona (Feigin, 1985). El efecto de la salinidad sobre el consumo de ni-tratos varía considerablemente con la especie y las condiciones experimentales. En especies del Ricinus, la salinidad tiene efecto para asignar la reducción de nitratos en una alta proporción en las raíces (Peuke et al., 1996). Cramer et al. (1995) concluyen que el consumo en la raíces de NO3
- en solución es inhibida por la salinización; con-secuentemente, las concentraciones de NO3
- en las hojas y tallos con alta actividad del nitrato reductasa es reducida por la salinidad, en compa-ración con las plantas sin tratamiento salino.
En brócoli, la concentración de fósforo en el tejido foliar fue alta (0,54%) al inicio del culti-vo y disminuyó a 0,39%; el rango óptimo para
COMpORTAMIENTO DE LOS NUTRIENTES EN TEj IDO FOL IAR

REV. COLOMB. CIENC. HORTIC.
70
este nutriente varía entre 0,45 y 0,8%; en los primeros 51 ddt, el contenido de P estuvo entre los rangos óptimos sugeridos por Castellanos (1998) (figura 1B). En repollo, el comportamien-to de la concentración de P (figura 2B) fue alto a los 19 ddt (0,54%), disminuyó hasta los 47 ddt (0,42%) y nuevamente aumentó hasta los 68 ddt (0,48%). El rango óptimo de P en tejido foliar es de 0,4-0,7% (Marambio, 2004), por consiguiente la concentración del P estuvo entre los rangos óp-timos. El P es un constituyente de ácidos nuclei-cos, fosfolípidos, fosfoproteínas, dinucleótidos y adenosin trifosfato. De ahí que es requerido para diferentes procesos, incluyendo almacenamiento y transferencia de energía, fotosíntesis, la regu-lación de enzimas y el transporte de carbohidra-tos (Hu y Schmidhalter, 2005). Papadopoulous y Rendig (1983), Sharpley et al. (1992), Grattan y Grieve (1999) reportan que disminuciones en el P total en el tejido de las plantas tienden a estar relacionadas con incrementos en la salinidad y, en particular, con las concentraciones de Ca2+ en el sustrato. Esto puede ser atribuido a la for-mación de fosfatos de Ca2+, así la concentración de Ca2+ se incrementa en el suelo haciéndolo no disponible para la planta. Patel y Pandey (2007) encontraron en plántulas de Cassia montana que el P y Ca2+ son factores limitantes para el cre-cimiento a salinidades de 10 dS m-1. Grattan y Grieve (1999) mencionan que en suelos salinos la disponibilidad del fosfato es reducida princi-palmente por procesos de absorción y por la baja solubilidad de los minerales de Ca2+ y P.
El potasio es un elemento esencial para la síntesis de proteínas, enzimas glicolíticas y fotosíntesis, un potencial osmótico durante la expansión ce-lular y turgor; también es un competidor de Na+ bajo condiciones salinas (Marschner, 2002). En brócoli, el comportamiento del K+ en tejido foliar presentó una alta concentración en el muestreo inicial (2,94%), disminuyó progresivamente has-ta los 51 ddt (1,95%) y aumentó levemente hasta los 61 ddt (2,06%). Es posible que esta disminu-ción en los contenidos de K+ esté asociada con el efecto competitivo con el Na+, pero también con
el hecho conocido de que esta especie es altamen-te consumidora de K+ en las etapas intermedias del cultivo. La concentración ideal de K+ en el te-jido foliar se ubica en el rango de 3,5-6,0% (Cas-tellanos, 1998). De acuerdo con nuestros resulta-dos, se presentaría una deficiencia marcada en la concentración de este elemento en el tejido foliar (figura 1C). En repollo, la concentración de K+ en el tejido foliar (figura 2C) a los 19 ddt fue 2,2% y aumentó progresivamente a los 68 ddt hasta 3,23%, siendo éste elemento deficiente durante todo el ensayo, ya que el rango óptimo de K+ se encuentra entre 4,5 y 7,5% (Marambio, 2004). El K+ es el soluto inorgánico más importante en plantas y como tal hace una mayor contribu-ción al bajo potencial osmótico en la estela de la raíz, que es un indispensable prerrequisito para conducir la presión de turgor en el transporte de solutos en el xilema y el balance hídrico en las plantas (Marschner, 2002). La deficiencia de K+ inducida por Na+ está implicada en la reducción del crecimiento y rendimiento de varios cultivos, incluyendo tomate (López y Satti, 1996; Song y Fujiyama, 1996), espinaca (Chow et al., 1990), hinojo (Graifenberg et al., 1996), y maíz (Botella et al., 1997). La reducción en el consumo de K+ en plantas por Na+ es un proceso competitivo y ocurre aún a pesar de todo si la solución es domi-nada por sales de Na+, de Cl- o de SO4
2-
En brócoli, la concentración de calcio en tejido foliar indica un alto porcentaje (2,44%) al inicio del cultivo, disminuyendo progresivamente has-ta los 47 días (0,98%) y aumentando levemente hasta el momento de la cosecha (1,37%). En los primeros 23 ddt, la concentración de Ca2+ total en la hoja se encontró en el rango óptimo (2-4%) (Castellanos, 1998), mientras que al final del cul-tivo estuvo en un rango de deficiencia, indicando cierta influencia de la salinidad del suelo en la absorción de Ca2+ por parte de la planta (figura 1D). En repollo, la concentración de Ca2+ (figura 2D) en el tejido foliar fue alto a los 19 ddt (2,6%) y disminuyó hasta la cosecha a los 68 días con 1,07%, valores considerados ligeramente por de-bajo del rango de suficiencia (1,9-6,0%) determi-
CARRANZA/LANCHERO/MIRANDA

Vol. 2 - No. 1 - 2008
71
nado por Marambio (2004). El Ca2+ desempeña un papel esencial en la integridad estructural y funcional de las membranas de las plantas, esta-bilidad de la pared celular, regulación de trans-porte de iones y permeabilidad de membranas, control del intercambio iónico, así como la ac-tividad enzimática de la pared celular (Rengel, 1992; Marschner, 2002). En las plantas, el Na+ puede desplazar el Ca2+ de las membranas ce-lulares, modificando la absorción de nutrientes, como el K+. Sin embargo, se ha observado que si existe un aporte suplementario de Ca2+ en el proceso de absorción de K+ funciona bien, en de-trimento del Na+. Así, la fertilización con Ca2+ mejora la producción agrícola al incrementar la absorción de K+ y reducir el flujo de NO3, PO4 (Azcón-Bieto y Talón, 2008). Concentraciones adecuadas de Ca2+ pueden tener un papel im-portante en la tolerancia de las plantas a con-diciones salinas (Collins et al., 2008). LaHaye y Epstein (1969) explicaron el papel crucial del Ca2+ en la regulación de la salinidad en las plan-tas y específicamente en el transporte selectivo o exclusión de Na+ y otros iones minerales por las membranas celulares de las plantas. Diversos estudios reportan que la adición de Ca2+ al me-dio de crecimiento atenúa el estrés por salinidad (Rengel, 1992). Ashraf y Naqvi (1992) reportan que suplementar con Ca2+ en la presencia de sa-linidad mejoró el crecimiento de Brassica juncea y B. napus, pero no de B. carinata y B. rapa, lo que indica una respuesta diferencial entre los genoti-pos a la suplementación del Ca2+. Ciertamente, altos niveles de Na inhiben la absorción de Ca2+ y K+, que resulta en un antagonismo entre Na+ y K+ (Benlloch et al., 1994). Los cationes Na+ y K+ son conocidos por ser el mayor componente inorgánico del potencial osmótico (Asch et al., 1999). En Brassicas Ashraf y McNeilly (2004) sugieren utilizar una relación alta K+/Na+ en el tejido como criterio para tolerancia a salinidad.
La concentración de magnesio en tejido foliar de brócoli fue creciente en los primeros 20 ddt
(0,54%), descendiendo hasta los 52 ddt (0,11%) y aumentando ligeramente hasta los 61 ddt (0,14%). Por lo tanto, en los primeros 33 ddt la concentra-ción total de Mg2+ se ubica dentro de los rangos óptimos de suficiencia (0,25-0,5%) (Castellanos, 1998). En los días posteriores, se presentó en un rango de deficiencia hasta la cosecha (figura 1E). En repollo, la concentración de Mg2+ (figura 2E) fue alta a los 19 ddt (0,5%) y disminuyó hasta los 68 ddt (0,25%). En general su concentración estuvo en los rangos óptimos de suficiencia (0,2-0,7%) sugeridos por Marambio (2004). Ruiz et al. (1997) encontraron que la salinidad por NaCl re-dujo la concentración foliar de Mg2+ en cítricos. Hu y Schmidhalter (1997) demostraron que la concentración de Mg2+ en hojas de trigo se redu-jo, pero podría ser aumentado para un nivel de salinidad dado, por incrementos en la concentra-ción de Mg2+ en la solución nutritiva.
El boro es un elemento esencial requerido para el crecimiento normal de las plantas superiores. La deficiencia de B es un problema extendido en muchos cultivos agrícolas (Shorrocks, 1997). En el tejido foliar de brócoli, la concentración de B (figura 1F) fue alta a los 19 ddt (25,91 mg kg-1), se redujo levemente hasta los 40 ddt (17,38 mg kg-1) y aumentó hasta los 61 ddt, obteniendo una concentración de 25,5 mg kg-1. Los cambios en las concentraciones de B pueden estar relacionados con los altos requerimientos de este nutriente por las Brassicas en general. Estudios previos mues-tran que la deficiencia de B disminuye la capa-cidad fotosintética de las plantas (Kastori et al., 1995; Zhao y Oosterhuis, 2002, 2003). En repollo (figura 2F), a los 19 ddt la concentración de B fue de 19,14 mg kg-1, aumentó hasta los 47 ddt (28,97 mg kg-1) y disminuyó hasta los 68 ddt (23,83 mg kg-1). La deficiencia de B es un principal problema que impide el crecimiento del cultivo y general-mente conduce al rápido cese de la elongación de la raíz, reduce la expansión foliar, principalmen-te debido a la reducción en la expansión celular (Dell y Huang, 1997; Marschner, 2002).
COMpORTAMIENTO DE LOS NUTRIENTES EN TEj IDO FOL IAR

REV. COLOMB. CIENC. HORTIC.
72
Finalmente, la salinidad presente en el suelo y en el agua de riego disminuyeron en brócoli la concentración de N, P, Ca2+ y Mg2+ en el tejido foliar en la etapa intermedia y al final del ciclo del cultivo, coincidiendo con los resultados obte-nidos por Cuartero et al. (1992) y Pérez-Alfocea et al. (1996). En tomate, en el cual la salinidad
Figura 1. Concentraciones de los nutrientes minerales en tejido foliar en brócoli. A) nitrógeno; B) fósforo; C) potasio; D) calcio; e) magnesio; F) boro.
CARRANZA/LANCHERO/MIRANDA
disminuyó en las hojas concentraciones de K+, Ca2+, Mg2+ y NO3
-, las plantas pueden tomar más K+, Ca2+, Mg2+ y NO3- desde el medio si las relaciones Na+/K+, Na+/Ca2+, Na+/Mg2+ y el equilibrio de los demás nutrientes son similares al tratamiento control; esto es más evidente en el desarrollo activo de hojas jóvenes.

Vol. 2 - No. 1 - 2008
73
ConClUSIoneS
•La presencia de sales tanto en el suelo como en el agua de riego influyó en la absorción, trans-porte y distribución de los elementos minera-les en brócoli y repollo.
•En brócoli, las concentraciones de N, P, Ca y Mg en tejido foliar estuvieron en los rangos adecuados los primeros 51 ddt y posteriormen-te estuvieron en deficiencia.
Figura 2. Concentraciones de los nutrientes minerales en tejido foliar en repollo. A) nitrógeno; B) fósforo; C) potasio; D) calcio; e) magnesio; F) boro.
COMpORTAMIENTO DE LOS NUTRIENTES EN TEj IDO FOL IAR

REV. COLOMB. CIENC. HORTIC.
74
•En repollo, los contenidos de P, Ca y Mg en te-jido foliar se presentaron en los rangos adecua-dos, por ende en esta Brassica se observó cierto nivel de tolerancia a la salinidad, la cual se ma-nifestó en un buen estatus de estos nutrientes.
•En todo el ciclo productivo del brócoli y repo-llo, se observó deficiencia de K en tejido foliar, mostrando que la salinidad incidió en la dis-ponibilidad de este elemento en la solución del suelo y en la absorción por las plantas.
reFerenCIAS BIBlIoGrÁFICAS
Asch, F.; M. Dingkuhn; C. Wittstock y K. Doerffling. 1999. Sodium and potassium uptake of rice pani-cles as affected by salinity and season in relation to yield and yield components. Plant Soil 207, 133-145.
Ashraf, M. y M.I. Naqvi. 1992. Growth and ion uptake of 4 Brassica species as affected by Na+/Ca2+ ratio in saline sand culture. Z. Pflanzenernähr. Bodenkd. 155, 101-108.
Ashraf, M. 2004. Some important physiological selec-tion criteria for salt tolerance in plants. Depart-ment of Botany, University of Agriculture, Islam-abad, Pakistan.
Ashraf, M. y T. McNeilly. 2004. Salinity tolerance in Brassica oilseeds. Crit. Rev. Plant Sci. 23, 157-174.
Asohofrucol. 2007. Brócoli y repollo. En: http://frutasyhor-talizas.com.co; consulta: 15 de diciembre de 2007.
Azcón-Bieto, J. y M. Talón. 2008. Fundamentos de fi-siología vegetal. 2da ed. McGraw-Hill Interameri-cana, Madrid.
Benlloch, M.; M.A. Ojeda; J. Ramos y A. Rodríguez-Navarro. 1994. Salt sensitivity and low discrimi-nation between potassium and sodium in bean plants. Plant Soil 166, 117-23.
Botella, M.A.; V. Martínez; M. Nieves y A. Cerdá. 1997. Effect of salinity on the growth and nitrogen up-take by wheat seedlings, J. Plant Nutr. 20, 793-804.
Castellanos, J.Z. 1998. El seguimiento de la nutrición del cultivo de brócoli en los sistemas de fertirrigación. Instituto Nacional de Investigaciones Forestales y Agropecuarias. En: http://www.ppi-far.org; con-sulta: 10 de diciembre de 2006.
CCI. 2006. Entorno nacional. pp. 41-44. En: Plan hortíc-ola nacional. Corporación Colombia Internacional, Bogotá.
Chow, W.S.; M.C. Ball y J.M. Anderson. 1990. Growth and photosynthetic responses of spinach to salin-ity: Implications of K nutrition for salt tolerance. Aust. J. Plant Physiol. 17, 563-578.
Collins, R.P.; P.J.C. Harris; M.J. Bateman y J. Henderson. 2008. Effect of calcium and potassium nutrition on yield, ion content, and salt tolerance of Brassica campestris (rapa). J. Plant Nutr. 31, 1461-1481.
Cramer, M.D.; A. Schierholt; Y.Z. Wang y S.H. Lips. 1995. The influence of salinity on the utilization of root anaplerotic carbon and nitrogen metabolism in tomato seedlings. J. Exp. Bot. 46, 1569-1577.
Cuartero, J.; A.R. Yeo y T.J. Flowers. 1992. Selection of donors for salt-tolerance in tomato using physi-ological traits. New Phytol. 121, 63-69.
Dell, B. y L.B. Huang. 1997. Physiological response of plants to low boron. Plant Soil 193, 103-120.
Feigin, A. 1985. Fertilization management of crops irri-gated with saline water. Plant Soil 89, 285-299.
Graifenberg, A.; L. Botrini; L. Giustiniani y M. Lipucci Di Paola. 1996. Salinity affects growth, yield and elemental concentration of fennel. HortScience 31, 1131-1134.
Grattan, S. y C. Grieve. 1999. Salinity-mineral nutrient relations in horticultural crops. Scientia Hort. 78, 127-157.
Greenway, H. y R. Munns. 1980. Mechanisms of salt tolerance in non-halophytes. Annu. Rev. Plant Physiol. 31, 149-190.
Hu, Y. y U. Schmidhalter. 1997. Interactive effects of sa-linity and macronutrient level on wheat. II. Com-position. J. Plant Nutr. 20, 1169-1182.
Hu, Y. y U. Schmidhalter. 2005. Drought and salinity: a comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 168, 541-549.
CARRANZA/LANCHERO/MIRANDA

Vol. 2 - No. 1 - 2008
75
ICA. 1989. El análisis de suelos, plantas y aguas para riego. Manual de Asistencia Técnica No. 47. Bogotá.
IGAC. 1990. Métodos analíticos del Laboratorio de Sue-los. 5a ed. Bogotá.
LaHaye, P.A. y E. Epstein. 1969. Salt toleration by plants: Enhancement with calcium. Science 166, 395-396.
López, M.V. y S.M.E. Satti. 1996. Calcium and potas-sium-enhanced growth and yield of tomato under sodium chloride stress. Plant Sci. 114, 19-27.
Kastori, R., M. Plesnicar, D. Pankovic y Z. Sakac. 1995. Photosynthesis, chlorophyll fluorescence and sol-uble carbohydrates in sunflower leaves as affected by boron deficiency. J. Plant Nutr. 18, 1751-1763.
Marambio, C.C. 2004. Curvas de acumulación de mate-ria seca y nutrientes en cultivares de repollo de Bru-selas. Tesis de Doctorado. Facultad de Agronomía, Universidad Católica de Chile, Santiago.
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, London.
Munns, R. 2003. Comparative physiology of salt and water stress. Plant Cell Environ. 25, 239-250.
Papadopoulous, I. y V.V. Rendig. 1983. Interactive ef-fects of salinity and nitrogen on growth and yield of tomato plants. Plant Soil 73, 47-57.
Patel, A.D. y A.N. Pandey. 2007. Effect of soil salinity on growth, water status and nutrient accumulation in seedlings of Cassia Montana (Fabaceae). J. Arid Environ. 70, 174-182.
Pérez-Alfocea, F.; M.E. Balibrea; A. Santa Cruz y M.T. Estaño. 1996. Agronomical and physiological char-acterization of salinity tolerance in a commercial tomato hybrid. Plant Soil 180, 251-257.
Peuke, A.D., J. Glaab, W.M. Kaiser y W.D. Jeschke. 1996. The uptake and flow of C, N and ions between roots and shoots in Ricinus communis L. IV. Flow and metabolism of inorganic nitrogen and malate depending on nitrogen nutrition and salt treat-ment. J. Exp. Bot. 47, 377-385.
Rengel, Z. 1992. The role of calcium in salt toxicity. Plant Cell Environ. 15, 625-632.
Salisbury, F. y C.W. Ross. 1994. Fisiología vegetal. Grupo Editorial Iberoamericana, México.
Ruiz, D.; V. Martínez y A. Cerda. 1997. Citrus response to salinity: growth and nutrient uptake. Tree Physiol. 17, 141-150.
Sharpley, A.N., J.J. Meisinger, J.F. Power y D.L. Suarez. 1992. Root extraction of nutrients associated with long-term soil management. Adv. Soil Sci. 19, 151-217.
Shorrocks, V.M. 1997. The occurrence and correction of boron deficiency. Plant Soil 193, 121-48.
Semillas Arroyave. 2007. Repollo Delus. En: http://www.semillasarroyave.com; consulta: 11 de diciem-bre de 2007.
Sen, D.N.; P.K. Kasera y S. Mohammad. 2002. Biology and physiology of saline plants. pp. 563-580. En: Pessaraki, M (ed.). Handbook of plant and crop physiology. 2nd ed. Marcel Dekker, New York, NY.
Song, J.Q. y H. Fujiyama. 1996. Difference in response of rice and tomato subjected to sodium saliniza-tion to the addition of calcium. Soil Sci. Plant Nutr. 42, 503-510.
USDA. 2006. Key to soil taxonomy by soil survey staff. United States Government Printing Office, Washington.
Volkmar, K.M.; Y. Hu y H. Steppuhn. 1998. Physiologi-cal responses of plants to salinity: a review. Can. J. Plant Sci. 78, 19-27.
Zhao, D. y D.M. Oosterhuis. 2002. Cotton carbon ex-change, nonstructural carbohydrates, and boron distribution in tissues during development of bo-ron deficiency. Field Crops Res. 78, 75-77.
Zhao, D. y D.M. Oosterhuis. 2003. Cotton growth and physiological responses to boron deficiency. J. Plant Nutr. 26, 855-867.
COMpORTAMIENTO DE LOS NUTRIENTES EN TEj IDO FOL IAR

lIllIAnA HoYoS-CArvAJAl1,4
GlorIA DUQUe2
SerGIo orDUZ P.3
Plantulas de tomate. Foto: álvarez-Herrera.
Antagonismo in vitro de Trichoderma spp. sobre aislamientos de Sclerotinia spp. y Rhizoctonia spp.
Antagonism of Trichoderma spp. against isolates of Sclerotinia spp. and Rhizoctonia spp. in vitro
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).2 Colegio Mayor de Antioquia, Medellín (Colombia).3 Facultad de Ciencias, Universidad Nacional de Colombia, Medellín (Colombia).4 Autor para correspondencia. [email protected]
Cultivo de habichuela.Foto: C. Carranza
reSUMen
El género Trichoderma consta de diversas especies de hongos que tienen la capacidad de parasitar a otros hongos que causan enfermedades en plantas. En el presente estudio, el objetivo fue evaluar la actividad micopara-sítica in vitro de 73 aislamientos de Trichoderma spp. sobre tres aislamientos del fitopatógeno Sclerotinia spp. provenientes de habichuela, repollo y pimentón y dos aislamientos de Rhizoctonia provenientes de arroz y de fríjol; se cuantificó evaluando el porcentaje de inhibición del crecimiento micelial o antagonismo siguiendo la metodología de cultivos duales y la calificación de micoparasitismo planteados por Royse y Ries (1978). Para analizar los datos obtenidos en los ensayos, se llevó a cabo la prueba de Kruskall-Wallis, detectando diferen-cias estadísticas significativas en el grado de parasitismo de aislamientos de Trichoderma spp. sobre los hongos fitopatógenos evaluados. Los aislamientos de Trichoderma con alto grado de micoparasitismo fueron aquellos capaces de combatir a los fitopatógenos en cuatro de las cinco pruebas realizadas, entre los cuales sobresalen T. asperellum T51 y T. harzianum T53. El aislamiento T. harzianum T21demostró tener un grado de antagonismo muy bajo en todas las pruebas realizadas.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 76-86, 2008

Vol. 2 - No. 1 - 2008
77
Palabras clave adicionales: control biológico, hongo fitopatógeno, actividad microparasítica, T. asperellum T51, T. harzianum T53.
ABStrACt
The genus Trichoderma includes various species of fungi that are able to parasitize other fungi, some of which cause plant disease. The goal of the present study was to evaluate, in vitro, the parasitic activity of 73 fungal isolates of Trichoderma spp. on three isolates of the phytopathogens Sclerotinia spp. obtained from green beans, cabbage and pepper, and two isolates of Rhizoctonia spp. obtained from rice and beans. We studied percentage inhibition of mycelial growth or antagonism, using the methodology of dual culture and the classification of mycoparasitism of Royse y Ries (1978). Data was subjected to the Kruskall-Wallis test to detect statistically significant differences in the degree of parasitism of isolates of Trichoderma spp. on the evaluated phyto-pathogenic fungi. T. asperellum T51 and T. harzianum T53 stood out using the criterion of successful control of the phytopathogens in four of five tests. In contrast, the isolate T. harzianum T21 demonstrated very little antagonism in all tests.
ANTAGONISMO IN V ITRO DE Tr i choderma spp .
Additional keywords: biological control, phytopathogenic fungi, mycoparasitic activity, T. asperellum T 51, T. harzianum T 53.
Fecha de recepción: 08-04-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
El manejo de enfermedades de plantas ocasiona-das por patógenos del suelo son verdaderos retos en la agricultura mundial que por su complejidad requieren un trato más cuidadoso que en el caso de los patógenos de la aerobiota (Castro, 1995). Las enfermedades no sólo tienen el potencial de destruir enteramente las cosechas, aun en los casos en que no causan pérdidas totales, por lo general reducen en forma crónica el rendimiento de la mayoría de los cultivos, obligando a tomar medidas de combate que aumentan los costos de producción y afectan la calidad y la durabili-dad de los productos cosechados, de manera que constituyen una de las principales causas de ines-tabilidad en la empresa agrícola y del déficit ali-mentario mundial (Strange y Scott, 2005). Más
aún aquellas causadas por patógenos formadores de esclerocios, los cuales son agrupaciones com-pactas de micelio, ricos en materiales de reserva que pueden permanecer en estado de dorman-cia, hasta 10 años (Schwartz y Pastor-Corrales, 1988). En este grupo de patógenos se encuentran los géneros Rhizoctonia spp. y Sclerotinia spp.
Rhizoctonia solani Khum tiene un micelio incolo-ro en las primeras etapas de formación y poste-riormente se torna amarillo o café cuando ma-dura; consta de células largas con ramificaciones que crecen en ángulo recto con respecto a la hifa principal (Ulloa y Hanlin, 2002). Los síntomas más comunes sobre su hospedero son el ahoga-miento de las plántulas y la pudrición de la raíz,

