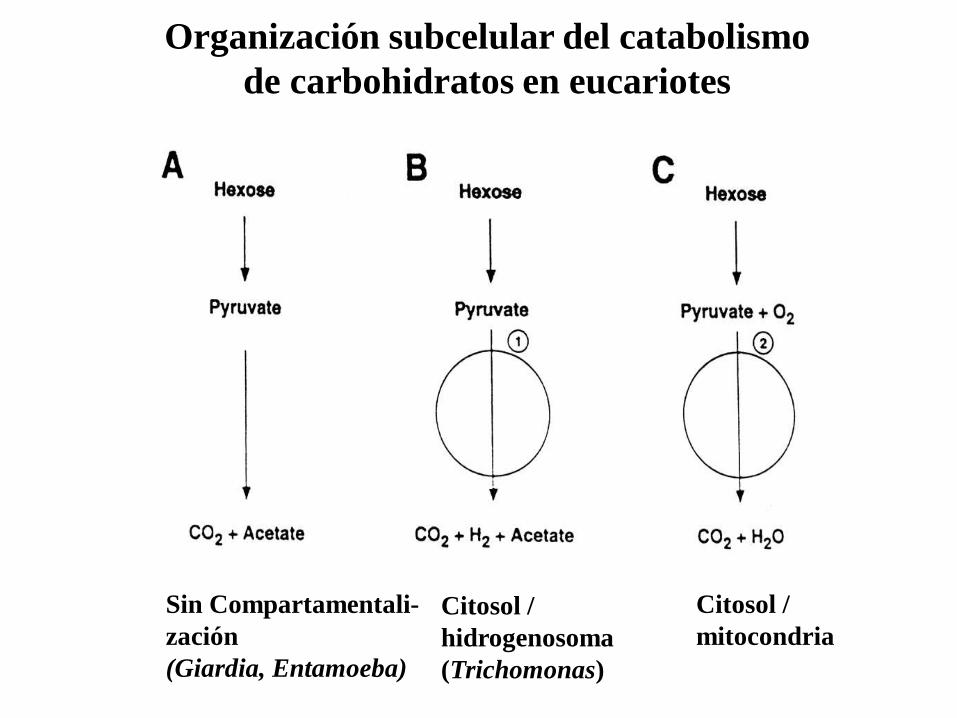
Organización subcelular del catabolismo
de carbohidratos en eucariotes
Sin Compartamentali-
zación
(Giardia, Entamoeba)
Citosol /
hidrogenosoma
(Trichomonas)
Citosol /
mitocondria

GLUCOLISIS
Plasmodium
Trypanosoma brucei,
forma sanguínea.
Trichomonas
vaginalis
Entamoeba
histolytica

METABOLISMO DE LOS CARBOHIDRATOS.
Trypanosomas y Leishmanias:
Consumo igual o menor de glucosa en anaerobiosis comparado con el consumo
aeróbico: falta de efecto Pasteur o aún efecto Pasteur inverso.
Producción y liberación al medio de catabolitos de la glucosa parcialmente
reducidos (= fermentación) aún en presencia de oxígeno: succinato,
acetato, glicerol, L-alanina, etanol. (= fermentación aeróbica de la glucosa).
Carecen de reserva de glucógeno, al igual que otros hemoparásitos. Las
Leishmanias contienen un polímero de manosa, el manógeno, que cumple una función
similar.
Trichomonas vaginalis:
Producción de H2 sólo en anaerobiosis. Producción de glicerol, lactato y acetato
tanto en aerobiosis como en anaerobiosis.
Giardia lamblia:
Producción de etanol, CO2 y acetato tanto en aerobiosis como en anaerobiosis.
Entamoeba histolytica:
Producción de etanol y CO2 en anaerobiosis, y de etanol, CO2 y acetato en
aerobiosis.
Plasmodium falciparum:
Las formas eritrocíticas producen lactato y no tienen un Ciclo de Krebs funcional.
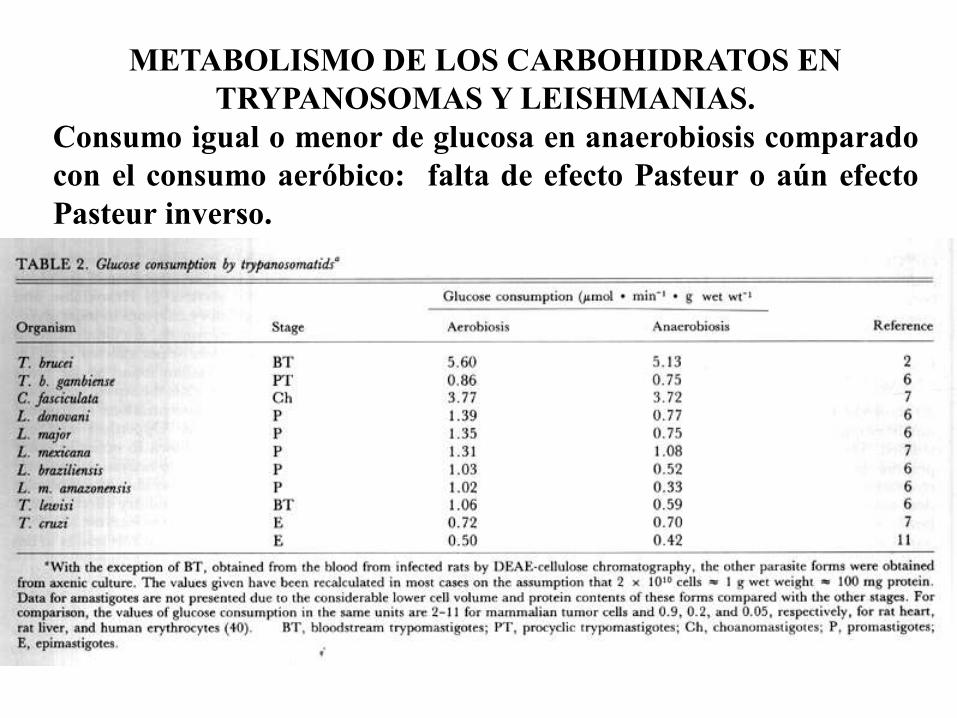
METABOLISMO DE LOS CARBOHIDRATOS EN
TRYPANOSOMAS Y LEISHMANIAS.
Consumo igual o menor de glucosa en anaerobiosis comparado
con el consumo aeróbico: falta de efecto Pasteur o aún efecto
Pasteur inverso.

Producción y liberación al medio de catabolitos de la glucosa parcialmente
reducidos (= fermentación) aún en presencia de oxígeno: succinato,
acetato, glicerol, L-alanina, etanol. (= fermentación aeróbica de la glucosa).
Carecen de reserva de glucógeno, al igual que otros hemoparásitos. Las
Leishmanias contienen un polímero de manosa, el manógeno, que cumple
una función similar.
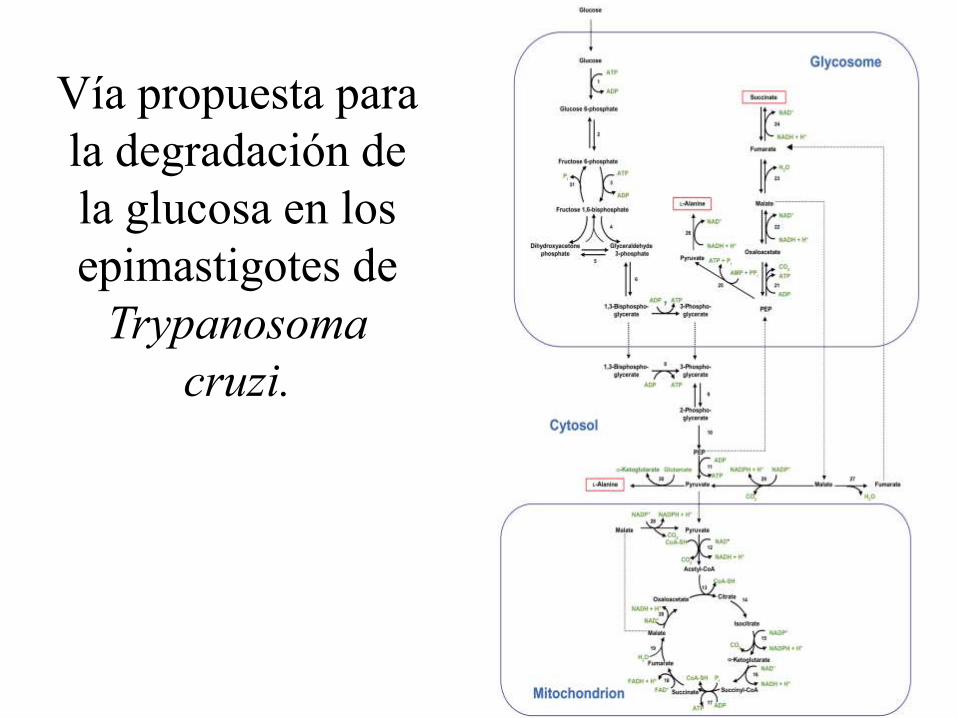
Vía propuesta para
la degradación de
la glucosa en los
epimastigotes de
Trypanosoma
cruzi.
PEP Piruvato
PEP Piruvato L-Alanina
Succinato
Fumarato
L-malato
Oxalacetato
CAT y
transaminación
Glucosa
Fructosa 1,6-bisfosfato
Gliceraldehído 3-fosfato
1,3-bisfosfoglicerato
3-fosfoglicerato
3-fosfoglicerato
2 ATP
2 ADP
ADP
ATP
NAD+
NADH
NADH
NAD+
NADH
NAD+
ADP
ATP
AMP ATP NADH NAD+
b-oxidación de ácidos grasos
Síntesis de esteroides
Metabolismo de purinas
y pirimidinas
PPi
ATP
PK
PPDK
Glicosoma
Citosol
PEPCK
CO2

