Embed Size (px)
Citation preview

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
1
RReepprroodduucccciióónn SSeelleeccttiivvaa ddee EEssppeecciieess FFoorreessttaalleess ddee RRááppiiddoo CCrreecciimmiieennttoo CCoonn éénnffaassiiss eenn eell ggéénneerroo PPooppuulluuss
por
FRANCISCO ZAMUDIO A. (PHD)
y
FERNANDO GUERRA G.
Universidad de Talca
Facultad de Ciencias Forestal
2 Norte 685, Talca. Chile Fono: 56 – 71 – 200379 Fax: 56 – 71 – 200455 / 200428
Talca, Junio de 2002
Palabras claves: Mejoramiento-genético, reproducción-selectiva, árboles, Populus

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
2
CCoonntteenniiddooss
Capítulo 1 - Introducción 4
Capítulo 2 - Generalidades del género Populus 8
Capítulo 3 - Información necesaria para desarrollar una estrategia de
mejoramiento genético 13
3.1 Objetivo 14
3.2 Variación genética 15
3.3 Biología de las especies 19
Capítulo 4 - Métodos de mejoramiento y selección 21
4.1 Selección masal 23
4.2 mejoramiento por retrocruzamientos 23
4.3 generación de variedades híbridas 24
4.4 selección recurrente 25
4.4.1 Selección recurrente simple (SRS) 27
4.4.2 Selección recurrente para capacidad combinatoria general (SR-CCG) 28
4.4.3 Selección recurrente para capacidad combinatoria específica (SR-CCE) 30
4.4.4 Selección recurrente recíproca (SRR) 32
4.5 La importancia del tiempo 33
4.5.1 Secuencia lógica y práctica en el establecimiento de ensayos de progenie 33
4.6 tipos de selección asociados a los métodos 38
4.6.1 Selección tipo masal o individual 38

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
3
4.6.2 Selección tipo familiar 38
4.6.3 Selección tipo ensayo de progenie 39
4.6.4 Selección tipo dentro de familia 40
4.6.5 Selección tipo entre familias más selección dentro de familias 40
4.7 selección para varios caracteres 41
4.7.1 Selección en tandem 41
4.7.2 Selección independiente 42
4.7.3 Indice de selección 42
Capítulo 5 - Tipos de poblaciones 44
5.1 población base 45
5.2 población reproductora 45
5.3 población de producción 46
Capítulo 6 - Estrategias de selección y reproducción para el álamo 47
6.1 reproducción y selección clonal 47
6.2 reproducción selectiva de generaciones múltiples 50
6.2.1 interacción genotipo - ambiente 51
6.3 sistemas de manejo de una población reproductiva de álamos 52
6.3.1 Sistema de reproducción de población simple 54
6.3.2 Sistema de reproducción de poblaciones múltiples 55
6.3.3 Sistema de reproducción selectiva de poblaciones replicadas 59
Capítulo 7 - Selección clonal 61
Bibliografía citada 65

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
4
CCaappííttuulloo 11
IINNTTRROODDUUCCCCIIÓÓNN
EEl mejoramiento genético de árboles forestales tiene un rol primordial en el manejo
forestal porque contribuye al aumento de la productividad, lo cual significa un
suplemento a los beneficios derivados de las prácticas silviculturales, tales como
podas y raleos, así como del mejoramiento del sitio y de la calidad ambiental.
El mejoramiento genético se realiza mediante la reproducción selectiva de
genotipos superiores. Por lo tanto, el propósito del mejoramiento genético es triple:
1. identificar individuos,
2. realizar cruzamientos entre ellos y
3. obtener progenies que generen poblaciones forestales con
características superiores a las que presentaban las poblaciones
anteriormente establecidas en el mismo sitio.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
5
Habitualmente, el incremento en productividad de árboles mejorados supera con
creces el incremento en costo involucrado en la obtención de la semilla
genéticamente mejorada.
El mejoramiento asociado a características tales como resistencia a
enfermedades y al ataque de insectos, así como algunas propiedades de la madera
(densidad de la madera, longitud de fibras y traqueidas, etc.), tiene asociado un fuerte
control genético. Esto representa una ventaja puesto que asegura una ganancia
genética sostenida en el tiempo.
La selección recurrente es el método más común utilizado en la generación de
nuevos árboles con características genéticas superiores. Una de las condiciones
básicas para su éxito es la presencia de un monto sustancial de variación genética en
los caracteres de valor económico que van a ser mejorados. Un ciclo completo de
selección, la cual es la unidad básica de cualquier sistema de selección, se muestra en
la Figura 1 (según Namkoong et al. 1988). El ciclo básico está compuesto de dos
poblaciones, base y seleccionada, y dos actividades, selección y cruzamientos. Este
ciclo se repite secuencialmente en el tiempo.
El mejorador forestal puede variar las técnicas utilizadas para la selección de
genotipos superiores, manipular el número de poblaciones involucradas en cada ciclo
y puede tratar el número de ciclos concurrentes en forma distinta.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
6
Figura 1. Modelo básico de un ciclo completo de selección. (Fuente: Namkoong et al, 1998).
Según Zobel y Talbert (1984), existen dos aspectos esenciales para que un
programa de mejoramiento genético (PMG) sea exitoso. El primero esta relacionado
con la obtención de una ganancia en cantidad y cualidad de los productos deseados
tan rápido y tan eficiente como sea posible. Esto se puede obtener a través de la
aplicación intensiva de los principios genéticos a los programas forestales
operacionales, lo cual resultará en rodales de mejor calidad, mejor adaptados, y de
mayor rendimiento. En efecto, la máxima ganancia genética puede ser lograda
mediante el uso de unos pocos de los mejores individuos (genotipos elite) para que
sirvan como padres y suministren las progenies que serán propagadas y plantadas con
fines operacionales. El segundo aspecto esta relacionado con la necesidad de largo
plazo de proveer una base genética lo más amplia posible, lo que es esencial para un
progreso continuado en el mejoramiento, a través de sucesivas generaciones.
Población
Selección Cruzamientos
Población seleccionada
Próximo ciclo

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
7
El desarrollo de estrategias de reproducción selectiva (mejoramiento genético) de
generaciones avanzadas no es una ciencia exacta. Una estrategia es dinámica y debe
evolucionar y cambiar con el tiempo (White et al 1993). Como fue mencionado por
Shelbourne et al (1986):
Una estrategia de mejoramiento genético es un plan de batalla que permite
asegurar ganancias genéticas cercanas al óptimo, en el corto y largo plazo,
enfrente de muchas incertidumbres. Debido a que la intuición y el juicio
subjetivo tienen un rol no menor en ello, el desarrollo de la estrategia es
correctamente visto como un arte.
En este documento se discuten una serie de aspectos relacionados con el
diseño y desarrollo de estrategias de mejoramiento genético forestal, con un énfasis
en la generación de variedades de álamo con características de interés económico
superiores.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
8
CCaappííttuulloo 22
GGEENNEERRAALLIIDDAADDEESS DDEELL GGÉÉNNEERROO PPOOPPUULLUUSS
EEl género Populus comprende cerca de 30 especies y se distribuye ampliamente en
Europa, Norte América y Asia (FAO, 1979). Los álamos crecen naturalmente en una
diversidad de sitios desde regiones boreales a subtropicales, y desde áreas
montañosas hasta riberas de ríos. También pueden formar grandes rodales naturales,
como ocurre aún en bosques boreales y a lo largo de valles fluviales. Otras veces, los
álamos pueden formar pequeños rodales, hileras o pequeños grupos de árboles
(Zsuffa et al, 1996)
Este género tiene la capacidad de formar híbridos ínter e intraespecíficos en
forma espontanea y controlada, lo cual ha permitido la creación de un alto número de
subespecies e híbridos artificiales (Orlovic et al, 1998). Esto ha resultado en una gran
variabilidad natural que permite a las especies e híbridos de Populus crecer en una
variedad de sitios forestales. Además de su gran adaptabilidad, la cual es significativa
para el mejoramiento genético, los álamos se distinguen por su rápido crecimiento y
facilidad de propagación (Cain y Ormond, 1984)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
9
Los álamos han sido útiles al ser humano y han sido cultivados desde tiempos
históricos. Ellos pueden crecer en muchos tipos de sitios y sirven como una excelente
fuente para un amplio rango de productos obtenibles de su madera, especialmente en
zonas temperadas. Los álamos también juegan un rol significativo en la protección
ambiental, especialmente en la protección del suelo contra el efecto erosivo del viento
y del agua (Zsuffa et al, 1996). Este tipo de árbol de rápido crecimiento puede
aumentar la diversidad forestal y enriquecer el paisaje, aumentando con ello la
apariencia estética del campo en una variedad de formas.
La introducción de variedades de álamo al país se inició con la conquista
española del territorio en el siglo XVI. Desde entonces, el álamo ha estado presente
en el desarrollo arquitectónico de nuestras ciudades, principalmente a través de su uso
como un árbol ornamental. El álamo ha sido también un componente casi esencial del
paisaje de nuestros campos y áreas rurales, en donde este tipo de árbol ha sido usado
principalmente para proteger cultivos agrícolas del efecto del viento y generar sombra
al ganado.
Sólo durante el siglo XX se reconoció que el álamo puede ser una fuente
interesante de bienes de consumo en Chile. Aunque el uso principal de la madera de
álamo en nuestro país ha estado orientado a la producción de unos pocos bienes, de
los cuales destaca la elaboración de fósforos. Hoy en día, se está reconociendo que la
madera de álamo tiene el potencial de ser usada para obtener bienes que podrían ser
destinados a mercados particularmente rigurosos y cuya demanda por productos
forestales secundarios o finales de alta calidad es bastante estable en el tiempo. Aquí
nos podemos referir al mercado europeo y norte americano.
Las condiciones para que el cultivo intensivo de álamo tenga éxito se han ido
desarrollando sistemáticamente en los últimos 10 años. Existe un programa nacional
de diversificación forestal orientado a aumentar el número de especies forestales que

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
10
conformen el futuro patrimonio económico forestal del país. Además, la Comisión
Nacional del Alamo (CNA), desde su creación en 1995, ha venido desarrollando
esfuerzos sostenidos para fomentar la forestación con híbridos de álamo. Aquí es
necesario enfatizar que existen vastas áreas de la zona centro y sur de Chile (incluso
de la IV Región) que presentan condiciones climáticas, de suelo y suministro de agua
que permiten el cultivo intensivo de álamo. Además, existe la ventaja que el álamo
puede ser manejado en forma de monocultivo o asociado a una diversidad de cultivos
agrícolas, en un sistema de manejo agroforestal. Esto último podría significar una
fuente extra de ingresos a una gran cantidad de agricultores medianos y pequeños.
Dentro de la comunidad forestal en Chile existe la idea ya establecida que el
género Populus debiera constituirse en el tercer género forestal de valor económico y
estratégico para el país, después de los géneros Pinus y Eucalyptus. Para que esto se
materialice resulta fundamental que la diversidad genética del género se incremente
significativamente en nuestro país. De esta manera se garantizará el cumplimiento de
dos aspectos esenciales para el éxito en el desarrollo de un programa de mejoramiento
de toda especie forestal de rápido crecimiento. Estos son:
1. Que la probabilidad de seleccionar híbridos de un valor genético y
comercial superior sea alta y la probabilidad que ocurra daños
ecológicos y económicos asociados a una reducida base genética sea
baja.
2. Que el máximo grado de mejoramiento en la productividad asociado al
cultivo intensivo de álamo sea obtenido en un mínimo tiempo posible.
Hasta la actualidad, los híbridos de Populus introducidos en Chile han sido
generados genéticamente en otros países. Los cruzamientos realizados para obtener
los híbridos se realizaron con árboles seleccionados bajo condiciones edafoclimáticas
específicas. Es probablemente que estas condiciones ambientales no sean