REV. COLOMB. CIENC. HORTIC.
78
así como la pudrición y el cáncer del tallo de las plantas adultas y en proceso de crecimiento. Sin embargo, en algunos hospederos también produ-ce pudrición de los órganos vegetales de almace-namiento, así como los tizones o manchas del follaje, especialmente del que se encuentra cerca del suelo (Agrios, 2005). El hongo se disemina principalmente por el agua de riego, por imple-mentos agrícolas y por material de propagación vegetativa (Gilligan et al., 1996).
Sclerotinia sclerotiorum produce hifas blancas en sus estados iniciales; más tarde el micelio se va compactando y toma un aspecto marrón o ne-gro para formar los esclerocios, los cuales son de principal importancia en la supervivencia y epidemiología de Sclerotinia sclerotiorum (Cundom et al., 2002). Los síntomas que produce Scleroti-nia varían un poco con el hospedero o la parte afectada de él y con las condiciones del medio ambiente. Las enfermedades causadas por Scle-rotinia se conocen bajo una amplia variedad de nombres, siendo las más comunes: la pudrición algodonosa, moho blanco o pudrición blanda aguanosa, pudriciones del tallo y maderamen, marchitez, ahogamiento, pudrición de la corona, tizón de las inflorescencias, nudo rosa, cáncer del tallo, pudriciones de la raíz y corona, entre otras (Agrios, 2005).
En cuanto a métodos de control utilizados para combatir estos hongos, se ha centrado en el uso de agroquímicos, los cuales contribuyen a la acu-mulación de residuos tóxicos en las cosechas y en el ambiente, con serias consecuencias para la salud humana. Además, los pesticidas no permi-ten un control efectivo de muchas enfermedades producidas por fitopatógenos del suelo por no ser selectivos. Una alternativa de control ha sido ba-sada en el control biológico, entendido como la reducción de la densidad o de las actividades pro-ductoras de enfermedades de un patógeno o pa-rásito, en su estado activo o durmiente, lograda de manera natural o a través de la manipulación del ambiente, del hospedero o de antagonistas del
patógeno o plaga que se quiere controlar (Chin-cholkar y Mukerji, 2007). Esta forma de control de algunos problemas fitosanitarios ha tomado importancia en los últimos años, fundamentada principalmente en la selección de organismos del suelo con propiedades antagónicas sobre otros organismos que producen enfermedades en las plantas (Fravel, 2005), ya que la utilización de microorganismos antagónicos o enmiendas or-gánicas se constituyen en una alternativa de control de estos fitopatógenos para mejorar la nutrición y resistencia de las plantas, así como disminuir la incidencia de enfermedades (Nico et al., 2005). El control biológico surge como una alternativa ante la necesidad de reducir los pro-ductos químicos, conservando la sanidad de los cultivos. Además, el componente más significa-tivo del suelo que afecta la supervivencia de los esclerocios parece ser el biológico; entre los cua-les se encuentran más de 30 especies de hongos y bacterias señaladas como antagonistas (Cundom et al., 2002).
Los antagonistas utilizados para control de en-fermedades son generalmente saprófitos, debido a su facilidad de adaptación al medio y a su alta capacidad de competencia por nutrientes frente a otros organismos, a su versatilidad y fácil mani-pulación. Entre estos se halla el género Trichoder-ma, que produce tres tipos de propágulos: hifas, clamidosporas y conidios, las cuales son activas contra fitopatógenos en diferentes fases del ciclo de vida, desde la germinación de la espora has-ta la esporulación, además ésta reúne una serie de características en su interacción directa con el fitopatógeno, que, según Harman (2000) y Howell (2006), hace de este organismo un buen agente antagonista de hongos fitopatógenos. En-tre ellas su capacidad micoparasítica, la cual pue-de ser definida como el uso del patógeno como alimento por su antagonista, en el que general-mente se ven implicadas enzimas extracelulares como quitinasas, celulasas, β-1-3-glucanasas y proteasas que lisan o digieren las paredes de los hongos; su capacidad de producción de antibióticos,
HOyOS-CARVAjAL/DUQUE/ORDUZ p.

Vol. 2 - No. 1 - 2008
79
referida a la producción de sustancias tóxicas para otros organismos, las cuales actúan en ba-jas concentraciones (menores a 10 mg L-1); y a su alta competencia por nutrientes y espacio, entendida como el desigual comportamiento de dos o más organismos ante un mismo requerimiento, siem-pre y cuando la utilización del mismo por uno de los organismos reduzca la cantidad disponi-ble para los demás. Un factor esencial para que exista competencia es que se halla en cantidades insuficientes de un elemento, si hay exceso no hay competencia. La competencia más común es por nutrientes, oxígeno o espacio y, por último, la desactivación de las enzimas de los patógenos (Benítez et al., 2004).
El género Trichoderma es un Ascomycete, pertene-ciente al orden Hypocreales Mitospóricos, a la fa-milia Hypocreaceae, cuyo estado anamorfo es Tri-choderma, y su telomorfo es Hypocrea. Este hongo es cosmopólita en suelo y se halla en madera en descomposición y en material vegetal, las especies de Trichoderma son frecuentes componentes domi-nantes de la microflora del suelo en una amplia variedad de hábitats, esta capacidad es atribuida al diverso potencial metabólico de las especies de Trichoderma y a su agresiva competencia natural (Samuels, 2006). Trichoderma spp. es bien conoci-do por la producción de enzimas líticas y la pene-tración de hifas de éste en el hongo fitopatógeno, tal fenómeno ha sido considerado como la base del antagonismo. Además, se ha demostrado que la interacción de Trichoderma spp. con su huésped es específica y controlada por componentes pre-sentes en la pared celular del fitopatógeno, lo cual hace que estos microorganismos sean reconocidos y posteriormente atacados.
El objetivo de este trabajo fue evaluar la capaci-dad antagónica de 73 aislamientos de Trichoderma spp de la colección de hongos filamentosos de la Unidad de Biotecnología y Control Biológico de la CIB sobre algunas especies de fitopatógenos tales como Sclerotinia spp. y Rhizoctonia spp.
MAterIAleS Y MÉtoDoS
Hongos antagonistas empleados: colección de 73 aislamientos de Trichoderma previamen-te identificados (Hoyos-Carvajal et al., 2008), pertenecientes a la colección de hongos filamen-tosos de la Corporación para Investigaciones Biológicas, en Medellín (tabla 1) procedentes de Colombia, Ecuador y Perú, los cuales se cultiva-ron en medio selectivo para Trichoderma (Elad et al., 1980) y en Agar Papa Dextrosa (PDA), según se requiriera.
Hongos fitopatógenos: colección de hongos fitopatógenos formadores de esclerocios los cua-les se encontraban en la Unidad de Biotecnología y Control Biológico de la CIB; Sclerotinia prove-niente de habichuela (Phaseolus vulgaris), Scleroti-nia proveniente de repollo (Brassica oleracea var. capitata), Sclerotinia proveniente de pimentón (Capsicum spp.), Rhizoctonia proveniente de arroz (Oriza sativa), Rhizoctonia proveniente de fríjol (Phaseolus vulgaris); la cual fue recuperada de plántulas de fríjol, su identificación se realizó de acuerdo con las características descritas por Bar-nett y Hunter (1998). Estos aislamientos fueron cultivados en PDA (tabla 2).
Pruebas de antagonismo in vitro: cada uno de los aislamientos de Trichoderma spp. y cada fito-patógeno se evaluó según la metodología descri-ta a continuación. En una caja de Petri con PDA se pone un disco de 5 mm de diámetro de un cultivo del hongo fitopatógeno durante el tiem-po necesario para que este inicie su crecimiento micelial. Transcurrido éste periodo de acondicio-namiento, se siembra al lado opuesto, a una dis-tancia de 3 cm, un disco de 5 mm de diámetro de un cultivo de cada aislamiento de Trichoderma spp. Se prepararon tres réplicas por cada pare-ja de disco y se dejaron en incubación hasta la toma de datos; bajo condiciones de 25 a 30ºC. Como control, se utiliza un disco de micelio del patógeno sin el antagonista.
ANTAGONISMO IN V ITRO DE Tr i choderma spp .
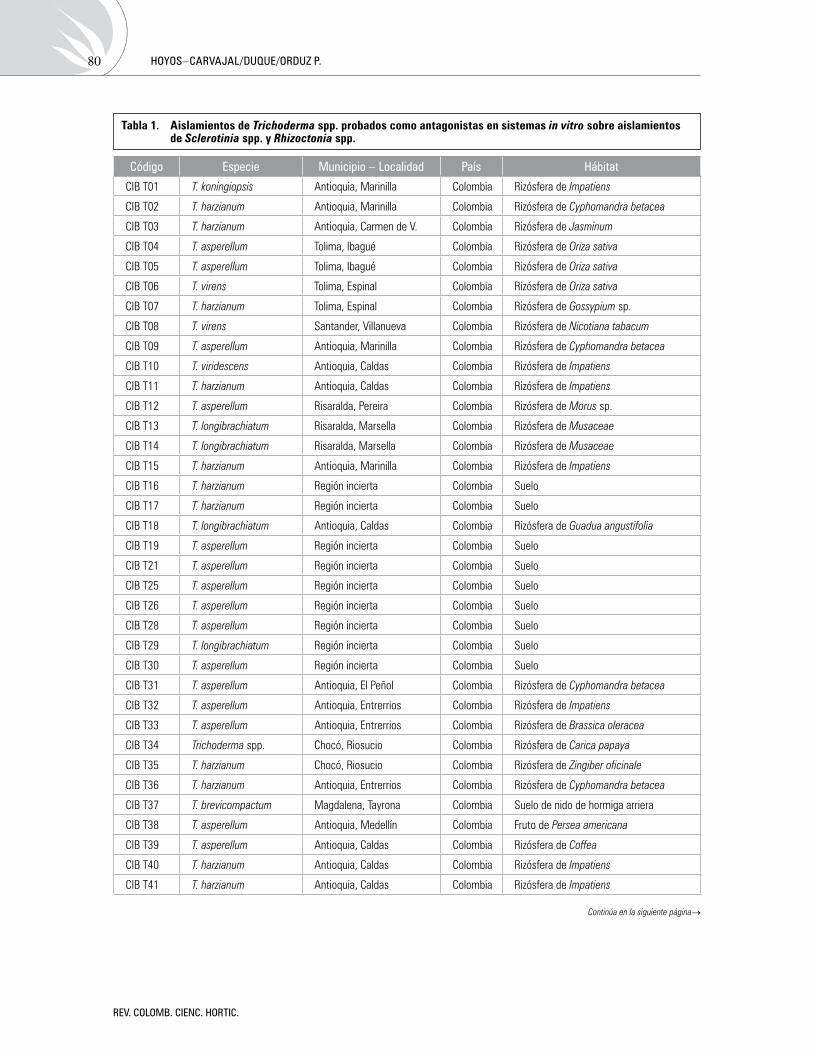
REV. COLOMB. CIENC. HORTIC.
80
Código Especie Municipio - Localidad país Hábitat
CIB T01 T. koningiopsis Antioquia, Marinilla Colombia Rizósfera de Impatiens
CIB T02 T. harzianum Antioquia, Marinilla Colombia Rizósfera de Cyphomandra betacea
CIB T03 T. harzianum Antioquia, Carmen de V. Colombia Rizósfera de Jasminum
CIB T04 T. asperellum Tolima, Ibagué Colombia Rizósfera de Oriza sativa
CIB T05 T. asperellum Tolima, Ibagué Colombia Rizósfera de Oriza sativa
CIB T06 T. virens Tolima, Espinal Colombia Rizósfera de Oriza sativa
CIB T07 T. harzianum Tolima, Espinal Colombia Rizósfera de Gossypium sp.
CIB T08 T. virens Santander, Villanueva Colombia Rizósfera de Nicotiana tabacum
CIB T09 T. asperellum Antioquia, Marinilla Colombia Rizósfera de Cyphomandra betacea
CIB T10 T. viridescens Antioquia, Caldas Colombia Rizósfera de Impatiens
CIB T11 T. harzianum Antioquia, Caldas Colombia Rizósfera de Impatiens
CIB T12 T. asperellum Risaralda, Pereira Colombia Rizósfera de Morus sp.
CIB T13 T. longibrachiatum Risaralda, Marsella Colombia Rizósfera de Musaceae
CIB T14 T. longibrachiatum Risaralda, Marsella Colombia Rizósfera de Musaceae
CIB T15 T. harzianum Antioquia, Marinilla Colombia Rizósfera de Impatiens
CIB T16 T. harzianum Región incierta Colombia Suelo
CIB T17 T. harzianum Región incierta Colombia Suelo
CIB T18 T. longibrachiatum Antioquia, Caldas Colombia Rizósfera de Guadua angustifolia
CIB T19 T. asperellum Región incierta Colombia Suelo
CIB T21 T. asperellum Región incierta Colombia Suelo
CIB T25 T. asperellum Región incierta Colombia Suelo
CIB T26 T. asperellum Región incierta Colombia Suelo
CIB T28 T. asperellum Región incierta Colombia Suelo
CIB T29 T. longibrachiatum Región incierta Colombia Suelo
CIB T30 T. asperellum Región incierta Colombia Suelo
CIB T31 T. asperellum Antioquia, El Peñol Colombia Rizósfera de Cyphomandra betacea
CIB T32 T. asperellum Antioquia, Entrerrios Colombia Rizósfera de Impatiens
CIB T33 T. asperellum Antioquia, Entrerrios Colombia Rizósfera de Brassica oleracea
CIB T34 Trichoderma spp. Chocó, Riosucio Colombia Rizósfera de Carica papaya
CIB T35 T. harzianum Chocó, Riosucio Colombia Rizósfera de Zingiber oficinale
CIB T36 T. harzianum Antioquia, Entrerrios Colombia Rizósfera de Cyphomandra betacea
CIB T37 T. brevicompactum Magdalena, Tayrona Colombia Suelo de nido de hormiga arriera
CIB T38 T. asperellum Antioquia, Medellín Colombia Fruto de Persea americana
CIB T39 T. asperellum Antioquia, Caldas Colombia Rizósfera de Coffea
CIB T40 T. harzianum Antioquia, Caldas Colombia Rizósfera de Impatiens
CIB T41 T. harzianum Antioquia, Caldas Colombia Rizósfera de Impatiens
tabla 1. Aislamientos de Trichoderma spp. probados como antagonistas en sistemas in vitro sobre aislamientos de Sclerotinia spp. y Rhizoctonia spp.
Continúa en la siguiente página→
HOyOS-CARVAjAL/DUQUE/ORDUZ p.
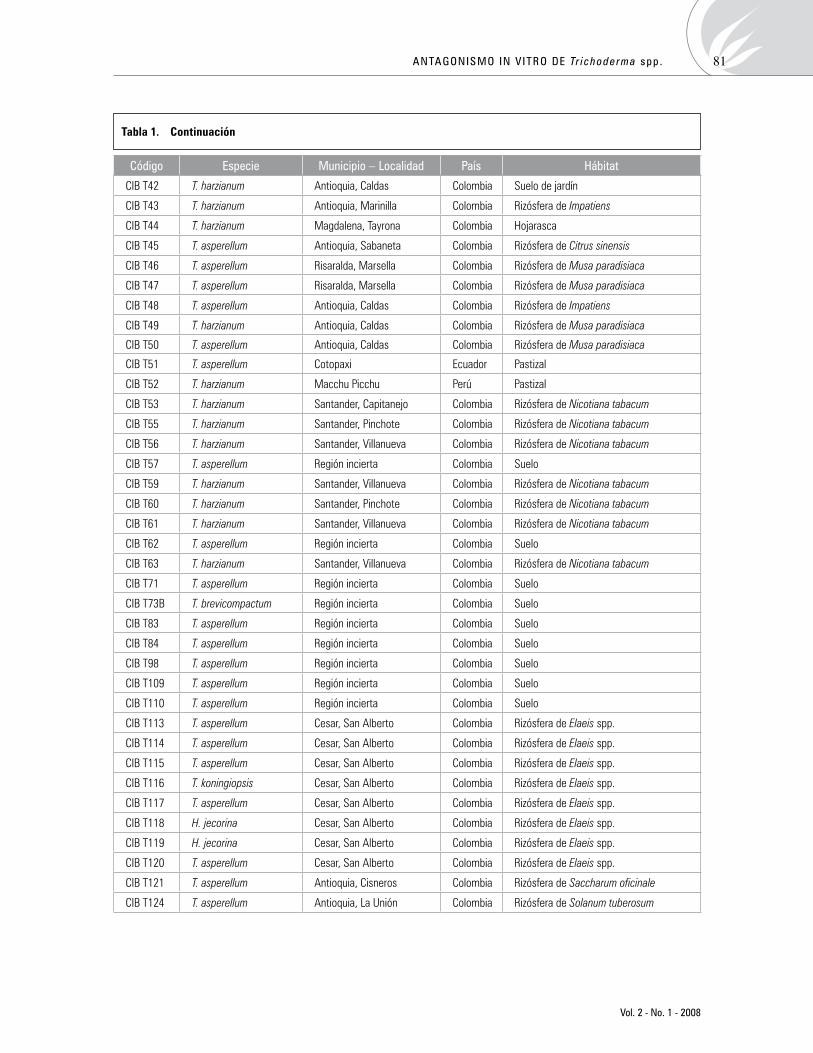
Vol. 2 - No. 1 - 2008
81
Código Especie Municipio - Localidad país Hábitat
CIB T42 T. harzianum Antioquia, Caldas Colombia Suelo de jardín
CIB T43 T. harzianum Antioquia, Marinilla Colombia Rizósfera de Impatiens
CIB T44 T. harzianum Magdalena, Tayrona Colombia Hojarasca
CIB T45 T. asperellum Antioquia, Sabaneta Colombia Rizósfera de Citrus sinensis
CIB T46 T. asperellum Risaralda, Marsella Colombia Rizósfera de Musa paradisiaca
CIB T47 T. asperellum Risaralda, Marsella Colombia Rizósfera de Musa paradisiaca
CIB T48 T. asperellum Antioquia, Caldas Colombia Rizósfera de Impatiens
CIB T49 T. harzianum Antioquia, Caldas Colombia Rizósfera de Musa paradisiaca
CIB T50 T. asperellum Antioquia, Caldas Colombia Rizósfera de Musa paradisiaca
CIB T51 T. asperellum Cotopaxi Ecuador Pastizal
CIB T52 T. harzianum Macchu Picchu Perú Pastizal
CIB T53 T. harzianum Santander, Capitanejo Colombia Rizósfera de Nicotiana tabacum
CIB T55 T. harzianum Santander, Pinchote Colombia Rizósfera de Nicotiana tabacum
CIB T56 T. harzianum Santander, Villanueva Colombia Rizósfera de Nicotiana tabacum
CIB T57 T. asperellum Región incierta Colombia Suelo
CIB T59 T. harzianum Santander, Villanueva Colombia Rizósfera de Nicotiana tabacum
CIB T60 T. harzianum Santander, Pinchote Colombia Rizósfera de Nicotiana tabacum
CIB T61 T. harzianum Santander, Villanueva Colombia Rizósfera de Nicotiana tabacum
CIB T62 T. asperellum Región incierta Colombia Suelo
CIB T63 T. harzianum Santander, Villanueva Colombia Rizósfera de Nicotiana tabacum
CIB T71 T. asperellum Región incierta Colombia Suelo
CIB T73B T. brevicompactum Región incierta Colombia Suelo
CIB T83 T. asperellum Región incierta Colombia Suelo
CIB T84 T. asperellum Región incierta Colombia Suelo
CIB T98 T. asperellum Región incierta Colombia Suelo
CIB T109 T. asperellum Región incierta Colombia Suelo
CIB T110 T. asperellum Región incierta Colombia Suelo
CIB T113 T. asperellum Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T114 T. asperellum Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T115 T. asperellum Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T116 T. koningiopsis Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T117 T. asperellum Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T118 H. jecorina Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T119 H. jecorina Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T120 T. asperellum Cesar, San Alberto Colombia Rizósfera de Elaeis spp.
CIB T121 T. asperellum Antioquia, Cisneros Colombia Rizósfera de Saccharum oficinale
CIB T124 T. asperellum Antioquia, La Unión Colombia Rizósfera de Solanum tuberosum
tabla 1. Continuación
ANTAGONISMO IN V ITRO DE Tr i choderma spp .

REV. COLOMB. CIENC. HORTIC.
82
Lectura de ensayos: en cada ensayo se evalúan los criterios de porcentaje de inhibición del cre-cimiento micelial del patógeno (Royse y Ries, 1978). Después de sembrar el antagonista en las cajas, la actividad de los aislamientos de Tricho-derma se clasifica de acuerdo con el grado basado en una escala de 1 a 5 en la cual 1 es el 100% de micoparasitismo y 5 es 0% de micoparasitismo. Un aislamiento se considera antagonista contra el patógeno si el promedio es ≥3. A un trata-miento que obtenga un promedio entre 2 y 3 no se le asigna clasificación.
Análisis estadísticos: para determinar dife-rencias en el efecto de las cepas de Trichoderma sobre la inhibición del radio micelial del patóge-no (P≤0,05) se analiza con la prueba de Krus-kall-Wallis (estadística no paramétrica) y para detectar diferencias entre tratamientos se hacen gráficos de Box and Wisker plots, en los que se determina cuáles de los tratamientos (aislamien-tos de Trichoderma) presentan mayor grado de micoparasitismo.
reSUltADoS Y DISCUSIÓn
Según la técnica descrita por Royse y Ries (1978) para la realización de pruebas de antagonismo, debe haber un periodo de acondicionamiento del hongo fitopatógeno, el cual se ve reflejado en el crecimiento sobre el medio de cultivo y varía con respecto a la tasa de crecimiento de cada fitopa-
tógeno; para Sclerotinia 48 h (los tres aislamien-tos), para Rhizoctonia de arroz menos de 12 h, y para Rhizoctonia de fríjol 24 h.
En los resultados obtenidos de los análisis esta-dísticos en la prueba de Kruskall-Wallis, un valor de P≤0,05 indica que en todos los tratamientos se encontraron diferencias significativas, como se muestra en la tabla 3. Según los análisis Box and Wister plots, se encontró que los aislamien-tos T. asperellum T51 y T. harzianum T53 resul-taron tener antagonismo in vitro sobre cinco de los seis aislamientos de fitopatógenos evaluados. Estos aislamientos antagónicos pertenecen a especies diferentes y proceden de hábitats muy diferentes del que fueron tomados los fitopató-genos. También puede afirmarse que la mayoría de hongos con actividad antagónica específica sobre los diferentes fitopatógenos corresponde a aislamientos de T. asperellum, especie que consti-tuye el 49% de aislamientos probados, así como T. harzianum, que es el 31,5%.
En contraste, el aislamiento que demostró tener un grado de antagonismo mas bajo en las prue-bas realizadas corresponde a T. harzianum T21, lo que indica que aislamientos de una misma especie de Trichoderma son específicos en su ac-tividad micoparasítica, por tanto, no se puede generalizar al decir que determinadas especies de Trichoderma son buenas antagonistas, ya que esto depende del aislamiento en particular y su capacidad de atacar de forma concreta a cepas
Código Género Hospedero localidad
Sp 1 Sclerotinia spp. Pimentón (Capsicum annuum var. annuum) Rionegro, Antioquia
Sh 2 Sclerotinia spp. Habichuela (Phaseolus vulgaris var.vulgaris ) Santuario, Antioquia
Sr 3 Sclerotinia spp. Repollo (Brassica oleracea var. capitata) Santuario, Antioquia
Ra 1 Rhizoctonia spp. Arroz (Oriza sativa) Palmira, Cali
Rf 2 Rhizoctonia spp. Fríjol (Phaseolus vulgaris) Santuario, Antioquia
tabla 2. Aislamientos de Sclerotinia spp. y Rhizoctonia spp. usados para pruebas de antagonismo con Trichoderma spp.
HOyOS-CARVAjAL/DUQUE/ORDUZ p.