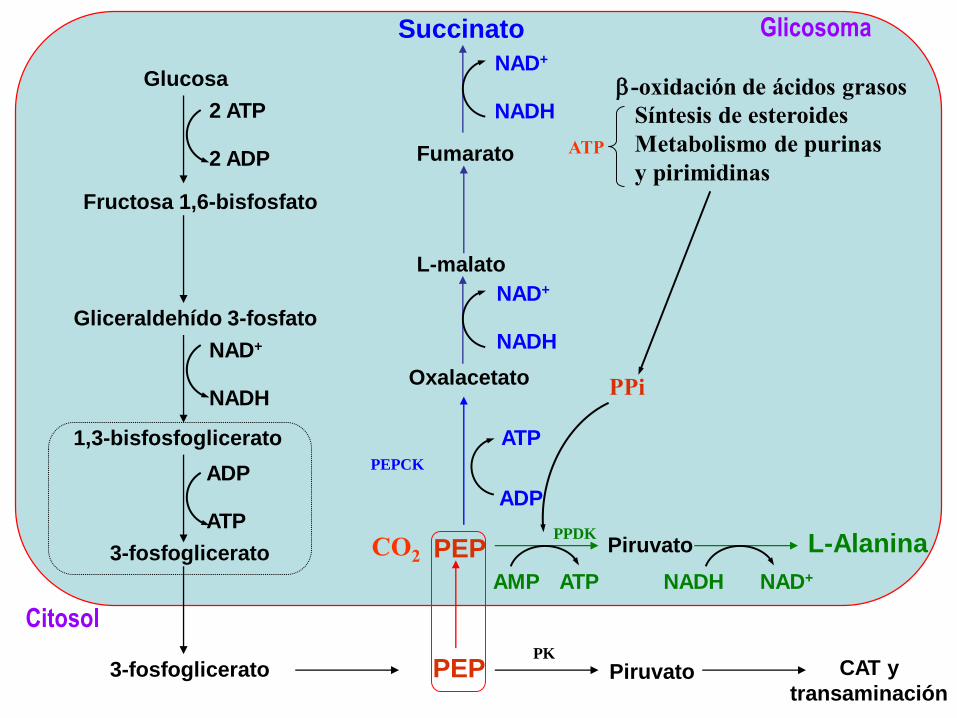
Resumen del Metabolismo Energético en epimastigotes de T.
cruzi.
1) Se ha estudiado el transporte de la glucosa en epimastigotes y
trypomastigotes sanguíneos; los amastigotes intracelulares no transportan
glucosa y consumirían esencialmente aminoácidos, y talvez lípidos.
2) Los productos finales del catabolismo de la glucosa por los epimastigotes
son el succinato y la L-alanina; el acetato no proviene de la glucosa. Una
concentración alta de CO3H- favorece la producción de succinato, y la
inhibición de la PEPCK la producción de L-alanina.
3) La falta del efecto Pasteur se debería a la falta del control de la HK (que no es
inhibida por G-6-P) y de la PFK (que no es inhibida por ATP, citrato o Pi y es
activada por AMP).
4) Otras dos diferencias con otros organismos son que la PFK, siendo
dependiente de ATP, tiene la máxima similitud de secuencia con las PFKs
dependientes de PPi, y que la PK es activada por F 2,6-bisP, que contrarresta
su inhibición por ATP.
5) No hay una prueba definitiva de que el Ciclo de Krebs sea completamente
funcional.
6) El Complejo I de la cadena respiratoria no sería funcional, y la oxidación del
NADH se haría a través de la fumarato reductasa dependiente de NADH,
dando succinato que entraría por el Complejo II, funcional, como los
complejos III y IV y la F0F1-ATP sintasa.
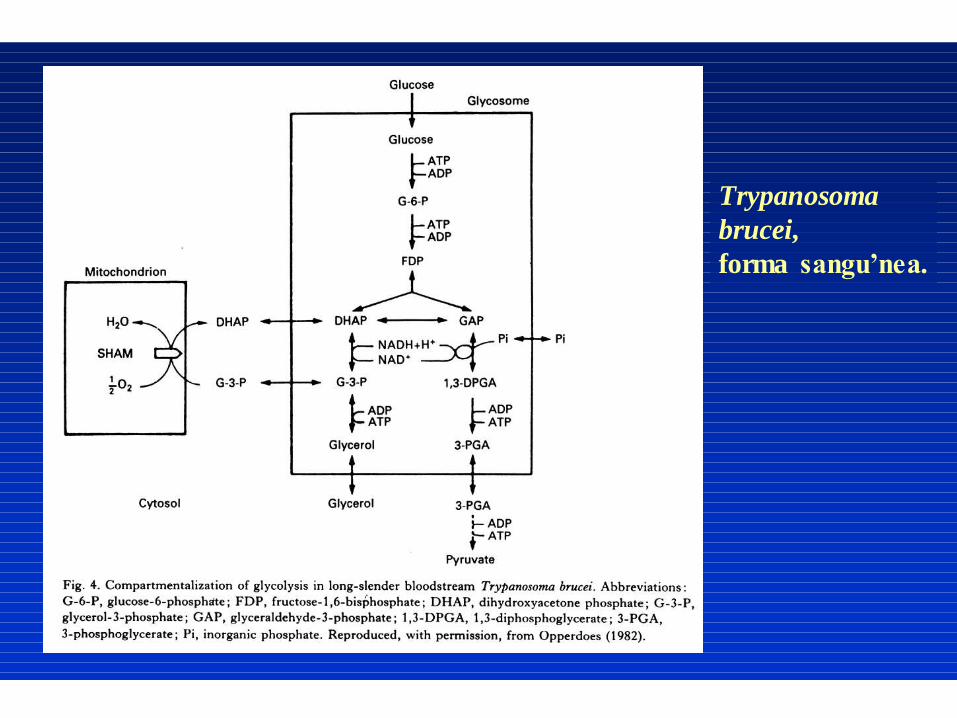
Trypanosoma
brucei,
forma sangu’nea.
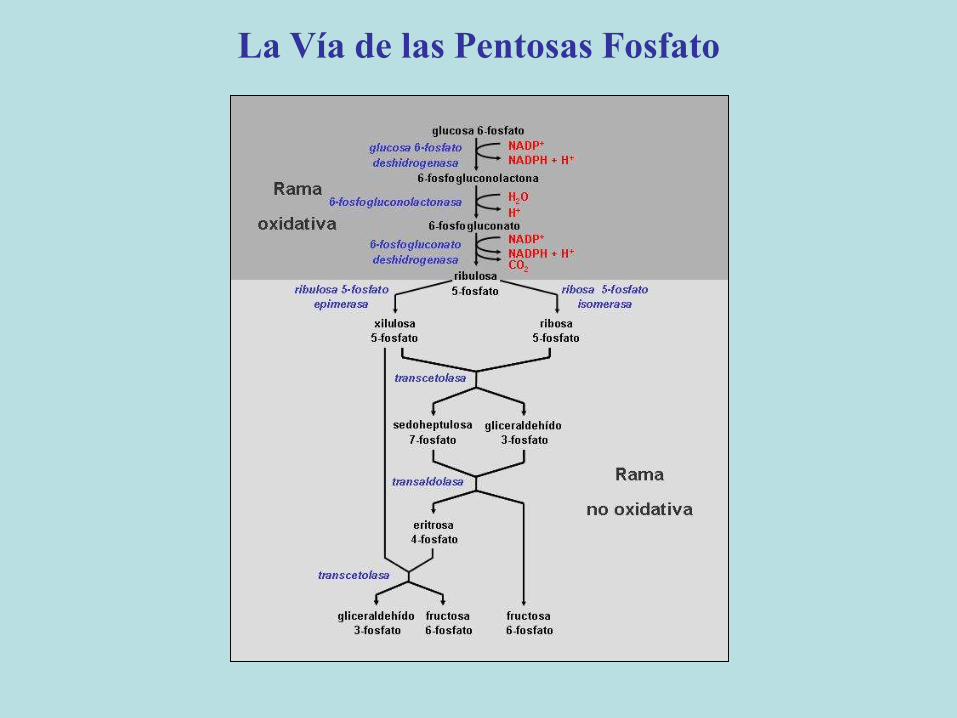
La Vía de las Pentosas Fosfato

