
UNIVERSIDAD DE SANTIAGO DE COMPOSTELA
Departamento de Química Analítica, Nutrición y Bromatología
CARACTERIZACIÓN DE MARCADORES PEPTÍDICOS ESPECÍFICOS
PARA LA IDENTIFICACIÓN DE ESPECIES DE LANGOSTINOS DE
INTERÉS COMERCIAL
por
Ignacio Ortea García
Tesis doctoral
Santiago de Compostela
2009


D. Alberto Cepeda Sáez, Catedrático de Universidad y Director del Departamento de Química
Analítica, Nutrición y Bromatología de la Universidad de Santiago de Compostela,
INFORMA:
Que D. Ignacio Ortea García ha realizado, bajo la dirección de la Dra. M. Pilar Calo Mata,
perteneciente a este departamento, y los Drs. José M. Gallardo Abuín y Benito Cañas Montalvo en el
Instituto de Investigaciones Marinas de Vigo (C.S.I.C.), el trabajo “Caracterización de marcadores
peptídicos específicos para la identificación de especies de langostinos de interés comercial” que
presenta para optar al grado de Doctor en Bioquímica.
Y para que así conste, firmo el presente informe en Santiago de Compostela, a 24 de Julio de
2009.
D. Alberto Cepeda Sáez


El Dr. José Manuel Gallardo Abuín, Profesor de Investigación del Consejo Superior de
Investigacines Científicas, con destino en el Instituto de Investigaciones Marinas de Vigo, la Dra. M.
Pilar Calo Mata, Profesora Contratada Doctora del Departamento de Química Analítica, Nutrición y
Bromatología de la Universidad de Santiago de Copostela, y el Dr. Benito Cañas Montalvo, Profesor
Contratado Doctor del Departamento de Química Analítica de la Universidad Complutense de Madrid,
INFORMAN:
Que la presente memoria, titulada “Caracterización de marcadores peptídicos específicos
para la identificación de especies de langostinos de interés comercial”, que presenta el Licenciado en
Bioquímica D. Ignacio Ortea García para optar al grado de Doctor en Bioquímica, ha sido realizada bajo
su dirección en el Instituto de Investigaciones Marinas del CSIC de Vigo y reúne los requisitos
necesarios para ser defendida ante el tribunal calificador, por lo que autorizamos su presentación en la
Universidad de Santiago de Compostela.
Y para que conste a los efectos oportunos firman el presente informe en Vigo, a 24 de Julio de
2009.
Fdo. Dr. José Manuel Gallardo Abuín Fdo. Dra. M. Pilar Calo Mata
Fdo. Dr. Benito Cañas Montalvo


El trabajo de investigación que se presenta en esta memoria se ha desarrollado en el Departamento
de Tecnología de los Alimentos del Instituto de Investigaciones Marinas (IIM) de Vigo, perteneciente al
Consejo Superior de Investigaciones Científicas (IIM), y ha sido financiado por los proyectos:
“Aplicación de métodos basados en la genómica y en la proteómica a la autentificación de
especies comerciales de crustáceos pertenecientes al Orden Decapada”, concedido por el INIA
(CAL03-030-C2-2).
“Definición de marcadores moleculares específicos en especies de langostinos de interés
comercial para el sector pesquero”, concedido por la Xunta de Galicia
(PGIDIT04RMA261004PR).
Para la realización de este trabajo, D. Ignacio Ortea García ha sido beneficiario de una beca-
contrato del programa de Formación de Profesorado Universitario (FPU) del Ministerio de Educación y
Ciencia.
Los resultados obtenidos de este trabajo han dado lugar hasta la fecha de su presentación a una
patente y a cinco publicaciones científicas, además de otros dos artículos que se encuentran enviados:
• Ortea, I., Gallardo, J.M., Medina, I., Barros, L. Procedimiento y kit para la identificación de las
principales especies comerciales de langostinos y camarones. Patente de invención (nº
P200930424), Consejo Superior de Investigaciones Científicas.
• Ortea, I., Cañas, B., Calo-Mata, P., Barros-Velázquez, J., Gallardo, J.M. (2009). Arginine kinase
peptide mass fingerprinting as a proteomic approach for species identification and taxonomic
analysis of commercially-relevant shrimp species. Journal of Agricultural and Food
Chemistry, 57, 5665-5672.
• Ortea, I., Barros, L., Cañas, B., Calo-Mata, P., Barros-Velázquez, J., Gallardo, J.M. (2009). A
method to compare MALDI-TOF MS PMF and its application in Phyloproteomics. En: S.
Omatu et al. (Eds). Distributed Computing, Artificial Intelligence, Bioinformatics, Soft
Computing, and Ambient Assisted Living. Lecture Notes in Computer Science 5518, pp.
1147-1153. Springer-Verlag Berlin Heidelberg.
• Ortea, I., Barros, L., Gallardo, J.M. (2009). Closely related shrimp species identification by
MALDI-TOF Mass Spectrometry. Journal of Aquatic Food Product Technology, 18:1, 146-
155.
• Ortea, I., Barros, L., Gallardo, J.M. (2008). Secuenciación de novo de péptidos diferenciadores
del decápodo de interés comercial Pleoticus muelleri. Proteómica, 1, 55-56.

• Ortea, I., Piñeiro, C., Barros, L., Gallardo, J.M. (2006). Definición de marcadores peptídicos
específicos en especies de langostinos de interés comercial. Alimentaria, 373, 98-99.
• Ortea, I., Cañas, B., Calo-Mata, P. Barros-Velázquez, J., Gallardo, J.M. Identification of
commercial prawn and shrimp species of food interest by native isoelectric focusing. Food
Chemistry. Enviado.
• Ortea, I., Cañas, B., Gallardo, J.M. Mass Spectrometry Characterization of Species-Specific
Peptides from Arginine Kinase for the Identification of Commercially Relevant Shrimp
Species. Journal of Proteome Research. Enviado.
Del presente trabajo también se han derivado cinco comunicaciones orales y tres pósters que han
sido presentados en diferentes congresos.
Algunos de los resultados obtenidos del presente trabajo se recogen en la página web del proyecto
“Definición de marcadores moleculares específicos en especies de langostinos de interés comercial para
el sector pesquero”, (PGIDIT04RMA261004PR), www.crusgenprot.org.

Agradecimientos
Han pasado cuatro años (y pico) desde que empecé este proyecto. Por eso, ahora que este período
llega a su fin, es tiempo de echar la vista atrás y recordar a los que, con su ayuda, han contribuido a
llevarlo a buen puerto. Seguramente me olvide de muchos, pues mi memoria ya no es la que era, y
además intentaré ser breve, así que pido perdón de antemano si algún lector ávido de encontrar su
nombre en estas líneas no se siente justamente recompensado.
Le estoy profundamente agradecido a mi director de tesis y principal artífice de que este trabajo se
haya llevado a cabo, el Dr. José Manuel Gallardo Abuín, por haberme dado la oportunidad de entrar en
su proyecto, por confiar plenamente en mí, por orientarme, por enseñarme a perseverar, por motivarme,
y por supuesto por su intuición. Gracias por brindarme toda tu ayuda y por enseñarme tanto.
Quiero agradecer a mi director de tesis, el Dr. Benito Cañas Montalvo, por haberme enseñado
desde cero qué es esto que damos en llamar “Proteómica”. Seguramente no me equivoco si digo que
tengo el mejor maestro. Gracias por tu dedicación y tus consejos. Gracias por tus correcciones y tu
ingenio. Y por echarme una mano cada vez que necesito tu ayuda.
Agradecer también a mi otra directora, la Dra. Pilar Calo Mata, por dar una dimensión nueva a mi
tesis. Gracias por tu trabajo, por tu optimismo, por tranquilizarme cuando las cosas no salen todo lo bien
que esperamos, y sobre todo, gracias por tus ideas.
A mi compañera de grupo Lorena, un agradecimiento especial por su ayuda con la parte
experimental, por su dedicación, por enseñarme tantas técnicas, y por sus buenos consejos.
A mi otra compañera de grupo, y mi otra maestra, Mónica, por enseñarme, por guiarme y por
animarme continuamente.
Me gustaría agradecer al Dr. Jorge Barros su disponibilidad para ayudar en todo momento. A los
Dres. Isabel Medina y Santiago Aubourg, por su colaboración, su interés, y por haberme acogido en el
grupo de Química de Productos Marinos. Extiendo el agradecimiento a los otros jefes del departamento,
los Dres. Carmen Sotelo y Ricardo Pérez-Martín.
Un agradecimiento afectuoso para la Dra. Carmen Piñeiro, además de por orientarme en mis
primeros pasos, por su tesón a la hora de reclamar mi beca, sin el cual quién sabe dónde estaría yo ahora.
A todos mis compañeros de laboratorio y café, los que están y los que ya se fueron: Ana, María,
Jacobo, Vanesa, Rodri, Manu, Maria José, Marcos, Hugo, Gloria, Jose, Elsi, Charo, Cris, Vicky,
Maribel, Salomé, Chus, Cruz, Helena, Susana, Marta, y Bea de informática, además de a todos los que
han pasado de prácticas. Gracias por enriquecerme con vuestra compañía en el chollo, por las jornadas
deportivas, por las salidas nocturnas, y por supuesto, ¡por los ágapes!

A todo el personal del IIM, especialmente a Juan Luís, a Marcos Villafín y a Paco.
A la Dra. Concha Gil y a su grupo de la Unidad de Proteómica de la Complutense, Pilar, Montse,
Antonio y María Luisa, por acogerme y enseñarme durante esos dos meses y pico de estancia, por
vuestro cariño y por lo mucho que me reía en el café con las anécdotas de Antonio.
A los Dres. Jesús Vázquez y Anabel Marina, del Servicio de Proteómica del C.B.M.S.O., por
acogerme durante mi primera y breve estancia formativa en Madrid.
Gracias a la Dra. Vanesa Díaz, antes en la Unidad de Espectrometría de Masas de la Universidad
de Oviedo, por su ayuda con el MALDI-TOF. A Estefanía, del dpto. de Química Analítica de la
Complutense, por su ayuda con las digestiones con ultrasonidos. A Ananías, de la USC, por su trabajo
con el ADN. Y a Karola, de la USC, por compartir la búsqueda de software filoproteómico.
Gracias a los de Gijón que vinieron a visitarme a Vigo: Juan y Silvia –en su luna de miel-, Omar,
Manuel, Campo, Gon y Santi. Y a Víctor por ir a visitarme a Madrid. A Marco, Gon y David H., por
acogerme en sus casas en Madrid en varias ocasiones.
Y un millón de gracias a Eva. No solo por los geles. Gracias por tu apoyo, tu paciencia y tu
cariño. Gracias por escucharme y por aguantarme. Gracias por animarme continuamente. Gracias por
estar ahí.

A mis padres,
sin ellos no habría llegado hasta aquí.


ÍNDICE
ÍNDICE .................................................................................................................................................... 1
ABREVIATURAS ................................................................................................................................... 5
INTRODUCCIÓN ................................................................................................................................ 9
1. VISIÓN GLOBAL DEL MERCADO DE LOS PRODUCTOS PESQUEROS........................... 11
2. LANGOSTINOS Y GAMBAS ......................................................................................................... 12
2.1. Sistemática .................................................................................................................................. 12
2.2. Biología........................................................................................................................................ 13
2.3. Importancia económica ............................................................................................................. 14
3. AUTENTICIDAD Y CONTROL DE ALIMENTOS DE ORIGEN MARINO ........................... 17
4. MÉTODOS DE IDENTIFICACIÓN DE ESPECIES EN PRODUCTOS
PESQUEROS......................................................................................................................................... 21
4.1. Métodos de identificación basados en el análisis del ADN ..................................................... 22
4.2. Métodos de identificación basados en el análisis de proteínas ............................................... 26
5. PROTEÓMICA ................................................................................................................................. 28
5.1. Separación de proteínas y péptidos .......................................................................................... 30
5.1.1. Técnicas electroforéticas ...................................................................................................... 30
5.1.1.1. SDS-PAGE .................................................................................................................... 30
5.1.1.2. Isoelectroenfoque .......................................................................................................... 30
5.1.1.3. 2-DE .............................................................................................................................. 31
5.1.2. Técnicas cromatográficas ..................................................................................................... 31
5.2. Espectrometría de masas ........................................................................................................... 32
5.2.1. MALDI-TOF ........................................................................................................................ 34
5.2.2. ESI-IT ................................................................................................................................... 38
5.3. Espectrometría de masas en tándem (MS/MS) ....................................................................... 41
5.3.1. Secuenciación de péptidos mediante MS/MS....................................................................... 43
5.4. Identificación de proteínas y péptidos...................................................................................... 46
5.4.1. Huella peptídica (PMF) ........................................................................................................ 46
5.4.2. Fragmentación de péptidos ................................................................................................... 47
5.5. Bioinformática ............................................................................................................................ 49
5.6. Proteómica y especies marinas.................................................................................................. 50
OBJETIVOS ........................................................................................................................................ 53
MATERIALES Y MÉTODOS ........................................................................................................ 57
1. Material biológico.............................................................................................................................. 59
2. Análisis taxonómico de ejemplares de referencia ........................................................................... 64
1

Índice
2.1. Identificación morfológica y obtención de secuencias de referencia........................................... 64
2.2. Identificación genética ................................................................................................................. 65
3. Extracción de las proteínas sarcoplásmicas .................................................................................... 65
4. Isoelectroenfoque nativo ................................................................................................................... 66
5. Electroforesis bidimensional............................................................................................................. 67
6. Digestión de las proteínas en gel....................................................................................................... 68
7. Mapeo peptídico mediante espectrometría de masas MALDI-TOF............................................. 69
7.1. Obtención de PMF ....................................................................................................................... 69
7.2. Procesamiento de los espectros de MALDI-TOF ........................................................................ 70
8. Análisis filogenético de los espectros PMF ...................................................................................... 70
9. Análisis de ADN................................................................................................................................. 71
9.1. Extracción del ADN..................................................................................................................... 71
9.2. Amplificación del ADN ............................................................................................................... 72
9.3. Secuenciación............................................................................................................................... 73
9.4. Análisis filogenético..................................................................................................................... 73
10. Caracterización de péptidos mediante espectrometría de masas en tándem ............................. 73
10.1. LC-ESI-IT-MS/MS .................................................................................................................... 73
10.2. nESI-IT-MS/MS......................................................................................................................... 75
10.3. Procesamiento de los datos de MS/MS...................................................................................... 75
11. Desarrollo de un procedimiento de análisis rápido por MS para la identificación
de las principales especies comerciales de langostinos ....................................................................... 76
11.1. Digestión en solución tradicional............................................................................................... 76
11.2. Digestión en solución ultrarrápida con HIFU ............................................................................ 76
11.3. Separación y monitorización de péptidos mediante SMIM ....................................................... 77
RESULTADOS Y DISCUSIÓN ...................................................................................................... 79
Capítulo 1: Identificación de especies de langostinos y gambas de interés alimentario por
isoelectroenfoque ................................................................................................................................ 81
Capítulo 2: Identificación de especies de langostinos próximas por MALDI-TOF
MS..................................................................................................................................................... 109
Capítulo 3: Huella peptídica de arginina quinasa, enfoque proteómico para la identificación y análisis
taxonómico de especies de langostinos de interés comercial............................................................ 125
Capítulo 4: Caracterización por espectrometría de masas de péptidos de arginina quinasa específicos
para especies de langostinos comerciales.......................................................................................... 147
Capítulo 5: Procedimiento y kit para la identificación rápida de las principales especies comerciales
de langostinos y camarones............................................................................................................... 189
DISCUSIÓN....................................................................................................................................... 251
2