Vol. 2 - No. 1 - 2008
83
determinadas de hongos fitopatógenos. Esto es soportado por numerosos reportes de literatu-ra en los cuales diversas especies de Trichoderma han sido reportadas como antagonistas en hon-gos formadores de esclerocios y es claro que son atributos de aislamientos particulares, no de especies (Elad et al., 1980; Elad 1983; Hjeljord y Tronsmo, 1998; Howell, 2006).
En cuanto a T. brevicompactum T37, T. koningiopsis y T. reseii T118 y T119, presentan antagonismo contra un único aislamiento de los fitopatóge-nos probados. De éstas, T. brevicompactum es una especie recientemente descrita en la literatura (Krauss et al., 2004), que se halla restringida al Caribe y de la cual no existen antecedentes como micoparásito de fitopatógenos.
En este estudio se probaron 73 aislamientos de Trichoderma spp. mediante ensayos in vitro que presentaron grado de micoparasitismo variable, calificado con la escala diseñada para tal efecto (figura 1), en los cuales se observó que la tasa de crecimiento del fitopatógeno no afecta la activi-dad micoparasítica de las especies de Trichoderma spp., es decir, una invasión temprana del medio
de crecimiento por parte del fitopatógeno no im-pide o compite por crecimiento con Trichoderma, por el contrario, este crecimiento micelial precoz del hongo fitopatógeno puede ser rápidamen-te colonizado y parasitado por Trichoderma spp. Pero esta observación sólo puede llevarse a cabo excediendo los tiempos de lectura recomendados por los autores de la escala de micoparasitismo (Royse y Ries, 1978), como se observó en los en-sayos con los aislamientos de Rhizoctonia spp.
Aunque generalmente las especies de Trichoder-ma son de crecimiento rápido y en pocas horas pueden cubrir la superficie de la caja de Petri, que un aislamiento sea de crecimiento lento no lo excluye como buen antagonista, ya que según Cundom et al. (2002), dicha característica, la tasa de crecimiento, no es obstáculo para que en ta-les hongos puedan encontrarse aislamientos de buen potencial antagónico.
Por tanto, las pruebas in vitro para determinar la capacidad antagónica de un microorganismo con respecto a otro no representan necesariamente el grado de antagonismo y de control biológico en condiciones naturales, pero reflejan la capacidad
Código Género valor P Aislamientos de Trichoderma con mayor grado de micoparasitismo1
Sp 1 Sclerotinia spp. 0,00T2, T4, T5, T6, T7, T9, T16, T17, T25, T26, T28, T31, T32, T33, T34, T35, T36, T40, T46, T50, t51, t53, T56, T83.
Sh 2 Sclerotinia spp. 169x10-12 T2, T4, T5, T6, T9, T16, T17, T25, T26, T28, T31, T32, T33, T34, T35, T36, T40, T46, T50, t51, t53.
Sr 3 Sclerotinia spp. 3.818x10-10 T3, T7.
Ra 1 Rhizoctonia spp. 4.61x10-5T3, T4, T6, T7, T10, T11, T12, T14, T17, T19, T25, T26, T28, T32, T33, T34, T35, T36, T37, T40, T46, T47, T49, T50, T51, T53, T55, T57, T59, T60, T71, T84, T98, T110, T113, T114, T115, T119, T123.
Rf 2 Rhizoctonia spp. 3.229x10-10 T43, t51, t53, T55, T56, T62, T63, T98, T114, T115, T116, T117, T118, T119, T123.
tabla 3. Aislamientos de Trichoderma con alto grado de antagonismo in vitro sobre Sclerotinia spp. y Rhizoctonia spp. según prueba Kruskall-Wallis.
1 Según escala Royse y Ries (1978).
Aislamientos en negrilla son antagónicos para la mayoría de fitopatógenos evaluados.
ANTAGONISMO IN V ITRO DE Tr i choderma spp .

REV. COLOMB. CIENC. HORTIC.
84
y la variabilidad genética del antagonista y la del fitopatógeno de invadir o crecer en una caja de Petri o en condiciones in vitro. Es de anotar que di-chas pruebas y la escala planteada permiten bási-camente cuantificar competencia por nutrientes en el medio controlado, mas no cuantificar como tal el micoparasitismo o antagonismo; porque en
tal caso se podría afirmar que si un aislamiento fitopatogénico tiene una tasa de crecimiento mas rápida que el aislamiento del Trichoderma, enton-ces éste sería antagonista evaluado como grado 3, 4 y 5 en la escala de Royse y Ries (1978) (ta-bla 4 y figura 1). Las pruebas de antagonismo in vitro permiten realizar la selección preliminar de
aislamientos con rápido crecimiento en sistemas controlados, es decir, miden competencia, no así micoparasitismo, para el cual se requieren ob-servaciones microscópicas en las que se vea una interacción directa entre estructuras del antago-nista y del fitopatógeno y, en virtud a esto, eva-luar la capacidad micoparasítica. Invariablemen-te, las pruebas de antagonismo in vitro requieren ser corroboradas en condiciones de invernadero y campo, porque no siempre los hongos que actúan como antagonistas bajo condiciones controladas tienen la capacidad potencial de usarse en el con-trol biológico, como se ha demostrado en otros experimentos con aislamientos usados en este es-tudio (Hoyos-Carvajal et al., 2008). Esto se debe a que el hongo antagonista puede modificar o ac-tuar diferente de acuerdo con las características del suelo y las sucesiones microbianas particula-res en sistemas in vivo (Garveba et al., 2004).
Figura 1. Colonización Trichoderma spp. (a) sobre micelio de Sclerotinia (f).
f
a
↓
↓
Grado MicoparasitismoPorcentaje de cubrimiento
del antagonista de la superficie del medio
1 El antagonista cubre en un 100% el micelio del fitopatógeno.
100 %
2El antagonista cubre en un 75% el micelio
del fitopatógeno.75%
3 El antagonista cubre en un 50% el micelio
del fitopatógeno.50%
4El antagonista cubre en un 25% el micelio
del fitopatógeno.25%
tabla 4. escala de micoparasitismo, según royse y ries (1978).
HOyOS-CARVAjAL/DUQUE/ORDUZ p.

Vol. 2 - No. 1 - 2008
85
AGrADeCIMIentoS
A la Unidad de Biotecnología y Control Biológico de la Corporación para Investigaciones Biológicas (CIP), Medellín, por la financiación de este estudio.
reFerenCIAS BIBlIoGrÁFICAS
Agrios, G. 2005. Fitopatología. 2a ed. Editorial Limusa, México D.F.
Barnett, H.L. y B.B. Hunter. 1998. Illustred genera of im-perfect fungi. 4th ed. American Phytopathological Society, New York, NY.
Benítez, T.; A.M. Rincón; M.C. Limón y A.C. Codón. 2004. Biocontrol mechanisms of Trichoderma strains. Intl. Microbiol. 7(4), 249-260.
Castro, B.L. 1995. Antagonismo de algunos aislamientos de Trichoderma koningii, originados en suelo colom-biano contra Rosellinia bunodes, Sclerotinia sclerotium y Pythium ultimum. Fitopatol. Colomb. 19(2), 7-18.
Chincholkar, S. y K.J. Mukerji. 2007. Biological control of plant diseases. Haworth Press, New York, NY.
Cundom, M.A.; S.M. Mazza de Gaiad; M.A. Mazzati de Castañon; S.A. Gutiérrez de Arriola y M. Coutin-ho. 2002. Actividad antagónica in vitro de Hongos saprófitos sobre Sclerotinia sclerotiorum. En: http://www.unne.edu.ar/cyt/agrarias/a-037.pdf; consul-ta: abril de 2008.
Elad, Y.; I. Chet y J. Katan. 1980.Trichoderma harzianum: a biocontrol agent effective against Sclerotinia rolfsii and Rhizoctonia solani. Phytopathol. 70(2), 119-121.
Elad, Y.; I.Chet; P. Boyle y Y. Henis. 1983. Parasitismo of Trichoderma spp. on Rhizoctonia solani and Sclerotium rolfsii scanning electron mycroscopy and fluores-cence microscopy. Phytopathol. 73, 85-88
Fravel, D.R. 2005. Commercialization and implementa-tion of biocontrol. Annu. Rev. Phytopathol. 43(2), 337-359.
Garveba, P.; J.A. van Veen y J.D. van Elsas. 2004. Micro-bial diversity in soil: Selección of microbial popula-tions by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 42(2), 243-270
Gilligan, C.A.; S.A. Simons y H.A. Hide. 1996. Inoculum density and spacial pattern of Rhizoctonia solani in field crops of Solanum tuberosum: effects of crop-ping frequency. Plant Pathol. 45(2), 232-244.
Harman, G. 2000. Myth and dogmas of biocontrol chang-es in perceptions derived from research on Trichoder-ma harzianum T22. Plant. Dis. 84(4), 377-393.
Hjeljord, L. y A. Tronsmo. 1998. Trichoderma and Glio-cladium in biological control: an overview. pp. 131-151. En: Harman, G.E. y C.P. Kubicek (eds.). Trichoderma and Gliocladium. Vol. 2. Enzymes, bio-logical control and commercial applications. Taylor & Francis, London.
Hoyos-Carvajal, L.; P. Chaparro; M. Abramsky; I. Chet y S. Orduz. 2008. Evaluación de aislamientos de Trichoderma spp. contra Rhizoctonia solani y Sclero-tium rolfsii bajo condiciones in vitro y de inverna-dero. Agron. Colomb. 26(3), 451-458.
Howell, C.R. 2006. Understanding the mechanisms em-ployed by Trichoderma virens to effect biological con-trol of cotton diseases. Phytopathol. 96(2), 178-180
Kraus, G.F.; I. Druzhinina; W. Gams; J. Bissett; D. Zafari; G. Szakacs; A. Koptchinski; H. Prillinger; R. Zare y C.P. Kubiek. 2004. Trichoderma brevicompactum sp. nov. Mycol. 96(5), 1059-1073.
Nico, A.I.; C.I. Mónaco; G. Dal Bello y H. Alippi. 2005. Efecto de la adición de enmiendas orgánicas al
ANTAGONISMO IN V ITRO DE Tr i choderma spp .

REV. COLOMB. CIENC. HORTIC.
86
suelo sobre la capacidad patogénica de Rhizoctonia solani: ii. Micoflora asociada y antagonismo in vitro de los aislados más frecuentes. Rev. Inv. Agrop. 34(1), 29-44.
Royse, D.J. y S.M. Ries. 1978. The influence of fungi isolated from peach twigs on the pathogenecity of Cytospora cincta. Phytopathol. 68, 603-607
Samuels, G.J. 2006. Trichoderma: Systematics, the sexual state, and ecology. Phytopathol. 96(2), 195-206.
Schwartz, H.F. y M. Pastor-Corrales. 1988. El moho blanco del frijol y su control. Guía de estudio. Cen-tro Internacional de Agricultura Tropical (CIAT), Cali, Colombia.
Strange, R.N. y P.R. Scott. 2005. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 43(2), 83-116.
Ulloa, M. y R.T. Hanlin. 2000. Illustrated dictionary of mycology. APS Press, St. Paul, MI.
HOyOS-CARVAjAL/DUQUE/ORDUZ p.

JeIMY M. PoveDA1
JoHn W. MArtÍneZ1, 2
Plantación de papa en el municipio de tunja, vereda Puente de Boyacá.Foto: j.M. poveda.
Abundancia y diversidad de Bacillus thuringiensis de diferentes hábitat en tres municipios de Boyacá, Colombia
Abundance and diversity of Bacillus thuringiensis in different habitats within three municipalities of Boyaca, Colombia
1 Grupo Manejo Biológico de Cultivos, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Co-lombia (UPTC), Tunja (Colombia).
2 Autor para correspondencia. [email protected]
reSUMen
La actual orientación de la agricultura hacia la sostenibilidad y los estudios recientes sobre el impacto social y económico que ha causado el abuso de plaguicidas químicos ha generado un creciente interés por promover desarrollos en el campo de los bioplaguicidas que contribuyan a la solución de problemas empleando estrate-gias acordes con las condiciones de cada región. Bacillus thuringiensis (Bt) es un microorganismo cosmopolita que se constituye en una herramienta valiosa para el control biológico de plagas. Asimismo, es reconocido entre los plaguicidas microbianos de mayor importancia comercial debido a su toxicidad específica contra insectos blanco e inocuidad para el hombre y los animales. El objetivo de este trabajo fue comparar la distri-bución, abundancia y diversidad de Bt provenientes de diferentes hábitats existentes en tres municipios de la provincia Centro de Boyacá (Chíquiza, Siachoque y Tuta), con el fin de iniciar una colección de este micro-organismo en el laboratorio de Control Biológico de la UPTC. De 102 aislamientos se obtuvieron 51 cepas de Bt de las cuales el 78% provenía de ecosistemas agrícolas. Del total de bacterias viables de Bt en los bosques fueron obtenidas 0,25-2,90 105 ufc/mL y 0,09-2,98 105 ufc/mL en cultivos de maíz, con índices de diversidad de 1,00. Los resultados mostraron la presencia de Bt en todos los municipios, aun en aquellos sitios perturbados constantemente por las labores agrícolas.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 87-97, 2008

REV. COLOMB. CIENC. HORTIC.
88
ABStrACt
The current orientation of agriculture toward sustainable development, and studies related to economic and social impacts of chemical pesticide abuse have increased interest in biopesticide technology. This has led to the creation of novel solutions specific the phytosanitary problems of each region. Bacillus thuringiensis (Bt) is a cosmopolitan microorganism that has provided the most valuable biological pest control tool worldwide, due to its specificity and low risk to humans and animals. The objective of the present work was to compare distribution, abundance and diversity of Bt from different habitats in three localities (Chiquiza, Siachoque y Tuta) of the central province of Boyaca, Colombia, with the goal of initiating a Bt collection in the Laboratory of Biological Control of UPTC. Starting from a 102 isolates, 51 Bt strains were obtained, out of which 78% originated from agricultural systems. Viable Bt bacteria were obtained from forests (0.25 to 2.90 105 cfu/mL), and maize crops (0.09 to 2.98 105 cfu/mL), with diversity indexes of 1.00. These results indicated that the Bt is present in the three localities studied, even in sites usually disturbed by agricultural activities.
Palabras clave adicionales: bacterias del suelo, biodiversidad, bioinsecticida, control biológico.
Additional keywords: soil bacteria, biodiversity, bioinsecticides, biological control.
Fecha de recepción: 10-03-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
Boyacá es un departamento netamente agrícola con diversidad de suelos que surte de alimentos a una buena parte de la región central y norte de Colombia. Uno de los factores que más dificul-ta la producción agrícola en el departamento es el manejo de los problemas fitosanitarios, cau-sados por enfermedades y plagas, cuyo control representa un alto porcentaje en la estructura de costos del cultivo. Existe en las zonas una arrai-gada tradición de uso de agroquímicos, los cua-les, además de ser ineficientes en muchos casos, generan resistencia en los insectos plaga, afectan al medio ambiente, contaminan las cosechas y, en muchos casos, tienen efectos directos en la salud de los productores o consumidores.
En un mercado internacional, en el que la com-petencia y las restricciones frente a la presencia
de residuos químicos adquieren gran importan-cia, la agricultura colombiana y boyacense debe promover el uso de alternativas de producción limpia que partan de nuestra diversidad, per-mitan la producción sostenible de los cultivos, garanticen la protección del medio ambiente, fa-vorezcan la calidad y con un beneficio común. Actualmente, se conocen estrategias biológicas de manejo de plagas desarrolladadas a partir de la exploración de la diversidad microbiana de di-ferentes regiones.
Bacillus thuringiensis (Bt) es una bacteria entomo-patógena del suelo, aeróbica facultativa, gram positiva, caracterizada por la producción de un cuerpo paraesporal o cristal de proteínas conoci-das como delta-endotoxinas o proteínas Cry. Es-tos cristales se forman durante el proceso de es-
pOVEDA/MARTÍNEZ

Vol. 2 - No. 1 - 2008
89
porulación y en ellas reside la actividad tóxica de Bt hacia larvas de diferentes órdenes de insectos, cuando son ingeridas (Hofte y Whiteley, 1989; Feitelson et al., 1992; Cerón et al., 1995; Ibarra et al., 2003; Tamez-G. et al., 2005). Dichos cristales presentan diferentes formas: bipiramidales, esfé-ricas, cuadradas, entre otras (Lynch y Baumann, 1985; Höfte y Whiteley, 1989; Rodríguez et al., 1991). Se ha demostrado que las inclusiones de esta bacteria contienen, principalmente, proteí-nas de 25 a 140 KDa (Restrepo et al., 1997).
Dadas las características de Bt como agente de control biológico y debido a su presencia cosmo-polita, sus esporas se han aislado de diversas fuen-tes como suelo (Martin y Travers, 1989; Hossain et al., 1997; Bravo et al., 1998), agua (Iriarte et al., 2000), hojas de plantas (Wlodzimierz y Elzbieta, 1999), insectos muertos (Chilcot y Wigley, 1993; Kaelin et al., 1994; Apoloyo et al., 1995; Kim et al., 1998), productos almacenados (Meadows et al., 1992; Kaelin et al., 1994; Xavier et al., 2007) y telarañas. Los bioinsecticidas derivados de esta bacteria se han utilizado por más de 40 años (Ibarra y López, 2000).
El uso de productos a base de Bt se ha genera-lizado principalmente en cultivos de hortalizas, cereales y algodón, representando más del 90% de los bioplaguicidas que se comercializan en el mundo (Yara et al., 1999; Vallejo y Orduz, 1996; Bravo y Cerón, 2004; Adams et al., 1996), cons-tituyéndose en el insecticida de origen biológico de mayor importancia y eficiencia en el control biológico de diversas plagas agrícolas en el mun-do (Feitelson et al., 1992); todo esto ha conducido al establecimiento de colecciones que suman cer-ca de 40.000 aislados, concentradas en grandes firmas privadas que controlan el mercado mun-dial (Lambert y Peferoen, 1992).
Algunos de los factores que han hecho posible su éxito en la agricultura, además de su aproba-ción por la Organización Mundial de la Salud, son su alta biodegradabilidad, alta especificidad
(Deacon, 1983) e inocuidad para mamíferos, otros vertebrados, plantas e inclusive otros in-sectos benéficos (Shelton et al., 2002; Ibarra et al., 2003), siendo una alternativa promisoria y ambientalmente segura en el control de insectos plaga nocivos.
Por otro lado, la aparición de resistencia a algunas toxinas de Bt (Restrepo et al., 1998) y el control ineficiente de los insecticidas químicos (Pérez et al., 1997), indica que la utilización masiva de bioinsecticidas a base de Bt debe estar apoyada por productos que puedan competir, en precio y disponibilidad, con los insecticidas químicos (Vallejo y Orduz, 1996); esto ha llevado tanto a las industrias como a los laboratorios y centros de investigación a enfocar sus esfuerzos en la búsqueda de nuevas moléculas, especialmente de bacterias entomopatógenas, para el desarrollo de alternativas novedosas de control (Bravo et al., 1998; Silva et al., 1999), lo cual ha contribuido a ampliar el conocimiento de la distribución eco-lógica de este microorganismo en la naturaleza (Uribe et al., 2003). Actualmente, los volúmenes de uso de esta bacteria están limitados al número de insectos susceptibles a los productos comer-ciales existentes, necesitándose aislar y probar nuevas cepas y aislamientos.
En Colombia, algunos grupos de investigación han caracterizado cepas nativas de Bt, enfocan-do el trabajo hacia lepidópteros como Spodopetra frugiperda, Heliothis virescens (Cerón et al., 1995) y T. solanivora (Martínez et al., 2002) y dípteros implicados en problemas de salud pública como Aedes aegyptii y Culex quinquefasciatus (Díaz et al., 1993). Cada vez más se reconoce el papel e importancia de la biodiversidad en el funciona-miento de los sistemas agrícolas; sin embargo, hasta el momento, trabajos sistemáticos sobre caracterización de la diversidad de cepa nativas de Bt en Boyacá no se han desarrollado, limitan-do la generación de nuevos registros de Bt para Boyacá y Colombia que puedan llegar a ser de importancia económica regional o nacional.
ABUNDANCIA y D IVERS IDAD DE Bac i l l us thu r i ng iens i s

REV. COLOMB. CIENC. HORTIC.
90
El objetivo de este trabajo fue comparar la dis-tribución, abundancia y diversidad de Bt prove-nientes de diferentes hábitats existentes en tres municipios de la provincia Centro de Boyacá
MAterIAleS Y MÉtoDoS
Recolección de muestras. Un total de 32 mues-tras de suelo fueron recolectadas en bolsas plás-
ticas estériles, en 12 diferentes hábitas existen-tes en zonas de bosques naturales, pantanos y zonas agrícolas de los municipios de Chiquiza, Siachoque, y Tuta de la Provincia, Centro del departamento de Boyacá (tabla 1). Las muestras se recolectaron eliminando totalmente la vege-tación que cubría el suelo en cada sitio, tomando para cada muestra, con una cuchara diferente, 250 g de suelo, hasta una profundidad de 7 cm. Las muestras se llevaron al laboratorio, donde se
origenMuestrade suelo
no. de bacterias x 106/ml antes de la pasteurización
no. de esporas formadas
x 105/ml
no. de B. thuringiensis
x 105/ml
Porcentaje de B.
thuringiensis
Índice de diversidad1
Bosquesnaturales
1 0,31 2,87 (9,3) 2,90 9,3 1,00 (5/5)2 0,69 1,61 (2,4) 0,25 0,4 0,50 (1/2)3 0,20 3,01 (15,1) 1,29 6,5 0,29 (2/7)4 0,17 0,71 (4,3) 0,00 0,0 0,00 (0/0)5 0,33 1,22 (8,9) 0,00 0,0 0,00 (0/1)6 0,40 0,48 (1,2) 0,00 0,0 0,00 (0/0)
Pantanos 7 0,48 1,32 (2,8) 1,31 2,8 1,00 (3/3)
Maíz
8 0,18 2,91 (16,1) 2,91 16,1 1,00 (8/8)9 0,25 2,08 (8,5) 1,05 4,3 0,67 (2/3)10 0,17 2,97 (18,0) 2,98 18,0 1,00 (2/2)11 0,65 2,76 (4,2) 0,09 0,1 022 (2/9)
trigo 12 2,86 1,14 (0,4) 0,00 0,0 0,00 (0/1)
Avena13 2,89 0,67 (0,2) 0,00 0,0 0,00 (0/1)14 0,34 1,20 (3,6) 0,16 0,5 0,43 (3/7)
leguminosas15 0,25 2,98 (11,9) 0,00 0,0 0,00 (0/3)16 0,82 2,74 (3,3) 0,00 0,0 0,00 (0/1)17 1,26 0,63 (0,5) 0,63 0,5 1,00 (2/2)
Papa
18 0,26 2,16 (8,3) 0,05 0,2 0,50 (1/2)19 0,30 0,15 (0,5) 0,59 2,0 0,67 (2/3)20 0,83 0,44 (0,5) 0,16 0,2 0,50 (2/4)21 0,11 0,05 (0,4) 0,00 0,0 0,00 (0/0)22 1,21 0,46 (0,4) 0,46 0,4 1,00 (2/2)
Pastos23 0,28 0,76 (2,8) 0,23 0,8 0,33 (1/3)24 0,40 2,41 (6,0) 0,06 0,2 0,33 (2/6)
rábano25 2,88 1,25 (0,4) 0,00 0,0 0,00 (0/1)26 0,61 1,17 (1,9) 0,18 0,3 0,50 (1/2)27 0,82 1,80 (2,2) 0,30 0,4 0,50 (1/2)
Cebolla cabezona
28 1,31 1,04 (0,8) 0,49 0,4 0,50 1/2)29 1,19 2,70 (2,3) 0,00 0,0 0,00 (0/1)
Frutales30 0,56 2,07 (3,7) 0,22 0,4 0,40 (2/5)31 0,33 1,43 (4,3) 0,40 1,2 0,50 (4/8)
Calabaza 32 1,31 0,36 (0,3) 1,09 0,8 0,33 (2/6)
1 Índice de diversidad (X/Y) es la diversidad de colonias de B. thuringiensis (X) sobre la diversidad de colonias de Bacillus sp. (Y).
tabla 1. Abundancia y diversidad de aislamientos de B. thuringiensis en Boyacá, Colombia.
pOVEDA/MARTÍNEZ