PARASITOS
ANAEROBIOS
(MICROAEROFILOS)
Trichomonas vaginalis -
Tritrichomonas foetus
Entamoeba histolytica
Giardia intestinalis (Giardia lamblia)

Protozoarios microaerofilos carentes de mitocondrias.
Los mas importantes son:
Entamoeba histolytica, agente de la amebiasis. Afecta al intestino
delgado, causando la disentería amebiana, y puede atravesar la
pared intestinal e invadir órganos como el hígado, produciendo el
absceso amebiano hepático, mortal si no es tratado a tiempo.
Carece de mitocondrias y no tiene otras organelas, como el
hidrogenosoma. Forma de resistencia: el quiste, eliminado en las
heces. Frecuente en países tropicales.
Giardia intestinalis o Giardia lamblia, agente de la giardiasis.
Forma de resistencia: el quiste, eliminado en las heces. Presente
en todos los países, aún los mas desarrollados.
Trichomonas vaginalis y Tritrichomonas foetus, parásitos del ser
humano y del ganado vacuno, respectivamente. Sin forma de
resistencia; transmisión directa, por contagio venéreo.

METABOLISMO
DE LA
GLUCOSA POR
ENTAMOEBA sp.
Todo el
metabolismo tiene
lugar en el citosol.

Enzimas que utilizan pirofosfato: presentes en general en
plantas y bacterias, y tambien en algunos Protozoarios.
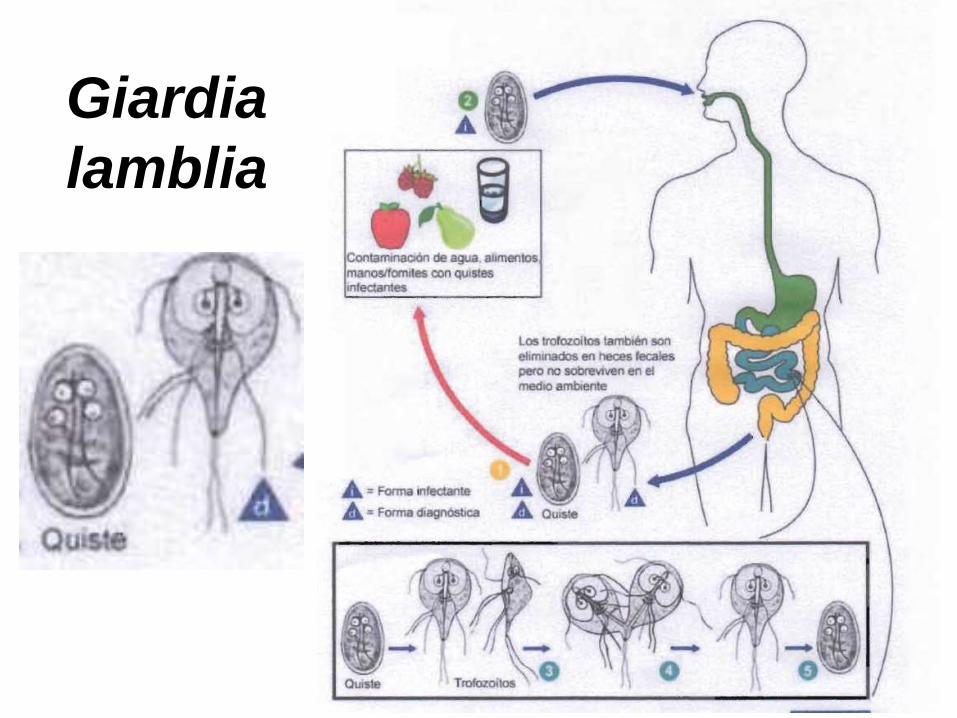
Giardia
lamblia
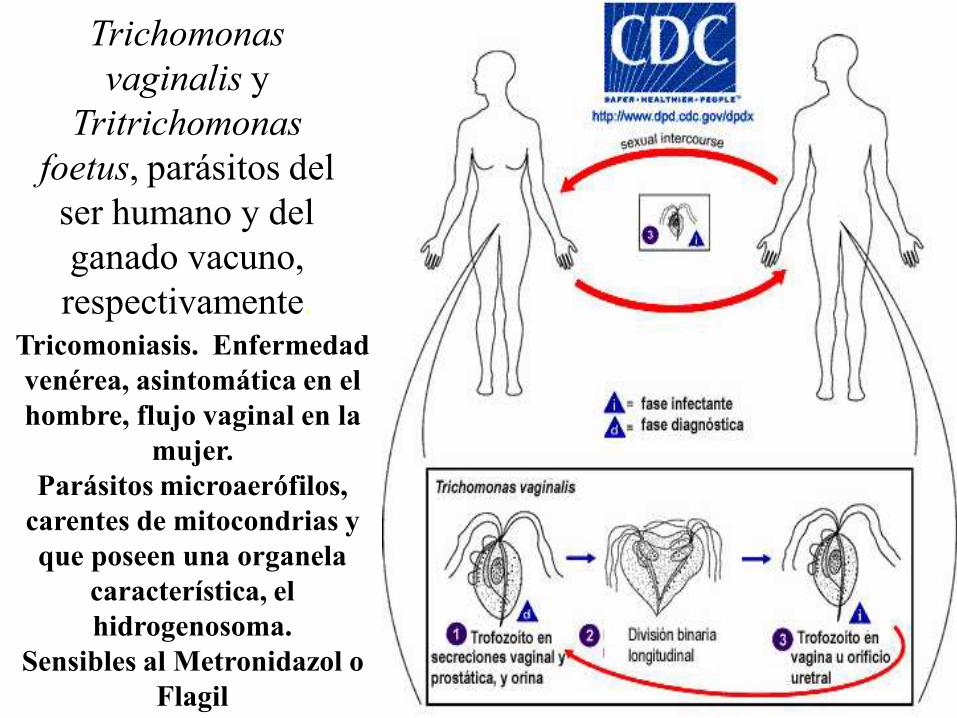
Trichomonas
vaginalis y
Tritrichomonas
foetus, parásitos del
ser humano y del
ganado vacuno,
respectivamente.Tricomoniasis. Enfermedad
venérea, asintomática en el
hombre, flujo vaginal en la
mujer.
Parásitos microaerófilos,
carentes de mitocondrias y
que poseen una organela
característica, el
hidrogenosoma.
Sensibles al Metronidazol o
Flagil
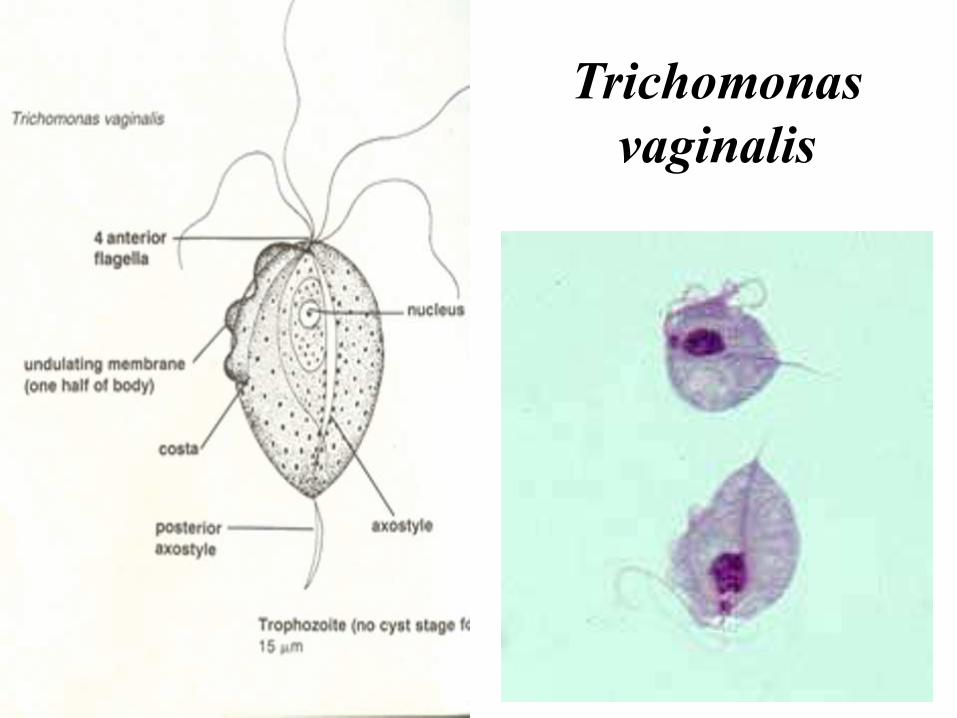
Trichomonas
vaginalis
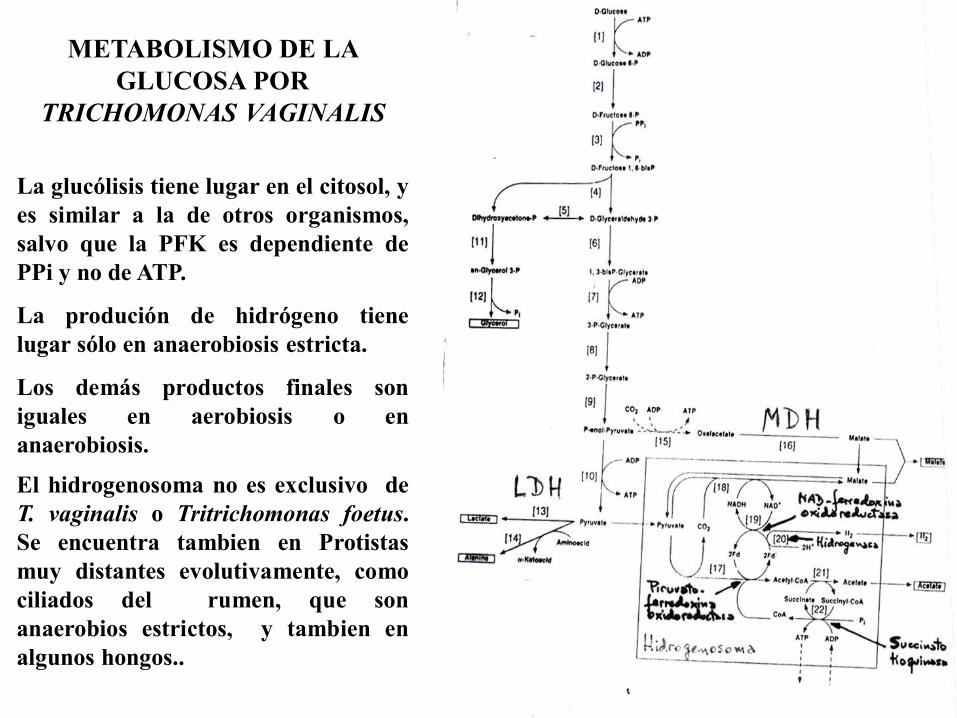
METABOLISMO DE LA
GLUCOSA POR
TRICHOMONAS VAGINALIS
La glucólisis tiene lugar en el citosol, y
es similar a la de otros organismos,
salvo que la PFK es dependiente de
PPi y no de ATP.
La produción de hidrógeno tiene
lugar sólo en anaerobiosis estricta.
Los demás productos finales son
iguales en aerobiosis o en
anaerobiosis.
El hidrogenosoma no es exclusivo de
T. vaginalis o Tritrichomonas foetus.
Se encuentra tambien en Protistas
muy distantes evolutivamente, como
ciliados del rumen, que son
anaerobios estrictos, y tambien en
algunos hongos..
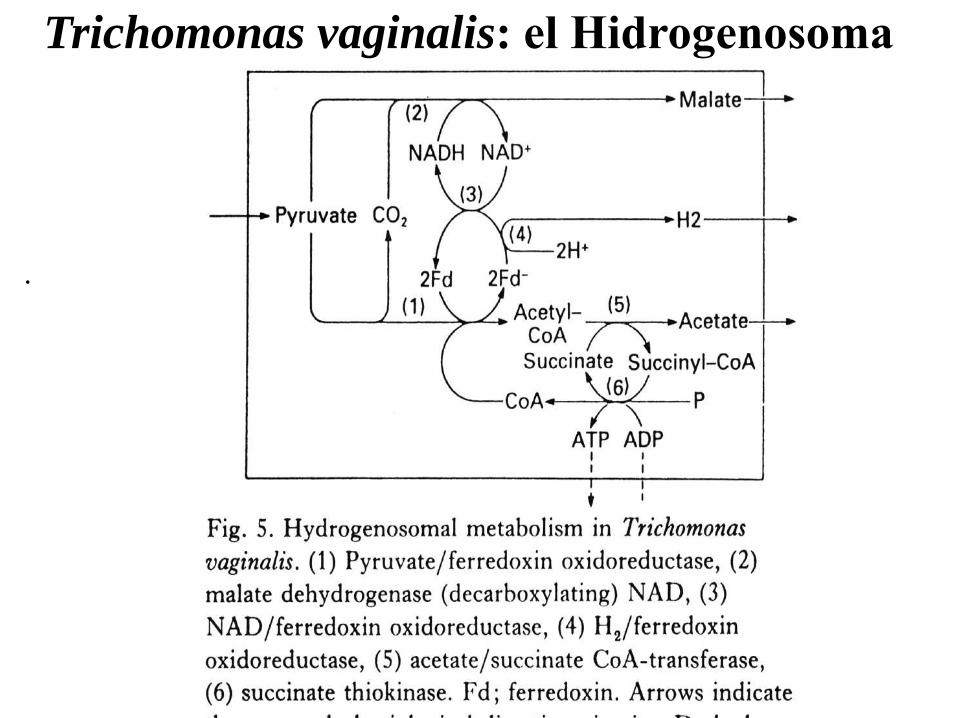
Trichomonas vaginalis: el Hidrogenosoma
.

