Embed Size (px)
Citation preview

–
N°9 Mayo-Junio 2004
• REVISTA DE BIOQUÍMICA ONLINE
DE CÓMO LA FUNCIÓN MUSCULAR REGULA LA EXPRESIÓN DE GENES EN LA CÉLULA.
Enrique Jaimovich. U. de Chile
• PIONEROS DE LA BIOQUÍMICA Matrimonio Gerty y Carl Cori.
• CIENCIA AL DIA • Se crea un 'chip' con todo el genoma de una bacteria. • Crisis energetica en Chile • Potenciando la tolerancia a la salinidad en Plantas. • Srta. Kaguya: ¿Serán prescindibles los hombres?
• : TECNICA DE UN BIOQUÍMICO
Western Blot
• BIOQUIMICA PATOLOGICA Enfermedad alzheimer segunda parte.

3 CARTA DEL DIRECTOR
4-12 CIENCIA AL DIA SE CREA UN 'CHIP' CON TODO EL GENOMA DE UNA BACTERIA.
CRISIS ENERGETICA EN CHILE. POTENCIANDO LA TOLERANCIA A LA SALINIDAD EN PLANTAS.
SRTA. KAGUYA: ¿SERÁN PRESCINDIBLES LOS HOMBRES?
13-14 CIENCIA EN CHILE. DE CÓMO LA FUNCIÓN MUSCULAR
REGULA LA EXPRESIÓN DE GENES EN LA CÉLULA. Enrique Jaimovich. U. de Chile
15-16 PIONEROS DE LA BIOQUIMICA GERTY Y CARL CORI
17-21 BIOQUIMICA PATOLOGICA ALZHEIMER SEGUNDA PARTE
22-23 TECNICA DE UN BIOQUIMICO WESTERN BLOT
25 TRIBUNA DEL ESTUDIANTE LEONARDO PARRA
Breve de Arte. Promoción 2002. ANÓNIMO
Primer Año 2004.
35 HUMOR GRAFICO
34 PERSONAJE DEL MES RICARDO HERRERA.
Promoción 2000
24 TRIBUNA DEL PROFESOR ROMANCE DE UN SUSTRATO.
Pedro valencia Araya. Bioquimico. PUCV
32 ENTREVISTA PATRICIA EWERT
27-31 BIOINFORMATICA
49GALERIA FOTOGRAFICA
26 TRIBUNA DEL TESISTA CLAUDIA PINO.
Promoción 1998. Bioquímica.
33 EXALUMNOS: ALLISSON ASTUYA VILLALON
25 AGENDA BIOQUIMICA

CARTA DEL DIRECTOR
DIRECTOR CARLOS LIZAMA
EDITORES KELLY CAUTIVO CARLOS LIZAMA
REDACCION KELLY CAUTIVO CARLOS LIZAMA ALVARO GONZALEZ PABLO TAPIA
REPORTEROS KELLY CAUTIVO PABLO TAPIA CARLOS LIZAMA ALVARO GONZALEZ DISEÑO GRAFICO KELLY CAUTIVO CARLOS LIZAMA PABLO TAPIA NUESTRA PORTADA
E-MAIL: [email protected]
. Secuencia de tres imágenes de microscopía confocal de un miotubo (célula de músculo esquelético de rata en cultivo) cargado con el colorante fluorescente sensible a calcio fluo-3. La primera imagen muestra la fluorescencia basal, la segunda muestra la fluorescencia aumentada en el citoplasma y en los núcleos celulares, 6 s luego de despolarizar la membrana con alto potasio y la tercera imagen muestra el retorno a condiciones cercanas a la basal 30 s después. Las imágenes fueron tratadas con una escala de pseudocolor que se muestra en la parte inferior de la figura.
Estimados lectores junto con saludarlos queremos expresarles nuestras disculpas ya que debido a problemas de fuerzas mayores cuya solución no estaba en nuestras manos, este numero tardo un poco mas en salir. Esperamos que esten semestre recien pasado todos hayan salido con un saldo positivo en lo que concierne a sus ramos, y los que no bueno obligados a estudiar un poco mas. Como siempre queremos invitarlos a participar en nuestra revistan, ya sea enviando noticicias, temas de interes, sus opiniones y otros. Además recuerden que pueden enviarnos mail solicitando ayuda respecto a cualquier asignatura. Les comunico además que este semestre comienza el ciclo de seminarios de nuestra carrera que apartir del viernes 13 a las 11:45 am parte. Por ultimo, permitanme comunicarle la lamentable noticia de la muerte de Francis Crick uno de los descubridores de la doble helice del ADN, quien fallecio de un cancer de colon, a los 88 años de edad. Por esto, les pedimos un minuto de reflección por uno de los padres de la genetica que se nos va, y en honor a el nuestra revista traera un articulo especial hacerca de Watson y Crick en el proximo número. Bueno, esperando que disfruten de este número y que nos envien sus mails me despido.
Carlos Lizama V Alumno Tesista
“Siempre recordaré a Francis por su inteligencia extraordinariamente centrada y por las muchas maneras en que me demostró su bondad y colaboró para elevar mi autoconfianza" James Watson.

Un investigador español crea un 'chip' con todo el genoma de una bacteria. El Centro Nacional de Biotecnología ha albergado la iniciativa de desarrollo del primer biochip capaz de contener la totalidad de los genes de un microoorganismo simple, la proteobacteria Pseudomonas putida KT2440, y bajo condiciones ambientales muy diversas. Este proyecto de expresión génica podría tener relevantes aplicaciones biotecnológicas.
Un equipo español coordinado por el investigador Fernando Rojo, del Centro Nacional de Biotecnología (CNB) del Consejo Superior de Investigaciones Científicas, ha conseguido culminar el desarrollo de un chip para analizar la expresión de todos los genes del genoma de la Pseudomonas putida KT2440. Este microorganismo pertenece al género de las bacterias aerobias Gram negativas y suelen causar infecciones urinarias, ya que son patógenos ocasionales localizados en suelo y agua. El hallazgo tiene una especial relevancia para la comunidad investigadora, ya que se trata del primer biochip de ADN que incluye todos los genes de una bacteria totalmente desarrollado en España y por científicos españoles. Sus posibles aplicaciones son realmente extensas. En primer lugar, este nuevo microarray de ADN permitirá abordar de una manera global y con mucha más rapidez problemas que hasta ahora sólo se habían investigado con enfoques parciales y de bajo rendimiento, aumentando la profundidad, alcance y fecundidad de los estudios. En segundo lugar, los investigadores del CNB utilizarán el chip para analizar la expresión diferencial del genoma del microorganismo en distintas condiciones ambientales relevantes para aplicaciones biotecnológicas con el objetivo de exprimir todo su potencial. Por último, los resultados obtenidos ayudarán a los investigadores a entender cómo se regula la expresión de los genes implicados en la biodegradación de hidrocarburos y compuestos aromáticos. También facilitará el análisis del metabolismo del nitrógeno cuando las bacterias crecen en suelos contaminados, con lo que se podrá optimizar el rendimiento de los procesos de biodegradación y biotransformación, así como desarrollar estirpes para aplicaciones fitosanitarias. El estudio global del funcionamiento de las bombas de expulsión de fármacos y solventes, tanto en estas bacterias como en otras pseudomonas, mejorará el diseño de mecanismos de inhibición de estas bombas, que están implicadas, por ejemplo, en la multirresistencia frente a los antibióticos y otros aspectos de interés clínico. El nuevo chip permite analizar la expresión de los 5.350 genes detectados hasta el momento en esta bacteria, así como los 140 genes detectados en el plásmido TOL presente en Pseudomonas putida mt-2, bacteria de la que deriva la estirpe KT2440. Para comprender el funcionamiento y comportamiento de los seres vivos es esencial poder analizar la expresión de sus distintos genes en diferentes condiciones. Un organismo simple como una bacteria puede tener entre 500 y 8.000 genes, mientras que el ser humano tendría, según las estimaciones más recientes, algo más de 30.000. Analizar la expresión de un gen concreto es ya una tarea habitual en los laboratorios de biología molecular, pero analizar la expresión de todos y cada uno de los genes de un organismo, o incluso de grandes grupos de genes, es una labor difícil de abarcar. Tanto la bacteria Pseudomonas putida KT2440 como el plásmido TOL tienen una gran
importancia biotecnológica. Se trata de una bacteria del suelo, fácil de manipular genéticamente y muy utilizada como hospedador en el análisis y manipulación de genes. En 1981 el Comité de ADN Recombinante de los Institutos de Salud de Estados Unidos certificó que esta bacteria no es patógena para animales y plantas, y que se puede considerar un hospedador seguro para genes de diverso origen. Esta cepa se utiliza actualmente en aplicaciones biotecnológicas muy variadas, como la síntesis industrial de compuestos químicos. Además, es la bacteria modelo utilizada desde hace varios años en una extensa mayoría de estudios sobre el modo en que las bacterias degradan compuestos tóxicos. La Pseudomonas putida KT2440 es el hospedador natural del plásmido TOL pWW0, que codifica una ruta para degradación de tolueno y xilenos, la ruta de biodegradación mejor caracterizada en la actualidad desde el punto de vista bioquímico y genético. La bacteria es además capaz de colonizar el sistema radicular de plantas y formar biopelículas. Las bases moleculares de esta colonización por bacterias del género Pseudomonas son de momento desconocidas, pero su elucidación abrirá un amplio abanico de posibilidades para su uso como biopesticidas. De hecho, Pseudomonas putida KT2440 está considerada como biopesticida potencial, tanto para las raíces como para las hojas, y es también un importante sistema modelo para estudiar los mecanismos bacterianos de resistencia a solventes orgánicos.

Crisis energética en Chile: recursos energéticos abundantes y limpios.
Ha llegado el momento de tomar una decisión vital que puede cambiar el futuro de Chile como nación energéticamente dependiente. Disponemos de los recursos naturales y de los grupos humanos capaces de
llevar a cabo esta tarea. ¿Qué estamos esperando?
En el tema de la energía los chilenos debemos aprender de las paradojas. Que el gas sea un recurso disponible en enormes yacimientos en el subsuelo de Argentina y Bolivia, no garantiza que Chile, como uno de sus principales consumidores, tenga el suministro asegurado. Eso ha quedado muy claro las últimas semanas. Resulta paradojal, también, que de tanto acostumbrarnos a nuestra geografía, a paisajes donde el viento azota pampas, cordilleras y valles y donde el agua brota a cientos de grados desde napas subterráneas, sean pocas las iniciativas nacionales en que se piensa incorporar seriamente a estos recursos como fuentes de autonomía energética. Desde hace varios años, un grupo de investigadores de la Pontificia Universidad Católica de Valparaíso (PUCV) ha demostrado la existencia de vastos yacimientos de
hidratos de gas metano congelado en el lecho marino de nuestras costas. Los investigadores afirman que los depósitos gasíferos serían capaces de satisfacer las exigencias energéticas de Chile durante los próximos dos siglos… a lo menos. El año 2001 FONDEF de CONICYT adjudicó a la Universidad de Magallanes un proyecto destinado a proveer de energía eólica a la Decimosegunda Región. Sus principales beneficiarios serán los ganaderos y agricultores de la zona, que podrán disponer de la energía del viento como un insumo abundante y barato. Les permitirá bombear agua desde napas subterráneas. Estimaciones conservadoras afirman que la energía del viento podría generar un 20% de la electricidad mundial hacia el año 2040. De hecho, Dinamarca ya produce más de un 10% de su electricidad con energía eólica y los planes del gobierno danés apuntan a que la cifra alcance el 50% para el año 2030. Según la European Wind Energy Association, con la energía eólica el planeta podría evitar que asombrosas cantidades de CO2 vayan a parar a la atmósfera: unos 70 millones de toneladas de CO2 menos hacia el año 2005, lo que subiría a unos 270 millones al 2010 y a la increíble cifra de 1.780 millones de toneladas menos de CO2 al 2020. Este promisorio futuro dependerá, eso sí, de la cantidad de turbinas instaladas y de su capacidad de generación. En términos de capacidad eléctrica instalada, los estudios indican que las turbinas eólicas pasarán de los 18.000 mw actuales a unos 70.000 mw hacia el 2010, lo que se dispararía a 1.200.000 mw al 2020 y a 3.000.000 mw al año 2040. En esos años el costo de generación, se estima, bajará de los actuales US$ 0,04 por kwh a la mitad (US$ 0,02 por kwh) lo que sitúa a la alternativa eólica entre las fuentes de generación energética más baratas. Al 2020 se estima que la capacidad instalada en Latinoamérica en energía eólica será de unos 72.000 mw, con lo cual se podría deducir que en Chile la capacidad eólica podría llegar a unos 4.000 mw a esa fecha, lo que equivale, por ejemplo, a la mitad de la capacidad eléctrica total instalada al año 2000. Por otro lado, se tiene la energía geotermoelectrica que se ha mantenido como una idea postergada. Chile tiene un buen potencial geotérmico porque está situado sobre el Cinturón de Fuego del Pacífico. En nuestro país, las fuentes de energía geotérmica radican mayoritariamente en el norte del país. La paradoja parece imponerse nuevamente, pues es el norte la zona más afectada por las restricciones de gas impuestas por el gobierno argentino. Hoy los sistemas eléctricos deben elegir entre aumentar sus costos o dejar de satisfacer la demanda. Lo más probable es que en las regiones nortinas sea el petróleo el combustible utilizado para generar electricidad. Sin embargo, podrían desarrollarse otras alternativas de largo aliento y profundo potencial. Justamente, en las montañas del norte grande brotan fluidos con temperaturas de hasta 260°C, que podrían permitir no sólo la generación eléctrica sino también la producción de agua fresca, la recuperación de elementos químicos contenidos en los fluidos geotermales y la explotación de importantes recursos minerales no-metálicos. Estas aplicaciones necesitan emanaciones de alta temperatura. Sin embargo, en Chile existen muchas áreas de temperatura baja-media (hasta 180°C), en las que su uso se ha restringido a la recreación y a propósitos médicos. Hoy, los investigadores piensan que es esencial que las investigaciones se dirijan a la promoción de diferentes usos de la energía geotérmica como en otras partes del mundo.

Para medir el potencial de la energía geotérmica y estudiar su real impacto, el profesor Lahsen trabaja desde hace varios años con un grupo de investigadores de la Universidad de Chile.
Gracias a un proyecto financiado por FONDEF el año 1999 a la Universidad de Chile, se realizaron las primeras exploraciones abarcando toda la zona desde Talca a Osorno. Respecto de las prospecciones, Lahsen relata: "Encontraremos los lugares más aptos para iniciar estudios detallados y contar así con todos los antecedentes acerca de los recursos geotérmicos disponibles en esas zonas. Después, ENAP -empresa que apoya el estudio y que participa en el proyecto-podrá iniciar, sola o en conjunto con otras compañías privadas, la construcción de la primera central de energía geotérmica en el país". Lahsen estima un potencial de energía geotérmica utilizable en la generación de electricidad del orden de 16.000 MW, esto es, aproximadamente el doble de la capacidad eléctrica instalada actualmente en el país. Sólo a través de la energía geotérmica podríamos llegar a ser un país autoabastecido en el mediano plazo.
Los costos de construcción de una planta de energía geotérmica se comparan con los de una central hidroeléctrica. La gran diferencia es que se trata de un recurso completamente renovable, que tiene una duración mínima de 50 años y que no presenta impactos negativos para el medio ambiente. Incluso, en algunos países que utilizan activamente este recurso, el agua y el vapor son reinyectados en el subsuelo para ser utilizados nuevamente. El proyecto, concluido el año pasado, ha permitido a Lahsen determinar la potencialidad de energía geotérmica de la zona central-sur de Chile, asociada al arco volcánico de la cadena andina. La potencialidad de la zona norte está clara desde los años 70, época en la que incluso se iba a construir una planta que no llegó a concretarse, debido al complicado entorno político económico. De acuerdo con las características físicas y geoquímicas de las áreas termales investigadas, se han establecido las posibilidades de utilización, ya sea para ser empleadas en forma directa como calor o para la generación de electricidad. El grupo de investigadores ya tendría seleccionado y en estudio los dos sistemas geotermales más promisorios de esas áreas, para fines de generación eléctrica. Evidencia sobre la viabilidad de este tipo de proyectos existe y en forma abundante y suficiente. Sin embargo, hasta el año 2000 no hubo una señal clara de parte de las autoridades que permitiera considerar esta opción dentro de un marco legal definido. La Comisión Nacional de Energía (CNE) incluyó recién el año 2000 y por primera vez en su historia, en su plan anual de obras, a esa fuente energética como una opción de desarrollo. También ENAP actúa como una contraparte sigilosa que prefiere que su apoyo a este tipo de iniciativas se mantenga en el silencio. Tras la promulgación de la Ley sobre Concesiones de Energía Geotérmica, en enero del 2000, se han otorgado 12 concesiones de exploración (entre ellas las de CODELCO, ENAP -en sociedad con la minera y la francesa CFG-, a CORFO y a la U. de Chile). Según la CNE, ha habido nuevas solicitudes aunque ahora son de compañías netamente privadas. La similitud que hay en este negocio en la etapa de exploración petrolera hizo que ENAP fuera una de las empresas que lideran la búsqueda de reservorios. Tiene seis concesiones de exploración en Chile. De hecho, hay un estudio para una central en la zona de Calabozo, frente a Talca. La empresa estatal está asociada con una compañía francesa para este proyecto. La iniciativa contempla una central operativa hacia 2009. Aportaría 300 megawatts (MW), equivalentes a una central de ciclo combinado a gas. Sin embargo lo que falta, además de perforaciones para medir el verdadero potencial geotérmico, es una decisión férrea de las autoridades para apoyar el tema. Evidencias de la viabilidad de estas iniciativas existe y en grandes cantidades. Una central geotermoeléctrica funciona como una planta termoeléctrica convencional, la que a través de combustibles como carbón, petróleo o gas natural produce el vapor que luego mueve las turbinas para generar electricidad. En el caso de la energía geotérmica, la caldera de vapor se encuentra en forma natural bajo la superficie de la tierra. Estos reservorios, ubicados hasta una profundidad de unos 4 Km., son económicamente explotados mediante pozos que extraen el agua caliente o vapor que accionan las turbinas para generar la electricidad. Hasta ahora, y a pesar de la urgencia dejada de manifiesto por el racionamiento argentino de gas natural hacia Chile, no ha existido la voluntad política para apoyar la constitución real de este tipo de centrales. Cuenta pendiente de la clase política con el desarrollo nacional. Pero si lo expuesto hasta ahora parece promisorio, el recurso que se lleva los mayores méritos en términos de expectativas y potencial es el de los hidratos metanos submarinos.