Índice
CONCLUSIONES ............................................................................................................................ 259
BIBLIOGRAFÍA .............................................................................................................................. 263
3


ABREVIATURAS
2-DE: Electroforesis bidimensional
ACN: Acetonitrilo
ADN: Ácido desoxirribonucleico
ADNmt: ADN mitocondrial
ADNn: ADN nuclear
AFLP: Polimorfismos de longitud de fragmentos amplificados (Amplified Fragment Lenght
Polymorphism)
AK: Arginina quinasa (Arginine Kinase)
amu: Unidad de masa atómica (atomic mass unit)
APS: Persulfato amónico
BLAST: Basic Local Alignment Search Tool
BSA: Albúmina de suero bovino
C-terminal: Carboxilo-terminal
CHCA: Ácido α-ciano-4-hidroxicinámico
CID: Disociación inducida por colisión (Collision Induced dissociation)
cyt b: Citocromo b
CZE: Capillary Zone Electrophoresis
Da: Dalton (unidad de masa atómica)
DHB: Ácido 2,5-dihidroxibenzoico
dNTP: Deoxirribonucleótido trifosfato
DTT: Ditiotreitol
ELISA: Enzyme-linked immunosorbent assay
ESI: Ionización por electrospray (Electrospray Ionization)
FINS: Forensically informative nucleotide sequencing
FWHM: Anchura de pico a media altura (Full Width at Half Maximum)
HIFU: Ultrasonidos focalizados de alta intensidad (High Intensity Focused Ultrasound)
HPLC: Cromatografía líquida de alta eficacia (High Performance Liquid Chromatography)
ICR: Resonancia ciclotrónica de iones (Ion Ciclotron Resonance)
IEF: Isoelectroenfoque
IPG: Gradiente inmobilizado de pH (Immobilized pH Gradient)
IT: Trampa iónica (Ion Trap)
LC: Cromatografía líquida (Liquid Chromatography)
LC-MS: Cromatografía líquida acoplada a espectrometría de masas
LHICA: Laboratorio de Higiene, Inspección y Control de Alimentos
5

Abreviaturas
LIT: Trampa iónica lineal (Linear Ion Trap)
MALDI: Ionización/desorción por láser asistida por matriz (Matriz Assisted Laser
Desorption/Ionization)
microESI: Microelectrospray o microspray
Mr: Masa molecular relativa
MS: Espectrometría de masas (Mass Spectrometry)
MS/MS: Espectrometría de masas en tándem (Tandem Mass Spectrometry)
MSn: Espectrometría de masas en tándem múltiple
m/z: Relación masa/carga
N-terminal: Amino-terminal
NCBI: National Center for Biotechnology Information
NCBInr: Base de datos no redundante del NCBI
nESI: Nanoelectrospray o nanospray
PCR: Reacción en cadena de la polimerasa (Polimerase Chain Reaction)
PFF: Huella de fragmentación peptídica (Peptide Fragment Fingerprinting)
pI: Punto isoeléctrico
PMF: Mapa o huella de masas peptídicas (Peptide Mass Fingerprinting)
PMSF: Fluoruro de fenil-metil-sulfonilo
ppm: Partes por millón
Q: Cuadrupolo
RAPD: ADN polimórfico amplificado al azar (Randomly Amplified Polymorphic DNA)
RF: Radiofrecuencia
RFLP: Polimorfismos de longitud de los fragmentos de restricción (Restriction Fragment
Lenght Polymorphism)
RP: Fase reversa (Reverse Phase)
SCP: Proteína sarcoplásmica de unión a calcio (Sarcoplasmic Calcium-binding Protein)
SCX: Intercambio catiónico fuerte (Strong Cation Exchange)
SDS: Dodecil sulfato sódico (Sodium Dodecyl Sulfate)
SDS-PAGE: Electroforesis en gel de poliacrilamida con SDS (SDS Polyacrylamide Gel
Electrophoresis)
SMIM: Monitorización de la fragmentación de un ión seleccionado (Selected MS/MS Ion
Monitoring)
SSCP: Polimorfismos de conformación de cadena sencilla (Single-Strand Conformational
Polymorphism)
TEMED: N’N’N’N’-tetra-metil-etilendiamina
TFA: Ácido trifluoroacético
6

Abreviaturas
Th: Thomson, unidad de m/z
TOF: Tiempo de vuelo (Time of Flight)
7


INTRODUCCIÓN

Introducción
1. VISIÓN GLOBAL DEL MERCADO DE LOS PRODUCTOS
PESQUEROS
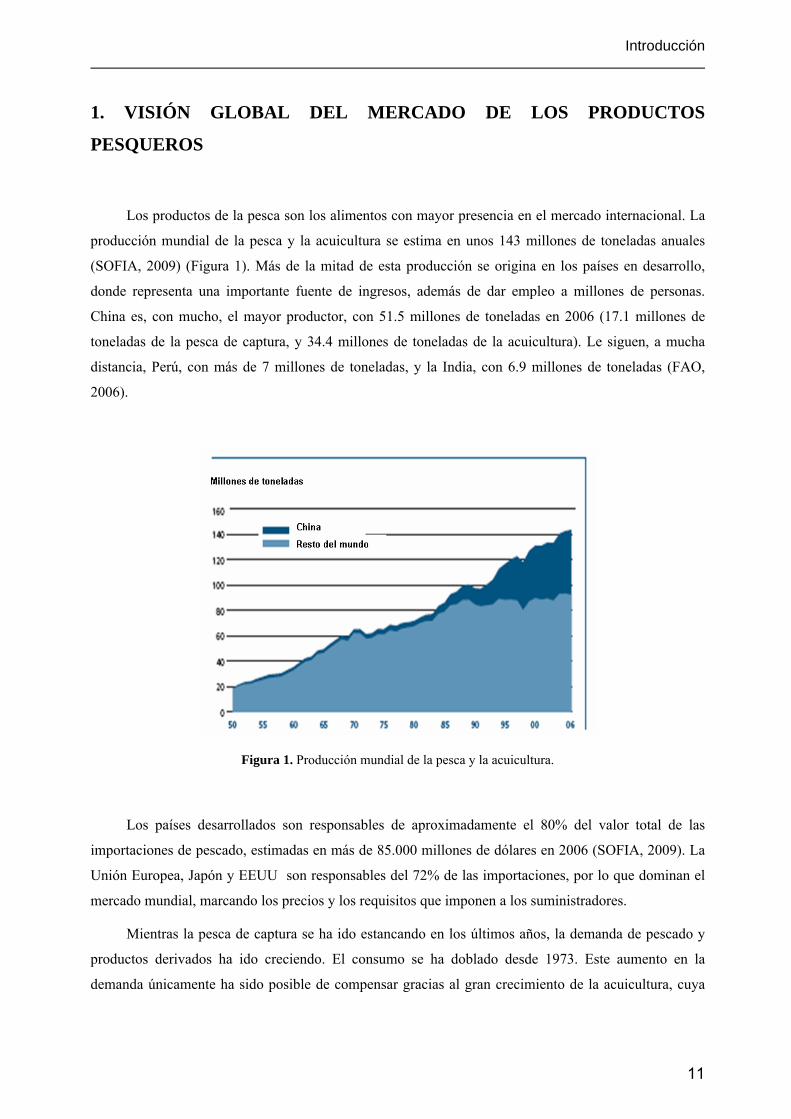
Los productos de la pesca son los alimentos con mayor presencia en el mercado internacional. La
producción mundial de la pesca y la acuicultura se estima en unos 143 millones de toneladas anuales
(SOFIA, 2009) (Figura 1). Más de la mitad de esta producción se origina en los países en desarrollo,
donde representa una importante fuente de ingresos, además de dar empleo a millones de personas.
China es, con mucho, el mayor productor, con 51.5 millones de toneladas en 2006 (17.1 millones de
toneladas de la pesca de captura, y 34.4 millones de toneladas de la acuicultura). Le siguen, a mucha
distancia, Perú, con más de 7 millones de toneladas, y la India, con 6.9 millones de toneladas (FAO,
2006).
Figura 1. Producción mundial de la pesca y la acuicultura.
Los países desarrollados son responsables de aproximadamente el 80% del valor total de las
importaciones de pescado, estimadas en más de 85.000 millones de dólares en 2006 (SOFIA, 2009). La
Unión Europea, Japón y EEUU son responsables del 72% de las importaciones, por lo que dominan el
mercado mundial, marcando los precios y los requisitos que imponen a los suministradores.
Mientras la pesca de captura se ha ido estancando en los últimos años, la demanda de pescado y
productos derivados ha ido creciendo. El consumo se ha doblado desde 1973. Este aumento en la
demanda únicamente ha sido posible de compensar gracias al gran crecimiento de la acuicultura, cuya
11

Introducción
producción, estimada en unas 51.7 millones de toneladas en 2006 (el 67% de ellas, de China), representa
el 36% de la producción de pescado total, frente al 3.9% que representaba en 1970 (SOFIA, 2009).
En lo referente a la utilización de los productos pesqueros, de las 140 millones de toneladas
obtenidas en el año 2006, el 77% fueron dedicadas al consumo humano directo, lo que equivale a un
consumo per cápita aparente de unos 16.7 kg/año (SOFIA, 2009). El resto se destinó principalmente a la
fabricación de harinas y aceites de pescado. El consumo medio en la Unión Europea se estimó para el
período 2003-2005 en 25.7 kg/año, y para España en 42.6 kg/año.
2. LANGOSTINOS Y GAMBAS
2.1. SISTEMÁTICA
Los crustáceos (Crustacea Brünnich, 1772), un subphylum de los artrópodos, son un grupo de
inmensa riqueza taxonómica y diversidad morfológica. Cuenta con unas 52.000 especies conocidas,
aunque el número real podría ser 10 veces mayor (Martin y Davis, 2001; Bowman y Abele, 1982).
Dentro de los crustáceos, el orden Decapoda Latreille, 1802 constituye el grupo más diversificado,
comprendiendo más de 10.000 especies descritas, agrupadas en unas 120 familias y 1.200 géneros
(Bowman y Abele, 1982). Los decápodos, entre los que se incluyen gambas, langostinos, cangrejos y
langostas, son reconocidos como los crustáceos más típicos y de mayor valor comercial (Yamauchi,
2004).
Dentro del orden Decapoda, el suborden Dendrobranchiata Bate, 1888 incluye la mayoría de las
gambas y langostinos comerciales (Figura 2), siendo la superfamilia Penaeoidea Rafinesque, 1815 la de
mayor importancia económica, ya que representa aproximadamente el 70% de la producción mundial,
fundamentalmente debido al género Penaeus. La referencia estándar para la clasificación de este grupo
de decápodos (el suborden Dendrobranchiata) es, actualmente, el estudio taxonómico de Pérez-Farfante
y Kensley (1997), en el que se recogen los 56 géneros y 500 especies que lo componen.
Otro grupo importante de gambas, tras las penaeoideas, son las carideas (infraorden Caridea Dana,
1852), responsables de aproximadamente otro 18% de la producción, fundamentalmente debido al
género boreal Pandalus.
12

Introducción
Figura 2. Clasificación de los principales grupos de gambas y langostinos de importancia económica.
El grupo más importante, bajo el punto de vista económico (Lavery y col., 2004), es el género
Penaeus sensu lato Fabricius, 1978. Este género constituye un conjunto problemático de especies, sobre
las que todavía no existe consenso sobre sus relaciones filogenéticas. Los seis grupos morfológicos que
anteriormente se trataban como subgéneros fueron elevados a la categoría de géneros por Pérez-Farfante
y Kensley (1997) en la revisión más reciente de la taxonomía de los Penaeoidea. Este cambio, según
Lavery y col. (2004), puede resultar prematuro, por lo que es necesario realizar estudios más profundos.
2.2. BIOLOGÍA
Los langostinos y gambas presentan un alto grado de diversidad en cuanto al hábitat que ocupan.
Así, aunque la gran mayoría son marinos, unas pocas familias han llegado a colonizar hábitats salobres o
incluso de agua dulce. En general habitan en aguas litorales a poca profundidad, distribuyéndose desde
la misma playa hasta unos 100 metros de profundidad, aunque algunas especies pueden ser bastante más
profundas (Sardà, 1993). El ciclo reproductivo es diverso y complejo e incluye aspectos migratorios
locales entre mar abierto y zonas litorales más o menos cerradas, tales como estuarios, bahías y lagunas
(Figura 3). La puesta tiene lugar en mar abierto y las larvas, entre 10 y 12 fases de vida pelágica
(planctónica), se acercan a la costa. Las postlarvas, todavía de vida pelágica, penetran en las zonas
lagunares o quedan cerca de la costa, donde cubren su fase juvenil para retornar al mar a finales de dicha
fase o en las primeras de adulto. Los adultos son bentónicos, necesitando un sustrato fangoso o arenoso
donde enterrarse durante el día. La alimentación suele ser crepuscular o nocturna, depredando sobre
13

Introducción
cualquier tipo de alimento a su alcance, normalmente sobre otros crustáceos y pequeños moluscos, como
bivalvos, que capturan por rastreo del sustrato.
Figura 3. Ciclo biológico del langostino.
2.3. IMPORTANCIA ECONÓMICA
Los crustáceos, incluyendo los langostinos y gambas, han sido una fuente de alimento para los
humanos desde la antigüedad. Ya se encuentran referencias a las gambas en las culturas china y
japonesa antiguas. La primera referencia conocida de las gambas penaeoideas en la literatura occidental
es la de la especie Mellicertus kerathurus, nombrada como “Squilla crangone” por Guillaume Rondelet
en 1554 en su Libri de Piscibus Marinis (Rondelet, 1554) (Figura 4).
14

