
< 1 >
El origen del lenguaje humano [*]
Ansi mismo era necesario que el dicho agujero de
la nuez se (errase al tiempo que la vianda se tra-
ga porque no entre por el garguero que sería cosa
enojosa y perjudicial, de la cual tenemos expe-
riencia cada día cuando nos damos priesa al tra-
gar y nos entra algo por el garguero, porque le
sigue tras ello tos muy molesta y congojosa.
Bernardino Montaña de Monserrate,
Libro de la anathomia del hombre.
El anillo del rey Salomón
OS seres humanos somos los únicos organismos que
hablamos. Es decir, transmitimos a nuestros semejan-
tes, y recibimos de ellos, cualquier tipo de informa-
ción nueva, codificando deliberadamente nuestros mensajes
en combinaciones (palabras) de sonidos preestablecidos (sí-
labas). El resto de los animales sólo son capaces de inter-
cambiar informaciones muy concretas sobre algunos aspec-
tos de su vida, empleando para ello un sistema limitado de
sonidos y gestos que no están codificados de manera inten-
cionada.
Konrad Lorenz se refiere, en un delicioso libro titulado en
* El texto corresponde al Capítulo 16 de “La especie elegida. La larga
marcha de la evolución humana” (por Juan Luis Arsuaga e Ignacio Mar-
tínez), Ed. Temas de Hoy, 2001.
L

< 2 >
castellano El anillo del rey Salomón, a la leyenda de que el
rey Salomón poseía un anillo que le permitía hablar con las
bestias. Lorenz se ufanaba de que, sin necesidad del anillo, él
también era capaz de entender el sencillo vocabulario de los
animales; pero añadía que éstos no tienen un verdadero len-
guaje, sino que cada individuo posee de manera innata un
código de señales formado por voces y movimientos expre-
sivos que otro ejemplar de la misma especie es capaz de en-
tender, también de manera innata. Sin embargo, y ahí está la
diferencia fundamental con nuestro lenguaje, los animales
emiten esas señales como autómatas cuando se encuentran
en un determinado estado de ánimo, incluso aunque no haya
nadie para presenciarlas. Lorenz expresaba esta idea dicien-
do que con sus sonidos los animales no emiten «palabras»,
sino «interjecciones».
Según el Diccionario de la Real Academia Española, inter-
jección es «una voz que expresa alguna emoción súbita o un
sentimiento profundo, como asombro, sorpresa, dolor, mo-
lestia, amor...». Y eso es exactamente lo que, según Lorenz,
expresan los animales.
Estas ideas parecen ser ciertas para la mayor parte de los
animales, pero quizá no sean tan exactas en el caso de los
primates. Dorothy Cheney y Robert Seyfarth han estudiado
el mono tota (Cercopithecus aethiops) en su medio natural
en África, observando que estos animales, además de emitir
señales en forma de sonidos o gestos que avisan sobre su
motivación o estado de ánimo, también informan sobre de-
terminados aspectos del medio. Por ejemplo, tienen diferen-
tes vocalizaciones para señalar la presencia de distintos de-
predadores: serpiente, águila o leopardo. Las reacciones que
provocan estas llamadas en los oyentes son diferentes en
cada caso: se suben a los árboles si el aviso es de un leopar-
do, se esconden en los matorrales o miran hacia arriba si se
trata de un águila y se yerguen y escrutan el herbaje cuando
se advierte de la presencia de una serpiente. Es decir, que
cada llamada tiene un significado distinto que desencadena
una respuesta diferente; no son simples gritos de miedo ante

< 3 >
la presencia de un depredador.
Además, las investigaciones de Cheney y Seyfarth con
otras llamadas relacionadas con la vida social de los total han
descubierto que estos monos asocian vocalizaciones cuyos
significados son similares, aunque sean acústicamente muy
distintas (de la misma manera que nosotros haríamos con las
palabras «coche» y «automóvil»).
Por otra parte, en los años sesenta y setenta la idea de la
comunicación directa con los animales más parecidos a no-
sotros, los chimpancés y gorilas, fue tomada muy en serio en
algunos programas de investigación. Ya que chimpancés y
gorilas no pueden pronunciar físicamente las palabras, se les
facilitó la tarea de comunicarse con nosotros enseñándoles el
lenguaje de los sordomudos, un lenguaje de gestos, que sí
podían reproducir. Los chimpancés y gorilas resultaron dis-
cípulos aventajados en esto y pusieron de manifiesto su ca-
pacidad de asociar ideas, que nosotros expresaríamos con
palabras, a gestos o a fichas de diferentes formas, con dibu-
jos y colores (iconos).
Un bonobo llamado Kanzi, del que ya hemos hablado en
relación con su capacidad de tallar instrumentos de piedra,
entiende más de ciento cincuenta palabras del inglés habla-
do. Y Kanzi no ha sido el único chimpancé que ha mostrado
ciertas capacidades lingüísticas. Washoe fue el primer chim-
pancé que aprendió una serie de signos del lenguaje de los
sordomudos (ciento treinta y dos signos tras algo más de
cuatro años de adiestramiento), Sarah dio muestras de habili-
dad para detectar el orden de objetos empleados como pala-
bras, y Lana completaba adecuadamente oraciones construi-
das con caracteres representativos de palabras, atendiendo al
significado y orden de las mismas.
Una pregunta clásica en relación con los sistemas de comu-
nicación de los animales es si éstos pueden engañar a sus
congéneres en su propio beneficio. El saber mentir los haría
más «humanos», ya que indicaría que no son meros autóma-
tas, sino que son capaces de controlar sus expresiones. Pues