Diversos estudios demuestran que el mar esconde la mayor reserva de energía del planeta: los hidratos de gas metano, principal fuente de gas natural. De hecho, ellos representan el 55 % de las reservas del planeta, lo que hace prever que serán la energía del futuro, la que reemplazará a la era del petróleo a partir de la mitad del presente siglo, cuando se agoten los yacimientos del combustible fósil. Los hidratos de gas, que contienen grandes concentraciones de metano -principal componente del gas natural- se encuentran ubicados en los fondos marinos de la mayoría de los márgenes continentales, incluyendo la costa chilena. Debido a que muchos países no tienen fuentes de energía tradicionales, como el petróleo, pero sí tienen costas marítimas, la potencial explotación de hidratos de metano en sus aguas jurisdiccionales a mediados de este siglo, podría provocar un vuelco en la estructura económica mundial. Los hidratos de gas constituyen una fuente energética de gran proyección
mundial, con reservas estimadas que prácticamente duplican las reservas convencionales actualmente reconocidas para los recursos energéticos fósiles. Dadas las enormes expectativas que abre su exploración, fue que FONDEF de CONICYT apoyó la realización del proyecto como "Hidratos de Gas Submarinos, una Nueva Fuente de Energía para el Siglo XXI", el cual busca, en una primera etapa, caracterizar y evaluar la presencia del recurso y sus implicancias medioambientales en las costas chilenas. De acuerdo al equipo de investigadores, el resultado del estudio, en conjunto con la compilación de información previa, "darán una acabada caracterización de los hidratos de gas en el margen chileno". Para la realización de las prospecciones e identificación de los recursos se cuenta con la colaboración de centros de investigación y expertos de nivel mundial en el tema, lo que acarreará una transferencia tecnológica invaluable en estas áreas. Los hidratos de metano se localizan en la geoesfera submarina de poca profundidad, sistema finamente balanceado, en equilibrio con todos sus componentes tales como sedimentos, el agua de poros, los flujos de fluidos, la presión, la temperatura, el agua que cubre la capa de sedimentos con hidratos, etc. De ahí el enorme desafío que implica su posible extracción, ya que la remoción de cualquier componente de este equilibrio puede desestabilizar todo el sistema dando lugar a daños irreparables. Los factores desestabilizantes, que pueden ser perturbaciones naturales o asociadas con la explotación de los hidratos, podrían potencialmente afectar el clima y el ambiente geológico a una escala global catastrófica. Los hidratos de gas han sido asociados con problemas ambientales, principalmente, debido a que el metano es muy eficiente en la generación del efecto invernadero, con un potencial de alrededor de 10 veces al del dióxido de carbono. Por otra parte, se ha logrado establecer que avalanchas submarinas ocurren en acumulaciones inestables de sedimentos del fondo oceánico. En algunos casos, estas remociones en masa pueden ser causadas por la descomposición de hidratos de gas y la expansión o liberación del gas resultante. De acuerdo a Esteban Morales, los resultados de este proyecto serán esenciales para la definición de normas medioambientales que aseguren una explotación sustentable del recurso. "El proyecto entregará, además, parámetros para la evaluación de los riesgos asociados a la ocurrencia, por ejemplo; de avalanchas submarinas y localización de zonas con eventuales fugas naturales de gases asociados a los hidratos de metano, desencadenadas por actividad sísmica natural". Aunque en Chile existiría lo necesario como para acabar con la dependencia energética extranjera, aún no se cuenta con la tecnología adecuada para explotarlo. Sin embargo, ya existen experiencias piloto en Japón que arrojan nuevas luces al respecto. Se espera que los resultados del proyecto estén disponibles a partir de septiembre de este año. A través de ellos, se podrá visualizar tanto la ubicación exacta de los yacimientos como también la cantidad de reservas existentes. La crisis que vive Chile producto de los recortes en el suministro de gas argentino ha dejado valiosa evidencia al descubierto. Hoy estamos más conscientes que nunca que tenemos recursos de sobra para ser autosuficientes utilizando, además, energías limpias y baratas. También es un hecho que existen en Chile grupos humanos altamente especializados que han creado las capacidades para facilitar la toma de decisiones basadas en criterios científicos y tecnológicos. Pero lo preocupante es que también esta crisis ha demostrado que ha sido la falta de voluntad política la que ha atrasado las inversiones en el tema. Todos los esfuerzos estuvieron concentrados por un largo tiempo en la opción fácil de comprar gas barato a Argentina y para ello se hicieron cuantiosas inversiones. Si el mismo nivel de inversiones hechas para transportar gas a través de Los Andes, amén de los esfuerzos por cambiar casi por completo los sistemas de distribución y uso, se realizara en alguna o en todas las energías descritas, Chile sería otro muy distinto de aquí a diez años, a lo sumo.

INGENIERIA GENETICA.
Potenciando la tolerancia a la salinidad en Plantas.
A través de una investigación realizada por el laboratorio de Biología Celular y Genética de la Universidad de Talca, encabezado por el doctor Simón Ruiz, se han lanzado a la identificación de los mecanismos que permiten a las plantas resistir altos grados de salinidad y a tratar de transferir esas propiedades a diferentes
cultivos. El paso crucial en la investigación consiste en determinar del rol que juegan los genes aislados en la tolerancia al estrés
salino observado en esta planta silvestre. Una de las estrategias más utilizada para tales efectos es el silenciamiento de los genes
en estudio.
Aproximadamente el 20% de la superficie terrestre utilizada para el cultivo en el mundo se encuentra afectado por niveles de salinidad que superan la tolerancia de las especies de cultivos tradicionales. Este porcentaje va en aumento a una tasa del 0,5% anual, debido fundamentalmente a bajas precipitaciones, alta superficie de evaporación, irrigación con aguas salinas y por las prácticas tradicionales de cultivo que favorecen el incremento de la concentración de sales en el suelo. Este estrés salino afecta directamente el rendimiento de los cultivos, inhibe su
óptimo desarrollo y en algunos casos puede conducir a la muerte de la planta. Una de las sales que causa mayor perjuicio es el cloruro de Sodio (NaCl). En los últimos años, el estrés salino en plantas provocado por NaCl ha sido un tema de estudio recurrente, tanto desde el punto de vista fisiológico, molecular y genético. Desde la perspectiva fisiológica el resultado de altos niveles de cloruro de sodio sobre las plantas considera dos componentes principales: un componente osmótico y uno iónico.
El primero se refiere a la privación del agua y constituye un factor común tanto para estrés salino como para estrés por sequía y frío. El segundo se relaciona con el efecto tóxico que provoca la disminución de la razón Potasio (K+)/ Sodio (Na+) a nivel celular y es propio de este tipo de estrés. El aumento en los niveles de NaCl en el medio lleva consigo un incremento en el influjo de Na+ a las raíces y su distribución por toda la planta. El ingreso sin control de iones Na+ por transporte pasivo ocurre como resultado de la generación de un gradiente electroquímico muy elevado entre ambos lados de la membrana celular de las raíces. Los iones Na+ ingresan al interior del citoplasma gracias a la presencia de canales y transportadores catiónicos en la membrana plasmática, los cuales son bastante inespecíficos frente a un alza en la razón Na+/K+ en el medio. La disminución en el contenido de iones K+ en el citoplasma trae como consecuencia una desestabilización en el potencial de membrana, la inactivación de enzimas y un efecto perjudicial sobre una serie de procesos fisiológicos. El restablecimiento de la homeostasis iónica, no es un problema de fácil solución para las plantas, ya que a diferencia de las células animales, ellas carecen de transportadores de Sodio, tales como las Na+-ATPasas o las Na+/K+-ATPasas, por lo que deben recurrir a las H+-ATPasas o H+-pirofosfatasas para generar un gradiente electroquímico de protones que permitan el intercambio de H+ por Na+ y también de otros iones y metabolitos.
Halófitas y Glicófitas
Las plantas pueden ser clasificadas en dos grupos, según su capacidad de tolerar la salinidad: Halófitas y Glicófitas. Las plantas halófitas, a diferencia de las glicófitas, se caracterizan por poseer una alta tolerancia a la salinidad, condición que en algunos casos esta dada por la presencia de estructuras especializadas, como glándulas excretoras o vesículas almacenadoras de sales. Sin embargo, en la mayoría de los casos las diferencias entre halófitas y glicófitas esta determinada por la expresión diferencial de genes, que de forma coordinada provocan cambios en el metabolismo celular, protegiendo la planta de la acción osmótica y tóxica de la sal asegurando un adecuado nivel de división y expansión celular, durante el tiempo que perdure el estrés.
Otro importante grupo de genes son los encargados de la homeostasis de agua dentro de la planta, entre ellos destacan los genes para acuaporinas y los que codifican para las proteínas encargadas de regular el cierre estomático, los cuales suelen ser los mismos genes que participan en el estrés por sequía y frío, ya que son inducidos por el efecto osmótico. En la acción indirecta, han sido identificados un importante número de genes cuyos productos son de eliminación de radicales libres y que actúan después de la formación de elementos reactivos de oxígeno (ROS), el cual es un efecto bastante común de diversos tipos de estrés abiótico. Estos productos génicos están participando activamente en la protección del efecto nocivo de los elementos ROS, en la membrana citoplasmática y en diversos organelos, tales como: mitocondrias y cloroplastos.
De acuerdo a los antecedentes previos la tolerancia al estrés salino aparece como un problema multifactorial y por lo tanto de difícil solución, más aún cuando los grados de tolerancia a la salinidad de una planta puede ser muy distintos en los diferentes estados específicos de su ontogenia, tales como: germinación y emergencia, crecimiento y sobrevivencia de la plántula, y crecimiento vegetativo y reproductivo.
Lycopersicon chilense

Los primeros avances conducentes a la generación de variedades cultivables de una relativa tolerancia a la salinidad, vino desde la genética clásica asociada a técnicas de biología molecular que permitieron la identificación de QTLs (quantitative trait loci) específicos de la tolerancia a la sal, en tomate y arroz. Sin embargo, los programas de mejoramiento genético de las especies cultivables, solo han podido lograr variedades moderadamente sensibles al estrés salino en todos los estados de desarrollo de la planta. En contraposición a lo observado y descrito, el grupo de investigación del Dr. Eduardo Blumwald de la Universidad de California Davis, ha logrado aumentar la tolerancia a este tipo de estrés en plantas transgénicas de tomate, canola y A. thaliana, con la sobreexpresión de un único gen que codifica para un antiporte vacuolar de Na+/H+. En estas plantas, el Sodio es acumulado en hojas, observándose además que los frutos y las semillas no son afectadas en su calidad. Similares resultados, respecto del aumento de la tolerancia a salinidad, ha sido mostrada en Arabidopsis thaliana, cuando es sobreexpresado el gen SOS1, el cual corresponde a un antiporter Na+/H+ de la membrana plasmática, y también cuando se ha realizado la sobreexpresión del gen AVP1, el cual codifica para H+-pirofosfatasa vacuolar. Estos resultados, sugieren que la forma más rápida de incrementar la tolerancia a la sal consiste en aumentar la razón K+/ Na+ en el citosol de las células vegetales, lo cual se consigue mediante la extrusión del Sodio y/o por la acumulación de éste en las Vacuolas. Se ha sugerido que una estrategia alternativa consistiría en aumentar la proporción de transportadores más selectivos de K+ en la raíz de la planta. Un modelo chileno Lycopersicon chilense, es una planta silvestre de tomate que se distribuye entre los 100 y los tres mil metros de altitud en el desierto de Atacama, y que dadas las condiciones extremas de su hábitat, ha logrado subsistir en condiciones de alta salinidad del suelo y de escasez hídrica. Dadas las características de esta planta, en el laboratorio de Biología Celular y Genética de la Universidad de Talca, nos pareció un excelente modelo donde estudiar los genes de expresión diferencial que se estaban manifestando durante el estrés por NaCl. En el año 2002 iniciamos esta investigación, cofinanciada por la Fundación de Innovación Agraria (FIA). Una de las primeras cuestiones a definir fue si esta planta absorbía el Sodio o bien gran parte de él era excluido por las raíces, ya que esta última posibilidad es una de las alternativas observadas en plantas halófitas. Nuestros resultados mostraron que efectivamente el Sodio era abundantemente absorbido y que mayoritariamente era acumulado en las hojas, lo cual sugería que esta planta desarrollaba mecanismos moleculares eficaces para contrarrestar el efecto osmótico y tóxico ocasionado por la sal. Con estos antecedentes la pesquisa de genes diferencialmente expresados se realizó mediante las técnicas de differential display y PCR-Select. Así, se identificaron más 20 genes implicados en distintos procesos metabólicos y de regulación de estructuras celulares, seleccionándose aquellos que por las características funcionales de sus productos génicos aparecían como más interesantes por su implicancia en procesos y estructuras claves para el óptimo funcionamiento de la planta. Entre los genes seleccionados, cabe mencionar los que se encuentran participando en el mantenimiento de la funcionalidad de la maquinaría fotosintética, la regulación del cierre estomático, la impermeabilización de la hoja, la regulación del gradiente electroquímico de protones y los de síntesis y acumulación de osmolitos compatibles. Algunos de ellos son también expresados cuando las plantas son sometidas a estrés por deshidratación, con lo cual parecen responder al efecto osmótico, aspecto común de ambos tipos de estrés. Sin embargo, el aislamiento de los genes resultó un poco más complejo de lo esperado, ya que la mayoría de ellos corresponden a familias multigénicas. En estas familias, hay genes que son regulados de manera tejido-especifico, otros durante determinados estados del desarrollo de la planta y además, están los activados por el estrés que a su vez son expresados de manera tejido-especifico.
Lycopersicon chilense