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
11
exactamente iguales a las condiciones presentes en los sitios donde el cultivo de
álamo es posible en Chile, o bien donde los híbridos de álamo ya se encuentran
establecidos en el país. Además, es posible que los criterios de selección de los
individuos padres usados en la hibridación fueron orientados a lograr objetivos
productivos propios de las instituciones que generaron los híbridos. Esta introducción
de híbridos al país se ha realizado a pesar de que nuestras condiciones ambientales y
nuestros propios objetivos productivos o de manejo forestal pueden ser distintos a los
de dichas instituciones.
Según algunos expertos, las zonas aptas para introducir álamo en Chile van desde
la IV hasta la XI regiones, ambas inclusive, lo cual indiscutiblemente implica la
existencia de una amplia variación en las condiciones ambientales (clima y suelo)
presentes en aquellos sitios donde el cultivo intensivo de álamo es posible. Esto
último justifica entonces que nuestro país tenga su propio programa de hibridación
para el genero Populus, puesto que:
1. nos permitirá generar híbridos de crecimiento rápido y particularmente
adaptados a condiciones edafoclimáticas específicas de los diferentes
sitios donde se establezca álamo en el futuro, o este ya se encuentre
establecido;
2. además nos permitirá independizarnos de la "compra obligada" de
híbridos genéticamente generados en el extranjero y
3. este programa le daría a Chile la flexibilidad necesaria para poder
responder adecuadamente, mediante la generación de nuevos híbridos, a
los posibles cambios del mercado forestal, los cuales también obligarían
a un cambio en las estrategias productivas ligadas al manejo forestal del
Alamo.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
12
La Facultad de Ciencias Forestales de la Universidad de Talca está
desarrollando un programa de selección genética de híbridos de Populus con
características superiores en crecimiento y calidad de la madera. Esto nos ha
permitido la interacción con instituciones lideres en el mejoramiento genético de este
género, como son la Poplar Molecular Genetics Cooperative (PMGC), de la
University of Washington, de Estados Unidos, y el Institute National de la Recherche
Agronomique (INRA), de Francia. Uno de los resultados obtenidos de esta relación ha
sido el aprendizaje y desarrollo de protocolos para la hibridación de Populus spp.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
13
CCaappííttuulloo 33
IINNFFOORRMMAACCIIÓÓNN NNEECCEESSAARRIIAA PPAARRAA DDEESSAARRRROOLLLLAARR UUNNAA EESSTTRRAATTÉÉGGIIAA DDEE MMEEJJOORRAAMMIIEENNTTOO GGEENNÉÉTTIICCOO
LLa siguiente información es necesaria para desarrollar una estrategia de mejoramiento genético
forestal:
1. el objetivo del mejoramiento;
2. el grado y patrón de variación y
3. la biología de la especie

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
14
3.1 Objetivo
El propósito del mejoramiento debe ser cuidadosamente considerado y claramente
establecido puesto que influirá en los métodos de selección y reproducción utilizados
en diversas formas. Ejemplos de objetivos pueden ser:
1. mejorar la producción de madera utilizable para fines estructurales;
2. mejorar el volumen de trozos aserrables o debobinables;
3. incrementar la cantidad y calidad de la pulpa;
4. desarrollar resistencia a enfermedades y plagas; etc.
Teóricamente, las opciones disponibles son múltiples, pero en la práctica, sólo se
podrá alcanzar un progreso satisfactorio si el programa se concentra en un número
reducido de caracteres. Estos deberán ser caracteres con un valor económico
importante y que presenten el potencial de ser mejorados mediante una combinación
de selección genética y silvicultura.
El mejoramiento genético es difícil de justificar y puede ser, incluso, superfluo
si algún carácter puede ser fácilmente modificable mediante algún proceso
tecnológico alternativo.
La elección del (o los) carácter(es) puede que no sea fácil y se requerirá de un
estrecho trabajo de equipo antes de tener una clara idea del objetivo del
mejoramiento. La discusión estratégica de qué mejorar deberá incluir especialistas en
tecnología de la madera, silvicultura, economía y manejo, protección forestal y
genética.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
15
3.2 Variación genética
El grado y patrón de variación genética determina el nivel de mejoramiento que es
posible alcanzar y como puede ser logrado.
Mientras mayor sea la variación genética y mayor el control ejercido por los
genes sobre los caracteres de interés, mayor será la posibilidad de obtener un
mejoramiento de éstos.
La variabilidad genética es compleja, pero su magnitud y tipo pueden ser
determinados mediante ensayos genéticos de campo. La variación genética puede ser
dividida en los componentes aditivo y no aditivo, de manera que la varianza genética es
igual a la varianza aditiva más la varianza no aditiva. En términos más simples, la
varianza aditiva se debe al efecto acumulativo de los alelos en todos los loci1 génicos
que influencian un carácter. La varianza genética no aditiva puede ser dividida en dos
tipos, de dominancia y de efectos epistáticos. La varianza de dominancia se debe a la
interacción de alelos2 específicos en un locus, mientras que la varianza de epistasis se
debe a las interacciones entre loci. Por ahora es suficiente saber que la porción aditiva es
la que tiene valor para los programas de mejoramiento genético que se basan
principalmente en la propagación por semilla. La variación no aditiva puede ser
explotada sólo a través del uso de programas de mejoramiento más especializados que
envuelven el desarrollo de cruzamientos intra o interespecíficos, o el uso de
propagación vegetativa para la producción comercial de árboles.
La mayor parte de las características de valor económico en árboles forestales
están bajo algún grado de control genético aditivo. Características tales como la
densidad básica, la rectitud de fuste y otras rasgos de calidad en árboles tienen un
1 Loci: (plural de locus) son las distintas posiciones en uno o varios cromosomas en las cuales se encuentran los genes que controlan un carácter en particular. Los genes están formados por secuencias de ADN que codifican para la expresión del carácter.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
16
componente de varianza aditiva mayor que las características de crecimiento (Burdon
y Low 1992). Aunque estos últimos son controlados hasta cierto grado por efectos
genéticos aditivos, ellos tienen una considerable varianza no aditiva asociada.
Por lo tanto, todo programa de selección y reproducción debe incluir el ensayo
de progenies de fenotipos seleccionados para determinar el valor genético real
del árbol.
La respuesta a la selección de características con considerable varianza no
aditiva, tales como crecimiento, es generalmente menos satisfactorio que para las
características relacionadas con la calidad de la madera, las cuales están usualmente
bajo un control genético aditivo más fuerte (Zobel y Jett 1995).
Aunque no ha sido cuantificado en detalles, parece ser que la mayoría de las
características de adaptabilidad están fuertemente heredadas de una manera aditiva. Por
lo tanto, se puede obtener una excelente ganancia a través del desarrollo de progenies
que crezcan adecuadamente en sitios marginales y submarginales. Esto es a través de la
selección de aquellos individuos que crecen bien en dichos lugares y del uso de sus
semillas para reforestación en áreas con similares características. El caso de la
resistencia a pestes involucra tanto la varianza aditiva como no aditiva, dependiendo de
la peste y la especie forestal involucrada.
2 alelo: una de varias formas alternativas de un gene que ocupan un locus dado en un cromosoma.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
17
La naturaleza ha creado la variación necesaria para su uso en un programa de
mejoramiento (Figura 2). Según Zobel y Talbert (1984), el mayor objetivo del
mejorador forestal es que los cuatro pasos siguientes se cumplan efectivamente:
1. la determinación del monto, tipo y causas de la variación genética
dentro de la(s) especie(s) de interés forestal;
2. el empaquetamiento de las características deseadas dentro de nuevos
individuos generados mediante cruzamientos selectivos;
3. la producción masiva de estos individuos mejorados con el propósito de
establecer plantaciones comerciales o la reforestación con otros fines y
4. desarrollo y mantención de una población genética base lo más amplia
posible para satisfacer las necesidades de las generaciones de selección
más avanzadas.
Figura 2. La fotografía muestra a un álamo (P. trichocarpa) que representa a un individuo con el arreglo genético adecuado para ser incluido en un programa de reproducción selectiva (fotografía es una gentileza del instituto INRA, Francia)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
18
A medida que los resultados del mejoramiento son alcanzados y los árboles
mejorados son obtenidos a través de generaciones sucesivas, los
procedimientos y estrategias utilizados para mantener y aprovechar al máximo
la variabilidad genética deberán ser inevitablemente más sofisticados y
científicamente estructurados, que aquellos utilizadas inicialmente.
En los PMG3 que utilizan la hibridación y/o propagación vegetativa, los cuatro
pasos anteriormente señalados aun se mantienen. La ventaja especial de la
propagación vegetativa es que el “nuevo arreglo genético”, o progenie mejorada y
obtenida mediante la selección y cruzamiento de árboles selectos, puede ser
reproducido muchas veces, y cada copia (propágulo) es genéticamente idéntica a otra.
Luego, la variación observada entre copias vegetativas (ramets) del mismo híbrido
(ortet) se debe exclusivamente a la variación ambiental (Figura 3). La propagación
vegetativa permite una ganancia genética rápida y sustancial porque todos los tipos de
variación genética pueden ser capturadas. Cuando el mejoramiento forestal se basa
únicamente en la reproducción por semilla, sólo una porción de la variación genética
de los árboles usados como padres será pasada a las progenies.
La hibridación tiene una ventaja comparativa porque permite la creación de
algo enteramente diferente. Esto se produce mediante la recombinación de la
variabilidad producida en la naturaleza, la cual esta empacada en un “nuevo
producto,” el árbol híbrido. Luego, la hibridación puede hacer posible crear un árbol
con buenas características para:
1. adaptarse a ambientes difíciles,
2. resistir el ataque de pestes y enfermedades,
3. obtener productos específicamente deseados y
3 PMG: programa de mejoramiento genético

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
19
4. obtener una productividad en biomasa y/o madera superior a la
registrada por ambos padres.
Figura 3. Fotografía obtenida del vivero experimental de álamo establecido por la Universidad de Talca, como parte del proyecto FONDEF D98I1086. Se observa la variación genética presente en el grado de susceptibilidad al ataque de roya, durante la temporada 2000-2001. A la izquierda, un híbrido severamente atacado por roya. A la derecha, un híbrido genéticamente diferente y totalmente inmune al ataque del hongo.
3.3 Biología de las especies
Finalmente, la biología de las especies debe ser tomada en cuenta. Esto incluye
aspectos tales como:
1. Tipo de floración: unisexual o bisexual;
2. Tipo de especie: monoica o dioica;
3. Modo de polinización predominante: autopolinización o polinización
cruzada, favorecida por el viento o insectos;
4. Habilidad para realizar injertos y formar raíces, tanto en púas apicales
como en estacas obtenidas de ramas;

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
20
5. Edad en la que se obtiene la madurez sexual;
6. Efectividad de la polinización controlada;
7. Presencia o ausencia de los genes que controlan los caracteres de
interés en la especie, tales como densidad de la madera, resistencia a
enfermedades, etc.;
8. Composición de la varianza genética (principalmente aditiva, o ¿es la
varianza no-aditiva también importante?)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
21
CCaappííttuulloo 44
MMÉÉTTOODDOOSS DDEE MMEEJJOORRAAMMIIEENNTTOO YY SSEELLEECCCCIIÓÓNN
LLas condiciones necesarias para que el mejoramiento genético tenga éxito son dos:
1. la existencia de variación genética en el carácter de interés y
2. la efectividad de escoger los padres adecuados para ser usados en los
cruzamientos (Namkoong et al 1988).
Las primeras técnicas de mejoramiento genético estuvieron basadas
generalmente en la colecta de semilla, originada por polinización abierta, desde
árboles padres con buen fenotipo (“árboles plus”). Estos lotes de semillas fueron
luego directamente utilizadas para el establecimiento de plantaciones, o más
comúnmente fueron establecidas en ensayos de progenie con el objeto de seleccionar
aquellos árboles madre que produjeran las mejores descendencias (en ensayos
retrospectivos).