APICOMPLEXACoccidios: Toxoplasma gondii y
Eimeria spp. (parásitos de aves y conejos)
Plasmodium spp.
Piroplasmas: Babesia spp. y Theileria
spp. (parásitos de mamíferos, en particular el
ganado)
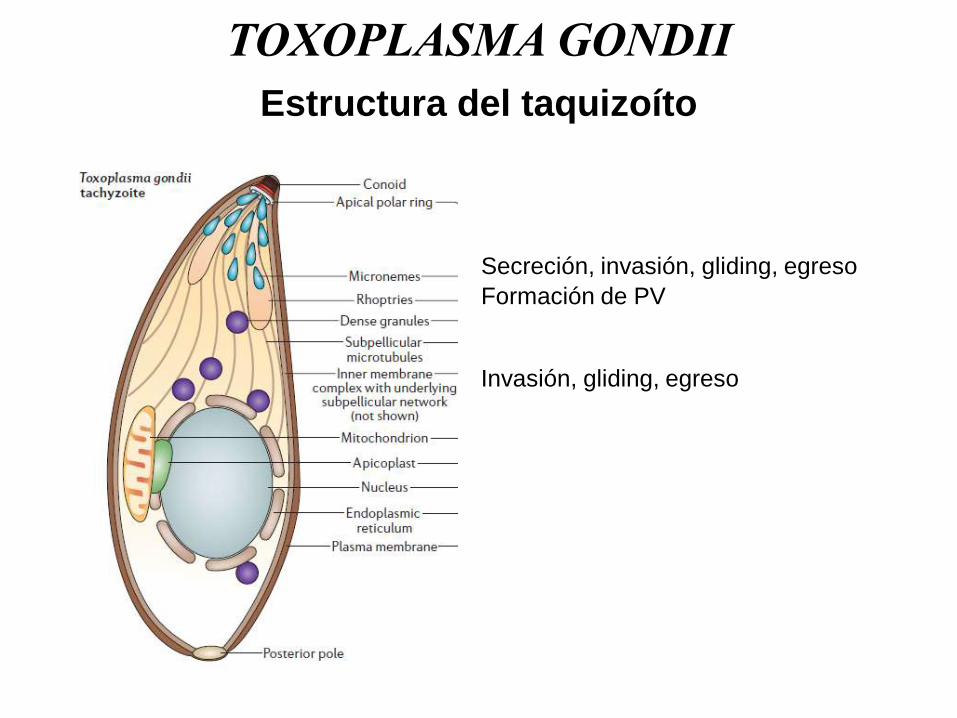
Estructura del taquizoíto
Formación de PV
Secreción, invasión, gliding, egreso
Invasión, gliding, egreso
Baum J., et al., Nat. Rev. Microbiol., 2006.
TOXOPLASMA GONDII
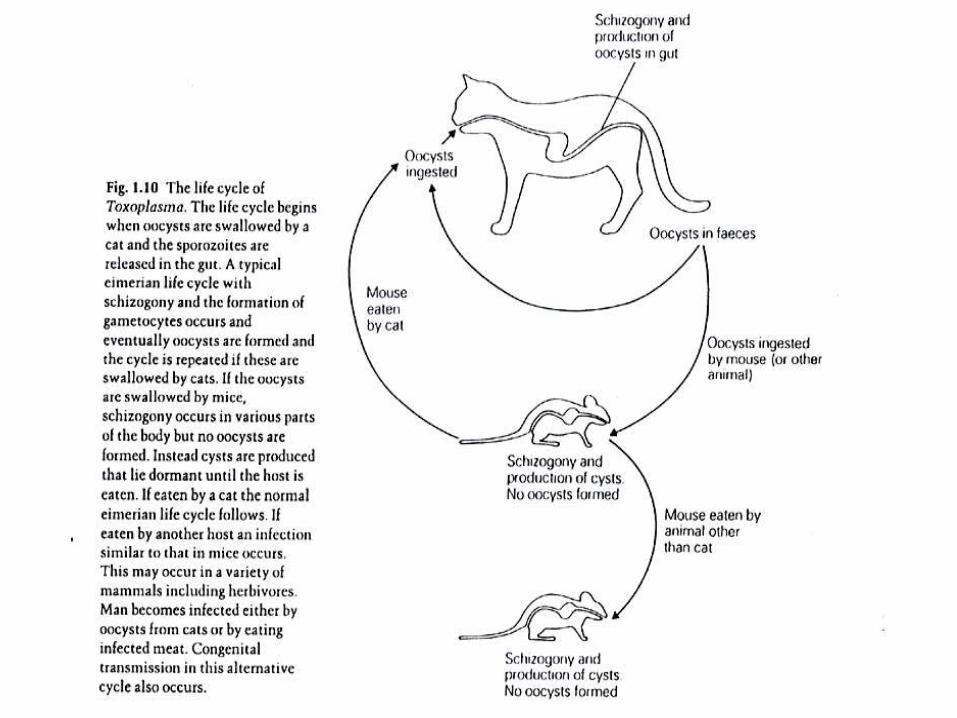
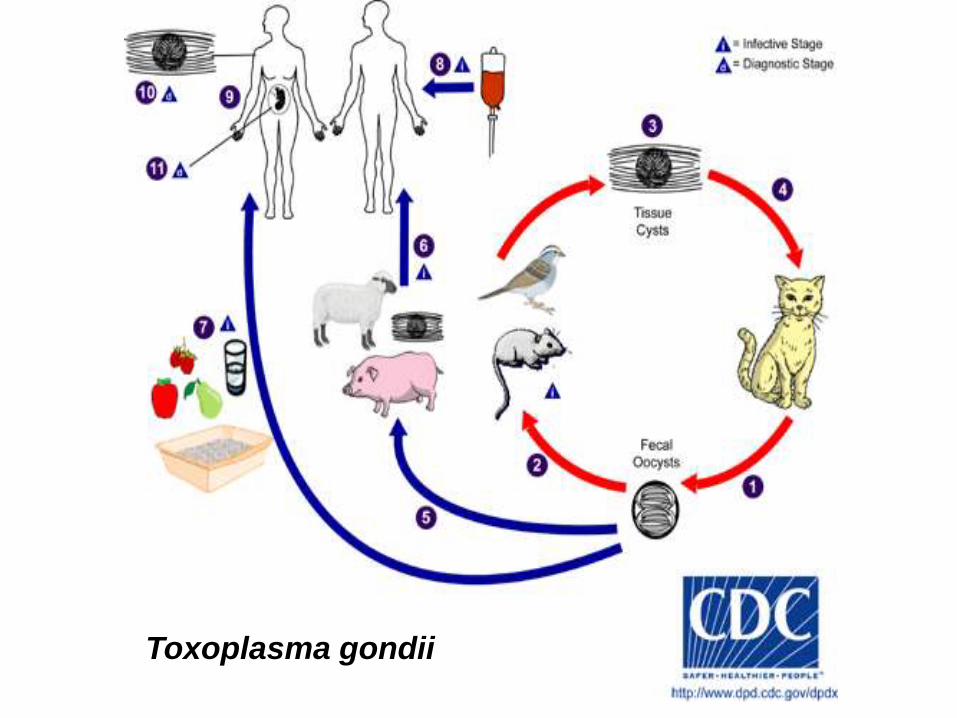
Toxoplasma gondii
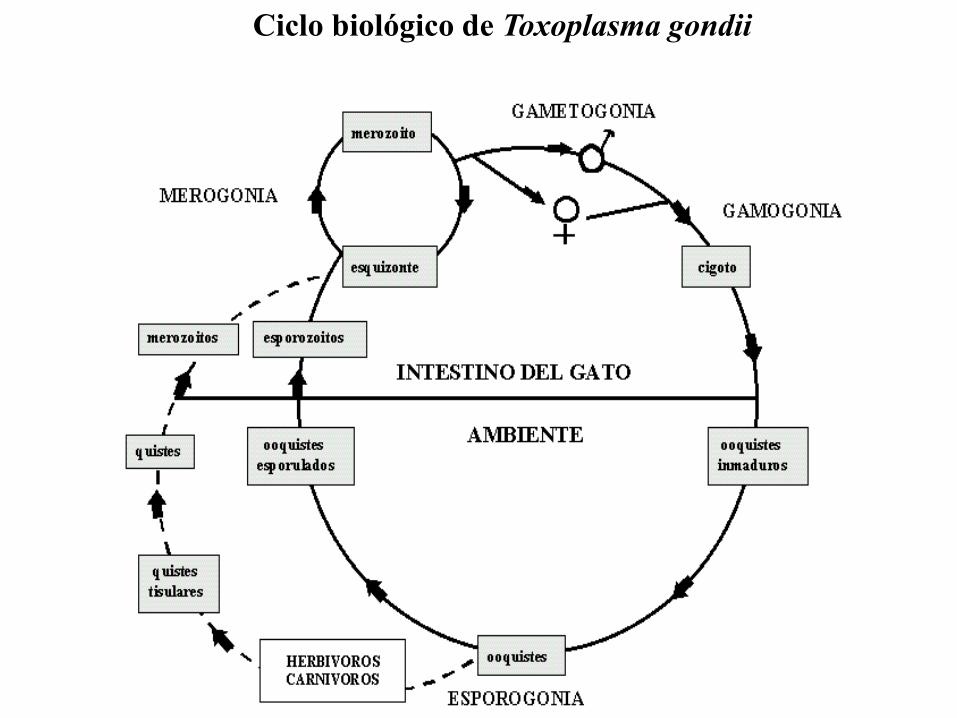
Ciclo biológico de Toxoplasma gondii
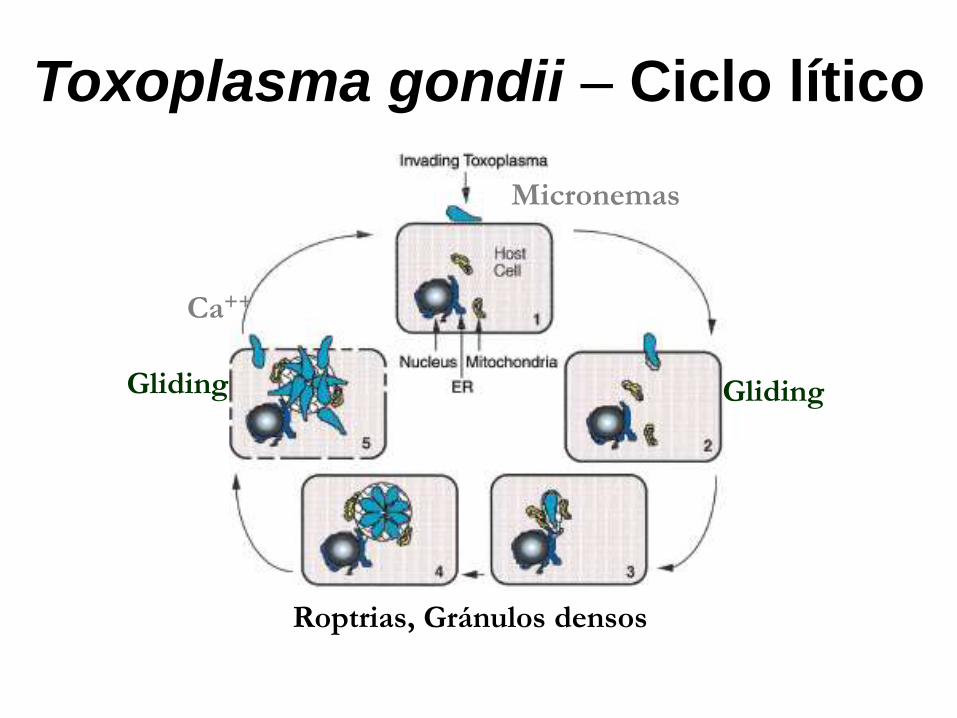
Toxoplasma gondii – Ciclo lítico
Black, M.W. Mol. Biol. Rev., 2000.
Ca++
Gliding Gliding
Roptrias, Gránulos densos
Micronemas
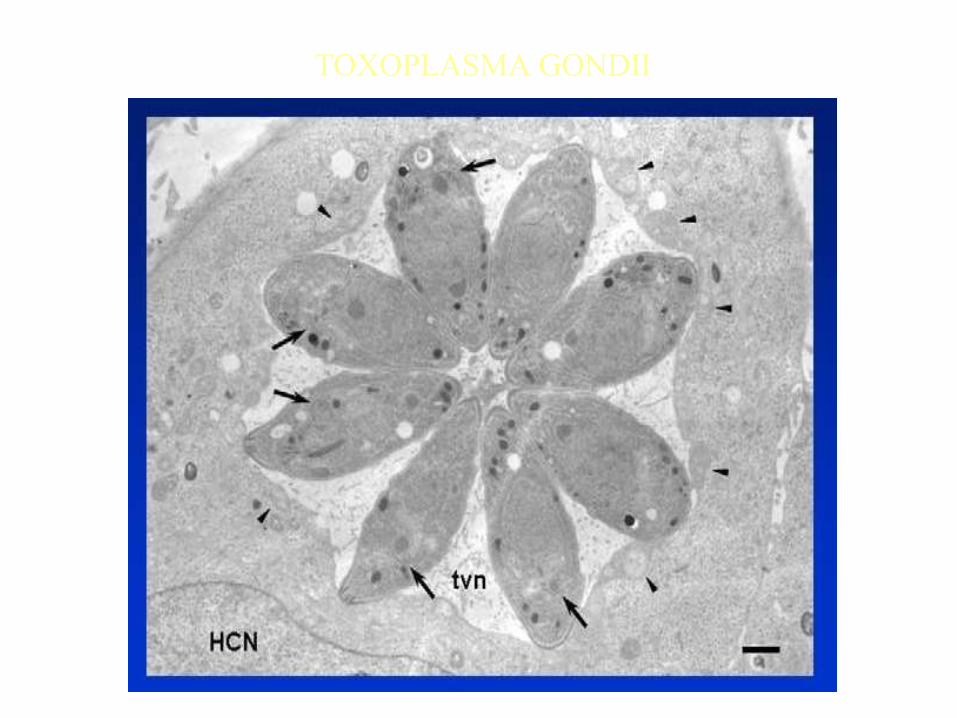
TOXOPLASMA GONDII
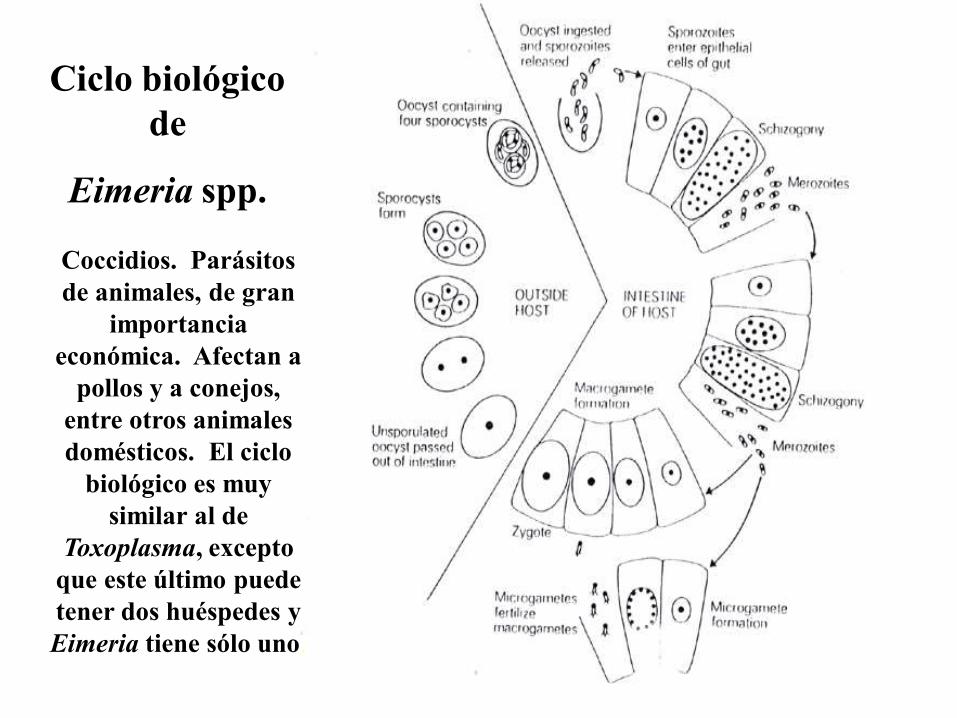
Ciclo biológico
de
Eimeria spp.
Coccidios. Parásitos
de animales, de gran
importancia
económica. Afectan a
pollos y a conejos,
entre otros animales
domésticos. El ciclo
biológico es muy
similar al de
Toxoplasma, excepto
que este último puede
tener dos huéspedes y
Eimeria tiene sólo uno.