Introducción
Figura 4. Página 547 del Libri de Piscibus Marinis, en la que se
recoge la primera referencia conocida a una gamba penaeoidea.
En la actualidad, dentro del mercado de los productos pesqueros, las gambas y langostinos son el
producto más importante en valor económico, representando el 17% del total del mercado internacional
de productos pesqueros en 2006 (SOFIA, 2009). Le siguen los salmónidos (11%), los pescados de fondo
(10%, merluza, bacalao, abadejo), y los túnidos (8%). La producción mundial de langostinos y gambas
en el año 2006 fue de unos 6.5 millones de toneladas, equivalentes a un valor de más de 24.000 millones
de dólares. (FAO, 2006). Esta producción se ha incrementado en los últimos años, coincidiendo con la
gran expansión de la acuicultura de este grupo, que ha crecido muy rápidamente desde 1997 (creció un
340% en peso y un 205% en valor en el periodo 1997-2006). En el 2006, más del 47% (unos 3.1
millones de toneladas) de la producción total de langostinos y gambas fue de acuicultura.
En el año 2006, las principales especies comercializadas dentro de este grupo fueron Litopenaeus
vannamei (2.133.000 t, 7.774M dolares) y Penaeus monodon (658.000 t, 3.122M dolares) en cultivo, y
Pandalus borealis (388.000 t) y P. monodon (244.000 t) en captura (FIGIS FAO, 2006).
15
Introducción
España es el tercer mayor importador de langostinos y gambas del mundo, tras EEUU y Japón, y
el primero de la UE, seguido por Francia. El descenso en las capturas de nuestra flota por un lado, y el
incremento en la demanda y en la producción de nuestra industria de transformación por otro, han ido
derivando hacia una dependencia creciente del abastecimiento exterior de materia prima (Libro Blanco
de la Pesca, MAPA). Así, en 2006 España importó una cantidad record de 179.500 toneladas de gambas
(Tabla 1), lo que supone un 10% en peso y un 18% en valor del total de las importaciones españolas de
productos pesqueros (FISH INFOnetwork). Estas cifras suponen un aumento del 15% respecto al 2005,
siguiendo la tendencia creciente de los años anteriores.
Tabla 1. Importaciones de langostinos en España en los últimos años.
Importaciones (miles de toneladas) Importaciones (millones de euros)
2001 2002 2003 2004 2005 2006 2001 2002 2003 2004 2005
133 128 143 145 156 179
883 769 859 790 845
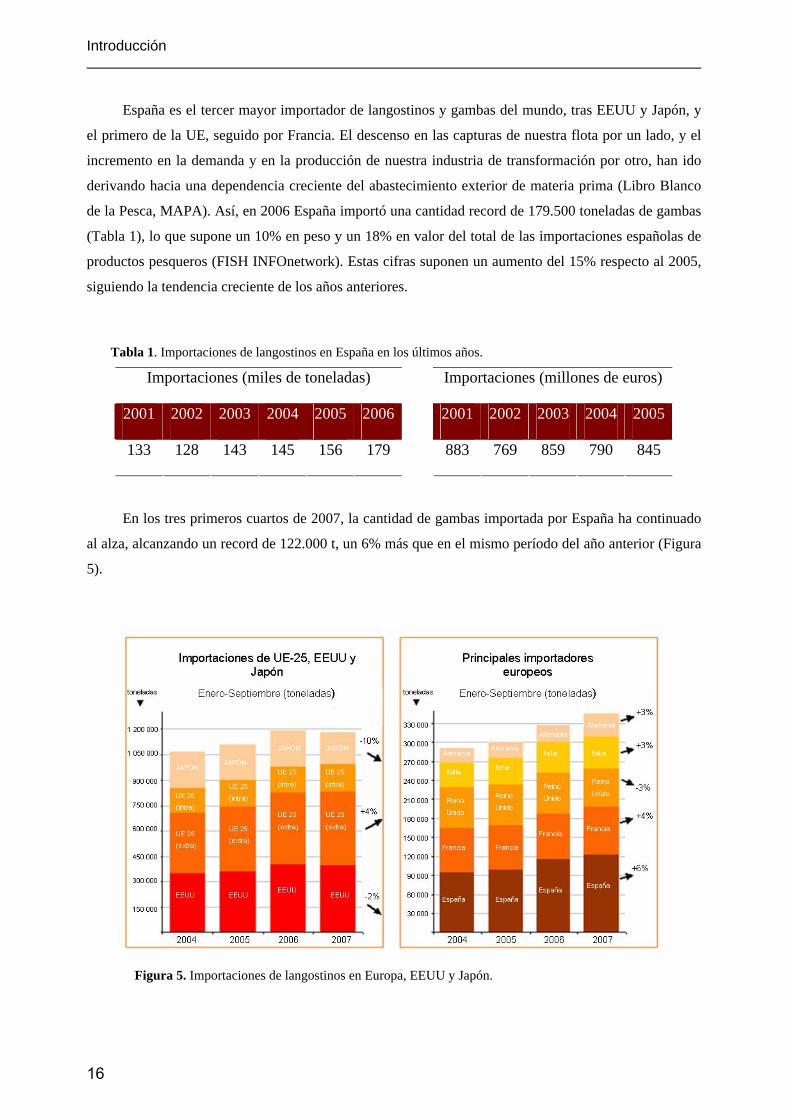
En los tres primeros cuartos de 2007, la cantidad de gambas importada por España ha continuado
al alza, alcanzando un record de 122.000 t, un 6% más que en el mismo período del año anterior (Figura
5).
Figura 5. Importaciones de langostinos en Europa, EEUU y Japón.
16
Introducción
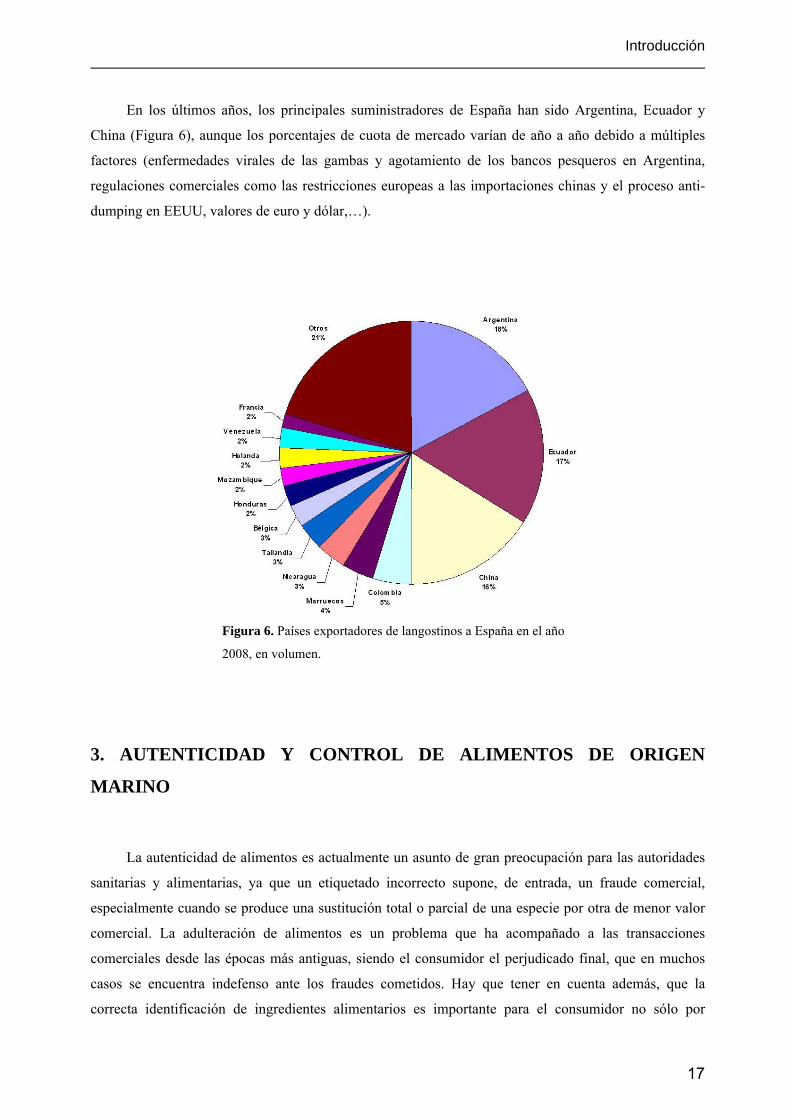
En los últimos años, los principales suministradores de España han sido Argentina, Ecuador y
China (Figura 6), aunque los porcentajes de cuota de mercado varían de año a año debido a múltiples
factores (enfermedades virales de las gambas y agotamiento de los bancos pesqueros en Argentina,
regulaciones comerciales como las restricciones europeas a las importaciones chinas y el proceso anti-
dumping en EEUU, valores de euro y dólar,…).
Figura 6. Países exportadores de langostinos a España en el año
2008, en volumen.
3. AUTENTICIDAD Y CONTROL DE ALIMENTOS DE ORIGEN
MARINO
La autenticidad de alimentos es actualmente un asunto de gran preocupación para las autoridades
sanitarias y alimentarias, ya que un etiquetado incorrecto supone, de entrada, un fraude comercial,
especialmente cuando se produce una sustitución total o parcial de una especie por otra de menor valor
comercial. La adulteración de alimentos es un problema que ha acompañado a las transacciones
comerciales desde las épocas más antiguas, siendo el consumidor el perjudicado final, que en muchos
casos se encuentra indefenso ante los fraudes cometidos. Hay que tener en cuenta además, que la
correcta identificación de ingredientes alimentarios es importante para el consumidor no sólo por
17

Introducción
razones económicas, sino también por razones sanitarias, debido a la posible introducción inadvertida de
componentes alergénicos, tóxicos o que puedan alterar los hábitos dietéticos de algún sector de la
población. Hay que señalar que los crustáceos son uno de los grupos de alimentos que producen alergias
alimentarias con mayor prevalencia (Sampson, 2004). Estas reacciones alérgicas pueden ser provocadas
por trazas de la proteína alergénica en el alimento, y provocan alteraciones gastrointestinales, cutáneas y
respiratorias, llegando a producir reacciones potencialmente mortales, como el shock anafiláctico
(Sicherer y col., 2004).
La adulteración de alimentos no sólo perjudica al consumidor final, sino también a la industria
transformadora, que puede verse obligada a comprar materia prima de calidad inferior a la declarada, lo
que conlleva pérdida económica, disminución de la calidad del producto, e insatisfacción del cliente. Por
ello, la industria transformadora debe establecer los métodos más adecuados para controlar las materias
primas recibidas de los proveedores.
En los productos pesqueros, la identificación de las diferentes especies es de gran importancia, ya
que en muchas ocasiones las características morfológicas externas han sido eliminadas. Además, cuando
la relación filogenética entre las especies es muy próxima, mayor es la similitud morfológica, por lo que
difícilmente se logra la identificación. La globalización del mercado de la pesca debida al desarrollo de
las tecnologías de conservación, elaboración y distribución de alimentos, sumado a la entrada de nuevas
especies como consecuencia de la búsqueda de nuevos caladeros, ha provocado que en la actualidad se
comercialicen gran cantidad de especies provenientes de todo el mundo, lo que exige un riguroso control
del origen.
El correcto etiquetado de los alimentos es fundamental, ya que supone la primera información que
se le ofrece al consumidor, por lo que ha de ser comprensible, preciso y verdadero. Para asegurar el
control del origen frente a sustituciones de especies que puedan originar fraudes al consumidor, la UE ha
establecido la directiva 2000/13/CE, relativa a la aproximación de las legislaciones de los Estados
miembros en materia de etiquetado, presentación y publicidad de los productos alimenticios, que fue
modificada por la Directiva 2003/89/CE, que especifica que el etiquetado de los productos alimenticios
no debe inducir a error al comprador sobre sus características, en particular sobre su naturaleza,
identidad, cualidades, composición, cantidad, duración, origen o procedencia y modo de fabricación u
obtención. En España, el Real Decreto 1380/2002, de 20 de diciembre (modificado por el Real Decreto
1702/2004), y el Real Decreto 121/2004, de 23 de enero, regulan la identificación y el etiquetado de los
productos de la pesca, la acuicultura y el marisqueo, congelados o ultracongelados el primero, y vivos,
frescos, refrigerados o cocidos el segundo. En esta legislación se establece que dichos productos deberán
llevar en el envase y/o embalaje correspondiente en lugar bien visible una etiqueta en la que se
contemplará, en caracteres legibles e indelebles, las siguientes especificaciones:
a) Denominación comercial y científica de la especie.
18

Introducción
b) Método de producción:
- Pesca extractiva o pescado.
- Pescado en aguas dulces.
- Criado o acuicultura.
- Marisqueo.
c) Zona de captura (Tabla 2) o de cría. Para los productos pescados en aguas dulces y para los
productos cultivados, se indicará el país donde se haya pescado o cultivado.
Tabla 2. Zonas de captura para pesca extractiva recogidas en el RD 1380/2002.
Zona de captura Definición de la zona
Atlántico Noroeste Zona FAO nº. 21 Atlántico Noreste Zona FAO nº. 27 Mar Báltico Zona FAO nº- 27, IIId Atlántico Centro-Oeste Zona FAO nº. 31 Atlántico Centro-Este Zona FAO nº. 34 Atlántico Suroeste Zona FAO nº. 41 Atlántico Sureste Zona FAO nº. 47 Mar Mediterráneo Zona FAO nº. 37.1, 37.2 y 37.3 Mar Negro Zona FAO nº. 37.4 Océano Índico Zona FAO nº. 51 y 57 Océano Pacífico Zona FAO nº. 61, 67, 71, 77, 81 y 87 Antártico Zona FAO nº. 48, 58 y 88
Además, a los efectos de poder conocer la trazabilidad de un producto, estas especificaciones
deberán estar disponibles en cada fase de comercialización del producto, desde la primera venta hasta el
consumidor final, incluyendo el transporte y la distribución.
La denominación comercial y científica de la especie a que se refieren dichas especificaciones,
deberá ser la establecida por Resolución de la Secretaría General del Mar. La Resolución de 4 de marzo
de 2009, de la Secretaría General del Mar, establece el listado de denominaciones comerciales de
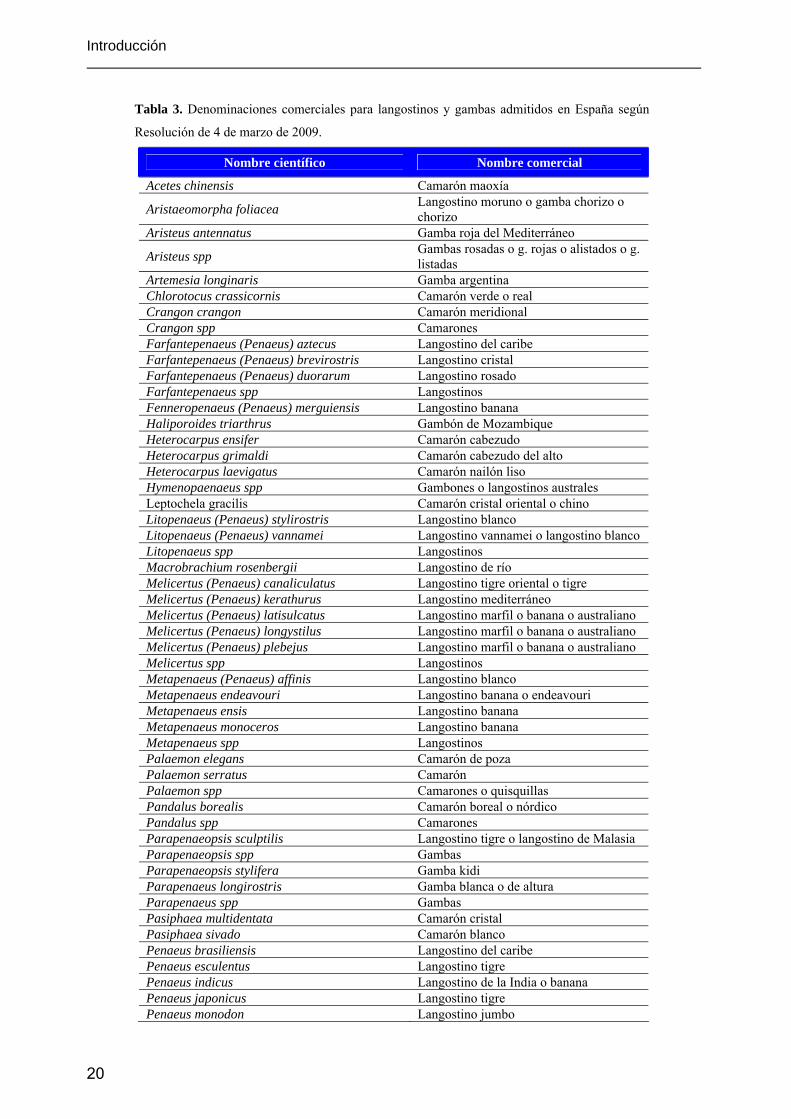
especies pesqueras y de acuicultura admitidas en España. En la tabla 3 se recogen las denominaciones
comerciales para especies de langostinos y gambas admitidos en España según dicha Resolución.
19
Introducción
Tabla 3. Denominaciones comerciales para langostinos y gambas admitidos en España según
Resolución de 4 de marzo de 2009.
Nombre científico Nombre comercial
Acetes chinensis Camarón maoxía
Aristaeomorpha foliacea Langostino moruno o gamba chorizo o chorizo
Aristeus antennatus Gamba roja del Mediterráneo
Aristeus spp Gambas rosadas o g. rojas o alistados o g. listadas
Artemesia longinaris Gamba argentina Chlorotocus crassicornis Camarón verde o real Crangon crangon Camarón meridional Crangon spp Camarones Farfantepenaeus (Penaeus) aztecus Langostino del caribe Farfantepenaeus (Penaeus) brevirostris Langostino cristal Farfantepenaeus (Penaeus) duorarum Langostino rosado Farfantepenaeus spp Langostinos Fenneropenaeus (Penaeus) merguiensis Langostino banana Haliporoides triarthrus Gambón de Mozambique Heterocarpus ensifer Camarón cabezudo Heterocarpus grimaldi Camarón cabezudo del alto Heterocarpus laevigatus Camarón nailón liso Hymenopaenaeus spp Gambones o langostinos australes Leptochela gracilis Camarón cristal oriental o chino Litopenaeus (Penaeus) stylirostris Langostino blanco Litopenaeus (Penaeus) vannamei Langostino vannamei o langostino blanco Litopenaeus spp Langostinos Macrobrachium rosenbergii Langostino de río Melicertus (Penaeus) canaliculatus Langostino tigre oriental o tigre Melicertus (Penaeus) kerathurus Langostino mediterráneo Melicertus (Penaeus) latisulcatus Langostino marfil o banana o australiano Melicertus (Penaeus) longystilus Langostino marfil o banana o australiano Melicertus (Penaeus) plebejus Langostino marfil o banana o australiano Melicertus spp Langostinos Metapenaeus (Penaeus) affinis Langostino blanco Metapenaeus endeavouri Langostino banana o endeavouri Metapenaeus ensis Langostino banana Metapenaeus monoceros Langostino banana Metapenaeus spp Langostinos Palaemon elegans Camarón de poza Palaemon serratus Camarón Palaemon spp Camarones o quisquillas Pandalus borealis Camarón boreal o nórdico Pandalus spp Camarones Parapenaeopsis sculptilis Langostino tigre o langostino de Malasia Parapenaeopsis spp Gambas Parapenaeopsis stylifera Gamba kidi Parapenaeus longirostris Gamba blanca o de altura Parapenaeus spp Gambas Pasiphaea multidentata Camarón cristal Pasiphaea sivado Camarón blanco Penaeus brasiliensis Langostino del caribe Penaeus esculentus Langostino tigre Penaeus indicus Langostino de la India o banana Penaeus japonicus Langostino tigre Penaeus monodon Langostino jumbo
20

