
Directores Dra. Ana María Kettlun Dra. María Antonieta Valenzuela Departamento de Bioquímica y Biología Molecular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile
UNIVERSIDAD DE CHILE FACULTAD DE CIENCIAS QUÍMICAS Y FARMACÉUTICAS DEPARTAMENTO DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR
“ANÁLISIS DE LAS MODIFICACIONES POST-
TRADUCCIONALES DE LA PROTEÍNA TAU PRESENTE EN LÍQUIDO CEFALORRAQUÍDEO DE PACIENTES CON
PARAPARESIA ESPÁSTICA TROPICAL”
Memoria para optar al título de Bioquímico
EMILIO JAVIER ORTIZ RIAÑO
Santiago de Chile, 2006
Patrocinante Dra. Ana María Kettlun Departamento de Bioquímica y Biología Molecular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile

ii
“Hoy doy un nuevo paso, en la senda de mi sueño”
Dedicada a mis padres y hermana que sin su apoyo, nada de esto sería posible

iii
Agradecimientos
Quisiera partir agradeciendo a la Dra. María Antonieta Valenzuela quién fue
fundamental, no sólo en lograr que yo sacara adelante esta tesis, si no que además en el
apoyo incondicional durante todo este tiempo, su confianza plena en mis actividades y esas
agradables tardes conversando, compartiendo una agradable comida. Agradecer también a
la Dra. Ana María Kettlun por sus comentarios y apoyo personal, a la Dra. Lorena García
quién me formó en el laboratorio y siempre tenía tiempo para escuchar y a la profesora
Lucía Collados por su buena disposición y humor.
También me quiero referir a la gente que trabajó conmigo en el laboratorio, mi
querida amiga Carolina, con la cuál compartimos dos años de trabajo en el laboratorio y
otros cinco más de clases, pero más importante una linda amistad, Carlos que le dio otro
toque al laboratorio y con el que compartimos un muy buen paseo. Como olvidar a la
querida Jésica, que siempre tenía algo para regalonearme.
A los vecinos del laboratorio, Ares, Ale y Javier c, los profesores Daniela y Sergio.
No puedo dejar de nombrar a mis compañeros de curso, años inolvidables y buenos
amigos, Coty, Leidy, Monse, Coté, Miguel, Jose, Roger, Majóo , Christian, Víctor, Erick.
Mis amigos de toda la vida, Beto, Huevo, Poroto, Fabi, Pipe y Maka.
Agradezco también a mi familia por su apoyo incondicional, mi abueli quién
siempre tuvo fe ciega en mi. No puedo olvidar a quienes participaron directamente en esta
tesis siendo familiares míos: Cristóbal quien me enseño a ocupar photoshop (sólo para los
accesorios de las fotos, no se malentienda) y Constanza quién generosamente me ayudó a
diseñar el poster que presenté en Pinamar.

iv
Y por supuesto a las personas que saben que las llevo en el corazón, a todos ustedes
Muchas Gracias.

v
Financiamiento
Esta tesis fue realizada en el Laboratorio de Bioquímica y Biología Molecular de la facultad
de Ciencias Químicas y Farmacéuticas de la Universidad de Chile financiada por el
proyecto FONDECYT N° 1050784, titulado “El desarrollo de Paraparesia en los pacientes
infectados con HTLV-I se relaciona con alteración de los mecanismos de
fosforilación/desfosforilación en proteínas del citoesqueleto de los axones largos que
influyen en el transporte axónico” y para los estudios por Espectrometría de Masas por el
proyecto Mecesup UCH0115, titulado “Red nacional de programas de doctorado en
biociencias moleculares: una articulación multidisciplinaria, interregional, horizontal y
dinámica”.

vi
Indice General
Agradecimientos ..................................................................................................................iii Financiamiento ..................................................................................................................... v Indice General......................................................................................................................vi Indice de Figuras y Tablas ................................................................................................viii Abreviaturas ........................................................................................................................ ix Resumen ................................................................................................................................ x Summary ............................................................................................................................xiii Introducción .......................................................................................................................... 1
Proteína Tau........................................................................................................................ 3 Quinasas y fosfatasas que actúan en Tau y sus residuos específicos ................................. 6 Estudios en líquido cefalorraquídeo ................................................................................. 10 HIPOTESIS ...................................................................................................................... 12 Objetivo general ............................................................................................................... 12 Objetivos específicos........................................................................................................ 13
Determinar Tau y sus formas fosforiladas en LCR ...................................................... 13 Aislar proteína Tau para estudios de Espectrometría de Masas ................................... 13
Materiales y Métodos. ....................................................................................................... 14 Reactivos. ......................................................................................................................... 14 Muestras biológicas. ......................................................................................................... 15
Diagnóstico de la infección por HTLV-I en los pacientes. .......................................... 16 Determinación de proteínas .............................................................................................. 16 Inmunoprecipitación......................................................................................................... 16
Inmunoprecipitación mediante agarosa unida a proteína A/G ..................................... 17 Inmunoprecipitación mediante “Sepharose” activada unida covalentemente al anticuerpo anti-tau policlonal. ...................................................................................... 17
Aislamiento de fosfopéptidos mediante su interacción con Ga(III) ................................. 18 Determinación de fosfoserina, fosfotreonina, tau y residuos fosforilados específicos de tau mediante “immunowestern blot”. ............................................................................... 19 Deshibridización de las membranas de nitrocelulosa....................................................... 20 Espectrometría de Masas. ................................................................................................. 20
Preparación de las muestras.......................................................................................... 20 Análisis por MALDI-TOF............................................................................................ 22
Resultados ........................................................................................................................... 24 Análisis en Líquido Cefalorraquídeo................................................................................ 24
Determinación del contenido de albúmina sérica en el LCR. ...................................... 24 Inmunodetección de Tau en LCR................................................................................. 26 Análisis de Tau en muestras de LCR de controles y pacientes. ................................... 27 Presencia de fosfoserina y fosfotreonina en la proteína Tau de LCR .......................... 29 Análisis de residuos específicos fosforilados de Tau en LCR...................................... 30
Aislamiento de Tau desde el LCR.................................................................................... 34 Aislamiento de Tau mediante cromatografía de afinidad con Ga(III).......................... 34 Aislamiento de Tau mediante cromatografía de inmunoafinidad ................................ 36
Inmunoprecipitación de Tau mediante Proteína A/G-Agarosa. ............................... 36

vii
Presencia de fosfoserina y fosfotreonina en la proteína Tau de inmunoprecipitados con Proteína A/G-Agarosa................................................... 38 Análisis de residuos fosforilados específicos en inmunoprecipitados con Proteína A/G-Agarosa......................................................................................................... 38
Inmunoprecipitación de Tau mediante “Sepharose ” activada con CNBr................ 40 Análisis mediante Espectrometría de Masas. ................................................................... 42
Discusión.............................................................................................................................. 44 Análisis en Líquido Cefalorraquídeo................................................................................ 44 Aislamiento de proteína Tau desde el LCR...................................................................... 47 Análisis de muestras inmunoprecipitadas por Espectrometría de Masas ......................... 49
Conclusiones........................................................................................................................ 51 Referencias .......................................................................................................................... 52

viii
Indice de Figuras y Tablas
Figura 1 Esquema vía córticoespinal y su relación con Retrotracción Axonal. ..................... 2 Figura 2 Distribución de Tau en microtúbulos....................................................................... 4 Figura 3 Residuos fosforilables en “Taupatías” y quinasas asociadas. .................................. 9 Figura 4 Presencia de HSA en LCR. .................................................................................... 25 Figura 5 Búsqueda de condiciones para detección de Tau en LCR. .................................... 26 Figura 6 Detección de Tau en muestras de LCR. ................................................................. 27 Figura 7 Distribución de los niveles de Tau en los grupos................................................... 28 Figura 8 Inmunoreactividad de LCR con anticuerpos que reconocen residuos fosforilados ............................................................................................................ 29 Figura 9 Detección de nivel de fosforilación en residuos específicos.................................. 31 Figura 10 Niveles de distintos residuos fosforilados en proteína Tau.................................. 32 Figura 11 Detección de Tau y treonina fosforilada en columna
cromatográfica de Galio(III)................................................................................ 35 Figura 12 Interferencia del anticuerpo primario presente en inmunoprecipitado con A/G
agarosa y su no detección al revelar con un anticuerpo policlonal ..................... 37 Figura 13 Bandas inmunoreactivas contra anticuerpos policlonales contra Tau utilizando
cantidades variables de inmunoprecipitados. ...................................................... 37 Figura 14 Detección de Tau y residuos serina y treonina fosforilados en
inmunoprecipitados con Proteína A/G-agarosa................................................... 39 Figura 15 Detección de residuos específicos fosforilados de Tau en
inmunoprecipitados con Proteína A/G-agarosa................................................... 39 Figura 16 Detección proteica en los inmunoprecipitados con Proteína A/G-agarosa.......... 40 Figura 17 Presencia de Tau en IPP mediante “Sepharose” activada con CNBr................... 41 Figura 18 Tinción Proteica de IPP con “Sepharose” activada con CNBr. ........................... 41 Figura 19 Comparación de las Huellas peptídicas por MALDI-TOF de Tau
inmunoprecipitado con un estándar..................................................................... 43 Tabla 1 Resumen de los análisis estadísticos de los niveles de Tau fosforilado en
residuos específicos ............................................................................................. 33

ix
Abreviaturas
AD Enfermedad de Alzheimer ALS Esclerosis lateral amiotrófica AMPc Adenosinmonofosfato cíclico AP-1 Proteína Activadora 1 Cdk4, 5 y 6 Quinasa dependiente de Ciclina 4, 5 y 6 CK-1 Quinasa de Caseína 1 CNBr Bromocianógeno EC Electroforesis capilar ELISA Ensayo Inmunoabsorbente unido a enzima GSK3-α y -β Quinasa de la glicógeno sintasa 3-α y -β HSA Albúmina sérica humana HSP Paraplegia Espástica Hereditaria HTLV-I Virus linfotrópico humano-I IgG Inmunoglobulina G Ipp Inmunoprecipitado kDa KiloDalton LCR Líquido cefalorraquídeo m/z Relación masa/carga MALDI-TOF Desorción/Ionización mediante Láser asistido por Matriz-
Tiempo de Vuelo MAP Proteínas asociadas a Microtúbulo MAPK Proteína quinasa activada por mitógeno Min Minutos MS Espectrometría de Masas NF-κB Factor Nuclear kappa B PHF Filamentos Helicoidales Pareados PKA Proteína quinasa A PKC Proteína quinasa C PMSF Fluoruro de p-metilsulfonilo PP2A Fosfatasa 2 A SDS Dodecilsulfato de sodio SDS-PAGE Electroforesis en geles de poliacrilamida en presencia de SDS SEM Error estándar media Ser Serina SFR Factor de respuesta a Suero SNC Sistema nervioso central TBS-T Tampón Tris salino con Tween 20 T-CD4+ Linfocitos T-CD4+ Tre Treonina TSP/HAM Paraparesia espástica tropical o mielopatía asociada al virus
linfotrófico humano tipo I

x
Resumen
La enfermedad neurológica conocida como Paraparesia Espástica Tropical, mielopatía
asociada al HTLV-I (TSP/HAM) se caracteriza por la degeneración axonal de los haces
córtico-espinales. Aunque no se conoce mucho como el HTLV-I afecta selectivamente a los
axones más largos del sistema nervioso central (SNC), es la proteína viral Tax secretada
desde los linfocitos T-CD4+ el principal candidato por su actividad estimuladora de
factores transcripcionales a través de varias vías de señalización. Esta proteína, se encuentra
presente en forma crónica en el líquido cefalorraquídeo (LCR), pudiendo alterar el
transporte axonal actuando extracelularmente, siendo afectados principalmente los axones
más largos por su mayor vulnerabilidad.
Estudios in vitro de la proteína Tax muestran que se une a varias proteínas celulares
que regulan vías relacionadas con la transcripción y el citoesqueleto, incluyendo fosfatasas
y quinasas, por lo que se sugiere que en esta patogenia podría haber un desbalance en el
nivel de fosforilación/desfosforilación de las proteínas del citoesqueleto.
Se han propuesto como biomarcadores de varias enfermedades neurodegenerativas los
niveles de Tau total y/o fosfo-Tau en el LCR. Por lo tanto, los objetivos de esta
investigación fueron comparar la fosforilación de Tau presente en el LCR de pacientes con
TSP/HAM y sujetos controles, a fin de aclarar si estas proteínas están involucradas en la
patogenia de esta enfermedad, y aislar la proteína Tau desde el LCR para posteriores
estudios de proteómica de sus fosfoformas.
Los niveles de Tau en el LCR y la evaluación del patrón molecular de fosforilación de
residuos específicos como Tre181, Ser199, Tre205, Tre231, Ser262, Ser356, Ser396,
Ser404 y Ser422 fue realizada por “immunowestern blot” utilizando anticuerpos

xi
policlonales monoespecíficos. Con todos los anticuerpos ensayados se observó la presencia
de una banda principal de 52 kDa.
Los niveles de Tau en el LCR de los TSP/HAM no fueron significativamente distintos
de los de controles, y aunque no fue significativamente diferente (p = 0,06), se podría
sugerir sólo una anormal hiperfosforilación de Tre181. Si se confirmara este mayor nivel
fosforilación en Tre181 trabajando con un número mayor de pacientes o utilizando otros
métodos más confiables, sugeriría un aumento en la actividad o una sobre-expresión de dos
quinasas dirigidas a Ser/Tre-Pro, GSK3-α y β, Cdk5.
Una caracterización completa de la fosforilación de los distintos residuos de Tau en el
LCR de los controles y pacientes con TSP/HAM requirió la aplicación adicional de la
técnica analítica de Espectrometría de Masas (MS). Por lo tanto, se trató de aislar Tau
desde LCR utilizando una columna de afinidad por fosfopéptidos de Ga(III), y dos métodos
de inmunoprecipitación, uno usando proteínas A/G acopladas a agarosa y una segunda con
unión covalente de los anticuerpos (“Sepharose” activada con CNBr). Ni la columna de
Ga(III) ni el primer método de inmunoprecipitación fueron adecuados para posterior
análisis por MS. Después de demostrar por SDS/PAGE que el método que usaba los
anticuerpos unidos covalentemente mostraba una banda principal de 52 kDa, se eluyó la
proteína del gel, tripsinizó y analizó por MS MALDI-TOF. Se observó la posible presencia
de proteínas asociadas al citoesqueleto que podrían haber coinmunoprecipitados con Tau.
Estos resultados no fueron confirmatorios por el bajo “score” observado.
En conclusión, dos de los resultados muestran diferencias entre TSP/HAM y varias
enfermedades neurodegenerativas: el no aumento en los niveles de Tau total, y la posible

xii
hiperfosforilación de sólo Tre181. Fue importante lograr aislar Tau desde el LCR para
futuros estudios de MS.

