
INSTITUTO POLITÉCNICO NACIONAL
ESCUELA SUPERIOR DE MEDICINA SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN
Análisis isobolográfico del efecto analgésico de la combinación gabapentina-metamizol administrada por vía oral y e spinal
en modelos de dolor inflamatorio y neuropático
TESIS
QUE PARA OBTENER EL GRADO DE DOCTOR EN INVESTIGACIÓN EN MEDICINA
PRESENTA
M. en C. Luis Fernando Ortega Varela
ASESORES: DR. VINICIO GRANADOS SOTO DR. JORGE E. HERRERA ABARCA
México, DF, marzo de 2008



Las series experimentales del presente trabajo se realizaron en el
laboratorio 12 del Departamento de Farmacobiología del Cinvestav, Sede Sur a
cargo del Dr. Vinicio Granados Soto. Algunos estudios piloto fueron efectuados
previamente en el laboratorio de Farmacología del Instituto de Investigaciones
Qímico-Biológicas de la UMSNH.
Durante el desarrollo de esta investigación se contó con el apoyo
otorgado por el CONACYT mediante la beca con número de registro 172076.

AGRADECIMIENTOS
Al Dr. Vinicio Granados Soto porque su política de “equal opportunity job
offer” me permitió darle un vistazo a las grandes ligas de la investigación.
También por convertir hasta los jueves pozoleros en cátedra sobre los
receptores TRPV.
Al Dr. Jorge E. Herrera por desempolvar para mí su “cachucha básica” y
porque sigue abriendo puertas que me parecían cerradas.
A los Doctores Gerardo Reyes, Javier Mancilla y Guillermo Ceballos
porque sus comentarios enriquecieron este trabajo y mis perspectivas.
A mis compañeros del Laboratorio 12 por todo lo aprendido: A Teremix
por chutarse mis borradores, a Nadia por las cirugías con Chava Flores de
fondo, a Deysi por su lema, al Rocha por su excelente manejo de la
herramienta word art y por los ciberconsejos, a Lupita, Ivonne, Mónica,
Rosaura, Luis Joaquín, Carlos, Jair y Adrián. Todavía espero que mañana se
les olvide.
A Oliva por ser superviviente en un programa al que inicialmente se
presentaron 16 morelianos.
A los Doctores Héctor Urquiza, Francisco Javier Larios, Francisco Javier
Flores Murrieta y Víctor Manuel Farías Rodríguez por su genuino interés en
este trabajo experimental.
A Tere Fraga, Madeline y Efraín por su encomiable ayuda en los pilotos
en el IIQB.
A todo el personal del Cinvestav Sede Sur, en especial a los adscritos al
bioterio, por ser tan amables con este polizón.

Sumo y sigo. A Irma, Vicky, Angélica y Edgardo por solapar mis
ausencias laborales en el laboratorio de química.
Al consejo técnico de la Preparatoria Melchor Ocampo de la UMSNH,
especialmente a Damián Arévalo, por encontrar un resquicio legal contra el
portazo de la burocracia.
A mis Alumnos, por su enorme disposición y por cargar con mis
neurosis.
A Mónica y Cristina por no perderme la pista, a Judith por los cafés
motivacionales. A Erasto por el tráfico de artículos. A Héctor y Ulises, por las
odiseas.
A la presidenta de la Fundación Bety Varela, y a sus dos bellísimas
vicepresidentas, por haberme otorgado una beca que incluyó hospedaje y
cariño.
A los Migues por prescribir inhibidores de proteasa cibernéticos a mis
archivos computacionales.
A Ricardo Ortega e Irma Varela porque nunca han cesado de decirme
“tihuí-tihuí”.
A toda mi familia, sí a toda!, por ser para conmigo más entusiastas que
la porra de doña Cholita.

“El dolor es el aguijón que nos previene del peligro”
H. G. Wells (en la isla del Dr. Moreau)

I
ÍNDICE
Página
ABREVIATURAS IV
LISTA DE FIGURAS Y TABLAS VIII
RESUMEN XI
ABSTRACT XlI
1. INTRODUCCIÓN 1
1.1. Dolor 1
1.1.1. Clasificación del dolor 2
1.1.2. Nocicepción 4
1.1.3. Sensibilización periférica 7
1.1.4. Sensibilización central 9
1.1.5. Inflamación 13
1.1.5.1. Dolor inflamatorio 13
1.1.5.1.1. Mediadores pronociceptivos 14
1.1.5.1.1.1. Protones 14
1.1.5.1.1.2. Cininas 15
1.1.5.1.1.3. Prostanoides 17
1.1.5.1.1.4. Adenosín trifosfato 18
1.1.5.1.1.5. Serotonina 19
1.1.5.1.1.6. Histamina 21
1.1.5.1.1.7. Neurocininas 21
1.1.5.1.1.8 Especies reactivas de oxígeno 22
1.1.5.1.1.8.1. Óxido nítrico 23
1.1.5.1.1.9 Mediadores inflamatorios de
las células inmunes 23
1.1.5.1.2. Modificaciones transcripcionales 24
1.1.5.2. Tratamiento del dolor inflamatorio 26
1.1.6. Dolor neuropático 27
1.1.6.1. Mecanismos del dolor neuropático 28
1.1.6.1.1. Aumento de descargas aferentes 28
1.1.6.1.2. Facilitación descendente 29
1.1.6.1.3. Aumento de dinorfina espinal 30
1.1.6.1.4. Ramificación colateral 30
1.1.6.1.5. Participación de factores neutróficos 31

II
1.1.6.2. Tratamiento del dolor neuropático 32
1.2. Combinaciones analgésicas 34
1.2.1. Análisis isobolográfico 35
1.3. Gabapentina 37
1.3.1. Mecanismo de acción de gabapentina 38
1.4. Metamizol 40
1.4.1. Metabolitos del metamizol 41
1.4.2. Empleo del metamizol 43
1.4.3. Efectos adversos 43
1.4.4. Mecanismo de acción de metamizol 44
2. PLANTEAMIENTO DEL PROBLEMA 47
3. HIPÓTESIS 49
4. OBJETIVOS 50
4.1. Objetivo general 50
4.1.1. Objetivos particulares 50
5. MATERIAL Y MÉTODOS 51
5.1. Animales de experimentación 51
5.2. Fármacos 51
5.3. Modelo inflamatorio 51
5.3.1. Prueba de la formalina 51
5.4. Modelo de neuropatía 54
5.4.1. Prueba de alodinia táctil 55
5.5. Cirugía espinal 57
5.6. Diseño del estudio 58
5.6.1. Eficacia individual y de la combinación 58
5.7. Mecanismo de acción 60
5.8. Análisis de datos 61
5.8.1. Análisis de los datos de formalina 61
5.8.2. Análisis de los datos de alodinia táctil 62
5.8.3. Análisis isobolográfico 63
5.8.4. Análisis estadístico 65
6. RESULTADOS 66
6.1. Modelo inflamatorio 66
6.1.1. Efecto de gabapentina, metamizol y su combinación
por vía oral en el modelo de formalina 66

III
6.1.2. Efecto espinal de gabapentina, metamizol y
la combinación en la prueba de la formalina 74
6.2. Modelo neuropático 80
6.2.1. Efecto antialodínico de gabapentina, metamizol y
la combinación por vía oral 81
6.2.2. Efecto antialodínico de Gabapentina, metamizol y
la combinación por vía espinal 83
6.3. Mecanismo de acción 86
6.3.1. Administración de naltrexona 88
6.3.2. Administración de L-NAME 88
6.3.3. Administración de glibenclamida 89
7. DISCUSIÓN 91
7.1 Modelo inflamatorio 91
7.1.1. Efecto antinociceptivo de gabapentina 91
7.1.2. Efecto antinociceptivo de metamizol 91
7.1.3. Efecto antinociceptivo de la combinación
de gabapentina y metamizol 92
7.2. Modelo neuropático 93
7.2.1. Efecto antialodínico de gabapentina 93
7.2.2. Efecto antialodínico de metamizol 94
7.2.3. Efecto antialodínico de la combinación
de gabapentina y metamizol 94
7.3. Mecanismo de acción 95
7.3.1. Mecanismos de acción de gabapentina 95
7.3.2. Mecanismos de acción de metamizol 96
7.3.3. Combinaciones de gabapentina y metamizol 98
7.3.4. Mecanismo de la combinación de gabapentina
y metamizol 99
8. CONCLUSIONES 101
9. BIBLIOGRAFÍA 102
ANEXOS XIII

IV
ABREVIATURAS
A2
Receptor 2 de adenosina
AA Amino antipirina
AAAP
4-acetil amino antipirina
AMPc
Adenosín monofosfato cíclico
ATP
Adenosín trifosfato
AINEs
Analgésicos anti-inflamatorios no esteroideos
AMPA
Ácido 2-amino 4-hidroximetil isooxazol propiónico
ANOVA
Análisis de varianza
ASICs
Canales de sodio sensibles a ácido
B1
Receptor uno de bradicinina
B2
Receptor dos de bradicinina
BDNF
Factor neurotrófico derivado del cerebro
Ca+2
Calcio
CCK
Colecistocinina
CGRP
Gen relacionado con el gen de la calcitonina
CI50
Concentración inhibitoria 50
Cox
Ciclooxigenasa
Cox 1
Ciclooxigenasa 1
Cox 2
Ciclooxigenasa 2
Cox 3 (Cox 1B)
Ciclooxigenasa 3
CCDV Canales de calcio dependientes de voltaje
CDR Curva dosis respuesta
DAG
Diacil glicerol
DE
Dosis efectiva

V
ABREVIATURAS
DE30
Dosis efectiva 30
DE50
Dosis efectiva 50
DLF
Funículo dorsolateral
DMSO Dimetil sulfóxido
E
Experimental
EP
Receptor de prostaglandina
FAAP
4-formil amino antipirina
g
Gramos
GABA
Ácido gamma amino butírico
Gbp
Gabapentina
Gli
Glibenclamida
H+
Protones
IAAS Estudio internacional de agranulocitosis y anemia aplástica
IASP Asociación internacional para el estudio del dolor
IL-1
Interleucina 1
IL-6
Interleucina 6
IL-8 Interleucina 8
IP
Receptor de prostaciclina
IP3
Inositol trifosfato
It
Vía intratecal
K+
Potasio
K+ATP
Canales de potasio sensibles a ATP
kg
Kilogramos
L-NAME
Metil éster de la L-nitro arginina
L-NOARG L-nitroarginina

VI
ABREVIATURAS LTB4
Leucotrieno B4
MAA
4 metil amino antipirina
Met
Metamizol
Mg+2
Magnesio
mg
Miligramos
Min
Minutos
ml
Mililitros
Na+
Sodio
NLT
Naltrexona
NK1
Receptor de neurocininas 1
NK2
Receptor de neurocininas 2
NK3
Receptor de neurocininas 3
NKA
Neurocinina A
OMS (WHO)
Organización Mundial de la Salud
ON
Óxido nítrico
P2x3
Receptor purinérgico 2x3
PGE2
Prostaglandina E2
PKC
Proteína cinasa C
po
Vía oral
pH Potencial de hidrógeno
RNAm Ácido ribonucléico mensajero
RVM
Médula rostroventromedial
SHAM
Cirugía de maniobras simuladas

VII
ABREVIATURAS
SNRI
Inhibidores de la recaptura de noradrenalina y serotonina
SNS
Canal de sodio neuronal sensorial
SNS-2
Canal de sodio neuronal sensorial 2
SP
Sustancia P
T
Teórico
TCA
Antidepresivos tricíclicos
TNFα
Factor de necrosis tumoral alfa
TrkA
Receptor tirocina cinasa A del factor de crecimiento neuronal
TRPV1
Receptor de potencial transitorio V1
Veh
Vehículo
γ
Índice de interacción
µg
microgramos
µl
microlitros
5-HT1
Receptor 1 de serotonina
5-HT2
Receptor 2 de serotonina
5-HT3
Receptor 3 de serotonina
5-HT1D
Receptor 1d de serotonina
5-HT7
Receptor 7 de serotonina
%EMP
Porcentaje de efecto máximo posible

VIII
FIGURAS Y TABLAS
FIGURAS
Página
Figura 1 Visión de descartes sobre el dolor
1
Figura 2 Tipos de dolor 5
Figura 3 Nocicepción
6
Figura 4 Sensibilización periférica 10
Figura 5 Sensibilización central
12
Figura 6 Dolor inflamatorio
26
Figura 7 Tratamiento del dolor inflamatorio
27
Figura 8 Mecanismos celulares de dolor neuropático
32
Figura 9 Tratamiento del dolor neuropático
34
Figura 10 Isobolograma
37
Figura 11 Estructura de GABA y gabapentina
38
Figura 12 Fórmula del metamizol 41
Figura 13 Metabolitos del metamizol
42
Figura 14 Prueba de la formalina 52
Figura 15 Secuencia de los experimentos en formalina
53
Figura 16 Curso temporal de formalina 53
Figura 17 Cirugía de Kim y Chung
54
Figura 18 Descripción de la inducción neuropática 55
Figura 19 Prueba de alodinia táctil 56
Figura 20 Secuencia de la prueba de alodinia táctil 56
Figura 21 Cirugía espinal 57
Figura 22 Grupos experimentales en el modelo inflamatorio 59
Figura 23 Grupos experimentales en el modelo neuropático 59
Figura 24 Secuencia de la prueba de alodinia táctil en presencia de inhibidores 60

IX
FIGURAS
Página
Figura 25 Grupos experimentales en presencia de inhibidores 61
Figura 26 Análisis isobolográfico
64
Figura 27 Cursos temporales en formalina 67
Figura 28 Curvas dosis respuesta para gabapentina oral en formalina 69
Figura 29 Curvas dosis respuesta para metamizol oral en formalina 70
Figura 30 Curva dosis respuesta para la combinación oral en formalina 72
Figura 31 Isobolograma de la interacción antinociceptiva de gabapentina y metamizol por vía oral
73
Figura 32 Curvas dosis respuesta para gabapentina espinal en formalina 75
Figura 33 Curvas dosis respuesta para metamizol espinal en formalina 76
Figura 34 Curvas dosis respuesta para la combinación espinal en formalina 78
Figura 35 Isobolograma de la interacción antinociceptiva de gabapentina y metamizol por vía espinal
79
Figura 36 Curso temporal de la prueba de alodinia táctil 80
Figura 37 Curva dosis respuesta del efecto antialodínico de metamizol, gabapentina y la combinación por vía oral
82
Figura 38 Curva dosis respuesta del efecto antialodínico de gabapentina espinal 83
Figura 39 Curva dosis respuesta del efecto antialodínico de metamizol espinal 85
Figura 40 Curva dosis respuesta del efecto antialodínico de la combinación espinal 86
Figura 41 Isobolograma de la interacción antialodínica de gabapentina y metamizol por vía espinal
87
Figura 42 Administración de naltrexona 88
Figura 43 Administración de L-NAME 89
Figura 44 Administración de glibenclamida 90

X
TABLAS
Página
Tabla 1 Clasificación del dolor 3
Tabla 2 Combinaciones de gabapentina
40
Tabla 3 Combinaciones de metamizol
47
Tabla 4 Combinaciones para el isobolograma por vía oral en la prueba de formalina
71
Tabla 5 Combinaciones para el isobolograma por vía espinal en la prueba de formalina
77
Tabla 6 Combinaciones para el isobolograma por vía espinal en la prueba de alodinia táctil
85

XI
RESUMEN
Recientemente se ha extendido el empleo de la terapia de combinación en un intento de mejorar los tratamientos contra el dolor. La mezcla de dos agentes analgésicos se llega a utilizar con la intención de obtener un mayor efecto al tiempo que disminuye la presencia de efectos no deseados. Con el propósito de analizar, mediante el análisis isobolográfico, la interacción de gabapentina y metamizol en un modelo de dolor inflamatorio (prueba de la formalina) y otro de dolor neuropático (alodinia inducida por el modelo de Kim y Chung), se utilizaron grupos de 6 animales para evaluar los efectos antinociceptivos y antialodínicos de la gabapentina, el metamizol y su combinación por vía oral y espinal. Además se evaluaron algunos de los posibles mecanismos de acción de la combinación. En el modelo inflamatorio, la gabapentina, el metamizol y la combinación tuvieron un efecto antinociceptivo dependiente de la dosis en la fase II de la prueba de la formalina al administrarse por ambas vías de administración. Se determinaron las DE30 para cada fármaco y se construyeron los isobologramas. Los valores de la DE30 teórica para la combinación fueron de 104.6 ± 5.5 µg y 78.8 ± 5.5 mg/kg para las vías espinal y oral respectivamente, que fueron significativamente distintos a los obtenidos de manera experimental (36.8 ± 3.1 µg y 15 ± 1.2 mg/kg) indicando una interacción sinérgica. La dosis de la combinación oral fue 5 veces menor a lo teóricamente calculado, mientras que por vía espinal se requirió un tercio de la DE30 teórica. En el modelo neuropático, tanto la gabapentina como la combinación mostraron un efecto antialodínico dependiente de la dosis por vía oral y espinal. El metamizol no tuvo eficacia por vía oral, al igual que la mayoría de los AINEs. Sin embargo, presentó un efecto significativo por vía intratecal. La DE30 teórica de la combinación fue de 118.4 ± 12 µg, mientras que la DE30 experimental tuvo un valor de 66.2 ± 10.1 µg indicando que la interacción resulta supra-aditiva. Para evaluar el efecto antialodínico de la combinación oral se administró una dosis alta de metamizol con dosis crecientes de gabapentina, registrándose la presencia de sinergismo a dosis altas de gabapentina. Para evaluar el posible mecanismo de acción de la combinación de gabapentina y metamizol, ésta se evaluó con el antagonista opioide naltrexona, el inhibidor de la sintasa de óxido nítrico L-NAME y el bloqueador de los canales de potasio sensibles a ATP glibenclamida. El pretratamiento con los 3 inhibidores redujo de manera signifcativa el efecto de la combinacion, implicando al menos la participación de los receptores opioides, la vía del óxido nítrico-GMPc y los canales de potasio sensibles a ATP en el efecto antinociceptivo y antialodínico de la combinación. Los datos sugieren que dosis bajas de gabapentina y metamizol presentan una interacción sinérgica en la reducción de las conductas nociceptivas y alodínicas, sugiriendo que el empleo de la combinación puede ser útil en el tratamiento del dolor inflamatorio y neuropático en humanos.

XII
ABSTRACT
Currently the combination therapy is commonly used in attempt to improve pain management. A mixture of two analgesic agents is often employed to produce greater analgesic effects with the minor appearance of side effects. This study was designed to evaluate the possible antinociceptive and antiallodynic interaction between gabapentin and metamizol on formalin-induced nociception and spinal nerve ligation model of tactile allodynia (Kim and Chung model). Gabapentin, metamizol or a fixed ratio combination of both drugs were assessed after intrathecal and oral administration in rats. Isobolographic analyses were employed to define the nature of the interaction between drugs. In the inflammatory model, gabapentin, metamizol and the gabapentin-metamizol combination yielded a dose-dependent antinociceptive effect in the phase II of the formalin test when administered by the two different routes. ED30 values were estimated for the individual drugs and isobolograms were constructed. Theoretical ED30 values for the combination estimated from the isobolograms were 104.6 ± 5.5 µg and 78.8 ± 5.5 mg/kg for the intrathecal and oral administration routes, respectively. These values were significantly higher than the experimentally obtained ED30 values (36.8 ± 3.1 µg and 15 ± 1.2 mg/kg) indicating a synergistic interaction. Systemic administration resulted in the highest synergism, being about fivefold, while spinal route increased about threefold, respectively. In the neuropathic model, tactile allodynia was evaluated with calibrated von Frey filaments. The percent of maximum possible effect was deteminated and gabapentin and gabapentin-metamizol combinations showed a dose dependent antiallodynic effect after oral and intrathecal administration. Metamizol had no efficacy by the oral administration route, as most NSAIDs do, but surprisingly it yielded significant effect by spinal route. Theoretical ED30 for the spinal combination estimated from the isobologram was 118.4 ± 12 µg whereas that experimental ED30 value was 66.2 ± 10.1 µg indicating a supra-additive interaction. A single high dose of metamizol was co-administrated with increasing doses of gabapentin to evaluate their antiallodynic effect. The orally administered combination showed a synergistic interaction at the highest gabapentin doses tested. To asses the possible mechanism of action of the gabapentin-metamizol combination, groups were pretreated with the opioid antagonist naltrexone, the NOS inhibitor L-NAME and the K+
ATP channel blocker glibenclamide. The spinal antiallodynic combination effect was abolished with the pretreatment of all inhibitors, indicating the participation of at least opioid receptors, ON-GMPc pathway and K+
ATP channels in the antinociceptive and antiallodynic effect of the combination. Data confirm that low doses of the gabapentin and metamizol can interact synergistically to reduce nociceptive and allodynic behaviors suggesting that this combination could be useful to treat inflammatory and neuropathic pain in humans.

1
1. INTRODUCCIÓN
1.1. El dolor
El dolor tiene un papel destacado entre los sistemas de “vigilancia” de la
homeostasis del organismo ya que despierta y retiene nuestra atención en el punto
exacto en donde se está desarrollando un evento que represente un riesgo para los
individuos (Wall, 1999). Conforme crecemos, las reacciones clásicas de remover el
estímulo lesivo, adoptar una postura que evite nuevas lesiones y optimice la
recuperación y buscar seguridad, alivio y cura se van haciendo más sutiles con la
experiencia, pero también más elaboradas y sofisticadas. Así pues, el dolor tiene la
función básica y fundamental de “dar aviso” sobre amenazas a nuestra
supervivencia y/o bienestar (Figura 1), pero clínicamente se convierte en un
síntoma que nos permite detectar e incluso determinar el grado de evolución de
diferentes enfermedades.
Figura 1. Visión de Descartes sobre el dolor, de su “tratado del hombre” (Treatise De l’homme, Descartes, 1644), en la que describe partes de la piel que al recibir un estímulo nocivo se comportan como “el extremo de una cuerda que acciona una campana” alertando al cerebro sobre el daño. Tomado de Brooks y Tracey (2005).

2
El concepto clásico del dolor fue establecido por el comité de taxonomía de
la asociación internacional para el estudio del dolor (IASP). Ésta lo define como
una experiencia sensorial y emocional desagradable asociada a un daño
tisular real o potencial o que puede ser descrito e n términos de la magnitud
del daño (IASP, 1986). Esta definición se considera la más adecuada ya que en
ella se indica que el dolor no es sólo una respuesta sensorial al daño tisular sino
que puede producirse sin causa somática que lo justifique. Al mismo tiempo está
integrado por un componente emocional subjetivo que lo establece como una
experiencia individual que involucra factores ambientales, culturales, sociales,
espirituales y psicológicos (Anand y Craig, 1996). De esta forma, cada individuo
aprende del dolor a través de experiencias relacionadas a lesiones sufridas durante
su desarrollo y al igual que el tacto, el gusto, la visión y audición es una sensación
que no requiere ser aprendida para que pueda ocurrir (Merskey, 1991).
1.1.1. Clasificación del dolor
El dolor se ha clasificado de varias maneras, debido a que puede variar en
intensidad (leve, moderado o severo), cualidad (urente, lancinante, punzante o
quemante), duración (transitorio, intermitente o persistente) o referencia (superficial
o profundo, localizado o difuso) (Dagnino, 1994). Sin embargo, una clasificación
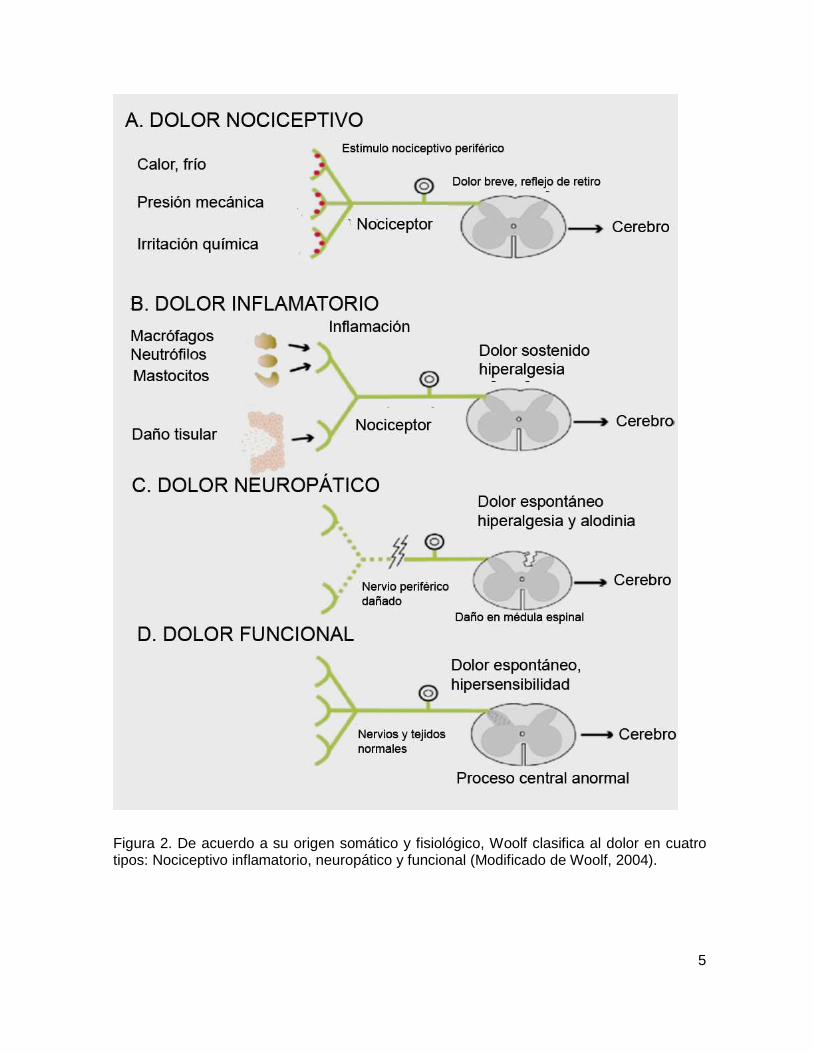
aceptada es la establecida por Woolf (2004) puesto que considera que existen 4
tipos de dolor (Tabla 1) de acuerdo a su origen somático y mecanismo fisiológico;
nociceptivo, inflamatorio, neuropático y funcional (Figura 2).
3
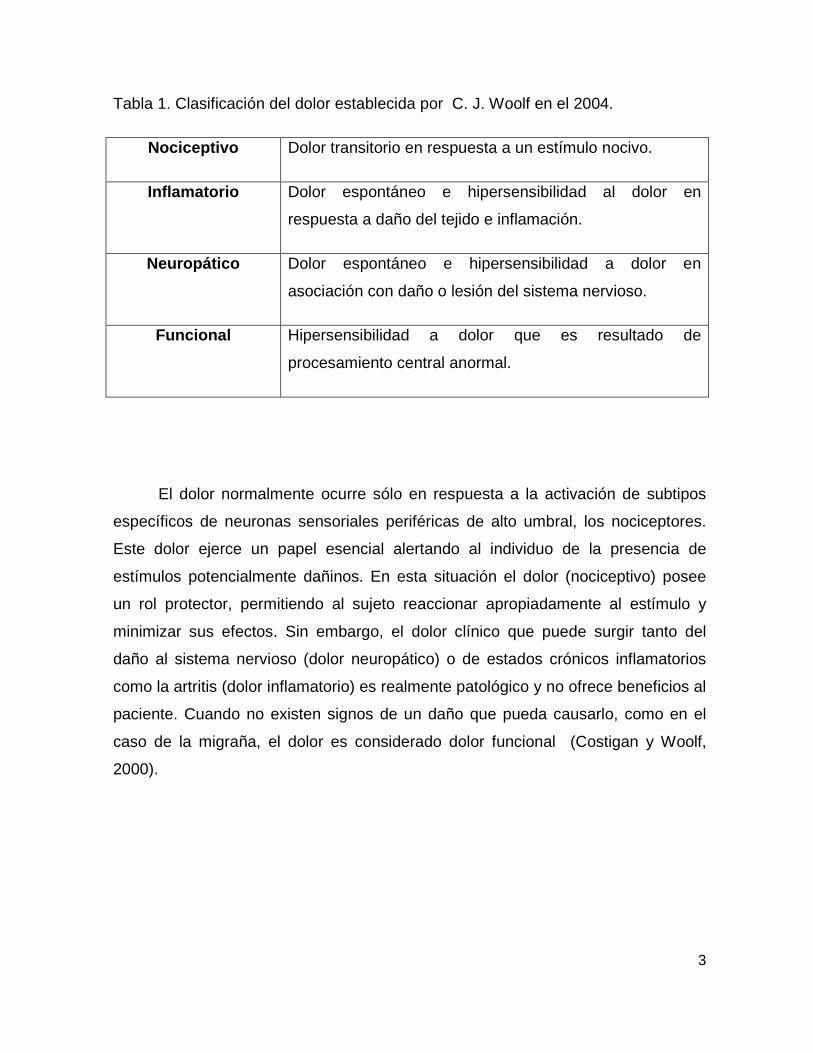
Tabla 1. Clasificación del dolor establecida por C. J. Woolf en el 2004.
Nociceptivo Dolor transitorio en respuesta a un estímulo nocivo.
Inflamatorio Dolor espontáneo e hipersensibilidad al dolor en
respuesta a daño del tejido e inflamación.
Neuropático Dolor espontáneo e hipersensibilidad a dolor en
asociación con daño o lesión del sistema nervioso.
Funcional Hipersensibilidad a dolor que es resultado de
procesamiento central anormal.
El dolor normalmente ocurre sólo en respuesta a la activación de subtipos
específicos de neuronas sensoriales periféricas de alto umbral, los nociceptores.
Este dolor ejerce un papel esencial alertando al individuo de la presencia de
estímulos potencialmente dañinos. En esta situación el dolor (nociceptivo) posee
un rol protector, permitiendo al sujeto reaccionar apropiadamente al estímulo y
minimizar sus efectos. Sin embargo, el dolor clínico que puede surgir tanto del
daño al sistema nervioso (dolor neuropático) o de estados crónicos inflamatorios
como la artritis (dolor inflamatorio) es realmente patológico y no ofrece beneficios al
paciente. Cuando no existen signos de un daño que pueda causarlo, como en el
caso de la migraña, el dolor es considerado dolor funcional (Costigan y Woolf,
2000).

4
1.1.2. Nocicepción
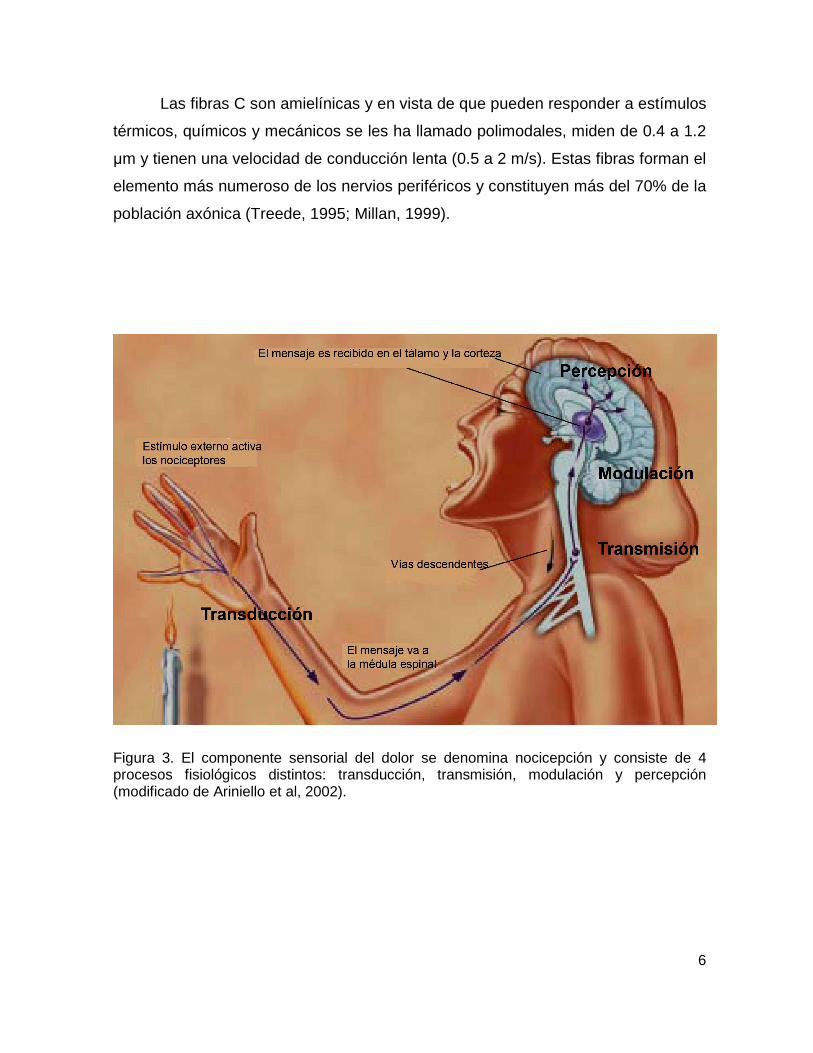
La nocicepción es un término neurofisiológico que se refiere a los
mecanismos neurales por los cuales se detectan los estímulos nocivos y consiste
en 4 procesos fisiológicos distintos: transducción, transmisión, percepción y
modulación (D’Amours y Ferrante, 1997). La transducción describe la conversión
de estímulos nocivos en señales eléctricas por las terminaciones nerviosas
periféricas. La transmisión es la propagación de las señales eléctricas a lo largo de
las vías nociceptivas. La percepción es el proceso por el cual se integran los
impulsos nociceptivos con factores cognoscitivos y emocionales para crear la
experiencia subjetiva de dolor. Además, se puede presentar la alteración de las
señales nociceptivas dentro del asta dorsal de la médula espinal y en la corteza
cerebral, en las que se entrecruzan terminaciones nerviosas de diversos tipos que
liberan neurotransmisores que inhiben, amplifican o alteran la señal sensorial. A
este último proceso se le conoce como modulación (Figura 3).
Los estímulos nociceptivos se transmiten de la periferia hasta la médula
espinal a través de los nervios periféricos que están formados por fibras de
diámetro y velocidad de conducción distintos. En función de estos dos parámetros
dichas fibras se clasifican en tres tipos: A, B y C.
Las de tipo A se subdividen en fibras α (alfa), β (beta), γ (gama) y δ (delta).
Las neuronas aferentes primarias propioceptivas Aβ son gruesas (más de 10 µm de
diámetro), mielinizadas y de conducción rápida (30 a 100 m/s). Las fibras aferentes
Aδ tienen como función la detección y transmisión de estímulos térmicos y
mecánicos. Estas fibras son mielinizadas parcialmente, delgadas (2-6 µm) y de
conducción intermedia (12 a 30 m/s). Las fibras de tipo B se relacionan con
estímulos neurovegetativos.
5
Figura 2. De acuerdo a su origen somático y fisiológico, Woolf clasifica al dolor en cuatro tipos: Nociceptivo inflamatorio, neuropático y funcional (Modificado de Woolf, 2004).
6
Las fibras C son amielínicas y en vista de que pueden responder a estímulos
térmicos, químicos y mecánicos se les ha llamado polimodales, miden de 0.4 a 1.2
µm y tienen una velocidad de conducción lenta (0.5 a 2 m/s). Estas fibras forman el
elemento más numeroso de los nervios periféricos y constituyen más del 70% de la
población axónica (Treede, 1995; Millan, 1999).
Figura 3. El componente sensorial del dolor se denomina nocicepción y consiste de 4 procesos fisiológicos distintos: transducción, transmisión, modulación y percepción (modificado de Ariniello et al, 2002).

7
En general se considera que las fibras Aδ y C son las encargadas directas
de la neuroconducción del dolor, sobresaliendo el papel de las fibras C en virtud
de su gran número, ya que aunque su velocidad de conducción es lenta pueden
transmitir una cantidad enorme de información nociceptiva. Por otro lado, las fibras
Aδ suelen clasificarse funcionalmente en dos categorías: Las fibras Aδ tipo I, que
se activan a temperaturas mayores de 52°C y las fib ras Aδ tipo II que lo hacen a
temperaturas mayores de 42°C. Las fibras nociceptiv as aferentes primarias Aδ y C
establecen su primer relevo en las astas dorsales de la médula espinal a través de
las raíces posteriores, conectándose con la capa marginal y la substancia
gelatinosa. Las fibras Aβ detectan estímulos táctiles inocuos, pero en ciertas
condiciones patológicas pueden ramificar colateralmente a las regiones externas
de las astas dorsales de forma que los estímulos inocuos se pueden interpretar
como dolorosos (Diamond et al, 1992; Baba et al, 1999).
1.1.3. Sensibilización periférica
Una explicación para la hipersensibilidad al dolor después de una lesión es
que la sensibilidad de los nociceptores de alto umbral cambia. Esta idea fue
propuesta de manera vigorosa por Thomas Lewis en la década de 1930 sugiriendo
además que el reflejo desde el axón era responsable de ampliar la sensibilización
desde la zona de daño tisular (hiperalgesia primaria) hacia regiones vecinas en las
que la respuesta está aumentada y el tejido sin daño (hiperalgesia secundaria)
(Lewis, 1942). A mediados de los 60’s Bessou y Perl (1969) demostraron en
receptores polimodales aislados que los estímulos térmicos intensos alteran la
sensibilidad de una gran proporción de fibras sensoriales a un estímulo térmico
subsecuente. Este trabajo se ha replicado extensamente por lo que actualmente no
quedan dudas de que el daño tisular y la respuesta inflamatoria que provoca da
inicio a cambios locales en la sensibilidad de las fibras nociceptivas, fenómeno que
se conoce como sensibilización periférica (Treede et al, 1992).

8
Una lesión tisular produce cambios profundos en el ambiente químico de las
terminales de los nociceptores periféricos (Woolf, 2004). Al dañarse, las células
liberan sustancias contenidas como adenosín trifosfato (ATP), iones hidrógeno (H+)
y potasio (K+). Además, las células (macrófagos, mastocitos, neutrófilos, etc)
reclutadas a la región del daño liberan citocinas y factores de crecimiento (Miao y
Levine, 1999). Algunos de estos factores actúan directamente en la terminal del
nociceptor para activarlo produciendo dolor, por lo que se denominan
pronociceptivos. Otros contribuyen a que la terminal se vuelva hipersensible a un
estímulo subsecuente, estableciéndose como sustancias sensibilizantes. Lo que
viene a complicar este aspecto es que tanto las sustancias pronociceptivas (ATP,
iones H+ y K+), como las sensibilizantes (sustancia P, prostaglandinas, péptido
relacionado con el gen de la calcitonina, etc.) actúan de manera sinérgica. De
forma que para lograr de manera óptima la sensibilización periférica se requiere la
presencia de un “cocktail” o “sopa” de esos diferentes mediadores (Handwerker,
1991; Woolf, 1995; Pertoldi y Benedetto, 2005). Los agentes sensibilizantes, como
la prostaglandina E2, reducen el umbral de activación de los nociceptores
aumentando la respuesta de la terminal al unirse de manera específica a
receptores situados en la membrana de la neurona aferente primaria. Estos
receptores están acoplados a cinasas intracelulares que se encuentran en el
citoplasma. La sensibilización periférica ocurre inicialmente mediante la activación
de diferentes tipos de estas cinasas que a su vez fosforilan múltiples proteínas,
entre ellas canales iónicos dependientes de voltaje y canales iónicos operados por
ligando. Esto ocasiona un incremento de corrientes de sodio (Na+) y calcio (Ca+2)
aunado a una disminución de corrientes de potasio (K+) (Gold et al, 1998; Millan,
2002).

9
Seguido de estos cambios tempranos (segundos) comienzan a producirse
modificaciones transcripcionales (minutos). Estas modificaciones se deben a la
acción de proteínas cinasas activadas que fosforilan factores de transcripción y
neurotrofinas como el factor de crecimiento neural, el cual se internaliza junto con
su receptor de membrana TrkA al núcleo celular donde inicia estos cambios (Leslie
et al, 1995). Los cambios en la transcripción refuerzan la excitabilidad neuronal ya
que se incrementa la síntesis de moléculas involucradas en la transmisión
nociceptiva como sustancia P y péptido relacionado con el gen de la calcitonina
(Costigan y Woolf, 2000) que no sólo afectan a las fibras C, sino también a las
fibras Aβ y Aδ que pueden modificar su fenotipo e inervación en el asta dorsal.
Un ejemplo de lo que ocurre durante la sensibilización periférica lo ofrecen
los receptores vainilloides TRPV1 (receptores de potencial transitorio V1). Estos
receptores son canales catiónicos que se activan por calor nocivo (42°C) y son
modulados por el pH. En condiciones de dolor nociceptivo se activan a 42°C, pero
una vez que las terminales periféricas se sensibilizan este tipo de canales puede
activarse a temperatura ambiente.
1.1.4. Sensibilización central
A principios de los años ochenta, un estudio sobre la plasticidad cerebral en
el sistema somatosensorial condujo al descubrimiento de que las lesiones tisulares
accionan un aumento en la excitabilidad de las neuronas de la médula espinal, un
fenómeno que ahora se conoce como sensibilización central (Woolf, 1983). Más
adelante se pudo saber que estos cambios plásticos se originan por la acumulación
de potenciales sinápticos subumbrales y que los receptores al ácido N-metil-D-
aspártico (NMDA) son una pieza clave para su desarrollo (Inturrisi, 1994; Ma y
Woolf, 1995; Pertoldi y Benedetto, 2005).

10
Figura 4. Sensibilización periférica. La figura muestra diferentes receptores de transducción de señal y canales iónicos situados en la terminal periférica de un nociceptor. En condiciones de dolor nociceptivo (A), estos receptores responden a estímulos térmicos, químicos y mecánicos de alto umbral. Si los estímulos persisten en intensidad y duración (B), aparte de los mediadores nociceptivos, sustancias sensibilizantes como la prostaglandina E2 (PGE2) o el factor de crecimiento neuronal (NGF) activan cinasas intracelulares que promueven fosforilaciones que reducen el umbral de los receptores e incrementan su excitabilidad (modificado de Woolf, 2004).
El estímulo normal nociceptivo de las fibras C y Aδ resulta en la liberación de
glutamato desde sus terminales centrales. El glutamato se une a varios receptores
con distinta afinidad, entre los que se incluyen los ionotrópicos AMPA y NMDA y
algunos metabotrópicos (Miller y Woolf, 1996). El glutamato se une inicialmente al
receptor AMPA, debido a que bajo circunstancias normales (potencial de reposo)
los receptores NMDA están bloqueados por un ión Mg+2. Los receptores AMPA se
activan por la unión de glutamato de forma que el influjo de iones Na+, K+ y Ca+2 a
través de sus canales resulta en una despolarización de las células de las astas
dorsales (figura 5). Esto se manifiesta en potenciales excitatorios postsinápticos
rápidos y señales que le dan duración, intensidad y localización al estímulo nocivo
periférico. La plasticidad y subsecuente sensibilización del sistema nervioso central
que incluyen la alteración de las funciones químicas, electrofisiológicas y

11
farmacológicas (Carlton et al, 1998; Winkelstein, 2004). Estos cambios causan una
percepción exagerada de los estimulos dolorosos (hiperalgesia) y una percepción
dolorosa de estímulos táctiles normales (alodinia) (Li et al, 1999; Kidd y Urban,
2001; Graven-Nielsen y Arentd-Nielsen, 2002).
La sumación temporal o “wind up” es un mecanismo espinal en el que la
estimulación intensa y sostenida asociada al daño tisular induce una suma
temporal de las despolarizaciones postsinápticas, que se experimenta como un
aumento del dolor (Gracely et al, 2003; Meeus, 2007). En 1965, experimentos en
animales mostraron por primera vez que la estimulación repetitiva de las fibras C
podían resultar en un aumento progresivo de las descargas eléctricas de las
neuronas de segundo orden en la médula espinal (Mendell y Wall, 1965). Este
mecanismo de amplificación produce un dolor sordo y está más relacionado con los
estados de dolor crónico, es transmitido a través de las fibras C no mielinizadas a
las neuronas nociceptivas de las astas dorsales.
El establecimiento del “wind up” resulta en la remoción del magnesio de los
receptores NMDA, permitiendo la entrada de más Ca+2 a la terminal postsináptica
(Inturrisi, 1994). Esto a su vez, activa cascadas de transducción de señal sensibles
a Ca+2 que conducen a la fosforilación de canales iónicos y receptores (entre los
que se incluyen los propios receptores NMDA), iniciando un prolongado incremento
en la excitabilidad de las neuronas espinales (Dubner y Ruda, 1992; Yoshimura y
Nishi, 1993; Woolf y Salter, 2000). Dicha entrada de Ca+2 a las neuronas
sensoriales de las astas dorsales induce la activación de la sintasa del óxido nítrico
y la posterior síntesis del óxido nítrico (Meller y Gebhart, 1993). El óxido nítrico
puede afectar las terminales de los nociceptores y aumentar la liberación de
neuropéptidos (en particular la sustancia P) desde las neuronas presinápticas,
contribuyendo con ello al desarrollo de la hiperalgesia (Luo y Cizcova, 2000). La
sustancia P es un neurotransmisor nociceptivo importante, disminuye el umbral de
excitabilidad sináptica y desencadena sinapsis interespinales que normalmente se
mantienen silentes (Liu et al, 1994). Más aún, la sustancia P puede extender su
efecto al recorrer grandes distancias desde su neurona de origen, ampliando

12
los campos receptivos y activando las neuronas de rango amplio y dinámico
incluso con estímulos no-nociceptivos (Staud, 2002; Meeus, 2007).
Si la serie de estímulos nocivos es lo suficientemente persistente ocurrirán
cambios en la transcripción dependientes de la actividad que redundan en un dolor
también persistente para los pacientes (Mannion et al, 1999; Ohtori et al, 2002). De
tal suerte que los estímulos relativamente breves (con duración de décimas de
segundo), sobre todo de las fibras C, pueden iniciar cambios muy rápidos en la
excitabilidad de la membrana al tiempo que progresivamente aumentan su
excitabilidad durante el curso del propio estímulo y promueven cambios que
perduran hasta por varias horas después de éste (Costigan y Woolf, 2000; Meeus,
2007).
Figura 5. Sensibilización central. En condiciones de dolor nociceptivo (A) la terminal central de los nociceptores libera aminoácidos excitatorios hacia las neuronas de transmisión de la médula espinal. Los potenciales de acción generados por glutamato en los receptores AMPA y kainato se conducen a centros superiores. Si la estimulación nociceptiva se mantiene (B) aumenta la concentración de los aminoácidos excitatorios y se agrega la liberación de sustancias sensibilizantes (como sustancia P y el factor neurotrófico derivado del cerebro) al medio promoviendo la actividad de cinasas que permiten la activación de los receptores NMDA y cambios en la transcripción. Esto permite amplificar las señales nociceptivas de tal forma que los pulsos que en condiciones nociceptivas tienen duración de segundos, pueden tener efectos por horas cuando la terminal central está sensibilizada (modificado de Woolf, 2004).

13
1.1.5. Inflamación
La inflamación puede definirse como una reacción defensiva local integrada
por alteración, exudación y proliferación. La reacción es desencadenada por
estímulos nocivos de muy diversa naturaleza: físicos, químicos y microorganismos
como bacterias, hongos y parásitos. El aspecto macroscópico de este proceso fue
caracterizado por Celso por cuatro signos, que se conocen hoy como los signos
cardinales de la inflamación: rubor y tumor con calor y dolor (Nathan, 2002).
La respuesta inflamatoria es una habilidad esencial para la supervivencia
como forma de enfrentar a organismos patógenos del medio y controlar las lesions
tisulares. Sin embargo, una inflamación exacerbada y sostenida puede presentarse
sin aparente beneficio e incluso implicar severas consecuencias (Pepper, 2005). El
fenómeno de la inflamación puede ocurrir en 3 distintas fases temporales, cada
una aparentemente mediada por distintos mecanismos: Una fase aguda
caracterizada por la presencia de vasodilatación y aumento de la permeabilidad
capilar, una fase subaguda que implica la infiltración de células leucocitarias y una
fase crónica proliferativa en la que se observa degeneración tisular y fibrosis
(Meunier, 2004).
1.1.5.1. Dolor inflamatorio
Los tejidos cutáneos y somáticos están inervados por neuronas aferentes
primarias que hacen sinapsis en la médula espinal. Las funciones de estas
neuronas incluyen la detección de estímulos nocivos y la subsiguiente transmisión
de información codificada al cerebro. El dolor es un proceso perceptivo que surge
como respuesta a dichas funciones. Muchos mecanismos están involucrados en la
promoción y resolución del proceso inflamatorio, tras el daño tisular los receptores
activados inician varias cascadas de transducción de señales que resultan en la
modulación postraduccional de moléculas efectoras asociadas a alteraciones en
los sistemas neuronales implicados en la transmisión nociceptiva, que se
exacerban con cambios tardíos en la transcripción de genes (Costigan y Woolf,

14
2000; Meunier et al, 2004).
1.1.5.1.1. Mediadores pronociceptivos
La inflamación resulta de la liberación de una compleja mezcla de citocinas y
otros agentes neuroactivos provenientes tanto de células inflamatorias como de no
inflamatorias reclutadas en el sitio de daño (Kidd y Urban, 2001). Estos agentes
actúan químicamente en receptores específicos y canales iónicos en las terminales
periféricas de los nociceptores (Figura 6). Estos mediadores tienen un gran impacto
cualitativo y cuantitativo en la producción de la inflamación así como en la actividad
de las fibras aferentes primarias (Dray, 1995; Millan, 1999, Ma y Quirion, 2007).
1.1.5.1.1.1. Protones
La producción de protones aumenta en la inflamación y se involucra en la
hiperalgesia inflamatoria y en la sensación desagradable debida a la hipoxia/anoxia
causada por el ejercicio muscular. De hecho, el dolor punzante producido por
inyecciones intradérmicas de soluciones ácidas y la disminución del pH aumenta
los efectos de otros mediadores inflamatorios (Bevan y Geppetti, 1994; Dray,
1994). Las soluciones ácidas administradas de manera exógena producen un
rápido pero transitorio aumento en la permeabilidad de cationes en las membranas,
lo que incrementa la activación sostenida de los nervios y la mecanosensibilidad
(Steen et al, 1992). El mecanismo por el que los protones inducen la activación de
las neuronas sensoriales está mediado por los receptores TRPV1. El receptor de
capsaicina TRPV1 es un canal iónico que se encuentra específicamente en los
nociceptores. Este receptor puede ser activado por la acidificación que causan los
protones, la capsaicina, y también por calor nocivo. Todos estos factores pueden
causar dolor in vivo. Algunos estudios con ratones deficientes en receptores
TRPV1 han demostrado que estos canales son esenciales para originar
hiperalgesia térmica. Un mecanismo que caracteriza al dolor inflamatorio iniciado
por el daño tisular es la sensibilización de los receptores TRPV1 (Numazaki y
Tominaga, 2004). Otros estudios indican que los receptores TRPV1 participan en la

15
nocicepción sólo cuando existen condiciones de acidez extrema (Shimada et al,
2004) y que los principales receptores a la disminución del pH en condiciones
inflamatorias en los nociceptores humanos son los llamados canales iónicos
sensibles al ácido (ASICs).
1.1.5.1.1.2. Cininas
Las cininas ejercen numerosos efectos proinflamatorios como la liberación
de prostanoides, citocinas y radicales libres desde varios tipos de células.
Estimulando las neuronas simpáticas postganglionares (Hall y Geppetti, 1995) que
afectan el calibre de los vasos sanguíneos. Las cininas degranulan los mastocitos
para liberar histamina y otros mediadores de la inflamación, causando
extravasación plasmática por la contracción de las células del endotelio vascular.
Las cininas son sustancias algogénicas potentes e inducen dolor por la activación
directa de los nociceptores, sensibilizándolos al calor y los estímulos mecánicos
(Beck y Handwerker, 1974; Rang et al, 1994). Se presenta un fuerte sinergismo
entre las acciones de bradicinina y otras sustancias algógenas como las
prostaglandinas y la serotonina. En la sangre la bradicinina se forma como parte de
la cascada de la coagulación por el proceso enzimático de un precursor cininógeno
de alto peso molecular. En otros tejidos, la kalidina de escaso peso molecular,
también se convierte en bradicinina por acción enzimática (Wang et al, 2006).
El efecto de las cininas está mediado por 2 receptores, B1 y B2 (Hall, 1992;
Dray et al, 1993; Menke et al, 1994). Los agonistas endógenos para el receptor B2
son la bradicinina y la kalidina. Estudios con antagonistas selectivos de los
receptores B2 confirman que la bradicinina es un mediador importante del dolor y
sus antagonistas atenúan el dolor y la hiperalgesia que se asocia a varias
condiciones inflamatorias (Perkins et al, 1993; Steranka et al, 1998; Wang et al
2006). Esos datos también indican que los antagonistas B2 tienen un potencial
analgésico como terapéuticos.