Introducción
Tabla 3. Continuación.
Nombre científico Nombre comercial Penaeus notialis Langostino blanco Penaeus semisulcatus Langostino banana Penaeus spp Langostinos Pleoticus muelleri Gambón o langostino austral Pleoticus spp Gambones o langostinos australes Plesionika edwardsii Camarón soldado Plesionika heterocarpus Camarón flecha Plesionika martia Camarón marcial Plesionika narval Camarón narval Plesionika spp Camarones Plesionika williansi Camarón rayado de guinea Plesiopenaeus edwardsianus Carabinero Pleuroncodes monodon Sastre rojo Pontophilus spinosus Camarón espinoso Processa edulis Camaroncillo Processa spp Camarones cabezones Solenocera melantho Gamba cabezón Solenocera membranacea Gamba del Atlántico Solenocera spp Gambas Trachypenaeus spp. Gambas
Se observa que existe gran diversidad de especies comerciales de langostinos y gambas,
destacando que bajo los términos “langostino banana”, “langostino blanco” o “langostino tigre” se
encuadran distintas especies, e incluso distintos géneros.
La autenticidad alimentaria tiene actualmente gran importancia en el sector alimentario, debido a
que la escasez de especies de elevado valor comercial incita a las sustituciones fraudulentas por otras de
menor valor, y en mayor medida cuando se trata de productos transformados o elaborados, en los que las
características externas han sido eliminadas. Por ello, es necesario disponer de metodologías analíticas
adecuadas para identificar los componentes presentes en los alimentos, asegurando el cumplimiento de
la legislación y de los requisitos impuestos por la industria y los consumidores.
4. MÉTODOS DE IDENTIFICACIÓN DE ESPECIES EN PRODUCTOS
PESQUEROS
Farfante y Kensley (1997) proporcionan las claves para identificar morfológicamente las cerca de
500 especies de gambas penaeoideas y sergestoideas. Sin embargo, debido a la gran similitud
morfológica que se observa cuando la relación filogenética entre las especies es muy próxima, llegar a
una correcta identificación es en ocasiones muy complicado, y solamente taxónomos expertos son
21

Introducción
capaces de identificar estas especies mediante observación directa. Además, una vez procesado el
producto, muchas de las características anatómicas son eliminadas, por lo que la identificación
fenotípica se hace imposible (Civera y Parisi, 1991; Mafra y col., 2008). Por ello es necesario recurrir a
técnicas basadas en el análisis molecular.
Los marcadores moleculares más utilizados para la identificación de las especies pesqueras
presentes en los alimentos son los ácidos nucleicos y las proteínas. Los estudios publicados utilizando el
análisis de los lípidos son escasos (Medina y col., 1995; Joensen y Grahl-Nielsen, 2000; Medina y col.,
2003), ya que la composición lipídica varía en función de muchos otros factores además de la especie,
como son el tipo de alimentación, la etapa de crecimiento, la temperatura ambiental, la salinidad, etc.
(Joensen y col., 2000). El análisis de la composición lipídica sí ha demostrado ser más útil para
distinguir entre pescado salvaje y de acuicultura (Chen y col., 1995; Tritt y col., 2005).
4.1. MÉTODOS DE IDENTIFICACIÓN BASADOS EN EL ANÁLISIS DEL ADN
La identificación genética de especies se basa en la existencia de polimorfismos en el ácido
desoxirribonucleico (ADN), es decir, variaciones genéticas que tienen lugar como resultado de
mutaciones en el código genético. De los distintos marcadores de ADN utilizados para la identificación
de especies, el ADN mitocondrial (ADNmt) tiene varias ventajas sobre el ADN nuclear (ADNn). El
ADNmt es más pequeño y más simple que el ADNn (no tiene intrones, pseudogenes, ADN repetitivo ni
transposones); es relativamente fácil de extraer; no presenta reorganizaciones genéticas como
recombinación; al ser de herencia materna los individuos sólo tienen un alelo, evitándose ambigüedades
en la secuencia debido a heterocigosis; tiene un número de copias más alto; y, finalmente, tiene una tasa
de mutación más alta, por lo que se acumulan las suficiente mutaciones para discriminar incluso entre
especies muy próximas. Esta última ventaja también conlleva un inconveniente, y es que el ADNmt
presenta un grado de variabilidad intraespecífica que hay que tener en cuenta.
Dentro del ADNmt, el gen más comúnmente utilizado en estudios de identificación de especies ha
sido el del citocromo b (cyt b) (Rasmussen y Morrissey, 2008). Otros marcadores de ADNmt utilizados
en identificación de especies son el gen 12S rRNA y el gen 16S rRNA. Hay además otros marcadores de
ADNmt, menos empleados en la identificación de especies marinas, como la región de control
mitocondrial o los genes de la citocromo oxidasa III y la citocromo oxidasa I.
Para detectar estos polimorfismos genéticos especie-específicos, la mayoría de los trabajos se
centran en el uso de la reacción en cadena de la polimerasa (PCR) (Mullis y col., 1986). El ADN es
extraído del organismo bajo estudio, y el fragmento de ADN de interés es amplificado por PCR. Esta
técnica consiste en múltiples ciclos en los que se copia una cadena de ADN que actúa como molde, a
22

Introducción
partir de oligonucleótidos sintéticos que flanquean ese ADN molde, uniéndose a él y actuando como
primers o cebadores. El resultado es la síntesis de millones de copias del fragmento de ADN. Estos
productos de la amplificación, o amplicones, pueden entonces ser analizados para encontrar los
polimorfismos característicos utilizando gran variedad de métodos, como secuenciación con mapeo
filogenético o FINS (forensically informative nucleotide sequencing), multiplex-PCR, polimorfismos de
longitud de los fragmentos de restricción (RFLP), polimorfismos de conformación de cadena sencilla
(SSCP), ADN polimórfico amplificado al azar (RAPD), polimorfismos de longitud de fragmentos
amplificados (AFLP), PCR en tiempo real, y PCR lab-on-a-chip.
En la tabla 4 se recoge un compendio de estudios de identificación de especies pesqueras en los
que se han utilizado métodos genéticos.
A pesar de la gran cantidad de técnicas disponibles, la mayoría de los estudios de identificación de
especies pesqueras basados en ADN han sido llevados a cabo o bien utilizando RFLP o bien mediante la
secuenciación de fragmentos de ADNmt (fundamentalmente del gen cyt b), previa amplificación por
PCR.
El gen 16S rRNA y el gen de la citocromo oxidasa I, del ADNmt, también han sido descritos
como buenos marcadores entre algunas especies de crustáceos, aunque la mayoría de estos estudios se
centran en estudios de poblaciones, filogeografía y relaciones filogenéticas (Maggioni y col., 2001;
Baldwin y col., 1998; Quan y col., 2004; Lavery y col., 2004; Voloch y col., 2005), no en diferenciación
de especies. Brzezinski (2005) ha propuesto un método basado en RFLP para la detección de ADN de
crustáceos en general, pero sin diferenciar entre especies de langostinos. Más recientemente, se han
propuesto tres métodos basados en el análisis del ADN para la identificación de especies de langostinos
y camarones. En el primero (Khamnamtong y col., 2005) se cita un método para la identificación de
especies de langostinos peneideos en base al polimorfismo del gen 16S rRNA, utilizando RFLP y SSCP,
previa amplificación por PCR. En este estudio se obtuvieron patrones de bandas para la identificación de
las especies Penaeus monodon, Litopenaeus vannamei y Fenneropenaeus merguiensis. Sin embargo, en
el caso de otras dos especies (P. semisulcatus y Marsupenaeus japonicus) no lograron dilucidar de qué
especie se trataba ya que presentaban los mismos patrones de corte. En el segundo de los estudios
(Pascoal y col., 2008a) se recoge un método basado en la región de ADNmt 16S rRNA/tRNAVal, que ha
sido patentado (Calo-Mata y col., Patente Nº PCT/ES2007/070212), para la identificación de especies de
langostinos de la superfamilia Penaeoidea utilizando la combinación de PCR y RFLP, en el que es
necesario el corte con 3 enzimas de restricción distintos para conseguir una correcta identificación de las
especies. Estos mismos autores han publicado un método similar basado en el gen mitocondrial del cyt b
(Pascoal y col., 2008b), para la identificación de seis especies de langostinos peneideos.
23

Introducción
Tabla 4. Resumen de los métodos genéticos utilizados en identificación de especies pesqueras.
Técnica Especies Referencias Primers especie-específicos y PCR multiplex
Tiburones Shivji y col., 2002 Abercrombie y col., 2005 Clarke y col., 2006 Magnussen y col., 2007
Pez espada Hsieh y col., 2004 Lenguado y fletán Céspedes y col., 1999a Salmónidos Greig y col., 2002 Perca, mero y cherna Asensio y col., 2001a Mero Asensio, 2008 Atún y bonito Lockley y Bardsley, 2000a
Salmón, bacalao y arenque Bartlett y Davidson, 1992 Gádidos Calo-Mata y col., 2003 Tiburones Blanco y col., 2008 Sardinas Jerome y col., 2003 Anchoas Santaclara y col., 2006 Cefalópodos Chapela y col., 2002 y 2003 Pargo rojo Marko y col., 2004
Secuenciación y análisis filogenético (FINS)
Gádidos y merlúcidos Pepe y col., 2005 RFLP Escombroides Ram y col., 1996
Quinteiro y col., 1998 Chow y col., 2003 Horstkotte y Rehbein, 2003 Takeyama y col., 2001
Peces planos Céspedes y col., 98a,b Sotelo y col., 2001 Comesaña y col., 2003
Gádidos Calo-Mata y col., 2003 Aranishi y col., 2005a,b Pepe y col., 2005
Salmónidos Rusell y col., 2000 Caballa Aranishi, 2005 Salmón y trucha Carrera y col., 1999 Almeja Fernández y col., 2002a Sables Chakraborty y col., 2005 Lenguado y fletán Céspedes y col., 2000 Túnidos Lin y Hwang, 2007 Moluscos Colombo y col., 2002 Anguila Rehbein y col., 2002 Langostinos Pascoal y col., 2008a,b
Khamnamtong y col., 2005 RFLP+lab-on-a-chip Salmon y trucha Dooley y col., 2005a Pescados blancos Dooley y col., 2005b SSCP Salmon, trucha, anguila,
esturión Rehbein y col., 1997
Túnidos Rehbein y col., 1999a Weder y col., 2001 Colombo y col., 2005
Peces planos Céspedes y col., 1999b Perca, mero y cherna Asensio y col., 2001b Almejas Fernández y col., 2002b Bacalao Comi y col., 2005 Merluza Chapela y col., 2007 Langostinos Khamnamtong y col., 2005
24

Introducción
Tabla 4. Continuación.
Técnica Especies Referencias RAPD Pez gato Liu y col., 1998a Tilapia Ahmed y col., 2004 Mejillón Rego y col., 2002 Arowana asiática Yue y col., 2002 Pez sapo Ramella y col., 2005 Salmónidos Jin y col., 2006
Yamazaki y col., 2005 Barbos Callejas y Ochando, 2001 AFLP Pez gato Liu y col., 1998b Ostras Yu y Guo, 2003
Li y Guo, 2004 Trucha Young y col, 1998 Atún Han y Ely, 2002 Salmón y trucha Zhang y Cai, 2006 Varias especies Maldini y col., 2006 PCR en tiempo real Bacalao Sotelo y col., 2003 Mero Trotta y col., 2005 Eglefino Hird y col., 2005 Túnidos López y Pardo, 2005
Aunque los métodos basados en ADN tienen algunas ventajas -las moléculas de ADN son en
general más termoestables que las proteínas, el ADN es igual en todos los tejidos de un organismo, y no
depende de diferencias en la expresión-, también presentan inconvenientes (Lockley y Bardsley, 2000b;
Rasmussen y Morrissey, 2008). Así, la secuenciación, paso necesario en algunas de estas estrategias -
sobre todo cuando se estudian especies que no están muy presentes en las bases de datos, como ocurre
con las especies marinas-, es una técnica cara y lenta, lo que impide su utilización rutinaria en muchos
laboratorios (Dooley y col., 2005a). En cuanto a la técnica de RFLP, el mayor inconveniente es la
posibilidad de variaciones intraespecíficas, es decir, que en una misma especie nos encontremos
patrones de restricción distintos. Además no está garantizado que cada especie produzca un patrón de
restricción único, es decir, puede haber más de una especie con el mismo patrón de restricción. La
técnica de SSCP requiere gran reproducibilidad en cuanto a las condiciones de los análisis, y muestras
de las especies de referencia tienen que ser analizadas en el mismo gel que las muestras problema. Los
RAPD presentan como mayor inconveniente la reproducibilidad del método, necesitando de condiciones
muy estables para asegurar una correcta identificación de las muestras problemas. En cuanto a los
AFLP, la técnica es muy laboriosa, y requiere ADN de gran calidad y elevado tamaño. Además hay que
tener en cuenta que la alta sensibilidad de la PCR implica que es muy susceptible a la contaminación, y
muchos compuestos presentes en alimentos pueden inhibir la amplificación del ADN (Teletchea y col.,
2005).
25