< 4 >
bien, hay numerosas observaciones sobre chimpancés en
libertad que engañan a sus compañeros, en contextos muy
diversos, con sus gestos, posturas y expresiones faciales.
Nuestros pecados son también los suyos.
Los resultados de todas estas investigaciones son muy valio-
sos porque han descubierto una incipiente destreza lingüísti-
ca en los primates que era negada en años anteriores, pero
han resultado decepcionantes en tanto que ninguno de ellos
nos ha comunicado información relevante alguna sobre sí
mismos. Los monos tocas parecen disponer de un limitado
repertorio de «palabras» que emplean en situaciones muy
concretas, y los chimpancés han demostrado ser muy compe-
tentes manejando símbolos e incluso unos consumados men-
tirosos; pero eso es todo. Quizá el mito del anillo del rey
Salomón sea, después de todo, sólo eso, un mito.
Puesto que el lenguaje humano es tan diferente del de nues-
tros parientes vivos, la cuestión de su origen y desarrollo
sólo puede ser abordada desde el campo de la paleontología.
La paleoneurología trata de determinar las capacidades men-
tales de una especie fósil a través de las impresiones que el
cerebro deja sobre la superficie interna del cráneo. Dos áreas
de la corteza cerebral, ambas en el hemisferio cerebral iz-
quierdo, están estrechamente relacionadas con el habla en los
humanos (figura 8.1). El área de Broca, situada en la tercera
circunvolución frontal (a la altura de la sien), es la encargada
de la construcción y planificación sintáctica; es decir, tradu-
ce los mensajes en una secuencia ordenada de movimientos
de los músculos que intervienen en la producción del habla.
Una lesión a este nivel perturba la capacidad de hablar y es-
cribir, pero no la comprensión del lenguaje hablado y se
puede seguir leyendo. Por su parte, el área de Wernicke,
situada entre la circunvolución temporal superior y el lóbulo
parietal (un poco por detrás y encima del oído), es la encar-
gada de la codificación y descodificación de los mensajes.
Una lesión en el área de Wernicke inhabilita para la correcta
comprensión y producción del lenguaje, hablado o escrito.

< 5 >
Según Phillip Tobias, la región inferior del lóbulo parietal
relacionada con el área de Wernicke está más desarrollada en
los fósiles de Homo habilis de Olduvai que en los australopi-
tecos, parántropos y antropomorfos. Por otra parte, el área de
Broca aparece netamente expandida tanto en los representan-
tes de Homo habilis/Homo rudolfensis como en los de Homo
ergaster. El desarrollo de esta área en los primeros humanos
es mucho mayor que el que presentan los australopitecos y
parántropos, en los que sólo está esbozada.
Es decir, que las regiones de la corteza cerebral más direc-
tamente relacionadas con la producción del lenguaje humano
ya estaban bien desarrolladas en los primeros representantes
de nuestro género. ¿Significa esto que aquellos humanos ya
poseían la capacidad de hablar? Aunque ésta es la conclusión
a la que llegan la mayor parte de los especialistas dedicados
al estudio de los moldes endocraneales de los homínidos
primitivos, existe un punto de vista contrapuesto.
En su libro The Wisdom of the Bones (La sabiduría de los
huesos), Alan Walker, el principal responsable del estudio
del Niño del Turkana, rechaza que este individuo, y por ex-
tensión todos los de su especie, fuera capaz de hablar. Para
llegar a tal conclusión, Walker se basa en que el canal medu-
lar de las vértebras torácicas del Niño del Turkana es muy
estrecho. Esta situación es la común entre los antropomorfos
pero no en los humanos modernos, que tenemos un canal medular ensanchado. Como consecuencia del reducido diá-
metro del canal medular, Walker defiende la hipótesis de que
la médula espinal del ejemplar fósil contenía menos neuro-
nas que la de los humanos modernos, por lo que la región
torácica del Niño del Turkana estaría menos inervada que la
nuestra. La única explicación plausible a este hecho, según
Walker, es la de aceptar que la musculatura torácica rela-
cionada con la respiración no era capaz de realizar los preci-
sos movimientos inspiratorios y espiratorios que requiere el
habla humana. ¿Cómo explicar entonces el gran desarrollo
del área de Broca que refleja la superficie endocraneal del
Niño del Turkana? A partir de los resultados de las modernas