16
Los receptores B2 se expresan de manera conjunta en los nociceptores con
los receptores TRPV1. Los receptores inducibles B1, que se regulan a la alta en la
inflamación, también se co-localizan con estos receptores vainilloides en las
neuronas sensoriales, por lo que se sugiere que los receptores de potencial
transitorio V1 podrían en parte ser responsables de las conductas nociceptivas
inducidas por la bradicinina (Ferreira et al, 2003). No existe evidencia de una
activación directa de las neuronas nociceptivas a través del receptor B1, al parecer
la hiperalgesia mediada por estos receptores es por la liberación de otros
mediadores como las prostaglandinas en macrofágos y leucocitos.
Los receptores B2 están presentes en las neuronas sensoriales y se acoplan
a las proteínas G para inducir la activación de la fosfolipasa C y generar diacil
glicerol (DAG) e inositol trifosfato (IP3) por el rompimiento de fosfolípidos de la
membrana. El IP3 estimula la liberación de Ca+2 almacenado intracelularmente
mientras que el DAG activa la proteína cinasa C (PKC) para fosforilar proteínas que
incluyen receptores celulares y canales iónicos. La PKC tiene un papel clave en la
excitación de las fibras aferentes ejercida por la bradicinina que se asocia a un
aumento al paso de iones, principalmente de sodio (Rang et al, 1994; Thayer et al,
1998).
Las neuronas sensoriales tienen una alta concentración de cloruro (Cl-)
debido a la presencia del cotransportador de sodio, potasio y cloruro (NKCC1). Las
corrientes entrantes de Cl- aumentan la excitabilidad neuronal que se exacerba aún
más debido al aumento de la conductancia de calcio activado por el cloruro en
células neuronales, pero también en células no neuronales lo que podría implicar
una excitación indirecta de los nociceptores causada por la cascada de efectos que
origina la bradicinina (England et al, 2001; Oh and Weinreich, 2004).

17
1.1.5.1.1.3. Prostanoides
Los prostanoides (prostaglandinas, leucotrienos e hidroxiácidos) están entre
los mediadores más importantes de la hiperalgesia inflamatoria, se generan a partir
del ácido araquidónico por la actividad de las enzimas ciclooxigenasa y
lipoxigenasa. Las prostaglandinas actúan por medio de varios receptores
acoplados con segundos mensajeros (Coleman et al, 1994; Bingham et al, 2006),
pero los receptores EP para la PGE2 e IP para la prostaciclina son probablemente
los más importantes por sus efectos en las neuronas sensoriales. De hecho, un
subtipo de receptor, el EP3, ha sido recientemente identificado en la mayoría de las
neuronas sensoriales no mielinizadas. Las prostaglandinas usualmente no evocan
dolor cuando se inyectan en la piel de manera intradérmica (Crunkhorn y Willis,
1971). No obstante, se ha reportado que la prostaglandina E1 y la prostaciclina
incrementan de manera directa la actividad de los nociceptores (Birell et al, 1991;
Zeilhofer, 2007) y la prostaglandina E2 estimula la liberación de sustancia P en
cultivos de celulas neuronales. Estos efectos despolarizantes pueden deberse a un
aumento en la conductancia de sodio en la membrana. De manera más usual, las
prostaglandinas sensibilizan las neuronas sensoriales reduciendo su umbral de
activación y exacerbando sus respuestas a otros estímulos (Dray, 1994; Levine et
al, 1993; Rang et al, 1994; Zeilhofer, 2007).
En los mamíferos existen 2 genes que codifican diferentes ciclooxigenasas.
La ciclooxigenasa 1 es prácticamente ubicua y se expresa de manera constitutiva
en los tejidos para efectuar diversas funciones fisiológicas, mientras que la
ciclooxigenasa 2 generalmente está presente en muy bajos niveles pero es
altamente inducible y se incrementa por muchos tipos de estímulos como puede
ser la presencia de citocinas o factores de crecimiento (Chandrasekharan, 2004), lo
que ocurre en la inflamación.
Los analgésicos no esteroideos poseen sus propiedades analgésicas y
antipiréticas por el bloqueo de las enzimas ciclooxigenasas y los compuestos
selectivos para la Ciclooxigenasa 2 producen analgesia con una enorme reducción

18
en los efectos colaterales gastrointestinales, pero de manera decepcionante
recientemente se ha descrito que poseen una potencial toxicidad cardiovascular
(Zeilhofer, 2007).
Algunos estudios han postulado la existencia de una variante de la
ciclooxigenasa 1 en caninos con una selectividad distinta a la inhibición por varios
analgésicos no esteroideos, como el acetaminofén y el metamizol, llamada
ciclooxigenasa 3 (Chandrasekharan, 2002). En contraparte, otros estudios han
cuestionado la presencia de esta enzima en tejidos humanos y la existencia de una
selectividad diferencial con la ciclooxigenasa 1 (Dinchuk et al, 2003; Qin et al,
2005).
La administración intradérmica del leucotrieno B4 (LTB4), un producto de la
vía de la lipoxigenasa, también disminuye los umbrales nociceptivos (Levine et al,
1993; Rang et al, 1994). El LTB4 produce directamente hiperalgesia al disminuir el
umbral de los receptores térmicos y mecánicos de las fibras C. Recientemente se
ha descrito el papel del LTB4 como un mediador crítico en el desarrollo de la
hiperalgesia mecánica inducida por la inyección intraplantar de ovoalbúmina como
antígeno (Cunha et al, 2003).
1.1.5.1.1.4. Adenosín trifosfato
La capacidad del adenosín trifosfato (ATP) para producir dolor se conoce
desde hace alrededor de 40 años (Collier et al, 1966; Kennedy et al, 2003), pero
los receptores y los mecanismos a través de los que interviene en la señalización
sensorial apenas comienzan a discernirse. Los efectos fisiológicos y
farmacológicos del ATP se producen mediante la actividad de los receptores P2X y
P2Y. Al respecto existe un creciente interés desde 1995, cuandó se clonó el
receptor P2X3 y se pudo saber que se expresaba en altas concentraciones
selectivamente en las neuronas nociceptivas (Chen et al, 1995; Fukui et al, 2006).

19
Inicialmente, se propuso que el Adenosín trifosfato liberado desde las
células dañadas tendría un papel importante en el dolor nociceptivo (Burnstock y
Wood, 1996), pero reportes recientes del empleo de ratones knock out y
antagonistas selectivos de los receptores P2X3, indican que el ATP está más
involucrado en condiciones crónicas, como lo son el desarrollo de dolor inflamatorio
y neuropático (Souslova et al, 2000; Jarvis et al, 2002). Sugiriendo que el adenosín
trifosfato liberado tónicamente de las neuronas dañadas o células vecinas
promueven la activación sostenida de los receptores P2X3 en las neuronas
sensoriales provocando potenciales de acción intensos que viajan a hacia el
sistema nervioso central (Kennedy et al, 2003), lo que se experimenta
concientemente como dolor crónico.
Aunque la atención está dirigida principalmente hacia los receptores P2X3, los
receptores P2Y también se expresan en las neuronas nociceptivas y se les atribuye
un rol más modesto en la nocicepción. Los receptores P2Y están acoplados a
proteínas G, lo que conduce a la activación de la fosfolipasa C, la liberación de
calcio intracelular y la posterior activación de la proteína cinasa C (Abbracchio et al,
2006). La mediación de estos efectos indica el potencial uso antinociceptivo de
antagonistas de estos receptores.
1.1.5.1.1.5. Serotonina
Está bien establecido que la serotonina se involucra en el procesamiento del
dolor. La actividad serotonérgica está mediada por al menos 12 tipos de receptores
distintos distribuidos de manera heterogénea, algunos con actividad pronociceptiva
y otros con capacidad analgésica (Giordano y Schultea, 2004). La mayoría de
estos receptores están metabotrópicamente ligados a proteínas G, por lo que
activan cascadas de segundos mensajeros para modular la actividad neuronal
La excepción la constituyen los receptores 5-HT3 que están asociados a un
canal de Na+. Lo que facilita la presencia de corrientes de sodio resistentes a
tetrodotoxina dependientes de voltaje que son específicas de las neuronas

20
sensoriales de calibre pequeño (Cárdenas et al, 1997; Millan, 1999; Bhargava y
Saha, 2001), disminuyendo su umbral de activación y aumentando la magnitud de
sus descargas.
En lo que se refiere a los receptores metabotrópicos de la serotonina, las
alteraciones que son promovidas en la excitabilidad pueden estar mediadas por
activación de la enzima adenilato ciclasa y la generación de AMPc. El AMPc
desencadena una serie de fosforilaciones mediadas por la proteína cinasa A (PKA)
que al actuar sobre canales de potasio disminuyen su conductancia contribuyendo
a la sensibilización de las aferentes primarias (Doak y Sawynok, 1997; Sasaki et al,
2003). Los receptores 5-HT2, 5-HT4 y 5-HT6 están acoplados a este sistema de
transducción de señal como factores pronociceptivos (Ponimaskin et al, 2002), a
pesar de que no hay evidencias contundentes de la expresión del receptor 5-HT6
en el ganglio de la raíz dorsal (Doak y Sawynok, 1997) y de que autores como
Dawson et al (2000) señalan que los antagonistas 5-HT6 aumentan la
concentración extracelular de glutamato y aspartato in vivo.
Por otro lado, la concepción original de que los receptores 5-HT1
promueven acciones pronociceptivas en las aferentes primarias (Levine et al,
1993), se ha vuelto endeble en función de que se ha descubierto que estos
receptores se acoplan negativamente a la adenilato ciclasa (Del mar et al, 1994).
Los receptores 5-HT1D pueden reducir la excitabilidad nociceptiva, la vasodilatación
y la extravasación plasmática que induce la liberación de neuropéptidos. Varios
agentes antimigrañosos actúan mediante este receptor (Buzzi y Moskowitz, 1990;
Bardin et al, 2000). Se ha observado que los receptores 5-HT5, presentan una
gran homología con los receptores 5-HT1D, pero parecen tener un perfil
farmacológico distinto debido a que no inhiben a la adenilato ciclasa y su expresión
mayoritaria es en sitios supraespinales (Plassat et al, 1993).

21
De manera reciente, algunos reportes en la literatura describen la
participación periférica y espinal de los receptores 5-HT7 en la hiperalgesia
inducida por la inflamación en el modelo de la formalina (Rocha-González et al,
2005).
1.1.5.1.1.6. Histamina
La histamina se puede liberar por la degranulación de los mastocitos
inducida por otros mediadores como la sustancia P. Su acción puede producir
comezón en bajas concentraciones y dolor cuando las concentraciones son altas
(Simone et al, 1991; Watanabe y Yanai, 2001).
Estudios efectuados con ratones knockout empleando modelos de dolor
como el tail flick, hot plate y la prueba de formalina indican que la histamina
exacerba la respuesta a los estímulos nociceptivos. Debido a que en todos estos
paradigmas los ratones knockout fueron menos sensibles que los de la cepa
silvestre (Mobarakeh et al, 2001). Las neuronas sensoriales liberan histamina y por
acción de sus receptores H1 se eleva la permeabilidad al calcio, lo que provoca la
liberación de prostaglandinas y otros péptidos conduciendo a la presencia de
hiperalgesia y otros efectos inflamatorios (Rang et al, 1994). Esto refuerza la idea
establecida en la clínica de forma empírica, de que los antihistamínicos ayudan a la
disminución del dolor si se administran de manera preventiva (Watanabe y Yanai,
2001). Asimismo, existen datos que muestran efectos sinérgicos de la morfina
cuando se coadministra con bloqueadores de los receptores H1 y H2. (Mobarakeh
et al, 2002).
1.1.5.1.1.7. Neurocininas
Normalmente algunos neuropéptidos (neruocininas, CGRP) son liberados
de las terminales nerviosas para ejercer efectos tróficos eferentes en sus sitios
blanco (Rang et al, 1994; Woolf, 2004). Sin embargo, durante la inflamación las
neurocininas sustancia P y neurocinina A (NKA) contribuyen de manera directa e
indirecta a la inflamación neurogénica, la presencia de hiperalgesia en la periferia

22
y los cambios en la excitabilidad de la médula espinal en sus astas dorsales.
Algunos estudios en los que se emplearon antagonistas indican que los efectos de
las neurocininas son mediados a través de la activación de receptores específicos
(Maggi et al, 1992; Hill, 2000). Actualmente los receptores NK1 y NK2 más que el
NK3, se involucran en los efectos proinflamatorios e hiperalgésicos de las
neurocininas. Estudios recientes indican que los receptores NK inducen la
activación de la proteína cinasa C, ejerciendo un rol crítico en la modulación de la
excitabilidad de las neuronas sensoriales (Ferreira et al, 2005)
Estos compuestos pueden también despolarizar de manera directa las
neuronas nociceptivas al reducir la permeabilidad al potasio (Dray y Pinock, 1982)
por medio de los receptores NK1 que están presentes en las terminales de las
fibras aferentes primarias. El papel de las neurocininas en la hiperalgesia
inflamatoria tiene el soporte experimental de que los antagonistas de los receptores
NK1 producen analgesia conductual, a pesar de ello, algunos estudios clínicos han
fallado al tratar de demostrar eficacia en diferentes tipos de dolor (Birch et al, 1992;
Hill, 2000).
1.1.5.1.1.8. Especies reactivas de oxígeno
Las especies reactivas de oxígeno han recibido poca atención con respecto
a la activación de las fibras aferentes durante el daño tisular y la inflamación. Las
especies reactivas de oxígeno incluyen al peróxido de hidrógeno y los iones
superóxido e hidroxilo. Son productos normales de las reacciones celulares de
transferencia de electrones que regulan en gran medida la transcripción genética.
Normalmente la producción de estas especies está finamente controlada por la
actividad antioxidante de la catalasa y la superóxido dismutasa.
Se sabe que el peróxido de hidrógeno aumenta los efectos de otros
mediadores inflamatorios incluyendo la bradicinina y la prostaglandina E2, mientras
que el óxido nítrico, induce un dolor quemante retardado bajo inyección
intradérmica (Holthusen y Arndt, 1994; Salvemini et al, 2006) y se ha postulado que

23
los donadores de óxido nítrico activan directamente fibras sensoriales cerebrales,
causando la liberación del vasodilatador CGRP. Estos datos Sugieren la
contribución del óxido nítrico a la migraña y otros dolores de cabeza (Olesen et al,
1994; Ciancarelli et al, 2007).
1.1.5.1.1.8.1. Óxido nítrico
La vía más importante de la formación de óxido nítrico es a partir de la L-
arginina. Esta ocurre en varios tipos celulares pero particularmente en las neuronas
de tamaño pequeño y mediano (Verge et al, 1992; Luo y Cizcova, 2000) tras la
activación de la sintasa de óxido nítrico por calcio y otros cofactores. El óxido
nítrico (ON) altera procesos celulares principalmente por la activación de la
guanilato ciclasa. En relación con esto, el aumento en la producción de GMPc
ocurre en células satélite en el ganglio de la raíz dorsal después de la
administración de donadores del óxido nítrico. Durante la inflamación, una forma
inducible y dependiente de calcio de la sintasa de óxido nítrico se presenta (Verge
et al, 1992; Dray 1994; Cheshire, 2001) lo que aumenta la síntesis de ON. Esta
sintasa inducible parece ser importante en la regulación de la actividad de la
ciclooxigenasa 2 y la posterior producción de prostanoides inflamatorios. Además,
el óxido nítrico puede alterar la respuesta de las neuronas sensoriales a otros
mediadores inflamatorios como la bradicinina. Recientemente, se ha sugerido que
la formación de óxido nítrico contribuye a las descargas ectópicas inducidas por las
lesiones de los nervios periféricos y que el aumento de la excitabilidad en estas
condiciones se podría reducir con el empleo de inhibidores selectivos de la sintasa
de óxido nítrico (Verge et al, 1992; Olesen, 1994; Hoheisel y Mense, 2000;
Cheshire, 2001).
1.1.5.1.1.9. Mediadores inflamatorios de las celula s inmunes
Varias citocinas, como las interleucinas (IL-1, IL6, IL8) y el factor de necrosis
tumoral alfa (TNFα), se liberan desde varias células inmunes y pueden producir una
profunda hiperalgesia (Flatters et al, 2003). La cual es mediada por varios

24
mecanismos indirectos entre los que se encuentran la liberación de prostanoides y
el aumento de la expresión de receptores a bradicinina (Dray, 1995; Ferreira,
2003).
Por otro lado, las neurotrofinas, en especial el factor de crecimiento neuronal
(NGF), en condiciones inflamatorias aumentan su producción debido a la
presencia de otros mediadores inflamatorios como la interleucina 1 y el factor de
necrosis tumoral. El factor de crecimiento neuronal incrementa a su vez la síntesis
del péptido relacionado con el gen de la calcitonina y la sustancia P, llegando a
regular varias proteínas como el receptor de capsaicina, canales de sodio y
canales iónicos sensibles a ácidos (ASICs). De esta manera, los anticuerpos anti-
NGF pueden reducir la hiperalgesia y los cambios neurogénicos inducidos por el
factor de crecimiento neuronal y la inflamación (Sah et al, 2003; Pezet y Mc Mahon,
2006).
1.1.5.1.2. Modificaciones transcripcionales
Aparte de los cambios en la traducción de la señal, la sopa inflamatoria
puede inducir cambios en la transcripción de las fibras aferentes primarias.
Moléculas de señal son liberadas de las células inmunes, el prototipo es el factor
de crecimiento neuronal (NGF) que está sustancialmente regulado a la alta en la
inflamación. Una vez liberado se une a su receptor trk A, el complejo de unión se
internaliza y transporta retrógradamente al cuerpo celular donde inicia cambios en
la transcripción de genes por activación de una cascada de cinasas (Woolf, 1996;
Kidd et al 2004). Estos cambios a la larga refuerzan el estado sensibilizado de la
neurona y se regulan a la alta canales catiónicos (TRPV1, SNS y SNS-2)
aumentando la capacidad de despolarización.

25
Figura 6. Durante la inflamación se liberan diversas sustancias mediadoras que incluyen péptidos (bradicinina), lípidos (prostaglandinas), neurotransmisores (serotonina, 5-HT y adenosín trifosfato, ATP), neurotrofinas (NGF) y reducción de pH. Estos factores excitan o sensibilizan (disminuyen el umbral) las terminales de los nociceptores al interactuar con sus receptores en esas neuronas. Ejemplos de estos factores y sus blancos moleculares están indicados en el cuadro. La activación del nociceptor transmite el estímulo hacia centros superiores iniciando además la inflamación neurogénica en la que neurotransmisores como el péptido relacionado con el gen de la calcitonina (CGRP) y la substancia P inducen vasodilatación y extravasación, así como la activación de células no neuronales como mastocitos y neutrófilos, los cuales adicionan sustancias a la sopa inflamatoria (modificado de Julius y Basbaum, 2001).
La actividad eléctrica en los nociceptores por sí sola es capaz de cambiar los
niveles de transcripción de genes de las fibras aferentes primarias. Ejemplo de ello
es la estimulación directa del nervio ciático con un estimulador calibrado por 30
minutos que induce el aumento de los niveles de ARNm (ácido ribonucléico
mensajero) del factor neurotrófico derivado del cerebro (BDNF) en el ganglio de la
raíz dorsal luego de 2 horas. El mismo protocolo aplicado a una fibra Aβ no
aumenta la transcripción del BDNF (Mannion et al, 1999).

26
Durante el curso de la inflamación, las fibras A del ganglio de la raíz dorsal
comienzan a expresar neuropéptidos y neuromoduladores asociados a fibras C
(como la sustancia P y el factor neurotrófico derivado del cerebro), en términos de
sus acciones centrales este cambio de fenotipo hace que las fibras Aβ de bajo
umbral y las Aδ de alto umbral comiencen a comportarse como las fibras C (Baba
et al, 1999; Dubner, 2004). Estos eventos pueden ser considerados como una
respuesta fisiológica protectora ya que el sistema nociceptivo retorna a la
normalidad una vez que el daño tisular ha sido reparado. Sin embargo, en
condiciones crónicas puede producirse dolor espontáneo de carácter intermitente o
persistente que también involucra complejos cambios en el procesamiento
periférico y central de la señalización. En estas circunstancias, la relevancia
fisiológica de la nocicepción es menos clara y se pierde la función protectora de la
actividad de las fibras aferentes primarias (Dray et al, 1994; Kidd et al, 2004).
1.1.5.2. Tratamiento del dolor inflamatorio
En 1982, la organización mundial de la salud (OMS) y un panel de expertos
establecieron un método de fácil aplicación para el tratamiento del dolor
inflamatorio conocido como “escalera del dolor” (Figura 7). Este modelo de
tratamiento se ha aceptado en todo el planeta y consiste en tomar como primer
escalón el empleo de acetaminofén, aspirina u otros medicamentos anti-
inflamatorios no esteroideos (AINEs) para el dolor de leve a moderado. Los
fármacos adyuvantes pueden emplearse en todo momento para potenciar el efecto
analgésico. Si el dolor se incrementa se deben incluir opioides leves como codeína
e hidrocodona. Cuando éstos no produzcan alivio, deben sustituirse por opioides
potentes como la morfina (WHO, 1990; De Lima, 2004).

27
Figura 7. Recomendaciones de la OMS para el tratamiento del dolor inflamatorio (modificado de De Lima, 2004).
1.1.6. Dolor neuropático
El dolor neuropático es una condición crónica causada por lesión del sistema
nervioso. A diferencia del dolor agudo que es protector, el dolor neuropático
persiste y no tiene un propósito útil afectando severamente la calidad de vida de los
pacientes (Sah et al, 2003). El dolor neuropático es una consecuencia de varios
tipos de procesos patológicos que afectan a las neuronas del sistema nervioso
central o periférico (Zhuo, 2007). Este tipo de dolor es parte de un amplio rango de
padecimientos como pueden ser traumatismos, diabetes, neuralgia postherpética,
cáncer, etc. Los pacientes con dolor neuropático pueden presentar diferentes
subtipos de dolor (Field et al, 2002) por lo que en la clínica se considera muy difícil
de tratar. La IASP define al dolor neuropático como “el dolor iniciado o causado
por una lesión primaria o disfunción del sistema ne rvioso”. Esta definición
guía la atención clínica y la terapia del dolor neuropático e incluye tanto las
situaciones en las que existe lesión del nervio y las debidas a un trauma o
enfermedad (IASP, 1986).

28
Clasificar al dolor neuropático en base a su mecanismo o de acuerdo a
consideraciones anatómicas ha resultado controvertido porque no existe una
descripción uniforme de éste. El dolor neuropático comprende dolor espontáneo
estimulo-dependiente e independiente del estímulo caracterizado como
hiperalgesia y alodinia (Zhuo, 2007). Muchos pacientes que experimentan daño en
los nervios no desarrollan dolor neuropático. Sin embargo, quienes lo desarrollan
sufren síndromes severos de dolor anormal y de larga duración tras el daño neural
central o periférico. La susceptibilidad individual al padecimiento no está bien
comprendida actualmente por lo que es imposible predecir cuál paciente
desarrollará dolor neuropático, peor aún, la eficacia de un tratamiento en particular
no siempre está relacionada a su etiología. Así que diferentes causas y
mecanismos fisiológicos distintos pueden existir entre los diferentes pacientes
manifestando síntomas clínicos similares (Zimmermann, 2001).
1.1.6.1. Mecanismos del dolor neuropático
Como el dolor neuropático puede surgir desde varias etiologías y presenta
diferentes síntomas, se han propuesto diversos mecanismos para su
manifestación.
1.1.6.1.1. Aumento de descargas aferentes
El dolor neuropático comienza y se mantiene debido a descargas anormales
de alta intensidad de los potenciales de acción de los tejidos nerviosos. Estas
descargas promueven cambios plásticos en la médula espinal y centros superiores
que conducen a la recurrencia del dolor en lo que se conoce como “síndromes de
dolor regional complejo” (Bridges et al, 2001; Schaible, 2007). El aumento de
descargas aferentes desde focos ectópicos al sitio de lesión o al ganglio de la raíz
dorsal correlaciona con la hipersensibilidad táctil y térmica en los modelos
animales. Sin embargo, el aumento de la actividad disminuye rápido tras un
periodo de 3 a 8 días, mientras que el dolor neuropático permanece por meses tras
la lesión indicando que otros mecanismos contribuyen a su mantenimiento (Liu et

29
al, 2000). El cambio en la excitabilidad podría deberse a incrementos importantes
en la densidad de canales de Na+, principalmente resistentes a tetrodotoxina (Lai et
al, 2001).
Una consecuencia importante de la lesión al nervio y el aumento de las
descargas es el desarrollo de sensibilización espinal, que se presume como
resultado la liberación aumentada de aminoácidos excitatorios desde las neuronas
aferentes primarias hiperactivas, que activan a los receptores espinales de estos
aminoácidos provocando un aumento en la respuesta a los estímulos nociceptivos
(Devor, 1991; Schaible, 2007).
1.1.6.1.2. Facilitación descendente
Se ha sugerido que el aumento anormal del dolor tras una lesión es mediado
a través de la activación de procesos descendentes de facilitación que surgen de la
médula rostral ventromedial como resultado de cambios neuroplásticos tras la
lesión. Las células de “encendido” de la médula rostral ventromedial responden a
estímulos nocivos con aumento en sus disparos activando rutas de facilitación del
dolor (Tortoricci et al, 1994; Ossipov et al, 2000). Para algunos autores, la
presencia de la facilitación descendente desde la médula rostroventromedial es un
requisito para el mantenimiento de la sensibilización central (Vera-Portocarrero,
2006). Lesiones en el funículo dorsal ipsilateral (DLF) principal conducto de
proyección desde la médula rostral ventromedial así como inyecciones de
lidocaína en la médula rostral ventromedial en animales de experimentación son
capaces de bloquear el dolor neuropático establecido (Ossipov et al, 2000; Sah et
al, 2003). El aumento de los disparos en las células de encendido de la médula
rostral ventromedial induce a su vez una elevación de las concentraciones de
aminoácidos excitatorios, que puede promover la apoptosis de interneuronas
gabaérgicas, exacerbando el dolor neuropático por pérdida de los mecanismos de
inhibición central (Moore et al, 2002; Vera-Portocarrero, 2006).

30
1.1.6.1.3. Aumento de dinorfina espinal
Varios estudios han demostrado una asociación cercana entre la lesión de
fibras periféricas y una regulación a la alta de dinorfina espinal (Kajander, 1990; Lai
et al, 2006). El aumento de las concentraciones de dinorfina espinal (que llega a
ser de hasta 7 veces su concentración habitual), se desarrolla lentamente
alcanzando un nivel máximo a los 10 días lo que indica un posible papel de la
dinorfina espinal en el mantenimiento neuroplástico más que en el inicio del dolor
neuropático.
La dinorfina espinal facilita la liberación de sustancia P y el péptido
relacionado con el gen de la calcitonina evocados por capsaicina mediante un
mecanismo no opioide. Estos mediadores sensibilizan a los receptores NMDA y
aumentan la liberación de glutamato. La dinorfina también promueve un aumento
de las concentraciones de calcio que no es bloqueado por naloxona (Lai et al,
2003). Estudios recientes sugieren que la dinorfina A puede contribuir al
mantenimiento del dolor neuropático al unirse a receptores espinales de
bradicinina, ya que el empleo de antagonistas B1 y B2 revierten la persistencia de
los procesos alodínicos, siempre y cuando exista una elevación sostenida de
dinorfina (Lai et al, 2006).
1.1.6.1.4. Ramificación colateral
Debido a que el procesamiento sensorial de estimulación táctil ligero está
mediado normalmente por las fibras Aβ, una hipótesis que ha recibido atención es
la idea de que en respuesta a daño del nervio, esas fibras forman ramificaciones
colaterales que luego hacen sinapsis anormal en neuronas de segundo orden del
tracto espinotalámico (Woolf et al, 1995; Lekan et al, 1997; Macías et al, 2006). Sin
embargo, para otros autores la ramificación de las fibras propuestas Aβ no
corresponde temporalmente con el desarrollo y mantenimiento del dolor
neuropático tras la lesión al nervio. Además, se ha determinado que marcadores
inmuno-histológicos que se pensaba marcaban fibras Aβ específicamente, también

31
marcan fibras C tras lesión (Bao et al, 2002; Sah et al, 2003) por lo que esta
hipótesis puede resultar insatisfactoria.
1.1.6.1.5. Participación de factores neurotróficos
Otro fenómeno que está captando la atención de los investigadores es la
participación de factores neurotróficos en el desarrollo del dolor neuropático. Las
neurotrofinas ayudan a la supervivencia neuronal y el crecimiento durante el
desarrollo del sistema nervioso, mantienen la integridad estructural y funcional del
sistema nervioso adulto y regulan la plasticidad del sistema nervioso tras daño o
enfermedad. Está reportado que en tejidos dañados el aumento del transporte
retrógrado del factor de crecimiento neuronal concurre, dando como resultado un
incremento en los niveles de sustancia P y el péptido relacionado con el gen de la
calcitonina indicando que el factor de crecimiento neuronal periférico contribuye
sustancialmente al desarrollo del dolor inflamatorio y tal vez al neuropático (Lewin y
Mendell, 1993, Wild et al, 2007).
De igual manera se ha postulado que el factor neurotrófico derivado del
cerebro (BDNF) actúa como modulador del dolor central, ya que su liberación
promueve la fosforilación y potenciación de los receptores NMDA en las neuronas
espinales. Experimentos con anticuerpos anti BDNF han mostrado reducción de la
hiperalgesia táctil y térmica así como de la alodinia mecánica (Zhou et al, 2000;
Macías et al, 2006). De esta forma, Anticuerpos monoclonales humanos para estos
factores neurotróficos pueden tener potencial terapéutico en el tratamiento del dolor
neuropático (Wild et al, 2007).

32
Figura 8. Mecanismos celulares de dolor neuropático. La lesión a un nervio periférico produce descargas ectópicas espontáneas que se inician en el sitio dañado, en el ganglio de la raíz dorsal (DRG) o en fibras adyacentes; además el proceso es exacerbado por el aumento de las concentraciones de sustancias como la dinorfina, colecistocinina (CCK) y neurotrofinas. A su vez, cambios en el fenotipo celular de las neuronas en la médula rostroventromedial (RVM), promueven una facilitación descendente del proceso doloroso (modificado de Sah et al 2003).
1.1.6.2. Tratamiento del dolor neuropático
Actualmente no se cuenta con un tratamiento completamente satisfactorio para el
dolor neuropático. Sin embargo, la disminución del dolor, la recuperación
terapéutica del sueño, el mantenimiento de las funciones y el mejoramiento general
de la calidad de vida son los objetivos primordiales del manejo terapéutico (Galluzi,
2005). La primera dificultad en el tratamiento farmacológico de estos padecimientos
recae en la apreciación de que los analgésicos convencionales, opioides y AINEs
ofrecen un efecto terapéutico limitado. La segunda es que la respuesta al uso de
tratamientos antinociceptivos no convencionales no resulta lo suficientemente
eficaz (McQuay, 2001). Aunque en la actualidad los tratamientos se consideran
clínicamente significativos en tan sólo el cincuenta por ciento de los casos (Bridges
et al, 2001), se ha optado por recurrir al uso de otro tipo de fármacos,

33
antidepresivos y anticonvulsivantes, como adyuvantes para el manejo de este tipo
de dolor, con relativo éxito debido a su alta incidencia de efectos adversos (King,
2007). Gilron et al (2006) han descrito un algoritmo de atención básica para el
tratamiento del dolor neuropático en el que uno de los tratamientos de primera
elección, lo constituye la gabapentina (Figura 9).
Tratamiento del dolor neuropático
Figura 9. Algoritmo para el tratamiento del dolor neuropático (Modificado de Gilron et al,
2006).

34
1.2. Combinaciones analgésicas
El empleo de una combinación de analgésicos ofrece varios beneficios
potenciales comparado con un agente individual. La inclusión de analgésicos
combinados en una sola formulación farmacéutica, puede facilitar la prescripción y
el apego al tratamiento al reducir la cantidad de medicamentos que los pacientes
llegan a consumir para el manejo del dolor (Raffa, 2001). Combinar productos
farmacológicos con diferentes mecanismos de acción provee la cobertura de un
amplio espectro de tipos de dolor, al tiempo que ofrece la posibilidad de que la
acción de los fármacos se presente de manera superior a una forma aditiva, es
decir, sinérgica. Además, al ser empleadas dosis más pequeñas de cada
analgésico que conforma la combinación, puede resultar en una menor incidencia
de sus efectos adversos individuales, mejorando los perfiles de seguridad.
Las terapias de combinación son recomendadas ampliamente por diversas
organizaciones médicas, como la organización mundial de la salud (OMS) (Schug
et al 1990). Las ventajas de combinar analgésicos han sido reconocidas por más
de un siglo, sin embargo la evidencia experimental al respecto aún es escasa y
muchas veces inadecuada debido a las limitaciones en los diseños de los estudios
experimentales y de las continuas dificultades para encontrar modelos en los que
se pueda evaluar adecuadamente el dolor (Raffa, 2001). A su vez, medir las
interacciones entre los fármacos que se combinan también ha representado
dificultad pero el desarrollo de técnicas de modelado farmacológico y de nuevos
tipos de análisis estadístico para comparar de manera precisa la eficacia de las
combinaciones se han convertido en herramientas actuales de gran utilidad
(Tallarida, 1996). Combinar analgésicos de eficacia probada es una estrategia
adecuada para conseguir algunas ventajas terapéuticas como pueden ser el
facilitar el apego al tratamiento, simplificar la prescripción, mejorar la eficacia sin
elevar los efectos adversos o disminuir los efectos adversos sin pérdida de la
eficacia (Raffa, 2001).

35
La mayoría de las combinaciones resultan en un efecto analgésico
puramente aditivo. Sin embargo, en ciertos casos el resultado de las
combinaciones es un efecto analgésico inesperadamente superior al aditivo
(sinérgico) (Bolan et al, 2002). La presencia del sinergismo ha quedado
demostrada debido a la interacción entre analgésicos diferentes (Tallarida, 1999;
Argüelles y Granados-Soto 2005), enantiómeros del mismo fármaco (Raffa et al
1993) o empleando el mismo fármaco combinando diferentes vías de
administración (Raffa et al, 2000; Pozos-Guillen et al, 2006).
Un hecho consistente derivado de este tipo de estudios es que el sinergismo
es una propiedad que no sólo depende de la actividad intrínseca de los fármacos
constituyentes, sino que también depende de la proporción de cada fármaco en la
combinación. Otro hecho notable es que el sinergismo puede ocurrir en
combinaciones en las que uno de los constituyentes carece de eficacia como
agente individual.
1.2.1. Análisis isobolográfico
Una herramienta importante para discernir el tipo de interacción presente en
las combinaciones es el isobolograma (Tallarida, 2000). Este es un método gráfico
que recurre al empleo de pares de dosis equiefectivas para obtener un
determinado nivel de efecto (isobolas) y ofrece una evaluación visual del
comportamiento de la combinación al establecer una línea de aditividad que surge
del nivel de efecto determinado para cada fármaco individual (DE50, DE30, etc)
como puntos axiales de un plano cartesiano. La citada línea formada entre estos
dos puntos corresponde a todas las posibles combinaciones entre los dos fármacos
que teóricamente presentarían ese efecto en forma aditiva. Esta línea teórica de
aditividad permite la comparación con datos obtenidos al evaluar esos pares de
dosis de manera experimental, determinando si la combinación se comporta de
manera aditiva, antagónica o sinérgica (Figura 10).
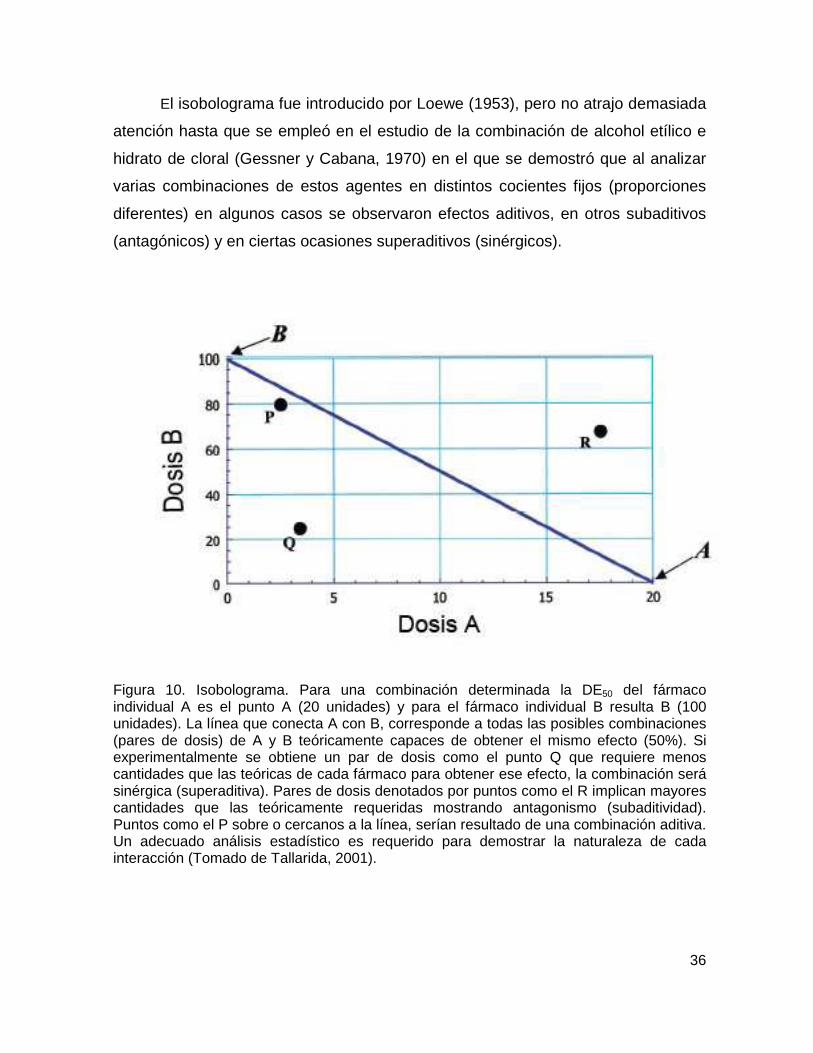
36
El isobolograma fue introducido por Loewe (1953), pero no atrajo demasiada
atención hasta que se empleó en el estudio de la combinación de alcohol etílico e
hidrato de cloral (Gessner y Cabana, 1970) en el que se demostró que al analizar
varias combinaciones de estos agentes en distintos cocientes fijos (proporciones
diferentes) en algunos casos se observaron efectos aditivos, en otros subaditivos
(antagónicos) y en ciertas ocasiones superaditivos (sinérgicos).
Figura 10. Isobolograma. Para una combinación determinada la DE50 del fármaco individual A es el punto A (20 unidades) y para el fármaco individual B resulta B (100 unidades). La línea que conecta A con B, corresponde a todas las posibles combinaciones (pares de dosis) de A y B teóricamente capaces de obtener el mismo efecto (50%). Si experimentalmente se obtiene un par de dosis como el punto Q que requiere menos cantidades que las teóricas de cada fármaco para obtener ese efecto, la combinación será sinérgica (superaditiva). Pares de dosis denotados por puntos como el R implican mayores cantidades que las teóricamente requeridas mostrando antagonismo (subaditividad). Puntos como el P sobre o cercanos a la línea, serían resultado de una combinación aditiva. Un adecuado análisis estadístico es requerido para demostrar la naturaleza de cada interacción (Tomado de Tallarida, 2001).

37
1.3. Gabapentina
La gabapentina es un medicamento generalmente utilizado para el
tratamiento de las convulsiones parciales (McNamara, 1988; Taylor, 1995) cuya
estructura consiste en una molécula de GABA ligada de manera covalente a un
anillo de ciclohexano lipófilo (Figura 11). Se diseñó para ser agonista GABA de
actividad central con una alta solubilidad en lípidos para cruzar la barrera
hematoencefálica. Sin embargo, se ha observado que la gabapentina no se une a
los receptores de GABA y su incremento de la descarga promovida por GABA es
un proceso aún no plenamente identificado.
Figura 11. Estructura del ácido-γ-amino-butírico GABA y de su análogo gabapentina.
Algunas investigaciones han demostrado que la gabapentina resulta exitosa
al prevenir la hiperalgesia en varios modelos animales de dolor (Sutton et al, 2002)
y en humanos está reportada su actividad anti-hiperalgésica en la atención clínica
de numerosas condiciones de dolor neuropático (Magnus, 1999). Más aún, la
gabapentina no presenta interacciones medicamentosas importantes, por lo que
resulta un fármaco recomendable para emplearse en combinaciones terapéuticas
(Tatum et al, 2000). En varios modelos la gabapentina produce actividad ansiolítica
en dosis mínimas (3-30 mg) cuyo efecto es comparable con el de las
benzodiacepinas.

38
La gabapentina se absorbe bien después de su administración oral y no
sufre metabolismo en humanos. Se excreta sin cambios en orina. Su vida media
cuando se administra como monoterapia es de 5 a 9 horas. La administración
concurrente de gabapentina no afecta las concentraciones plasmáticas de
fenitoína, carbamazepina, fenobarbital o valproato (McNamara, 2001).
1.3.1. Mecanismo de acción de gabapentina
El mecanismo de acción de la gabapentina se ha explorado ampliamente,
reportándose diversas maneras en las que puede lograr su efecto. La gabapentina
produce su efecto analgésico mediante la inhibición de la activación de canales de
Ca+2 dependientes de voltaje (CCDV) a través de la unión a su subunidad α2δ (Gee
et al, 1996; Stefani et al, 1998. Sutton et al, 2002). Pero también están planteadas
otras posibilidades, por ejemplo, la D-serina, un agonista del sitio de unión a glicina
de los receptores NMDA, es capaz de revertir el efecto antinociceptivo de la
gabapentina en la prueba de la formalina (Carlton y Zhou, 1998). De igual manera,
la gabapentina posee la capacidad de inhibir in vitro la transmisión glutamatérgica
sináptica en las láminas superficiales de la médula espinal, pero no en las
profundas, de ratas normales (Shimoyama et al 2000). La gabapentina también
tiene la capacidad de disminuir la síntesis del glutamato en tejido cerebral, al ser un
inhibidor competitivo de la aminotransferasa de aminoácidos ramificados (leucina,
isoleucina y valina) y de inhibir la liberación excesiva de glutamato, al menos in
vitro, al modular la liberación endógena de este aminoácido evocada por corrientes
de K+ (Dooley et al, 1999).
De la misma manera, está bien establecido que la gabapentina puede ser
capaz de conducir a la apertura de canales de potasio (dependientes de ATP y
activados por calcio) lo cual induce una hiperpolarización de las neuronas
sensoriales que reduce las conductas nociceptivas en animales de
experimentación (Freiman et al, 2001; Mixcoatl-Zecuatl et al, 2006).
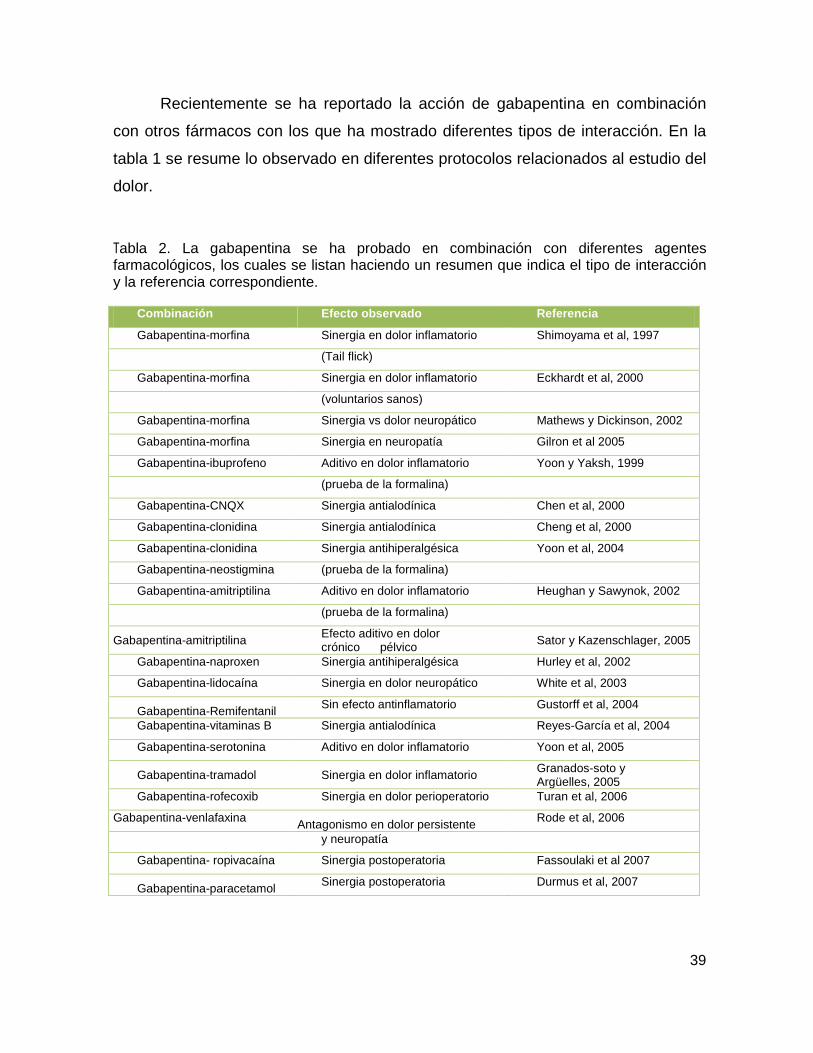
39
Recientemente se ha reportado la acción de gabapentina en combinación
con otros fármacos con los que ha mostrado diferentes tipos de interacción. En la
tabla 1 se resume lo observado en diferentes protocolos relacionados al estudio del
dolor.
Tabla 2. La gabapentina se ha probado en combinación con diferentes agentes farmacológicos, los cuales se listan haciendo un resumen que indica el tipo de interacción y la referencia correspondiente.
Combinación Efecto observado Referencia
Gabapentina-morfina Sinergia en dolor inflamatorio Shimoyama et al, 1997
(Tail flick)
Gabapentina-morfina Sinergia en dolor inflamatorio Eckhardt et al, 2000
(voluntarios sanos)
Gabapentina-morfina Sinergia vs dolor neuropático Mathews y Dickinson, 2002
Gabapentina-morfina Sinergia en neuropatía Gilron et al 2005
Gabapentina-ibuprofeno Aditivo en dolor inflamatorio Yoon y Yaksh, 1999
(prueba de la formalina)
Gabapentina-CNQX Sinergia antialodínica Chen et al, 2000
Gabapentina-clonidina Sinergia antialodínica Cheng et al, 2000
Gabapentina-clonidina Sinergia antihiperalgésica Yoon et al, 2004
Gabapentina-neostigmina (prueba de la formalina)
Gabapentina-amitriptilina Aditivo en dolor inflamatorio Heughan y Sawynok, 2002
(prueba de la formalina)
Gabapentina-amitriptilina Efecto aditivo en dolor crónico pélvico Sator y Kazenschlager, 2005
Gabapentina-naproxen Sinergia antihiperalgésica Hurley et al, 2002
Gabapentina-lidocaína Sinergia en dolor neuropático White et al, 2003
Gabapentina-Remifentanil Sin efecto antinflamatorio Gustorff et al, 2004
Gabapentina-vitaminas B Sinergia antialodínica Reyes-García et al, 2004
Gabapentina-serotonina Aditivo en dolor inflamatorio Yoon et al, 2005
Gabapentina-tramadol Sinergia en dolor inflamatorio Granados-soto y Argüelles, 2005
Gabapentina-rofecoxib Sinergia en dolor perioperatorio Turan et al, 2006
Gabapentina-venlafaxina Antagonismo en dolor persistente Rode et al, 2006
y neuropatía
Gabapentina- ropivacaína Sinergia postoperatoria Fassoulaki et al 2007
Gabapentina-paracetamol Sinergia postoperatoria Durmus et al, 2007

40
1.4. Metamizol
El metamizol es un analgésico anti-inflamatorio no esteroideo (AINE)
ampliamente utilizado en Europa y América Latina que tiene fuerte actividad
analgésica, antipirética y espasmolítica, pero escasa actividad anti-inflamatoria. A
pesar de ser una molécula sintetizada hace más de 100 años en 1897 (Levy,
2000), su mecanismo y sitio de acción, son aún materia de controversia
(Shimada,1994; Alves et al, 2002; Hinz et al, 2007).
Se presenta como un polvo blanco, inodoro, amargo, muy soluble en agua y
alcohol metílico e insoluble en éter. Su pH en solución acuosa es neutro y su pKa
es de 2.1. Las soluciones acuosas pueden colorearse de amarillo por exposición al
aire y la luz (Levy et al, 1985). Su denominación química es 2,3-dimetil-fenil-5
pirazolona 4-metilamino metanosulfonato sódico (aunque puede ser magnésico),
(figura 12). Su peso molecular es 336.3 y su fórmula condensada es: C13 H16 N3
SO3 . H20 Na.
Figura 12. Fórmula semidesarrollada del metamizol, un derivado de las pirazolonas.