EQUIPO DE TRABAJO: Dra. Marcela Salazar, doctora Mariana Rodríguez, doctor Gerardo Tapia, Licenciada Mónica Yañez, doctor Simón Ruiz, Jesica Valdés, técnico de laboratorio; Nilo Mejía, estudiante de doctorado; y doctor José Loyola.
El paso crucial en la investigación descrita, consiste en la determinación del rol que juegan los genes aislados en la tolerancia al estrés salino observado en esta planta silvestre. Uno de las estrategias más utilizada para tales efectos, es el silenciamiento de los genes en estudio, procedimiento que tradicionalmente se ha realizado utilizando los mismos genes pero con posición antisentido, de tal manera, que al transcribirse el gen en estudio, también se transcribe el antisentido, formándose un RNA duplex, formado por ambos RNAs mensajeros que no puede ser traducido por los ribosomas y termina siendo degradado por las RNAasas intracelulares (enzimas que hidrolizan RNA). La otra forma de estudiar el efecto de un gen en particular, es obtener su sobreexpresión en plantas transgénicas de su misma especie o en plantas heterólogas. Dicha sobreexpresión se obtiene, regularmente, colocando la región codificante del gen bajo el control de un promotor constitutivo como es el 35S del Virus del Mosaico de la Coliflor. Sin embargo, no es el método más apropiado cuando se estudian genes de expresión tejido-específico, por lo que la transformación debe ser realizada con el propio gen, lo cual implica un caracterización más acabada de su promotor y de los elementos en cis reguladores de su expresión. La otra posibilidad es colocar la región codificante del gen bajo la regulación de un promotor de otro gen que también se exprese en el mismo tejido y del cual se conozca como puede ser inducido exógenamente. Varios de los genes aislados se encuentran en la fase de estudio de su rol en la tolerancia al estrés salino, por cuanto para su análisis se están utilizando algunas de las estrategias anteriormente mencionadas. Los resultados que aquí se obtengan servirán como base para comprender en parte como algunas plantas halófitas logran dar solución al problema de la salinidad del suelo, el conocimiento aquí generado lo podemos utilizar en función del incremento de la tolerancia al estrés salino de las plantas de importancia económica y así aumentar la actual superficie cultivable o al menos contrarrestar la pérdida anual de tierras aptas para cultivo.
La reunión de receptores de membrana acelera la apoptosis. Un equipo del Centro de Investigación del Cáncer (CIC), de Salamanca, trabaja en una nueva vía para acelerar la apoptosis en células tumorales, después de que hayan observado que es posible actuando directamente sobre la maquinaria de la muerte celular. El enfoque se centra en la concentración de receptores de muerte celular en los microdominios de membrana denominados rafts. El objetivo es diseñar fármacos capaces de concentrar un número elevado de receptores de muerte en esa zona para producir una señal apoptótica con gran potencia que acabe con la célula rápidamente. Frente a los antitumorales actuales, que acaban provocando apoptosis, pero indirectamente y tras varios pasos de señalización complejos que implican la participación de sensores, que a menudo están mutados en la célula tumoral, los fármacos proapoptóticos permiten obviar la cadena de sensores (por ejemplo, p53, mutado en más del 50 por ciento de los tumores), que trasladan la señal de daño celular a muerte o apoptosis de la célula. Lo que hacen es "dar directamente a la célula la señal de que tiene que morir". De esta forma se obvian muchas mutaciones de genes importantes presentes en cáncer, pues se trata de actuar directamente en la maquinaria de muerte celular programada. El equipo de Salamanca, que ya ha obtenido resultados preliminares positivos con un éter lípido antitumoral, trabaja en el desarrollo de nuevos proapoptóticos, denominados así, según el experto, porque van dirigidos a la eliminación por apoptosis de forma directa. El trabajo se trata de la intervención a nivel de los receptores de muerte celular, entre los que destacan dos proteínas: Fas y el receptor de Trail. En cuanto al mecanismo de muerte descubierto, el investigador ha señalado que se conocen determinados receptores de muerte celular, bien descritos a nivel bioquímico, que están en la membrana y no tienen actividad enzimática, a diferencia de otro tipo de receptores. Esos receptores de muerte funcionan como una especie de pegamento de otras proteínas, de forma que a su alrededor se va formando un complejo o andamiaje. Al final de este complejo existe una caspasa, molécula fundamental para apoptosis, ya que se trata de una proteasa que tras su activación rompe proteínas clave para la vida de la célula, de forma muy específica y en sitios muy concretos. El grupo de Mollinedo ha logrado ya con un determinado compuesto atraer receptores de muerte dispersos y concentrarlos en los dominios rafts de membrana, observando que esa agregación dispara una señal muy potente de muerte. Lo que no se sabe aún es cómo la sustancia logra unificar los receptores en esa zona. En este sentido, por el momento, el investigador no concede tanta importancia a la sustancia utilizada como al nuevo mecanismo descrito para acabar con células tumorales, pues se trata de "una nueva forma de atacar tumores vía apoptosis". Los resultados de laboratorio son más "claros y notorios en leucemias", aunque se están tratando de extrapolar a tumores sólidos, que suelen ser más complejos, ya que su mecanismo de muerte presenta algunas variaciones respecto a los cánceres hematológicos.
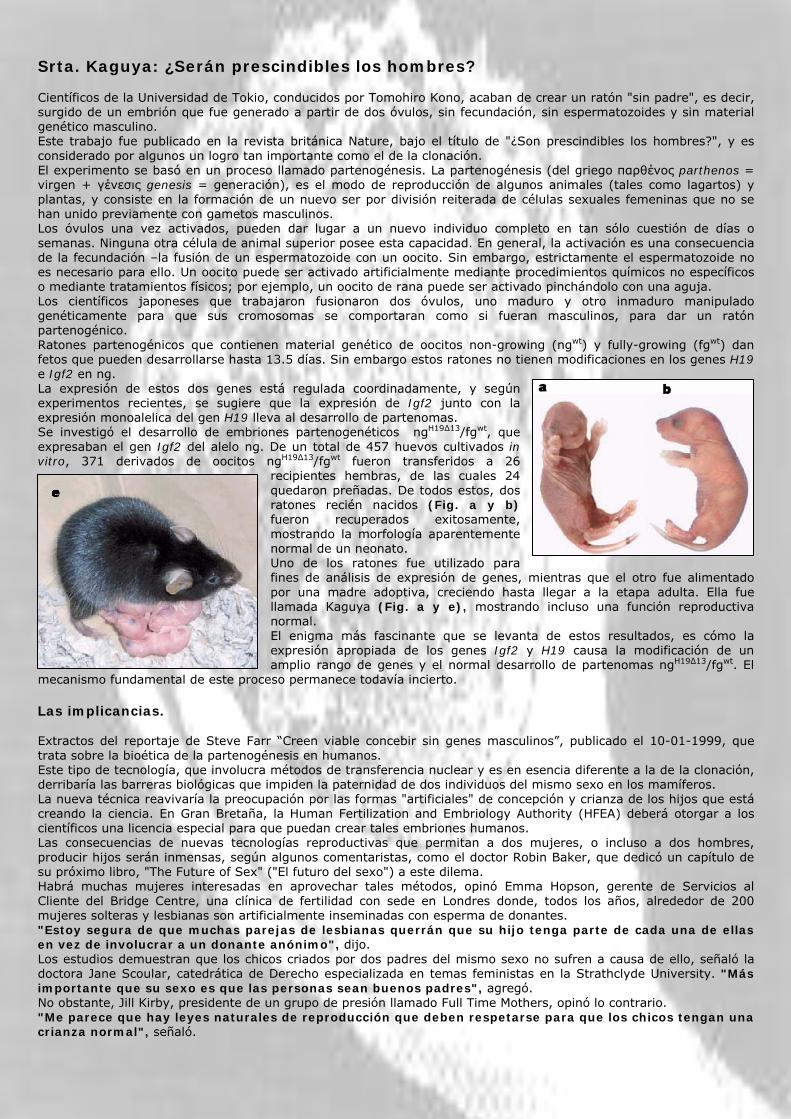
Srta. Kaguya: ¿Serán prescindibles los hombres? Científicos de la Universidad de Tokio, conducidos por Tomohiro Kono, acaban de crear un ratón "sin padre", es decir, surgido de un embrión que fue generado a partir de dos óvulos, sin fecundación, sin espermatozoides y sin material genético masculino. Este trabajo fue publicado en la revista británica Nature, bajo el título de "¿Son prescindibles los hombres?", y es considerado por algunos un logro tan importante como el de la clonación. El experimento se basó en un proceso llamado partenogénesis. La partenogénesis (del griego παρθένος parthenos = virgen + γένεσις genesis = generación), es el modo de reproducción de algunos animales (tales como lagartos) y plantas, y consiste en la formación de un nuevo ser por división reiterada de células sexuales femeninas que no se han unido previamente con gametos masculinos. Los óvulos una vez activados, pueden dar lugar a un nuevo individuo completo en tan sólo cuestión de días o semanas. Ninguna otra célula de animal superior posee esta capacidad. En general, la activación es una consecuencia de la fecundación –la fusión de un espermatozoide con un oocito. Sin embargo, estrictamente el espermatozoide no es necesario para ello. Un oocito puede ser activado artificialmente mediante procedimientos químicos no específicos o mediante tratamientos físicos; por ejemplo, un oocito de rana puede ser activado pinchándolo con una aguja. Los científicos japoneses que trabajaron fusionaron dos óvulos, uno maduro y otro inmaduro manipulado genéticamente para que sus cromosomas se comportaran como si fueran masculinos, para dar un ratón partenogénico. Ratones partenogénicos que contienen material genético de oocitos non-growing (ngwt) y fully-growing (fgwt) dan fetos que pueden desarrollarse hasta 13.5 días. Sin embargo estos ratones no tienen modificaciones en los genes H19 e Igf2 en ng. La expresión de estos dos genes está regulada coordinadamente, y según experimentos recientes, se sugiere que la expresión de Igf2 junto con la expresión monoalelica del gen H19 lleva al desarrollo de partenomas. Se investigó el desarrollo de embriones partenogenéticos ngH19∆13/fgwt, que expresaban el gen Igf2 del alelo ng. De un total de 457 huevos cultivados in vitro, 371 derivados de oocitos ngH19∆13/fgwt fueron transferidos a 26
recipientes hembras, de las cuales 24 quedaron preñadas. De todos estos, dos ratones recién nacidos (Fig. a y b) fueron recuperados exitosamente, mostrando la morfología aparentemente normal de un neonato. Uno de los ratones fue utilizado para fines de análisis de expresión de genes, mientras que el otro fue alimentado por una madre adoptiva, creciendo hasta llegar a la etapa adulta. Ella fue llamada Kaguya (Fig. a y e), mostrando incluso una función reproductiva normal. El enigma más fascinante que se levanta de estos resultados, es cómo la expresión apropiada de los genes Igf2 y H19 causa la modificación de un amplio rango de genes y el normal desarrollo de partenomas ngH19∆13/fgwt. El
mecanismo fundamental de este proceso permanece todavía incierto. Las implicancias. Extractos del reportaje de Steve Farr “Creen viable concebir sin genes masculinos”, publicado el 10-01-1999, que trata sobre la bioética de la partenogénesis en humanos. Este tipo de tecnología, que involucra métodos de transferencia nuclear y es en esencia diferente a la de la clonación, derribaría las barreras biológicas que impiden la paternidad de dos individuos del mismo sexo en los mamíferos. La nueva técnica reavivaría la preocupación por las formas "artificiales" de concepción y crianza de los hijos que está creando la ciencia. En Gran Bretaña, la Human Fertilization and Embriology Authority (HFEA) deberá otorgar a los científicos una licencia especial para que puedan crear tales embriones humanos. Las consecuencias de nuevas tecnologías reproductivas que permitan a dos mujeres, o incluso a dos hombres, producir hijos serán inmensas, según algunos comentaristas, como el doctor Robin Baker, que dedicó un capítulo de su próximo libro, "The Future of Sex" ("El futuro del sexo") a este dilema. Habrá muchas mujeres interesadas en aprovechar tales métodos, opinó Emma Hopson, gerente de Servicios al Cliente del Bridge Centre, una clínica de fertilidad con sede en Londres donde, todos los años, alrededor de 200 mujeres solteras y lesbianas son artificialmente inseminadas con esperma de donantes. "Estoy segura de que muchas parejas de lesbianas querrán que su hijo tenga parte de cada una de ellas en vez de involucrar a un donante anónimo", dijo. Los estudios demuestran que los chicos criados por dos padres del mismo sexo no sufren a causa de ello, señaló la doctora Jane Scoular, catedrática de Derecho especializada en temas feministas en la Strathclyde University. "Más importante que su sexo es que las personas sean buenos padres", agregó. No obstante, Jill Kirby, presidente de un grupo de presión llamado Full Time Mothers, opinó lo contrario. "Me parece que hay leyes naturales de reproducción que deben respetarse para que los chicos tengan una crianza normal", señaló.

Este tipo de investigaciones, junto con la clonación y otros métodos de reproducción asistida, podría terminar produciendo chicos con anormalidades que aparecieran más tarde en la vida, advirtió el doctor Richard Nicholson, editor del Bulletin of Medical Ethics. "Parece que llegamos a una etapa en la que se permite a los investigadores hacer lo que quieren sin que nadie les pregunte por qué -observó-. Más bien, la gente debería interrogarse sobre si realmente necesitamos este tipo de conocimientos."
Mayor Información: Birth of parthenogenetic mice that can develop to adulthood. Nature, April 22, 2004; 428(6985): 860-4. http://www.lanacion.com.ar/04/04/22/sl_594803.asp http://www.lanacion.com.ar/99/01/10/g09.htm
El gen 'snail' hace resistentes a las células cancerígenas. El gen 'Snail', que actúa de forma natural y beneficiosa durante el desarrollo embrionario, puede activarse patológicamente durante la edad madura, facilitando a diversos tumores la capacidad de formar metástasis y confiriendo a sus células resistencia a la destrucción. Esta ha sido la conclusión de un trabajo, coordinado por la investigadora española Ángela Nieto, del Consejo Superior de Investigaciones Científicas (CSIC) en el Instituto Cajal, en Madrid, que se publica en el último número de la revista 'Genes & Development'. El estudio tiene implicaciones para conocer el comportamiento tanto de las células embrionarias como de las cancerígenas. Para Ángela Nieto, "abre importantes expectativas en el tratamiento del cáncer". En concreto, destaca el conocimiento que aporta sobre el proceso de malignización tumoral que resulta en la formación de metástasis. El equipo de Nieto, que lleva doce años trabajando en el aislamiento y caracterización de este tipo de genes, ya había demostrado que la principal función del 'Snail' es inducir el movimiento de las células desde su origen a otras zonas, a veces alejadas. En el año 2000 publicaron los resultados de un trabajo que mostraba que las protéinas sintetizadas por este gen estaban implicadas en conferir a las células la propiedad de moverse. Tanto durante el desarrollo embrionario como durante la progresión de los tumores en la metástasis hay células que se sueltan y se desplazan para llegar a otras partes. Dos años más tarde, al estudiar biopsias de pacientes con tumores de mama se encontró una relación entre el gen 'Snail' y los tumores con metástasis. "Cuando esta proteína se expresa, la célula adquiere la capacidad para moverse y migrar a otro sitios, una propiedad positiva durante la formación del embrión, ya que permite la construcción de nuevos tejidos, pero perjudicial para la progresión del tumor", señala Nieto. Nuevas dianas El nuevo trabajo publicado explica que el gen 'Snail', además de posibilitar el movimiento, es el factor que las mantiene vivas durante el recorrido. "Las proteínas codificadas hacen que las células sean resistentes a la muerte al mantener activas todas las cascadas de supervivencia, formadas por la activación de las kinasas, entre otras moléculas". El conocimiento de este gen, que según aclara la investigadora, no muta, sino que see activa patológicamente en los tumores para desarrollar la misma función que en las células en desarrollo, será de gran ayuda en el desarrollo de nuevas terapias. "Lo importante ahora es encontrar represores naturales o artificiales que inactiven la proteína para que las células por un lado no se desprendan y no se forme metástasis y por otro sean más sensibles a la muerte. Hay varios grupos de investigación que trabajan en ello en todo el mundo". (Genes Dev. 2004; 18:1131-143)