Vol. 2 - No. 1 - 2008
91
secaron a temperatura ambiente; posteriormen-te, se homogenizaron con rodillo y se tamizaron en tamices de 850 y 500 micrones. Las muestras procesadas se almacenaron a una temperatura de 4°C, hasta su respectivo análisis. En un formato individual y para cada muestra, se recopilaron datos como: vereda, finca, propietario, tipo de cultivo, estado de desarrollo, labores culturales y riego.
Aislamiento de bacterias. El aislamiento de Ba-cillus thuringiensis y otras formas esporales de bac-terias se hizo de acuerdo con el método descrito por Travers et al. (1987). En agua destilada estéril se suspendió un gramo de cada muestra de suelo y se pasteurizó a 80°C por media hora, seguida-mente se realizaron diluciones y se sembraron en agar nutritivo e incubaron a 30°C durante 24 h. Para la selección de cepas de Bacillus thuringiensis, se suspendieron 0,5 g de cada muestra de suelo procesada, en matraz de 20 mL, con 10 mL de caldo Luria Bertani (LB) y Acetato de Na 0,25M (Travers et al., 1987) previamente esterilizados. Posteriormente, se llevó este material a agitación continua a 200 rpm, durante 12 h a 29°C. Pasado este tiempo, las muestras se calentaron a 80°C durante 3 min en baño maría. De esta suspen-sión, se realizaron diluciones desde 10-1 hasta 10-4 con agua destilada estéril, las cuales fueron sembradas por duplicado en medio LB (10 g L-1 triptona, 5 g L-1 extracto de levadura, 10 g L-1 NaCl y 15 g L-1 de agar) e incubadas a 30ºC por 48 h. El recuento de bacterias viables esporuladas se realizó después de la pasteurización y el re-cuento de colonias de B. thuringiensis después de llevar a cabo el método de selección por acetato.
Identificación de Bacillus thuringiensis. Se determinaron las colonias características de la bacteria, realizando tinciones simples con cristal violeta (0,5%) y verde de malaquita, detectando por microscopía a 100X células vegetativas, espo-ras o cristales (Porcar y Juárez, 2004). Finalmen-te, las colonias típicas de Bt se repicaron en me-dio LB y se incubaron nuevamente a 30°C, hasta alcanzar esporulación, para verificar la presencia del cuerpo paraesporal y definir su forma.
Conservación de cepas. Las cepas de Bacillus thuringiensis identificadas y caracterizadas se conservaron en tiras de papel filtro, siguiendo la metodología descrita por Porcar y Juárez (2004), en viales ámbar de 20 mL y sellados con agra-fes. El material se almacenó en cajas de cartón para protección contra la luz y permanecen en el Laboratorio de Control Biológico de la UPTC en Tunja, Colombia.
reSUltADoS Y DISCUSIÓn
Se determinó la presencia de cepas nativas de B. thuringiensis, obtenidas en 32 muestras de suelo, recolectadas en 12 hábitats diferentes de los mu-nicipios de Chíquiza, Siachoque y Tuta, situados en la Provincia Centro del departamento de Bo-yacá, municipios seleccionados por su diversidad de agroecosistemas. El aislamiento de las cepas nativas de Bt se hizo en seis muestras de suelo provenientes de zonas protegidas (Bosques Na-turales), pantanos (una muestra) y 25 muestras provenientes de áreas agrícolas (tabla 1). El mayor recuento de bacterias viables antes de la pasteu-rización (~2,80 106 ufc/mL) se encontró en áreas cultivadas con trigo, avena y rábano, mientras que en el cultivo de papa se determinó el menor valor de bacterias viables (0,11 106 ufc/mL).
Los resultados evidencian que las áreas de bosques presentan el contenido más alto de bacterias espo-ruladas (0,48-3,01 105 ufc/mL) con un 1,2 - 15,1%, seguido por los suelos cultivados con maíz (2,08-2,97 105 ufc/mL) con 4,2% y 18,0% (tabla 1), En cuanto a B. thuringiensis, se observaron 0,25-2,90 105 ufc/mL, para bosques, y 0,09-2,98 105 ufc/mL para maíz con un 0,4-9,3% y 0,1-18,0%, del total de bacterias viables, respectivamente. Estos ran-gos son inferiores comparados con los reportados por Chatterjee et al. (2007) en tierras de la India con rangos entre 4,23-6,52 105 ufc/mL.
En zonas protegidas y áreas cultivadas con maíz, papa y frutales, se encontró la mayor abundan-cia de esporas y bacterias de B. thuringiensis, mientras que en zonas cultivadas con trigo no se
ABUNDANCIA y D IVERS IDAD DE Bac i l l us thu r i ng iens i s

REV. COLOMB. CIENC. HORTIC.
92
encontraron cepas productoras del cuerpo para-esporal (tabla 2).
En la tabla 1 se evidencia que las cepas de B. thu-ringiensis predominaron entre las cepas de Baci-llus sp. con índice de diversidad de 1,00 para las muestras 1, 7, 8, 10, 17 y 22 pertenecientes a sue-los de bosques, pantanos, maíz, leguminosas y papa, información que se asemeja a la reportada por Phyllis et al. (1989). Los índices de diversidad de las demás muestras oscilan entre 0,0 y 0,67, datos similares a los índices reportados para suelos de España que oscilan entre 0,067 y 0,11 (Quesada-Moraga et al., 2004). La mayor diver-sidad de colonias del genero Bacillus se encontró en suelos cultivados con maíz nueve colonias (ta-bla 1), seguida por suelos cultivados con frutales ocho colonias y suelos cultivados con avena y áreas de bosques naturales siete colonias; la más alta diversidad de colonias de B. thuringiensis se encontró en áreas de maíz, ocho colonias, segui-
das por cinco colonias en áreas de bosques, fru-tales, cuatro colonias, y pantanos, tres colonias; se obtuvieron pocas colonias de Bt de los suelos de leguminosas, papa, rábano y cebolla cabezo-na (tabla 1). Se obtuvieron 51 cepas de Bt, de las cuales el 78% provenían de ecosistemas agríco-las, destacándose las áreas cultivadas con maíz con 27%, similar a Valicente y Barrero (2003), que aislaron la mayoría de cepas de Bt en zonas productoras de maíz en Brasil. El 22% restante correspondió a hábitats de bosques naturales y pantanos; es importante mencionar que tanto el 50% de las muestras de suelo, pertenecientes a bosques naturales, como el 72% de las muestras, pertenecientes a zonas agrícolas, presentaron al menos una cepa de Bt. Por otro lado, en el ecosis-tema de pantanos el 100% registró presencia de Bt (tabla 2).
La mayor presencia de Bt en ecosistemas agrícolas coincide con lo reportado por Xavier et al. (2007);
OrigenNo. de muestras de suelo No. de colonias
pH del sueloRecolectadas Con Bt (%) Examinadas
Bt(% del total)
Bosques naturales 6 3 (50) 15 8 (16) 4,20
Pantanos 1 1 (100) 3 3 (6) 5,09
Maíz 4 4 (100) 22 14 (27) 5,82
Avena 2 1(50) 8 3 (6) 4,60
Leguminosas 3 1 (33) 6 2 (4) 4,67
Papa 5 4 (80) 11 7(14) 3,94
Pastos 2 2 (100) 9 3 (6) 4,06
Rábano 3 2 (67) 5 2 (4) 4,32
Cebolla cabezona 2 1 (50) 3 1 (2) 4,31
Frutales 2 2 (100) 13 6 (12) 6,04
Calabaza 1 1 (100) 6 2(4) 5,16
Trigo 1 0 (0) 1 0 (0) 4,22
Subtotal 25 18 (72) 84 40 (78)
total 32 22 (69) 102 51 (100)
tabla 2. Distribución de Bacillus thuringiensis aislados de las muestras de suelos boyacenses, colombianos.
pOVEDA/MARTÍNEZ
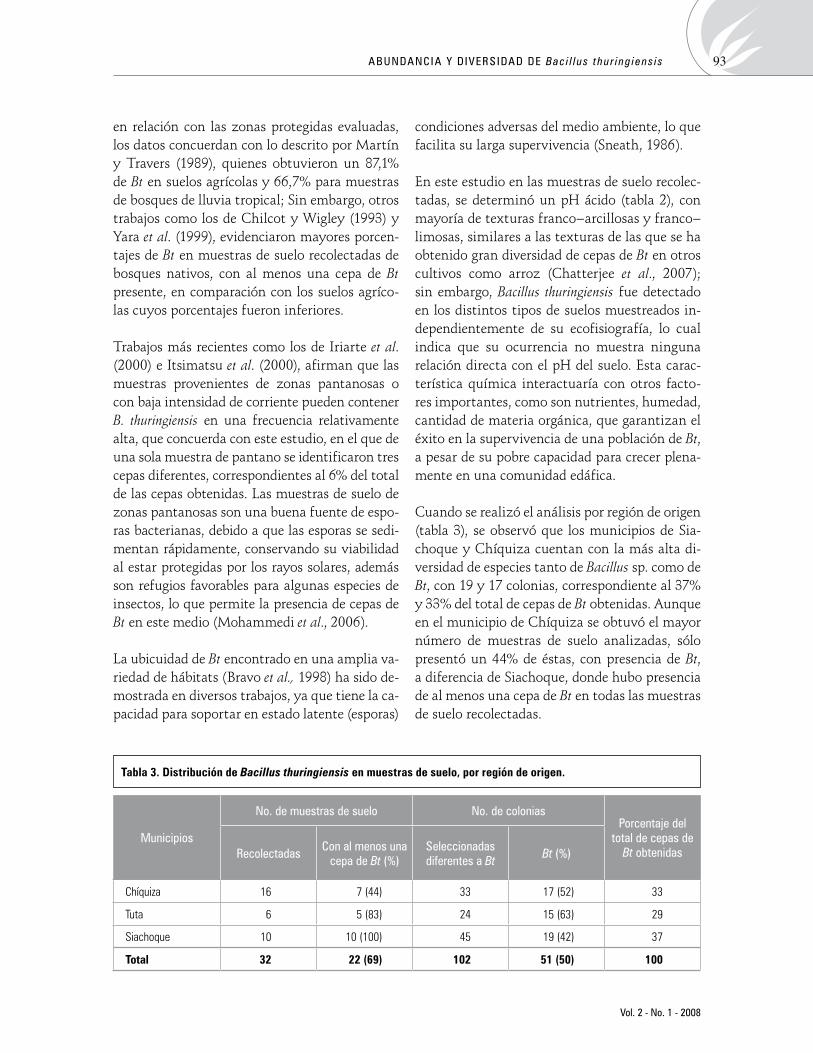
Vol. 2 - No. 1 - 2008
93
en relación con las zonas protegidas evaluadas, los datos concuerdan con lo descrito por Martín y Travers (1989), quienes obtuvieron un 87,1% de Bt en suelos agrícolas y 66,7% para muestras de bosques de lluvia tropical; Sin embargo, otros trabajos como los de Chilcot y Wigley (1993) y Yara et al. (1999), evidenciaron mayores porcen-tajes de Bt en muestras de suelo recolectadas de bosques nativos, con al menos una cepa de Bt presente, en comparación con los suelos agríco-las cuyos porcentajes fueron inferiores.
Trabajos más recientes como los de Iriarte et al. (2000) e Itsimatsu et al. (2000), afirman que las muestras provenientes de zonas pantanosas o con baja intensidad de corriente pueden contener B. thuringiensis en una frecuencia relativamente alta, que concuerda con este estudio, en el que de una sola muestra de pantano se identificaron tres cepas diferentes, correspondientes al 6% del total de las cepas obtenidas. Las muestras de suelo de zonas pantanosas son una buena fuente de espo-ras bacterianas, debido a que las esporas se sedi-mentan rápidamente, conservando su viabilidad al estar protegidas por los rayos solares, además son refugios favorables para algunas especies de insectos, lo que permite la presencia de cepas de Bt en este medio (Mohammedi et al., 2006).
La ubicuidad de Bt encontrado en una amplia va-riedad de hábitats (Bravo et al., 1998) ha sido de-mostrada en diversos trabajos, ya que tiene la ca-pacidad para soportar en estado latente (esporas)
condiciones adversas del medio ambiente, lo que facilita su larga supervivencia (Sneath, 1986).
En este estudio en las muestras de suelo recolec-tadas, se determinó un pH ácido (tabla 2), con mayoría de texturas franco-arcillosas y franco-limosas, similares a las texturas de las que se ha obtenido gran diversidad de cepas de Bt en otros cultivos como arroz (Chatterjee et al., 2007); sin embargo, Bacillus thuringiensis fue detectado en los distintos tipos de suelos muestreados in-dependientemente de su ecofisiografía, lo cual indica que su ocurrencia no muestra ninguna relación directa con el pH del suelo. Esta carac-terística química interactuaría con otros facto-res importantes, como son nutrientes, humedad, cantidad de materia orgánica, que garantizan el éxito en la supervivencia de una población de Bt, a pesar de su pobre capacidad para crecer plena-mente en una comunidad edáfica.
Cuando se realizó el análisis por región de origen (tabla 3), se observó que los municipios de Sia-choque y Chíquiza cuentan con la más alta di-versidad de especies tanto de Bacillus sp. como de Bt, con 19 y 17 colonias, correspondiente al 37% y 33% del total de cepas de Bt obtenidas. Aunque en el municipio de Chíquiza se obtuvó el mayor número de muestras de suelo analizadas, sólo presentó un 44% de éstas, con presencia de Bt, a diferencia de Siachoque, donde hubo presencia de al menos una cepa de Bt en todas las muestras de suelo recolectadas.
Municipios
No. de muestras de suelo No. de coloniasporcentaje del
total de cepas de Bt obtenidasRecolectadas Con al menos una
cepa de Bt (%)Seleccionadas diferentes a Bt Bt (%)
Chíquiza 16 7 (44) 33 17 (52) 33
Tuta 6 5 (83) 24 15 (63) 29
Siachoque 10 10 (100) 45 19 (42) 37
total 32 22 (69) 102 51 (50) 100
tabla 3. Distribución de Bacillus thuringiensis en muestras de suelo, por región de origen.
ABUNDANCIA y D IVERS IDAD DE Bac i l l us thu r i ng iens i s

REV. COLOMB. CIENC. HORTIC.
94
Esta menor presencia puede estar relacionada con la existencia de una agricultura intensiva de mo-nocultivos, con gran uso de productos químicos para mantener su producción y controlar plagas, lo que, probablemente afecta de manera negativa la diversidad microbiológica de sus suelos.
Por otro lado, en el municipio de Tuta se aisló el 63% de las colonias de Bacillus thuringiensis; sin em-bargo, este municipio presenta el menor índice de diversidad comparado con los otros municipios.
La selección de cepas de Bt, por sus características macroscópicas y microscópicas; reveló la presen-cia de una inclusión o cristales en los esporangios (células bacilares con una espora en su interior), siendo estos de forma bipiramidal, triangular, redonda, cuadrada y amorfa (tabla 4). La gran mayoría de los aislamientos con estos tipos de cristales corresponden a muestras de cultivos de cereales obtenidos de zonas agrícolas y otras de áreas protegidas (tabla 1).
Es importante señalar que la forma bipiramidal de los cristales fácilmente observable (figura 1), predomina en suelos de bosque natural y suelos cultivados con maíz y papa, lo cual concuerda con los últimos estudios realizados en varias re-giones de Colombia por Armengol et al. (2007), donde éste es el tipo de cristal más frecuente, con el 60% de abundancia; igualmente, la forma bipiramidal es muy común al ser aislada de co-mida para animales (Meadows et al., 1992). Un alto porcentaje de aislamientos presentaron in-clusiones paraesporales de forma triangular-re-donda no muy frecuentes en la morfología típica de B. thuringiensis en cuanto al cristal se refiere; la gran mayoría de estos aislamientos corresponde a zonas de bosques, pantanos, cultivos de avena, leguminosas, pastos y calabaza. En general, los cristales cuadrados fueron menos frecuentes, sin embargo el tipo cuadrado-redondo se observó en aislamientos obtenidos de suelos cultivados con frutales y calabaza, con 67% y 50% del to-tal, respectivamente (tabla 4).
OrigenNúmero de cepas (% del tipo de cristal) Total de
cepasB A-O A-T C-T C-O T-O T-B-C T-C-O
Bosque natural 3 (38) 1 (13) - 1 (13) - 3 (38) - - 8 (100)
Pantanos 1 (33) - - - - 1 (33) 1 (33) - 3 (100)
Maíz 6 (43) 3 (21) 1 (7) 1 (7) 2 (14) 1 (7) - - 14 (100)
Avena - - - 1 (33) 1 (33) 1 (33) - - 3 (100)
Leguminosas - 1 (50) - - - 1 (50) - - 2 (100)
Papa 3 (43) - 1 (14) 1 (14) - 1 (14) - 1 (14) 7 (100)
Pastos 1 (33) - - - 1 (33) 1 (33) - - 3 (100)
Rábano - - - 1 (50) - - 1 (50) - 2 (100)
Cebolla cabezona - - - 1(100) - - - - 1 (100)
Frutales - 1 (17) - - 4 (67) 1 (17) - - 6 (100)
Calabaza - - - - 1 (50) 1 (50) - - 2 (100)
total 14 (27) 6 (12) 2 (4) 6 (12) 9 (18) 11(22) 2 (4) 1 (2) 51 (100)
tabla 4. Distribución de los tipos morfológicos de cuerpos paraesporales por lugar de origen.
Clasificación Tipos Morfológicos de inclusiones paraesporales.
B, Bipiramidal; A-O Amorfo-Redondo; A-T amorfo-triangular; C-T cuadrado-triangular; C-O cuadrado-redondo; T-O triangular-redondo; T-B-C triangular-bipiramidal y cuadrado o rectangular; T-C-O triangular-cuadrado-redondo.
pOVEDA/MARTÍNEZ

Vol. 2 - No. 1 - 2008
95
En un aislamiento obtenido del municipio de Siachoque, especialmente de cultivos de papa, se observaron inclusiones paraesporales de tipo triangular, cuadrado y redondo (figura 2), carac-terística poco común y escasamente registrados en la literatura (Yara et al., 1999). Encontrar ce-pas con características poco comunes en suelos Boyacenses muestra la gran diversidad de Bt. De esta forma, nuestros resultados contribuyen al conocimiento de la diversidad de Bt en Colom-bia, donde existen muy pocas colecciones carac-terizadas, en comparación con el resto de Améri-ca latina (Bravo et al., 1998).
cuenta que estudios en diferentes ecosistemas de Colombia han reportado porcentajes del 82% (Uribe et al., 2003).
Este estudio es un aporte importante al inicio de un censo de la diversidad de Bt en Boyacá, con el fin de contar con nuevos genes y proteínas ac-tivas para el desarrollo futuro de bioplaguicidas para el control de plagas de importancia agrícola de la región.
Aunque en los ecosistemas agrícolas como maíz, frutales, pastos, papa y calabaza se determinó diversidad, los bosques naturales y pantanos fue-ron una buena fuente de diversidad microbiana, que debe seguir siendo protegida y explorada.
Esta investigación constituyó la base para la creación de una colección de referencia, a partir de la obtención y caracterización de aislamien-tos de Bt de suelos.
Figura 1. visualización microscópica de una cepa nativa de Bacillus thuringiensis proveniente de cultivos de maíz. Cristales de forma bipiramidal (B).
Figura 2. visualización microscópica de una cepa nativa de Bacillus thuringiensis que presenta tres tipos diferentes de cristal paraesporal: triangular (t); Cuadrado, (C), redondo (o). Igualmente se observa presencia de esporas (e).
ConClUSIoneS
Este estudio mostró que el 69% de las mues-tras de suelo analizadas en la Provincia Centro del departamento de Boyacá contiene al menos una cepa de Bt, valor considerable, teniendo en
AGrADeCIMIentoS
Este proyecto fue desarrollado con el apoyo de Colciencias y la DIN (UPTC Tunja), por medio del programa de jóvenes investigadores. Los autores agradecen al Dr. Jairo Cerón y la Dra. María Victo-ria Grosso del Grupo de Biopesticidas del Instituto de Biotecnología de la Universidad Nacional de Colombia, Bogotá, por sus invaluables aportes para la realización de este trabajo.
ABUNDANCIA y D IVERS IDAD DE Bac i l l us thu r i ng iens i s

REV. COLOMB. CIENC. HORTIC.
96
Adams, I.F.; C.l. Liu; S. Macintosh y R.L. Stamen. 1996. Diversity and biological activity of Bacillus thuring-iensis. pp. 360-386. En: Coppin, E.L. (ed.). Crop protection agents from nature. Natural products and analogues. The Royal Society of Chemistry, Cambridge, UK.
Apoloyo, C.I.; L. Drif; J.M. Vassal; H. Debarjac; J.P. Bossy; F. Leclant y R. Frutos. 1995. Isolation of múltiple subspecies of Bacillus thuringiensis from a population of the European sunflower moth, Homoeosoma nebulella. Appl. Environ. Microbiol. 61(2), 4243-4347.
Armengol, G.; M.C. Escobar; M.E. Maldonado y S. Or-duz. 2007. Diversity of Colombian strains of Bacil-lus thuringiensis against dipteran and lepidopteran insects. J. Appl. Microbiol. 1(62), 77-88.
Bravo, A.; S. Sarabia; L. Lopez; E. Ontiveros; C. Abarca; A. Ortiz; M. Ortiz; L. Lina; F. Villalobos; G. Peña; M.A. Núñez; M. Soberon y R. Quintero. 1998. Characterization of cry genes in a Mexican Bacillus thuringiensis strain Collection. Appl. Environ. Mi-crobiol. 64, 4965-4972.
Bravo, A. y J. Cerón (eds.). 2004. Bacillus thuringiensis en el control biológico. Universidad Nacional de Co-lombia, Ed. Buena Semilla, Bogotá.
Cerón, J.; A. Ortiz; R. Quintero; L. Queresa y A. Bravo. 1995. Specific PCR primers directed to indentify Cry I and Cry III genes within a Bacillus thuringi-ensis strain collection. Appl. Environ. Microbiol. 61(1), 3826-3831.
Chatterjee, S.N.; T. Bhattacharya; T.K. Dangar y G. Chandra. 2007. Ecology and diversity of Bacillus thuringiensis in soil environment. Afric. J. Biotech. 13(6), 1587-1591.
Chilcot, C.N. y P.J. Wigley. 1993. Isolation and toxicity of Bacillus thuringiensis from soil and insect habi-tats in New Zeland. J. Invert. Pathol. 61, 244-247.
Deacon, J.W. 1983. Microbial control of plant pests and diseases. American Society for Microbiology, Washington, DC.
Díaz, T.; N. Restrepo; S. Orduz y W. Rojas. 1993. Dis-tribución y aislamiento de Bacillus thuringiensis en Colombia. Rev. Coloma. Entomol. 19(2), 35-40.
Feitelson, J.S.; J. Payne y L. Kim. 1992. Bacillus thuringien-sis, Insects and Beyond. BioTechnol. 4(10), 271-276.
Höfte, H. y H.R. Whiteley. 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53, 242-255.
Hossain, M.A.; S. Ahmed y S. Hoque. 1997. Abundance and distribution of Bacillus thuringiensis in the ag-ricultural soil of Banglasdesh. J. Invert. Pathol. 70, 221-225.
Ibarra, J.E. y M. López. 2000. Bacterias entomopatóge-nas. Cap. 23. pp. 281-295. En: Badii, M.; A.E. Flores y L.J. Galán Wong (eds.). Fundamentos y perspec-tivas de control biológico. Universidad Autónoma de Nuevo León, Monterrey, México.
Ibarra, J.E.; M. Soberón y A. Bravo. 2003. La biotec-nología y el control biológico de insectos. pp. 27-52. En: Bolívar Z., F.G. y L. Herrera E. (eds.). Fronteras de la biología en los inicios del siglo XXI. Módulo 3: Biotecnología agrícola. El Colegio Nacional, México D.F.
Iriarte, J.; M. Porcar.; M. Lecadet y P. Caballero. 2000. Isolation and characterization of Bacillus thuringi-ensis strains from aquatic environments in Spain. Curr. Microbiol. 40, 402-408.
Itsimatsu, T.; E. Mizuki; K. Nishimura; T. Akao; H. Sayito; K. Higuchi y M. Ohba. 2000. Occurrence of Bacillus thuringiensis in fresh waters of Japan. Curr. Microbiol. 40, 217-220.
Kaelin, P.; M. Morel y F. Gadani. 1994. Isolation of Bacil-lus thuringiensis from stored tabaco and Lasioderma serricorne (F.). Appl. Environ. Microbiol. 60(1), 19-25.
Kim, H.S.; D.W. Lee; S.D. Woo; Y.M. Yu y S.K. Kang. 1998. Seasonal distribution and characterization of Bacillus thuringiensis isolated from sericultural en-vironments in Korea. J. Gen. Appl. Microbiol. 44, 133-138.
Lambert, B. y M. Peferoen. 1992. Insecticidal promise of Bacillus thuringiensis. Facts and mysteries about a successful biopesticide. Bio Science 2(42), 112-121.
Lynch, M.J. y P. Baumann. 1985. Immunological com-parisons of the crystal protein from strains of Ba-cillus thuringiensis. J. Invert. Pathol. 46, 47-57.
Martin, P. y R. Travers. 1989. Worldwide abundance and distribution of Bacillus thuringiensis isolates. Appl. Environ. Microbiol. 55(10), 2437-2442.
reFerenCIAS BIBlIoGrÁFICAS
pOVEDA/MARTÍNEZ