41
1.4.1. Metabolitos del metamizol
El metamizol es un profármaco que se hidroliza espontáneamente de forma
no enzimática dando lugar a diversos metabolitos. Entre sus derivados
pirazolónicos activos el metabolito 4-metil-amino-antipirina es farmacológicamente
el más importante (Vlahov, 1990; Hinz et al, 2007). La 4-metil-amino-antipirina
puede transformarse a otros metabolitos como la 4 formil amino antipirina y la 4-
amino-antipirina. Esta última puede acetilarse para formar la 4-acetil-amino-
antipirina (Noda et al, 1976; Volz y Kellner, 1980; Pierre et al, 2007).
Figura 13. Principales metabolitos del metamizol. Fórmulas semidesarrolladas de la 4
metil amino antipirina (MAA) y la 4 amino pirina (AA).
4 metil amino antipirina 4 amino pirina

42
Tras su administración oral, intramuscular o intravenosa es muy difícil
detectar al metamizol en plasma como tal, excepto durante breves lapsos antes de
su paso por el hígado donde se metaboliza. In vitro, la vida media de degradación
del metamizol es de 16 minutos (Volz y Kellner, 1980). El metabolito principal del
metamizol, la 4 metil amino antipirina, provoca una inhibición marcada de las
ciclooxigenasas 1 y 2. In vitro, se ha reportado que no existe diferencia significativa
en la selectividad del fármaco hacia ambas enzimas, 2.5 vs 4.65 µmol/l
respectivamente, con base en sus concentraciones inhibitorias 50 (CI50) (Hinz et al,
2007). Opuesto a ello, otros estudios muestran que los valores in vivo de CI50 para
la ciclooxigenasa 1 en plaquetas humanas están muy por encima de sus
concentraciones farmacológicas (Campos et al, 1999). Que aunado al hecho de
que el metamizol contrarresta la hiperalgesia y el edema en el modelo de la
carragenina (Brune y Alperman, 1983; Lorenzetti y Ferreira 1985; Hinz y Brune,
2007), condiciones inflamatorias dependientes de las prostaglandinas derivadas de
la ciclooxigenasa 2, podrían otorgarle una relativa selectividad al metamizol para la
ciclooxigenasa 2. Sin embargo, un estudio reciente indica para el metamizol una
inhibición en voluntarios sanos similar para ambas isoformas (Ciclooxigenasas 1 y
2), mostrando una reducción de las prostaglandinas derivadas por la
ciclooxigenasa 1 parecida a la ejercida por varios analgésicos no esteroideos
tradicionales como la aspirina, haciendo hincapié en que el escaso daño
gastrointestinal inducido por el metamizol se debe más a factores fisicoquímicos (al
ser un AINE no acídico) que a su selectividad por la ciclooxigenasa 2 (Hinz et al,
2007).
Por otro lado, la actividad atípica del metamizol y sus derivados metabólicos
podría deberse a su acción en un sitio diferente al de los analgésicos no
esteroideos convencionales. Existe un reporte reciente que sugiere que tanto la 4-
metil-amino-antipirina como la 4-amino-antipirina inhiben la actividad de las
ciclooxigenasas al secuestrar radicales que son claves para el inicio de su acción
catalítica a través de la inducción de cambios en el estado de oxidación de las
proteínas Ciclooxigenasas (Pierre et al, 2007).

43
1.4.2. Empleo del metamizol
El metamizol se utiliza fundamentalmente como analgésico y antipirético ya
que posee un efecto anti-inflamatorio moderado. Existen reportes en la literatura
que muestran una actividad analgésica más potente del metamizol comparado con
aspirina (Lorenzetti y Ferreira, 1985). Se emplea sobre todo contra dolores de
intensidad moderada tanto agudos como crónicos. Además ejerce una acción
relajante de la fibra muscular lisa por lo que es útil en dolores de tipo cólico, sólo o
asociado a otros espasmolíticos o anticolinérgicos. Llega a emplearse incluso
contra malestares como la migraña y el cáncer (Levy et al, 1995; Hackental, 1997)
Para el tratamiento del dolor agudo puede administrarse por vía oral, intramuscular,
intravenosa o rectal. En el tratamiento del dolor agudo postoperatorio, se
administran 2g de manera endovenosa en intervalos de 6-8 horas (Ready, 1992).
1.4.3. Efectos adversos
Su acción sobre la mucosa gástrica es moderada, cuando se compara con
otros AINEs el riesgo relativo de hemorragia digestiva es similar al del paracetamol
(1.6 veces mayor que el ibuprofeno) y muy inferior al del diclofenaco y aspirina
(que resulta 7 veces mayor) o del piroxicam (19 veces mayor) (Henry et al, 1996).
Como el resto de los AINEs puede afectar al riñón por su inhibición de la síntesis
de prostaglandinas, lo que puede tener trascendencia clínica en aquellas
situaciones en que está comprometida la función renal. A dosis elevadas produce
hipotensión, efecto que podría estar en relación con la relajación de la fibra lisa
muscular (Shimada et al, 1994).
La mayor preocupación con el uso de metamizol son algunos reportes en la
literatura médica sobre casos de agranulocitosis y otras discracias sanguíneas
asociados con el uso de este medicamento. Se ha demostrado que el metamizol y
su metabolito activo 4-metil-amino-antipirina, al igual que la aspirina y el
diclofenaco, no afectan el proceso de diferenciación de los granulocitos ni inducen
apoptosis de los granulocitos ya diferenciados (García-Martínez et al, 2003). En

44
1986 se publicó el estudio epidemiológico más grande sobre el metamizol, el
llamado Estudio Boston o International Aplastic Anemia and Agranulocytosis Study
(IAAS, 1986). Este estudio arrojó una incidencia de tan sólo 1.2 casos de
agranulocitosis por un millón de personas expuestas al metamizol hasta por una
semana (Arcila-Herrera et al, 2004).
1.4.4. Mecanismo de acción de metamizol
Aunque el metamizol tiene una amplia prescripción a lo largo de varios
países como México, Brasil, Israel, Rusia y España, su mecanismo de acción
permanece elusivo (Hinz et al, 2007). El metamizol es un derivado de las
pirazolonas que bloquea el paso limitante en la síntesis de prostaglandinas, la
conversión de ácido araquidónico a prostaglandina H2 (Campos et al, 1999), la cual
es catalizada por varias isoenzimas llamadas ciclooxigenasas. La ciclooxígenasa 1
es una enzima constitutiva expresada en varios tejidos, mientras que la
ciclooxigenasa 2 es inducible y está asociada primordialmente con inflamación. De
manera adicional, recientemente se ha demostrado la existencia de la
ciclooxigenasa 3 (Chandrasekharan et al, 2002). Siendo el metamizol uno de los
fármacos cuya acción es capaz de inhibir a la ciclooxigenasa 3. La reducción en la
síntesis de prostaglandinas por acción del metamizol ha sido reportada tanto en el
sistema nervioso periférico como en el central (Volz y Kernel, 1980; Shimada et al,
1994).
El metamizol causa una inhibición dependiente de la dosis de la
hiperalgesia inducida por la inyección intratecal de glutamato en ratón,
involucrando su interacción, al menos en parte, con aminoácidos excitatorios
(Beirith et al 1998). De igual manera, la antinocicepción inducida por el metamizol
puede revertirse al inhibir la sintasa de óxido nítrico o la guanilato ciclasa soluble
sugiriendo que este fármaco es capaz de activar la vía óxido nítrico-GMPc-canales
de K+ (Duarte et al 1992; Granados-Soto et al, 1997), con la controvertida
participación de los canales de K+ sensibles a ATP (Alves y Duarte, 2002) o de los
canales de K+ activados por calcio y dependientes de voltaje (Ortíz et al, 2002) en

45
la reducción de las conductas nociceptivas en rata.
Pero el metamizol también tiene la capacidad de inducir antinocicepción en
ausencia de inflamación, por lo que se ha sugerido la participación de mecanismos
centrales en su actividad analgésica (Hernández y Vanegas, 2001).
Microinyecciones de metamizol en la sustancia gris periacueductal disminuyen la
respuesta de axones espinales ascendentes que activan de manera subsecuente a
fibras C periféricas, lo que produce una antinocicepción debida a los sistemas
descendentes opioides (Akman et al, 1996; Hernández y Vanegas, 2001) que
puede ser revertida por naloxona (Tortoricci y Vanegas, 2000). Recientemente,
registros de pulsos neuronales han determinado que actuando sobre la sustancia
gris periacueductal el metamizol tiene la capacidad de activar los sistemas
descendentes de control del dolor cuya función inhibitoria sobre las astas dorsales
es ejercida por medio de la médula rostral ventromedial, de forma que cuando la
inflamación y la sensibilización central están completamente desarrolladas la
acción central de este fármaco es suficiente para revertir esta hiperexcitabilidad y
presumiblemente la hiperalgesia y la alodinia (Vázquez et al, 2007).
El metamizol también se ha estudiado en combinación con distintos
fármacos en la búsqueda de la potenciación de su efecto analgésico. En la tabla 2
se muestra un resumen con los reportes más representativos.

46
Tabla 3. Se listan las interacciones farmacológicas de metamizol en combinación
con diferentes fármacos, en la búsqueda de la potenciación de su efecto analgésico.
Combinación Efecto observado Referencia
Metamizol-verapamilo Sinergia antihiperalgésica Vlaskovska et al, 1989
Metamizol-cafeína No hay potenciación Fialip et al, 1989
Metamizol-metadona Sinergia dolor perioperatorio Lehmann et al, 1990
Metamizol-morfina Sinergia en dolor visceral Taylor et al, 1998
Metamizol-morfina Sinergia antihiperalgésica Aguirre-Bañuelos y Granados-Soto, 1999
Metamizol-morfina Sinergía en dolor inflamatorio Hernández-Delgadillo et al, 2002
Metamizol-mianserina Sinergia en dolor inflamatorio Pakulska y Czarnecka, 2002
(tail-flick)
Metamizol-tramadol Sinergia en dolor postoperatorio Stamer et al, 2003
Metamizol-clonidina Dolor visceral, sinergia ip Miranda y Pinardi, 2004
efecto aditivo it

47
2. PLANTEAMIENTO DEL PROBLEMA
El dolor asociado a los padecimientos crónicos tiene importantes
implicaciones sociales tanto por la repercusión en la calidad de vida de los
pacientes, como por lo elevado de los costos directos e indirectos en el ámbito
asistencial (Bolten et al, 1998), lo que adquiere mayor relevancia si se
considera que este tipo de enfermedades están en permanente incremento. De
esta manera, el dolor como problema de salud pública exige la búsqueda de
mejores recursos terapéuticos para obtener tratamientos más racionales y
eficaces.
Entre los diferentes fármacos utilizados para aliviar el dolor, los más
relevantes en la clínica son los opioides que ofrecen el mejor efecto disponible
pero a la vez presentan más desventajas, entre las que sobresalen la falta de
efectividad contra el dolor neuropático, la depresión respiratoria, la
constipación, pero sobre todo el desarrollo de tolerancia (McQuay, 2001). Estas
limitantes han contribuido a un replanteamiento en el empleo de los
analgésicos basado en el concepto de “terapias de combinación” (Shimoyama
et al, 1997). Cuando dos o más fármacos se administran de manera simultánea
sus efectos pueden manifestarse de manera independiente sin la ocurrencia de
interacción (aditividad). En ocasiones los efectos observados pueden ser
diferentes a los esperados y la sinergia o el antagonismo pueden llegar a
presentarse (Raffa, 2001).
Las ventajas de combinar analgésicos no han sido lo suficientemente
reconocidas por la falta de estudios que evalúen adecuadamente este tipo de
interacciones farmacológicas. Sin embargo, cada vez se recurre con mayor
frecuencia al empleo de un método convencional, el isobolograma, que
consiste en establecer la relación dosis-respuesta de cada fármaco de manera
individual y de la combinación en cocientes fijos, para determinar gráficamente
el tipo de interacción presente en cada combinación farmacológica (Tallarida,
2000).

48
El metamizol es un AINE empleado comúnmente en Europa y América
Latina contra el dolor moderado a severo, por ser más potente que la aspirina y
el acetaminofén. La gabapentina es un anticonvulsivante del que se han
reportado efectos antinociceptivos en diferentes modelos en animales y en la
clínica se está convirtiendo en uno de los fármacos de elección contra el dolor
neuropático.
Ambos compuestos han mostrado efectos sinérgicos cuando han sido
coadministrados con otros analgésicos (Shimoyama et al 1997; Taylor et al,
1998; Stamer et al, 2003; Granados-Soto y Argüelles, 2005), pero no se tiene
registro de una terapia de combinación entre gabapentina y metamizol por lo
que explorar el efecto de su co-administración por diferentes vías de
administración podría implicar un novedoso recurso terapéutico contra el dolor
inflamatorio y neuropático.

49
3. HIPÓTESIS
La combinación de la gabapentina y el metamizol produce un efecto
analgésico superior al inducido por los fármacos individuales en modelos de
dolor inflamatorio y neuropático en la rata.

50
4. OBJETIVOS
4.1. Objetivo general
Evaluar el efecto antinociceptivo de la gabapentina y el metamizol
administrados de manera individual o en combinación por vía oral ó espinal en
modelos de dolor inflamatorio y neuropático en la rata.
4.1.1. Objetivos particulares
1. Evaluar la eficacia de la gabapentina y el metamizol por separado y en
combinación, en la vía oral y espinal con la prueba de formalina y el modelo de
Kim y Chung en la rata.
2. Determinar la interacción entre gabapentina y metamizol administrados por
vía oral o espinal en los citados modelos de dolor, mediante el análisis
isobolográfico.
3. Evaluar algunos de los posibles mecanismos de la combinación
gabapentina-metamizol.

51
5. MATERIAL Y MÉTODOS
5.1. Animales de experimentación
Se utilizaron ratas Wistar hembra de 7 (140-160 g) y 10 semanas (180-220
g) provenientes del bioterio del Departamento de Farmacobiología del Cinvestav,
Sede Sur. Las ratas se mantuvieron en un ciclo de 12 horas luz/oscuridad, con
acceso a agua y alimento ad-libitum, salvo en los experimentos de formalina en
que el alimento se retiró 8 horas antes de las evaluaciones. Todos los
experimentos se llevaron a cabo de acuerdo a los lineamientos éticos para la
investigación del dolor en animales conscientes (Zimmermann, 1983). Cada rata
se evaluó sólo una vez y se sacrificó en una cámara con dióxido de carbono al
final del experimento. De manera adicional, el comité institucional de uso y
cuidado de los animales (Cinvestav) aprobó el estudio.
5.2. Fármacos
La gabapentina fue un regalo de Merck S.A. de C.V. (México, D.F.). El
metamizol (dipirona), la glibenclamida y la naltrexona (clorhidrato de naltrexona)
se adquirieron en Sigma (St. Louis MO, EUA). El NG-L-nitro-arginina- metil-éster
(L-NAME) se compró en RBI (Natick, MA, EUA). La ketamina se adquirió en Galen
(México, D.F.) y la xilazina en Fort Dodge (lowa, EUA). Se prepararon soluciones
de gabapentina, metamizol, L-NAME y naltrexona con solución salina al 0.9%
obtenida de Abbott (México, D.F.) como vehículo. El dimetilsulfóxido (DMSO) fue
el vehículo empleado en las soluciones de glibenclamida.
5.3. Modelo inflamatorio
5.3.1. La prueba de la formalina
La nocicepción se evaluó mediante la prueba de la formalina (Wheeler-
Aceto y Cowan, 1991). Los animales de experimentación se colocaron en una caja
de acrílico transparente que funge como cámara de observación. Primeramente se
sometieron a un periodo de adaptación por 30 minutos, tras el cual se retiraron

52
para la administración intraplantar de 50 µl de formalina al 1% con una aguja del
número 30 en la cara dorsal de la pata posterior derecha. Las ratas se colocaron
nuevamente en la cámara, que contiene espejos en la parte de atrás, para que
puedan ser observadas desde todos los ángulos (Figura 14). Se realizó una
observación inmediata en la que se cuantificó durante un minuto el número de
sacudidas de la pata inyectada y se repitió el proceso cada 5 minutos hasta
completar una hora (Figura 15).
Figura 14. Modelo de la formalina. La administración subcutánea de formaldehído al 1% en la cara dorsal de la pata posterior derecha de las ratas induce una conducta de sacudidas que son evaluadas por periodos de un minuto cada 5 minutos hasta completar una hora. La reversión del número de sacudidas tras la previa administración de un analgésico se interpreta como antinocicepción.
Las sacudidas se caracterizaron como flexiones breves y rápidas de la pata
inyectada. Asimismo, se ha reportado que este comportamiento inducido por
formalina es de carácter bifásico, comprendiendo una fase aguda (0-10 min), un
breve periodo de quiescencia y una respuesta tónica prolongada (15-60 min). La
fase inicial se relaciona al estímulo nociceptivo directo y se conoce como
neurogénica, la segunda fase se denomina inflamatoria pues responde al proceso
inflamatorio subsecuente (Figura 16).
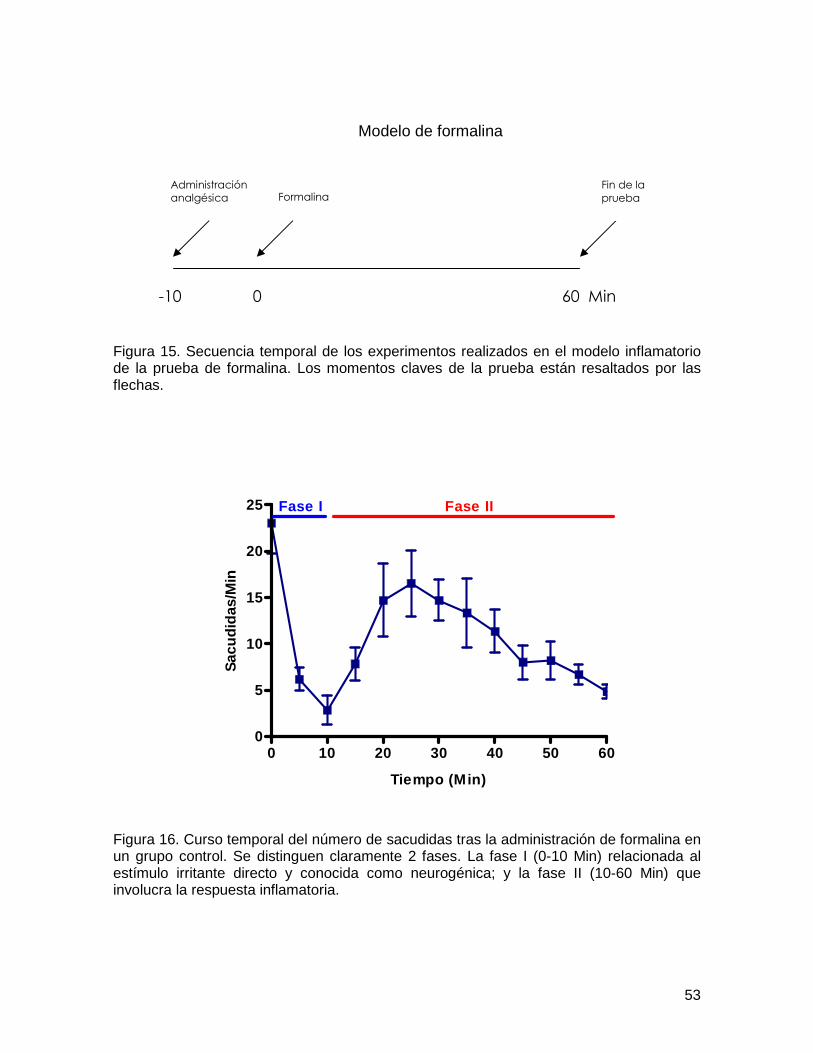
53
Modelo de formalina
Figura 15. Secuencia temporal de los experimentos realizados en el modelo inflamatorio de la prueba de formalina. Los momentos claves de la prueba están resaltados por las flechas.
0 10 20 30 40 50 600
5
10
15
20
25 Fase I Fase II
Tiempo (Min)
Sac
udid
as/M
in
Figura 16. Curso temporal del número de sacudidas tras la administración de formalina en un grupo control. Se distinguen claramente 2 fases. La fase I (0-10 Min) relacionada al estímulo irritante directo y conocida como neurogénica; y la fase II (10-60 Min) que involucra la respuesta inflamatoria.
-10 0 60 Min
Administración analgésica Formalina
Fin de la
prueba

54
5.4. Modelo de neuropatía
Para la inducción de la neuropatía se recurrió al empleo del procedimiento
quirúrgico descrito por Kim y Chung (1992). Las ratas se anestesiaron con una
mezcla de ketamina/xilazina (45-12 mg/kg, ip). Tras la preparación quirúrgica y
exposición de la columna vertebral dorsal, los nervios espinales izquierdos L5 y L6
fueron expuestos y firmemente ligados con sutura de seda 6-0 distal al ganglio de
la raíz dorsal, próximo a la formación del nervio ciático (Figuras 17 y 18).
Figura 17. Cirugía de Kim y Chung. La ligadura de los nervios espinales se lleva a cabo con ayuda de un microscopio quirúrgico.
En los animales del grupo control, los nervios de las ratas operadas se
expusieron pero no se ligaron. Se cerraron las incisiones y los animales
dispusieron de 12 días para recuperarse de la cirugía. Las ratas que presentaron
deficiencias motoras se descartaron de las pruebas.
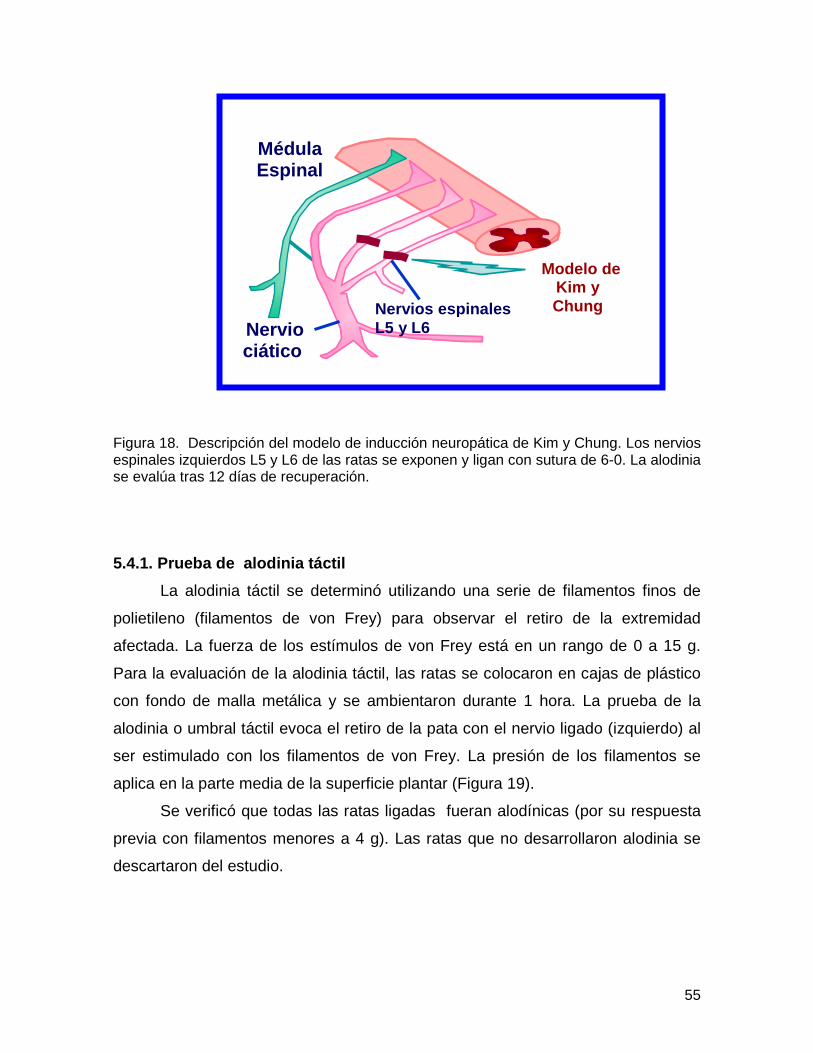
55
Figura 18. Descripción del modelo de inducción neuropática de Kim y Chung. Los nervios espinales izquierdos L5 y L6 de las ratas se exponen y ligan con sutura de 6-0. La alodinia se evalúa tras 12 días de recuperación.
5.4.1. Prueba de alodinia táctil
La alodinia táctil se determinó utilizando una serie de filamentos finos de
polietileno (filamentos de von Frey) para observar el retiro de la extremidad
afectada. La fuerza de los estímulos de von Frey está en un rango de 0 a 15 g.
Para la evaluación de la alodinia táctil, las ratas se colocaron en cajas de plástico
con fondo de malla metálica y se ambientaron durante 1 hora. La prueba de la
alodinia o umbral táctil evoca el retiro de la pata con el nervio ligado (izquierdo) al
ser estimulado con los filamentos de von Frey. La presión de los filamentos se
aplica en la parte media de la superficie plantar (Figura 19).
Se verificó que todas las ratas ligadas fueran alodínicas (por su respuesta
previa con filamentos menores a 4 g). Las ratas que no desarrollaron alodinia se
descartaron del estudio.
Nervios espinales L5 y L6 Nervio
ciático nerve
Médula Espinal
Modelo de Kim y Chung

56
Figura 19. Prueba de alodinia táctil. Los animales se colocan en contenedores de acrílico con una malla de acero inoxidable como fondo y se evalúan con filamentos de von Frey. Los filamentos están graduados logarítmicamente en gramos de presión. El retiro de la pata por acción del filamento se considera como respuesta positiva.
Prueba de alodinia táctil
Figura 20. Secuencia del proceso de las series experimentales efectuadas para la prueba de alodinia táctil. El experimento duró 4 horas. Cada flecha indica la realización de una medición con los filamentos de von Frey.
0 15 30 60 90 120 150 180 210 240 Min
Administración
analgésica
Fin de la
prueba
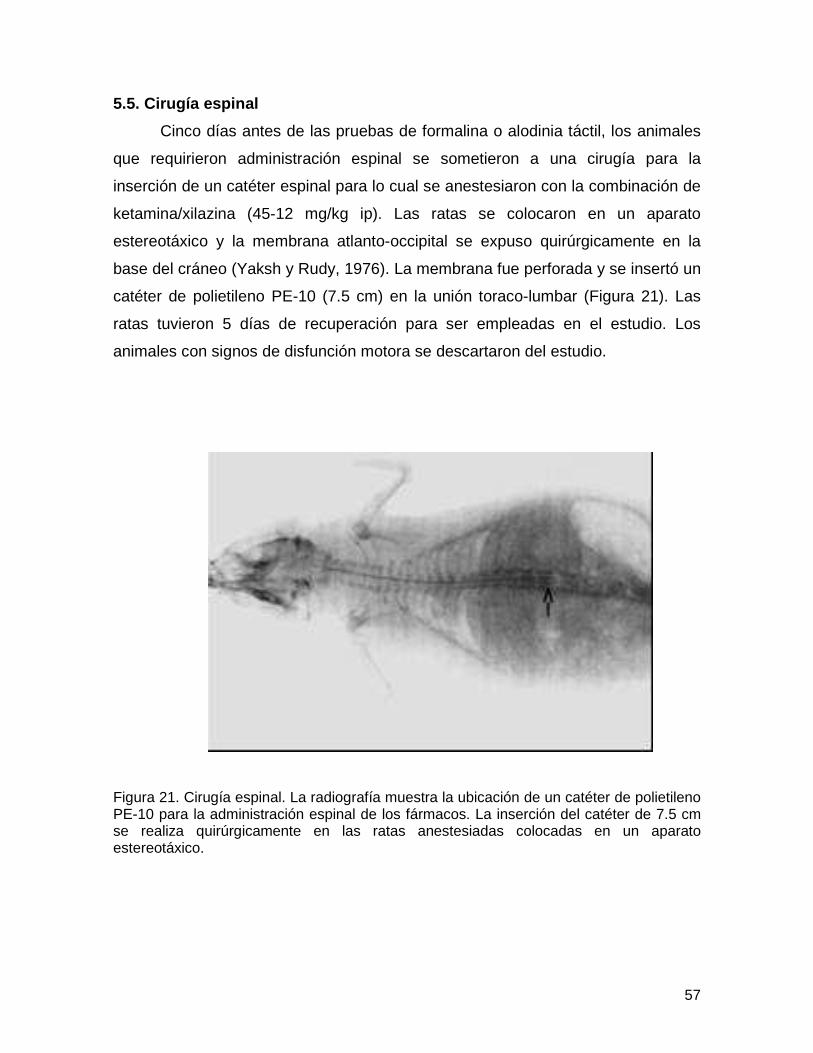
57
5.5. Cirugía espinal
Cinco días antes de las pruebas de formalina o alodinia táctil, los animales
que requirieron administración espinal se sometieron a una cirugía para la
inserción de un catéter espinal para lo cual se anestesiaron con la combinación de
ketamina/xilazina (45-12 mg/kg ip). Las ratas se colocaron en un aparato
estereotáxico y la membrana atlanto-occipital se expuso quirúrgicamente en la
base del cráneo (Yaksh y Rudy, 1976). La membrana fue perforada y se insertó un
catéter de polietileno PE-10 (7.5 cm) en la unión toraco-lumbar (Figura 21). Las
ratas tuvieron 5 días de recuperación para ser empleadas en el estudio. Los
animales con signos de disfunción motora se descartaron del estudio.
Figura 21. Cirugía espinal. La radiografía muestra la ubicación de un catéter de polietileno PE-10 para la administración espinal de los fármacos. La inserción del catéter de 7.5 cm se realiza quirúrgicamente en las ratas anestesiadas colocadas en un aparato estereotáxico.

58
5.6. Diseño del estudio
5.6.1. Eficacia individual y de la combinación
Se emplearon los mismos rangos de dosis para ambos paradigmas
experimentales, el inflamatorio y el neuropático. Las ratas recibieron por vía oral
(4 ml/kg del fármaco) gabapentina o metamizol en dosis crecientes (30, 100, 300 y
600 mg/kg) o el mismo volumen de vehículo (salina). En la vía espinal, los
animales recibieron solución salina (vehículo) o dosis crecientes de los fármacos
gabapentina (25, 50, 100 y 200 µg en 10 µl) y metamizol (50, 100, 200 y 400 µg en
10 µl). Las dosis de las combinaciones partieron del cálculo de las respectivas
DE30 de los fármacos en cada esquema experimental, empleándose fracciones de
éstas calculadas por el análisis isobolográfico. Este valor (DE30) se tomó como
base para los isobologramas en lugar de la DE50 en virtud de que en un estudio
piloto el metamizol no siempre tuvo un valor máximo por encima del 50% de
efecto.
Para el caso del modelo inflamatorio, las administraciones se efectuaron 10
minutos antes de la inyección de formalina al 1%. En lo referente al modelo de
neuropatía, se realizó una prueba de alodinia táctil como control y la siguiente se
efectuó 15 minutos después de la administración farmacológica.
Se emplearon grupos de seis animales por grupo para realizar las curvas
dosis-respuesta de metamizol y gabapentina de manera individual y con base en
esos datos la curva dosis-respuesta de cada combinación (Figuras 22 y 23). Sin
embargo, como se detallará en la sección de resultados, los datos obtenidos por
vía oral en la prueba de alodinia táctil, obligaron a efectuar una estrategia diferente
al isobolograma tradicional.
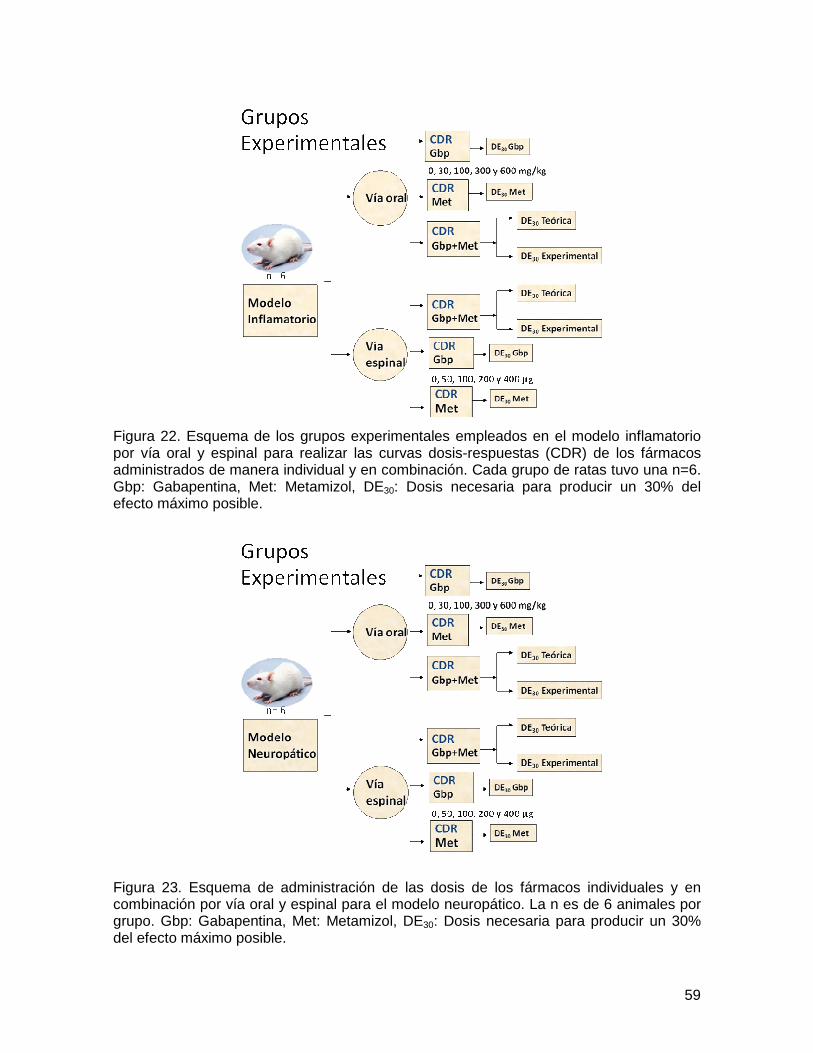
59
Figura 22. Esquema de los grupos experimentales empleados en el modelo inflamatorio por vía oral y espinal para realizar las curvas dosis-respuestas (CDR) de los fármacos administrados de manera individual y en combinación. Cada grupo de ratas tuvo una n=6. Gbp: Gabapentina, Met: Metamizol, DE30: Dosis necesaria para producir un 30% del efecto máximo posible.
Figura 23. Esquema de administración de las dosis de los fármacos individuales y en combinación por vía oral y espinal para el modelo neuropático. La n es de 6 animales por grupo. Gbp: Gabapentina, Met: Metamizol, DE30: Dosis necesaria para producir un 30% del efecto máximo posible.

60
5.7. Mecanismo de acción
Con el fin de obtener información acerca del posible mecanismo de acción
de la combinación farmacológica en cuestión, se recurrió al empleo de algunos
inhibidores de las vías analgésicas más comunes. De esta forma, la DE30 obtenida
de manera experimental para la combinación se aplicó a las ratas por vía espinal
en el modelo de dolor neuropático 10 minutos después de la administración de los
siguientes inhibidores:
a) Naltrexona (50 µg; antagonista no selectivo de los receptores opioides)
para la vía opioide.
b) L-NAME (50 µg; inhibidor inespecífico de la sintasa de ON) para la vía
del óxido nítrico/GMPc.
c) Glibenclamida (50 µg; bloqueador de los canales de K+ sensibles a ATP)
para analizar la influencia de los canales de potasio sensibles a ATP.
Prueba de alodinia táctil
Figura 24. Secuencia temporal de las series experimentales de alodinia táctil que involucraron pretratamiento con inhibidores para analizar los posibles mecanismos de acción de la combinación. Cada flecha indica la realización de una evaluación de alodinia empleando los filamentos de von Frey.
-10 0 15 30 60 90 120 150 180 210 240 Min
Administración
analgésica Registros de alodinia táctil
Fin de la
prueba
Pretratamiento
inhibidores

61
Los tres inhibidores empleados se administraron en una concentración de
50 µg. La dosis de los inhibidores se obtuvo de estudios previos realizados en el
laboratorio (Mixcoatl-Zecuatl et al, 2004; Mixcoatl-Zecuatl et al, 2006; Ambríz-
Tututi y Granados-Soto, 2007).
Figura 25. Esquema de administración de la exploración del mecanismo antialodínico espinal de la combinación, tras el pretratamiento con los inhibidores, se administró una dosis alta de los fármacos individuales o en combinación para evaluar su capacidad de revertir la alodinia. DE30: Dosis necesaria para producir un 30% del efecto máximo posible. La n es de 6 animales por grupo.
5.8. Análisis de datos
En todos los casos los datos se presentan como el valor de la media ± el
error estándar de 6 animales por grupo.
5.8.1. Análisis de los datos de formalina
Se construyeron curvas del número de sacudidas/minuto en función del
tiempo (Figura 16). De los cursos temporales anteriores se calculó el área bajo la
curva mediante el método del trapezoide y se consideró como una expresión
global de la intensidad y duración del efecto. Las áreas bajo la curva se
transformaron a porcentaje del efecto máximo posible (%EMP) en las curvas

62
dosis-respuesta, éstas expresan el porcentaje de inhibición de sacudidas con
respecto al control del periodo en particular (fase I o II) calculado con la fórmula
siguiente:
X = Área bajo la curva del promedio de sacudidas en el grupo control. Xi = Área bajo la curva del promedio de sacudidas en el tratamiento respectivo.
Un valor cercano al 0% indica que el número de sacudidas en el grupo
tratado es igual al grupo control, es decir no hay efecto antinociceptivo y 100%
significa que las ratas no sacudieron la pata.
5.8.2. Análisis de los datos de alodinia táctil
Se determinó el 50% de umbral de retiro empleando el método de Dixon
(1980). Este valor se define como la intensidad de estímulo requerido para
producir una respuesta en 50% de las aplicaciones para cada animal. En el
paradigma arriba-abajo, la prueba se inició con el filamento de 4.31 g, a la mitad
de la serie de 20 filamentos de von Frey con incrementos logarítmicos de rigidez o
dureza; ascendiendo o descendiendo en función de la respuesta del animal
(Dixon, 1980; Chaplan et al, 1994).
Se midió el umbral de retiro tomando como respuesta positiva el retiro de la
pata estimulada en un lapso de 10 segundos, denotándose con un símbolo “x”. La
respuesta negativa es aquella en la que al estimular la pata ésta no es retirada y
fue registrada con un símbolo “o”. Después del primer cambio se llevó a cabo la
estimulación otras 4 veces, tomando una serie de 6 patrones de respuesta y el
último filamento utilizado. La estimulación se llevó a cabo a tiempos de 0, 15, 30,
45, 60, 90, 120, 150, 180 y 240 minutos después de administrar los fármacos
(Figura 20). Los patrones de resultado de respuesta se tabularon y el 50% de
umbral de respuesta se calculó usando la fórmula:

63
Donde:
Xf: Valor del último filamento de von Frey usado (en unidades log)
k: Factor de corrección basado en los patrones de respuesta de la tabla de calibración y al valor tabulado en base al patrón de respuestas positivas y negativas
δδδδ: Diferencia de promedio entre estímulos (unidades log)
5.8.3. Análisis isobolográfico
Este método está basado en la selección particular de un nivel de efecto,
que usualmente es el 50% del efecto máximo, aunque pueden emplearse distintos
niveles de efecto si son obtenidos por cada fármaco (Tallarida et al, 1997). En el
presente trabajo el nivel elegido fue el de 30%, por lo que las dosis efectivas 30
(DE30) tanto de gabapentina como de metamizol se determinaron a través de
curvas dosis respuesta. Una combinación aditiva correspondiente al efecto
determinado se constituye con 2 partes que aportan una fracción de dicho efecto
determinado. Para nuestro caso se eligió que cada fracción fuera de 0.5 de forma
que la dosis efectiva 30 teórica de la combinación elegida (DE30T) contiene un 0.5
de la DE30 de gabapentina y el restante 0.5 de la DE30 de metamizol.
DE30T = (0.5) (DE30 de gabapentina) + (0.5) (DE30 de metamizol)
Posteriormente se crea una curva dosis respuesta experimental que
contiene combinaciones de ambos fármacos en la proporción establecida y se
calcula la dosis correspondiente al valor determinado del 30% (DE30E), que luego
se compara con la DE30T. El isobolograma muestra los resultados en una gráfica
cuyas coordenadas representan la contribución de cada fármaco (Figura 26). La
línea que conecta como interceptos a las DE30 de gabapentina y metamizol
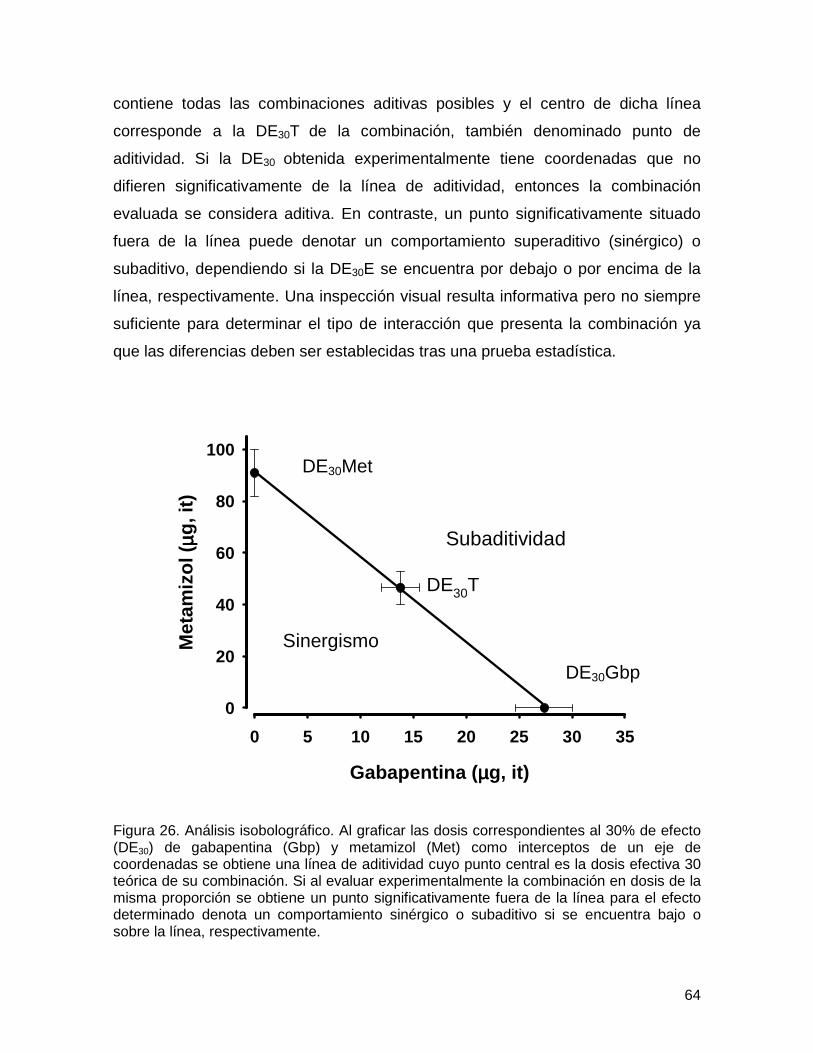
64
contiene todas las combinaciones aditivas posibles y el centro de dicha línea
corresponde a la DE30T de la combinación, también denominado punto de
aditividad. Si la DE30 obtenida experimentalmente tiene coordenadas que no
difieren significativamente de la línea de aditividad, entonces la combinación
evaluada se considera aditiva. En contraste, un punto significativamente situado
fuera de la línea puede denotar un comportamiento superaditivo (sinérgico) o
subaditivo, dependiendo si la DE30E se encuentra por debajo o por encima de la
línea, respectivamente. Una inspección visual resulta informativa pero no siempre
suficiente para determinar el tipo de interacción que presenta la combinación ya
que las diferencias deben ser establecidas tras una prueba estadística.
Gabapentina ( µµµµg, it)
0 5 10 15 20 25 30 35
Met
amiz
ol (
µµ µµg, i
t)
0
20
40
60
80
100
DE30T
Sinergismo
Figura 26. Análisis isobolográfico. Al graficar las dosis correspondientes al 30% de efecto (DE30) de gabapentina (Gbp) y metamizol (Met) como interceptos de un eje de coordenadas se obtiene una línea de aditividad cuyo punto central es la dosis efectiva 30 teórica de su combinación. Si al evaluar experimentalmente la combinación en dosis de la misma proporción se obtiene un punto significativamente fuera de la línea para el efecto determinado denota un comportamiento sinérgico o subaditivo si se encuentra bajo o sobre la línea, respectivamente.
DE30Met
DE30Gbp
Subaditividad

65
5.8.4. Análisis estadístico
Los datos de las curvas dosis respuesta se compararon con sus respectivos
controles por análisis de varianza de una vía, seguido por la prueba de Dunnett
para comparar las diferencias con los diferentes tratamientos. La significancia
estadística entre la DE30 teórica y la experimental se determinó mediante una
prueba de t de “Student”. En los experimentos de exploración del mecanismo de
acción de la combinación se utilizó t de “Student”. En todos los casos, el nivel de
significancia elegido fue p < 0.05.

