Embed Size (px)
Citation preview


Vertebrados indicadores del estado de conservación del hayedo de Oianleku
Jorge González-Esteban Idoia Villate DESMA Estudios Ambientales SL
Enero 2007
La complejidad estructural en el hayedo de Oianleku 2
Las salamandras y la complejidad estructural 16
Los paseriformes nidificantes y la complejidad estructural 24
Los micromamíferos y la complejidad estructural 32
Conclusiones 36

La complejidad estructural en el hayedo de Oianleku
La fragmentación y la pérdida de hábitat pueden ocurrir en los bosques a varias
escalas espaciales. En primer lugar, a gran escala, en la escala "de paisaje", en
donde puede darse la desaparición directa del hábitat propiamente dicho. En
segundo término, aún manteniéndose intacta la cubierta forestal, áreas continuas
naturales de distintos tipos de bosque o de estados sucesionales pueden estar
siendo fragmentadas. Finalmente, en la escala más reducida, dentro de un
determinado tipo de bosque pueden perderse algunos de los elementos florísticos y
estructurales (Angelstam 1996). Por ello la conservación del hábitat precisa
estrategias de gestión a diferentes escalas, que van desde unos pocos metros
cuadrados hasta miles de hectáreas (desde árboles individuales hasta grandes
reservas). Lindenmayer y Franklin (2002) proponen para alcanzar este objetivo las
siguientes directrices: el mantenimiento de la conectividad a través del paisaje, el
mantenimiento de la heterogeneidad del paisaje, el mantenimiento de la complejidad
estructural y la diversidad de especies vegetales a escala de mancha o de parcela y
el mantenimiento de la integridad de los ecosistemas acuáticos, incluyendo los
procesos hidrológicos y geomorfológicos.
A pequeña escala (cientos o algunos miles de hectáreas), a una mayor
complejidad estructural de las comunidades vegetales le corresponde una mayor
diversidad animal (Perry 1994). Aunque actualmente existen pocas dudas acerca de
la veracidad de esta afirmación, pocos trabajos han estudiado la relación entre la
estructura del bosque y la diversidad animal. En el grupo de los vertebrados la
atención se ha centrado sobre todo en las aves, desarrollándose la mayor parte de
los estudios en bosques boreales y templados de Norteamérica.
El propósito del presente trabajo es describir y analizar dicha relación en el
hayedo de Oianleku, contando con algunas especies de vertebrados como reflejo de
la diversidad animal.
Material y métodos Los trabajos se han desarrollado en el hayedo de Oianleku (que cuenta con una
extensión aproximada de 200 ha).
2

La complejidad estructural es un rasgo común a todos los bosques templados
del mundo y altos niveles de heterogeneidad espacial son característicos de todos
los bosques maduros-viejos (Lindenmayer y Franklin 2002). La complejidad
estructural a escala de mancha o de rodal incluye una serie rasgos y elementos
estructurales, tales como: árboles de diferentes edades y tamaños, árboles vivos en
distinto estado de decaimiento, árboles muertos aún en pie (que en la literatura
científica se conocen como snags), árboles caídos (que en la literatura científica se
conocen como logs), claros.
En el hayedo pueden identificarse fácilmente estos elementos. Su
cuantificación se ha realizado en una serie de parcelas. Para garantizar la
aleatoriedad del muestreo se seleccionaron las parcelas utilizando como referencia
el retículo UTM (cuadrícula de 250 m de lado). El centro de las parcelas de muestreo
se hizo coincidir con el centro de las cuadrículas. La figura 1 recoge la disposición de
las parcelas seleccionadas. La caracterización de la complejidad estructural de cada
parcela se realizó conforme al protocolo que se detalla a continuación.
En primer lugar sobre un transecto lineal de 100 m de longitud, cuyo punto
medio coincide con el centro de la parcela, y siguiendo el método PCQ (Point-
Centered Quarter; Cottam y Curtis 1956) se han muestreado los árboles vivos. Se
registró la especie a la que pertenecen, su diámetro a una altura de 1.4 m (dbh) y su
estado de decaimiento. Se han considerado como árboles todos aquellos cuyo dbh
es superior a 10 cm. Para valorar el estado de decaimiento de los árboles vivos se
ha seguido el criterio elaborado por Carey y Haley (1981):
[1]: la corona presenta una forma regular, sin que se observen ramas muertas de
más de 10 cm de diámetro.
[2]: el árbol presenta 1 o 2 ramas grandes muertas (de más de 10 cm de diámetro).
[3]: el árbol presenta 3 o más ramas grandes muertas (de más de 10 cm de
diámetro); una parte importante de la corona (> 1/3) está muerta o ha desaparecido.
[4]: el árbol presenta una porción del tronco muerta.
[5]: todo el árbol está muerto, pero permanece aún en pie. Es lo que se conoce
como un snag.
En segundo lugar, se anotó el número de snags presentes en dos parcelas
circulares de 25 m de radio centradas en los extremos del transecto. De los snags se
registró la especie a la que pertenecen, su dbh, su altura y su estado de pudrición.
Se han considerado únicamente aquellos que presentaban un dbh superior a 10 cm. 3

Para valorar el estado de pudrición de los snags se ha seguido el criterio elaborado
por Goodburn y Lorimer (1998). El grado de descomposición de la madera se ha
valorado con la ayuda de una varilla metálica de 5 mm de grosor y punta
redondeada (no afilada). Las clases de pudrición consideradas son las siguientes:
[1]: el árbol está muerto, pero la corteza permanece intacta; no se aprecian aún
signos de pudrición
[2]: la corteza comienza a desprenderse y se aprecian algunos signos de pudrición;
la varilla no penetra en el tronco más allá de 1-2 cm.
[3]: la pudrición se aprecia claramente; la varilla se introduce con facilidad, sin
alcanzar aún la parte interna del tronco
[4]: la pudrición se extiende por todo el tronco; la varilla lo atraviesa fácilmente; ha
comenzado el desprendimiento de madera podrida
[5]: el árbol muestra poca integridad estructural; ha perdido gran parte de su
volumen.
El volumen de los snags se ha calculado asumiendo que todos ellos
presentan forma cilíndrica.
Posteriormente se ha calculado el volumen de los troncos muertos que
descansan sobre el suelo (logs). Se han considerado como logs todos los troncos de
diámetro superior a 10 cm y longitud superior a 1 m, que se encuentran sobre el
suelo o suspendidos por uno de sus extremos sin que el ángulo que forman con el
suelo llegue a superar los 45o. Los logs se han muestreado sobre un transecto lineal
de 100 m, cuyo punto medio coincide con el centro de la parcela. Dado que en
función de las características del área a estudiar la distribución de los logs puede
seguir una orientación determinada, la orientación del transecto fue determinada al
azar. En los puntos en el que el transecto lineal tocaba a los logs se anotó la
especie, su diámetro y el estado de pudrición. Para valorar el estado de pudrición de
los logs se ha seguido el criterio elaborado por Pyle y Brown (1998):
[1]: la corteza permanece intacta; no se aprecian aún signos de pudrición
[2]: sin corteza o con poca corteza; la superficie está dura aunque puede haber
comenzado el proceso de pudrición interna
[3]: sin corteza; la superficie está húmeda y se desprende al golpearla; podemos
introducir el dedo fácilmente, aunque aún presenta cierto grado de firmeza
4

[4]: el log se aplasta o rompe fácilmente y presenta sección ovalada o aplastada; al
apretar con el dedo exhuda humedad; comienza a aparecer en su entorno serrín o
pequeños fragmentos de madera podrida
[5]: el log está en su mayor parte hecho serrín.
El volumen fue estimado usando la fórmula descrita por Lofroth (1992):
V = (Л2Σd2) / 8L
donde V es volumen en m3/ha, d es diámetro (cm) de cada log y L es la longitud del
transecto (en este caso 100 m).
La tabla 1 recoge la descripción de los parámetros utilizados.
La información acumulada en los últimos 20 años acerca de los requerimientos
ecológicos de la fauna forestal (principalmente en Norteamérica y norte de Europa)
ha consolidado la idea de que la biodiversidad está fuertemente ligada a la
heterogeneidad espacial y a la complejidad estructural, y que por ello es necesario
realizar una gestión activa encaminada al mantenimiento de ambas en los
ecosistemas forestales. En esta línea han proliferado en la última década trabajos
que pretenden describir de forma estandarizada para cada tipo de bosque las
características del bosque old-growth (Batista y Platt, 1997; Greenberg et al. 1997;
Kennedy y Nowacki 1997), con objeto de conocer su disponibilidad y frenar su
progresiva desaparición.
Existe cierta controversia a la hora de definir lo que se entiende por bosque
old-growth, debido principalmente a la necesidad de establecer unos criterios que
permitan comparar bosques de distintas regiones. La definición más simple se
refiere a ellos como bosques relativamente viejos y relativamente inalterados. Dicho
de otra forma, la alta complejidad estructural y las características funcionales
asociadas con el bosque old-growth se han desarrollado en largos períodos de
tiempo con las mínimas perturbaciones (tanto de origen natural como humano)
(Tyrrell et al. 1998).
En este caso se ha contado como referencia de la complejidad estructural
propia de un bosque natural, viejo y no explotado con los resultados de un trabajo
similar realizado en el hayedo de Bértiz (Navarra) (Villate y González-Esteban,
2002). En este hayedo (de aproximadamente 1800 ha) la explotación maderera cesó
en 1900 y durante todo el siglo XX no ha sufrido intervenciones de importancia
(Esparza 2000). Si comparamos los valores de complejidad estructural obtenidos en 5

Bértiz con los que ofrece la literatura científica para distintos tipos de bosque old-
growth (tablas 2, 3, y 4) observamos que, si bien existe una notable variación, los
bosques old-growth muestran al igual que Bértiz gran cantidad de madera muerta y
una alta densidad de árboles vivos de gran diámetro y de árboles vivos con altos
niveles de decaimiento.
Resultados
Utilizando los valores de Bértiz como referencia se puede describir Oianleku como
un hayedo de baja complejidad estructural en el que destacan tres rasgos: la baja
densidad de árboles, el gran tamaño de éstos y el número de árboles en avanzado
estado de decaimiento (tabla 5).
La carencia más notable, en cuanto a complejidad se refiere, es la de árboles
muertos, tanto logs como snags, no destacando ninguna de las clases de pudrición y
tamaño. El volumen total de madera muerta (aprox 12 m3/ha) está dentro del rango
de variación que ofrecen distintos inventarios forestales en Europa (Vallauri et al.
2002).
La madera muerta se incorpora al bosque debido a perturbaciones naturales
(vendavales, fuego, insectos, sequía), como resultado de la competencia entre
árboles por el espacio, o como resultado final del proceso de envejecimiento propio
del ciclo vital de la especie. En las masas sujetas a aprovechamiento maderero, el
conjunto de tratamientos que se realizan sobre cada rodal va configurando su
complejidad estructural. En este caso, la práctica del trasmocheo ha reducido
considerablemente la acción de los mecanismos de incorporación de árboles
muertos modelando una masa excesivamente monótona.
Cabe destacar no obstante que la complejidad que actualmente ofrece
Oianleku (decaimiento, pequeña cantidad de madera muerta) es producto del
abandono del trasmocheo. Muy probablemente, a principios del siglo XX, cuando
este hayedo se encontraba en explotación, su complejidad era aún menor.
Referencias Angelstam, P., 1996. The ghost of forest past. Natural disturbance regimes as a basis dor reconstruction for biologically diverse forests in Europe. In: Conservation of faunal diversity in forested landscapes. R.M. DeGraaf, R.I. Miller (Eds.). London, Chapman & Hall, 287-337.
6