xiii
Summary
Post-translational modifications analysis of Tau Protein of
Cerebrospinal fluid in Patients with TSP/HAM.
The neurological disorder known as tropical spastic paraparesis / HTLV-I-associated
myelopathy (TSP/HAM) is characterized by axonal degeneration of the cortico-spinal
tracts. Although the selectivity of HTLV-I towards the longest axons of the central nervous
system (CNS) is not fully understood, the main candidate is the secreted viral protein Tax
from lymphocytes T-CD4+ that shows a stimulatory activity of transcriptional factors
acting through several signaling pathways is the main candidate. This protein, chronically
present in CSF, could extracellularly alter axonal transport. The longest axons are mainly
affected because they are more vulnerable.
In vitro studies on Tax protein have shown the binding of this protein to many cellular
proteins that regulate transcription and cytoskeleton related pathways including
phosphatases and kinases, therefore an imbalance in the level of cytoskeleton
phosphorylated/unphosphorylated proteins could be involved in this pathogeny.
Levels of total Tau and/or phospho-Tau in the CSF have been proposed as biomarkers
of various neurodegenerative diseases. Hence, the aims of this investigation were to
compare Tau phosphorylation present in CSF of TSP/HAM patients and control subjects, to
elucidate if these proteins are involved in the pathogeny of this disease, and to isolate Tau
protein from CSF for further proteomic studies of its phosphoforms.
Levels of Tau in CSF and evaluation of the molecular pattern of phosphorylation of
selected residues such as Thr181, Ser199, Thr205, Thr231, Ser262, Ser356, Ser396, Ser404

xiv
and Ser422 were performed by immunowestern blot, using monospecific polyclonal
antibodies. With all the antibodies tested we observed a main band with 52 kDa. The CSF
levels of Tau in TSP/HAM were not significantly different from those of controls. Only an
abnormal hyperphosphorylation on Thr181, although it was not significantly different (p =
0.06), could be suggested in TSP/HAM patients. If a high phosphorylation level in Thr181
is confirmed by working with a larger number of patients or other more reliable methods, it
would suggest an increase in activity or over-expression of two Ser/Thr-pro-directed
kinases GSK3-α and β, and Cdk5.
A complete characterization of Tau phosphorylation sites in CSF from control and
TSP/HAM patients required the use of the additional analytical technique of Mass
Spectrometry (MS). Therefore, we tried to isolate Tau from CSF samples using the
phosphopeptide affinity column with Ga(III), and two immunoprecipitation methods, one
using protein A/G-coupled agarose beads and a second one with covalently bound
antibodies (Sepharose activated with CNBr). Neither the Ga(III) column nor the first
immunoprecipitation method were adequated for MS analysis. After demonstrating by
SDS/PAGE that the method that used covalently bound antibodies showed a main band of
52 kDa, the sample was eluted, trypsinized and analyzed in a Mass Spectrometer MALDI-
TOF. This preliminary study suggested the presence of Tau and of some other cytoskeleton
associated proteins that could have been coimmunoprecipited with Tau. The low “score”
obtained did not allow reach confirmatory results.
In conclusions, two of the results showed differences with various neurodegenerative
diseases: lack of changes in Tau levels in CSF, and possible hyperphosphorylation of only

xv
threonine 181. It was important to have been able to isolate a Tau form from CSF for
further studies of MS.

1
Introducción
La Paraparesia Espástica Tropical o Mielopatía asociada al virus linfotrófico
humano, TSP/HAM de sus siglas en inglés, es una enfermedad neurológica asociada
etiológicamente a la infección del retrovirus HTLV-I (virus linfotrópico humano de las
células T tipo I) (Gessain y cols, 1985; Osame y cols, 1985), que afecta crónica y
progresivamente al Sistema Nervioso Central (SNC). Se caracteriza por debilidad crónica
progresiva de las extremidades inferiores, hiperreflexia generalizada, vejiga neurogénica,
espasticidad variable, signo de Babinski, con leve o ninguna pérdida sensorial (García-
Vallejo y cols, 2005; Cartier y Ramírez, 2005). Los estudios neuropatológicos muestran
pérdida de mielina y degeneración del haz córticoespinal configurando el fenómeno
conocido como retrotracción axonal, en los axones más largos de esta vía (Cartier y cols,
1997), lo cuál se aprecia en la Figura 1. Este fenómeno se ve respaldado por el
compromiso específico y ascendente de la vía además del hallazgo de dilataciones axonales
con neurofibrillas empaquetadas, cambios mitocondriales y presencia de amiloide en los
axones (Liberski y cols, 1994, 1999; Umehara y cols, 2000). Chile tiene una de las más
altas prevalencias de la infección, siendo cercana al 0,3%, que la hace bastante importante,
junto a otras enfermedades más conocidas como lo son el Chagas 0,6%; VIH 0,075%;
hepatitis B 0,05%; hepatitis C 0,25% (Vásquez 2003). El serotipo nacional corresponde a
la cepa Cosmopolitan y en el subgrupo Transcontinental (Ramírez y cols, 2002).

2
A) B)
Figura 1: Esquema vía córticoespinal y su relación con Retrotracción Axonal. (A) Se aprecia el haz córticoespinal desde su nacimiento en la Corteza Cerebral hasta dónde realiza sinapsis en la Médula Espinal. Se destaca el cruce de las fibras a nivel del Bulbo. Modificado de Bhatnagar, Neuroscience. (B) Se esquematiza el resultado de la retrotracción que involucra daño en la regeneración axonal, produciéndose liberación de componentes citoplasmáticos y restos de mielina. Modificado de Strittmatter, 2002.

3
El ingreso del virus al organismo es a través de vía sanguínea, leche materna o
semen, infectando linfocitos T CD4+ (Igakura y cols, 2003). Se necesita el contacto celular
de un linfocito a otro, para la infección, lo que se logra por la formación de un sincicio, en
dónde las partículas virales son traspasadas gracias a la modificación y polarización del
citoesqueleto de la célula huésped, ya que la infección no se propaga por viriones libres
(Igakura y cols, 2003). Esto indica la capacidad de modificación de la estructura celular
dada por la infección. El genoma viral codifica para varias proteínas que regulan la
transcripción viral y la de la célula huésped, dentro de las más importantes se encuentra la
proteína Tax que es capaz de regular la transducción de señales, ciclo celular y apoptosis
(Arima 2001; Iwanaga 2001; Wu y cols, 2004). Cuatro son las vías de transducción
principalmente afectadas por Tax: CREB/ATF, NF-κB, AP-1 y SFR (Li y Gaynor, 1999;
Jeang 2001; Yoshida 2001; Jeang y cols, 2004). Al inmunoprecipitar Tax desde cultivos
celulares de linfocitos se demostró que tiene la capacidad de unirse a diversas quinasas
dependientes de ciclina entre las cuáles están Cdk4, Cdk6 y PKC, teniendo la capacidad de
activarlas (Haller y cols, 2002). Por otra parte, al interactuar con subunidades catalíticas de
la fosfatasa 2A (PP2A) provoca la inactivación de ésta (Fu y cols, 2003). Esta acción
sinérgica, que afecta el estado de fosforilación/desfosforilación explicaría la capacidad de
activar factores transcripcionales (Prajapati y Gaynor, 2002).
Proteína Tau
La proteína Tau es la principal proteína asociada a los microtúbulos (MAP),
inicialmente aislada como co-producto de purificación de la tubulina (Billingsley y cols,
1997). En el ser humano proviene de un sólo gen que se encuentra en el brazo largo del

4
cromosoma 17 (posición 17q21). Contiene 16 exones que mediante “splicing” alternativo
produce 6 isoformas de Tau en el cerebro adulto, generando proteínas cuyas masas
moleculares varían entre los 45 y 65 kDa (Buée y cols, 2000; Gong y cols, 2005). Es
posible encontrar éstas y más isoformas en otras regiones (distribución espacial) y estadíos
del desarrollo neuronal (distribución temporal) (Avila, 2004; Billingsley, 1997). Su
principal función es estimular el ensamblaje de los microtúbulos y estabilizar su estructura.
Tau posee una región repetida de unión a microtúbulos en la región Carboxilo Terminal
proveniente de los exones 9 al 12, la cuál a su vez se ha dividido en dos regiones, la
verdadera región de unión a microtúbulos y la región acídica Carboxilo Terminal. La
porción Amino Terminal se proyecta sobre la superficie de los microtúbulos, en dónde
interacciona con otros elementos del citoesqueleto y de la membrana plasmática. A su vez
ésta se ha subdividido en dos regiones siendo una la zona con una alta proporción de
residuos ácidos y la otra región rica en prolinas (Avila y cols, 2004).
Es una proteína que se encuentra principalmente en el axón, aunque como se dijo
anteriormente es posible encontrarla en otras regiones de la neurona, e incluso en algunos
casos patológicos se ha descrito en diferentes tipos de células gliales (Avila y cols, 2004).
Es una proteína hidrofílica altamente asimétrica, la cuál ha logrado ser vista por
microscopía electrónica (Figura 2).
Figura 2: Distribución de Tau en microtúbulos. La figura muestra con flechas la ubicación visible de la proteína Tau, de origen porcino, que esta unida en forma saturante a los microtúbulos. Imagen obtenida por microscopía electrónica. Extraído de Hirokawa y cols, 1988.

5
Se han realizado investigaciones que relacionan a Tau con el transporte mismo en la
neurona. En estos estudios la sobrexpresión de Tau inhibe el transporte de peroxisomas,
mitocondrias y vesículas derivadas de Golgi, dependiente de quinesina, hacia los axones
(Stamer y cols, 2002). Además, influye en las velocidades de unión y liberación de las
proteínas desde los microtúbulos, haciendo que el movimiento retrógrado mediado por
dineína predomine en el transporte de organelos y vesículas (Trinczek y cols, 1999). De
esta manera se genera un retardo en el transporte y alteración en el aporte de material y
energía hacia el cono de crecimiento. De forma contraria, si los niveles de Tau se
disminuyen considerablemente, usando oligonucleótidos antisentido, se logran ver
alteraciones a nivel del crecimiento de neuritas (Avila y cols, 2004).
A pesar de que en el cerebro adulto se han descrito 6 isoformas de Tau, es posible
encontrar un bandeo que no corresponde exactamente con los valores de masa molecular
obtenidos por la predicción aminoacídica de sus genes (Billingsley, 1997). Esta no
correspondencia viene dada por la gran cantidad de modificaciones post-traduccionales que
puede sufrir la proteína entre las cuáles es posible incluir fosforilación, glicosilación,
ubiquitinación, desamidación, oxidación y nitración siendo la más estudiada la fosforilación
(Avila y cols, 2004; Gong y cols, 2005).
Tau es una proteína que se encuentra extensamente fosforilada en dónde la mayoría
de los sitios corresponden a motivos de serina y treonina vecinos a prolina (Tseng y cols,
1999) Existen 79 sitios putativos de fosforilación en residuos de serina y treonina en la
isoforma de Tau de mayor masa molecular encontrada en el SNC, la cuál posee 441
residuos, y que de ahora en adelante dará la pauta en la numeración de los residuos mismos.
Es bien sabido que la fosforilación de Tau es la principal forma en la cual se regula su
actividad de unión a microtúbulos e incluso su unión a membranas. Los niveles normales

6
de fosforilación permiten una óptima funcionalidad de Tau, y si es que esta se encuentra
hiperfosforilada pierde su actividad biológica, siendo capaz de formar agregados en
filamentos en cerebros de pacientes con enfermedad de Alzheimer (AD) u otras
“Taupatías” (Evans y cols, 2000; Hasegawa, 2006; Iwatsubo, 2006). Se da el nombre de
“Taupatías” a las enfermedades en dónde se encuentran niveles alterados en la fosforilación
de la proteína entre las cuales se encuentran Esclerosis Lateral Amiotrófica (ALS),
síndrome de Down, demencia frontotemporal con Parkinsonismo, degeneración
corticobasal, entre otras (Buée y cols, 2000; Avila y cols, 2004). Es entonces importante
conocer cuáles son las quinasas o fosfatasas que actúan sobre Tau, para producir las
fosforilaciones en los residuos específicos, que a su vez producen efectos distintos sobre el
ensamblaje de los microtúbulos. Esto tanto en condiciones normales como patológicas.
Quinasas y fosfatasas que actúan en Tau y sus residuos específicos
Es sabido que la fosforilación en residuos específicos de Tau es un mecanismo que
regula la formación y estabilización de los microtúbulos tanto in vitro como in vivo. Existen
dos grandes grupos de quinasas: las dirigidas y las no dirigidas por prolina (Evans y cols
2000; Tsujio, 2000; Sengupta y cols, 1998).
Quinasas dirigidas por prolina: Dentro de este grupo podemos encontrar
principalmente a la quinasa que fosforila la glicógeno sintasa -3 tanto α como β (GSK-3α,
GSK-3β) y la Cdk-5.
A) GSK-3α es capaz de fosforilar a Tau en los residuos serina 199, 202, 235 y 396,
y que bajo estimulación con Heparina se puede fosforilar los sitios treonina 212, 231, y
serina 324 y 356 (Billingsley, 1997).