66
6. RESULTADOS 6.1. Modelo Inflamatorio
La inyección de formalina produjo la conducta nociceptiva típica
previamente descrita. El curso temporal mostró una respuesta bifásica que
incluye una conducta nociceptiva inmediata o fase I (0-10 minutos) y después de
un breve periodo quiescente, otra que inició de manera gradual y continuó durante
todo el periodo de observación (fase II).
La figura 27 muestra el curso temporal de las sacudidas por minuto del
grupo control y de las dosis más altas de gabapentina y metamizol empleadas en
este estudio por vía oral y espinal. La administración oral (Figura 27A) y espinal
(Figura 27B) de gabapentina y metamizol redujo el número de sacudidas inducidas
por la inyección de formalina, lo que se interpreta como antinocicepción.
6.1.1. Efecto de gabapentina, metamizol y su combinación por vía oral en el modelo de formalina
Al administrar los fármacos por vía oral en el modelo inflamatorio se
observó un efecto antinociceptivo en la fase I que no fue dependiente de la dosis.
Para la gabapentina se utilizaron por vía oral dosis de 10-300 mg/kg que
resultaron diferentes del grupo control en ambas fases de la prueba. El mayor
efecto observado en la fase I fue de 27% ± 4.2. En contraste, la gabapentina
redujo la nocicepción en la fase II de manera dependiente de la dosis registrando
una eficacia de hasta 53.1% ± 3.5 (Figura 28).
En lo referente a la curva dosis respuesta del metamizol, se emplearon por
vía oral dosis de 30-600 mg/kg. Todos los tratamientos en ambas fases resultaron
diferentes del vehículo (solución salina). Sin embargo, en la fase I no se observó
una relación directamente proporcional entre la dosis y el porcentaje de
antinocicepción, alcanzándose un efecto similar para las cuatro dosis evaluadas,
que fue del 21.4% (300 mg/kg) al 26.4% (600 mg/kg). La fase II de la prueba
mostró un efecto antinociceptivo máximo para el metamizol oral de 42.9% ± 4.1 y
los efectos observados fueron dependientes de la dosis (Figura 29).
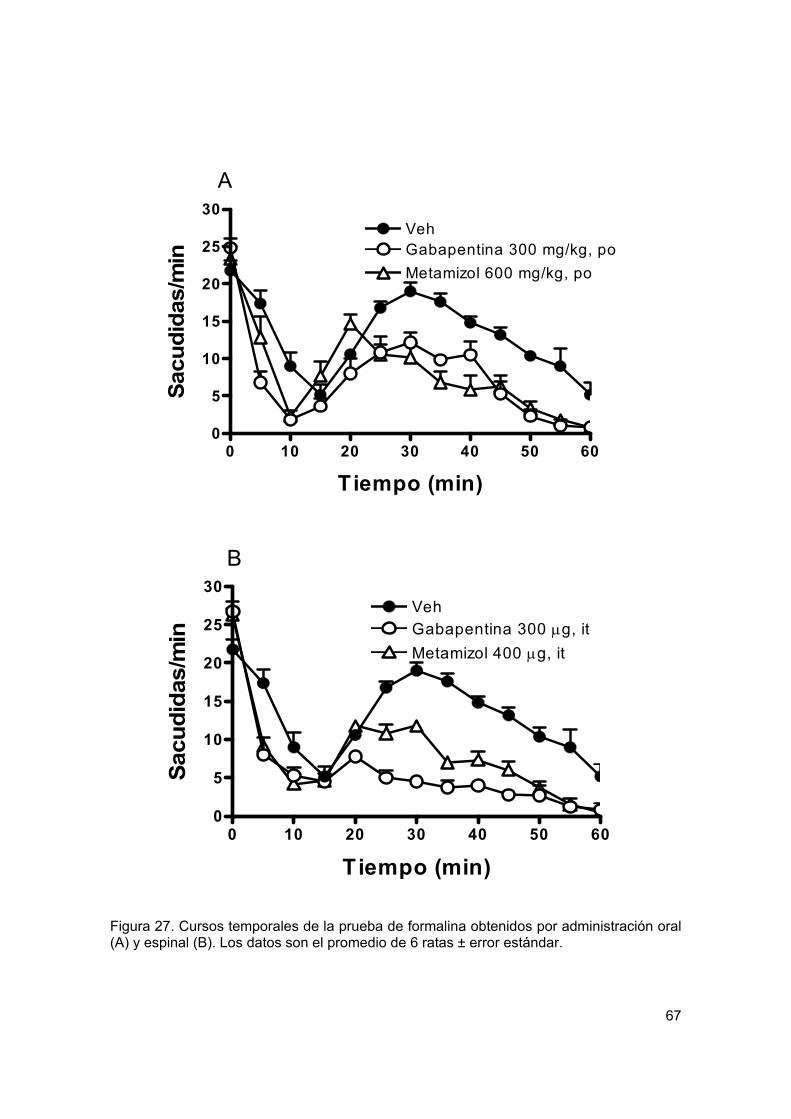
67
Figura 27. Cursos temporales de la prueba de formalina obtenidos por administración oral (A) y espinal (B). Los datos son el promedio de 6 ratas ± error estándar.
0 10 20 30 40 50 600
5
10
15
20
25
30VehGabapentina 300 mg/kg, poMetamizol 600 mg/kg, po
A
Tiempo (min)
Sacu
dida
s/m
in
0 10 20 30 40 50 600
5
10
15
20
25
30VehGabapentina 300 µg, itMetamizol 400 µg, it
B
Tiempo (min)
Sacu
dida
s/m
in

68
Los valores obtenidos a partir de los experimentos de gabapentina y
metamizol administrados individualmente por vía oral se emplearon para el
análisis isobolográfico. Se obtuvieron los valores de las respectivas DE30 de cada
fármaco en la fase II y se establecieron las dosis de cada uno para efectuar la
curva dosis respuesta de la combinación en una proporción de uno a uno (0.5:0.5).
Las dosis empleadas corresponden a fracciones de la proporción establecida y se
detallan en la tabla 3. Luego, las dosis de la combinación oral calculadas por el
isobolograma se probaron experimentalmente en el modelo de la formalina. La
figura 30 describe los resultados en ambas fases de la prueba. El efecto de la
combinación durante la fase I no fue dosis dependiente y el porcentaje de
antinocicepción máximo fue de 25.6%. En la segunda fase de la prueba se
registraron efectos dosis dependientes y la dosis de la combinación que
teóricamente estaba calculada para el efecto del 30%, alcanzó experimentalmente
un efecto del 56.3% (Figura 30).
La dosis efectiva 30 teórica (DE30T) de la combinación fue de 78.8 mg/kg,
mientras que el de la DE30E fue de 15 mg/kg que al ser diferentemente
significativo del teórico muestra que la combinación resulta sinérgica por esta vía
de administración. El índice de interacción resultante fue de 0.19 lo que muestra
que la dosis de la combinación fue 5 veces menor a lo calculado (Figura 31).
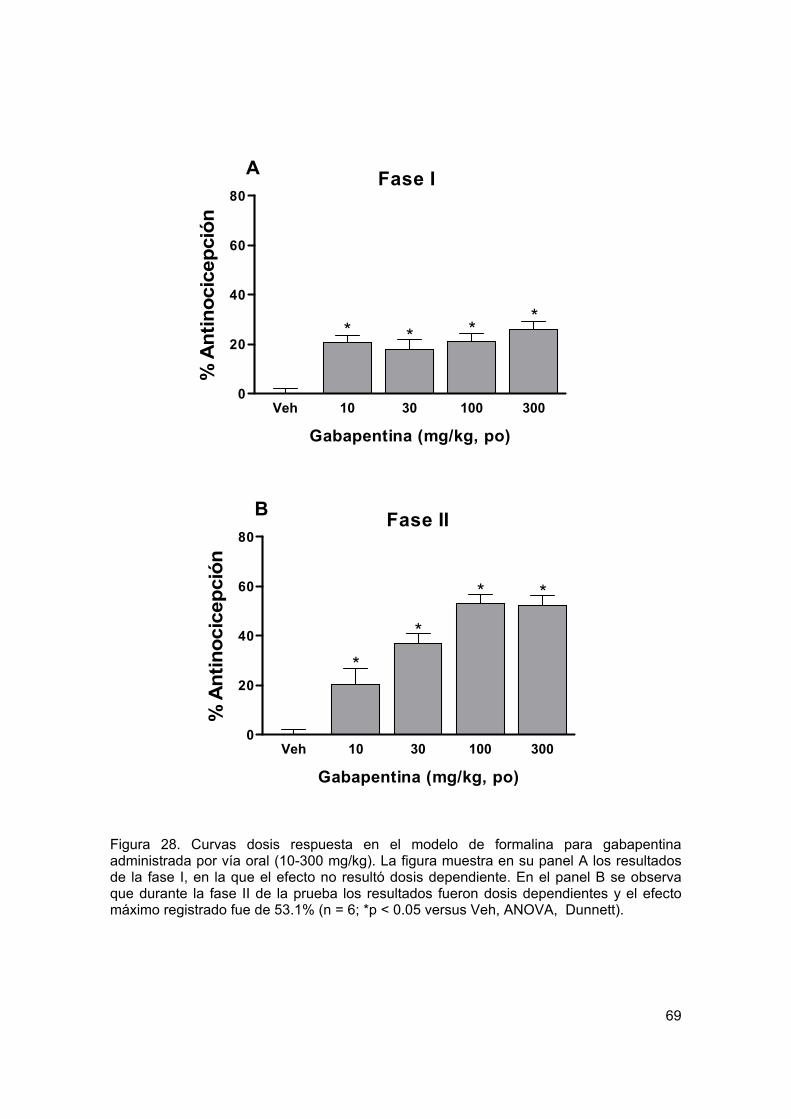
69
Figura 28. Curvas dosis respuesta en el modelo de formalina para gabapentina administrada por vía oral (10-300 mg/kg). La figura muestra en su panel A los resultados de la fase I, en la que el efecto no resultó dosis dependiente. En el panel B se observa que durante la fase II de la prueba los resultados fueron dosis dependientes y el efecto máximo registrado fue de 53.1% (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 10 30 100 3000
20
40
60
80
*
*
* *
Fase IIB
Gabapentina (mg/kg, po)
% A
ntin
ocic
epci
ón
Veh 10 30 100 3000
20
40
60
80
* * **
Fase IA
Gabapentina (mg/kg, po)
% A
ntin
ocic
epci
ón
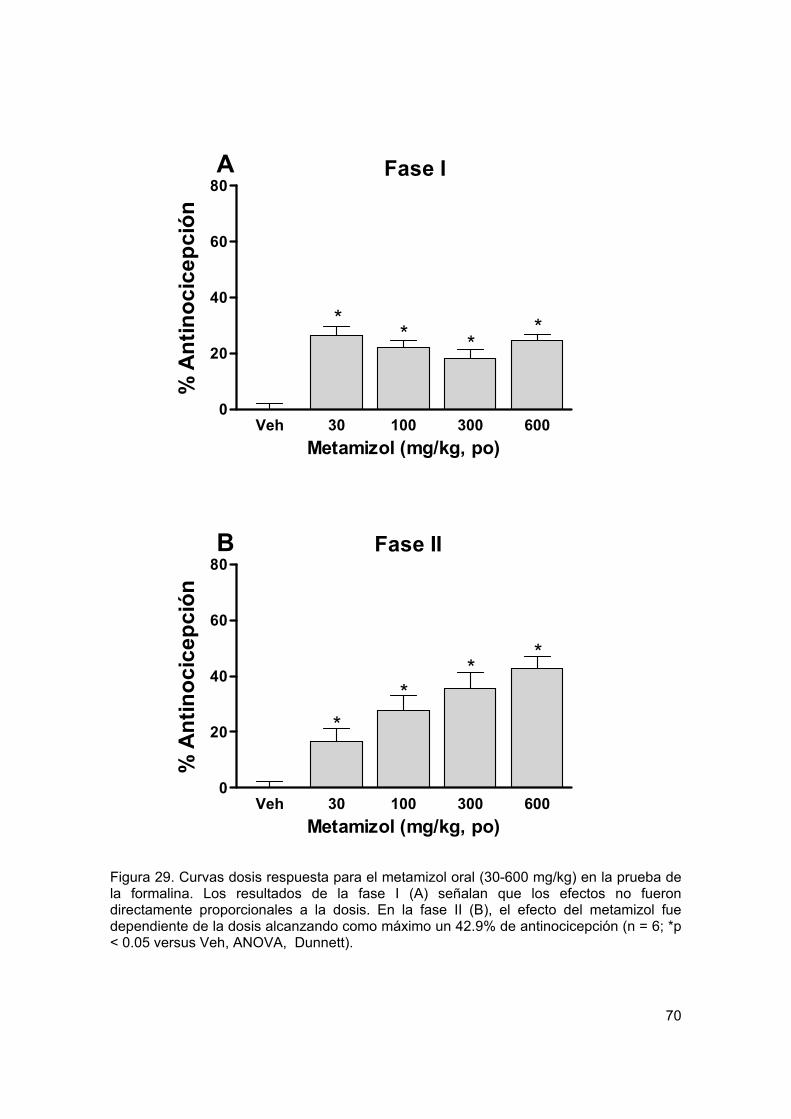
70
Figura 29. Curvas dosis respuesta para el metamizol oral (30-600 mg/kg) en la prueba de la formalina. Los resultados de la fase I (A) señalan que los efectos no fueron directamente proporcionales a la dosis. En la fase II (B), el efecto del metamizol fue dependiente de la dosis alcanzando como máximo un 42.9% de antinocicepción (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 30 100 300 6000
20
40
60
80
**
**
B
Metamizol (mg/kg, po)
% A
ntin
ocic
epci
ón
Fase II
Veh 30 100 300 6000
20
40
60
80
** *
*
A
Metamizol (mg/kg, po)
% A
ntin
ocic
epci
ónFase I
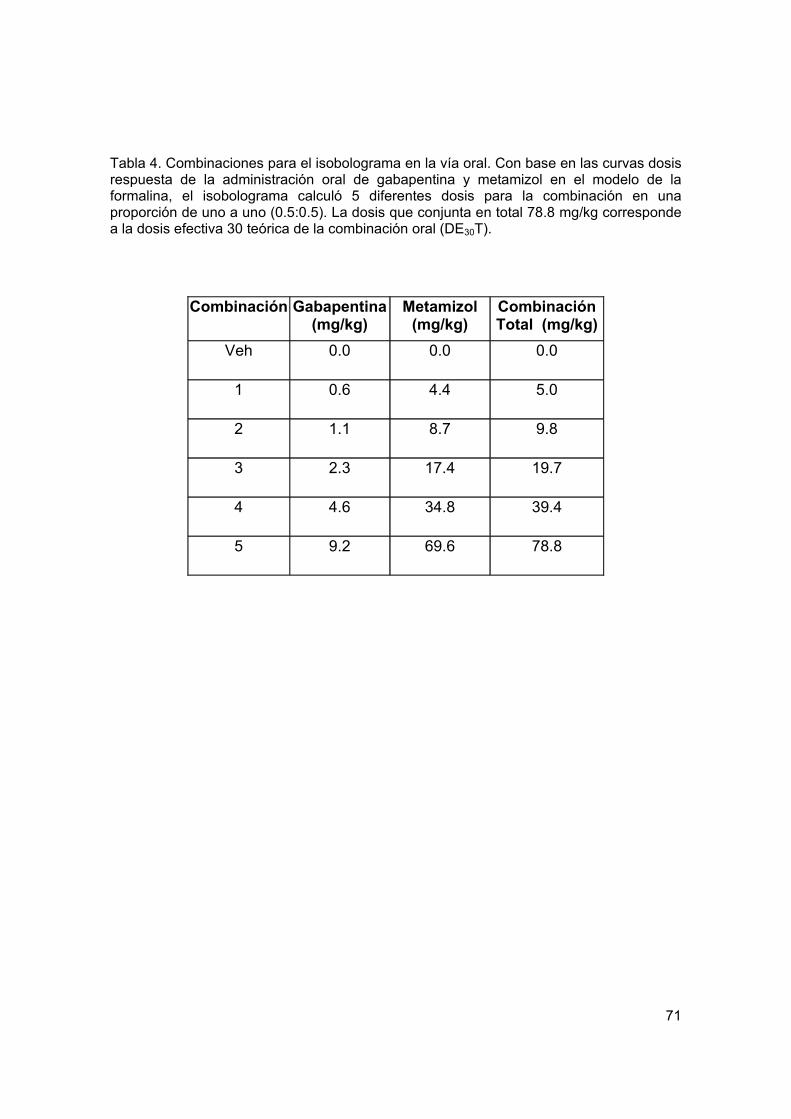
71
Tabla 4. Combinaciones para el isobolograma en la vía oral. Con base en las curvas dosis respuesta de la administración oral de gabapentina y metamizol en el modelo de la formalina, el isobolograma calculó 5 diferentes dosis para la combinación en una proporción de uno a uno (0.5:0.5). La dosis que conjunta en total 78.8 mg/kg corresponde a la dosis efectiva 30 teórica de la combinación oral (DE30T).
Combinación Gabapentina(mg/kg)
Metamizol (mg/kg)
Combinación Total (mg/kg)
Veh 0.0 0.0 0.0
1 0.6 4.4 5.0
2 1.1 8.7 9.8
3 2.3 17.4 19.7
4 4.6 34.8 39.4
5 9.2 69.6 78.8
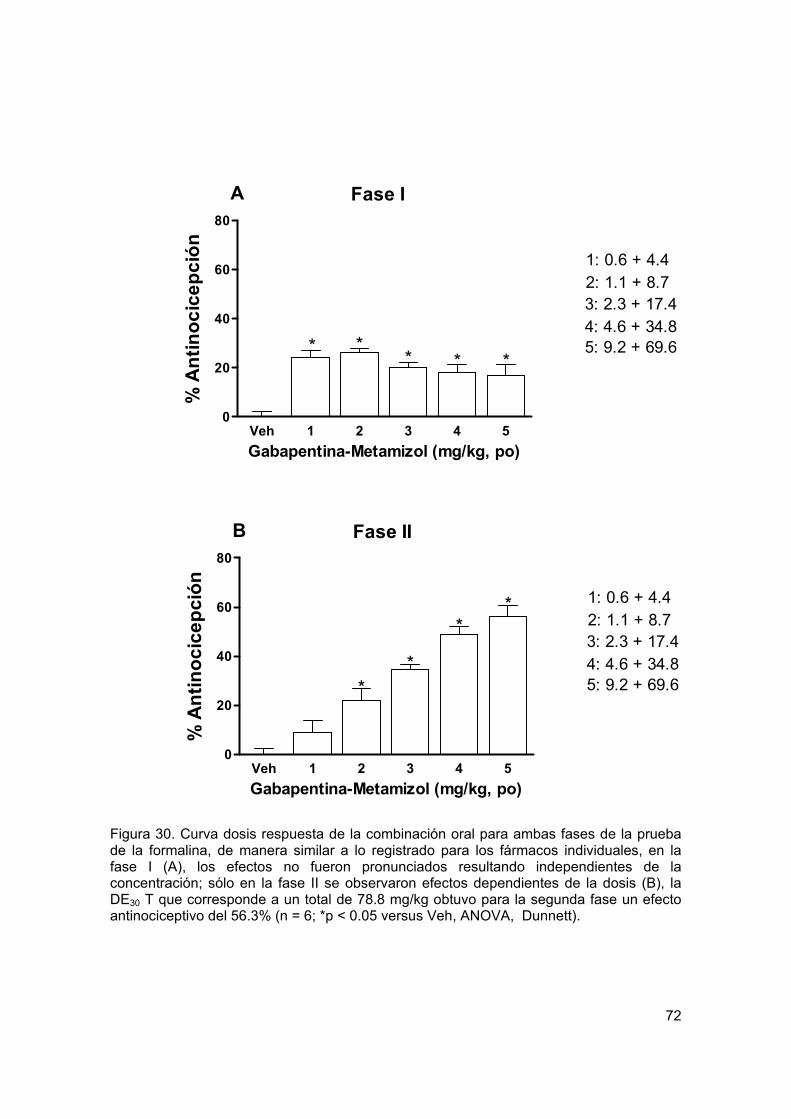
72
Figura 30. Curva dosis respuesta de la combinación oral para ambas fases de la prueba de la formalina, de manera similar a lo registrado para los fármacos individuales, en la fase I (A), los efectos no fueron pronunciados resultando independientes de la concentración; sólo en la fase II se observaron efectos dependientes de la dosis (B), la DE30 T que corresponde a un total de 78.8 mg/kg obtuvo para la segunda fase un efecto antinociceptivo del 56.3% (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 1 2 3 4 50
20
40
60
80
**
**
Fase IIB
1: 0.6 + 4.42: 1.1 + 8.73: 2.3 + 17.44: 4.6 + 34.85: 9.2 + 69.6
Gabapentina-Metamizol (mg/kg, po)
% A
ntin
ocic
epci
ón
Veh 1 2 3 4 50
20
40
60
80
** **
Fase IA
1: 0.6 + 4.42: 1.1 + 8.73: 2.3 + 17.44: 4.6 + 34.85: 9.2 + 69.6
Gabapentina-Metamizol (mg/kg, po)
% A
ntin
ocic
epci
ón
*
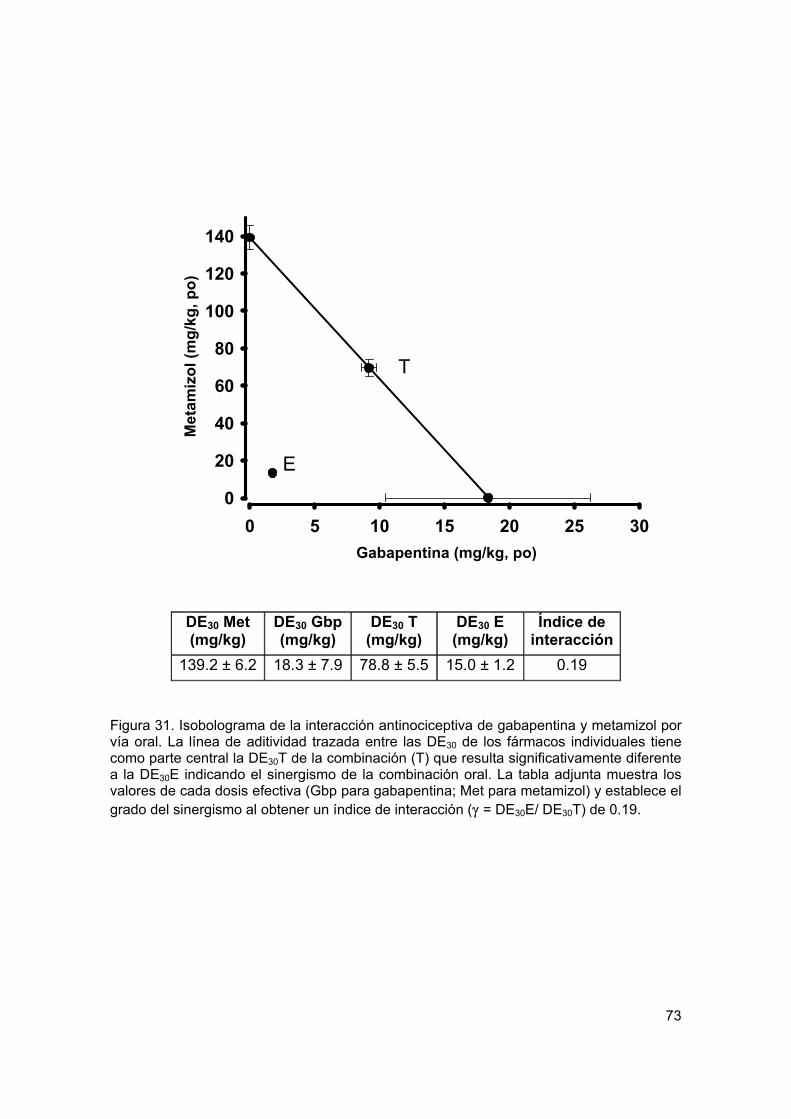
73
DE30 Met (mg/kg)
DE30 Gbp (mg/kg)
DE30 T (mg/kg)
DE30 E (mg/kg)
Índice de interacción
139.2 ± 6.2 18.3 ± 7.9 78.8 ± 5.5 15.0 ± 1.2 0.19
Figura 31. Isobolograma de la interacción antinociceptiva de gabapentina y metamizol por vía oral. La línea de aditividad trazada entre las DE30 de los fármacos individuales tiene como parte central la DE30T de la combinación (T) que resulta significativamente diferente a la DE30E indicando el sinergismo de la combinación oral. La tabla adjunta muestra los valores de cada dosis efectiva (Gbp para gabapentina; Met para metamizol) y establece el grado del sinergismo al obtener un índice de interacción (γ = DE30E/ DE30T) de 0.19.
Gabapentina (mg/kg, po)0 5 10 15 20 25 30
Met
amiz
ol (m
g/kg
, po)
0
20
40
60
80
100
120
140
T
E

74
6.1.2. Efecto espinal de gabapentina, metamizol y la combinación en la prueba de formalina La administración intratecal de gabapentina redujo la nocicepción inducida
por formalina en los cuatro niveles de dosis que fueron empleados (25-200 µg). En
la fase uno los efectos observados no fueron dependientes de la dosis y se
registró como efecto máximo el 31% ± 3.1 con la dosis de 100 µg. En la fase II el
efecto máximo detectado fue de 68.2% de antinocicepción ± 4.1 el cual
correspondió a la dosis más alta (200 µg) debido a que en esta fase la
gabapentina espinal tuvo una eficacia dependiente de la dosis (Figura 32).
Con respecto a la administración intratecal de metamizol, las dosis
empleadas (50-400 µg) resultaron diferentes del control de solución salina. En la
fase neurogénica (fase I) los tratamientos tuvieron un efecto que no puede
considerarse dependiente de la dosis y la respuesta más pronunciada fue de 31%
± 5.5 para la dosis de 200 µg. En la fase II el metamizol redujo de manera dosis
dependiente la conducta de sacudidas y se alcanzó un efecto de hasta 48.5% con
la dosis de 400 µg, como se puede observar de mejor manera en la figura 33.
La curva dosis respuesta de la combinación espinal se efectuó en la fase II
de la prueba (los valores de la fase I se ignoraron al no existir una relación directa
entre la cantidad de dosis y el porcentaje de antinocicepción) utilizando las dosis
descritas en la tabla 4. La dosis más elevada (104.8 µg) corresponde a la dosis
efectiva 30 teórica (DE30T). Una vez que las dosis de la combinación fueron
establecidas por los cálculos del análisis isobolográfico, éstas se sometieron
experimentalmente a la prueba de la formalina. Los resultados mostrados en la
figura 34 indican que la combinación no tuvo un efecto dependiente de la dosis en
la fase inicial de la prueba y el mayor efecto que se pudo observar fue de 27% ±
3.5.
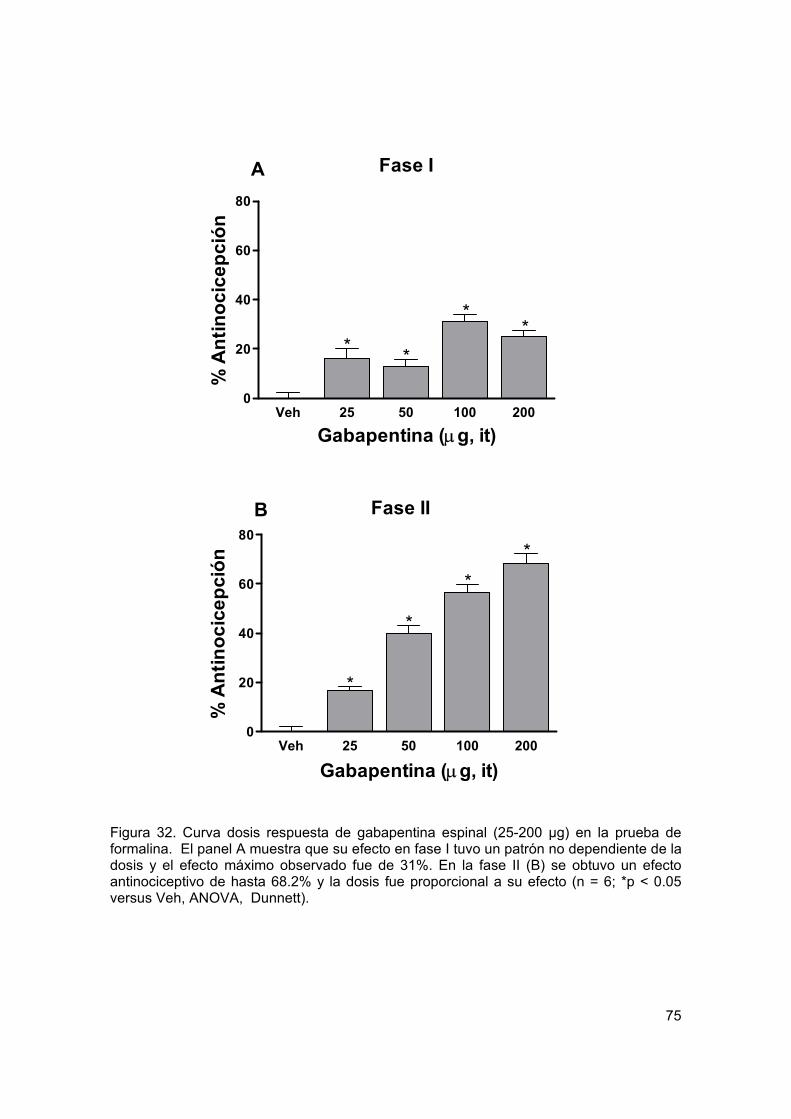
75
Fase I
Veh 25 50 100 2000
20
40
60
80
* **
*
A
Gabapentina (µg, it)
% A
ntin
ocic
epci
ón
Figura 32. Curva dosis respuesta de gabapentina espinal (25-200 µg) en la prueba de formalina. El panel A muestra que su efecto en fase I tuvo un patrón no dependiente de la dosis y el efecto máximo observado fue de 31%. En la fase II (B) se obtuvo un efecto antinociceptivo de hasta 68.2% y la dosis fue proporcional a su efecto (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 25 50 100 2000
20
40
60
80
*
*
**
Fase IIB
Gabapentina (µg, it)
% A
ntin
ocic
epci
ón
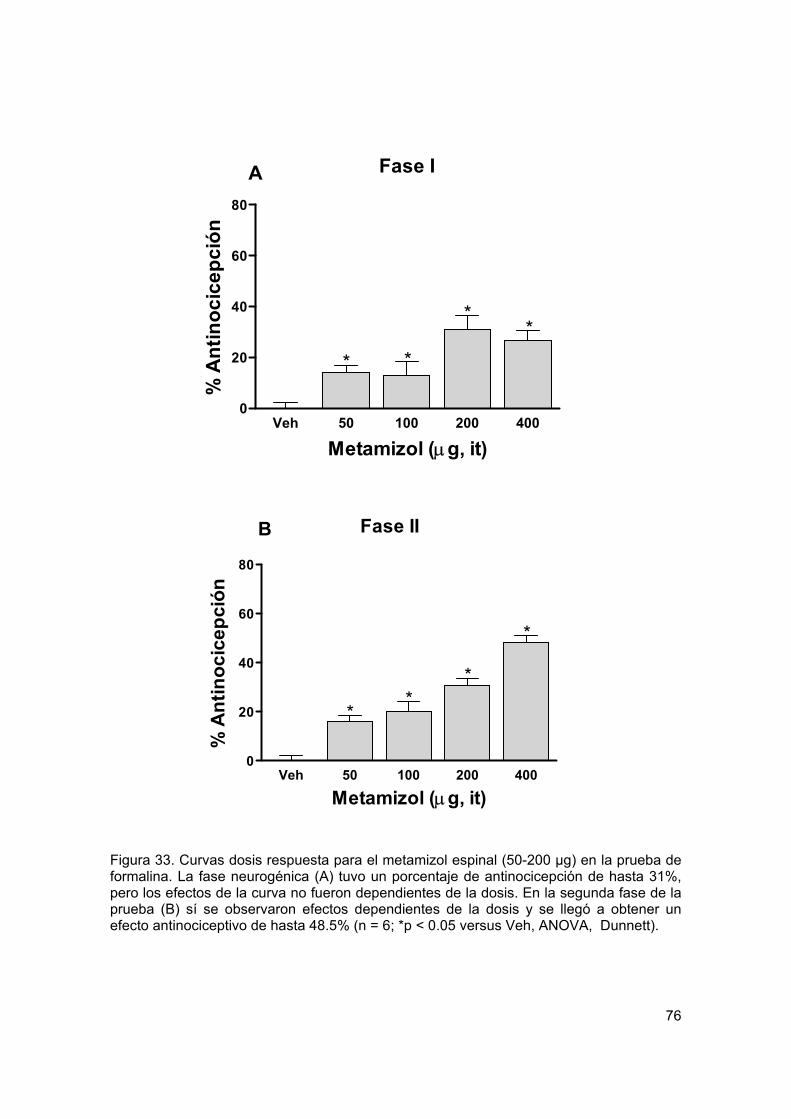
76
Fase I
Veh 50 100 200 4000
20
40
60
80
*
**
*
A
Metamizol (µg, it)
% A
ntin
ocic
epci
ón
Figura 33. Curvas dosis respuesta para el metamizol espinal (50-200 µg) en la prueba de formalina. La fase neurogénica (A) tuvo un porcentaje de antinocicepción de hasta 31%, pero los efectos de la curva no fueron dependientes de la dosis. En la segunda fase de la prueba (B) sí se observaron efectos dependientes de la dosis y se llegó a obtener un efecto antinociceptivo de hasta 48.5% (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 50 100 200 4000
20
40
60
80
**
*
*
Fase IIB
Metamizol (µg, it)
% A
ntin
ocic
epci
ón
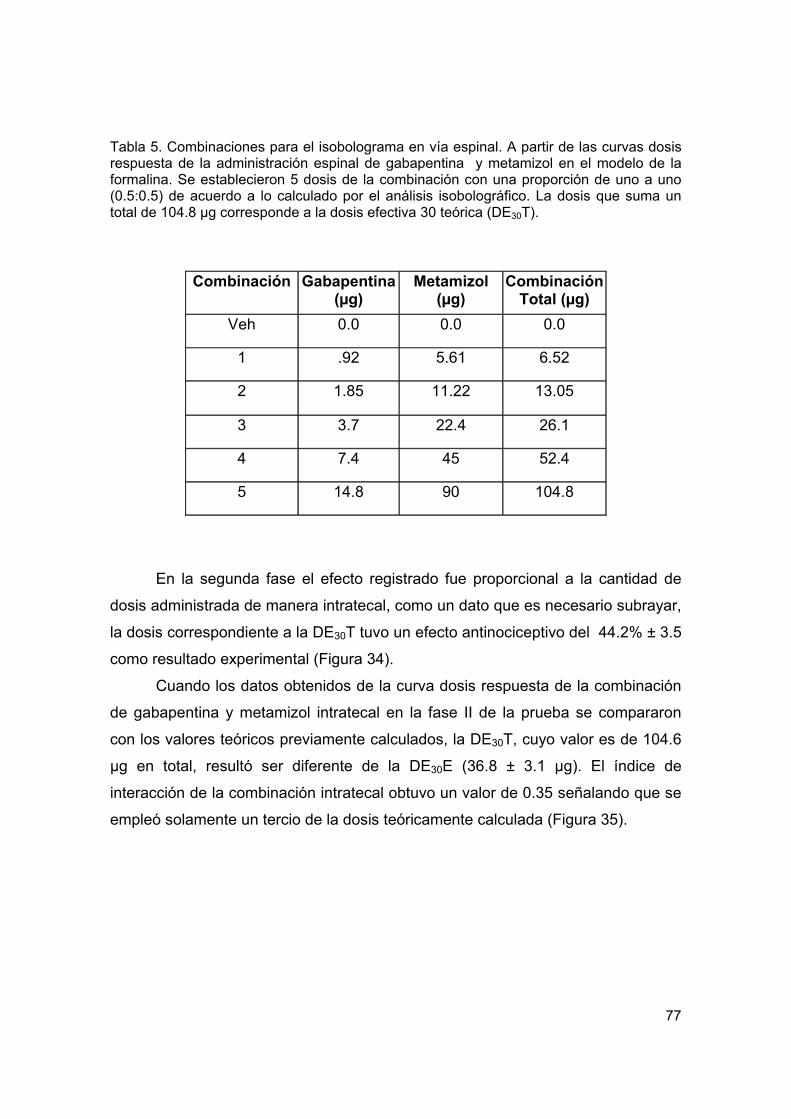
77
Tabla 5. Combinaciones para el isobolograma en vía espinal. A partir de las curvas dosis respuesta de la administración espinal de gabapentina y metamizol en el modelo de la formalina. Se establecieron 5 dosis de la combinación con una proporción de uno a uno (0.5:0.5) de acuerdo a lo calculado por el análisis isobolográfico. La dosis que suma un total de 104.8 µg corresponde a la dosis efectiva 30 teórica (DE30T).
Combinación Gabapentina
(µg) Metamizol
(µg) Combinación
Total (µg) Veh 0.0 0.0 0.0
1 .92 5.61 6.52
2 1.85 11.22 13.05
3 3.7 22.4 26.1
4 7.4 45 52.4
5 14.8 90 104.8
En la segunda fase el efecto registrado fue proporcional a la cantidad de
dosis administrada de manera intratecal, como un dato que es necesario subrayar,
la dosis correspondiente a la DE30T tuvo un efecto antinociceptivo del 44.2% ± 3.5
como resultado experimental (Figura 34).
Cuando los datos obtenidos de la curva dosis respuesta de la combinación
de gabapentina y metamizol intratecal en la fase II de la prueba se compararon
con los valores teóricos previamente calculados, la DE30T, cuyo valor es de 104.6
µg en total, resultó ser diferente de la DE30E (36.8 ± 3.1 µg). El índice de
interacción de la combinación intratecal obtuvo un valor de 0.35 señalando que se
empleó solamente un tercio de la dosis teóricamente calculada (Figura 35).
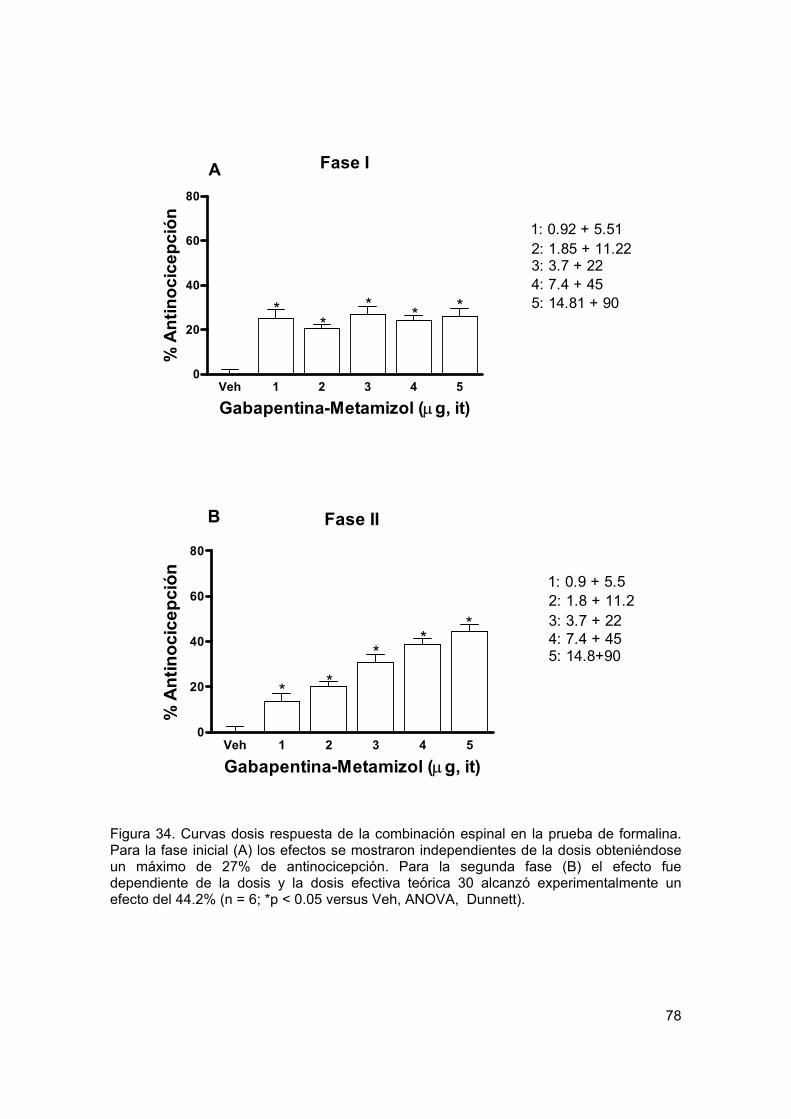
78
Fase I
Veh 1 2 3 4 50
20
40
60
80
****
*
A
1: 0.92 + 5.512: 1.85 + 11.223: 3.7 + 224: 7.4 + 455: 14.81 + 90
Gabapentina-Metamizol (µg, it)
% A
ntin
ocic
epci
ón
Figura 34. Curvas dosis respuesta de la combinación espinal en la prueba de formalina. Para la fase inicial (A) los efectos se mostraron independientes de la dosis obteniéndose un máximo de 27% de antinocicepción. Para la segunda fase (B) el efecto fue dependiente de la dosis y la dosis efectiva teórica 30 alcanzó experimentalmente un efecto del 44.2% (n = 6; *p < 0.05 versus Veh, ANOVA, Dunnett).
Veh 1 2 3 4 50
20
40
60
80
*
**
**
Fase IIB
1: 0.9 + 5.52: 1.8 + 11.23: 3.7 + 224: 7.4 + 455: 14.8+90
Gabapentina-Metamizol (µg, it)
% A
ntin
ocic
epci
ón
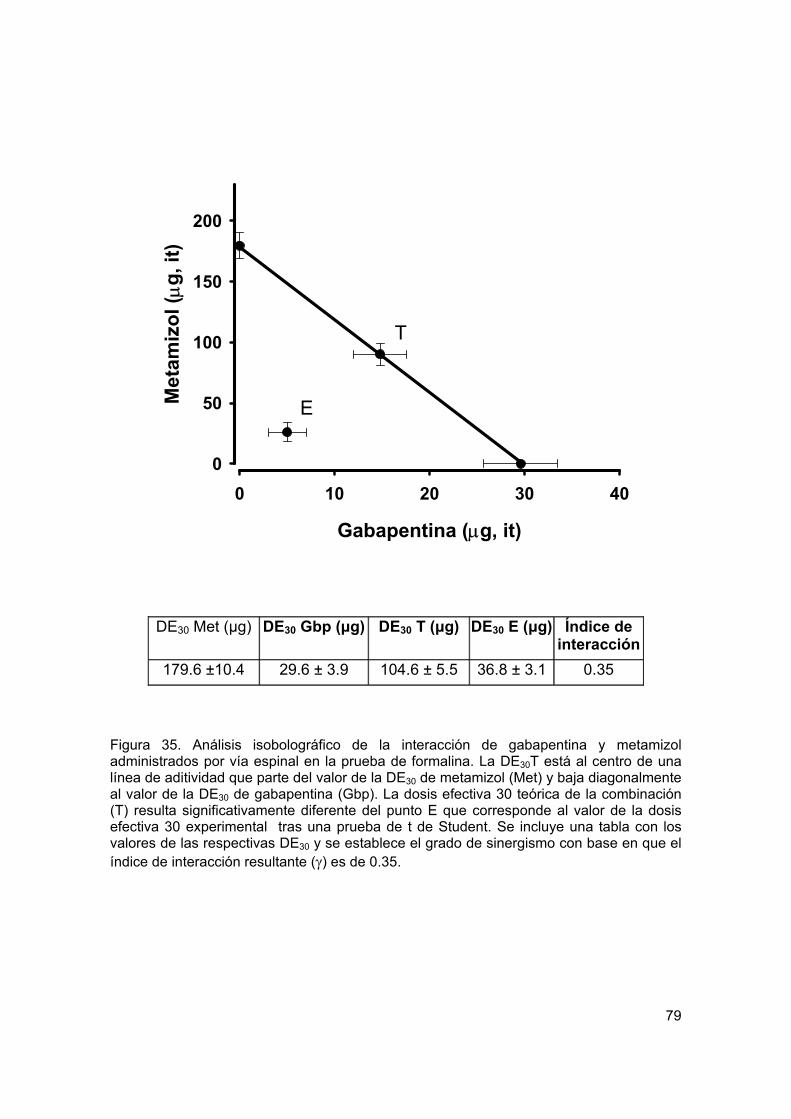
79
Gabapentina (µg, it)
0 10 20 30 40
Met
amiz
ol ( µ
g, it
)
0
50
100
150
200
T
E
DE30 Met (µg) DE30 Gbp (µg) DE30 T (µg) DE30 E (µg) Índice de interacción
179.6 ±10.4 29.6 ± 3.9 104.6 ± 5.5 36.8 ± 3.1 0.35 Figura 35. Análisis isobolográfico de la interacción de gabapentina y metamizol administrados por vía espinal en la prueba de formalina. La DE30T está al centro de una línea de aditividad que parte del valor de la DE30 de metamizol (Met) y baja diagonalmente al valor de la DE30 de gabapentina (Gbp). La dosis efectiva 30 teórica de la combinación (T) resulta significativamente diferente del punto E que corresponde al valor de la dosis efectiva 30 experimental tras una prueba de t de Student. Se incluye una tabla con los valores de las respectivas DE30 y se establece el grado de sinergismo con base en que el índice de interacción resultante (γ) es de 0.35.
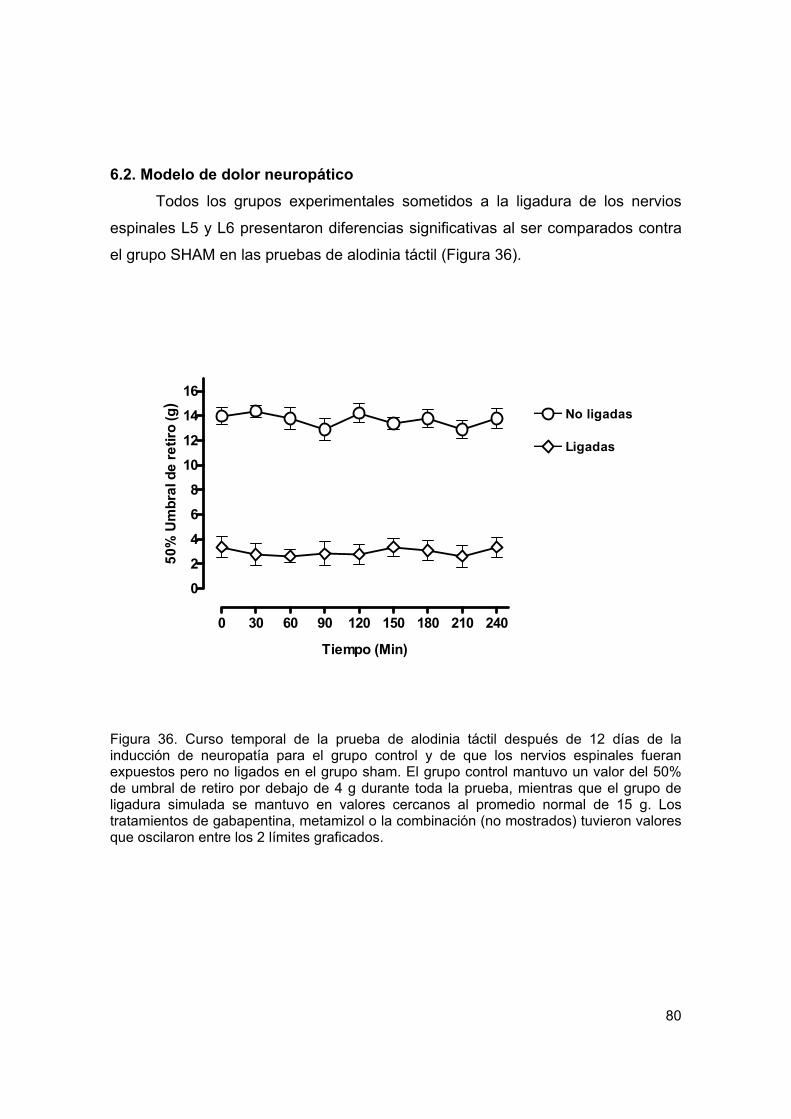
80
6.2. Modelo de dolor neuropático Todos los grupos experimentales sometidos a la ligadura de los nervios
espinales L5 y L6 presentaron diferencias significativas al ser comparados contra
el grupo SHAM en las pruebas de alodinia táctil (Figura 36).
0 30 60 90 120 150 180 210 240
0
2
4
6
8
10
12
14
16
Ligadas
No ligadas
Tiempo (Min)
50%
Um
bral
de
retir
o (g
)
Figura 36. Curso temporal de la prueba de alodinia táctil después de 12 días de la inducción de neuropatía para el grupo control y de que los nervios espinales fueran expuestos pero no ligados en el grupo sham. El grupo control mantuvo un valor del 50% de umbral de retiro por debajo de 4 g durante toda la prueba, mientras que el grupo de ligadura simulada se mantuvo en valores cercanos al promedio normal de 15 g. Los tratamientos de gabapentina, metamizol o la combinación (no mostrados) tuvieron valores que oscilaron entre los 2 límites graficados.

