Embed Size (px)
Citation preview

UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS Fundada en 1551
FACULTAD DE CIENCIAS BIOLÓGICAS
E.A.P. DE CIENCIAS BIOLÓGICAS
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA”
TESIS :
Para optar el Título Profesional de :
BIÓLOGO
Con mención en :
GENÉTICA
AUTOR
Ángela Renee Arias Ramírez
LIMA – PERÚ
2002

Este trabajo se realizó en el Laboratorio de Recursos Genéticos y
Biotecnología de la Facultad de Ciencias Biológicas de la Universidad
Nacional Mayor de San Marcos y en el Laboratorio del Dr. César
Fuertes Ruitón en el Instituto de Química Orgánica Aplicada a la
Farmacia de la Facultad de Farmacia y Bioquímica de la Universidad
Nacional Mayor de San Marcos.

AGRADECIMIENTOS
Al profesor Rolando Estrada, por todo el apoyo y cariño que me brindó
mientras estuve en el LRGB.
Al Dr. César Fuertes, investigador del Instituto de Química Orgánica
Aplicada a la Farmacia, de la Facultad de Farmacia y Bioquímica sin
cuya asesoría, apoyo y enseñanzas este trabajo no hubiera podido
llevarse a cabo.
A mis padres, Juan y Hortensia por su amor, apoyo y paciencia.
A mis hermanos Marina, Nina, Juana. Lena, Paul y Eva por su amor
y apoyo incondicional.
A Juan Diego y Alessia que vinieron a iluminar nuestras vidas y
llenarlas de esperanza y alegría.
A mi mamá Eloy, a mi mamá Jessi y a mi papá Cesar, y a mis primos
aunque sería más adecuado decir hermanos.
A Verónica, Delcy y Enrique, por enseñarme a querer y a sentirme
querida como nunca antes y a Luchito Alanya con todo mi cariño.

A Mary Gálvez y Judith Toledo, mis maestras y amigas a las que
siempre llevo en el corazón y a Miles por su amistad sincera y su
entusiasmo contagiante.
A mis amigos de la Facultad de Ciencias Biológicas, en especial a
Nancy, Maggi, Miriam, María Esther, Ingrid, al profesor Mariano, a
la profesora Caleen Távara, y a todos los amigos que conocí en esta
casa de estudios y que no puedo enumerar por razones de espacio.
A mis amigos del LRGB, los “dinosaurios” César, Fredy, Rafo, Pepe,
Flor, Margarita, y Augusto que me hicieron sentir bienvenida y en mi
segundo hogar, a los que siguieron llegando: Héctor, Indira, Mariellix,
Alberto, Elisa, Gilmar, Roberto, Gerardo, Nancy, las profesoras
Vidalina y Yolanda para compartir preocupaciones y alegrías, los
chicos que hoy están empezando su camino en especial a Arturo, Alex,
Ivett y Joel .

INDICE
I.- RESÚMEN I.1.- ABSTRACT II.- INTRODUCCIÓN II.1.- OBJETIVOS
II.1.1.- GENERALES
II.1.2.- ESPECÍFICOS
III.- ANTECEDENTES
III.1.- DESCRIPCIÓN DE LA PLANTA
III.1.1.- CLASIFICACIÓN TAXONÓMICA
III.1. 2.- DESCRIPCIÓN BOTÁNICA
III.1.3.- NÚMERO DE CROMOSOMAS
III.1.4.- DISTRIBUCIÓN GEOGRÁFICA
III.1.5.- IMPORTANCIA NUTRICIONAL
III.2.- METABOLITOS SECUNDARIOS
III.2.1.- GLUCOSINOLATOS
III.2.1.1.- EL SISTEMA MIROSINASA – GLUCOSINOLATO
III.2.2.- ALCALOIDES
III.3.- CULTIVO DE TEJIDOS
III.3.1.- CULTIVO DE CALLOS
III.3.2.- HORMONAS Y REGULADORES DE CRECIMIENTO
III.3.2.1- AUXINAS
III.3.2.2.- CITOCININAS
III.3.3.- EL EXPLANTE
III.3.4.- CULTIVO DE TEJIDOS Y METABOLITOS
SECUNDARIOS
III.4.- CROMATOGRAFÍA
III.5.- ELECTROFORESIS EN GEL
IV.- MATERIAL Y MÉTODOS
IV.1.- INDUCCIÓN DE CALLOS
IV.1.1.- MATERIAL VEGETAL

IV.1.1.1.- DESINFECCIÓN
IV.1.1.2.- CONDICIONES DE CULTIVO
IV.1.2.- EXPLANTES
IV.1.3.- MEDIO DE CULTIVO PARA
LA INDUCCIÓN DE CALLOS
IV.1.4.- INDUCCIÓN DE CALLOS
IV.2.- DETECCIÓN DE METABOLITOS SECUNDARIOS
IV.2.1.- EXTRACCIÓN Y DETECCIÓN DE GLUCOSINOLATOS
IV.2.2.- EXTRACCIÓN Y DETECCIÓN DE ALCALOIDES
IV.3.- DETECCIÓN DE MIROSINASAS
IV.3.1.- ELECTROFORESIS
IV.3.2.- EXTRACCIÓN DE LAS MUESTRAS
IV.3.3.- DETECCIÓN DE ACTIVIDAD
V.- RESULTADOS
V.1.- INDUCCIÓN DE CALLOS
V.2.- DETECCIÓN DE METABOLITOS SECUNDARIOS
V.2.1.- GLUCOSINOLATOS
V.2.2.- ALCALOIDES
V.3.- DETECCIÓN DE MIROSINASAS
VI.- DISCUSIÓN
VI.1.- INDUCCIÓN DE CALLOS
VI.2.- DETECCIÓN DE METABOLITOS SECUNDARIOS
VI.3.- DETECCIÓN DE MIROSINASAS
VII.- CONCLUSIONES VIII.- RECOMENDACIONES IX.- REFERENCIAS BIBLIOGRÁFICAS X.- APÉNDICE

ABREVIATURAS IAA Ácido indol acético
IBA Ácido indolbutírico
NAA Ácido Naftalenacético
2,4D Ácido 2,4-diclorofenoacético
BAP Bencil amino purina
KIN Kinetina
ZEA Zeatina
2ip 2-isopentenidenina

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO I
RESÚMEN
Lepidium peruvianum Chacón (maca), es una crucífera altoandina, que crece a
entre los 3,500 y 4500 m.s.n.m. Originaria de la meseta del Bombón, en los
departamentos de Junín y Pasco; por sus cualidades medicinales y su alto valor
nutritivo, es una planta de alto interés económico, cuyo cultivo se ha extendido a
otras regiones de nuestro país. En el presente trabajo se estudió la susceptibilidad
de la especie a las técnicas del cultivo de tejidos como herramienta de producción
de metabolitos secundarios. Se realizó la inducción de callos en diferentes
explantes de L. peruvianum utilizando la auxina 2,4-D y la citoquinina Kinetina, en
un factorial de medios con diferentes concentraciones auxina/citoquinina. Se
obtuvieron callos en la mayoría de los medios usados, la relación de hormonas más
eficiente fue 1 µM auxina/citoquinina. Se evaluó la presencia de glucosinolatos y
alcaloides en los callos obtenidos y se compararon con muestras control de
hipocótilos de maca. Se observó la presencia variable de dos fracciones de
glucosinolatos en los callos, en la mayoría de los casos las manchas tuvieron una
coloración más intensa en los callos que en los controles. De otro lado se observó
una alta variabilidad en la presencia de alcaloides y otros metabolitos no
identificados en los callos obtenidos en este trabajo. También se evaluó
cualitativamente la presencia de mirosinasas en los callos obtenidos, observándose
bandas positivas en los callos y las muestras de plantas de maca.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
ABSTRACT
Lepidium peruvianum Chacón, is a Cruciferae native from the Andes. It grows
between 3,500 and 4500 m. Original from the Bombón plateau, located at the
Peruvian localities Junín and Pasco. It became in a crop with a high economical
value, due its medicinal and nutritional properties. Actually, it is extended to other
regions of the country. The main objective of this research is to study the tissue
culture ability of the crop to use in vitro tissues as a tool for secondary metabolite
production.
Leaves, petioles, roots and hypocotils of L. peruvianum were tested as explants to
induce calli. Different concentrations of 2,4-D and Kinetin, in MS basic medium were
tested. Calli were induced in most of the media tested, the most efficient hormone
ratio auxin/citokinin was 1. It was evaluated the presence of glucosionlatos and
alkaloids in the callus induced and compared to maca hypocotils as control sample.
Two glucosinolatos fractions were obtained from calli analyzed. It was found one or
two fractions according to the callus and in most of the cases the concentration was
higher in callus than in control. In the other hand, it was observed a high variability in
the alkaloid fractions and other unidentified metabolites extracted from the calli
evaluated in this work.
It was also evaluated the presence of myrosinases in the calli studied, and it was
found positive bands either in callus as in maca hypocotils.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO II
INTRODUCCIÓN
Lepidium peruvianum Chacón, "maca", es una crucífera calificada como una
de las raíces y tubérculos andinos (RTA) de más alto contenido proteico. Crece
entre los 3,500 y 4500 m.s.n.m (King, 1988). Originaria de la meseta del Bombón,
entre los departamentos de Junín y Pasco, es la única Brassicacea domesticada de
los andes y el único cultivo que reemplaza el cultivo de las papas amargas en esas
altitudes (regiones suni y puna de los departamentos de Junín y Pasco), soportando
condiciones ambientales extremas, como cambios de temperatura de 18° C a - 10°
C en un solo día, fuertes vientos, y poca disponibilidad de agua. La maca es una
pequeña planta arrosetada de hojas postradas, cuya raíz tuberosa es rica en
azúcares, otros carbohidratos, proteínas y minerales esenciales, lo que la convierte
en una excelente fuente alimenticia, siendo utilizada principalmente por las
poblaciones rurales de la sierra central, mientras que fuera de su área de cultivo
tradicional aún no es conocida completamente (Aliaga 1999, National Research
Council. 1989)
En la última década del siglo XX, la maca que se consideraba como un cultivo
en proceso de extinción (Castro de León 1990) ha empezado a cobrar importancia
gracias al redescubrimiento de su alto valor nutritivo, energético y medicinal. Una de
las principales propiedades medicinales atribuidas a la maca es la de potenciador de
la fertilidad, y su acción sobre la conducta sexual, propiedad por la que ya era usada
en el incanato (Johns, 1980).
Se han realizado muchos estudios sobre la actividad fertilizante de la maca,
hallándosele propiedades como potenciador de la fertilidad en ratas (Chacón, 1961 y
Ninanya 1995), propiedades estrogénicas marcadas, (Lamas y Upumayta, 1993),

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
como potenciador de la actividad espermática (Yauri y Valerio, 1991). Estos efectos
están relacionados a la presencia de glucosionlatos aromáticos (Johns, 1980), y de
alcaloides en la planta (Chacón, 1992). Sin embargo los metabolitos de “maca” no
han sido estudiados a profundidad, desconociéndose las propiedades de muchos
de ellos.
Los glucosionlatos han sido descritos desde hace mucho tiempo, junto con las
mirosinasas (las enzimas que los degradan) como un sistema característico del
orden de las Capparales. Este sistema está involucrado en la defensa de la planta
contra insectos y patógenos. Además, los productos de la degradación de los
glucosionlatos están relacionados con efectos negativos en el ser humano, como la
probable inhibición del yodo en la tiroides producida por los tiocianatos, derivados de
la hidrólisis de los glucosionlatos por acción de la mirosinasa (Daxenbichler y colb.,
1964). Pero otro lado, recientemente estos metabolitos han adquirido interés debido
a sus propiedades anticarcinogénicas (Quiros, 1999).
Anteriores estudios realizados en los alcaloides de Lepidium peruvianum Ch.
muestran que estos compuestos tienen efectos estimulantes sobre la fertilidad
femenina (Chacón, 1990). Por lo descrito anteriormente se eligieron pues los
alcaloides y glucosionlatos para evaluar la posibilidad de utilizar el cultivo de tejidos
vegetales para la producción y estudio de los metabolitos secundarios de maca,
debido a que los cultivos de tejidos y suspensiones celulares pueden ser
manipulados para obtener una gran variedad de productos naturales usados en las
industrias de alimentos y farmacéutica. De hecho esta técnica ya ha sido utilizada a
escala industrial en la producción de metabolitos secundarios (Nuñez y Ochoa,
2000).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
II.1.- OBJETIVOS
II.1.1.- GENERALES
• Evaluar la producción de metabolitos secundarios de Lepidium peruvianum
Ch. bajo condiciones de cultivo in vitro.
II.1.2.- ESPECÍFICOS
• Determinar la composición de un medio de cultivo óptimo para la inducción de
callos en diferentes explantes de L. peruvianum Ch.
• Determinar la presencia de glucosinolatos y alcaloides en callos de L.
peruvianum Ch.
• Comparar la presencia de glucosinolatos y alcaloides en callos de L.
peruvianum Ch. con la que se presenta in vivo.
• Determinar la presencia de mirosinasas en callos de L. peruvianum Ch. y
compararla con la que se presenta in vivo.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO III
ANTECEDENTES
III.1.- DESCRIPCIÓN DE LA PLANTA.-
El género Lepidium incluye aproximadamente 175 especies, distribuidas en
prácticamente todos los continentes, excepto la Antártida, siendo uno de los más
grandes dentro de las crucíferas. Las especies de América del Norte, Australia y
Europa han sido estudiadas extensamente, pero se sabe muy poco de las
especies endémicas Andinas (Quiros 1999).
Gloria Chacón (1990) menciona que en el Perú, de acuerdo a Ferreira
(1986), existían 11 especies clasificadas del género Lepidium a la que agrega
Lepidium peruvianum Chacón como una nueva especie.
De acuerdo a Rea (1992, citado por Quiros y colb. 1996) la maca fue
domesticada hace cerca de 2000 años en San Blas, Junín. Siendo, según Bonnier
(1986, citado por Chacón 1990), una de las primeras plantas domesticadas en el
Perú.
La maca parece haber jugado un papel importante en la economía
prehispánica, y durante los primeros cien años de la colonia formó parte de los
tributos exigidos por el Encomendador (Chacón, 1990).
III.1.1.- CLASIFICACIÓN TAXONÓMICA.
DIVISION : ANGIOSPERMAE
CLASE : DICOTYLEDONEAE
SUBCLASE : ARCHICHLAMYDEAE

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
ORDEN : PAPAVERALES
FAMILIA : BRASSICACEAE
GENERO : Lepidium
ESPECIE : Lepidium peruvianum Chacón. Sp. Nov. Antes L.
meyenii Walp.
NOMBRE COMUN : MACA, MACA-MACA, MAINO, AYAK CHICHIRA,
AYAC WILLKU, GINSENG PERUANO.
III.1. 2.- DESCRIPCIÓN BOTÁNICA
La maca, según Aliaga (1988), es una planta de comportamiento bienal
en la sierra alta, (no se tienen referencias respecto a su comportamiento en
menores altitudes) siendo autógama, cleistógama, con una fase reproductiva de
5 meses con una floración que dura dos meses. Las flores se abren en series
sucesivas, presentando cuatro etapas: flor cerrada con anteras sin dehiscencia,
flores cerradas con anteras con dehiscencia, flor abierta con estructuras florales
en pleno desarrollo, flor abierta con estructuras florales entrando en
senescencia. Esta fase reproductiva es precedida por una fase vegetativa de 8
meses, que se inicia con la siembra de la semilla, en la que se produce el
ensanchamiento de los hipocótilos. El cultivo de estas dos fases se da
generalmente entre los meses de setiembre y junio en dos años consecutivos
(Aliaga, 1999).
La raíz es napiforme (Beltrán y colb. Citado por Obregón, 1998), descrita
como tubérculo hipocotíleo por Piñas (1994) recibiendo también el nombre de
hipocótilo (Aliaga 1995, 1997; Tello y colb. 1992; Quiros, 1996), según León
(1964), éste es un eje carnoso derivado del hipocótilo cuya parte superior

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
termina en una superficie plana de la que brotan de 6 a 15 hojas; mientras la
inferior es cónica y se alarga en una raíz ancha y fuerte con dos áreas
irregulares en la mitad inferior, de donde nacen las raicillas.
Los colores de maca más frecuentes son: amarillo, morado, medio
morado (la parte superior morada y la inferior blanca) y negro (León, 1964) pero
se pueden encontrar también maca de color rosado, crema-morado, amarillo-
negro, plomo, y raras veces morado-negro (Aliaga, 1999), el color preferido por
los campesinos y el mercado es el color amarillo (León, 1964 y Aliaga, 1999).
El tallo es acaule, mientras las hojas son arrosetadas, pecioladas,
compuestas bipinnatisectas, con foliolos opuestos y dimórficas: En la fase
vegetativa son grandes (10 – 15 cm de largo) y muy reducidas (menos de 5 cm)
en la fase reproductiva (Tello y colb. 1992 y Aliaga, 1999).
La inflorescencia es un racimo compuesto, raramente simple (Piñas
1994), con el eje floral corto. Las flores, pequeñas y blancas, son perfectas, con
cuatro sépalos, cuatro pétalos, 2 estambres fértiles y cuatro estaminodios y un
ovario súpero bicarpelar y bilocular, con un óvulo en cada lóculo de
placentación axilar. Su fórmula floral es: K4C4A2-4G2 (León, 1964 y Aliaga,
1995).
El fruto es una silícula de dehiscencia longitudinal con 2 semillas ovoides
de 2-3 mm de ancho y 4 – 5 mm de largo (Beltrán y colb. Citado por Obregón,
1998) el color de las semillas varía del amarillo-naranja al marrón oscuro (Aliaga,
1999).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
III.1.3.- NÚMERO DE CROMOSOMAS
Tello y colb. (1992) sugirieron que la maca era un hexaploide de n = 8
con 54 – 56 cromosomas, pero en 1996, Quiros y colb. observaron, en células
madres de plantas en floración, 32 pares de cromosomas; concluyendo que la
maca es un poliploide disómico del número cromosómico básico del género: n
= 8, siendo así un octoploide: 2n x 8 = 64 cromosomas.
III.1.4.- DISTRIBUCIÓN GEOGRÁFICA
A pesar que la maca crece a altitudes mayores de los 3500 m.s.n.m e
inclusive en regiones en las que ni siquiera las papas amargas pueden
sobrevivir, altitudes que se encuentran a lo largo de la Cordillera de los Andes
(National Research Council. 1989); el cultivo de la maca (luego de la colonia),
estuvo restringido a los departamentos de Junín y Pasco, en las regiones suni y
puna, principalmente en las laderas circundantes al lago de Junín; a altitudes
de 4100 – 4450 m.s.n.m. (Tello 1992); y en menor grado en otras localidades de
estos departamentos desde los 3400 m.s.n.m.
Los datos de los cronistas y las memorias de tributos, sugieren que la
distribución del cultivo era amplia en los territorios altoandinos, pero el área de
cultivo de la planta disminuyó progresivamente hasta que entre 1777 y 1778,
Hipólito Ruiz lo circunscribe la región de Chinchaycocha (Castro, 1990). Esta
tendencia se mantuvo hasta la década de los ochenta, en la que el área de
cultivo de la planta era de apenas 25 Has (Aliaga, 1999). En la década de los 90
esta tendencia se revirtió al crecer la demanda de la planta, y su cultivo se
extendió a otros departamentos del Perú, de tal manera que actualmente la
maca se está cultivando en Puno, Ayacucho, Huancavelica, Huánuco y
Apurímac (Aliaga, 1999).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
III.1.5.- IMPORTANCIA NUTRICIONAL
La maca es la única especie del género Lepidium cuya raíz es utilizada
como alimento, aunque las hojas de otras especies son usadas como verduras
(National Research Council. 1989), además, dentro de los tubérculos y raíces
andinos, es una de las especies con mayor valor alimenticio; con un alto
contenido de aminoácidos esenciales, comparables con los recomendados por
la FAO – OMS, siendo altos sus valores de lisina, tirosina y fenilalanina. La
fracción grasa es rica en ácido linoleico utilizado por el organismo para la
biosíntesis de ácidos grasos esenciales. Por otro lado, los niveles de minerales
de la planta son altos comparados con otras especies (Pizza y colb. 1998). (Ver
apéndice)
III.2.- METABOLITOS SECUNDARIOS.
Se definen como metabolitos secundarios a aquellos compuestos que no
tienen una función reconocida en el mantenimiento de los procesos fisiológicos
fundamentales de los organismos que los sintetizan (Robert y colb. 1991).
Entre las funciones que cumplen estos metabolitos están: proteger la planta
de los depredadores, competir ventajosamente con otras plantas, atraer
polinizadores y simbiontes y brindarles protección contra diferentes tipos de estrés a
los que se vean sometidas (Loyola y Vargas, 1985, citados por Robert y colb. 1991).
Las evidencias de una estricta regulación metabólica de estos productos nos
indicaría que cumplen un rol importante en la sobrevivencia de las plantas

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
III.2.1.- GLUCOSINOLATOS.
Los glucosinolatos son una clase tioglucósidos restringidos a las
dicotiledóneas limitados a pocas familias y en algunos casos a ciertos géneros y
especies (Kjaer, 1973), los que son almacenados en diferentes tejidos de la
planta, siendo una fuente significativa del contenido de azufre y minerales de la
planta (Josefsson, 1970; citado por Rodman, 1981), se encuentran en muchos
cultivos comerciales y junto con la enzima que los degrada, la mirosinasa,
conforman un sistema que define fitoquímicamente al orden de las Capparales
(Hegnauer, 1986; citado por Bones y Rossiter, 1996); estando presente en
muchas brassicaceas económicamente importantes, como la mostaza,
calabazas, brócoli, nabo, etc. El daño mecánico, infecciones o ataque de pestes
produce, como una respuesta de defensa de la planta, el rompimiento celular
que exponen los glucosinolatos almacenados a la acción de las enzimas que los
degradan, las mirosinasas, produciendo isotiocianatos, nitrilos,
cianoepitioalcanos y tiocinatos dependiendo del pH y/u otros factores (Fig 1)
(Bones y Rossiter, 1996). Estos compuestos disminuyen la palatibilidad de las
hojas para herbívoros no específicos como aves y babosas, pero aumenta la
susceptibilidad del tejido a insectos específicos como los escarabajos alados,
(Mithen y col. 1995)
La primera revisión de la presencia de glucosinolatos en varias especies
fue hecha por Kjaer en 1960, con actualizaciones periódicas (Xenophon y colb.,
1981), en 1973, Kjaer puntualizó: “entre el vasto número de productos
secundarios vegetales, aquellos que presentan una distribución discontinua junto
con un llamado inmediato a los sentidos humanos (olor, sabor, color) han
adquirido, por razones obvias, una posición prominente en discusiones acerca
de los méritos de tales productos para propósitos de clasificación y, más
fundamentalmente, consideraciones filogenéticas. Los glucosinolatos ........

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
constituyen un grupo con algunas de las características anteriores.”....... “La
complejidad sistemática de numerosos taxa productores de glucosinolatos, por
ejemplo dentro de Cruciferae, es un reto para nuestro entendimiento de la
posición biológica e importancia de este grupo de constituyentes vegetales
químicamente bien definido y en constante crecimiento”.
Hace ya varias décadas, se ha deseado reducir el contenido de
tioglucósidos en Brassicáceas mediante métodos de cultivo, pues la presencia
de algunos de ellos son indeseables por algunos efectos tóxicos de los
productos de su degradación (Josefsson, 1967), tales como nitrilos,
isotiocianatos, tiocianatos, epitionitrilos y vinil oxazolidinetionas (Bones y
Rossiter, 1996), así como por su sabor; mientras que otros vienen siendo
estudiados por sus propiedades terapéuticas. Actualmente, la concentración de
glucosinolatos puede ser manipulada genéticamente, disminuyéndola para
mejorar la palatabilidad del cultivo, como en el caso de la colza, o,
aumentándola como se trata de hacer en brócoli con el glucosinolato
glucorafanina, que al hidrolizarse forma sulfurofano con actividad
anticancerígena (Quiros, 1999).
Las especies del género Lepidium son usualmente ricas en bencil
isotiocianatos (Daxenbichler y colb., 1964) y en el caso de la maca, Jonhs
(1981), sugiere que las propiedades potenciadoras de la fertilidad se deben a los
productos de los glucosinolatos de maca, los isotiocianatos aromáticos,
bencilisotiocianato y el p-metoxibencil isotiocianato, el último de los cuales
también se encuentra en mashua.
Químicamente, los glucosinolatos son compuestos hidrosolubles, tienen
el mismo núcleo el cual consiste en un tioglucósido unido al carbón de una
oxima sulfonada y un grupo funcional R que deriva de aminoácidos siendo éste

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
el carácter variable (Bones y Rossiter, 1996) y de cuya estructura depende la
actividad biológica del glucosinolato al determinar la naturaleza de los productos
resultantes del daño de los tejidos de la planta, (Mithen y colb., 1995). La
naturaleza de este grupo R los divide en dos grupos: los glucosinolatos
aromáticos (alquil, alquenil, hidroxialquenil, w- metiltioalkenil) y los alifáticos
(bencil, bencil sustituido), el grupo sulfato le imparte fuertes propiedades ácidas,
por lo que los glucosinolatos se presentan como aniones contrabalanceados por
un catión, la glucosa y sus formas aniónicas, los hacen compuestos no volátiles
e hidrofílicos, se han identificado más de 100 glucosinolatos; todos ellos se
diferencian sólo en la cadena lateral R. (Bones y Rossiter, 1996). Por el gran
número de glucosinolatos, se usan nombres semisistemáticos, en los que el
nombre de la cadena R es seguido por el sufijo “glucosinolato”,
Los glucosinolatos provienen del metabolismo de los α- aminoácidos,
Valina, Alanina, Leucina, Isoleucina, Fenilalanina, Tirosina y Triptofano (Kjaer,
1973), en una serie de reacciones en las que el grupo carboxilo se pierde y el
carbón α se transforma en el carbón central del glucosinolato (Larsen, 1981),
aunque solo siete corresponden a aminoácidos proteicos (antes mencionados),
los restantes derivan de modificaciones de la cadena lateral, que probablemente
a nivel de glucosinolato o derivan de aminoácidos no proteicos (Chapple, y colb.,
1994 y Larsen, 1981).
Para el aislamiento y análisis de los glucosinolatos se han desarrollado
muchas técnicas, como la cromatografía en papel Kjaer 1970, cromatografía de
capa fina, cromatografía de gases, además de procesos que involucra el estudio
de los productos de la degradación enzimática, aunque la gran variabilidad de
los productos de la degradación enzimática pueden causar resultados erróneos,
(Larsen, 1981).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
El tratamiento de los glucosinolatos con AgNO3 o Hg(Oac)2 resulta en la
producción de α-D-glucosa y derivados de la plata o el mercurio los que
nuevamente pueden ser descompuestos en nitrilos, (Larsen, 1981)
Los glucosinolatos se encuentran usualmente en toda la planta en
vacuolas celulares, pero presumiblemente en las semillas maduras y dormantes
ricas en glucosinolatos y que presumiblemente no presentan vacuolas acuosas
aún no se ha definido su localización celular (Rodman, 1981).
Lockwood y Belkhiri, (1991), revisaron la clasificación de crucíferas de
Argelia, de acuerdo a sus glucosinolatos, reclasificando el género Isatis y
reubicando el género Sinapsis fuera de Brassicaria, esto demuestra pues, el
valor taxonómico de estos compuestos.
III.2.1.1.- EL SISTEMA MIROSINASA – GLUCOSINOLATO
Las enzimas que hidrolizan los glucosinolatos son las mirosinasas y estas
están invariablemente depositadas en toda la naturaleza con el sustrato. (Kjaer,
1973), así, sin excepción conocida, los glucosinolatos están acompañados en la
planta por esta enzima. Las mirosinasas se encuentran en idioblastos
especializados ricos en proteínas llamados células de mirosina (Rodman, 1981)
que se encuentran en el tejido parenquimático (Bonnes y Rossiter, 1996), éstos
se tiñen de rojo intenso con el reactivo de Millon. (Fahn, 1979; citado por
Rodman, 1981) de tal manera que los dos componentes se encuentran
separados hasta que se produce daño tisular o autolisis, generando durante la
masticación los productos de defensa de la planta (Purrington, 2000).
Este sistema fue observado por primera vez en semillas de mostaza,
(Kjaer, 1968) posteriormente fue encontrado en todas las Brassicáceas, y en la

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
familias Caparaceae, Tropaeolaceae, Caricaceae, entre otras. Por otro lado se
ha encontrado en los hongos Aspergillus sydowi y Aspergillus niger , en la
bacteria Enterobacter cloacae, en tejidos de mamíferos y en los áfidos que
atacan crucíferas Lipaphis erisimi y Brevicoryne brassicae. Pero estas enzimas
parecen no estar relacionadas con las enzimas de las plantas (Bones y Rossiter,
1996).
S – Glu
R – C
NOSO3
H2O mirosinasa
S
R –C + D - Glu
NOSO3
Isotiocianatos nitrilos cianoepitioalcanos tiocianato
FIG. 1.- Hidrólisis de los glucosinolatos por la mirosinasa. (Fuente: Bonnes y
Rossiter. 1996. The Myrosinase – Glucosinolate System, its Organization and
Biochemistry. Physiologia Plantarum. 97:194 -208 )
La degradación de los glucosinolatos mediada por la mirosinasa puede
producir: a) isocianatos, cuando la hidrólisis se produce en un pH normal; b)
nitrilos, cuando la hidrólisis ocurre a pH ácido; c) cianoepitioalcanos, cuando la
mirosinasa se presenta con una pequeña proteína conocida como proteína
epitioespecificadora; d) tiocianatos, que son producidos solo por tres de los
glucosionatos que se dan en la naturaleza, los allyl, bencil, y 4-
(metiltio)butilglucosinolatos.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
MacGibbon y Allison en 1970 detectaron por electroforesis en gel
separando varias isoenzimas de mirosinasas, ellos trabajaron con 7 especies de
Rhoeadales, cada especie tenia un patrón diferente con 1 a 4 bandas. También
encontraron que este patrón variaba de acuerdo a la parte de la planta de la cual
se hicieran los extractos.
Diferentes estudios de las mirosinasas demuestran que el patrón de estas
enzimas puede variar no solo de acuerdo a la especie, sino también de acuerdo
al órgano y edad de la planta, pero se sabe poco de la razón fisiológica de esta
diferencia.(Bonnes y Rossiter, 1996). Se ha postulado que las isoenzimas
particulares corresponden a condiciones endógenas encontradas en las plantas,
o al organismo blanco o a los glucosinolatos que predominan en el
tejido.(Bonnes y Rossiter, 1996).
III.2.2.- ALCALOIDES
Los alcaloides forman una amplia y variada familia de metabolitos
secundarios de moléculas no relacionadas entre sí, siendo de gran importancia
económica debido a sus propiedades farmacológicas, las mismas que han sido
usadas desde la prehistoria hasta la actualidad. Son los metabolitos mas
frecuentes en el reino vegetal, habiéndose encontrado cerca de 10,000
alcaloides en aproximadamente 20 por ciento de plantas con flores,
principalmente dicotiledóneas herbáceas (Hopkins, 1999).
Se clasifican como alcaloides a aquellas sustancias básicas con uno o
más átomos de nitrógeno en su sistema cíclico, que manifiestan actividad
farmacológica (Lock, 1994)
La mayoría de los alcaloides provienen del metabolismo de los
aminoácidos, principalmente tirosina, triptofano, arginina y lisina, y aunque