7
B) GSK-3β es una de las quinasas más descrita que fosforilan a Tau. Su actividad
influye de manera importante en la capacidad de Tau de promover el ensamblaje de los
microtúbulos, pero no se afecta significativamente la afinidad de Tau por los microtúbulos
(Godemann y cols, 1999). Estas observaciones sugieren que existen sitios ubicados fuera de
la región de unión a microtúbulos que son capaces de regular el ensamblaje (Utton y cols,
1997). Apoyándose por la información que indica que los sitios de fosforilación afectados
en Tau por la GSK-3β corresponden a las serinas 199, 202, 235, 396, y 404 pero no así la
serina 262, es decir, estaría afectando sólo los sitios que se encuentran fuera de la región de
unión a microtúbulos (Godemann y cols, 1999, Gong y cols, 2005). Esta enzima es inhibida
usando litio en el medio de ensayo y en su presencia se ve una disminución de la
fosforilación en los residuos treonina 181 y serina 396 y 404 (Xie y cols, 1998) Además
ésta enzima posee la capacidad de actuar a nivel del transporte celular siendo importante en
el movimiento anterógrado de organelos (Tatebayashi y cols, 2004).
C) Cdk-5 está muy asociada con la regulación del ciclo celular, estando
especialmente relacionada con citoesqueleto y sistemas de membrana, su función principal
se ha descrito en el sistema nervioso (Smith y Tsai 2002). Se sabe que juega un papel
importante en la patogenia de enfermedades como el Alzheimer y ALS, entre otras (Zheng
y cols 2005). Contribuye a la fosforilación de la proteína Tau humana en los residuos serina
202, 235, 404 y treonina 181 y 205 (Wada y cols, 1998; Alvarez y cols, 1999; Gong y cols,
2005).
Cdk5 se relaciona a estados patológicos ya que su inhibición previene la muerte
neuronal a causada por β-amiloide en modelos celulares de AD, afectando señales que se
relacionan directamente con el crecimiento axonal (Maccioni y cols, 2001; Stamer y cols,

8
2002; Kesavapany y cols, 2004). Por esto se sugiere que Cdk5 podría ser regulador de las
proteínas asociadas a microtúbulos y por ende de la dinámica del citoesqueleto (Pigino y
cols, 1997).
Quinasas no dirigidas por prolina: en este grupo se encuentra principalmente la
proteína quinasa A (PKA) la cuál fosforila a Tau principalmente en el residuo serina 214, y
en menor medida en serina 262, 324, 356, 409 y 416. Estas fosforilaciones in vitro
disminuyen la capacidad de promover la nucleación y el crecimiento de los microtúbulos.
In vivo sólo se aprecia que la fosforilación en serina 262 y 356 son reguladas por esta
enzima (Xie y cols, 1998). Es de considerable interés el residuo serina 262 ya que se
encuentra en la zona de unión a microtúbulos y esta presente hiperfosforilado en los
filamentos helicoidales pareados (PHF), una estructura ampliamente descrita en AD
(Billingsley y cols, 1997).
En la búsqueda de los residuos cuya fosforilación participaría activamente en la
regulación de la actividad de Tau, se han logrado varios avances. Al probar variadas
combinaciones de quinasas se ha determinado cuáles son los residuos clave en la unión a
microtúbulos y cuáles a pesar de no modificar la unión si son capaces de disminuir el
ensamblaje de éstos. Es así como se sabe que tanto la treonina 231 como la serina 262 son
necesarias para una máxima inhibición de la capacidad de Tau de ensamblar a los
microtúbulos, y en menor medida las serinas 214, 396 y 404 (Seubert y cols, 2005;
Sengupta y cols, 1998, Schneider y cols, 1999). Además, se han relacionado epítopes con
algunos estados patológicos como lo es la serina 422 que se encuentra principalmente
fosforilada en muchas de las “Taupatías” (Bussière y cols, 1999).

9
En la Figura 3A se representan la posición de los residuos fosforilables en la proteína
Tau. Las quinasas descritas que fosforilan a los distintos residuos se muestran en la Figura
3B.
A)
B)
Figura 3: Residuos fosforilables en “Taupatías” y quinasas asociadas. (A) Disposición espacial de los residuos en la proteína, sale indicado los dominios de unión a microtúbulos, además de los residuos comercialmente disponibles para análisis de fosforilación. (Modificado de catálogo Biosource International, Camarillo, CA, USA) (B) Para cada sitio viene indicado cuáles quinasas los fosforilan. (Modificado de Gong y cols, 2005)

10
En el balance de los procesos de fosforilación /desfosforilación es importante
considerar las acción de las fosfatasas. La principal fosfatasa que actúa a nivel de Tau es la
PP2A la cuál posiblemente en los estados patológicos se encuentra inhibida, generando la
hiperfosforilación y posterior acumulación de Tau (Gong y cols, 2000; Xie y cols, 1998;
Avila y cols, 2004; Gong y cols, 2005)
Estudios en líquido cefalorraquídeo
Es importante buscar una fuente de información fidedigna para poder comprender la
patogenia de la enfermedad. Dado que las proteínas son responsables de la mayoría de los
procesos en un organismo, cambios en su concentración o en su estructura (incluyendo
éstas modificaciones post-traduccionales) podrían reflejar los efectos de la enfermedad,
haciendo de éstas un blanco atractivo en la investigación. Una alternativa es el uso de
fluidos corporales, ya que son de fácil acceso (comparado con una biopsia) y son una fuente
de información rica en proteínas y que posee componentes de todo lo que se encuentra en
su recorrido a través de la circulación por el cuerpo. El análisis del líquido cefalorraquídeo
(LCR) es el más conveniente método para estudiar la biología de las enfermedades
neurodegenerativas, ya que refleja las proteínas del SNC bajo condiciones normales como
también en muchas de éstas enfermedades. Siendo de gran valor en etapas tempranas de las
enfermedades, cuando los síntomas clínicos son vagos y el diagnóstico se dificulta.
Estudios en el LCR son hasta ahora una de las principales herramientas de diagnóstico para
una gran variedad de condiciones que afectan tanto el sistema nervioso central como
periférico (Davidsson y Sjögren, 2005).

11
En el LCR de pacientes con TSP/HAM se ha descrito la presencia de la proteína
viral Tax. (Cartier y cols, 2005). Esta proteína Tax generada por los linfocitos infectados es
capaz de afectar vías transduccionales que se pueden cruzar con las principales quinasas y
fosfatasas que actúan sobre Tau (Haller y cols, 2002; Fu y cols, 2003). Con estos
antecedentes es nuestro interés el poder relacionar a futuro las fosforilaciones presentes en
la proteína Tau con las vías específicas afectadas por la infección del virus en TSP/HAM.
Esto podría dar cuenta de una alteración del citoesqueleto neuronal modificando el
transporte axonal llevando finalmente a una retrotracción axonal de las neuronas del haz
córticoespinal. En el LCR se ha descrito la presencia de la proteína Tau fosforilada y no
fosforilada lo que permitiría estudiar en nuestros pacientes la alteración en su nivel de
fosforilación y relacionar la proteína Tau total con la Tau fosforilada en residuos
específicos (Davidsson y Sjögren 2005). También es importante aislar las distintas formas
de Tau para poder analizarlas mediante Espectrometría de Masas, técnica de preferencia
para estudios de proteómica (Reynolds y cols, 2000; Davidsson y Sjögren 2005;
D’Ambrosio y cols, 2006).
Nuestro grupo propone que en TSP/HAM hay alteraciones en las fosforilaciones en
los residuos de serina y treonina de Tau lo que podría dar cuenta de un daño axonal por
algún efecto de proteínas virales secretadas como Tax que es capaz de activar quinasas e
inhibir fosfatasas (Haller y cols, 2002; Fu y cols, 2003).
La propuesta de un daño a nivel de transporte axonal se ve además respaldada por
los antecedentes de la Paraparesia Espástica Hereditaria (HSP) con características clínicas e
histológicas idénticas a la producida por el virus en los individuos que desarrollan la
paraparesia (Evans y cols, 2005). En el caso de HSP, una de las principales causas es la
mutación del gen que expresa la Espastina, proteína clave en la dinámica de los

12
microtúbulos, sugiriéndose que su mal funcionamiento se relacionaría con la degeneración
axonal, encontrándose además alteración en el transporte axonal de proteínas y organelos
(Ferreirinha y cols, 2004; Evans y cols, 2005).
El estudiar en forma más específica las modificaciones post-traduccionales de Tau
permitiría posteriormente relacionar estos cambios con los niveles o actividades quinásicas,
permitiendo comprender mejor la acción viral sobre el SNC.
HIPOTESIS
En base a estos antecedentes se plantea la siguiente Hipótesis:
La retrotracción y pérdida axonal que se observa en los haces córtico-espinales de pacientes
infectados con HTLV-I que desarrollan paraparesia espástica, se relaciona con un aumento
en la fosforilación de proteína Tau del citoesqueleto axonal, lo que podría originar una
disminución del transporte axonal y una inhibición de la regeneración axonal.
Objetivo general
Determinar si en pacientes con TSP/HAM hay variaciones en el estado de fosforilación de
la proteína Tau deducido de su presencia en líquido cefalorraquídeo.

13
Objetivos específicos
Determinar Tau y sus formas fosforiladas en LCR
1.1.- Buscar las condiciones para determinar los niveles de Tau en LCR mediante
inmunoanálisis.
1.2.- Comparar los niveles de Tau en muestras de pacientes y controles.
1.3.- Determinar la colocalización de Tau con la presencia de residuos de serina y treonina
fosforilados por “immunowestern blot”.
1.4.- Determinar la presencia de fosforilaciones en residuos específicos de serina y treonina
por “immunowestern blot” en las muestras de LCR.
1.5.- Comparar los niveles de los diversos residuos fosforilados en serina y treonina en
muestras de LCR de TSP/HAM y controles a fin de asociarlo al aumento de algunas
actividades quinásicas.
Aislar proteína Tau para estudios de Espectrometría de Masas
2.1.- Aislar de LCR de pacientes y controles la proteína Tau para comparar el grado de
inmunoreactividad con anticuerpos anti-fosfoserina, anti-fosfotreonina y residuos
fosforilados específicos de Tau.
2.2.- Realizar “immunowestern blot” con las muestras inmunoprecipitadas para su posterior
análisis por Espectrometría de Masas.

14
Materiales y Métodos.
Reactivos.
AMERSHAM BIOSCIENCES, Uppsala, Sweden CNBr-Activated Sepharose
BIORAD, Hércules, CA, USA Membrana de Nitrocelulosa BIOSOURCE International, Camarillo, CA, USA Anticuerpo monoclonal para TAU (AHB0042) Anticuerpo policlonal para TAU pT181 (44-732Z) Anticuerpo policlonal para TAU pS199 (44-734ZG) Anticuerpo policlonal para TAU pT205 (44-738ZG) Anticuerpo policlonal para TAU pT231 (44-746ZG) Anticuerpo policlonal para TAU pS262 (44-750ZG) Anticuerpo policlonal para TAU pS356 (44-751Z) Anticuerpo policlonal para TAU pS396 (44-752ZG) Anticuerpo policlonal para TAU pS404 (44-758ZG) Anticuerpo policlonal para TAU pS422 (44-764ZG) CHEMICON Internacional, Temecula, CA, USA Anticuerpo policlonal para Fosfoserina (AB1603) Anticuerpo policlonal para Fosfotreonina (AB1607) Solución para deshibridización, Re-Blot Plus Mild (2500-10 ml) “Quick blocker” EASTMAN KODAK Company, Rochester, New York, USA. Película BIOMAX MR. MERCK, Darmstadt, Alemania Metanol Acido acético Etanol PIERCE, IL, USA. 2° anticuerpo anti-conejo conjugado con peroxidasa. 2° anticuerpo anti-ratón conjugado con peroxidasa. Super Signal West Femto Chemiluminescent Substrate. CLXposure film XR-B “Phosphopeptide Isolation Kit”

15
SANTA CRUZ BIOTECHNOLOGY, CA, USA. Protein A/G PLUS agarose
SIGMA CHEMICAL Co, S Louis, Mo, USA. Trizma base NaCl Glicina Acrilamida Bisacrilamida Persulfato de amonio Tween 20 Azul de Coomassie R Azul de Coomassie G250 US BIOLOGICAL, Swampscott, MA, USA Anticuerpo policlonal para TAU (T1032) Muestras biológicas.
Las muestras de LCR fueron obtenidas en el Hospital del Salvador mediante
punción lumbar atraumática de pacientes con TSP/HAM como parte de procedimientos de
diagnóstico normales. Los dos grupos compuestos de 6 pacientes y 6 controles fueron
debidamente informados sobre el protocolo de investigación aprobado por el Comité de
ética del Hospital del Salvador y todos aceptaron libremente participar en el estudio. El
grupo control correspondió a muestras de maternidad o bien pacientes que requirieron
punción lumbar como parte del diagnóstico, sin alteraciones neurológicas de tipo
inflamatorio o infeccioso. El rango etario del grupo control comprendió desde los 45 hasta
los 67 años y el del grupo TSP/HAM desde los 47 a los 65 años. El LCR se mantuvo
congelado a -20°C hasta el análisis.