Batista, W.B., Platt, W.J., 1997. An old-growth definition for Southern mixed hardwood forests. Gen. Tech. Rep. SRS-9. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 11 p. Carey, A.B., Healy, W.M., 1981. Cavities in trees around spring seeps in the maple-beech-birch forest type. Northeast. For. Exp. Stn., Broomall, Pa. Research paper, NE-480. 7 p. Cottam, G., Curtis, J.T., 1956. The use of distance measures in phytosociological sampling. Ecology, 37:451-460. Esparza, A., 2000. El Señorío de Bértiz. Cuadernos del Centro de Estudios Bidasoa, 3. Goodburn, J.M., Lorimer, C.G., 1998. Cavity trees and coarse woody debris in old-growth and managed Northern Harwood Forests in Wisconsin and Michigan. Can. J. For. Res., 28: 427-438. Greenberg, C.H., McLeod, D.E., Loftis, D.L., 1997. An old-growth definition for western and mixed mesophytic forests. Gen. Tech. Rep. SRS-16. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 14 p. Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V., Latton, J.D., Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R., Lienkaemper, G.W., Cromack, K. Jr., Cummins, K.W., 1986. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research, 15:133-302. Kennedy, H. E., Nowacki, G.J., 1997. An old-growth definition for seasonally wet oak-hardwood woodlands. Gen. Tech. Rep. SRS-8. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 6 p. Lindenmayer, D.B., Franklin, J.F., 2002. Conserving Forest Biodiversity. A comprehensive multiscaled approach. Washington, Island Press. Lofroth, E., 1992. Measuring habitat elements at the stand level. En: Proc. Methodologies for monitoring wildlife diversity in BC forests. Wildlife Branch. Ministry of Environment, Victoria. Meadows, J. S.; Nowacki, G. J. 1996. An old-growth definition for eastern riverfront forests. Gen. Tech. Rep. SRS-4. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 7 p. Outcalt, K. W. 1997. An old-growth definition for sand pine forests. Gen. Tech. Rep. SRS-12., Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 8 p. Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins University Press. Pyle, C., Brown, M.M. 1998. A rapid system of decay classification for hardwood logs of the eastern deciduous forest floor. Journal of the Torrey Botanical Society, 125(3): 237-245. Siitonen, J., 2001. Forest management, coarse woody debris and saproxylic organism: Fennoscandian boreal forests as an example. En: Ecology of woody debris in boreal forests. Ecological Bulletins, 49:11-41. Spetich, M.A., Shifley, S.R., Parker, G,R., 1999. Regional distribution and dynamics of coarse woody debris in midwestern old-growth forests. Forest Science, 45:303-313. Tyrrell, L. E.; Nowacki, G. J.; Crow, T. R.; Buckley, D. S.;Nauertz, E. A.; Niese, J. N.; Rollinger, J. L.; Zasada, J. C.1998. Information about old growth for selected forest type groups in the eastern United States. Gen. Tech. Rep. NC-197. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 507 p.
7

Vallauri, D., André, J., Blondel, J., 2002. Le bois mort, un attribut vital de la biodiversité de la forêt naturelle, une lacune des forêts gérées.Rapport Scientifique. WWW-France, 31 pp. Villate, I., González-Esteban, J., 2002. Incidencia en la comunidad de vertebrados de los elementos de diversidad forestal relacionados con la gestión del bosque. Gobierno de Navarra. White, D.L., Lloyd, F.T., 1998. An old-growth definition for dry and dry-mesic oak-pine forests. Gen. Tech. Rep. SRS-23. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station. 42 p.
8