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
algunos alcaloides se encuentran en varios géneros o aún en una familia, la
mayoría de las especies presentan un patrón único, genéticamente
determinado. Por otro lado su distribución está restringida a determinados
órganos de la planta, como raíces, hojas o frutos jóvenes (Hopkins, 1999).
No se ha determinado aún cual es la función de los alcaloides en la
planta, aunque se le asignan rol defensivo, debido a que la mayoría son
amargos, lo cual es considerado universalmente como un carácter repelente.
Todos son biológicamente activos, muchos significativamente tóxicos y otros con
propiedades antibióticas. Frecuentemente los tejidos que acumulan alcaloides
son los más vulnerables, o son tejidos periféricos que pueden ser atacados por
herbívoros (Hopkins, 1999), pero el hecho que cerca de 80% de las plantas no
contienen alcaloides hace suponer que estos no son vitales para todos los
organismos vivientes (Lock, 1994).
Para la extracción de alcaloides se aprovecha su carácter básico,
utilizando soluciones acuosas o alcohólicas ligeramente ácidas, para obtener el
extracto crudo de alcaloides; pudiendo previamente liberarlos con soluciones de
amoniaco y carbonato de sodio, para extraerlos luego con solventes
orgánicos.(Lock, 1994)
Las técnicas de separación de alcaloides más comunes son la
cromatografía de capa fina o delgada aunque en los noventa se generalizó el
uso de Sephadex LH-20 por elución y la cromatografía líquida de alta
perfomance (HPLC). Los sistemas solventes son muy variados y el agente
revelador de uso general es el reactivo de Dragendorff, cuya aplicación produce
manchas generalmente de color naranja (Lock 1994).
La Dra. Gloria Chacón, (1961, 1990) demostró la presencia de alcaloides

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
en Lepidium peruvianum, para demostrar la acción fecundadora del extracto
alcaloideo de maca, lo inoculó en ratas obteniendo una marcada estimulación
de la maduración los folículos de Graaf y engrosamiento del endometrio.
Posteriormente Yllesca, (1994), en un estudio fitoquímico de la planta encontró
3 alcaloides en los extractos metanólico y butanólico de maca de los ecotipos
amarillo, rojo y negro. En 1993, Garró y col. obtuvieron cuatro fracciones de
alcaloides por cromatografía de capa fina. Por otro lado Dini y colb., (1994),
detectaron por cromatografía de capa fina de un extracto alcalino de polvo de
maca realizado en cloroformo 3 fracciones positivas al reactivo de Dragendorff.
Sin embargo la presencia de alcaloides en maca aún es discutida y no se ha
determinado su estructura.
III.3.- CULTIVO DE TEJIDOS
El cultivo de tejidos como técnica consiste en aislar una porción de planta
y proporcionarle las condiciones apropiadas para que las células expresen su
potencial intrínseco o inducido. Es en realidad un conjunto de diversas técnicas
mediante las cuales un explante, es decir una parte separada de la planta, se
cultiva asépticamente en un medio de composición definida y se incuba en
condiciones ambientales controladas. (Roca y Mroginski, 1991).
Los principios de esta técnica están contenidos en la teoría celular de
Schleiden y Schwann (1839, citados por Gautheret, 1982) que postula que la
célula es capaz de tener autonomía y totipotencia, lo que fue demostrado en
células somáticas con la observación de la formación y cicatrización de callos en
áreas donde la planta sufre cortes (Gautheret, 1982).
Los primeros intentos de establecer cultivos de tejidos vegetales fueron
realizados por el botánico alemán G. Haberlandt (1902, citado por Gautheret

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
1982), quien trabajó con células de palizada, pelos glandulares y pelos de
estambres de Tradescantia; pero aunque logró que las células sobrevivieran
entre 20 a 27 días, no tuvo crecimiento celular, probablemente debido a que
eligió nutrientes relativamente simples y a que eligió células altamente
diferenciadas (Dodds y Roberts, 1982). Recién en 1934, mucho después que en
1912 Carrel diseñara el método para cultivar células animales; que Gautheret
logra los primeros resultados exitosos en el cultivo de tejidos del cambium en
Acer pseudoplatanus, Salix capraea y Sambucus nigra (Gautheret 1982) y White
demuestra el crecimiento ilimitado de las puntas cortadas de raíces de tomate
(Dodds y Roberts 1982).
Los primeros logros del cultivo de tejidos fueron los de formación de
callos indiferenciados, que podían crecer ilimitadamente a través de subcultivos,
por lo que se puso mucho énfasis en la determinación de los requerimientos
nutricionales para el crecimiento sostenido (Dodds y Roberts 1982). Para
entonces ya se había descubierto la auxina IAA (Went, 1926, citado por
Gautheret, 1982) y en las décadas siguientes se estudiaron diversas sustancias
que favorecían el crecimiento celular de los tejidos cultivados in vitro (Gautheret
1982) se descubrió la kinetina y otras hormonas que recibieron el nombre de
citoquininas (Dodds y Roberts 1982) , hasta que en 1962, Murashigue y Skoog
propusieron una solución de minerales asociada a auxinas y citoquininas que
permite el crecimiento de la mayoría de tejidos (Gautheret 1982).
También se inició una serie de estudios en organogénesis e histogénesis,
que sentaron las bases de la micropropagación cuando Ball en 1946 descubrió el
principio de la propagación vegetativa determinando cuales eran los tipos de
meristemos que eran capaces de generar plantas completas. En 1964, Morel
aplicó estos principios en la propagación clonal de orquídeas. (Gautheret 1982).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Muir (1953 – 1954, citado por Dodds y Roberts 1982) encontró que los
callos que eran trasladados a un medio líquido se convertían en suspensiones
celulares, desarrollándose luego muchas técnicas de cultivo de células aisladas y
de otro tipo de cultivos, como el de anteras, de microsporas, el aislamiento y
cultivo de protoplastos y otros. (Dodds y Roberts 1982).
Actualmente el cultivo de tejidos se aplica entre otros:
a) Estudios básicos de fisiología, genética, bioquímica y ciencias afines.
b) Bioconversión y producción de compuestos útiles.
c) Incremento de la variabilidad genética.
d) Obtención de plantas libres de patógenos.
e) Propagación de plantas.
f) Conservación e intercambio de germoplasma.
III.3.1.- CULTIVO DE CALLOS
El establecimiento del cultivo de tejidos depende del tipo de explante
usado, y la elección de éste dependerá del objetivo perseguido y la especie
vegetal utilizada. Para la obtención de callos se puede utilizar cualquier tipo de
explante que contenga células nucleadas, y éste se seleccionará en función a
razones prácticas como disponibilidad, facilidad de manipulación,
homogeneidad, baja contaminación, respuesta in vitro; esta respuesta puede
variar con el genotipo de las plantas, su estado de desarrollo, edad ontogénica y
tamaño del explante (Roca y Mroginski, 1991). A las dos o tres semanas de
incubación empieza a formarse, en las regiones de corte, el callo, una masa
amorfa desdiferenciada de células parenquimáticas de pared delgada (Dodds y
Roberts 1982), y sigue creciendo cubriendo el explante en unos casos, y en
otros desintegrándose a medida que el callo crece (Nuñez y Ochoa, 2000). Este
crecimiento depende de la relación entre la planta de origen, el medio de cultivo,

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
las condiciones ambientales y el tiempo de incubación; algunos callos crecen
fuertemente lignificados y de textura dura, mientras otros se rompen fácilmente
en pequeños fragmentos, estos callos son llamados friables (Dodds y Roberts,
1995), estos callos son los mejores para la iniciación de cultivos celulares.
El establecimiento de un callo a partir de un explante, puede dividirse en
tres fases de desarrollo: Inducción, en la que el metabolismo es estimulado
antes de la mitosis; división celular en el que las células del explante revierten a
un estado meristemático; y por ultimo diferenciación celular y expresión de
algunas vías metabólicas que llevan a la formación de metabolitos secundarios
(Dodds y Roberts, 1995).
La formación de callos en plantas intactas, normalmente está controlada
por hormonas endógenas, las auxinas y citoquininas, por lo que para su
inducción in vitro en explantes vegetales sin causar estrés, depende de la
introducción de reguladores de crecimientos en el medio de cultivo (Nuñez y
Ochoa, 2000).
Los otros factores más importantes para el éxito de esta tecnología son:
el explante y el medio de cultivo, es decir, los componentes esenciales y
opcionales que contenga. Los nutrientes esenciales consisten en las sales
inorgánicas, fuentes de carbono y energía, vitaminas y fitohormonas (Gamborg y
Shyluk, 1981). Otros componentes como compuestos nitrogenados, ácidos
orgánicos y sustancias complejas pueden ser importantes pero son opcionales.
Después de que el callo ha crecido por un tiempo en asociación con el
tejido original, es necesario subcultivar el callo a medio fresco. El crecimiento en
el mismo medio por un tiempo prolongado lleva al agotamiento de los

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
nutrientes, la gradual desecación del agente gelificante y a la acumulación de
metabolito que pueden resultar tóxicos para el callo (Dodds y Roberts, 1995).
Los subcultivos sucesivos usualmente son llevados a cabo cada seis
semanas, con cultivos mantenidos en medio agar a 25°C o más, pero en
realidad este tiempo varía con la tasa de crecimiento del callo (Dodds y Roberts,
1995).
Carhuaz en 1997 (comunicación personal), indujo callos en maca,
utilizando un factorial de cuatro citoquininas (BAP, KIN, ZEA y 2ip) y cuatro
auxinas (IIA, IBA, NAA y 2,4-D) a concentraciones de 0.1, 1.0, 10.0, 100 ìM,
concluyendo que el 2,4-D en el rango de 0.1 – 10.0 ìM el regulador m ás
efectivo para promover la formación de callos, coincidiendo con Gamborg y colb.
(1981), quienes indican un rango óptimo de 1- 5 ìM para la acción del 2,4-D y
con Flick y colb. (1983), que también señalan al 2,4-D y a la kinetina como los
reguladores más efectivos para la inducción de callos en Brassicáceas.
III.3.2.- HORMONAS Y REGULADORES DE CRECIMIENTO
Se llama hormona o fitohormona a aquellas sustancias orgánicas
naturales que a bajas concentraciones ejercen una profunda influencia en los
procesos fisiológicos de la planta (Hopkins, 1999), mientras que las sustancias
sintéticas con las mismas propiedades son llamadas reguladores de crecimiento,
y no son consideradas como hormonas vegetales (Dodds y Roberts, 1995).
Se conocen cinco grupos de hormonas: auxinas, giberelinas, citoquininas,
ácido absícico y etileno (Hopkins, 1999). De estas las auxinas y citoquininas son
las mas usadas en el cultivo de callos, mientras las giberelinas son usadas con
poca frecuencia y generalmente en cultivos de meristemos apicales. De igual

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
modo el ácido absícico y el etileno se usan con poca frecuencia (Dodds y
Roberts, 1995).
En la familia Brassicaceae se encuentran muchas especies
económicamente importantes, en muchas de ellas se han inducido callos,
incluyendo a Brassica oleracea, B. napus y Arabidopsis thaliana, los
requerimientos para inducir callos en esta familia pueden ser cubiertos con
muchas hormonas, con un amplio rango de concentraciones, estas
concentraciones y la hormona a ser elegida, así como la respuesta de
crecimiento varía con la especie y el genotipo. Sin embargo, usualmente se
utiliza una auxina y una citoquinina, (Flick y colb., 1983), dentro de las
citoquininas la más usada es la Kinetina, mientras que la auxina utilizada con
más frecuencia es 2,4-D, de igual modo, la relación auxina:citoquinina más
eficiente para la inducción de callos es de 1 (Flick y colb. (1983).
III.3.2.1- AUXINAS
Las auxinas (del griego auxein = incrementar) fueron identificadas por
primera vez por Went en 1926, quien describió la acción elongadora del IAA en
coleóptidos de avena. Posteriormente se hallaron mas auxinas naturales, de las
cuales la más usada en cultivo de tejidos es el IAA, y otras sintéticas o
reguladores de crecimiento de las cuales las más usadas son el 2,4-D, el NAA y
el IBA (Krikorian, 1991).
Los efectos de las auxinas más importantes para el cultivo de tejidos son
el crecimiento celular y la elongación celular. Algunos tejidos sólo forman callos
en respuesta a una auxina en particular, siendo a veces necesario utilizar altas
concentraciones de auxina para la iniciación del callo, aunque en el caso del 2,4-
D, se ha observado efectos inhibitorios a concentraciones mayores a 1 mg/litro.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Este regulador es la auxina más efectiva para la proliferación de callos
usándose a concentraciones de 10-7 – 10-5 M y aún en ausencia de una
citoquinina exógena (Yeoman y Forche, 1980).
Además de inducir la proliferación celular, el 2,4-D tiende a suprimir la
morfogénesis del callo, por lo que su uso no es recomendado para estudios de
diferenciación (Yeoman y Forche, 1980). En Arabidopsis thaliana, la brassicacea
mas estudiada, la auxina mas usada en la obtención de callos en diferentes
explantes es el 2,4-D, sea junto a una citoquinina o a leche de coco (Morris y
Altmann, 1994).
III.3.2.2.- CITOCININAS
En 1954 Skoog y Miller descubrieron un compuesto muy activo en la
promoción de la citocinesis, el que se formaba por la descomposición parcial de
ADN, llamándolo cinetina (Kinetina, KIN) y propusieron el término cinina para
denominar las sustancias naturales y sintéticas que presentaban el mismo tipo
de actividad biológica. Posteriormente para evitar confusiones con la sustancia
usada en cultivo de tejidos animales, se optó por el nombre citocinina para
estas sustancias (Krikorian, 1991).
Las citoquininas se caracterizan por su habilidad para estimular la división
celular en el cultivo de tejidos cultivo, induciendo la división celular sincrónica a
las 18 h (Szweykowska, 1974), y, en presencia de una auxina, estimular la
formación de brotes e inhibir el enraizamiento (Dodds y Roberts, 1995).
Las citoquininas mas usadas son la kinetina y la zeatina, siendo
compuestos sintéticos, mientras la zeatina es una citoquinina natural (Dodds y
Roberts, 1995).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Hay excepciones en el requerimiento de auxinas y citoquininas, así
algunos cultivos no requieren de una auxina exógena y otros requieren de una
auxina y no de una citoquinina (Dodds y Roberts, 1995). Se ha observado
además que en todos los casos las citoquininas a altas concentraciones (10 –25
mg/l) inhiben la formación de callos (Yeoman y Forche, 1980).
En Lepidium peruvianum, Carhuaz (1997, comunicación personal) utilizó
un factorial de medios con cuatro auxinas (IAA, IBA, NAA, y 2,4-D) y cuatro
citoquininas ( BAP, KIN, ZEA, Y 2ip) a concentraciones de 0.1, 1.0, 10.0 y 100
µM, utilizando como explantes, la raíz, hoja, peciolo e hipocótilo.
III.3.3.- EL EXPLANTE
Los requerimientos hormonales para la iniciación del callo dependen del
origen del tejido aislado (explante), los explantes que contienen células del
cambium pueden formar callos sin adición de reguladores de crecimiento; pero
la mayoría de los explantes requiere de uno o más reguladores de crecimiento
para iniciar la formación de callos (Dodds y Roberts, 1995). Esta respuesta
también depende del estado fisiológico del tejido y del tiempo de escisión,
(Yeoman y Forche, 1980).
Los explantes pueden ser clasificados de acuerdo a sus requerimientos
exógenos de la siguiente manera: 1) auxinas 2) citoquininas 3) auxinas y
citoquininas 4) extractos naturales complejos (Dodds y Roberts, 1995).
III.3.4.- CULTIVO DE TEJIDOS Y METABOLITOS SECUNDARIOS
Las plantas siguen siendo una importante fuente comercial de metabolitos
secundarios, pero en algunos casos estas plantas no han sido sujetas a

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
programas intensivos de mejoramiento genético y se presentan además
problemas técnicos y económicos para su cultivo (Dodds y Roberts, 1995).
El cultivo de tejidos puede ser una alternativa para producir metabolitos
secundarios cuya extracción a partir de las plantas que los producen puede
resultar difícil o económicamente inviable, sea por el largo tiempo de espera
necesario, por el riesgo de sobreexplotación al que se expone el cultivo; por las
pequeñas cantidades de compuesto presente en la planta o por los controles a
los que se someten a las plantas productoras (Robert y colb., 1991). Por otro
lado el cultivo de tejidos puede ser utilizado para mejorar el cultivo de estas
plantas (Dodds y Roberts, 1995).
La noción de que una línea celular altamente productiva podía ser
seleccionada para generar compuestos útiles fue introducida en la década de
1970, en la década de los ochenta, las Universidades de Kyoto y Kitasato en
cooperación con la industria privada, produjeron shikonina de Lithospermum
erythrorhyzon y saponinas de ginseng respectivamente (Hara, 1996).
La producción de shikonina, un pigmento que se extrae de las raíces de la
planta asiática Lithospermum erythrorhyzon es un ejemplo de la efectividad del
uso del cultivo de tejidos vegetales en la producción de metabolitos secundarios.
En la planta el rendimiento de shikonina, además de ser muy bajo, depende de
la distribución geográfica y el clima, habiendo sufrido una rápida disminución en
su población. Luego de establecer una estrategia de producción in vitro, la
empresa petroquímica Mitsui del Japón logró la primera producción industrial del
compuesto, vendiendo a 4000 dólares el kilo de shikonina producida in vitro
(Robert y colb., 1991).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
En la década de los noventa el número de compuestos producidos por
cultivo de tejidos aumentó y la mejora de la tecnología Hara permitió obtener
compuestos raros en los recursos vegetales naturales. Un ejemplo de esta
tecnología es la producción comercial, por Kao, de un polisacárido tuberoso
usado en cosméticos (Hara, 1996).
Actualmente existen al menos tres programas de búsqueda de
compuestos con actividad biológica, llevados a cabo por las industrias
farmacéuticas, cada año varios cientos de plantas son colectados y se intenta en
ellas la inducción de callos, con un éxito de cerca al 50%. Cerca del 10% de los
callos muestra alguna forma de actividad ( Schripsema, y colb., 1996).
Todo el proceso desde la colección hasta la obtención de un compuesto
puro es estimado entre los 2 y 4 años. Algunos resultados de estas búsquedas
han sido publicados y patentados, como el alcaloide jatrorrhizina y los
dehidrodiconiferil alcohol glucósidos aislados de cultivos celulares de
Plagiorhegma dubium Maxim (Schripsema y colb., 1996).
La búsqueda de nuevos compuestos a nivel de callos tiene la ventaja de
reducir el número de cultivos de interés, muchas veces el paso de callos a
suspensiones celulares representa una reducción considerable de los niveles de
ciertos metabolitos secundarios, ocurriendo algunas veces la pérdida completa
de productividad para ciertos productos, lo que no significa que los cultivos
celulares no sean capaces de producir compuestos de interés, por el contrario,
Ruyter y Stöckingt (1989 citados por Schripsema y colb., 1996) demostraron que
los cultivos celulares son una excelente fuente de nuevos compuestos.
Las ventajas esperadas del cultivo de tejidos en la producción de
metabolitos secundarios son las siguientes:

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• Producción a escala industrial
• Producción de sustancias difíciles de obtener por extracción o por síntesis
química
• Reducción de los costos de producción a largo plazo
• Eliminación de la dependencia en materia de importación
• Producción continua y controlada de sustancias independiente- mente de los
factores del medio ambiente.
• Precios estables
• Posibilidad de realizar la producción cerca de las fábricas de procesamiento
final y de los mercados.
Se ha estudiado también la posibilidad de aumentar significativamente la
productividad del cultivo usando un medio de inducción (para la producción de
los metabolitos secundarios) (Becker, 1987).
III.4.- CROMATOGRAFÍA
La cromatografía es la técnica analítica usada para la separación de
mezclas y sustancias, por adsorción selectiva (Microsoft® Encarta® Online
Encyclopedia 2001) y fue descubierta por el botánico ruso Michael Tswett en
1903, quien descubrió que los pigmentos de las plantas podían separarse al
filtrarlos con éter de petróleo a través de una columna de carbonato de calcio.
Pero el desarrollo de esta técnica empieza realmente en 1931, cuando,
posteriormente otros investigadores introducen otros materiales como las
columnas de silicagel, papel (Randerath, 1968).
Actualmente se usa un amplio rango de técnicas cromatográficas de
acuerdo a los adsorbentes usados, la cromatografía de columna utiliza sílica,
alúmina y sílica gel; en la cromatografía de capa fina el adsorbente se encuentra

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
sobre un vidrio o una película plástica, mientras que en la cromatografía de
papel, una muestra líquida fluye sobre el papel adsorbente. Otras técnicas
cromatográficas son: la cromatografía gas-líquida, que permite la separación de
sustancias que pueden ser volatizadas; la cromatografía iónica donde un gas
puede ser descompuesto en iones que interactúan con el adsorbente; la
cromatografía de permeación de gel usa un adsorbente con poros de tamaño
uniforme, y finalmente, la cromatografía líquida de alto perfomance (HPLC)
donde el adsorbente es líquido, esta técnica es ampliamente usada
actualmente (Microsoft® Encarta® Online Encyclopedia 2001).
III.5.- ELECTROFORESIS EN GEL
La electroforesis en gel es un método de separación, en el que las
partículas cargadas migran hacia un electrodo de carga opuesta bajo la influencia
de un campo eléctrico aplicado externamente. Este movimiento se realiza en
interacción con la matriz circundante, la cual actúa como un filtro molecular,
generándose como consecuencia, tasas diferenciales de migración para las
proteínas contenidas en una muestra (Garfin, 1990).
Los primeros trabajos de electroforesis fueron realizados en soportes de
papel y acetato de celulosa, y se utilizaron principalmente para separar
aminoácidos, péptidos y mezclas proteicas mal separadas por cromatografía
(Freifelder, 1979), los soportes pueden ser relativamente inertes, como papel,
acetato de celulosa, silicagel, alúmina y celulosa y actuar como tamices
moleculares como son los geles de almidón, agarosa y poliacrilamida (Hames,
1981, citado por Gálvez, 1990).
El método más utilizado es la electroforesis en geles de poliacrilamida
(PAGE), por proporcionar un tamaño de poro controlable, ser repetible y de alta
resolución (Garfin, 1990).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Los geles de poliacrilamida se forman por la polimerización del monómero
de acrilamida y un co-monómero de ligamiento cruzado, la N,N’-metilen-
bisacrilamida. Esta polimerización es catalizada por un sistema generador de
radicales libres, compuesto por persulfato de amonio, que actúa como iniciador,
y un acelerador, el tetrametiletilendiamida TEMED, que causa la formación de
radicales libres a partir del persulfato de amonio, los que a su vez catalizan la
polimerización del gel (Garfin, 1990). Otro agente donante de radicales libres es
la riboflavina, pero a diferencia del persulfato de amonio requiere de luz y
oxígeno, agentes que convierten la convierten a su forma leuco, la cual inicia la
polimerización (Gálvez, 1990).
Los poros se forman por la naturaleza del enlace cruzado entre la
acrilamida y la bisacrilamida, y su tamaño disminuye a medida que la
concentración de acrilamida aumenta, determinando sus propiedades como
tamiz.
La separación por electroforesis en gel esta basada en los tamaños,
formas y cargas netas de las macromoléculas, los sistemas diseñados para
proteínas nativas se utilizan principalmente para separar y categorizar mezclas
de proteínas; pero no para medir la pureza de una preparación o el peso
molecular de una muestra desconocida, pues estos sistemas no diferencian entre
los efectos del tamaño, forma y carga en la movilidad electroforética, debido a
que proteínas con diferentes pesos moleculares pueden tener la misma
movilidad; para diferenciar estos efectos se utiliza la electroforesis denaturante
(SDS – PAGE)(Garfin, 1990).
La separación por electroforesis en gel esta basada en los tamaños,
formas y cargas netas de las macromoléculas, los sistemas diseñados para
proteínas nativas se utilizan principalmente para separar y categorizar mezclas

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
de proteínas; pero no para medir la pureza de una preparación o el peso
molecular de una muestra desconocida, pues estos sistemas no diferencian entre
los efectos del tamaño, forma y carga en la movilidad electroforética, debido a
que proteínas con diferentes pesos moleculares pueden tener la misma
movilidad; para diferenciar estos efectos se utiliza la electroforesis denaturante
(Garfin, 1990), en la cual se utilizan detergentes iónicos como el SDS (sodium
dodecyl sulfato) el mismo que neutraliza la cargas de la proteína de tal forma
que las proteínas migran por efecto de su peso molecular, las muestras en este
caso deben ser tratadas con SDS y un tiol, como el mercaptoetanol. (Gálvez,
1990).
El método más popular es el SDS-PAGE desarrollado por Laemmli en
1970 (Garfin, 1990), el cual consiste en un método discontinuo con dos tipos de
geles: un gel de separación y uno de concentración, ambos geles tienen
diferentes porosidades y pH, sirviendo el segundo como un medio anticonvectivo,
donde la muestra se apila antes de pasar al gel de separación que se halla
inmediatamente debajo y que posee un tamaño de poro más pequeño. Además,
se utilizan diferentes buffers para el gel y los electrodos, esta discontinuidad
permite concentrar grandes volúmenes de muestra en el gel de concentración,
resultando en una mejor resolución (Laemmli, 1970).