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
22
Variantes a estos esquemas se produjeron con la posibilidad de injertar
material vegetativo de los árboles plus y establecer los injertos en huertos semilleros
clonales. Posteriormente, los huertos semilleros pudieron ser raleados sobre la base
del desempeño de las progenies en los ensayos de campo. Luego del raleo selectivo,
los huertos pudieron ser utilizados como una fuente de semilla originada de material
parental que demostró ser genéticamente superior en los ensayos de progenie.
Además del uso de la selección intraespecífica, la generación de híbridos
interespecíficos ha sido bastante importante en silvicultura (Fowler, 1978). El uso de
híbridos se facilita cuando ellos pueden ser propagados vegetativamente, como es el
caso de los álamo en distintas partes del mundo (Stettler et al 1996). Incluso cuando
la reproducción por semilla es considerada como prioritaria, los híbridos pueden
demostrar ser útiles en programas operacionales, particularmente cuando las
plantaciones comerciales deben ser establecidas en condiciones ambientales difíciles.
Los métodos de reproducción más importantes usados con especies forestales
que pueden ser reproducidas por polinización cruzada son (Shelbourne 1969; Allard
1960):
1. Selección masal,
2. Mejoramiento mediante retrocruzamientos,
3. Generación de variedades híbridas,
4. Selección recurrente

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
23
4.1 Selección masal
En este método, los individuos seleccionados generan semilla mediante polinización
abierta. La semilla es colectada y mezclada, no es establecida en un ensayo de
progenie y es directamente utilizada en el establecimiento de la siguiente generación
de plantaciones.
La selección está basada solamente en el fenotipo de la madre, puesto que no
hay control sobre la polinización y, por lo tanto, el grado de parentesco con los
árboles padre.
En términos simples, la semilla (de polinización abierta) se colecta en los
mejores árboles en el bosque y se usa para establecer las plantaciones desde las
cuales, a su debido tiempo, los mejores árboles serán nuevamente seleccionados.
La efectividad de este procedimiento está determinada por la heredabilidad4
del carácter involucrado y la intensidad de selección. Si la heredabilidad es alta, la
identificación del genotipo de los árboles madre será precisa y las progenies
obtenidas serán notablemente mejoradas.
4.2 mejoramiento por retrocruzamientos
Este método es particularmente útil en cultivos agrícolas para transferir genes
específicos a una buena variedad que es deficiente en una o unas pocas
características. En este método, los retrocruzamientos son hechos con el “parental”
más deseable mientras que la selección se practica respecto de los caracteres
transferidos desde el padre/madre donante.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
24
Las perspectivas para el uso de retrocruzamientos son bastante limitadas con
las especies forestales, principalmente porque una sola generación de selección puede
variar entre 4 a 20 años. Sin embargo, en especies con maduración sexual temprana,
los retrocruzamientos podrían ser aplicados, siempre y cuando el carácter transferido
también puede ser evaluado a una edad bastante temprana.
4.3 generación de variedades híbridas
Las variedades híbridas dependen para su superioridad del vigor híbrido o heterosis5
asociada con los híbridos de la generación F1. Líneas consanguíneas, clones, razas,
variedades o incluso especies pueden ser cruzadas para obtener el efecto heterótico.
Una vez que la combinación híbrida adecuada ha sido identificada, los genotipos
parentales deben ser mantenidos sin cambio, de manera que el mismo híbrido pueda
ser producido repetidamente.
A su vez, el híbrido obtenido, genéticamente superior a las líneas parentales,
puede ser considerado como un producto final, por lo tanto no es conveniente
que vuelva a ser utilizado en futuros cruzamientos, ya sea con alguno de los
padres o con otro híbrido F1, de manera que de origen a una progenie F2.
4 Heredabilidad: la relativa importancia de la herencia en la determinación de los valores fenotípicos particulares de un carácter. Expresa la proporción de la variación total en una población que es atribuible a los efectos promedios de los genes, los cuales generan las diferencias genéticas entre individuos. 5 Heterosis: el fenómeno de mayor capacidad adaptativa, desarrollo y supervivencia de organismos híbridos, particularmente aquellos formados mediante el cruzamiento de líneas altamente consanguíneas. Heterosis es usualmente el reflejo de un incremento en la heterosigosidad, y el incremento en vigor se piensa que se debe al enmascaramiento de alelos recesivos, los cuales son deletéreos en el estado homocigótico.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
25
Esto se debe a que cualquier nuevo cruzamiento tiene asociada la probabilidad que el
arreglo particular de alelos que confieren la heterosis al híbrido con características
notables puede ser destruida debido a:
1. la segregación independiente de los alelos, durante la meiosis, y
2. la recombinación aleatoria de los zigotos durante la fecundación.
Cruzamientos interespecíficos han sido usados en un número limitado de
PMG forestal, como una forma de obtener un mejoramiento genético que no es
posible con otros métodos. Esto usualmente toma la forma de una combinación de
caracteres útiles de una especie con aquellos de otra. El resultado esperado es que los
caracteres tengan, en general, un valor intermedio en el híbrido, así como la presencia
de vigor híbrido.
La generación de variedades híbridas puede ser desarrollada mediante el
sistema de reproducción selectiva de poblaciones múltiples, el cual se discute en la
sección 6.3. Un ejemplo gráfico de un ciclo de selección recurrente recíproca para
poblaciones reproductivas múltiples se muestra en la Figura 9.
4.4 selección recurrente
La mayor parte de los PMG forestal son diseñados para entregar una ganancia
continua a través de varios ciclos de mejoramiento. Los métodos de selección
presentados hasta ahora están centrados en el mejoramiento que puede ser obtenido
en una generación, pero ellos son también aplicables a programas que involucran
varias generaciones. El procedimiento de selección que involucra varios ciclos de
selección y cruzamiento es conocida como selección recurrente (Fig. 1). Varios
esquemas de selección recurrente han sido diseñados para plantas y animales de
manera que se pueda utilizar la capacidad combinatoria general y, en algunos casos,

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
26
la capacidad combinatoria específica. El sistema más utilizado en el mejoramiento
genético forestal es la selección recurrente simple, la cual es explicada más abajo.
Con este sistema, los PMG comienzan la selección de árboles en rodales naturales o
plantaciones sin mejoramiento en base a sus valores fenotípicos.
Los árboles seleccionados son luego cruzados y sus progenies son luego
establecidas de manera tal que puedan ser usadas como una fuente de
selección para la segunda generación de selección.
En muchos casos, la selecciones de la segunda generación son hechas en base a
valores familiares e individuales (ver sección 4.5). Estas selecciones son luego
cruzadas, creando una nueva generación de progenies que pueden ser usadas como
una fuente de selecciones para la próxima generación.
Este sistema es uno que se repite por si mismo. Las selecciones son hechas en
una población base (ver Capítulo 5), cruzadas en una forma determinada, y las
progenies resultantes sirven como una población base para la próxima
generación de reproducción.
Es importante mencionar aquí que cuando se estructuran varios ciclos de
selección, un cuidado extremo debe ser puesto en el tipo de resultado esperado, de
manera que los recursos genéticos estén disponibles para realizar el programa de
selección recurrente en forma exitosa.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
27
El mejorador forestal debe tener las siguientes dos metas en mente cuando
comienza un PMG forestal:
1. obtener el máximo de ganancia genética y en la forma más rápida
posible, a través de la selección y producción de semilla mejorada y
2. mantener una base genética grande y lo suficientemente diversa para
permitir que el PMG sea exitoso en forma continua por varias
generaciones.
Shelbourne (1969) realizó una revisión del método de selección recurrente y
describió cuatro tipos específicos, dependiendo de la forma en que los individuos con
características deseadas son identificados. Estos son:
1. Selección recurrente simple (identificación basada en el fenotipo
parental);
2. Selección recurrente para capacidad combinatoria general (basada en
el desempeño promedio de cada progenie6 obtenida del cruzamiento de
un árbol madre con varios padres evaluadores);
3. Selección recurrente para capacidad combinatoria específica (basada en
el desempeño promedio de cada progenie obtenida del cruzamiento de
un árbol madre con un solo padre) y
4. Selección recurrente recíproca.
6 En el contexto de este documento, una progenie se refiere al conjunto de descendientes obtenidos mediante cruzamientos dirigidos o polinización abierta.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
28
4.4.1 Selección recurrente simple (SRS)
Los genotipos deseables son seleccionados en base a su propio fenotipo y son
intercruzados sin restricción para producir las poblaciones que serán utilizadas en el
segundo ciclo de selección (Fig. 1). Por lo tanto, este proceso es diferente de la
selección masal, puesto que tanto el árbol padre como la madre son seleccionados.
Ciclos adicionales de selección pueden ser efectuados mientras haya un mejoramiento
efectivo. Para una intensidad de selección dada, el mejoramiento genético esperado
será el doble que el registrado mediante la selección masal.
En el mejoramiento forestal, la selección de árboles plus, su propagación en
un HSC7 y la cosecha de la semilla representa un solo ciclo de SRS. Los propágulos
clonales pueden interpolinizarse al azar en el HSC y la semilla producida puede ser
usada para establecer plantaciones desde las cuales se efectúe el próximo ciclo de
selección. Como no se realizan cruzamientos predeterminados ni tampoco ensayos de
progenies, la SRS, al igual que la SM, sólo es eficiente para caracteres con una
heredabilidad lo suficientemente alta para realizar selecciones precisas en búsqueda
de un genotipo superior a partir de la evaluación fenotípica. Los otros tipos de
selección recurrentes son más adecuados para la selección de caracteres con una baja
heredabilidad, puesto que ellos involucran el ensayo de cruzamientos para medir la
capacidad combinatoria.
4.4.2 Selección recurrente para capacidad combinatoria general8 (SR-CCG)
Esto involucra el cruzamiento de individuos fenotípicamente seleccionados con un
grupo de padres utilizados como “evaluadores”, los que poseen una base genética
7 HSC: huerto semillero clonal 8 CCG (Capacidad combinatoria general): el desempeño promedio de una progenie o un individuo cuando es cruzado con un número determinado de otros individuos de la población.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
29
amplia. En el mejoramiento genético forestal, esta práctica puede incluir alguno de
los siguientes esquemas de cruzamiento:
1. El cruzamiento de las selecciones con un grupo estándar de individuos
“evaluadores”;
2. El policruzamiento de individuos selectos con una mezcla de polen de
varios individuos selectos (polimix);
3. La colecta de semilla originada de polinización abierta entre los
individuos selectos con la población no seleccionada o
4. El uso de rametos vegetativos de los individuos selectos (esto entrega
una evaluación adecuada de la CCG de los individuos, asumiendo que
los efectos genéticos no aditivos no son muy grande)
La medición de los descendientes, luego de ser plantados en un diseño de campo
adecuado, entregará promedios del tipo:
1. medio-hermanos,
2. hermanos-completos o
3. clonal,
los cuales serán usados para obtener estimaciones de la CCG. Los individuos con los
valores más altos de CCG son luego propagados e intercruzados. La semilla originada
de estos intercruzamientos es mezclada y usada para formar la población del siguiente
ciclo de selección.
Este procedimiento corresponde al uso de un HSC con ensayo de progenie, el cual
es de uso general en los PMG forestal. Árboles superiores son seleccionados sobre la
base de su propio fenotipo en el bosque y propagados vegetativamente en un huerto o
banco clonal.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
30
La semilla es colectada de los árboles parentales selectos luego de efectuar alguna
de las siguientes modalidades de cruzamiento en el HSC:
1. polinización abierta,
2. cruzamientos controlados específicos o
3. policruzamientos.
El resultado será la generación de familias de medio-hermanos o hermanos-
completos, las cuales son establecidas en los ensayos de campo.
La fase de ensayo de progenie implica que los individuos seleccionados son
evaluados por su capacidad de generar progenies superiores antes de ser utilizados en
huertos semilleros, como productores de la semilla que será utilizada con fines
comerciales (Fig. 4).
4.4.3 Selección recurrente para capacidad combinatoria específica9 (SR-CCE)
La SR-CCE es posible para aquellas especies en que el mejoramiento ha avanzado a
un nivel tal que una o más líneas establecidas o variedades estándar son utilizadas
consistentemente como padres, o madres, en un PMG basado en la hibridación
intraespecífica. En este sistema de mejoramiento se pretende mejorar la productividad
de líneas híbridas mediante el uso de las variedades estándar como “evaluadores”, los
cuales son usados recurrentemente en cruzamientos con las líneas “no estándar”. Los
potenciales padres son seleccionados en base al desempeño de las progenies híbridas.
Estos son cruzados entre ellos para producir la próxima generación de padres
9 CCE (Capacidad combinatoria específica): el desempeño promedio de la progenie de un cruzamiento entre dos padres específicos que son diferentes de lo que sería esperado sobre la base de sus capacidades combinatorias generales solamente. Estas pueden ser positivas o negativas. La CCE siempre está referida a un cruzamiento específico y nunca a un padre en particular, por sí mismo.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
31
potenciales, los cuales serán incluidos en las líneas “no estándar”. En cualquier
generación, los padres selectos, o un subconjunto de ellos, podrán ser usados para la
producción de semilla comercial.
Este procedimiento también requiere del establecimiento de ensayos de
progenie para poder evaluar la efectividad de los cruzamientos (Fig. 4). En
condiciones de campo, se cuantifica el desempeño de los árboles incluidos en las
líneas no estándar, así como la efectividad de los parentales “evaluadores.”
Figura 4. Ciclo simple de selección con una fase de ensayo de progenie desarrollado en forma previa al establecimiento de huertos semilleros y la selección final. (Fuente: Namkoong et al 1998).
Selección
Muestreo y preselección
Ensayo de progenie Intercruzamientos
Población Forestal
Población forestal seleccionada o clones de
la población seleccionada
Información