Tabla 1. Descripción de las variables utilizadas en la caracterización de la
complejidad estructural del bosque.
Variable
(abreviatura)
Descripción
DA Diámetro medio de los árboles vivos medido a 1.4 m de altura (cm)
NA Número de árboles vivos por ha
NA30 Número de árboles vivos (por ha) con dbh > 30 cm
NA60 Número de árboles vivos (por ha) con dbh > 60 cm
NA12 Número de árboles vivos (por ha) con un estado de decaimiento de clase 1 o 2
NA34 Número de árboles vivos (por ha) con un estado de decaimiento de clase 3 o 4
NS Número de snags por ha
NS30 Número de snags (por ha) con dbh > 30 cm
NS60 Número de snags (por ha) con dbh > 60 cm
NS12 Número de snags (por ha) con un estado de pudrición de clase 1 o 2
NS34 Número de snags (por ha) con un estado de pudrición de clase 3 o 4
VS Volumen total de los snags (m3 / ha)
VS12 Volumen de los snags con un estado de pudrición de clase 1 o 2 (m3 / ha)
VS34 Volumen de los snags con un estado de pudrición de clase 3 o 4 (m3 / ha)
VL Volumen total de los logs (m3 / ha)
VL30 Volumen de los logs con un diámetro mayor de 30 cm (m3 / ha)
VL12 Volumen de los logs con un estado de pudrición de clase 1 o 2 (m3 / ha)
VL34 Volumen de los logs con un estado de pudrición de clase 3 o 4 (m3 / ha)
9
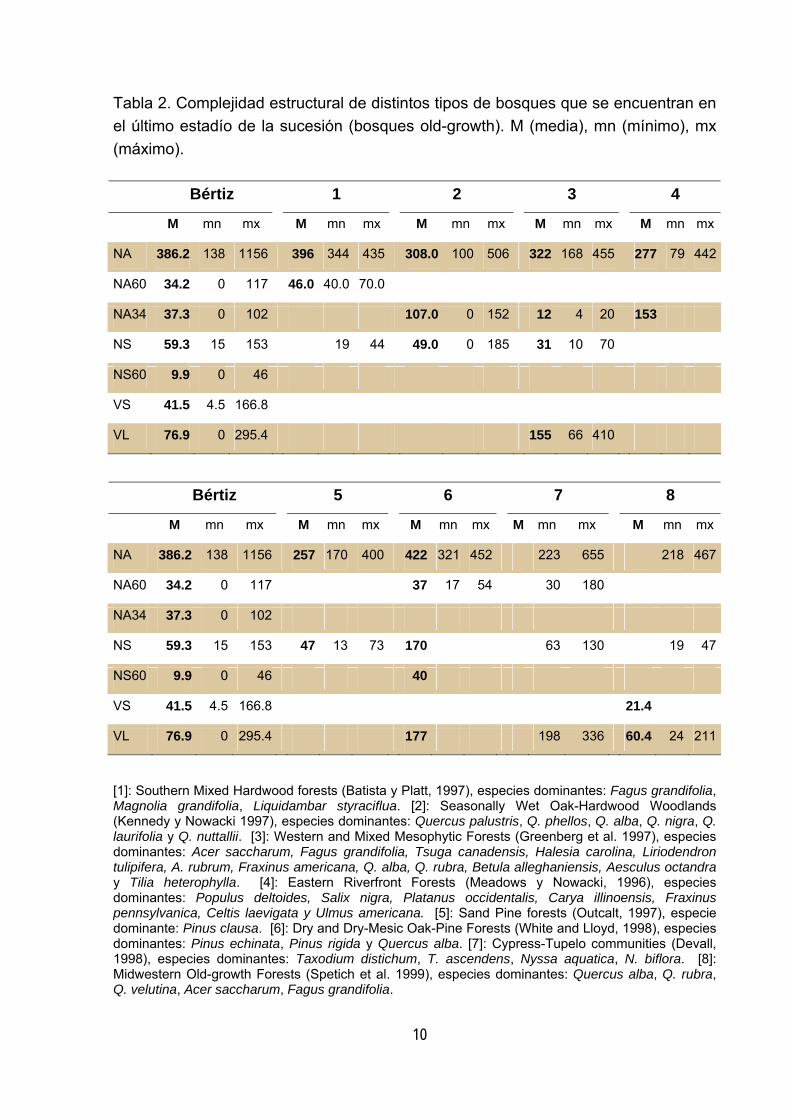
Tabla 2. Complejidad estructural de distintos tipos de bosques que se encuentran en el último estadío de la sucesión (bosques old-growth). M (media), mn (mínimo), mx (máximo).
Bértiz 1 2 3 4
M mn mx M mn mx M mn mx M mn mx M mn mx
NA 386.2 138 1156 396 344 435 308.0 100 506 322 168 455 277 79 442
NA60 34.2 0 117 46.0 40.0 70.0
NA34 37.3 0 102 107.0 0 152 12 4 20 153
NS 59.3 15 153 19 44 49.0 0 185 31 10 70
NS60 9.9 0 46
VS 41.5 4.5 166.8
VL 76.9 0 295.4 155 66 410
Bértiz 5 6 7 8
M mn mx M mn mx M mn mx M mn mx M mn mx
NA 386.2 138 1156 257 170 400 422 321 452 223 655 218 467
NA60 34.2 0 117 37 17 54 30 180
NA34 37.3 0 102
NS 59.3 15 153 47 13 73 170 63 130 19 47
NS60 9.9 0 46 40
VS 41.5 4.5 166.8 21.4
VL 76.9 0 295.4 177 198 336 60.4 24 211
[1]: Southern Mixed Hardwood forests (Batista y Platt, 1997), especies dominantes: Fagus grandifolia, Magnolia grandifolia, Liquidambar styraciflua. [2]: Seasonally Wet Oak-Hardwood Woodlands (Kennedy y Nowacki 1997), especies dominantes: Quercus palustris, Q. phellos, Q. alba, Q. nigra, Q. laurifolia y Q. nuttallii. [3]: Western and Mixed Mesophytic Forests (Greenberg et al. 1997), especies dominantes: Acer saccharum, Fagus grandifolia, Tsuga canadensis, Halesia carolina, Liriodendron tulipifera, A. rubrum, Fraxinus americana, Q. alba, Q. rubra, Betula alleghaniensis, Aesculus octandra y Tilia heterophylla. [4]: Eastern Riverfront Forests (Meadows y Nowacki, 1996), especies dominantes: Populus deltoides, Salix nigra, Platanus occidentalis, Carya illinoensis, Fraxinus pennsylvanica, Celtis laevigata y Ulmus americana. [5]: Sand Pine forests (Outcalt, 1997), especie dominante: Pinus clausa. [6]: Dry and Dry-Mesic Oak-Pine Forests (White and Lloyd, 1998), especies dominantes: Pinus echinata, Pinus rigida y Quercus alba. [7]: Cypress-Tupelo communities (Devall, 1998), especies dominantes: Taxodium distichum, T. ascendens, Nyssa aquatica, N. biflora. [8]: Midwestern Old-growth Forests (Spetich et al. 1999), especies dominantes: Quercus alba, Q. rubra, Q. velutina, Acer saccharum, Fagus grandifolia.
10
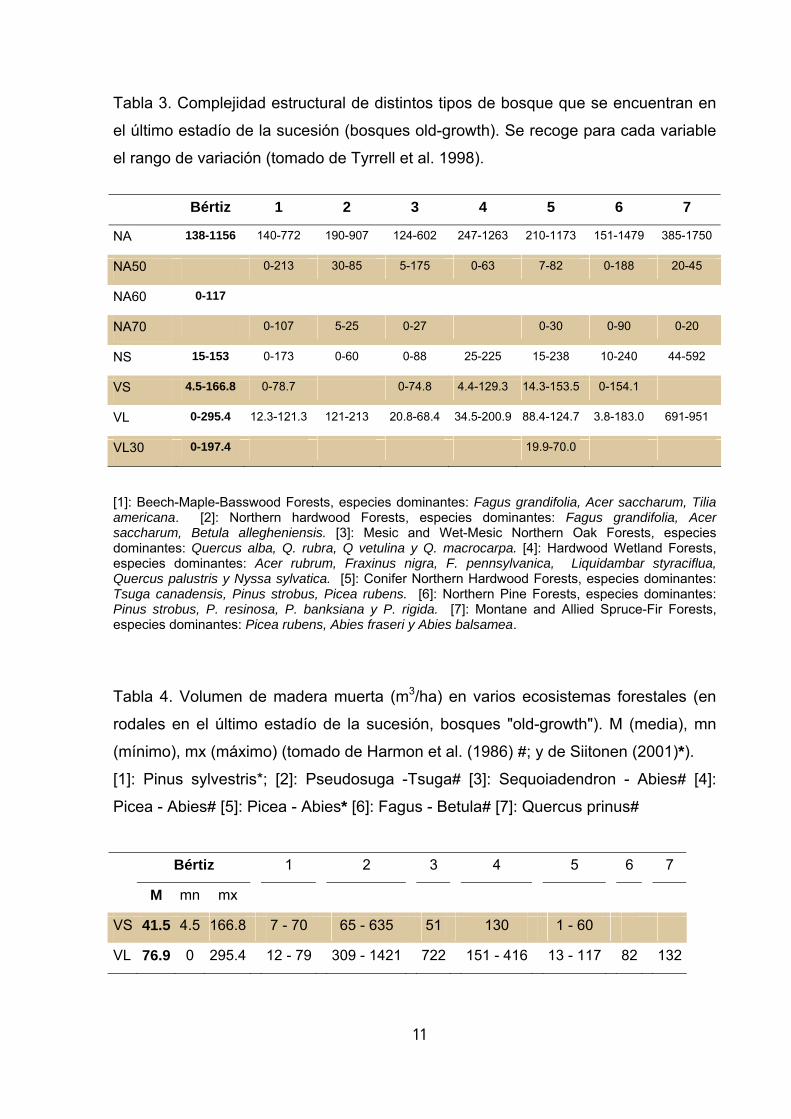
Tabla 3. Complejidad estructural de distintos tipos de bosque que se encuentran en
el último estadío de la sucesión (bosques old-growth). Se recoge para cada variable
el rango de variación (tomado de Tyrrell et al. 1998).
Bértiz 1 2 3 4 5 6 7
NA 138-1156 140-772 190-907 124-602 247-1263 210-1173 151-1479 385-1750
NA50 0-213 30-85 5-175 0-63 7-82 0-188 20-45
NA60 0-117
NA70 0-107 5-25 0-27 0-30 0-90 0-20
NS 15-153 0-173 0-60 0-88 25-225 15-238 10-240 44-592
VS 4.5-166.8 0-78.7 0-74.8 4.4-129.3 14.3-153.5 0-154.1
VL 0-295.4 12.3-121.3 121-213 20.8-68.4 34.5-200.9 88.4-124.7 3.8-183.0 691-951
VL30 0-197.4 19.9-70.0
[1]: Beech-Maple-Basswood Forests, especies dominantes: Fagus grandifolia, Acer saccharum, Tilia americana. [2]: Northern hardwood Forests, especies dominantes: Fagus grandifolia, Acer saccharum, Betula allegheniensis. [3]: Mesic and Wet-Mesic Northern Oak Forests, especies dominantes: Quercus alba, Q. rubra, Q vetulina y Q. macrocarpa. [4]: Hardwood Wetland Forests, especies dominantes: Acer rubrum, Fraxinus nigra, F. pennsylvanica, Liquidambar styraciflua, Quercus palustris y Nyssa sylvatica. [5]: Conifer Northern Hardwood Forests, especies dominantes: Tsuga canadensis, Pinus strobus, Picea rubens. [6]: Northern Pine Forests, especies dominantes: Pinus strobus, P. resinosa, P. banksiana y P. rigida. [7]: Montane and Allied Spruce-Fir Forests, especies dominantes: Picea rubens, Abies fraseri y Abies balsamea.
Tabla 4. Volumen de madera muerta (m3/ha) en varios ecosistemas forestales (en
rodales en el último estadío de la sucesión, bosques "old-growth"). M (media), mn
(mínimo), mx (máximo) (tomado de Harmon et al. (1986) #; y de Siitonen (2001)*). [1]: Pinus sylvestris*; [2]: Pseudosuga -Tsuga# [3]: Sequoiadendron - Abies# [4]:
Picea - Abies# [5]: Picea - Abies* [6]: Fagus - Betula# [7]: Quercus prinus#
Bértiz 1 2 3 4 5 6 7
M mn mx
VS 41.5 4.5 166.8 7 - 70 65 - 635 51 130 1 - 60
VL 76.9 0 295.4 12 - 79 309 - 1421 722 151 - 416 13 - 117 82 132
11
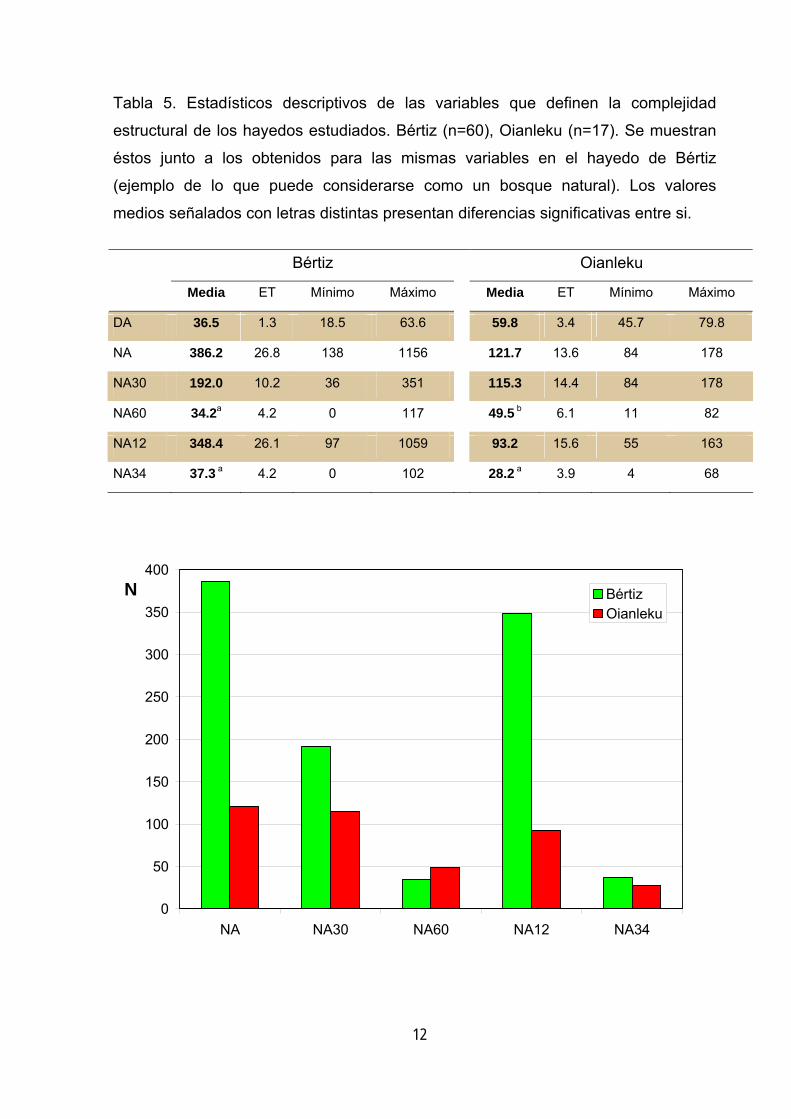
Tabla 5. Estadísticos descriptivos de las variables que definen la complejidad
estructural de los hayedos estudiados. Bértiz (n=60), Oianleku (n=17). Se muestran
éstos junto a los obtenidos para las mismas variables en el hayedo de Bértiz
(ejemplo de lo que puede considerarse como un bosque natural). Los valores
medios señalados con letras distintas presentan diferencias significativas entre si.
Bértiz Oianleku
Media ET Mínimo Máximo Media ET Mínimo Máximo
DA 36.5 1.3 18.5 63.6 59.8 3.4 45.7 79.8
NA 386.2 26.8 138 1156 121.7 13.6 84 178
NA30 192.0 10.2 36 351 115.3 14.4 84 178
NA60 34.2a 4.2 0 117 49.5 b 6.1 11 82
NA12 348.4 26.1 97 1059 93.2 15.6 55 163
NA34 37.3 a 4.2 0 102 28.2 a 3.9 4 68
0
50
100
150
200
250
300
350
400
NA NA30 NA60 NA12 NA34
N BértizOianleku
12
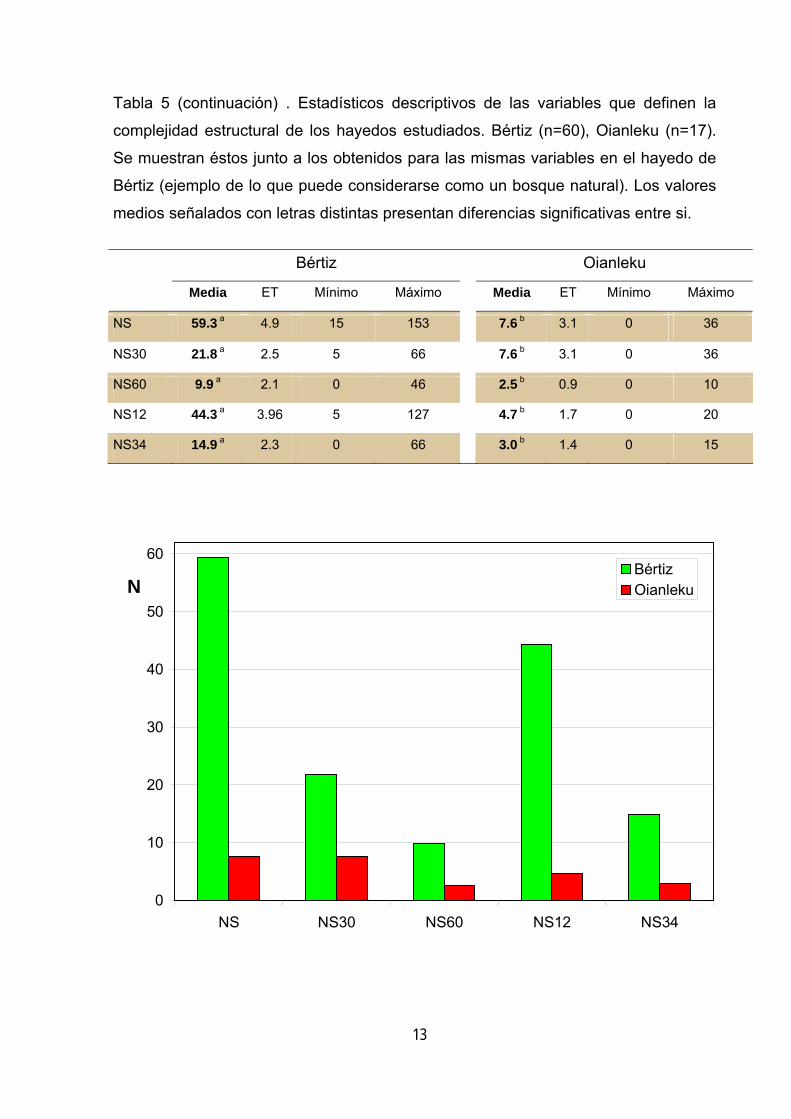
Tabla 5 (continuación) . Estadísticos descriptivos de las variables que definen la
complejidad estructural de los hayedos estudiados. Bértiz (n=60), Oianleku (n=17).
Se muestran éstos junto a los obtenidos para las mismas variables en el hayedo de
Bértiz (ejemplo de lo que puede considerarse como un bosque natural). Los valores
medios señalados con letras distintas presentan diferencias significativas entre si.
Bértiz Oianleku
Media ET Mínimo Máximo Media ET Mínimo Máximo
NS 59.3 a 4.9 15 153 7.6 b 3.1 0 36
NS30 21.8 a 2.5 5 66 7.6 b 3.1 0 36
NS60 9.9 a 2.1 0 46 2.5 b 0.9 0 10
NS12 44.3 a 3.96 5 127 4.7 b 1.7 0 20
NS34 14.9 a 2.3 0 66 3.0 b 1.4 0 15
0
10
20
30
40
50
60
NS NS30 NS60 NS12 NS34
NBértizOianleku
13
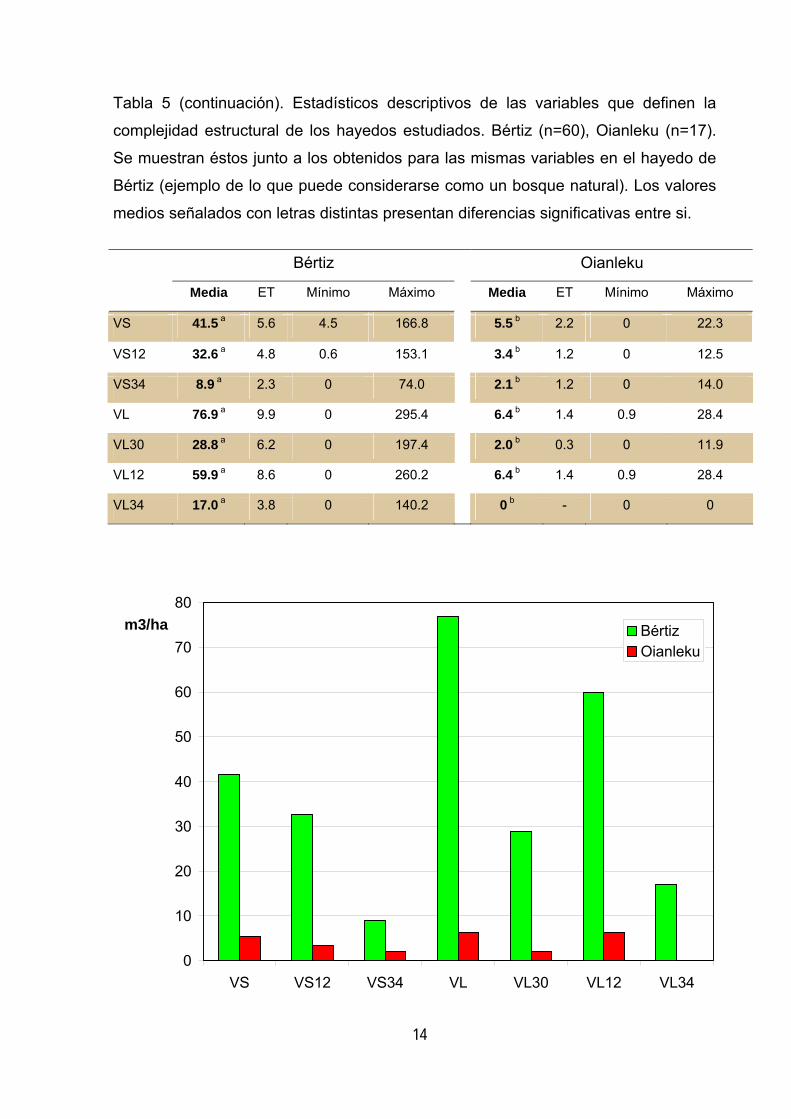
Tabla 5 (continuación). Estadísticos descriptivos de las variables que definen la
complejidad estructural de los hayedos estudiados. Bértiz (n=60), Oianleku (n=17).
Se muestran éstos junto a los obtenidos para las mismas variables en el hayedo de
Bértiz (ejemplo de lo que puede considerarse como un bosque natural). Los valores
medios señalados con letras distintas presentan diferencias significativas entre si.
Bértiz Oianleku
Media ET Mínimo Máximo Media ET Mínimo Máximo
VS 41.5 a 5.6 4.5 166.8 5.5 b 2.2 0 22.3
VS12 32.6 a 4.8 0.6 153.1 3.4 b 1.2 0 12.5
VS34 8.9 a 2.3 0 74.0 2.1 b 1.2 0 14.0
VL 76.9 a 9.9 0 295.4 6.4 b 1.4 0.9 28.4
VL30 28.8 a 6.2 0 197.4 2.0 b 0.3 0 11.9
VL12 59.9 a 8.6 0 260.2 6.4 b 1.4 0.9 28.4
VL34 17.0 a 3.8 0 140.2 0 b - 0 0
0
10
20
30
40
50
60
70
80
VS VS12 VS34 VL VL30 VL12 VL34
m3/ha BértizOianleku
14