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO IV
MATERIAL Y MÉTODOS
IV.1.- INDUCCIÓN DE CALLOS Para la inducción de callos se siguió un protocolo en base al trabajo
realizado por Carhuaz en 1997 (comunicación personal) según se describe a
continuación.
IV.1.1.- MATERIAL VEGETAL
Se utilizaron plántulas de maca amarilla, morada y negra, germinadas in vitro
en medio Murashigue y Skoog (1962), sin hormonas, suplementado con
vitaminas, mioinositol, pantetonato de calcio, 20% de sucrosa, y 2% de gelrite.
Las semillas fueron gentilmente proporcionadas por el Ing. Rolando Aliaga
(CIMA – UNA) y provinieron de la comunidad de Huayre – Junín.
IV.1.1.1.- DESINFECCIÓN
La desinfección de las semillas se realizó adecuando el protocolo de
desinfección de rutina usado en el LRGB, de la manera siguiente:
Inmersión en alcohol al 70% : 1 minuto
Inmersión en hipoclorito de Na al 2% : 10 minutos
Enjuague con agua destilada estéril : 1 minuto, cuatro veces
IV.1.1.2.- CONDICIONES DE CULTIVO
Las condiciones del cuarto de cultivo fueron: 18°C, 1,500 Lux, fotoperíodo
de 16 h de luz. A los 40 días, las plántulas tuvieron el tamaño de hoja necesario
para obtener dos explantes: apical y proximal.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Los colores del ecotipo de maca correspondiente a cada plántula de
origen fueron denominados utilizando la siguiente nomenclatura:
M1 = Ecotipo amarillo, M2 = Ecotipo morado, M3 = Ecotipo negro.
IV.1.2.- EXPLANTES
Los explantes obtenidos fueron los siguientes:
E1: Región apical de la hoja (Aprox. 0.5 cm de longitud x 0.5 en la base.)
E2: Región proximal de la hoja del mismo tamaño que el explante anterior.
Los bordes de las hojas fueron cortados para obtener una mayor área de
inducción de callos.
E3: Porción de peciolo aprox. 1 cm de longitud.
E4: Hipocótilo completo, incluyendo la roseta meristemática, la mayoría de
ellos medía tuvieron aproximadamente 0.4 cm.
E5: raíces segmentos sin puntas de aprox.1 cm se escindieron de la región
proximal.
IV.1.3.- MEDIO DE CULTIVO PARA LA INDUCCIÓN DE CALLOS:
Se utilizaron dos hormonas vegetales para la inducción de callos, una
auxina (2,4-D) en concentraciones de 0, 0.1, 0.5, 1 y 10 µM y una citoquinina
(kinetina) en concentraciones de 0, 0.1, 0.5, 1 y 10 µM; como medio básico se
utilizó el medio descrito por Murashige y Skoog (1962) al que se le agregaron
mioinositol 100 ppm, pantotenato de calcio 2 ppm, vitaminas: tiamina 2 ppm,
piridoxina 0.5 ppm, ácido nicotínico 0.1 ppm y glicina 0.5 ppm; 2% de sucrosa y
0.2% de gelrite, tal como se utiliza en rutina en el LRGB, obteniéndose un
factorial de 17 medios de inducción semisólidos con diferentes concentraciones
de hormona incluyendo el control negativo.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
V.1.4.- INDUCCIÓN DE CALLOS:
Se sembraron los explantes en placas petri descartables con 25 ml de
medio de inducción, haciendo cinco repeticiones por cada medio de inducción.
Las placas se mantuvieron en oscuridad a 25°C. realizándose evaluaciones a
partir de los 25 días de siembra con un intervalo de 10 días por evaluación
durante dos meses, estas evaluaciones fueron cualitativas para confirmar el
crecimiento de los callos.
A los dos meses de crecimiento se hizo el primer subcultivo en placas
petri descartables con medios de inducción frescos, a los cuatro meses se
evaluó el material midiendo el área ocupada por cada callo sobre una cuadrícula
milimetrada, para determinar así la masa relativa de los mismos y por
consiguiente el medio con mayor rendimiento en masa. Seguidamente, se
realizó el subcultivo de todos los callos en el medio K (ver apéndice), a partir de
entonces se realizaron subcultivos cada dos meses hasta los ocho meses
cuando se cosecharon los callos para la extracción de metabolitos secundarios.
IV.2.- DETECCIÓN DE METABOLITOS SECUNDARIOS
Se realizaron cromatografías de papel para la detección de glucosinolatos
y cromatografías de capa fina ascendente para la detección de alcaloides tanto
en los callos obtenidos como en los controles, no se utilizó una sola línea celular
debido a la pequeña cantidad de materia seca obtenida de los callos como
consecuencia de su alto porcentaje de humedad. Para la extracción de los
metabolitos se utilizaron 30 mg muestra estabilizada.
La estabilización se realizó secando los callos en una estufa a 37° C por
36 horas para luego molerlos y guardar los frascos en un lugar seco y sin luz.
Como control, se usaron muestras de hipocótilos amarillos morados y negros

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
provenientes de la localidad de Huayre y estabilizadas en las mismas
condiciones que los callos. Los callos se clasificaron de acuerdo al color y
naturaleza del explante del que provenían, así un callo proveniente de la zona
apical de la hoja de ecotipo amarillo se denominó M1E1 y así sucesivamente (ver
apéndice). Esta clasificación nos facilitó el manejo de las muestra en las
cromatografías.
Antes de la pulverización de las muestras se determinó el % de humedad.
IV.2.1.- EXTRACCIÓN Y DETECCIÓN DE GLUCOSINOLATOS
Para la extracción y cromatografía de glucosinolatos se adecuó el
protocolo de rutina utilizado por el Dr. César Fuertes en el Instituto de Química
Orgánica Aplicada a la Farmacia, en la Facultad de Farmacia y Bioquímica de la
Universidad Nacional Mayor de San Marcos (ver apéndice), como se describe a
continuación:
Para la extracción etanólica se tomaron 30 mg de cada tipo de callo y se
pusieron a macerar en 2 ml de etanol al 96% durante dos semanas. Se sembraron,
100 alicuotas de cada uno de estos extractos en hojas de papel Wathman N° 1 de
11 x 29 cm y se colocaron en un tanque de cromatografía de papel descendente,
dejándose desarrollar por 22 horas.
Como sistema solvente se usó una solución de n-butanol:etanol:agua (4:1:4)
descrito por Kjaer en 1968.
Después de retirar los papeles del tanque de cromatografía se secaron a
temperatura ambiente e inmediatamente se revelaron para lo cual se pulverizaron
primero con una solución de nitrato de plata:acetona 15 mg/ml, y luego de dejar
secar por unos minutos, con una solución de KOH:Etanol 20 mg/ml. Se dejaron
secar las hojas de papel unos minutos y se colocaron en una estufa caliente. Una

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
vez obtenidas las manchas indicadoras, se colocaron los cromatogramas en una
solución decolorante de hiposulfito de potasio al 5%, durante 20 hrs.
IV.2.2.- EXTRACCIÓN Y DETECCIÓN DE ALCALOIDES
Se utilizaron 30 mg de cada muestra, las que se pusieron a macerar en éter
etílico, realizándose tres extracciones sucesivas.
La cromatografía de capa fina se realizó de acuerdo a la técnica descrita por
Stahl (1958 y 1965), en cromatofolios de silicagel GF254 ( para las
cromatografías preliminares se usaron cromatofolios de 2 x 10 cm) y
cromatoplacas Kieselgel 60 G F254 (Merck) de 20 x 20 cm.
En las cromatografías preliminares se realizaron con las muestras de
hipocótilos de maca (controles) y se probaron las siguientes soluciones para el
sistema solvente:
Benzol : éter dietílico : metanol : amoniaco (75 : 75 : 6 : 0.7);
Cloroformo : metanol : amoniaco (2 : 0.12 : 0.02);
Cloroformo : metanol (2 : 1), (2 : 0.25), (2 : 0.15) y (2 : 0.12), siendo esta
última la que se utilizó en la estandarización de la cromatografía.
Se desarrollaron cromatografías de muestras acidificadas y sin acidificar.
Para el revelado de las placas se utilizó el reactivo de Dragendorff y
posteriormente el reactivo yodoplatinato de potasio.
Antes de revelar las placas se observaron en el espectro de luz UV.
En vista de la ausencia de resultados congruentes en la primera
cromatografía, se procedió a estandarizar la extracción en una muestra de

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
hipocótilos de varios colores, la cantidad de metabolitos no deseables para la
cromatografía fue bastante alta por lo que se procedió a lavar el extracto
acidificando el extracto etéreo con HCl acuoso, recuperándose la fase acuosa a
la cual, luego de ser alcalinizada con NaOH concentrado, se le agregó dos
volúmenes de éter etílico para recuperar la fase orgánica. Se cromatografió este
extracto en una cromatoplaca Kieselgel 60 F F254 (Merck) de 20 x 20 cm. Se
revelaron los bordes de la placa con el reactivo de Dragendorff modificado por
Munnier (1955) y el reactivo yodoplatinato de potasio, aislando luego las
regiones correspondientes a las manchas positivas.
Una vez obtenidas las fracciones positivas para alcaloides, se
cromatografiaron los extractos de callos y los controles, junto con el extracto
etéreo y sus cuatro fracciones, las que se usaron como un control extra.
IV.3.- DETECCIÓN DE MIROSINASAS IV.3.1.- ELECTROFORESIS.-
Se utilizó la electroforesis en geles de policrilamida (PAGE) para separar
las mirosinasas, los geles y buffers fueron preparados de acuerdo al sistema de
Laemmli (1970), pero eliminando el uso de SDS y mercaptoetanol tanto de las
soluciones de los geles como de las de los buffers para obtener una PAGE en
condiciones no denaturantes. La concentración de los geles fue de 8.6 y 7.5%.
Las corridas en los geles se llevaron a cabo con una corriente constante de 30
mA, el voltaje inicial fue de 80 V y el final de 250V. La duración de las corridas
fue de 5 horas.
IV.3.2.- EXTRACCIÓN DE LAS MUESTRAS.
Se llevó a cabo adaptando el protocolo de MacGibbon y Allison (1964): 0.5g
de muestra fueron molidos en un mortero frío con 2 ml de buffer Tris-Cl pH 6.8,