81
6.2.1. Efecto antialodínico de gabapentina, metamizol y la combinación por vía oral La administración oral de gabapentina redujo de manera dosis dependiente
la alodinia táctil en ratas neuropáticas. El efecto máximo alcanzado fue de 59.3%
para la dosis de 300 mg/kg (Figura 37).
Para el caso del metamizol, al no existir estudios de referencia se decidió
emplear las dosis de 100-600 mg/kg. Sin embargo, la eficacia del metamizol fue
prácticamente nula observándose un efecto del 7% sólo en la dosis de 600 mg/kg.
Dado que el metamizol careció de efecto antialodínico en la práctica por esta vía
de administración, el isobolograma convencional no pudo emplearse debido a que
está basado en el uso de concentraciones equiefectivas de los fármacos en
combinación. Sin embargo, existe una variante en la que se puede considerar a
uno de los fármacos como adyuvante de otro con eficacia probada para analizar
su interacción (Tallarida, 2003). Con base en este tipo de estudios, se decidió
emplear una dosis fija de metamizol (600 mg/kg) combinada con las dosis
crecientes de gabapentina empleadas previamente en la curva dosis respuesta
(30-300 mg/kg). La combinación oral tuvo un efecto significativamente superior a
la gabapentina individual, pero sólo en las dosis más altas (Figura 37). El efecto
máximo observado se elevó hasta un 77.9% ± 4 con la combinación (Figura 37).
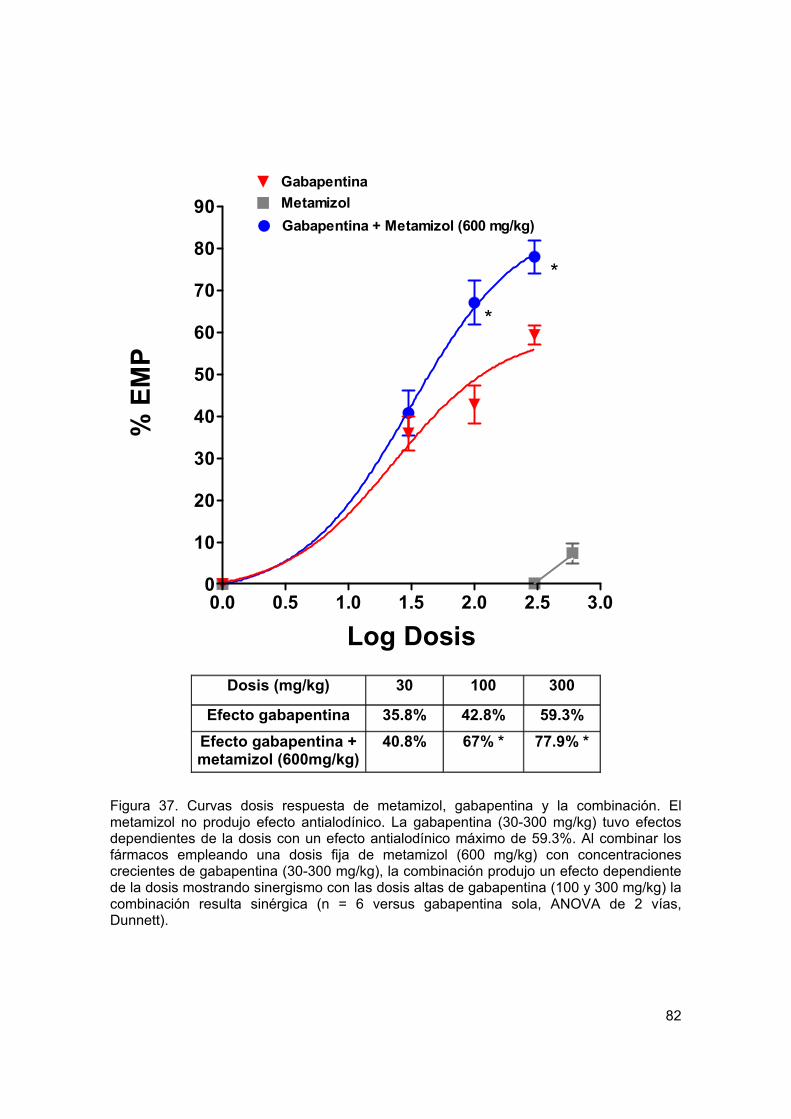
82
Figura 37. Curvas dosis respuesta de metamizol, gabapentina y la combinación. El metamizol no produjo efecto antialodínico. La gabapentina (30-300 mg/kg) tuvo efectos dependientes de la dosis con un efecto antialodínico máximo de 59.3%. Al combinar los fármacos empleando una dosis fija de metamizol (600 mg/kg) con concentraciones crecientes de gabapentina (30-300 mg/kg), la combinación produjo un efecto dependiente de la dosis mostrando sinergismo con las dosis altas de gabapentina (100 y 300 mg/kg) la combinación resulta sinérgica (n = 6 versus gabapentina sola, ANOVA de 2 vías, Dunnett).
Dosis (mg/kg) 30 100 300
Efecto gabapentina 35.8% 42.8% 59.3%
Efecto gabapentina + metamizol (600mg/kg)
40.8% 67% * 77.9% *
0.0 0.5 1.0 1.5 2.0 2.5 3.00
10
20
30
40
50
60
70
80
90Gabapentina
Gabapentina + Metamizol (600 mg/kg)Metamizol
*
*
Log Dosis
% E
MP
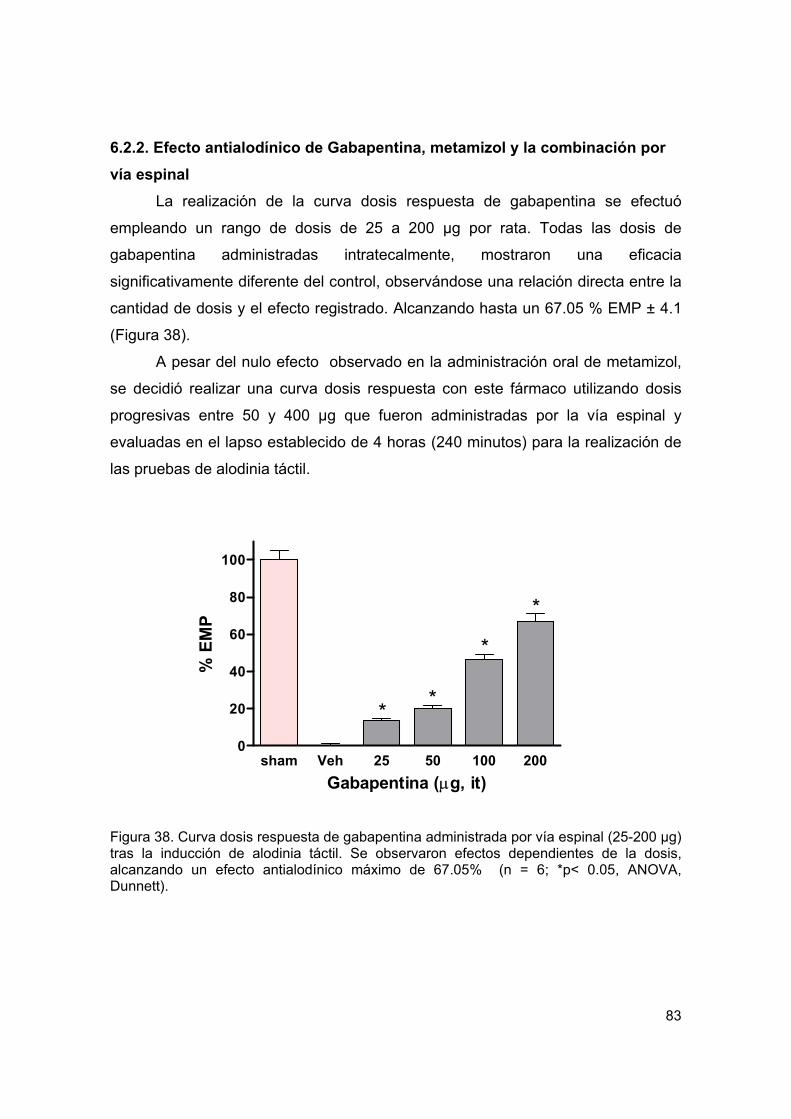
83
6.2.2. Efecto antialodínico de Gabapentina, metamizol y la combinación por vía espinal La realización de la curva dosis respuesta de gabapentina se efectuó
empleando un rango de dosis de 25 a 200 µg por rata. Todas las dosis de
gabapentina administradas intratecalmente, mostraron una eficacia
significativamente diferente del control, observándose una relación directa entre la
cantidad de dosis y el efecto registrado. Alcanzando hasta un 67.05 % EMP ± 4.1
(Figura 38).
A pesar del nulo efecto observado en la administración oral de metamizol,
se decidió realizar una curva dosis respuesta con este fármaco utilizando dosis
progresivas entre 50 y 400 µg que fueron administradas por la vía espinal y
evaluadas en el lapso establecido de 4 horas (240 minutos) para la realización de
las pruebas de alodinia táctil.
Figura 38. Curva dosis respuesta de gabapentina administrada por vía espinal (25-200 µg) tras la inducción de alodinia táctil. Se observaron efectos dependientes de la dosis, alcanzando un efecto antialodínico máximo de 67.05% (n = 6; *p< 0.05, ANOVA, Dunnett).
sham Veh 25 50 100 2000
20
40
60
80
100
* *
*
*
Gabapentina (µg, it)
% E
MP

84
El metamizol, de forma inesperada, presentó eficacia por esta vía de
administración (Figura 39). Los efectos de las cuatro dosis administradas tuvieron
diferencias significativas con el control y la curva dosis respuesta se mostró
dependiente de la dosis. De manera contrastante con lo descrito para la mayoría
de los analgésicos no esteroideos e incluso para lo observado con el mismo
fármaco en este trabajo por vía oral, el metamizol alcanzó un efecto de hasta
39.2% ± 3.5 como se puede observar en la figura 39.
De esta manera, al presentar ambos fármacos eficacia por la vía de
administración espinal, se pudo emplear el isobolograma convencional tomando
como base las DE30 de gabapentina y metamizol por vía espinal. Con este fin se
calcularon las dosis para la combinación que se enumeran en la tabla 5 y de las
que se describe su efecto en la figura 40.
Las cuatro dosis de los fármacos en combinación tuvieron efectos con
diferencia significativa al ser comparados contra el control. En conjunto, la eficacia
de la combinación de gabapentina y metamizol se mostró dependiente de la dosis
al administrarse por vía espinal. La DE30T de la combinación registró
experimentalmente un efecto de 39.17% EMP ± 3.5 (Figura 40).
Los datos de la curva dosis respuesta de la administración individual de los
fármacos y de la combinación espinal se utilizaron para el cálculo del análisis
isobolográfico. La dosis efectiva 30 teórica de la combinación (118.4 µg) resultó
significativamente diferente del valor de 66.2 µg correspondiente a la dosis
efectiva 30 experimental (DE30E). Al grado de requerir prácticamente la mitad de
la dosis teóricamente calculada al presentar un índice de interacción de 0.55
(Figura 41).
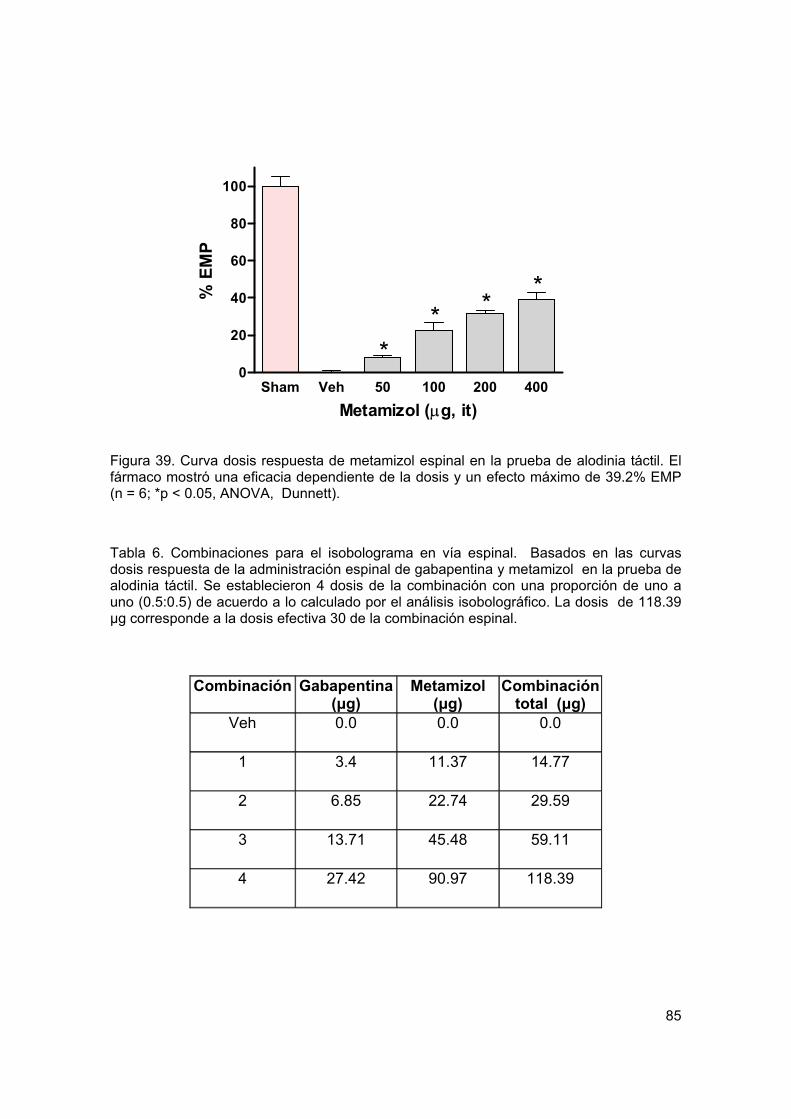
85
Figura 39. Curva dosis respuesta de metamizol espinal en la prueba de alodinia táctil. El fármaco mostró una eficacia dependiente de la dosis y un efecto máximo de 39.2% EMP (n = 6; *p < 0.05, ANOVA, Dunnett).
Tabla 6. Combinaciones para el isobolograma en vía espinal. Basados en las curvas dosis respuesta de la administración espinal de gabapentina y metamizol en la prueba de alodinia táctil. Se establecieron 4 dosis de la combinación con una proporción de uno a uno (0.5:0.5) de acuerdo a lo calculado por el análisis isobolográfico. La dosis de 118.39 µg corresponde a la dosis efectiva 30 de la combinación espinal.
Combinación Gabapentina(µg)
Metamizol (µg)
Combinación total (µg)
Veh 0.0 0.0 0.0
1 3.4 11.37 14.77
2 6.85 22.74 29.59
3 13.71 45.48 59.11
4 27.42 90.97 118.39
Sham Veh 50 100 200 4000
20
40
60
80
100
** *
*
Metamizol (µg, it)
% E
MP
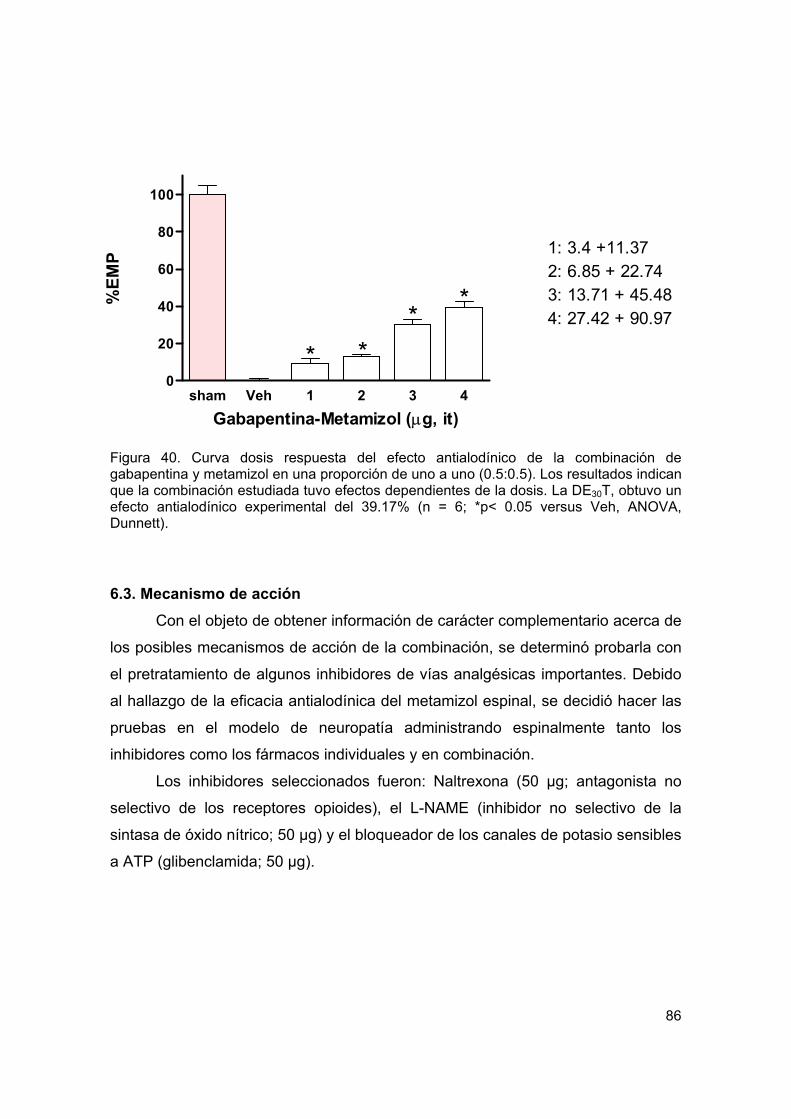
86
Figura 40. Curva dosis respuesta del efecto antialodínico de la combinación de gabapentina y metamizol en una proporción de uno a uno (0.5:0.5). Los resultados indican que la combinación estudiada tuvo efectos dependientes de la dosis. La DE30T, obtuvo un efecto antialodínico experimental del 39.17% (n = 6; *p< 0.05 versus Veh, ANOVA, Dunnett).
6.3. Mecanismo de acción Con el objeto de obtener información de carácter complementario acerca de
los posibles mecanismos de acción de la combinación, se determinó probarla con
el pretratamiento de algunos inhibidores de vías analgésicas importantes. Debido
al hallazgo de la eficacia antialodínica del metamizol espinal, se decidió hacer las
pruebas en el modelo de neuropatía administrando espinalmente tanto los
inhibidores como los fármacos individuales y en combinación.
Los inhibidores seleccionados fueron: Naltrexona (50 µg; antagonista no
selectivo de los receptores opioides), el L-NAME (inhibidor no selectivo de la
sintasa de óxido nítrico; 50 µg) y el bloqueador de los canales de potasio sensibles
a ATP (glibenclamida; 50 µg).
sham Veh 1 2 3 40
20
40
60
80
100
* **
*
1: 3.4 +11.372: 6.85 + 22.743: 13.71 + 45.484: 27.42 + 90.97
Gabapentina-Metamizol (µg, it)
%EM
P
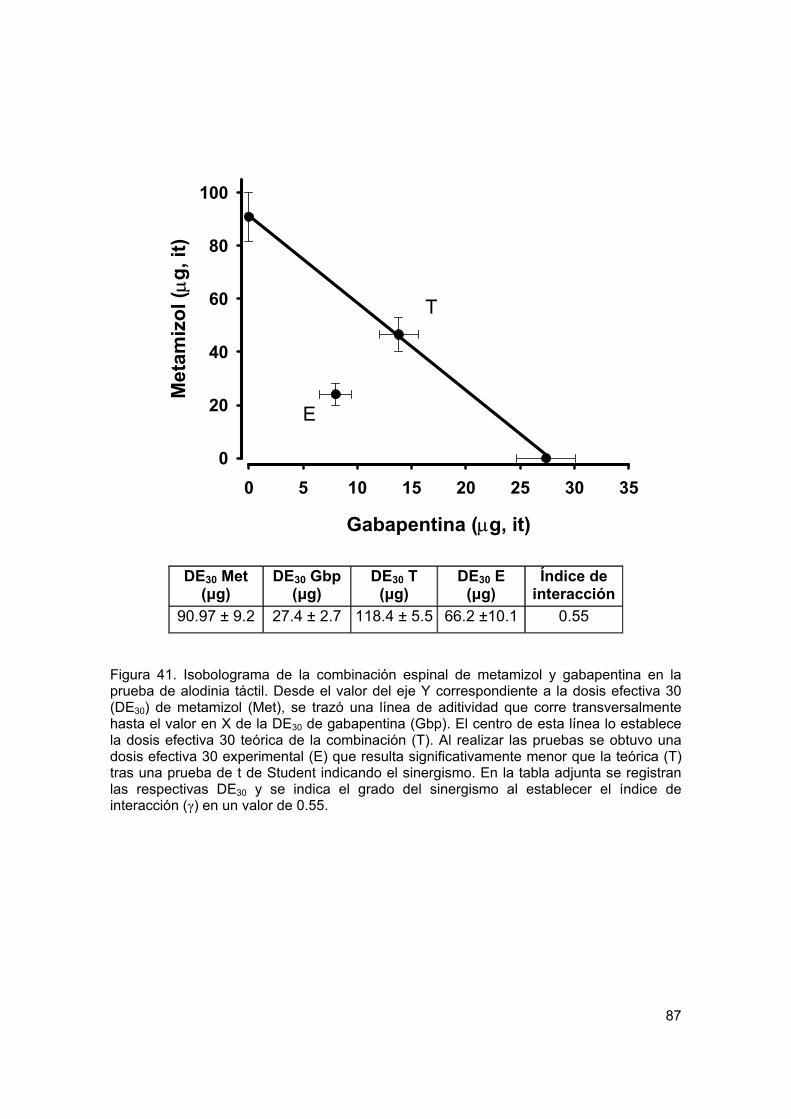
87
Gabapentina (µg, it)
0 5 10 15 20 25 30 35
Met
amiz
ol ( µ
g, it
)
0
20
40
60
80
100
T
E
DE30 Met (µg)
DE30 Gbp (µg)
DE30 T (µg)
DE30 E (µg)
Índice de interacción
90.97 ± 9.2 27.4 ± 2.7 118.4 ± 5.5 66.2 ±10.1 0.55 Figura 41. Isobolograma de la combinación espinal de metamizol y gabapentina en la prueba de alodinia táctil. Desde el valor del eje Y correspondiente a la dosis efectiva 30 (DE30) de metamizol (Met), se trazó una línea de aditividad que corre transversalmente hasta el valor en X de la DE30 de gabapentina (Gbp). El centro de esta línea lo establece la dosis efectiva 30 teórica de la combinación (T). Al realizar las pruebas se obtuvo una dosis efectiva 30 experimental (E) que resulta significativamente menor que la teórica (T) tras una prueba de t de Student indicando el sinergismo. En la tabla adjunta se registran las respectivas DE30 y se indica el grado del sinergismo al establecer el índice de interacción (γ) en un valor de 0.55.
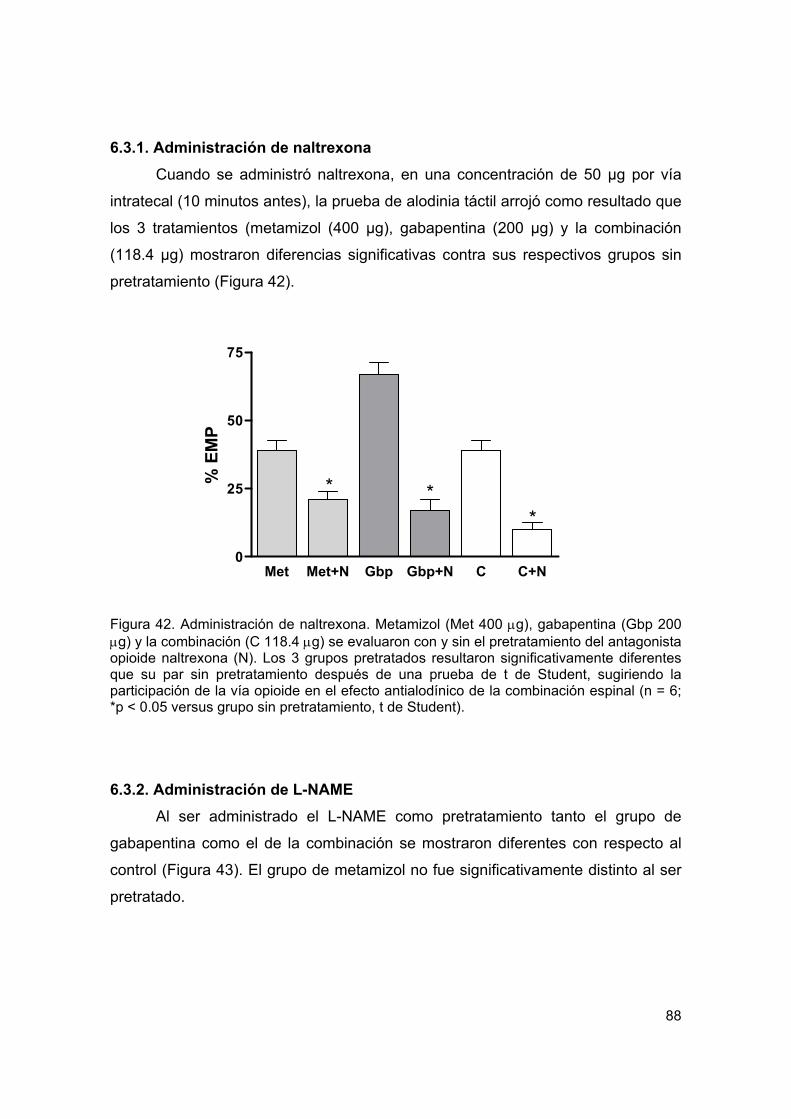
88
6.3.1. Administración de naltrexona
Cuando se administró naltrexona, en una concentración de 50 µg por vía
intratecal (10 minutos antes), la prueba de alodinia táctil arrojó como resultado que
los 3 tratamientos (metamizol (400 µg), gabapentina (200 µg) y la combinación
(118.4 µg) mostraron diferencias significativas contra sus respectivos grupos sin
pretratamiento (Figura 42).
Met Met+N Gbp Gbp+N C C+N0
25
50
75
**
% E
MP
*
Figura 42. Administración de naltrexona. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se evaluaron con y sin el pretratamiento del antagonista opioide naltrexona (N). Los 3 grupos pretratados resultaron significativamente diferentes que su par sin pretratamiento después de una prueba de t de Student, sugiriendo la participación de la vía opioide en el efecto antialodínico de la combinación espinal (n = 6; *p < 0.05 versus grupo sin pretratamiento, t de Student). 6.3.2. Administración de L-NAME Al ser administrado el L-NAME como pretratamiento tanto el grupo de
gabapentina como el de la combinación se mostraron diferentes con respecto al
control (Figura 43). El grupo de metamizol no fue significativamente distinto al ser
pretratado.
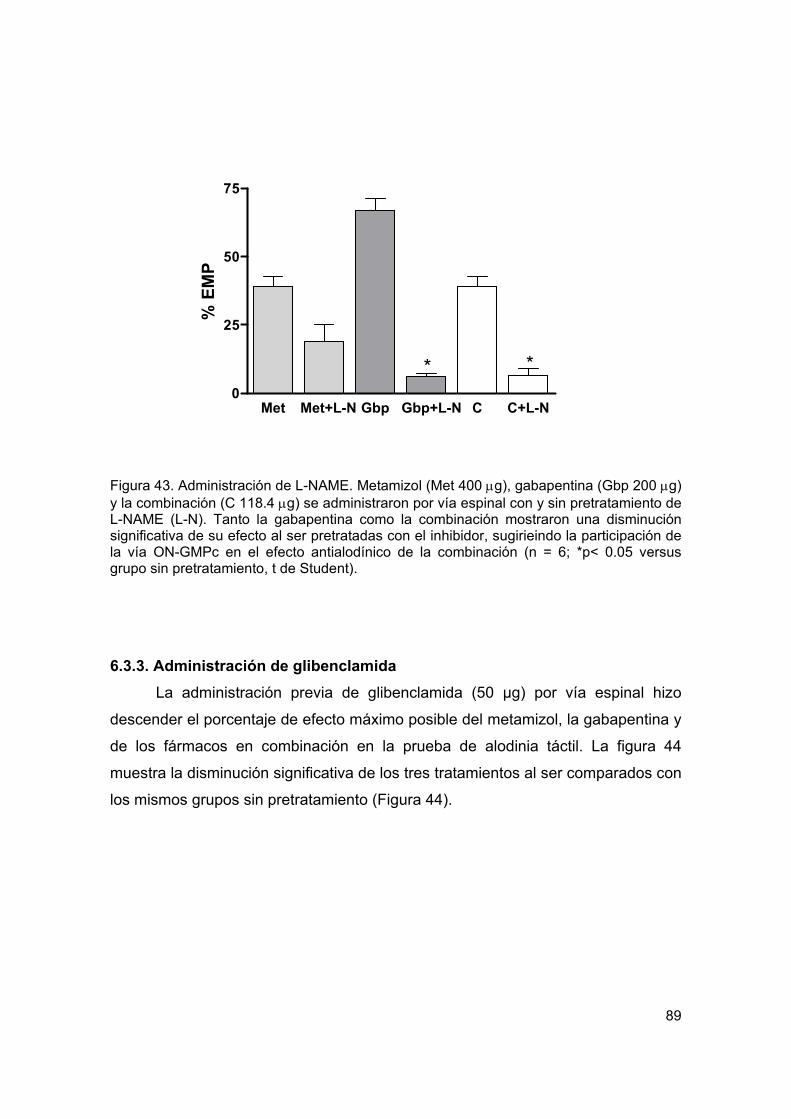
89
Figura 43. Administración de L-NAME. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron por vía espinal con y sin pretratamiento de L-NAME (L-N). Tanto la gabapentina como la combinación mostraron una disminución significativa de su efecto al ser pretratadas con el inhibidor, sugirieindo la participación de la vía ON-GMPc en el efecto antialodínico de la combinación (n = 6; *p< 0.05 versus grupo sin pretratamiento, t de Student). 6.3.3. Administración de glibenclamida La administración previa de glibenclamida (50 µg) por vía espinal hizo
descender el porcentaje de efecto máximo posible del metamizol, la gabapentina y
de los fármacos en combinación en la prueba de alodinia táctil. La figura 44
muestra la disminución significativa de los tres tratamientos al ser comparados con
los mismos grupos sin pretratamiento (Figura 44).
Met Met+L-N Gbp Gbp+L-N C C+L-N0
25
50
75
* *
% E
MP
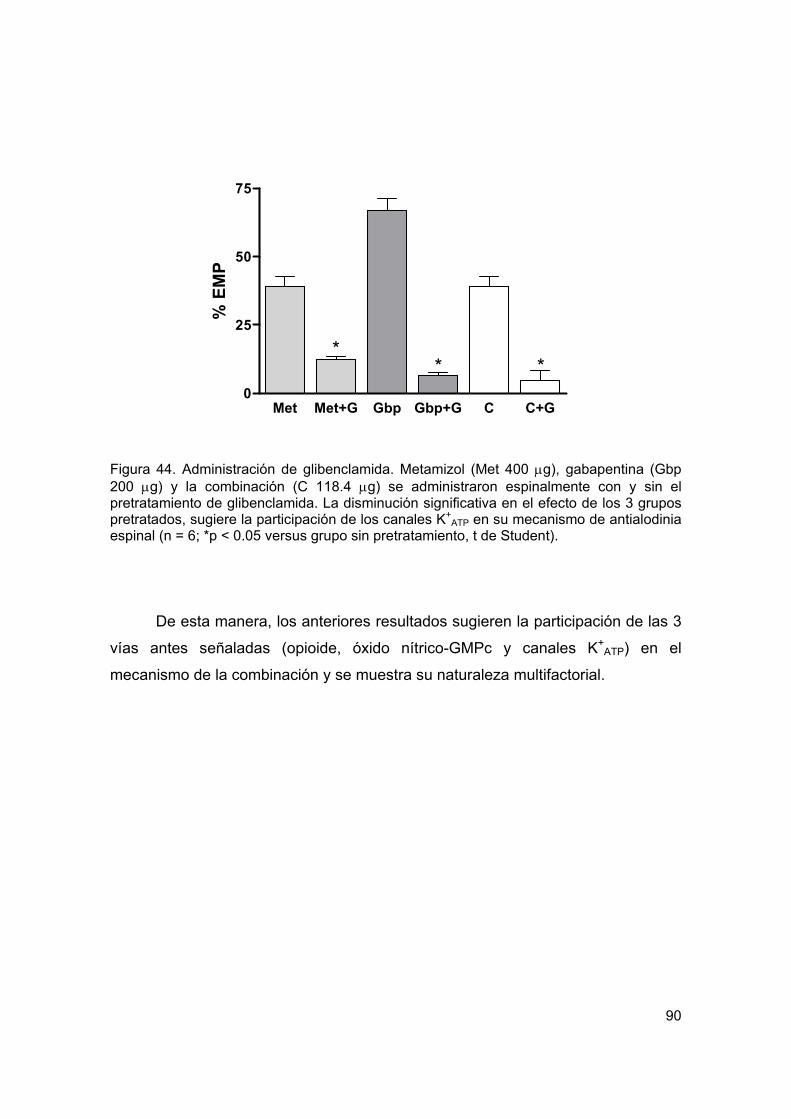
90
Figura 44. Administración de glibenclamida. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron espinalmente con y sin el pretratamiento de glibenclamida. La disminución significativa en el efecto de los 3 grupos pretratados, sugiere la participación de los canales K+
ATP en su mecanismo de antialodinia espinal (n = 6; *p < 0.05 versus grupo sin pretratamiento, t de Student).
De esta manera, los anteriores resultados sugieren la participación de las 3
vías antes señaladas (opioide, óxido nítrico-GMPc y canales K+ATP) en el
mecanismo de la combinación y se muestra su naturaleza multifactorial.
Met Met+G Gbp Gbp+G C C+G0
25
50
75
** *
% E
MP
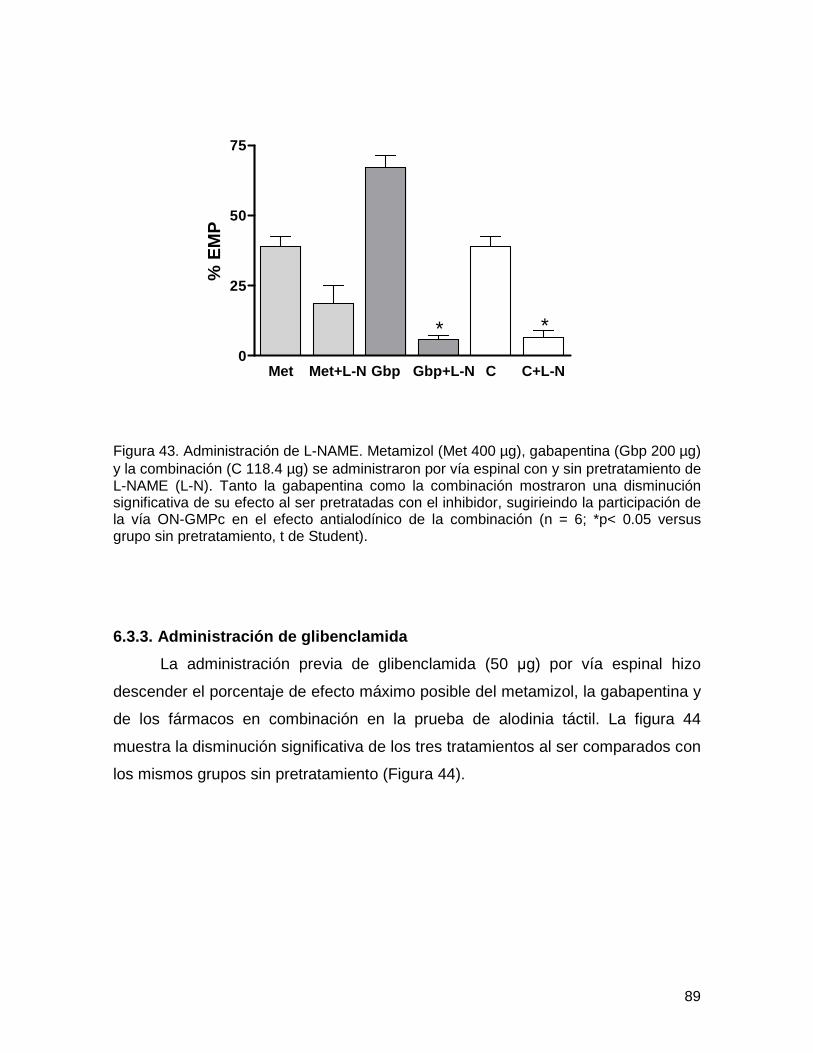
89
Figura 43. Administración de L-NAME. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron por vía espinal con y sin pretratamiento de L-NAME (L-N). Tanto la gabapentina como la combinación mostraron una disminución significativa de su efecto al ser pretratadas con el inhibidor, sugirieindo la participación de la vía ON-GMPc en el efecto antialodínico de la combinación (n = 6; *p< 0.05 versus grupo sin pretratamiento, t de Student).
6.3.3. Administración de glibenclamida
La administración previa de glibenclamida (50 µg) por vía espinal hizo
descender el porcentaje de efecto máximo posible del metamizol, la gabapentina y
de los fármacos en combinación en la prueba de alodinia táctil. La figura 44
muestra la disminución significativa de los tres tratamientos al ser comparados con
los mismos grupos sin pretratamiento (Figura 44).
Met Met+L-N Gbp Gbp+L-N C C+L-N0
25
50
75
* *
% E
MP
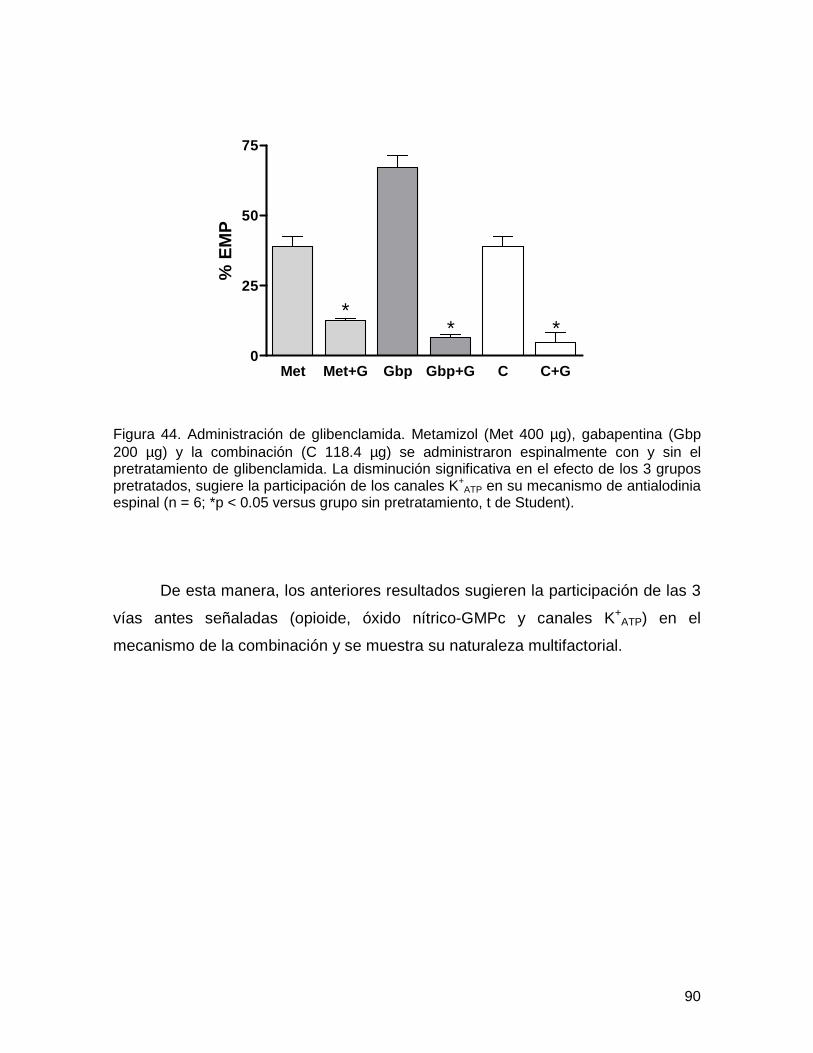
90
Figura 44. Administración de glibenclamida. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron espinalmente con y sin el pretratamiento de glibenclamida. La disminución significativa en el efecto de los 3 grupos pretratados, sugiere la participación de los canales K+
ATP en su mecanismo de antialodinia espinal (n = 6; *p < 0.05 versus grupo sin pretratamiento, t de Student).
De esta manera, los anteriores resultados sugieren la participación de las 3
vías antes señaladas (opioide, óxido nítrico-GMPc y canales K+ATP) en el
mecanismo de la combinación y se muestra su naturaleza multifactorial.
Met Met+G Gbp Gbp+G C C+G0
25
50
75
** *
% E
MP
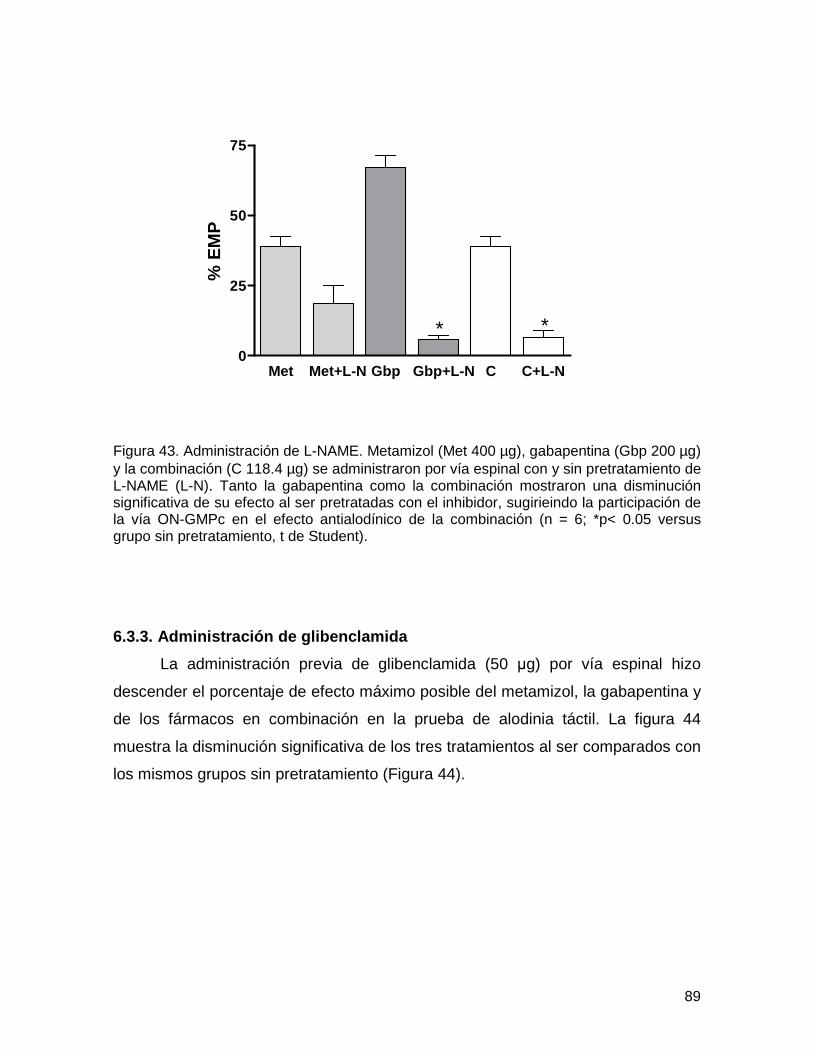
89
Figura 43. Administración de L-NAME. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron por vía espinal con y sin pretratamiento de L-NAME (L-N). Tanto la gabapentina como la combinación mostraron una disminución significativa de su efecto al ser pretratadas con el inhibidor, sugirieindo la participación de la vía ON-GMPc en el efecto antialodínico de la combinación (n = 6; *p< 0.05 versus grupo sin pretratamiento, t de Student).
6.3.3. Administración de glibenclamida
La administración previa de glibenclamida (50 µg) por vía espinal hizo
descender el porcentaje de efecto máximo posible del metamizol, la gabapentina y
de los fármacos en combinación en la prueba de alodinia táctil. La figura 44
muestra la disminución significativa de los tres tratamientos al ser comparados con
los mismos grupos sin pretratamiento (Figura 44).
Met Met+L-N Gbp Gbp+L-N C C+L-N0
25
50
75
* *
% E
MP
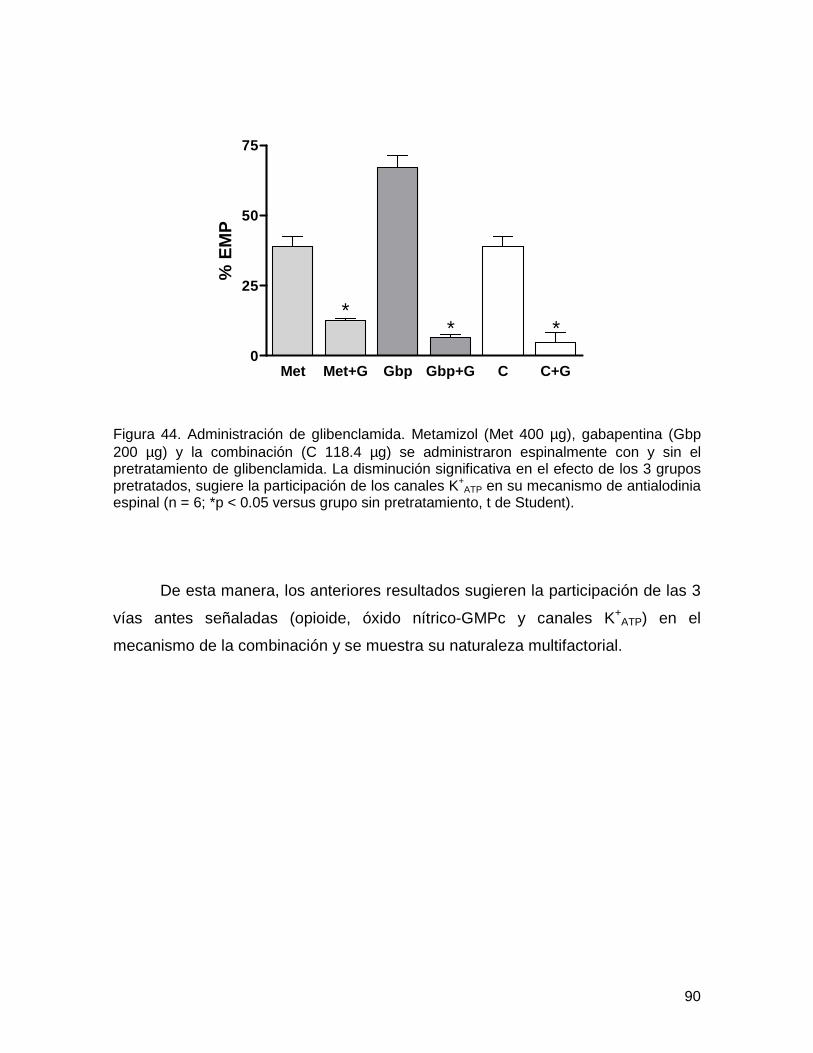
90
Figura 44. Administración de glibenclamida. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron espinalmente con y sin el pretratamiento de glibenclamida. La disminución significativa en el efecto de los 3 grupos pretratados, sugiere la participación de los canales K+
ATP en su mecanismo de antialodinia espinal (n = 6; *p < 0.05 versus grupo sin pretratamiento, t de Student).
De esta manera, los anteriores resultados sugieren la participación de las 3
vías antes señaladas (opioide, óxido nítrico-GMPc y canales K+ATP) en el
mecanismo de la combinación y se muestra su naturaleza multifactorial.
Met Met+G Gbp Gbp+G C C+G0
25
50
75
** *
% E
MP
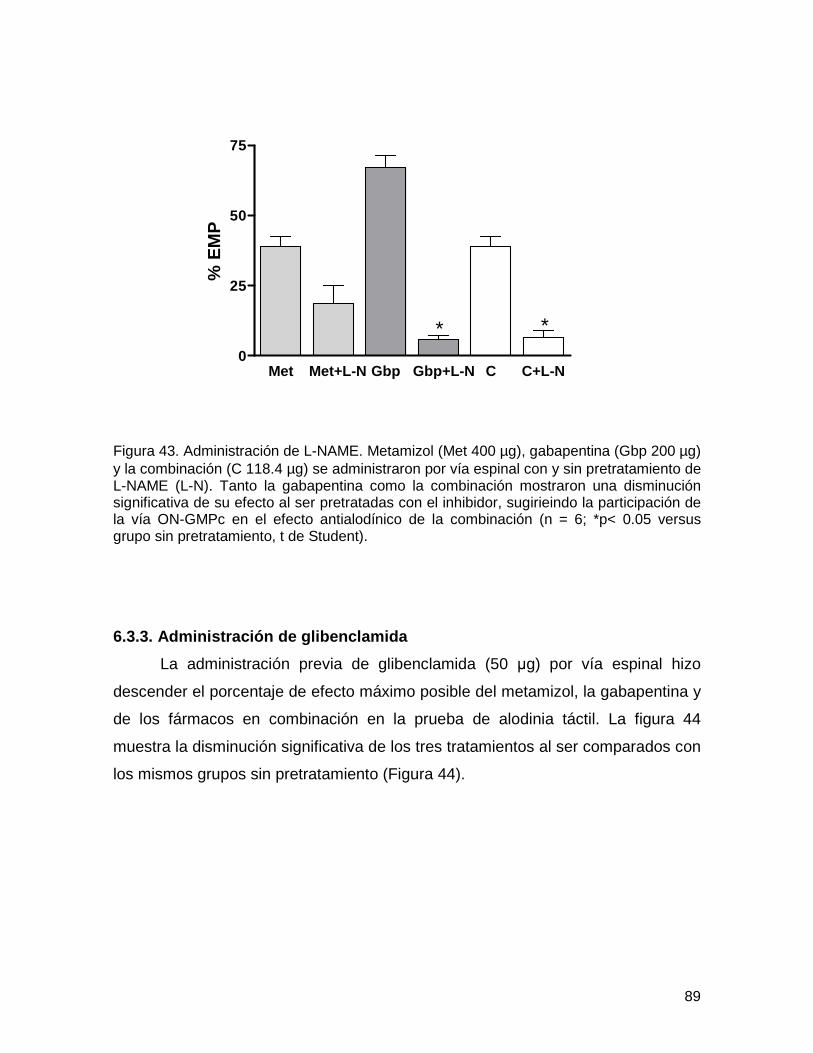
89
Figura 43. Administración de L-NAME. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron por vía espinal con y sin pretratamiento de L-NAME (L-N). Tanto la gabapentina como la combinación mostraron una disminución significativa de su efecto al ser pretratadas con el inhibidor, sugirieindo la participación de la vía ON-GMPc en el efecto antialodínico de la combinación (n = 6; *p< 0.05 versus grupo sin pretratamiento, t de Student).
6.3.3. Administración de glibenclamida
La administración previa de glibenclamida (50 µg) por vía espinal hizo
descender el porcentaje de efecto máximo posible del metamizol, la gabapentina y
de los fármacos en combinación en la prueba de alodinia táctil. La figura 44
muestra la disminución significativa de los tres tratamientos al ser comparados con
los mismos grupos sin pretratamiento (Figura 44).
Met Met+L-N Gbp Gbp+L-N C C+L-N0
25
50
75
* *
% E
MP
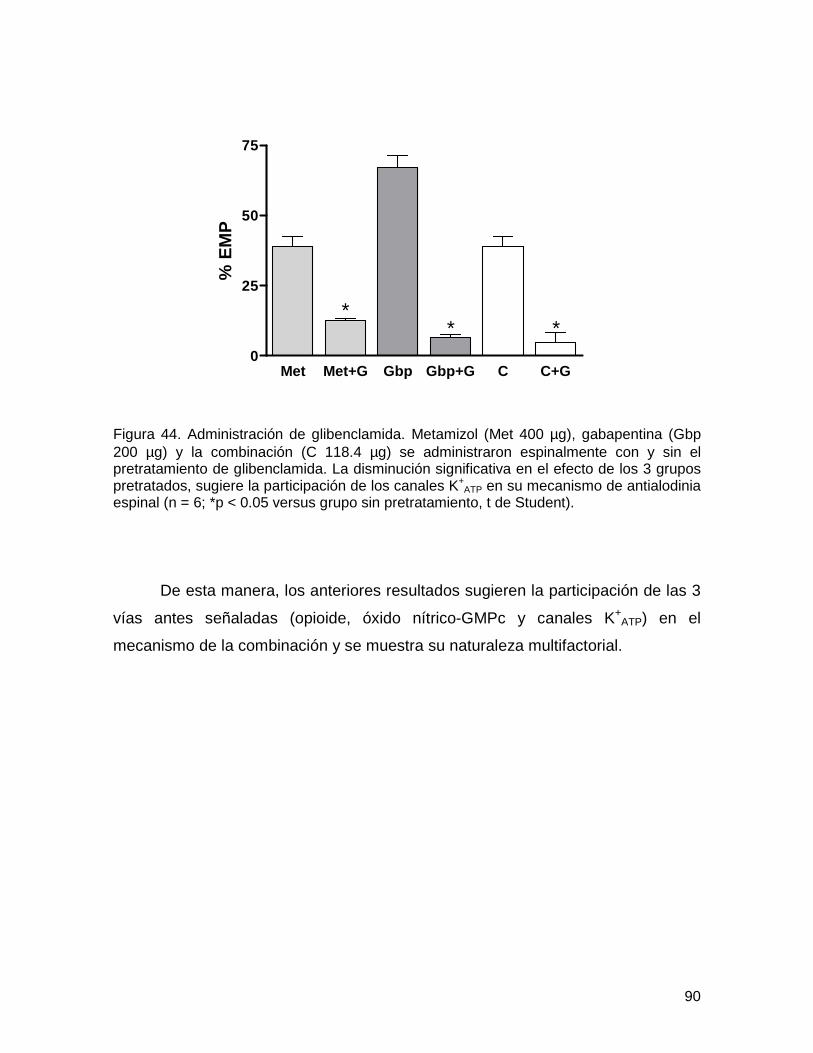
90
Figura 44. Administración de glibenclamida. Metamizol (Met 400 µg), gabapentina (Gbp 200 µg) y la combinación (C 118.4 µg) se administraron espinalmente con y sin el pretratamiento de glibenclamida. La disminución significativa en el efecto de los 3 grupos pretratados, sugiere la participación de los canales K+
ATP en su mecanismo de antialodinia espinal (n = 6; *p < 0.05 versus grupo sin pretratamiento, t de Student).
De esta manera, los anteriores resultados sugieren la participación de las 3
vías antes señaladas (opioide, óxido nítrico-GMPc y canales K+ATP) en el
mecanismo de la combinación y se muestra su naturaleza multifactorial.
Met Met+G Gbp Gbp+G C C+G0
25
50
75
** *
% E
MP

91
7. DISCUSIÓN
7.1. Modelo inflamatorio
7.1.1. Efecto antinociceptivo de gabapentina
La administración oral y espinal de gabapentina redujo de forma limitada
el número de sacudidas en la fase inicial de la prueba de formalina y los
efectos registrados no fueron proporcionales al nivel de dosis. No obstante, se
registró una antinocicepción dependiente de la dosis en la fase II de la prueba
al administrar gabapentina de manera oral e intratecal.
El presente trabajo concuerda con las referencias de la literatura ya que
está bien establecido que la gabapentina reduce las conductas nociceptivas
inducidas por la formalina tras su administración oral (Carlton y Zhuo, 1998;
Yoon y Yaksh, 1999b; Heughan y Sawynok, 2002; Granados-soto y Arguelles,
2005) y espinal (Shimoyama et al, 1997; Yoon y Yaksh, 1999a; Kaneko et al,
2000; Yoon et al, 2003; 2004; Granados-Soto y Argüelles, 2005).
Para este fármaco el mejor efecto fue observado por vía espinal (68.2%)
por un máximo de 53.1% correspondiente a la vía oral, un efecto espinal más
pronunciado ya ha sido reportado por otros autores (Yoon et al, 1999).
El hecho de que la gabapentina no presente una eficacia elevada ni
dependiente de la dosis en la fase inicial de la prueba podría indicar que este
fármaco más que disminuir directamente la actividad de los nociceptores es
capaz de reducir la respuesta inflamatoria inducida por la formalina, tal como lo
han descrito previamente Carlton y Zhuo (1998).
7.1.2. Efecto antinociceptivo de metamizol
Los datos presentes demuestran que la administración oral e intratecal
de metamizol produce un efecto antinociceptivo en ambas fases de la prueba
de la formalina en ratas. El efecto observado en la fase I de la prueba no
sobrepasó el 31% y por ambas vías de administración los efectos no fueron
dosis dependientes. Sin embargo, en la fase inflamatoria se registraron efectos
dependientes de la dosis en ambas vías de administración, de manera similar a
lo ocurrido con gabapentina, la administración espinal de metamizol rindió un
mejor efecto que la vía oral. Los efectos máximos obtenidos fueron 42.9 y

92
48.5% para la vía oral y espinal, respectivamente. Estas observaciones
concuerdan con estudios previos que muestran que el metamizol induce
antinocicepción tras su administración oral (Granados-Soto et al, 1995;
Lorenzetti y Ferreira, 1996; Beirith et al, 1998; Miranda y Pinardi, 2004) o
espinal (Beirith et al, 1998; Miranda y Pinardi, 2004; Vázquez et al, 2005) en
rata y ratón.
La carencia de un efecto pronunciado y dependiente de la dosis
mostrada por el metamizol en la fase I de formalina concuerda en parte con
varios estudios previos (Malmberg y Yaksh, 1992; Correa y Calixto, 1993) que
indican reiteradamente que los analgésicos anti-inflamatorios no esteroideos
son ineficaces para prevenir la respuesta neurogénica inducida por formalina
(fase I) y capsaicina. Sin embargo, otros autores han registrado que el
metamizol presenta una escasa eficacia pero dependiente de la dosis en la
fase inicial de la prueba de la formalina al administrarlo intraperitonealmente en
ratón (Beirith et al, 1998). Las diferencias se pueden deber a la especie y la vía
de administración. No obstante, el efecto discreto del metamizol en esta fase
de la prueba podría sugerir que a diferencia de otros analgésicos anti-
inflamatorios no esteroideos la analgesia del metamizol va más allá de su
capacidad bien conocida de inhibir la síntesis de las prostaglandinas (Hinz et al,
2007).
7.1.3. Efecto antinociceptivo de la combinación de gabapentina y
metamizol
Por vía oral, los efectos observados por la combinación en la fase I de la
prueba de la formalina fueron independientes de la dosis y el efecto no pasó
del 25% de antinocicepción. En la fase II de la prueba el efecto registrado fue
dosis dependiente con un máximo de 78.8%. De igual manera, la
administración espinal produjo un efecto dosis dependiente únicamente en la
fase II de la prueba, observándose un efecto de hasta 44.2%. Una vez
efectuados los correspondientes análisis isobolográficos los índices de
interacción obtenidos indicaron la presencia de sinergismo en ambos casos, el
valor de γ para la vía oral fue de 0.19 lo que implica que la dosis efectiva 30
experimental (DE30E) fue 5 veces menor a lo teóricamente calculado.