Introducción
4.2. MÉTODOS DE IDENTIFICACIÓN BASADOS EN EL ANÁLISIS DE PROTEÍNAS
Como en todo músculo contráctil (Goll y col., 1977), las proteínas del músculo del abdomen (la
parte comestible) de los decápodos se dividen en tres fracciones:
a) Proteínas sarcoplásmicas. Se extraen con agua o soluciones salinas de baja fuerza iónica, y
constituyen entre el 20 y el 35% del contenido total de proteínas del músculo. La mayoría
son enzimas responsables del metabolismo de la célula.
b) Proteínas miofibrilares. Constituyen entre el 65 y el 75% del total. Se extraen con
soluciones salinas fuertes. Miosina y actina son las dos proteínas mayoritarias en esta
fracción.
c) Proteínas del tejido conectivo. Representan entre el 3 y el 10% del total de proteínas. El
componente principal es el colágeno, que es insoluble en soluciones salinas en su estado
nativo, por lo que para solubilizarlo hay que desnaturalizarlo mediante calentamiento o
mediante soluciones ácidas o básicas.
Las diferencias genéticas entre especies han mostrado estar más desarrolladas en la fracción
sarcoplásmica, al ser proteínas responsables de reacciones enzimáticas muy diversificadas (Mackie,
1997), por lo que se han utilizado ampliamente en estudios de identificación de especies pesqueras.
Se han utilizado gran variedad de técnicas de identificación basados en el análisis de proteínas
(Mackie, 1997). Estas técnicas podrían clasificarse en 4 grandes grupos: (a) técnicas electroforéticas,
que incluyen la electroforesis en geles de poliacrilamida con dodecil sulfato sódico (SDS-PAGE), el
isoelectroenfoque (IEF), la electroforesis bidimensional (2-DE) y la electroforesis capilar (CZE); (b)
técnicas cromatográficas, específicamente cromatografía líquida de alta eficacia (HPLC); (c) técnicas
inmunológicas, que consisten generalmente en el empleo de anticuerpos policlonales o monoclonales en
ensayos ELISA (Enzyme-Linked ImmunoSorbent Assay); y (d) técnicas de espectrometría de masas
(MS). En la tabla 5 se recoge una revisión de las referencias que se pueden encuadrar en alguna de estas
técnicas.
Las técnicas electroforéticas han sido las más utilizadas, constituyendo el IEF de proteínas
sarcoplásmicas la mejor técnica de identificación (Mackie, 1997), siendo adoptada por la Association of
Analytical Communities (AOAC) como el único método oficial validado (AOAC, 1990), AOAC
Official MethodSM 980.16.
26

Introducción
Tabla 5. Resumen de los métodos basados en proteínas utilizados en identificación de especies pesqueras.
Técnica Especies Referencias SDS-PAGE Varios Cowie, 1968 Merluzas Mackie y Jones, 1978 Varios Scobbie y Mackie, 1988 Langostinos An y col., 1988
Civera y Parisi, 1991 Abadejo y merluza en surimi An y col., 1989a Varios Piñeiro y col., 1999a Varios Martínez y col., 2001 Varios Sotelo y col., 1992 Varios Rehbein y col., 1999b Varios Etienne y col., 2000 Pez globo Chen y Hwang, 2002 IEF Varios Lundstrom, 1979 Langostinos An y col., 1989b
Wei y col., 1990 Abadejo y merluza en surimi An y col., 1989a Varios Whitmore, 1990 Varios Sotelo y col., 1992 Congrio Valenzuela y col., 1999 Tiburones Weaver y col., 1999 Gádidos Piñeiro y col., 2000 Salmónidos y anguilas Mackie y col., 2000 Pez globo Chen y col., 2003 Perca Berrini y col., 2006 Varios Rehbein y col., 1999b, 2000 Varios Etienne y col., 2000 Peces planos y gádidos Tepedino y col., 2001 2-DE Varios Martínez y col., 1990 Bacalao, bacaladilla y eglefino Martínez y Friis, 2004 Pez globo Chen y col., 2004 Congrio Valenzuela y col., 1999 Peces planos Piñeiro y col., 1999b Perca Berrini y col., 2006 Gádidos Piñeiro y col., 1998 Merluzas Piñeiro y col., 2001 Mejillón López y col., 2002a CZE Bacalao y abadejo Leblanc y col., 1994 Peces planos Gallardo y col., 1995
Electroforesis
Congrio Valenzuela y col., 1999 Larraín y col., 2002
Varios Rehbein, 1990 Varios Osman y col., 1987 Varios Armstrong y Leach, 1992 Gádidos Piñeiro y col., 1997 Ballenas Ukishima y col., 1995 Especies de agua dulce Knuutinen y Harjula, 1998
Cromatografía
Varios Chou y col., 2007
27

Introducción
Tabla 5. Continuación.
Técnica Especies Referencias
Inmunoprecipitación y aglutinación
Arenques Mairs y Sindermann, 1962
Inmunodifusión e inmunodotting
Lenguado, fletán, bacalao y anguila
Domínguez y col., 1997
ELISA Sardina Taylor y Jones, 1992 Salmón, trucha y palometa Carrera y col., 1996, 1997 Mero, perca y cherna Asensio y col., 2003a, b Lenguado, fletán, solla y
platija Céspedes y col., 1999c
Pargo rojo Huang y col., 1995 Pez gato NcNulty y Klesius, 2005
Inmunológicas
Langostinos An y col, 1990 Mejillón López y col., 2002b Espectrometría de masas Merluzas Piñeiro y col., 2001
Carrera y col., 2006, 2007
En cuanto al grupo de los crustáceos decápodos, los estudios de identificación de especies basados
en el análisis de las proteínas son escasos: un estudio utilizando SDS-PAGE (Civera y Parisi, 1991), en
el que no se consiguen identificar especies próximas; y 3 estudios del grupo de An (An y col.,1988,
1989b; Wei y col., 1990), utilizando SDS-PAGE el primero e IEF los 2 últimos, en los que sólo se
incluyeron 3 especies minoritarias (Penaeus duorarum, Penaeus setiferus y Sicyonia brevirostris). El
mismo grupo desarrolló un anticuerpo monoclonal que reconoce una proteína específica de S.
brevirostris, y probaron su especificidad para detectar únicamente esta especie y no otras (An y col.,
1990). Debido a la escasez de estudios de autentificación de especies de decápodos, se hace necesario el
desarrollo de nuevas herramientas capaces de conseguir la identificación de las principales especies
comerciales de langostinos y gambas, de una forma rápida, fiable y sencilla. Como veremos, las
metodologías proteómicas pueden contribuir a este objetivo.
5. PROTEÓMICA
La proteómica es la disciplina que comprende el conjunto de metodologías orientadas al análisis y
estudio de proteínas a gran escala (Pandey y Mann, 2000). También podría definirse como el conjunto
de metodologías utilizadas para el estudio de un proteoma. Del término “proteoma” pueden actualmente
encontrarse diferentes definiciones, una de las más aceptadas es “el conjunto de proteínas de un
organismo, una célula, un orgánulo, etc., en un determinado momento, bajo precisas y determinadas
condiciones”.
28

Introducción
El esquema general de análisis de un proteoma implica una primera etapa de preparación de la
muestra, una segunda de separación de los analitos, un análisis de éstos por espectrometría de masas
(MS) y por último un análisis exhaustivo de los datos que se han generado. Para el estudio proteómico
llevado a cabo en la presente tesis, se seguirán dos aproximaciones diferentes:
a) Aproximación clásica.
Esta estrategia, la más utilizada hasta el momento, consiste en la separación electroforética de la
mezcla compleja de proteínas en geles de poliacrilamida, generalmente en dos dimensiones (2-DE),
aunque también puede utilizarse una única dimensión. Las proteínas separadas en el gel son
visualizadas mediante distintas técnicas de tinción, y los geles son escaneados y sometidos a análisis
de imagen mediante diferentes programas informáticos. Las proteínas de interés son digeridas en gel
con enzimas como la tripsina, y los péptidos resultantes de la digestión son analizados mediante MS.
En ocasiones, se recurre a la separación de la mezcla de péptidos mediante cromatografía líquida
(LC, Liquid Chromatography) acoplada al equipo de MS. La identificación de la proteína original se
lleva a cabo comparando el mapa o huella de masas peptídicas (PMF, Peptide Mass Fingerprinting),
o bien el espectro de fragmentación de uno o más de los péptidos, frente a valores teóricos
calculados a partir de las secuencias contenidas en las bases de datos.
b) Aproximación Shotgun o de segunda generación.
Mediante esta estrategia se estudian las proteínas contenidas en una muestra a partir del análisis
directo y rápido de la mezcla compleja de péptidos resultantes de su digestión enzimática (Link y
col., 1999; Wu y MacCoss, 2002). Se emplean por tanto los digeridos obtenidos a partir de mezclas
de proteínas, sin necesidad de una separación electroforética previa. Posteriormente, esta mezcla
compleja de péptidos se separa mediante cromatrografía acoplada a un espectrómetro de masas
como detector, que va fragmentando los péptidos según eluyen de la cromatografía. Típicamente
esta separación cromatográfica se realiza en dos dimensiones, atendiendo a dos propiedades
diferentes de los péptidos, por ejemplo una LC de intercambio catiónico fuerte y una LC en fase
reversa (RP, Reverse Phase). Pero también puede realizarse en una única dimensión, normalmente
una LC en RP. Finalmente se realiza la comparación bioinformática de las fragmentaciones
obtenidas con las fragmentaciones virtuales de los péptidos procedentes de las proteínas presentes en
las bases de datos. Esta aproximación conlleva un mayor grado de automatización y un menor
tiempo de análisis.
29

Introducción
5.1. SEPARACIÓN DE PROTEÍNAS Y PÉPTIDOS
Las principales técnicas de separación de los componentes de un proteoma son las técnicas
electroforéticas y las técnicas cromatográficas.
5.1.1. Técnicas electroforéticas
5.1.1.1. SDS-PAGE
La separación de proteínas en geles de poliacrilamida con dodecil sulfato sódico (SDS-PAGE,
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis) (Laemmli, 1970) se basa en la unión de
docecil sulfato sódico (SDS) a la proteína. El SDS es un detergente aniónico que se une a las proteínas
con una relación de una molécula de SDS por cada dos aminoácidos. Esto le confiere a la proteína una
carga neta negativa, proporcional a su masa molecular relativa (Mr). Para la separación de proteínas se
utilizan geles de poliacrilamida con distinto grado de entrecruzamiento según el tamaño de las proteínas
a separar. Sometidos a una diferencia de potencial, los complejos SDS-proteína migran a través del
tamiz molecular en función únicamente de la Mr, resolviéndose en bandas. Tras el revelado del gel
mediante tinción, se puede estimar la Mr de una proteína problema por comparación con una mezcla de
proteínas estándar de Mr conocidas, ya que la movilidad electroforética es proporcional al logaritmo del
Mr.
Ésta es una técnica sencilla, rápida y barata, que permite separar polipéptidos de entre 10 y 300
kDa. Sin embargo, el poder de resolución de estos geles de SDS-PAGE en una única dimensión es
limitado, y así bandas que aparentemente contienen una sola proteína pueden en realidad contener
varias. Por ello, la utilidad de este método de separación depende de la complejidad de la muestra, no
pudiendo resolver más de 80-100 proteínas diferentes en un mismo gel (López, 2007).
5.1.1.2. Isoelectroenfoque
En el isoelectroenfoque (IEF), las proteínas se separan en función de su punto isoeléctrico (pI), es
decir, el pH en el cual su carga neta es cero. Esta técnica de separación se basa en la propiedad
anfotérica de los aminoácidos: pueden protonarse o desprotonarse según el pH del medio. En medio
básico, los grupos ácidos se cargan negativamente, y en medio ácido, los grupos básicos se cargan
positivamente, siendo la carga neta de la proteína la suma de todas las cargas negativas y positivas. Al
aplicar un campo eléctrico en un gradiente de pH, las proteínas cargadas migran hacia el electrodo de
signo opuesto a su carga neta, así las proteínas migrarán hasta la zona con pH en el cual no presenten
carga neta (el pI), donde se detendrán.
30

Introducción
El IEF puede llevarse a cabo en gel o en solución. Actualmente se comercializan geles y tiras con
gradientes inmovilizados de pH (IPG, Immobilized pH Gradient) de distintos tamaños y rangos de
gradientes de pH, que permiten alcanzar excelentes resoluciones (ΔpI=0,001). Tras revelado del gel
mediante tinción, se puede estimar el pI de una proteína problema mediante comparación con una
mezcla de proteínas estándar de pI conocidos.
5.1.1.3. 2-DE
La 2-DE (O’Farrel, 1975; Klose, 1975) es, hasta el momento, la técnica que permite resolver el
mayor número de proteínas en una mezcla de complejidad elevada (cientos o miles de proteínas).
Consiste en combinar dos separaciones distintas, en la primera las proteínas se separan por IEF según su
pI, y en la segunda las proteínas se separan según su Mr en un gel SDS-PAGE. Tras la tinción con plata,
coomassie, sypro, etc., las proteínas separadas aparecen como manchas o spots.
El poder de resolución y la reproducibilidad de esta técnica se incrementó notablemente con la
introducción de los geles IPG para la primera dimensión. Sin embargo, esta metodología tiene algunos
inconvenientes, principalmente la dificultad para realizar análisis completamente reproducibles, además
de que es una técnica muy laboriosa, requiere tiempo y es difícil de automatizar.
Para el análisis y comparación de los geles 2-DE se han desarrollado programas informáticos de
análisis de imagen, como el PDQuest (Garrels, 1989) y el Melanie (Appel y col., 1991).
5.1.2. Técnicas cromatográficas
La separación mediante cromatografía líquida (LC) permite separar tanto péptidos como proteínas
en función de sus propiedades fisicoquímicas, así como concentrar la muestra y eliminar sales y
detergentes. Los métodos cromatográficos más comunes en la separación de péptidos y proteínas
utilizan sistemas de alta eficacia (HPLC, High Performace Liquid Chromatography). Los tipos de
separación cromatográfica más utilizados son la fase reversa (RP), el intercambio iónico, la exclusión
molecular y la afinidad.
Actualmente, la mayoría de las separaciones cromatográficas de péptidos se realizan en equipos de
HPLC con columnas capilares de RP, gracias a su gran poder de resolución, reproducibilidad y
compatibilidad con los equipos de MS, a los que se pueden acoplar (LC-MS). Aunque la LC
monodimensional es la más utilizada, la LC multidimensional está suponiendo una gran mejora en la
resolución de mezclas complejas de péptidos. (Link y col., 1999), y constituye habitualmente la base de
los estudios Shotgun. Comúnmente, la LC multidimensional consiste en dos separaciones ortogonales,
siendo intercambio catiónico fuerte (SCX) y RP las más utilizadas.
31