< 6 >
técnicas de exploración de la actividad cerebral (en concreto
de la conocida como PET, positron emission tomography o
tomografía por emisión de positrones»), que relacionan la
región de la corteza cerebral circundante al área de Broca
también con el manejo de la mano derecha, Walker ha pro-
puesto que el desarrollo de esta área en los primeros huma-
nos no fue una adaptación relacionada con el habla, sino con
la talla de la piedra.
En resumen, aunque los distintos estudios sobre las áreas de
la corteza cerebral de los primeros homínidos concuerdan en
señalar un desarrollo mayor de las áreas vinculadas con el
lenguaje (especialmente el área de Broca) en los primeros
humanos que en los australopitecos, parántropos y antropo-
morfos, no existe acuerdo sobre su significado fisiológico.
La solución a este problema puede estar en las investigacio-
nes realizadas para establecer la anatomía del aparato fona-
dor de los homínidos fósiles.
El primate atragantado La segunda vía de investigación sobre el origen del lenguaje
la constituye el estudio del conjunto de órganos responsables
de la emisión de los sonidos que componen el habla humana.
Aunque a primera vista pueda parecer que éste es un aspecto
secundario, argumentando que las habilidades mentales ne-
cesarias para disponer de un lenguaje complejo no dependen
de la capacidad fisiológica para producirlo, lo cierto es que
no es posible componer una música para la que no existen
instrumentos.
Los sonidos en los que se basa el lenguaje humano se produ-
cen y modulan en la serie de cavidades que constituyen el
tramo superior del conducto respiratorio y reciben el nombre
genérico de tracto vocal: la laringe, la faringe y las cavida-
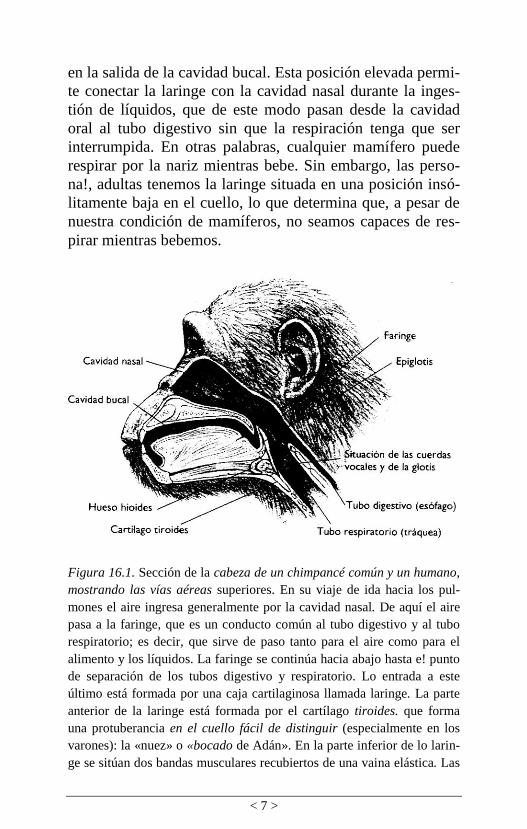
des nasal y oral (figura 16.1).
En todos los mamíferos, excepto en las personas adultas, la
laringe ocupa una posición alta en el cuello, situándose casi
< 7 >
en la salida de la cavidad bucal. Esta posición elevada permi-
te conectar la laringe con la cavidad nasal durante la inges-
tión de líquidos, que de este modo pasan desde la cavidad
oral al tubo digestivo sin que la respiración tenga que ser
interrumpida. En otras palabras, cualquier mamífero puede
respirar por la nariz mientras bebe. Sin embargo, las perso-
na!, adultas tenemos la laringe situada en una posición insó-
litamente baja en el cuello, lo que determina que, a pesar de
nuestra condición de mamíferos, no seamos capaces de res-
pirar mientras bebemos.
Figura 16.1. Sección de la cabeza de un chimpancé común y un humano,
mostrando las vías aéreas superiores. En su viaje de ida hacia los pul-
mones el aire ingresa generalmente por la cavidad nasal. De aquí el aire
pasa a la faringe, que es un conducto común al tubo digestivo y al tubo
respiratorio; es decir, que sirve de paso tanto para el aire como para el
alimento y los líquidos. La faringe se continúa hacia abajo hasta e! punto
de separación de los tubos digestivo y respiratorio. Lo entrada a este
último está formada por una caja cartilaginosa llamada laringe. La parte
anterior de la laringe está formada por el cartílago tiroides. que forma
una protuberancia en el cuello fácil de distinguir (especialmente en los
varones): la «nuez» o «bocado de Adán». En la parte inferior de lo larin-
ge se sitúan dos bandas musculares recubiertos de una vaina elástica. Las
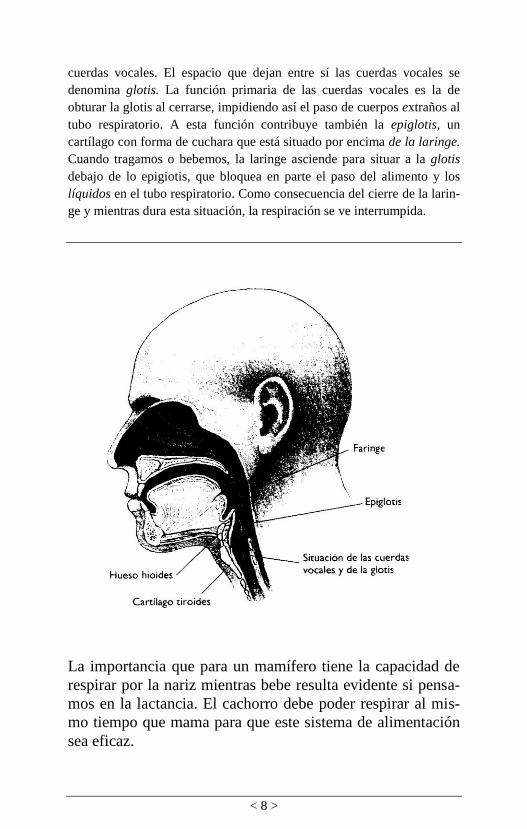
< 8 >
cuerdas vocales. El espacio que dejan entre sí las cuerdas vocales se
denomina glotis. La función primaria de las cuerdas vocales es la de
obturar la glotis al cerrarse, impidiendo así el paso de cuerpos extraños al
tubo respiratorio. A esta función contribuye también la epiglotis, un
cartílago con forma de cuchara que está situado por encima de la laringe.
Cuando tragamos o bebemos, la laringe asciende para situar a la glotis
debajo de lo epigiotis, que bloquea en parte el paso del alimento y los
líquidos en el tubo respiratorio. Como consecuencia del cierre de la larin-
ge y mientras dura esta situación, la respiración se ve interrumpida.
La importancia que para un mamífero tiene la capacidad de
respirar por la nariz mientras bebe resulta evidente si pensa-
mos en la lactancia. El cachorro debe poder respirar al mis-
mo tiempo que mama para que este sistema de alimentación
sea eficaz.