Vol. 2 - No. 1 - 2008
97
Martínez, W.; D. Uribe y J. Cerón. 2002. Efecto tóxico de proteínas Cry1 de B. thuringiensis en larvas Te-cia solanivora Povolny. p. 65. En: Memorias Primer Congreso Colombiano de Biotecnología. 26-28 de Junio 2002. Bogotá.
Meadows, M.P.; D.J. Ellis; J. Butt; P. Jarret y H.D. Burg-es. 1992. Distribution, frequency, and diversity of Bacillus thuringiensis an animal feed mill. Appl. En-viron. Microbiol. 58, 1344-1350.
Mohammedi, S.; S.S. Bala; S. Yan; R.D. Tyagi y J.R. Vale-ro. 2006. Molecular screening of Bacillus thuringien-sis strains from wastewater sludge for biopesticide production. Proc. Biochem. 41(4), 829-835.
Pérez, Y.O.; A. Rodríguez y A.M. Cotes. 1997. Método rápido y confiable para evaluar la actividad insecti-cida de cepas nativas de Bacillus thuringiensis contra Tecia solanívora (Polvony) (Lepidóptera: Gelechi-idae). Rev. Entomol. Colomb. 3-4(23), 113-118.
Phyllis, A.; W. Martin y R.S. Travers. 1989. Worldwide abundance and distribution of Bacillus thuringi-ensis isolates. Appl. Environ. Microbiol. 10(55), 2437-2442.
Porcar, M.M. y V. Juarez. 2004. Aislamiento y estableci-miento de una colección de Bacillus thuringiensis. Capítulo 6. pp. 151-176. En: Bravo, A. y J. Cerón. (eds.). Bacillus thuringiensis en el control biológico. Universidad Nacional de Colombia, Ed. Buena Se-milla, Bogotá.
Quesada-Moraga, E.; E. García-Tovar; P. Valverde-Gar-cía y C. Santiago-Álvarez. 2004. Isolation, geo-graphical diversity and insecticidal activity of Bacil-lus thuringiensis from soils in Spain. Microbiol. Res. 1(159), 59-71.
Restrepo, H.N.; D. Gutirerrez.; M.M. Patiño; I. Thiéry; A. Delécluse y S. Orduz. 1997. Clonaje, Expresión y Toxicidad de un gen tóxico de Bacillus thuringien-sis subsp. Rev. Colomb. Entomol. 23, 97-101.
Restrepo, H.N.; M. Patiño; S. Orduz; T. Diaz y W. Rojas. 1998. Caracterización de los genes de las toxinas de Bacillus thuringiensis ssp. Rev. Colomb. Entomol. 24, 7-12.
Rodríguez, M.; M.M. De la Torre y N.E. Urquijo. 1991. Bacillus thuringiensis: características biológicas y perspectivas de producción. Rev. Lat. Amer. Micro-biol. 33, 279-292.
Shelton, A.M.; J.Z. Zhao y R.T. Roush. 2002. Economic, ecological, food safety, and social consequences of the deployment of Bt transgenic plants. Annu. Rev. Entomol. 47, 845-881.
Silva, J.; M. De-Souza; S.J. Díaz y R. Bergmann. 1999. Characterization of Bacillus thuringiensis Subsp. Kurstaki strain S93 effective against the fall army-worm (Spodoptera frugiperda). Canad. J. Microbiol. 45, 464-471.
Sneath, P.H.A. 1986. Endospore-forming gram-positive rods and cocci. pp. 1104-1207. En: Butler, J.P. (ed). Bergey´s manual of systematic entomology. Vol 2. Williams and Wilkins, Baltimore, MD.
Tamez-G., P.; M. Iracheta; A.B. Pereyra; W.L. Galán; F.R. Gómez; G. Támez y P. Rodríguez. 2005. Caracter-ización de cepas mexicanas de Bacillus thuringiensis tóxicas para larvas de lepidópteros y coleópteros. Ciencia UANL 8(4), 447-482.
Travers, R.S.; P.A. Martin y C.F. Reicheldelfer. 1987. Se-lective process for efficient isolation of soil Bacillus spp. Appl. Environ. Microbiol. 53(6), 1263-1266.
Uribe, D.; W. Martínez y J. Cerón. 2003. Distribution and diversity of cry genes in native strains of Bacil-lus thuringiensis obtained from different ecosystems from Colombia. J. Invert. Pathol. 82(2), 119-127.
Valicente F.H. y M.R. Barreto. 2003. Bacillus thuringien-sis survey in Brazil: geographical distribution and insecticidal activity against Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Neotrop. Entomol. 32(4), 639-644.
Vallejo, L.F. y P.S. Orduz. 1996. Producción de un plagui-cida a base de Bacillus thuringiensis subsp. Kurstaki, a nivel de laboratorio. Rev. Colomb. Entomol. 22, 61-67.
Wlodzimierz, D. y L. Elzbieta. 1999. Biodiversity of Ba-cillus thuringiensis strains in the phylloplane and soil of Lower Silesia region (Poland). Acta Micro-biol. Polon. 48(4), 355-361.
Xavier, R.; C.M. Reena y S. Sreeramanan. 2007. Envi-ronmental distribution and diversity of insecticidal proteinas of Bacillus thuringiensis Berliner. Malays. J. Microbiol. 3(2), 1-6.
Yara, E.; J. Cerón y D. Uribe. 1999. Caracterización de aislamientos nativos de Bacillus thuringiensis prove-nientes de bosque natural y zonas de cultivos co-lombianos. Rev. Colomb. Entomol. 25, 185-190.
ABUNDANCIA y D IVERS IDAD DE Bac i l l us thu r i ng iens i s

MILENA ALFONSO1
ALEJANDRO COCA1
WILLIAM RAMÍREZ1
LILIANA HOYOS-CARVAJAL1,2
Plantulas de tomate. Foto: Álvarez-Herrera.
Aproximación de la dinámica poblacional de los microorganismos en diferentes sustratos empleados en el cultivo de rosa (Rosa spp. var. charlotte) en la sabana de bogotá
Overview of the population dynamics of microorganisms in different substrates used in rose (Rosa spp. var. Charlotte) cultivation on the Bogota Plateau
Cultivo de rosa var. Charlotte bajo invernadero en la Sabana de Bogotá.Foto: V.J. Flórez
RESUMEN
La identificación y estudio de la dinámica poblacional de los microorganismos permite reconocer y determinar sus cambios físicos o estructurales en una unidad de tiempo y espacio. En el caso de la rosa, son pocos los estu-dios que detallan esta información. Esta investigación evaluó la dinámica poblacional de los microorganismos (hongos y bacterias) en un cultivo experimental de rosa var. Charlotte ya establecido en la Sabana de Bogotá y dispuesto en cinco sustratos (suelo 100%, cascarilla de arroz 100%, mezcla de cascarilla de arroz y fibra de coco 65:35% (v/v), mezcla de cascarilla de arroz y fibra de coco 35:65% (v/v) y fibra de coco 100%) en un siste-ma con recircularización de nutrientes. Se realizaron dos muestreos con un mes de diferencia y las muestras de los sustratos fueron analizadas en el laboratorio, donde se caracterizaron e identificaron los microorganismos presentes. Se hicieron conteos a las 24 y 48 horas después de siembra en tres medios de cultivo (PDA, AN y D1M). Los resultados de la dinámica poblacional entre los dos muestreos determinaron una mayor actividad de hongos y bacterias en el segundo muestreo, indicando que los microorganismos cambian su actividad y en algunos casos la presencia o ausencia de ciertos géneros teniendo como variable el tiempo, siendo en el caso de los hongos los géneros Mucor y Cladosporium, los que mayor ocurrencia tuvieron en todos los tratamientos.
1 Grupo de Investigación en Fitopatología, Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).
2 Autor para correspondencia. [email protected]
reVisTA cOLOMbiANA de cieNciAs HOrTÍcOLAs - Vol. 2 - No. 1 - pp. 98-109, 2008

Vol. 2 - No. 1 - 2008
99
Palabras clave adicionales: microorganismos, dinámica poblacional, recirculación de nutrientes.
Additional keywords: microorganisms, population dynamics, nutrient recircularization.
Fecha de recepción: 30-04-2008 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
ABStrACt
Information on the distribution and population dynamics of microorganisms facilitates recognition of physical or structural changes that may occur in both time and space. In the case of the rose, a few studies are available that provide such information. In this study, the population dynamics of microorganisms (fungi and bacteria) associated with the established experimental rose variety Charlotte was evaluated on the Bogotá Plateau in five substrates: 100% soil, 100% rice husks, a 65:35% (v/v) mixture of rice husks and coconut fiber, a 35:65% (v/v) mixture of rice husks and coconut fiber, and 100% coconut fiber, all in a system with 100% nutrient recirculation. Samples were taken on two dates with a month between the samplings and the microorganisms were characterized and identified in the laboratory. Samples were plated in three culture media: PDA, AN and D1M and counted after 24 and 48 hours. Greater incidence of both fungi and bacteria from the second sampling indicated that the microorganisms increased activity over time, with the presence or absence of certain genera also varying over time. Among the fungi, the genera Mucor and Cladosporium had the highest incidence over all treatments.
El suelo es un ambiente complejo y dinámico en el cual la actividad biológica está en mayor me-dida dominada por la actividad de los microor-ganismos (Gomes et al., 2003). La diversidad mi-crobiana es fundamental para el funcionamiento del ecosistema, debido a la diversidad de procesos como la descomposición, ciclaje de nutrientes, agregación del suelo, antagonismos y patogenici-dad (Dubey et al., 2006). En el caso de la agricul-tura y campos afines, un estudio del papel de los microorganismos del sistema suelo-planta, y con mayor énfasis en la rizosfera, permite generar un mayor uso sostenible del mismo, encaminado a unos mayores rendimientos de los cultivos, los cuales estén relacionados a un manejo adecuado que favorezca el medioambiente.
El tipo de suelo ejerce un fuerte efecto sobre las co-munidades de la rizosfera, siendo principalmente
sus diferencias en pH, aireación y características físico-químicas las que generan comunidades de microorganismos definidas y espacialmente varia-bles. Asimismo, la disponibilidad de nutrientes minerales, como el nitrógeno, fosforo o hierro y la cantidad de carbono en el suelo pueden afec-tar también el número de microorganismos en la rizosfera y cambiar la composición de la comu-nidad de los mismos (Marschner et al., 2001). Por otra parte, los microorganismos asociados con la raíces de las plantas son importantes en la nutri-ción, promoción del crecimiento e interacciones por patogenicidad (Marschner et al., 2001). En algunos casos, la composición de la comunidad de la rizosfera puede estar influenciada por el ge-notipo de la planta, aunque existen excepciones (Cattelan et al., 1998). Los efectos de las espe-cies de plantas son probablemente debidos a las diferencias en los componentes de las células y
ApROxIMACIóN DE LA D INáMICA pOBLACIONAL DE LOS MICROORGANISMOS

REV. COLOMB. CIENC. HORTIC.
100
exudados de la raíz, los cuales pueden también variar con la zona radicular, edad de la planta, cantidad de nitrógeno e infección por hongos micorrízicos (Marschner et al., 2001).
Existen reportes de investigaciones sobre comu-nidades de microorganismos de la rizosfera in-teractuando en diferentes clases de suelos y con varias especies de plantas. En algunos casos, el efecto del tipo de suelo sobre los microorganis-mos fue mayor que el de las plantas (Buyer et al., 1999; Damastri et al., 1999) y en contraste de acuerdo con estudios realizados por Grayston et al. (1998) y Miethling et al. (2000), las espe-cies de plantas tuvieron un efecto mayor sobre la composición de la comunidad de la rizosfera que el tipo de suelo (Marschner et al., 2001).
El cultivo de la rosa, que en la actualidad ocupa el 36,5% del área total sembrada en Colombia y que forma parte del 48% de la exportación de flor cortada del país (Asocolflores, 2007), es afectado por varias enfermedades causadas por patógenos de suelo, los cuales producen pérdidas y sobre-costos en la producción cada año (Asocolflores, 2003). Los floricultores, ante esta problemática, han optado por emplear como alternativa al sue-lo otros medios de soporte, como cascarilla de arroz quemada, fibra de coco o mezcla de éstas, con el fin de disminuir la incidencia de enferme-dades. Estas opciones en la mayoría de los casos se han empleado sin el sistema de recirculariza-ción de nutrientes (Pizano, 2000). Sin embar-go, esta técnica tiene grandes ventajas, ya que permite que la solución madre sea desinfectada por métodos físicos o químicos (filtración, cale-facción, cloración entre otros) y algunos agentes biológicos (Steijl et al., 1999; Chave et al., 2008).
Actualmente son escasos los reportes de estu-dios que indiquen cómo es el comportamiento microbiológico en los sustratos empleados bajo el sistema de recirculación de nutrientes en culti-vos de rosa en la Sabana de Bogotá. Dado lo an-terior, el Grupo de Investigación de Fitopatología de la Facultad de Agronomía de la Universidad
Nacional de Colombia ha propuesto el siguien-te estudio, con el fin de determinar la dinámica poblacional de los microorganismos en los dife-rentes sustratos empleados de manera comercial en el cultivo de rosa en la Sabana de Bogotá, por medio de aislamientos, caracterizaciones y prue-bas bioquímicas.
MAterIAleS Y MÉtoDoS
El estudio se realizó en el Centro Multisectorial de Mosquera, Regional de Cundinamarca, en un cultivo tipo comercial de cuatro años de rosa variedad Charlotte, injertadas sobre el patrón ‘Natal Briar’, el cual estaba sembrado en materas con capacidad de 8 L y con cinco diferentes sus-tratos (tabla 1).
Recolección, cuantificación, aislamiento y caracterización de los microorganismos
Los muestreos se realizaron el 29 de septiembre y el 27 de octubre de 2008, con el fin de observar y describir los detalles de la dinámica poblacional de cada uno de los sustratos. Para la obtención de muestras, se tomó una submuestra por matera, recogiendo así una cantidad de sustrato conside-rable y representativo del mismo. Posteriormente se homogeneizó el material, conformando una muestra final de 500 g por cada sustrato.
El procesamiento de las muestras se realizó en el Laboratorio de Microbiología de la Facultad de Agronomía de la Universidad Nacional de Co-lombia, sede Bogotá. Los sustratos se secaron a
No. Tipo de sustrato
1 Suelo.
2 Cascarilla de arroz quemada 100%.
3 Cascarilla de arroz quemada más fibra de coco 65:35.
4 Cascarilla de arroz quemada más fibra de coco 35:65.
5 Fibra de coco 100%.
tabla 1. Sustratos usados en el estudio.
ALFONSO/COCA/RAMÍREZ/HOyOS-CARVAjAL

Vol. 2 - No. 1 - 2008
101
temperatura ambiente para eliminar el conteni-do de agua y hacer las disoluciones en seco. Lue-go, mediante métodos físicos (molido, macerado, tamizado y mezclado) cada material fue homo-genizado para garantizar la representatividad de los resultados obtenidos en el análisis. Para rea-lizar el análisis microbiológico, se empleó el mé-todo de las diluciones, ya que permite apreciar de manera cualitativa la actividad de las especies aisladas (Agrios, 2005).
Con la ayuda de una pipeta estéril, se tomaron 100 µL de la dilución correspondiente (10-3 para hongos y 10-5 para bacterias) de cada sustrato y se sirvió en una caja de Petri estéril con un medio de cultivo específico: PDA, agar nutritivo (AN) para hongos y bacterias y (D1M) específico a Agrobacterium tumefaciens. El material sembrado en las cajas se dispuso en una cámara de creci-miento a 22°C y se realizó un conteo a las 24, 48 y 72 h después de incubación, con el fin de observar si existía alguna diferencia significativa en cuanto a la cantidad de hongos y bacterias presentes por tratamiento.
Se llevó un registro fotográfico y se realizó la caracterización macroscópica de los hongos y bacterias, determinando a su vez el porcentaje de frecuencia de acuerdo con los géneros o mor-fotipos identificados, respectivamente. Para la caracterización del género de hongos, se utilizó la metodología de la impronta y la clave taxo-nómica de Barnett y Hunter (1998). En el caso de las bacterias, de acuerdo con las que mayor frecuencia presentaban (1 ó 2 morfotipos por tratamiento), se caracterizaron macroscópica y microscópica, adicionalmente se les realizó la prueba de tinción de Gram.
Análisis estadístico
El análisis estadístico del estudio para la evalua-ción en laboratorio, específicamente la cuanti-ficación según el caso de bacterias y hongos a las 24 y 48 h después de siembra (hds), estuvo basado en un diseño completamente al azar, con
cinco tratamientos (tipo de sustrato) y seis repe-ticiones por medio para un total de 90 cajas por muestreo. Los datos se sometieron a un análisis de varianza y una prueba de rango múltiple de Tukey (P≤0,05), utilizando como software de so-porte, SAS versión 9.1®.
reSUltADoS Y DISCUSIÓn
Cuantificación de los microorganismos
En los dos muestreos realizados se evidenció que la dinámica poblacional en el cultivo de rosa bajo los distintos sustratos es fluctuante, tenien-do como variable el tiempo (figura 1A y figura 1B). Se comprobó en este caso que las bacterias, debido a su cantidad en los conteos, tienen una mayor actividad en todos los sustratos evaluados respecto a la población de hongos. Según Olal-de y Aguilera (1998), las bacterias representan entre el 25% y 30% de la biomasa microbiana del suelo, comportándose como los organismos más numerosos del suelo (entre 106 y 107 bac-terias/g de suelo), mientras que los hongos, dado su mayor tamaño y presentando menor abun-dancia, evidencian la biomasa más significativa. En complemento, Blaine (1992) señala que las bacterias son numerosas por su pequeño tama-ño, pero los hongos son probablemente de igual o mayor importancia en muchos suelos por su biomasa significativa, asociación íntima con los sistemas radiculares y competencia saprofita con grandes detritus físicos y componentes estructu-rales complejos.
En el caso de los hongos, se observó en el segun-do muestreo un cambio evidente y pronunciado en cuanto a su población a las 48 h después de siembra, siendo los tratamientos de cascarilla de arroz 100% y fibra de coco 100% significativa-mente diferentes (P≤0,05) y en mayor cantidad en cuanto a ufc/g de hongos suelo respecto a los demás tratamientos (figura 1A). Este comporta-miento puede estar dado por intervención de fac-tores ambientales o condiciones físico-químicas
ApROxIMACIóN DE LA D INáMICA pOBLACIONAL DE LOS MICROORGANISMOS

REV. COLOMB. CIENC. HORTIC.
102
favorables (pH, porosidad, fertilización, entre otros) de estos sustratos para un desarrollo más acelerado de las colonias de hongos.
Las bacterias presentaron una tendencia similar, pero menos marcada, respecto al crecimiento de los hongos, indicando quizás una mayor estabi-lidad en cuanto a un cambio gradual en el tiem-po de las condiciones del sustrato (figura 1B). Se observa en este caso que en el medio de mezcla de cascarilla de arroz 35% y fibra de coco 65% presentan cierta cantidad mayor de bacterias tanto en el primer como en el segundo muestreo, siendo esto una posible indicación de una mayor actividad bacteriana en este sustrato. El notable crecimiento de número de bacterias en el sustra-to de cascarilla de arroz 100% en el muestreo 2 puede deberse en mayor medida a una condición bastante favorable (ambiente o sustrato) que ge-nera una mayor actividad de estos microorganis-mos en el tratamiento.
Caracterización e identificación de los hongos
Típicamente, los hongos edáficos son más abun-dantes en la proximidad de la superficie que en el subsuelo y lo son, asimismo, en los suelos ligeros
y bien aireados que en los pesados (Wild, 1989). Las características microscópicas permitieron identificar un total de siete géneros de hongos y no se logró identificar un género con caracte-rísticas de micelio septado y hialino, debido a la carencia de una estructura reproductiva o mice-lar propia para su clasificación (figura 3). En el primer muestreo, los géneros Mucor sp. y Clados-porium sp. resultaron ser los hongos con mayor presencia y porcentaje de ocurrencia en todos los sustratos evaluados (tabla 2). Esta alta frecuencia se relaciona posiblemente a una alta capacidad competitiva y colonizadora de ambos géneros. En la parte fitopatológica, las especies del género Mucor se caracterizan por un micelio bastante grueso y resultan ser solamente relevantes en la poscosecha de los alimentos, en la cual pueden generarse pudriciones del material vegetal cose-chado, generando grandes perdidas, teniendo en cuenta que las especies tienden a ser muy resis-tentes a los fungicidas (Smith, 1992).
La mayoría de las especies de Cladosporium sp. son saprófitos y crecen sobre una gran variedad de sustratos. Su caracterización microscópica se baso en la presencia de conidióforos oscuros con varias ramas cerca de la porción superior o media,
Figura 1. Cuantificación de hongos (A) y bacterias (B) a las 24 y 48 h después de siembra (hds) en materas de rosa con recirculación de nutrientes en Mosquera, Colombia. Promedios con letras distintas indican diferencias significativas según la prueba de tukey (P≤0,05).
ALFONSO/COCA/RAMÍREZ/HOyOS-CARVAjAL