Los únicos huéspedes definitivos de Toxoplasma gondii son miembros de la familia Felidae
(gatos domésticos y otros félidos). Los oocistos no esporulados se eliminan en la materia fecal
del gato. Aunque los oocistos se eliminan sólo durante 1-2 semanas, pueden eliminarse en
gran número. Los oocistos tardan 1-5 días para esporular en el ambiente y hacerse infectivos.
Los huéspedes intermedios naturales (que incluyen aves y roedores) se infectan después de
ingerir tierra, agua o material vegetal contaminado con oocistos. Los oocistos se transforman
en taquizoítos poco después de la ingestión. Estos taquizoítos se localizan en tejido muscular y
neural y se desarrollan como bradizoítos en quistes tisulares.
Los gatos se infectan después de consumir huéspedes intermedios que contienen quistes
tisulares. Los gatos pueden también infectarse directamente por ingestión de oocistos
esporulados. Los animales criados para consumo humano o salvajes pueden también
infectarse con quistes tisulares después de la ingestión de oocistos esporulados en el ambiente.
Los humanos pueden infectarse por una de varias rutas:
Comiendo carne mal cocida de animales que contienen quistes tisulares.
Consumiendo comida o agua contaminada con heces de gato o por muestras ambientales
contaminadas (por ejemplo tierra contaminada o al cambiar el contenido de la caja de un
gato mascota).
Por transfusión de sangre o transplante de órganos.
Transplacentariamente, de la madre al feto.
En el huésped humano, los parásitos forman quistes tisulares, comúnmente en el músculo
esquelético, miocardio, cerebro y ojos; estos quistes pueden permanecer a lo largo de toda la
vida del huésped. El diagnóstico se hace usualmente por serología, aunque los quistes
tisulares pueden ser observados en especímenes teñidos de biopsias. El diagnóstico de la
infección congénita puede lograrse detectando DNA de T. gondii en el flúido amniótico
usando métodos moleculares como la PCR.