93
Intratecalmente γ tuvo un valor de 0.35 señalando que la DE30E fue
aproximadamente un tercio de la dosis efectiva 30 teórica (DE30T).
Combinar analgésicos es una estrategia ideada con el fin de obtener
ventajas como facilitar el apego al tratamiento, simplificar la prescripción,
mejorar la eficacia sin el aumento de los efectos adversos o disminuir estos
efectos colaterales sin perder la eficacia (Raffa, 2001). El objetivo primordial del
presente estudio consistió en analizar la interacción entre gabapentina y
metamizol en varios niveles. Un sinergismo significativo se demuestra como
resultado al emplear la combinación por vía oral y espinal, gracias al uso del
análisis isobolográfico.
El hecho de que en el modelo inflamatorio se observó una interacción
sinérgica más importante tras la administración por vía oral sugiere una posible
ventaja en su uso comparado con la administración espinal. En este sentido,
Miranda y Pinardi (2004) han reportado que el metamizol al combinarse con
clonidina induce una interacción aditiva por vía oral, mientras que resulta
sinérgica al ser administrada intraperitonealmente.
7.2. Modelo Neuropático
7.2.1. Efecto antialodínico de gabapentina
Existen varios reportes en la literatura que indican que la administración
oral y espinal de gabapentina disminuye la alodinia inducida por ligadura de los
nervios espinales L5 y L6 en ratas (Jun y Yaksh, 1998; Field et al, 1999; Chen
et al, 2000; Patel et al, 2001). Los resultados del presente trabajo concuerdan
con lo reportado ya que se observó que la administración de gabapentina
revirtió la alodinia táctil en ratas con ligadura de los nervios espinales. El efecto
máximo observado por vía oral (59%) concuerda en términos generales con
otros estudios previos en los que ha sido empleada la gabapentina por esta vía
de administración (Fox et al, 2003; Reyes-García et al, 2004). Para la vía
espinal, los efectos de gabapentina fueron más pronunciados (67%) y sugieren
una participación significativa del componente espinal en el efecto antialodínico
de este fármaco, como lo han establecido previamente otros autores (Yoon et
al, 1999; Mixcoatl-Zecuatl et al, 2004; 2006; Cheng y Chiou, 2006). Los datos

94
aquí presentados confirman la eficacia antialodínica de gabapentina
administrada por vía oral y espinal.
7.2.2. Efecto antialodínico de metamizol
La administración oral de metamizol no mostró un efecto significativo en
el modelo de Kim y Chung. Este hecho podría constituir un argumento acorde
con la apreciación clínica de que los analgésicos anti-inflamatorios no
esteroideos tienen un efecto muy limitado en el tratamiento del dolor
neuropático (Mc Quay, 2001; Gilron, 2006). Sin embargo, la administración
espinal del metamizol presentó un efecto de 39%. Algunos estudios han
demostrado la utilidad de este tipo de fármacos cuando se administran antes,
pero no después, del establecimiento del dolor neuropático. Tal es el caso de
ketoprofeno (Ossipov et al, 2000) y meloxicam (Takeda et al, 2005). Estos
reportes sugieren que las prostaglandinas tendrían una participación importante
en el desarrollo, pero no en el mantenimiento, del dolor neuropático. La
presente investigación contradice esta hipótesis y sugiere que las
prostaglandinas espinales podrían tener un papel importante en el
mantenimiento del dolor neuropático. De acuerdo con esto se ha reportado un
aumento de la expresión de la ciclooxigenasa 2 en la médula espinal de ratas
sometidas a lesión del nervio ciático o de las raíces espinales L5 y L6 de 2 a 4
semanas posterior al daño (Ma y Eisenach, 2003). De esta manera, se podría
sugerir que el efecto antialodínico espinal del metamizol se debe a la inhibición
de la ciclooxigenasa 2 en este sitio. El hecho de que el metamizol inhibe a la
ciclooxigenasa 2 (Campos et al, 1999) apoya esta sugerencia. Dado que el
metamizol presenta otros mecanismos de acción, no se puede descartar que
dichos mecanismos también participen en su efecto antialodínico espinal.

95
7.2.3. Efecto antialodínico de la combinación de ga bapentina y metamizol
Debido a que el metamizol no produjo efecto antialodínico por vía oral, el
análisis isobolográfico convencional no pudo realizarse para esta vía de
administración. Por lo tanto, se eligió una estrategia diferente donde se utilizan
dosis crecientes de gabapentina (30-300 mg/kg) combinadas con una dosis fija
de metamizol (600 mg/kg). Esta estrategia es usual y un ejemplo al respecto es
el uso de glucosamina como adyuvante de ibuprofeno (Tallarida et al, 2003).
En estas condiciones el metamizol aumentó el efecto máximo de gabapentina
de 59.3% a 77.9% presentando un sinergismo por vía oral. En contraste, la
eficacia antialodínica no prevista del metamizol por vía espinal facilitó el hecho
de que pudiera efectuarse un análisis isobolográfico. Al comparar las dosis
efectivas 30 teórica y experimental se obtuvo un valor de γ de 0.55 lo que
indica un sinergismo por vía espinal.
Para nuestro conocimiento este es el primer estudio sobre la
combinación de gabapentina con un analgésico no esteroideo como el
metamizol. Los resultados encontrados con la combinación por vía oral y
espinal justifican la realización de estudios clínicos con el fin de demostrar su
utilidad en el tratamiento del dolor neuropático en humanos.
7.3. Mecanismo de acción
7.3.1. Mecanismos de gabapentina
El efecto de gabapentina espinal se redujo significativamente después
de la administración intratecal de naltrexona, L-NAME y glibenclamida. Estos
datos sugieren la participación de los receptores opioides, la vía del óxido
nítrico y los canales de potasio sensibles a ATP. Los resultados de la presente
investigación confirman lo descrito por Yoon et al (2002) donde se demuestra
que la naloxona bloquea parcialmente el efecto antinociceptivo de gabapentina
espinal en el modelo de la formalina. Además, los datos son similares a los
reportados por Mixcoatl-Zecuatl et al (2004; 2006) que registran que el L-NAME
y la glibenclamida bloquean el efecto antialodínico de la gabapentina
administrada por la misma vía. Estos hallazgos también son acordes con lo
descrito por Ortiz et al (2006) quienes reportan que la gabapentina tiene la
capacidad de activar la vía del óxido nítrico-GMPc-canales K+ a nivel periférico.

96
En marcado contraste, también se ha reportado que el efecto antihiperalgésico
de gabapentina es insensible a la naloxona (Field et al, 1997). Las diferencias
en este sentido pueden deberse al modelo de dolor utilizado. En resumen, el
efecto antialodínico de gabapentina se podría deber a la activación de la vía
óxido nítrico-GMPc-canales K+ lo que conduciría a la hiperpolarización de las
neuronas sensoriales y consecuentemente a la reducción del dolor en
animales.
Por otro lado, este fármaco también es capaz de activar otros blancos
terapéuticos. Es posible que el efecto antinociceptivo y antialodínico de la
gabapentina resulte de su unión a la subunidad α2δ de los canales de calcio
dependientes de voltaje tipo L como ha sido previamente descrito (Gee et al,
1996) o de los canales tipo N (Saegusa et al, 2001). Se ha reportado que la
subunidad α2δ aumenta su expresión después del daño al nervio (Luo et al,
2001; 2002). Tomados juntos, estos datos sugieren que la eficacia de
gabapentina se debe al bloqueo de esta subunidad sobreexpresada. Además,
la gabapentina puede interactuar de manera indirecta con los receptores NMDA
debido a que la D-Serina, agonista del sitio a glicina de los receptores NMDA,
puede revertir la actividad anti-hiperalgésica de la gabapentina (Singh et al,
1996). La gabapentina también es capaz de bloquear la hiperalgesia térmica
inducida por la administración intratecal de NMDA (Partridge et al, 1998).
Recientemente, un estudio electrofisiológico demostró que la gabapentina
inhibe de manera presináptica la neurotransmisión glutamatérgica en las astas
dorsales (Shimoyama y Shimoyama, 2000).
7.3.2. Mecanismos de metamizol
El efecto de metamizol espinal se redujo significativamente después de
la administración intratecal de naltrexona y glibenclamida. Debido a que la
naltrexona y la glibenclamida son inhibidores de los receptores opioides
(Yaksh, 1987; Shaiova et al, 2007) y canales de potasio sensibles a ATP,
respectivamente (Amoroso et al, 1990; Davis et al, 1991; Mixcoatl-Zecuatl et al,
2004), los datos sugieren la participación de los receptores opioides y los
canales de potasio sensibles a ATP en su efecto antialodínico. Los resultados
aquí expuestos están de acuerdo con reportes previos que demuestran que la
naloxona, otro antagonista de los receptores opioides, bloquea el efecto

97
antinociceptivo de metamizol en diferentes modelos (Taylor et al, 1998;
Vanegas y Tortorici, 2002; Vázquez et al, 2005; Hernández-Delgadillo y Cruz,
2006). Además, se ha reportado que la microinyección directa de metamizol en
la sustancia gris periacueductal (Carlsson y Jurna, 1987; Hernández y
Vanegas, 2001) así como en el núcleo de raphé magno (Tortorici y Vanegas,
1994; Vázquez et al, 2005) induce efectos antinociceptivos que conducen a la
estimulación del sistema inhibitorio descendente, tal como lo hacen los
opioides. La reducción del efecto del metamizol con el pretratramiento de
naltrexona en la presente investigación resulta acorde con dichos reportes.
Los datos también concuerdan con el reporte de Alves y Duarte (2002)
que demuestra que la glibenclamida bloquea el efecto antinociceptivo de
metamizol sugiriendo la participación de los canales de potasio sensibles a
ATP. En el mismo sentido, se ha reportado que el metamizol genera una
corriente de potasio mediante la apertura de estos canales en células de
músculo liso vascular (Valenzuela et al, 2005). En contraste, otro estudio
descarta la participación de los canales de potasio sensibles a ATP y sugiere la
participación de los canales de potasio activados por calcio de conductancia
alta y baja (Ortiz et al, 2003). En el protocolo presente no se observó un
bloqueo del L-NAME al efecto antialodínico de metamizol en el modelo de Kim
y Chung. En este sentido, un estudio publicado por Reis et al (2003) destaca
que el efecto anticonvulsivo del metamizol, en un modelo de convulsiones
audiogénicas, no pudo ser abatido por el pretratamiento del L-NOARG (otro
inhibidor de la sintasa de óxido nítrico). No obstante, también está reportado
que el L-NAME es capaz de bloquear las acciones antinociceptivas del
metamizol (Granados-Soto et al, 1995; Lorenzetti y Ferreira, 1996) en dolor
inflamatorio. Las diferencias observadas con estos últimos reportes pueden
deberse a la vía de administración y principalmente al tipo de dolor.
Los datos reunidos sugieren que la acción antialodínica de metamizol se
debe al menos a una acción tipo opioide y a la apertura de canales de potasio
sensibles a ATP. Pero no permiten descartar la participación de otros
mecanismos.

98
La eficacia de este compuesto se relaciona principalmente con su
capacidad de interferir con la vía del ácido araquidónico al inhibir a las ciclo-
oxigenasas (1, 2, y 3) y la subsecuente producción de prostaglandinas a nivel
periférico y central (Shimada et al, 1994; Campos et al, 1999; Chandrasekharan
et al, 2002; Pierre et al, 2007). Se ha sugerido que el efecto del metamizol se
debe a la acción inhibitoria de su metabolito 4-metil amino antipirina de manera
equipotente de las isoformas 1 y 2 (Hinz et al, 2007). Sin embargo, el perfil de
eficacia mostrado por el metamizol en ese estudio parece ser más parecido al
de los inhibidores selectivos de la ciclooxigenasa 2. Mientras que su perfil
reducido de efectos adversos se debe a su escasa acidez más que a la
inhibición diferencial sobre la ciclooxigenasa 1 en voluntarios (Hinz et al, 2007).
En marcado contraste, se ha reportado que el metamizol inhibe
preferentemente a la ciclooxigenasa 2 en función de sus concentraciones
inhibitorias 50 en humanos (12 contra 486 µg/ml; Campos et al, 1999). Otros
autores han sugerido que el metamizol debe su efecto a la inhibición selectiva
de la ciclooxigenasa 3 (Chandrasekharan et al, 2002), ahora conocida como
ciclooxigenasa 1B.
El metamizol también parece estar involucrado en la modulación de la
liberación o actividad de los aminoácidos excitatorios en la médula espinal, ya
que interfiere parcialmente con el sitio de unión a glutamato en membranas de
neuronas de rata y ratón. Además de que el metamizol puede reducir de
manera dependiente de la dosis la hiperalgesia inducida por la inyección de
glutamato en ratón (Beirith et al, 1998).
7.3.3. Combinaciones de gabapentina y metamizol
Para nuestro conocimiento, este es el primer reporte de una
combinación farmacológica entre gabapentina y metamizol. No obstante, la
gabapentina previamente se ha coadministrado con ibuprofeno (Yoon y Yaksh,
1999), naproxeno (Hurley et al, 2002), diclofenaco (Picazo et al, 2006),
amitriptilina (Heughan y Sawynok, 2002), serotonina (Yoon et al, 2005),
clonidina y neostigmina (Yoon et al, 2004), tramadol (Granados-soto y
Arguelles, 2005) y morfina (Shimoyama et al, 1997; Meymandi et al, 2006)
observándose efectos sinergísticos en modelos de dolor inflamatorio. Además,
en dolor neuropático la gabapentina se ha combinado con CNQX (antagonista

99
de los receptores AMPA, Chen et al, 2000), CI-1021 (antagonista de los
receptores NK1, Field et al, 2002), morfina (Matthews y Dickenson, 2002),
vitaminas B (Reyes-García et al, 2004) y donepezil (inhibidor de la
colinesterasa, Hayashida et al, 2007).
Por otro lado, el metamizol ha mostrado efectos supra-aditivos al
combinarse con vitaminas del complejo B (Muller et al, 1985), verapamilo
(Vlaskovska et al, 1989), metadona (Lehmann et al, 1990), morfina (Aguirre-
Bañuelos y Granados-Soto, 1999; Hernández-Delgadillo et al, 2002;
Hernández-Delgadillo y Cruz, 2006), tramadol (Stamer et al, 2003) y clonidina
(Miranda y Pinardi, 2004).
Estos reportes reunidos, apoyan la idea de que ambos fármacos tienen
un enorme potencial de combinación en función de que poseen múltiples
mecanismos de acción.
7.3.4. Mecanismo de la combinación de gabapentina y metamizol
La coadministración de gabapentina y metamizol produce una
interacción sinérgica significativa en las diferentes combinaciones analizadas
en el protocolo presente. Además, el efecto antialodínico de la combinación
gabapentina-metamizol se redujo significativamente después de la
administración intratecal de naltrexona, L-NAME y glibenclamida. Estos datos
sugieren la participación de los receptores opioides, la vía del óxido nítrico y los
canales de potasio sensibles a ATP en el efecto antialodínico de la
combinación. Para nuestro conocimiento este es el primer reporte del efecto
antinociceptivo y antialodínico de la combinación así como de su bloqueo por
los inhibidores antes mencionados. Sin embargo, con los datos que aquí se
exponen no se puede descartar la posible participación de otros mecanismos
en la eficacia de la combinación. La interacción sinérgica podría tener origen en
la multiplicidad de acciones y sitios de acción de ambos fármacos, de hecho, el
tipo de actividad y los sitios de acción en apariencia son complementarios. La
unión de gabapentina a la subunidad α2δ de los canales de calcio dependientes
de voltaje (Taylor et al, 1998), combinada con la activación de receptores
opioides, la vía del óxido nítrico y la apertura de canales de potasio sensibles a
ATP por acción de ambos fármacos podría ser la base de la interacción supra-
aditiva entre estos compuestos. Además, tanto gabapentina como metamizol

100
comparten la capacidad de interaccionar, al menos de manera indirecta, con la
liberación de los aminoácidos excitatorios, como el glutamato (Beirith et al,
1998; Shimoyama y Shimoyama, 2000). Por lo que está claro que el
mecanismo de acción de la combinación es de origen multifactorial.
.

101
7. CONCLUSIONES
1. La gabapentina y el metamizol reducen de manera significativa y
dependiente de la dosis la fase II de la prueba de la formalina, por vía
oral y espinal.
2. La combinación de gabapentina y metamizol presenta una interacción
antihiperalgésica sinérgica mayor en la vía oral que en la espinal en la
prueba de formalina.
3. La gabapentina disminuye signifcativamente la alodinia inducida por el
modelo de Kim y Chung de manera dosis dependiente por vía oral y
espinal.
4. El metamizol oral no presenta efecto en este modelo de ligadura de los
nervios espinales L5 y L6 como la mayoría de los AINEs. Sin embargo,
mostró eficacia al administrarse por vía espinal.
5. La combinación de gabapentina y metamizol presenta efectos
antialodínicos supra-aditivos tras su administración oral y espinal.
6. El efecto de la combinación de gabapentina y metamizol es producto de
un mecanismo de acción multifactorial en el que al menos interviene la
activación de los receptores opioides, la vía del óxido nítrico-GMPc y la
apertura de canales de potasio sensibles a ATP.
7. El empleo de la combinación de gabapentina y metamizol puede ser útil
en el tratamiento del dolor inflamatorio y neuropático en humanos.

102
8. BIBLIOGRAFÍA
Abbracchio MP, Verderio C: Pathophysiological roles of P2 receptors in glial cells. Novartis Found Symp 276: 91-103, 2006. Ambriz-Tututi M, Granados-Soto V: Oral and spinal melatonin reduces tactile allodynia in rats via activation of MT2 and opioid receptors. Pain 132: 273-280, 2007. Aguirre-Bañuelos P y Granados-Soto V: Evidence for a peripheral mechanism of action for the potentiation of the antinociceptive effect of morphine by dipyrone. J Pharmacol Toxicol Methods 42: 79-85, 1999. Amoroso S, Schmid-Antomarchi H, Fosset M, Lazdunski M: Glucose, sulfonylureas, and neurotransmitter release: role of ATP-sensitive K+ channels. Science 247: 852-854. 1990. Akman H, Aksu F, Gültekin I, Ozbek H, Oral U, Doran F, Baysal F: A possible central antinociceptive effect of dipyrone in mice. Pharmacology 53: 71-78, 1996. Alves D, Duarte I: Involvement of ATP-sensitive K+ channels in the peripheral antinociceptive effect induced by dipyrone. Eur J Pharmacol 444: 47-52, 2002. Anand KJS, Craig KD: New perspectives on the definition of pain. Pain 67: 3-6, 1996. Arcila-Herrera H, Barragán-Padilla S, Borbolla-Escoboza JR, Canto-Solis A, Castañeda-Hernández G, de León-Gonzalez M, Genis-Rondero MA, Granados-Soto V, Gutiérrez-Garcia JL, Hernández-Hernández S, Kassian-Rank A, Lara-Perera V, Vargas-Correa JB: Consensus of a group of Mexican experts: efficacy and safety of metamizol (Dipirone). Gac Med Mex 140: 99-101, 2004. Ariniello L, McComb M and Carey J: Brain Facts. Society for Neurosciences: 1-63, 2002. Baba H, Doubell TP, Woolf CJ: Peripheral inflammation facilitates Abeta fiber-mediated synaptic input to the substantia gelatinosa of the adult rat spinal cord. J Neurosci 19: 859-867, 1999. Bao L, Wang HF, Cai HJ, Tong YG, Jin SX, Lu YJ, Grant G, Hokfelt T, Zhang X. Peripheral axotomy induces only very limited sprouting of coarse myelinated afferents into inner lamina II of rat spinal cord. Eur J Neurosci 16: 175-185, 2002.
Bardin L, Lavarenne J, Eschalier A: Serotonin receptor subtypes involved in the spinal antinociceptive effect of 5-HT in rats. Pain 86:11-18, 2000.

103
Beck PW, Handwerker HO: Bradykinin and serotonin effects on various types of cutaneous nerve fibers. Pflugers Arch 347: 209-222, 1974. Beirith A, Santos AR, Rodrigues AL, Creczynski-Pasa TB, Calixto JB: Spinal and supraspinal antinociceptive action of dipyrone in formalin, capsaicin and glutamate tests. Study of the mechanism of action. Eur J Pharmacol 345: 233-245, 1998. Bessou P, Perl ER: Response of cutaneous sensory units with unmyelinated fibers to noxious stimuli. J Neurophysiol 32:1025-1043, 1969. Bevan S, Geppetti P: Protons: small stimulants of capsaicin-sensitive sensory nerves. Trends Neurosci 12: 509-512, 1994. Bevan S, Yeats J: Protons activate a cation conductance in a sub-population of rat dorsal root ganglion neurones. J Physiol 433: 145-161, 1991.
Bhargava VK, Saha L: Serotonergic mechanism in imipramine induced antinociception in rat tail flick test. Indian J Physiol Pharmacol 45: 107-110, 2001.
Bingham S, Beswick PJ, Blum DE, Gray NM, Chessell I: The role of the cylooxygenase pathway in nociception and pain. Semin Cell Dev Biol 17: 544-554, 2006. Birch PJ, Harrison SM, Hayes AG, Rogers H, Tyers MB: the non-peptide NK1 receptor antagonist (±)-CP- 96, 345, produces antinociceptive and anti-oedema effects in the rat. Br J Pharmacol 105: 508-510, 1992. Birell GJ, McQueen DS, Iggo A, Colman RA, Grubb BD: PGI2-induced activation and sensitization of articular mechanonociceptors. Neurosci lett 124: 5-8, 1991. Bleehan T, Keele CA: Observations on the algogenic actions of adenosine compounds on the human blister base preparation. Pain 3: 367-377, 1977. Bolan EA, Tallarida RJ, Pasternak GW. Synergy between mu opioid ligands: evidence for functional interactions among mu opioid receptor subtypes. J Pharmacol Exp Ther 303: 557-562, 2002. Bolten W, Kempel-Waibel A, Pförringer W: Analysis of the cost of illness in backache. Med Klin (Munich) 93: 388-393, 1998. Bridges D, Thompson SW, Rice AS: Mechanisms of neuropathic pain. Br J Anaesth 87: 12-26, 2001. Brooks J, Tracey I: From nociception to pain perception: imaging the spinal and supraspinal pathways. J Anat 207: 19-33, 2005.

104
Brune K, Alpermann H: Non-acidic pyrazoles: inhibition of prostaglandin production, carrageenan oedema and yeast fever. Agents Actions 13: 360-363, 1983. Burnstock G, Wood JN: Purinergic receptors: their role in nociception and primary afferent neurotransmission. Curr Opin Neurobiol 6: 526-532, 1996. Buzzi MG, Moskowitz MA: The antimigraine drug, sumatriptan (GR43175), selectively blocks neurogenic plasma extravasation from blood vessels in dura mater. Br J Pharmacol 99: 202-226, 1990. Campos C, de Gregorio R, García-Nieto R, Gago F, Ortiz P, Alemany S Regulation of cyclooxygenase activity by metamizol. Eur J Pharmacol 378: 339-347, 1999. Cárdenas CG, Del Mar LP, Cooper BY, Scroggs RS: 5HT4 receptors couple positively to tetrodotoxin-insensitive sodium channels in a subpopulation of capsaicin-sensitive rat sensory neurons. J Neurosci 17: 7181-7189, 1997. Carlsson KH, Jurna I: The role of descending inhibition in the antinociceptive effects of the pyrazolone derivatives, metamizol (dipyrone) and aminophenazone ("Pyramidon"). Naunyn Schmiedebergs Arch Pharmacol 335: 154-159, 1987. Carlton SM, Zhou S, Coggeshall RE: Evidence for the interaction of glutamate and NK1 receptors in the periphery. Brain Res 90: 160-169, 1998. Carlton SM, Zhou S: Attenuation of formalin-induced nociceptive behaviors following local peripheral injection of gabapentin. Pain 76: 201-207, 1998. Cesare P, McNaughton P: Peripheral pain mechanisms. Curr Opin Neurobiol 7: 493-499, 1997. Chandrasekharan NV, Dai H, Roos KL, Evanson NK, Tomsik J, Elton TS, Simmons DL: COX-3, a cyclooxygenase-1 variant inhibited by acetaminophen and other analgesic/antipyretic drugs: Cloning, structure, and expression. Proc Natl Acad Sci USA 99: 13926-13931, 2002. Chandrasekharan NV, Simmons DL: The cyclooxygenases. Genome Biol 5: 241-248, 2004. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL: Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods 53: 55-63, 1994. Chen CC, Akopian AN, Sivilotti L, Colquhoun D, Burnstock G, Wood JN: A P2X purinoceptor expressed by a subset of sensory neurons. Nature 377: 428-431, 1995.

105
Chen SR, Eisenach JC, McCaslin PP, Pan HL: Synergistic effect between intrathecal non-NMDA antagonist and gabapentin on allodynia induced by spinal nerve ligation in rats. Anesthesiology 92: 500-506, 2000. Cheng JK, Pan HL, Eisenach JC Antiallodynic effect of intrathecal gabapentin and its interaction with clonidine in a rat model of postoperative pain. Anesthesiology 92: 1126-1131, 2000. Cheng JK, Chiou LC: Mechanisms of the antinociceptive action of gabapentin. J Pharmacol Sci 100: 471-486, 2006. Cheshire DR: Use of nitric oxide synthase inhibitors for the treatment of inflammatory disease and pain. I Drugs 4: 795-803, 2001. Ciancarelli I, Tozzi-Ciancarelli M, Spacca G, Di Massimo C, Carolei A: Relationship between biofeedback and oxidative stress in patients with chronic migraine. Cephalalgia (en prensa), 2007. Coleman RA, Smith WL, Narumiya S: International Union of Pharmacology classification of prostanoid receptors: properties, distribution, and structure of the receptors and their subtypes. Pharmacol Rev 46: 205-229, 1994. Collier HB, Duchon G: Determination of traces of calcium in commercial adenosine triphosphate (ATP). Anal Biochem 15: 367-368, 1966. Correa CR, Calixto JB: Evidence for participation of B1 and B2 kinin receptors in formalin-induced nociceptive response in the mouse. Br J Pharmacol 110: 193-198, 1993. Costigan y Woolf 1999 Transcriptional and posttranslational plasticity and the generation of inflammatory pain. Proc Natl Acad Sci USA 96: 7723-7730, 1999. Costigan M, Woolf CJ: Pain: molecular mechanisms. J Pain 1: 35-44, 2000. Crunkhorn P, Willis AL: Interaction between prostaglandins E and F given intradermally in the rat. Br J Pharmacol 41: 507-512, 1971. Cunha JM, Sachs D, Canetti CA, Poole S, Ferreira SH, Cunha FQ: The critical role of leukotriene B4 in antigen-induced mechanical hyperalgesia in immunised rats. Br J Pharmacol 139: 1135-1145, 2003. Dagnino J: Boletín de la Escuela de Medicina. Universidad Católica de Chile. 23: 148-151, 1994. D´Amours RH, Ferrante FM : Postoperative pain management. J Orthop Sports Phys Ther 24: 227-236, 1996.

106
Davis NW, Standen NB, Stanfield PR: ATP-dependent K+ channels of muscle cells-their properties, regulation, and functions. J Bioenerg Biomembr 23: 509-535, 1991. Dawson LA, Nguyen HQ, Li P: In vivo effects of the 5-HT6 antagonist SB-271046 on striatal and frontal cortex extracellular concentrations of noradrenaline, dopamine, 5-HT, glutamate and aspartate. Br J Pharmacol 130: 23-26, 2000. De lima L, Bruera E: The role of internacional treaties in the opioid availability process: Relationship between INCB, national goverments, the pharmaceutical Industry and Physicians. Prog Pal Care 8: 128-131, 2000. Del Mar LP, Cárdenas CG, Scroggs RS: Serotonin inhibits high-threshold Ca2+ channel currents in capsaicin-sensitive acutely isolated adult rat DRG neurons. J Neurophysiol 72: 2551-2554, 1994. Devor M: Neuropathic pain and injured nerve: peripheral mechanisms. Br Med Bull 47: 619-630, 1991. Diamond J, Foerster A, Holmes M, Coughlin M: Sensory nerves in adult rats regenerate and restore sensory function to the skin independently of endogenous NGF. J Neurosci 12: 1467-1476, 1992. Dinc E, Baleanu D and Onur F: Spectrophotometric multicomponent analysis of a mixture of metamizol, acetaminophen and caffeine in pharmaceutical formulations by two chemometric techniques. J Pharm Biomed Anal 26: 949-957, 2001. Dinchuk JE, Liu RQ, Trzaskos JM: COX-3: in the wrong frame in mind. Immunol Lett 86: 121, 2003. Dixon WJ: Efficient analysis of experimental observations. Ann Rev Pharmacol Toxicol 20: 441-462, 1980. Doak GJ, Sawynok J: Formalin-induced nociceptive behavior and edema: involvement of multiple peripheral 5-hydroxytryptamine receptor subtypes. Neuroscience 80: 939-949, 1997. Dooley DJ, Mieske CA, Borosky SA: Inhibition of K+-evoked glutamate release from rat neocortical and hippocampal slices by gabapentin. Neurosci Lett 18; 280: 107-110, 2000. Dray A, Pinnock RD: Effects of substance P on adult rat sensory ganglion neurones in vitro. Neurosci Lett 33: 61-66, 1982. Dray A: Neuropharmacological mechanisms of capsaicin and related substances. Biochem Pharmacol 44: 611-615, 1992.

107
Dray A, Perkins M: Bradykinin and inflammatory pain. Trends Neurosci 16: 99-104, 1993. Dray A: Tasting the inflammatory soup: Role of peripheral neurones. Pain Reviews 1: 153-171, 1994. Dray A : Inflammatory mediators of pain. Br J Anaesth 75: 125-131, 1995. Dawson LA, Nguyen HQ, Li P: In vivo effects of the 5-HT6 antagonist SB-271046 on striatal and frontal cortex extracellular concentrations of noradrenaline, dopamine, 5-HT, glutamate and aspartate. Br J Pharmacol 130: 23-26, 2000. Duarte ID, dos Santos IR, Lorenzetti BB, Ferreira SH: Analgesia by direct antagonism of nociceptor sensitization involves the arginine-nitric oxide-cGMP pathway. Eur J Pharmacol 217: 225-227, 1992. Dubner R, Ruda MA: Activity-dependent neuronal plasticity following tissue injury and inflammation. Trends Neurosci 15: 96-103, 1992. Dubner R: The neurobiology of persistent pain and its clinical implications. Suppl Clin Neurophysiol 57: 3-7, 2004. Durmus M, Kadir But A, Saricicek V, Ilksen Toprak H, Ozcan Ersoy M: The post-operative analgesic effects of a combination of gabapentin and paracetamol in patients undergoing abdominal hysterectomy: a randomized clinical trial. Acta Anaesthesiol Scand 51: 299-304, 2007. Eckhardt K, Ammon S, Hofmann U, Riebe A, Gugeler N, Mikus G: Gabapentin enhances the analgesic effect of morphine in healthy volunteers. Anesth Analg 91: 185-191, 2000. England S, Heblich F, James IF, Robbins J, Docherty RJ: Heblich et al, 2001; Bradykinin evokes a Ca2+-activated chloride current in non-neuronal cells isolated from neonatal rat dorsal root ganglia. J Physiol 530: 395-403, 2001. Fassoulaki A, Melemeni A, Stamatakis E, Petropoulos G, Sarantopoulos C: A combination of gabapentin and local anaesthetics attenuates acute and late pain after abdominal hysterectomy. Eur J Anaesthesiol 24: 521-528, 2007 Ferreira MA: Cytokine expression in allergic inflammation: systematic review of in vivo challenge studies. Mediators Inflamm 12: 259-67, 2003. Ferreira J, Trichês KM, Medeiros R, Calixto JB: Mechanisms involved in the nociception produced by peripheral protein kinase C activation in mice. Pain 117: 171-181, 2005.

108
Fialip J, Porteix A, Marty H, Eschalier A, Duchêne-Marullaz P: Lack of importance of caffeine as an analgesic adjuvant of dipyrone in mice. Arch Int Pharmacodyn Ther 302: 86-95, 1989. Field MJ, McCleary S, Hughes J, Singh L: Gabapentin and pregabalin, but not morphine and amitriptyline, block both static and dynamic components of mechanical allodynia induced by streptozocin in the rat. Pain 80: 391-398, 1999.
Field MJ, Gonzalez MI, Tallarida RJ, Singh L: Gabapentin and the neurokinin1 receptor antagonist CI-1021 act synergistically in two rat models of neuropathic pain. J Pharmacol Exp Ther 303: 730-735, 2002. Flatters SJ, Fox AJ, Dickenson AH: Spinal interleukin-6 (IL-6) inhibits nociceptive transmission following neuropathy. Brain Res 984: 54-62, 2003. Fox A, Gentry C, Patel S, Kesingland A, Bevan S: Comparative activity of the anti-convulsants oxcarbazepine, carbamazepine, lamotrigine and gabapentin in a model of neuropathic pain in the rat and guinea-pig. Pain 105: 355-362, 2003. Freiman TM, Kukolja J, Heinemeyer J, Eckhardt K, Aranda H, Rominger A, Dooley DJ, Zentner J, Feuerstein TJ: Modulation of K+-evoked [3H]-noradrenaline release from rat and human brain slices by gabapentin: involvement of KATP channels. Naunyn Schmiedebergs Arch Pharmacol 363: 537-542, 2001. Fukui M, Nakagawa T, Minami M, Satoh M, Kaneko S: Inhibitory role of supraspinal P2X3/P2X2/3 subtypes on nociception in rats. Mol Pain 2: 19, 2006.
Galluzi KE: Management of neuropathic pain. J Am Osteopath Assoc 105: S12-S19, 2005.
García-Martínez JM, Fresno Vara JA, Lastres P, Bernabéu C, Betés PO, Martín-Pérez J: Effect of metamizol on promyelocytic and terminally differentiated granulocytic cells. Comparative analysis with acetylsalicylic acid and diclofenac. Biochem Pharmacol 65: 209-217, 2003.
Gee NS, Brown JP, Dissanayake VU, Offord J, Thurlow R, Woodruff GN The novel anticonvulsant drug, gabapentin (Neurontin), binds to the alpha2delta subunit of a calcium channel. J Biol Chem 271: 5768-5776, 1996. Gessner PK, Cabana BE: A study of the interaction of the hypnotic effects and of the toxic effects of chloral hydrate and ethanol. J Pharmacol Exp Ther 174: 247-259, 1970. Gilron I, Bailey JM, Tu D, Holden RR, Weaver DF, Houlden RL: Morphine, gabapentin, or their combination for neuropathic pain. N Engl J Med 352: 1324-1334, 2005.

109
Gilron I, Watson CP, Cahill CM, Moulin DE: Neuropathic pain: a practical guide for the clinician. CMAJ 175: 265-275, 2006. Giordano J, Schultea T: Serotonin 5-HT3 receptor mediation of pain and anti-nociception: implications for clinical therapeutics. Pain Physician 7: 141-147, 2004. Gold MS, Levine JD, Correa AM: Modulation of TTX-R INa by PKC and PKA and their role in PGE2-induced sensitization of rat sensory neurons in vitro. J Neurosci 18: 10345-10355, 1998. Gracely RH, Grant MA, Giesecke T: Evoked pain measures in fibromyalgia. Best Pract Res Clin Rheumatol 17: 593-609, 2003. Granados-Soto V, Flores-Murrieta FJ, Castañeda-Hernández G, López-Muñoz FJ: Evidence for the involvement of nitric oxide in the antinociceptive effect of ketorolac.Eur J Pharmacol 277: 281-284, 1995. Granados-Soto 1997 Granados-Soto V, Rufino MO, Gomes Lopes LD, Ferreira SH: Evidence for the involvement of the nitric oxide-cGMP pathway in the antinociception of morphine in the formalin test. Eur J Pharmacol 340: 177-180, 1997. Granados-soto V, Arguelles CF: Synergic antinociceptive interaction between tramadol and gabapentin after local, spinal and systemic administration. Pharmacology 74: 200-208, 2005. Graven-Nielsen T, Arendt-Nielsen L: Peripheral and central sensitization in musculoskeletal pain disorders: an experimental approach. Curr Rheumatol Rep 4: 313-321, 2002. Gustorff B, Hoechtl K, Sycha T, Felouzis E, Lehr S, Kress HG: The effects of remifentanil and gabapentin on hyperalgesia in a new extended inflammatory skin pain model in healthy volunteers. Anesth Analg 98: 401-407, 2004. Hackental E: Paracetamol and metamizol in the treatment of chronic pain syndromes. Schmerz 11: 269-275, 1997. Hall JM: Bradykinin receptors: Pharmacological properties and biological roles. Pharmacol Therapeutics 56: 131-190, 1992. Hall JM, Geppetti P: Kinins and kinin receptors in the nervous system. Neurochem Int 26: 17-26, 1995. Handwerker HO, Forster C, Kirchhoff C: Discharge patterns of human C-fibers induced by itching and burning stimuli. J Neurophysiol 66: 307-315, 1991.

110
Hayashida K, Parker R, Eisenach JC: Oral gabapentin activates spinal cholinergic circuits to reduce hypersensitivity after peripheral nerve injury and interacts synergistically with oral donepezil. Anesthesiology 106: 1213-1219, 2007. Henry D, Lim LL, Garcia Rodriguez LA, Perez Gutthann S, Carson JL, Griffin M, Savage R, Logan R, Moride Y, Hawkey C, Hill S, Fries JT: Variability in risk of gastrointestinal complications with individual non-steroidal anti-inflammatory drugs: results of a collaborative meta-analysis. BMJ 312: 1563-1566, 1996. Hernández-Delgadillo GP, Ventura Martínez R, Díaz Reval MI, Domínguez Ramírez AM, López-Muñoz FJ: Metamizol potentiates morphine antinociception but not constipation after chronic treatment. Eur J Pharmacol 441: 177-183, 2002. Hernández-Delgadillo GP, Cruz SL: Endogenous opioids are involved in morphine and dipyrone analgesic potentiation in the tail flick test in rats. Eur J Pharmacol 546: 54-59, 2006. Hernández N, Vanegas H: Antinociception induced by PAG-microinjected dipyrone (metamizol) in rats: involvement of spinal endogenous opioids. Brain Res 896: 175-178, 2001. Heughan CE, Sawynok J: The interaction between gabapentin and amitriptyline in the rat formalin test after systemic administration. Anesth Analg 94: 975-980, 2002. Hill R: NK1 (substance P) receptor antagonists--why are they not analgesic in humans? Trends Pharmacol Sci 21: 244-246, 2000. Hinz B, Cheremina O, Bachmakov J, Renner B, Zolk O, Fromm MF, Brune K: Dipyrone elicits substantial inhibition of peripheral cyclooxygenases in humans: new insights into the pharmacology of an old analgesic. FASEB J 21: 2343-2351, 2007. Hinz B, Brune K: Antipyretic analgesics: nonsteroidal antiinflammatory drugs, selective COX-2 inhibitors, paracetamol and pyrazolinones. Handb Exp Pharmacol 177: 65-93, 2007. Hoheisel U, Mense S: The role of spinal nitric oxide in the control of spontaneous pain following nociceptive input. Prog Brain Res 129: 163-72, 2000. Holthusen H, Arndt JO: Nitric oxide evokes pain in humans on intracutaneous injection. Neurosciences Letters 165: 71-74, 1994. Hurley RW, Chatterjea D, Rose Feng M, Taylor CP, Hammond DL: Gabapentin and pregabalin can interact synergistically with naproxen to produce antihyperalgesia. Anesthesiology 97: 1263-1273, 2002.

111
IAAS 1986: Risks of agranulocytosis and aplastic anemia. A first report of their relation to drug use with special reference to analgesics. The International Agranulocytosis and Aplastic Anemia Study. JAMA 256: 1749-1757, 1986, IASP. Pain terms: a current list with definitions and notes on usage. Pain Suppl 3: S215-S221, 1986. Inturrisi CE: The role of N-methyl-D-aspartate (NMDA) receptors in pain and morphine tolerance. Minerva Anestesiol 60: 401-403, 1994. Jarvis MF, Burgard EC, McGaraughty S, Honore P, Lynch K, Brennan TJ, Subieta A, Van Biesen T, Cartmell J, Bianchi B, Niforatos W, Kage K, Yu H, Mikusa J, Wismer CT, Zhu CZ, Chu K, Lee CH, Stewart AO, Polakowski J, Cox BF, Kowaluk E, Williams M, Sullivan J, Faltynek C: A-317491, a novel potent and selective non-nucleotide antagonist of P2X3 and P2X2/3 receptors, reduces chronic inflammatory and neuropathic pain in the rat. Proc Natl Acad Sci USA 24: 17179-17184, 2002. Julius D, Basbaum AI: Molecular mechanisms of nociception. Nature 413: 203-210, 2001. Jun JH, Yaksh TL: The effect of intrathecal gabapentin and 3-isobutyl gamma-aminobutyric acid on the hyperalgesia observed after thermal injury in the rat. Anesth Analg 86: 348-354, 1998. Kajander KC, Sahara Y, Iadarola MJ, Bennett GJ: Dynorphin increases in the dorsal spinal cord in rats with a painful peripheral neuropathy. Peptides 11: 719-728, 1990. Kaneko M, Mestre C, Sánchez EH, Hammond DL: Intrathecally administered gabapentin inhibits formalin-evoked nociception and the expression of Fos-like immunoreactivity in the spinal cord of the rat. J Pharmacol Exp Ther 292: 743-751, 2000. Kennedy C, Assis TS, Currie AJ, Rowan EG: Crossing the pain barrier: P2 receptors as targets for novel analgesics. J Physiol. 553: 683-694, 2003. Kidd BL Urban LA: Mechanisms of inflammatory pain. Br J Anaesth Jul; 87 (1): 3-11, 2001. Kidd BL, Photiou A, Inglis JJ: The role of inflammatory mediators on nociception and pain in arthritis. Novartis Found Symp 260: 122-33, 2004. Kim SH, Chung JM: An experimental model for periphery neuropathy produced by segmental spinal nerve ligation in the rat. Pain 50: 355-363, 1992. King, JC: Therapeutic options for neuropathic pain. J Trauma 62: S7, 2007.

112
Kis B, Snipes JA, Busija DW: Acetaminophen and the cyclooxygenase-3 puzzle: sorting out facts, fictions, and uncertainties. J Pharmacol Exp Ther 315: 1-7, 2005. Kocsis JD, Honmou O: Gabapentin increases GABA-induced depolarization in rat neonatal optic nerve. Neurosci Lett 14; 169: 181-184, 1994. Lai J, Ossipov MH, Vanderah TW, Malan TP Jr, Porreca F: Neuropathic pain: the paradox of dynorphin. Mol Interv 1: 160-167, 2001. Lai J, Hunter JC, Porreca F. The role of voltage-gated sodium channels in neuropathic pain. Curr Opin Neurobiol 13: 291-297, 2003. Lai J, Luo MC, Chen Q, Ma S, Gardell LR, Ossipov MH, Porreca F: Dynorphin A activates bradykinin receptors to maintain neuropathic pain. Nat Neurosci 9: 1534-1540, 2006. Lehmann KA, Abu-Shibika M, Horrichs-Haermeyer G: Postoperative pain therapy with 1-methadone and metamizole. A randomized study within the scope of intravenous on-demand analgesia. Anasth Intensiv Ther Notfall Med 25: 152-159, 1990. Lekan HA, Chung K, Yoon YW, Chung JM, Coggeshall RE: Loss of dorsal root ganglion cells concomitant with dorsal root axon sprouting following segmental nerve lesions. Neuroscience 81: 527-534, 1997. Leslie TA, Emson PC, Dowd PM, Woolf CJ: Nerve growth factor contributes to the up-regulation of growth-associated protein 43 and preprotachykinin A messenger RNAs in primary sensory neurons following peripheral inflammation. Neuroscience 67: 753-761, 1995. Levine JD, Fields HL, Basbaum AI: Peptides and the primary afferent nociceptor. J Neurosci 13: 2273-2286, 1993. Levy M, Flusser D, Zylber-Katz E, Granit L: Plasma kinetics of dipyrone metabolites in rapid and slow acetylators. Eur J Clin Pharmacol 27: 453-458, 1984. Levy M, Zylber-Katz E, Rosenkranz B: Clinical pharmacokinetics of dipyrone and its metabolites. Clin Pharmacokinet 28: 216-234, 1995. Levy M : Hypersensitivity to pyrazolones. Thorax 2: S72-S74, 2000. Lewin GR, Mendell LM : 1993 Nerve growth factor and nociception. Trends Neurosci 16: 353-359, 1993. Lewis, Thomas: Pain. New York: The Macmillan company, 1-192, 1942.

113
Li J, Simone DA, Larson AA: Windup leads to characteristics of central sensitization. Pain 79: 75-82, 1999. Liu CN, Devor M, Waxman SG, Kocsis JD: Subthreshold oscillations induced by spinal nerve injury in dissociated muscle and cutaneous afferents of mouse DRG. J Neurophysiol 87: 2009-2017, 2002. Loewe S: The problem of synergism and antagonism of combined drugs. Arzneimittelforschung 3: 285-290, 1953. Lopez-Muñoz FJ, Villalón CM, Terrón JA, Salazar LA: Analgesic interactions produced by combinations of dipyrone and morphine in the rat. Proc West Pharmacol Soc 37: 17-19, 1994. Lorenzetti BB, Ferreira SH: Mode of analgesic action of dipyrone: direct antagonism of inflammatory hyperalgesia. Eur J Pharmacol 114: 375-381, 1985. Lorenzetti BB, Ferreira SH: Activation of the arginine-nitric oxide pathway in primary sensory neurons contributes to dipyrone-induced spinal and peripheral analgesia. Inflamm Res 45: 308-311, 1996. Luo ZD, Cizkova D: The role of nitric oxide in nociception. Curr Rev Pain 4: 459-466, 2000. Luo ZD, Chaplan SR, Higuera ES, Sorkin LS, Stauderman KA, Williams ME, Yaksh TL: Upregulation of dorsal root ganglion alpha2delta calcium channel subunit and its correlation with allodynia in spinal nerve-injured rats. J Neurosci 21: 1868-1875, 2001. Luo ZD, Calcutt NA, Higuera ES, Valder CR, Song YH, Svensson CI, Myers RR Injury type-specific calcium channel alpha 2 delta-1 subunit up-regulation in rat neuropathic pain models correlates with antiallodynic effects of gabapentin. J Pharmacol Exp Ther 303: 1199-1205, 2002. Ma Q, Woolf CJ: Noxious stimuli induce an N-methyl-D-aspartate receptor-dependent hypersensitivity of the flexion withdrawal reflex to touch: implications for the treatment of mechanical allodynia. Pain 61: 383-390, 1995. Ma W, Eisenach JC: Intraplantar injection of a cyclooxygenase inhibitor ketorolac reduces immunoreactivities of substance P, calcitonin gene-related peptide, and dynorphin in the dorsal horn of rats with nerve injury or inflammation. Neuroscience 121: 681-690, 2003. Ma W, Quirion R: Inflammatory mediators modulating the transient receptor potential vanilloid 1 receptor: therapeutic targets to treat inflammatory and neuropathic pain. Expert Opin Ther Targets 11: 307-320, 2007.