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
32
Este método debe desarrollarse con el cuidado de no producir poblaciones
mejoradas con una base genética muy reducida, lo cual es considerado insatisfactorio
para el mejoramiento genético forestal de generaciones avanzadas.
4.4.4 Selección recurrente recíproca (SRR)
Este método ha sido propuesto como un procedimiento útil en la selección simultanea
de CCG y CCE. Se requiere dos poblaciones genéticamente heterogéneas (A y B) y,
en lo posible, deberían ser genéticamente divergentes, aunque no sexualmente
aisladas. Además, deberían tener características tecnológicas y un crecimiento
deseables. Un grupo de individuos selectos de la población A es cruzado con un
muestra de similar tamaño de la población B. De la misma forma, un grupo selecto de
la población B es cruzado con la población A. Los individuos en cada población que
muestran la más alta CCG con la otra población, basada en el desempeño de los
cruces de ensayo, son luego cruzados entre ellos dentro de la población y los
descendientes resultantes de cada población son utilizados para realizar el siguiente
ciclo de selección.
Las poblaciones desarrolladas mediante varios ciclos de SRR podrían ser
cruzadas en cualquier momento y, dependiendo del monto de divergencia en
frecuencias génicas y por lo tanto en heterocigosidad obtenida, los híbridos
resultantes serán más o menos heteróticos. Este esquema construirá dos poblaciones
que son altas en CCG para las características seleccionadas y, si la heterosis inter-
racial existe para características tales como crecimiento, estas poblaciones se tornarán
progresivamente más divergentes en sus frecuencias génicas. De esta manera, se
alcanzará un máximo de heterosis cuando estas poblaciones son cruzadas. Además, el
híbrido obtenido será generalmente intermedio en características entre las dos
poblaciones.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
33
En los PMG que trabajan con especies exóticas de rápido crecimiento, dos
razas (procedencias) de una especie, cada una con un buen desempeño pero
originarias de partes distantes del rango de distribución natural, pueden ser
combinadas para generar un híbrido heterótico o un híbrido que tolerará las
condiciones ambientales contrastantes de cada raza.
4.5 La importancia del tiempo
4.5.1 Secuencia lógica y práctica en el establecimiento de ensayos de progenie
Uno de los aspectos más importantes en un programa de reproducción selectiva de
generaciones avanzadas es el TIEMPO. Tomará años hacer la selección, los
cruzamientos y el establecimiento de los ensayos de progenie requeridos para obtener
el mejoramiento deseado. Cuando los beneficiarios del mejoramiento genético
reforestan grandes áreas anualmente, cada año que las plantas sin mejoramiento son
usadas para establecer rodales comerciales representa una pérdida en el futuro
retorno. Por lo que la fase operacional de un PMG está siempre bajo una fuerte
presión para producir retornos tempranos.
En todo PMG, el tiempo requerido debe balancear las posibles ganancias tanto
en el corto como en el largo plazo. El objetivo clave del mejoramiento genético
clásico ha sido obtener el máximo de ganancia por unidad de tiempo.
Para la mayor parte de las especies forestales, el tiempo que media entre dos
generaciones de selección es extenso, a causa de la larga edad de rotación y los diez o
veinte años que generalmente se requieren para obtener una floración abundante.
Aunque, corrientemente se realizan esfuerzos para acortar la edad de floración.
Resultados llamativos fueron observados por el primer autor en el sudeste de los

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
34
Estados Unidos, donde empresas forestales han desarrollado invernaderos
climáticamente controlados. Por ejemplo, algunos de estos resultados muestran que
clones de Pinus taeda pueden empezar a producir floración tres meses después de
establecidos en el invernadero.
Otro factor crítico es la necesidad de evaluar características tan importantes
como la tasa de crecimiento, la adaptabilidad, la resistencia a enfermedades y plagas
y la forma del árbol a una edad temprana. Generalmente se entiende que, para la
mayoría de las especies forestales, no es satisfactorio evaluar las características de
crecimiento antes de cumplida la mitad de la rotación (edad de cosecha), aunque
ciertas características de la forma, madera, y adaptabilidad pueden ser evaluadas con
precisión a una edad temprana.
El tiempo requerido en regenerar las masas forestales es una de las limitantes
mayores para el desarrollo de un PMG, aunque ciertos atajos se pueden desarrollar.
Un buen ejemplo es el establecimiento de huertos semilleros, dentro de la fase
operativa de un PMG. El mejor método científico para establecer un HSC que arroje
la máxima ganancia en producción es la evaluación cuidadosa de las progenies de
árboles parentales selectos a través de ensayos de progenies seguidos por el
establecimiento de HSC de producción, desde los cuales la semilla mejorada puede
ser obtenida. Esto es lo que habitualmente se entiende como una secuencia lógica
para el desarrollo del método de selección recurrente que incluya el establecimiento
de HSC y ensayos de progenie (ver sección 4.4.2). La secuencia lógica también está
representada por la Fig. 4.
Aunque puede entregar la máxima ganancia genética, esta secuencia lógica
considera varios años de atraso que incluyen el período requerido para establecer y
evaluar los ensayos, lo que incluso puede tomar hasta la mitad del período de rotación
forestal para algunas características. Esta pérdida de tiempo, antes que los árboles

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
35
finalmente seleccionados (genéticamente superiores) sean establecidos en los huertos
y la semilla este disponible, no puede ser aceptada cuando ésta se necesita
inmediatamente para un programa de plantaciones a gran escala. Para superar este
inconveniente, la secuencia de etapas involucrada en el método de selección
recurrente puede sufrir algunas modificaciones. Normalmente, los mejores fenotipos
son seleccionados (árboles plus) e inmediatamente establecidos en el huerto antes que
el valor genético de sus genotipos este determinado.
Rápidamente después que (o al mismo tiempo que) el HSC se ha establecido
con los mejores fenotipos, los ensayos de progenie se pueden establecen para
determinar el "valor" genético de los padres usados en el huerto semillero inicial. En
cuanto los datos de las progenies son disponibles, el huerto puede ser "depurado" de
los padres indeseables (genotípicamente inferiores).
Durante el periodo de ensayo, la semilla para plantación que ha sido obtenida
en el huerto es considerablemente mejor que la semilla de origen natural, pero
no es tan buena como aquella proveniente de un huerto en el que sólo se
mantienen aquellos árboles seleccionados que ya demostraron ser
genéticamente superiores.
A esta alternativa se le puede denominar como secuencia práctica del método de
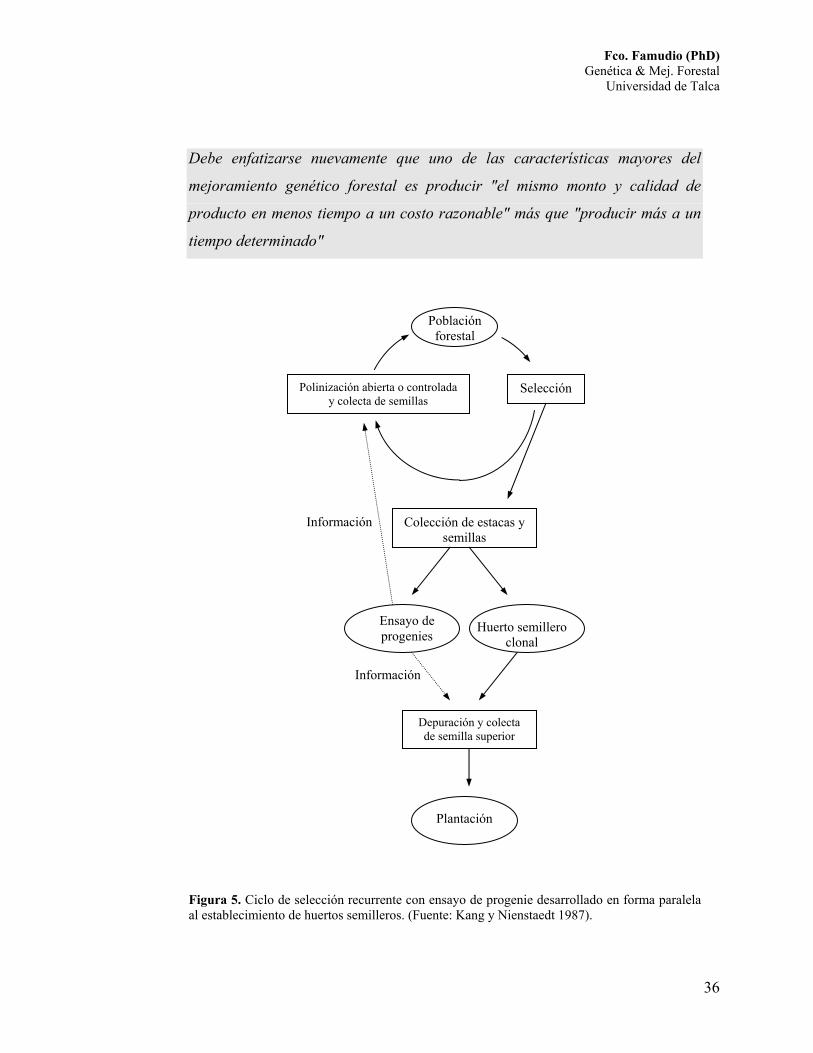
selección recurrente y está gráficamente representada en la Figura 5.
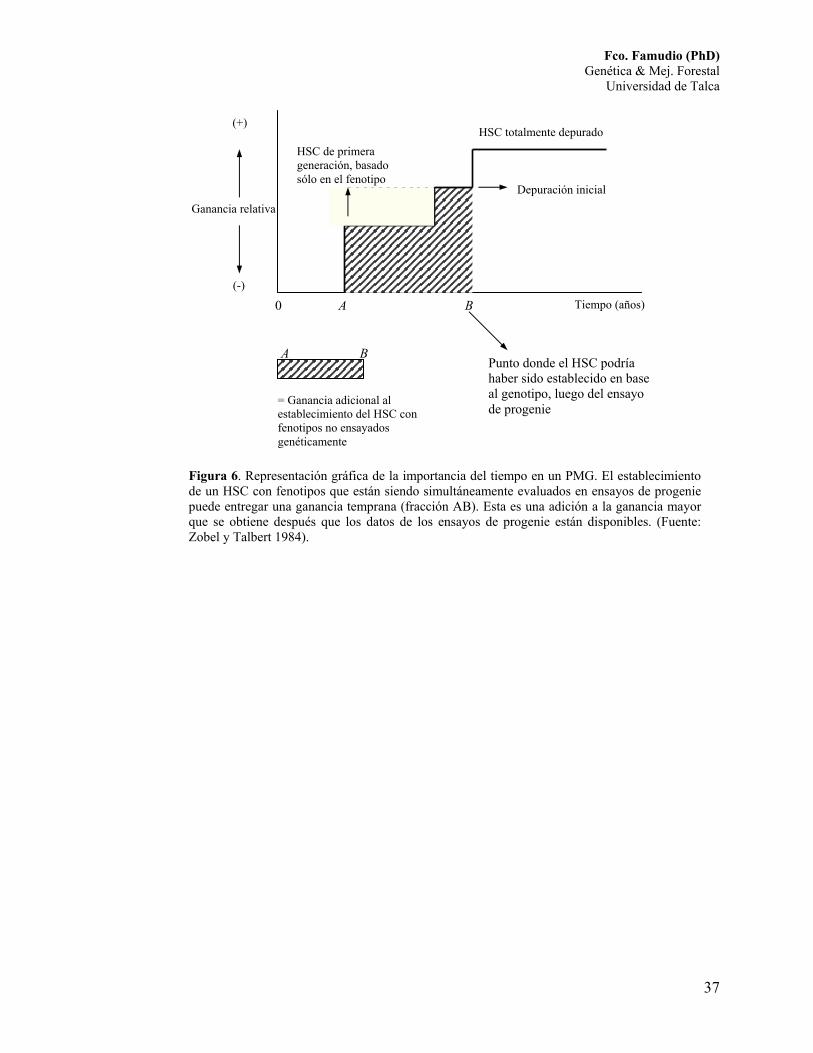
El valor del tiempo puede ser observado en la Figura 6. Una ganancia menor
pero temprana se obtienen cuando el huerto se establece con padres escogidos sólo
por su apariencia fenotípica comparada con la ganancia obtenida cuando la calidad
genética de los padres es conocida. La ganancia se incrementará a medida que la
información proveniente de los ensayos de progenie se hace disponible y la
"depuración" se realiza en el huerto semillero.
Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
36
Debe enfatizarse nuevamente que uno de las características mayores del
mejoramiento genético forestal es producir "el mismo monto y calidad de
producto en menos tiempo a un costo razonable" más que "producir más a un
tiempo determinado"
Figura 5. Ciclo de selección recurrente con ensayo de progenie desarrollado en forma paralela al establecimiento de huertos semilleros. (Fuente: Kang y Nienstaedt 1987).
Selección Polinización abierta o controlada y colecta de semillas
Población forestal
Colección de estacas y semillas
Depuración y colecta de semilla superior
Huerto semillero clonal
Plantación
Ensayo de progenies
Información
Información
Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
37
Figura 6. Representación gráfica de la importancia del tiempo en un PMG. El establecimiento de un HSC con fenotipos que están siendo simultáneamente evaluados en ensayos de progenie puede entregar una ganancia temprana (fracción AB). Esta es una adición a la ganancia mayor que se obtiene después que los datos de los ensayos de progenie están disponibles. (Fuente: Zobel y Talbert 1984).
Ganancia relativa
A B
HSC totalmente depurado
Depuración inicial
HSC de primera generación, basado sólo en el fenotipo
Punto donde el HSC podría haber sido establecido en base al genotipo, luego del ensayo de progenie
Tiempo (años) 0
A B
= Ganancia adicional al establecimiento del HSC con fenotipos no ensayados genéticamente
(+)
(-)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
38
4.6 tipos de selección asociados a los métodos
Existen diversos tipos de selección, pero en esta sección se tratarán algunos de los
más relevantes y comúnmente encontrados en la literatura.
4.6.1 Selección tipo masal o individual
En el método de mejoramiento mediante la selección masal (ver sección 4.1), la
técnica de selección de individuos tiene el mismo nombre que el método. La
selección masal, también denominada selección individual (Falconer y Mackay
1998), involucra:
escoger individuos solamente sobre la base de sus fenotipos, sin poner atención
en el desempeño de los ancestros, descendientes, u otro tipo de parientes.
La selección masal trabaja bien para caracteres altamente heredables, donde el
fenotipo es un buen reflejo del genotipo. Es el único tipo de selección que puede ser
usado en rodales naturales o en plantaciones donde el pedigree es desconocido. La
selección masal es raramente utilizada cuando los pedigrees son conocidos, como es
el caso de los ensayos genéticos de generaciones avanzadas, debido a que una mayor
ganancia genética puede ser obtenida usando otros métodos.
4.6.2 Selección tipo familiar
Este tipo de selección involucra:
La elección de familias enteras en base a sus valores fenotípicos. No hay
selección de individuos dentro de las familias y los valores de árbol individual
son utilizados sólo para calcular los promedios familiares