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
se centrifugó a 10,000 rpm por 4 min, se tomaron 80 µl del sobrenadante a los
que se añadieron mercaptoetanol 5 µl, glicerol 10 µl y 5µl de un stock de azul de
bromofenol preparado para los procedimientos de rutina del LRGB. Se
colocaron 50 µl de muestra en cada pocillo del gel.
Se utilizaron hipocótilos y hojas de campo.
IV.3.3.- DETECCIÓN DE ACTIVIDAD.-
Se adaptó la modificación del método de detección de actividad basado
en la formación de bandas de BaSO4, descrito por MacGibbon y Allison (1970),
esta modificación fue gentilmente proporcionada por el Dr. Atle Bones en una
comunicación personal (1999). Para adaptar este protocolo se utilizó un extracto
etanólico de semillas de mostaza, preparado moliendo 100 g de semillas y
macerándolas en 250 ml de alcohol etílico al 96%.
Cuando los geles fueron removidos, fueron colocados entre dos placas de vidrio
e incubados a 50°C en una solución acuosa de ácido ascórbico 1mM; Acetato de
Bario 10mM; Buffer Tris-Cl 0.5 M; pH 6.8 200 ml/L; extracto etanólico de semillas
de mostaza 300 ml/L.
Cuando se hicieron evidentes las bandas, los geles fueron fotografiados con una
cámara digital DAGEMIT CCD100 en blanco y negro.
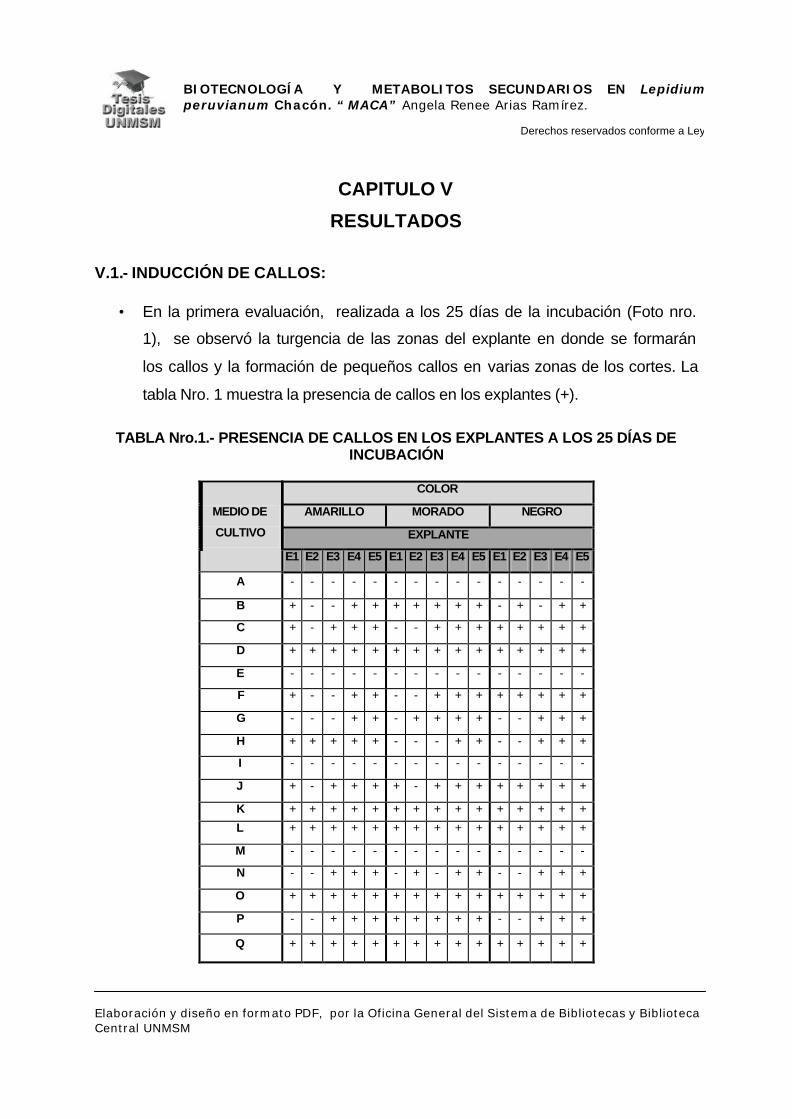
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO V
RESULTADOS
V.1.- INDUCCIÓN DE CALLOS:
• En la primera evaluación, realizada a los 25 días de la incubación (Foto nro.
1), se observó la turgencia de las zonas del explante en donde se formarán
los callos y la formación de pequeños callos en varias zonas de los cortes. La
tabla Nro. 1 muestra la presencia de callos en los explantes (+).
TABLA Nro.1.- PRESENCIA DE CALLOS EN LOS EXPLANTES A LOS 25 DÍAS DE
INCUBACIÓN
COLOR
AMARILLO MORADO NEGRO
EXPLANTE
MEDIO DE
CULTIVO
E1 E2 E3 E4 E5 E1 E2 E3 E4 E5 E1 E2 E3 E4 E5
A - - - - - - - - - - - - - - -
B + - - + + + + + + + - + - + +
C + - + + + - - + + + + + + + +
D + + + + + + + + + + + + + + +
E - - - - - - - - - - - - - - -
F + - - + + - - + + + + + + + +
G - - - + + - + + + + - - + + +
H + + + + + - - - + + - - + + +
I - - - - - - - - - - - - - - -
J + - + + + + - + + + + + + + +
K + + + + + + + + + + + + + + +
L + + + + + + + + + + + + + + +
M - - - - - - - - - - - - - - -
N - - + + + - + - + + - - + + +
O + + + + + + + + + + + + + + +
P - - + + + + + + + + - - + + +
Q + + + + + + + + + + + + + + +
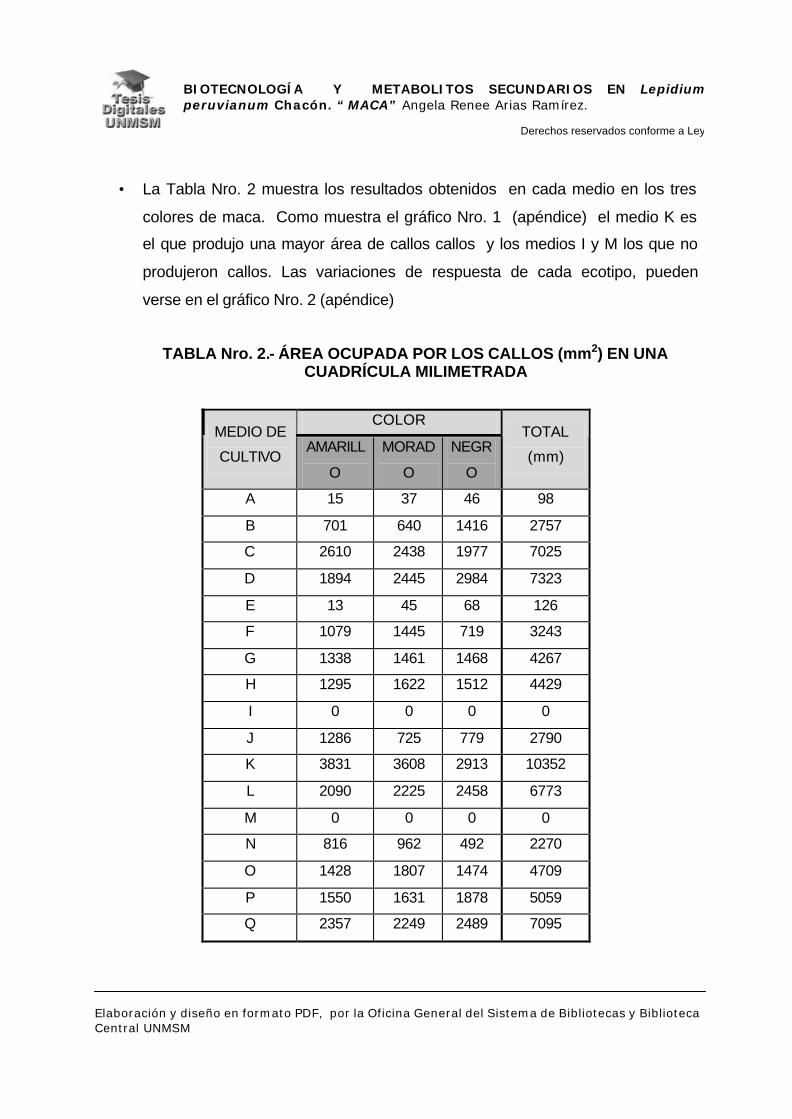
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 2 muestra los resultados obtenidos en cada medio en los tres
colores de maca. Como muestra el gráfico Nro. 1 (apéndice) el medio K es
el que produjo una mayor área de callos callos y los medios I y M los que no
produjeron callos. Las variaciones de respuesta de cada ecotipo, pueden
verse en el gráfico Nro. 2 (apéndice)
TABLA Nro. 2.- ÁREA OCUPADA POR LOS CALLOS (mm2) EN UNA CUADRÍCULA MILIMETRADA
COLOR MEDIO DE
CULTIVO AMARILL
O
MORAD
O
NEGR
O
TOTAL
(mm)
A 15 37 46 98
B 701 640 1416 2757
C 2610 2438 1977 7025
D 1894 2445 2984 7323
E 13 45 68 126
F 1079 1445 719 3243
G 1338 1461 1468 4267
H 1295 1622 1512 4429
I 0 0 0 0
J 1286 725 779 2790
K 3831 3608 2913 10352
L 2090 2225 2458 6773
M 0 0 0 0
N 816 962 492 2270
O 1428 1807 1474 4709
P 1550 1631 1878 5059
Q 2357 2249 2489 7095
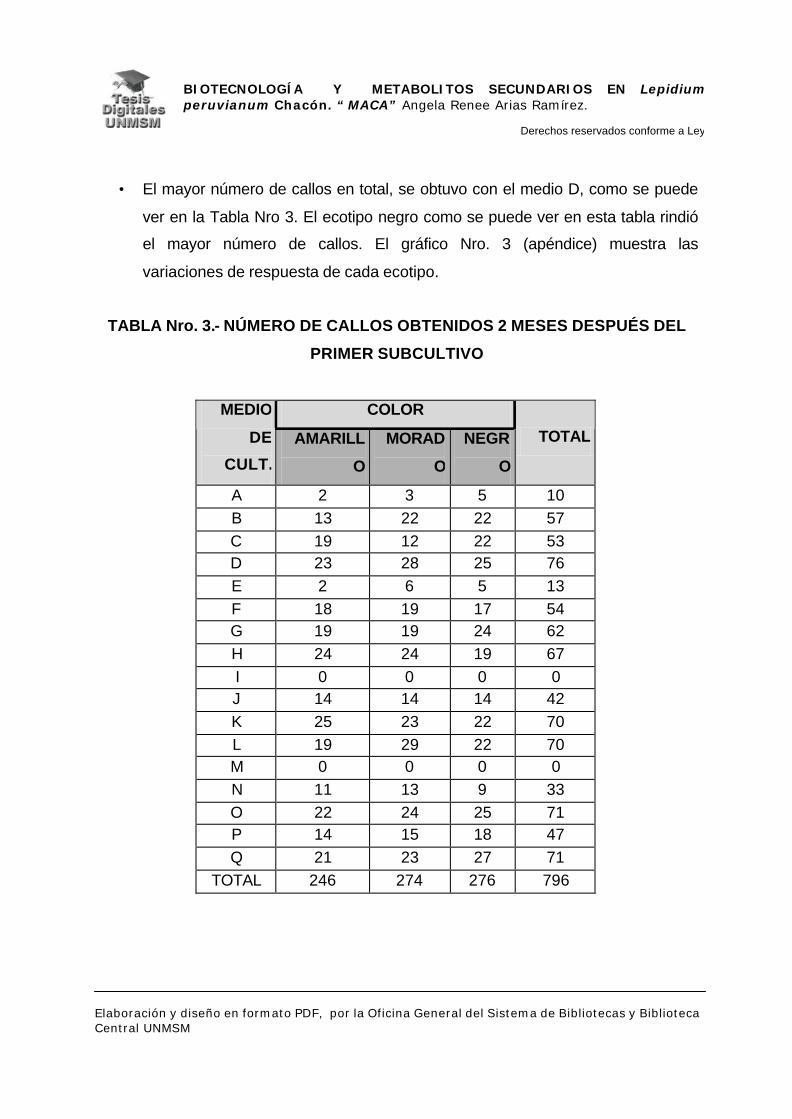
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• El mayor número de callos en total, se obtuvo con el medio D, como se puede
ver en la Tabla Nro 3. El ecotipo negro como se puede ver en esta tabla rindió
el mayor número de callos. El gráfico Nro. 3 (apéndice) muestra las
variaciones de respuesta de cada ecotipo.
TABLA Nro. 3.- NÚMERO DE CALLOS OBTENIDOS 2 MESES DESPUÉS DEL
PRIMER SUBCULTIVO
COLOR MEDIO
DE
CULT.
AMARILL
O
MORAD
O
NEGR
O
TOTAL
A 2 3 5 10 B 13 22 22 57 C 19 12 22 53 D 23 28 25 76 E 2 6 5 13 F 18 19 17 54 G 19 19 24 62 H 24 24 19 67 I 0 0 0 0 J 14 14 14 42 K 25 23 22 70 L 19 29 22 70 M 0 0 0 0 N 11 13 9 33 O 22 24 25 71 P 14 15 18 47 Q 21 23 27 71
TOTAL 246 274 276 796
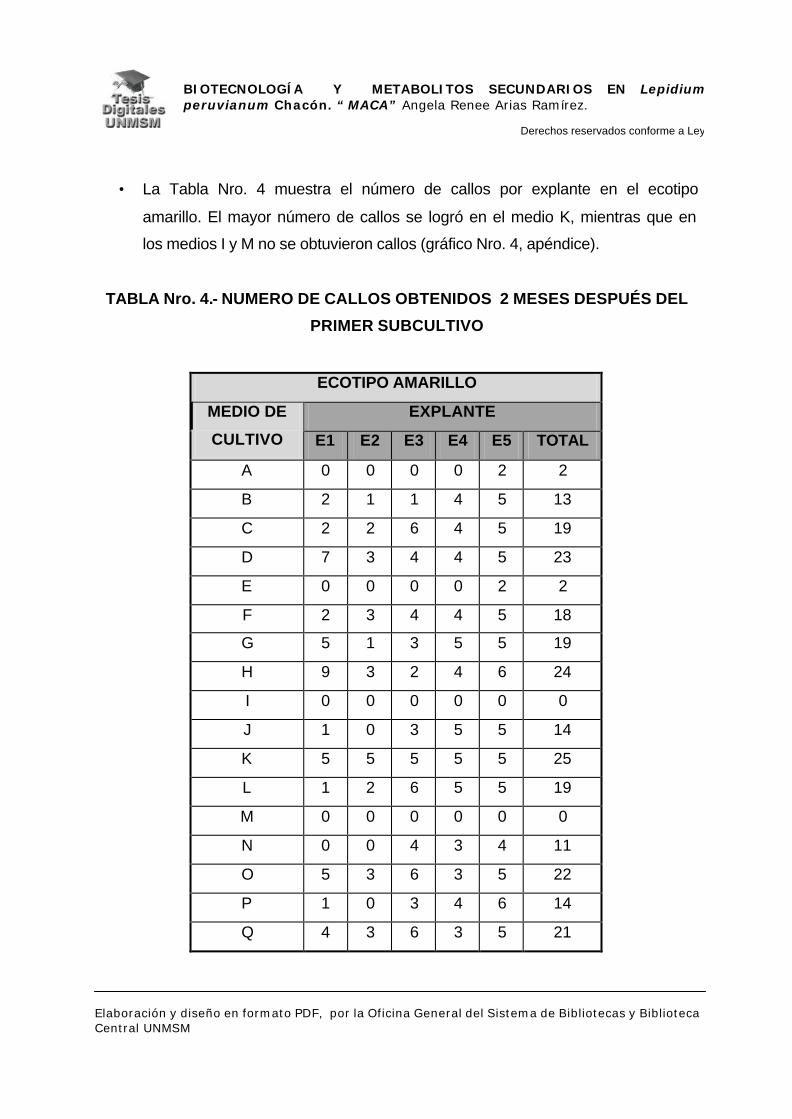
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 4 muestra el número de callos por explante en el ecotipo
amarillo. El mayor número de callos se logró en el medio K, mientras que en
los medios I y M no se obtuvieron callos (gráfico Nro. 4, apéndice).
TABLA Nro. 4.- NUMERO DE CALLOS OBTENIDOS 2 MESES DESPUÉS DEL
PRIMER SUBCULTIVO
ECOTIPO AMARILLO
EXPLANTE MEDIO DE
CULTIVO E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 2 2
B 2 1 1 4 5 13
C 2 2 6 4 5 19
D 7 3 4 4 5 23
E 0 0 0 0 2 2
F 2 3 4 4 5 18
G 5 1 3 5 5 19
H 9 3 2 4 6 24
I 0 0 0 0 0 0
J 1 0 3 5 5 14
K 5 5 5 5 5 25
L 1 2 6 5 5 19
M 0 0 0 0 0 0
N 0 0 4 3 4 11
O 5 3 6 3 5 22
P 1 0 3 4 6 14
Q 4 3 6 3 5 21
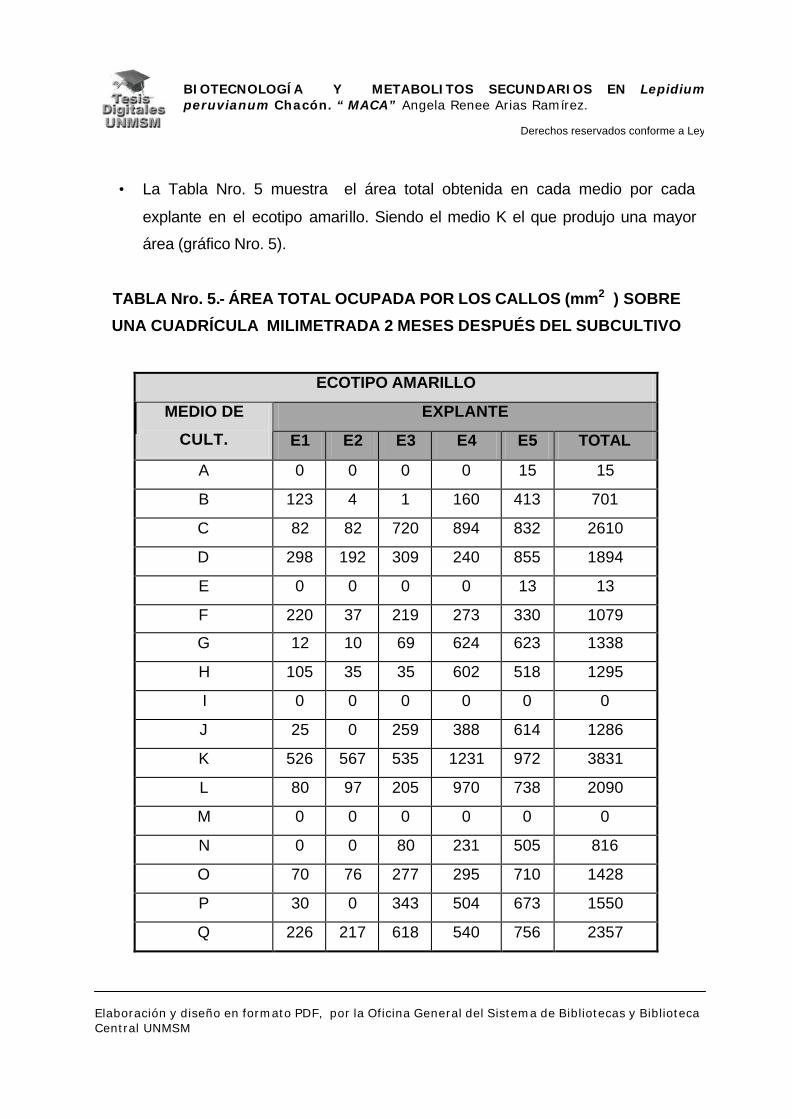
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 5 muestra el área total obtenida en cada medio por cada
explante en el ecotipo amarillo. Siendo el medio K el que produjo una mayor
área (gráfico Nro. 5).
TABLA Nro. 5.- ÁREA TOTAL OCUPADA POR LOS CALLOS (mm2 ) SOBRE
UNA CUADRÍCULA MILIMETRADA 2 MESES DESPUÉS DEL SUBCULTIVO
ECOTIPO AMARILLO
EXPLANTE MEDIO DE
CULT. E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 15 15
B 123 4 1 160 413 701
C 82 82 720 894 832 2610
D 298 192 309 240 855 1894
E 0 0 0 0 13 13
F 220 37 219 273 330 1079
G 12 10 69 624 623 1338
H 105 35 35 602 518 1295
I 0 0 0 0 0 0
J 25 0 259 388 614 1286
K 526 567 535 1231 972 3831
L 80 97 205 970 738 2090
M 0 0 0 0 0 0
N 0 0 80 231 505 816
O 70 76 277 295 710 1428
P 30 0 343 504 673 1550
Q 226 217 618 540 756 2357
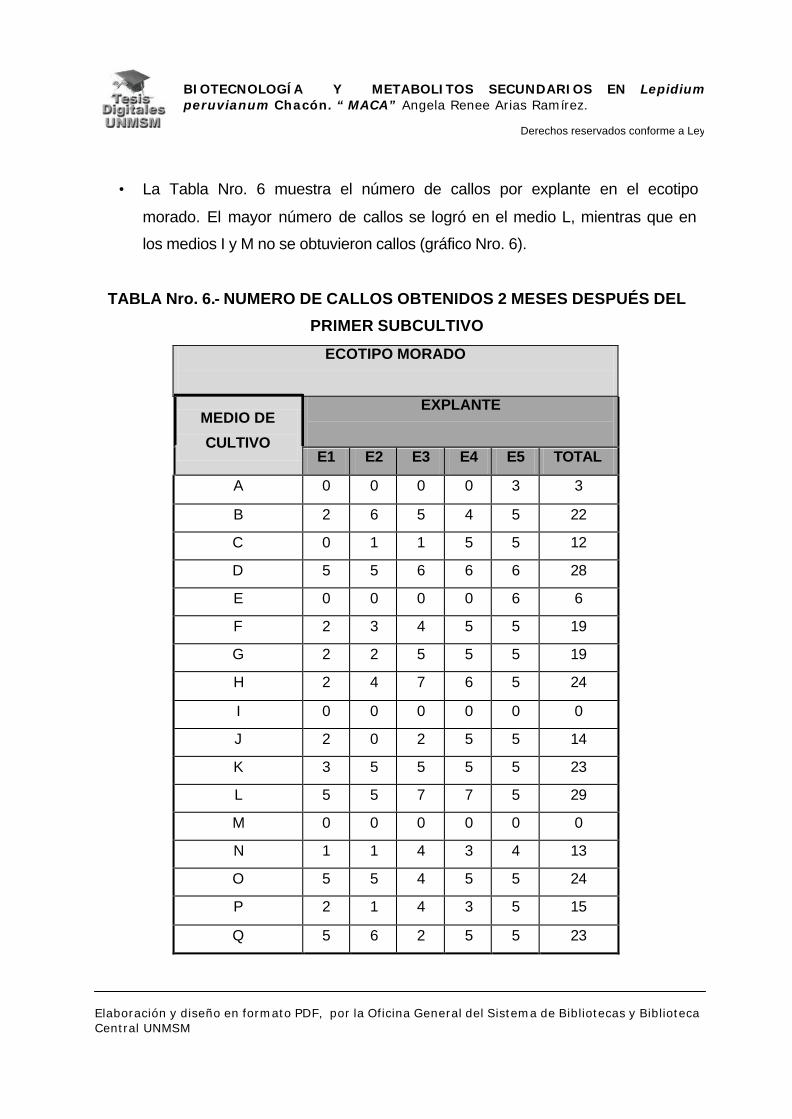
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 6 muestra el número de callos por explante en el ecotipo
morado. El mayor número de callos se logró en el medio L, mientras que en
los medios I y M no se obtuvieron callos (gráfico Nro. 6).
TABLA Nro. 6.- NUMERO DE CALLOS OBTENIDOS 2 MESES DESPUÉS DEL
PRIMER SUBCULTIVO
ECOTIPO MORADO
EXPLANTE
MEDIO DE
CULTIVO E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 3 3
B 2 6 5 4 5 22
C 0 1 1 5 5 12
D 5 5 6 6 6 28
E 0 0 0 0 6 6
F 2 3 4 5 5 19
G 2 2 5 5 5 19
H 2 4 7 6 5 24
I 0 0 0 0 0 0
J 2 0 2 5 5 14
K 3 5 5 5 5 23
L 5 5 7 7 5 29
M 0 0 0 0 0 0
N 1 1 4 3 4 13
O 5 5 4 5 5 24
P 2 1 4 3 5 15
Q 5 6 2 5 5 23
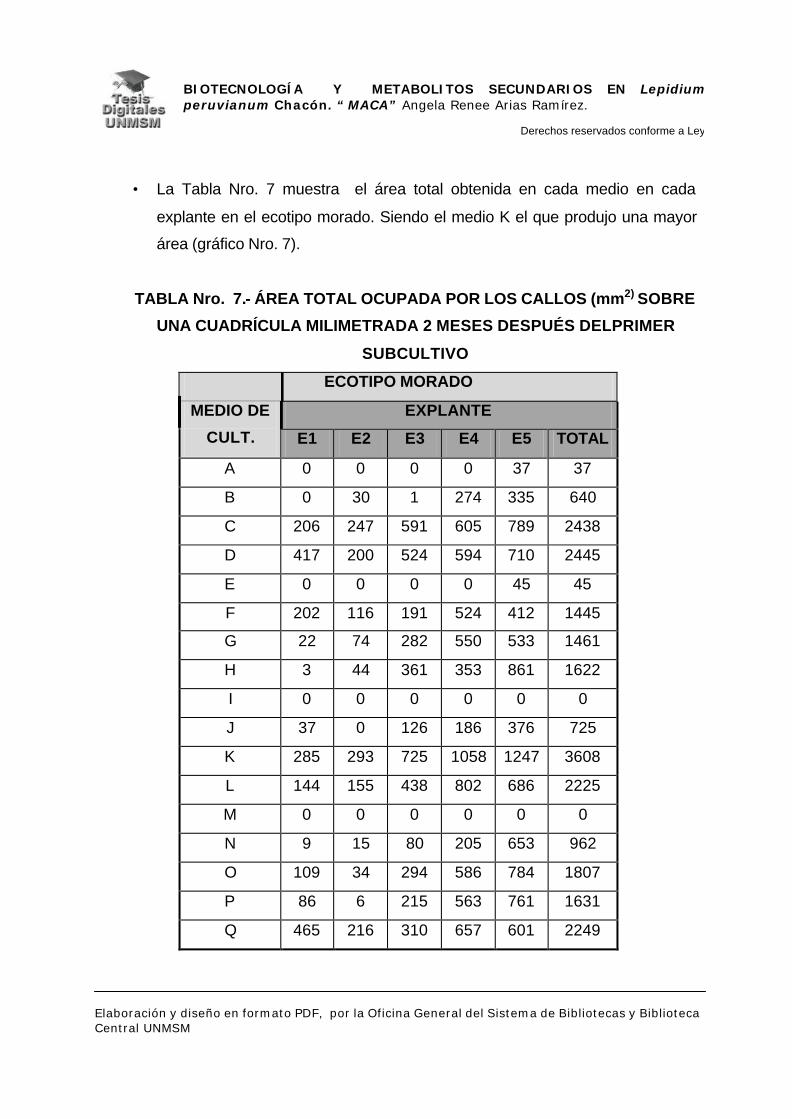
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 7 muestra el área total obtenida en cada medio en cada
explante en el ecotipo morado. Siendo el medio K el que produjo una mayor
área (gráfico Nro. 7).
TABLA Nro. 7.- ÁREA TOTAL OCUPADA POR LOS CALLOS (mm2) SOBRE
UNA CUADRÍCULA MILIMETRADA 2 MESES DESPUÉS DELPRIMER
SUBCULTIVO
ECOTIPO MORADO
EXPLANTE MEDIO DE
CULT. E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 37 37
B 0 30 1 274 335 640
C 206 247 591 605 789 2438
D 417 200 524 594 710 2445
E 0 0 0 0 45 45
F 202 116 191 524 412 1445
G 22 74 282 550 533 1461
H 3 44 361 353 861 1622
I 0 0 0 0 0 0
J 37 0 126 186 376 725
K 285 293 725 1058 1247 3608
L 144 155 438 802 686 2225
M 0 0 0 0 0 0
N 9 15 80 205 653 962
O 109 34 294 586 784 1807
P 86 6 215 563 761 1631
Q 465 216 310 657 601 2249
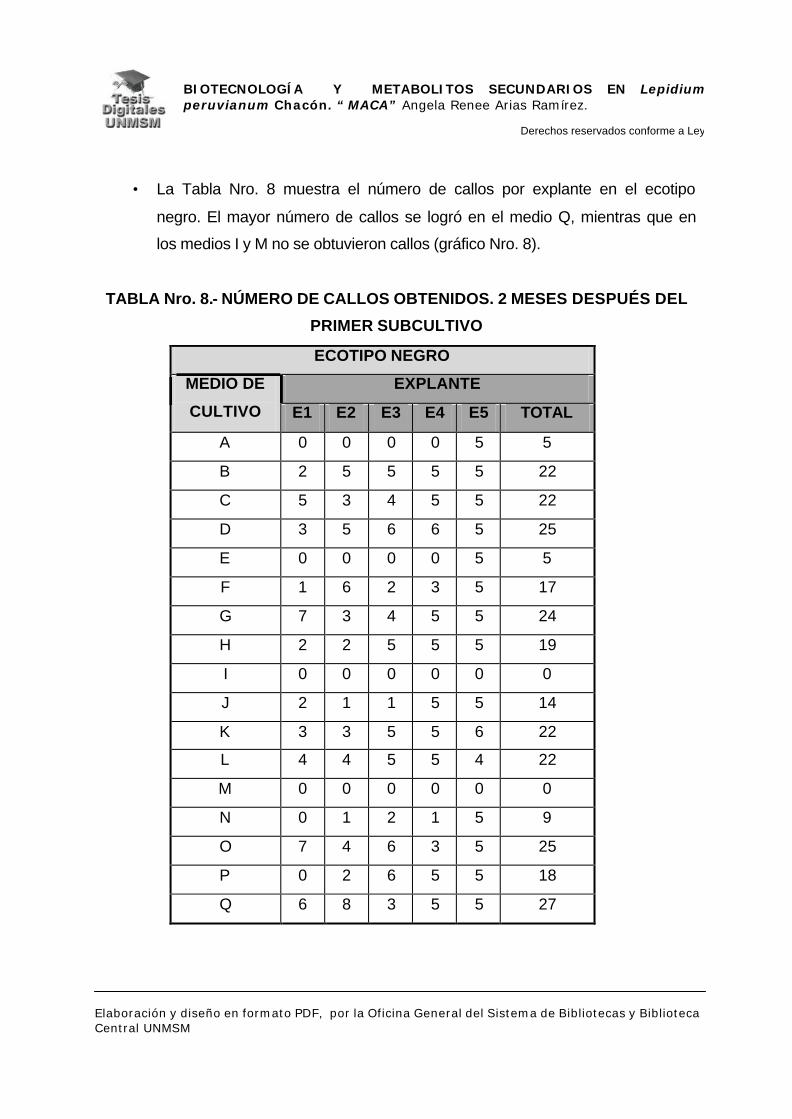
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 8 muestra el número de callos por explante en el ecotipo
negro. El mayor número de callos se logró en el medio Q, mientras que en
los medios I y M no se obtuvieron callos (gráfico Nro. 8).
TABLA Nro. 8.- NÚMERO DE CALLOS OBTENIDOS. 2 MESES DESPUÉS DEL
PRIMER SUBCULTIVO
ECOTIPO NEGRO
EXPLANTE MEDIO DE
CULTIVO E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 5 5
B 2 5 5 5 5 22
C 5 3 4 5 5 22
D 3 5 6 6 5 25
E 0 0 0 0 5 5
F 1 6 2 3 5 17
G 7 3 4 5 5 24
H 2 2 5 5 5 19
I 0 0 0 0 0 0
J 2 1 1 5 5 14
K 3 3 5 5 6 22
L 4 4 5 5 4 22
M 0 0 0 0 0 0
N 0 1 2 1 5 9
O 7 4 6 3 5 25
P 0 2 6 5 5 18
Q 6 8 3 5 5 27
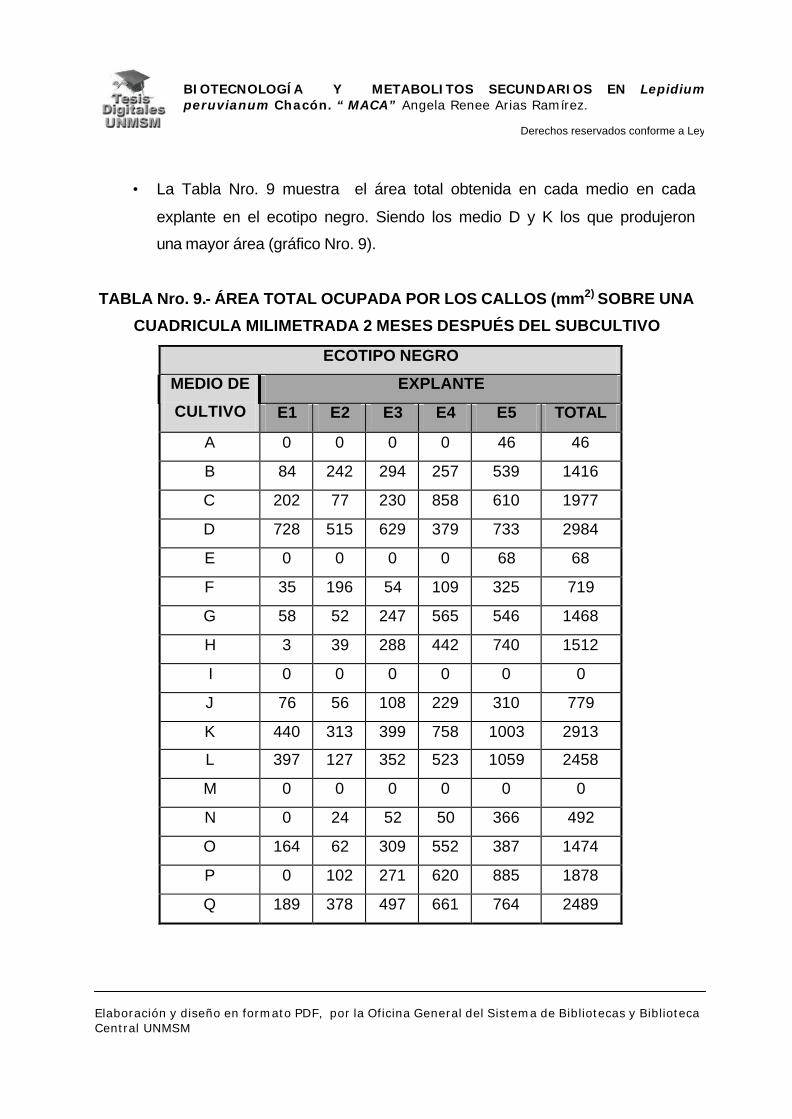
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• La Tabla Nro. 9 muestra el área total obtenida en cada medio en cada
explante en el ecotipo negro. Siendo los medio D y K los que produjeron
una mayor área (gráfico Nro. 9).
TABLA Nro. 9.- ÁREA TOTAL OCUPADA POR LOS CALLOS (mm2) SOBRE UNA
CUADRICULA MILIMETRADA 2 MESES DESPUÉS DEL SUBCULTIVO
ECOTIPO NEGRO
EXPLANTE MEDIO DE
CULTIVO E1 E2 E3 E4 E5 TOTAL
A 0 0 0 0 46 46
B 84 242 294 257 539 1416
C 202 77 230 858 610 1977
D 728 515 629 379 733 2984
E 0 0 0 0 68 68
F 35 196 54 109 325 719
G 58 52 247 565 546 1468
H 3 39 288 442 740 1512
I 0 0 0 0 0 0
J 76 56 108 229 310 779
K 440 313 399 758 1003 2913
L 397 127 352 523 1059 2458
M 0 0 0 0 0 0
N 0 24 52 50 366 492
O 164 62 309 552 387 1474
P 0 102 271 620 885 1878
Q 189 378 497 661 764 2489
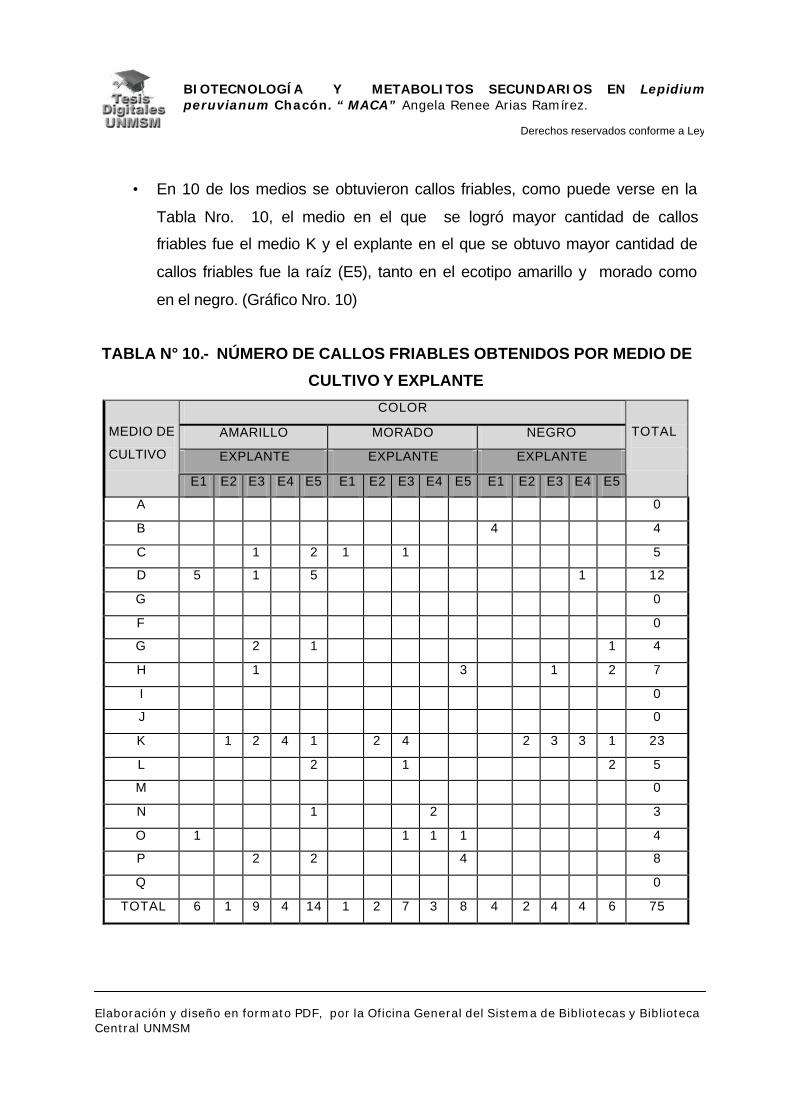
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• En 10 de los medios se obtuvieron callos friables, como puede verse en la
Tabla Nro. 10, el medio en el que se logró mayor cantidad de callos
friables fue el medio K y el explante en el que se obtuvo mayor cantidad de
callos friables fue la raíz (E5), tanto en el ecotipo amarillo y morado como
en el negro. (Gráfico Nro. 10)
TABLA N° 10.- NÚMERO DE CALLOS FRIABLES OBTENIDOS POR MEDIO DE
CULTIVO Y EXPLANTE
COLOR
AMARILLO MORADO NEGRO
EXPLANTE EXPLANTE EXPLANTE
MEDIO DE
CULTIVO
E1 E2 E3 E4 E5 E1 E2 E3 E4 E5 E1 E2 E3 E4 E5
TOTAL
A 0
B 4 4
C 1 2 1 1 5
D 5 1 5 1 12
G 0
F 0
G 2 1 1 4
H 1 3 1 2 7
I 0
J 0
K 1 2 4 1 2 4 2 3 3 1 23
L 2 1 2 5
M 0
N 1 2 3
O 1 1 1 1 4
P 2 2 4 8
Q 0
TOTAL 6 1 9 4 14 1 2 7 3 8 4 2 4 4 6 75
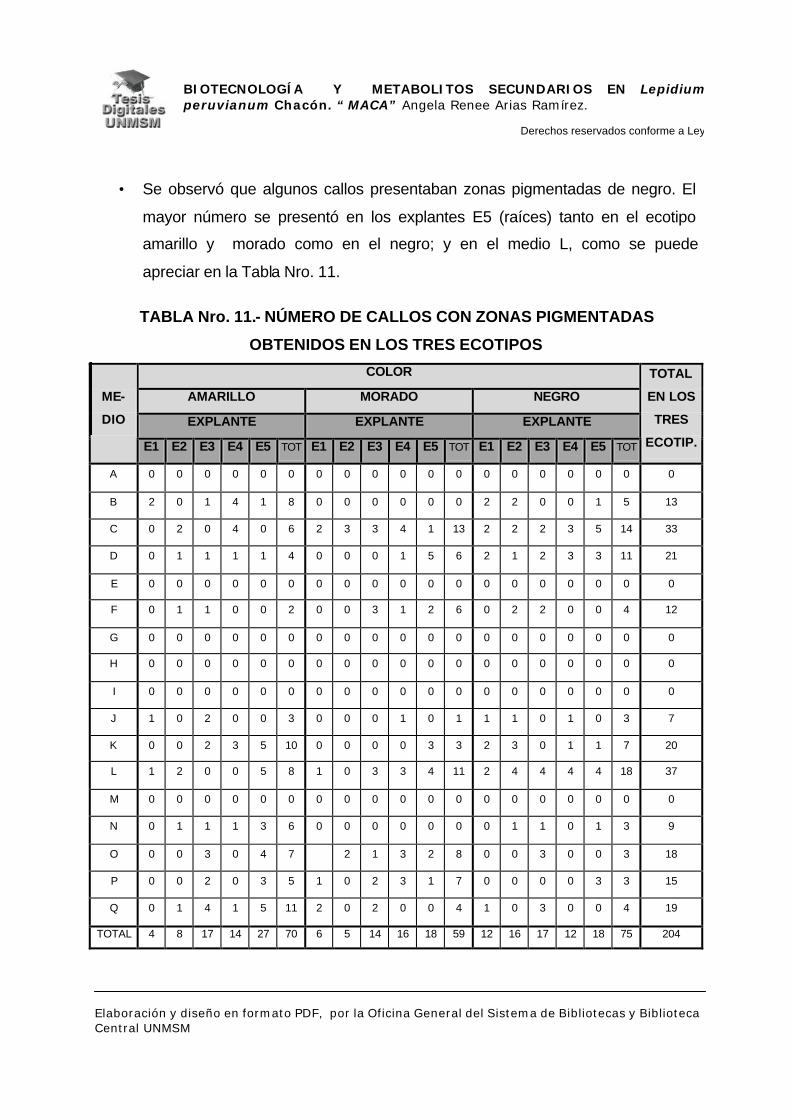
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• Se observó que algunos callos presentaban zonas pigmentadas de negro. El
mayor número se presentó en los explantes E5 (raíces) tanto en el ecotipo
amarillo y morado como en el negro; y en el medio L, como se puede
apreciar en la Tabla Nro. 11.
TABLA Nro. 11.- NÚMERO DE CALLOS CON ZONAS PIGMENTADAS
OBTENIDOS EN LOS TRES ECOTIPOS
COLOR
AMARILLO MORADO NEGRO
EXPLANTE EXPLANTE EXPLANTE
ME-
DIO
E1 E2 E3 E4 E5 TOT E1 E2 E3 E4 E5 TOT E1 E2 E3 E4 E5 TOT
TOTAL
EN LOS
TRES
ECOTIP.
A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
B 2 0 1 4 1 8 0 0 0 0 0 0 2 2 0 0 1 5 13
C 0 2 0 4 0 6 2 3 3 4 1 13 2 2 2 3 5 14 33
D 0 1 1 1 1 4 0 0 0 1 5 6 2 1 2 3 3 11 21
E 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
F 0 1 1 0 0 2 0 0 3 1 2 6 0 2 2 0 0 4 12
G 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
H 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
I 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
J 1 0 2 0 0 3 0 0 0 1 0 1 1 1 0 1 0 3 7
K 0 0 2 3 5 10 0 0 0 0 3 3 2 3 0 1 1 7 20
L 1 2 0 0 5 8 1 0 3 3 4 11 2 4 4 4 4 18 37
M 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
N 0 1 1 1 3 6 0 0 0 0 0 0 0 1 1 0 1 3 9
O 0 0 3 0 4 7 2 1 3 2 8 0 0 3 0 0 3 18
P 0 0 2 0 3 5 1 0 2 3 1 7 0 0 0 0 3 3 15
Q 0 1 4 1 5 11 2 0 2 0 0 4 1 0 3 0 0 4 19
TOTAL 4 8 17 14 27 70 6 5 14 16 18 59 12 16 17 12 18 75 204
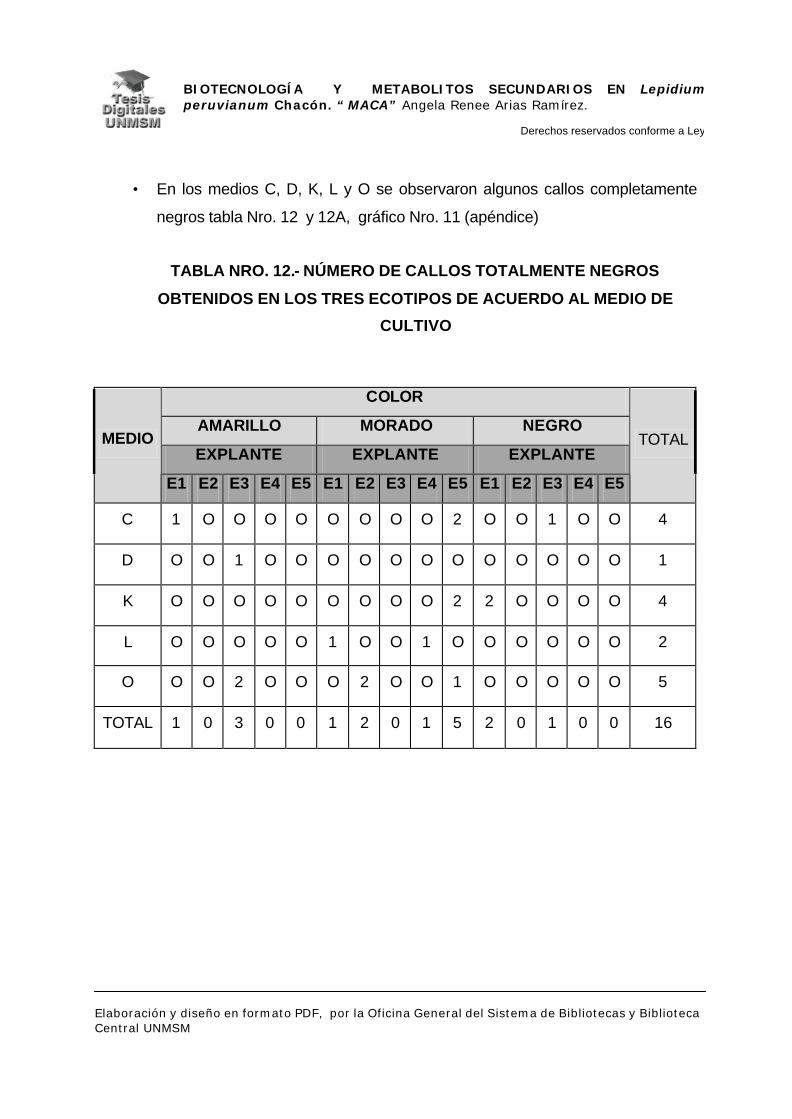
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
• En los medios C, D, K, L y O se observaron algunos callos completamente
negros tabla Nro. 12 y 12A, gráfico Nro. 11 (apéndice)
TABLA NRO. 12.- NÚMERO DE CALLOS TOTALMENTE NEGROS
OBTENIDOS EN LOS TRES ECOTIPOS DE ACUERDO AL MEDIO DE
CULTIVO
COLOR
AMARILLO MORADO NEGRO
EXPLANTE EXPLANTE EXPLANTE MEDIO
E1 E2 E3 E4 E5 E1 E2 E3 E4 E5 E1 E2 E3 E4 E5
TOTAL
C 1 O O O O O O O O 2 O O 1 O O 4
D O O 1 O O O O O O O O O O O O 1
K O O O O O O O O O 2 2 O O O O 4
L O O O O O 1 O O 1 O O O O O O 2
O O O 2 O O O 2 O O 1 O O O O O 5
TOTAL 1 0 3 0 0 1 2 0 1 5 2 0 1 0 0 16
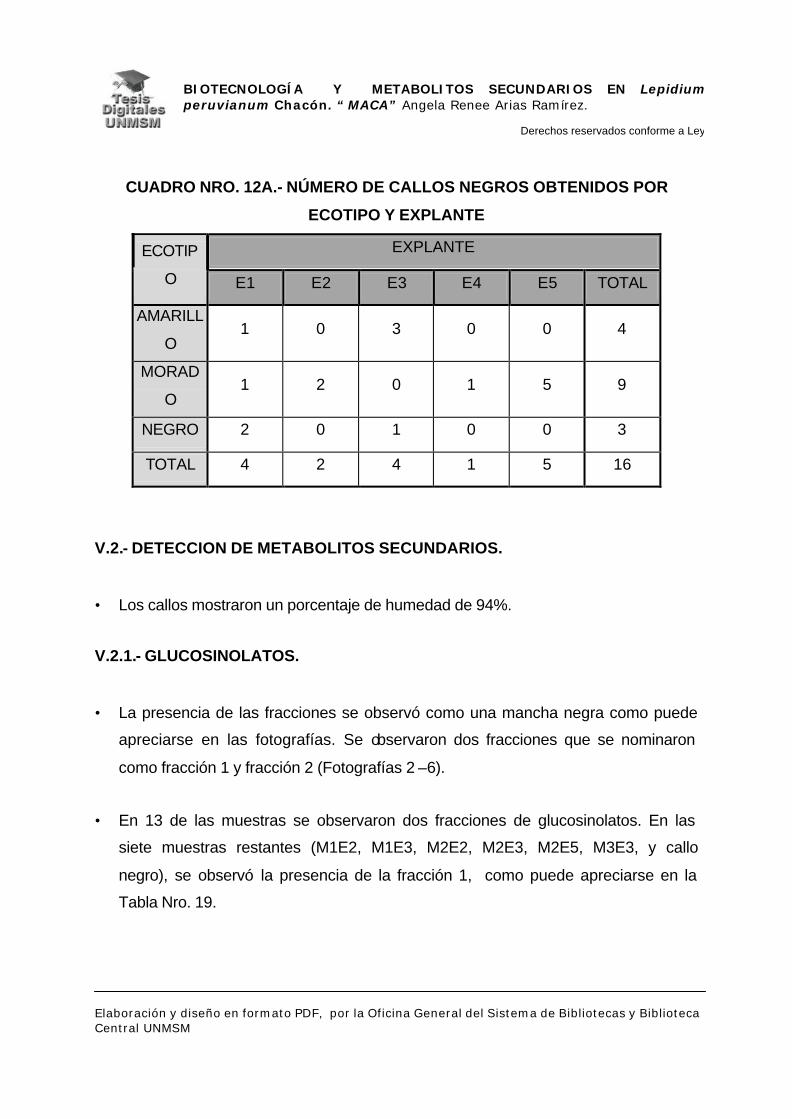
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CUADRO NRO. 12A.- NÚMERO DE CALLOS NEGROS OBTENIDOS POR
ECOTIPO Y EXPLANTE
EXPLANTE ECOTIP
O E1 E2 E3 E4 E5 TOTAL
AMARILL
O 1 0 3 0 0 4
MORAD
O 1 2 0 1 5 9
NEGRO 2 0 1 0 0 3
TOTAL 4 2 4 1 5 16
V.2.- DETECCION DE METABOLITOS SECUNDARIOS.
• Los callos mostraron un porcentaje de humedad de 94%.
V.2.1.- GLUCOSINOLATOS.
• La presencia de las fracciones se observó como una mancha negra como puede
apreciarse en las fotografías. Se observaron dos fracciones que se nominaron
como fracción 1 y fracción 2 (Fotografías 2 –6).
• En 13 de las muestras se observaron dos fracciones de glucosinolatos. En las
siete muestras restantes (M1E2, M1E3, M2E2, M2E3, M2E5, M3E3, y callo
negro), se observó la presencia de la fracción 1, como puede apreciarse en la
Tabla Nro. 19.
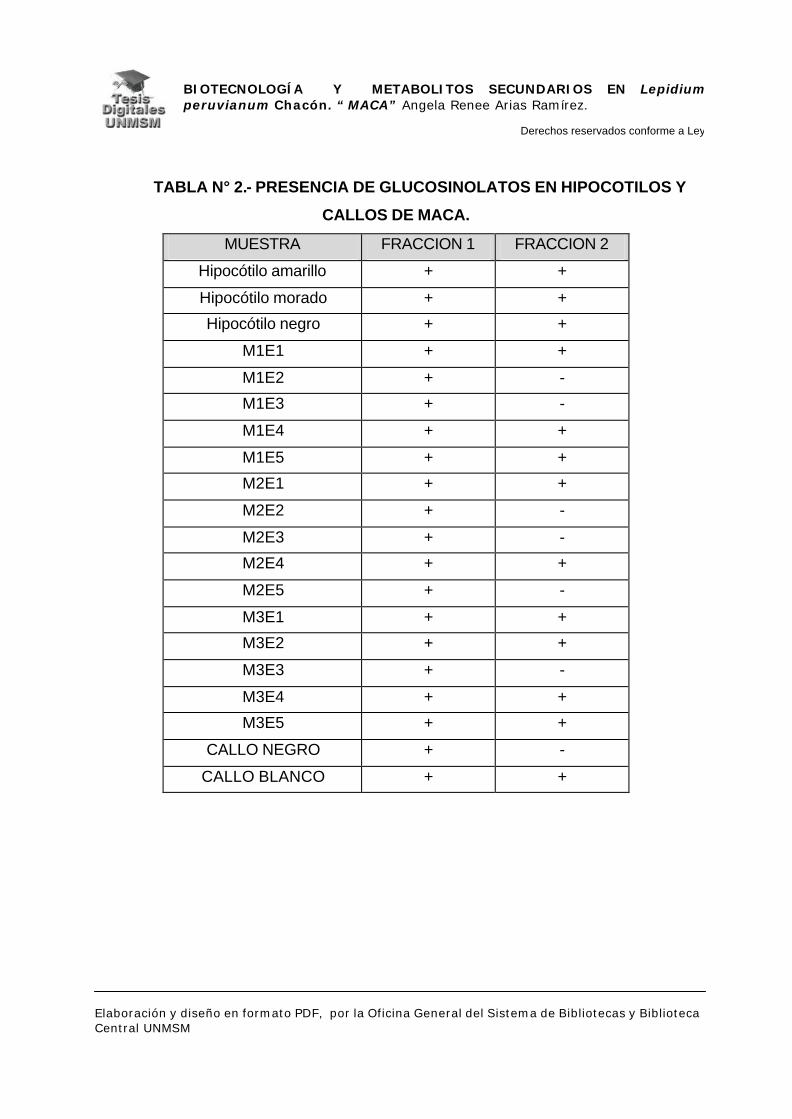
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
TABLA N° 2.- PRESENCIA DE GLUCOSINOLATOS EN HIPOCOTILOS Y
CALLOS DE MACA.
MUESTRA FRACCION 1 FRACCION 2
Hipocótilo amarillo + +
Hipocótilo morado + +
Hipocótilo negro + +
M1E1 + +
M1E2 + -
M1E3 + -
M1E4 + +
M1E5 + +
M2E1 + +
M2E2 + -
M2E3 + -
M2E4 + +
M2E5 + -
M3E1 + +
M3E2 + +
M3E3 + -
M3E4 + +
M3E5 + +
CALLO NEGRO + -
CALLO BLANCO + +