De cómo la función muscular regula la expresión de genes en la célula. Enrique JaimovichCentro FONDAP de Estudios Moleculares de la Célula, Instituto de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile. [email protected] El Centro de Estudios Moleculares de la Célula El Centro FONDAP de Estudios Moleculares de la Célula (CEMC, www.cmcmed.cl) tiene como objetivo central desarrollar investigación en la frontera de la ciencia en el área de Transducción de Señales, que esté fuertemente ligada a los Programas de Doctorado del área biomédica de la Universidad de Chile (Doctorados en Ciencias Biomédicas, Ciencias Médicas, Farmacología y Bioquímica) y que apoye también el entrenamiento posdoctoral. Su especial naturaleza básico-clínica, permite al Centro organizar cursos, seminarios y talleres que reúnen a científicos con un amplio espectro de intereses, proporcionando las bases para nuevas iniciativas que el Centro promueve en la Facultad de Medicina, en sus Institutos y en el entorno del campus biomédico. El Centro está abocado al estudio de uno de los temas centrales de la biología celular moderna – la transducción de señales - entendiendo como tal el conjunto de mecanismos básicos que utilizan las células para descifrar las señales que reciben de su entorno y traducirlas en respuestas específicas. Este Centro agrupa a un conjunto de investigadores y co-investigadores (más de 20 en total) que plantean preguntas de relevancia para comprender las funciones cognitivas, la generación y acción de las hormonas, la fisiología del músculo, la muerte celular, el envejecimiento y los mecanismos moleculares que subyacen el cáncer. Los temas específicos de investigación del Centro incluyen el estudio de los mecanismos de transducción de señales involucrados en la señalización celular mediada por calcio en músculo esquelético, cardiaco y neuronas (Cecilia Hidalgo, Enrique Jaimovich y Sergio Lavandero), en la generación de hormonas (Luigi Devoto) y células tumorales (Andrew Quest), y en la muerte celular provocada por estrés oxidativo (Andrés Stutzin).
Laboratorio de ADAPTACION MUSCULAR El investigador principal de este grupo se ha dedicado durante muchos años al estudio de señales rápidas en células de músculo esquelético. La mayor parte de este trabajo ha estado dedicado a la comprensión de la transformación de señales eléctricas en cambios de calcio intracelular. El primer artículo de esta serie (1976) describió la presencia de receptores de tetrodotoxina en el sistema de túbulos transversales, estableciendo la evidencia para la propagación de un potencial de acción dependiente de sodio hacia el centro de la fibra. La densidad de receptores era diferente a la de la superficie sugiriendo una composición diferente para los dos sistemas de membranas. Las propiedades diferentes de los canales sensibles a potencial del túbulo transversal fueron estudiadas durante algún tiempo y en un trabajo publicado en 1983, se encontró que los receptores de dihidropiridina (rDHP), un marcador para los canales de calcio de tipo L, estaban localizados exclusivamente
en la membrana del túbulo transversal en gran cantidad. Estos trabajos (con más de 500 citas en la literatura) generaron la pregunta sobre la función de dichos receptores, ya que las corrientes de calcio en el músculo esquelético son muy pequeñas y aparentemente no necesarias para el acoplamiento excitación-contracción. El trabajo de varios laboratorios durante la década siguiente demostró que los rDHP juegan un rol central como sensor de potencial en el modelo actual de acoplamiento excitación-contracción, asociándose a los receptores de ryanodina, canales de liberación de calcio del retículo sarcoplasmático. Entre 1984 y 1989 varios artículos demostraron la presencia en las membranas de músculo esquelético de la maquinaria metabólica necesaria para producir inositol trifosfato (IP3). El IP3 se demostró capaz de liberar calcio de compartimentos intracelulares (1990), pero no se pudo demostrar un rol para el IP3 en el acoplamiento excitación-contracción. La respuesta a este problema necesitó de estudios en células intactas y elegimos el músculo esquelético en cultivo (tanto cultivo primario como líneas celulares, algunas desarrolladas en nuestros laboratorios) como modelo. Una serie de publicaciones recientes sugieren un rol para el IP3 en la regulación del calcio nucleoplasmático, lo que gatilla la expresión temprana de genes en estas células. Es muy interesante el hecho de que los rDHPs aparecen nuevamente en un papel como sensores de potencial, ahora para el acoplamiento excitación-transcripción. El descubrimiento exitoso de este mecanismo se logró uniendo a un equipo de investigadores con diferentes capacidades expertas. La transducción de señales que conduce a la adaptación del músculo esquelético con el ejercicio ha sido abordada por un grupo de laboratorios liderado por Enrique Jaimovich; su laboratorio se especializa en la medición de señales mediadas por calcio en células musculares en cultivo y lo integran además Jorge Hidalgo en los aspectos electrofisiológicos y José Luis Liberona en aspectos bioquímicos. Los laboratorios asociados incluyen los de María Angélica Carrasco, quien montó un laboratorio de Biología Molecular para este propósito, Jeanne Powell (Smith College, Massachussets) quien incorporó la inmunocitoquímica y los procedimientos de transfección celular a nuestro laboratorio, Paul Allen (Harvard, Boston) quien nos proveyó de líneas celulares que no expresan receptores de ryanodina y Jordi Molgó (Gif-sur-Yvette, Francia), quien contribuyó a nuestros estudios en músculo adulto. Se han agregado los laboratorios de Nora Riveros en la expresión de genes musculares y Edio Maldonado en los mecanismos de transcripción. La siguiente etapa es más ambiciosa e implica el comprender los mecanismos moleculares de la regulación de la transcripción y la expresión génica en una célula excitable. Mecanismo de excitación-transcripción En el músculo esquelético, el aumento y la disminución de su uso así como el daño, son formas conocidas de adaptación. Sin embargo, los mecanismos celulares

responsables de estos cambios no se conocen. El proceso de adaptación muscular está supeditado a la actividad nerviosa y a la interacción nervio-músculo. Constituye, por lo tanto, un modelo de estudio tanto para la plasticidad como para la regeneración de células excitables. El ión calcio es un mensajero intracelular que está involucrado en procesos asociados a la regulación de genes en muchos tipos celulares. Sin embargo, dada su preponderancia en la contracción muscular, no se ha estudiado en profundidad su participación en otros procesos. Las células musculares en cultivo permiten simular muchas de las funciones presentes en el músculo adulto como son su crecimiento, diferenciación, adaptación y contracción. Nuestro grupo ha descrito que en miotubos (fibras musculares multinucleadas en cultivo) la elevación transitoria de Ca2+ intracelular producto de la despolarización de la membrana puede ser modelada por dos procesos separables de forma espacial, cinética y farmacológica. Sólo el componente temprano está asociado con la contracción. El componente tardío, que dura varios segundos, representa un tipo de señal diferente. Nos proponemos estudiar la secuencia de eventos que ocurren en la célula muscular inmediatamente después de la estimulación. Como estímulo se ha usado además de la despolarización de la membrana la exposición a varios agentes, incluyendo hormonas esteroidales y no esteroidales. Así hemos logrado identificar parte del mecanismo de transducción en la membrana plasmática. Los resultados más recientes indican la existencia de una serie de eventos que siguen a un corto tiempo de despolarización por alto potasio externo. Estos son: i). Entre 2 y 10 segundos post despolarización hay un aumento transitorio en la masa de IP3, ii). Entre 5 y 25 segundos hay un cambio transitorio lento de Ca2+ que se propaga como una onda por el miotubo, y que muestra un aumento de Ca2+ en los núcleos que parece estar asociado con receptores a IP3 presentes en la membrana nuclear, iii).Entre 30 segundos y 10 minutos hay fosforilación de quinasas activadas por mitógenos (ERK 1/2) y fosforilación del factor de transcripción CREB, y iv). Entre 10 a 15 minutos hay un aumento en el nivel de expresión de RNA mensajero de los genes tempranos c-fos, c-jun y egr-1. Una batería de experimentos con protocolos para la inhibición específica de cada uno de los pasos de estas secuencias así como el uso de líneas celulares con modificaciones en proteínas claves involucradas en la señalización por Ca2+, han permitido establecer vínculos entre los pasos secuenciales desencadenados por la despolarización. La transcripción regulada por calcio está siendo estudiada en núcleos aislados y se avanza en la identificación de los genes tardíos regulados por este mecanismo. Dado que el mismo tipo de cascadas de señales parece activarse por neurotransmisores así como por hormonas y agentes oxidantes, el conjunto de los resultados permitirá aclarar los mecanismos generales basados en estas cascadas que asocian la estimulación de la membrana plasmática con la regulación de la transcripción de genes en células musculares esqueléticas. El siguiente esquema resume parte de nuestros hallazgos, los que nos permiten sentar las bases de un mecanismo de acoplamiento entre excitación y transcripción en células musculares esqueléticas.
Temas de futuras tesis y/o unidades de
investigación 1. Mecanismo de regulación de la transcripción in
vitro por calcio e inositol trifosfato. 2. Regulación de la expresión génica por hormonas
en células musculares. Análisis de “gene arrays” 3. Rol de los sub-tipos de receptores de IP3 en la
localización de señales de calcio en células musculares
4. Señales de calcio en la unión neuromuscular, rol en la regulación génica en músculo adulto.
5. Mecanismos de translocación nuclear de la PKC y su rol en el control de la expresión génica
6. Translocación de factores de transcripción inducida por estimulación eléctrica. Estudio inmunohistoquímico.
Publicaciones recientes Araya, R., Liberona, J.L., Cárdenas, J.C., Riveros, N., Estrada, M., Powell, J.A. Carrasco, M.A. and Jaimovich, E. (2003) Dihydropyridine receptors as voltage sensors for a depolarization-evoked, IP3R-mediated, slow calcium signal in skeletal muscle cells. J. Gen Physiol 121:3-16 Carrasco, M.A., Riveros, N., Ríos, J., Müller, M., Torres, F. Pineda, J., Lantadilla, S. and Jaimovich, E. (2003). Depolarization induced slow calcium transients activate early genes in skeletal muscle cells. Am J. Physiol (Cell Physiol.) 284:C1438-C1447 Estrada M, Espinosa A, Müller M and Jaimovich E. (2003) Testosterone stimulates intracellular calcium release and mitogen-activated protein kinases in skeletal muscle cells. Endocrinology. 144: 3586-3597. Powell, JA, Molgó, J, Adams, DS, Colasante, C, Williams, A, Bohlen, M. and Jaimovich E. (2003). IP3 Receptors and Associated Ca2+ Signals Localize to Satellite Cells and to Components of the Neuromuscular Junction in Skeletal Muscle. J. Neuroscience, 23: 8185-8192. Ibarra C , Estrada M, Carrasco L, Liberona JL, Chiong M, Díaz-Araya G, Cardenas C, Jaimovich E and Lavandero S (2004) IGF-1 Induces an Inositol 1,4,5-Trisphosphate-Dependent Increase in Nuclear and Cytosolic Calcium in Cultured Rat Cardiac Myocytes J. Biol. Chem 79:7554-7565.
Eltit JM, Hidalgo J. and Jaimovich E. (2004) Slow calcium signals after tetanic electrical stimulation in skeletal myotubes. .Biophysical J., en prensa.
Ca2+
Núcleo
SR
Túbulo-TPLC
rDHP
IP3R
IP3R
Ca2+
IP3
DAG
CRE c-fos c-jun
P-CREB
Gi
PKC
Ras
raf
MEK
ERK 1/2
P-ERK 1/2
PKC
Ca2+
Núcleo
SR
Túbulo-TPLC
rDHP
IP3R
IP3R
Ca2+
IP3
DAG
CRE c-fos c-jun
P-CREB
Gi
PKC
Ras
raf
MEK
ERK 1/2
P-ERK 1/2
PKC
MATRIMONIO GERTY Y CARL CORI. “Es nuestra creencia que el arte y la ciencia pueden crecer y convertirse en una sociedad que acaricie la libertad y que demuestre el respecto por las necesidades, la felicidad y la dignidad de seres humanos. Mi esposa y yo estamos orgullosos de haber sido honrados por un país que sobresalga en todas estas calidades y estamos felices de ser huéspedes en esta ciudad hermosa y hospitalaria.”
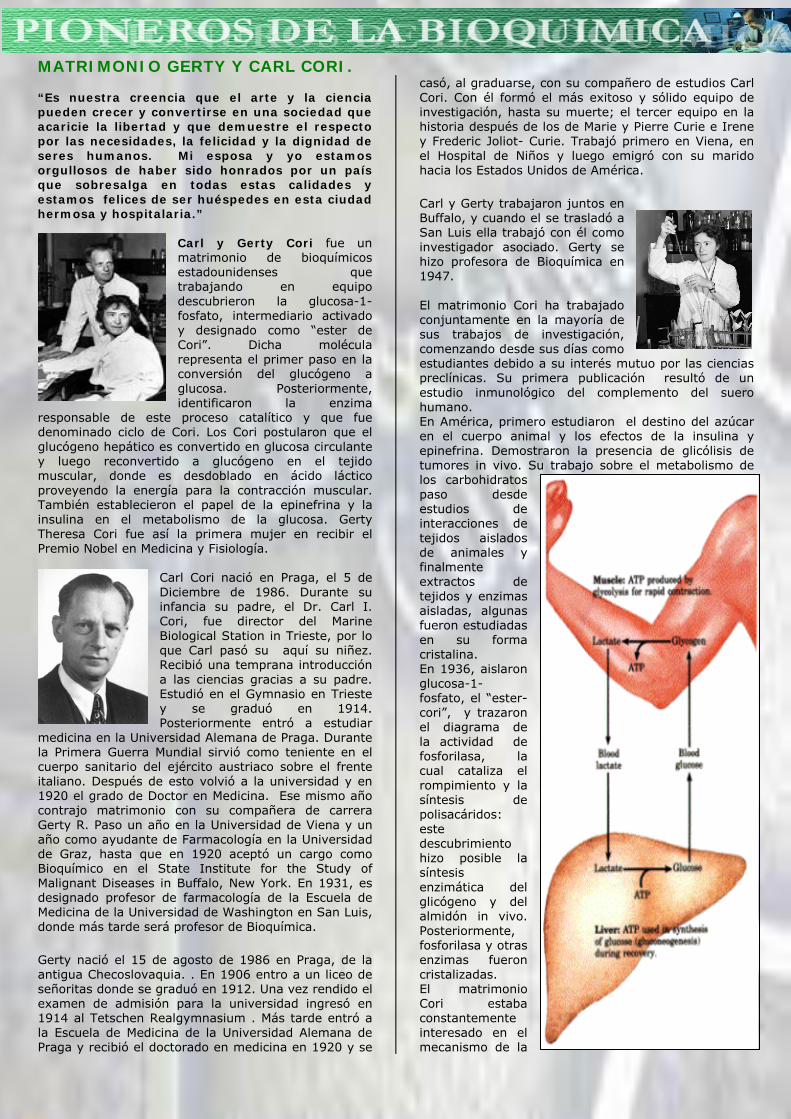
Carl y Gerty Cori fue un matrimonio de bioquímicos estadounidenses que trabajando en equipo descubrieron la glucosa-1-fosfato, intermediario activado y designado como “ester de Cori”. Dicha molécula representa el primer paso en la conversión del glucógeno a glucosa. Posteriormente, identificaron la enzima
responsable de este proceso catalítico y que fue denominado ciclo de Cori. Los Cori postularon que el glucógeno hepático es convertido en glucosa circulante y luego reconvertido a glucógeno en el tejido muscular, donde es desdoblado en ácido láctico proveyendo la energía para la contracción muscular. También establecieron el papel de la epinefrina y la insulina en el metabolismo de la glucosa. Gerty Theresa Cori fue así la primera mujer en recibir el Premio Nobel en Medicina y Fisiología.
Carl Cori nació en Praga, el 5 de Diciembre de 1986. Durante su infancia su padre, el Dr. Carl I. Cori, fue director del Marine Biological Station in Trieste, por lo que Carl pasó su aquí su niñez. Recibió una temprana introducción a las ciencias gracias a su padre. Estudió en el Gymnasio en Trieste y se graduó en 1914. Posteriormente entró a estudiar
medicina en la Universidad Alemana de Praga. Durante la Primera Guerra Mundial sirvió como teniente en el cuerpo sanitario del ejército austriaco sobre el frente italiano. Después de esto volvió a la universidad y en 1920 el grado de Doctor en Medicina. Ese mismo año contrajo matrimonio con su compañera de carrera Gerty R. Paso un año en la Universidad de Viena y un año como ayudante de Farmacología en la Universidad de Graz, hasta que en 1920 aceptó un cargo como Bioquímico en el State Institute for the Study of Malignant Diseases in Buffalo, New York. En 1931, es designado profesor de farmacología de la Escuela de Medicina de la Universidad de Washington en San Luis, donde más tarde será profesor de Bioquímica.
Gerty nació el 15 de agosto de 1986 en Praga, de la antigua Checoslovaquia. . En 1906 entro a un liceo de señoritas donde se graduó en 1912. Una vez rendido el examen de admisión para la universidad ingresó en 1914 al Tetschen Realgymnasium . Más tarde entró a la Escuela de Medicina de la Universidad Alemana de Praga y recibió el doctorado en medicina en 1920 y se
casó, al graduarse, con su compañero de estudios Carl Cori. Con él formó el más exitoso y sólido equipo de investigación, hasta su muerte; el tercer equipo en la historia después de los de Marie y Pierre Curie e Irene y Frederic Joliot- Curie. Trabajó primero en Viena, en el Hospital de Niños y luego emigró con su marido hacia los Estados Unidos de América.
Carl y Gerty trabajaron juntos en Buffalo, y cuando el se trasladó a San Luis ella trabajó con él como investigador asociado. Gerty se hizo profesora de Bioquímica en 1947. El matrimonio Cori ha trabajado conjuntamente en la mayoría de sus trabajos de investigación, comenzando desde sus días como estudiantes debido a su interés mutuo por las ciencias preclínicas. Su primera publicación resultó de un estudio inmunológico del complemento del suero humano. En América, primero estudiaron el destino del azúcar en el cuerpo animal y los efectos de la insulina y epinefrina. Demostraron la presencia de glicólisis de tumores in vivo. Su trabajo sobre el metabolismo de los carbohidratos paso desde estudios de interacciones de tejidos aislados de animales y finalmente extractos de tejidos y enzimas aisladas, algunas fueron estudiadas en su forma cristalina. En 1936, aislaron glucosa-1-fosfato, el “ester-cori”, y trazaron el diagrama de la actividad de fosforilasa, la cual cataliza el rompimiento y la síntesis de polisacáridos: este descubrimiento hizo posible la síntesis enzimática del glicógeno y del almidón in vivo. Posteriormente, fosforilasa y otras enzimas fueron cristalizadas. El matrimonio Cori estaba constantemente interesado en el mecanismo de la