16
Diagnóstico de la infección por HTLV-I en los pacientes.
En el laboratorio del Doctor Eugenio Ramírez en el Instituto de Salud Pública, se
determinó la presencia de anticuerpos contra HTLV-I (Gallo y cols, 1991; Galeno y cols,
1994).
Determinación de proteínas
Este análisis se realizó utilizando un sistema de Electroforesis Capilar Ion Analyzer
(Waters Quanta 4000) con software Millenium (Waters) para el manejo de la información.
Para llevar a cabo esta técnica se utilizó un capilar de sílica fundida (68 cm de distancia al
detector y 50 μm de diámetro interno), una solución de electrolito de corrida consistente en
H3PO4 150 mM, pH 1.5. La detección ultravioleta se llevó a cabo a 185 nm (longitud de
onda correspondiente al enlace peptídico) y la separación se realizó a 18 kV y 25 ºC con
inyecciones de muestra de 10 segundos. Entre cada muestra la columna capilar se lavó en
forma secuencial con inyecciones de 3 minutos cada una de NaOH 0,5 M, agua destilada y
solución de electrolito de corrida. El estándar utilizado fue Albúmina Sérica Humana
(HSA).
Inmunoprecipitación.
Se realizaron dos métodos de inmunoprecipitación distintos, para los cuáles la
metodología ocupada fue la siguiente:

17
Inmunoprecipitación mediante agarosa unida a proteína A/G
El procedimiento de inmunoprecipitación se realizó de la siguiente manera: a 50 μL
de LCR cuya concentración fuese de 0,4 mg/mL se le agregó 1 μL de inhibidores de
proteasas: aprotinina, leupeptina, pepstatina y PMSF quedando a una concentración final de
2 μg/mL, 20 μg/mL, 2 μg/mL, 1 μmol/mL respectivamente y 3 μL de anticuerpo
monoclonal anti tau, esta mezcla se dejó agitando en un rotoagitador con ángulo de 45º a
4ºC toda la noche. Al día siguiente a cada mezcla se le agregó 40 μL de Proteína A/G
Agarosa ambientada con tampón de inmunoprecipitación (Tris-HCl 50 mM pH a 7,5 con
NaCl 150 mM). Esto se realizó lavando 4 veces con 300 μL, centrifugando cada vez a 1000
x g microfuga (mikro22R, marca Hettich) por 1 min 4ºC. Esta nueva mezcla se mantuvo en
el rotoagitador 2,5 horas a temperatura ambiente. Posteriormente se guardó el sobrenadante
y la agarosa fue lavada 5 veces con 700 μL de tampón de inmunoprecipitación, el
precipitado se resuspendió en tampón 1x (Laemmli, 1970), luego se centrifugó a 1000 x g y
se usó el sobrenadante para análisis.
Inmunoprecipitación mediante “Sepharose” activada unida covalentemente al
anticuerpo anti-tau policlonal.
Este procedimiento se efectuó según instrucciones del fabricante. El primer paso
consistió en activar la “Sepharose” con HCl 1 mM, utilizándose 200 veces en volumen la
cantidad pesada (0,1 g con 20 mL), en varias alícuotas para lograr una correcta hidratación
(15 min) una vez lograda la hidratación se puede guardar a 4°C. Paralelamente se disolvió
el anticuerpo (5 μL) en 15 μL de solución de acoplamiento NaHCO3 0,1M a pH 8,3 con

18
NaCl 0,5M. Se tomó un alícuota de 25 μL de “Sepharose” activada y se ambientó,
rápidamente, 2 veces con la solución de acoplamiento para posteriormente mezclarla con la
solución en la que se encontraba el anticuerpo, dejándose en rotoagitador toda la noche. Al
día siguiente se sacó el sobrenadante y se enjuagó con solución de acoplamiento (2 veces
con 5 volúmenes) y se bloquearon los grupos reactivos remanentes con Tris-HCl 0,1M a
pH 8,0 durante 2 horas. Pasado este tiempo se lavó el medio con al menos 3 ciclos de
alternancia de pH (5 volúmenes), centrifugando a 1000 x g cada vez. El ciclo consistió en
acetato 0,1 M a pH 4,0 con NaCl 0,5 M y Tris-HCl 0,1 M a pH 8,0 con 0,5 M NaCl,
quedando lista la resina para la inmunoprecipitación. La “Sepharose” se mezcló con 60 μL
de LCR concentrado y se dejó en el rotoagitador toda la noche a 4°C, además se agregó el
mismo volumen de tampón de inmunoprecipitación (Tris-HCl 50 mM, NaCl 150 mM, pH
7,5) para que se mezclara con más facilidad la muestra con la matriz. Después de esta
incubación se centrifugó a 1000 x g, se eliminó el sobrenadante y la “Sepharose” se lavó
unas 5 veces con el tampón de inmunoprecipitación, luego de eso a la muestra se le agregó
60 μL de glicina 0,2 M a pH 2,5 con NaCl 150 mM. Se agitó por 15 min a T° ambiente y se
extrajo por centrifugación a 1000 x g y la “Sepharose” se conservó para una futura
separación de la proteína de interés.
Aislamiento de fosfopéptidos mediante su interacción con Ga(III)
El aislamiento de fosfopéptidos se llevó a cabo siguiendo las instrucciones del kit
comercial “Phosphopeptide Isolation Kit”, consistente en columnas “Mini-spin SwellGel®
Gallium-Chelated Disc”, con la capacidad de unir grupos fosfato, y específicamente 150 μg
de fosfopéptidos. La primera etapa involucra una unión fosfopéptido-SwellGel® a pH 2,0-

19
5,5 (5% ácido acético), seguida de dos lavados consecutivos a pH 2,0-3,5 (lavado 1: 0,1%
ácido acético; lavado 2: 0,1% ácido acético y 10% acetonitrilo, pH 3,0) para finalmente,
eluir los fosfopéptidos de la columna a un pH mayor que 6,0 usando 0,1M bicarbonato de
amonio, para la elución.
Determinación de fosfoserina, fosfotreonina, tau y residuos fosforilados
específicos de tau mediante “immunowestern blot”.
El “immunowestern blot” se efectuó según Towbin y cols (1979) empleando muestras
de 10 μl de LCR desnaturadas hirviendo 5 min con tampón 4x (Laemmli, 1970), y/o las
muestras inmunoprecipitadas ya tratadas con este tampón, éstas fueron separadas por
electroforesis en gel de poliacrilamida al 12%. Las proteínas fueron transferidas a
membrana de nitrocelulosa (BioRad, tamaño de poro 0,45 μm), la que posteriormente fue
bloqueada por una hora con “Quick Blocker” al 6%. El reconocimiento de fosfoserina,
fosfotreonina, tau y los residuos fosforilados específicos de ésta, se llevó a cabo ocupando,
por separado, anticuerpos tanto monoclonales como policlonales para cada epítope, diluidos
de la siguiente manera: tau monoclonal 1:3000, tau policlonal 1:1000 fosfoserina,
fosfotreonina y residuos específicos fosforilados de tau 1:5000 en TBS-T (Tris-HCl 20 mM
pH 7,6, NaCl 137 mM, Tween-20 al 0,1%), e incubando las membranas por toda la noche a
4°C. Posteriormente se lavó la membrana con tampón TBS-T (4 veces x 5 min). Se incubó
por 1 hora con anticuerpo policlonal anti-ratón o anti-conejo (según corresponda)
conjugado con peroxidasa diluido 1:10.000 en TBS-T con Tween al 0,1%. La reacción

20
positiva se identificó mediante quimioluminiscencia exponiendo la membrana a la placa
fotográfica (el tiempo fue determinado según la intensidad de la señal).
Deshibridización de las membranas de nitrocelulosa.
Para el estudio de la posible colocalización de la inmunoreactividad de la proteína
tau con fosfotreonina, fosfoserina y/o residuo específico fosforilado, se procedió a
deshibridizar cada membrana nitrocelulosa luego de haber sido revelada con el agente
quimioluminiscente. La deshibridización se realizó incubando, a temperatura ambiente con
agitación durante 15 min, la membrana de nitrocelulosa en 10 ml de solución de
deshibridización 1x (1 ml de “Re-Blot Plus 10x mild” más 9 ml de agua destilada). Luego
se lavó la membrana con TBS-T x 2 veces, y se bloqueó posteriormente por una hora con
“Quick Blocker” al 6%. Por último se incubaba la membrana con la dilución indicada del
anticuerpo correspondiente.
Espectrometría de Masas.
Este análisis se realizó en muestras de LCR inmunoprecipitadas con la técnica de
“Sepharose” activada con CNBr, las que se sometieron a electroforesis en condiciones
desnaturantes, posteriormente se eluyeron del gel, se digirieron con tripsina y se analizaron
en el espectrómetro MALDI-TOF (Bruker).
Preparación de las muestras
Las proteínas obtenidas por la inmunoprecipitación, una vez corridas en los geles de
poliacrilamida fueron teñidas con azul de Coomassie coloidal (G250) (metanol al 34%,

21
ácido fosfórico al 3% y sulfato de amonio 1,28 M) (Kang y cols, 2002). Las bandas fueron
seleccionadas y cortadas del gel para su posterior hidrólisis enzimática y análisis en el
espectrómetro de masas. La hidrólisis enzimática fue realizada con tripsina mediante el
protocolo descrito a continuación.
Una vez cortada la banda de proteína de interés del gel utilizando una hoja de bisturí,
cuidando eliminar al máximo la poliacrilamida (exceso de gel), se transfirió la banda a un
tubo Eppendorf de 0,5 mL. Para la eliminación del colorante unido a las proteínas se agregó
150 μL de NH4HCO3 200 mM preparado en acetonitrilo 50% v/v agitando brevemente, y se
mantuvo durante 10 min a 30ºC. El sobrenadante se eliminó y se repitió este paso al menos
dos veces (hasta que la banda del gel perdió su coloración azul), finalmente se centrifugó
para remover el exceso de solución de distinción.
Lograda la decoloración del gel se agregó 50 μL de ditiotreitol 10 mM preparado en
NH4HCO3 200 mM y se agitó brevemente antes de incubar a 37ºC durante 20 min. Luego
se removió el sobrenadante, y se centrifugó para quitar el exceso de solución.
Posteriormente se adicionó 50 μL de yodoacetamida 55 mM preparado en NH4HCO3,
agitándose brevemente e incubándose a 37ºC durante 20 min (en oscuridad), este paso se
realiza para lograr la modificación química los residuos cisteína mediante la alquilación del
residuo. El exceso de yodoacetamida fue extraído mediante centrifugación y a continuación
se agregó 100 μL de acetonitrilo 100% y se volvió a incubar durante 5 min a 37ºC. Se
removió el sobrenadante y se repitió este último paso dos veces, pero esta vez con 50 μL de
acetonitrilo 100%. Centrifugándose al final para poder extraer el exceso de solución. A
esta altura la banda se sigue viendo blanca opaca y disminuida de tamaño. Se dejó secar a
temperatura ambiente durante 10 min, y luego se colocó en hielo.

22
Para comenzar la tripsinización de la banda se agregó 8 μL de tampón de digestión
NH4HCO3 50 mM pH 7,8 que contiene 2 ng/μL de tripsina y se incubó en hielo durante 45
min, se agregó entre 20 y 40 μL de NH4HCO3 50 mM pH 7,8 de modo de cubrir la banda
recortada, y se dejó durante toda la noche (8-16 h) a 37ºC, cuidando de minimizar el
volumen agregado. Al día siguiente se recuperó el sobrenadante y se transfirió a un nuevo
tubo Eppendorf de 0,5 mL, se le agregó 20 μL de acetonitrilo al 60% v/v / ácido fórmico
0,1% v/v sobre la banda, se le aplicó vórtex durante 1-2 min y se centrifugó, luego se
incubó durante 10 min a 30ºC, volviéndose a recuperar el sobrenadante y mezclándose con
el obtenido de la digestión. Estos pasos se repiten nuevamente. Finalmente, los
sobrenadantes combinados fueron secados en SpeedVac a temperatura ambiente (no
exceder 30ºC) durante 2-4 h. Siendo resuspendidos en 10-20 μL de ácido fórmico 0,1% v/v,
y sonicados durante 5 min.
Análisis por MALDI-TOF
Para este análisis se utilizó una matriz de ácido α-ciano-4-hidróxicinámico (CHCA)
preparada en acetonitrilo/ácido fórmico 0,1 v/v 1:1 a una concentración de 10 mg/mL. Esta
matriz se usó para diluir la muestra. Se mezcló 1 μL de muestra con 1 μL de matriz y los 2
μL resultantes, se depositaron en una placa porta muestra micro-scout. Además, se realizó
una segunda dilución, mezclando 1 μL de muestra con 10 μL de matriz y se depositando 2
μL en la placa porta muestra.
Los espectros se obtuvieron en un equipo Microflex MALDI-TOF (Bruker
Daltonics, Inc., MA-USA). Previo a la generación de los espectros se realizó una
calibración del equipo con un estándar externo correspondiente a una mezcla de péptidos de

23
masas 1.000-3.000 Da. Para la obtención de espectros se trabajó en modo ion positivo y
detección en reflexión. La ionización se realizó mediante la utilización de pulsos de láser
ultravioleta para lo cual se utilizó un láser de nitrógeno de longitud de onda de 337 nm con
una energía de pulso de 150 μJ y frecuencia de 20 Hz. Y se trabajó bajo los siguientes
parámetros: voltaje de fuente de iones 1, 19,00 kV; voltaje de fuente de iones 2, 16,30 kV;
voltaje de lentes, 9,75 kV; voltaje de reflector, 20 kV y tiempo de extracción pulsada de
iones, 100 ns. Los espectros finales corresponden a espectros sumados correspondientes a
15 barridos de 30 disparos de láser aplicados en diferentes puntos tomados al azar. Para el
análisis de los espectros se utilizó el programa flexAnalysis (Bruker Daltonics), y se utilizó
el algoritmo SNAP (señal/ruido, 6) para la detección de la señales m/z. La identificación se
realizó utilizando tres bases de datos: Mascot, MS-Fit y ProFound. Para la supresión de
señales de la matriz se utilizó modo de deflexión de hasta 500 Da.
En estos análisis se contó con la colaboración del Dr. Andrés Barriga, encargado de
los Espectrómetros de Masas (Facultad de Ciencias Químicas y Farmacéuticas,
CEPEDEQ).