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
V.2.2.- ALCALOIDES.
En las cromatografías preliminares se observó la separación de cuatro
fracciones al usar como solvente la solución Cloroformo:metanol 2:0.12. Las
muestras acidificadas desarrollaron una sola fracción, mientras que las sin
acidificar mostraron cuatro fracciones.
Los resultados de la primera cromatografía (de todas las muestras) se
observan en la fotografía Nro. 7.
En el extracto lavado de hipocótilos de maca se obtuvieron cuatro
fracciones que fueron positivas tanto para el reactivo de Dragendorff.
Estas fracciones también fueron positivas para el reactivo yodoplatinato de
potasio, la fracción N°1 de color amarillo fue la de mayor Rf, y se presentó en la
mancha más grande, la fracción N° 2 de color plomizo debajo de ésta se halla
en cantidades pequeñas. La fracción N° 3 presentó un color marrón oscuro. La
fracción N° 4 que se encontró en el punto de siembra presentó un color azul
cemento (Fotografía Nro 8).
En la segunda cromatografía de todas las muestras, se observó la
presencia de las fracciones 1, 3 y 4, la segunda fracción no se observó en
ninguna de las muestras, incluyendo el extracto patrón, (fotografía Nro.9).
Al observar la cromatoplaca en la lámpara de luz ultravioleta, los patrones
de extractos de callos presentaron una gran cantidad de fracciones de
metabolitos mostrando patrones diferentes para cada muestra (Fotografía Nro
10). Las fracciones correspondientes a los alcaloides no mostraron fluorescencia.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
V.3.- DETECCION DE MIROSINASAS
Se observó un patrón de una sola banda por muestra (foto), las bandas
se observaron demasiado cerca del borde superior del gel por lo que no se
midieron los Rf.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO VI
DISCUSIÓN
VI.1.- INDUCCIÓN DE CALLOS.-
1. La respuesta a las diferentes concentraciones de hormonas fue variable. La
mayor producción de callos se obtuvo con los medios K,( 0.5/0.5 ìM 2,4-D/ìM
KIN), D ( 1/0 ìM 2,4-D/ìM KIN) y Q (10/10 ìM 2,4-D/ìM KIN) los medios C (
0.5/0 ìM 2,4-D/ìM KIN), L ( 1/0.5 ìM 2,4-D/ìM KIN) y P ( 1/1 ìM 2,4-D/ìM KIN)
tuvieron una respuesta menor a los anteriores pero superior a los demás. Del
mismo modo los medios M ( 0/1 ìM 2,4-D/ìM KIN) e I ( 0/0.5 ìM 2,4-D/ìM KIN)
no produjeron ninguna respuesta callogénica, mientras que el medio E ( 0/0.1 ìM
2,4-D/ìM KIN) produjo una bajísima respuesta. De todo esto y de la observación
del comportamiento de los explantes en los demás medios, podemos inferir que
la presencia del 2,4-D es de mayor importancia para la inducción de callos que la
KIN, siendo inclusive suficiente para la callogénesis. Esta observación es
congruente con el análisis estadístico realizado para esta prueba, el cual nos
muestra que el 2,4-D tiene un efecto estadísticamente significativo en la
inducción de callos. Siendo su significancia mayor que la de la auxina y la
interacción de ambas hormonas. Este dominancia ya fue señala da en otras
ocasiones, Dodds ( 1982) hace referencia a su uso junto con el agua de coco
para inducir la proliferación celular, al parecer esta alta eficiencia del 2,4-D en la
inducción de callos se debe a la alta disponibilidad de auxinas en los tejidos de la
mayoría de las dicotiledóneas ( Jacobsen, 1983)
2. En crucíferas, los callos son generalmente iniciados y mantenidos en medios que
presentasn una relacion auxina, citoquinina mayor que 1. en nuestro caso en
presencia de la KIN, la concentración de 2,4-D que dio mejores resultados fue la
de 0.5 ìM, ya que en concentraciones mayores el rendimiento del medio

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
disminuye, aun cuando la relación con la citoquinina es 1/1. Por otro lado,
podemos ver que la presencia de la KIN potencia la efectividad del 2,4-D siempre
y cuando su concentración no exceda la del 2,4-D, pues en estos casos el
rendimiento del medio disminuye. Esta disminución de la actividad podría estar
relacionada a la toxicidad del 2,4- D, pues como reportan Carew y Krueger (1977,
citados por Yerman. 1980 ) tiene efectos inhibitorios a concentraciones de 1 mgr.
por litro. De la misma la disminución en la producción de callos cuando la
relación auxina/citoquinina disminuye pueden explicarse por la actividad
inhibitoria que tienen las citoquininas cuando se presentan en altas
concentraciones (Yeoman y Forsche, 1980).
3. El medio que produjo mayor número de callos fue el medio D, seguido por los
medios O, Q, K y L, el rendimiento de masa en los medios D y Q está relacionado
al número de callos generado más que al tamaño de los mismos. La significancia
de cada una de las hormonas en el número de callos producidos es mayor para
el 2,4-D que para la KIN, sin embargo la combinación de ambas hormonas no
tiene un efecto significativo en el número de callos obtenidos. (ver apéndice:
análisis estadístico) lo cual nos indicaría que cuando el objetivo es lograr un
mayor número de líneas celulares quizá sería conveniente experimentar con una
sola hormona, especialmente el 2,4 –D.
4. La mayor efectividad del 2,4 –D corresponde a la esperada, pues es congruentes
con los resultados obtenidos por Carhuaz en Lepidium peruvianum (1997, sin
publicar) y por los resultados obtenidos en el cultivo de tejidos de diferentes
especies de la familia Cruciferae. (Flick y colb. 1983)
5. La respuesta de los ecotipos a cada uno de los medios de cultivo no fue
uniforme, como puede observarse en los gráficos 2C, 2D y 2E, siendo el ecotipo
negro el que presento mayor diferencia, sin embargo estas diferencias no son

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
demasiado grandes.
6. La masa de callos obtenidas en cada uno de los ecotipos fue mayor en el medio
K en todos los casos, mientras que los medios con los que se obtuvo un mayor
número de callos fue diferente para cada ecotipo (K, L y Q para amarillo, morado
y negro respectivamente) lo que nos muestra que la respuesta de los explantes
respecto al número de callos por explante no es uniforme. Al analizar la
respuesta de los explantes por ecotipo, se ha observado que las raíces tienen
una respuesta significativamente diferente a los otros explantes. Mientras que
para la respuesta de los explantes en relación del área ocupada por los callos se
observa una respuesta descendente de raíz a hoja. Es así que en ambos casos
las raíces son los tejidos de mayor potencial callogenético.
7. El medio A no contiene hormonas, sin embargo en este medio se produjo una
pequeña cantidad de callos, estos podrían deberse a la presencia de hormonas
endógenas en este explante que permiten la callogénesis como respuesta al
corte, esto sería coherente con la observación de que este es el explante que
produce mayor número y masa de callos en todos los ecotipos y medios y al
hecho de que en las raíces se producen auxinas endógenas (Hopkins 1999).
8. No se observaron diferencias significativas entre los dos tipos de explantes de
hoja, lo cual es coherente con los resultados obtenidos en otros trabajos, de otro
lado el medio Q fue el que produjo el mayor número de callos en estos
explantes.
9. Los peciolos e hipocótilos tuvieron una respuesta mucho mayor a los
tratamientos siendo el hipocótilo el más susceptible de los dos, esto tendría
relación con la presencia de tejido meristemático en este explante.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
10. El medio que produjo mayor cantidad de explantes friables fue el medio K,
aunque la cantidad obtenida de este tipo de callos fue bastante pequeña, lo cual
se podría deber a una predisposición de la especie o al tipo de actividad de las
hormonas usadas, de todos modos a diferencia del número de callos obtenido y
el área ocupada por los callos, en este caso puede observarse que la interacción
de las hormonas tiene un efecto más significativo que el de las hormonas solas.
11. Los explantes que produjeron la mayor cantidad de callos friables fueron los
provenientes de las raíces, mientras que los provenientes de los hipocótilos
produjeron menor cantidad de callos friables que los pecíolos, siendo igual que
en los explantes de la región terminal de la hoja. En este caso no se observaron
diferencias significativas entre los explantes de hoja terminal y proximal, aunque
el menos susceptible fue el explante proximal. Si bien la raíz y el hipocótilo
produjeron mayor número de callos friables, la diferencia entre ambos explantes
no fue significativa.
12. El efecto del color del ecotipo en la obtención de callos friables fue
significativamente diferente para el color amarillo, mientras que entre el el ecotipo
morado y negro, las diferencias no fueron significativas.
13. La presencia de callos con pigmentación negra puede explicarse por la presencia
de metabolitos secundarios en los callos, el número obtenido en los explantes
provenientes del ecotipo amarillo fue similar al obtenido del ecotipo negro, y
ambos fueron superiores a la cantidad obtenida en el ecotipo morado, por otro
lado la cantidad de callos totalmente negros obtenido en el ecotipo morado fue
mucho mayor que en los ecotipos negro y amarillo. Esto nos indicaría que esta
pigmentación no está relacionada con el color del ecotipo del que proviene el
explante.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
VI.2.- DETECCION DE METABOLITOS SECUNDARIOS
13. Los callos de Lepidium peruvianum Ch. sí producen glucosinolatos. Pero la
producción de estos metabolitos varía de acuerdo al callo, pudiendo ser mayor o
menor. Se encontraron callos en los que se formaron solo una de las fracciones.
Esto demuestra que el cultivo de callos de maca, puede ser utilizado para hallar
líneas que produzcan un solo tipo de glucosinolato, o los produzcan en
cantidades mayores a las obtenidas en campo.
14. Los explantes provenientes de peciolo(E3) y de la parte proximal de la hoja (E2)
mostraron una sola fracción de glucosinolatos en casi todos los casos, esto
sugeriría que el origen de los explantes estaría involucrado en la cantidad y tipo
de glucosinolatos producidos. Para poder comprobar esta posibilidad se tendría
que realizar pruebas con un tamaño de muestra estadísticamente significativa.
15. El origen de los explantes no parece ser la causa de la variabilidad de la
producción de callos. Pues los callos con diferentes patrones cromatográficos
provienen de diferentes tipos de explante y de plántulas de diferentes ecotipos.
Sin embargo en este caso también se observa una variabilidad alta en el
explante E5.
16. Los Rf de cada una de las fracciones es muy similar pero no igual para cada
extracto, esto puede deberse a que, por el tamaño de la cámara, las muestras no
se pudieron desarrollar en una sola cromatografía por lo que no estuvieron
sujetas a condiciones idénticas.
17. El mayor tamaño de las manchas de glucosinolatos en los extractos de callo,
indican una mayor concentración del metabolito en éstos. Esto nos brinda muy
buenas expectativas para su producción in vitro.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
18. En los cromatogramas de glucosinolatos se observan manchas débiles a lo largo
del cromatograma, estas manchas fueron asumidas como porciones de
glucosionatos la por la mirosinasa, (Kjaer, 1970). Sin embargo, Genyi y col.
reportaron en el año 2001, la presencia de otros glucosinolatos en maca, si bien
en pequeñas cantidades. Debido a esto estas no podemos asegurar que
manchas correspondan a artefactos o a esos glucosinolatos.
19. Los patrones cromatográficos de alcaloides muestran una gran variabilidad, así
como los patrones de los metabolitos con fluorescencia a la luz UV, esto nos
indicaría que los callos tienen una gran variabilidad respecto a su producción de
metabolitos, lo que sería de gran utilidad en el caso de necesitarse la explotación
de alguno de estos. Los extractos que mostraron una presencia notablemente
mayor de una de las fracciones de alcaloides fueron los de los callos M3E3 y
M3E4.
20. La fracción N° 2 del extracto de alcaloides, no es detectable cuando se
cromatografiaron cantidades pequeñas de extracto, por lo que su ausencia en la
cromatografía de los extractos de callos y controles no indican ausencia
necesaria del metabolito.
21. El hecho de que algunos extractos de callos muestren mayor presencia de
algunas fracciones de callos y dada la pequeña cantidad en la que parecen
encontrarse en los hipocótilos, el cultivo de callos o suspensiones celulares
puede ser utilizada para lograr mayores cantidades que permitan el estudio de
las propiedades y actividad biológica de estos compuestos.
22. La poca cantidad de alcaloides que presenta la planta, puede ser aumenta en el
cultivo de tejidos utilizando callos con mayor tiempo de incubación, pues, la
producción de metabolitos secundarios aumenta al iniciarse la fase estacionaria

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
(Yeoman y colb. 1980), y la de los alcaloides es inversamente proporcional a la
tasa de crecimiento (Carew y Krueger, 1977). Esta propiedad debe ser tomada
en cuenta tanto en el caso de desear producir alcaloides como otros metabolitos.
23. El hecho de que en algunos callos se observe manchas mas intensas que en el
hipocótilo y que en otros casos se observen fracciones que no se presentan en
las muestras de hipocótilos, nos indicaría el gran potencial de la planta, pues
usualmente los cultivos de tejidos vegetales producen metabolitos en pequeñas
cantidades ( Yanadi y Fujita, 1983), por otro lado la alta variabilidad de la
producción de metabolitos merece que se realicen investigaciones posteriores
para determinar si estas variaciones son fisiológics o genéticas, dado del
potencial mitogénico del 2,4-D y el aumento de la variabilidad producido por la
tecnica de cultivo de tejidos (Reisch, 1983).
VI.3.- DETECCION DE MIROSINASAS
24. La detección de mirosinasas fue cualitativa, con la finalidad de determinar
únicamente la presencia de mirosinasas en los callos e hipocótilos, pues para
una evaluación cuantitativa, se requeriría de mayores estudios de la presencia de
esta enzima en esta especie.
25. El que las bandas de mirosinasas no migraran mucho más allá de la parte
superior del gel, a pesar de dejarlas correr por encima del tiempo requerido de
acuerdo a la migración del frente de corrida, nos indicaría que estas proteínas
tienen un gran peso molecular, esto estaría en concordancia con los resultados
obtenidos por MacGibbon y Allison (1970) en otras especies y a la vez nos
indicaría que la mirosinasas de maca son mas grandes que las de otras especies
estudiadas.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO VII
CONCLUSIONES
1. Lepidium peruvianum es susceptible a la inducción de callos con auxinas y
citoquininas.
2. La auxina 2,4-D es eficiente en la inducción de callos, siendo su concentración
óptima la de 0.5 ìM.
3. La relación óptima 2,4-D/KIN es de 1
4. El comportamiento de los callos de L. peruvianum es altamente variable respecto
a la presencia de metabolitos secundarios.
5. Lepidium peruvianum es una especie que presenta condiciones óptimas para
realizar estudios de producción de metabolitos secundarios in vitro.
6. La enzima mirosinasa sí está presente en los callos y plantas de campo de L.
peruvianum. Estas enzimas son de gran tamaño molecular.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO VIII
RECOMENDACIONES
1. Recomendamos iniciar un estudio del metabolismo secundario de callos de L.
peruvianum, “maca” que nos permita determinar los factores que induzcan la
producción de metabolitos in vitro.
2. También es necesario profundizar el estudio de alcaloides de maca, para
determinar la estructura y actividad biológica de los mismos.
3. Realizar estudios histoquímicos que determinen la distribución de los
glucosinolatos y mirosinasas en la planta.
4. Sería recomendable realizar la caracterización de las mirosinasas de L.
peruvianum y compararlas con las descritas en otras especies.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CAPITULO IX
REFERENCIAS BIBLIOGRAFICAS
ALIAGA, R. 1995. Biología Floral de la Maca (Lepidium meyenii Walp.) Tesis
departamento de Horticultura, Facultad de Agronomía, Universidad Nacional Agraria
La Molina. Lima.
ALIAGA, R. 1999. Guía para el cultivo, aprovechamiento y conservación de la maca
(Lepidium meyenii) Convenio Andrés Bello. Santafé de Bogotá.
ARBIZU, C., HERMAN, M. 1995. Algunos factores limitantes en el uso de Raíces y
Tubérculos Andinos y sus prioridades de investigación. Taller Internacional sobre el
Agrosistema Andino. Lima, marzo - abril 1992. pp 223 -229.
BECKER, H. 1987. Regulation of secondary metabolism in plant cell cultures. En
Plant Tissue and Cell Culture, (Ed. por C.E.Green, D.A. Somers, W.P. Hackett y D.D.
Biesboer). Alan R. Liss. Inc. New York.
BONES M. Y ROSSITER J. 1996. The Myrosinase - Glucosinolate System, Its
Organisation and Biochemistry. Physiologia Plantarum. 97: 194 - 208.
CASTRO DE LEON, M. 1990. Un Cultivo en Extinción: El Caso de la Maca. Perú
Indig. 12(28): 85-94.
CHACON, G. 1961. Estudio Fitoquímico de Lepidium meyenii Walp. Tesis de
Bachiller en Ciencias Biológicas, Facultad de Ciencias de Universidad Nacional
Mayor de San Marcos. Lima.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
CHACON DE POPOVICH, G. 1990. La maca (Lepidium peruvianum Chacón sp.
nov.) y su hábitat. Rev. Per. Biol. 3(2):169-272.
CHAPPLE C., SHIRLEY B., SOOK M., HAMMERSCHMIDT R., SOMERVILLE S.
1994. Secondary Metabolism in Arabidopsis. En Arabidopsis ( Ed. E. M.
Meyerowitz y C.R. Somerville) Cold Spring Harbor Laboratory Press. Pp 989 – 1029.
DAXENBINCHLER M., Van ETTEN C.H., BROWN, F. y QUENTIN J. 1964.
Oxazolidinethiones and Volatile Isothiocyanates in Enzyme-Treated Seed Meal from
65 Species of Cruciferae. Journal of Agricultural and Food Chemistry. 12 (2):127-
130.
DINI, A.; MIGLIUOLO, G.; RASTRELLI, L.; SATURNINO, P.; SCHETTINO, O. 1994.
Chemical composition of Lepidium meyenii. Food Chemistry. 49: 347-349.
DODDS, J. H. y ROBERTS, L. W. 1982. Experiments in Plant Tissue Culture.
Cambridge University Press. N. Y.
FLICK, C.E., EVANS, D. A., y SHARP W. R. 1983. Organogénesis. En: Handbook
of Plant Cell Culture. Vol. 1. ( Ed. por D. A. Evans, W. R. Sharp, P. V. Amirato,
Y.Yamada). Collier Macmillan Publishers. Londres. Pp. 13 - 81.
FREIFELDER. 1979. Métodos y técnicas en bioquímica. Ed. Reverté. Barcelona.
GALVEZ M.E. 1990. Electroforesis de isoenzimas y proteínas totales en clones de
Ullucus tuberosus Loz. “olluco”. Informe de prácticas pre-profesionales para optar el
título de biólogo en la especialidad de genética. UNMSM. Lima.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
GAMBORG, L. O. Y SHYLUK J. P. 1981. Nutrition, Media and Characteristics of
Plant Cell and Tissue Culture. En: Plant Tissue Culture. Methods and Applications in
Agriculture. (Ed. T. A. Thorpe). Academic Press. N.Y.
GARFIN, D. 1990. One - Dimensional Gel Electroforesis. En Methods in Enzimology.
Vol 182. Guide to Protein Purification. (Ed. por M. Deutscher). Academic Press.
California.
GARRO, V., LEON, E., JULCA, B. 1993. Extracción, separación e identificación por
cromatografía de alcaloides de Lepidium meyenii Walp. ("Maka"). Instituto de
Química Orgánica Aplicada a la Farmacia y Bioquímica. UNMSM. VI Congreso
Peruano de Farmacia y Bioquímica.
GENYI L., AMMMERMANN, U., y QUIROS, C. 2001. Glucosinolate contents in
maca (lepidium peruvianum Chacón) seeds, sprouts, mature plants and several
derived products. Ecomomyc Botany 55 (2):255 – 262.
GRESSHOFF, P. and MOHAPATRA, S. 1981.Legume Cell and Tissue Culture. En
Tissue Culture of Economically Important Plants. Proceedings Of The International
Symposium Held at National University Of Singapore. Singapore.
HARA, Y. 1996. Research on the Production of Useful Compounds by Plant Cell
Cultures in Japan. En Plant Cell Culture and Secondary Metabolism. CRC Press
HOPKINS, W. 1999. Introduction to Plant Physiology. 2nd Ed. John Wiley & Sons.
New York.
JACOBSEN H.J. 1983. Biochemical mechanisms of plant hormona activity. En:
Handbook of Plant Cell Culture. Vol. 1. ( Ed. por D. A. Evans, W. R. Sharp, P. V.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Amirato, Y.Yamada). Collier Macmillan Publishers. Londres.
JAIME, J.A. 1994. Los tubérculos y granos andinos en el Perú. Folleto UNAS. Tingo
María.
JOHNS, T. 1980. Ethnobotany and phytochemistry of Tropaeolum tuberosum and
Lepidium meyenii from Andean South America. Faculty of graduate Studies
University of British Columbia.
JOSEFSSON, E. 1967.Distribution of Thioglucosides in different parts of Brassica
Plants. Phytochemistry, Vol. 6. Pp. 1617 – 1627.
KING. S. R. 1987. Four Endemic Andean tuber Crops: Promising Food Resources
for Agricultural Diversification. Mountain Research and Development. 7(1):43-52.
KING, S. R. 1988. Economic Botany of the Andean tuber Crop Complex: Lepidium
meyenii, Oxalis tuberosa, Tropaeolum tuberosum, and Ullucus Tuberosus. Thesis for
Doctor of Philosophy. NY
KJAER, A. 1968. Glucosinolates in Lepidium bonaeriense L. Phytochemistry 7:1663 -
1666.
KJAER, A. 1970. Glucosinolates in Some New World Species of Capparidaceae.
Phytochemistry. Vol 9. Pp 601 – 602.
KJAER, A. 1970. Glucosinolates in the Caricaceae. Phytochemistry. Vol 9. pp 591-
593.
KJAER, A. 1973. The Natural Distribution of Glucosinolates: A Uniform Group of
Sulphur - Containing Glucosides. Nobel 25. Chemistry in Botanical Classification:

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
229 - 234.
KRIKORIAN, A. D. 1991. Medios de cultivo: generalidades, composición y
preparación. En: Cultivo de Tejidos en la Agricultura. (Ed. por W. M. Roca y L. A.
Mroginski). CIAT. Cali. Pp 41 – 78.
LAEMMLI, U.K. (1970) Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227, 680-685
LARSEN, P.O. 1981. The biochemistry of plants. Vol 7. Academic Press. 501 - 525.
LEON, J. 1964. Plantas alimenticias andinas. Bol. Tec. del Instituto Interamericano
de Ciencias Agrícolas Zona Andina. Lima. 6: 43 - 46.
LOCK DE UGAZ, O. 1994. Investigación fitoquímica. Métodos en el estudio de
Productos naturales. PUCP. Fondo Editorial. Lima.
LOCKWOOD, G.; y BELKHIRI, A. 1991. Glucosinolate Spectrum of Some Algerian
Cruciferae. Pl. Syst. Evol. 176: 11 – 20.
MACGIBBON, D.B. y ALLISON, R.M. 1970. A method for the separation and
detection of plant glucosinolases (myrosinases). Phytochemistry. Vol 9: 541 – 544.
MICROSOFT® ENCARTA® ONLINE ENCYCLOPAEDIA 2001. "Chromatography,"
http://encarta.msn.com © 1997-2001 Microsoft Corporation. All rights reserved.
MITHEN R., CLARKE J., LISTER C. y DEAN C. 1995. Genetics of Aliphatic
Glucosinolates. III. Side Chain Structure of Aliphatic Glucosinolates in Arabidopsis
thaliana. Heredity 74: 210-215.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
MORRIS, P y ALTOONA, T. 1994. Tissue Culture and Transformation. En
Arabidopsis. (Ed. Por E.M. Meyerowitz y C. R. Somerville). Cold Spring Harbor
Laboratory Press. Portland Or.
NATIONAL RESEARCH COUNCIL. 1989. Lost Crops of the Incas: Little-known
plants of the Andes with Promise for Worldwide Cultivation. National Academy Press.
Washington, D.C.
NUÑEZ, P. y OCHOA, A. 2000. In vitro mass Cultivation of Cells and Tissues. En
Molecular Biotechnology for Plant Food Production. (Ed. por O. Paredes – López).
Ed. Technomic. Lancaster.
OBREGON, L. 1998. Maca, Planta Medicinal y Nutritiva. Instituto de Fitoterapia
Americano. Lima.
PETIARD, V.; COURTOIS, D.; GUERITTE, F.; LANCLOIS, N. y MOMPON, B. 1982.
New Alkaloids in Plant Tissue Cultures. 5th International Congress of Plant Tissue
and Cell Culture. Tokio.
PIÑAS, J. 1994. Los Tubérculos y Granos Andinos en el Perú. Folleto Universidad
Agraria de la Selva.
PIZZA, C.; DE FEO, V.; DE SIMONE, F. RASTRELLI, L. 1998. Qualita’ Nutrizionali
dei Tuberi Andini. En La Maca “Il Ginseng Delle Ande” E Altre Radici E Tuberi
Andini. Contributo alla conoscenza e valorizzazione delle risorse vegetali e animali
dell’America Latina. Quaderni IILA . Serie scienza. IILA. Roma.
PURRINGTON, C. 2000. Costs of Resistance. Current Opinion in Plant Biology
3:305-308.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
QUIROS, C.; EPPERSON, A; HU, J; y HOLLE, M. 1996. Physiological studies and
determination of chromosome number in maca, Lepidium Meyenii (Brassicaceae).
Economic Botany 50(2):216-223.
QUIROS, C. y ALIAGA, R. 1996. Maca Lepidium meyenii Walpers. En Andean Roots
and Tubers: Ahipa, Arracacha, Maca and Yacón. IPGRI. Roma.
QUIROS, C. 1999. Genética de la maca y especies relacionadas. Curso Taller
Internacional sobre Maca. Universidad Nacional Agraria La Molina. Lima – Perú.
RANDERATH, K. 1968. Thin-layer chromatography. Verlag Chemie – Academic
Press. Weiheim/Bergstr.
REISCH, R. 1983. Genetic variability in regenerated plants. En: Handbook of Plant
Cell Culture. Vol. 1. ( Ed. por D. A. Evans, W. R. Sharp, P. V. Amirato, Y.Yamada).
Collier Macmillan Publishers. Londres.
ROCA, W. M. Y MROGINSKI, L. A. 1991. Establecimiento de un laboratorio para el
cultivo de tejidos vegetales. En: Cultivo de Tejidos en la Agricultura. (Ed. por W.M.
Roca y L.A. Mroginski). CIAT. Cali. 211 – 238.
ROBERT, M.L.; REYES, J.; LOYOLA, V.M. 1991. Biosíntesis y bioconversión de
metabolitos secundarios por células cultivadas in vitro. En: Cultivo de Tejidos en la
Agricultura. (Ed. por W.M. Roca y L.A. Mroginski). CIAT. Cali. 211 – 238.
RODMAN, J. 1981. Divergence, Convergence, and Parallelism in Phytochemical
Characters: the Glucosinolate-Myrosinase System. En Phytochemistry and
Angiosperm phylogeny. (Ed por Young, D. A. y Seijler D.J). Praeyer Publishers. N.Y.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
SCHRIPSEMA, J; FUNG, S; y VERPOORTE,R. 1996. Screening of Plant Cell
Cultures for New Industrially Interesting Compounds. En Plant cell culture secondary
metabolism. Toward Industrial Application. (Ed. Por F. Dicosmo y M. Misawa). CRC.
SZWEYKOWSKA, A. 1974. The Role of Cytokinins in the Control of Cell Growth and
Differentiation in Culture. En Tissue Culture and Plant Science. (Ed. Por H.E.
Street). Academic Press. Londres.
TELLO, J., HERMAN, M., Y CALDERON, A. 1992. La maca (Lepidium meyenii
Walp); cultivo alimenticio potencial para las zonas altoandinas. Boletín de Lima. 81:
59 – 66.
XENOPHON, H; MACLEOD, J y MOREAU, M. 1981. Glucosinolates of Nine
Cruciferae and Two Capparaceae Species. Phytochemistry . Vol 20. Nro. 10. Pp
2355 – 2358.
YEOMAN, M & FORCHE,E. 1980. Cell Proliferation and Growth in Callus Cultures.
International review of Cytology, Supplement 11A. 1-24.
YLLESCA, M. 1994. Estudio químico y fitoquímico comparativo de 3 ecotipos de
Lepidium meyenii Walp. "Maca", procedente de Carhuamayo (Junín). Cátedra en
Bromatología. Facultad de Farmacia y Bioquímica. Universidad Nacional Mayor de
San Marcos. Lima.
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
APÉNDICE
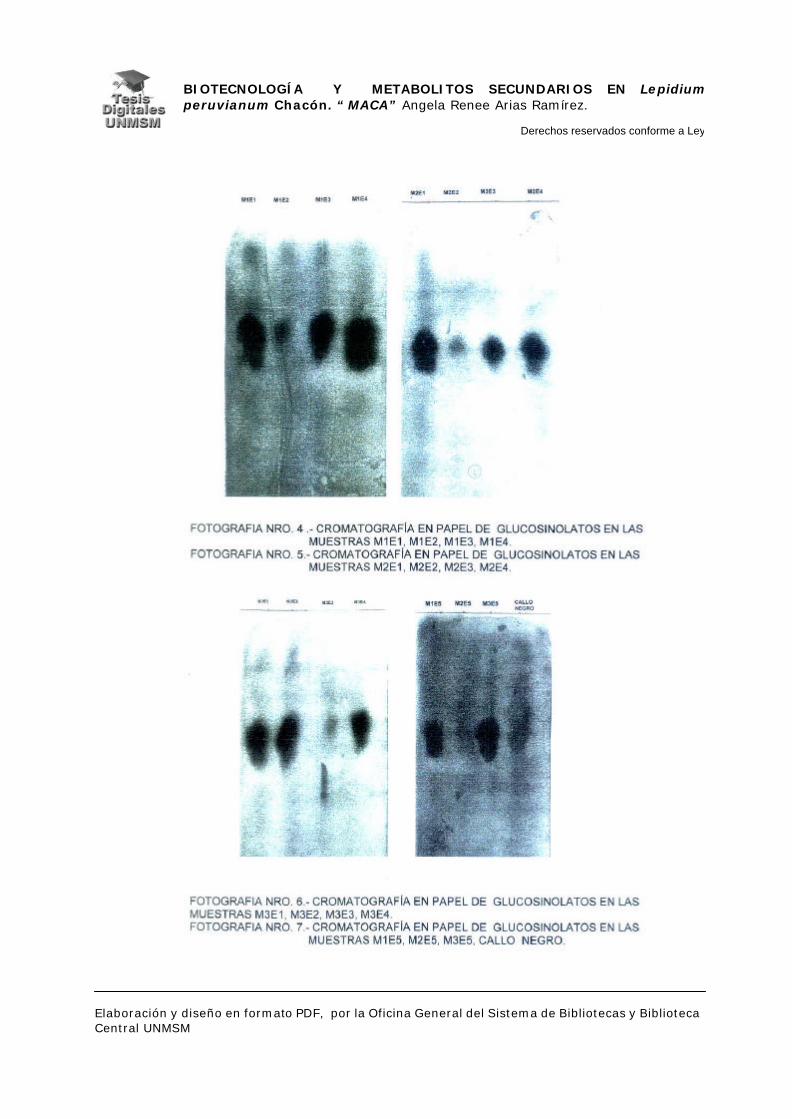
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
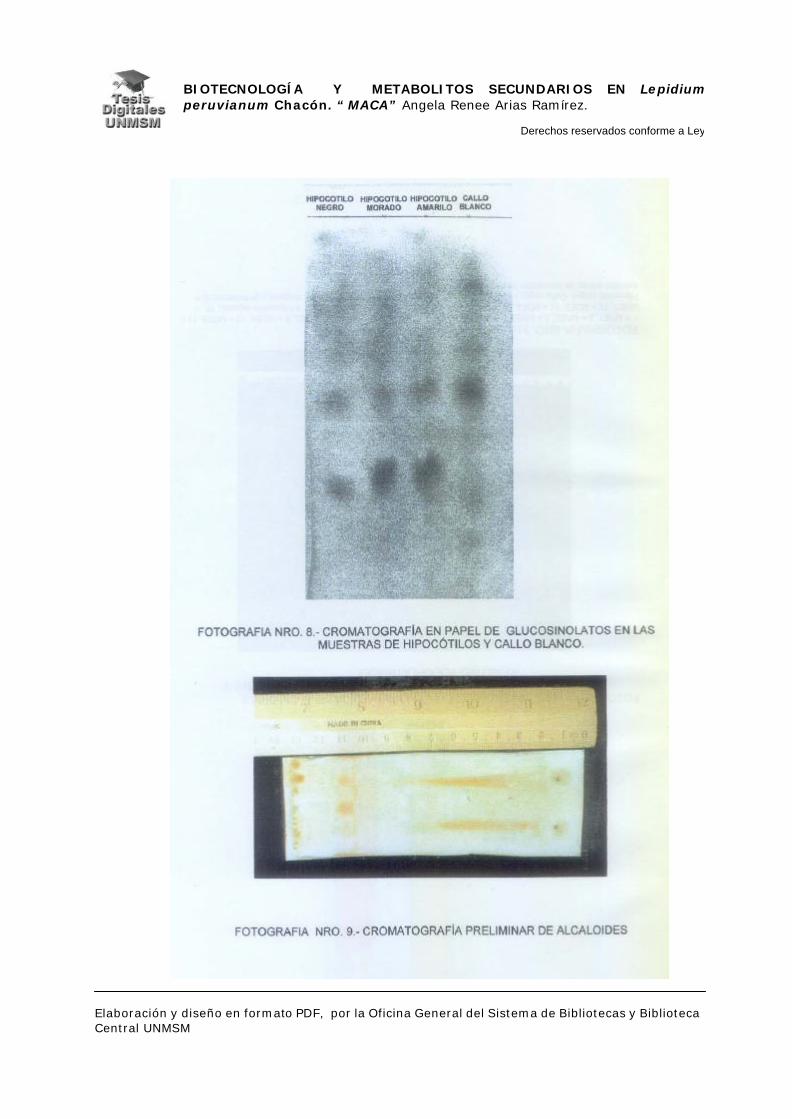
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
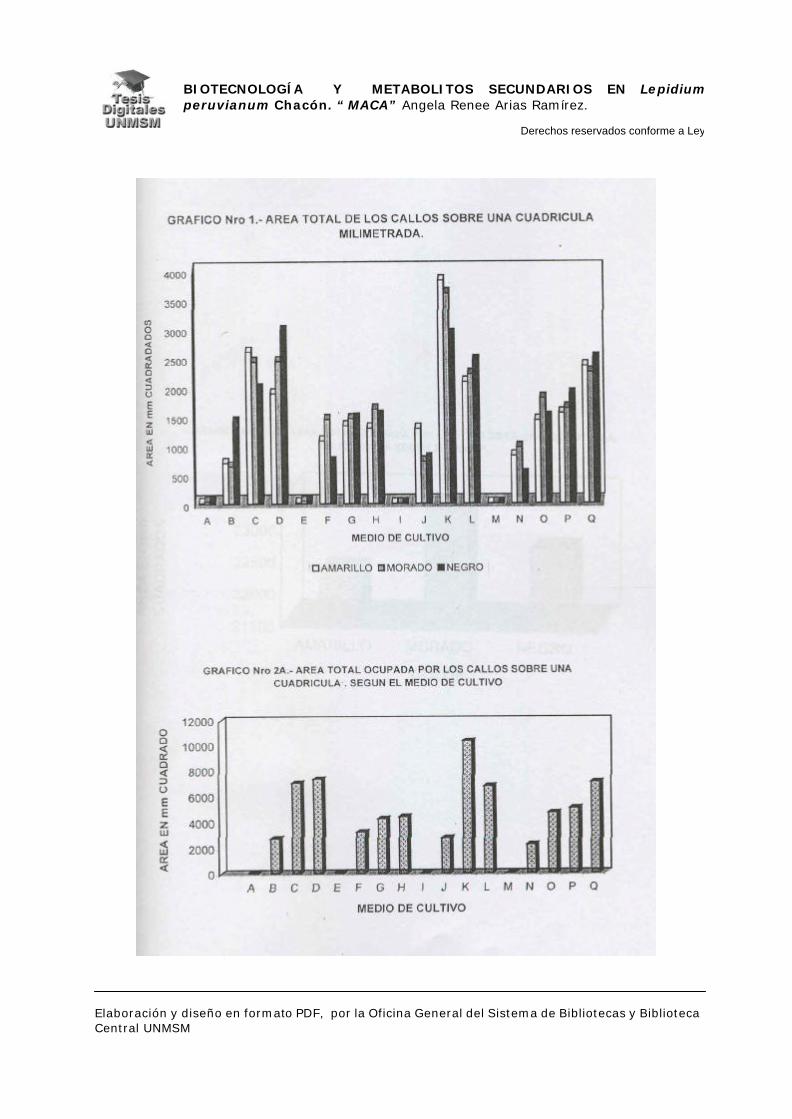
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
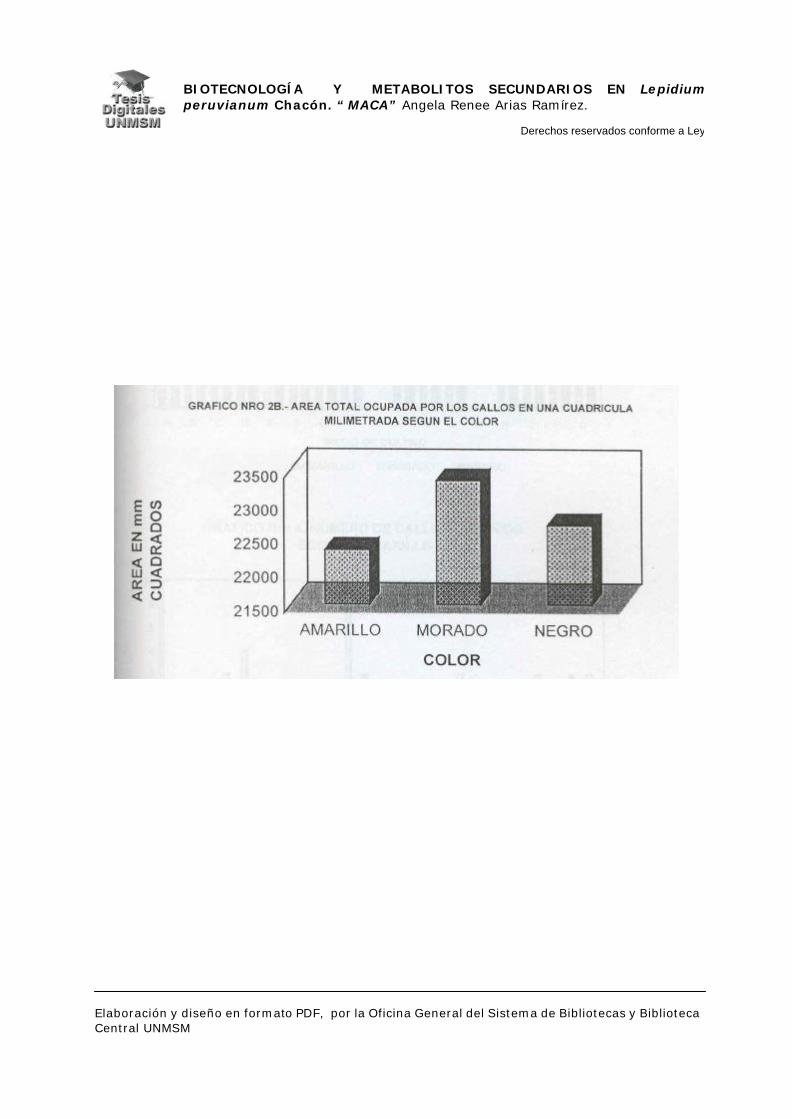
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
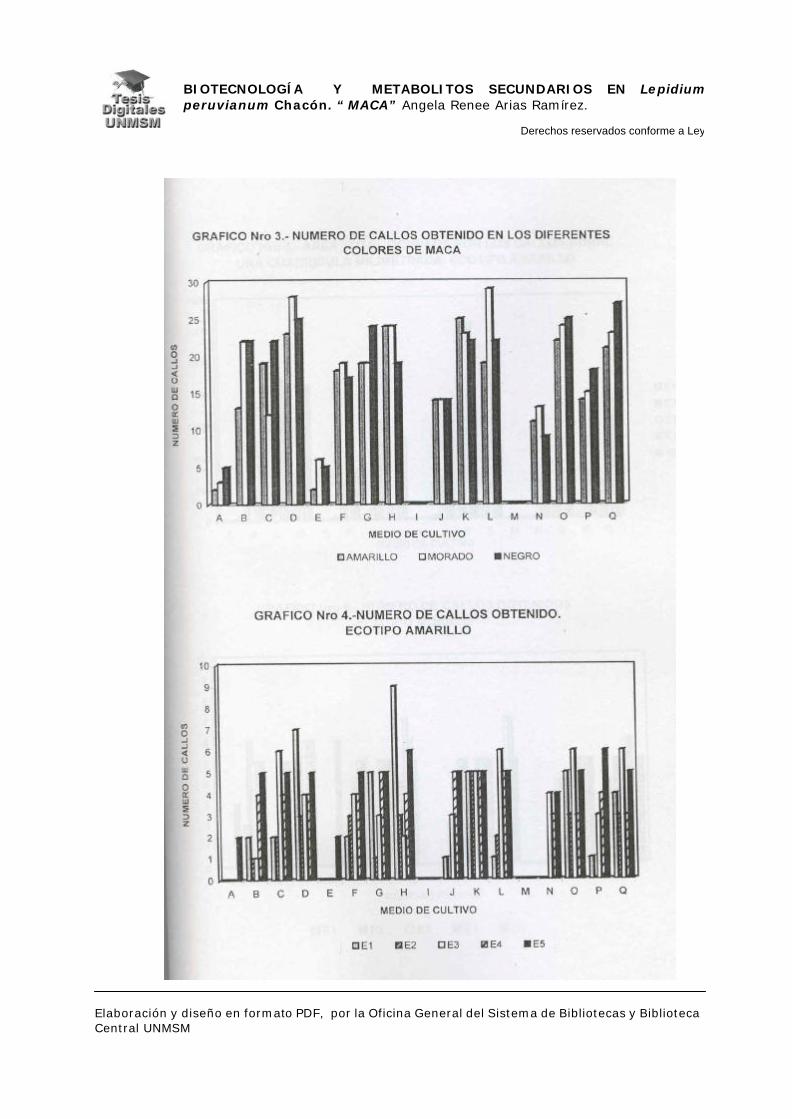
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
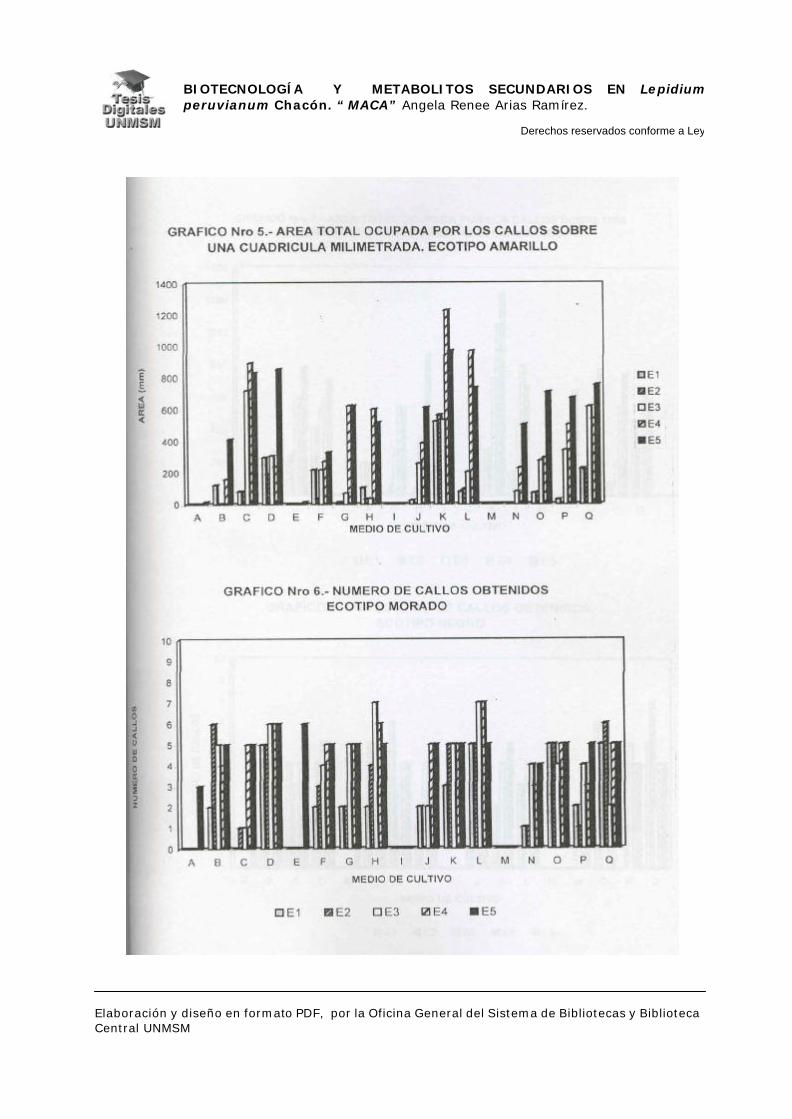
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
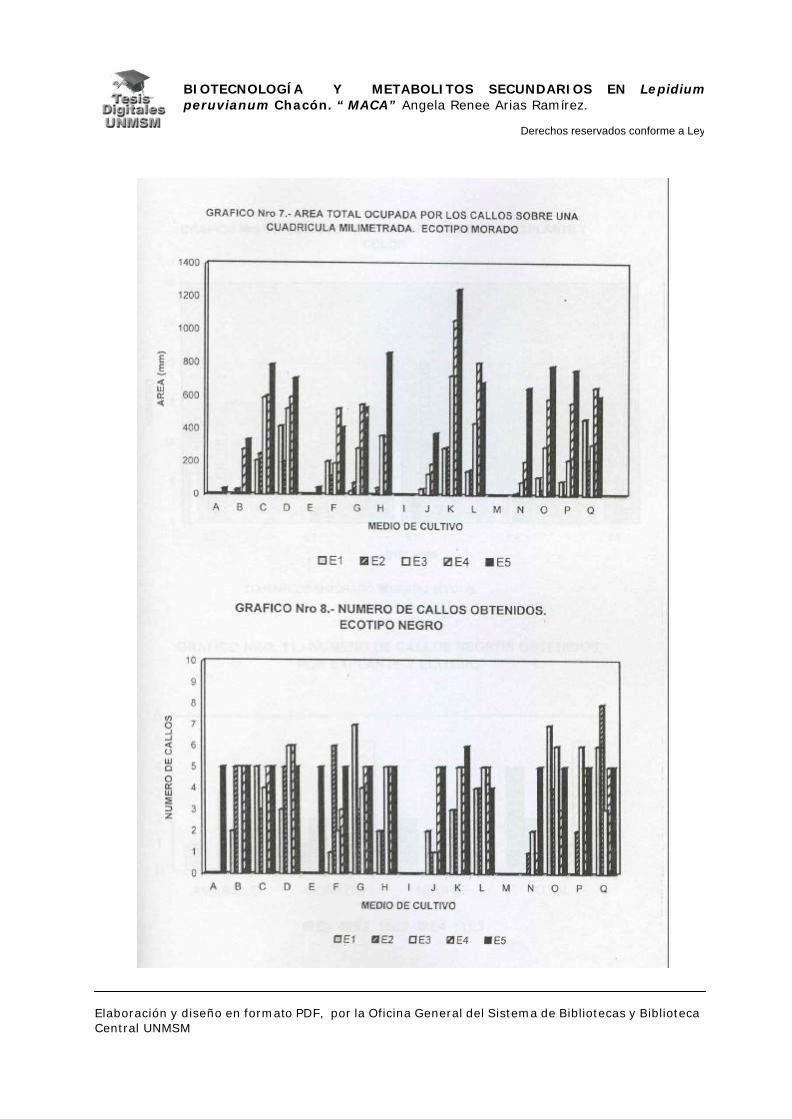
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
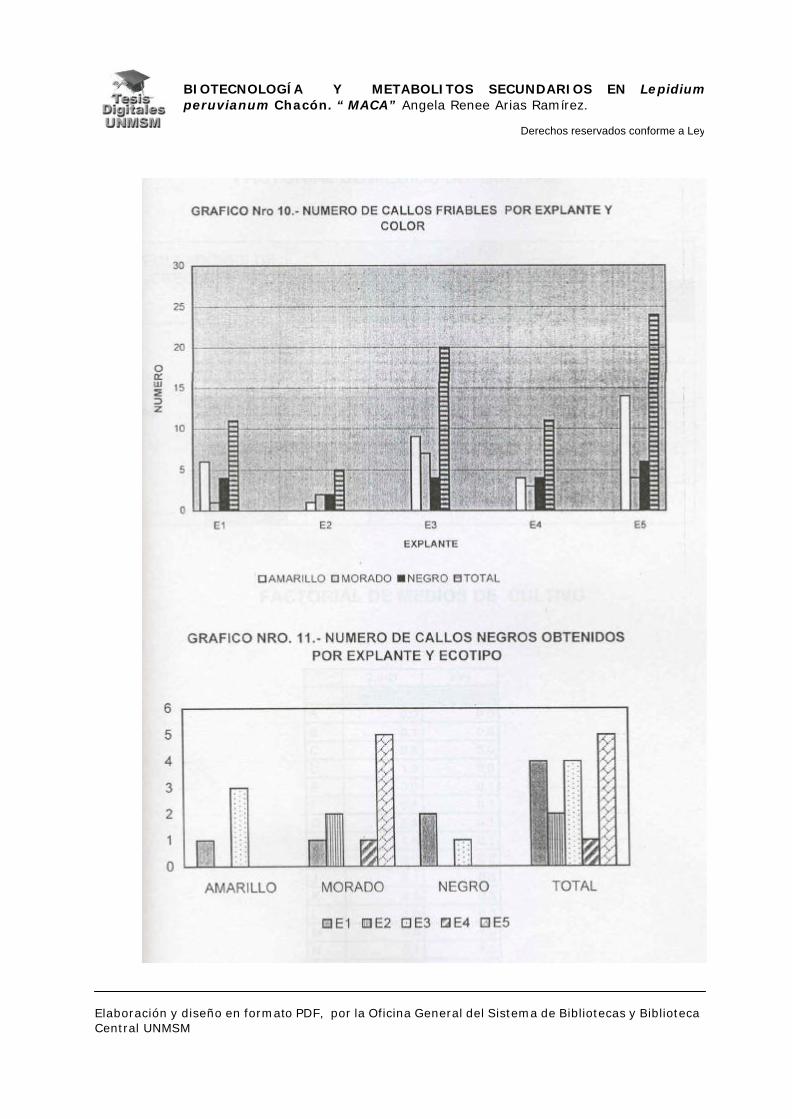
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
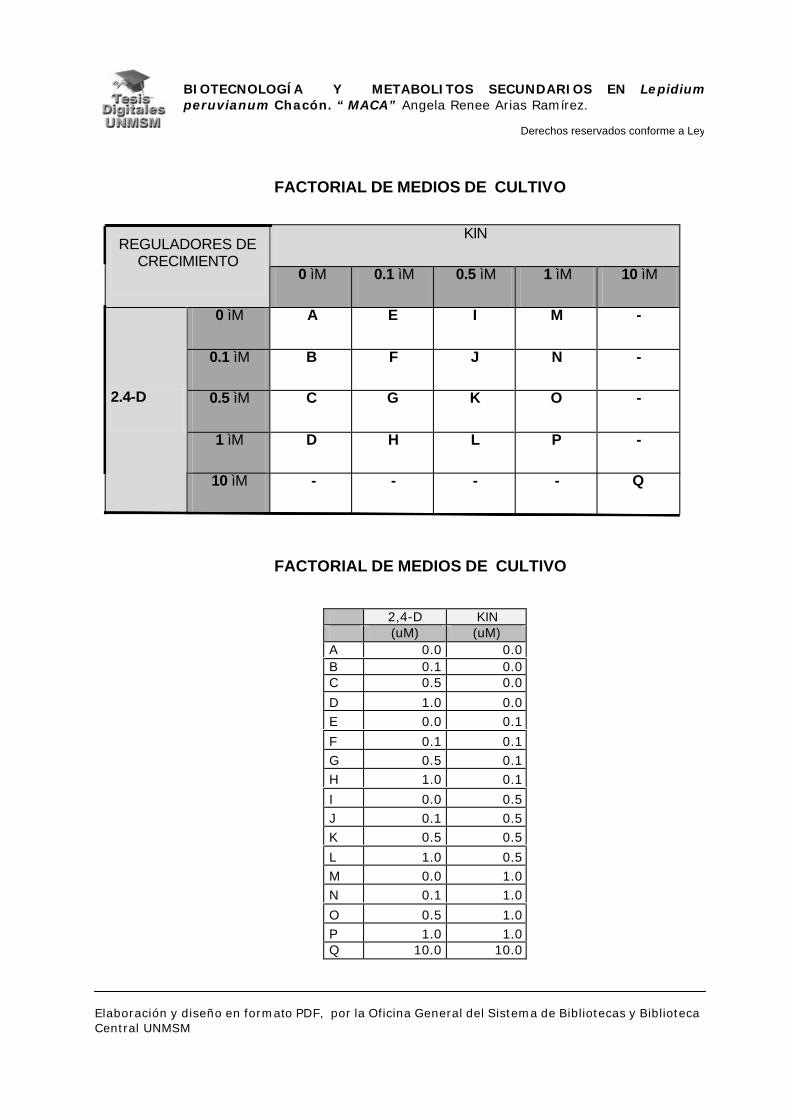
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
FACTORIAL DE MEDIOS DE CULTIVO
FACTORIAL DE MEDIOS DE CULTIVO
KIN REGULADORES DE
CRECIMIENTO 0 ìM 0.1 ìM 0.5 ìM 1 ìM 10 ìM
0 ìM A E I M -
0.1 ìM B F J N -
0.5 ìM C G K O -
1 ìM D H L P -
2.4-D
10 ìM - - - - Q
2,4-D KIN (uM) (uM) A 0.0 0.0 B 0.1 0.0 C 0.5 0.0
D 1.0 0.0 E 0.0 0.1
F 0.1 0.1 G 0.5 0.1 H 1.0 0.1
I 0.0 0.5 J 0.1 0.5 K 0.5 0.5
L 1.0 0.5 M 0.0 1.0 N 0.1 1.0
O 0.5 1.0 P 1.0 1.0 Q 10.0 10.0
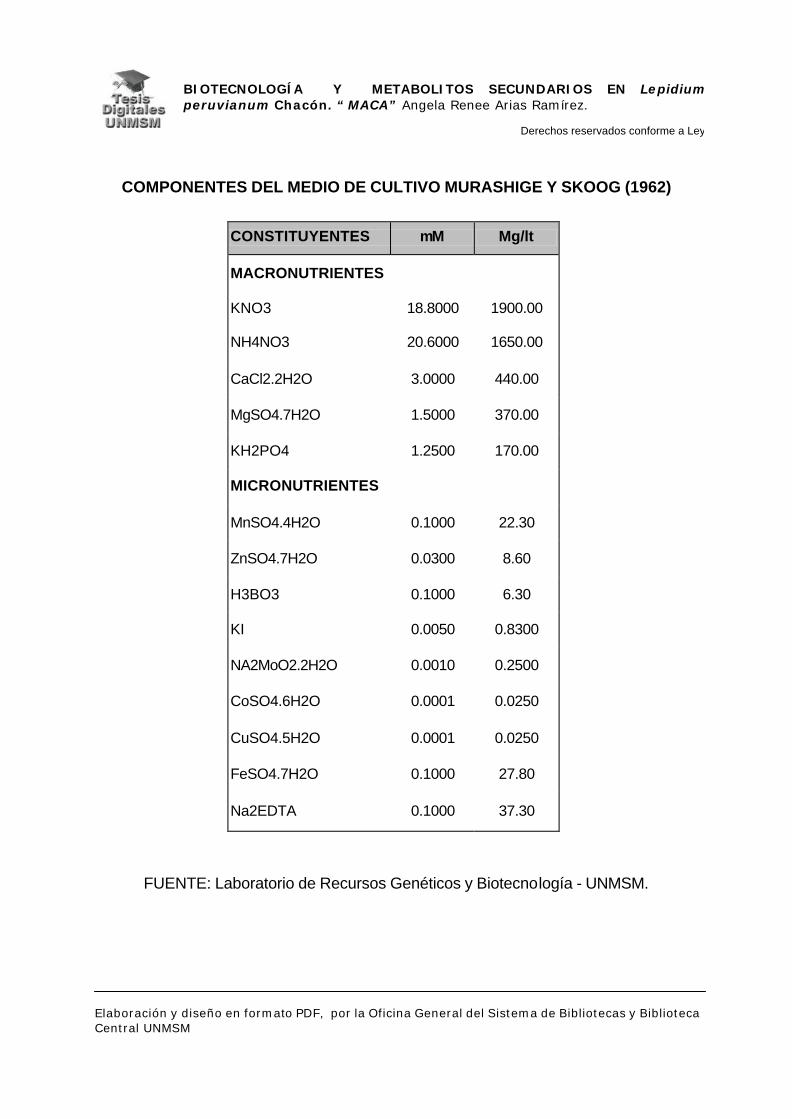
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
COMPONENTES DEL MEDIO DE CULTIVO MURASHIGE Y SKOOG (1962)
CONSTITUYENTES mM Mg/lt
MACRONUTRIENTES
KNO3 18.8000 1900.00
NH4NO3 20.6000 1650.00
CaCl2.2H2O 3.0000 440.00
MgSO4.7H2O 1.5000 370.00
KH2PO4 1.2500 170.00
MICRONUTRIENTES
MnSO4.4H2O 0.1000 22.30
ZnSO4.7H2O 0.0300 8.60
H3BO3 0.1000 6.30
KI 0.0050 0.8300
NA2MoO2.2H2O 0.0010 0.2500
CoSO4.6H2O 0.0001 0.0250
CuSO4.5H2O 0.0001 0.0250
FeSO4.7H2O 0.1000 27.80
Na2EDTA 0.1000 37.30
FUENTE: Laboratorio de Recursos Genéticos y Biotecnología - UNMSM.
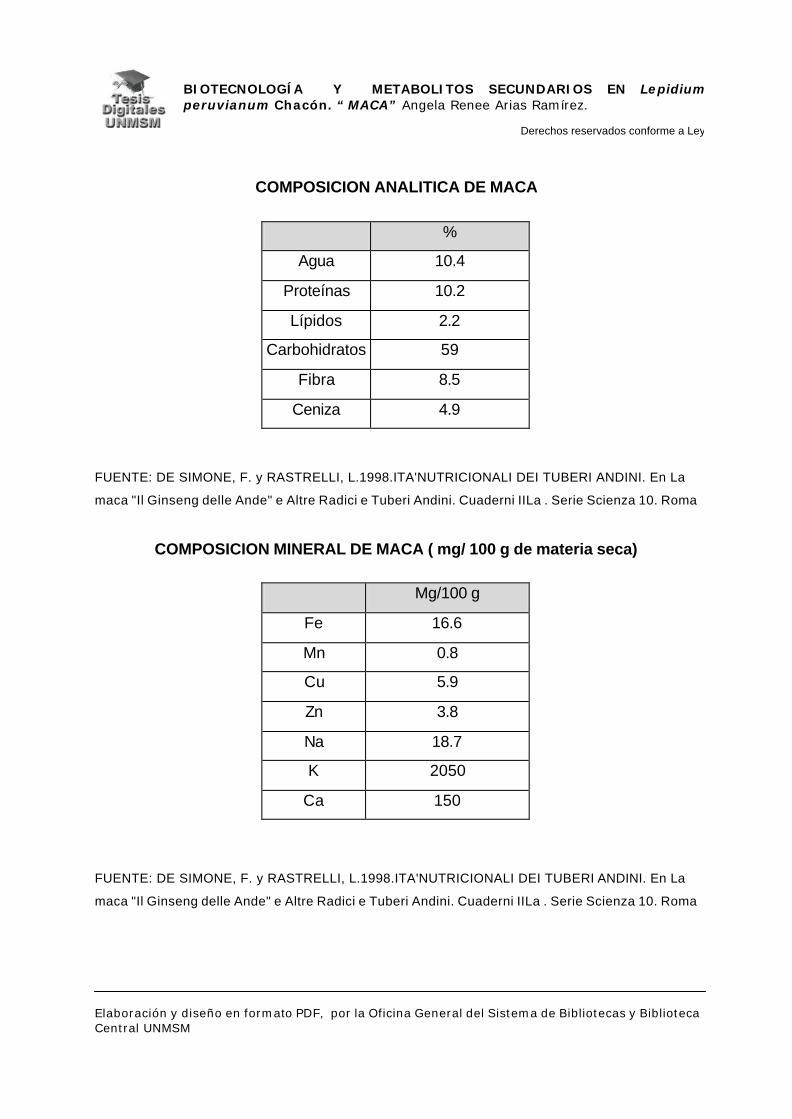
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
COMPOSICION ANALITICA DE MACA
%
Agua 10.4
Proteínas 10.2
Lípidos 2.2
Carbohidratos 59
Fibra 8.5
Ceniza 4.9
FUENTE: DE SIMONE, F. y RASTRELLI, L.1998.ITA'NUTRICIONALI DEI TUBERI ANDINI. En La
maca "Il Ginseng delle Ande" e Altre Radici e Tuberi Andini. Cuaderni IILa . Serie Scienza 10. Roma
COMPOSICION MINERAL DE MACA ( mg/ 100 g de materia seca)
Mg/100 g
Fe 16.6
Mn 0.8
Cu 5.9
Zn 3.8
Na 18.7
K 2050
Ca 150
FUENTE: DE SIMONE, F. y RASTRELLI, L.1998.ITA'NUTRICIONALI DEI TUBERI ANDINI. En La
maca "Il Ginseng delle Ande" e Altre Radici e Tuberi Andini. Cuaderni IILa . Serie Scienza 10. Roma
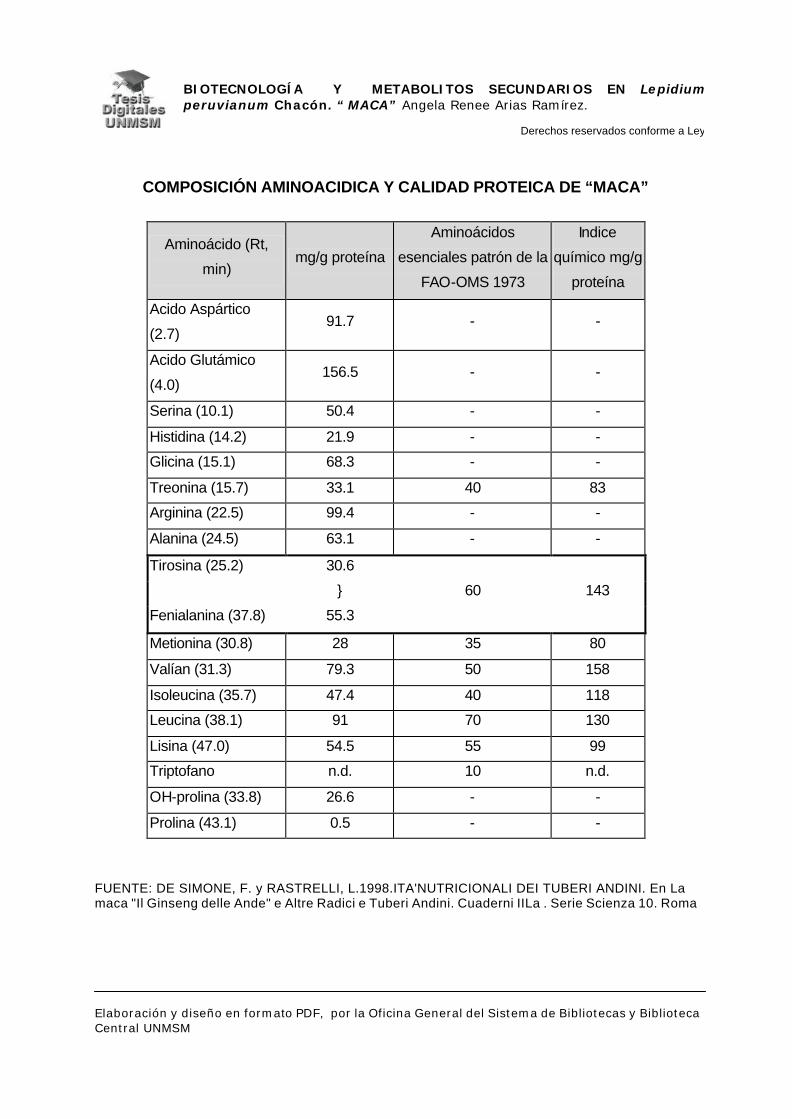
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
COMPOSICIÓN AMINOACIDICA Y CALIDAD PROTEICA DE “MACA”
Aminoácido (Rt,
min) mg/g proteína
Aminoácidos
esenciales patrón de la
FAO-OMS 1973
Indice
químico mg/g
proteína
Acido Aspártico
(2.7) 91.7 - -
Acido Glutámico
(4.0) 156.5 - -
Serina (10.1) 50.4 - -
Histidina (14.2) 21.9 - -
Glicina (15.1) 68.3 - -
Treonina (15.7) 33.1 40 83
Arginina (22.5) 99.4 - -
Alanina (24.5) 63.1 - -
Tirosina (25.2) 30.6
} 60 143
Fenialanina (37.8) 55.3
Metionina (30.8) 28 35 80
Valían (31.3) 79.3 50 158
Isoleucina (35.7) 47.4 40 118
Leucina (38.1) 91 70 130
Lisina (47.0) 54.5 55 99
Triptofano n.d. 10 n.d.
OH-prolina (33.8) 26.6 - -
Prolina (43.1) 0.5 - -
FUENTE: DE SIMONE, F. y RASTRELLI, L.1998.ITA'NUTRICIONALI DEI TUBERI ANDINI. En La maca "Il Ginseng delle Ande" e Altre Radici e Tuberi Andini. Cuaderni IILa . Serie Scienza 10. Roma