acción de las hormonas y habían realizado varios estudios sobre la pituitaria. Observaron que la marcada disminución en glicógeno y la baja de azúcar en la sangre en ratas hipofisectomisadas ocurrían con un incremento concominante en el índice de oxidación de la glucosa. Posteriormente, por un estudio de la acción de las hormonas sobre hexokinasa, ellos observaron que algunos extractos pituitarios inhiben a esta enzima in vivo e in vitro y que la insulina contrarresta esta inhibición. Además, de su propio trabajo altamente original, el matrimonio Cori siempre ha sido una fuente de inspiración para sus colegas en los distintos centros activos de investigación bioquímica que han dirigido. Ellos han contribuido en muchos artículos para The Journal of Biological Chemestry y otros periódicos científicos.
Carl Cori y Gerty Cori fueron miembros de la American Society of Biological Chemistry, de la National Academy of Sciences, de la American Society y de la American Philosophical Society. Ellos fueron
presentados conjuntamente con el Midwest Award (American Chemical Society) en 1946 y
al Squibb Award en Endocrinologia en 1947. Además Gerty Cori recibió el Garvan Medal en 1948, el St. Louis Award en 1948 y fue nombrada Doctor honorario de Ciencias para la Universidad de Boston (1948), Smith College (1949), Yale (1951), Colombia (1954), y Rochester (1955). Carl Cori, miembro del Royal Society (Londres) y de la American Association for the Advancement of Science, también recibió la Medalla Willard Gibbs en 1948, el Sugar Research Foundation Award (1947,1950) y Doctor honorario de Ciencias para Western Reserve University (1946), Yale (1946), Boston (1948) y Cambridge (1949). Fue Presidente del IV Congreso Internacional de Bioquímica. (Viena, 1958). Carl y Gerty se nacionalizaron norteamericanos en 1928. El 26 de Octubre de 1957 muere la Dra Gerty Cori y en 1960, Carl contrae matrimonio por segunda vez con Anne Fitz-Gerald Jones. “Nuestra colaboración comenzó hace 30 años cuando seguíamos siendo estudiantes de medicina en la universidad de Praga y ha continuado desde entonces. Nuestros esfuerzos han sido en gran parte complementarios, y el uno sin el otro no habría logrado conseguir lo que logramos en combinación.. Mientras aun muy jóvenes tuvimos la fortuna de ir a los Estados Unidos. Nuestro país adoptado nos ha tratado con extrema generosidad y ha sido de gran importancia para nuestro desarrollo científico y nuestra perspectiva el vida.”
Su único hijo, Tom Cori nació cuando ella tenía cuarenta años. Las expectativas de sus padres pesaron en la decisión del muchacho, que de adolescente era apasionado por el béisbol, pero luego se inclinó por la Química y pasó a presidir una empresa de productos químicos. Los Cori desarrollaron el fundamento de cómo se alimentan las células y transforman la energía. El concepto es ahora parte del estudio básico de Ciencias, por lo que es difícil verlo como el descubrimiento revolucionario de su época, en 1920. Estudiaron el papel de los azúcares en el cuerpo animal y los efectos de la insulina y adrenalina. Su trabajo con el metabolismo de carbohidratos pasó de estudios de animales completos al aislamiento de tejidos, y luego a extractos de tejido y encimas. Fueron pioneros en la investigación de enzimas, las proteínas que permiten a las células funcionar, crecer y reproducirse. Sus estudios de hormonas contribuyeron a la comprensión del papel de la pituitaria y de desórdenes metabólicos, como la diabetes. Además de su intenso trabajo, los Cori fueron una fuente de inspiración y empuje para sus colegas en los centros de investigación bioquímica que dirigieron. El cuerpo de profesores del Carolina Institute decidió entregar el Premio Nobel de 1947 en Fisiología y Medicina de manera compartida. Una parte fue otorgada al Matrimonio Cori por el descubrimiento de el curso de la conversión catalítica de glucógeno, y la otra parte fue dada al Profesor Bernardo Houssay. Ellos han revelado las reacciones enzimáticas entre glucosa y glucógeno, y han mostrado que estas reacciones son controladas por factores fisiológicos.

ENFERMEDAD DE ALZHEIMER: SEGUNDA PARTE Mecanismos patogénicos. La hipótesis del amiloide. El estudio de las anomalías anatómicas presentes en la enfermedad de Alzheimer ha permitido discernir los mecanismos patogénicos de la enfermedad. Las placas seniles están formadas por una proteína fundamental (Ab) de 40-42 aminoácidos, siendo la enfermedad de Alzheimer una amiloidosis. Por amiloide se entiende el acúmulo de diferentes proteínas, que tienen en común su estructura bplegada, que le confiere una afinidad especial por el colorante rojo Congo, en las preparaciones histológicas. El material amiloide se puede depositar de forma generalizada por todo el organismo (amilodosis sistémicas, como la amiloidosis secundaria a infecciones crónicas) y amiloidosis confinadas a un determinado órgano (amiloidosis localizadas, como la angiopatía amiloide cerebral, en la que hay acúmulo de material amiloide en la capa media de las arterias cerebrales y leptomeníngeas). En todas las amiloidosis existe una proteina fundamental, que en el caso del Alzheimer es la proteina Ab. En 1984 Glenner y Wong aislaron y secuenciaron la proteína fundamental de las placas seniles, una proteína de 40 a 42 aminoácidos que llamaron b proteína, y que más tarde pasó a denominarse A4, por su tamaño de 4 Kilodalton, o finalmente Ab o beta amiloide. Una vez conocida su secuencia, se pudo construir una sonda de ADN, que se hibridó con su complementario en el genoma, encontrándose que la proteína Ab, era solo una parte de una proteína de mayor tamaño, llamada proteína precursora de amiloide, de la que se deriva mediante proteolisis. La APP, se codifica por un gen localizado en el cromosoma 21, formado por 18 exones, existiendo 7 isoformas, que se producen por procesamientos alternativos de estos exones. Estas isoformas tienen un tamaño de 365, 563, 695, 714,751 y 770 aminoácidos. De todas ellas parece que las isoformas 751 y 770 son las que más importancia tienen en la enfermedad de Alzheimer, dado que el resto o no contienen la secuencia Ab y por lo tanto no pueden generar placas seniles o no se expresan en el cerebro. No se conoce con exactitud cual es la función de la APP in vivo, habiéndose descrito diferentes funciones como inhibidor de proteasas, lo que hace que cuando se ramifique la neurona, las proteasas, que inicialmente abren el camino a esta ramificación cuando son inhibidas se permite la formación de sinapsis estables. A nivel extracerebral la función inhibidora de proteasas interviene en la formación de los coágulos
sanguíneos. Otras posibles funciones descritas de la proteína precursora de amiloide son la función como receptor asociado a la proteína G, un papel neurotrófico y de estímulo de crecimiento celular en medio de cultivo. La APP es un proteína transmembrana, localizada especialmente en las prolongaciones de las neuronas. Su degradación se produce "segregándose" parte de la APP al medio extracelular, por lo que se ha denominado a las proteinas que la degradan "secretasas". La principal vía de rotura de la APP es a través de la a secretasa, que rompe la APP en la zona de peptido Ab, segregándose por tanto restos de APP que no se agregan y por tanto no amiloidogénicos. Existen otras dos secretasas (b secretasa y g secretasa) que rompen la APP en los extremos del peptido Ab, segregándose restos de APP amiloidogénicos. Por tanto, el estímulo de
la vía a secretasa, evita la formación de depósitos beta amiloides, mientras que la inhibición de esta vía metabólica o el estímulo de la vía de la b y g secretasa origina deposito Ab. Una vez formado el depósito de amiloide, este depósito parece que ejerce su toxicidad por varios mecanismos, desde aumento de los procesos de oxidación de las membranas a estímulo glutaminérgico y entrada de calcio en la célula.
Las hormonas esteroideas como el 17b-estradiol y la dihydroepiandrostendiona (DHEA) también parecen regular el metabolismo de la APP en células en cultivo. Posiblemente, a través del aumento de la actividad de la proteína kinasa C que inducen los estrógenos, estos compuestos podrían reducir el depósito de Ab. De otra parte, la proteína tau, constituyente de los ovillos neurofibrilares, es una proteína codificada por un gen localizado en el cromosoma 17, formado por 13 exones, existiendo 6 isoformas. Su función es la estabilización de los microtúbulos que forman el citoesqueleto de la neurona. Durante años, se ha discutido cual de las dos alteraciones, las placas seniles o la degeneracion neurofibrilar, era la lesión fundamental en la enfermedad de Alzheimer y cual era la lesión secundaria. Sin embargo, los hallazgos de los últimos años, fundamentalmente de la genética del Alzheimer y de las demencias frontales, parecen indicar que es la placa senil la alteración primaria en estos pacientes.
Hoy en día se piensa que en la Enfermedad de Alzheimer, la alteración fundamental está en el procesamiento de la APP (teoría amiloidogénica o de la cascada de amiloide), siendo la degeneración neurofibrilar un fenómeno secundario. Neurotransmisión. El sistema colinérgico en la Enfermedad de Alzheimer. La pérdida de memoria es la alteración más precoz en el Alzheimer. La base biológica de esto es que los circuitos neuronales implicados en el aprendizaje y memoria son los más precoz e intensamente lesionados. Dos circuitos neurales involucrados en las diversas facetas de la memoria, son los que presentan la principal afectación: la conexión del hipocampo y el cortex entorinal con el resto del cortex y la del sistema colinérgico del cerebro basal anterior con el cortex. De los múltiples sistemas de neurotransmisores implicados en la fisiopatología de la enfermedad de Alzheimer, el mejor caracterizado es el
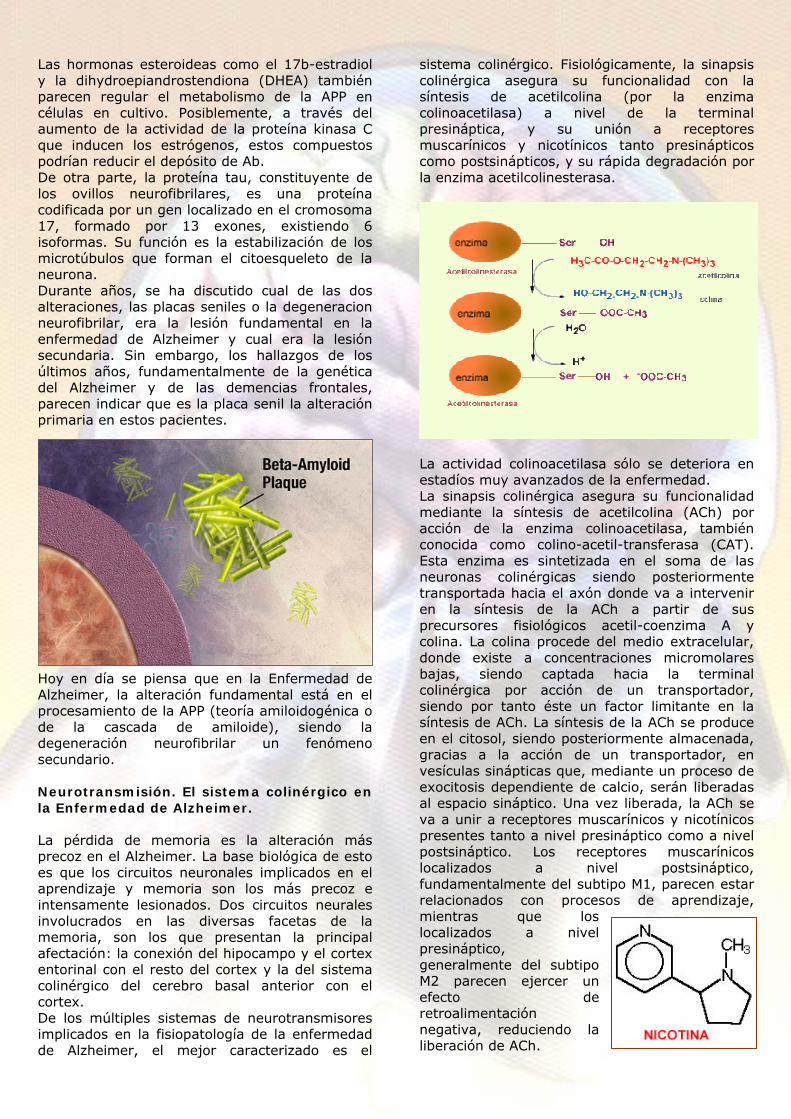
sistema colinérgico. Fisiológicamente, la sinapsis colinérgica asegura su funcionalidad con la síntesis de acetilcolina (por la enzima colinoacetilasa) a nivel de la terminal presináptica, y su unión a receptores muscarínicos y nicotínicos tanto presinápticos como postsinápticos, y su rápida degradación por la enzima acetilcolinesterasa.
La actividad colinoacetilasa sólo se deteriora en estadíos muy avanzados de la enfermedad. La sinapsis colinérgica asegura su funcionalidad mediante la síntesis de acetilcolina (ACh) por acción de la enzima colinoacetilasa, también conocida como colino-acetil-transferasa (CAT). Esta enzima es sintetizada en el soma de las neuronas colinérgicas siendo posteriormente transportada hacia el axón donde va a intervenir en la síntesis de la ACh a partir de sus precursores fisiológicos acetil-coenzima A y colina. La colina procede del medio extracelular, donde existe a concentraciones micromolares bajas, siendo captada hacia la terminal colinérgica por acción de un transportador, siendo por tanto éste un factor limitante en la síntesis de ACh. La síntesis de la ACh se produce en el citosol, siendo posteriormente almacenada, gracias a la acción de un transportador, en vesículas sinápticas que, mediante un proceso de exocitosis dependiente de calcio, serán liberadas al espacio sináptico. Una vez liberada, la ACh se va a unir a receptores muscarínicos y nicotínicos presentes tanto a nivel presináptico como a nivel postsináptico. Los receptores muscarínicos localizados a nivel postsináptico, fundamentalmente del subtipo M1, parecen estar relacionados con procesos de aprendizaje, mientras que los localizados a nivel presináptico, generalmente del subtipo M2 parecen ejercer un efecto de retroalimentación negativa, reduciendo la liberación de ACh.
Con respecto a los receptores nicotínicos, existe una amplia variedad de ellos que ejercen múltiples funciones a nivel del Sistema Nervioso Central (SNC), si bien en términos generales puede decirse que los receptores nicotínicos presinápticos ejercen un papel modulador de la liberación de neurotransmisores, mientras que los postsinápticos median procesos de transmisión sináptica excitatoria.
Los receptores muscarínicos parecen estar disminuidos y se sabe que los
bloqueantes muscarínicos (p. ej.
escopolamina) provocan un deterioro
de la cognición y, por tanto un agonista muscarínico podría resultar útil para mejorar el aprendizaje en estos pacientes. En particular, estudios in vitro sobre material de autopsias e in vivo usando tomografía de emisión de positrones han demostrado una disminución temprana de los niveles de receptores nicotínicos de acetilcolina que contienen subunidades α4, α3, y α7.Tiene especial interés la observación de que sólo los niveles de RNA mensajero de la subunidad α7 se encuentran alterados, lo que induce a pensar en una asociación particular entre los receptores que contienen esta subunidad y la patología. Por consiguiente, el desarrollo de ligandos selectivos por subtipos de receptores nicotínicos es un desafío importante, tanto desde el punto de vista de una mayor comprensión de la enfermedad en sí como de su eventual tratamiento. Así, a los datos epidemiológicos que apuntan a un efecto neuroprotector de la nicotina se suman estudios que demuestran que la neuroprotección in vitro por nicotina frente al daño inducido por Aβ es bloqueada por un antagonista de receptores α7. Entre los agonistas nicotínicos naturales se cuentan, además de la nicotina, la epibatidina, que es el veneno de una ranita de la selva ecuatoriana y la citisina, alcaloide característico de muchas plantas de la familia de las leguminosas. En los últimos años se ha investigado un número considerable de agonistas nicotínicos sintéticos. Estos compuestos, sin embargo, mantienen un patrón común a los agonistas naturales: tienen alta afinidad por los receptores que contienen subunidades α4 y afinidad mucho menor por los que contienen subunidades α7. Al postularse la "teoría colinérgica del Alzheimer" (Bartus et al., 1982) se pensó que, de forma similar a lo que ocurría en otras enfermedades neurodegenerativas como la enfermedad de Parkinson. Sin embargo, los primeros intentos de tratamiento de pacientes de Alzheimer con precursores colinérgicos (principalmente colina y
lecitina) no resultaron satisfactorios, no pudiéndose confirmar la utilidad clínica de estos agentes mediante ensayos clínicos controlados. "Teoría colinérgica del Alzheimer”:
1- Diferentes estudios postmortem de unión de [3H]-nicotina a membranas y rodajas cerebrales (autorradiografía), estudios de imagen con tomografía de emisión de positrones (PET) utilizando [11C]-nicotina y estudios inmunohistoquímicos utilizando anticuerpos selectivos, muestran una reducción en la densidad de receptores nicotínicos en diferentes áreas cerebrales. (Perry et al., 1995; Nordberg et al., 1993, 1995; Shiver et al., 1999; Martín-Ruiz et al., 1999; Wevers et al., 1999; Guan et al., 2000; Burghaus et al., 2000). 2- Se ha observado una disminución del número de placas necróticas en cerebros de fumadores, en comparación con los de no fumadores (Ulrich et al., 1997), si bien no está claramente establecido si el hábito de fumar puede ser considerado o no como un factor de protección frente al desarrollo de la enfermedad, encontrándose tanto datos que sugieren una correlación negativa entre el hábito de fumar y el desarrollo de la enfermedad (Brenner et al., 1993), como estudios que sugieren la existencia de una correlación positiva entre estos dos factores (Shalat et al., 1987). 3- Por otro lado, en pacientes fumadores existe una mayor densidad de receptores nicotínicos cerebrales, al mismo tiempo que se observa una menor neurodegeneración (Nordberg et al., 1995). Esta acción "neuroprotectora" de la nicotina se relacionaría con la capacidad de ésta de inducir la síntesis de factores de crecimiento nervioso o de reducir la citotoxicidad para glutamato; recientemente se ha descrito que este papel neuroprotector de los agentes nicotínicos estaría mediado por los receptores del subtipo α7 (Donelly-Roberts et al., 1996; Kaneko et al., 1997; Shimohama et al., 1998)