< 9 >
Para el lector perspicaz no habrá pasado desapercibido que
nuestros bebés también pueden respirar por la nariz mientras
maman o se beben su biberón. Su sagacidad le habrá llevado
asimismo a suponer correctamente que los lactantes huma-
nos tienen la laringe en la misma posición que el resto de los
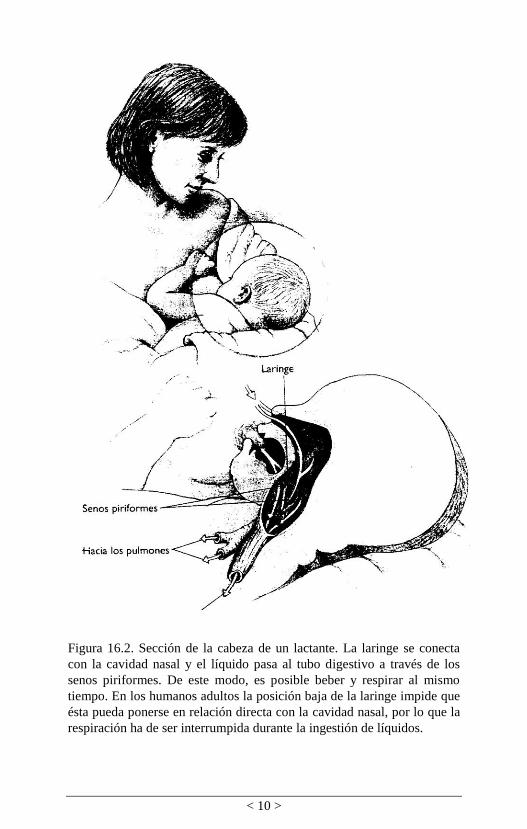
mamíferos (figura 16.2). El descenso de laringe en nuestra
especie se produce hacia los dos años de vida. A partir de
este momento, no sólo perdemos la facultad de respirar
mientras bebemos, sino que la insólita situación de la laringe
humana hace posible la obstrucción del conducto respiratorio
por el alimento, ya que la epiglotis no alcanza a obliterarlo
por completo (figura 16.1). Atragantarse no es una broma,
uno puede morir por ello.
Pero si nuestro tramo respiratorio superior ha perdido efi-
cacia en este aspecto (y también para la respiración y el olfa-
to), ¿cuál es la contrapartida? La respuesta está en la existen-
cia en nuestra especie de una faringe más larga que la de
ningún otro mamífero, que nos capacita para modular una
amplia serie de sonidos diferentes.
La producción del habla
En contra de lo que muchas personas creen, la mayoría de
los sonidos básicos que forman el habla humana no se origi-
nan directamente como tales en las cuerdas vocales. En la
producción de las vocales (y también de las consonantes
sonoras, pero para simplificar la explicación en adelante nos
referiremos sólo a las vocales) las cuerdas participan gene-
rando (al abrirse y cerrarse con rapidez al paso de soplos
periódicos de aire) un sonido «base», o tono laríngeo, que es
siempre el mismo, independientemente de la vocal que pro-
nunciemos.
< 10 >
Figura 16.2. Sección de la cabeza de un lactante. La laringe se conecta
con la cavidad nasal y el líquido pasa al tubo digestivo a través de los
senos piriformes. De este modo, es posible beber y respirar al mismo
tiempo. En los humanos adultos la posición baja de la laringe impide que
ésta pueda ponerse en relación directa con la cavidad nasal, por lo que la
respiración ha de ser interrumpida durante la ingestión de líquidos.

< 11 >
El tono laríngeo está formado por una frecuencia principal y
una serie de frecuencias «acompañantes» o armónicos. Si el
conjunto de cavidades situadas por encima de la laringe (fa-
ringe, cavidad nasal y cavidad oral) no tuviera ninguna inter-
vención en la producción de los sonidos del habla humana, el
sonido que oiríamos estaría formado sobre todo por el co-
rrespondiente a la frecuencia principal del tono laríngeo; la
mayor parte de sus armónicos son demasiado débiles como
para percibirlos.
Sin embargo, esto no ocurre así debido al fenómeno conoci-
do como resonancia, según el cual un cuerpo (resonador)
puede producir vibraciones como consecuencia de la vibra-
ción de otro cuerpo cercano. Ésta es la causa de que los cris-
tales de las ventanas vibren (resuenen) con determinados
ruidos del tráfico. Ahora bien, no todos los cuerpos que ac-
túan como resonadores reproducen las mismas vibraciones,
cada tipo de resonador responde ante unas vibraciones de-
terminadas. De manera que si hacemos pasar un sonido com-
puesto de varias frecuencias (como es el caso del tono larín-
geo) a través de un resonador, éste reproducirá, e incluso
ampliará, ciertas frecuencias pero no otras. Como resultado
de este fenómeno, obtendremos un filtrado acústico en el que
algunas frecuencias se habrán amplificado mientras que otras
se habrán extinguido; el sonido resultante será diferente del
original. Según sea el resonador que empleemos, obtendre-
mos sonidos distintos a partir de un mismo sonido base.
Pues bien, el tracto vocal humano puede adoptar diferentes
configuraciones, cada una de las cuales actúa como un reso-
nador distinto que filtra de un modo específico el tono larín-
geo producido en las cuerdas vocales, dando lugar a los dis-
tintos sonidos vocálicos. Para realizar este filtrado deben
producirse estrechamientos y ensanchamientos a determina-
das distancias de la fuente de producción del tono laríngeo,
tanto en la faringe como en la cavidad bucal.
Por ejemplo, para producir el sonido [a], la lengua se aplana
en la cavidad bucal y se desplaza hacia atrás para estrechar la
faringe. Al mismo tiempo, la laringe se eleva para acortar la