24
Resultados
Primer Objetivo: Determinar en LCR Tau y sus formas fosforiladas
Análisis en Líquido Cefalorraquídeo.
Para llevar a cabo este primer objetivo se efectuaron las siguientes determinaciones en
las muestras de LCR de individuos controles y pacientes TSP/HAM: concentración de
proteínas, detección de la presencia de isoformas de proteína Tau, visualización de
fosforilaciones de la proteína Tau en serina y treonina e inmunodetección de residuos
fosforilados específicos.
Determinación del contenido de albúmina sérica en el LCR.
Esta determinación fue realizada mediante el método de Electroforesis Capilar,
midiendo la presencia del enlace peptídico (185 nm) refiriéndose a una curva estándar con
albúmina sérica humana (HSA). La Figura 4 muestra un electroferograma representativo
de LCR (Fig. 4A) junto con un estándar de HSA (Fig. 4B). El promedio de la
concentración de HSA de los controles y su desviación estándar fue 0,21 ± 0,13 mg/mL y el
del grupo patológico fue 0,24 ± 0,06 mg/mL. El LCR es un ultrafiltrado del plasma que se
produce en los plexos coriónicos por lo que la HSA es la proteína mayoritaria. (Blennow y
col., 1993). Resultados previos del laboratorio han mostrado una muy buena correlación
entre las mediciones de HSA y la determinación de proteínas totales por los métodos
clásicos como el de Lowry (Valenzuela y col., 1999). Las ventajas de emplear la
Electroforesis Capilar sobre los otros métodos clásicos incluyen su sensibilidad, rapidez del

25
ensayo, un consumo sólo de nanolitros de la muestra la que además no se destruye ni se
manipula, pudiéndose recuperar para otro análisis. Los resultados encontrados indican que
la concentración de proteínas presente en el LCR de los pacientes está dentro de los valores
normales que se describen entre 0,15-0,45 mg/mL (Blennow y cols, 1993).
A)
B)
Figura 4: Presencia de HSA en LCR. (A) Electroferograma de una muestra representativa de LCR realizada mediante EC según las condiciones descritas en la sección Materiales y Métodos. (B) En esta figura se muestra un electroferograma de un estándar de HSA e IgG medidos mediante EC en las condiciones descritas en la sección de Materiales y Métodos.
IgG

26
Inmunodetección de Tau en LCR.
La detección de Tau se realizó mediante “immunowestern blot” utilizando
anticuerpos policlonales monoespecíficos. Inicialmente se buscaron las condiciones
óptimas de su detección en LCR, utilizando el anticuerpo a las diluciones propuestas por el
fabricante (USBiological), se ensayaron diversas diluciones anticuerpo secundario para
evitar interferencia en la señal. En la Figura 5 se observa una tenue banda alrededor de la
zona de 50 kDa al utilizar el anticuerpo secundario (anti-IgG de conejo hecho en cabra) a
diluciones menores de 1:20.000, lo que implicó realizar los experimentos al menos con esta
dilución por una parte, para evitar la interferencia y por otra, observar una señal medible.
Este anticuerpo según el fabricante (USBiological) reconoce bandas de Tau entre 40 y 67
kDa. También se efectuó un barrido de la cantidad de LCR a utilizar escogiéndose trabajar
con 7,5 microlitros de muestra.
Figura 5: Búsqueda de condiciones para detección de Tau en LCR. “Immunowestern blot” de una muestra de LCR revelado con anticuerpos policlonales a una dilución de 1:1.000, utilizando distintas diluciones del anticuerpo secundario, y el control en ausencia del anticuerpo primario. La flecha indica la posición del estándar de masa molecular de 60,4 kDa.

27
Análisis de Tau en muestras de LCR de controles y pacientes.
El resultado del “immunowestern blot” de 6 muestras de LCR de controles y 6 de
TSP/HAM se muestra en la Figura 6. A fin de poder hacer un análisis densitométrico de
películas fotográficas donde las intensidades de las señales no estuvieran saturando dichas
películas, hubo que buscar las condiciones más adecuadas para el revelado. Por esto el
revelado se hizo en tiempo breve lo que significó que sólo la banda de 52 kDa fue
prácticamente la única detectada. Más adelante se muestra que esta es la banda más
característica de Tau y además se deduce de los experimentos descritos posteriormente en
el aislamiento por immunoafinidad y por el reconocimiento por anticuerpos contra residuos
específicos fosforilados en serina y treonina. La aplicación de estos anticuerpos en
determinaciones en muestras de LCR de pacientes con diversas patologías se encuentra
ampliamente reportada en la literatura.
Figura 6: Detección de Tau en muestras de LCR. “Immunowestern blot” de muestras de LCR controles (C1-C6) y de muestras de pacientes (P1-P6). Las masas moleculares señaladas corresponden a los valores calculados para estas tres señales inmunoreactivas contra anticuerpos policlonales anti-Tau.
La comparación de los niveles relativos de Tau de las muestras de controles y
pacientes se realizó a través de un análisis densitométrico de las películas fotográficas. Los
resultados de las distintas muestras de ambos grupos, expresados en unidades arbitrarias de

28
pixeles (UA)/mg HSA, se encuentran en la Figura 7. Estos datos mostraron tener una
distribución normal y las varianzas de ambos grupos fueron similares, por lo que utilizó una
Prueba T. Este análisis estadístico no mostró diferencias significativas de la señal medida
de Tau entre el grupo TSP/HAM y el control, siendo el valor promedio con su desviación
estándar del grupo control de 20.649 ± 4.213 UA/mg HSA, y el del grupo TSP/HAM de
20.771 ± 16.087.
Figura 7: Distribución de los niveles de Tau en los grupos. En el gráfico se muestra la cantidad de Tau, expresada como UA/mg de HSA, de ambos grupos. En cada caso se muestra el promedio señalizado como una barra.

29
Presencia de fosfoserina y fosfotreonina en la proteína Tau de LCR
Se analizó la posible colocalización de la proteína Tau con las bandas detectadas con
los anticuerpos anti-fosfoserina y anti-fosfotreonina para ver la presencia de estos sitios
fosforilados en estos residuos. Sin embargo, como se ve en la Figura 8 se observa la
presencia de una banda principal muy amplia e intensa centrada en una masa molecular
alrededor de 67 kDa. Con estos resultados fue imposible relacionar las bandas del LCR
obtenidas en los “immunowestern blot” contra fosfoserina y fosfotreonina con las bandas
correspondientes a Tau.
Figura 8: Inmunoreactividad de LCR con anticuerpos que reconocen residuos fosforilados. “Immunowestern blot” de LCR revelado con anticuerpos contra fosfoserina y fosfotreonina. Se indica la banda correspondiente al estándar de 60,7 kDa presente en el carril de los estándares.

30
Análisis de residuos específicos fosforilados de Tau en LCR.
Diversas enfermedades neurodegenerativas se han asociado a una hiperfosforilación
de algunos residuos de serina y/o treonina, por lo que se investigó la presencia de al menos
9 residuos específicos fosforilados de Tau mediante el uso de los correspondientes
anticuerpos monoespecíficos para ellos comercialmente disponibles. Realizando
“immunowestern blot” se encontró que existía reacción positiva para los 9 residuos
investigados, siendo éstos treonina 181 (T181), serina 199 (S199), treonina 205 (T205),
treonina 231 (T231), serina 262 (S262), serina 356 (S356), serina 396 (S396), serina 404
(S404) y serina 422 (S422). Al utilizar estos anticuerpos sólo se observó una banda de 52
kDa similar a la isoforma de Tau mayoritaria (ver Figura 9). El control negativo del
“immunowestern blot” en ausencia del primer anticuerpo no dio reacción positiva, por lo
que se siguió utilizando las mismas condiciones que las del anticuerpo anti-Tau policlonal,
es decir con el segundo anticuerpo a dilución 1:20.000.
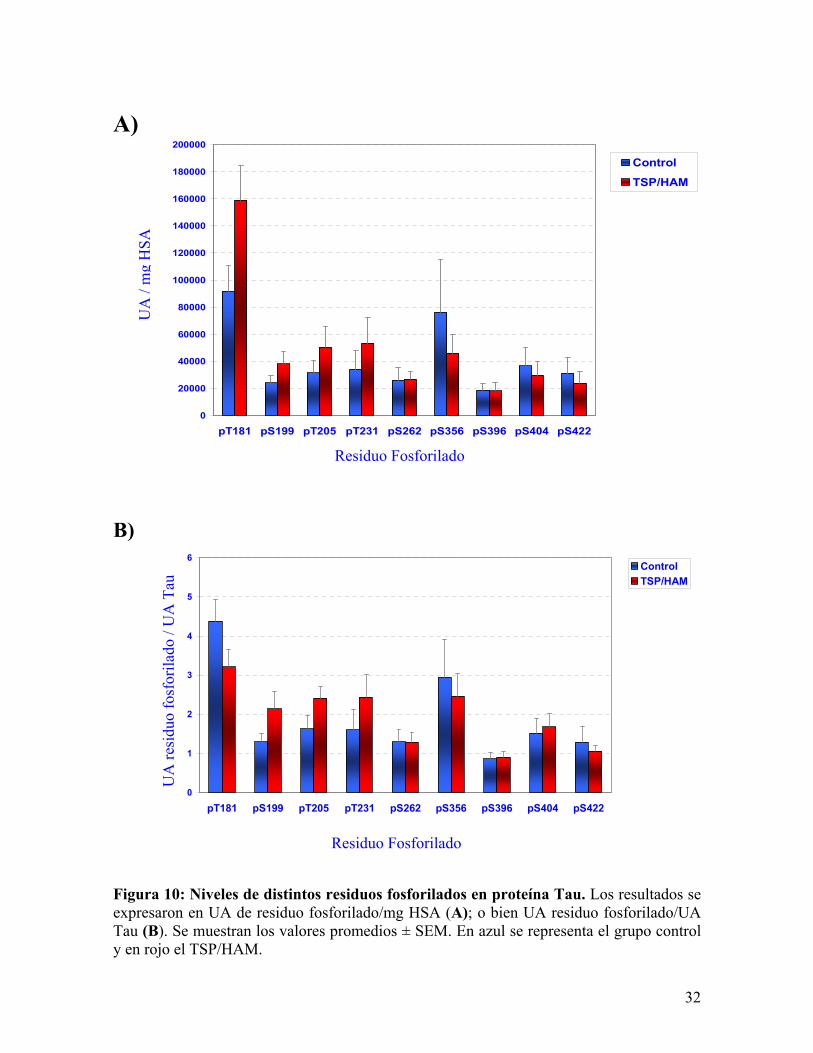
Una vez obtenidas las películas fotográficas se procedió a realizar un análisis
densitométrico de éstas. Los promedios de los valores expresados como UA/mg HSA y su
desviación estándar media (SEM) se muestran en la Figura 10A. Los resultados también se
expresaron en función de la concentración de Tau, mostrándose en la Figura 10B los
promedios de los valores UA residuo fosforilado/UA de Tau ± SEM. Esta comparación
entre las muestras de controles y pacientes se hizo ensayando en la misma membrana las 6
muestras controles y 6 patológicas. Las diferencias experimentales producto de la
electrotransferencia, tratamiento con los anticuerpos primarios y secundarios, y finalmente
el revelado con el sustrato quimioluminiscente, se caracterizan por una gran variabilidad de

31
membrana en membrana. Lo que hace imprescindible que todas las muestras sean
separadas en el mismo gel y tengan idéntico tratamiento posterior.
Figura 9: Detección de nivel de fosforilación en residuos específicos. “Immunowestern blot” de 6 muestras de LCR de controles (C1-C6) y de 6 pacientes (P1-P6). Se señalan los residuos que están reconociendo los anticuerpos y además la correspondiente inmunoreactividad frente anticuerpos policlonales contra Tau total.
32
UA
resi
duo
fosf
orila
do /
UA
Tau
Residuo Fosforilado
0
1
2
3
4
5
6
pT181 pS199 pT205 pT231 pS262 pS356 pS396 pS404 pS422
ControlTSP/HAM
UA
/ m
g H
SA
Residuo Fosforilado
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
pT181 pS199 pT205 pT231 pS262 pS356 pS396 pS404 pS422
Control
TSP/HAM
A) B)
Figura 10: Niveles de distintos residuos fosforilados en proteína Tau. Los resultados se expresaron en UA de residuo fosforilado/mg HSA (A); o bien UA residuo fosforilado/UA Tau (B). Se muestran los valores promedios ± SEM. En azul se representa el grupo control y en rojo el TSP/HAM.