Introducción
5.2. ESPECTROMETRÍA DE MASAS
La espectrometría de masas (MS) se ha convertido desde finales de los años 80 en el método de
elección para la identificación, caracterización y cuantificación de proteínas, debido a su rapidez,
sensibilidad y precisión. Esta tecnología permite producir iones en fase gaseosa a partir de moléculas
orgánicas o inorgánicas, separarlos en función de su relación masa/carga (m/z) y medir su intensidad.
Mediante esta técnica puede determinarse el peso molecular y la abundancia de los componentes de una
mezcla. Los iones analizados pueden corresponder a moléculas enteras, aductos o fragmentos de
moléculas. Además, el análisis de los fragmentos iónicos formados a partir de un determinado ión
molecular o precursor, proporciona información sobre su estructura química.
Un espectrómetro de masas está formado por tres partes fundamentales: la fuente de iones, el
analizador de masas, y el detector (Figura 7). Las muestras a analizar se
Figura 7. Representación esquemática de los principales componentes de un
espectrómetro de masas.
introducen en la fuente de iones donde los componentes de la muestra son convertidos en iones en fase
gaseosa. Estos iones son transferidos al analizador, donde son acelerados y separados según su relación
m/z, utilizando distintos principios físicos según el tipo de analizador (Tabla 6). La fuente de ionización
y el analizador de masas definen el tipo de espectrómetro de masas, existiendo diversos tipos de ambos y
dando lugar a numerosas combinaciones. Los analizadores se clasifican en cuatro grupos: de sectores
32
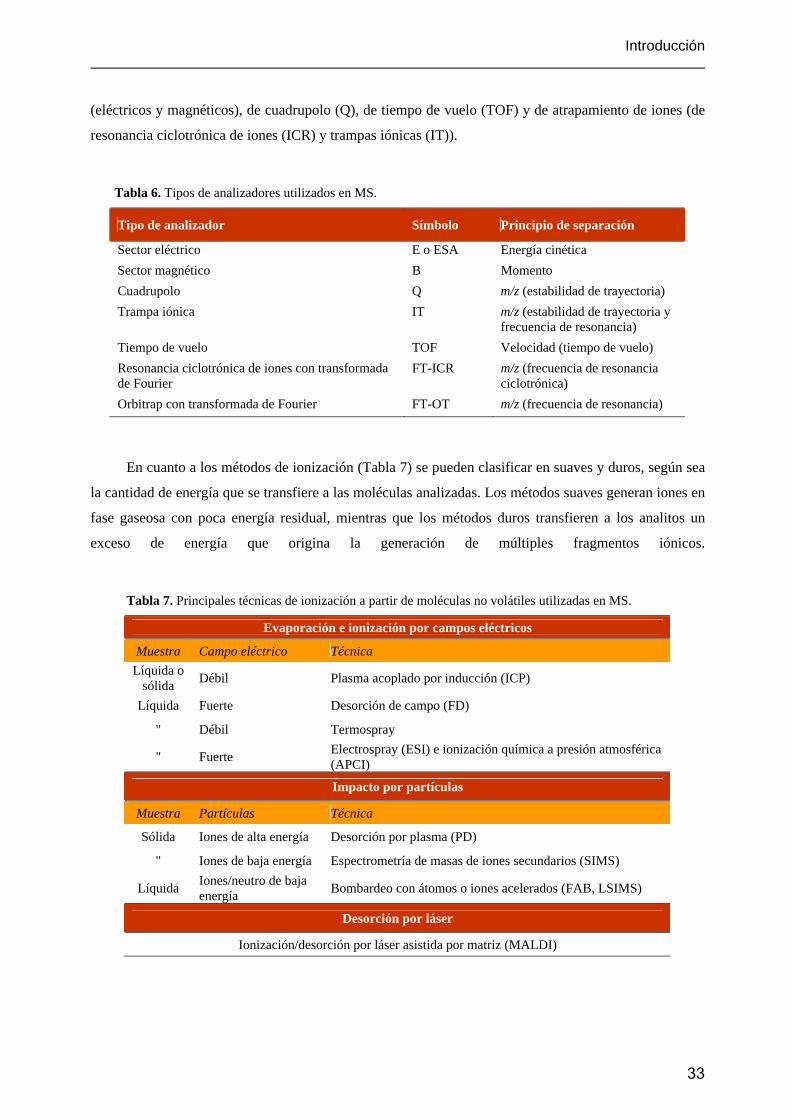
Introducción
(eléctricos y magnéticos), de cuadrupolo (Q), de tiempo de vuelo (TOF) y de atrapamiento de iones (de
resonancia ciclotrónica de iones (ICR) y trampas iónicas (IT)).
Tabla 6. Tipos de analizadores utilizados en MS.
Tipo de analizador Símbolo Principio de separación
Sector eléctrico E o ESA Energía cinética Sector magnético B Momento Cuadrupolo Q m/z (estabilidad de trayectoria) Trampa iónica IT m/z (estabilidad de trayectoria y
frecuencia de resonancia) Tiempo de vuelo TOF Velocidad (tiempo de vuelo) Resonancia ciclotrónica de iones con transformada de Fourier
FT-ICR m/z (frecuencia de resonancia ciclotrónica)
Orbitrap con transformada de Fourier FT-OT m/z (frecuencia de resonancia)
En cuanto a los métodos de ionización (Tabla 7) se pueden clasificar en suaves y duros, según sea
la cantidad de energía que se transfiere a las moléculas analizadas. Los métodos suaves generan iones en
fase gaseosa con poca energía residual, mientras que los métodos duros transfieren a los analitos un
exceso de energía que origina la generación de múltiples fragmentos iónicos.
Tabla 7. Principales técnicas de ionización a partir de moléculas no volátiles utilizadas en MS.
Evaporación e ionización por campos eléctricos
Muestra Campo eléctrico Técnica Líquida o
sólida Débil Plasma acoplado por inducción (ICP)
Líquida Fuerte Desorción de campo (FD)
" Débil Termospray
" Fuerte Electrospray (ESI) e ionización química a presión atmosférica (APCI)
Impacto por partículas
Muestra Partículas Técnica
Sólida Iones de alta energía Desorción por plasma (PD)
" Iones de baja energía Espectrometría de masas de iones secundarios (SIMS)
Líquida Iones/neutro de baja energía Bombardeo con átomos o iones acelerados (FAB, LSIMS)
Desorción por láser
Ionización/desorción por láser asistida por matriz (MALDI)
33

Introducción
La aparición a finales de los años 80 de la ionización por electrospray (ESI) (Fenn y col., 1989) y
la ionización/desorción por láser asistida por matriz (MALDI) (Karas y Hillenkamp, 1988; Tanaka y
col., 1988) permitieron el estudio mediante MS de moléculas que resultaban hasta entonces difíciles de
ionizar y volatilizar sin que fueran destruidas o deterioradas debido a su gran tamaño y polaridad. Estas
técnicas de ionización suave permiten la generación de iones a partir de analitos no volátiles y de
elevado peso molecular, como las proteínas y péptidos, sin una fragmentación significativa. Además, la
formación de iones multicargados a partir de péptidos y proteínas en la ionización por ESI permite su
detección y fragmentación en aparatos con un rango de masas limitado, como los cuadrupolos y las
trampas iónicas, dando lugar a patrones de fragmentación fácilmente interpretables. Debido a su
eficiencia, estas dos técnicas son las preferidas en la actualidad para el análisis de proteínas y péptidos,
habiendo reemplazado a la secuenciación de Edman como método de identificación y caracterización.
En 2002 se concedió el Premio Nobel en Química a John Fenn y Koichi Tanaka por el desarrollo del
ESI y la base para el posterior desarrollo del MALDI, respectivamente. En este trabajo utilizaremos
métodos de espectrometría de masas combinando ionización por MALDI y analizador de TOF, e
ionización por ESI y analizador de IT, por lo que estos dos sistemas se describen más detalladamente a
continuación.
5.2.1. MALDI-TOF
En la técnica de ionización por MALDI (Karas y Hillenkamp, 1988; Tanaka y col., 1988) se
utilizan pulsos de luz láser de nanosegundos en la frecuencia del ultravioleta (nitrógeno a 337 nm es el
más utilizado) o del infrarrojo lejano para desorber e ionizar una muestra previamente cristalizada en
una placa junto a un compuesto denominado matriz. La matriz tiene varias funciones: incorporar y
dispersar las moléculas de analito, preservarlas de recibir una excesiva cantidad de energía, y absorber la
energía del láser para conseguir una transferencia controlada y eficaz de ésta.
El progreso de la técnica MALDI ha sido fundamentalmente experimental, ya que los procesos
fisicoquímicos en los que se basa están todavía sujetos a debate. Los principales hechos empíricos
encontrados han sido:
- el espectro se caracteriza por una escasa fragmentación.
- prácticamente solo se observan iones monocargados.
- se pueden detectar iones positivos o negativos dependiendo de la polaridad que se elija en el
campo de extracción.
Uno de los mecanismos más aceptados para explicar la ionización por MALDI es un proceso
fotoquímico, según el cual la radiación láser es absorbida por las moléculas de la matriz generándose
una nube en la que hay iones, moléculas neutras y agrupamientos de estas. Los iones de los analitos se
34

Introducción
formarían por protonación, desprotonación, o cationización tras el choque con iones de la matriz en la
fase gaseosa (Figura 8). Se producen así iones monocargados estables, aunque también se observan
iones con dos o más cargas en el caso de proteínas con pesos moleculares superiores a 5 kDa. Con esta
técnica pueden detectarse proteínas de más de 300 kDa.
Figura 8. Esquema de la ionización MALDI.
En la tabla 8 se muestran algunas de las matrices más utilizadas actualmente y los compuestos
para los que se recomiendan. La mayor parte de ellas tienen grupos aromáticos, para absorber la
radiación láser, y un grupo hidroxilo (-OH) en posición orto respecto a un grupo carbonilo (-COOH, -
CO, -CONH2), para facilitar la transferencia protónica. Las matrices más utilizadas para el análisis de
proteínas y péptidos son el α–ciano-4-hidroxicinámico (CHCA), el ácido 2,5-dihidroxibenzoico (DHB)
y el ácico sinapínico.
La ionización MALDI es relativamente menos sensible a la contaminación por sales, tampones,
detergentes y otros contaminantes que otros tipos de ionización, lo que es una ventaja cuando se
analizan extractos de proteínas o péptidos obtenidos de muestras procedentes de alimentos.
35

Introducción
Tabla 8. Matrices empleadas para MALDI.
Matriz Estructura Campo de aplicación
Ácido sinapínico (SA)
Péptidos y proteínas > 10.000 u
Ácido α–ciano-4-hidroxicinámico (α-ciano o CHCA)
Péptidos y proteínas < 10.000 u
Ácido 3-hidroxipicolínico (3-HPA)
Oligonucléotidos > 3.500 u
2,4,6-trihidroxiacetofenona (THAP)
Oligonucleótidos < 3.500 u
Ácido 2,5-dihidroxibenzoico (DHB)
Péptidos, carbohidratos neutros o básicos, moléculas pequeñas, polímeros sintéticos polares
Ditranol
Polímeros sintéticos aromáticos no polares
Ácido trans-3-indoleacrílico (IAA)
Polímeros sintéticos apolares
Tradicionalmente, la fuente de ionización MALDI se suele acoplar con analizadores de tiempo de
vuelo (TOF), constituyendo este sistema (MALDI-TOF) un método estándar para el análisis de péptidos
y proteínas, caracterizándose por su alta sensibilidad, robustez, sencillez de manejo y capacidad de
automatización, así como por su alta tolerancia a contaminantes. En un espectrómetro MALDI-TOF los
iones formados en la fuente se aceleran mediante la aplicación de un campo eléctrico para, a
continuación, ser separados en un tubo de vuelo a alto vacío, en el que se desplazan libremente sin ser
sometidos a ningún campo eléctrico o magnético, gracias a la energía cinética que adquirieron en la
fuente (Figura 9). Estos iones adquieren la misma energía cinética durante la aceleración, pero su
velocidad, y por tanto el tiempo que tardan en atravesar el tubo de vuelo, varía. Este tiempo depende de
su relación m/z y de su energía cinética. Como la energía cinética es constante para un voltaje de
36
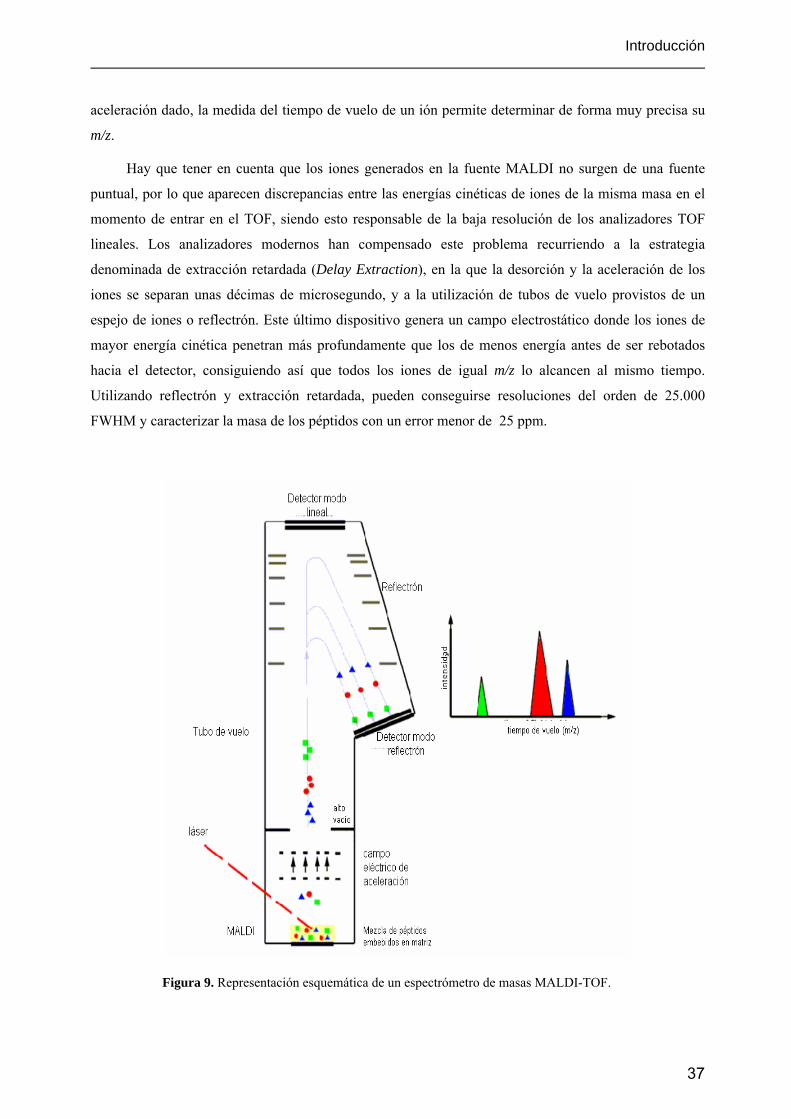
Introducción
aceleración dado, la medida del tiempo de vuelo de un ión permite determinar de forma muy precisa su
m/z.
Hay que tener en cuenta que los iones generados en la fuente MALDI no surgen de una fuente
puntual, por lo que aparecen discrepancias entre las energías cinéticas de iones de la misma masa en el
momento de entrar en el TOF, siendo esto responsable de la baja resolución de los analizadores TOF
lineales. Los analizadores modernos han compensado este problema recurriendo a la estrategia
denominada de extracción retardada (Delay Extraction), en la que la desorción y la aceleración de los
iones se separan unas décimas de microsegundo, y a la utilización de tubos de vuelo provistos de un
espejo de iones o reflectrón. Este último dispositivo genera un campo electrostático donde los iones de
mayor energía cinética penetran más profundamente que los de menos energía antes de ser rebotados
hacia el detector, consiguiendo así que todos los iones de igual m/z lo alcancen al mismo tiempo.
Utilizando reflectrón y extracción retardada, pueden conseguirse resoluciones del orden de 25.000
FWHM y caracterizar la masa de los péptidos con un error menor de 25 ppm.
Figura 9. Representación esquemática de un espectrómetro de masas MALDI-TOF.
37