Figura 1. Distribución de las parcelas seleccionadas para estudiar la complejidad
estructural en el hayedo de Oianleku.
15

Las salamandras y la complejidad estructural
Los anfibios desempeñan un importante papel en las redes tróficas de los bosques
templados, canalizando una parte significativa del flujo de energía que discurre a
través de la comunidad de vertebrados (Perry 1994). Además explotan recursos
alimenticios que por su tamaño o por su modo de vida resultan inaccesibles a aves y
mamíferos (Burton y Likens 1975).
En los hayedos acidófilos estudiados en el presente trabajo, a pesar de que la
humedad se mantiene alta a lo largo de todo el año, solo dos especies de anfibios, el
sapo común Bufo bufo y la salamandra Salamandra salamandra, pueden
considerarse como propias de su fauna. Y es únicamente esta última la que cuenta
con poblaciones numerosas y estables en este tipo de bosque.
Se ha detectado en las últimas décadas un declive global de las poblaciones
de anfibios (Alford y Richards 1999), asociado en la mayor parte de las ocasiones a
la alteración directa del hábitat que produce el aprovechamiento de los recursos
naturales (Griffiths 1995, Lizana y Barbadillo 1997). La información disponible acerca
del efecto de la silvicultura y de la gestión forestal sobre los anfibios pone de
manifiesto que los estadíos de bosque maduro y bosque viejo son capaces de
albergar comunidades de vertebrados más ricas y diversas, en las que los anfibios
están bien representados (Welsh y Lind 1988, Butts y McComb 2000). Se observa
también en estos trabajos que la reducción de la complejidad estructural producto
del aprovechamiento maderero incide negativamente sobre las poblaciones de
anfibios y concretamente sobre las de distintas especies de salamandras
(deMaynadier y Hunter 1995, Herbeck y Larsen 1999, Andrew y Pollock 1999,
Harpole y Haas 1999).
En el presente trabajo se estudia la relación entre distintos parámetros de las
poblaciones de salamandras y la complejidad estructural del hayedo.
Material y métodos. El estudio se desarrolló en los hayedos de Bértiz y Oianleku. En ellos se
seleccionaron 31 parcelas circulares (19 en Bértiz y 12 en Oianleku) de 25 m de
radio en las que se caracterizó la complejidad estructural y se estudiaron distintos
16

parámetros de la población de salamandras. La selección de las parcelas se realizó
priorizando el obtener un conjunto de muestras en el que estuviera representada la
heterogeneidad estructural propia del hayedo. Todas ellas distantes más de un
kilómetro de un curso de agua permanente y más de 250 m de un curso de agua
temporal.
En cada parcela se realizó un muestreo nocturno entre los meses de junio y
septiembre. En dichos muestreos dos personas batieron las parcelas capturando los
animales que podían observarse a simple vista, bien en terreno abierto o refugiados
en determinados elementos estructurales (árboles vivos, snags, logs). Dado que la
actividad de las salamandras depende de la temperatura y de la humedad ambiental
(Salvador y García-Paris 2001) los muestreos se realizaron en noches lluviosas y
con una temperatura superior a los 15oC. Para evitar recapturas en un mismo
muestreo los animales fueron retirados de la parcela tras la toma de datos y
liberados en ella al finalizar el muestreo.
La caracterización de la complejidad estructural de cada parcela se realizó
conforme a los protocolos de trabajo descritos en el capítulo anterior.
Resultados La densidad de salamandras es significativamente mayor en el hayedo que cuenta
con alta complejidad estructural (Bértiz, 589.5 individuos/ha) que en el que cuenta
con baja complejidad (Oianleku, 77.1 individuos/ha) (U Mann-Whitney = 6.5,
p<0.001).
Analizando el conjunto de las muestras se observa que existe una marcada
correlación positiva entre la abundancia de salamandras y la mayor parte de las
variables que definen la abundancia de madera muerta, así como con la abundancia
de grandes árboles y de árboles con valores de decaimiento altos (tabla 6). Idéntico
comportamiento se observa al contrastar el peso total de las salamandras
capturadas con las variables de caracterización (tabla 6). Ambos parámetros,
número de salamandras y peso total de salamandras presentan una elevada
correlación positiva entre sí (r Pearson = 0.87, p < 0.001, n = 31). Únicamente se
observa correlación negativa con la abundancia de árboles y con la abundancia de
árboles con valores bajos de decaimiento.
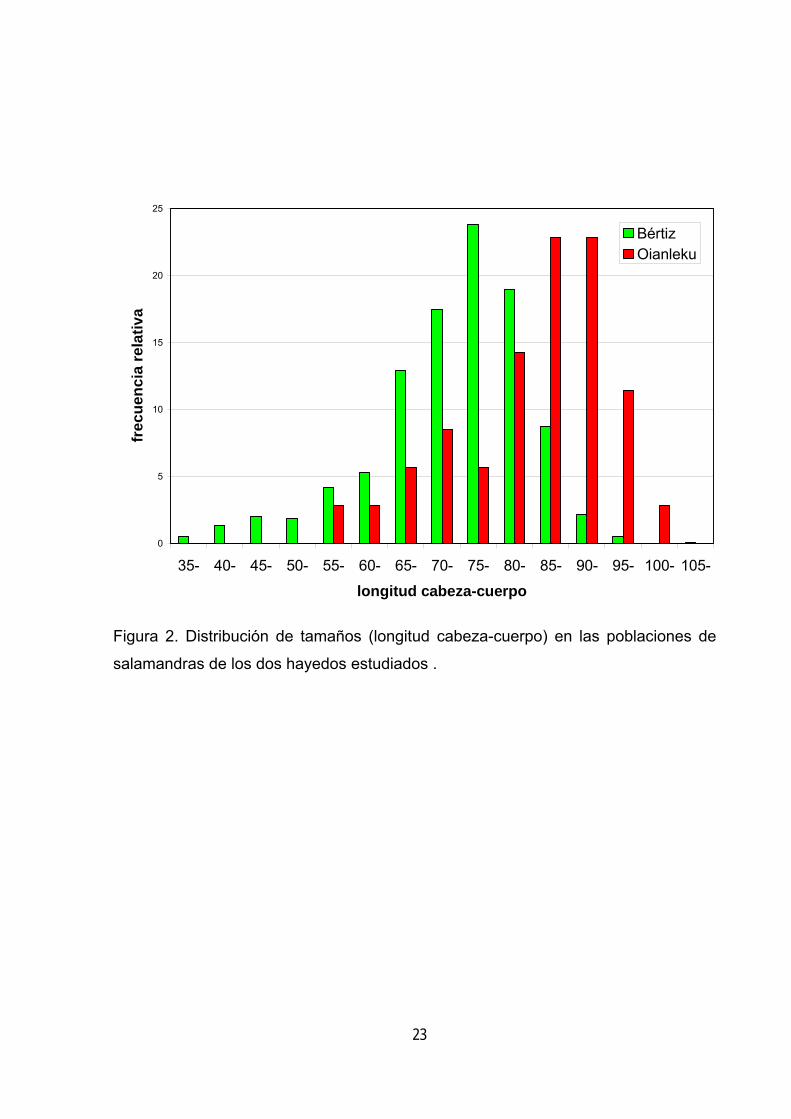
Por último, se ha contrastado la distribución de tamaños de las salamandras
en ambos hayedos. En Bértiz el rango de tamaños es mayor, faltando en Oianleku la 17