Causada por parásitos sanguíneos del género Plasmodium. Hay
aproximadamente 156 especies de Plasmodium que pueden
infectar varias especies de vertebrados. Son muy específicos de la
especie que invaden. Cuatro especies son consideradas
verdaderos parásitos de humanos, pues utilizan humanos casi
exclusivamente como un hospedador intermedio natural: P.
falciparum (terciana maligna), P. malariae (cuartana), P. vivax
(terciana benigna), P. ovale (terciana ovale). Sin embargo, hay
informes periódicos de plasmodios de simios encontrados en
humanos, en la mayoría de los casos implicando a P. knowlesi. No
se ha determinado aún si P. knowlesi puede ser transmitido
naturalmente de humano a humano por el mosquito, sin
intervención del hospedador intermedio natural (macacos,
género Macaca). Por lo tanto, P. knowlesi es aún considerado
como una malaria zoonótica.
PLASMODIUM spp. Y LA MALARIA
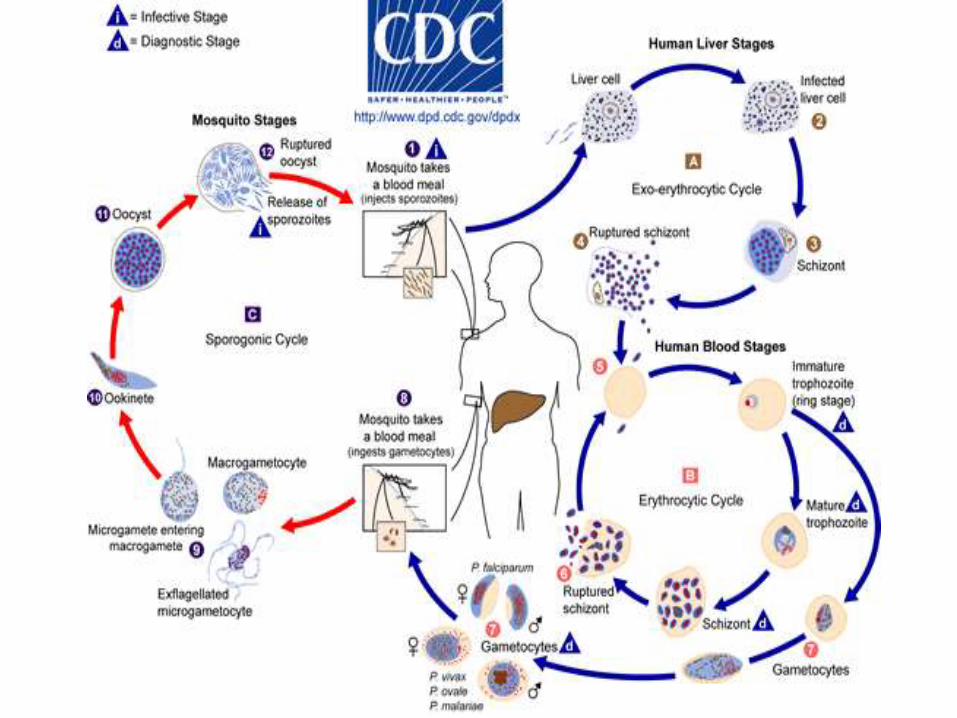
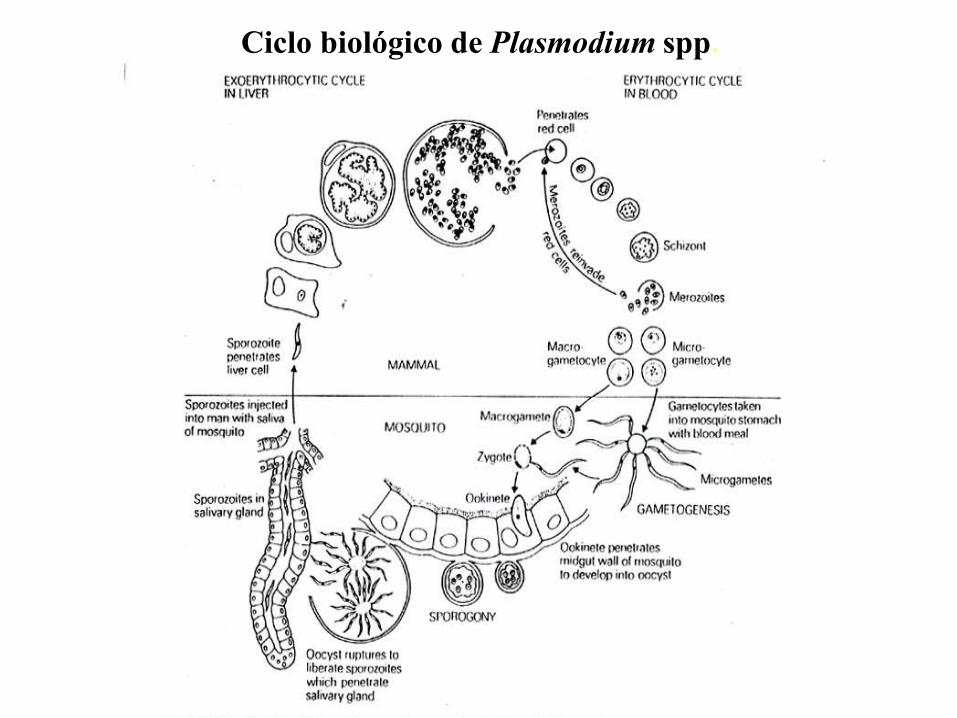
Ciclo biológico de Plasmodium spp.