33
Estos datos presentaron una distribución normal y varianzas de ambos grupos
similares aplicándose, en consecuencia el análisis estadístico a través de la Prueba T. En la
Tabla 1 se resume el análisis estadístico, donde se puede ver que sólo en el caso del residuo
treonina 181 el valor de P es el más cercano a la significancia (0,063)
Tabla 1: Resumen de los análisis estadísticos de los niveles de Tau fosforilado en residuos específicos. En esta tabla se muestran los valores de P (significancia) para los casos estudiados y graficados anteriormente. El n fue de 6 de controles y de 6 patológicos.

34
Segundo Objetivo: Aislar proteína Tau para estudios de Espectrometría de Masas
Aislamiento de Tau desde el LCR
El segundo Objetivo específico de este trabajo fue dirigido para poder efectuar en un
futuro un análisis proteómico de Tau en el LCR. Se han realizado muchos estudios de
proteómica en LCR porque refleja las proteínas del SNC secretadas desde el tejido
neuronal, y se han dirigido principalmente a la búsqueda de biomarcadores en
enfermedades neurodegenerativas (Davidsson y Sjögren, 2005). En este caso, el interés fue
confirmar los resultados obtenidos mediante el uso de anticuerpos específicos contra
diversos residuos de serina y treonina fosforilados, a través de análisis por Espectrometría
de Masas, técnica que permite determinar la presencia de modificaciones post-
traduccionales en la proteína Tau, como la fosforilación de residuos específicos de Tau.
Esto hizo necesario aislar Tau del LCR descartando otras proteínas interferentes por
lo que se emplearon dos estrategias de cromatografía de afinidad: a) una de ellas fue con
una resina unida a Ga(III) que interacciona selectivamente con grupos cargados
negativamente, por lo que se considera selectiva para unir proteínas o péptidos fosforilados;
b) la otra estrategia se basó en la unión específica a anticuerpos.
Aislamiento de Tau mediante cromatografía de afinidad con Ga(III)
En la Figura 11A se muestra un “immunowestern blot” de dos eluciones consecutivas
desde una columna cromatográfica de Ga(III) revelado con anticuerpos policlonales anti-
Tau. Ambos patrones de bandas de 75, 52 y 25 kDa fueron similares entre sí y además a
los que se han podido observar en LCR (se compara con un LCR concentrado ocho veces

35
mediante “speed vac”). Para ver si las proteínas retenidas por esta columna de afinidad por
metales correspondían a proteínas fosforiladas se reveló el “immunowestern blot” con
anticuerpos contra fosfotreonina, usando como control el anticuerpo policlonal contra el
residuo fosforilado en serina 262 de Tau (Fig. 11B). Igual con las muestras de LCR se
observa una gran banda inmunoreactiva contra anti-fosfotreonina y además otras bandas de
mayor y menor masa molecular (ver Fig.8), a diferencia del uso del anticuerpo específico
para la serina 262 fosforilada de tau donde sólo se detecta la banda de proteína de 52 kDa
(ver Fig. 9).
A) B)
Figura 11: Detección de Tau y treonina fosforilada en columna cromatográfica de Galio(III). (A) En la figura se muestran el primero y segundo eluido de las columnas de Ga(III) comparados con un LCR, concentrado 8 veces mediante “speed vac”. (B) Una de las mismas muestra eluidas de la columna reveladas con anticuerpos contra treonina-fosforilada y con anticuerpos contra anticuerpos que reconocen específicamente a la Ser262 de Tau fosforilada.

36
Aislamiento de Tau mediante cromatografía de inmunoafinidad
La inmunoprecipitación de la proteína Tau proveniente del LCR se realizó: A) con un
anticuerpo monoclonal contra Tau unido a la fase sólida de proteína A/G-agarosa; B)
uniendo covalentemente un anticuerpo policlonal contra Tau a CNBr-“Sepharose”.
Inmunoprecipitación de Tau mediante Proteína A/G-Agarosa.
El “immunowestern blot” de las muestras eluídas de la agarosa mostró que al revelar
con el mismo anticuerpo monoclonal utilizado para la inmunoprecipitación aparecía una
banda a 52 kDa (Fig. 12A). Al hacer el control sin el primer anticuerpo y revelando sólo
con el anticuerpo secundario contra ratón aparece una fuerte señal a 50 kDa similar a la
observada al revelar con el anticuerpo primario y secundario. Esta banda immunoreactiva
frente el anticuerpo secundario, por su masa molecular, debe corresponder a la subunidad
pesada de IgG del anticuerpo primario. Esta señal no se observó al revelar con un
anticuerpo secundario contra conejo. Igualmente esta interferencia se suprimió al revelar
con un anticuerpo policlonal contra tau (Figura 12B).
Al usar concentraciones mayores de anticuerpo primario y cantidades crecientes de
inmunoprecipitado es posible detectar más isoformas y posibles productos de degradación
(Figura 13).
Es importante rescatar la posibilidad de analizar los residuos fosforilados de estas
muestras inmunoprecipitadas a través del sistema Proteína A/G-agarosa, dado que para sus
detecciones se utilizan anticuerpos policlonales y por ende, no existe la interferencia
anteriormente descrita. Por lo tanto, se efectuaron los análisis para determinar si la banda
inmunoreactiva contra anticuerpos anti-Tau eran reconocidos por anticuerpos anti-

37
fosfoserina y fosfotreonina, y posteriormente con anticuerpos contra los 9 residuos
fosforilados de Tau.
A) B)
Figura 12: Interferencia del anticuerpo primario presente en inmunoprecipitado con A/G agarosa y su no detección al revelar con un anticuerpo policlonal. (A) Tanto en presencia como ausencia del primer anticuerpo (monoclonal antiTau) se apreció reacción positiva en los inmunoprecipitados al revelar con el mismo anticuerpo monoclonal con que se precipitó. (B) Esta interferencia se eliminó al revelar el mismo inmunoprecipitado con un anticuerpo policlonal contra el residuo fosforilado 181 de Tau.
Figura 13: Bandas inmunoreactivas contra anticuerpos policlonales contra Tau utilizando cantidades variables de inmunoprecipitados. Las flechas indican las bandas mayoritarias que pueden corresponder a Tau o sus productos de degradación. Se agregaron al “immunowestern blot” 3 y 5 μL de inmunoprecipitados.

38
Presencia de fosfoserina y fosfotreonina en la proteína Tau de inmunoprecipitados
con Proteína A/G-Agarosa.
Al analizar los inmunoprecipitados para ver las proteínas que poseen sitios fosforilados en
serina y treonina, se aprecian dos bandas de 52 y 75 kDa (Figura 14). Siendo sólo la de 52
kDa detectada por el anticuerpo anti-Tau monoclonal. Cabe destacar que las cadenas
pesadas de los anticuerpos no sufren modificaciones post-traduccionales del tipo
fosforilación, por lo que es descartable que la banda que aparece a la altura de Tau sea el
anticuerpo monoclonal presente fosforilado. Estos resultados apuntan a que la banda
correspondiente a Tau de 52 kda presenta fosforilaciones en serina y treonina.
Análisis de residuos fosforilados específicos en inmunoprecipitados con Proteína
A/G-Agarosa.
Se procedió a estudiar los inmunoprecipitados en 4 pacientes y 4 controles mediante
“immunowestern blot”, lo que se muestra en la Figura 15. Se puede apreciar que en todos
los casos sólo se observó una banda inmunoreactiva con una masa molecular de 52 kDa. En
la Figura 16 se muestra la tinción con plata de estas muestras separadas mediante SDS-
PAGE, la banda principal en la zona de 50 kDa (que debe corresponder a la IgG del
anticuerpo primario) no permite distinguir la banda de 52 kDa que correspondería a Tau.
La interferencia producida por la presencia del anticuerpo primario utilizado para
inmunoprecipitar nos impidió emplear el método con el uso de la proteína A/G agarosa
para aislar la banda correspondiente Tau desde el gel de poliacrilamida para su posterior
análisis por Espectrometría de Masas. Por esto se utilizó otra técnica de
inmunoprecipitación en que el anticuerpo se une covalentemente a la matriz.

39
Figura 14: Detección de Tau y residuos serina y treonina fosforilados en inmunoprecipitados con Proteína A/G-agarosa. “Immunowestern blot de muestras de controles (C1-C3) y TSP/HAM (P1-P3). En la imagen se aprecian las bandas correspondientes a fosfoserina y fosfotreonina de los inmunoprecipitados, comparando con la banda reconocida con el anticuerpo contra Tau. En la parte superior se aprecia al grupo al que pertenecen las muestras.
Figura 15: Detección de residuos específicos fosforilados de Tau en inmunoprecipitados con Proteína A/G-agarosa. Se aprecia en la figura los diversos “immunowestern blot” revelados con anticuerpos contra los residuos fosforilados de Tau correspondientes que se señalan a la izquierda. Las muestras son de 4 controles (C1-C4) y 4 pacientes (P1-P4).

40
Figura 16: Detección proteica en los inmunoprecipitados con Proteína A/G-agarosa. Mediante tinción con Plata se ven los productos proteicos obtenidos en la inmunoprecipitación. En la parte superior esta indicado al grupo al cuál pertenecen las muestras que es similar al de la Fig. 14 (C1-C4, controles; P1-P4, TSP/HAM).
Inmunoprecipitación de Tau mediante “Sepharose ” activada con CNBr
Este procedimiento de inmunoprecipitación requiere de una gran cantidad de muestra
según instrucciones del fabricante, por lo que se realizó exclusivamente para poder obtener
muestras para Espectrometría de Masas. Los análisis por “immunowestern blot”
demostraron la ausencia de contaminación con los anticuerpos primarios (policlonales
monoespecíficos) lo que se muestra en la Figura 17. En la Fig. 17 se confirma la ausencia
del primer anticuerpo al no observarse bandas cuando “immunowestern blot” se revela sin
el anticuerpo primario.
Posteriormente, fue importante determinar si la banda principal de 52 kDa reconocida
por “immunowestern blot” tenía una concentración de proteínas adecuada para su posterior
análisis por Espectrometría de Masas el gel de poliacrilamida se tiñó con azul de

41
Coomassie coloidal, mostrando que si había una masa suficiente para los estudios de
proteómica (Figura 18).
Se visualizan otras bandas tanto de menor como de mayor masa molecular que la de
52 kDa. Estas otras bandas débiles observadas pueden corresponder en el caso de la de
menor masa molecular a un producto de degradación de Tau o bien a otra proteína que
coinmuprecipitó con Tau. También las de masa superior podrían corresponder a otras
formas de Tau (producto de “splicing” alternativo o hiperfosforilación), pero no se puede
descartar que también correspondan a proteínas que hayan coimmunoprecipitado.
Figura 17: Presencia de Tau en IPP mediante “Sepharose” activada con CNBr. (A) Inmunoreactividad contra anti-Tau policlonal en el ipp y ausencia de reconocimiento por el anticuerpo secundario al revelar el “immunowestern blot” sin adicionar el anticuerpo policlonal primario contra Tau. (B)
Figura 18: Tinción Proteica de IPP con “Sepharose” activada con CNBr. La banda más importante se ve en la zona de 52 kDa hablando de una importante banda inmunoprecipitada correspondiente a la masa molecular de Tau.

42
Análisis mediante Espectrometría de Masas.
El producto de inmunoprecipitación mediante “sepharose” activada con CNBr
mostró una banda principal de 52 kDa por SDS-PAGE, la cuál fue recortada y procesada
para luego ser analizada por el equipo de Espectrometría de Masas, MALDI-TOF. La
huella peptídica resultante de ésta banda se muestra en la Figura 19A. Los valores de
intensidad indicados en el gráfico corresponden a unidades arbitrarias de intensidad (UA).
En este gráfico se eliminaron las señales de m/z menores a 850,0 ya que esa zona
corresponde a las señales obtenidas por la matriz. En la Figura 19B se aprecia el patrón de
péptidos obtenidos para el estándar de Tau.
Como resultado de la identificación se encontró que existían 3 péptidos coincidentes
en ambos espectros, según análisis obtenido en la base de datos del servidor Mascot. Sin
embargo el “score” alcanzado con este único análisis no permitió identificar con certeza la
presencia de la proteína Tau, además aunque este espectro fue muy similar al obtenido con
un estándar de proteína Tau, se encontró un desfase de 30 Da en los valores de m/z en las
señales más intensas. Los péptidos identificados corresponden a sitios consensuados de
MAP

43
A)
B)
Figura 19. Comparación de las Huellas peptídicas por MALDI-TOF de Tau inmunoprecipitado con un estándar. (A) Espectro de Tau inmunoprecipitado utilizando “Sepharose” CNBr y sometido a tripsinización (B) Espectro de Tau estándar sometido a tripsinización. Se indican los péptidos coincidentes entre las muestras.