fracción de la población de menor tamaño (por debajo de 55 mm) (figura 2). Además
la población de salamandras del hayedo pobre en madera muerta presenta una
mayor proporción de salamandras grandes y el tamaño medio de los animales es
significativamente mayor (t = 5.12, p < 0.001).
Los resultados obtenidos destacan la correlación positiva entre la abundancia de
salamandras y las características del hayedo maduro-viejo (tipo de bosque que
contaría con gran cantidad de madera muerta, con algunos árboles vivos de gran
diámetro y con árboles vivos en distintos estados de decaimiento). Varios estudios
han mostrado esta relación (Welsh y Lind 1988, Herbeck y Larsen 1999),
destacando la importancia de la complejidad estructural del bosque, que proporciona
un rango más estrecho y estable de humedad y temperatura, y apuntando
principalmente al volumen de madera muerta como factor determinante de la
abundancia de salamandras o de la riqueza en especies de este grupo de anfibios
(Petranka et al. 1994, Butts y McComb 2000, Villate y González-Esteban 2002). En
estos trabajos se destaca sobre todo la importancia de los logs, elementos que
pueden considerarse como un hábitat crítico para las salamandras terrestres ya que
su gran capacidad de retener agua las protege de la desecación, al tiempo que
facilita la termoregulación y sirve de sustrato de alimentación (Maser y Trappe 1984,
Aubry et al. 1988). En algunos trabajos se destaca la preferencia de las salamandras
por las parcelas que cuentan con logs de mayor tamaño y por aquellas en los que
existen logs que presentan un estado de pudrición más avanzado (Maser y Trappe
1984, Harmon et al. 1986, Corn y Bury 1991), ya que éstos proporcionan más
cobertura, una oportunidad más larga para ser usados, mejores lugares de refugio y
mayor abundancia de presas.
En el presente trabajo el tamaño o el estado de pudrición de la madera
muerta no determina la abundancia de salamandras y el volumen de snags
desempeña un papel tan importante como el volumen de logs. Salamandra
salamandra se comporta como un consumidor generalista de invertebrados del suelo
(principalmente coleópteros, miriápodos, dermápteros, opiliones, oligoquetos (Bas
1983, Salvador y García-Paris 2001)) para los que la madera muerta resulta un
recurso muy atractivo y probablemente crítico (sobre todo en el hayedo acidófilo en
donde el estrato vegetal inferior está pobremente representado y la capa de
hojarasca, donde existe, es de escaso espesor). Probablemente las salamandras 18

satisfagan sus necesidades de alimento sin depender de la fauna de invertebrados
característica de un determinado estado de pudrición de la madera muerta. Por otra
parte, a simple vista, tanto logs como snags, proporcionan gran cantidad de huecos
donde refugiarse independientemente de su estado de pudrición. Cabe pensar
además que los elementos estructurales en avanzado estado de pudrición han de
resultar, por frágiles, menos seguros como refugio frente a los depredadores.
Las salamandras son marcadamente territoriales (Bas 1983), por lo que en los
hayedos con poca madera muerta la competencia por este valioso recurso puede
estar ocasionando las diferencias observadas en cuanto a la distribución de
tamaños. En estos hayedos las salamandras más pequeñas no tendrían acceso a
los lugares más seguros y con mayor cantidad de alimento y su mortalidad sería
mayor que en los hayedos estructuralmente complejos. De este modo los hayedos
pobres en madera muerta contarían con poblaciones pequeñas y además
envejecidas, y por tanto la probabilidad de persistencia de dichas poblaciones sería
menor.
Las salamandras son un importante componente ecológico de las
comunidades del suelo del bosque. Actuan como depredadores sobre una amplia
variedad de invertebrados realizando una eficiente conversión de nutrientes y
biomasa de baja calidad. La concentración media de proteinas en las salamandras
terrestres es del 50% (Burton y Likens 1975), lo que las convierte en una fuente de
energía de alta calidad para sus depredadores. Por otra parte, las salamandras son
los vertebrados más numerosos en el hayedo y a pesar de su lento crecimiento y
baja tasa metabólica (Bas 1983) su producción anual de biomasa debe ser superior
a la de aves y micromamíferos, tal como se ha comprobado en especies similares
(Burton y Likens 1975).
La dependencia que muestran las salamandras terrestres de las condiciones
microclimáticas del ecosistema forestal las hacen sensibles a las alteraciones. Esta
circunstancia, junto con su gran abundancia en bosques con altos niveles de
complejidad estructural y la facilidad del censado de sus poblaciones, permiten
señalar a esta especie como un buen indicador de los efectos de la gestión forestal
sobre el hayedo y de los procesos y tiempos de recuperación asociados con los
aprovechamientos de este tipo de bosque.
19

Referencias Alford, R.A., Richards, S.J., 1999. Global amphibian declines: a problem in applied ecology. Ann. Rev. Ecol. Syst., 30:133-165. Andrew, N.A., Pollock, K.H., 1999. Clearcuts, salamanders, and field studies. Conservation Biology, 13:206-208. Aubry, K.B., Jones, L.C., Hall, P.A., 1988. Use of woody debris by plethodontid salamanders in Douglas-fir forests in Washington. En: Management of amphibians, reptiles, and small mammals in North America. USDA, Gen. Tech. Report RM-166, 32-37. Bas, S., 1983. Estudio de la situación microevolutiva y de la ecología de Salamandra salamandra (L.) en el N. OE. ibérico. Tesis Doctoral, Universidad de Santiago de Compostela. Burton, T.M., Likens, G.E., 1975. Salamander population and biomass in the Hubbard Brook Experimental Forest, New Hampshire. Copeia,100:541-546. Butts, S.R., McComb, W.C., 2000. Associations of forest-floor vertebrates with coarse woody debris in managed forests of Western Oregon. J. Wildl. Manage., 64:95-104. Corn, P.S., Bury, R.B., 1991. Terrestrial amphibian communities in the Oregon Coast range. En: Wildlife and vegetation of unmanaged Douglas-fir forests. L.F. Ruggiero, K.B. Aubry, A.B. Carey, M.H. Huff (Eds.). PNW-GTR-285. U.S. Forest Service, Portland, Oregon, 305-318. Villate, I., González-Esteban, J., 2002. Incidencia en la comunidad de vertebrados de los elementos de diversidad forestal relacionados con la gestión del bosque. Servicio de Conservación de la Biodiversidad. Gobierno de Navarra. Griffiths, R.A., 1995. Newts and salamanders of Europe. London, T&AD Poyser Ltd. Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V., Latton, J.D., Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R., Lienkaemper, G.W., Cromack, K. Jr., Cummins, K.W., 1986. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research, 15:133-302. Harpole, D.H., Haas, C.A., 1999. Effects of seven silvicultural treatments on terrestrial salamanders. Forest Ecol. Manage., 114:349-356. Herbeck, L.A., Larsen, D.R, 1999. Plethodontid salamander response to silvicultural practices in Missouri Ozark forests. Conserv. Biol., 13:623-632. Lizana, M., Barbadillo, L.J., 1997. Legislación, protección y estado de conservación de los anfibios y reptiles españoles. En: Distribución y biogeografía de los anfibios y reptiles en España y Portugal. J.M. Pleguezuelos (Ed.). Granada, Asociación Herpetológica Española, Universidad de Granada, 477-516. Maser, C. Trappe, J.M., 1984. The seen and unseen world of the fallen tree. PNW-GTR-164. U.S. Forest Service, Portland, Oregon. deMaynadier, P.G. y Hunter, M.L. Jr., 1995. The relationship between forest management and amphibian ecology: a review of the North American literature. Environ. Rev., 3:230-261. Petranka, J.W., Brannon, M.P., Hopey, M.E. , Smith, C.K., 1994. Effects of timber harvesting on low elevation populations of southern Appalachian salamanders. For. Ecol. Manage., 67:135-147. Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins University Press.
20

Salvador, A., García-Paris, M., 2001. Anfibios españoles. Talavera de la Reina, Canseco Editores. Welsh, H.H., Lind, A.J., 1988. Old growth forests and the distribution of the terrestrial herpetofauna. En: Management of amphibians, reptiles, and small mammals in North America. USDA, Gen. Tech. Report RM-166, 439-455.
21
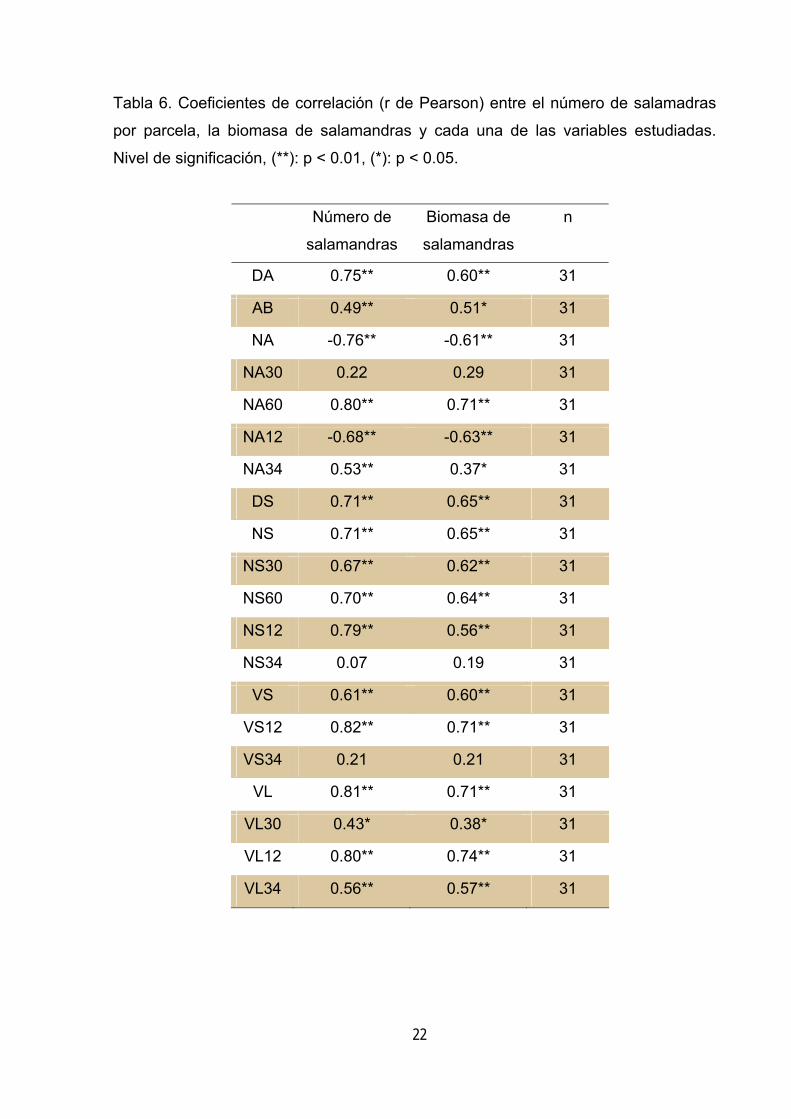
Tabla 6. Coeficientes de correlación (r de Pearson) entre el número de salamadras
por parcela, la biomasa de salamandras y cada una de las variables estudiadas.
Nivel de significación, (**): p < 0.01, (*): p < 0.05.
Número de
salamandras
Biomasa de
salamandras
n
DA 0.75** 0.60** 31
AB 0.49** 0.51* 31
NA -0.76** -0.61** 31
NA30 0.22 0.29 31
NA60 0.80** 0.71** 31
NA12 -0.68** -0.63** 31
NA34 0.53** 0.37* 31
DS 0.71** 0.65** 31
NS 0.71** 0.65** 31
NS30 0.67** 0.62** 31
NS60 0.70** 0.64** 31
NS12 0.79** 0.56** 31
NS34 0.07 0.19 31
VS 0.61** 0.60** 31
VS12 0.82** 0.71** 31
VS34 0.21 0.21 31
VL 0.81** 0.71** 31
VL30 0.43* 0.38* 31
VL12 0.80** 0.74** 31
VL34 0.56** 0.57** 31
22
0
5
10
15
20
25
35- 40- 45- 50- 55- 60- 65- 70- 75- 80- 85- 90- 95- 100- 105-
longitud cabeza-cuerpo
frec
uenc
ia re
lativ
a
BértizOianleku
Figura 2. Distribución de tamaños (longitud cabeza-cuerpo) en las poblaciones de
salamandras de los dos hayedos estudiados .
23