114
Macías MY, Syring MB, Pizzi MA, Crowe MJ, Alexanian AR, Kurpad SN: Pain with no gain: allodynia following neural stem cell transplantation in spinal cord injury. Exp Neurol 201: 335-348, 2006. Maggi CA, Patacchini A, Rovero P, Giancheti A: Tachykinin receptor and tachykinin receptor antagonists. J Autonom Pharm 13: 23-93, 1992. Magnus L: Nonepileptic uses of gabapentin. Epilepsia 1999; 40: S66-S72, 1999. Malmberg AB Yaksh TL: Hyperalgesia mediated by spinal glutamate or substance P receptor blocked by spinal cyclooxygenase inhibition. Science 28: 1276-1279, 1992. Mannion RJ, Costigan M, Decosterd I, Amaya F, Ma QP, Holstege JC, Ji RR, Acheson A, Lindsay RM, Wilkinson GA, Woolf CJ: Neurotrophins: peripherally and centrally acting modulators of tactile stimulus-induced inflammatory pain hypersensitivity. Proc Natl Acad Sci USA 96: 9385-9390, 1999. Matthews EA, Dickenson AH: A combination of gabapentin and morphine mediates enhanced inhibitory effects on dorsal horn neuronal responses in a rat model of neuropathy. Anesthesiology 96: 633-640, 2002. McNamara JO: Emerging insights into the genesis of epilepsy. Nature; 399: A15-22, 1999. McNamara JO: Drugs Effective in the Therapy of the Epilepsies. In: The Pharmacological Basis of Therapeutics 10ª Ed. Editado por Goodman Gilman A, Hardman J.H. and Limbird L. E. McGraw-Hill Companies. Pergamon Press, New York. Chicago, San Francisco, Lisbon, London, Madrid, Mexico City, Milan, New Delhi, San Juan, Seoul, Singapore, Sydney, Toronto: 538-539, 2001. McQuay H: Opioids in chronic non-malignant pain. BMJ 322: 1134-1135, 2001. Meeus M, Nijs J: Central sensitization: a biopsychosocial explanation for chronic widespread pain in patients with fibromyalgia and chronic fatigue syndrome. Clin Rheumatol 26: 465-473, 2007. Meller ST, Gebhart GF: Nitric oxide (NO) and nociceptive processing in the spinal cord. Pain 52: 127-136, 1993. Menke JG, Borkowski JA, Bierilo KK, MacNeil T, Derrick AW, Schneck KA, Ransom RW, Strader CD, Linemeyer DL, Hess JF: Expression cloning of a human B1 bradykinin receptor. J Biol Chem 269: 21583-21586, 1994. Meunier A, Braz J, Cesselin F, Hamon M, Pohl M: Inflammation et doleur: Thérapie génique expérimentale. Medecine/sciences 2004; 20: 325-30.

115
Meymandi et al, 2006 Gabapentin enhances the analgesic response to morphine in acute model of pain in male rats. Pharmacol Biochem Behav 85: 185-189, 2006. Merskey H: The definition of pain. Eur. J. Psychiatry 6: 153-159, 1991. Miao FJ, Levine JD: Neural and endocrine mechanisms mediating noxious stimulus-induced inhibition of bradykinin plasma extravasation in the rat. J Pharmacol Exp Ther 291: 1028-37, 1999. Millan MJ: The induction of pain: an integrative review. Prog Neurobiol 57: 1-164, 1999. Millan MJ: Descending control of pain. Prog Neurobiol 66: 355-474, 2002. Miller BA, Woolf CJ: Glutamate-mediated slow synaptic currents in neonatal rat deep dorsal horn neurons in vitro. J Neurophysiol 76: 1465-1476, 1996. Miranda HF, Pinardi G: Isobolographic analysis of the antinociceptive interactions of clonidine with nonsteroidal anti-inflammatory drugs. Pharmacol Res 50: 273-278, 2004. Mitchell A, Akarasereenont P, Thiermermann C, Flower RJ, Vane JR: Selectivity of nonsteroidal antiinflammatory drugs as inhibitors of constitutive and inducible cycklooxigenase. Proc Natl Acad Sci USA 90: 11693-11697, 1993. Mixcoatl-Zecuatl T, Medina-Santillán R, Reyes-García G, Vidal-Cantú GC, Granados-Soto V: Effect of K+ channel modulators on the antiallodynic effect of gabapentin. Eur J Pharmacol 484: 201-208, 2004. Mixcoatl-Zecuatl T, Flores-Murrieta FJ, Granados-Soto V: The nitric oxide-cyclic GMP-protein kinase G-K+ channel pathway participates in the antiallodynic effect of spinal gabapentin. Eur J Pharmacol 531: 87-95, 2006. Montes A, Warner W, Puig MM: Use of intravenous patient-controlled analgesia for the documentation of synergy between tramadol and metamizol. Br J Anaesth 85: 217-223, 2000. Mobarakeh JI, Sakurada S, Katsuyama S, Kutsuwa M, Kuramasu A, Lin ZY, Watanabe T, Hashimoto Y, Yanai K: Role of histamine H(1) receptor in pain perception: a study of the receptor gene knockout mice. Eur J Pharmacol 391: 81-89, 2000. Mobarakeh JI, Sakurada S, Hayashi T, Orito T, Okuyama K, Sakurada T, Kuramasu A, Watanabe T, Yanai K: Enhanced antinociception by intrathecally-administered morphine in histamine H1 receptor gene knockout mice. Neuropharmacology 42: 1079-1088, 2002.

116
Müller-Fassbender HR, Schlegel H, Sinterhauf U: Treatment of radicular compression syndrome. An open comparative study with pirprofen and a metamizole-vitamin B complex. Fortschr Med 103: 193-196, 1985. Nathan C: Points of control in inflammation. Nature 420: 846-852, 2002. Neumann S, Doubell TP, Leslie T, Woolf CJ: Inflammatory pain hypersensitivity mediated by phenotypic switch in myelinated primary sensory neurons. Nature 384: 360-364, 1996. Noda A, Goromaru T, Tsubone N, Matsuyama K, Iguchi S: In vivo formation of 4-formylaminoantipyrine as a new metabolite of aminopyrine. I. Chem Pharm Bull (Tokyo) 24: 1502-1505, 1976. Numazaki M, Tominaga M: Nociception and TRP Channels. Curr Drug Targets CNS Neurol Disord 3: 479-485, 2004. Oh EJ, Weinreich D: Bradykinin decreases K+ and increases Cl- conductances in vagal afferent neurones of the guinea pig. J Physiol 558: 513-526, 2004. Ohtori S, Takahashi K, Chiba T, Yamagata M, Sameda H, Moriya H: Substance P and calcitonin gene-related peptide immunoreactive sensory DRG neurons innervating the lumbar intervertebral discs in rats. Ann Anat 184: 235-240, 2002. Olesen J : Understanding the biologic basis of migraine. N Engl J Med 331: 1713-1714, 1994. Ortíz MI, Torres-López JE, Castañeda-Hernández G, Rosas R, Vidal-Cantú GC, Granados-Soto V: Pharmacological evidence for the activation of K+ channels by diclofenac. Eur J Pharmacol 438: 85-91, 2002. Ortíz MI, Castañeda-Hernández G, Granados-Soto V: Possible involvement of potassium channels in peripheral antinociception induced by metamizol: lack of participation of ATP-sensitive K+ channels. Pharmacol Biochem Behav 74: 465-470, 2003. Ortíz MI, Medina-Tato DA, Sarmiento-Heredia D, Palma-Martínez J, Granados-Soto V: Possible activation of the NO-cyclic GMP-protein kinase G-K+ channels pathway by gabapentin on the formalin test. Pharmacol Biochem Behav 83: 420-427, 2006. Ossipov MH, Hong Sun T, Malan P Jr, Lai J, Porreca F: Mediation of spinal nerve injury induced tactile allodynia by descending facilitatory pathways in the dorsolateral funiculus in rats. Neurosci Lett 290: 129-132, 2000. Partridge BJ, Chaplan SR, Sakamoto E, Yaksh TL: Characterization of the effects of gabapentin and 3-isobutyl-gamma-aminobutyric acid on substance P-induced thermal hyperalgesia. Anesthesiol 88: 196-205, 1998.

117
Patel S, Naeem S, Kesingland A, Froestl W, Capogna M, Urban L, Fox A: The effects of GABAB agonists and gabapentin on mechanical hyperalgesia in models of neuropathic and inflammatory pain in the rat. Pain 90: 217-226, 2001. Plassat JL, Boschert U, Amlaiky N, Hen R: The mouse 5HT5 receptor reveals a remarkable heterogeneity within the 5HT1D receptor family. EMBO J 11: 4779-4786, 1992. Pepper I: Apuntes de inflamación. En Mecanismos de enfermedad y reacción del organismo. Universidad de Chile: 1-19, 2005. Perkins MN, Campbell E, Dray A: Antinociceptive activity of the bradykinin B1 and B2 receptor antagonists, des-Arg9, [Leu8]-BK and HOE 140, in two models of persistent hyperalgesia in the rat. Pain 53: 191-197, 1993. Perl ER : Somatic sensation: transfer and processing of information. 1.Peripheral receptors. Electroencephalogr Clin Neurophysiol 27: 650-651, 1969. Pertoldi S, Di Benedetto P: Shoulder-hand syndrome after stroke. A complex regional pain syndrome. Eur Medicophys 41: 283-292, 2005. Pezet S, McMahon SB: Neurotrophins: mediators and modulators of pain. Annu Rev Neurosci 29: 507-538, 2006. Picazo A, Castañeda-Hernández G, Ortíz MI: Examination of the interaction between peripheral diclofenac and gabapentin on the 5% formalin test in rats. Life Sci 79: 2283-2287, 2006. Pierre SC, Schmidt R, Brenneis C, Michaelis M, Geisslinger G, Scholich K: Inhibition of cyclooxygenases by dipyrone. Br J Pharmacol 151: 494-503, 2007. Ponimaskin EG, Profirovic J, Vaiskunaite R, Richter DW, Voyno-Yasenetskaya TA: 5-Hydroxytryptamine 4(a) receptor is coupled to the Galpha subunit of heterotrimeric G13 protein. J Biol Chem 277: 20812-9, 2002. Pozos-Guillén AJ, Aguirre-Bañuelos P, Arellano-Guerrero A, Castañeda-Hernández G, Hoyo-Vadillo C, Pérez-Urizar J: Isobolographic analysis of the dual-site synergism in the antinociceptive response of tramadol in the formalin test in rats. Life Sci 79: 2275-2282, 2006. Qin N, Zhang SP, Reitz TL, Mei JM, Flores CM: Cloning, expression, and functional characterization of human cyclooxygenase-1 splicing variants: evidence for intron 1 retention. J Pharmacol Exp Ther 315: 1298-305, 2005. Raffa RB, Friderichs E, Reimann W, Shank RP, Codd EE, Vaught JL, Jacoby HI, Selve N: Complementary and synergistic antinociceptive interaction between the enantiomers of tramadol. J Pharmacol Exp Ther 267: 331-340, 1993.

118
Raffa RB, Stone DJ Jr, Tallarida RJ: Discovery of "self-synergistic" spinal/supraspinal antinociception produced by acetaminophen (paracetamol). J Pharmacol Exp Ther 295: 291-294, 2000. Raffa RB: Pharmacology of oral combination analgesics: Rational therapy for pain. J Clin Pharm and Ther 26: 257-264, 2001.
Rang HP, Urban L: New molecules in analgesia. Br J Anaesth 75: 145-156, 1995.
Ready LB: The acute pain service. Acta Anaesthesiol Belg 43: 21-27, 1992. Reis GM, Doretto MC, Duarte ID, Tatsuo MA: Do endogenous opioids and nitric oxide participate in the anticonvulsant action of dipyrone?. Braz J Med Biol Res 36: 1263-1268, 2003. Reyes-García G, Caram-Salas NL, Medina-Santillán R, Granados-Soto V: Oral administration of B vitamins increases the antiallodynic effect of gabapentin in the rat. Proc West Pharmacol Soc 47: 76-79, 2004. Rocha-González HI, Meneses A, Carlton SM, Granados-Soto V: Pronociceptive role of peripheral and spinal 5-HT7 receptors in the formalin test. Pain 117: 182-192, 2005. Rode F, Broløs T, Blackburn-Munro G, Bjerrum OJ: Venlafaxine compromises the antinociceptive actions of gabapentin in rat models of neuropathic and persistent pain. Psychopharmacology (Berl) 187: 364-375, 2006. Rosenkranz B, Lehr KH, Mackert G, Seyberth HW: Metamizole-furosemide interaction study in healthy volunteers. Eur J Clin Pharmacol 42: 593-598, 1992. Saegusa H, Kurihara T, Zong S, Kazuno A, Matsuda Y, Nonaka T, Han W, Toriyama H, Tanabe T: Suppression of inflammatory and neuropathic pain symptoms in mice lacking the N-type Ca2+ channel. EMBO J 20: 2349-2356, 2001. Sah DW, Ossipov MH, Porreca F: Neurotrophic factors as novel therapeutics for neuropathic pain. Nat Rev Drug Discov 2: 460-472, 2003. Salvemini D, Doyle TM, Cuzzocrea S: Superoxide, peroxynitrite and oxidative/nitrative stress in inflammation. Biochem Soc Trans 34: 965-970, 2006.
Sasaki M, Obata H, Saito S, Goto F: Antinociception with intrathecal alpha-methyl-5-hydroxytryptamine, a 5-hydroxytryptamine 2A/2C receptor agonist, in two rat models of sustained pain. Anesth Analg 96: 1072-1078, 2003.
Sator-Katzenschlager SM, Scharbert G, Kress HG, Frickey N, Ellend A, Gleiss A, Kozek-Langenecker SA: Chronic pelvic pain treated with gabapentin and

119
amitriptyline: a randomized controlled pilot study. Wien Klin Wochenschr 117: 761-768, 2005. Schaible HG: Peripheral and central mechanisms of pain generation. Handb Exp Pharmacol 177: 3-28, 2007. Schug SA, Zech D, Dorr U. Cancer pain management according to WHO analgesics guidelines. J Pain Simpt Manage 5: 27-32, 1990. Shaiova L, Rim F, Friedman D, Jahdi M: A review of methylnaltrexone, a peripheral opioid receptor antagonist, and its role in opioid-induced constipation. Palliat Support Care 5: 161-166, 2007.
Shimada SG, Otterness IG, Stitt JT: study of the mechanism of action of the mild analgesic dipyrone. Agents Actions 41: 188-192, 1994.
Shimada S, Yamamura H, Ueda T, Yamamoto T, Ugawa S: Functional analysis of acid sensing ion channels. Nihon Shinkei Seishin Yakurigaku Zasshi 24: 243-246, 2004.
Shimoyama M, Shimoyama N, Inturrisi CE, Elliott KJ: Gabapentin enhances the antinociceptive effects of spinal morphine in the rat tail-flick test. Pain 72: 375-382, 1997. Shimoyama M, Shimoyama N, Hori Y: Gabapentin affects glutamatergic excitatory neurotransmission in the rat dorsal horn. Pain 85: 405-414, 2000. Simone DA, Alreja M, LaMotte RH: Psychophysical studies of the itch sensation and itchy skin ("alloknesis") produced by intracutaneous injection of histamine. Somatosens Mot Res 8: 271-279, 1991. Singh L, Field MJ, Ferris P, Hunter JC, Oles RJ, Williams RG, Woodruff GN: The antiepileptic agent gabapentin (Neurontin) possesses anxiolytic-like and antinociceptive actions that are reversed by D-serine. Psychopharmacology (Berl) 127: 1-9, 1996. Solaro C, Messmer Uccelli M, Uccelli A, Leandri M, Mancardi GL: Low-dose gabapentin combined with either lamotrigine or carbamazepine can be useful therapies for trigeminal neuralgia in multiple sclerosis. Eur Neurol 44: 45-48, 2000. Souslova V, Cesare P, Ding Y, Akopian AN, Stanfa L, Suzuki R, Carpenter K, Dickenson A, Boyce S, Hill R, Nebenuis-Oosthuizen D, Smith AJ, Kidd EJ, Wood JN: Warm-coding deficits and aberrant inflammatory pain in mice lacking P2X3 receptors. Nature 407: 1015-1017, 2000. Staud R, Smitherman ML: Peripheral and central sensitization in fibromyalgia: pathogenetic role. Curr Pain Headache Rep 6: 259-266, 2002.

120
Stamer UM, Höthker F, Lehnen K, Stüber F: Postoperative analgesia with tramadol and metamizol. Continual infusion versus patient controlled analgesia. Anaesthesist 52: 33-41, 2003. Steen KH, Reeh PW, Anton F, Handwerker HO: Protons selectively induce lasting excitation and sensitization to mechanical stimulation of nociceptors in rat skin, in vitro. J Neurosci 12: 86-95, 1992. Stefani A, Spadoni F, Giacomini P, Lavaroni F, Bernardi G: The effects of gabapentin on different ligand- and voltage-gated currents in isolated cortical neurons. Epilepsy Res 43: 239-248, 2001. Steranka LR, Manning DC, DeHaas CJ, Ferkany JW, Borosky SA, Connor JR, Vavrek RJ, Stewart JM, Snyder SH: Bradykinin as a pain mediator: receptors are localized to sensory neurons, and antagonists have analgesic actions. Proc Natl Acad Sci USA 85: 3245-3249, 1988. Sutton KG, Martin DJ, Pinnock RD, Lee K, Scott RH: Gabapentin inhibits high-threshold calcium channel currents in cultured rat dorsal root ganglion neurones. Br J Pharmacol 135: 257-265, 2002. Takeda K, Sawamura S, Tamai H, Sekiyama H, Hanaoka K: Role for cyclooxygenase 2 in the development and maintenance of neuropathic pain and spinal glial activation. Anesthesiology 103: 837-844, 2005. Tallarida RJ, Raffa RB. Testing for synergism over a range of fixed ratio drug combinations: Replacing the isobologram. Life Sci 58: 23-28, 1996. Tallarida RJ, Stone DJ Jr, Raffa RB: Efficient designs for studying synergistic drug combinations. Life Sci 61: PL 417-425, 1997. Tallarida RJ: Drug synergism and Dose-Effect Data Analysis. CRC press Boca Raton FL: 1-72, 2000. Tallarida RJ: Drug synergism: its detection and applications. J Pharmacol Exp Ther 298: 865-872, 2001. Tallarida RJ, Cowan A, Raffa RB: Antinociceptive synergy, additivity, and subadditivity with combinations of oral glucosamine plus nonopioid analgesics in mice. J Pharmacol Exp Ther 307: 699-704, 2003. Tatum WO 4th, Galvez R, Benbadis S, Carrazana E: New antiepileptic drugs: Into the new millennium. Arch Fam Med 9: 1135-1141, 2000. Taylor CP: Mechanisms of action of gabapentin. Rev Neurol (Paris): 153: S39-45, 1997.

121
Taylor CP, Gee NS, Su TZ, Kocsis JD, Welty DF, Brown JP, Dooley DJ, Boden P, Singh L: A summary of mechanistic hypotheses of gabapentin pharmacology. Epilepsy Res 29: 233-249, 1998. Taylor J, Mellström B, Fernaud I, Naranjo JR: Metamizol potentiates morphine effects on visceral pain and evoked c-Fos immunoreactivity in spinal cord. Eur J Pharmacol 351: 39-47, 1998. Tortorici V, Vanegas H: Putative role of medullary off-and on-cells in the antinociception produced by dipyrone (metamizol) administered systemically or microinjected into PAG. Pain 57: 197-205, 1994. Tortoricci V, Vanegas H: Opioid tolerance induced by metamizol (dipyrone) microinjections into the periaqueductal grey of rats. Eur J Neurosci 12: 4074-4040, 2000. Treede RD, Meyer RA, Raja SN, Campbell JN: Peripheral and central mechanisms of cutaneous hyperalgesia. Prog Neurobiol 38: 397-421, 1992. Treede RD: Peripheral acute pain mechanisms. Ann Med 27: 213-216, 1995. Turan A, White P, Karamanlioglu B, Memis D, Tasdogan M, Pamukçu Z, Yavuz E: An lternative to the cyclooxygenase-2 inhibitors for perioperative pain management. Anesth Analg 102: 175-181, 2006. Valenzuela F, García-Saisó S, Lemini C, Ramírez-Solares R, Vidrio H, Mendoza-Fernández V: Metamizol acts as an ATP sensitive potassium channel opener to inhibit the contracting response induced by angiotensin II but not to norepinephrine in rat thoracic aorta smooth muscle. Vascul Pharmacol 43: 120-127, 2005. Vázquez E, Hernández N, Escobar W, Vanegas H: Antinociception induced by intravenous dipyrone (metamizol) upon dorsal horn neurons: involvement of endogenous opioids at the periaqueductal gray matter, the nucleus raphe magnus, and the spinal cord in rats. Brain Res 1048: 211-217, 2005. Vázquez E, Escobar W, Ramírez K, Vanegas H: A nonopioid analgesic acts upon the PAG-RVM axis to reverse inflammatory hyperalgesia. Eur J Neurosci 25: 471-479, 2007. Vera-Portocarrero LP, Zhang ET, Ossipov MH, Xie JY, King T, Lai J, Porreca F: Descending facilitation from the rostral ventromedial medulla maintains nerve injury-induced central sensitization. Neuroscience 140: 1311-1320, 2006. Verge VM, Xu Z, Xu XJ, Wiesenfeld-Hallin Z, Hökfelt T: Marked increase in nitric oxide synthase mRNA in rat dorsal root ganglia after peripheral axotomy: in situ hybridization and functional studies. Proc Natl Acad Sci USA 89: 11617-11621, 1992.

122
Vlahov V, Badian M, Verho M, Bacracheva N: Vlahov 1990 Pharmacokinetics of metamizol metabolites in healthy subjects after a single oral dose of metamizol sodium. Eur J Clin Pharmacol 38: 61-65, 1990. Vlaskovska M, Surcheva S, Ovcharov R: Importance of endogenous opioids and prostaglandins in the action of analgin (metamizole) and verapamil. Farmakol Toksikol 52: 25-29, 1989. Volz M, Kellner HM: Kinetics and metabolism of pyrazolones (propyphenazone, aminopyrine and dipyrone). Br J Clin Pharmacol 10: 299S-308S, 1980. Walker KM, Urban L, Medhurst SJ, Patel S, Panesar M, Fox AJ, McIntyre P: The VR1 antagonist capsazepine reverses mechanical hyperalgesia in models of inflammatory and neuropathic pain. J Pharmacol Exp Ther 304: 56-62, 2003. Wall PD: Introduction to the fouth edition. In: Wall PD, Melzack R: Textbook of pain. Churchill Livingstone (Londres): 1-8, 1999. Wang PH, Cenedeze MA, Pesquero JB, Pacheco-Silva A, Câmara NO: Influence of bradykinin B1 and B2 receptors in the immune response triggered by renal ischemia-reperfusion injury. Int Immunopharmacol 6:1960-5, 2006. Watanabe T, Yanai K: Studies on functional roles of the histaminergic neuron system by using pharmacological agents, knockout mice and positron emission tomography. Tohoku J Exp Med 195: 197-217, 2001. Wei EP, Moskowitz MA, Boccalini P, Kontos HA: Calcitonin gene-related peptide mediates nitroglycerin and sodium nitroprusside-induced vasodilation in feline cerebral arterioles. Circ Res 70: 1313-1319, 1992. Wheeler-Aceto H, Cowan A: Standardization of the rat paw formalin test for the evaluation of analgesics. Psychopharmacology (Berl) 104: 35-44, 1991. White WT, Patel N, Drass M, Nalamachu S: Lidocaine patch 5% with systemic analgesics such as gabapentin: a rational polypharmacy approach for the treatment of chronic pain. Pain Med 4: 321-30, 2003. Wild KD, Bian D, Zhu D, Davis J, Bannon AW, Zhang TJ, Louis JC: Antibodies to nerve growth factor reverse established tactile allodynia in rodent models of neuropathic pain without tolerance. J Pharmacol Exp Ther 322: 282-287, 2007. WHO: World Health Organization. Cancer pain relief and palliative care: Report of a WHO expert committee. Geneva: WHO 1990 (technical report series num 804). Winkelstein BA: Mechanisms of central sensitization, neuroimmunology & injury biomechanics in persistent pain: implications for musculoskeletal disorders. J Electromyogr Kinesiol 14: 87-93, 2004.

123
Woolf CJ: Evidence for a central component of post-injury pain hypersensitivity. Nature 306: 686-688, 1983. Woolf CJ, Thompson SW: The induction and maintenance of central sensitization is dependent on N-methyl-D-aspartic acid receptor activation; implications for the treatment of post-injury pain hypersensitivity states. Pain 44: 293-299, 1991. Woolf CJ, Shortland P, Reynolds M, Ridings J, Doubell T, Coggeshall RE: Reorganization of central terminals of myelinated primary afferents in the rat dorsal horn following peripheral axotomy. J Comp Neurol 360: 121-134, 1995. Woolf CJ: Phenotypic modification of primary sensory neurons: the role of nerve growth factor in the production of persistent pain. Philos Trans R Soc Lond B Biol Sci 351: 441-448, 1996. Woolf CJ, Salter MW: Neuronal plasticity: increasing the gain in pain. Science 288: 1765-1769, 2000. Woolf CJ: Pain: Moving from symptom control toward mechanism-specific pharmacologic management. Ann Intern Med 140: 441-451, 2004. Yaksh TL, Rudy TA: Analgesia mediated by a direct spinal action of narcotics. Science 192: 1357-1358, 1976. Yaksh TL: Spinal opiates: a review of their effect on spinal function with emphasis on pain processing. Acta Anaesthesiol Scand Suppl 85: 25-37, 1987. Yoon MH, Yaksh TL: The effect of intrathecal gabapentin on pain behavior and hemodynamics on the formalin test in the rat. Anesth Analg. 1999 Aug; 89 (2): 434-9The effect of intrathecal gabapentin on pain behavior and hemodynamics on the formalin test in the rat. Anesth Analg 89: 434-439, 1999a. Yoon MH, Yaksh TL: Evaluation of interaction between gabapentin and ibuprofen on the formalin test in rats. Anesthesiology 91: 1006-1013, 1999b. Yoon MH, Choi JI, Jeong SW: Spinal gabapentin and antinociception: mechanisms of action. J Korean Med Sci 18: 255-261, 2003. Yoon MH, Choi JI, Kwak SH: Characteristic of interactions between intrathecal gabapentin and either clonidine or neostigmine in the formalin test. Anesth Analg 98: 1374-1379, 2004. Yoon MH, Bae HB, Choi JI: Antinociceptive interactions between intrathecal gabapentin and MK801 or NBQX in rat formalin test. J Korean Med Sci 20: 307-312, 2005.

124
Yoshimura M, Nishi S: Blind patch-clamp recordings from substantia gelatinosa neurons in adult rat spinal cord slices: pharmacological properties of synaptic currents. Neuroscience 53: 519-526, 1993. Zeilhofer HU: Prostanoids in nociception and pain. Biochem Pharmacol 73: 165-174, 2007. Zhou Q, Le Grevés P, Ragnar F, Nyberg F: Intracerebroventricular injection of the N-terminal substance P fragment SP (1-7) regulates the expression of the N-methyl-D-aspartate receptor NR1, NR2A and NR2B subunit mRNAs in the rat brain. Neurosci Lett 291: 109-112, 2000. Zhuo M: Neuronal mechanism for neuropathic pain. Mol Pain 3: 14, 2007. Zimmermann M: Ethical guidelines for investigations of experimental pain in conscious animals. Pain 16: 109-110, 1983. Zimmermann M: Pain mechanisms and mediators in osteoarthritis. Semin Arthritis Rheum 18: 22-29, 1989. Zimmermann M: Pathobiology of neuropathic pain. Eur J Pharmacol 429: 23-37, 2001.

XIII
ANEXOS
Ponencias presentadas
LF Ortega-Varela, J Herrera, H Rocha-González, R Medina-
Santillán, G Reyes-García, V Granados-Soto. Interacción sinérgica de la
combinación gabapentina-metamizol en la prueba de formalina en ratas.
Memorias del XXVII Congreso Nacional de Farmacología. Villahermosa,
Tabasco. AMEFAR-UJAT, 2004.
LF Ortega-Varela, HI Rocha-Gonzalez, R Medina-Santillán, G
Reyes-García, V Granados-Soto. Synergistic interaction between
gabapentin and metamizol in the rat formalin test. 47th annual meeting of
the Western Pharmacology Society. Waikiki, Hawai, EUA ,2004.
LF Ortega-Varela, T Fraga-Bravo, JE Herrera-Abarca, H Urquiza-
Marín, V Granados-Soto. The use of a local combination of gabapentin and
metamizol shows synergistic effect in the rat formalin test. Society for
Neuroscience 35th annual Meeting. Washington, DC, EUA 2005.
Ortega-Varela LF, Caram N, Herrera-Abarca J, Granados V.
Preclinical asessment of the intrarhecal combination of gabapentin and
metamizol to relief neuropathic pain. 49th annual meeting of the Western
Pharmacology Society and XXIX Congreso Nacioanal de Farmacología.
Puerto Vallarta, Jalisco 2006.
Ortega-Varela LF, Herrera-Abarca JE, Mixcoatl-Zecualt T,
Granados-Soto V. The intrathecal combination of gabapentin and metamizol
reveals a multifactorial mechanism of action. Banff 2007 pharmacology and
therapeutics conference. Banff, Canadá.
Ortega-Varela LF y Granados-Soto V. El metamizol espinal
revierte la alodinia inducida en el modelo de Chung. XXX Congreso
Nacional de Farmacología. Culiacán, Sinaloa 2007.

XIV
PUBLICACIONES
Ortega-Varela LF, Herrera JE, Medina-Santillán R, Reyes-García
G, Rocha-González HI, Granados-Soto V: Synergistic interaction
between gabapentin and metamizol in the rat formalin test. Proc West
Pharmacol Soc 4: 80-83, 2004.
Ortega-Varela LF, Herrera J, Caram-Salas NL, Rocha-González
HI, Granados-Soto V: Isobolographic Analyses of the Gabapentin-
Metamizol combination after local peripheral, intrathecal and oral
administration in the rat. Pharmacology 79: 214-222, 2007.
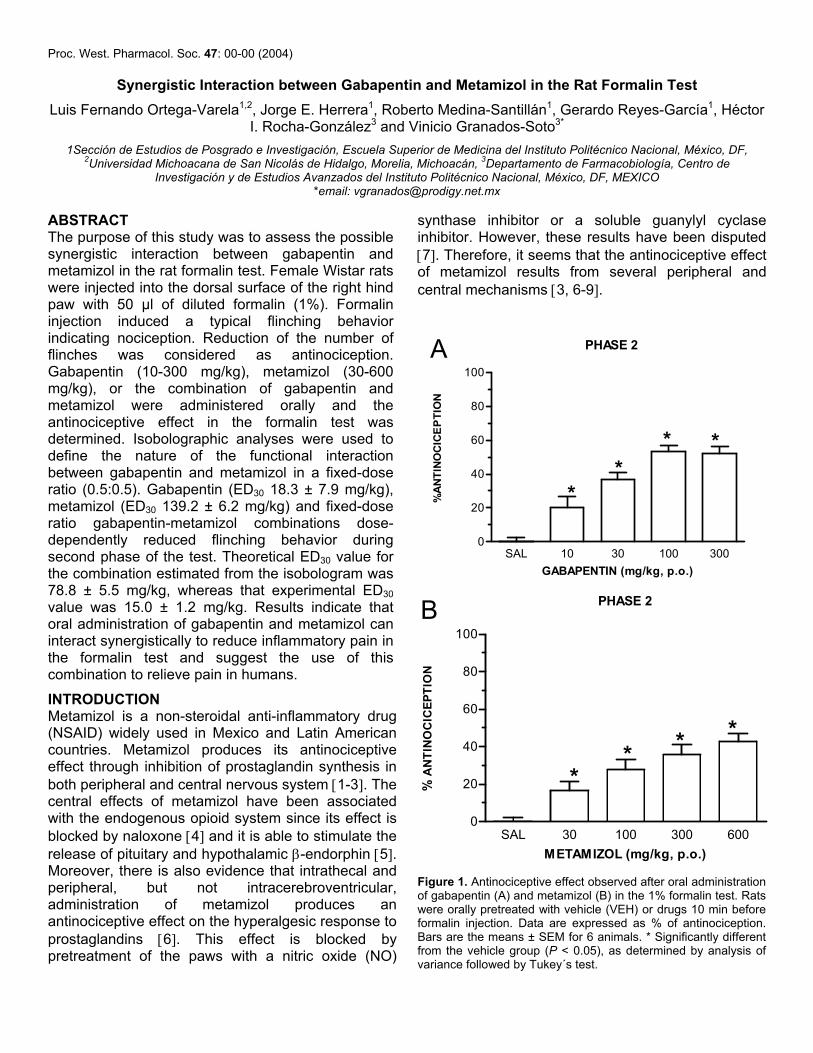
Proc. West. Pharmacol. Soc. 47: 00-00 (2004)
Synergistic Interaction between Gabapentin and Metamizol in the Rat Formalin Test Luis Fernando Ortega-Varela1,2, Jorge E. Herrera1, Roberto Medina-Santillán1, Gerardo Reyes-García1, Héctor
I. Rocha-González3 and Vinicio Granados-Soto3* 1Sección de Estudios de Posgrado e Investigación, Escuela Superior de Medicina del Instituto Politécnico Nacional, México, DF,
2Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, 3Departamento de Farmacobiología, Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, México, DF, MEXICO
*email: [email protected]
ABSTRACT The purpose of this study was to assess the possible synergistic interaction between gabapentin and metamizol in the rat formalin test. Female Wistar rats were injected into the dorsal surface of the right hind paw with 50 µl of diluted formalin (1%). Formalin injection induced a typical flinching behavior indicating nociception. Reduction of the number of flinches was considered as antinociception. Gabapentin (10-300 mg/kg), metamizol (30-600 mg/kg), or the combination of gabapentin and metamizol were administered orally and the antinociceptive effect in the formalin test was determined. Isobolographic analyses were used to define the nature of the functional interaction between gabapentin and metamizol in a fixed-dose ratio (0.5:0.5). Gabapentin (ED30 18.3 ± 7.9 mg/kg), metamizol (ED30 139.2 ± 6.2 mg/kg) and fixed-dose ratio gabapentin-metamizol combinations dose-dependently reduced flinching behavior during second phase of the test. Theoretical ED30 value for the combination estimated from the isobologram was 78.8 ± 5.5 mg/kg, whereas that experimental ED30 value was 15.0 ± 1.2 mg/kg. Results indicate that oral administration of gabapentin and metamizol can interact synergistically to reduce inflammatory pain in the formalin test and suggest the use of this combination to relieve pain in humans.
INTRODUCTION Metamizol is a non-steroidal anti-inflammatory drug (NSAID) widely used in Mexico and Latin American countries. Metamizol produces its antinociceptive effect through inhibition of prostaglandin synthesis in both peripheral and central nervous system [1-3]. The central effects of metamizol have been associated with the endogenous opioid system since its effect is blocked by naloxone [4] and it is able to stimulate the release of pituitary and hypothalamic β-endorphin [5]. Moreover, there is also evidence that intrathecal and peripheral, but not intracerebroventricular, administration of metamizol produces an antinociceptive effect on the hyperalgesic response to prostaglandins [6]. This effect is blocked by pretreatment of the paws with a nitric oxide (NO)
synthase inhibitor or a soluble guanylyl cyclase inhibitor. However, these results have been disputed [7]. Therefore, it seems that the antinociceptive effect of metamizol results from several peripheral and central mechanisms [3, 6-9]. Figure 1. Antinociceptive effect observed after oral administration of gabapentin (A) and metamizol (B) in the 1% formalin test. Rats were orally pretreated with vehicle (VEH) or drugs 10 min before formalin injection. Data are expressed as % of antinociception. Bars are the means ± SEM for 6 animals. * Significantly different from the vehicle group (P < 0.05), as determined by analysis of variance followed by Tukey´s test.
PHASE 2
SAL 30 100 300 6000
20
40
60
80
100
** * *
B
METAMIZOL (mg/kg, p.o.)
%A
NTI
NO
CIC
EPTI
ON
SAL 10 30 100 3000
20
40
60
80
100
PHASE 2
**
* *
GABAPENTIN (mg/kg, p.o.)
%A
NTI
NO
CIC
EP
TIO
N
A

Gabapentin, a structural analog of γ-aminobutyric acid (GABA), is a novel anticonvulsant used effectively for the treatment of refractory partial seizures with and without secondarily generalized tonic-clonic seizures. Several studies indicate that, besides its antiepileptic activity, gabapentin is an effective antihyperalgesic agent. Systemic or intrathecal administration of gabapentin attenuates nociceptive behaviors in animals and pain in humans that arise following peripheral nerve injury [10-11]. However, there is a considerable incidence of side effects at analgesic doses in humans [11-12]. Several studies have been done in rats as an attempt to reduce incidence of side effects for this drug in relieving pain [13-14]. The current study was designed to assess the oral efficacy of gabapentin and metamizol to reduce formalin-induced pain. In addition, the possible synergistic interaction between these drugs was also assessed in rats. METHODS: Female Wistar rats of 7-8 weeks (weight range, 180-200 g) from our own breeding facilities were used in this study. Rats had free access to drinking water, but food was restricted 12 h before the experiment. All experiments followed the Guidelines on Ethical Standards for Investigation of Experimental Pain in Animals [15]. Additionally, the study was approved by the Institutional Animal Care and Use Committee (Centro de Investigación y Estudios Avanzados del Instituto Politécnico Nacional, México, DF, MEXICO). Evaluation of antinociceptive activity: Antinociception was assessed by the formalin test. Rats were placed in an open Plexiglas® observation chamber for 30 min to allow them to accommodate to their surroundings, then they were removed for formalin administration. The rats were injected into the dorsal surface of the right hind paw with 50 µl of dilute formalin (1%), using a 30-gauge needle. The animal was then returned to the chamber for observation. A mirror was placed behind the chamber to enable unhindered observation of the formalin-injected paw. The rats were observed for nociceptive behavior immediately after formalin injection. Nociceptive behavior was quantified as the number of flinches of the injected paw during 1 min-periods every 5 min up to 60 min after injection. Flinching was readily discriminated and was characterized as rapid and brief withdrawal or flexing of the injected paw. As reported, formalin-induced flinching behavior was biphasic [16]. The initial acute phase (0-10 min) is followed by a relatively short quiescent period, which is then followed by a prolonged tonic response (15-60 min). At the end of the experiment, rats were killed in a CO2 chamber. Drugs: Metamizol was purchased from Sigma (St. Louis, MO, USA). Gabapentin was kindly provided by Merck S.A. de C.V. (Mexico City). All drugs were dissolved in saline. Study design: Rats received the oral administration of saline (4 ml/kg) or increasing doses of gabapentin (10, 30, 100 and 300 mg/kg), metamizol (30, 100, 300 and 600 mg/kg) or dose-fixed ratio gabapentin-metamizol combinations (0.5:0.5), 10 min before formalin injection. After formalin injection, flinching behavior was assessed for the next 60 min. Data analysis and statistics: All results are presented as means ± SEM for at least 6 animals per group. The area under the
number of flinches against time curves (AUC) was calculated by the trapezoidal rule [17]. Dose-response curves for each compound tested were established based on the percent maximum possible effect (expressed as % Antinociception) calculated from AUC of the phase 2 of each individual rat: % Antinociception = ((vehicle-post-compound)/vehicle) x 100. For evaluation of the interaction between gabapentin and metamizol, isobolograms were constructed using ED30 values obtained when the drugs were administered alone or combined. We used ED30 instead of ED50 values because neither drug was able to reach more than 50% of antinociception in our model. The construction of the dose-response curves and the determination of doses producing 30% maximum possible effect (ED30) were computed. To perform the isobolographic analysis, gabapentin and metamizol were administered in combination as fixed ratios of equieffective ED30 dose for each drug (gabapentin/metamizol =0.5:0.5). The ED30 values (± SEM) for gabapentin and metamizol alone were plotted on the x and y axes, respectively, and the theoretical additive point was calculated according to Tallarida et al. [17]. From the dose-response curve of the combined drugs, the ED30 value of the total dose of the combination was calculated. Statistical significance between the theoretical additive point and the experimentally derived ED30 value was evaluated using Student´s t test. An experimental ED30 significantly less than the theoretical additive ED30 (P < 0.05) was considered to indicate a synergistic interaction between gabapentin and metamizol.
RESULTS: Formalin administration produced a typical pattern of flinching behavior. The first phase started immediately after administration of formalin and then diminished gradually in about 10 min. The second phase started at 15 min and lasted until 1 hr. Oral administration of metamizol (30-600 mg/kg) dose-dependently reduced flinching behavior induced by formalin during phase 2 (Figure 1) reaching a maximum effect of 43% (ED30 139.2 ± 6.2 mg/kg). In addition, oral administration of gabapentin (10-300 mg/kg) significantly reduced formalin-induced nociceptive behavior in the rat, reaching a maximum effect of about 52% (ED30 18.3 ± 7.9 mg/kg).
Oral co-administration of gabapentin and metamizol induced a dose-dependent increase in the % antinociception (Figure 2). The antinociceptive effect of the combination was observed mainly on the second phase of the test. Theoretical ED30 value for the combination gabapentin and metamizol estimated from the isobolograms was 78.8 ± 5.5 mg/kg. This value was significantly higher than experimental ED30 value which was 15.0 ± 1.2 mg/kg. As shown in figure 3, the experimentally derived ED30 (± SEM) in the isobologram is below the theoretically additive dose line, indicating a significant synergistic interaction between gabapentin and metamizol in the formalin test (P < 0.05).
DISCUSSION: Oral administration of metamizol produced a dose-related antinociception in the
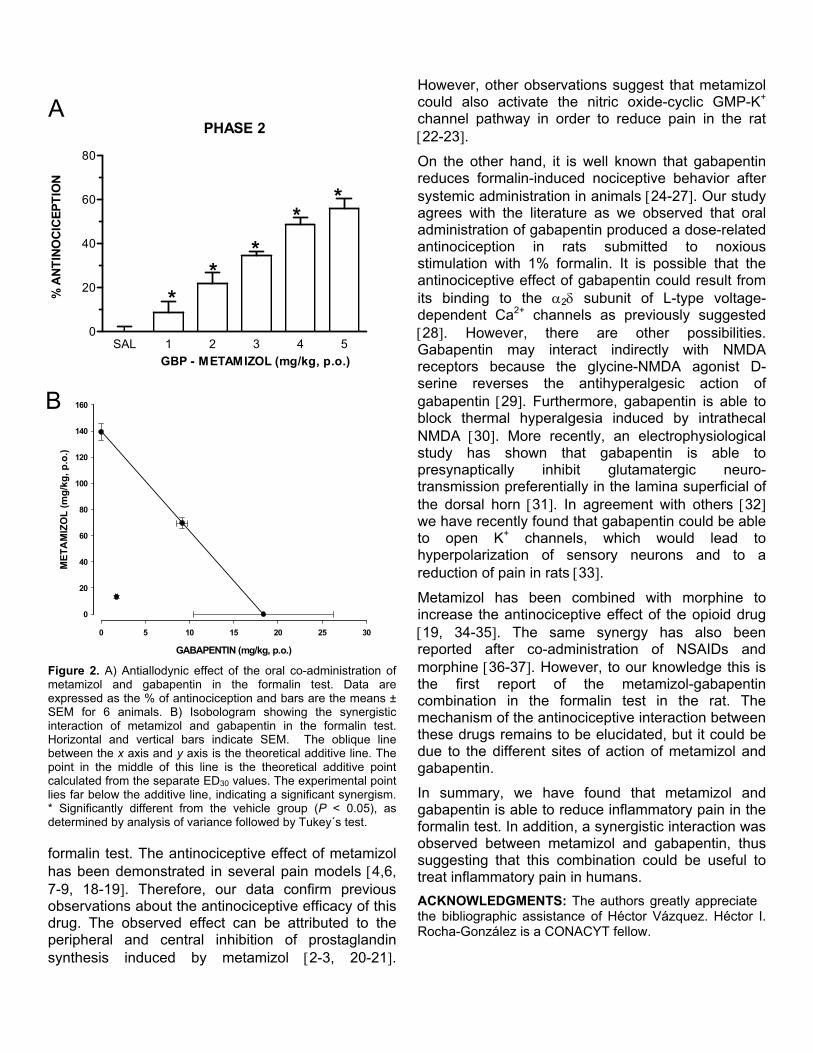
Figure 2. A) Antiallodynic effect of the oral co-administration of metamizol and gabapentin in the formalin test. Data are expressed as the % of antinociception and bars are the means ± SEM for 6 animals. B) Isobologram showing the synergistic interaction of metamizol and gabapentin in the formalin test. Horizontal and vertical bars indicate SEM. The oblique line between the x axis and y axis is the theoretical additive line. The point in the middle of this line is the theoretical additive point calculated from the separate ED30 values. The experimental point lies far below the additive line, indicating a significant synergism. * Significantly different from the vehicle group (P < 0.05), as determined by analysis of variance followed by Tukey´s test.
formalin test. The antinociceptive effect of metamizol has been demonstrated in several pain models [4,6, 7-9, 18-19]. Therefore, our data confirm previous observations about the antinociceptive efficacy of this drug. The observed effect can be attributed to the peripheral and central inhibition of prostaglandin synthesis induced by metamizol [2-3, 20-21].
However, other observations suggest that metamizol could also activate the nitric oxide-cyclic GMP-K+ channel pathway in order to reduce pain in the rat [22-23].
On the other hand, it is well known that gabapentin reduces formalin-induced nociceptive behavior after systemic administration in animals [24-27]. Our study agrees with the literature as we observed that oral administration of gabapentin produced a dose-related antinociception in rats submitted to noxious stimulation with 1% formalin. It is possible that the antinociceptive effect of gabapentin could result from its binding to the α2δ subunit of L-type voltage-dependent Ca2+ channels as previously suggested [28]. However, there are other possibilities. Gabapentin may interact indirectly with NMDA receptors because the glycine-NMDA agonist D-serine reverses the antihyperalgesic action of gabapentin [29]. Furthermore, gabapentin is able to block thermal hyperalgesia induced by intrathecal NMDA [30]. More recently, an electrophysiological study has shown that gabapentin is able to presynaptically inhibit glutamatergic neuro-transmission preferentially in the lamina superficial of the dorsal horn [31]. In agreement with others [32] we have recently found that gabapentin could be able to open K+ channels, which would lead to hyperpolarization of sensory neurons and to a reduction of pain in rats [33].
Metamizol has been combined with morphine to increase the antinociceptive effect of the opioid drug [19, 34-35]. The same synergy has also been reported after co-administration of NSAIDs and morphine [36-37]. However, to our knowledge this is the first report of the metamizol-gabapentin combination in the formalin test in the rat. The mechanism of the antinociceptive interaction between these drugs remains to be elucidated, but it could be due to the different sites of action of metamizol and gabapentin.
In summary, we have found that metamizol and gabapentin is able to reduce inflammatory pain in the formalin test. In addition, a synergistic interaction was observed between metamizol and gabapentin, thus suggesting that this combination could be useful to treat inflammatory pain in humans. ACKNOWLEDGMENTS: The authors greatly appreciate the bibliographic assistance of Héctor Vázquez. Héctor I. Rocha-González is a CONACYT fellow.
PHASE 2
SAL 1 2 3 4 50
20
40
60
80
**
**
*
GBP - METAMIZOL (mg/kg, p.o.)
% A
NTI
NOC
ICEP
TIO
N
A
GABAPENTIN (mg/kg, p.o.)
0 5 10 15 20 25 30
MET
AM
IZO
L (m
g/kg
, p.o
.)
0
20
40
60
80
100
120
140
160B

REFERENCES 1. Abbate, R., Gori, A.M., Pinto, S., Attanasio, M.,
Paniccia, R., Coppo, M., Castellani, S., Giusti, B., Boddi, M., and Neri, S.G.G.: Prostaglan. Leuk. Essen. Fatty Acids 41: 89-93 (1990).
2. Shimada, S.G., Otterness, I.G., and Stitt, J.T.: Agents Actions 41: 188-192 (1994).
3. Campos, C., de Gregorio, R., García-Nieto, R., Gago, F., Ortiz, P., and Alemany, S.: Eur. J. Pharmacol. 378: 339-347 (1999).
4. Tortorici, V., Vasquez, E., and Vanegas, H.: Brain Res. 725: 106-110 (1996).
5. Vlaskovska, M., Surcheva, S., and Ovcharov, R.: Farmakol. Toksikol. 52: 25-29 (1989).
6. Lorenzetti, B.B. and Ferreira, S.H.: Inflamm. Res. 45: 308-311 (1996).
7. Beirith, A., Santos, A.R.S., Rodrigues, A.L.S., Creczynski-Pasa, T.B., and Calixto, J.B.: Eur. J. Pharmacol. 345: 233-245 (1998).
8. Granados-Soto, V., Flores-Murrieta, F.J., Castaneda-Hernandez, G., and López-Muñoz, F.J.: Eur. J. Pharmacol. 277: 281-284 (1995).
9. Laird, J.M.A., Roza, C., and Olivar, T.: Inflamm. Res. 47: 389-395 (1998).
10. Mellick, G.A. and Mellick, L.B.: J. Pain Symp. Manage. 10: 265-266 (1995).
11. Rowbotham, M., Harden, N., Stacey, B., Bernstein, P., and Magnus-Miller, L.: JAMA 280: 1837-1842 (1998).
12. Rice, A.S.C. and Maton, S.: Pain 94: 215-224 (2001). 13. Chen, S.R., Eisenach, J.C., McCaslin, P.P., and Pan,
H.L.: Anesthesiology 92: 500-506 (2000). 14. Field, M.J., Gonzalez, M.I., Tallarida, R.J., and Singh,
L.: J. Pharmacol. Exp. Ther. 303: 730-735 (2002). 15. Zimmermann, M.: Pain 16: 109-110 (1991). 16. Wheeler-Aceto, H., Cowan, A.: Psychopharmacol. 104:
35-44 (1991). 17. Tallarida, R.J.: Drug Synergism and Dose-Effect Data
Analysis. New York, Chapman and Hall/CRC, pp. 1-72 (2000).
18. Carlsson, K.H., Monzel, W., and Jurna, I.: Pain 32: 313-326 (1988).
19. Aguirre-Bañuelos, P. and Granados-Soto, V.: J. Pharmacol. Toxicol. Methods 42: 79-85 (1999).
20. Vane, J.R.: Nature 231: 232-235 (1971). 21. Ferreira, S.H., Lorenzetti, B.B., Correa, F.M.A.: Eur. J.
Pharmacol. 53: 39-48 (1978).
22. Alves, D. and Duarte, I.D.G.: Eur. J. Pharmacol. 444 : 47-52 (2002).
23. Ortiz, M.I., Castañeda-Hernández, and G., Granados-Soto, V.: Pharmacol. Biochem. Behav. 74: 465-70 (2003).
24. Shimoyama, M., Shimoyama, N., Davies, A.M., Inturrisi, C.E., and Elliot, K.J.: Neurosci. Lett. 222: 65-67 (1997).
25. Yoon, M.Y. and Yaksh, T.L.: Anesthesiology 91: 1006-1013 (1999).
26. Kaneko, M., Mestre, C., Sánchez, E.H., and Hammaond, D.L.: J. Pharmacol. Exp. Ther. 292: 743-751 (2000).
27. Hwang, J.H. and Yaksh, T.L.: Regional Anesth. 22: 249-256 (1997).
28. Gee, N.S., Brown, J.P., Dissanayke, V.U.K., Offord, J., Thurlow, R., and Woodruff, G.N.: J. Biol. Chem. 271: 5768-5776 (1996).
29. Singh, L., Field, M.J., Ferris, P., Hunter, J.C., Oles, R.J., Williams, R.G., and Woodruff, G.N.: Psychopharmacol. 127: 1-9 (1996).
30. Partridge, B., Chaplan, S.R., Sakamoto, E., and Yaksh, T.L.: Anesthesiology 18: 196-205 (1998).
31. Shimoyama. M., Shimoyama. N., and Hori, Y.: Pain 85: 405-414 (2000).
32. Freiman, T.M., Kukolja, J., Heinemeyer, J., Eckhardt, K., Aranda, H., Rominger, A., Dooley, D.J., Zentner, J., and Feuertein, T.J.: Naunyn-Schmiedeberg´s Arch. Pharmacol. 363: 537-542 (2001).
33. Mixcoatl-Zecuatl, T., Medina-Santillán, R., Reyes-García, G., Vidal-Cantú, G.C., and Granados-Soto, V.: Eur. J. Pharmacol. In Press (2004).
34. Taylor, J., Mellstrom, B., Fernaud, I., and Naranjo, J.R.: Eur. J. Pharmacol. 351: 39-47 (1998).
35. Hernández-Delgadillo, G.P., Ventura Martínez, R., Díaz Reval, M.I., Domínguez Ramírez, A.M., and López-Muñoz, F.J.: Eur. J. Pharmacol. 441: 177-83 (2002).
36. Malmberg, A.B. and Yaksh, T.L.: Anesthesiology 79: 270-281 (1993).
37. Maves, T.J., Pechman, P.S., Meller, S.T., and Gebhart, G.F.: Anesthesiology 80:1094-1001 (1994).