Vol. 2 - No. 1 - 2008
103
agrupados o simples, con conidias oscuras de una a dos celulas, variables en forma y tamaño, ovoi-de a cilíndrico e irregular, las cuales concuerdan con lo descrito por Barnett y Hunter (1998).
En cuanto a otras identificaciones, se destaca la presencia de Pythium sp. y Trichoderma sp. En el primer caso, resulta relevante por su efecto pa-togénico y oportunista en el establecimiento de plántulas en semilleros (Agrios, 2005) y en el segundo caso como un potencial colonizador de superficies y medios y en adición antagonista de otros hongos (Ezziyyani et al., 2004). Pythium co-loniza materia orgánica fresca alcanzando altos niveles de inóculo en 24-48 h; sin embargo, es un mal competidor con otros hongos y bacterias. En algunos casos, los suelos forestales han resul-tado ser supresivos a las especies patogénicas de Pythium debido a competencia, en particular con Mucorales, de forma que Pythium está aparente-mente ausente (Smith, 1992).
En menor medida, se encontraron los géneros de Rhizopus sp. y Papulaspora sp., siendo el primero un Mucoral que presenta especies que resultan importantes debido a los efectos negativos (po-dredumbres severas) que pueden generar en la
producción de hortalizas y frutas (Smith, 1992). En el caso del segundo organismo, es poco lo que se discute en la literatura, pero entre sus carac-terísticas microscópicas están esporas asexuales carentes, micelio claro y producción de agrupa-ciones compactas de pequeñas células o bulbilos, los cuales parecieran esclerotizados y sirven para reproducir el hongo (Barnett y Hunter, 1998).
En el segundo muestreo se identificaron cua-tro géneros de hongos y uno sin identificar, el cual coincidió con las características hialino-septadas del hongo sin clasificación del primer muestreo (tabla 3). Prevalecieron nuevamente Cladosporium sp. y Mucor sp., ratificando su ca-pacidad competitiva y presencia característica de los sustratos utilizados en el cultivo de rosa. Por otra parte, se determinó en mayor frecuencia, respecto al primer muestreo, la presencia de Pe-nicillium sp. en los sustratos de cascarilla de arroz más fibra de coco en proporción 65:35 y 35:65 y en fibra de coco 100%.
En caso contrario al primer muestreo, el género Mucor sp. en el suelo (tratamiento 1), indican-do como se nombró anteriormente una posible competencia en el medio PDA. Respecto a la pre-
GéneroTratamiento
Suelo CA 100% CA 65% FC 35% CA 35% y FC 65% FC 100%
Cladosporium sp. - + (f=69,3%) + (f=26,7%) + (f=50%) -
Mucor sp. - + (f=4,0%) + (f=66,7%) + (f=6,2%) + (f=100%)
Papulaspora sp. - + (f=26,7%) - - -
Penicillium sp. - - - +(f=43,8%) -
Pythium sp. + (f=55,6%) - - - -
Rhizopus sp. - - + (f=6,6%) - -
Trichoderma sp. + (f=33,3%) - - - -
Sin definir septado hialino + (f=11,1%) - - - -
f = porcentaje de ocurrencia a las 120 hds.
CA, cascarilla de arroz; FC, fibra de coco.
tabla 2. Presencia (+) y ausencia (-) de especies de hongos caracterizados sustratos de rosa en materas con recirculación de nutrientes para un primer muestreo en Mosquera, Colombia.
ApROxIMACIóN DE LA D INáMICA pOBLACIONAL DE LOS MICROORGANISMOS
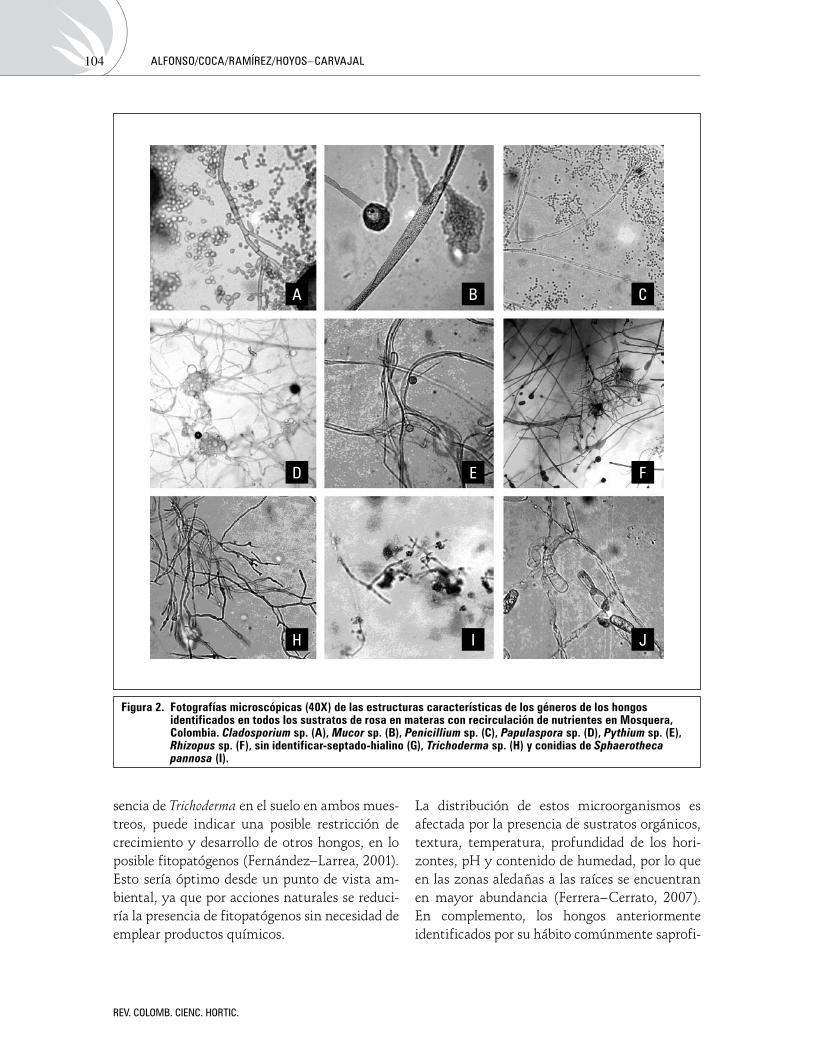
REV. COLOMB. CIENC. HORTIC.
104
sencia de Trichoderma en el suelo en ambos mues-treos, puede indicar una posible restricción de crecimiento y desarrollo de otros hongos, en lo posible fitopatógenos (Fernández-Larrea, 2001). Esto sería óptimo desde un punto de vista am-biental, ya que por acciones naturales se reduci-ría la presencia de fitopatógenos sin necesidad de emplear productos químicos.
La distribución de estos microorganismos es afectada por la presencia de sustratos orgánicos, textura, temperatura, profundidad de los hori-zontes, pH y contenido de humedad, por lo que en las zonas aledañas a las raíces se encuentran en mayor abundancia (Ferrera-Cerrato, 2007). En complemento, los hongos anteriormente identificados por su hábito comúnmente saprofi-
Figura 2. Fotografías microscópicas (40X) de las estructuras características de los géneros de los hongos identificados en todos los sustratos de rosa en materas con recirculación de nutrientes en Mosquera, Colombia. Cladosporium sp. (A), Mucor sp. (B), Penicillium sp. (C), Papulaspora sp. (D), Pythium sp. (e), Rhizopus sp. (F), sin identificar-septado-hialino (G), Trichoderma sp. (H) y conidias de Sphaerotheca pannosa (I).
A B C
D E F
H I j
ALFONSO/COCA/RAMÍREZ/HOyOS-CARVAjAL

Vol. 2 - No. 1 - 2008
105
to en el suelo pueden ser eficaces transformado-res de sustratos edáficos en tejidos microbianos; algunos pueden asimilar entre el 30% y el 50% del carbono presente en la materia orgánica que descomponen, lo que representa una tasa de con-versión muy superior a las bacterias, que se sitúa solamente entre el 5% y el 20% (Wild, 1989).
Caracterización de las bacterias
La cantidad y el tipo de bacterias están determi-nados por su contenido de arcilla, humedad, ai-reación, temperatura, contenido de materia orgá-nica y pH, así como por cultivo, estación del año, profundidad, abundancia de protozoarios y otros alimentos que se nutren de ellas (Ferrera-Cerra-to, 2007). Las primeras bacterias caracterizadas en el muestreo 1 resultaron ser en su mayoría ma-croscópicamente de color amarillo (tabla 4) y mi-croscópicamente cocos (tabla 5). Caso contrario en el muestreo 2, en el cual la población de bacilos fue relativamente mayor y macroscópicamente su aspecto variaba de color amarillo a blancuzco.
Este cambio evidencia que la dinámica en la población de bacterias en el cultivo de rosas va-ria en cuanto a las formas, de coco a bacilos y quizás en visceversa, teniendo como variable de cambio el tiempo. En un contexto mayor, estos resultados van con la idea de que la mayoría de las bacterias del suelo son gram-negativas (Fe-rrera-Cerrato, 2007).
La observación directa de la flora bacteriana edá-fica enseña que en casi todos los suelos predo-minan los cocos pequeños y bacilos cortos. La determinación inicial del comportamiento de bacterias gram+ o gram- durante un tiempo de evaluación (un mes), permite tener una idea sobre la dinámica poblacional de las bacterias y nos permite inferir que tanto el ambiente como el sustrato van a estar sujetos a cambios de orden climático, químico o mecánico, los cuales van a afectar en definitiva la población bacteriana, desde el número hasta la diversidad.
ConClUSIoneS
El estudio de la dinámica poblacional permitió conocer que, teniendo como variable el tiempo, la población tanto de bacterias como de hongos puede fluctuar, en este caso aumentar, debido seguramente a condición favorable para el creci-miento y desarrollo de uno o varios géneros en el espacio.
Un porcentaje de ocurrencia relativamente alto por parte de los géneros Mucor y Cladosporium presente en todos los sustratos permite estable-cer una alta competencia y alta participación de estos en los procesos de formación del suelo. Asi-mismo, una posible interacción entre el género Mucor y Phytium, indicando la capacidad del pri-mero de poder suprimir la actividad del segundo,
GéneroTratamiento
Suelo CA 100% CA 65% FC 35% CA 35% FC 65% FC 100%
Cladosporium sp. + (f=40,0%) + (f=42,9%) + (f=54,4%) + (f=75,5%) + (f=62,8%)
Mucor sp. + (f=30,9%) + (f=50,4%) + (f=14,0%) + (f=9,4%) + (f=31,9%)
Penicillium sp. - - + (f=25,0%) + (f=1,6%) + (f=1,1%)
Trichoderma sp. + (f=1,8%) - - -
Sin definir septado hialino + (f=27,3%) +(f=6,7%) + (f=6,6%) + (f=13,5%) + (f=4,2%)
tabla 3. Presencia (+) y ausencia (-) de especies de hongos caracterizadas en los diferentes tratamientos en el muestreo 2.
f = porcentaje de ocurrencia a las 120 hds. CA, Cascarilla de arroz; FC, fibra de coco.
ApROxIMACIóN DE LA D INáMICA pOBLACIONAL DE LOS MICROORGANISMOS

REV. COLOMB. CIENC. HORTIC.
106
Mue
stre
oTr
atam
ient
oM
orfo
tipo
Cara
cter
izaci
ón m
acro
scóp
ica
Cara
cter
izaci
ón
mic
rosc
ópic
a
Colo
rFo
rma
Bord
esTa
mañ
oSu
perfi
cie
Aspe
cto
Elev
ació
nCo
nsis
tenc
iaFo
rma
Tinc
ión
1
Suel
o1A
Blan
caIrr
egul
arOn
dula
doGr
ande
Rugo
saBr
illant
ePl
ana
Cero
saBa
cilo
Gram
-
CA 1
00%
2AAm
arilla
ver
deCi
rcul
arEn
tero
Gran
deLis
aM
ucos
aPl
ana
Cero
saCo
coGr
am+
2BAm
arilla
pál
ida
Circ
ular
Ente
roGr
ande
Lisa
Brilla
nte
Umbo
nada
Cero
saCo
coGr
am+
CA 6
5% F
C 35
%3A
Amar
illa v
erde
Circ
ular
Ente
roGr
ande
Lisa
Muc
osa
Conv
exa
Cero
saDi
ploc
oGr
am-
CA 3
5% F
C 65
%4A
Amar
illo o
cre
Circ
ular
Ente
roM
edia
naLis
aOp
aca
Conv
exa
Cero
saDi
ploc
oGr
am-
FC 1
00%
5ACa
fé c
laro
Irreg
ular
Ente
roGr
ande
Lisa
Brilla
nte
Plan
aCe
rosa
Coco
Gram
-
5BBl
anco-c
rem
aIrr
egul
arEn
tero
Med
iana
Lisa
Brilla
nte
Conv
exa
Cero
saCo
coGr
am+
2
Suel
o6A
Amar
illo c
laro
Circ
ular
Ente
roM
edia
naLis
aBr
illant
eCo
nvex
aCe
rosa
Baci
loGr
am-
6BBl
anco
Circ
ular
Ente
roGr
ande
Lisa
Seca
, opa
caPl
ana
Cero
saCo
coGr
am-
CA 1
00%
7AAm
arillo
ver
doso
Circ
ular
Ente
roM
edia
naLis
aBr
illant
eCo
nvex
aCe
rosa
Coco
Gram
-
7BBl
anco
Circ
ular
Ente
roM
edia
naLis
aBr
illant
eCo
nvex
aCe
rosa
Baci
loGr
am-
CA 6
5% F
C 35
%8A
Amar
illo v
erdo
soCi
rcul
arEn
tero
Med
iana
Lisa
Brilla
nte
Conv
exa
Cero
saBa
cilo
Gram
-
8BBl
anco
Circ
ular
Ente
roGr
ande
Lisa
Brilla
nte
Conv
exa
Cero
saBa
cilo
Gram
-
CA 3
5% F
C 65
%9A
Amar
illo n
aran
jaCi
rcul
arEn
tero
Med
iana
Lisa
Brilla
nte
Umbo
nada
Cero
saBa
cilo
Gram
-
9BBl
anco
Circ
ular
Ente
roM
edia
naLis
aBr
illant
eCo
nvex
aCe
rosa
Baci
loGr
am-
FC 1
00%
10A
Amar
illo q
uem
ado
Circ
ular
Ente
roM
edia
naLis
aBr
illant
eCo
nvex
aCe
rosa
Baci
loGr
am+
10B
Amar
illo v
erdo
soCi
rcul
arEn
tero
Med
iana
Lisa
Brilla
nte
Conv
exa
Cero
saCo
coGr
am-
10C
Blan
coIrr
egul
arOn
dula
doGr
ande
Rugo
saSe
caUm
bona
daCe
rosa
Dipl
oco
Gram
-
tabl
a 4.
Ca
ract
eriz
ació
n m
acro
scóp
ica
y m
icro
scóp
ica
de lo
s m
orfo
tipos
de
bact
eria
s co
n m
ayor
frec
uenc
ia e
n su
stra
tos
de ro
sa e
n m
ater
as c
on re
circ
ulac
ión
de
nutr
ient
es p
ara
dos
mue
stre
os e
n M
osqu
era,
Col
ombi
a.
ALFONSO/COCA/RAMÍREZ/HOyOS-CARVAjAL

Vol. 2 - No. 1 - 2008
107
y de esta forma la presencia de Phytium, sólo se pudo ver expresada en las cajas de Petri en ausen-cia de colonias de Mucor.
Aunque la identificación de bacterias no se rea-lizó, la caracterización y uso de tinción de gram permitió observar diferencias claras en cuanto a
Muestreo
Número de bacterias presentes según su forma
CocoGram+
CocoGram-
BaciloGram+
BaciloGram-
1 3 3 0 1
2 0 4 1 6
tabla 5. Cantidad de bacterias con mayor frecuencia presentes en cada uno de los muestreos teniendo en cuenta todos los tratamientos.
presencia y ausencia de diferentes formas (cocos o bacilos) bacterianas de acuerdo con el tiempo de muestreo. Esto permite establecer al tiempo como una variable indirecta en la cual se percibe un cambio en cuanto a la diversidad y frecuencia sin excluir que está directamente influido por condiciones climáticas, físico-químicas del sus-trato y manejo del cultivo.
Con fines fitopatológicos, se recomienda detallar con mayor profundidad la especie perteneciente a los géneros de hongos encontrados e identificar las bacterias que en el ensayo se marcaron como ba-cilos y en mayor medida las gram negativa, ya que son las que mayor daño pueden generar al cultivo de rosa. Si es posible, con el ámbito de entender con un nivel más preciso y mejor la diversidad mi-crobial del suelo el estudio se vería muy enriqueci-do con introducción de técnicas moleculares.
AGrADeCIMIentoS
Los autores agradecen a la Universidad Nacional de Colombia y al personal de invernaderos del pro-yecto “Producción más limpia de rosa y clavel con dos técnicas de cultivo sin suelo en la Sabana de Bogotá” y laboratorios, así como a los profesores Víctor Flórez y Diego Nieto por su colaboración en la realización y análisis del presente estudio.
ApROxIMACIóN DE LA D INáMICA pOBLACIONAL DE LOS MICROORGANISMOS

REV. COLOMB. CIENC. HORTIC.
108
Agrios, G. 2005. Plant pathology. 5th ed. Elsevier Aca-demic Press, New York, NY.
Asocolflores. 2003. Asociación Colombiana de Exporta-dores de Flores. Boletín 176, Bogotá.
Asocolflores. 2007. Estadísticas 2007: Floricultura Co-lombiana. En: Asocolflores, http://www.asocol-flores.org/asocolflores/servlet/Download?idExternalFile=236&name=HOJA-DATOS-2008-7.pdf. 4 p.; consulta: mayo de 2008.
Barnett, H.L. y B.B. Hunter. 1998. Illustrated genera of imperfect fungi. 4th ed. American Phytopathologi-cal Society, New York, NY.
Blaine, F. 1992. Soil microbial ecology: applications in agricultural and environmental management. Marcel Dekker, New York, NY.
Bergey, D.H. 1984. Bergey’s manual of systematic bacte-riology. Williams and Wilkins, Baltimore, MD.
Buyer, J.S.; D.P. Roberts y E. Russek-Cohen. 1999. Mi-crobial community structure and function in the spermosphere as affected by soil and seed type. Can. J. Microbiol. 48(11), 955-964.
Cattelan, A.J.; P.G. Hartel y J.J. Fuhrmann. 1998. Bacte-rial composition in the rhizosphere of nodulating and non-nodulating soybean. Soil Sci. Soc. Amer. J. 62(6), 1549-1555.
Chave, M., P. Dabert, R. Brun, J.J. Godon y C. Poncet. 2008. Dynamics of rhizoplane bacterial communi-ties subjected to physicochemical treatments in hydroponic crops. Crop Prot. 27(3), 418-426.
Damastri, C.; L. Chiarini; C. Cantale; A. Bevivno y S. Tabacchioni. 1999. Soil type and maize cultivar af-fect the genetic diversity of maize-associated Burk-holderia cepacia populations. Microb. Ecol. 38(3), 273-284.
Dubey, S.K.; A.K. Tripathi y S.N. Upadhyay. 2006. Ex-ploration of soil bacterial communities for their potential as bioresource. Biores. Technol. 97(17), 2217-2224.
Ezziyyani, M.; C. Pérez; A.S. Ahmed; M.E. Requena y M.E. Candela. Trichoderma harzianum como biofun-gicida para el biocontrol de Phytophthora capsici en plantas de pimiento (Capsicum annuum L.). Ana-les de Biología 26(1), 35-45.
Fernández-Larrea, O. 2001. Microorganismos antagoni-stas para el control fitosanitario. Man. Integr. Plag. 62(2), 96-100.
Ferrera-Cerrato, R. 2007. Microbiología agrícola. Trillas, México D.F.
Gomes, N.; O. Fagbola; R. Costa; N. Rumjanek; A. Bu-chner; L. Mendona-Hagler y K. Smalla. 2003. Dy-namics of fungal communities in bulk and maize rhizosphere soil in the tropics. Appl. Environ. Mi-crobiol. 69(7), 3758-3766.
Grayston, S.J.; S. Wang; C.D. Campbell y A.C. Edwards. 1998. Selective influence of plant species on mi-crobial diversity in the rhizosphere. Soil Biol. Bio-chem. 30(3), 369-378.
Marschner, P.; C. Yang; R. Lieberei y D.E. Crowley. 2001. Soil and plant specific effects on bacterial commu-nity composition in the rhizosphere. Soil Biol. Bio-chem. 33(11), 1437-1445.
Miethling, R.; G. Wieland; H. Backhaus y C.C. Tebbe. 2000. Variation of microbial rhizosphere commu-nities in response to crop species, soil origin, and inoculation with Sinorhizobium meliloti L33. Mi-crob. Ecol. 40(1), 43-56.
Pizano, M. 2000. Clavel. Ediciones HortiTecnia, Bogotá.
Olalde, V. y L.I. Aguilera. 1998. Microorganismos y bio-diversidad. Terra Latinoamericana 16(3), 289-292.
Steijl, H., G.J. Niemann y J.J. Boon. 1999. Changes in chemical composition related to fungal infection and induced resistance in carnation and radish in-vestigated by pyrolysis mass spectrometry. Physi-ol. Mol. Plant Pathol. 55(5), 297-311.
Smith, I. 1992. Manual de enfermedades de las plantas. Ediciones Mundi-Prensa, Madrid.
Wild, A. 1989. Condiciones del suelo y desarrollo de las plan-tas según Russel. Ediciones Mundi-Prensa, Madrid.
reFerenCIAS BIBlIoGrÁFICAS
ALFONSO/COCA/RAMÍREZ/HOyOS-CARVAjAL

AlFreDo JArMA-oroZCo1
Estudios de adaptación y manejo integrado de estevia (Stevia rebaudiana Bert.): nueva alternativa agroindustrial del Caribe colombiano. Una revisión
Environmental suitability and integrated management studies in stevia (Stevia rebaudiana Bert.): a new agro-industrial alternative for the Colombian Caribbean. A review
Plantulas de tomate. Foto: álvarez-Herrera.
Cultivo de Stevia rebaudiana en Cerrito, valle del Cauca, Colombia. Foto: F. Guerrero
1 Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). Grupo Regional de Investigación Parti-cipativa de los Pequeños Productores de la Costa Atlántica. [email protected]
Palabras clave adicionales: radiación, área foliar, nutrición, propagación, patógenos, producción de cultivos, edulcorantes naturales.
reSUMen
En el sector rural del Caribe colombiano, la escasez de alternativas rentables de producción ha generado una reducción en el área cultivada en 380.000 ha, con la pérdida de más de cien mil empleos en la última década. Ante este escenario, la Universidad de Córdoba (Colombia) estudia la adaptación y manejo de nuevas especies agroindustriales que por su rentabilidad minimicen la migración de productores a las ciudades y contribuyan a mejorar la calidad de vida. Hace siete años se iniciaron las primeras investigaciones con estevia en aspectos como ecofisiología, nutrición, fitopatología, entre otros. Los resultados indican que estevia puede cultivarse con éxito en esta región del país. Esta planta contiene glucósidos bajos en calorías, cuyo poder edulcorante en estado puro y cristalino puede llegar a ser entre 200 y 400 veces mayor que el azúcar de caña. Adicionalmente, muchas de las labores que demanda su producción la convierten en un sistema altamente generador de empleo rural, además de los beneficios que su uso trae a la salud. El presente documento resume algunos de los resul-tados más importantes de las investigaciones que ha adelantado la Universidad de Córdoba con esta especie.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No. 1 - pp. 110-121, 2008

REV. COLOMB. CIENC. HORTIC.
110
Additional keywords: radiation, leaf area, nutrition, propagation, pathogens, crop production, natural sweeteners.
Fecha de recepción: 11-12-2007 Aprobado para publicación: 03-06-2008
IntroDUCCIÓn
ABStrACt
Lack of profitable production alternatives in the Colombian Caribbean rural sector has generated a reduction in the cultivated area of roughly 380,000 ha, with the loss of more than 100,000 jobs in the last decade. Faced with this situation, Cordoba University (Colombia) has been studying the adaptation and integrated management of new plant species that might offer profitable alternatives to the agricultural industry and thereby minimize migration to the cities and contribute to improved quality of their life. For the last seven years, the Faculty of Agricultural Sciences has conducted studies with stevia including aspects of plant physiology, nutrition, phytopathology and propagation, among others. The results indicate that this crop can be cultivated successfully in the Colombian Caribbean. This plant contains low caloric glycosides, whose sweeting power in pure and crystal-clear state can reach values of between 200 and 400-times that of sugarcane. In addition to its health benefits for the consumer, the farm work required for stevia production is a strong generator of employment opportunities. The present document covers some of the most important results of research that has been carried out in Cordoba University with this crop.
La estevia es una planta selvática subtropical del alto Paraná, nativa del noroeste de la provincia de Misiones, en el Paraguay, donde era utilizada por los nativos del lugar como edulcorante y cu-rativa. En 1899, el sabio Moisés Santiago Bertoni obtuvo plantas por primera vez de los indígenas nativos, las cuales cultivó y finalmente en 1905 la clasificó (Ministerio Agricultura y Ganadería del Paraguay, 1996).
Los componentes edulcorantes de las hojas de es-tevia son glucósidos de diterpeno sintetizados, al menos en los estados iniciales, usando la misma ruta del ácido giberélico a partir del mevalonato (Kasahara et al., 2002; Hsieh y Goodman, 2005; Guevara et al., 2005), con la diferencia que en es-tevia, el kaureno, precursor de dichas hormonas, se convierte a esteviol en el retículo endoplásmi-co. Numerosos investigadores han reportado la ruta de síntesis de los edulcorantes más impor-
tantes de S. rebaudiana (Totté et al., 2000; Bran-dle et al., 2002; Geuns, 2003; Totté et al., 2003; Brandle, 2005).
Se conoce actualmente que la estevia tiene va-rios componentes edulcorantes, entre los cuales la concentración en la hojas de los cuatro más importantes son: esteviósido (5-10%), rebaudió-sido A (2-4%), rebaudiósido C (1-2%) y dulcósi-do A (0,5-1%) (Rirdc, 2005). El edulcorante es un polvo cristalino blanco que se extrae de las hojas, 100% natural, no tiene calorías y puede llegar a ser hasta 300 veces más dulce que la sacarosa y, comparativamente, es similar al as-partame (Prakash et al., 2008); las hojas pueden utilizarse en su estado natural y sólo son nece-sarias cantidades pequeñas del producto, gracias a su gran poder edulcorante. Puede tener efectos benéficos sobre la salud humana (Tadhani et al., 2007); puede ser utilizado por diabéticos tipo II,
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
111ESTUDIOS DE ADApTACIóN y MANEjO INTEGRADO DE ESTEV IA
ya que no se acumula en el cuerpo y reduce los niveles de glucosa en la sangre (Jeppesen et al., 2000, 2002; Geuns et al., 2007; Brandle y Telmer, 2007), no aporta calorías al ser metabolizado, antihipertensivo (Chan et al., 2000; Lee et al., 2001; Hsieh et al., 2003; Gregersen et al., 2004), antiácido, cardiotónico, no produce caries, al no ser fermentado por las bacterias orales, presen-ta actividad antirotavirus (Das et al., 1992) y se distingue de los edulcorantes artificiales por no tener sabor metálico y no ser cancerígeno (Bran-dle, 2005; Galván et al., 2003; Jarma-Orozco et al., 2005; Jarma-Orozco et al., 2003).
Con la difusión de las informaciones sobre la na-turaleza y los usos potenciales de los principios edulcorantes de esta planta, su cultivo comer-cial adquirió importancia variable en Brasil, Ja-pón, Corea, Taiwán, Tailandia, Indonesia, Laos, Malasia, China, Filipinas, Estados Unidos (Ca-lifornia) y, lógicamente, en el Paraguay (Marín, 2004). El 5% de la producción mundial actual se concentra en el noroeste asiático (Koyama et al., 2003). Japón es el país con el mayor número de fábricas procesadoras y extractoras de esteviósi-do (Schwebel, 2005).
En Colombia se tienen informes sobre la intro-ducción de estevia al Valle del Cauca y Antioquia iniciando la década del noventa, desconociéndo-se su procedencia y año de introducción. En la actualidad hay registros de la siembra de esta especie en Antioquia, Córdoba, Tolima, Huila, Valle del Cauca y Meta.
En el cultivo de estevia se utilizan cerca de 245 jornales permanentes por ha y año, por lo que puede plantearse como alternativa para genera-ción de empleo, además de contribuir a la diver-sificación de los renglones agrícolas de la región y el país (Secretaría de Agricultura de Antioquia, 2000). Sin embargo, por ser un cultivo de recien-te introducción, las técnicas de manejo, los ge-notipos utilizados, los requerimientos hídricos y nutricionales y, en general, el desconocimiento del mismo, han sido limitantes para una produc-ción sostenible y eficiente (Eafit, 2004).
El éxito de cualquier cultivo depende en princi-pio de los conocimientos que sobre éste se ten-gan. Es clave entender la fisiología de la especie en función del ambiente para implementar las prácticas de cultivo que optimicen la produc-ción. Diversos factores como la oferta de la ra-diación, el tipo de suelos, la nutrición, la densi-dad de poblaciones, etc. desempeñan un papel fundamental y determinante, porque a partir de su interacción se sintetizarán gran cantidad de compuestos metabólicos.
En Colombia, la Universidad de Córdoba ha ve-nido desarrollando durante siete años trabajos de investigación con esta especie, que apuntan a definir paquetes tecnológicos de manejo agronó-mico (requerimientos nutricionales, respuesta a la radiación, tipos de suelo, entre otros) y cuyos resultados muestran que es viable la posibilidad de cultivarse en la costa caribeña colombiana. Sin embargo, aún faltan estudios científicos que indiquen cómo crece y se desarrolla la especie ante diversos factores y la manera en que éstos afectan la calidad de los edulcorantes (Jarma-Orozco, 2006). Por esta razón, este documento presenta una relación de los principales avances en la investigación que ha adelantado la Univer-sidad de Córdoba en estevia con miras de deter-minar la factibilidad agronómica del cultivo en la región del Caribe húmedo de Colombia y defi-nir las prioridades de investigación.
rADIACIÓn
Con el propósito de evaluar el efecto de cuatro niveles de radiación incidente (RI) sobre el com-portamiento fisiológico de estevia, se realizó una investigación entre julio de 2002 y abril de 2003 en la Facultad de Ciencias Agrícolas de la Uni-versidad de Córdoba (Montería). Para lograrlo, se planteó un diseño completo al azar con arreglo factorial de 4 x 2, en el que el factor A correspon-dió a los niveles de RI (19, 24, 56 y 100%) y el factor B a los genotipos conocidos en Colombia como ‘Morita 1’ y ‘Morita 2’. Los mayores regis-tros de RI se dieron a las 11:00 horas, observán-