Los paseriformes nidificantes y la complejidad estructural
El papel de la comunidad de aves forestales como indicador de las condiciones del
bosque ha sido objeto de controversia (Morrison et al. 1992). No obstante, las
especies especialistas muestran una mayor sensibilidad a los cambios en el
ecosistema y pueden ser utilizadas como indicadoras y en líneas generales
parámetros como la abundancia, la riqueza y la diversidad de algunos grupos de
especies de aves se ven significativamente afectados por las alteraciones que sufre
el bosque (Szaro y Balda 1982). Estos cambios generalmente reducen la
complejidad estructural del ecosistema y sus efectos sobre las aves que nidifican en
él se encuentran bien documentados (Perry 1994, Hall et al. 1997, Bunnell et al.
1997), sobre todo porque las aves son fácilmente identificables y cuentan con
poblaciones con un número de efectivos suficiente como para realizar análisis
estadísticos y construir modelos predictivos.
En el presente trabajo se estudia la relación entre distintos parámetros que
definen la riqueza y diversidad de la comunidad de paseriformes y la complejidad
estructural del hayedo.
Material y métodos El estudio se desarrolló en los hayedos de Bértiz, Añarbe y Oianleku. En ellos se
seleccionaron 72 parcelas circulares de 50 m de radio en las que se caracterizó la
complejidad estructural y se cuantificó la comunidad de paseriformes nidificantes
(excepto córvidos) mediante estaciones de censo. La selección de las parcelas se
realizó priorizando el obtener un conjunto de muestras en el que estuviera
representada la heterogeneidad estructural propia del hayedo. Las unidades de
muestreo están separadas entre sí un mínimo de 200 m y al menos 150 m del borde
del bosque.
El muestreo se realizó durante los meses de junio y julio. Todos los censos se
han realizado entre las 7:00 y las 11:00 de la mañana, en días sin lluvia, viento o
niebla. Cada unidad de muestreo ha sido censada 2 veces con un intervalo mínimo
de 10 días y siempre a distinta hora.
24

Para aumentar la probabilidad de detección cada unidad de muestreo ha sido
censada por dos observadores. Se han anotado todas las aves vistas u oídas dentro
de la unidad de muestreo. El protocolo seguido ha sido adaptado de Gunn et al.
(2000). Una vez alcanzada la posición central de la parcela se dejó transcurrir un
período de cinco minutos antes de comenzar el censo. No se tuvieron en cuenta las
aves observadas durante estos primeros cinco minutos. El censado comenzaba con
cinco minutos de silencio, tras los cuales se procedía a emitir durante otros cinco
minutos un reclamo grabado, acabando el censo con otros cinco minutos de silencio.
El reclamo utilizado ha sido las mobbing calls (llamadas de hostigamiento colectivo)
del carbonero común. Se ha empleado un reproductor de casetes provisto de 2
amplificadores de 2 W cada uno. Este método proporciona un mayor rendimiento de
observaciones en medios en los que la detectabilidad de las aves es baja (Hurd,
1996; Cassady et al., 1998; Gunn et al., 2000), como es el caso del hayedo, en
donde el dosel supera en ocasiones los 30 m de altura. En todos los casos el
número de especies observadas en los períodos de emisión del reclamo ha sido
superior al observado en el período inicial de silencio. En ensayos previos se
comprobó que todas las especies propias del hayedo responden a las mobbing calls
del carbonero común aproximándose a la fuente del sonido y emitiendo llamadas
similares. Gunn et al. (2000) estiman que este método atrae a las aves en un radio
de aproximadamente 100 m.
La abundancia y riqueza de paseriformes nidificantes se ha estimado a partir
del número de especies (NSP), el número total de parejas (NPAR) y la diversidad
(índice de Shannon-Weaver) (DVD).
La caracterización de la complejidad estructural de cada parcela se realizó
conforme a los protocolos de trabajo descritos en el primer capítulo.
Resultados Mediante las estaciones de censo se han obtenido observaciones de 15 especies
(tabla 7). En primer lugar cabe señalar que los tres parámetros estudiados ofrecen
valores significativamente más altos en el hayedo de mayor complejidad estructural
(figura 3). Por otra parte, el número de especies y de parejas tienen un
comportamiento similar en cuanto a su correlación con las variables que caracterizan
la complejidad estructural del hayedo (tabla 8). Estas correlaciones indican que la
25

comunidad de paseriformes es más rica en el hayedo que presenta mayor cantidad
de madera muerta (tanto en forma de snags, como en forma de logs) (tabla 8).
Para estudiar la relación entre las distintas especies y la complejidad
estructural del hayedo se ha realizado un análisis de correspondencias a partir de la
matriz de frecuencias de las especies de aves en cada uno de los hayedos. En este
análisis no han intervenido las especies que han proporcionado un escaso número
de observaciones (chochín, mito, zorzal charlo, mirlo común, colirrojo real). La
distribución de especies y bosques sobre el plano definido por los dos factores
extraidos (figura 4) permiten destacar la asociación de un grupo de especies con
Bértiz, el hayedo estructuralmente más complejo. Estas son el carbonero palustre, el
trepador, el petirrojo, el agateador común y el herrerillo capuchino.
Los resultados obtenidos muestran como al aumentar la complejidad estructural del
hayedo (sobre todo de madera muerta, tanto logs como snags) aumenta la
abundancia y la riqueza de la comunidad de paseriformes nidificantes. Numerosos
estudios han puesto de manifiesto dicha relación (por ejemplo, Balda 1975, Wetmore
et al. 1985), estando orientados la mayor parte de estos trabajos a estudiar la
comunidad de paseriformes en distintos estadíos seriales o en distintas etapas de
las diferentes modalidades del aprovechamiento maderero. En ellos al igual que en
el presente trabajo se destaca que si bien existe una tendencia general a que los
estadíos seriales más avanzados cuenten con una mayor abundancia, riqueza y
diversidad de aves, existen diferencias entre las distintas especies en cuanto a
selección de hábitat. Por ello a la hora de utilizar la abundancia o la diversidad de los
paseriformes forestales como un indicador de las condiciones del bosque es
necesario identificar las especies sensibles a las modificaciones del ecosistema
propias de la sucesión vegetal o de la gestión forestal.
En los hayedos estudiados, cinco especies aparecen ligadas a la masa
estructuralmente más compleja y cada una de estas especies explota dentro del
hayedo recursos tróficos netamente distintos (Snow y Perrins 1998) por lo que cabe
pensar que la abundancia de éstas en su conjunto puede ser un buen indicador de
las condiciones naturales del hayedo.
26