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Pharmacology 2007;79:214–222 DOI: 10.1159/000101390
Isobolographic Analyses of the Gabapentin-Metamizol Combination after Local Peripheral, Intrathecal and Oral Administration in the Rat
Luis F. Ortega-Varela a, b Jorge E. Herrera a Nadia L. Caram-Salas c
Héctor I. Rocha-González c Vinicio Granados-Soto c, d
a Sección de Estudios de Posgrado e Investigación, Escuela Superior de Medicina del Instituto Politécnico Nacional,
México, D.F. ; b Escuela Preparatoria Melchor Ocampo, Universidad Michoacana de San Nicolás de Hidalgo,
Morelia, Michoacán ; c Departamento de Farmacobiología, Centro de Investigación y Estudios Avanzados, Sede Sur,
México, D.F. , and d Centro de Investigación y de Estudios Avanzados, Unidad Monterrey, Monterrey, N.L. , Mexico
ergism. Data confirm that low doses of the gabapentin and metamizol can interact synergistically to reduce formalin-induced nociceptive behavior suggesting that this combina-tion could be useful to treat inflammatory pain in humans.
Copyright © 2007 S. Karger AG, Basel
Introduction
Currently, combination therapy is commonly used in an attempt to improve pain management. The potential advantage of using combination therapy is that analgesic effects can be maximized while the incidence of adverse side effects could be minimized. Moreover, the multiplic-ity of mechanisms involved in pain [1] suggests that com-bination therapy can improve pain management [2] .
In addition to the inhibitory action on the prostaglan-din synthesis [3, 4] , it has been reported that certain non-steroidal anti-inflammatory drugs (NSAIDs) are able to produce part of their antinociceptive effect through the activation of the nitric oxide-cyclic GMP pathway [5–8] . A prototype drug for these effects is metamizol (dipy-rone). There is evidence to support that metamizol pro-duces its antinociceptive effect through inhibition of prostaglandin synthesis in both the peripheral and cen-
Key Words Antinociception � Synergy � Isobolograms � Gabapentin � Metamizol � Formalin test
Abstract This study was designed to evaluate the possible antinoci-ceptive interaction between gabapentin and metamizol on formalin-induced nociception. Gabapentin, metamizol or a fixed dose-ratio combination of both drugs were assessed after local peripheral, intrathecal and oral administration in rats. Isobolographic analyses were employed to define the nature of the interaction between drugs. Gabapentin, met-amizol and gabapentin-metamizol combinations yielded a dose-dependent antinociceptive effect when administered by the three different routes. ED 30 values were estimated for the individual drugs and isobolograms were constructed. Theoretical ED 30 values for the combination estimated from the isobolograms were 21.11 8 1.17 � g/paw, 104.6 8 5.5 � g/rat and 78.8 8 5.5 mg/kg for the local peripheral, intra-thecal and oral administration routes, respectively. These values were significantly higher than the experimentally ob-tained ED 30 values which were 11.3 8 1.5 � g/paw, 36.8 8 3.1 � g/rat and 15 8 1.2 mg/kg indicating a synergistic inter-action. Systemic administration resulted in the highest syn-
Received: August 24, 2006 Accepted: December 22, 2006 Published online: March 29, 2007
Vinicio Granados-Soto, PhD Departamento de Farmacobiología, Cinvestav, Sede Sur Calzada de los Tenorios 235, Col. Granjas Coapa 14330 México, D.F. (Mexico) Tel. +52 55 5061 2868, Fax +52 55 5061 2863, E-Mail [email protected]
© 2007 S. Karger AG, Basel 0031–7012/07/0794–0214$23.50/0
Accessible online at: www.karger.com/pha

Isobolographic Analyses of Gabapentin-Metamizol Combination
Pharmacology 2007;79:214–222 215
tral nervous system [9–11] . Moreover, the central effects of metamizol have been associated with the endogenous opioid system since its effect is blocked by naloxone [12] and it is able to stimulate the release of pituitary and hy-pothalamic � -endorphin [13] . In contrast, there is also evidence that intrathecal and local peripheral, but not in-tracerebroventricular, administration of metamizol re-duces prostaglandin-induced hyperalgesia [14] . This ef-fect can be blocked by pretreatment of the paws with a nitric oxide synthase inhibitor or a soluble guanylyl cy-clase inhibitor suggesting the activation of the nitric ox-ide-cyclic GMP pathway. More recently it has been sug-gested that activation of the nitric oxide-cyclic GMP pathway by metamizol leads to the opening of K + chan-nels [15, 16] which in turns leads to hyperpolarization and antinociception. Therefore, it seems that the antino-ciceptive effect of metamizol results from several periph-eral and central mechanisms [8, 14–17] and it could be a rational target for combination.
On the other hand, gabapentin is a novel anticonvul-sant used effectively for the treatment of neuropathic pain in humans [18, 19] . However, studies in rats have also shown that local peripheral, intrathecal or systemic administration of gabapentin attenuates nociceptive be-haviors that arise following tissue injury [20–25] . Not-withstanding its efficacy, there is a considerable incidence of side effects at doses that produce relief of pain in hu-mans [18, 19] . Therefore, the purpose of this study was to assess the possible synergistic interaction between gaba-pentin and metamizol in the rat formalin test at local pe-ripheral, intrathecal and oral administration by isobolo-graphic analyses.
Material and Methods
Animals Experiments were performed on adult female Wistar rats
(body weight range 180–220 g) of 6–7 weeks of age. Animals were obtained from our own breeding facilities and had free access to food and drinking water before experiments. All experiments fol-lowed Guidelines on Ethical Standards for Investigation of Ex-perimental Pain in Animals [26] . Additionally, this study was ap-proved by the Institutional Animal Care and Use Committee (Centro de Investigación y de Estudios Avanzados, México, D.F., Mexico).
Measurement of Antinociceptive Activity Antinociception was assessed using the formalin test [27] . The
rats were placed in open Plexiglas observation chambers for 30 min to allow them to acclimate to their surroundings; then they were removed for formalin administration. Rats were gently re-strained while the dorsum of the hind paw was injected with 50
� l of diluted formalin (1%) into the dorsal surface of the right hind paw with a 30-gauge needle. The animals were returned to the chambers and nociceptive behavior was observed immedi-ately after formalin injection. Mirrors were placed in each cham-ber to enable unhindered observation. Nociceptive behavior was quantified as the number of flinches of the injected paw during 1-min periods every 5 min, up to 60 min after injection [27] . Flinching was readily discriminated and was characterized as rapid and brief withdrawal, or as flexing of the injected paw. We decided to evaluate flinching because it is a simple and reliable parameter of pain behavior and one producing high scores [27] . Formalin-induced flinching behavior was biphasic [27] . The ini-tial acute phase (0–10 min) was followed by a relatively short qui-escent period, which was then followed by a prolonged tonic re-sponse (15–60 min). At the end of the experiment the rats were sacrificed in a CO 2 chamber.
Spinal Surgery For spinal administration, chronic catheterization of the spi-
nal subarachnoid space was performed as described by Yaksh and Rudy [28] . The rats were anesthetized with a ketamine/xylazine mixture (45–12 mg/kg, i.p.), placed in a stereotaxic head-holder, and the atlanto-occipital membrane exposed. The latter mem-brane was pierced, and a polyethylene catheter (PE-10, 7.0 cm length) was inserted intrathecally and advanced caudally to the level of the thoracolumbar junction. The wound was sutured and animals were allowed to recover from surgery for at least 5 days before testing. Rats showing any signs of motor impairment were euthanized in a CO 2 chamber.
Drugs Gabapentin was kindly provided by Merck SA de C.V. (Méxi-
co, D.F.). Metamizol (dipyrone) was purchased from Sigma (St. Louis, Mo., USA). Both drugs were dissolved in saline.
Study Design For the local peripheral study, rats received a subcutaneous
injection (50 � l) in the dorsal surface of the right hind paw of ve-hicle or increasing doses of either gabapentin (10–300 � g/paw), metamizol (10–300 � g/paw), or the gabapentin-metamizol com-bination (as indicated in tables 1 , 2 ) 10 min before formalin injec-tion in the same paw (ipsilateral). To assess if the antinociceptive effect of drugs was due to a local action, formalin was adminis-tered in one paw and the greatest dose of the tested drugs (300 � g/paw) in the contralateral paw. For the spinal study, rats re-ceived an intrathecal injection (10 � l) of saline or increasing dos-es of either gabapentin (25–200 � g/rat), metamizol (50–400 � g/rat), or the gabapentin-metamizol combination (as indicated in tables 1 , 2 ) 10 min before formalin injection. In the systemic study, animals received gabapentin (10–300 mg/kg), metamizol (30–600 mg/kg), or the metamizol-gabapentin combinations ( tables 1, 2 ) orally 10 min before formalin injection.
For all routes of administration, doses and time of administra-tion were selected on the basis of a pilot study as well as on previ-ous studies in our model. Rats in all groups were observed regard-ing behavioral or motor function changes induced by the treat-ments. This was assessed, but not quantified, by testing the animals’ ability to stand and walk in a normal posture, as pro-posed elsewhere [29] .
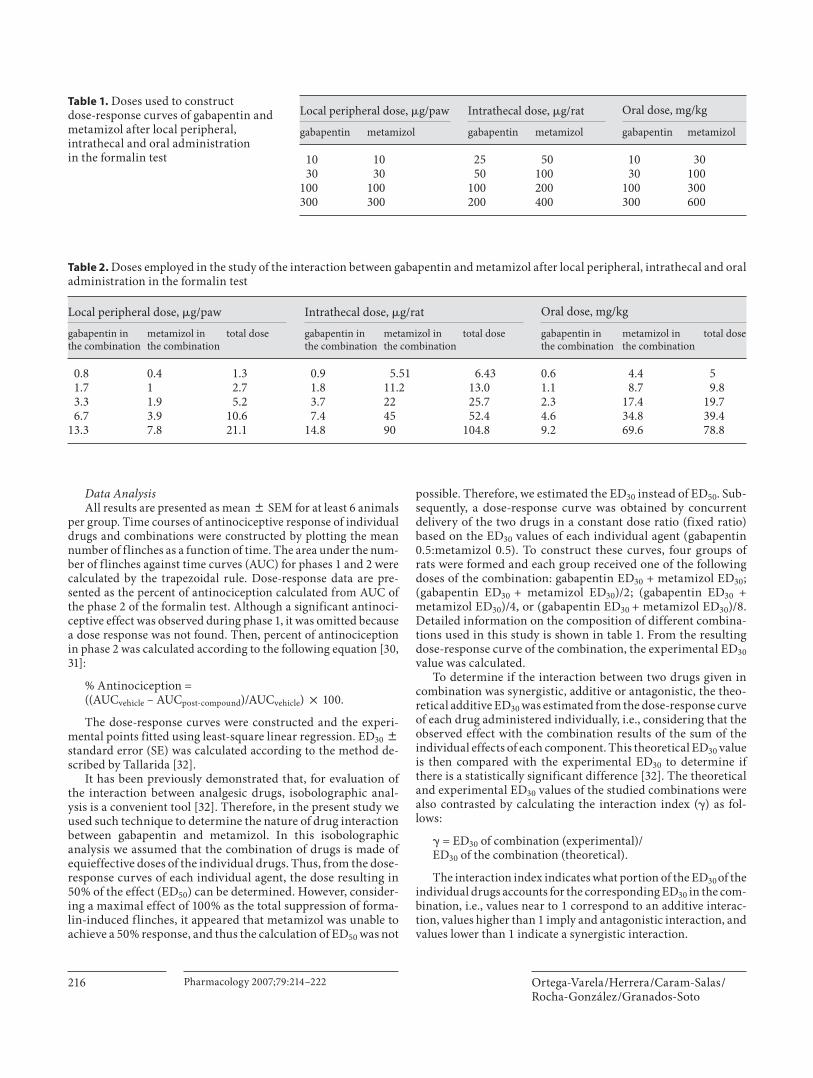
Ortega-Varela /Herrera /Caram-Salas /Rocha-González /Granados-Soto
Pharmacology 2007;79:214–222 216
Data Analysis All results are presented as mean 8 SEM for at least 6 animals
per group. Time courses of antinociceptive response of individual drugs and combinations were constructed by plotting the mean number of flinches as a function of time. The area under the num-ber of flinches against time curves (AUC) for phases 1 and 2 were calculated by the trapezoidal rule. Dose-response data are pre-sented as the percent of antinociception calculated from AUC of the phase 2 of the formalin test. Although a significant antinoci-ceptive effect was observed during phase 1, it was omitted because a dose response was not found. Then, percent of antinociception in phase 2 was calculated according to the following equation [30, 31] :
% Antinociception = ((AUC vehicle – AUC post-compound )/AUC vehicle ) ! 100.
The dose-response curves were constructed and the experi-mental points fitted using least-square linear regression. ED 30 8 standard error (SE) was calculated according to the method de-scribed by Tallarida [32] .
It has been previously demonstrated that, for evaluation of the interaction between analgesic drugs, isobolographic anal - ysis is a convenient tool [32] . Therefore, in the present study we used such technique to determine the nature of drug interaction between gabapentin and metamizol. In this isobolographic analysis we assumed that the combination of drugs is made of equieffective doses of the individual drugs. Thus, from the dose-response curves of each individual agent, the dose resulting in 50% of the effect (ED 50 ) can be determined. However, consider-ing a maximal effect of 100% as the total suppression of forma-lin-induced flinches, it appeared that metamizol was unable to achieve a 50% response, and thus the calculation of ED 50 was not
possible. Therefore, we estimated the ED 30 instead of ED 50 . Sub-sequently, a dose-response curve was obtained by concurrent delivery of the two drugs in a constant dose ratio (fixed ratio) based on the ED 30 values of each individual agent (gabapentin 0.5:metamizol 0.5). To construct these curves, four groups of rats were formed and each group received one of the following doses of the combination: gabapentin ED 30 + metamizol ED 30 ; (gabapentin ED 30 + metamizol ED 30 )/2; (gabapentin ED 30 + metamizol ED 30 )/4, or (gabapentin ED 30 + metamizol ED 30 )/8. Detailed information on the composition of different combina-tions used in this study is shown in table 1 . From the resulting dose-response curve of the combination, the experimental ED 30 value was calculated.
To determine if the interaction between two drugs given in combination was synergistic, additive or antagonistic, the theo-retical additive ED 30 was estimated from the dose-response curve of each drug administered individually, i.e., considering that the observed effect with the combination results of the sum of the individual effects of each component. This theoretical ED 30 value is then compared with the experimental ED 30 to determine if there is a statistically significant difference [32] . The theoretical and experimental ED 30 values of the studied combinations were also contrasted by calculating the interaction index ( � ) as fol-lows:
� = ED 30 of combination (experimental)/ ED 30 of the combination (theoretical).
The interaction index indicates what portion of the ED 30 of the individual drugs accounts for the corresponding ED 30 in the com-bination, i.e., values near to 1 correspond to an additive interac-tion, values higher than 1 imply and antagonistic interaction, and values lower than 1 indicate a synergistic interaction.
Local peripheral dose, �g/paw Intrathecal dose, �g/rat Oral dose, mg/kg
gabapentin metamizol gabapentin metamizol gabapentin metamizol
10 10 25 50 10 3030 30 50 100 30 100
100 100 100 200 100 300300 300 200 400 300 600
Table 1. Doses used to constructdose-response curves of gabapentin and metamizol after local peripheral, intrathecal and oral administrationin the formalin test
Table 2. Doses employed in the study of the interaction between gabapentin and metamizol after local peripheral, intrathecal and oral administration in the formalin test
Local peripheral dose, �g/paw Intrathecal dose, �g/rat Oral dose, mg/kg
gabapentin inthe combination
metamizol inthe combination
total dose gabapentin inthe combination
metamizol inthe combination
total dose gabapentin inthe combination
metamizol inthe combination
total dose
0.8 0.4 1.3 0.9 5.51 6.43 0.6 4.4 51.7 1 2.7 1.8 11.2 13.0 1.1 8.7 9.83.3 1.9 5.2 3.7 22 25.7 2.3 17.4 19.76.7 3.9 10.6 7.4 45 52.4 4.6 34.8 39.4
13.3 7.8 21.1 14.8 90 104.8 9.2 69.6 78.8
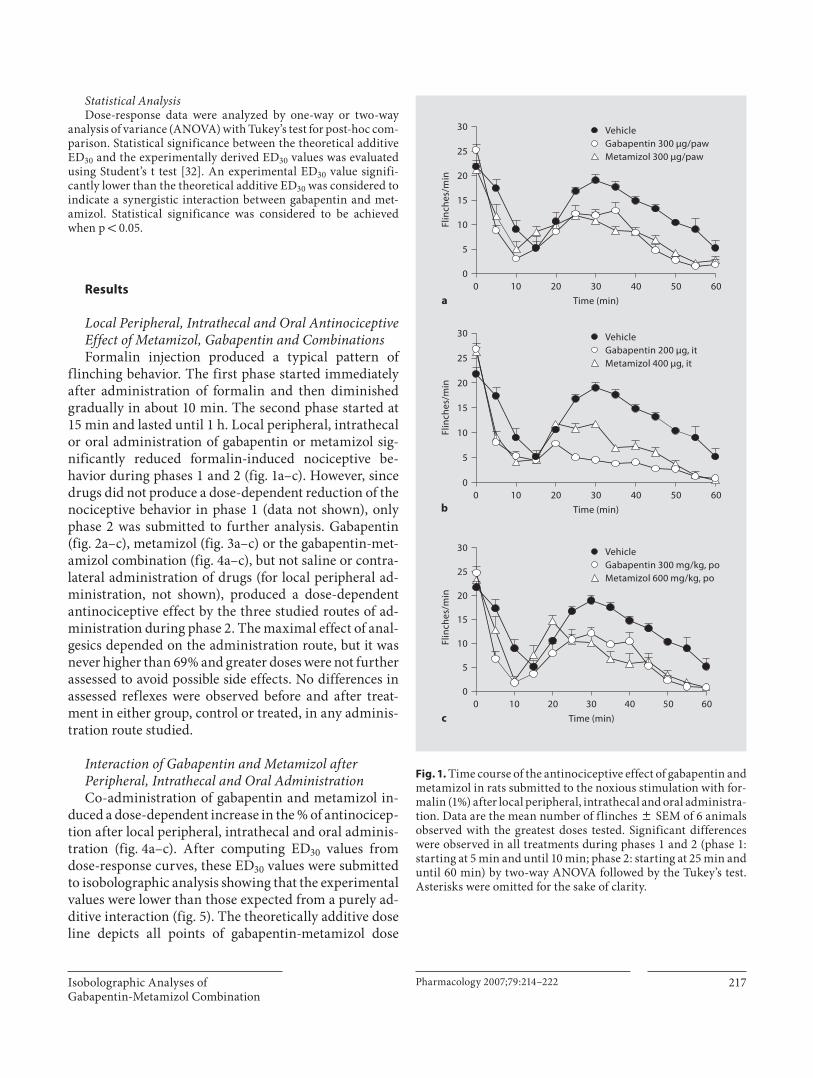
Isobolographic Analyses of Gabapentin-Metamizol Combination
Pharmacology 2007;79:214–222 217
Statistical Analysis Dose-response data were analyzed by one-way or two-way
analysis of variance (ANOVA) with Tukey’s test for post-hoc com-parison. Statistical significance between the theoretical additive ED 30 and the experimentally derived ED 30 values was evaluated using Student’s t test [32] . An experimental ED 30 value signifi-cantly lower than the theoretical additive ED 30 was considered to indicate a synergistic interaction between gabapentin and met-amizol. Statistical significance was considered to be achieved when p ! 0.05.
Results
Local Peripheral, Intrathecal and Oral Antinociceptive Effect of Metamizol, Gabapentin and Combinations Formalin injection produced a typical pattern of
flinching behavior. The first phase started immediately after administration of formalin and then diminished gradually in about 10 min. The second phase started at 15 min and lasted until 1 h. Local peripheral, intrathecal or oral administration of gabapentin or metamizol sig-nificantly reduced formalin-induced nociceptive be-havior during phases 1 and 2 ( fig. 1 a–c). However, since drugs did not produce a dose-dependent reduction of the nociceptive behavior in phase 1 (data not shown), only phase 2 was submitted to further analysis. Gabapentin ( fig. 2 a–c), metamizol ( fig. 3 a–c) or the gabapentin-met-amizol combination ( fig. 4 a–c), but not saline or contra-lateral administration of drugs (for local peripheral ad-ministration, not shown), produced a dose-dependent antinociceptive effect by the three studied routes of ad-ministration during phase 2. The maximal effect of anal-gesics depended on the administration route, but it was never higher than 69% and greater doses were not further assessed to avoid possible side effects. No differences in assessed reflexes were observed before and after treat-ment in either group, control or treated, in any adminis-tration route studied.
Interaction of Gabapentin and Metamizol after Peripheral, Intrathecal and Oral Administration Co-administration of gabapentin and metamizol in-
duced a dose-dependent increase in the % of antinocicep-tion after local peripheral, intrathecal and oral adminis-tration ( fig. 4 a–c). After computing ED 30 values from dose-response curves, these ED 30 values were submitted to isobolographic analysis showing that the experimental values were lower than those expected from a purely ad-ditive interaction ( fig. 5 ). The theoretically additive dose line depicts all points of gabapentin-metamizol dose
30
Fli
nc
he
s/m
in
Vehicle
Gabapentin 300 µg/paw
Metamizol 300 µg/paw25
20
15
10
5
0
0 10 20 30 40 50 60
Time (min)
Vehicle
Gabapentin 200 µg, it
Metamizol 400 µg, it
Vehicle
Gabapentin 300 mg/kg, po
Metamizol 600 mg/kg, po
a
30
30
Fli
nc
he
s/m
inF
lin
ch
es/
min
25
25
20
20
15
15
10
10
5
5
0
0
0
0
10
10
20
20
30
30
40
40
50
50
60
60
Time (min)
Time (min)
b
c
Fig. 1. Time course of the antinociceptive effect of gabapentin and metamizol in rats submitted to the noxious stimulation with for-malin (1%) after local peripheral, intrathecal and oral administra-tion. Data are the mean number of flinches 8 SEM of 6 animals observed with the greatest doses tested. Significant differences were observed in all treatments during phases 1 and 2 (phase 1: starting at 5 min and until 10 min; phase 2: starting at 25 min and until 60 min) by two-way ANOVA followed by the Tukey’s test. Asterisks were omitted for the sake of clarity.
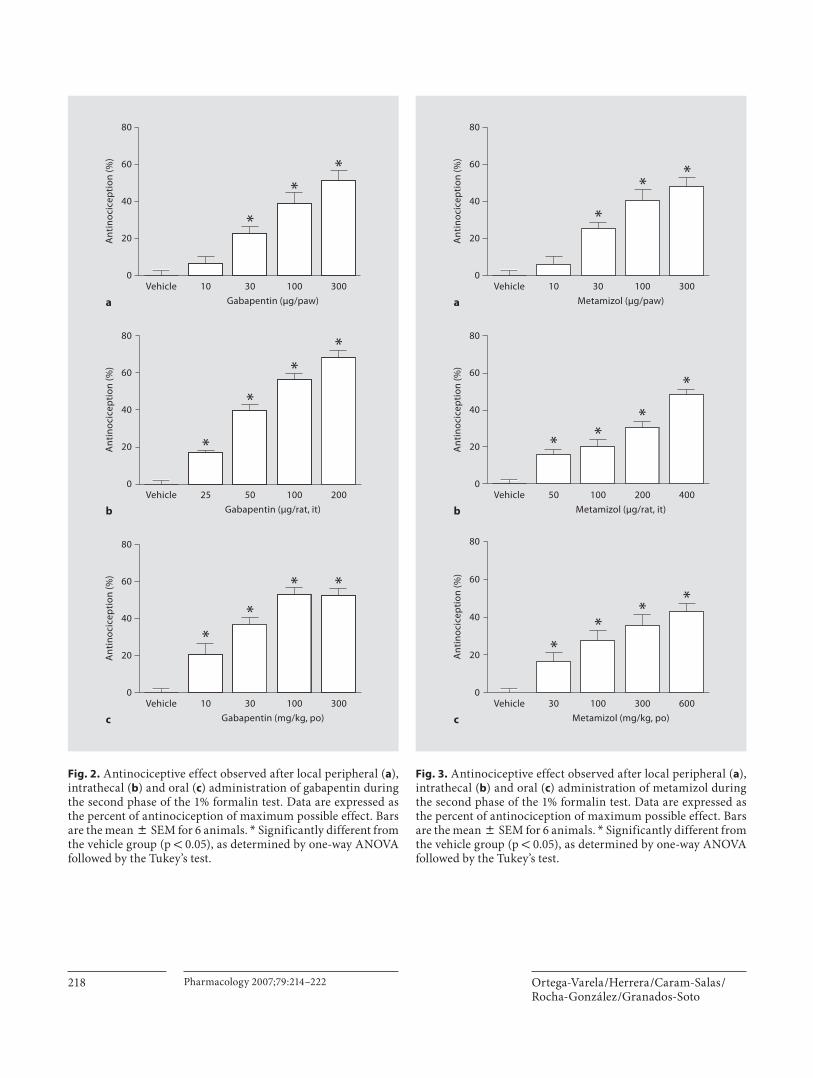
Ortega-Varela /Herrera /Caram-Salas /Rocha-González /Granados-Soto
Pharmacology 2007;79:214–222 218
80
An
tin
oc
ice
pti
on
(%)
60
40
20
0
a
Vehicle 10 30 100 300
Gabapentin (µg/paw)
*
*
*
*
*
*
*
80
80
An
tin
oc
ice
pti
on
(%)
An
tin
oc
ice
pti
on
(%)
60
60
40
40
20
20
0
0
b
c
Vehicle
Vehicle
25
10
50
30
100
100
200
300
Gabapentin (µg/rat, it)
Gabapentin (mg/kg, po)
*
*
* *
Fig. 2. Antinociceptive effect observed after local peripheral ( a ), intrathecal ( b ) and oral ( c ) administration of gabapentin during the second phase of the 1% formalin test. Data are expressed as the percent of antinociception of maximum possible effect. Bars are the mean 8 SEM for 6 animals. * Significantly different from the vehicle group (p ! 0.05), as determined by one-way ANOVA followed by the Tukey’s test.
80
An
tin
oc
ice
pti
on
(%)
60
40
20
0
a
Vehicle 10 30 100 300
Metamizol (µg/paw)
*
*
80
80
An
tin
oc
ice
pti
on
(%)
An
tin
oc
ice
pti
on
(%)
60
60
40
40
20
20
0
0
b
c
Vehicle
Vehicle
50
30
100
100
200
300
400
600
Metamizol (µg/rat, it)
Metamizol (mg/kg, po)
*
*
*
**
*
**
*
Fig. 3. Antinociceptive effect observed after local peripheral ( a ), intrathecal ( b ) and oral ( c ) administration of metamizol during the second phase of the 1% formalin test. Data are expressed as the percent of antinociception of maximum possible effect. Bars are the mean 8 SEM for 6 animals. * Significantly different from the vehicle group (p ! 0.05), as determined by one-way ANOVA followed by the Tukey’s test.
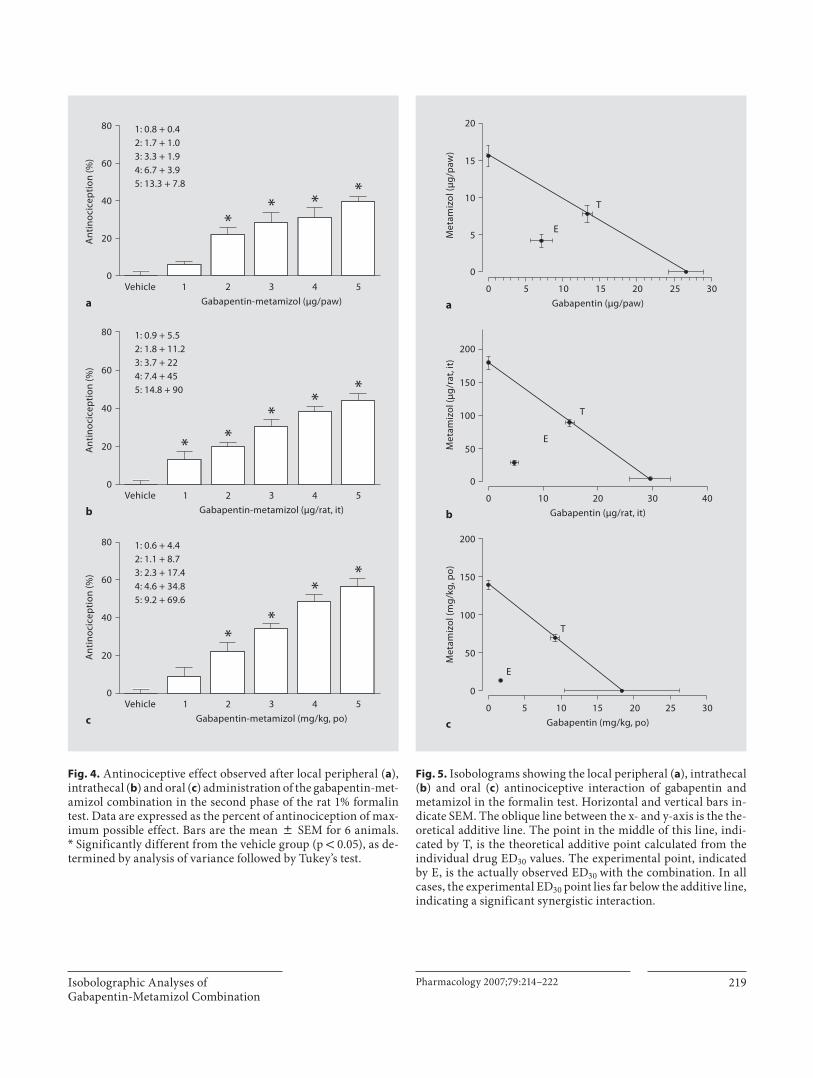
Isobolographic Analyses of Gabapentin-Metamizol Combination
Pharmacology 2007;79:214–222 219
80
An
tin
oc
ice
pti
on
(%)
60
40
20
0
a
Vehicle 1
Gabapentin-metamizol (µg/paw)
80
80
An
tin
oc
ice
pti
on
(%)
An
tin
oc
ice
pti
on
(%)
60
60
40
40
20
20
0
0
b
c
Vehicle
Vehicle
Gabapentin-metamizol (µg/rat, it)
Gabapentin-metamizol (mg/kg, po)
2 3 4 5
*
1: 0.8 + 0.4
2: 1.7 + 1.0
3: 3.3 + 1.9
4: 6.7 + 3.9
5: 13.3 + 7.8
***
**
*
*
1 2 3 4 5
*
1 2 3 4 5
*
**
*
1: 0.9 + 5.5
2: 1.8 + 11.2
3: 3.7 + 22
4: 7.4 + 45
5: 14.8 + 90
1: 0.6 + 4.4
2: 1.1 + 8.7
3: 2.3 + 17.4
4: 4.6 + 34.8
5: 9.2 + 69.6
Fig. 4. Antinociceptive effect observed after local peripheral ( a ), intrathecal ( b ) and oral ( c ) administration of the gabapentin-met-amizol combination in the second phase of the rat 1% formalin test. Data are expressed as the percent of antinociception of max-imum possible effect. Bars are the mean 8 SEM for 6 animals. * Significantly different from the vehicle group (p ! 0.05), as de-termined by analysis of variance followed by Tukey’s test.
E
T
200
Me
tam
izo
l(m
g/k
g,p
o)
150
100
50
0
c
10 15 25 30
Gabapentin (mg/kg, po)
E
T
50 20
T
E
200
20
Me
tam
izo
l()
µg
/ra
t,it
Me
tam
izo
l ()
µg
/pa
w
150
15
100
10
50
5
0
0
b
a
10
10
40
15 25
30
30
Gabapentin (µg/rat, it)
Gabapentin (µg/paw)
5
0
0
20
20
Fig. 5. Isobolograms showing the local peripheral ( a ), intrathecal ( b ) and oral ( c ) antinociceptive interaction of gabapentin and metamizol in the formalin test. Horizontal and vertical bars in-dicate SEM. The oblique line between the x- and y-axis is the the-oretical additive line. The point in the middle of this line, indi-cated by T, is the theoretical additive point calculated from the individual drug ED 30 values. The experimental point, indicated by E, is the actually observed ED 30 with the combination. In all cases, the experimental ED 30 point lies far below the additive line, indicating a significant synergistic interaction.

Ortega-Varela /Herrera /Caram-Salas /Rocha-González /Granados-Soto
Pharmacology 2007;79:214–222 220
combinations yielding an effect of 30% according to an additive interaction. For the three routes of administra-tion, the experimental ED 30 values of the gabapentin-metamizol combination clearly were situated below theo-retically additive dose line, indicating a synergistic inter-action between gabapentin and metamizol after local peripheral, intrathecal and oral administration ( fig. 5 a–c). Comparison of experimental and theoretical ED 30 val-ues by the Student’s t test yielded statistically significant differences (p ! 0.05) for all routes ( table 3 ). Analysis of the interaction index showed that the best synergism was obtained after oral administration ( � = 0.19).
Discussion
Antinociceptive Effect of Gabapentin It is well known that gabapentin is able to reduce for-
malin-induced nociceptive behavior after local peripheral [33] , spinal [23, 25] and systemic [24, 34, 35] administra-tion. Our study agrees with the literature as we observed that either local peripheral, intrathecal or oral adminis-tration of gabapentin produced a dose-related antinoci-ception in rats submitted to noxious stimulation with 1% formalin. It is possible that the antinociceptive effect of gabapentin could result from its binding to the � 2 � sub-unit of L-type voltage-dependent Ca 2+ channels as previ-ously reported [36] . However, this drug could activate other targets. Gabapentin may interact indirectly with NMDA receptors because the glycine-NMDA agonist D -serine reverses the antihyperalgesic action of gabapentin [20] . Furthermore, gabapentin is able to block thermal hy-peralgesia induced by intrathecal NMDA [22] . More re-cently, an electrophysiological study has shown that gaba-
pentin is able to presynaptically inhibit glutamatergic neurotransmission in the dorsal horn [37] . Accordingly with other authors, we have recently found that gabapen-tin could be able to activate the nitric oxide-cyclic GMP-protein kinase G-K + channel pathway at the periphery [38] , which would lead to hyperpolarization of sensory neurons and to a reduction of inflammatory pain in rats.
Antinociceptive Effect of Metamizol The present data demonstrates that local peripheral, in-
trathecal or oral administration of metamizol is able to produce a dose-dependent antinociceptive effect during both phases of the formalin test. These observations agree with previous studies showing that metamizol produces antinociception after local peripheral [14–16, 39] , intrathe-cal [14, 17, 40] and systemic [8, 40, 41] administration in rats and mice. The antinociceptive effects of this drug could be related with peripheral and central cyclooxygen-ase inhibition [9–11] . However, there are other possibilities. The antinociceptive effect of metamizol has been related with activation of the nitric oxide-cyclic GMP-K + channel pathway in both the periphery and spinal cord [14–16] , as well as with a central opioidergic pathway leading to stim-ulation of inhibitory descendent system [12, 41, 42] . More-over, metamizol seems to modulate the excitatory amino acid release and/or action at the spinal cord [17] .
Antinociceptive Effect of the Gabapentin-Metamizol Combination Combining analgesics is a strategy intended to achieve
some therapeutic goals such as facilitating compliance, simplifying prescription, and improving efficacy without increasing adverse effects or decreasing adverse effects without loss of efficacy [2] . The primary objective of this
Local peripheraladministrationED30, �g/paw
Intrathecal administrationED30, �g/rat
Oral administrationED30, mg/kg
Gabapentin 26.682.4 29.683.9 18.387.9Metamizol 15.681.4 179.6810.4 139.286.2Theoretical combination 21.181.1 104.685.5 78.885.5Experimental combination 11.381.5* 36.883.1* 1581.2*Interaction index 0.53 0.35 0.19
ED30: effective dose resulting in a 30% reduction on control response. Data are the mean 8 SE of the estimate.
* Significantly different from the theoretical combination data (р < 0.05), by the Stu-dent’s t test.
Table 3. Effect of local peripheral, intrathecal and oral administrationof gabapentin and metamizol alone or in combination in the formalin test

Isobolographic Analyses of Gabapentin-Metamizol Combination
Pharmacology 2007;79:214–222 221
References
1 Millan MJ: The induction of pain: an inte-grative review. Prog Neurobiol 1999; 57: 1–164.
2 Raffa RB: Pharmacology of oral analgesic combination analgesics rational therapy for pain. J Clin Pharm Ther 2001; 26: 257–264.
3 Vane JR: Inhibition of prostaglandin synthe-sis as a mechanism of action for aspirin-like drugs. Nature 1971; 231: 232–235.
4 Ferreira SH: Prostaglandins, aspirin-like drugs and analgesia. Nature 1972; 240: 200–203.
5 Lorenzetti BB, Ferreira SH: Mode of analge-sic action of dipyrone: direct antagonism of inflammatory hyperalgesia. Eur J Pharma-col 1985; 114: 375–381.
6 Duarte IDG, Dos Santos IR, Lorenzetti BB, Ferreira SH: Antinociception by direct an-tagonism of nociceptor sensitization in-volves the L -arginine: nitric oxide: cGMP pathway. Eur J Pharmacol 1992; 217: 225–227.
7 Tonussi CR, Ferreira SH: Mechanism of di-clofenac analgesia: direct blockade of in-flammatory sensitization. Eur J Pharmacol 1994: 251: 173–179.
8 Granados-Soto V, Flores-Murrieta FJ, Casta-ñeda-Hernández G, López-Muñoz FJ: Evi-dence for the involvement of nitric oxide in the antinociceptive effect of ketorolac in the rat. Eur J Pharmacol 1995; 277: 281–284.
9 Abbate R, Gori AM, Pinto S, Attanasio M, Paniccia R, Coppo M, Castellani S, Giusti B, Boddi M, Neri SGG: Cycloxygenase and li-poxygenase metabolite synthesis by poly-morphonuclear neutrophils: in vitro effect of dipyrone. Prostaglandins Leukot Essent Fat-ty Acids 1990; 41: 89–93.
10 Shimada SG, Otterness IG, Stitt JT: A study of the mechanism of action of the mild anal-gesic dipyrone. Agents Actions 1994; 41: 188–192.
11 Campos C, de Gregorio R, Garcia-Nieto R, Gago F, Ortiz P, Alemany S: Regulation of cyclooxygenase activity by metamizol. Eur J Pharmacol 1999; 378: 339–347.
12 Tortorici V, Vasquez E, Vanegas H: Naloxone partial reversal of the antinociception pro-duced by dipyrone microinjected into peri-aqueductal gray of rats. Possible involvement of medullary off- and on-cells. Brain Res 1996; 725: 106–110.
13 Vlaskovska M, Surcheva S, Ovcharov R: Im-portance of endogenous opioids and prosta-glandins in the action of analgin (met-amizole) and verapamil. Farmakol Toksikol 1989; 52: 25–29.
14 Lorenzetti BB, Ferreira SH: Activation of the arginine-nitric oxide pathway in primary sensory neurons contributes to dipyrone-in-duced spinal and peripheral analgesia. In-flamm Res 1996; 45: 308–311.
15 Alves DP, Duarte IDG: Involvement of ATP-sensitive K + channels in the peripheral anti-nociceptive effect induced by dipyrone. Eur J Pharmacol 2002; 444: 45–72.
16 Ortiz MI, Castañeda-Hernández G, Grana-dos-Soto V: Possible involvement of potas-sium channels in peripheral antinociception induced by metamizol: lack of participation of ATP-sensitive K + channels. Pharmacol Biochem Behav 2003; 74: 465–470.
17 Beirith A, Santos ARS, Rodrigues ALS, Cre-cynski-Pasa TB, Calixto JB: Spinal and su-praspinal antinociceptive action of dipyrone in formalin, capsaicin and glutamate tests. Study of the mechanism of action. Eur J Pharmacol 1998; 345: 233–245.
study was to understand the nature of the interaction be-tween gabapentin and metamizol at several levels. A sig-nificant synergy between gabapentin and metamizol was demonstrated at local peripheral, intrathecal and system-ic levels by the isobolographic analysis. The interaction of these drugs has not been previously examined; how-ever, gabapentin has been co-administered with ibupro-fen [24] , naproxen [43] , amitriptyline [34] , serotonin [44] and tramadol [35] yielding synergistic effects in inflam-matory pain models.
On the other hand, metamizol has shown synergism in combination with morphine [39, 45] , tramadol [46] and B vitamins [47] . Co-administration of gabapentin and metamizol produced a significant synergistic inter-action at the different levels assessed. The mechanism of this antinociceptive interaction remains to be elucidated. However, the mechanism of the synergy could be due to the different actions and sites of action of both drugs. Ac-tually the sites and mechanisms at which both drugs act appear to be complementary. Binding of gabapentin to � 2 � subunit combined with the inhibition of prostaglan-din synthesis by metamizol may be the basis of the syn-ergistic interaction of these two drugs. However, with the present experiments we cannot ruled out that other mechanisms could participate in this interaction.
Interaction index ( � ) provides a measure of the degree of synergism [32] . Therefore, the fact that the best syner-gism was seen after oral administration suggests a pos-sible advantage in their use compared to local peripheral or intrathecal administration. In this sense, Miranda and Pinardi [40] have reported that metamizol and clonidine produce an additive interaction after intrathecal admin-istration, whereas that a synergistic interaction is ob-served after intraperitoneal administration.
In summary, gabapentin and metamizol produced an-tinociception in the rat formalin test at different levels of pain transmission. In addition, a synergistic interaction was observed between gabapentin and metamizol at the local peripheral, spinal and systemic level, suggesting that this combination could be useful to treat inflamma-tory pain in humans.
Acknowledgements
The authors greatly appreciate the technical and bibliograph-ical assistance of Guadalupe C. Vidal-Cantú and Héctor Vazquez, respectively. Luis F. Ortega-Varela is a Conacyt Fellow. This work is part of the PhD dissertation of Luis F. Ortega-Varela.

Ortega-Varela /Herrera /Caram-Salas /Rocha-González /Granados-Soto
Pharmacology 2007;79:214–222 222
18 Rowbotham M, Harden N, Stacey B, Bern-stein P, Magnus-Miller L: Gabapentin for the treatment of postherpetic neuralgia. JAMA 1998; 280: 1837–1842.
19 Rice AS, Maton S, Postherpetic Neuralgia Study Group: Gabapentin in postherpetic neuralgia: a randomised, double-blind, pla-cebo-controlled study. Pain 2001; 94: 215–224.
20 Singh L, Field MJ, Ferris P, Hunter JC, Oles RJ, Williams RG, Woodruff GN: The anti-epileptic agent gabapentin (Neurontin) pos-sesses anxiolytic-like and antinociceptive actions that are reversed by D -serine. Psy-chopharmacology (Berl) 1996; 127: 1–9.
21 Jun JH, Yaksh TL: The effect of intrathecal gabapentin and 3-isobutyl � -aminobutyric acid on the hyperalgesia observed after ther-mal injury in the rat. Anesth Analg 1998; 86: 48–54.
22 Partridge BJ, Chaplan SR, Sakamoto E, Yaksh TL: Characterization of the effects of gabapentin and 3-isobutyl- � -aminobutyric acid on substance P-induced thermal hyper-algesia. Anesthesiology 1998; 88: 196–205.
23 Yoon MH, Yaksh TL: The effect of intrathe-cal gabapentin on pain behavior and hemo-dynamics on the formalin test in the rat. Anesth Analg 1999; 89: 434–439.
24 Yoon MH, Yaksh TL: Evaluation of interac-tion between gabapentin and ibuprofen on formalin test in rats. Anesthesiology 1999; 91: 1006–1013.
25 Kaneko M, Mestre C, Sanchez EH, Ham-mond DL: Intrathecally administered gaba-pentin inhibits formalin-evoked nocicep-tion and the expression of Fos-like immu -n oreactivity in the spinal cord of the rat. J Pharmacol Exp Ther 2000; 292: 743–751.
26 Zimmermann M: Ethical guidelines for in-vestigation of experimental pain in con-scious animals. Pain 1983; 16: 109–110.
27 Wheeler-Aceto H, Cowan A: Standardiza-tion of the rat paw formalin test for the eval-uation of analgesics. Psychopharmacology 1991; 104: 35–44.
28 Yaksh TL, Rudy TA: Chronic catheterization of the spinal subarachnoid space. Physiol Be-hav 1976; 17: 1031–1036.
29 Chen SR, Pan HL: Spinal endogenous acetyl-choline contributes to the analgesic effect of systemic morphine in rats. Anesthesiology 2001; 95: 525–530.
30 Argüelles CF, Torres-López JE, Granados-Soto V: Peripheral antinociceptive action of morphine and the synergistic interaction with lamotrigine. Anesthesiology 2002; 96: 921–925.
31 Jiménez-Andrade JM, Ortiz MI, Pérez-Uri-zar J, Aguirre-Bañuelos P, Granados-Soto V, Castañeda-Hernández G: Synergistic effects between codeine and diclofenac after local, spinal and systemic administration. Phar-macol Biochem Behav 2003; 76: 463–471.
32 Tallarida RJ: Drug Synergism and Dose-Ef-fect Data Analysis, ed 1. New York, Chapman & Hall/CRC, 2000, pp 1–72.
33 Carlton SM, Zhou S: Attenuation of forma-lin-induced nociceptive behaviors following local peripheral injection of gabapentin. Pain 1998; 76: 201–207.
34 Heughan CE, Sawynok J: The interaction be-tween gabapentin and amitriptyline in the rat formalin test after systemic administra-tion. Anesth Analg 2002; 94: 975–980.
35 Granados-Soto V, Argüelles CF: Synergistic antinociceptive interaction between trama-dol and gabapentin after local, spinal and systemic administration. Pharmacology 2005; 74: 200–208.
36 Gee NS, Brown JP, Dissanayake VU, Offord J, Thurlow R, Woodruff GN: The novel anti-convulsant drug, gabapentin (Neurontin), binds to the � 2 � subunit of a calcium chan-nel. J Biol Chem 1996; 271: 5768–5776.
37 Shimoyama M, Shimoyama N, Hori Y: Ga-bapentin affects glutamatergic excitatory neurotransmission in the rat dorsal horn. Pain 2000; 85: 405–414.
38 Ortiz MI, Medina-Tato DA, Sarmiento-He-redia D, Palma-Martínez J, Granados-Soto V: Possible activation of the NO-cyclic GMP-protein kinase G-K + channels pathway by ga-bapentin on the formalin test. Pharmacol Biochem Behav 2006; 83: 420–427.
39 Aguirre-Bañuelos P, Granados-Soto V: Evi-dence for a peripheral mechanism of action for the potentiation of the antinociceptive ef-fect of morphine by dipyrone. J Pharmacol Toxicol 1999; 42: 79–85.
40 Miranda HF, Pinardi G: Isobolographic analysis of the antinociceptive interactions of clonidine with non-steroidal anti-inflam-matory drugs. Pharmacol Res 2004; 50: 273–278.
41 Vazquez E, Hernandez N, Escobar W, Vane-gas H: Antinociception induced by intrave-nous dipyrone (metamizol) upon dorsal horn neurons: involvement of endogenous opioids at the periaqueductal gray matter, the nucleus raphe magnus, and the spinal cord in rats. Brain Res 2005; 1048: 211–217.
42 Carlsson KH, Jurna I: The role of descending inhibition in the antinociceptive effects of the pyrazolone derivatives, metamizole (di-pyrone) and aminophenazone (Piramidon). Naunyn Schmiedebergs Arch Pharmacol 1987; 335: 149–154.
43 Hurley RW, Chatterjea D, Rose Feng M, Tay-lor CP, Hammond DL: Gabapentin and pre-gabalin can interact synergistically with naproxen to produce antihyperalgesia. An-esthesiology 2002; 97: 1263–1273.
44 Yoon MH, Choi JI, Park HC, Bae HB, Jeong SW, Jeong CY: Analysis of interactions be-tween serotonin and gabapentin or adenos-ine in the spinal cord of rats. Pharmacology 2005; 74: 15–22.
45 Hernández-Delgadillo GP, Ventura-Mar-tínez R, Díaz-Reval MI, Domínguez-Ramírez AM, López-Muñoz FJ: Metamizol potenti-ates morphine antinociception but not con-stipation after chronic treatment. Eur J Phar-macol 2002; 441: 177–183.
46 Poveda R, Planas E, Pol O, Romero A, San-chez S, Puig MM: Interaction between met-amizol and tramadol in a model of acute vis-ceral pain in rats. Eur J Pain 2003; 7: 439–448.
47 Terán-Rosales F, Medina-Santillán R, Reyes-García G, Granados-Soto V: Synergistic an-tinociceptive interaction between acetamin-ophen or metamizol and B vitamins in the formalin test. Drug Dev Res 2006; 66: 286–294.
Recommended

