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
39
La selección familiar trabaja bien con caracteres de baja heredabilidad, donde los
fenotipos individuales no son un buen reflejo del genotipo. Cuando los promedios
familiares se basan en un gran número de individuos, la varianza ambiental tiende a
ser reducida, y los promedios familiares se vuelven un buen estimador del valor
genético promedio. La selección familiar es escasamente aplicada como único tipo de
selección en un PMG forestal. Esto se debe a que una mayor ganancia genética puede
ser lograda a partir de otros métodos que incluyen la selección familiar como parte
del método. La selección familiar también puede conducir a un incremento de la tasa
de consanguinidad porque familias enteras pueden ser descartadas, por lo tanto se
reduce la base genética de la población.
4.6.3 Selección tipo ensayo de progenie
Este tipo de selección involucra:
la selección de árboles parentales en base al desempeño de las progenies.
Primero se establece el ensayo de progenie con las descendencias de los
árboles candidatos (por ejemplo, árboles plus originados de poblaciones
forestales sin mejoramiento), las progenies crecen y se desarrollan, estas son
evaluadas y, finalmente, se selecciona aquellos candidatos cuyas progenies
presentaron las mejores características en comparación con el resto del
ensayo.
Este tipo de selección puede ser muy precisa, debido que permite estimar en forma
directa los valores reproductivos10 que serán usados en el proceso de selección. Esto
es lo que ocurre cuando los padres de un HSC son establecidos en ensayos de
progenie (ver selección recurrente para capacidad combinatoria general en sección
10 Valor reproductivo: es una propiedad genética del individuo, en referencia a una población en particular. Consiste en la suma de los efectos genéticos aditivos de todos los alelos, en todos los loci, que controlan un carácter en un individuo.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
40
3.4) y los huertos son raleados de aquellos padres que demostraron ser genéticamente
pobres. Generalmente, los ensayos de progenie no son la forma inicial de selección
para todos los PMG. Si el método de selección comienza con el ensayo de progenie,
la longitud del intervalo entre dos generaciones de selección se alarga
considerablemente, lo que significa una pérdida crítica en tiempo. Otros tipos de
selección son usualmente más eficientes en maximizar el monto de ganancia genética
por unidad de tiempo.
4.6.4 Selección tipo dentro de familia
Los individuos aquí son escogidos sobre la base de sus desviaciones del promedio
familiar, y los valores familiares per se no son ponderados cuando se realizan las
selecciones. De todos los tipos de selección, éste entrega la tasa más baja de
consanguinidad. Si esta tasa es alta, se transforma en un problema mayor para un
PMG.
4.6.5 Selección tipo entre familias más selección dentro de familias
Este tipo de selección en dos etapas involucra:
la selección entre familias seguido por la selección de individuos dentro de
familias.
Esta selección trabaja bien con bajas heredabilidades y es el tipo predominante de
selección utilizado en los PMG más avanzados. Consiste en escoger las mejores
familias seguido de la selección de los mejores individuos dentro de ellas. Un
refinamiento de este tipo de selección es la “selección combinada”, en la cual se
calcula un índice que pondera todos lo individuos sobre la base de sus valores

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
41
familiares combinados con sus valores fenotípicos individuales. Los coeficientes de
ponderación, o peso, usados en el índice dependen de la heredabilidad del carácter.
Más peso es dado al promedio familiar para caracteres con baja heredabilidad,
mientras que una mayor ponderación es otorgada al valor individual cuando la
heredabilidad del carácter alta.
4.7 selección para varios caracteres
La mayor parte de los PMG están orientados a mejorar varios caracteres al mismo
tiempo. Esto requiere que la información obtenida de diferentes caracteres sea
incluida en el proceso de selección. Cualquiera de los tipos de selección mencionados
arriba podría ser utilizado para desarrollar información sobre caracteres individuales,
pero la información muchas veces debe ser manipulada para desarrollar un esquema
de selección de multicaracteres. Esencialmente, se han desarrollado tres sistemas para
la selección de caracteres múltiples.
4.7.1 Selección en tandem
En este sistema, la selección y posterior cruzamiento es realizado para un carácter
primero hasta que se obtiene un nivel de mejoramiento deseado. Usualmente, el
primer carácter es el más importante, aunque la obtención del grado de mejoramiento
puede tomar más de una generación. Luego, los esfuerzos de selección y cruzamiento
se concentran en otro carácter. Este método de mejorar varios caracteres en tandem es
raramente utilizado debido a las presiones de tiempo y la necesidad de mejorar varios
caracteres simultáneamente. El uso prioritario de la selección en tandem es cuando un
carácter tiene una importancia primordial, tal como la resistencia a enfermedades.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
42
4.7.2 Selección independiente
Este método de selección múltiple involucra la determinación de valores mínimos
para cada carácter de interés. Los individuos deben cumplir con el criterio mínimo si
ellos van a ser seleccionados. La figura 7 (extraída de Zobel y Talbert 1984) muestra
gráficamente este procedimiento.
Figura 7. El método de selección independiente se ilustra como si fuese aplicado a 2 caracteres. Sólo aquellos individuos que superan un mínimo estándar para ambos caracteres son seleccionados. (Fuente: Zobel y Talbert 1984)
4.7.3 Índice de selección
Esta es una forma de selección de caracteres múltiples que combina información de
todos los caracteres de interés en un solo índice. Esto permite al mejorador asignar un
valor total a cada individuo. Además de información genética, también incluye una
ponderación económica a cada una de las características en consideración. En su
forma más completa y compleja, un índice de selección combina tanto información
familiar como individual para todos los caracteres en un solo índice.
Caracter B
Caracter A
Valores mínimos de selección

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
43
El valor del índice para cada individuo es derivado a través de una ecuación
de regresión múltiple, en la cual los coeficientes dependen de las heredabilidades, la
correlación genética entre caracteres, y el peso económico de cada carácter.
Teóricamente, el índice de selección es el método de selección múltiple que entrega
la mayor ganancia genética total para todos lo caracteres combinados (Namkoong et
al 1988, Falconer y Mackay 1998, White y Hodge 1989) Un problema mayor consiste
en determinar los pesos económicos adecuados. El uso de índices de selección donde
los pesos económicos son marcadamente incorrectos puede conducir a programas de
selección muy ineficientes.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
44
CCaappííttuulloo 55
TTIIPPOOSS DDEE PPOOBBLLAACCIIOONNEESS
UUna de las características más notables del mejoramiento genético denominado de
“generaciones avanzadas” es la separación de las poblaciones forestales que lo
componen, de acuerdo a su función particular en el PMG. Quizás Namkoong et al
(1966) fueron los primeros en reconocer poblaciones con diferentes funciones. Van
Buijtenen (1975) clasificó las poblaciones en cuatro tipos: población reproductiva,
población de ensayo de progenie, población productora de semilla y rodales
forestales. Aquí se debe notar que la clasificación de las poblaciones se basa en su
función y no en su subdivisión física. Por ejemplo, una población física simple puede
desempeñar varias funciones, mientras que la población reproductiva puede estar
subdividida en subpoblaciones. A continuación se describen las poblaciones que
según Zobel y Talbert (1984) deben estar siempre presentes en cualquier momento de
un ciclo de selección y reproducción.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
45
5.1 población base
Consiste del conjunto de árboles desde los cuales el mejorador forestal escoge para
realizar las selecciones para la próxima generación de reproducción. En la primera
generación de mejoramiento, la población base usualmente está formada por árboles
creciendo en rodales naturales o plantaciones sin mejoramiento. En generaciones
avanzadas, la población base es muy a menudo el o los ensayos genéticos que
contienen las progenies de los árboles seleccionados de la generación previa. La
identidad de al menos uno y usualmente ambos padres de todas las progenies
creciendo en el ensayo genético es conocida. La población base comúnmente
contiene entre cientos a varios miles de genotipos.
5.2 población reproductora
Consiste de un subconjunto de individuos de la población base, la cual es
seleccionada por sus características deseables y servirá como padres para la próxima
generación de reproducción. Los incrementos en la ganancia genética que resultan de
proseguir a la siguiente generación de mejoramiento son el resultado de los esfuerzos
de selección orientados a configurar la población reproductora.
Se deberá llegar a un acuerdo entre incrementar el diferencial de selección,
que entrega una ganancia genética mayor a costa de una población reproductora de
menor tamaño, y la necesidad de mantener una población reproductora lo
suficientemente grande, de manera que la variación genética sea mantenida a un nivel
que permita un progreso genético continuado en futuras generaciones. En la práctica,
se utiliza una intensidad de selección moderada en la configuración de la población
reproductora. Usualmente, 200 o más árboles seleccionados son incluidos en esta
población en cada generación (Zobel y Talbert 1984)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
46
5.3 población de producción
Esta población es usada estrictamente para producir semillas o propágulos
vegetativos para ser utilizados en programas de reforestación operacionales. En
algunos casos, la población de producción consiste del mismo set de genotipos de la
población reproductora, pero usualmente la población de producción es un
subconjunto altamente seleccionado de esta última (genotipos elite) Entre 20 a 30
genotipos pueden ser incluidos en la población de producción. Estos individuos
pueden ser los mejores en la población reproductora y son las mejores selecciones
que pueden ser obtenidas en cada generación. Ellos son escogidos para maximizar la
ganancia genética en plantaciones forestales comerciales.
Una población de producción pequeña y altamente seleccionada es un punto
final desde una perspectiva reproductiva, debido principalmente a su reducida base
genética. Si esta población se utilizara para una posterior selección genética, se
produciría un decrecimiento en los esfuerzos de mejoramiento y muy rápidamente
llevaría a un incremento significativo de la consanguinidad, mediante la endogamia.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
47
CCaappííttuulloo 66
EESSTTRRAATTEEGGIIAASS DDEE SSEELLEECCCCIIÓÓNN YY RREEPPRROODDUUCCCCIIÓÓNN PPAARRAA EELL ÁÁLLAAMMOO
6.1 reproducción y selección clonal
Dos son las características que han predominado en la historia de la reproducción
selectiva del álamo:
1. la reproducción selectiva interespecífica y
2. la selección clonal (Bisoffi y Gullberg 1996).
Híbridos interespecíficos se encuentran a menudo en la naturaleza donde el rango
de distribución de dos especies se traslapa (Eckenwalder 1984). Muchos de estos
híbridos espontáneos comenzaron a ser usados tanto en Europa como en
Norteamérica a partir del siglo 18, aunque los primeros cruzamientos dirigidos fueron
hechos sólo a comienzos del siglo 20 (Bisoffi y Gullberg 1996). La generación de
híbridos interespecíficos fue también el objetivo de los primeros programas de
reproducción artificial siguiendo los métodos de selección modernos (Screiner 1970).