Referencias Balda, R.P., 1975. Vegetation structure and breeding bird diversity. En: Management of forests and range habitats for nongame birds. D.R. Smith (tech. co-ord.) USDA Forest Service, Gen. Tech. Rep., WO-1, 59-80. Bunnell, F.L., Kremsater, F.L., Wells, R.W., 1997. Likely consequences of forest management on terrestrial, forest-dwelling vertebrates in Oregon. Centre for Applied Conservation Biology, University of British Columbia, RM-7. Cassady, C., Bélisle, M., Desrochers, A. & Hannon, S. 1998. Winter responses of forest birds to habitat corridors and gaps. Conservation Ecology (online), 2(2):13. Gunn, J.S., Desrochers, A., Villard, M.A., Bourque, J. & Ibarzabal, J. 2000. Playbacks of mobbing calls of black-capped chickadees as a method to estimate reproductive activity of forest birds. J. Field Ornithol., 71(3): 472-482. Hall, L.S., Morrison, M.L., Block, W.M., 1997. Songbird status and roles. En: Songbird ecology in Southwestern Ponderosa Pine Forests: A literature review. Block, W.M., Finch, D.M. (Eds.). USDA, Forest Service, Gen. Tech. Rep., RM-292. Hurd, C.R. 1996. Interspecific attraction to the mobbing calls of black-capped chickadees (Parus atricapillus). Behav. Ecol. Sociobiol., 38: 287-292. Morrison, M.L., Marcot, B.G., Mannan, R.W., 1992. Wildlife-habitat relationships: concepts and applications. University of Wisconsin Press, Madison. Perry, D.A., 1994. Forest Ecosystems. Baltimore and London, The Johns Hopkins University Press. Snow, D.W., Perrins, C.M., 1998. The birds of the Western Palearctic. Concise Edition. Oxford, Oxford University Press. Szaro, R.C., Balda R.P., 1982. Selection and monitoring of avian indicator species: an example from a ponderosa pine forest in the southwest. USDA Forest Service, Gen. Tech. Rep., RM-89. Wetmore, S.P., Keller, A., Smith, G.E.J., 1985. Effects of logging on bird populations in British Columbia as determined by a modified point count method. Can. Field Naturalist, 99:224-233.
27
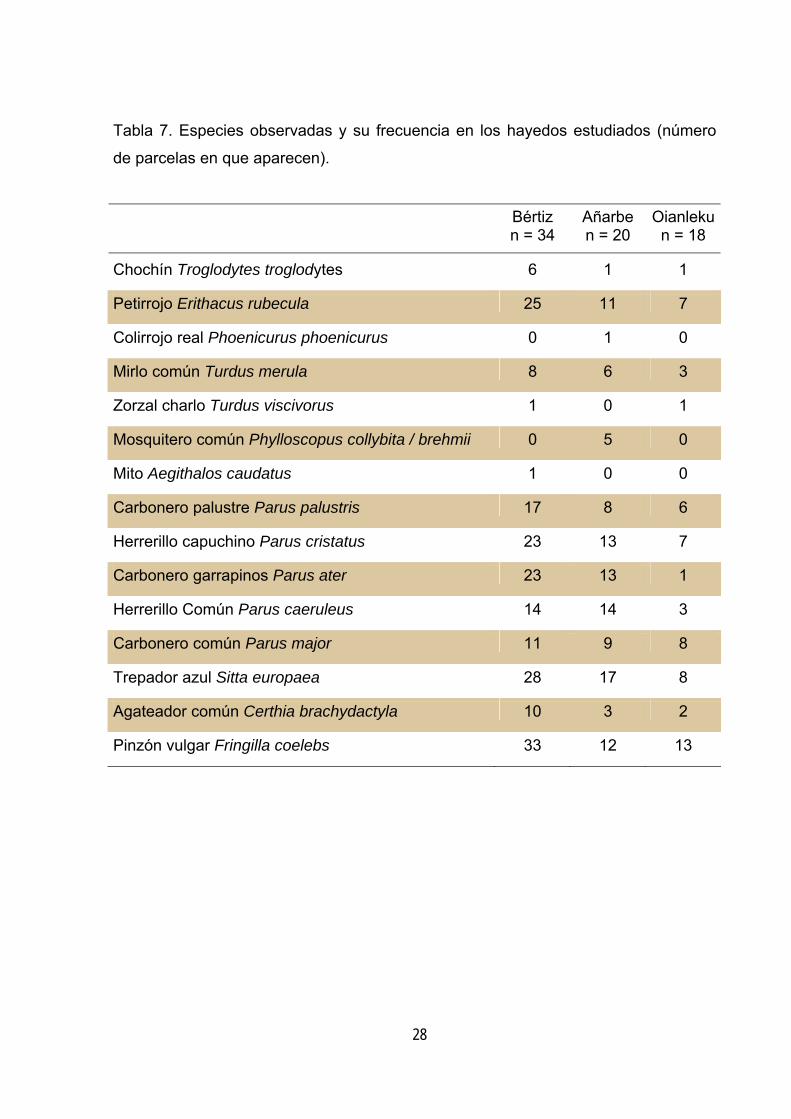
Tabla 7. Especies observadas y su frecuencia en los hayedos estudiados (número
de parcelas en que aparecen).
Bértiz n = 34
Añarbe n = 20
Oianlekun = 18
Chochín Troglodytes troglodytes 6 1 1
Petirrojo Erithacus rubecula 25 11 7
Colirrojo real Phoenicurus phoenicurus 0 1 0
Mirlo común Turdus merula 8 6 3
Zorzal charlo Turdus viscivorus 1 0 1
Mosquitero común Phylloscopus collybita / brehmii 0 5 0
Mito Aegithalos caudatus 1 0 0
Carbonero palustre Parus palustris 17 8 6
Herrerillo capuchino Parus cristatus 23 13 7
Carbonero garrapinos Parus ater 23 13 1
Herrerillo Común Parus caeruleus 14 14 3
Carbonero común Parus major 11 9 8
Trepador azul Sitta europaea 28 17 8
Agateador común Certhia brachydactyla 10 3 2
Pinzón vulgar Fringilla coelebs 33 12 13
28
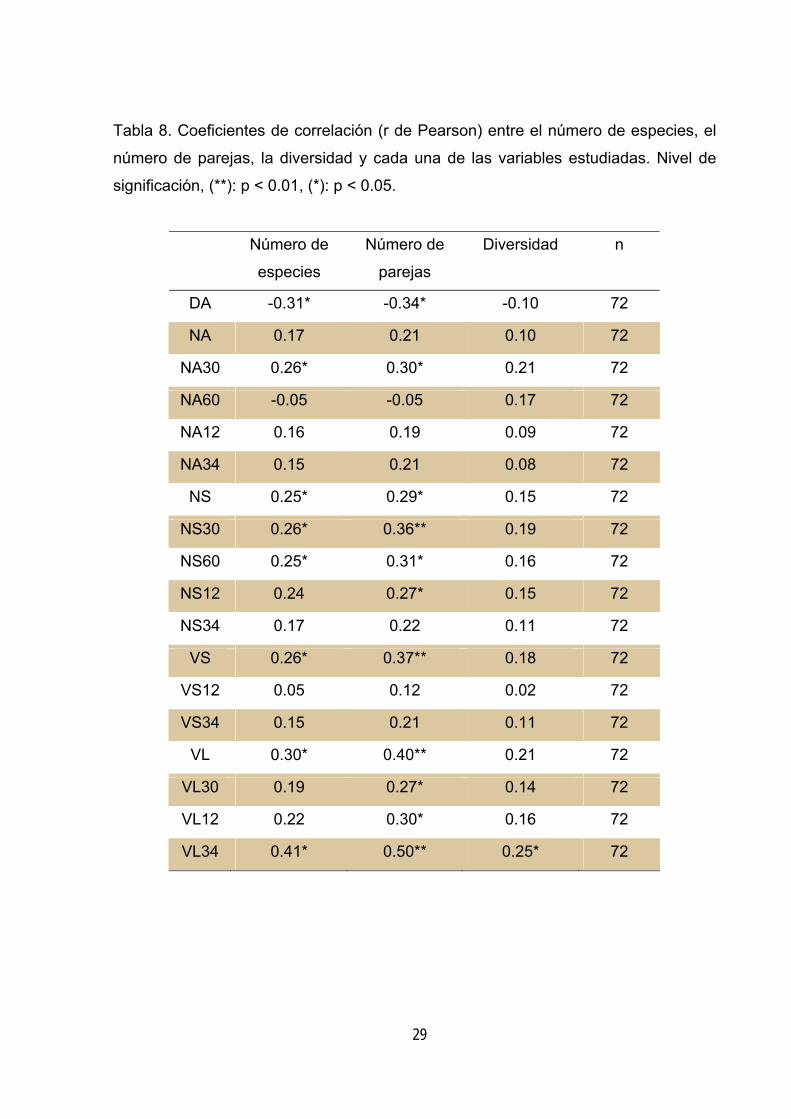
Tabla 8. Coeficientes de correlación (r de Pearson) entre el número de especies, el
número de parejas, la diversidad y cada una de las variables estudiadas. Nivel de
significación, (**): p < 0.01, (*): p < 0.05.
Número de
especies
Número de
parejas
Diversidad n
DA -0.31* -0.34* -0.10 72
NA 0.17 0.21 0.10 72
NA30 0.26* 0.30* 0.21 72
NA60 -0.05 -0.05 0.17 72
NA12 0.16 0.19 0.09 72
NA34 0.15 0.21 0.08 72
NS 0.25* 0.29* 0.15 72
NS30 0.26* 0.36** 0.19 72
NS60 0.25* 0.31* 0.16 72
NS12 0.24 0.27* 0.15 72
NS34 0.17 0.22 0.11 72
VS 0.26* 0.37** 0.18 72
VS12 0.05 0.12 0.02 72
VS34 0.15 0.21 0.11 72
VL 0.30* 0.40** 0.21 72
VL30 0.19 0.27* 0.14 72
VL12 0.22 0.30* 0.16 72
VL34 0.41* 0.50** 0.25* 72
29
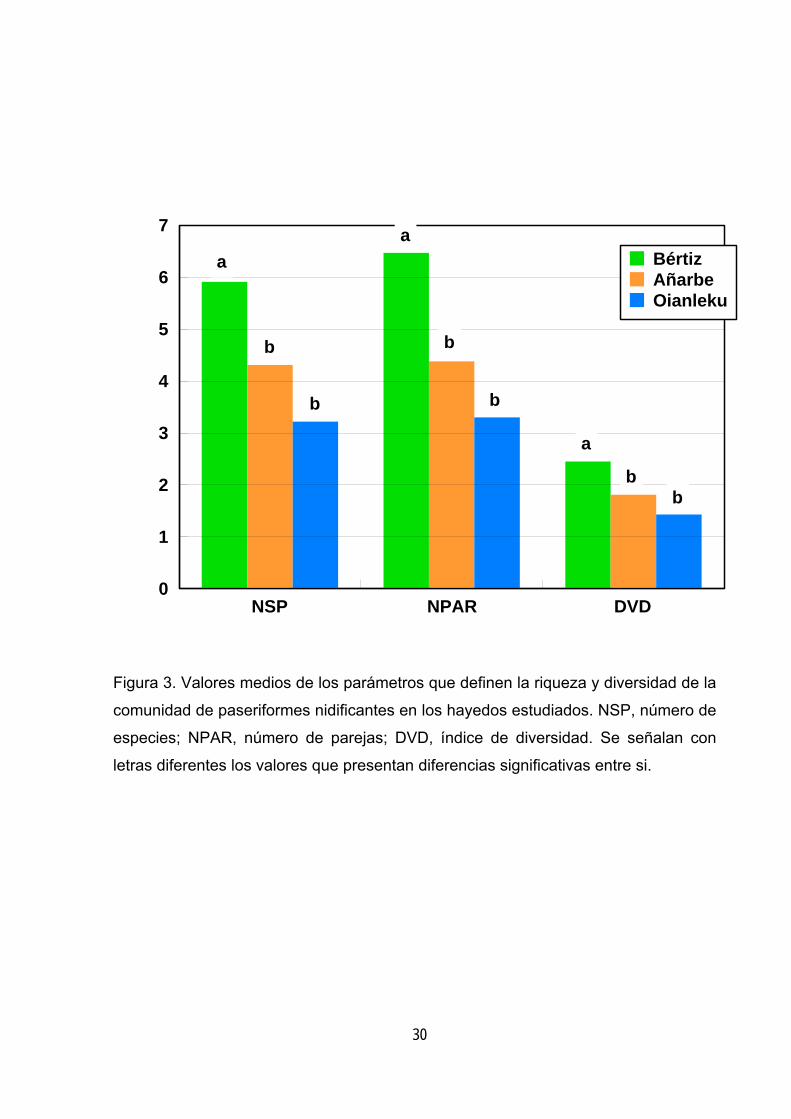
NSP NPAR DVD0
1
2
3
4
5
6
7
BértizAñarbeOianleku
a
a
a
b
bb
b
bb
Figura 3. Valores medios de los parámetros que definen la riqueza y diversidad de la
comunidad de paseriformes nidificantes en los hayedos estudiados. NSP, número de
especies; NPAR, número de parejas; DVD, índice de diversidad. Se señalan con
letras diferentes los valores que presentan diferencias significativas entre si.
30
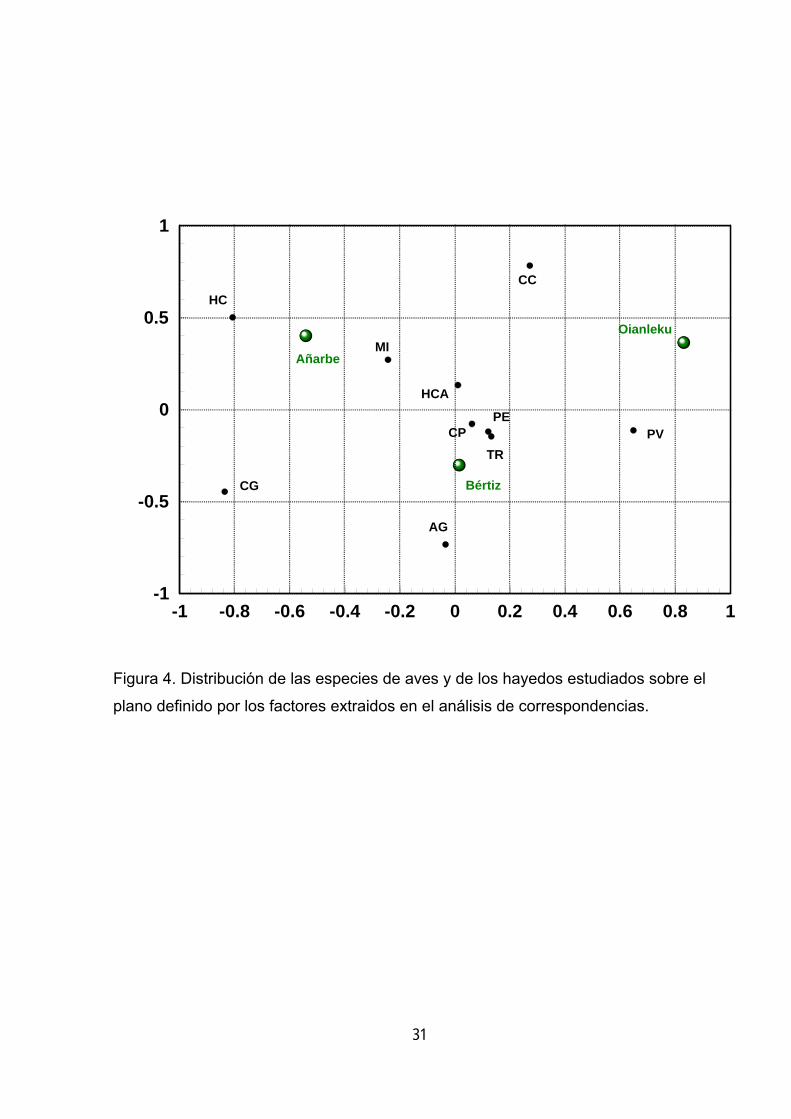
-1 -0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8 1-1
-0.5
0
0.5
1
AG
PV
CCHC
CG
MI
HCA
CPTR
PE
Añarbe
Bértiz
Oianleku
Figura 4. Distribución de las especies de aves y de los hayedos estudiados sobre el
plano definido por los factores extraidos en el análisis de correspondencias.
31