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
PROTOCOLO DE EXTRACCION Y CROMATOGRAFIA EN PAPEL DE
GLUCOSINOLATOS
Extracción
Macerar las muestras en etanol al 96% por 3 días como mínimo.
Filtrar
Guardar la solución y con el residuo hacer una nueva extracción con etanol.
Filtrar y guardar con la solución anterior.
Cromatografía en papel
Cortar las hojas de papel de 11 x 29 cm. Hacer tres dobleces a 2, 4 y cm del borde
del papel para que sirvan de soporte de la hoja en el tanque de cromatografía.
A 5 cm del borde hacer una marca con lápiz y hacer pequeñas equis en ella donde
se va a sembrar la muestra.
Colocar una alicuota de cada muestra en cada marca, dejar secar, repetir 40 veces.
Colocar los papeles en las bandejas de soporte del tanque de cromatografía, y
sujetar con las varillas respectivas. Agregar el solvente a las bandejas y tapar
herméticamente el tanque.
Dejar desarrollar durante la noche.
Retirar del tanque de cromatografía y secar.
Pulverizar con una solución de nitrato de plata : acetona 15 mg/ml, dejar secar unos
minutos
Pulverizar con una solución de KOH : Etanol 20 mg/ml, dejar secar unos minutos.
Colocar los cromatogramas en una estufa caliente.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Retirar de la estufa cuando se forman las manchas negras, (indicadoras de la
presencia de glucosinolatos).
Sumergir en una solución de hiposulfito de potasio al 5% por 20 - 24 para
decolorar.
Secar Fotocopiar inmediatamente.
Preparación del solvente Hacer una solución de n-butanol:etanol:agua 4:1:4 en una pera de decantación.
La mezcla se realiza en orden: mezclar primero el n-butanol y el etanol, agitar y
agregar el agua, agitar bien y dejar reposar sin mover 24 hr.
Se obtienen dos fases: una acuosa y una orgánica (superior), desechar la fase
acuosa.
Preparación del revelador Solución de nitrato de plata : acetona (para 100 ml.) Preparar una solución saturada de nitrato de plata y agua: diluir 1.5 gr. de nitrato de
plata en 0.5 ml. de agua destilada, si no se disuelve calentar ligeramente a baño
María
Disolver en 80 ml. de acetona y agregar agua destilada hasta disolver el precipitado
que se haya formado.
Usar inmediatamente.
Solución de hidróxido de potasio : etanol. Solución al 2% (para 100 ml)
Disolver 2 gr de KOH en 0.5 ml de agua destilada y completar a 100 ml con etanol.
Preparación de la solución de hiposulfito de sodio al 5% (decolorante)
Disolver 2 gr de hiposulfito de sodio en 400 ml de agua destilada.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
PROTOCOLO DE EXTRACCION Y CROMATOGRAFIA DE ALCALOIDES
Extracción
A.- extracto crudo:
Colocar las muestra estabilizada en un frasco y humedecer con amoniacoy dejar en
oscuridad por 48 hr.
Agregar éter etílico agitar y reposar varios días.
Filtrar o centrifugar.
B.- extracto lavado:
Macerar la muestra con una solución de agua : etanol (4:1).
Acidificar hasta pH 3 ó 4 con HCl.
Centrifugar o filtra para recuperar la fase acuosa.
Alcalinizar la fase acuosa con NaOH u otro álcali, hasta pH 8 ó 9.
Poner en una pera de decantación y agregar una cantidad similar de eter etilico,
agitar y recuperar la fase orgánica.
Repetir esta extracción.
Cromatografía de capa fina ascendente.
Sembrar las muestras en un cromatograma de silicagel
Colocar el solvente en el tanque de cromatografía de capa fina ascendente, cerrar
herméticamente y dejar saturar.
Colocar la cromatoplaca en el tanque, cerrar herméticamente y dejar desarrollar.
Cuando el solvente llega al tope de la placa, sacar la cromatoplaca del tanque, dejar
secar y pulverizar con el revelador.

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
Preparación del solvente
Mezclar en el tanque de cromatografía cloroformo:metanol u otra sol en las
proporciones adecuadas en nuestro caso la proporción fue 2 : 0.12.
Preparación del revelador
Reactivo de Dragendorff (modificado por Munnier)
SOLUCION I.- Disolver 17 gr de subnitrato de bismuto y 200 gr. de ácido tartárico en
800 ml de agua destilada.
SOLUCION II.- Disolver 160 gr de yoduro de potasio en 400 ml de agua destilada.
Mezclar las soluciones I y II en una proporción 1 : 1
PARA USAR: disolver 100 gr. De ácido tartárico en 50 ml de esta mezcla y 500 ml
de agua destilada.
El stock puede almacenarse durante meses. La solución diluida puede almacenarse
por unas semanas.
Reactivo Yodoplatinado
Diluir 3ml de ácido cloroplatínico al 10% en 97 ml de agua destilada.
Agregar 100 ml de solución de yoduro de potasio al 6%.
Este reactivo puede almacenarse por largo tiempo.
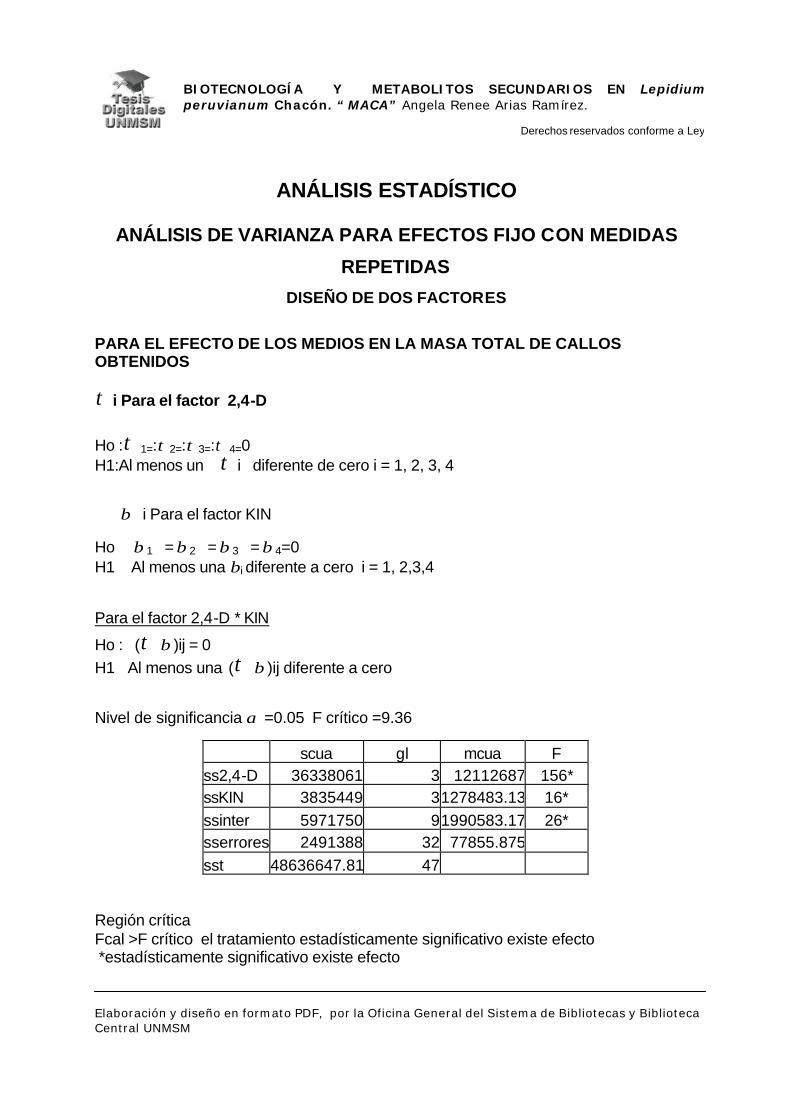
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
ANÁLISIS ESTADÍSTICO
ANÁLISIS DE VARIANZA PARA EFECTOS FIJO CON MEDIDAS
REPETIDAS
DISEÑO DE DOS FACTORES
PARA EL EFECTO DE LOS MEDIOS EN LA MASA TOTAL DE CALLOS OBTENIDOS τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i = 1, 2, 3, 4
β i Para el factor KIN
Ho β 1 = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i = 1, 2,3,4
Para el factor 2,4-D * KIN
Ho : (τ β )ij = 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Región crítica Fcal >F crítico el tratamiento estadísticamente significativo existe efecto *estadísticamente significativo existe efecto
scua gl mcua F ss2,4-D 36338061 3 12112687 156* ssKIN 3835449 31278483.13 16* ssinter 5971750 91990583.17 26* sserrores 2491388 32 77855.875 sst 48636647.81 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL NUMERO DE CALLOS OBTENIDO Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3,4
Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Región crítica Fcal >F crítico el tratamiento estadísticamente significativo existe efecto *estadísticamente significativo existe efecto
Scua gl mcua f
ss2,4-D 3071 3 1024 29.3*
ssKIN 112.5625 3 37.52083333 17*
ssinter 284 9 94.673611112.705639747
sserrores 11197 32 349.9121094
sst 14665 47
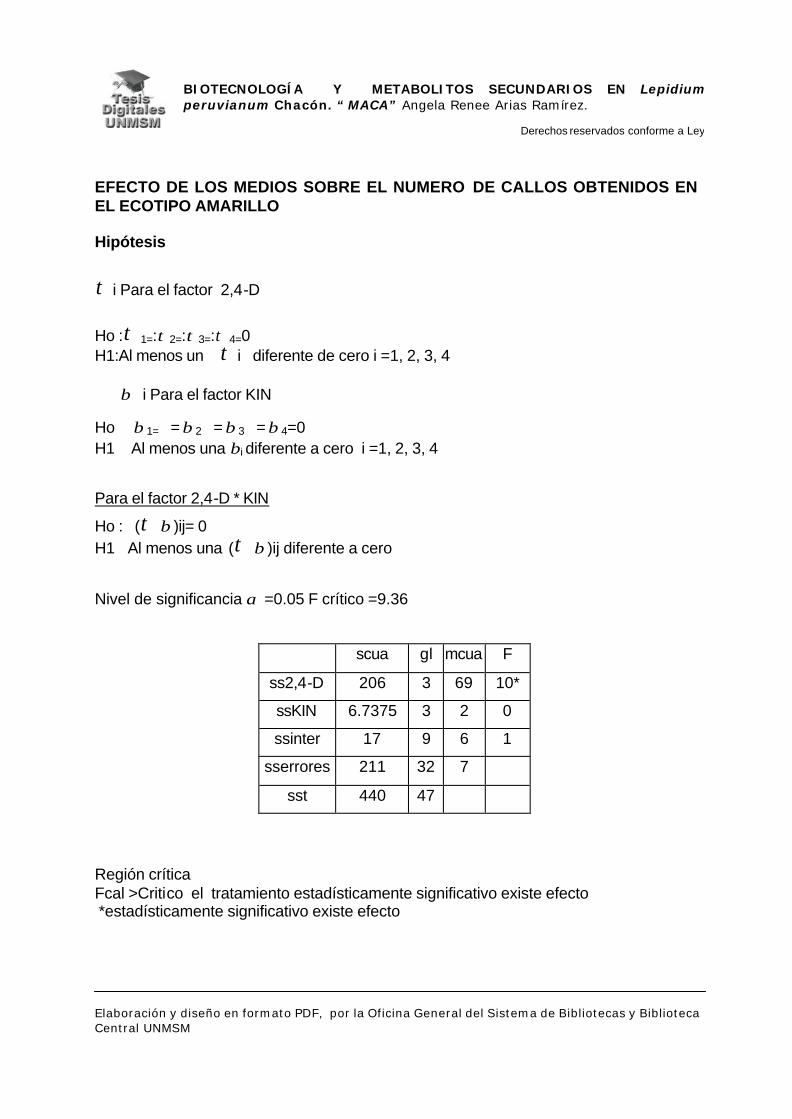
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL NUMERO DE CALLOS OBTENIDOS EN EL ECOTIPO AMARILLO Hipótesis
τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4
Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Región crítica Fcal >Critico el tratamiento estadísticamente significativo existe efecto *estadísticamente significativo existe efecto
scua gl mcua F
ss2,4-D 206 3 69 10*
ssKIN 6.7375 3 2 0
ssinter 17 9 6 1
sserrores 211 32 7
sst 440 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL ÁREA OCUPADA POR LOS CALLOS EN EL ECOTIPO AMARILLO Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4 Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero Nivel de significancia α =0.05 F crítico =9.36
Regios crítica Fcal >F crítico el tratamiento estadísticamente significativo existe efecto *estadísticamente significativo existe efecto
scua gl mcua f ss2,4-D 2350703 3 783568 13*
SsKIN 399929 3 133310 2
ssinter 542059 9 180686 3
sserrores 3988904.4 64 62326.6313 Sst 7281595.55 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL NUMERO DE CALLOS OBTENIDOS EN EL ECOTIPO MORADO Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4 Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero Nivel de significancia α =0.05 F crítico =9.36
Región crítica Fcal >F crítico el tratamiento estadísticamente significativo existe efecto *estadísticamente significativo existe efecto
scua gl mcua f ss2,4-D 213 3 71 31*
ssKIN 8 3 3 1
ssinter 50 9 17 7
sserrores 148 32 2 sst 419 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL ÁREA OCUPADA POR LOS CALLOS EN EL ECOTIPO MORADO Hipótesis
τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4
Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Región crítica Fcal >F crítico el tratamiemto estadísticamente significativo exlste efecto *estadísticamente significativo exlste efecto
scua gl mcua f ss2,4-D 3E+06 3 875861 14*
ssKIN 1E+05 3 49762 1
ssinter 6E+05 9 189788 3
sserrores4E+06 64 64090 sst 7E+06 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL NUMERO DE CALLOS OBTENIDOS EN EL ECOTIPO NEGRO Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4
Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Region crítica Fcal >F crítico el tratamiemto estadísticamente significativo exlste efecto *estadísticamente significativo exlste efecto
scua gl mcua f ss2,4-D 207 3 69 38*
ssKIN 13 3 4 15*
ssinter 17 9 6 4
sserrores 927 64 14 sst 1165 47