El ciclo biológico del parásito de la malaria involucra dos huéspedes. Durante su
alimentación con sangre, una hembra infectada del mosquito Anopheles inocula
esporozoitos al hospedador humano. Los esporozoitos infectan hepatocitos y maduran
dando esquizontes, que se rompen liberando merozoitos. (En los casos de P. vivax y P.
ovale un estadio durmiente [hipnozoitos] puede persistir en el hígado y causar
relapsos invadiendo la sangre semanas, o aún años, después de la infección.) Después
de la replicación inicial en el hígado (esquizogonia exo-eritrocítica), los parásitos se
replican asexualmente en los eritrocitos (esquizogonia eritrocítica). Los merozoitos
infectan los glóbulos rojos y se transforman en trofozoítos en estadío de anillo, los
cuales luego maduran dando esquizontes, que se rompen liberando merozoitos.
Algunos parásitos se diferencian a estadíos sexuales eritrocíticos (gametocitos). El
estadío sanguíneo de los parásitos es responsable de las manifestaciones clínicas de la
enfermedad.
Los gametocitos, masculinos (microgametocitos) y femeninos (macrogametocitos), son
ingeridos por un mosquito Anopheles mientras se alimenta con la sangre del humano
infectado. La multiplicación de los parásitos en el mosquito se denomina ciclo
esporogónico. En el estómago del mosquito las microgametas penetran en las
macrogametas generando zigotas. Las zigotas adquieren motilidad y se elongan
transformándose en oocinetos, los cuales invaden la pared del intestino medio del
mosquito, donde se desarrollan como oocistos. Los oocistos crecen, se rompen, y
liberan esporozoitos, los que se dirigen a las glándulas salivales. La inoculación de los
esporozoitos a un nuevo hospedador humano perpetúa el ciclo biológico de la malaria.
PIROPLASMAS
Parásitos de animales, con particular importancia para el ganado
vacuno. Transmitidos por garrapatas.
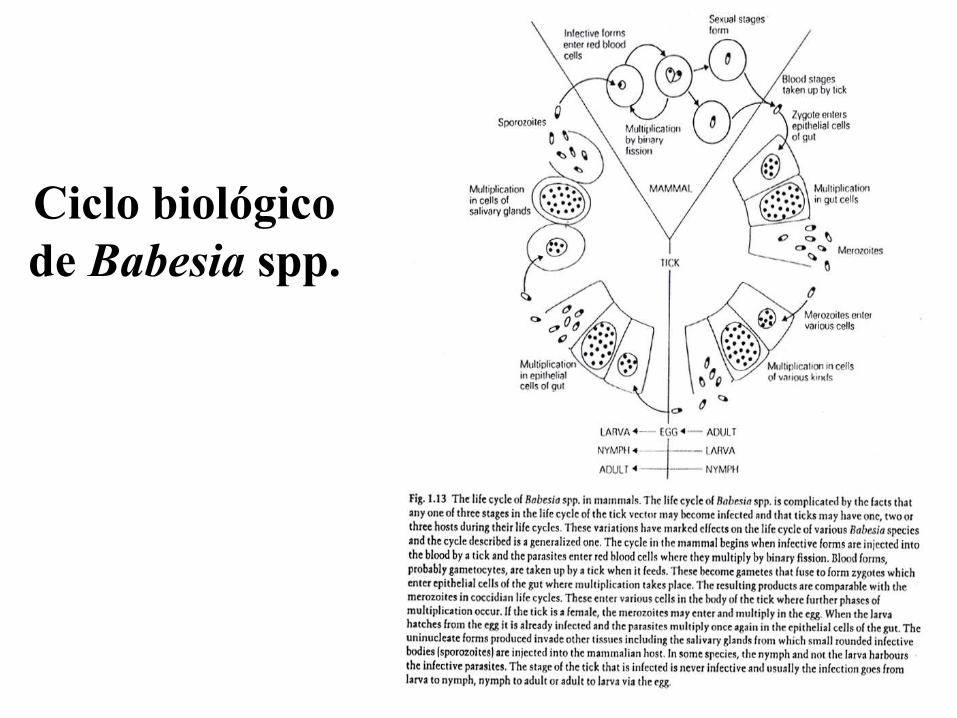
Ciclo biológico
de Babesia spp.
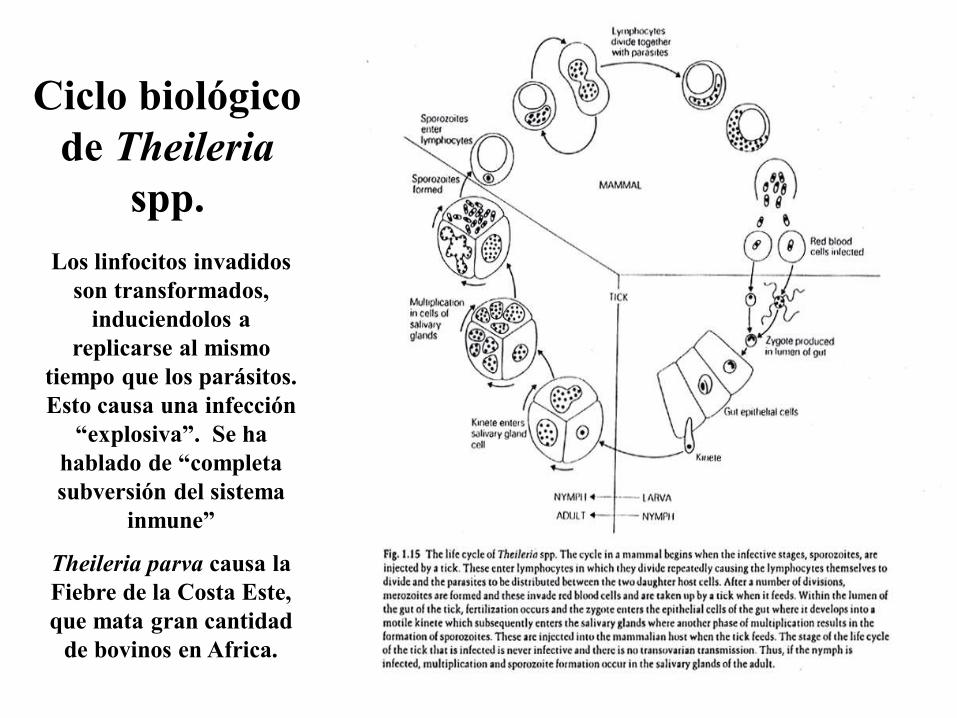
Ciclo biológico
de Theileria
spp.
Los linfocitos invadidos
son transformados,
induciendolos a
replicarse al mismo
tiempo que los parásitos.
Esto causa una infección
“explosiva”. Se ha
hablado de “completa
subversión del sistema
inmune”
Theileria parva causa la
Fiebre de la Costa Este,
que mata gran cantidad
de bovinos en Africa.
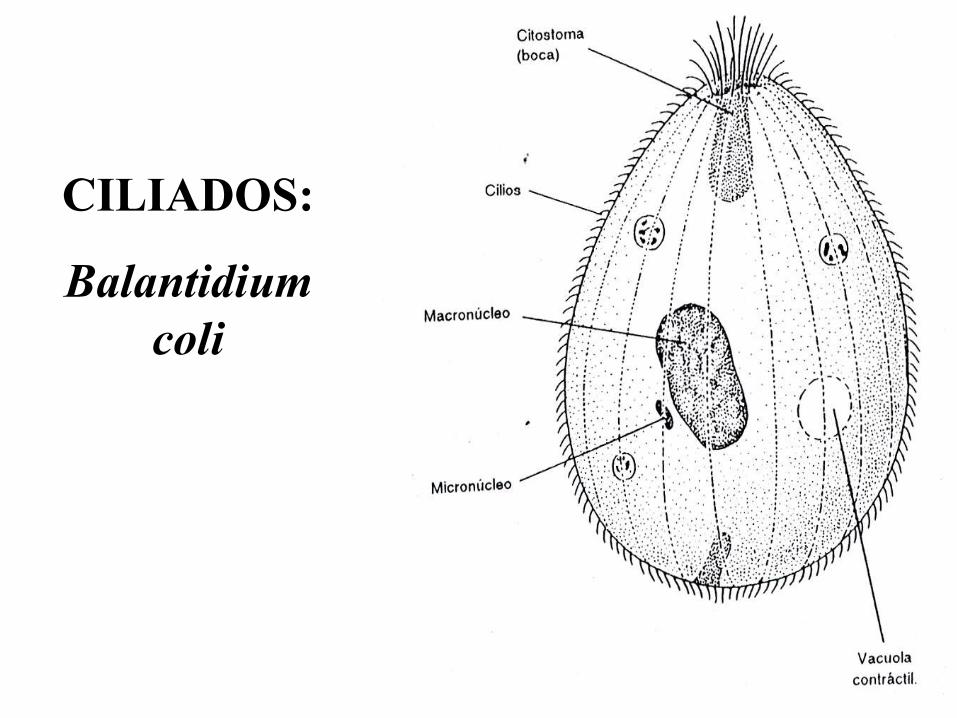
CILIADOS:
Balantidium
coli