< 12 >
distancia entre las cuerdas vocales y las zonas de estrecha-
miento (faringe) y ensanchamiento (cavidad bucal). La situa-
ción contraria se presenta al producir el sonido [i]. Ahora, la
lengua se eleva, estrechando la cavidad bucal, y la faringe se
ensancha. En el caso de la [u], tanto la cavidad bucal como la
faringe se ensanchan, mientras que la parte posterior de la
lengua asciende para producir un estrechamiento en la parte
posterior del paladar.
Para que esta compleja gimnasia articulatoria tenga lugar es
preciso que la cavidad bucal y la faringe puedan actuar como
dos conductos independientes, lo que requiere que la faringe
tenga una cierta longitud y que, además, esté dispuesta en
ángulo respecto de la cavidad bucal. Dicho de otro modo,
hace falta que la laringe esté situada en una posición baja en
el cuello (figura 16.1).
Los sonidos vocálicos formados de este modo son mati-
zados en la cavidad bucal mediante movimientos de la len-
gua, los labios y el paladar blando (donde está la úvula o
campanilla), dando lugar a las consonantes. Para que la len-
gua alcance con rapidez y precisión los lugares adecuados
para la producción de las consonantes es preciso que no sea
demasiado larga. Esto es debido a que algunos de los mús-
culos que mueven la lengua se insertan en el hueso hioides,
situado en la parte inferior y posterior de la misma (figura
16.1). Cuanta más longitud tenga la lengua, más largos serán
estos músculos y más lentos e imprecisos se tornarán sus
movimientos.
Es fácil estimar la longitud de la lengua a partir de un
cráneo fósil, ya que ésta es proporcional a la longitud del
paladar óseo. Sin embargo, deducir la posición de la laringe
es harina de otro costal. Como ya hemos indicado, este ór-
gano está constituido por cartílagos y sostenido por músculos
y ligamentos, ninguno de los cuales fosiliza.

< 13 >
Hablan los fósiles
Desde mediados de la década de los setenta el lingüista Phi-
llip Lieberman y el anatomista J'effrey Laitman han encabe-
zado una serie de investigaciones destinadas a reconstruir la
morfología del tramo superior del conducto respiratorio en
los homínidos fósiles. Como resultado de sus estudios, con-
cluyeron que una serie de rasgos de la base del cráneo po-
dían usarse para averiguar la posición de la laringe en el cue-
llo v así establecer las habilidades fonéticas de los homínidos
fósiles. Entre estas características, la que ha gozado de un
mayor crédito en la comunidad científica ha sido el grado de
flexión de la base del cráneo.
Si seccionásemos un cráneo humano por su plano medio o de
simetría (el que divide al cráneo en dos mitades equivalen-
tes), encontraríamos que el perfil de su borde inferior presen-
ta una marcada inflexión situada entre el foramen magnum y
la parte posterior del paladar. Pues bien, los humanos recién
nacidos y los antropomorfos en general tienen una base del
cráneo poco flexionada. En los humanos la flexión de la base
del cráneo se va acentuando durante la infancia hasta alcan-
zar su máximo en el estado adulto.
Puesto que los recién nacidos humanos y los antropomorfos
comparten una base del cráneo poco flexionada junto con
una posición elevada de la laringe, y dado que en los huma-
nos el proceso de descenso de la laringe es acompañado por
el incremento de la flexión basicraneal, parece existir una
clara relación entre la posición de la laringe y el grado de
flexión basicraneal. Esta relación también se ha observado
mediante la experiencia en ratas a las que se les produjo qui-
rúrgicamente un aumento de la flexión basicraneal.
Con este argumento, Laitman y sus colaboradores han reali-
zado diversos estudios en diferentes homínidos fósiles, lle-
gando a una serie de conclusiones sobre sus aparatos fonado-
res. Según estos investigadores, en los australopitecos, pa-
rántropos y Homo habilis la laringe debió de ocupar una po-