REV. COLOMB. CIENC. HORTIC.
112
dose valores de 3.885, 2.132, 891,5 y 741,3 µmol m-2 s-1 para los niveles 100, 56, 24 y 19%, respec-tivamente (Rengifo et al., 2003). Los principales resultados indicaron que el área foliar por planta de ‘Morita 2’ fue significativamente superior al de ‘Morita 1’ (P≤0,05), independientemente del nivel de radiación (tabla 1) e igualmente los altos niveles de RI (100 y 56%) registraron la mayor acumulación de biomasa de las hojas, siendo su-perior en ‘Morita 2’ durante todo el ciclo de eva-luación (figura 1). Por su parte, la interacción que evaluó la tasa absoluta de crecimiento (TAC) in-dicó que de manera consistente con lo anterior, ‘Morita 2’ expuesta al 100% de radiación inci-dente presentó valores superiores de este índice respecto a ‘Morita 1’.
Los resultados indicarían que estevia se com-portaría como especie heliófila para estas con-diciones de Colombia, aunque sería importante encontrar los puntos de saturación lumínica, al considerar que en la región se presentan radia-ciones muy altas (como las registradas a un nivel de 100%), las cuales pueden causar daños al sis-tema oxidativo de la planta, aparentemente de manera más importante en ‘Morita 1’ respecto a ‘Morita 2’. Algunos autores han indicado en otras especies que los factores de cultivo pueden afectar diferencialmente la expresión del área fo-liar y la asimilación de biomasa en función de la radiación interceptada y de la especie (Lizaso et al., 2003; Mansab et al., 2003; Blanco y Folegatti 2005; Tewolde et al., 2005;). El comportamiento
GenotipoDías después del trasplante
60 150 225‘Morita 1’ 71 a* 239 a 878 a
‘Morita 2’ 186 b 399 b 1700 b
*: Promedios con letras distintas en una misma columna indican diferencia significativa de acuerdo al test protegido de DMS (P≤0,05).
tabla 1. Área foliar total por planta (cm2) de dos genotipos de estevia en el valle del Sinú, Colombia.
Figura 1. Masa seca de hojas por planta de dos genotipos de estevia (M1 = Morita 1; M2 = Morita 2) bajo cuatro niveles de radiación incidente (%) en el valle del Sinú del Caribe colombiano, según Jarma-orozco et al. (2005).
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
113
diferencial entre los genotipos evaluados supone que las divergencias genéticas favorecen a ‘Mo-rita 2’ respecto a su eficiencia en la síntesis de asimilados bajo condiciones de alta radiación, que son propias de esta región del país (Jarma-Orozco et al., 2005, 2006).
nUtrICIÓn
En una investigación desarrollada en 2002, se cuantificó el efecto de la aplicación de dosis com-binadas de los elementos N, P y K sobre variables morfológicas y fisiológicas de ‘Morita 2’ y se de-terminaron los contenidos de estos elementos en la planta en diferentes estados de desarrollo. Se utilizó un diseño completamente aleatorio con tres repeticiones y los análisis se hicieron bajo la metodología de superficie respuesta (Galván et al., 2003).
Los principales hallazgos indican que, para la producción de 2,5 t ha-1 de hoja seca, las plantas cultivadas bajo la oferta ambiental de Córdoba requieren aproximadamente la mitad de N y P y la cuarta parte de K a los 60 días después del trasplante (tabla 2), en comparación con algunos reportes de investigadores de Brasil, que indican que a esta edad el cultivo requiere 325, 47 y 330 kg ha-1 de N, P y K, respectivamente (De Lima y Malavolta, 1997; De Lima et al., 1997 a y 1997 b). Las diferencias podrían obedecer a que a los 60 ddt la edad fisiológica de las plantas fue dife-rencial, adicionalmente a que los requerimientos
nutricionales varían con la oferta ambiental de cada región, incluyendo los tipos de suelos (Peng et al., 2002; Vergara et al., 2003; Yan et al., 2006). En 2002, Marschner indicó que los estudios de nutrición deben estar dirigidos al establecimien-to de la relación entre el suministro de nutrien-tes y el crecimiento de las plantas en términos de producción de materia seca.
Por otra parte, las respuestas observadas en la acumulación de biomasa de las hojas por planta (figura 2) mostraron que, ante la ausencia de P, los niveles altos de K pueden incidir de manera negativa sobre esta biomasa, pero que a niveles medios y altos de P el aumento del peso seco de las hojas es directamente proporcional al incre-mento en los niveles de K, lo que sin duda sugiere la importancia de mantener niveles adecuados de los dos elementos en el plan de nutrición. Asimis-mo, se pueden analizar los resultados en el senti-do que no hay respuesta importante al P cuando la aplicación de K es cero, pero es alta y positiva cuando los niveles de K disponible son altos.
ÁreA FolIAr
Con el propósito de ajustar un modelo estadís-tico para estimar el área foliar (Af) sin utilizar muestreos destructivos Espitia et al. (2006) eva-luaron los genotipos ‘Morita 1’ y ‘Morita 2’ bajo condiciones de campo; en cada uno se tomaron 20 ramas de 25 cm de largo distribuidas aleatoria-mente dentro del dosel. Cada rama se dividió en
ElementoContenido nutricional (mg/planta)
Requerimientos nutricionales (kg ha-1)
población: 100 mil plantas/ha
para 1,0 t hojas secas
para 2,5 t hojas secas
N 149,0 15 70,0 175
P 19,4 2 9,2 23
K 71,2 7 32,0 80
tabla 2. Contenido nutricional de estevia (60 ddt) en Montería (Córdoba, Colombia) y requerimientos nutricionales de n, P y K.
ESTUDIOS DE ADApTACIóN y MANEjO INTEGRADO DE ESTEV IA

REV. COLOMB. CIENC. HORTIC.
114
tres tercios: inferior, medio y superior. De cada tercio se tomaron cinco muestras (repeticiones) de 20 hojas cada una. A cada hoja se le midió el largo y ancho máximo. Con los datos de largo del tercio inferior (LTI), medio (LTM), superior (LTS), el ancho del tercio inferior (ATI), medio (ATM) y superior (ATS) de las hojas y utilizando el método gravimétrico, se realizó un análisis de regresión lineal múltiple para definir el modelo de predicción del Af.
Los resultados demostraron que es posible utili-zar el largo y ancho máximo de la hoja para es-timar el Af. Los modelos resultantes fueron: √Af = 0,288 + 0,093LTI + 0,525ATI + 0,171LTS (R2 = 0,9604) para ‘Morita 1’ y √Af = 0,319 + 0,223ATI + 0,332ATM + 0,067LTS + 0,257ATS (R2 = 0,9213) para ‘Morita 2’.
ProPAGACIÓn
La variabilidad genética de estevia en Colombia es limitada si se considera que en el país sólo se
cultivan los genotipos ‘Morita 1’ y ‘Morita 2’; por ello, es importante evaluar técnicas biotecnoló-gicas para desarrollar protocolos de regeneración de plantas que puedan constituirse en solucio-nes para la generación de variabilidad genética en esta especie. En un trabajo adelantado por Suárez y Salgado (2008), se trataron secciones de entrenudos de plantas de estevia establecidas en condiciones in vitro con diferentes concentra-ciones de bencilaminopurina (BAP) combinadas con varias cantidades de ácido naftalenacético (ANA), con el propósito de evaluar su efecto so-bre el porcentaje de inducción de callo, friabili-dad y formación de órganos. Posteriormente, los callos inducidos fueron multiplicados y se obser-vó el efecto de cuatro combinaciones de ANA y BAP sobre el incremento de masa fresca. Final-mente, los tallos proliferados fueron transferidos sobre medio de regeneración suplementado con diferentes cantidades de BAP o en combinación con ANA con el fin de evaluar la regeneración de brotes. Todos los tratamientos fueron dis-tribuidos utilizando un diseño completamente aleatorio con un mínimo de 15 repeticiones por
Figura 2. Masa seca de hojas en estevia (‘Morita 2’), en función de dosis combinadas de P y K en el valle del Sinú del Caribe Colombiano, según Galván et al. (2003).
Dosis (kg ha-1)
1,79
2,17
2,56
2,94
8743
0
47
93
140
0K
Mas
a se
ca (g
/pla
nta)
P
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
115
tratamiento. Los resultados permitieron obser-var que la presencia de auxina en el medio de cultivo es necesaria y suficiente para inducir la formación de callos en los explantes de estevia, por cuanto al menos el 80% desarrolló callos en presencia de ANA o ANA + BAP, lo cual con-cuerda con los resultados de otros investigadores cuando evaluaron el efecto de las auxinas en pre-sencia de BAP en la formación de tejido calloso en otras especies (Solange et al., 2002; Sánchez et al., 2005). Por otro lado, menos de un 5% de los explantes cultivados en la dosis máxima de BAP (4 mg dm-3) desarrollaron algún tipo de cultivo (figura 3).
En cuanto a la regeneración de brotes vía orga-nogénesis, ocurrió de forma independiente a la presencia de reguladores de crecimiento en el medio, y estos tampoco afectaron el incremento de masa fresca de callo ni tuvieron efecto sobre la regeneración de plantas a partir del callo pro-liferado. Los resultados permiten recomendar un suplemento combinado de 1,0 mg dm-3 ANA + 4,0 mg dm-3 BAP para obtener plantas de estevia mediante organogénesis.
En otros trabajos de propagación adelantados por Caraballo (2006), tendientes a multiplicar la especie por vía sexual a través del rompimiento de latencia de las semillas, se evaluó el efecto de diferentes tipos de luz, temperaturas, hormonas y sus combinaciones. Los resultados indicaron
que ninguno de los factores estudiados presentó efectos positivos significativos sobre la germina-ción de las semillas. El autor concluyó que estos resultados pudieron obedecer al bajo porcentaje de viabilidad que presenta la semilla sexual de esta especie, cuyos porcentajes de germinación varían entre el 10% y 38% y a que la longevi-dad de los aquenios es corta (a los cuatro meses, el porcentaje de germinación se reduce entre un 40% y 70%, después de ocho meses es casi nulo). Adicionalmente a lo anterior, es probable que es-tos resultados obedezcan también a que la plan-ta presenta semillas estériles con apariencia de semillas normales, lo cual reduce el porcentaje de germinación de la misma (Felippe et al., 1971; De Vargas, 1980; Sagakuchi y Tatsuiko, 1982; Jordan, 1984; Caraballo, 2006).
Por otra parte, los estudios de propagación asexual con inducción hormonal por ácido in-dolbutírico (AIB) mostraron que el área foliar, la longitud de raíces y la masa seca de las hojas de ‘Morita 2’ fueron superiores a los de ‘Morita 1’, independientemente de las concentraciones de AIB aplicadas. Sin embargo, ‘Morita 1’ registró mayor longitud de raíces, lo cual probablemente estuvo relacionado con los contenidos endógenos de auxinas de la variedad. Las concentraciones de AIB no afectaron la tasa absoluta de crecimiento (TAC) en ninguno de los dos genotipos (Jarma-Orozco, 2005).
PoBlACIoneS
Para determinar el efecto de diversas poblaciones por hectárea sobre los rendimientos de biomasa del cultivo con base en la oferta ambiental de Córdoba, se han adelantado trabajos con mate-rial vegetativo de estevia entre 2005 y 2007 con 62.500, 125.000 y 250.000 plantas/ha para ‘Mo-rita 1’ y ‘Morita 2’ (Barbosa y Janna, 2007). Los resultados preliminares de esta investigación per-miten inferir que, para las condiciones del Caribe colombiano en Córdoba, el comportamiento de ‘Morita 1’ y ‘Morita 2’ es diferente. Así, se pudo
Figura 3. organogénesis de estevia; callo (izquierda), regeneración de tallos (centro), regeneración de raíces (derecha), según Suárez y Salgado (2008).
A B C
ESTUDIOS DE ADApTACIóN y MANEjO INTEGRADO DE ESTEV IA

REV. COLOMB. CIENC. HORTIC.
116
determinar que ‘Morita 1’ presenta mejores res-puestas a poblaciones altas (250.000 plantas/ha) en tanto que ‘Morita 2’ mantiene los rendimien-tos de hoja seca por hectárea en poblaciones in-termedias y altas (figura 4).
HonGoS FItoPAtÓGenoS
Muestreos realizados por el Laboratorio de Fito-patología de la Universidad de Córdoba, Mon-tería, en los cultivos de estevia, han permitido determinar un complejo de hongos fitopatóge-nos que pueden ser limitantes de la producción si no se da un manejo adecuado de los mismos (Jarma-Orozco et al., 2003). En dicho estudio se aislaron patógenos pertenecientes a los géneros Fusarium, Sclerotium, Alternaria, Botriodiplodia, Co-lletotrichum, Cladosporium, Cercospora, Choanepho-ra, Corinespora y Curvularia; estos tres últimos no presentan reportes de incidencia en estevia en el país (figura 5).
La determinación de estos patógenos muestra el riesgo potencial: eventualmente pueden pre-sentarse de forma epidémica en el cultivo, esti-mulados por las condiciones de alta humedad ambiental, propias de la región, y por grandes extensiones del mismo; por ello, es de importan-cia disponer de una mayor oferta de materiales genéticos para un futuro.
USoS
Adicionalmente a los trabajos anteriores, el gru-po de investigadores de la Universidad de Cór-doba ha estudiado algunos aspectos como el uso de los edulcorantes de estevia en mermeladas y néctares de guayaba (De Paula et al., 2003). Los resultados más importantes de este trabajo indi-caron que es posible obtener productos vegeta-les como néctar y mermelada de guayaba dulce edulcorados con estevia, en los cuales se susti-tuye un porcentaje de sacarosa con adecuadas
Figura 4. rendimiento de hoja seca de dos genotipos de estevia ante tres densidades de población de plantas.
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
117
características fisicoquímicas, microbiológicas y sensoriales. Sin embargo, sus grados Brix no serán suficientes para mantener el néctar estable por un periodo largo de tiempo, debido a que la estabilidad y conservación del mismo depende, en gran proporción, de su contenido de sólidos solubles y estos fueron bajos en los productos ex-perimentales. Por otra parte, los investigadores informaron que con la utilización de estevia se podrán reemplazar los edulcorantes artificiales como sacarina y aspartame, empleados común-mente para endulzar alimentos procesados, lo-grando con ello incursionar en el mercado con un producto que cumple con la misma función de forma natural. Ambos productos, mermelada y néctar de guayaba dulce, fueron satisfactoria-mente aceptados por los consumidores.
ConClUSIoneS
Córdoba ofrece condiciones ambientales que le permiten potencializarse como un gran produc-tor de estevia en Colombia. Factores como la oferta de luz y suelos aptos pueden convertirse en puntos estratégicos a favor de su explotación. Sin embargo, la escasa oferta de variabilidad genéti-ca, las altas humedades relativas en determinadas épocas del año y su efecto sobre la aparición de enfermedades pueden ser limitantes serias.
Las investigaciones adelantadas por la Universi-dad de Córdoba, aunque no todas relacionadas en este documento, pueden considerarse como un primer paso en la identificación de tecnolo-gías competitivas, que hagan de la agroindustria de la estevia un negocio rentable y a su vez gene-re bienestar social en la región.
Por último, es importante recordar que el éxito de cualquier cultivo depende en gran parte de la oferta ambiental, en la que se establezcan el ge-notipo y las tecnologías para su producción. Una combinación adecuada de estos factores se logra cuando se tienen conocimientos objetivos de la especie, los cuales deben estar basados en resul-tados de investigaciones científicas que poste-riormente puedan escalarse en el sistema de pro-ducción. Aunque aún falta mucho por investigar en el manejo de esta nueva especie en el país, los avances en este proceso de la Universidad de Córdoba pueden ser una motivación para animar a otras entidades que también realizan investiga-ciones en el sector agroindustrial de Colombia a reunir esfuerzos y recursos para trabajar en equi-po en pro de una producción e industrialización sostenible de la estevia en el país.
Figura 5. Algunas estructuras de géneros de hongos fitopatógenos aislados en el laboratorio de Fitopatología de la Universidad de Córdoba a partir de hojas y tallos de estevia. a) Fusarium, b) Choanesphora, c) Curvularia y d) Corynespora, según espitia et al. (2008).
a b
c d
ESTUDIOS DE ADApTACIóN y MANEjO INTEGRADO DE ESTEV IA

REV. COLOMB. CIENC. HORTIC.
118
Barbosa, C. y J. Janna. 2007. Comportamiento del cre-cimiento de dos clones de Stevia rebaudiana Bert. bajo tres arreglos poblacionales en las condiciones ambientales del Sinú Medio. Trabajo de grado. Fac-ultad de Ciencias Agrícolas, Universidad de Cór-doba, Montería, Colombia.
Blanco, F. y M. Folegatti. 2005. Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Sci. Agric. 62, 305-309.
Brandle, J. 2005. Stevia, nature’s natural low calorie sweetener. En: http://www.certifiedorganicfood stevia FAQ; consulta: junio de 2006.
Brandle, J.; A. Richman; A. Swanson y B. Chapman. 2002. Leaf ESTs from Stevia rebaudiana: a resource for gene discovery in diterpene synthesis. Plant Mol. Biol. 50, 613-622.
Brandle, J.E. y P.G. Telmer. 2007. Steviol glycoside bio-synthesis. Phytochem. 68, 1855-1863.
Caraballo, U. 2006. Manejo de propágalos de Stevia rebaudiana (Bertoni) para optimizar su estableci-miento bajo condiciones del Caribe Colombiano. Tesis de maestría. Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería, Colombia.
Chan, P.; B. Tomlinson; Y. Chen; J. Liu; M. Hsieh y J. Cheng. 2000. A double blind placebo-controlled study of the effectiveness and tolerability of oral stevioside in human hypertension. Brit. J. Clin. Pharmacol. 50, 215-220.
Das, S.; A. Das; R. Murphy; I. Punwani; M. Nasution y A. Kinghorn. 1992. Evaluation of the cariogenic potential of the intense natural sweeteners stevio-side and rebaudioside A. Caries Res. 26, 363-366.
De Lima, O. y E. Malavolta. 1997. Sintomas de desor-dens nutricionais em estévia Stevia Rebaudiana (Bert.) Bertoni. Sci. Agric. 54(1-2), 53-61.
De Lima, O.; E. Malavolta; J. De Sena y J. Carneiro. 1997a. Absorção e acumulação de nutrientes em estévia Stevia rebaudiana (Bert.) Bertoni: I. Macro-nutrientes. Sci. Agric. 54(1-2), 23-30.
De Lima, O.; E. Malavolta; J. De Sena y J. Carneiro 1997b. Absorção e acumulação de nutrientes em Estévia Stevia rebaudiana (Bert.) Bertoni: II. Micro-nutrientes. Sci. Agric. 54(1-2), 14-22.
De Paula, C.; A. Carmona y G. Lombana. 2003. Determi-nación de las condiciones de utilización del edulcor-ante natural Stevia rebaudiana Bert., en el proceso de elaboración de derivados vegetales (mermelada y néctar de guayaba dulce). Trabajo de grado. Es-cuela de Ingeniería de Alimentos, Universidad de Córdoba, Montería, Colombia.
De Vargas, R. 1980. Informe sobre viaje al Japón para observar la producción, comercialización e indus-trialización de la planta Stevia rebaudiana Bertoni. Asunción, Paraguay.
Eafit, Escuela de Administración y Finanzas. 2004. In-teligencia de mercados internacionales de S. rebau-diana. Departamento de Negocios Internacionales, Medellín, Colombia.
Espitia, M.; R. Montoya y A. Jarma. 2008. Stevia en el Caribe Colombiano. Facultad de Ciencias Agríco-las, Universidad de Córdoba, Montería, Colombia.
Espitia, M.; R. Montoya; J. Robles; C. Barbosa y C. Ver-gara, C. 2006. Modelo estadístico para estimación del área foliar en Stevia rebaudiana Bertoni en el Sinú Medio. Temas Agrarios 11(2), 45-51.
Felippe, G.; N. Lucas; L. Behar y M. Oliveira. 1971. Ob-servaçoes a respeito da germinaçao de Stevia rebau-diana Bert. Hoehnea 1, 81- 91.
Galván, L.; J. Guzmán; A. Jarma y E. Combatt. 2003. Determinación de los requerimientos nutricionales de Stevia rebaudiana, en el Medio Sinú. En: Memo-rias XXXIII Congreso Anual de Comalfi. Montería, Colombia.
Geuns, J. 2003. Stevioside. Phytochem. 64, 913-921.
Geuns, J.; J. Buyse; A. Vankeirsbilck y E. Temme. 2007. Metabolism of stevioside by healthy volunteers. Exp. Biol. Med. 232, 164-173.
Gregersen, S.; P. Jeppesen; J. Holst y K. Hermansen. 2004. Antihyperglycemic effects of stevioside in type 2 diabetic subjects. Metab. Clin. Exp. 53, 73-76.
Guevara-García, A.; C. San Román; A. Arroyo; M. Cor-tes; M. De la Luz Gutiérrez-Nava y P. León. 2005. Characterization of the arabidopsis clb6 mutant illustrates the importance of posttranscriptional regulation of the methyl-D-erythritol 4-phosphate pathway. Plant Cell 17(2), 628-643.
reFerenCIAS BIBlIoGrÁFICAS
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
119
Hsieh, M. y H. Goodman. 2005. The arabidopsis IspH homolog is involved in the plastid nonmevalonate pathway of isoprenoid biosynthesis. Plant Physiol. 138(2), 641-653.
Hsieh, M.; P. Chan; Y. Sue; J. Liu; T. Liang; T. Huang; B. Tomlinson; M. Chow; P. Kao y Y. Chen. 2003. Ef-ficacy and tolerability of oral stevioside in patients with mild essential hypertensiona two-year, ran-domized, placebo-controlled study. Clin. Ther. 25, 2797-2808.
Jarma-Orozco, A. 2005. Cinética del crecimiento. Semi-nario doctoral. Escuela de Posgrados, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá
Jarma-Orozco, A. 2006. Adaptación de dos clones de Stevia rebaudiana a tres ambientes del Caribe co-lombiano. Seminario doctoral. Escuela de Posgra-dos, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Jarma-Orozco, A.; R. Montoya y H. Aramendiz. 2003. Stevia rebaudiana, alternativa de sustitución de cul-tivos ilícitos en el Caribe colombiano. pp. 64-65. En: Memorias XXXIII Congreso de Comalfi. Mon-tería, Colombia.
Jarma-Orozco, A.; T. Rengifo y H. Aramendiz. 2005. Aspectos fisiológicos de estevia (Stevia rebaudiana Bertoni) en el Caribe colombiano. I. Efecto de la radiación incidente sobre el área foliar y la distribu-ción de biomasa. Agron. Colomb. 23(2), 207-216.
Jarma-Orozco, A.; T. Rengifo y H. Aramendiz. 2006. Aspectos fisiológicos de estevia (Stevia rebaudiana Bertoni) en el Caribe colombiano. II. Efecto de la radiación incidente sobre los índices de crecimien-to. Agron. Colomb. 24(1), 38-47.
Jeppesen, P., S. Gregersen., K. Alstrupp y K. Herman-sen. 2002. Stevioside induces antihyperglycaemic, insulinotropic and glucagonostatic effects in vivo: studies in the diabetic goto-Kakizaki (GK) rats. Phytomed. 9, 9-14.
Jeppesen, P., S. Gregersen; C. Poulsen y K. Hermansen. 2000. Stevioside acts directly on pancreatic a cells to secrete insulin: actions independent of cyclic adenosine monophosphate and adenosine triphos-phate-sensitive K+ channel activity. Metabolism 49, 208-214.
Jordán, M., F. 1984. Publicación miscelánea No. 11. El Ka´a He´e, Stevia rebaudiana (Bertoni). Análisis bibliográfico y anotaciones hortícolas. Ministerio de Agricultura y Ganadería, Asunción, Paraguay.
Kasahara, H.; A. Hanada; T. Kuzuyama; M. Takagi; Y. Kamiya y S. Yamaguchi. 2002. Contribution of the mevalonate and methylerythritol phosphate path-ways to the biosynthesis of gibberellins in arabi-dopsis. J. Biol. Chem. 277(47), 45188-45194.
Koyama, E.; N. Sakai; Y. Ohri; K., Kitazawa; O. Izawa; K. Kakegawa; A. Fujino y M. Ui. 2003. Absorption and metabolism of glycosidic sweeteners of stevia mixture and their aglycone, steviol, in rats and hu-mans. Food Chem. Toxicol. 41, 875-883.
Lee, C.; K. Wong; J. Liu; Y. Chen; J. Chen y P. Chan. 2001. Inhibitory effect of stevioside on calcium influx to produce antihypertension. Planta Medica 67, 796-799.
Lizaso, J.; W. Batchelor y M. Westgate. 2003. A leaf area model to simulate cultivar-specific expansion and se-nescence of maize leaves. Field Crops Res. 80, 1-17.
Mansab A.; D.L. Jeffers y P.R. Henderlong. 2003. Inter-relationship between leaf area, light interception and growth rate in a soybean-wheat system. Asian J. Plant Sci. 2(8), 605-612.
Marín, W. 2004. Sondeo de mercado de la Estevia. Insti-tuto de Investigación de Recursos Biológicos Alex-ander von Humboldt, Bogotá.
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, New York, NY.
Ministerio de Agricultura y Ganadería del Paraguay. 1996. Producción de Ka´a He´e. 3a. ed., Asunción.
Peng, S.; J. Huang; X. Zhong; J. Yang; G. Wang y Y. Zou. 2002. Research strategy in improving fertilizer ni-trogen use efficiency of irrigated rice in China. Sci. Agr. Sinica 35, 1095-1103.
Prakash, I.; G.E. DuBois; J.F. Clos; K.L. Wilkens y L.E. Fos-dick. 2008. Development of rebiana, a natural, non-caloric sweetener. Food Chem. Toxicol. 46, S75-S82
Rengifo, T.; A. Jarma y J. Clavijo. 2003. Crecimiento de Stevia rebaudiana bajo cuatro niveles de radiación incidente. p. 66. En: Memorias XXXIII Congreso de Comalfi. Montería, Colombia.
RIRDC, Rural Industries Research and Development Corporation. 2005. A new rural industry -Stevia- to replace imported chemical sweeteners. Publica-tion No. W02/022. En: http://www.rirdc.gov.au; consulta: agosto de 2006.
Sagakuchi, M. y K. Tatsuiko. 1982. As pesquisas japone-sas com Stevia rabaudiana (Bert.) Bertoni e o este-viosideo. Cienc. Cult. 34(2), 235-248.
ESTUDIOS DE ADApTACIóN y MANEjO INTEGRADO DE ESTEV IA