probablemente debido a la mayor permeabilidad al catión Ca2+ de este subtipo receptorial. 4- La neurotransmisión colinérgica nicotínica está también implicada en los procesos de aprendizaje y memoria y así se ha podido observar que la administración de agonistas nicotínicos mejoran el aprendizaje en modelos animales (Summers y Giacobini, 1995), así como una mejoría de las funciones cognitivas en pacientes con enfermedad de Alzheimer a los que se administró nicotina por vía sistémica (Newhouse et al., 1988; Shahakian et al., 1989), mientras que la administración de un antagonista de los receptores nicotínicos como la mecamilamina puede empeorar estas funciones en sujetos sanos (Newhouse et al., 1994). Respecto a otros neurotransmisores, el principal neurotransmisor implicado en el circuito hipocampo cortical, es el glutamato. En la enfermedad de Alzheimer se han descrito alteraciones en los niveles de glutamato. Galantamina para la enfermedad del Alzheimer.
La galantamina ((4aS,6R,8aS)-6-
hidroxi-3-metoxi-11-metil-5,6,9,10,11,12-
hexahidro-4aH-benzofuro [3a,3,2-e,f][2] benzazepina) es un alcaloide
(sustancia nitrogenada natural
derivada de plantas) con un grupo nitrógeno terciario que tiene una actividad anticolinesterásica. Fue aislado por primera vez en 1952 del bulbo del Galanthus woronowii. Actualmente, la galantamina se obtiene mediante síntesis de laboratorio. La galantamina se absorbe rápidamente en el tubo digestivo tras la administración oral y la concentración plasmática máxima se alcanza tras 1,2 horas. La biodisponibilidad oral absoluta es elevada (88,5-100%). Presenta una baja unión a proteínas plasmáticas (17,7%) a concentraciones terapéuticas. La vida media de la galantamina es de unas 7-8 horas, lo que permite ser administrado dos veces al día. Su novedoso mecanismo de doble acción sobre el sistema colinérgico central, inhibiendo la degradación de acetilcolina y potenciando la acción de ésta sobre los receptores nicotínicos, hacen de ella un fármaco con propiedades farmacologicas interesantes. La modulación de receptores y su capacidad para modificar las respuestas intracelulares abren un nuevo campo de investigación cuyos resultados preliminares son muy prometedores para comenzar a evaluar en la clínica un efecto
modificador del curso de la enfermedad. El comportamiento cinético y perfil de seguridad de galantamina en humanos hacen que sea cómodo su uso en una población anciana. Síndrome de Down. Virtualmente, todas las personas con síndrome de Down presentan en sus cerebros a la edad de 35-40 años lesiones características de Alzheimer. La beta-secretasa BACE2 (pepsina de la familia de las aspartil proteasas que forma amiloide beta a partir de la proteina precursora) se codifica en el cromosoma 21, extracopiado en el síndrome de Down, lo que levanta la posibilidad que tal proteasa contribuya a la aparición del fenotipo patológico del Alzheimer en los pacientes downianos por producir en los mismos más amiloide beta desde su nacimiento. Diagnóstico de la Enfermedad de Alzheimer
Actualmente la única manera definitiva para diagnosticar la enfermedad de Alzheimer es investigar sobre la existencia de placas y ovillos en el tejido cerebral. Pero para observar el tejido cerebral se debe esperar hasta que se haga la autopsia (examen del cuerpo que se hace después que la persona muere). Por consiguiente, los médicos deben hacer un diagnóstico de "posible" o "probable" enfermedad de Alzheimer en vida del paciente.
La corriente actual es que la Enfermedad de Alzheimer no es un diagnóstico de exclusión, sino que existe una capacidad para realizar este diagnóstico con seguridad en base a criterios de inclusión y exclusión. Los criterios NINCDS-ADRDA (siglas correspondientes a la comisión creada para generar los criterios de diagnóstico formada por el National Institute of Neurological, Communicative Disorders and Stroke y la Alzheimer's Disease and Related Disorders Association) que provee tres niveles de seguridad diagnóstica: Enfermedad de Alzheimer definida, cuando existe un cuadro clínico apropiado de demencia y confirmación autópsica o mediante biopsia, al que recientemente se ha añadido que
la existencia de una mutación en el gen de la APP o las presenilinas; Enfermedad de Alzheimer probable, que es el diagnóstico que se realiza habitualmente cuando hay una historia clínica característica y se
excluyen otros procesos con la exploración y los datos de laboratorio y Enfermedad de Alzheimer posible cuando la demencia presenta datos de atipicidad para la Enfermedad de Alzheimer, como
puede ser la alteración precoz en el habla o si existe otro proceso demenciante, aunque se

piense que la Enfermedad de Alzheimer es la causa principal de la demencia. En la práctica clínica diaria estos criterios únicamente se utilizan como referencia.
Las pruebas de imagen para el diagnóstico de la demencia senil son la resonancia magnética cerebral, la tomografia axial computarizada y las técnicas de medicina nuclear. En pacientes con Enfermedad de Alzheimer se detecta mediante resonancia magnética la existencia de atrofia cortical y subcortical en mayor o menor grado, que se manifiesta por aumento de tamaño de los surcos corticales, espacio subaracnoideo y sistema ventricular. Pero esta exploración es de alto costo y exige una gran colaboración del enfermo. Por ello, la principal prueba de imagen en el estudio del Alzheimer es actualmente la tomografía axial computarizada. Esta prueba tiene un costo menor y un tiempo de realización más bajo, lo que se adapta mejor al paciente con demencia. Además existen dos pruebas isotópicas aplicables al estudio de esta enfermedad: la tomografía de emisión de fotón único (SPECT) y la tomografía por emisión de positrones (PET). La PET permite evaluar los procesos bioquímicos y fisiológicos cerebrales de los pacientes con enfermedad de Alzheimer. La PET tiene mayor resolución y sensibilidad diagnóstica. Es una tecnica de exploracion por imagen basada en el metabolismo celular. Con ella pueden detectarse Enfermedad de Alzheimer en las primeras fases. Sin embargo, la corta vida media del isótopo utilizado hace que sea una prueba cara y disponible en pocos sitios.
Marcadores biológicos de la Enfermedad de Alzheimer
El diagnóstico de la Enfermedad de Alzheimer mediante criterios clínicos, aunque tiene una alta precisión, tiene un cierto grado de incertidumbre, que ha llevado a la búsqueda de marcadores biológicos (ver cuadro de texto) que sean capaces de aumentar esta precisión hasta hacer innecesarios los estudios neurohistológicos Los marcadores biológicos actualmente disponibles para el diagnóstico de la Enfermedad de Alzheimer incluyen los test genéticos, los estudios de proteínas del plasma y finalmente la determinación de péptidos en el LCR. Los test genéticos son los estudios de mutaciones de la APP y Ps, que restringidos a aquellas familias con Enfermedad de Alzheimer hereditaria, deben realizarse por su alto valor predictivo y la determinación del genotipo APOE que dado la mayor presencia del genotipo e4 en las formas de Enfermedad de Alzheimer esporádica puede usarse para el diagnóstico de la Enfermedad de Alzheimer en la población general. Los
marcadores biológicos de Enfermedad de Alzheimer del LCR se basan en el estudio de los niveles de proteínas implicadas en el mecanismo de la enfermedad. Los niveles de Ab42 en LCR son más bajos que los de los controles, disminuyendo aún más a medida que progresa la enfermedad, lo que se pone en relación con el depósito progresivo del péptido en el cerebro. Por el contrario, existe un aumento de los niveles de proteína tau en LCR en pacientes con Enfermedad de Alzheimer. La sensibilidad de estos marcadores varía entre un 60-90 por ciento y un 70-96 por ciento respectivamente. Otro marcador de esta enfermedad en el líquido cefalorraquídeo es la AD7C-NTP, una proteína transmembrana con propiedades de receptor cuyo cDNA se ha clonado recientemente. Su patrón de expresión es predominantemente neuronal y su expresión está aumentada en la Enfermedad de Alzheimer.
Más cerca de aclarar la causa del Alzheimer
Acaba de publicarse un trabajo que puede tener mucha trascendencia en el definitivo descubrimiento de la causa última de la enfermedad de Alzheimer y, en consecuencia, el hallazgo de un tratamiento curativo de la misma. Varios investigadores de la Feinberg School of Medicine de la Northwestern University, de Chicago, recurrieron a las técnicas de ingeniería genética, tan generalizadas hoy en biomedicina y sobre todo en neurociencias, para producir un modelo de Alzheimer en ratones lo que, evidentemente, no es lo ideal para explicar la enfermedad humana pero sí resulta una aproximación científicamente válida. Estos ratones, a medida que envejecen, van acumulando amiloide ß en sus cerebros lo mismo que ocurre en los pacientes con Alzheimer. Lo importante del trabajo que comento es que los ratones modelo de Alzheimer fueron manipulados genéticamente para que no expresaran ß-secretasa. Deprivados de esta enzima, estos animales no produjeron amiloide ß, no perdieron capacidades de aprendizaje y sus sinapsis funcionaron con normalidad a pesar de ir haciéndose viejas. Lo bueno además es que no apareció efecto adverso o indeseable de tipo alguno. Con lo cual hay firmes esperanzas de que pueda iniciarse pronto el ensayo clínico correspondiente en enfermos con productos inhibidores de la ß-secretasa. Cuadro de texto
Test en investigación (marcadores biológicos de la Enfermedad de Alzheimer)
En sangre: niveles de Ab42, proteína p97 y genotipo APOE
En LCR: niveles de Ab40 y Ab42, tau, ADC7-NTP Resonancia magnética espectroscópica

INTRODUCCIÓN Los términos " northern " y " western" derivan de " southern" (observen la diferencia) según lo inventado por E. M. Southern en los años 70. En una southern blot, los fragmentos del DNA primero son separados por tamaño usando electroforesis de gel. El DNA se corta generalmente con endonucleasas de restricción para crear los fragmentos de tamaños definidos. Los endonucleasas de restricción cortan solamente el DNA en sitios específicos, así tienen capacidad de crear fragmentos de DNA de longitud definida. Una muestra del DNA cortado se coloca en un extremo de un " gel " hecho de agarosa. Cuando se aplica un campo eléctrico, el DNA negativamente cargado emigra a través del gel hacia el lado positivo del aparato en electroforesis. Los segmentos de DNA más pequeños tienen más facilidad de moverse a través de la matriz de agarosa. De este modo las moléculas del DNA se pueden separar por tamaño. Después de electroforetizar, el gel se quita del aparato, y del DNA es transferido sobre una membrana. Membranas de nitrocelulosa fueron utilizadas originalmente, pero no se comportan de forma ideal; membranas de nylon se utilizan en vez de nitrocelulosa. La membrana, con los pedazos asociados del DNA, pueden ahora estar * rastreados * con una secuencia específica del DNA, la punta de prueba reconoce una secuencia única del DNA que atará a un sitio específico en la membrana. Las puntas de prueba se etiquetan generalmente de una cierta manera, a menudo con los isótopos radiactivos (32P o 35S, e.g.).
La membrana bloteada puede después ser colocada debajo de la película y ser desarrollada. Las bandas oscuras aparecerán donde están las secuencias radio marcadas del DNA pegadas a la membrana. "Northern blot" y " western " siguen el mismo procedimiento que la southern blot – fraccione por tamaño, transfiera a una membrana, después sondee - pero utilizan diversos materiales distintos. Los nombres fueron utilizados en " honor direccional " de southern blot " (¿quién dice que la ciencia no puede incitar la diversión en sí misma?).
En un northern blot las moléculas del RNA son identificadas. En un western, las proteínas son de tamaño fraccionado en un gel hecho de la acrilamida, transferido a una membrana, después sondeado comúnmente con los anticuerpos que deben reconocer proteínas específicas. WESTERN BLOTTING El análisis western blot puede detectar una proteína en una mezcla de cualquier número de proteínas. No importa si la proteína se haya sintetizado in vivo o in vitro. Este método es, sin embargo, dependiente en el uso de un anticuerpo de alta calidad dirigido contra una proteína deseada. Se debe poder producir por lo menos una porción pequeña de la proteína de un fragmento reproducido del DNA. Se utilizará este anticuerpo como punta de prueba para detectar la proteína del interés. El western blot indica cuánto se ha acumulado de la proteína en tales células. Si se está interesado en el índice de la síntesis de una proteína, la técnica Radio-Immune precipitation (RIP) puede ser un mejor análisis. También, si una proteína se degrada rápidamente, western blotting no la detectará bien; necesitará utilizar (RIP).

PROTOCOLO 1. Separe las proteínas usando electroforesis de gel de SDS-poliacrilamida (también conocido como SDS-PAGE). Esto separa las proteínas por tamaño. 2. Coloque una membrana de nitrocelulosa en el gel y, con electroforesis, conduzca las bandas de la proteína (polipéptido) sobre la membrana de nitrocelulosa. La carga negativa tendrá que estar en el lado del gel y la carga positiva en el lado de la membrana de nitrocelulosa para conducir las proteínas cargadas negativamente encima de la membrana cargada positivamente de nitrocelulosa. Esto le da una membrana de nitrocelulosa que se imprima con las mismas bandas de la proteína en gel. Hay que pasar con cierto ensayo y error para encontrar el pH óptimo, también se necesita estar seguro de que no haya burbujas de aire entre la nitrocelulosa y el gel o las proteínas no se transferirán. 3. Incube la membrana de nitrocelulosa con un anticuerpo primario. El anticuerpo primario, que es el anticuerpo específico mencionado arriba, se pega a su proteína y forma un complejo anticuerpo-proteína con la proteína de interés. 4. Incube la membrana de nitrocelulosa con un anticuerpo secundario. Este anticuerpo debe ser una conjugación del anticuerpo-enzima. El anticuerpo secundario debe ser un anticuerpo contra el anticuerpo primario. La enzima conjugada permitirá que se visualice el complejo anticuerpo-proteína. 5. Para ver realmente la enzima en acción, se necesitará incubarla en una mezcla de reacción que sea específica para su enzima. Si todo es trabajado correctamente, se ven bandas dondequiera que haya un complejo anticuerpo-enzima, o, es decir dondequiera que este la proteína. 6. Ponga la película de radiografía en el gel para detectar un flash de luz, que es emitida por la enzima. La reacción funciona generalmente dentro de una hora.
APLICACIONES: Mientras que ELISA mide el anticuerpo al virus entero y da un resultado "positivo", "negativo " o indeterminado, el western blot es una prueba más específica. Permite que uno visualice los anticuerpos dirigidos contra cada proteína viral. Por esta razón, es una prueba confirmativa para un ELISA positivo de VIH. El VIH, como cualquier otro virus, se compone de un número de diversas proteínas. El carril positivo del control del western blot contiene las proteínas de los sueros de los pacientes así como las proteínas del VIH. La positividad del VIH se puede por lo tanto confirmar solamente por la presencia de los tipos siguientes de proteínas:
gp160 viral envelope precursor (env)
gp120 viral envelope protein (env) binds to CD4
p24 viral core protein (gag)
p31 Reverse Transcriptase (pol)
Una proteína específica puede ser identificada y visualizada en un 'Western blot' empleando técnicas de inmunodetección. Estas técnicas emplean la capacidad de reconocimiento de los anticuerpos a sus antígenos. En 'Western blot' se emplea comunmente el sistema de inmunodetección indirecta.