< 14 >
sición elevada y sus capacidades fonéticas hubieron de ser
similares a las de los chimpancés. Por el contrario, hallaron
que los cráneos de Broken Hill y Steinheim (del Pleistoceno
Medio de África y Europa, respectivamente) presentaban
basicráneos flexionados, lo que implicaría laringes bajas y
capacidades fonéticas similares a las nuestras. Respecto de
los neandertales, llegaron a la conclusión de que su aparato
fonador sólo les permitiría articular un limitado repertorio de
vocales (entre las que no se encontrarían la [a], la [i] ni la
[u]), con lo que su lenguaje hablado sería rudimentario y
lento.
Sin embargo, nosotros hemos estudiado la región de la base
del cráneo en los únicos ejemplares de Homo habilis (OH
24) y de Horno ergaster (ER 3733) con el basicráneo bien
conservado, obteniendo unos valores de flexión basicraneal
superiores a los de los australopitecos, chimpancés y gorilas.
Estos resultados sugieren que el aparato fonador del HOMO
habilis y del Homo ergaster ya era parecido al nuestro (si
bien en el Homo habilis el paladar era proporcionalmente tan
largo como el de los chimpancés, lo que indicaría un reperto-
rio de consonantes más menguado), y refuerzan la hipótesis
que liga el origen de nuestro género con el de la palabra;
puesto que si estos primeros humanos no hablaban, es difícil
de explicar, por selección natural, la posición baja de sus
laringes.
Por otra parte, a muchos investigadores se les hace difícil de
aceptar que los neandertales hubieran reducido su capacidad
de hablar desde la condición presente en sus antecesores
(caso de Steinheim).
En respuesta a estas críticas, Laitman ha propuesto que los
neandertales vieron reducidas sus capacidades fonéticas co-
mo consecuencia de una adaptación más importante para su
supervivencia: la adecuación de sus vías aéreas superiores a
la necesidad de calentar y humedecer el frío y seco aire de
las épocas glaciales; respirar es más importante que hablar.
Por otra parte, Cristopher Stringer y Clive Gamble, en su

< 15 >
libro En busca de los neandertales aducen que los ante-
pasados de los neandertales (Steinheim y Petralona) carecían
de un lenguaje hablado como el nuestro, a pesar de tener las
bases anatómicas para producirlo, debido a limitaciones psí-
quicas derivadas de sus relativamente pequeños cerebros.
Sin embargo, la idea de que los neandertales no eran capaces
de hablar como nosotros comenzó a tambalearse cuando, a
mediados de los años ochenta, el paleoantropólogo Jean-
Louis Heim anunció que el cráneo del ejemplar neandertal
conocido como «el Viejo», de La Chapelle-aux-Saints, esta-
ba mal reconstruido por los primeros investigadores y que la
nueva reconstrucción realizada por él mostraba un mayor
grado de flexión basicraneal. Extremo éste que fue confirma-
do por David Frayer, quien midió la flexión de la nueva re-
construcción del ejemplar de La Chapelle-aux-Saints y en-
contró que era similar a la de una serie de cráneos medieva-
les. Puesto que este fósil era uno de los estudiados por Lait-
man, sus resultados fueron puestos en entredicho.
Por otra parte, en 1989 se halló en el yacimiento israelí de
Kebara un hueso hioides perteneciente a un ejemplar nean-
dertal, que es el único publicado de un homínido fósil. Como
ya hemos comentado, el hueso hioides presta inserción a la
musculatura de la lengua y su posición en el cuello está muy
relacionada con la propia de la laringe. El hioides de Kebara
presenta una morfología y dimensiones comparables con las
del hueso hioides de cualquiera de nosotros, lo que llevó al
equipo de científicos que lo estudiaron, encabezados por
Baruch Arensburg, a concluir que los neandertales eran ana-
tómicamente tan capaces de hablar como los humanos mo-
dernos. Esta afirmación ha sido contestada por Lieberman y
Laitman aduciendo que la morfología del hueso hioides no
es un dato relevante para establecer las capacidades fonéticas
de los homínidos. Desgraciadamente, en Kebara no se ha
hallado ningún cráneo, por lo que no es posible comparar la
morfología del hueso hioides con el grado de flexión basi-
craneal.
Lo cierto es que las investigaciones dedicadas a la recons-

< 16 >
trucción del aparato fonador de los neandertales se encuen-
tran en un punto muerto. Para unos, los estudios realizados
sobre la flexión basicraneal carecen de validez porque se
llevaron a cabo sobre ejemplares mal reconstruidos y prefie-
ren conceder crédito a los resultados del análisis del hioides
de Kebara. Otros niegan el valor de estos estudios y siguen
manteniendo las conclusiones alcanzadas por los análisis de
la flexión basicraneal de los neandertales.
La única manera de romper esta situación es la de encontrar
nuevo material fósil que incluya tanto basicráneos intactos
como huesos hioides. Podría parecer que un hallazgo de esta
naturaleza es prácticamente imposible, puesto que debe con-
jugar dos hechos muy improbables como son el hallazgo de
un cráneo intacto y el de un hueso hioides (sólo se conoce
uno en todo el registro fósil de los homínidos, el de Kebara).
Sin embargo, tal descubrimiento ha tenido lugar en fechas
recientes en el yacimiento de la Sima de los Huesos de la
Sierra de Atapuerca, donde hemos encontrado un cráneo con
su base prácticamente completa, el Cráneo 5 (figura 12.2),
así como la mayor parte de dos huesos hioides. Habrá que
esperar a que se culminen las investigaciones en curso sobre
este extraordinario material fósil para conocer más datos
sobre el origen del habla humana.
La selección de grupo y la extinción de los neandertales En el mundo en que vivimos estamos familiarizados con la
idea de que la comunicación y la información son la clave
del progreso y la base de nuestro actual desarrollo tecnológi-
co. Por eso la posesión del lenguaje articulado parece que
nos da una indiscutible superioridad sobre el resto de los
seres vivos en la lucha por la existencia. Pero si se reflexiona
un poco es fácil comprender que la habilidad lingüística no
le aprovecha de nada a un humano solitario enfrentado a la