REV. COLOMB. CIENC. HORTIC.
120
Sánchez, D.; K. Huamaní; E. Pascual; H. Sánchez y R. Es-trada. 2005. Efecto de las auxinas y citoquininas en la callogénesis sobre explantes nodales e internodales de Tropaeolum tuberosum (r. and p.) Mashua. p. 155. En: Memorias XIV Reunión Científica ICBAR, Lima.
Schwebel, R. 2005. Stevia, el edulcorante natural sudamer-icano con cero calorías. En: http://www.geocities.com/schwebel_rene/; consulta: junio de 2006.
Secretaría de Agricultura de Antioquia. 2000. Informe preliminar sobre adaptación de la especie Stevia rebaudiana en la región tropical. Gobernación de Antioquia, Medellín, Colombia.
Solange, E., J. Pereira; A. Ramalho; N. Arbex; M. Cardo-so y O. Alves. 2002. Indução de calos em erva-de-touro (Tridax procumbens L.) utilizando diferentes reguladores de crescimento e tipos de explantes. Cienc. Agrot. 26(2), 301-308.
Suárez, I. y J. Salgado. 2008. Propagación in vitro de Ste-via rebaudiana Bert. (Asteraceae-Eupatorieae) a tra-vés de organogénesis. Temas Agr. 13(1), 40-48.
Tadhani, M.; V. Patel y S. Rema. 2007. In vitro antioxi-dant activities of Stevia rebaudiana leaves and cal-lus. J. Food Compos. Anal. 20, 323-329.
Tewolde H.; K. Sistani; D. Rowe; A. Adeli y T. Tsegaye. 2005. Estimating cotton leaf area index nonde-structively with a light sensor. Agron. J. 97, 1158-1163.
Totté, N.; L. Charon; M. Rohmer; F. Compernolle; I. Baboeuf y J. Geuns. 2000. Biosynthesis of the di-terpenoid steviol, an entkaurene derivative from Stevia rebaudiana Bertoni, via the methylerythritol phosphate pathway. Tetrahedron Lett. 41, 6407-6410.
Totté, N.; W. Van den Ende; E. Van Damme; F. Comper-nolle; I. Baboeuf y J. Geuns. 2003. Cloning and het-erologous expression of early genes in gibberellin and steviol biosynthesis via the methylerythritol phosphate pathway in Stevia rebaudiana Bertoni. Can. J. Bot. 81(5), 517-522.
Vergara, C.; A. Jarma; J. Polo y L. Pastrana. 2003. Cre-cimiento y desarrollo de dos variedades de Stevia rebaudiana en tres tipos de suelo del Medio Sinú. p. 68. En: Memorias XXXIII Congreso Anual de Comalfi. Montería, Colombia.
Yan, X.; P. Wu; H. Ling; G. Xu; F. Xu y Q. Zhang. 2006. Plant nutriomics in China: An overview. Ann. Bot. 98, 473-482.
jARMA-OROZCO

Vol. 2 - No. 1 - 2008
121
pOLÍTICA EDITORIAL
La Revista Colombiana de Ciencias Hortícolas es el órgano de divulgación de la Sociedad Colombiana de Ciencias Hortícolas (SCCH – www.soccolhort.com), adscrita a la ISHS (Internacional Society for Horticultural Science – www.ishs.org), con frecuencia semestral en el territorio nacional. Constituye una publicación abierta a la dis-cusión y difusión de trabajos técnico-científicos en el área de las ciencias agrícolas, con énfasis en horticultura (frutales, hortalizas, ornamentales, hierbas aromáticas, viveros) y disciplinas afines, propuestos por autores nacionales e internacionales. Busca divulgar trabajos inéditos desarrollados por investigadores de diversas uni-versidades y centros de investigación del país y difundir y someter a discusión los avances científicos que se producen, con el fin de contribuir a la consolidación de una comunidad académica congregada en torno a las disciplinas afines a la horticultura.
Los artículos deben presentarse según las normas establecidas en las instrucciones para los autores. El Comité Editorial se reserva el derecho de aceptar o no las contribuciones que se hagan a la revista. Los manuscritos se enviarán a dos pares evaluadores expertos en el área específica del conocimiento, quienes decidirán si el artí-culo tiene el nivel científico necesario para publicarse. Si se llega a presentar alguna diferencia de conceptos en cuanto a la aprobación para publicación, el artículo será enviado a un tercer par, quien entregará la decisión final. Si el artículo se acepta para publicación, el autor debe realizar las correcciones sugeridas en el tiempo indicado por el editor.
La Revista Colombiana de Ciencias Hortícolas autoriza la reproducción y citación del contenido de la publi-cación, siempre y cuando se registre de forma explícita el nombre de la revista, nombre del autor (es), año, volumen, número y página del artículo consultado. Los artículos publicados en la Revista serán enviados en medio magnético a las diferentes bases de datos y sistemas de indexación (Publindex), para la divulgación de su contenido y se podrán consultar mediante Internet.
Las ideas y afirmaciones emitidas por los autores son responsabilidad de estos y no expresan necesariamente las opiniones y políticas del Comité Editorial de la SCCH, ni de las instituciones a las cuales están vinculados quienes las proponen.
INSTRUCCIONES pARA LOS AUTORES
Los artículos puestos a consideración del Comité Editorial de la Revista Colombiana de Ciencias Hortícolas deben ser inéditos; en consecuencia, aquellos manuscritos que hayan sido publicados en otras revistas o publicacio-nes técnico-científicas no serán aceptados.
La Revista Colombiana de Ciencias Hortícolas acepta artículos originales de los siguientes tipos, en concordancia con lo requerido por el programa Publindex de Colciencias para la categoría B:
• Artículo de investigación científica y tecnológica: documento que presenta de manera detallada los resultados ori-ginales de proyectos de investigación. La estructura generalmente utilizada tiene cuatro partes esenciales: introducción, metodología (materiales y métodos), resultados y discusión, y conclusiones.
• Artículo de reflexión: documento que presenta resultados de investigación desde una perspectiva analítica, interpretativa y crítica del autor, sobre un tema específico y recurriendo a fuentes originales.

REV. COLOMB. CIENC. HORTIC.
122
• Artículo de revisión: documento resultado de una investigación en la que se analizan, sistematizan e integran los resultados de investigaciones, publicados o no, sobre un campo en ciencia o tecnología, con el fin de dar cuenta de los avances y tendencia de desarrollo; se caracteriza por presentar un soporte bibliográfico cuida-doso no menor a 50 referencias.
Extensión y remisión del artículo y de la ilustraciónLa extensión del artículo de investigación no debe exceder las 5.200 palabras (incluido literatura y las tablas), excepto el artículo de revisión, que puede contener hasta 6.500 palabras; tamaño carta, escritas a doble espa-ciado, letra o fuente Times New Roman con tamaño de 12 puntos, márgenes de 3 cm en la parte superior, 2 cm en la inferior y 2,5 cm en las márgenes laterales derecha e izquierda. Las tablas y figuras (es decir, los grá-ficos, dibujos, esquemas, diagramas de flujo, fotos y mapas) deben presentarse en hojas independientes y con numeración consecutiva (Tabla 1 ... Tabla n. Figura 1 ... Figura n; etc.). Los textos y tablas deben presentarse en el procesador de palabra MS-Word®; las tablas y los diagramas de frecuencia (barras y torta) originales deben suministrarse en el archivo del manuscrito y también en su original de MS-Excel®; otras figuras, como fotografías sobre papel y dibujos, se pueden enviar en originales o escanearlas y remitirlas en el formato digital de compresión JPG (o JPEG), preferiblemente con una resolución de 600 x 600 dpi (mínimo 300 dpi). Como norma general, las tablas y figuras sólo se aceptan en blanco y negro. El artículo con sus archivos en Word, Excel y JPG debe ser enviado al correo electrónico: [email protected], al editor profesor Dr. Fánor Casierra, [email protected] o co-editor profesor Dr. Gerhard Fischer, [email protected].
Título y autoresEl título es en minúscula y no debe exceder las 15 palabras. Es obligatoria su respectiva traducción al idioma inglés. Cuando incluya nombres científicos de vegetales o animales, estos se deben escribir con letra cursiva (itálica) y en minúsculas, y sólo con mayúsculas la primera letra del género y el clasificador. Debajo del título en inglés se escribe el nombre(s) y apellido(s) de los autores, sin sus respectivos títulos académicos ni cargos laborales, en una línea horizontal y de acuerdo con su contribución en la investigación o preparación del artí-culo. En la parte inferior de la primera página, es decir, en aquélla que contiene el título del artículo, el nombre y la ciudad de ubicación de la entidad a la cual prestan sus servicios o del patrocinador para la realización del trabajo y el autor para correspondencia con su correo electrónico (e-mail).
Resumen, abstract y palabras clave adicionalesEl resumen debe escribirse en español, portugués o inglés, correspondiente al idioma del artículo. Los artí-culos en español o portugués deben contener un abstract en inglés, un artículo en inglés y un resumen en español. Ambos textos deben describir en forma breve el problema, los métodos utilizados, su justificación y los resultados obtenidos más relevantes, y no deben exceder de 250 palabras escritas en un único párrafo. Es obligatorio acompañar el resumen con máximo seis palabras clave, que no hayan sido usadas en el título, y de su traducción a inglés (additional key words), las que tampoco deben figurar en el título en inglés.
IntroducciónTexto que debe contener la situación actual del problema, su definición y la revisión de los trabajos previos relacionados con él; además, los objetivos y la justificación de la investigación. Es obligatorio acompañar los nombres vulgares con el nombre(s) científico(s) y la abreviatura(s) del clasificador en la primera mención en el artículo.
Materiales y métodosEn este apartado se deben describir de forma clara, concisa y secuencial, los materiales (vegetales, animales, implementos agrícolas o de laboratorio) utilizados en desarrollo del trabajo, además de los procedimientos o protocolos seguidos y el diseño escogido para el tratamiento estadístico de los datos.
Resultados y discusiónLos resultados deben presentarse de manera lógica, objetiva y secuencial mediante textos, tablas y figuras; estos dos últimos apoyos deben ser fáciles de leer y deben poderse interpretar de manera autónoma, aunque deben citarse siempre en el texto. Las gráficas serán bidimensionales y a una sola tinta, con porcentajes de negro para las variaciones de las columnas; las líneas de las curvas deben ser de color negro, punteadas o con-tinuas (- - - - ó --------), usando convenciones como: l, , s, ∆, etc.

Vol. 2 - No. 1 - 2008
123
Las tablas se deben elaborar con pocas columnas y renglones. La discusión de resultados debe ser completa y exhaustiva, contrastando los resultados obtenidos con la literatura más actual sobre el tema. En esta sección se relacionan los hallazgos más concluyentes de la investigación.
ConclusionesEn este apartado se relacionan los hallazgos más concluyentes de la investigación, es decir, aquellos que cons-tituyan un aporte significativo para el avance del campo temático explorado, además de un direccionamiento sobre futuras investigaciones.
AgradecimientosSi se considera necesario, se agradecen aquellas contribuciones importantes en la concepción, financiación o realización de la investigación: especialistas, firmas comerciales, entidades oficiales o privadas, asociaciones de profesionales y operarios.
Literatura citadaPara las citas bibliográficas que sustentan las afirmaciones en el texto, se utilizará el sistema [autor(es), año] de forma uniforme; cuando la publicación citada tenga tres o más autores, se debe mencionar el apellido del primer autor acompañado de la expresión latina et al., equivalente a “y otros”, en cursivas y con el año (ej: García et al., 2003). La lista completa con las referencias bibliográficas mencionadas se debe incluir al final del artículo. Los apellidos y nombres de todos los autores deben escribirse en redondas, en el orden alfabético de sus apellidos; cuando se citan varias publicaciones del mismo(s) autor(es) deben listarse en orden cronológico. Se prevén algunos casos:
• Para libros: Autor (es), año. Título del libro, edición, casa editora y ciudad de su sede, páginas consultadas (pp. # - #). Ejemplo: Agrios, G. 1996. Fitopatología. Segunda edición. Editorial Limusa, México D.F.
• Para capítulos de libros: Autor (es), año. Título del capítulo, páginas consultadas (pp. # - #). En: Apellidos y nombres de los compiladores o editores (eds.)., título del libro, edición, casa editora y ciudad de su sede. Ejemplo: Bernal, H. 1996. Capítulo 6: Evapotranspiración. pp. 112-125. En: Agrios, G. (ed.). Fitopatología. 2a ed. Editorial Limusa, México D.F.
• Para revistas: Autor (es), año. Título del artículo, nombre abreviado de la revista volumen(número), página-página. Ejemplo: García, S.; W. Clinton; L. Arreaza y R. Thibaud. 2004. Inhibitory effect of flowering and early fruit growth on leaf photosynthesis in mango. Tree Physiol. 24(3), 387-399.
• Para citas de internet: Autor (es), año. Título del artículo. En: Nombre de la publicación electrónica del website, portal o página y su URL, fecha de consulta. Ejemplo: Arafat, Y. 1996. Siembra de olivos en el de-sierto palestino. En: Agricultura Tropical, http://agrotropical.edunet.es; consulta: noviembre de 2003.

Del 8 al 12 de septiembre de 2009, Hotel panorama de la ciudad de paipa, Boyacá (Colombia)
InForMACIÓn e InSCrIPCIÓn
Sociedad Colombiana de Ciencias Hortícolas • http://www.soccolhort.com • e-mail: [email protected]
Teléfonos: (57-1) 316 5000 ext. 19041, 19036, 19043 • Fax: 3165498 • Móvil: (57) 310 202 5293, 315 854 4938, 311 624 0934
La Sociedad Colombiana de Ciencias Horticolas (SCCH) invita al
III ConGreSo ColoMBIAno De HortICUltUrA Y el SIMPoSIo InternACIonAl De CeBollA
Y AJo en el trÓPICo
Simposio Internacional de Cebolla y Ajo en el trópico10, 11 y 12 de septiembre de 2009
programa científico de dos días y medio, en el que se presentarán resultados en áreas como genética y mejoramiento, aspectos fisiológicos y ecofisiológicos, sistemas de producción, plagas y enfermedades, desórdenes fisiológicos, tecnología de poscose-cha, procesamiento, economía, mercadeo y desarrollo rural. El día 12 se realizará una jornada tecnológica dirigida a productores del departamento de Boyacá y otras regiones del país, y finalmente una gira técnica por zonas cercanas de paipa.
III Congreso Colombiano de Horticultura“Ciencia y tecnología hortícola frente al cambio económico y ambiental”
8, 9 y 10 de septiembre de 2009
El programa científico de dos días y medio es el escenario para el intercambio de experiencias y conocimientos en temas relacio-nados con el sector horticola: frutales, hortalizas, ornamentales, hierbas aromáticas y medicinales. El objetivo es que los partici-pantes obtengan información actualizada por especialistas en cada una de las temáticas, conozcan los avances en investigaciones tanto científicas como tecnológicas, y aporten a la planificación y ejecución de proyectos de investigación y extensión, con miras a enfrentar los nuevos cambios en la economía mundial e impacto ambiental.

Forma de pago suscriptores nacionalespago en efectivo o en cheque a nombre de la Sociedad Colombiana de Ciencias Hortícolas o por consignación nacional en el Banco de Bogotá, cuenta corriente 033811761, y envío posterior de copia de recibo y formato de suscripción diligenciado al fax (57+1) 316 5498.
Valor para 2008 (dos números, incluyendo costos postales para el envío): Colombia: $40.000 (pesos colombianos); América Latina y el Caribe: US$ 30; Estados Unidos y Canadá: US$ 50; otras regiones y países: US$ 65.
SUSCrIPCIÓn
revista Colombiana de Ciencias HortícolasSociedad Colombiana de Ciencias Hortícolas-Facultad de Agronomía Universidad Nacional de ColombiaA.A. 14490, Bogotá-ColombiaTeléfono: 571 - 316 5000 Ext. 19041/19043Tel/fax: 571 - 316 [email protected], [email protected]
Nombre / Name
Documento de identidad / Identification document
Dirección / Address Ciudad / City
Teléfono / Phone
Departamento / Department Zona postal / Zip code
País / Country Correo electrónico / E-mail
Profesión / Profession
Institución / Employer
Dirección de envío / Mailing address
Último ejemplar recibido / Last issue mailed.
Año / Year Volumen / Volume Número / Number Fecha de envío / Mailing date



Volumen 2 / N
o. 1 / Enero-Junio 2008Revista C
olombiana de C
iencias Hortícolas
Editorial 5
Seccióndefrutales
Efectodelatemperatura,inmersiónenaguayconcentracióndefitorreguladoressobrelagerminacióndesemillasdepapaya(Carica apayaL.)Effect of temperature, immersion in water and concentration of plant growth regulators on germination of papaya (Carica papaya L.) seedsArletteIvonneGilC.,DiegoMirandaL. 9
Característicasfísico-químicasdefrutosdefeijoainfluenciadasporlaposiciónenelcanopiPhysico-chemical characteristics of pineapple guava fruits as influenced by canopi positionRonaldR.Martínez-Vega,GerhardFischer,AníbalHerrera,BernardoChaves,OmarCamilo
Quintero 21
Influenciadelácidogiberélicoydelnitratodecalciosobreladuraciónposcosechadefrutosdefresa(Fragariasp.)Influence of gibberellin and calcium nitrate on post-harvest conservation of strawberry (Fragaria sp.) fruits
FánorCasierra-PosadayRicardoA.SalamancaG. 33
Respuestasfisiológicasymorfológicasdeplantasdemora(Rubussp.)sometidasaestrésporvientoinducidoPhysiological and morphological responses of blackberry (Rubus sp.) plants to wind-induced stress
FánorCasierra-Posada,OscarE.Aguilar-Avendaño 43
Seccióndehortalizas
Efectodediferentesláminasderiegoysustratosenlapropagacióndetomate(Solanum lycopersicumL.)Effect of different water levels and substrates on tomato (Solanum lycopersicum L.) propagation
YuliDeaquiz-Oyola,JavierÁlvarez-Herrera,AnaFraile 54
Comportamientodelosnutrientesentejidofoliarenbrócoli(Brassica oleraceavar.italica)‘Coronado’yrepollo(Brassica oleracea)hibrido‘Delus’cultivadosenlaSabanadeBogotáNutrient levels in foliar tissue of broccoli (Brassica oleracea var. italica) ‘Coronado’ and cabbage (Brassica oleracea) hybrid ‘Delus’ cultivated on the Bogota PlateauCarlosCarranza,OctavioLanchero,DiegoMiranda 66
Antagonismoin vitrodeTrichodermaspp.sobreaislamientosdeSclerotiniaspp.yRhizoctoniaspp.Antagonism of Trichoderma spp. against isolates of Sclerotinia spp. and Rhizoctonia spp. in vitro
LillianaHoyos-Carvajal,GloriaDuque,SergioOrduzP. 76
AbundanciaydiversidaddeBacillusthuringiensisdediferenteshábitatentresmunicipiosdeBoyacá,ColombiaAbundance and diversity of Bacillus thuringiensis in different habitats within three municipalities of Boyaca, ColombiaJeimyM.Poveda,JohnW.Martínez 87
Seccióndeornamentales
Aproximacióndeladinámicapoblacionaldelosmicroorganismosendiferentessustratosempleadosenelcultivoderosa(Rosaspp.var.Charlotte)enlaSabanadeBogotáOverview of the population dynamics of microorganisms in different substrates used in rose (Rosa spp. var. Charlotte) cultivation on the Bogota PlateauMilenaAlfonso,AlejandroCoca,WilliamRamírez,LilianaHoyos-Carvajal 98
Seccióndeotrasespecies
Estudiosdeadaptaciónymanejointegradodeestevia(Stevia rebaudianaBert.):nuevaalternativaagroindustrialdelCaribecolombiano.UnarevisiónEnvironmental suitability and integrated management studies in stevia (Stevia rebaudiana Bert.): a new agro-industrial alternative for the Colombian Caribbean. A review
AlfredoJarma-Orozco 109
Políticaeditorialeinstruccionesparalosautores 121





