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
48
La superioridad de los híbridos en F1 ha sido más bien asumida que probada,
pero estudios recientes confirman que el vigor híbrido existe (Bradshaw y Stettler
1996). La superioridad de híbridos F1 parece ser debido generalmente a la
complementación de genes dominantes o codominantes llevados separadamente por
las dos especies parentales en diferentes loci (Stettler et al 1988).
El segundo aspecto de la mayoría de los PMG de álamo ha sido el fuerte
componente de selección clonal (Thielges 1985). Esto se ha debido parcialmente a los
recursos limitados que han impuesto usualmente la maximización de los beneficios en
el corto plazo. Aunque otra razón se debe a que la evaluación precisa de clones ha
sido efectuada gracias a la relativa facilidad que presentan los álamos para ser
propagados tanto vegetativa como sexualmente.
El poder de la selección clonal ha sido demostrado por el impresionante
número de híbridos producidos por los mejoradores y cultivados a través del
mundo. Bissoffi (2001, comunicación personal) señala que alrededor de 280
cultivares o variedades son o han sido cultivadas en el mundo, la mayor parte
representadas por clones.
Métodos no convencionales de mejoramiento también han sido probados, pero
hasta ahora han tenido un impacto menor en la producción de variedades. Por
ejemplo, clones triploides11 de Populus tremula fueron producidos a comienzos de la
década de los 60, pero el interés por la poliploidia12 declinó en los 70, con la
excepción de algunos trabajos en el campo de la genética molecular (Bradshaw y
Stettler 1993). Interesantes resultados han sido obtenidos a través de la variación
somaclonal13, especialmente en el campo de la resistencia in vitro a factores adversos
11 Triploide: individuo con tres juegos de cromosomas, en oposición a uno diploide que posee el número normal de dos sets. Uno aportado por el árbol padre y el otro por el árbol madre. 12 Poliploidia: presencia de un número mayor del set doble de cromosomas que caracterizan a una especie 13 Variación somaclonal: variación genética producida por mutaciones que ocurren en el tejido vegetativo (somático), por lo tanto afectan sólo a los descendientes y no se transmite a través de la herencia.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
49
tales como herbicidas, toxinas y sales (Michler y Haissig 1988). Algunas técnicas
relacionadas, tales como el cultivo de protoplastos como una base para la hibridación
somática u otras aplicaciones, aunque han probado ser viable a un nivel experimental,
su aplicación en el mejoramiento genético aun aparece como muy distante.
La facilidad que presentan los híbridos de álamo para ser manipulados in vitro
ha motivado la investigación y aplicación de la tecnología recombinante del ADN14
por su habilidad de incorporar secuencias de ADN en un genotipo bien definido con
mínima disrupción de su organización genómica. En efecto, los álamos fueron los
primeros árboles forestales en ser genéticamente transformados con un gene de
importancia agronómica (Fillatti et al 1988).
Hoy en día existen varios grupos de investigación en el mundo usando los
álamos y la genética molecular para entender los mecanismos de expresión génica en
árboles (Han et al 1996). La identificación de genes de importancia para el
desempeño varietal también está siendo desarrollado desde años recientes.
Específicamente, los esfuerzos se han concentrado en genes involucrados en la
resistencia a herbicidas, insectos, enfermedades y estrés debido a factores abióticos, o
genes involucrados en ciertas vías bioquímicas tales como la síntesis de lignina.
Debemos esperar que este tipo de estudio tendrá un valor de importancia creciente en
el futuro inmediato.
Es evidente que la reproducción selectiva de álamo ha creado y ensayado
soluciones que no son sólo justificadas desde una perspectiva de corto plazo sino que
representan opciones metodológicas potencialmente útiles en el futuro. Actividades
tales como la generación de haploides15 y tetraploides son más bien clásicas, mientras
14 Esta tecnología consiste en el aislamiento del ADN a partir de individuos donantes, la separación de segmentos que contienen genes de interés relevante, la inserción de estos segmentos aislados en vectores artificialmente construidos y la inserción de éstos dentro del genoma (ADN) de individuos receptores 15 haploide: individuo que posee un solo juego de cromosomas, mientras que el número normal es de dos, uno de cada individuo parental

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
50
que la utilización de la variación somaclonal y la aplicación de las técnicas de
recombinación de ADN son más bien recientes. Todas estas pueden ser consideradas
como típicamente de largo plazo. Sin embargo, un componente necesario para la
aplicación sistemática de la reproducción selectiva ha sido sólo superficialmente
explorada, tal es el caso de la reproducción sistemática de generaciones múltiples
(Bisoffi y Gullberg, 1996).
A continuación se presentan algunos aspectos esenciales en el diseño de
estrategias de mejoramiento para álamo, así cómo en la determinación de la estructura
y desarrollo de la población reproductiva.
6.2 reproducción selectiva de generaciones múltiples
Puesto que el potencial genético de los álamos permite la reproducción
selectiva interespecífica y la selección clonal muestra ser una herramienta de
éxito, lo lógico ha sido que una estrategia de mejoramiento genético del
género Populus incluya la hibridación interespecífica y todas las
herramientas estadísticas y biotecnológicas que permiten optimizar las
técnicas de selección basadas en la reproducción vegetativa.
Según Bisoffi y Gullberg (1996), el punto de vista prevaleciente en los
mejoradores de álamo ha sido adoptar esquemas de reproducción selectiva enfocados
en una ganancia que sea obtenible en una o dos generaciones. Lo anterior debido a la
enorme variación genética generada en un segunda generación de selección y los
recombinantes que han podido ser obtenidos de tales trabajos han demostrado tener
un desempeño superior al esperado.
Por otra parte, las nuevas posibilidades de mejoramiento que pueden aportar
las nuevas tecnologías de manipulación del genoma podrían promover una falta de

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
51
interés por la reproducción selectiva que involucra generaciones múltiples. Los éxitos
recientes obtenidos con el uso de la técnica de recombinación de ADN en álamos
permitirían una construcción rápida de nuevos genotipos mediante la transferencia
génica, en vez de la selección a través de varias generaciones. Las técnicas mejoradas
para la selección precisa de genes valiosos, o combinaciones génicas, mediante el uso
de los marcadores moleculares también puede ayudar a desviar la atención del
mejoramiento de largo plazo.
Sin embargo, es poco probable que la aplicación mecánica de la reproducción
selectiva de corto plazo (que no es lo mismo que la selección recurrente, la cual
incluye distintos tipos de poblaciones, como las mencionadas en el Capítulo 5), o el
uso de las tecnologías de manipulación molecular del genoma, entreguen los mejores
resultados de largo plazo. Esto se debe a que los ambientes en que los álamos
habitualmente crecen tienden a ser extremadamente heterogéneos, tanto en el tiempo
(cambios climáticos estacionales, o anuales, y cíclicos de largo plazo) como en el
espacio (fluctuaciones en la disponibilidad de agua, nutrientes, oxígeno, etc.).
Además, como ha sido reportado para muchas especies forestales, a pesar del pobre
conocimiento de la estructura genómica, es posible cambiar el valor promedio de
caracteres de valor forestal a través de la reproducción selectiva de generaciones
múltiples (Zobel y Talbert 1984; Namkoong et al 1988).
6.2.1 Interacción genotipo - ambiente
La presencia de heterogeneidad ambiental puede dar origen a la interacción genotipo
x ambiente, la cual puede convertirse en un problema serio en la selección de híbridos
notables. Barnes et al (1984) mencionan que una interacción entre dos factores (por
ejemplo: efectos debido a la familia e híbrido o clon y el sitio de ensayo) puede ser
definida, en términos estadísticos y genéticos, como una pérdida de aditividad de los

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
52
efectos individuales (genotipo y ambiente), o bien como un cambio de orden de los
niveles de un primer factor a diferentes niveles del segundo factor.
En términos simples, la interacción genotipo x ambiente significa que el
desempeño relativo de clones, familias, procedencias, o especies, difiere
cuando ellos crecen en diferentes ambientes.
Esto puede involucrar un cambio real en el rango, que es el tipo de interacción más
importante para un mejorador forestal, o puede simplemente involucrar un cambio en
la variación de un ambiente a otro, sin que se produzca un cambio de rango (Kang y
Gauch 1998)
Debido a la interacción genotipo x ambiente, la extrapolación del fenotipo de
un ambiente a otro puede ser difícil, puesto que los genes involucrados y sus
expresiones pueden cambiar. Por lo tanto, la manipulación de genes individuales,
realizada a través de la tecnología recombinante del ADN, es una solución
insuficiente para aquellos caracteres que lo presentan. Luego, la reproducción
selectiva de generaciones múltiples específicamente orientada a varios ambientes-
objetivo puede ser una solución parcial mínima. Por lo tanto, los mejoradores de
álamo necesitan considerar la reproducción selectiva de largo plazo como una parte
integral de sus estrategias reproductivas. No importa todo lo remota que pueda
parecer la reproducción selectiva de generaciones múltiples, las decisiones tomadas o
no tomadas hoy van a limitar las futuras opciones si los aspectos ligados al
mejoramiento de largo plazo son ignorados (Bisoffi y Gullberg 1996)

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
53
6.3 sistemas de manejo de una población reproductiva de álamos
Existen al menos dos aproximaciones para mantener las poblaciones reproductivas de
álamo en un esquema de generaciones múltiples. En un primer caso, la población
reproductiva conforma una sola unidad integra, la cual es sometida, en un generación
de selección determinada, a esquemas de cruzamientos que llevan a configurar la
próxima generación de progenies selectas. En un segundo caso, la población
reproductiva esta conformada por poblaciones múltiples.
Existen varias desventajas en mantener una sola población reproductiva
grande. La eficiencia en la selección total es baja. Los árboles de la población
reproductiva deben ser usualmente plantados en una amplia área, lo cual hace que el
control ambiental sea difícil comparado con los cultivos agrícolas.
Una población reproductiva simple debe mostrar un amplio grado de
adaptabilidad a condiciones ambientales, lo cual puede ser posible con el sacrificio de
una disminución en la heredabilidad y la intensidad de selección. Esta deficiencia
puede ser corregida manteniendo una población reproductiva subdividida, en que
cada subpoblación puede ser establecida en cada tipo de macro-ambiente objetivo.
Además, independientemente del tamaño de la población, existe la probabilidad que
una población simple experimente una sola secuencia de ambientes y alguno
complejos génicos útiles en otras condiciones ambientales sean eliminados.
Por otra parte, el uso de una población reproductiva subdividida (réplicas o
sublineas) permite al mejorador aumentar la posibilidad de conservar y/o crear
variación genética para futuras generaciones. También el establecimiento de
poblaciones múltiples, es decir, subpoblaciones que no compartan ancestros comunes,
permitirá al mejorador preservar la variabilidad actual. La creación de nuevas