Introducción
5.2.2. ESI-IT
El ESI (Fenn y col., 1989) es una técnica de ionización a presión atmosférica en la que una
disolución de la muestra es nebulizada a la salida de un tubo capilar por la acción de un fuerte campo
eléctrico (Figura 10). De esta forma, la disolución se dispersa en forma de un spray muy fino, formado
por pequeñas gotas cargadas que contienen los iones de los analitos. Estos iones pueden pasar a fase
gaseosa bien a través de procesos de desolvatación de la gota, bien por desorción directa de los iones
desde la gota debido a la alta tensión electrostática en su interior. Este proceso de evaporación puede ser
ayudado por calor y/o por un flujo de gas (N2). Este tipo de ionización es extremadamente suave,
permitiendo no solo generar iones de la molécula intacta (sin fragmentar la molécula), sino también de
complejos formados a través de interacciones no covalentes.
Figura 10. Esquema de una fuente de ionización por electrospray.
La ionización por ESI produce característicamente iones multicargados, y tanto el tipo como el
número de iones producidos reflejan el equilibrio ácido-base que existe en la disolución. El grado de
carga de un determinado ión depende de su estructura (presencia de grupos ácidos o básicos) y del
disolvente utilizado. En el caso de péptidos y proteínas, el número de cargas depende de su longitud y
del número de residuos básicos de la molécula (His, Arg, Lys y el extremo amino-terminal (N-
terminal)), que son los candidatos a protonarse. Dado que al aumentar el tamaño de un péptido aumenta
también el número de residuos capaces de adquirir carga, la relación m/z de los iones producidos a partir
de péptidos y proteínas suelen encontrarse en el rango de 700 a 2000 Th, independientemente de la masa
38

Introducción
(Figura 11). Esto permite el análisis de iones de compuestos con pesos moleculares muy superiores al
límite de barrido de analizadores como los cuadrupolos o las trampas iónicas, cuyos límites no superan
unos pocos miles de Th.
nnM +=m/z
Figura 11. Ecuación que describe la distribución de las señales iónicas observadas por MS derivadas de una
molécula de masa M que puede tomar n protones para generar iones con n cargas.
Al ionizar por ESI péptidos procedentes de digestiones con tripsina, muy frecuente en estudios de
proteómica, los iones que suelen predominar son los de carga +2, aunque también se pueden generar
iones con carga +1, +3 o superiores. Dado que la tripsina rompe los enlaces peptídicos por el extremo
carboxilo-terminal (C-terminal) de las Lys y Arg, estos péptidos presentan al menos dos sitios básicos
susceptibles de protonarse, el extremo N-terminal y la cadena lateral de la Lys o Arg, con lo que
tendrían carga +2. Además, pueden generarse péptidos con más de 2 cargas, debido al contenido interno
de residuos de His, Arg, y/o Lys.
Las fuentes actuales, denominadas de microspray (microESI) (Emmett y Caprioli, 1994), son
fuentes con un capilar muy fino (75 μm) y en las que el flujo es muy pequeño (del orden de μL/min). A
este tipo de fuentes suele acoplarse una separación cromatográfica previa, con dos objetivos: eliminar
las sales de las muestras proteicas (ya que las sales disminuyen la sensibilidad del análisis); y separar los
péptidos o proteínas contenidos en la muestra.
Una última versión, las fuentes de nanospray (nESI) (Wilm y Mann, 1996), opera de forma similar
a la de microESI, pero trabajando a flujo más bajo aún (5-10 nL/min) y con un diámetro de capilar o
aguja mucho menor (1-10 μm). Debido a estas características, el spray estará formado por gotas mucho
más pequeñas (menos de 200 nm de diámetro), por lo que aumenta la eficiencia en la ionización y el
número de iones que entran hacia el analizador, aumentando la sensibilidad hasta un límite de detección
del orden de 50 attomoles. Además, el nESI presenta otras ventajas, como un menor consumo de
muestra, aumentando el tiempo de análisis. Con relación a las fuentes de microESI, el nESI es más
tolerante con la composición y el pH del medio, aunque es recomendable un desalado previo de la
muestra.
Las fuentes de ESI se pueden combinar con diferentes analizadores. Para los análisis de péptidos
realizados en la presente tesis se utilizó un analizador de trampa iónica acoplado a fuentes de micro y
nanoelectrospray, ya que esta combinación es una de las más sensibles y eficaces para ese propósito.
39

Introducción
El analizador de trampa iónica (Paul y Steinwedel, 1953; Stafford y col., 1984) es el análogo
tridimensional del analizador de cuadrupolo. Su desarrollo le valió a Paul el premio Nobel de Física en
1989. Consiste en un recinto definido por tres electrodos: un electrodo anular (ring electrode) y dos
electrodos de superficie hiperbólica denominados electrodos de cierre (endcap electrode) situados a
modo de tapaderas a cada lado del electrodo anular, formando una cavidad en la cual es posible
almacenar y analizar los iones (Figura 12). Estos electrodos de cierre tienen en el centro una pequeña
apertura que permite a los iones entrar o salir de la cavidad.
Figura 12. Esquema de una trampa iónica tridimensional.
La aplicación de un voltaje de radiofrecuencias (RF) al electrodo anular produce un campo
eléctrico tridimensional pulsante, que es el responsable del atrapamiento de los iones dentro de la
trampa. Los iones oscilan en órbitas estables que pueden mantenerlos confinados en el interior de la
trampa durante un cierto tiempo, y la presencia de un gas inerte (He) “enfría” los iones a través de
procesos de colisión, ayudando a estabilizar sus órbitas. Posteriormente, la intensidad de la corriente de
RF puede aumentarse de modo que los iones adquieren secuencialmente trayectorias inestables en
relación directa a su m/z, siendo expulsados desde la trampa hacia el detector. Los iones son expulsados
secuencialmente en función de su m/z, primero salen los iones de menor m/z y luego los de mayor. Así, a
partir del potencial necesario para la expulsión de un determinado ión puede determinarse de forma muy
precisa su valor m/z.
40

Introducción
Una reciente mejora son las trampas iónicas lineales (LIT), o trampas iónicas 2D, en las que los
iones quedan atrapados en el interior de un cuadrupolo por la acción combinada de un campo de RF y
otro creado por un voltaje de corriente continua. Estas trampas ofrecen mayor eficiencia en el
atrapamiento de iones y mayor sensibilidad.
Otro tipo de analizador que funcina atrapando iones es el recientemente desarrollado orbitrap. En
su interior, los iones son atrapados radialmente, entre un electrodo externo con forma de barril y un
electrodo interno central en forma de huso (Figura 13). Ambos electrodos forman un campo
electrostático en cuyo interior los iones oscilan alrededor del electrodo central, de forma que el valor m/z
de cada ión se calcula en función de su frecuencia de oscilación armónica. Este analizador proporciona
una alta resolución y una gran exactitud en la determinación de las masas de los iones (2-5 ppm de
error).
Figura 13. Esquema de un analizador tipo Orbitrap.
5.3. ESPECTROMETRÍA DE MASAS EN TÁNDEM (MS/MS)
Para obtener información sobre la secuencia de los péptidos o proteínas analizadas, es necesario
recurrir a la espectrometría de masas en tándem (MS/MS). Mediante esta técnica, un ión específico
generado en la fuente de ionización (ión precursor) es aislado en el interior del espectrómetro de masas
y, posteriormente, fragmentado (McLafferty, 1981). Del análisis de las m/z de los iones fragmento
producidos, se deriva la información estructural de la molécula.
Este procedimiento puede llevarse a cabo de dos formas: separando las operaciones de selección
del ión precursor y de análisis de los fragmentos en el espacio, o separándolas en el tiempo. El primer
procedimiento (separación en el espacio) implica la utilización de dos analizadores de masas dispuestos
en serie, entre los que se intercala una celda o cámara de colisión. El ión precursor se selecciona en el
primer analizador y se fragmenta mediante procesos de colisión en la cámara de colisión.
41

Introducción
Posteriormente, los iones fragmento son separados en el segundo analizador. Como ejemplo de estos
instrumentos se pueden destacar el triple cuadrupolo, el q-TOF y el TOF-TOF.
El segundo procedimiento (separación en el tiempo) utiliza un único analizador de masas donde
tienen lugar las 3 etapas -selección del ión precursor, fragmentación y análisis de los fragmentos- pero
secuencialmente, en momentos diferentes. Las trampas iónicas funcionan de esta manera. El ión
precursor se aísla mediante la aplicación de voltajes que desestabilizan y expulsan al resto de iones. Los
iones aislados se fragmentan y, una vez formados los iones fragmento, se realiza un barrido de RF en el
que estos iones fragmento van siendo expulsados secuencialmente en función de su m/z y son
detectados.
El mecanismo de fragmentación más utilizado en proteómica es la disociación inducida por
colisión (CID). Durante este proceso, el ión seleccionado es activado por colisiones con las moléculas de
un gas neutro (He, N2, Ar) que se encuentra en la celda de colisión, convirtiéndose una parte de su
energía cinética en energía interna. Esto da lugar a una reacción unimolecular que termina con la
fragmentación del ión, principalmente por sus enlaces peptídicos. De esta forma, el patrón de
fragmentación de un péptido analizado mediante MS/MS depende de la energía de colisión, de la
presión y el tipo de gas de colisión, y de la carga del péptido. Ese patrón es además característico de la
secuencia aminoacídica específica del péptido, de forma que ésta puede deducirse del análisis del
correspondiente espectro MS/MS.
Una de las características más interesantes de la trampa iónica es su capacidad para llevar a cabo
fragmentaciones sucesivas (MSn, espectrometría de masas en tándem múltiple). El proceso de
aislamiento, excitación y expulsión de los iones puede ser repetido n veces, seleccionando cada vez un
ión precursor entre los iones producidos en la fragmentación anterior (Figura 14). Este método es muy
Figura 14. Proceso de masas en tándem en una trampa iónica.
42

Introducción
específico y proporciona información estructural muy valiosa para la secuenciación de péptidos.
5.3.1. Secuenciación de péptidos mediante MS/MS
Durante el análisis MS/MS, los péptidos pueden fragmentarse por cualquiera de los enlaces de la
cadena lineal, generando fragmentos de diferentes tipos (Figura 15). Las series de fragmentos de tipo a,
b y c contienen el extremo N-terminal, mientras que las de tipo x, y y z contienen el C-terminal
(Roepstorff y Fohlman, 1984). Para nombrar los distintos fragmentos se utilizan los símbolos anteriores
con un subíndice que indica el número de aminoácidos que contienen.
Figura 15. Esquema de la fragmentación de péptidos mediante MS/MS.
La fragmentación mediante colisiones de baja energía solo es capaz de romper los enlaces más
débiles de la cadena lineal del péptido, predominando en este caso los iones y y b. Cuando el ión
fragmentado es monocargado, uno de los fragmentos (y o b) retiene ese carga formando un ión y siendo
detectado por el espectrómetro de masas, mientras el otro fragmento queda sin carga y por lo tanto no es
detectado. Los iones producidos por ESI pueden llevar más de una carga, lo que permite que ambos
fragmentos (y y b) reciban alguna carga y por tanto se detecten simultáneamente en un mismo evento de
fragmentación. Dentro de cada serie (y o b), las rupturas dan lugar a fragmentos cuyas m/z difieren en
masas que se corresponden con los residuos de los aminoácidos. Se entiende como masas de residuo
(Tabla 9) la correspondiente a la masa de un aminoácido menos la de la molécula de agua que se pierde
al formarse el enlace peptídico.
43
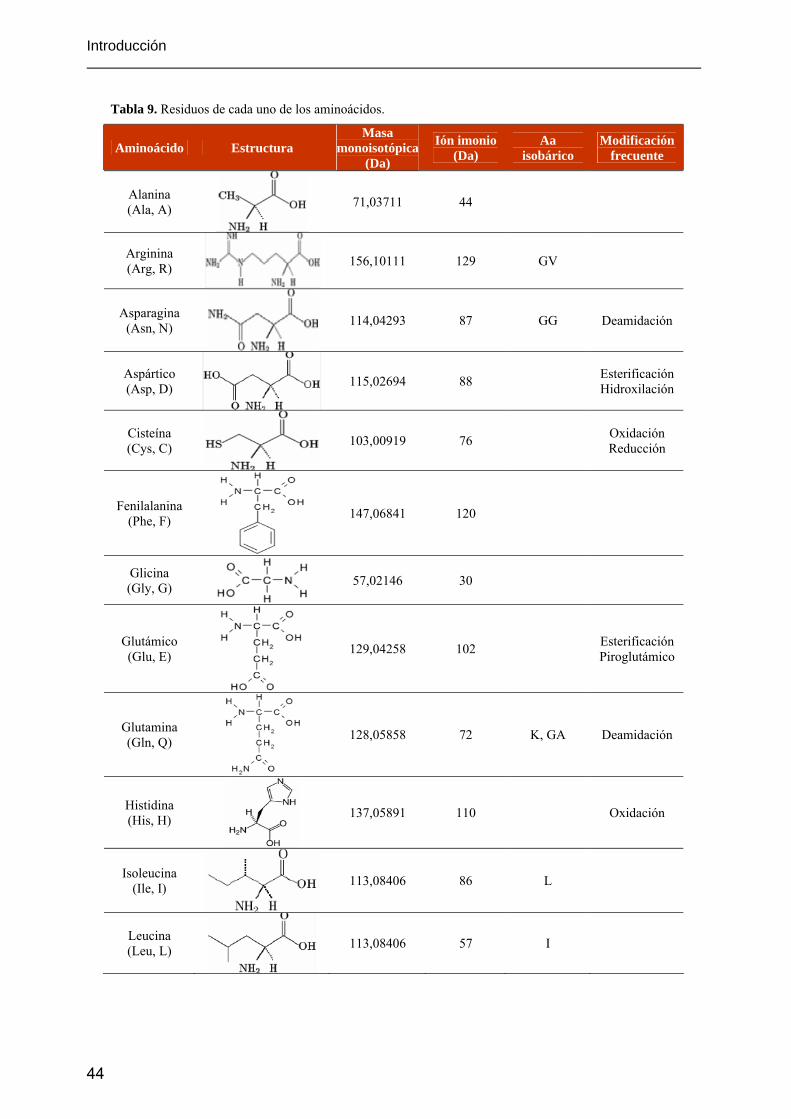
Introducción
Tabla 9. Residuos de cada uno de los aminoácidos.
Aminoácido Estructura Masa
monoisotópica (Da)
Ión imonio (Da)
Aa isobárico
Modificación frecuente
Alanina (Ala, A)
71,03711 44
Arginina (Arg, R)
156,10111 129 GV
Asparagina (Asn, N)
114,04293 87 GG Deamidación
Aspártico (Asp, D)
115,02694 88 Esterificación Hidroxilación
Cisteína (Cys, C)
103,00919 76 Oxidación Reducción
Fenilalanina (Phe, F)
147,06841 120
Glicina (Gly, G)
57,02146 30
Glutámico (Glu, E)
129,04258 102 Esterificación Piroglutámico
Glutamina (Gln, Q)
128,05858 72 K, GA Deamidación
Histidina (His, H)
137,05891 110 Oxidación
Isoleucina (Ile, I)
113,08406 86 L
Leucina (Leu, L)
113,08406 57 I
44

Introducción
Tabla 9. Continuación.
Aminoácido Estructura Masa
monoisotópica (Da)
Ión imonio (Da)
Aa isobárico
Modificación frecuente
Lisina (Lys, K)
128,09496 72 Q, GA Acilación Hidroxilación
Metionina (Met, M)
131,04049 104 Oxidación
Prolina (Pro, P)
97,05276 70 Hidroxilación
Serina (Ser, S)
87,03203 60 Fosforilación Sulfonación
Tirosina (Tyr, Y)
163,06333 136 Fosforilación Sulfonación
Treonina (Thr, T)
101,04768 74 Fosforilación Sulfonación
Triptófano (Trp, W)
186,07931 159 GE, AD, SV hidroxilación
Valina (Val, V)
99,06841 72
Pueden formarse también fragmentos iónicos derivados de la rotura de dos enlaces, denominados
fragmentos internos. Los más frecuentes son los iones imonio, que contienen un solo aminoácido
(Figura 15). Estos iones imonio son característicos de cada aminoácido, por lo que permiten un análisis
cualitativo de la composición del péptido estudiando la zona de baja m/z del espectro.
45