METABOLISMO DE PROTEINAS, AMINOACIDOS, PURINAS Y
PIRIMIDINAS.
PROTEINASAS.
Las proteinasas se clasifican, según su mecanismo de reacción, en
cistein proteinasas, serin proteinasas, treonin proteinasas (el proteasoma),
aspartil proteinasas y metaloproteinasas. Todas estas clases han sido
encontradas en Protozoarios parásitos, aunque no todas en todos ellos. Son
en general importantes no sólo para la nutrición del parásito, sino que en
muchos casos participan activamente en el mecanismo de patogénesis, y
puede considerarselas factores de virulencia.
En Trypanosomas, Leishmanias, Entamoeba, Giardia y Trichomonas,
predominan las cisteín proteinasas. Las Leishmanias poseen tambien una
metaloproteinasa de superficie cuantitativamente muy importante.
En Plasmodium falciparum la digestión de la hemoglobina del
glóbulo rojo parasitado se hace por la acción concertada de cisteín
proteinasas (las falcipaínas) y aspartil proteinasas (las plasmepsinas).
La inhibición de cistein proteinasas de Trypanosoma cruzi (la
cruzipaína) y de Plasmodium falciparum mata al parásito o impide etapas de
diferenciación en su ciclo biológico. Se están ensayando inhibidores de la
cruzipaína como eventuales agentes quimioterápicos contra la enfermedad
de Chagas.

CATABOLISMO DE AMINOACIDOS. Los protozoarios parásitos toman en general los aminoácidos del hospedador, y sus
capacidades sintéticas son limitadas. Los utilizan para la biosíntesis proteica, para otros
procesos biosintéticos y para la generación de energía por oxidación de la cadena
carbonada (en los casos en que hay un Ciclo de Krebs y una cadena respiratoria
funcionales).
Algunas de las vías metabólicas son apreciablemente diferentes de las presentes en el
hospedador. Por ejemplo, el catabolismo de los aminoácidos aromáticos (fenilalanina,
tirosina y triptofano) en los Trypanosomas es mucho mas simple que en los mamíferos, y
resulta en la liberación al medio de los derivados aromáticos del lactato, a través de solo
dos reacciones enzimáticas, una transaminación y una reducción.
BIOSINTESIS Y RECUPERACION DE PIRIMIDINAS Y PURINAS.Los parásitos tienden a simplificar su metabolismo, tomando del hospedador tantos
componentes esenciales ya formados como sea posible. Esto es particularmente claro en
el caso de las bases púricas, las cuales no son sintetizadas, sino que los organismos las
recuperan del medio. En el caso de las bases pirimidínicas, además de recuperación del
medio, la generalidad de los Protozoarios parásitos son capaces de sintetizarlas.
HEMOLos Trypanosomas y las Leishmanias son totalmente incapaces de sintetizar el hemo
necesario para sus citocromos y otras hemoproteínas, y deben obtenerlo del medio.
Los Plasmodios tienen la necesidad de evitar los efectos tóxicos de la acumulación de
hemo proveniente de la hemoglobina digerida, y para ello lo precipitan en forma de
pigmento malárico o hemozoína.

CONCEPTOS MAS IMPORTANTES.
Protozoarios: grupo polifilético no relacionado de organismos eucariotas inferiores,
unicelulares, que comparten algunas características morfológicas, reproductivas,
ecológicas y bioquímicas.
Enfermedad de Chagas, Enfermedad del Sueño y Leishmaniasis. Trypanosomátidos:
Un flagelo. Única mitocondria y kinetoplasto. Glicosoma. T. cruzi: formas
amastigote y trypomastigote sanguíneo en mamífero, epimastigote y trypomastigote
metacíclico en vinchuca. T. brucei: trypomastigotes sanguíneos delgado y grueso en
mamífero, Trypomastigote procíclico, epimastigote y trypomastigote metacíclico en
mosca. Variación antigénica. Leishmanias: promastigotes en insecto, amastigotes en
los macrófagos del mamífero. Metabolismo incompleto de la glucosa: fermentación
aeróbica. Falta de efecto Pasteur. Metabolismo de glucosa diferente en T. brucei
sanguíneo, con oxidasa terminal alternativa en la mitocondria.
Microaerófilos: Trichomonas vaginalis. Enfermedad venérea. Hidrogenosoma.
Giardia lamblia y Entamoeba histolytica: Enfermedades que causan.
Apicomplexa: Intracelulares. Complejo apical. Apicoplasto. Toxoplasma gondii:
taquizoítos y bradizoítos. Felinos como huésped definitivo. Transmisión por quistes
en material fecal del felino. Plasmodium falciparum y otros: Específicos de especie.
Vector: mosquito Anopheles. Ciclo esquizogónico en el mamífero y esporogónico
(sexual) en el mosquito.
Recommended

