< 17 >
naturaleza con sus únicos medios, y que esta característica
sólo tiene sentido en el interior del grupo al que necesaria-
mente pertenece. El lenguaje no es una propiedad del indivi-
duo, sino de la colectividad. No es que un individuo se co-
munique bien; sería mejor decir que un grupo está bien co-
municado. La capacidad para compartir y transmitir infor-
mación entre individuos y entre generaciones por la vía del
lenguaje confiere una gran ventaja adaptativa al grupo en su
conjunto, no al individuo aislado.
Esto que nos parece obvio plantea un arduo problema en
biología evolutiva, porque la teoría de la evolución por me-
dio de la selección natural, la teoría de Darwin aceptada uni-
versalmente con más o menos matices por todos los científi-
cos, se basa en la competencia entre individuos. Ya hemos
comentado que algunos autores han planteado una versión
reduccionista de la selección natural situándola a un nivel
inferior al del individuo, como es el nivel de los genes. En
las páginas de este libro hemos visto que la competencia se
puede en ocasiones situar al nivel de los espermatozoides,
también por debajo del nivel individual. El propio Darwin
propuso el mecanismo de la selección sexual para explicar
determinadas características de los individuos que no se pue-
den entender desde el punto de vista de la simple competen-
cia por los recursos del medio y la lucha por la superviven-
cia; a fin de cuentas de lo que se trata es de vivir para repro-
ducirse.
Sin embargo, la aparición de propiedades que caracterizan a
los grupos, como la comunicación, plantea el problema de la
selección natural a un nivel supraindividual, el nivel de gru-
po. Así, el lenguaje habría sido seleccionado porque los gru-
pos con un mayor nivel de comunicación interna eran más
competitivos, más eficaces en la explotación de los recursos
del medio y desplazaban a otros grupos. O en otras palabras,
un individuo con capacidad para producir y entender el len-
guaje articulado puede no ser más competitivo que otro indi-
viduo que no tiene esa facultad, pero un grupo con lenguaje
lo es más que un grupo que no lo tiene.

< 18 >
Que existe competencia entre grupos de la misma especie es
algo que se conoce desde antiguo para los mamíferos socia-
les, primates incluidos. Los chimpancés sin ir más lejos de-
fienden sus territorios y a veces invaden los ajenos o son
invadidos por los grupos vecinos. No sólo la comunicación
serviría para dar cohesión y más eficacia a los grupos (“la
información es poder” se dice en nuestra sociedad), sino que
los comportamientos de cooperación social («altruismo so-
cial») dentro del grupo serían importantísimos en la selec-
ción entre grupos. La cooperación social se extiende a mu-
chos campos, como la defensa del territorio o de los recur-
sos, la defensa contra los depredadores, la caza en grupo,
prestar cuidados a crías ajenas, compartir comida...
Ahora bien, si la competencia entre grupos es fácil de enten-
der, la cosa se complica a la hora de formular un modelo
realista de selección de grupos. Porque, inevitablemente,
tenemos que volver a hablar de genes. Partiendo de la base
de que el comportamiento cooperativo y la capacidad, men-
tal y fonética, para el lenguaje tienen una base genética, po-
demos acotar el problema considerando la existencia de un
gen para el comportamiento cooperativo y un gen para el
lenguaje (esperamos que el lector escrupuloso sepa perdonar
esta simplificación de las bases genéticas de la conducta).
Para que la selección entre grupos pueda producirse es preci-
so que dentro de cada grupo haya mucha homogeneidad ge-
nética, o sea, mucha consanguinidad. Dicho más técnicamen-
te, es necesario que la variabilidad dentro de cada grupo sea
mucho menor que la variabilidad entre los diferentes grupos.
Los grupos con elevadas frecuencias de los genes para el
lenguaje y para la cooperación serán más competitivos que
los demás; como la cooperación y la comunicación no se
rigen por la ley del todo o la nada, en realidad habría que
hablar de mayor o menor capacidad para lo uno y lo otro. En
el caso de la cooperación social existe el problema adicional
de que los individuos «egoístas» se beneficiarían del esfuer-
zo de los demás sin hacer ningún gasto, y por tanto podrían
resultar beneficiados por la selección natural y legar más

< 19 >
genes a la siguiente generación, con lo que el comportamien-
to «altruista» no podría llegar a imponerse nunca.
Una solución es que existan mecanismos de rechazo social
para impedir que los «egoístas» se reproduzcan, pero enton-
ces habría que preguntarse cómo fueron seleccionados estos
mecanismos, con lo que volvemos al principio del problema.
Los chimpancés forman grupos sociales en los que los ma-
chos están emparentados, pero no las hembras, que van de un
grupo a otro. Algo parecido, como vimos, puede imaginarse
para los primeros homínidos. Es decir, que el grupo social no
es un grupo reproductor cerrado, con lo que se pierde uni-
formidad genética al importarse hembras (genes en definiti-
va) de otros grupos. Así y todo se pueden formular modelos
matemáticos que hacen viable la selección al nivel de grupo
en determinadas condiciones. Pero también es posible ima-
ginar que nuestros antepasados llegaron a formar unidades
sociales mayores, que funcionaban a la vez como unidades
reproductoras y competían entre sí. En todo caso, no tene-
mos hoy una explicación mejor que la selección de grupo
para entender una de las más importantes de nuestras carac-
terísticas: el lenguaje articulado.
Otra cuestión en la que la selección en el ámbito de grupo
parece ser la única explicación viable es la del reemplaza-
miento de los neandertales por los humanos modernos. No
parece razonable pensar que dicha sustitución se produjera
por selección en el ámbito individual. Como ya hemos tenido
ocasión de comentar, los neandertales eran más fuertes físi-
camente y su anatomía estaba mejor adaptada al clima euro-
peo (figura 13.1), por lo que cabe pensar que eran individual-
mente tan aptos como los humanos modernos (si no más).
Casi con toda seguridad, el éxito de nuestros antepasados
radicó en alguna propiedad de grupo, pero ¿cuál?
Para un buen número de autores, la ausencia de un auténtico
lenguaje en los neandertales fue una de las principales causas
(tal vez la principal) de su sustitución por los humanos mo-
dernos. Sencillamente, el rudimentario lenguaje de los nean-