44
Discusión
Análisis en Líquido Cefalorraquídeo.
La búsqueda de Tau en el LCR se ha descrito ampliamente, habiendo sido el LCR una
excelente fuente de estudio para variadas enfermedades neurodegenerativas (Davidsson y
Sjögren, 2005). Hasta ahora hay muchos trabajos que buscan en LCR proteínas como
biomarcadores que diferencien diversas enfermedades del SNC por la capacidad de
entregar información sobre las proteínas que vienen del SNC donde suelen estar en
concentraciones medibles (Sjögren y cols, 2001; Riemenschneider y cols, 2003; Andreasen
y cols, 2003; Satoh y cols, 2006). En LCR se ha medido los niveles relativos de Tau y
además, se ha evaluado la fosforilación de algunos de sus residuos (Sjögren y cols, 2001;
Hampel y cols, 2005; Maccioni y cols, 2006).
Como primer paso se decidió comparar los niveles de Tau total entre los grupos
control y TSP/HAM encontrándose que no existen diferencias significativas entre ambos
grupos. Estos resultados difieren de lo descrito en otras enfermedades neurodegenerativas
como enfermedad de Alzheimer, enfermedad de Creutzfeldt-Jakob, esclerosis múltiple
(Kapaki y cols, 2000; Sjögren y cols, 2001; Avila y cols, 2004; Satoh y cols, 2006), en
cambio son similares a lo observado en pacientes con enfermedad de Parkinson (Molina y
cols, 1997).
Dado que la funcionalidad de Tau, como pieza clave en el ensamblaje de los
microtúbulos, está principalmente regulada por su fosforilación, el siguiente paso fue
comparar el grado de fosforilación de Tau. El primer acercamiento fue tratar de analizar si
era posible colocalizar en el LCR las proteínas inmunoreactivas con anticuerpos anti fosfo-
serina y anti fosfo-treonina con las bandas reconocidas por el anticuerpo contra Tau. Como

45
se ve en la Fig. 8, el patrón obtenido con los anticuerpos contra estos residuos fosforilados
impidió poder afirmar con certeza la presencia de las bandas reactivas contra Tau, esto
dado que la inmunorreacción importante se observaba en una banda de masa molecular
centrada en la zona entre 70-60 kDa que interfería con la banda de 52 kDa. Estos resultados
indican que hay varias otras proteínas aparte de Tau que se encuentran en el LCR y que
están fosforiladas en serina y treonina. No se descarta que una de ellas sea la albúmina
sérica (de masa molecular 67 kDa) por un trabajo de literatura que muestra que en estudios
in vitro se ha logrado fosforilarla con CK1 y con quinasa dependiente de AMPc (Martin y
Ekman, 1986) y más recientemente trabajos en nuestro laboratorio (Alberti, C.
Comunicación Personal.)
Fue entonces necesario obtener una aproximación más específica para estimar el
grado de fosforilación de Tau. Como ya fue citado anteriormente existe una regulación fina
sobre Tau en el ensamblaje de los microtúbulos, lo que viene dada por la fosforilación en
residuos específicos en distintas zonas de la proteína. Por lo tanto, averiguar que residuos
son los que están siendo afectados en la enfermedad TSP/HAM de manera distinta o similar
a otras enfermedades neurodegenerativas donde hay estudios previos permitiría comprender
las causas del daño axonal. A su vez encontrar en TSP/HAM un daño a nivel de transporte
axonal podría explicar el origen a la retrotracción axonal.
Al realizar los estudios con los 9 anticuerpos contra los residuos fosforilados de Tau,
se encontró que sólo la banda de 52 kDa es inmunoreactiva y colocaliza con la
correspondiente banda mayoritaria ya observada al revelar con anticuerpos contra Tau. El
único residuo específico que estuvo más cercano a alcanzar un aumento significativo (p =
0063) entre los grupos fue el residuo treonina 181 (Tabla 1) al expresar las unidades
arbitrarias de la banda immunoreactiva contra el anticuerpo específico por el contenido de

46
albúmina humana en la muestra de LCR. Tampoco, al expresar estas unidades arbitrarias en
función del contenido total de Tau, se obtuvo significancia estadística obteniéndose un
valor de p = 0,142. En consecuencia, para confirmar o descartar que hay un aumento
significativo en la fosforilación de treonina 181 es recomendable por una parte, hacer un
análisis con un número mayor de muestras de LCR de TSP/HAM y de controles y por otra,
cuantificar mediante ensayos de ELISA la concentración de treonina 181 fosforilada
presente por mol de Tau. Aunque este residuo se encuentra fuera del sitio de unión a
microtúbulos, se ha descrito su importancia como marcador temprano en la enfermedad de
Alzheimer (Schonknecht y cols, 2003) y en otras enfermedades neurodegenerativas como
esclerosis múltiple (Bartosik-Psujek y Stelmasiak, 2005) y enfermedad de Creutzfeldt-
Jakob (Satoh y cols, 2006). No es claro el efecto de esta fosforilación en las enfermedades
estudiadas, pero si las placas PHF, muestran Tau hiperfosforilado en treonina 181
(Vanmechelen y cols, 2000).
Se ha descrito que tanto la GSK-3β como la Cdk-5 son capaces de fosforilar a la
treonina 181, es decir en los pacientes con paraparesia espástica tropical podría existir un
aumento en la actividad de las enzimas o un incremento en su cantidad, lo que podría estar
determinando el aumento de su nivel de fosforilación o hiperfosforilación en Tau. Esto a su
vez podría estar asociado a la acción de la proteína Tax que como se mencionó en la
Introducción que es capaz de activar las vías transduccionales que aumentarían la actividad
de estas enzimas (Jeang y cols, 2004). Sería interesante entonces utilizar un modelo celular
neuronal que permitiría hacer estudios del efecto que produce la exposición a productos
secretados por linfocitos T infectados con el virus como Tax, lo que podría aclarar dos
preguntas que han surgido en esta memoria.

47
a) ¿Se produce un aumento en los niveles y/o actividad de la GSK-3β y/o de la Cdk-
5 en las células neuronales por efecto de proteínas virales? ¿Hay un aumento del
nivel de fosforilación en el residuo treonina 181 de Tau o de otros residuos en
dichas células expuestas al estímulo?
b) ¿Se producen cambios en el citoesqueleto axonal, por esta exposición a proteínas
virales? Si se observaran cambios en el citoesqueleto axonal ¿se producirá un
daño a nivel de transporte?
Aislamiento de proteína Tau desde el LCR
El siguiente paso en la evolución del trabajo fue intentar aislar la proteína Tau desde
LCR que fue reconocida con anticuerpos anti-Tau para realizar análisis por Espectrometría
de Masas. Estos análisis corresponden a una técnica de punta para estudios a nivel de
proteómica que permite conocer exactamente la presencia de modificaciones post-
traduccionales, entre ellas las fosforilaciones (D’Ambrosio y cols, 2006). Hay ya reportes
en literatura de estas determinaciones (Lund y cols, 2001; Yoshimura y cols, 2003).
Para lograrlo se intentaron 2 acercamientos:
a) Concentración de proteínas fosforiladas a través de unión por afinidad a
columnas de Galio (III)
b) Concentración a través de inmunoabsorción empleando dos tipos de
inmunoprecipitación, uno uniendo los anticuerpos no covalentemente a proteína
A/G-Agarosa y el otro, uniendo covalentemente los anticuerpos “Sepharose”
activada con CNBr.

48
La primera aproximación con la columna de afinidad por iones metálicos no resultó
ser adecuada, aunque los anticuerpos anti-tau reconocieron bandas, el revelado con
anticuerpos contra fosfo-treonina mostró que no era factible distinguir entre las distintas
bandas inmunoreactivas. Sin embargo, este resultado indica que esta columna de Ga(III)
retuvo proteínas fosforiladas (Posewitz y Tempst, 1999).
Al emplear las técnicas de inmunoprecipitación sólo resultó adecuada la que evitaba
la contaminación con los anticuerpos primarios utilizados para unirse la proteína A/G-
Agarosa, cuya banda se superponía en el “immunowestern blot” con la banda
correspondiente a la proteína Tau de 52 kDa (ver Fig. 12A). Sin embargo, en muestras
inmunoprecipitadas provenientes de LCR de pacientes y controles se demostró que había
fosforilación en los 9 residuos de Tau que habían sido estudiados en LCR. Como en este
caso el nivel de Tau se midió con el mismo anticuerpo monoclonal usado para
inmunoprecipitar, las bandas detectadas corresponden a una mezcla de Tau y de IgG, por lo
que no se hizo un análisis estadístico de estos resultados. Además, no se expresó por el
contenido de albúmina como en el caso del LCR porque la elución fue en tampón de
muestra de Laemmli (1970), no siendo factible cuantificar el contenido final de proteínas.
Para expresar los resultados por el contenido de Tau, se debió haber revelado con
anticuerpos policlonales en cuyo caso no hubiera interferido el anticuerpo monoclonal.
Para simplificar los análisis por Espectrometría de Masas se buscó un método de
separación que evitara la contaminación proteica con IgG, lo que se logró utilizando la otra
técnica de inmunoprecipitación donde el anticuerpo se encontraba unido covalentemente a
la matriz. Se demostró que esto efectivamente ocurría y no se detectaba su presencia en los
eluidos de esta fase sólida (“Sepharose”-CNBr). Como este método según describe el
fabricante requiere un mayor volumen de muestra, la cantidad de proteínas reconocida por

49
el anticuerpo contra Tau mostró una banda principal muy destacada en geles de
poliacrilamida teñidos con azul de Coomassie coloidal (ver Fig. 17), indicando que hay
suficiente cantidad de proteínas para los análisis de Espectrometría de Masas.
Análisis de muestras inmunoprecipitadas por Espectrometría de Masas
Los resultados preliminares del análisis por espectrometría de Masas de la muestra
inmunoprecipitada indican una similitud en la identificación de la proteína Tau. A pesar de
que el espectro obtenido para el estándar era muy similar al de la muestra, se observó una
diferencia de 30 Da en m/z, encontrándose valores menores en las muestras
inmunoprecipitadas. Una de las posibles explicaciones de este desfase en la huella peptídica
puede ser la oxidación en los residuos prolina que producen una disminución en el tamaño
de los péptidos igual a 30 Da. Esta disminución se encuentra descrita en el servidor Unimod
y hace referencia al paso de prolina a 2-pirrilidona mediante oxidación, perdiéndose dos
átomos de Hidrógeno, uno de Carbono y uno de Oxígeno (Berlett y Stadtman, 1997). La
proteína Tau posee varios residuos de prolina los que caracterizan los sitios de fosforilación
en los residuos serina y treonina por quinasas dirigidas a prolina (Tseng y cols, 1999), y
además posee una región rica en prolina (Avila y cols, 2004). Considerando que las
muestras han sido tomadas mediante punción lumbar, posteriormente centrifugadas para
eliminar células y mantenidas congeladas hasta su inmunoprecipitación, sin tomar
precaución para evitar una atmósfera oxidante, es muy probable las alteraciones en
prolinas.
El análisis de otros de los péptidos presentes en el espectro de MALDI sugiere la
presencia de las siguientes proteínas “Microtubule-Actin cross linking Factor 1”, “Tubulin-

50
Specific Chaperon B”, “serine/threonine PP2A regulatory subunit B” y PARK7. Todas
estas proteínas tienen relación con el citoesqueleto celular, lo cuál podría indicar la
presencia de co-inmunoprecipitación de éstas proteínas que estarían interactuando con Tau.
Este es un estudio preeliminar para estimar la posibilidad de aislar Tau y analizarla
mediante Espectrometría de Masas, por lo que se utilizó una muestra control, dejando a
futuro el análisis comparativo entre los grupos TSP/HAM y controles.

51
Conclusiones
1.- Se logró la determinación de Tau en el LCR no encontrándose diferencias significativas
en los niveles de Tau entre ambos grupos
2.- Se encontró que Tau está fosforilado en todos los residuos analizados, encontrándose
diferencia cercana a la significancia entre los grupos, sólo en el residuo treonina 181, lo
que se podría confirmar posteriormente analizando un número mayor de muestras y
utilizando otras técnicas de detección.
3.- Se aisló la proteína Tau, a través de inmunoprecipitación, para analizar su grado de
fosforilación en los distintos residuos y aplicarle Espectrometría de Masas.

52
Referencias
Alvarez, A., R. Toro, et al. (1999). "Inhibition of tau phosphorylating protein kinase cdk5 prevents beta-amyloid-induced neuronal death." FEBS Lett 459(3): 421-6.
Andreasen, N., E. Vanmechelen, et al. (2003). "Cerebrospinal fluid levels of total-tau, phospho-tau and A beta 42 predicts development of Alzheimer's disease in patients with mild cognitive impairment." Acta Neurol Scand Suppl 179: 47-51.
Arima, N. and C. Tei (2001). "HTLV-I Tax related dysfunction of cell cycle regulators and oncogenesis of adult T cell leukemia." Leuk Lymphoma 40(3-4): 267-78.
Avila, J., J. J. Lucas, et al. (2004). "Role of tau protein in both physiological and pathological conditions." Physiol Rev 84(2): 361-84.
Bartosik-Psujek, H. and Z. Stelmasiak (2006). "The CSF levels of total-tau and phosphotau in patients with relapsing-remitting multiple sclerosis." J Neural Transm 113(3): 339-45.
Berlett, B. S. and E. R. Stadtman (1997). "Protein oxidation in aging, disease, and oxidative stress." J Biol Chem 272(33): 20313-6.
Bhatnagar, S. (2002). Neuroscience, Lippincott Williams & Wilkins. Billingsley, M. L. and R. L. Kincaid (1997). "Regulated phosphorylation and
dephosphorylation of tau protein: effects on microtubule interaction, intracellular trafficking and neurodegeneration." Biochem J 323 ( Pt 3): 577-91.
Blennow, K., P. Fredman, et al. (1993). "Protein analysis in cerebrospinal fluid. III. Relation to blood-cerebrospinal fluid barrier function for formulas for quantitative determination of intrathecal IgG production." Eur Neurol 33(2): 134-42.
Buee, L., T. Bussiere, et al. (2000). "Tau protein isoforms, phosphorylation and role in neurodegenerative disorders." Brain Res Brain Res Rev 33(1): 95-130.
Bussiere, T., P. R. Hof, et al. (1999). "Phosphorylated serine422 on tau proteins is a pathological epitope found in several diseases with neurofibrillary degeneration." Acta Neuropathol (Berl) 97(3): 221-30.
Cartier, L. and E. Ramirez (2005). "Presence of HTLV-I Tax protein in cerebrospinal fluid from HAM/TSP patients." Arch Virol 150(4): 743-53.
Cartier, L. M., J. G. Cea, et al. (1997). "Clinical and neuropathological study of six patients with spastic paraparesis associated with HTLV-I: an axomyelinic degeneration of the central nervous system." J Neuropathol Exp Neurol 56(4): 403-13.
D'Ambrosio, C., A. M. Salzano, et al. (2006). "Analytical methodologies for the detection and structural characterization of phosphorylated proteins." J Chromatogr B Analyt Technol Biomed Life Sci: [Epub ahead of print].
Davidsson, P. and M. Sjogren (2005). "The use of proteomics in biomarker discovery in neurodegenerative diseases." Dis Markers 21(2): 81-92.
Evans, D. B., K. B. Rank, et al. (2000). "Tau phosphorylation at serine 396 and serine 404 by human recombinant tau protein kinase II inhibits tau's ability to promote microtubule assembly." J Biol Chem 275(32): 24977-83.
Evans, K. J., E. R. Gomes, et al. (2005). "Linking axonal degeneration to microtubule remodeling by Spastin-mediated microtubule severing." J Cell Biol 168(4): 599-606.