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DE LOS MEDIOS SOBRE EL ÁREA OCUPADA POR LOS CALLOS EN EL ECOTIPO NEGRO Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4 Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Region crítica Fcal >F crítico el tratamiemto estadísticamente significativo exlste efecto *estadísticamente significativo exlste efecto.
scua gl mcua f ss2,4-D 2E+06 3E+00 8E+05 60*
ssKIN 3E+05 3E+00 1E+05 70*
ssinter 2E+06 9E+00 8E+05 50*
sserrores 9E+05 6E+01 1E+04 sst 6E+06 5E+01
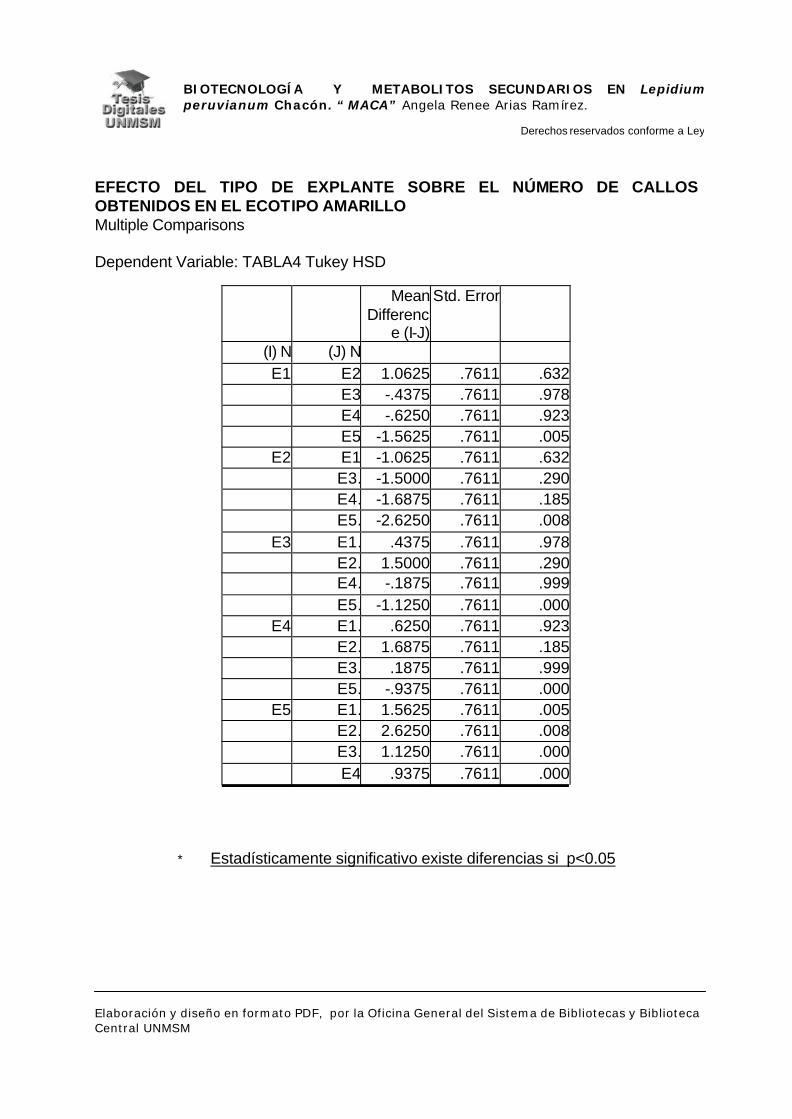
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL NÚMERO DE CALLOS OBTENIDOS EN EL ECOTIPO AMARILLO Multiple Comparisons Dependent Variable: TABLA4 Tukey HSD
* Estadísticamente significativo existe diferencias si p<0.05
Mean Differenc
e (I-J)
Std. Error
(I) N (J) NE1 E2 1.0625 .7611 .632
E3 -.4375 .7611 .978E4 -.6250 .7611 .923E5 -1.5625 .7611 .005
E2 E1 -1.0625 .7611 .632E3. -1.5000 .7611 .290E4. -1.6875 .7611 .185E5. -2.6250 .7611 .008
E3 E1. .4375 .7611 .978E2. 1.5000 .7611 .290E4. -.1875 .7611 .999E5. -1.1250 .7611 .000
E4 E1. .6250 .7611 .923E2. 1.6875 .7611 .185E3. .1875 .7611 .999E5. -.9375 .7611 .000
E5 E1. 1.5625 .7611 .005E2. 2.6250 .7611 .008E3. 1.1250 .7611 .000E4 .9375 .7611 .000
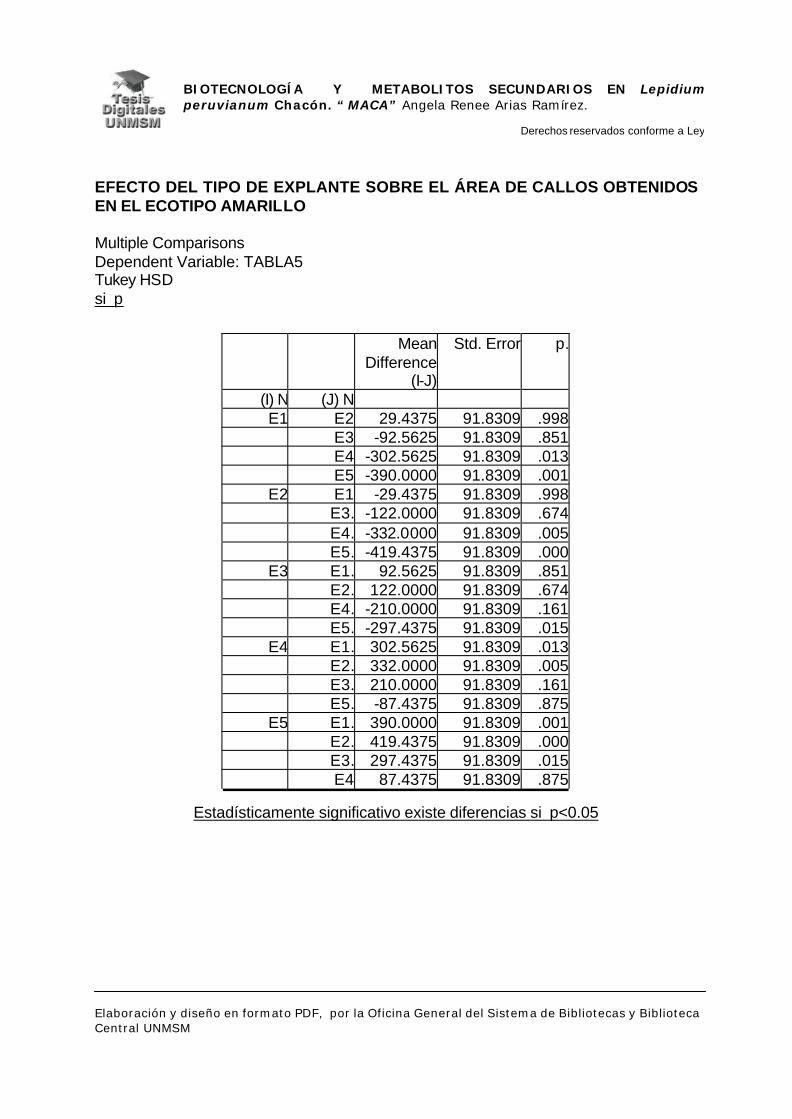
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL ÁREA DE CALLOS OBTENIDOS EN EL ECOTIPO AMARILLO Multiple Comparisons Dependent Variable: TABLA5 Tukey HSD si p
Estadísticamente significativo existe diferencias si p<0.05
Mean Difference
(I-J)
Std. Error p.
(I) N (J) NE1 E2 29.4375 91.8309 .998
E3 -92.5625 91.8309 .851E4 -302.5625 91.8309 .013E5 -390.0000 91.8309 .001
E2 E1 -29.4375 91.8309 .998E3. -122.0000 91.8309 .674E4. -332.0000 91.8309 .005E5. -419.4375 91.8309 .000
E3 E1. 92.5625 91.8309 .851E2. 122.0000 91.8309 .674E4. -210.0000 91.8309 .161E5. -297.4375 91.8309 .015
E4 E1. 302.5625 91.8309 .013E2. 332.0000 91.8309 .005E3. 210.0000 91.8309 .161E5. -87.4375 91.8309 .875
E5 E1. 390.0000 91.8309 .001E2. 419.4375 91.8309 .000E3. 297.4375 91.8309 .015E4 87.4375 91.8309 .875
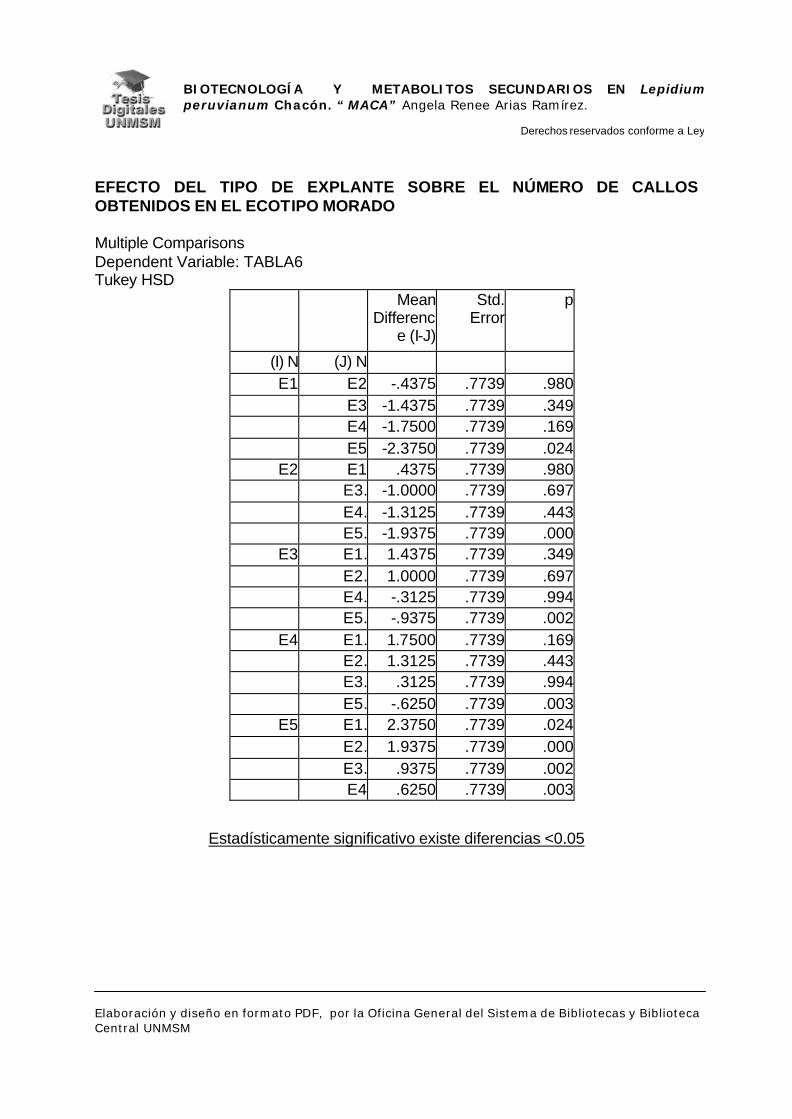
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL NÚMERO DE CALLOS OBTENIDOS EN EL ECOTIPO MORADO Multiple Comparisons Dependent Variable: TABLA6 Tukey HSD
Mean Differenc
e (I-J)
Std. Error
p
(I) N (J) NE1 E2 -.4375 .7739 .980
E3 -1.4375 .7739 .349E4 -1.7500 .7739 .169E5 -2.3750 .7739 .024
E2 E1 .4375 .7739 .980E3. -1.0000 .7739 .697E4. -1.3125 .7739 .443E5. -1.9375 .7739 .000
E3 E1. 1.4375 .7739 .349E2. 1.0000 .7739 .697E4. -.3125 .7739 .994E5. -.9375 .7739 .002
E4 E1. 1.7500 .7739 .169E2. 1.3125 .7739 .443E3. .3125 .7739 .994E5. -.6250 .7739 .003
E5 E1. 2.3750 .7739 .024E2. 1.9375 .7739 .000E3. .9375 .7739 .002E4 .6250 .7739 .003
Estadísticamente significativo existe diferencias <0.05
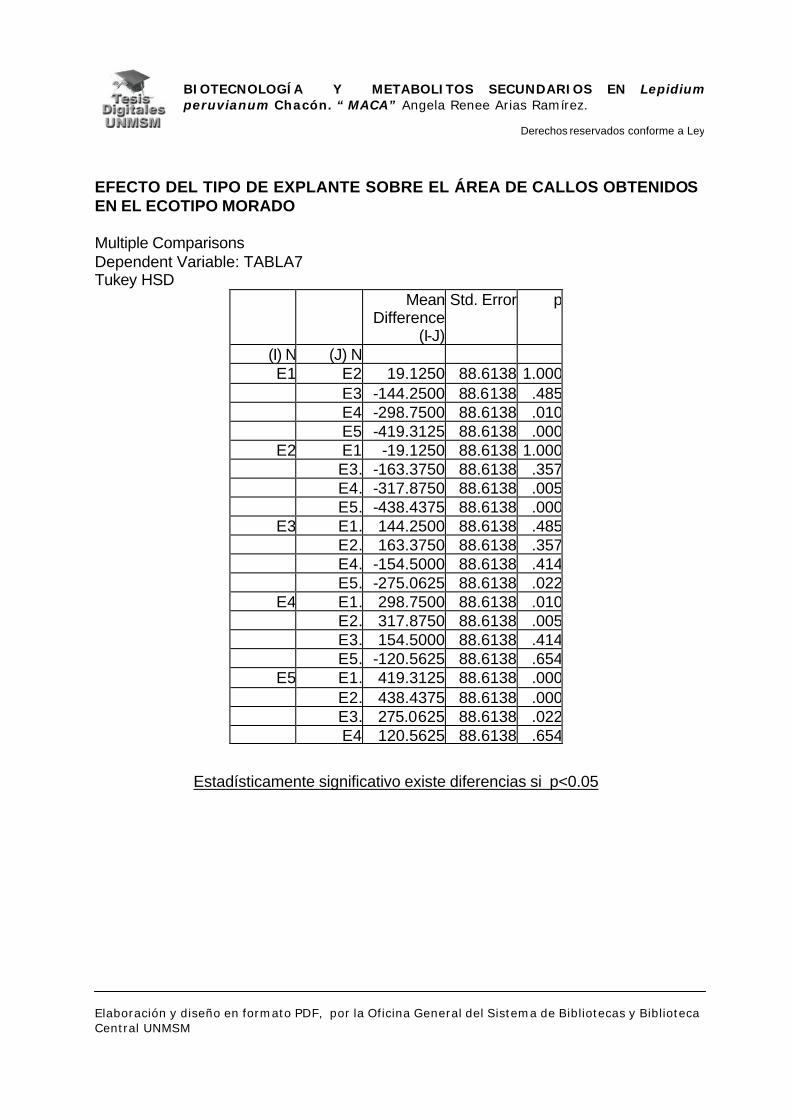
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL ÁREA DE CALLOS OBTENIDOS EN EL ECOTIPO MORADO Multiple Comparisons Dependent Variable: TABLA7 Tukey HSD
Mean Difference
(I-J)
Std. Error p
(I) N (J) NE1 E2 19.1250 88.6138 1.000
E3 -144.2500 88.6138 .485E4 -298.7500 88.6138 .010E5 -419.3125 88.6138 .000
E2 E1 -19.1250 88.6138 1.000E3. -163.3750 88.6138 .357E4. -317.8750 88.6138 .005E5. -438.4375 88.6138 .000
E3 E1. 144.2500 88.6138 .485E2. 163.3750 88.6138 .357E4. -154.5000 88.6138 .414E5. -275.0625 88.6138 .022
E4 E1. 298.7500 88.6138 .010E2. 317.8750 88.6138 .005E3. 154.5000 88.6138 .414E5. -120.5625 88.6138 .654
E5 E1. 419.3125 88.6138 .000E2. 438.4375 88.6138 .000E3. 275.0625 88.6138 .022E4 120.5625 88.6138 .654
Estadísticamente significativo existe diferencias si p<0.05
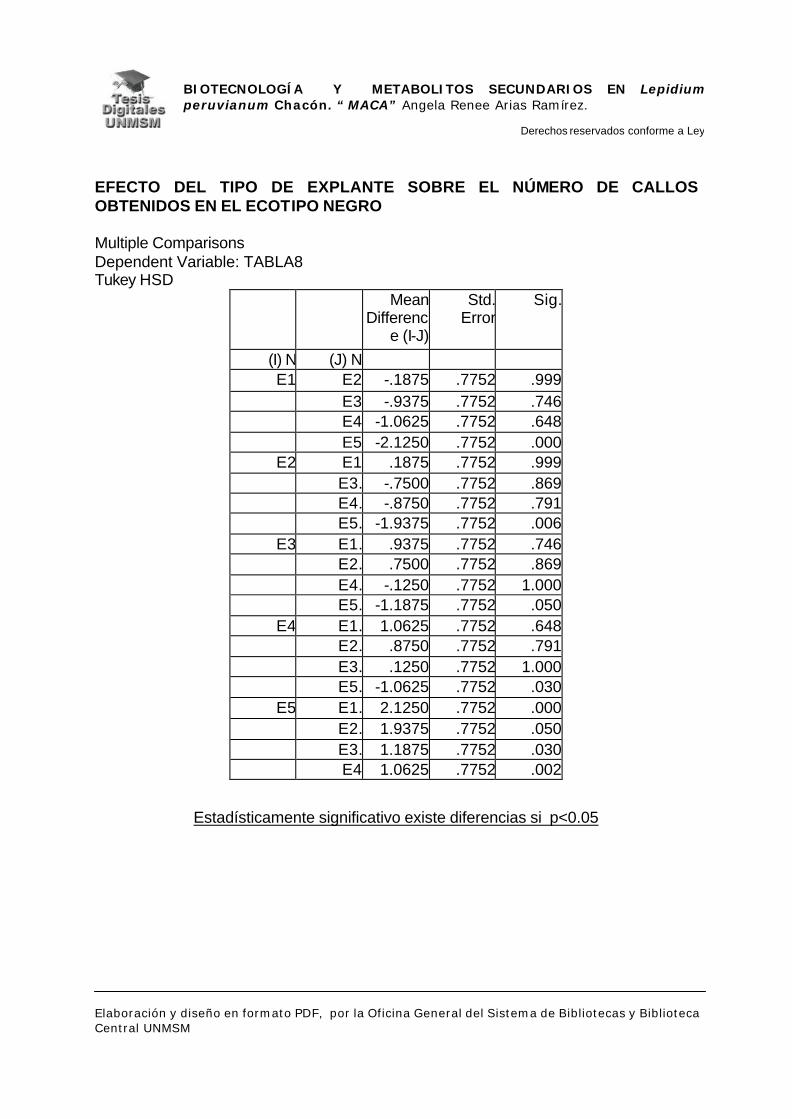
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL NÚMERO DE CALLOS OBTENIDOS EN EL ECOTIPO NEGRO Multiple Comparisons Dependent Variable: TABLA8 Tukey HSD
Mean Differenc
e (I-J)
Std. Error
Sig.
(I) N (J) NE1 E2 -.1875 .7752 .999
E3 -.9375 .7752 .746E4 -1.0625 .7752 .648E5 -2.1250 .7752 .000
E2 E1 .1875 .7752 .999E3. -.7500 .7752 .869E4. -.8750 .7752 .791E5. -1.9375 .7752 .006
E3 E1. .9375 .7752 .746E2. .7500 .7752 .869E4. -.1250 .7752 1.000E5. -1.1875 .7752 .050
E4 E1. 1.0625 .7752 .648E2. .8750 .7752 .791E3. .1250 .7752 1.000E5. -1.0625 .7752 .030
E5 E1. 2.1250 .7752 .000E2. 1.9375 .7752 .050E3. 1.1875 .7752 .030E4 1.0625 .7752 .002
Estadísticamente significativo existe diferencias si p<0.05
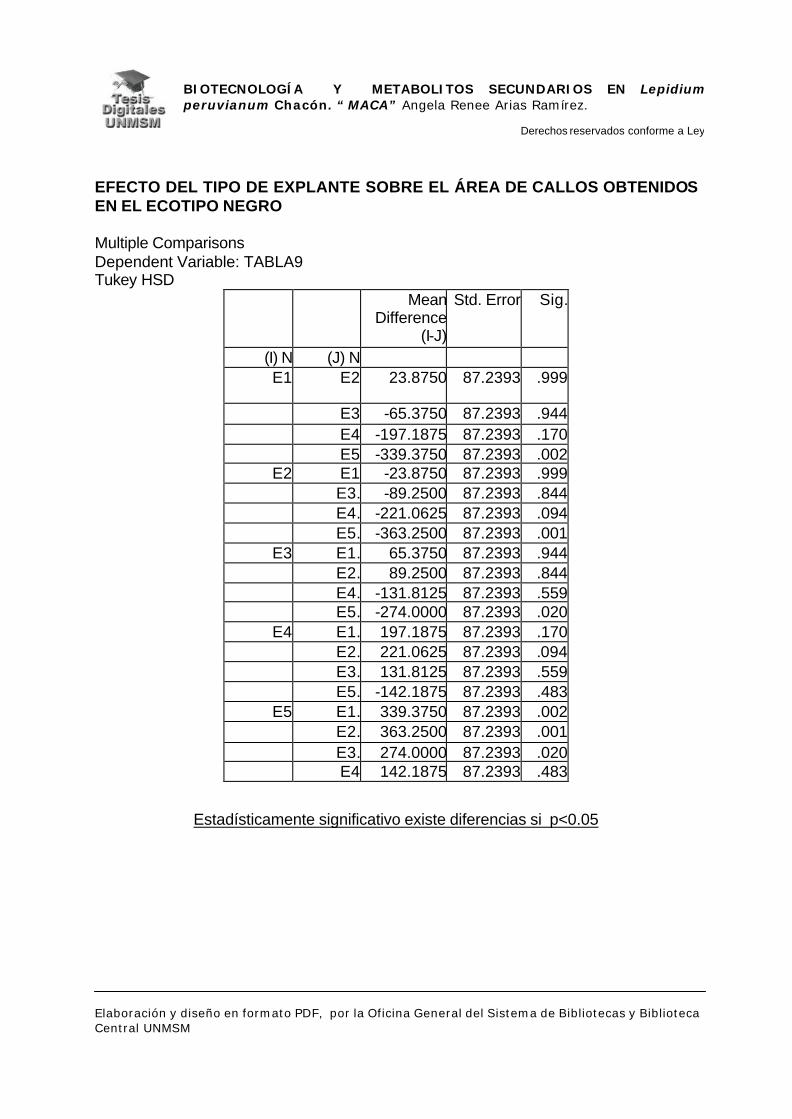
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE SOBRE EL ÁREA DE CALLOS OBTENIDOS EN EL ECOTIPO NEGRO Multiple Comparisons Dependent Variable: TABLA9 Tukey HSD
Mean Difference
(I-J)
Std. Error Sig.
(I) N (J) NE1 E2 23.8750 87.2393 .999
E3 -65.3750 87.2393 .944E4 -197.1875 87.2393 .170E5 -339.3750 87.2393 .002
E2 E1 -23.8750 87.2393 .999E3. -89.2500 87.2393 .844E4. -221.0625 87.2393 .094E5. -363.2500 87.2393 .001
E3 E1. 65.3750 87.2393 .944E2. 89.2500 87.2393 .844E4. -131.8125 87.2393 .559E5. -274.0000 87.2393 .020
E4 E1. 197.1875 87.2393 .170E2. 221.0625 87.2393 .094E3. 131.8125 87.2393 .559E5. -142.1875 87.2393 .483
E5 E1. 339.3750 87.2393 .002E2. 363.2500 87.2393 .001E3. 274.0000 87.2393 .020E4 142.1875 87.2393 .483
Estadísticamente significativo existe diferencias si p<0.05

BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
ANALISIS EDE VARIANZA PARA DOS EFECTOS FIJOS EFECTO DEL MEDIO DE CULTIVO EN LA PRODUCCIÓN DE CALLOS FRIALES Hipótesis τ i Para el factor 2,4-D
Ho :τ 1=:τ 2=:τ 3=:τ 4=0 H1:Al menos un τ i diferente de cero i =1, 2, 3, 4
β i Para el factor KIN
Ho β 1= = β 2 = β 3 = β 4=0 H1 Al menos una βi diferente a cero i =1, 2, 3, 4 Para el factor 2,4-D * KIN
Ho : (τ β )ij= 0 H1 Al menos una (τ β )ij diferente a cero
Nivel de significancia α =0.05 F crítico =9.36
Region crítica Fcal >F crítico el tratamiemto estadísticamente significativo exlste efecto *estadísticamente significativo exlste efecto
scua gl mcua f ss2,4-D 80 3 48.7430556 12*
ssKIN 14 3 34.57638889 11*
ssinter 87 9 45.8402778 15*
sserrores 119. 32 3.72916667 sst 299.8125 47
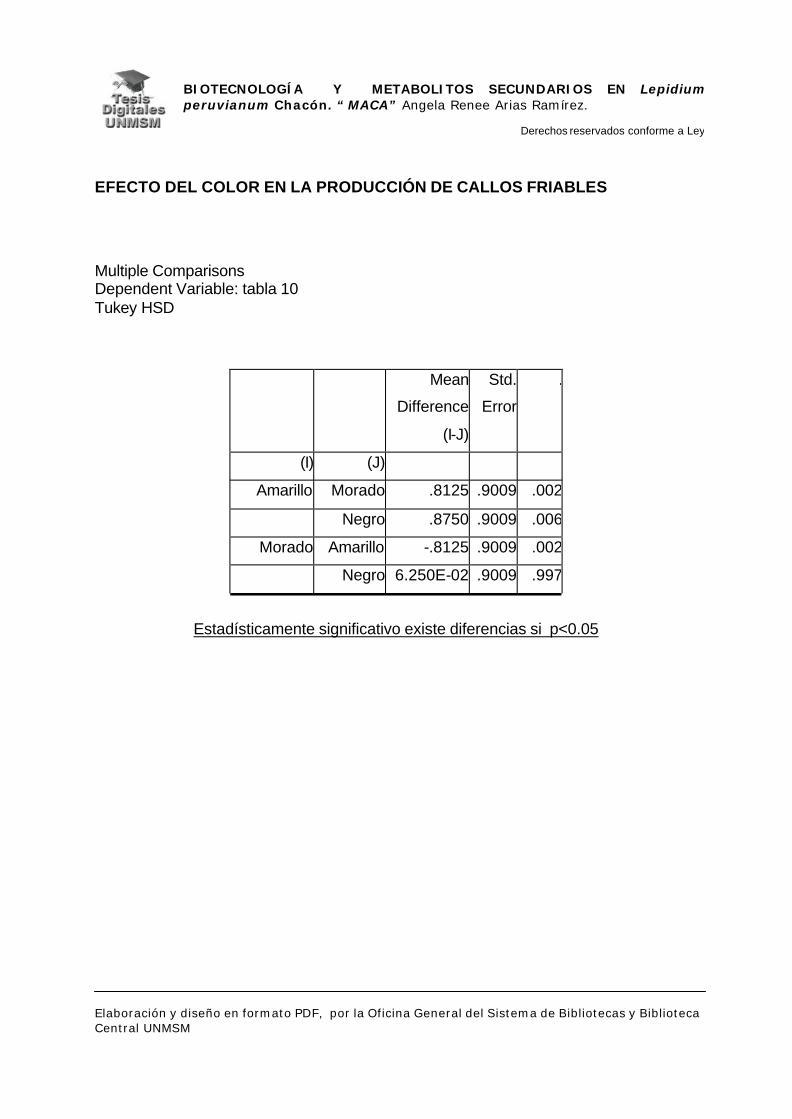
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL COLOR EN LA PRODUCCIÓN DE CALLOS FRIABLES
Multiple Comparisons Dependent Variable: tabla 10 Tukey HSD
Estadísticamente significativo existe diferencias si p<0.05
Mean
Difference
(I-J)
Std.
Error
.
(I) (J)
Amarillo Morado .8125 .9009 .002
Negro .8750 .9009 .006
Morado Amarillo -.8125 .9009 .002
Negro 6.250E-02 .9009 .997
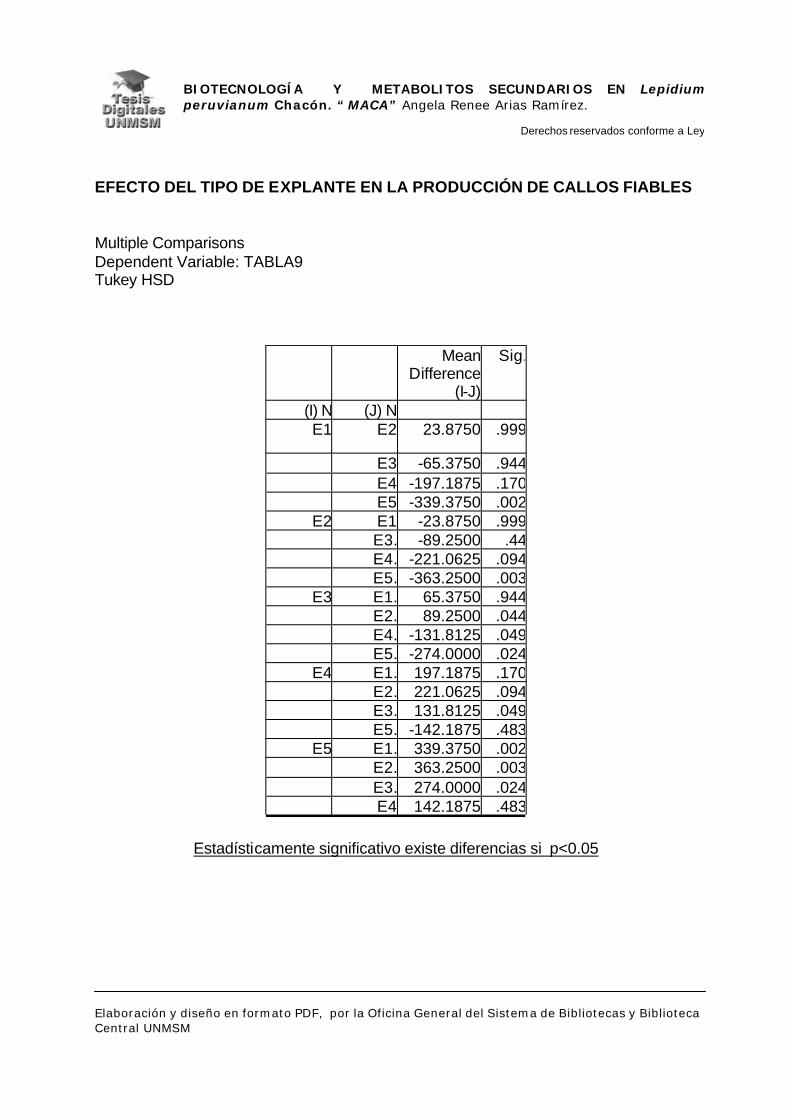
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
EFECTO DEL TIPO DE EXPLANTE EN LA PRODUCCIÓN DE CALLOS FIABLES Multiple Comparisons Dependent Variable: TABLA9 Tukey HSD
Estadísticamente significativo existe diferencias si p<0.05
Mean Difference
(I-J)
Sig.
(I) N (J) NE1 E2 23.8750 .999
E3 -65.3750 .944E4 -197.1875 .170E5 -339.3750 .002
E2 E1 -23.8750 .999E3. -89.2500 .44E4. -221.0625 .094E5. -363.2500 .003
E3 E1. 65.3750 .944E2. 89.2500 .044E4. -131.8125 .049E5. -274.0000 .024
E4 E1. 197.1875 .170E2. 221.0625 .094E3. 131.8125 .049E5. -142.1875 .483
E5 E1. 339.3750 .002E2. 363.2500 .003E3. 274.0000 .024E4 142.1875 .483
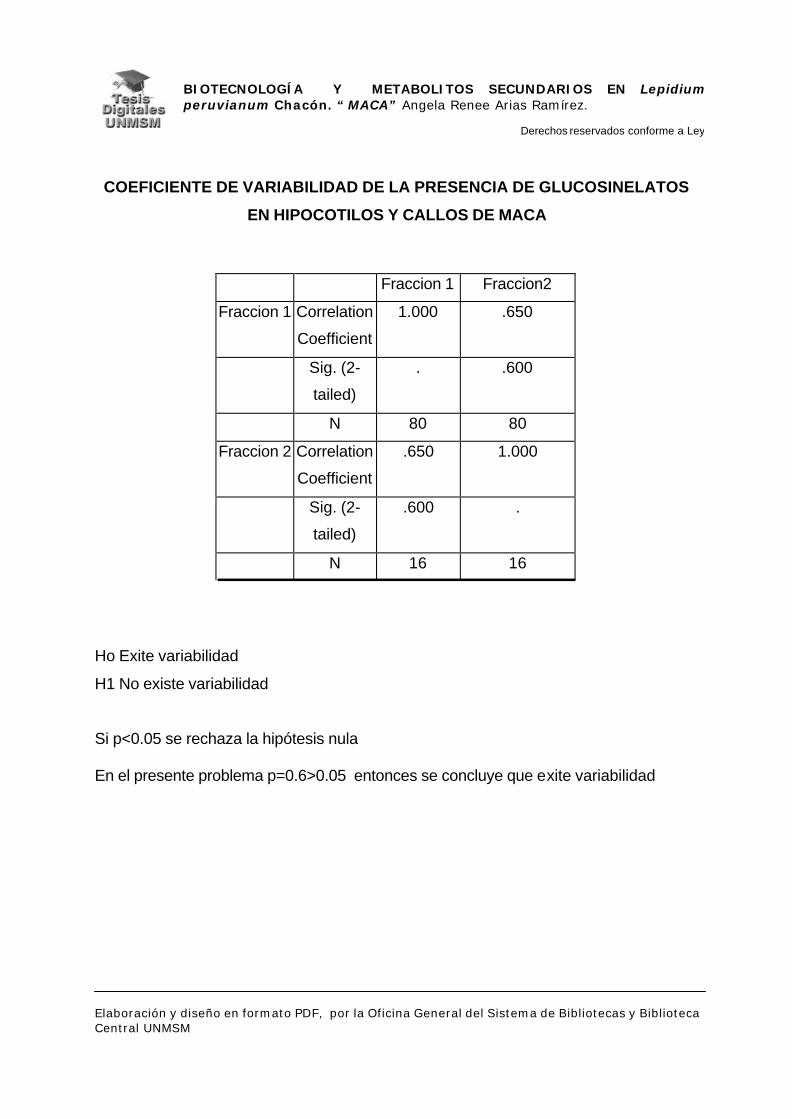
BIOTECNOLOGÍA Y METABOLITOS SECUNDARIOS EN Lepidium peruvianum Chacón. “ MACA” Angela Renee Arias Ramírez. Derechos reservados conforme a Ley
Elaboración y diseño en formato PDF, por la Oficina General del Sistema de Bibliotecas y Biblioteca Central UNMSM
COEFICIENTE DE VARIABILIDAD DE LA PRESENCIA DE GLUCOSINELATOS
EN HIPOCOTILOS Y CALLOS DE MACA
Ho Exite variabilidad
H1 No existe variabilidad
Si p<0.05 se rechaza la hipótesis nula En el presente problema p=0.6>0.05 entonces se concluye que exite variabilidad
Fraccion 1 Fraccion2
Fraccion 1 Correlation
Coefficient
1.000 .650
Sig. (2-
tailed)
. .600
N 80 80
Fraccion 2 Correlation
Coefficient
.650 1.000
Sig. (2-
tailed)
.600 .
N 16 16