ROMANCES DE UN SUSTRATO
Divagaba un sustrato en un mar de solución salina ubicada en el extenso citoplasma de una célula, cuando conoció a una enzima de formidable simpatía y espléndida conformación tridimensional, cuyos contornos se insinuaban en una estructura terciaria que produjo mas que un llamado de atención sobre el solitario sustrato. Enseguida, como si sus interacciones hubiesen sido de todo un tiempo de vida media, notaron que la atracción era demasiado fuerte y complementaria. Estaban hechos el uno para el otro. La enzima, llevada por un impulso totalmente espontáneo, abrazó al sustrato rodeándolo completamente a la vez que excluía de su corazón activo toda molécula que no tuviese las propiedades de su nuevo amor, el sustrato. Ahora, las moléculas de agua pasaban a ser meras espectadoras. Las fuerzas que les mantenían juntos hicieron que comenzara la más grande de las experiencias a una velocidad catalítica. Inmediatamente las interacciones electrostáticas convirtieron el encuentro en sensaciones vibratorias de un nivel casi cuántico del cual parecía que ninguna ley fisicoquímica les permitiría escapar. Compartieron en completitud todas sus propiedades. Cada enlace de hidrógeno fue suavemente acariciado, las interacciones de Van der Waals se presentaban sutilmente como un cosquilleo en cada átomo del sustrato, las atracciones iónicas fueron un ir y venir de electrones dispuestos a satisfacer cada necesidad de sus ansiosas electronegatividades. Nada podía ser mejor. Cada millonésima de segundo que pasaba por sus vidas medias fue plenamente disfrutado, eran la envidia de toda molécula, el sueño reactivo hecho una catalítica realidad. El clímax se acercaba avisado por los estados excitados y la placentera explosión de energía irradiada que se hizo sentir en cada una de las atónitas y expectantes moléculas de alrededor, las cuales no quedaron insensibles a tal cantidad de calor que fluía desde el interior de aquella rebosante enzima.
Todo indicaba que sería para siempre. De pronto el intercambio de electrones se detuvo, sus contornos moleculares ya no eran complementarios, las atracciones electrostáticas se habían convertido en repulsiones electrostáticas, algo había sucedido, ya nada era como antes. La acalorada enzima ya no reconocía a quién millonésimas de segundo antes hubiese sido la molécula que llenaba su sitio activo. Fue así como el sustrato, sintiendo que había sido utilizado solo con fines energéticos, era espontáneamente expulsado del interior de la enzima que millonésimas de segundo antes lo habían acogido con enlaces de magnitud culómbica. El decepcionado sustrato sentía que la experiencia lo había cambiado, sentía que sus propiedades reactivas eran diferentes de las que acostumbraba tener, ya no era el mismo de antes. Tratando de olvidar tan traumática experiencia, volvió a divagar por el extenso mar de aquel citoplasma que lo vio transformarse. Repentinamente, cuando ya olvidaba a aquella
enzima traicionera que se había aprovechado de su ∆G, notó que otra enzima, de una conformación
estructural diferente, lo observaba con ansiosa reactividad. Deseando no tener ningún tipo de relación química, el sustrato quiso ignorar los ofrecimientos de esa impúdica catalizadora. Todas son iguales,
decía. Pero para su sorpresa comenzaba a sentir nuevamente atracciones electrostáticas que al parecer eran más fuertes que las de su predecesora experiencia. No lo pudo evitar, enseguida, como si sus interacciones hubiesen sido de todo un tiempo de vida media, notaron que la atracción era demasiado fuerte y complementaria...
Pedro Valencia Araya.
Bioquímico.
Pedro Valencia Araya

No morirá la flor de la palabra
Podrá morir el rostro oculto de quien la nombra hoy Pero la palabra de la que vino desde el fondo de la historia y
la tierra Ya no podrá ser arrancada por
la soberbia del poder Nosotros nacimos de la noche,
En ella vivimos, moriremos en ella. Pero la luz será mañana para los demás,
Para todos aquellos que hoy lloran la noche, Para quienes se niega el día,
Para quienes es regalo la muerte, Para quienes esta prohibida la vida. Para todos la luz, para todos todo Para nosotros la alegre rebeldía,
Para nosotros nada…
Leonardo Parra. Promoción 2001. Bioquímica.
Bioquímica. ¿Por qué Bioquímica?, es la pregunta que a todos en un principio nos hicieron; tal vez porque siempre les ha gustado, o por asuntos de el puntaje, otros porque no sabían a que postular, en fin muchos casos hay del porque de Bioquímica. Hoy comparto con personas que vienen de demasiado lejos, por ejemplo, desde Arica, Puerto Montt, etc.; caso que es el mío también. Por esto el viajar a nuestras casas de repente se hace imposible, cabe solo esperar los fines de semanas largos que por cierto no son muchos. Pero para nosotros que recién estamos empezando, ¿será realmente lo que esperamos ser? Un punto importante es que nos guste la carrera, ¿pero que pasa si nos olvidamos de ella?... porque hay muchos factores que influyen en esto: están los amigos, el pololeo, los carretes, el vivir solos, el “estar libres”……para la mayoría es bacán quizás… ¿y para los otros? Seremos capaces todos de enfrentar un cambio tan radical o cuantos quedaran en el camino. ¿Será realmente Bioquímica?
Anónimo. Primer Año 2004.
Lunes 2 y Martes 3 de Agosto: Periodo de Matriculas 2º Semestre. Miércoles 4 de Agosto: Inicio clases 2º Semestre Viernes 6 de Agosto: Claustro Pleno. Viernes 13 de Agosto: Comienza el 2º Ciclo de Seminarios en Biotecnologia y Biomedicina. La primera expositora Invitada es la Dr. Claudia Ortiz, Bioquímica USACH Hora: 11:45 am Quinto Centenario. ENTRADA LIBERADA Jueves 19 de Agosto: Tocata Bioquímica , el objetivo es rreunir fondos para aquelos alumnos de nuestra carrera que iran al Congreso de Estudiantes de Bioquímica 2004 en Valdivia. Lugar : Cafeta CC Organiza Ana Maria Bravo Sepúlveda.

Hemos llegado a una etapa final de nuestra carrera en la cual debemos aplicar muchos conocimientos que nos han entregado, lamentablemente nos hemos encontrado con la gran dificultad de que la universidad no tiene lugares de Práctica designado y se debe ir solicitando en varios
hospitales si aceptan alumnos en Practica sin el convenio establecido; del cual la universidad obligatoriamente lo debe gestionar y no realizar las diligencias nosotros como estudiantes. Como consecuencia ya no se tiene la posibilidad de realizar la práctica en el Hospital Naval, tampoco en el Gustavo Fricke y para que nombrar el resto, que están regidos por la zona Valparaíso-San Antonio o Viña del Mar-Quillota, en la cual exigen tal trámite... Me siento desilusionada de esta situación, varios de nosotros realizamos la Practica este verano del 2004 en donde por buena disposición y gentileza no se complicaron por el convenio pero a la siguiente promoción les pedirán nuevamente tal gestión. Solo estoy pidiendo que por favor se nos tome en cuenta y nos puedan dar el valor que merecemos, pedir innovaciones para nuestros laboratorios y docentes es lo que necesitamos , no se trata de desvalorar a un profesor para exigir experiencia de laboratorio o de trabajo sino que es invertir en nuestra formación como profesionales, al igual que lo realizan nuestras familias y por que no decirlo varios de nosotros que trabajamos para atenuar los gastos que implica a la familia tener un hijo universitario..... No solo quiero sentir el interés de mi universidad para que me llame por teléfono y no olvide el pago de la mensualidad al final de un semestre siendo en mi caso un usuario responsable; sino que nuestra universidad nos de la infraestructura necesaria, los conocimientos, experiencia de tal manera que al salir a enfrentar una practica profesional, estemos mejor preparados. Se necesita un cambio de mentalidad de ambas partes con el fin de mejorar nuestra carrera y las relaciones y dejar de sentir que nos tilden de "poco comprometidos o conflictivos porque tenemos necesidades”. Por ejemplo este año han ingresado a la universidad varias personas discapacitadas donde solo el hecho de entrar a la casa central ya es un trauma, ya que no esta habilitada una bajada especial para el ingreso de sillas de ruedas o personas con muletas o no videntes, como también el respeto y solidaridad que podríamos tener cada uno de nosotros cuando
un estudiante minusválido cruza la calle para asistir a otro día de clases sintiendo que todo es una lucha contra el tiempo que ni siquiera el semáforo espera… también tienen la capacidad de estudio igual que todos, son necesidades básicas de cualquier ser humano digno de respeto .El hecho de que puedan desempeñarse como universitarios ya nos dan la lección mas grande del mundo; ya que nos demuestran valentía, tenacidad y ganas de vivir. Estas necesidades que se ven y son latentes, involucran a todos los estudiantes de la Universidad. Solo los invito a que podamos ser protagonistas de nuestro cambio y que nuestras peticiones sean atendidas dignamente. Quiero ser parte de la Universidad, y sentir que fue tomada en cuenta mi opinión y que no solo nos traten como un trámite, para eso están los bancos, entidades públicas, etc. Solo es un llamado a la conciencia de cada persona que ocupa una función, trabajo en la universidad, nosotros somos el futuro, no es pedir un cambio rotundo pero si volver a intentarlo.
Claudia Pino. Promoción 1998. Bioquímica.
______________________
Un tipo de estructura de ADN se asocia a linfoma no Hodgkin
Investigadores de la Universidad del Sur de California descubrieron en los cromosomas de linfocitos una estructura poco usual de ADN que crea en ellos "sitios frágiles" y predispone a las células que los incorporan a desarrollar un linfoma no Hodgkin.
La revista Nature publicó poco tiempo atrás este estudio que "asocia por primera vez una enfermedad a una desviación de la hélice Watson-Crick", según ha indicado el coordinador del trabajo, Michael Lieber. El sitio frágil en cuestión se localiza en el gen Bcl-2 situado en el cromosoma 18. El Bcl-2 suele intervenir en la inhibición de la apoptosis, pero cuando está sobreexpresado provoca un linfoma de células B denominado linfoma folicular.
(Nature 2004; 428: 88-93).

Introducción. Búsquedas por similitud. La comparación de secuencias homologas ha servido (y sirve) para establecer relaciones filogenéticas entre diferentes individuos, o entre secuencias de un mismo individuo. Estas son una fuente de información mucho mas completa que los caracteres homólogos anatómicos usados clásicamente con el mismo fin, pero que presentan mas problemas a la hora de seleccionar datos útiles. A medida que los métodos lo han ido permitiendo, se han analizado cada vez más mayor número de secuencias homólogas a modo de compararlas, típicamente diseñando sondas poco específicas para detectar estar secuencias, y después obteniendo datos tales como sitios de restricción o más tarde (con el abaratamiento de las técnicas de secuenciación directa) datos de secuencias completas. Pero los ya no tan recientes proyectos genoma de diferentes organismos han producido una gran cantidad de datos de secuencias, muchos de ellos por analizar. Es por ello por lo que se han creado sistemas de búsqueda por similitud, los cuales rastrean las secuencias almacenadas en busca de fragmentos con una cierta similitud (según el umbral que le indiquemos) a un fragmento que previamente hemos introducido. Los dos programas de estas características más usados son FASTA y BLAST. El primero de ellos está casi en desuso mientras que el segundo de ellos es un sistema muy extendido y es el objeto del presente texto. Estos sistemas de búsqueda no solo proporcionan datos para analizar la evolución de las secuencias, sino que al alinear las mismas y observar los dominios conservados podemos extrapolar funciones de proteínas hasta ahora desconocidas por comparación con dominios de función ya conocida. Muchos dominios proteicos se han caracterizado por similitud entre secuencias, y nunca se han descrito físicamente ni se conoce su función real, solo se sabe que están muy conservados entre secuencias (por ejemplo el motivo WCCH del elemento Pyret, en Nakayashiki, 2001) El programa BLAST. Algoritmos. El programa BLAST (Basic Local Alignment Search Tool) es un programa interactivo disponible vía web desarrollado por Altschul y colaboradores (1990) y actualmente se encuentra en prácticamente cualquier base de datos. No es necesario hacer más comentarios acerca de su historia ya que se escaparía de la línea de esta guía. Solo añadir que este programa es de dominio público y cualquiera puede acceder y hacer uso de él. Al ser usado en un trabajo publicado, la referencia a usar es la siguiente: ALTSCHUL, S. F., T. L. MADDEN, A. A. SCHÄFFER, J. ZHANG, Z. ZHANG, W. MILLER and D. J. LIPMAN (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acid Research 25(17): 3389-3402. En resumen, el sistema se basa en un algoritmo de búsqueda heurístico (busca secuencias significativamente similares a la que introducimos, usando potentes métodos estadísticos), el cual ha sido mejorado con el tiempo y actualmente permite el alineamiento con huecos (gaps) lo que permite hacer análisis más precisos. Como todo método estadístico, BLAST también tiene valores de significación, en este caso basados en el algoritmo de Karlin y Altschul (1990, 1993). Estos valores se discutirán mas adelante desde un aspecto práctico, no matemático. Ya que el desarrollo matemático del método no es objeto de esta guía, a partir de ahora me limitaré a comentar los aspectos estrictamente prácticos. Si desea profundizar en la estructura del método, puede recurrir a los artículos originales (véase apartado de referencias) o las numerosas guías que están disponibles en la red. Búsqueda con BLAST. Formatos y opciones. Ahora veremos como usar los BLAST que se encuentran en el NCBI (National Center for Biotechnology Information), pero todos los demás BLAST disponibles tienen un funcionamiento similar. Así que lo primero es ir al directorio blast del NCBI; http://www.ncbi.nlm.nih.com/blast Una vez allí nos encontraremos con una lista de diferentes programas blast, aquí comentaremos los más interesantes.
BLASTP: Compara una secuencia de aminoácidos contra bases de datos de proteínas. Otra utilidad es que al dar el resultado, el programa nos dice si ha encontrado en la proteína que le damos dominios que previamente han sido bien caracterizados.
BLASTN: Compara secuencias de nucleótidos contra bases de datos de nucleótidos. La principal utilidad es encontrar secuencias no traducidas a proteína, como por ejemplo rRNAs, secuencias reguladoras o simplemente DNA “basura”.
BLASTX: Compara las 6 posibles pautas de lectura (3 en cada cadena) de una secuencia nucleotídica que le demos con bases de datos de proteínas. La utilidad reside en que podemos encontrar cual es el sentido de traducción de una secuencia de nucleótidos, si este codifica para una proteína conocida o similar. Con este método podemos saltar codones de STOP (y con un poco de maña frameshifts) que truncarían la secuencia en otros métodos de búsqueda de ORFs como el ORF finder del NCBI.
TBLASTN: Compara una proteína que le demos con 6 pautas de lectura (que se traducen dinámicamente) de bases de datos de nucleótidos. Puede servir para rastrear un genoma en busca de homólogos de una proteína que acabamos de caracterizar. Muy útil por ejemplo en el estudio de evolución de elementos móviles (véase por ejemplo Marín y Llorens,2000).

TBLASTX: Compara las 6 pautas posibles de lectura de la secuencia de nucleótidos que le demos con las 6 pautas de lectura de las secuencias de las bases de datos de nucleótidos. Nunca la he utilizado en mi trabajo pero es una herramienta de detección muy potente.
PSI-BLAST (Position Specific Iterative BLAST): Es una herramienta muy potente a la hora de detectar funciones bioquímicas para proteínas desconocidas. Es sistema se basa en realizar una primera búsqueda, y después de alinear el resultado el programa calcula una matriz en la cual se da un valor a cada posición y aminoácido (PSI-BLAST es para buscar proteínas), esta matriz es la matriz PSSM (position specific scoring matrix), y es la que usa el programa para realizar una segunda búsqueda. Tras esta búsqueda calcula una nueva PSSM y realiza una nueva búsqueda, así hasta que nos interese. En esta guía no comentaré más acerca de PSI-BLAST, pero si está interesado puede leer una guía en NCBI (en http://www.ncbi.nlm.nih.gov/Education/BLASTinfo/psi1.html). PHI-BLAST es un sistema incluido en PSI-BLAST relacionado con la búsqueda de motivos, pero este no está aun muy desarrollado.
BLAST 2: Sequences. Usa cualquier tipo de BLAST para buscar similitudes entre dos secuencias. Este programa es muy útil a la hora de determinar la estructura de una secuencia, por ejemplo, para ver como se distribuyen las diferentes ORF en elementos móviles. Si navegan por la página BLAST del NCBI encontrará mas tipos de BLAST, pero suelen ser variaciones de los anteriores, y no merece la pena que los introduzca, aunque conforme adquieran soltura deberían probar trabajar con todos ellos, en un futuro podrían serles útiles y ahorrarán mucho tiempo si ya los conoce.
COMO REALIZAR UNA BUSQUEDA:
Introducir la secuencia problema en el campo search, o bien pegar desde otro archivo.Admite texto lineal y formato FASTA.
En el campo siguiente puedes elegir el tipo de BLAST, pero se supone que ya lo elegiste anteriormente.
En Set Subsequence indicar (si es necesario) desde que nucleótido (o aminoácido) a cuál de la secuencia introducida se realizará la búsqueda. Generalmente no se rellena ya que se suele introducir únicamente la secuencia que se quiere analizar.
El campo Choose database puede ser útil. Por defecto está marcado nr, es decir, la base de datos no redundante. Otras opciones posibles son bases de datos de ciertos organismos, de secuencias Alu, de datos estructurales de proteínas... pero la que quizás pueda importar más en principio a parte de nr, es la base de datos month, en la cual se encuentran las secuencias de reciente incorporación a GeneBank (durante el último mes).
El campo Genetic Codes, obviamente se refiere a que código genético quieres que use el programa (recuerda que TBLASTX trabaja con secuencias traducidas).
El botón BLAST! Inicia la búsqueda, pero abajo puedes modificas más opciones.
Opciones de BLAST
Limit entry by query admite restricciones a la hora de buscar.
Del resto de opciones, solo cabe destacar la matriz de sustitución, que por defecto es una BLOSUM62. En estudios muy finos de evolución molecular puede interesar cambiar la matriz, pero generalmente se ha convertido en una cuestión de preferencia personal según el investigador.