< 20 >
dertales limitaba su complejidad social, restringiendo su ca-
pacidad de transmitir información esencial para explotar los
recursos del medio. Y cuando los humanos modernos apare-
cieron en el horizonte, pertrechados con su elaborado len-
guaje, los neandertales fueron abocados a la extinción.
Aunque ésta de la superioridad lingüística de los humanos
modernos es una hipótesis atractiva, ya hemos visto que no
acaba de encajar con los fósiles. Aparte de que no esté claro
el tipo de lenguaje que tuvieron los neandertales, ya hemos
comentado que éstos no fueron «arrollados» por los humanos
modernos en un proceso rápido y universal, sino que la susti-
tución de unos por otros duró cerca de 10.000 años. Si los
humanos modernos tenían una ventaja tan abrumadora en
cuanto a complejidad social y explotación del medio, ¿por
qué tardaron tanto en reemplazar a los neandertales?
Nuestra opinión al respecto es que lo que permitió a nuestros
antepasados desplazar a los neandertales no fue la presencia
de una ventaja cualitativa, del tipo del lenguaje, sino más
bien de un mayor desarrollo de sus capacidades para explotar
los recursos; sencillamente, tenían más de lo mismo.
Los renglones torcidos de la selección natural La relación que mantuvieron entre sí los dos descubridores
de la selección natural fue ejemplar. Entre Charles Darwin y
Alfred Russel Wallace nunca hubo lugar para los celos pro-
fesionales, sino que imperó entre ellos el respeto mutuo y
una cálida amistad. Cuando el 26 de abril de 1882 la socie-
dad británica quiso reconocer la importancia del trabajo de
Darwin enterrándolo en la abadía de Westminster, al lado de
la tumba de sir Isaac Newton, Wallace estuvo entre los tres
amigos íntimos que, junto a miembros de la nobleza y auto-
ridades, transportaron el féretro de Darwin.
Sin embargo, y a pesar de su buena amistad y consideración
mutua, Darwin y Wallace mantuvieron puntos de vista con-
trapuestos acerca del origen de algunas de las características

< 21 >
más relevantes de nuestra especie, tales como la inteligencia
y el habla. Mientras Darwin contemplaba dichos rasgos co-
mo un resultado más del proceso evolutivo regido por la se-
lección natural, Wallace atribuía su origen a causas sobrena-
turales, colocando así el origen del ser humano más allá de la
acción de la selección natural.
Uno de los argumentos más profundos empleados por Dar-
win para probar la existencia del hecho evolutivo fue el de la
existencia de «chapuzas» en los seres vivos. Si los organis-
mos fueran el resultado de un acto directo de creación divi-
na, sus distintas partes deberían mostrarse «como recién sa-
lidas de fábrica»; es decir, que deben ser diseños específicos
para cumplir de manera eficaz una función determinada. Lo
que no esperaríamos encontrar, desde luego, son órganos que
aparezcan como una modificación, más o menos afortunada,
de otros que cumplen una función distinta en organismos
diferentes. Dicho de otro modo, lo esperable es que cada ser
vivo tenga sus propias piezas, perfectamente ajustadas al
desempeño de las funciones que tiene encomendadas.
La selección natural no planifica el cambio evolutivo, sim-
plemente elige entre lo que hay. Es decir, que preserva aque-
llas variaciones de los órganos existentes que confieren al-
guna ventaja a los individuos. De este modo, un órgano pue-
de verse modificado y acabar desempeñando una función
distinta de la que tenía. En este proceso, puede ocurrir que el
órgano en cuestión pierda eficacia en el desempeño de la
misión original, siempre y cuando esta pérdida se vea com-
pensada por la ventaja conferida por la nueva función.
Pero volvamos al tema de la voz humana. Cuando Wallace y
Darwin disputaban sobre la naturaleza de la selección natural
y su papel en el origen de los seres humanos, no se conocía
la base anatómica ni los mecanismos fisiológicos del habla.
Hoy comprendemos que esta cualidad humana está basada
en la posición baja de nuestra laringe, que a su vez es debida
a una modificación del modelo de vías respiratorias superio-
res que es común en el resto de los mamíferos. También sa-
bemos que esta peculiar posición de la laringe restringe

< 22 >
nuestra capacidad de tragar líquidos y respirar al mismo
tiempo, y que es la responsable de un fenómeno tan desagra-
dable, y peligroso, como es el atragantamiento. Sin embargo,
estos inconvenientes los consideramos como minucias al
lado de la gran ventaja que supone el disponer de un meca-
nismo que nos permite modular los sonidos que están en la
base de nuestro lenguaje; sin duda, el saldo es muy ventajo-
so. De este modo, en la anatomía de nuestro aparato fonador
puede reconocerse la huella de la selección natural y el rastro
de la historia evolutiva de nuestra especie.
Darwin puede descansar tranquilo al lado de Newton; una
vez más, tenía razón.
Recommended

