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
54
adaptaciones en algunas de las subpoblaciones a través de la selección puede mejorar
la posibilidades de superar los problemas ambientales imprevisibles (Bisoffi y
Gullberg 1996)
El programa de mejoramiento genético de álamo de Italia es un ejemplo, al
nivel de la especie, de cómo una población reproductiva múltiple puede otorgar
flexibilidad en la mantención de la variación genética (E. Bisoffi, comunicación
personal).
6.3.1 Sistema de reproducción de población simple
La Fig. 8 es una adaptación de la Fig. 1, la cual muestra el flujo de actividades
consideradas para un ciclo de selección, dentro de un sistema de selección recurrente,
para el caso de una población reproductiva simple. La selección puede ser realizada a
través de ensayos de progenie o selección genotípica-clonal. El procedimiento de
selección también puede estar basado en el tipo de selección individual (el más
simple), algún tipo de selección familiar (como se menciona en la sección 4.6), o en
algún procedimiento de selección de caracteres múltiples (ver sección 4.7). Los
cruzamientos pueden incluir todos, o un subconjunto, de los padres selectos. Las
variedades comerciales también pueden ser el total de la población seleccionada o, lo
más probable, un subconjunto de las familias o clones seleccionados. Las
intensidades de selección están marcadas como “m,” para las madres y “p,” para los
padres.
Según Bisoffi y Gullberg (1996), el sistema de reproducción de una población
simple, en un sentido riguroso, es improbable que exista en el mejoramiento genético
de álamo. La mantención de una población reproductiva dividida está a menudo
motivada por el hecho que la mayor parte de las poblaciones reproductivas utilizadas
en el mejoramiento de largo plazo son demasiado grandes para ser manejadas como

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
55
poblaciones simples. Independiente del o los objetivos del mejoramiento de álamos,
la red global de mejoradores de álamo es probable que replique la población
reproductiva, la divida en múltiples poblaciones, o aplique ambos criterios.
Figura 8. Desarrollo de una población reproductiva simple para el caso de especies del género Populus. (Adaptado de Bisoffi y Gullberg, 1996). Las tasas de selección para madres y padres son expresadas con las letras "m" y "p", respectivamente.
6.3.2 Sistema de reproducción de poblaciones múltiples
Los actividades relacionadas con la selección y los cruzamientos que buscan
aprovechar los efectos heteróticos, o la combinación de caracteres, son algo más fácil
de organizar si consideramos la reproducción selectiva en poblaciones múltiples. Si
N padres
N madres
mN madres
pN padres
Selección
Variedades comerciales
Selección
Selección Selección y cruzamientos intraespecíficos
Próximo ciclo

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
56
todos los esfuerzos de mejoramiento son confinados a una sola población
reproductiva o un par de poblaciones, la organización del mejoramiento se puede
complicar (Namkoong et al 1988)
En las operaciones forestales, los diversos y a veces heterogéneos
requerimientos demandados por los usuarios de productos forestales, o los objetivos
múltiples pueden ser alcanzados en varias formas. Por ejemplo, todos los
requerimientos o caracteres pueden ser combinados en un solo árbol objetivo y todos
los árboles pueden ser cruzados y manejados de la misma forma. Alternativamente,
un rodal puede consistir de árboles similares, pero los rodales pueden ser diseñados
para diferir en su origen genotípico (por ejemplo rodales clonales) y/o régimen
silvicultural.
Otra alternativa es mezclar diferentes genotipos en rodales simples de manera
que árboles distintos satisfagan varios objetivos. Obviamente, el arreglo de metas
para cada bloque o rodal puede variar también, de manera que diferentes mezclas
estarían presentes en los diversos bloques (rodales) de manejo.
En un sistema productivo forestal de objetivos múltiples es razonable pensar
que diferentes poblaciones reproductivas puedan entregar genotipos para diferentes
objetivos, los cuales pueden incluir:
1. generar linajes adaptados a condiciones ambientales específicas;
2. mejorar una forma especial de crecimiento;
3. aumentar la producción de biomasa;
4. mejorar la calidad de la madera;
5. aumentar la resistencia a plagas y/o enfermedades específicas.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
57
Si la acción de los genes es predominantemente aditiva, luego las poblaciones
reproductivas pueden generar poblaciones híbridas, las cuales presentarían un
desempeño promedio en la gran mayoría de los caracteres en relación con las
poblaciones parentales. Estas combinaciones híbridas de poblaciones cruzadas para
lograr diferentes objetivos corresponden una forma de selección independiente a
nivel de poblaciones (Namkoong et al 1988).
Un esquema de reproducción selectiva mediante poblaciones múltiples es
observado en la Fig. 9. De acuerdo con Kang y Nienstaedt (1987), las poblaciones
subdivididas pueden consistir de poblaciones con un origen genético distinto, como
puede ser el caso de dos especies de Populus, una colección de poblaciones
originadas de diferentes subconjuntos de la población base y/o copias de la misma
población base, las cuales comparten los mismos ancestros. En los dos últimos casos,
nos estamos refiriendo a una sola especie.
Cuando este sistema se utiliza con dos especies que no han sido usadas en
hibridación, ambos lados del sistema de parentesco híbrido deben ser desarrollados.
Un método puede ser esencialmente duplicar el método de selección recurrente
simple en dos poblaciones, pero se debe usar los intercruzamientos entre ellas para
probar el desempeño de los híbridos intraespecíficos o interespecíficos y para la
producción de las variedades comerciales híbridas. Aquí se busca maximizar el uso
de la heterosis. El programa de selección genética de álamo de Italia es un ejemplo de
este tipo de esquema (Bisoffi y Gullberg 1988).
Las poblaciones reproductivas son mantenidas genéticamente separadas. Dentro
de cada una de ellas se aplica un esquema de selección recurrente. Los padres selectos
son cruzados entre ellos mismos, dentro de cada población, para dar origen a la
próxima generación de selección recurrente. También se realiza una selección de los
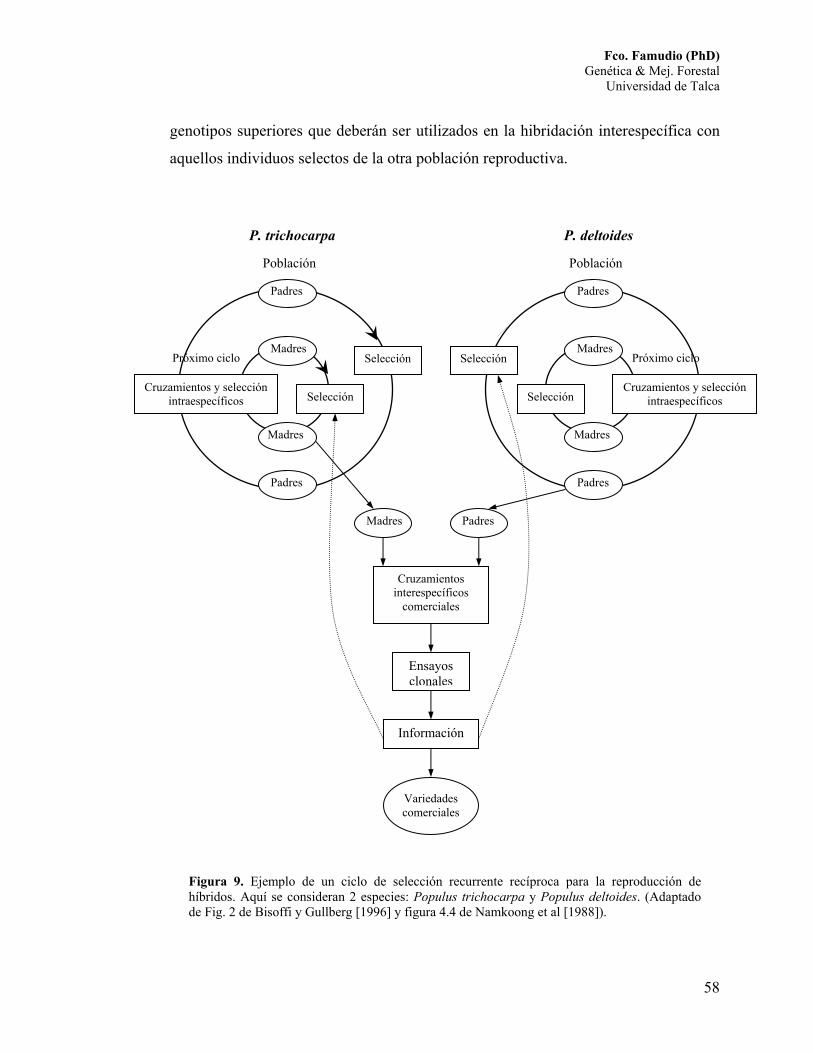
Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
58
genotipos superiores que deberán ser utilizados en la hibridación interespecífica con
aquellos individuos selectos de la otra población reproductiva.
Figura 9. Ejemplo de un ciclo de selección recurrente recíproca para la reproducción de híbridos. Aquí se consideran 2 especies: Populus trichocarpa y Populus deltoides. (Adaptado de Fig. 2 de Bisoffi y Gullberg [1996] y figura 4.4 de Namkoong et al [1988]).
Ensayos clonales
Variedades comerciales
P. trichocarpa P. deltoides
Padres
Selección
Cruzamientos y selección intraespecíficos
Próximo ciclo Madres
Padres
Madres
Selección
Población
Padres
Selección
Cruzamientos y selección intraespecíficos
Próximo ciclo Madres
Padres
Madres
Selección
Población
Madres Padres
Cruzamientos interespecíficos
comerciales
Información

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
59
Cuando se aplica este sistema a una especie, las dos o más subpoblaciones pueden
estar completamente aisladas entre ellas o conectadas a través de cruzamientos
selectivos (migración). El material comercial puede ser originado desde cada una de
las poblaciones simples y/o desde los híbridos producidos por los cruzamientos entre
las poblaciones simples. La selección dentro de cada población está generalmente
orientada a mejorar caracteres específicos de cada una. Los cruzamientos
intraespecíficos (entre las poblaciones simples) tiene como objetivo generar linajes
con características presentes en ambas (o más) poblaciones parentales. Aquí se busca
optimizar el uso de los efectos aditivos de genes presentes en ambas (o más)
poblaciones.
6.3.3 Sistema de reproducción selectiva de poblaciones replicadas
Una forma alternativa de manejo de poblaciones múltiples es la subdivisión de una
población reproductiva simple y grande en unidades independientes pequeñas. La
asignación inicial de los individuos miembros a cada grupo es arbitraria. La
reproducción selectiva dentro de cada subpoblación es mediante algún procedimiento
de selección recurrente y con el objetivo de mejorar los mismos caracteres.
Mediante la selección en subpoblaciones menores, se debe esperar que la
variación en la efectividad de la selección genere una variación interpoblacional en la
ganancia genética. Aunque el promedio de las subpoblaciones podría ser algo menor
que el progreso esperado de una sola población reproductiva simple, el mejorador
podría seleccionar las mejores subpoblaciones para lograr un mayor productividad
(Namkoong et al 1988). Además, el intercruzamiento entre estas subpoblaciones
seleccionadas, los beneficios de la recombinación de diferentes alelos de efecto
positivo podrían ser recuperados para el mejoramiento continuo de generaciones más
avanzadas.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
60
El método de poblaciones replicadas hace que el mejoramiento sea algo más
fácil de efectuar. Además, permite que se realicen ajustes a causa de que las
poblaciones múltiples pequeñas pueden ser más fáciles de manejar que una sola
población grande. En efecto, en las generaciones tempranas de mejoramiento
genético, cuando se puede tolerar un cierto grado de incertidumbre respecto a la
estructura final de la población reproductiva es mayor, el sistema de poblaciones
subdivididas entrega mayor flexibilidad para el manejo de la población reproductiva
total. Esto es esencialmente lo que Lowe y van Buijtenen (1984) llamaron sublineas.
En cualquier generación de selección, la división de la población reproductiva
en sublineas tiene la ventaja de controlar la consanguinidad en la población
reproductiva total. Cierto grado de consanguinidad es permitido dentro de cada
subpoblación, porque se realizan cruzamientos principalmente dentro de cada una de
ellas. Mientras que la consanguinidad entre las subpoblaciones es mantenida a un
mínimo. Esta situación se presenta en el caso que las sublineas presenten algún grado
de jerarquización, puesto que se permiten cruzamientos muy específicos, sólo entre
los individuos elite de cada una (ver por ejemplo White et al [1999]; White y Hodge
[1992]; Cotterill et al [1988]).
La principal utilidad de las sublineas es que la consanguinidad es evitada en
las poblaciones productoras de semilla, tales como huertos semilleros (Namkoong et
al 1988).