Formato de Salida
Las opciones del formato no alteran el análisis, sino el formato en que BLAST nos presenta los resultados. El único campo que nos puede interesar cambiar (al menos mientras nos familiarizamos con el programa) es el número de secuencias que queramos que nos muestre.
En ocasiones, para búsquedas finas nos puede interesar tener un mayor número de secuencias y de alineamientos, o tan solo mayor número de descripciones (conforme más use este programa, más partido le sacará a esta opción).
Una vez pulse el botón BLAST!, aparecerá de nuevo una pantalla con las opciones de formato, pero además aparecerán dos cosas nuevas, un botón de FORMAT! y un número de entrada (ID).
Al hacer click sobre el botón FORMAT! El programa nos dará el resultado de la búsqueda.
La pantalla también nos da el tiempo estimado que tardará en hacer la búsqueda. Si el tiempo es muy elevado, una opción es copiar el número de acceso que da (ID) y después recuperar el
resultado desde la dirección de BLAST del NCBI (http://www.ncbi.nlm.nih.com/blast) y en el link Retrieve results with a Request ID.
Obtención de Resultados.
En la pantalla de resultados aparecen al principio datos informativos acerca de la base de datos que se ha usado, la referencia bibliográfica a usar, el tamaño de la secuencia problema, y un gráfico que indica la distribución de las secuencias encontradas. El gráfico es mas que nada para dar una idea aproximada y sigue un código de colores según la similitud con de la
secuencia encontrada con la secuencia problema. Lo que nos interesa aparece a continuación del gráfico. Lo primero que se observa son las descripciones. Estas son simplemente el nombre y número de serie de las secuencias encontradas con similitud, seguidas de dos valores, el Store y el E-Value que se comentarán mas adelante en este punto. Desde la línea de descripciones podemos acceder directamente a la secuencia encontrada. El nombre de la secuencia tiene una serie de partes se pasara a describir: Identificador GI: Es un identificador asignado por el NCBI, de modo que cada secuencia tiene un gi único e irrepetible. La estructura es gi|gi_identifier, como por ejemplo: Identificador de base de datos: Indica de que base de datos ha sido obtenida la secuencia, seguido de un número de serie o el nombre de la secuencia, incluso puede figurar el locus, dependiendo de la base de datos como se muestra en la siguiente tabla. Después de las descripciones aparecen los alineamientos de las secuencias encontradas con la secuencia que hemos introducido. El alineamiento soporta huecos (gaps) y compara las secuencias según la matriz que hayamos elegido en el apartado de opciones de búsqueda. En cada alineamiento podemos encontrar los siguientes elementos:

Los dos valores que nos interesan para determinar si dos secuencias detectadas como “homólogas” por BLAST son significativamente similares, son el Score y el E-Value. Score: Este valor indica que grado de similitud existe entre las dos secuencias comparadas. A mayor valor mayor similitud. De todas formas no es un valor tan útil como el E-Value. E-Value: Indica la probabilidad de encontrar al azar en el genoma una secuencia igual a la encontrada, es decir, la probabilidad de que las dos secuencias comparadas no sean homólogas. Obviamente nos interesan valores muy bajos, por lo que se suele prefijar un umbral a partir del cual no tendremos en cuenta las secuencias encontradas. Generalmente valores superiores a 10-3 no son muy fiables, aunque depende del tipo de análisis. Estos valores pueden usarse como criba para determinar que secuencias consideramos homólogas y cuales no. De nuevo es una cuestión arbitraria y lo estricto que se sea depende del tipo de trabajo que se esté realizando. Consideraciones finales. Si nuestro objetivo era buscar concretamente una secuencia, basta con seleccionarla y copiarla en un archivo para trabajarla posteriormente, pero si nuestro trabajo era conseguir un grupo de secuencias similares (por ejemplo para análisis filogenético o de dominios conservados) deberemos hacer nuestra base de datos de secuencias. Esto es una parte un tanto pesada ya que debemos coger las secuencias que hayamos seleccionado, copiarlas, y pegarlas en un nuevo archivo, generalmente en formato FASTA (ya que es el más sencillo). Por si al tiempo de leer esta guía no está familiarizado con el formato FASTA, al costado derecho del texto se muestra la estructura típica de este; Los caracteres que acepta este formato son los mismos que acepta BLAST para realizar búsquedas, y son los que siguen (dependiendo de si trabajamos con aminoácidos o con nucleótidos);

A lo largo de esta guía le he mostrado como obtener resultados con BLAST, pero esto es solo el primer paso. Tras tener nuestra base de datos, hemos de analizar las secuencias con otros programas (que se escapan de este manual). Habrá observado que la mayoría de ejemplos se basan en TBLAST (aminoácidos) ya que es el programa con el que más he trabajado, pero el resto de programas BLAST tienen un funcionamiento similar, lo único que puedo decirle es que practique con todos ellos e irá adquiriendo cada vez más soltura. No hace falta decir que esto es un pequeño manual, y que BLAST tiene multitud más de opciones y posibilidades, pero esta guía tiene un carácter meramente introductorio. En el apéndice 2 se dan algunas direcciones donde se pueden conseguir guías más completas acerca de este poderoso sistema de búsqueda. Apéndice 1. Búsqueda de mRNAs La búsqueda de mRNAs puede realizarse desde BLAST, introduciendo una restricción en el campo Limit Entrez by Query del menu Options. El comando de restricción es; biomol_mrna[prop] Las secuencias que aparecen en el resultado pueden llevar o no información del tejido en el que se expresa, depende del autor. Otro comando interesante es el contrario, es decir, a partir de mRNA buscar la secuencia genómica. Para ello usaremos la restricción; biomol_genomic[prop] BIBLIOGRAFIA www.ncbi.nlm.nih.gov/Education/BLASTinfo/guide.html www.ncbi.nlm.nih.gov/BLAST www.ch.embnet.org/software/bBLAST.HTML www.igbmc.u-strasbg.fr:8080/DbClustal/dbclustal.html www.dna.affrc.go.jp/htdocs/Blats/blast.html
DEBIDO A LA COMPLEJIDAD DE CREAR ESTA SECCIÓN VA SER SUPER IMPORTANTE TU APORTE, YA QUE MUCHOS ALUMNOS NO MANEJAN NINGUN CONCEPTO COMPUTACIONAL, Y PARA ALGUNOS ALUMNOS AUN ES DIFICIL MANEJAR ALGUNOS CONCEPTOS BIOLOGICOS, FISICOS, ESTADISTICOS, QUIMICOS Y MATEMATICOS, QUE SON DE GRAN IMPRTANCIA PARA PODER ENTENDER ESTA CIENCIA YA QUE COMO HEMOS VISTO PARA PODER APLICARLA DE LA FORMA CORRECTA ES IMPORTANTE SABER UN POCO DE CADA CIENCIA. ES POR ESTO QUE NOSOTROS NOS DIRIJIMOS A USTEDES PIDIENDOLES LO SIGUIENTE:
1) CUALQUIER DUDA O CONSULTA DE ALGUN TERMINO EN ESPECIAL PUEDES ENVIARNOS UN MAIL Y EN LA PROXIMA REVISTA ESTA SERA CONTESTADA Y SI ES POSIBLE TU RESPUESTA SERA ENVIADA DE INMEDIATO.
2) SI QUIERES HACER ALGUNA SUGERENCIA DE ALGUN TEMA A TRATAR EN ESTA SECCION TAMBIEN LO PUEDES HACER. (YA SEA ALGUNA APLICACIÓN O ALGUNA HERRAMIENTA BIOINFORMATICA QUE NECESITES SABER USAR, SOLO ENVIANOS TU CONSULTA).
3) LA ESTRUCTURACION DE ESTA SECCIÓN PARTE CON UN ENTENDIMIENTO DE ALGUNOS CONCEPTOS BASICOS, QUE SE IRAN TRATANDO MAS ESPECIFICAMENTE A FUTURO, YA SEA CON HERRAMIENTAS BIOINFORMATICAS DISPONIBLES EN LA WEB U OTRAS QUE TU PUEDAS BAJAR A TU PC.
4) PROXIMAMENTE ESTA SECCIÓN ADEMAS SERA APOYADA POR BIBLIOGRAFIA, SI TU QUIERES APORTAR CON ALGO O CONSIDERAS QUE HAY ALGUN TERMINO QUE ESTA MAL APLICADO, SE PUEDE PONER EN DISCUSIÓN PARA QUE TODOS VAYAN APRENDIENDO Y ENTENDIENDO ESTA CIENCIA.
5) ESPERAMOS POR ULTIMO QUE PARTICIPES Y NOS AYUDES A DESARROLLAR ESTA SECCION COMO DEBE SER.

Entrevista a Patricia Ewert (Ex-alumna DE Bioquímica PUCV, actualmente estudiando bioquímica en la U. DE. CHILE) 1.- ¿Por qué te cambiaste de la Universidad Católica de Valparaíso a la Universidad de Chile? Creo que cuando me cambie me afectaron varios factores, que no necesariamente tenían que ver con el ámbito académico. Pero igual influenciaron en mí, que el cambio lo hacía a una universidad que tiene un prestigio, que las mallas hasta donde yo curse bioquímica en la UCV eran muy similares; ya que lo único que no me convalidaron fue bioinorgánica, bioquímica y analítica I. Además que no me atrasaría mas de lo que ya estaba (no me acuerdo bien si estos eran los nombres correctos de los ramos). Además que no me vine sola, lo hice con mi amiga (la Pauly), por lo tanto, eso igual influyo, por que yo creo que sola no lo hubiera hecho (ya me había afiatado a un grupo de amigos, por lo tanto igual era mas difícil). 2.- ¿Cuáles son las principales diferencias que hay en infraestructura entre las dos universidades, considerando la cantidad de alumnos que hay en cada una de ellas? Creo que hay de todo, tanto cosas buenas como malas. Te cuento que la impresión que tuve cuando recién llegue fue de guauuuuuu (jajajajaja), ya que me encontré con laboratorios mucho mas preparados en la parte física, imagínate que cada uno tenia su cajonera con su material a ocupar, etc. Pero de todas maneras hay falencias como uno biblioteca súper chica, que se comparte con los químicos farmacéuticos (120), la cantidad de computadores también es súper escasa. Lo bueno de estar acá, es que la facultad esta al frente de la de medicina, y si la biblioteca o los computadores de mi facultad estaba ocupados, tu ibas a la de medicina. En cuanto a las salas que mas se puede pedir si entran como 30. Dato: los baños de ustedes son un 7, nada más que decir. 3.- Ya que conoces las mallas académicas de ambas universidades ¿cuáles son las principales diferencias entre ambas? y ¿qué le cambiarías a la malla de Bioquímica de la PUCV? Aquí si que estamos mal, por que, te puedo responder de la malla hasta donde yo curse bioquímica por que de los ramos que seguían no me acuerdo. Pero haciendo una comparación, yo pienso que el ramo de introducción de primer año, por lo menos cuando yo lo curse, no servia para nada; acá ese ramo también se dicta pero con otro nombre, la diferencia esta en los temas a tratar, conoces los laboratorios y en que están trabajando cada uno de los profes. Parece que ustedes no tenían ingles, lo cual para alumnos que no poseen una base sólida es un gran inconveniente, ya que con todo lo que hay que leer. Encuentro que hace falta más biología en los primeros años. 4.- La carrera de Bioquímica de la Universidad de Chile, desde Enero del 2004, está acreditada por 6 años por la CNAP. ¿Cómo fue el proceso de acreditación de Bioquímica en tú universidad? (o sea, qué supiste del proceso en el momento, qué significó para ustedes. etc). Aquí el proceso de Acreditación fue súper normal, participaron todos los estamentos de la universidad, entiéndase profesores, alumnos de todos los años. La información de lo que se trataba, en que consistía, se entrego con mucha anticipación y se pudo dar una opinión muy acertada y sincera (entiéndase esto, como que no se puso nada de más ni de menos). Además sirvió para mejorar hartas cosas (como las del baño). 5.- ¿Qué es lo más destacable que, en tu opinión, tiene Bioquímica de la PUCV en comparación con la misma carrera en otras universidades? Creo que la PUCV tiene una visión mucho mas química, solo basta mirar en que están los profesores para darse cuenta. En cuanto a las relaciones humanas por lo menos hasta donde yo estuve eran mucho mejor, acá el asunto es mucho más competitivo, cada uno se las arregla como puede, y mi experiencia allá no fue así. 6.- ¿Qué características tiene el cuerpo académico de la U. de Chile? En la parte académica, que yo sepa son todos doctores, dependiendo del área que se trate. Las relaciones se van haciendo más cercanas a medida que pasan los cursos, imagínate que en cuarto y quinto año quedan tan pocos alumnos, y viéndose todos los días es casi imposible. También hay muchas clases con profesores invitados de todas las universidades (Juan Reyes es uno de ellos, en el ramo de subcelular). Te cuento que, creo haber tomado una muy buena decisión por la parte académica, ya que tu acá tienes la posibilidad de entrar en cualquier laboratorio que te interese, las tesis están abiertas tanto en la Chile, como en la Cato, incluso llaman a la U para pedir tesistas. Cosa que no veía allá; además muchos de los compañeros que tuve, los he visto acá. En cuanto a las relaciones humanas fue bastante difícil, ya que tenia grandes Amigo(a)s, y el ambiente era mucho mas relajado, para mi fue muy grato ver a la Paty y al memo, y sentir el abrazo sincero.

ALLISSON ASTUYA VILLALON.
Me presento soy Allisson Astuya Villalón, y estudie en la Cato antes de que fuera PUCV, y guardo muy buenos recuerdos de los profesores y por supuesto de mis compañeros con los que aún mantengo contacto (muchas veces gracias a ellos). Los que aún me recuerden sabrán que participe de la ANEB por largos años, he incluso nos toco organizar el mejor congreso de estudiantes que se vio por esos días. Al poco tiempo de egresar de Bioquímica, me vine a Concepción a desarrollar mi Doctorado en Ciencias Biológicas, área de especialización en Biología Celular y Molecular, y Hoy me encuentro terminando mi tesis la cual si todo sale bien defiendo a fin de año. Trabajo en el laboratorio de Neurobiología el cual dirige Francisco Nualart quien fue premiado con la distinción de Científico Joven el 2002. Trabajamos investigando la acción de vitamina C en tumores del sistema nervioso, aunque no es el único tema que se desarrolla en nuestro laboratorio.
Actualmente estamos trabajando para demostrar que la vitamina C cumple funciones importantes en tejido normal, sin embargo y este es el aporte, hemos visto que las células tumorales del sistema nervioso se transforma y disminuyen su capacidad de incorporar la vitamina C reducida y aumentan su capacidad de incorporar la forma oxidada a través de transportadores de glucosa GLUTs, lo que protegería al tumor contra tratamientos como radio y quimioterapia. Mi tesis fue respaldada por la beca de apoyo a la realización de tesis doctoral Conicyt. He participado en publicaciones tales como: 1: Medina RA, Meneses AM, Vera JC, Guzmán C, Nualart F, Astuya A, García MA, Kato S, Carvajal A, Pinto M, Owen GI. Estrogen and progesterone up-regulate glucose transporter expression in ZR-75-1 human breast cancer cells.. Endocrinology. 2003 Oct;144(10):4527-35. Epub 2003 Jun 26. 2: Castro M, Caprile T, Astuya A, Millan C, Reinicke K, Vera JC, Vasquez O, Aguayo LG, Nualart F. High-affinity sodium-vitamin C co-transporters (SVCT) expression in embryonic mouse neurons. J Neurochem. 2001 Aug;78(4):815-23. Este año fui contratada por la U de Conce para participar de los cursos de Biología Celular e Histología. Aunque llevo años impartiendo los prácticos de estos ramos y de otros como Fármacos y Bioquímica. Mis planes para un futuro próximo es volver a mi ciudad, porque pucha que se echa de menos, sin embargo, creo que antes de que esto se cumpla realizaré un postDoctorado, fuera de chile, aun estoy evaluando cuando y donde.

PERSONAJE DEL MES RICARDO HERRERA L
Nombre IUPAC : Alguien lo conoce Isotipos :Toxico, chanana, chancho. Medio : La torre, la locomotora I y II, el kamikaze, la Eskuba…
Características : Sin tollar, malo pa tomar, relajao y buena onda.
Localización subcelular: En la oficina (en cualquiera de sus sucursales, aunque es mas probable encontrarlo tomando), con alguna chanana.
Frase típica: L.Q.N.S.Vio: De vocalista con su grupo de Tagua-Tagua.
L.Q.N.S.Supo: Cuando lo quito la Toxica al Che, la pelea con el chupacabra en la Torre.
Regalo útil: Mayor virtud:


A veces tenemos que ponernos serios, pero a estos quien se las cree….
Este segundo semestre comenzamos con el segundo ciclo de seminarios, esperemos que el quinto centenario se llene, y que haya una buena ventilación.
Este es un homenaje a un compañero que ya no esta… Hay cosas que nunca vamos a olvidar….


![TEMAS DE BIOQUIMICA TEMA I: INTRODUCCION A LA BIOQUIMICA BIOQUIMICA [Tomo I].pdf · 1 TEMAS DE BIOQUIMICA TEMA I: INTRODUCCION A LA BIOQUIMICA Generalidades Concepto de materia, cuerpo,](https://img.pdfslide.es/doc/110x75/5a7707fe7f8b9a9c548dbcf5/temas-de-bioquimica-tema-i-introduccion-a-la-bioquimica-bioquimica-tomo-ipdfaa.jpg)



