53
Ferreirinha, F., A. Quattrini, et al. (2004). "Axonal degeneration in paraplegin-deficient mice is associated with abnormal mitochondria and impairment of axonal transport." J Clin Invest 113(2): 231-42.
Fu, D. X., Y. L. Kuo, et al. (2003). "Human T-lymphotropic virus type I tax activates I-kappa B kinase by inhibiting I-kappa B kinase-associated serine/threonine protein phosphatase 2A." J Biol Chem 278(3): 1487-93.
Galeno, H., E. Ramirez, et al. (1994). "Anti HTLV-I antibody titers in seropositive infected individuals." Rev Med Chil 122(9): 1004-7.
Gallo, D., L. M. Penning, et al. (1991). "Detection and differentiation of antibodies to human T-cell lymphotropic virus types I and II by the immunofluorescence method." J Clin Microbiol 29(10): 2345-7.
Garcia-Vallejo, F., M. C. Dominguez, et al. (2005). "Autoimmunity and molecular mimicry in tropical spastic paraparesis/human T-lymphotropic virus-associated myelopathy." Braz J Med Biol Res 38(2): 241-50.
Gessain, A., F. Barin, et al. (1985). "Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis." Lancet 2(8452): 407-10.
Godemann, R., J. Biernat, et al. (1999). "Phosphorylation of tau protein by recombinant GSK-3beta: pronounced phosphorylation at select Ser/Thr-Pro motifs but no phosphorylation at Ser262 in the repeat domain." FEBS Lett 454(1-2): 157-64.
Gong, C. X., T. Lidsky, et al. (2000). "Phosphorylation of microtubule-associated protein tau is regulated by protein phosphatase 2A in mammalian brain. Implications for neurofibrillary degeneration in Alzheimer's disease." J Biol Chem 275(8): 5535-44.
Gong, C. X., F. Liu, et al. (2005). "Post-translational modifications of tau protein in Alzheimer's disease." J Neural Transm 112(6): 813-38.
Haller, K., Y. Wu, et al. (2002). "Physical interaction of human T-cell leukemia virus type 1 Tax with cyclin-dependent kinase 4 stimulates the phosphorylation of retinoblastoma protein." Mol Cell Biol 22(10): 3327-38.
Hampel, H., K. Burger, et al. (2005). "Correlation of cerebrospinal fluid levels of tau protein phosphorylated at threonine 231 with rates of hippocampal atrophy in Alzheimer disease." Arch Neurol 62(5): 770-3.
Hasegawa, M. (2006). "Biochemistry and molecular biology of tauopathies." Neuropathology 26(5): 484-90.
Hirokawa, N., Y. Shiomura, et al. (1988). "Tau proteins: the molecular structure and mode of binding on microtubules." J Cell Biol 107(4): 1449-59.
Igakura, T., J. C. Stinchcombe, et al. (2003). "Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton." Science 299(5613): 1713-6.
Iwanaga, R., K. Ohtani, et al. (2001). "Molecular mechanism of cell cycle progression induced by the oncogene product Tax of human T-cell leukemia virus type I." Oncogene 20(17): 2055-67.
Iwatsubo, T. (2006). "Tauopathy: an overview." Neuropathology 26(5): 455-6. Jeang, K. T. (2001). "Functional activities of the human T-cell leukemia virus type I Tax
oncoprotein: cellular signaling through NF-kappa B." Cytokine Growth Factor Rev 12(2-3): 207-17.
Jeang, K. T., C. Z. Giam, et al. (2004). "Life, death, and tax: role of HTLV-I oncoprotein in genetic instability and cellular transformation." J Biol Chem 279(31): 31991-4.

54
Kang, D., Y. S. Gho, et al. (2002). "Highly Sensitive and Fast Protein Detection with Coomassie Brilliant Blue in Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis." Bull. Korean Chem. Soc. 23(11): 1511-1512.
Kapaki, E., G. P. Paraskevas, et al. (2000). "Increased cerebrospinal fluid tau protein in multiple sclerosis." Eur Neurol 43(4): 228-32.
Kesavapany, S., B. S. Li, et al. (2004). "Neuronal cyclin-dependent kinase 5: role in nervous system function and its specific inhibition by the Cdk5 inhibitory peptide." Biochim Biophys Acta 1697(1-2): 143-53.
Laemmli, U. K. (1970). "Cleavage of structural proteins during the assembly of the head of bacteriophage T4." Nature 227(5259): 680-5.
Li, X. H. and R. B. Gaynor (1999). "Regulation of NF-kappaB by the HTLV-1 Tax protein." Gene Expr 7(4-6): 233-45.
Liberski, P. P., J. Buczynski, et al. (1999). "Ultrastructural pathology of a Chilean case of tropical spastic paraparesis/human T-cell lymphotropic type I-associated myelopathy (TSP/HAM)." Ultrastruct Pathol 23(3): 157-62.
Liberski, P. P., P. Rodgers-Johnson, et al. (1994). "Ultrastructural pathology of human T-cell lymphotropic virus type I encephalomyelopathy in a white patient with adult T-cell leukemia/lymphoma." Ultrastruct Pathol 18(5): 511-8.
Lund, E. T., R. McKenna, et al. (2001). "Characterization of the in vitro phosphorylation of human tau by tau protein kinase II (cdk5/p20) using mass spectrometry." J Neurochem 76(4): 1221-32.
Maccioni, R. B., M. Lavados, et al. (2006). "Anomalously phosphorylated tau and Abeta fragments in the CSF correlates with cognitive impairment in MCI subjects." Neurobiol Aging 27(2): 237-44.
Maccioni, R. B., C. Otth, et al. (2001). "The protein kinase Cdk5. Structural aspects, roles in neurogenesis and involvement in Alzheimer's pathology." Eur J Biochem 268(6): 1518-27.
Martin, S. C. and P. Ekman (1986). "In vitro phosphorylation of serum albumin by two protein kinases: a potential pitfall in protein phosphorylation reactions." Anal Biochem 154(2): 395-9.
Molina, J. A., J. Benito-Leon, et al. (1997). "Tau protein concentrations in cerebrospinal fluid of non-demented Parkinson's disease patients." Neurosci Lett 238(3): 139-41.
Osame, M., K. Usuku, et al. (1986). "HTLV-I associated myelopathy, a new clinical entity." Lancet 1(8488): 1031-2.
Pigino, G., G. Paglini, et al. (1997). "Analysis of the expression, distribution and function of cyclin dependent kinase 5 (cdk5) in developing cerebellar macroneurons." J Cell Sci 110 ( Pt 2): 257-70.
Posewitz, M. C. and P. Tempst (1999). "Immobilized gallium(III) affinity chromatography of phosphopeptides." Anal Chem 71(14): 2883-92.
Prajapati, S. and R. B. Gaynor (2002). "Regulation of Ikappa B kinase (IKK)gamma /NEMO function by IKKbeta -mediated phosphorylation." J Biol Chem 277(27): 24331-9.
Ramirez, E., L. Cartier, et al. (2002). "Genetic characterization and phylogeny of human T-cell lymphotropic virus type I from Chile." Virus Res 84(1-2): 135-49.

55
Reynolds, C. H., J. C. Betts, et al. (2000). "Phosphorylation sites on tau identified by nanoelectrospray mass spectrometry: differences in vitro between the mitogen-activated protein kinases ERK2, c-Jun N-terminal kinase and P38, and glycogen synthase kinase-3beta." J Neurochem 74(4): 1587-95.
Riemenschneider, M., S. Wagenpfeil, et al. (2003). "Phospho-tau/total tau ratio in cerebrospinal fluid discriminates Creutzfeldt-Jakob disease from other dementias." Mol Psychiatry 8(3): 343-7.
Satoh, K., S. Shirabe, et al. (2006). "14-3-3 protein, total tau and phosphorylated tau in cerebrospinal fluid of patients with Creutzfeldt-Jakob disease and neurodegenerative disease in Japan." Cell Mol Neurobiol 26(1): 45-52.
Schneider, A., J. Biernat, et al. (1999). "Phosphorylation that detaches tau protein from microtubules (Ser262, Ser214) also protects it against aggregation into Alzheimer paired helical filaments." Biochemistry 38(12): 3549-58.
Schonknecht, P., J. Pantel, et al. (2003). "Levels of total tau and tau protein phosphorylated at threonine 181 in patients with incipient and manifest Alzheimer's disease." Neurosci Lett 339(2): 172-4.
Sengupta, A., J. Kabat, et al. (1998). "Phosphorylation of tau at both Thr 231 and Ser 262 is required for maximal inhibition of its binding to microtubules." Arch Biochem Biophys 357(2): 299-309.
Seubert, P., M. Mawal-Dewan, et al. (1995). "Detection of phosphorylated Ser262 in fetal tau, adult tau, and paired helical filament tau." J Biol Chem 270(32): 18917-22.
Sjogren, M., P. Davidsson, et al. (2001). "Both total and phosphorylated tau are increased in Alzheimer's disease." J Neurol Neurosurg Psychiatry 70(5): 624-30.
Smith, D. S. and L. H. Tsai (2002). "Cdk5 behind the wheel: a role in trafficking and transport?" Trends Cell Biol 12(1): 28-36.
Stamer, K., R. Vogel, et al. (2002). "Tau blocks traffic of organelles, neurofilaments, and APP vesicles in neurons and enhances oxidative stress." J Cell Biol 156(6): 1051-63.
Strittmatter, S. M. (2002). "Modulation of axonal regeneration in neurodegenerative disease: focus on Nogo." J Mol Neurosci 19(1-2): 117-21.
Tatebayashi, Y., N. Haque, et al. (2004). "Role of tau phosphorylation by glycogen synthase kinase-3beta in the regulation of organelle transport." J Cell Sci 117(Pt 9): 1653-63.
Towbin, H., T. Staehelin, et al. (1979). "Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications." Proc Natl Acad Sci U S A 76(9): 4350-4.
Trinczek, B., A. Ebneth, et al. (1999). "Tau regulates the attachment/detachment but not the speed of motors in microtubule-dependent transport of single vesicles and organelles." J Cell Sci 112 ( Pt 14): 2355-67.
Tseng, H. C., Q. Lu, et al. (1999). "Phosphorylated tau can promote tubulin assembly." Proc Natl Acad Sci U S A 96(17): 9503-8.
Tsujio, I., T. Tanaka, et al. (2000). "Inactivation of glycogen synthase kinase-3 by protein kinase C delta: implications for regulation of tau phosphorylation." FEBS Lett 469(1): 111-7.
Umehara, F., M. Abe, et al. (2000). "Axonal damage revealed by accumulation of beta-amyloid precursor protein in HTLV-I-associated myelopathy." J Neurol Sci 176(2): 95-101.

56
Utton, M. A., A. Vandecandelaere, et al. (1997). "Phosphorylation of tau by glycogen synthase kinase 3beta affects the ability of tau to promote microtubule self-assembly." Biochem J 323 ( Pt 3): 741-7.
Valenzuela, M. A., L. Cartier, et al. (1999). "Gelatinase activity of matrix metalloproteinases in the cerebrospinal fluid of various patient populations." Res Commun Mol Pathol Pharmacol 104(1): 42-52.
Vanmechelen, E., H. Vanderstichele, et al. (2000). "Quantification of tau phosphorylated at threonine 181 in human cerebrospinal fluid: a sandwich ELISA with a synthetic phosphopeptide for standardization." Neurosci Lett 285(1): 49-52.
Vasquez, P. (2003). "HTLV-I (Human T-cell Lymphotropic virus) something to say?" Rev Chil Infect 20(Supl 1): S34-S37.
Wada, Y., K. Ishiguro, et al. (1998). "Microtubule-stimulated phosphorylation of tau at Ser202 and Thr205 by cdk5 decreases its microtubule nucleation activity." J Biochem (Tokyo) 124(4): 738-46.
Wu, K., M. E. Bottazzi, et al. (2004). "Protein profile of tax-associated complexes." J Biol Chem 279(1): 495-508.
Xie, H., J. M. Litersky, et al. (1998). "The interrelationship between selective tau phosphorylation and microtubule association." Brain Res 798(1-2): 173-83.
Yoshida, M. (2001). "Multiple viral strategies of HTLV-1 for dysregulation of cell growth control." Annu Rev Immunol 19: 475-96.
Yoshimura, Y., T. Ichinose, et al. (2003). "Phosphorylation of tau protein to sites found in Alzheimer's disease brain is catalyzed by Ca2+/calmodulin-dependent protein kinase II as demonstrated tandem mass spectrometry." Neurosci Lett 353(3): 185-8.
Zheng, Y. L., S. Kesavapany, et al. (2005). "A Cdk5 inhibitory peptide reduces tau hyperphosphorylation and apoptosis in neurons." Embo J 24(1): 209-20.
Recommended

