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
61
CCaappííttuulloo 77
SSEELLEECCCCIIÓÓNN CCLLOONNAALL
LLa selección clonal es una parte esencial en un programa de mejoramiento de álamo.
En su forma más simple, se comienza con un material abundante y genéticamente
variado y se termina con un número limitado de cultivares clonales. Estos últimos son
establecidos comercialmente en gran escala. El origen del material es obviamente
importante para el éxito, pero no tiene relación con la mecánica del proceso selectivo.
El punto de inicio puede ser variado. Los clones ensayados pueden provenir
de semilla originada de polinización abierta o controlada, originada a partir de padres
selectos (generaciones avanzadas de mejoramiento) o no seleccionados, los cuales
pertenecen a poblaciones naturales (sin mejoramiento) o manejadas (rodales
comerciales). En este último caso, la selección clonal se desarrolla sin una estrategia
de mejoramiento genético, es decir, sin un método de selección (ver sección 3) y una
estructura de poblaciones (ver sección 4). También puede ocurrir que un programa de
selección clonal comience utilizando colecciones de clones ya establecidos. Este es el

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
62
caso si el objetivo es seleccionar clones para un nuevo ambiente y se utiliza, o ensaya,
clones que fueron seleccionados en otra región.
Aunque clones seleccionados podrían ser también valiosos como padres para
la reproducción selectiva de generaciones avanzadas, se debe recordar que las metas
económicas inmediatas son el principal motivo de la selección clonal. Una forma
adecuada de proceder consiste en que la selección clonal con fines económicos sea
hecha entre híbridos F1, mientras que se establecen poblaciones reproductivas
(orientadas a un mejoramiento de largo plazo) para cada una de las especies
parentales. Luego la selección de padres selectos, los que serán utilizados en
hibridación y generación de clones candidatos, se realiza dentro de cada población
reproductiva.
Bisoffi y Gullberg (1996) sugieren el siguiente criterio para la selección clonal
de álamo. Ellos señalan que aunque tienen más se 60 años de aplicación, aun es
largamente aplicable:
1. rápido crecimiento juvenil;
2. adaptabilidad amplia;
3. resistencia a enfermedades y el frío;
4. fuste recto y cilíndrico;
5. madera blanca, homogénea y resistente, adecuada para tableros,
partículas, debobinado y pulpa
El denominador común en todos los programas de reproducción selectiva es que
la productividad se incrementa mediante un mejoramiento de la tasa de crecimiento y
una reducción en las perdidas.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
63
Esto es equivalente a decir que la meta estratégica del mejoramiento genético
forestal es obtener una ganancia en cantidad y calidad de los productos
deseados tan rápido y tan eficiente como sea posible.
Luego, la tasa de crecimiento y la resistencia a factores adversos son los tipos de
caracteres más importantes de la reproducción selectiva (Thielges 1985).
Como se mencionó anteriormente, el mejoramiento genético debe generar una
ganancia genética rápida y eficiente, pero la base genética que sustenta el programa
de reproducción selectiva no debe ser reducida, por el contrario, ésta debe mantenerse
o ampliarse de una generación de selección a la siguiente. De este modo, se
maximizará la capacidad de respuesta a factores o situaciones externas imprevistas y
relacionadas con:
1. la presencia de nuevas plagas y enfermedades forestales;
2. los cambios en los requerimientos o la demanda del mercado;
3. los cambios en las tecnologías utilizadas en el procesamiento de la
madera;
4. las variaciones climáticas imprevisibles; etc.
Una estrategia de mejoramiento que tiene como resultado la selección de híbridos
interespecíficos, los que serán propagados en forma clonal y utilizados con fines
comerciales, resulta complicada porque debe incluir el desarrollo de una población
reproductiva para cada especie involucrada. Luego, el programa de reproducción
selectiva deberá ser dinámico y flexible en el tiempo. En efecto, será adecuado
realizar una revisión periódica de la estrategia, en la cual se incluya un análisis de los
factores externos imprevistos (arriba mencionados) y la modificación de las
actividades previamente incluidas, o la incorporación de nuevas actividades.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
64
Para organismos de larga vida, los cuales son normalmente cosechados al fin
de la rotación (en contraste con árboles frutales) y que, debido a su tamaño, necesitan
de mucho espacio por árbol, los siguientes aspectos son inclusive más cruciales que
para los cultivos agrícolas:
1. la elección del método de selección óptimo,
2. el estudio de las correlaciones juvenil-maduro,
3. el estudio de la interacción genotipo-ambiente y
4. el diseño experimental utilizado en los ensayos de campo
En una selección clonal genéticamente eficiente, una atención especial se debe
poner en aquellos caracteres que poseen los siguientes atributos:
1. un alto grado de estabilidad a través de la edad (por ejemplo:
resistencia a enfermedades) o
2. una importancia comercial relevante a una edad temprana.
Un aspecto de importancia relevante es que la selección debe comenzar en el
nivel del vivero, debido al limitado espacio necesitado por las plantas a este estado de
desarrollo.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
65
BBiibblliiooggrraaffííaa CCiittaaddaa
- Allard, R. W. 1960. Principles of plant breeding. John Wiley & Sons Inc. NY.
498 p.
- Barnes, R.D., Burley, J., Gibson, G.L., and Garcia de Leon, J.P. 1984.
Genotype-Environment interactions in tropical pines and their effects on the
structure of breeding populations. Silvae Genetica. 33 (6): 186-197.
- Bisoffi, S.; Gullberg, U. 1996. Poplar breeding and selection strategies. In:
Biology of Populus and its implications for management and conservation. Part I,
Chapter 6. Edted by R.F. Stettler, H.D. Bradshaw, Jr., P.E. Heilman, and T.M.
Hinckley. NRC Research Press, National Research Council of Canada, Ottawa,
ON. pp. 139-158.
- Bradshaw, H.D. Jr.; Stettler, R.F. 1993. Molecular genetics of growth and
development in Populus. I. Triploidy in hybrid poplars. Theor. Appl. Genet. 89:
301-307.
- Bradshaw, H.D. Jr.; Stettler, R.F. 1995. Molecular genetics of growth and
development in Populus. IV. Mapping QTLs with large effects on growth, form,
and phenology traits in a forest tree. Genetics. 139: 963-973.
- Burdon, R.D.; Low, C.B. 1992. Genetic survey of Pinus radiata. 6: Wood
properties: variation, heritabilities, and interrelationships with other traits.
N.Z.J.For Sci. 22: 228-245.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
66
- Cain, N.P.; Ormond, D.P. 1984. Hybrid vigor as indicated by early growth
characteristics of Populus deltoides, P. nigra, and P. x euroamericana. Can. J.
Bot. 62: 1-8.
- Cotterill, P.P.; Dean, C.; Cameron, J.; Brindbergs, M. 1989. Nucleus
breeding: a new strategy for rapid improvement under clonal forestry. In:
Breeding tropical trees: Population structure and genetic improvement strategies
in clonal and seedling forestry. Pro. IUFRO Conference. Pattaye, Thailand, Nov.
1988. Pp 39-51.
- Eckenwalder, J.E. 1984. Natural intersectional hybridization between North
American species of Populus (Salicaceae) in sections Aigeiros and Tacamahaca.
II. Taxonomy. Can. J. Bot. 62: 325-335.
- Screiner, E.J. 1970. Genetics of eastern cottonwood. U.S. Dep. Agric. For. Ser.
Res. Pap. WO-11. 24 pp.
- FAO. 1979. Poplars and willows. FAO Forestry Series. No 10. Food and
Agriculture Organization of the nited Nations, 1979, Rome, Italy. 328 pp.
- Falconer, D.S.; Mackay, T.F.C.1996. Introduction to quantitative genetics. 4rd Ed.
Longman Sci. And Tech. Harlow, UK. 438 pp.
- Fillatti, J.J.; Haissig, B.; McCown, B.; Comai, L. Reimenschneider, D. 1988.
Development of glyphosate-tolerant Populus plants through expression of a
mutant aroA gene from Salmonella typhimurium. In: Genetic manipulation of
woody plants. Ed. By J.W. Hanover and D.E. Keathley. Plenum Press, New York
and London. Pp 243-249
- Fowler, D.P. 1978. Population improvement and hybridization. Unasylva 30: 21-
26
- Han, K. H.; Gordon, M.P.; Strauss, S.H. 1996. Cellular and molecular biology
of Agrobacterium-mediated transformation of plants and its application to genetic
transformation of Populus. In: Biology of Populus and its implications for
management and conservation. Part I, Chapter 9. Edited by R.F. Stettler, H.D.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
67
Bradshaw, Jr., P.E. Heilman, and T.M. Hinckley. NRC Research Press, National
Research Council of Canada, Ottawa, ON. pp. 201-222.
- Kang, H.; Nienstaedt, H. 1987. Managing long-term breeding stock. Silvae
genetica 36: 30-39.
- Kang, M.; Gauch, H. G. 1996. Genotype-by-environment interaction. CRC
Press. Boca Raton. 416 pp.
- Lowe, W.J.; van Buijtenen, J.P. 1981. Tree improvement philosophy and
strategy for the western Gulf Tree Improvement Program. In: Proc. 15th North
Am. Quant. For. Gen. Workshop, pp 43-50.
- Michler, C.H.; Haissig, B.E. 1988. Increased herbicide tolerance of in vitro
selected hybrid poplars. In somatic cell genetics of woody plants. Ed. By M.R.
Ahuja. Kluwer Ac. Pub., Dordrecht, The Netherlands, pp 183-189.
- Namkoong, G.; Snyder, E.B.; Stonecypher, R.W. 1966. Heritability and gain
concepts for evaluating breeding systems such as seedling orchards. Silvae
genetica 15 (3): 76-84.
- Namkoong, G.; Kang, H.C.; Brouard, J.S. 1988. Tree breeding: Principles and
strategies. Monograph on theoretical and applied genetics 11. Springer-Verlag.
New York. 180 p.
- Orlovic, S.; Guzina, V.; Krstic, B.; Merkulov, L. 1998. Genetic variability in
anatomical, physiological and growth characteristics of hybrid poplar (Populus x
euramericana Dode (Guinier)) and eastern cottonwood (Populus deltoides Bartr.)
clones. Silvae Genetica 47 (4): 183-190.
- Shelbourne, C.J.A.; Burdon, R.D.; Carson, S.D.; Firth, A.; Vincent, T.G.
1986. Development plan for radiata pine breeding. New Zealand Forest Service.,
Rotorua, NZ. 142 pp.
- Shelbourne, C.J.A. 1969. Tree breeding methods. New Zealand Forest Service.
Technical paper N° 55. 43 pp.

Fco. Famudio (PhD) Genética & Mej. Forestal
Universidad de Talca
68
- Stettler R.F.; Fenn, R.C.; Heilman, P.E.; Stanton, B.J. 1988. Populus
trichocarpa x P. deltoides hybrids for short rotation culture: variation patterns and
4-year field performance. Can. J. For. Res. 18: 745-753.
- Stettler, R.F.; Zsuffa, L.; Wu, R. 1996. The role of hybridization in the genetic
manipulation of Populus. Part I, Chapter 4. Edited by R.F. Stettler, H.D.
Bradshaw, Jr., P.E. Heilman, and T.M. Hinckley. NRC Research Press, National
Research Council of Canada, Ottawa, ON. pp. 87-112.
- Thielges, B.A. 1985. Breeding poplars for disease resistance. FAO Forestry Paper
N° 56. Rome, Italy. 66 pp.
- van Buijtenen, J.P. 1975. Advanced generation breeding. Proc. of 13th Southern
Forest Tree Improvement Conference. Raleigh, NC, pp 63-72
- White, T.L.; Hodge, G.R. 1989. Predicting breeding values with applications in
forest tree improvement. Kluwer Ac. Pub. Dordrecht, The Netherlands. 367 p.
- White, T.L.; Hodge, G.R.; Powell, G.L. 1993. An advanced-generation tree
improvement plan for slash pine in the southeastern United States. Silvae genetica
42 (6): 359-371.
- White, T.L.; Matheson, A.C.; Cotterill, P.P.; Jhonson, R.G.; Rout, A.F.;
Boosma, D.B. 1999. A nucleus breeding plan for radiata pine in Australia. Silvae
genetica (3/4): 122-133.
- Zobel, B. Talbert, J. 1984. Applied forest tree improvement. John Wiley &
Sons. NY. 505 pp.
- Zobel, B.J.; Jett, J.B. 1995 Genetic if wood formation. Springer-Verlag. 337 pp.
- Zsuffa, L.; Giordano, E.; Pryor, L.D.; Stettler, R.F. 1996. Trends in poplar
culture: some global and regional perspectives. In: Biology of Populus and its
implications for management and conservation. Part II, Chapter 19. Edted by R.F.
Stettler, H.D. Bradshaw, Jr., P.E. Heilman, and T.M. Hinckley. NRC Research
Press, National Research Council of Canada, Ottawa, ON. pp. 515-539.





