Los micromamíferos y la complejidad estructural Los micromamíferos contribuyen a la biodiversidad de los bosques templados, tanto
a la diversidad de especies y formas de vida como a la diversidad funcional del
ecosistema forestal. Dentro de este grupo podemos encontrar especies
consumidoras de plantas (incluyendo semillas y frutos), de líquenes y hongos, de
invertebrados, y todas ellas sirven como alimento a una amplia variedad de aves,
reptiles y mamíferos. Además se ha comprobado que en algunos bosques su
presencia es necesaria para que se produzca la dispersión de las micorrizas (Carey
y Johnson 1995). Por ello, su abundancia y diversidad puede servir como un buen
indicador de los cambios que suceden en la estructura del bosque (Sullivan y
Sullivan 2001).
Varios trabajos se han ocupado de estudiar las modificaciones que sufren los
patrones de abundancia de las distintas especies como producto de las distintas
técnicas empleadas en el aprovechamiento maderero (Bayne y Hobson, 1998;
Waters y Zabel 1998; Sullivan et al 1999). En lineas generales los resultados de
estos trabajos coinciden al señalar que la diversidad y la abundancia se ven
afectadas y no existe un patrón de respuesta común para todas las especies.
En el presente trabajo se estudia la relación entre la abundancia y la riqueza
de la comunidad de micromamíferos y la complejidad estructural del hayedo.
Material y métodos El estudio se ha desarrollado en los hayedos de Bértiz y Oianleku. Con objeto de
contrastar la composición de la comunidad de micromamíferos y la complejidad
estructural se seleccionaron en cada uno de los hayedos 10 parcelas (del conjunto
de parcelas utilizadas en la caracterización). Las parcelas seleccionadas en cada
masa son representativas de un hayedo estructuralmente complejo (Bértiz) y de un
hayedo de complejidad baja (Oianleku). Dentro de cada parcela se eligieron 10
árboles con un dbh superior a 30 cm y sobre cada uno de ellos, a ras del suelo y a
una altura superior a 2 m, se colocaron sendos tubos de PVC de 10 cm de longitud
y 3 cm de anchura con cinta adhesiva de doble cara adherida a la parte superior
interna del tubo. En la cinta adhesiva quedan adheridos mechones de pelo de los
micromamíferos que los visitan. Los pelos han sido identificados siguiendo el 32

protocolo descrito por Teerink (1991). Los tubos permanecieron en los árboles
durante los meses de julio y agosto.
Resultados Los resultados muestran la mayor riqueza del hayedo complejo. En éste se
detectaron 5 especies (4 roedores y 1 insectívoro), mientras que en el hayedo pobre
en estructuras solo aparecen tres especies de roedores y ningún insectívoro (tabla
9). Utilizando como índices de abundancia el número de árboles y parcelas visitadas
por los micromamíferos se observa igualmente frecuencias mayores en el hayedo
complejo (tabla 9).
Estos resultados sugieren que una mayor oferta de estructuras se
corresponde con una comunidad de micromamíferos más rica. La información que
ofrecen distintos trabajos señala a la madera muerta como un importante
componente del hábitat de distintas especies de los géneros Sorex (Lee, 1995),
Peromyscus (McCay, 2000; Greenberg 2002) y Clethrionomys (Tallmon y Mills 1994;
Bowman et al. 2000). En nuestro caso el hayedo estructuralmente complejo ofrece
los recursos espaciales necesarios (refugio y vías de dispersión-colonización) para
que los roedores puedan aprovechar los años de cosecha abundante de hayuco y
sustratos de alimentación (logs) para los insectívoros.
Algunos de los trabajos que han estudiado el efecto de la reducción de la
complejidad estructural producto de los diferentes tratamientos selvícolas sobre la
comunidad de micromamíferos (Probst y Rakstad, 1987; Waters y Zabel, 1998;
Sullivan et al., 1999) muestran que no existe un patrón de respuesta común para
todas las especies: algunas se muestran indiferentes a los cambios, sin que sus
parámetros poblacionales se vean afectados, en otros casos su abundancia es
significativamente menor en masas jóvenes regulares e incluso existen especies que
se ven favorecidas. Sin embargo en el hayedo acidófilo la respuesta observada es
única: la disminución de la complejidad estructural conlleva la reducción de la
diversidad de la comunidad de micromamíferos y ésta es producto de la reducción
de los efectivos de todas las especies. Hay que tener en cuenta que la baja
diversidad florística del hayedo proporciona unos recursos tróficos y espaciales muy
limitados, que resultan críticos para la comunidad de micromamíferos. En el mejor de
los casos en el hayedo difícilmente se van a poder mantener, en grandes áreas de
forma continuada, poblaciones estables de 2 o 3 especies de micromamíferos, 33

conduciendo cualquier alteración notable de la complejidad estructural al
enrarecimiento o a la desaparición de la comunidad característica de este tipo de
bosque.
Referencias Bayne, E.M. y Hobson, K.A., 1998. The effects of habitat fragmentation by forestry and agriculture on the abundance of small mammals in the southern boreal mixedwood forest. Can. J. Zool., 76:62-69. Bowman, J.C., Sleep, D., Forbes, G.J., Edwards, M., 2000. The association of small mammals with coarse woody debris at log and stand scales. Forest Ecol. Manage., 129:119-124. Carey, A.B. y Johnson, M.L., 1995. Small mammals in managed, naturally young, and old-growth forests. Ecol. Apl., 5:336-352. Greenberg, C.H., 2002. Response of white-footed mice (Peromyscus leucopus) to coarse woody debris and microsite use in southern Appalachian treefall gaps. Forest Ecol. Manage., 164:57-66. Lee, S.D., 1995. Comparison of population characteristics of three species of shrews and the shrew-mole habitats with different amounts of coarse woody debris. Acta Theriol., 40:415-424. McCay, T.S., 2000. Use of woody debris by cotton mice (Peromyscus gossypinus) in a Southeastern pine forest. J. Mammal., 81:527-535. Probst, J.R. y Rakstad, D.S., 1987. Small mammal communities in three aspen stand-age classes. Canadian Field-Naturalist, 101:362-368. Sullivan, T. P, Lautenschlager, R.A., Wagner, R.G., 1999. Clearcutting and burning of northern spruce-fir forests: implications for small mammal communities. J. Appl. Ecol., 36:327-344. Sullivan, T.P., Sullivan, D.S., 2001. Influence of variable retention harvests on forest ecosystems. II. Diversity and population dynamics of small mammals. J. Appl. Ecol., 38: 1234-1252. Tallmon, D. y Mills, L.S., 1994. Use of logs within home ranges of California red-backed voles on a remnant of forest. J. Mammal., 75:97-101. Teerink, B.J. 1991. Hair of West-european Mammals. Atlas and identification key. Cambridge University Press. Cambridge: 1-224. Waters, J.R. y Zabel, C.J., 1998. Abundances of small mammals in fir forests in Northeastern California. J. Mammal., 79:1244-1253.
34
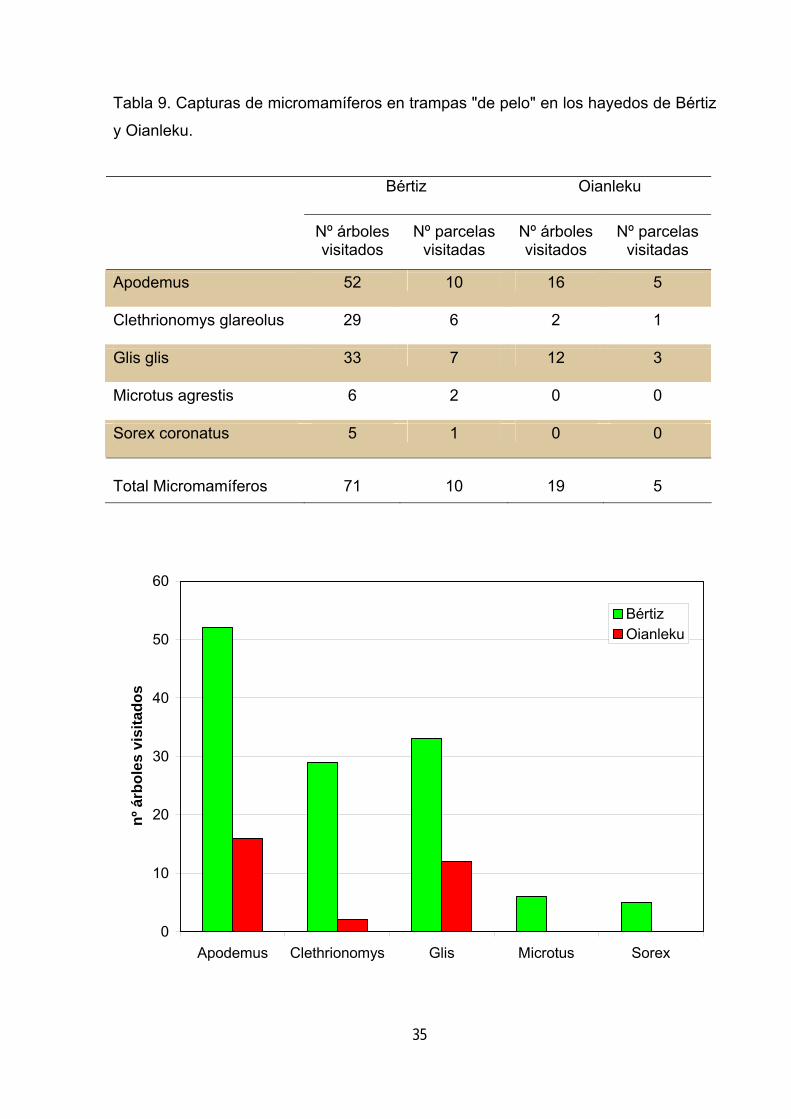
Tabla 9. Capturas de micromamíferos en trampas "de pelo" en los hayedos de Bértiz
y Oianleku.
Bértiz Oianleku
Nº árboles visitados
Nº parcelas visitadas
Nº árboles visitados
Nº parcelas visitadas
Apodemus 52 10 16 5
Clethrionomys glareolus 29 6 2 1
Glis glis 33 7 12 3
Microtus agrestis 6 2 0 0
Sorex coronatus 5 1 0 0
Total Micromamíferos
71
10
19
5
0
10
20
30
40
50
60
Apodemus Clethrionomys Glis Microtus Sorex
nº á
rbol
es v
isita
dos
Bértiz Oianleku
35

Conclusiones
1. El hayedo, a pesar de su baja diversidad florística, puede alcanzar en su último
estadío serial unos valores de complejidad estructural similares a los de otros
bosques templados.
2. Los tratamientos selvícolas utilizados en el hayedo de Oianleku han producido
una notable reducción de la complejidad estructural. Es especialmente significativa
la escasez de árboles muertos, circunstancia que afecta a los distintos tamaños y
clases de pudrición en los que puede presentarse este recurso.
3. Las especies de vertebrados estudiadas muestran unos patrones de abundancia,
riqueza y diversidad que fluctúan siguiendo una pauta similar. Estos parámetros
muestran valores significativamente mayores en el hayedo de mayor complejidad
estructural.
4. Los parámetros poblacionales de las especies de vertebrados estudiadas
muestran una notable sensibilidad a la variabilidad de la complejidad estructural del
hayedo. Circunstancia que permite señalarles como unos indicadores útiles de los
efectos de la gestión sobre el ecosistema.
5. En función de los resultados del presente estudio se puede afirmar, en el contexto
de la Directiva de Hábitats1, que el estado de conservación del hayedo de Oianleku
no puede considerase como favorable. El aprovechamiento maderero al que ha sido
sometido ha provocado una reducción de la complejidad estructural, que se traduce
en unos valores de biodiversidad, en cuanto a fauna de vertebrados, inferiores a los
que corresponden a un hayedo de su madurez.
36
1 Directiva 92/43/CEE del Consejo, de 21 de mayo de 1992, relativa a la conservación de los hábitats naturales y de la fauna y flora silvestres

