Introducción
En instrumentos donde pueden llevarse a cabo colisiones de alta energía (como los de sectores y
los TOF-TOF), el perfil de fragmentación es diferente, produciéndose también roturas en otros lugares
de la cadena lineal (dando lugar a iones a, x, c y z).
5.4. IDENTIFICACIÓN DE PROTEÍNAS Y PÉPTIDOS
Tradicionalmente, las proteínas se han identificado mediante diferentes procedimientos, como la
secuenciación del extremo N-terminal (degradación secuencial de Edman), detección con anticuerpos
específicos, deleción de genes, etc. Estas técnicas son lentas, laboriosas y caras. A partir de los años 80
comienza a utilizarse la MS para identificar proteínas, a raíz de la implantación de las nuevas técnicas de
ionización suave. Hoy en día la MS se ha convertido en el método de elección para la identificación,
caracterización y cuantificación de proteínas, debido a su rapidez, elevada capacidad de análisis y alta
sensibilidad y precisión. Existen dos aproximaciones para identificar proteínas y péptidos utilizando
MS:
5.4.1. Huella peptídica (PMF)
La técnica más utilizada, por su rapidez y sencillez, es la identificación de proteínas mediante
huella peptídica (PMF, Peptide Mass Fingerprinting), utilizando instrumentos de MS tipo MALDI-TOF
(Henzel y col., 1993; Pappin y col., 1993). A partir de la digestión en gel de un spot o banda de interés
con proteasas específicas como la tripsina, se obtiene una mezcla de péptidos, específicos y
reproducibles para cada proteína, que es directamente analizada mediante MALDI-TOF. Las enzimas
proteolíticas actúan sobre enlaces específicos, así la tripsina corta el enlace peptídico después de un
residuo de Lys o Arg (excepto cuando estos aminoácidos van seguidos de un residuo de Pro). Como las
proteínas presentan por término medio un residuo de Lys o Arg por cada 10 aminoácidos, los péptidos
obtenidos en la digestión tríptica tendrán, en su mayoría, masas comprendidas entre los 800 y 2000 Da.
El espectro de MALDI-TOF del digerido proporciona las masas de una serie de péptidos procedentes de
dicha proteína, y en condiciones óptimas es específico, permitiendo su identificación inequívoca. Los
espectros obtenidos experimentalmente se contrastan con los millones de espectros teóricos generados
por la digestión virtual de las secuencias contenidas en las bases de datos, mediante diversos programas
informáticos como MASCOT (Perkins y col., 1999) (Figura 16). A cada uno de los resultados se le
asigna un valor probabilístico, teniendo en cuenta principalmente el número de péptidos que coinciden
entre nuestro espectro experimental y el espectro virtual obtenido de la base de datos.
46

Introducción
Figura 16. Mapa peptídico e identificación de una proteína. El espectro de masas de los
péptidos trípticos (A), obtenido por MALDI-TOF, se contrasta con una base de datos de
proteínas. (B) Representación gráfica de un resultado de la búsqueda, con las secuencias de
los péptidos coincidentes en rojo.
5.4.2. Fragmentación de péptidos
La identificación se lleva a cabo a partir del espectro MS/MS de uno o más péptidos obtenidos por
digestión enzimática, generalmente con tripsina, de la proteína de interés, siguiendo una de estas
aproximaciones:
- Huella de fragmentación peptídica (PFF, Peptide Fragmentation Fingerprint). Se basa en la
comparación del espectro de fragmentación sin interpretar con los espectros teóricos de los
péptidos producidos por digestión virtual de las proteínas contenidas en las bases de datos
(Figura 17), con la ayuda de programas bioinformáticos como MASCOT (Perkins y col.,
1999) y SEQUEST (Eng y col., 1994).
- Etiqueta de secuencia (Peptide Sequence Tag). Se basa en la identificación del péptido
mediante búsqueda en las bases de datos a partir de la interpretación parcial de una pequeña
secuencia de aminoácidos, de la masa del péptido y de las masas de los fragmentos que
bordean a la secuencia parcial (Mann y Wilm, 1994).
47

Introducción
Figura 17. Visión general del proceso de huella de fragmentación
peptídica. El espectro MS/MS obtenido experimentalmente se compara
con los espectros teóricos de los péptidos producidos por digestión virtual
de las proteínas contenidas en las bases de datos.
En los casos en los que la búsqueda en las bases de datos no proporciona resultados, se hace
necesaria la interpretación de novo del espectro para obtener la secuencia peptídica. Esta secuenciación
de novo de péptidos consiste en la determinación de la secuencia de aminoácidos que lo forman, a partir
de la interpretación directa del espectro de fragmentación (Figura 18), basándose para ello en una serie
de reglas que cumplen los iones peptídicos cuando son fragmentados, comúnmente mediante CID. La
precisión de la secuencia deducida está limitada por las características del equipo de MS y por la calidad
de la fragmentación obtenida. Este método es independiente de las bases de datos, por lo que es el
procedimiento más adecuado a la hora de identificar y caracterizar proteínas cuyas secuencias no estén
descritas en dichas bases de datos. La secuenciación de novo también permite estudiar mejor aquellas
asignaciones que mediante los anteriores motores de búsqueda han dado resultados dudosos. Por otra
parte, hay que tener en cuenta que la secuenciación de novo de péptidos a partir de su espectro de
fragmentación es un procedimiento laborioso, que exige experiencia por parte del usuario, y que puede
48
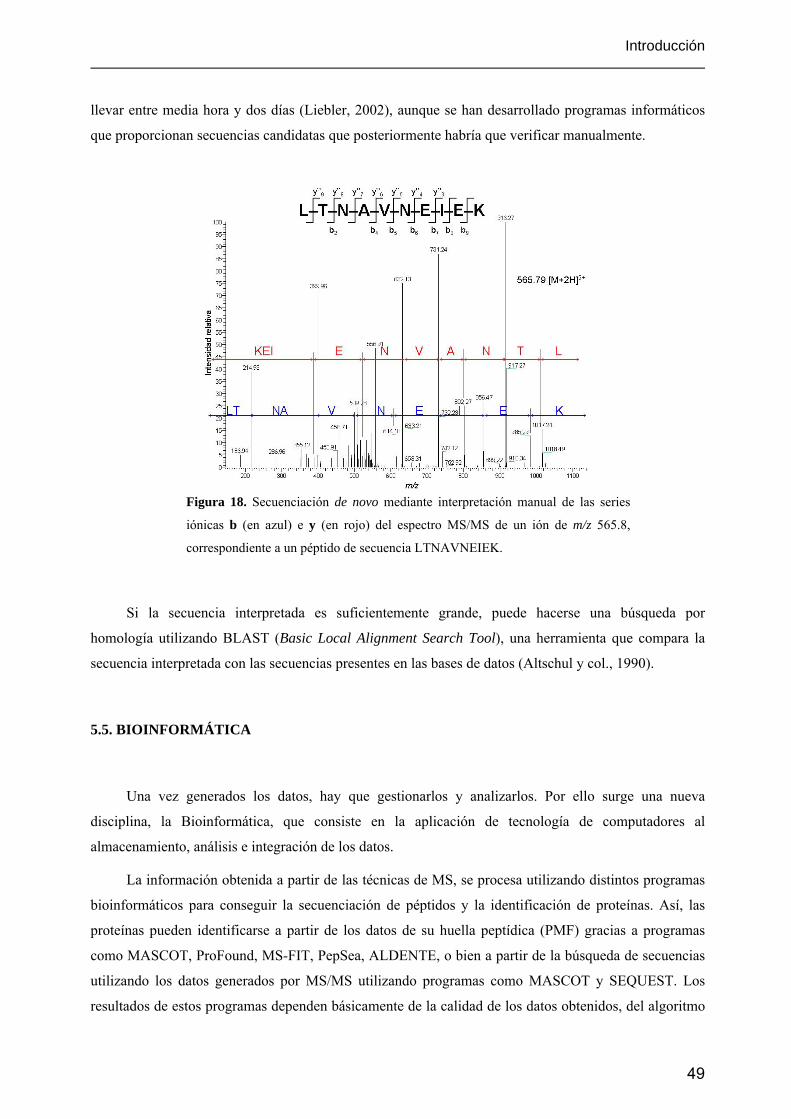
Introducción
llevar entre media hora y dos días (Liebler, 2002), aunque se han desarrollado programas informáticos
que proporcionan secuencias candidatas que posteriormente habría que verificar manualmente.
Figura 18. Secuenciación de novo mediante interpretación manual de las series
iónicas b (en azul) e y (en rojo) del espectro MS/MS de un ión de m/z 565.8,
correspondiente a un péptido de secuencia LTNAVNEIEK.
Si la secuencia interpretada es suficientemente grande, puede hacerse una búsqueda por
homología utilizando BLAST (Basic Local Alignment Search Tool), una herramienta que compara la
secuencia interpretada con las secuencias presentes en las bases de datos (Altschul y col., 1990).
5.5. BIOINFORMÁTICA
Una vez generados los datos, hay que gestionarlos y analizarlos. Por ello surge una nueva
disciplina, la Bioinformática, que consiste en la aplicación de tecnología de computadores al
almacenamiento, análisis e integración de los datos.
La información obtenida a partir de las técnicas de MS, se procesa utilizando distintos programas
bioinformáticos para conseguir la secuenciación de péptidos y la identificación de proteínas. Así, las
proteínas pueden identificarse a partir de los datos de su huella peptídica (PMF) gracias a programas
como MASCOT, ProFound, MS-FIT, PepSea, ALDENTE, o bien a partir de la búsqueda de secuencias
utilizando los datos generados por MS/MS utilizando programas como MASCOT y SEQUEST. Los
resultados de estos programas dependen básicamente de la calidad de los datos obtenidos, del algoritmo
49

Introducción
de búsqueda y de los parámetros utilizados para la búsqueda. Frecuentemente es necesaria una
verificación manual de los espectros MS/MS para asegurar que la asignación dada es correcta. Por otro
lado, la bioinformática también permite la secuenciación de novo a partir de espectros MS/MS mediante
la utilización de programas como DeNovoX (Thermo Fisher Scientific), PEAKS (Ma y col., 2003),
PepNovo (Frank y Pevzner, 2005) y SeqMS.
La bioinformática también ha propiciado la generación de programas de comparación automática
entre geles bidimensionales, destacando programas comerciales como PDQuest, Melanie, ImageMaster
2D y otros.
5.6. PROTEÓMICA Y ESPECIES MARINAS
La investigación en proteómica marina se centra actualmente en tres aspectos: estudios
medioambientales, estudios en microorganismos marinos, y estudios de interés alimentario (Tabla 10).
Dentro de estos últimos, las aplicaciones proteómicas podrían dividirse en tres grupos (Piñeiro y col.,
2003): aspectos nutricionales, seguridad alimentaria, y autentificación de especies.
Tabla 10. Resumen de las áreas de aplicación de la proteómica marina.
Área de aplicación Temática Referencias
Medioambiental Cangrejos como biomarcadores de contaminación
Gomiero y col., 2006
Estrés medioambiental en salmón Provan y col., 2006
Utilización de mejillón como biomonitorización de contaminación por petróleo
Mi y col., 2007
Biomarcadores de contaminación en mejillón
Knigge y col., 2004 Apraiz y col., 2006
Microorganismos marinos Revisión Schweder y col., 2008
Adaptación ecológica de patógenos Tunsjø y col., 2007
Ecología microbiana Thomas y col., 2007 Pedrós-Alió, 2006
Indicadores de contaminación marina Devereux y col., 2006
Proteínas con actividad patógena Evans y col., 2007 Descripción de subproteomas Redeby y col., 2006
Alimentos de origen marino Aspectos nutricionales Barnes y Kim, 2004 Kvasnicka, 2003
Seguridad y calidad alimentaria Erickson, 2005 Booy y col., 2005
Autentificación de especies
Piñeiro y col., 1998, 2001 López y col., 2002a, b Martínez y Friis, 2004 Carrera y col., 2006, 2007
50
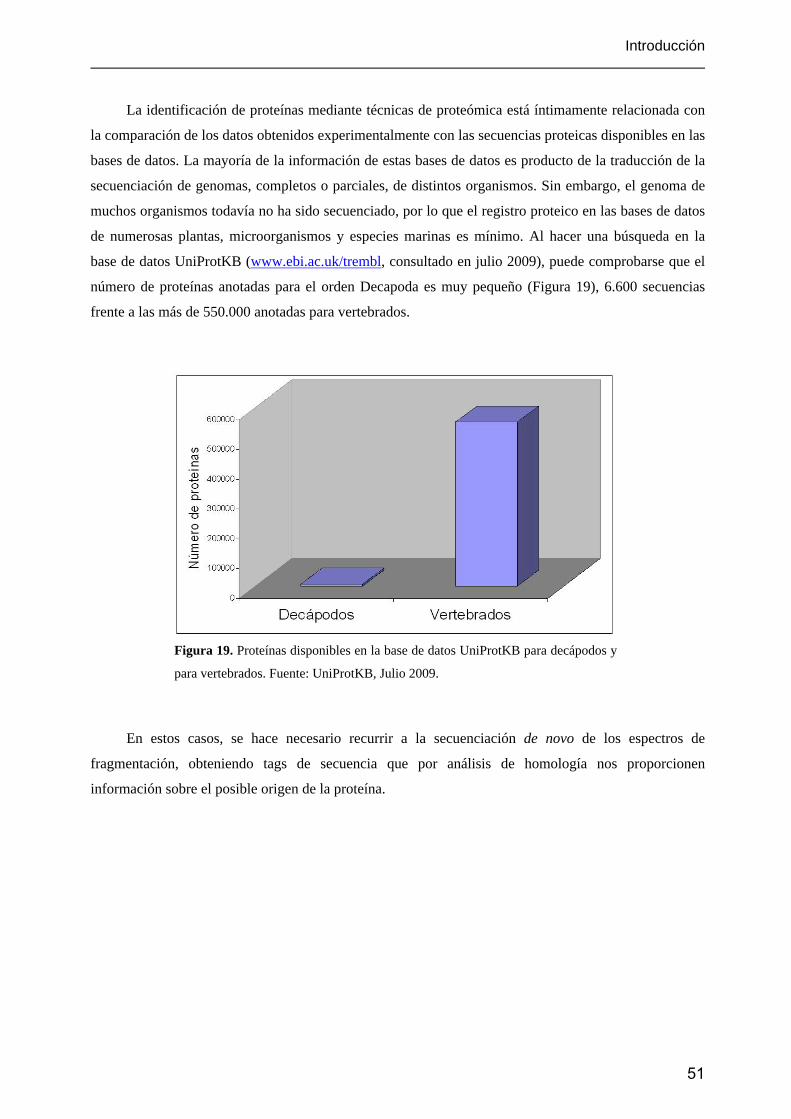
Introducción
La identificación de proteínas mediante técnicas de proteómica está íntimamente relacionada con
la comparación de los datos obtenidos experimentalmente con las secuencias proteicas disponibles en las
bases de datos. La mayoría de la información de estas bases de datos es producto de la traducción de la
secuenciación de genomas, completos o parciales, de distintos organismos. Sin embargo, el genoma de
muchos organismos todavía no ha sido secuenciado, por lo que el registro proteico en las bases de datos
de numerosas plantas, microorganismos y especies marinas es mínimo. Al hacer una búsqueda en la
base de datos UniProtKB (www.ebi.ac.uk/trembl, consultado en julio 2009), puede comprobarse que el
número de proteínas anotadas para el orden Decapoda es muy pequeño (Figura 19), 6.600 secuencias
frente a las más de 550.000 anotadas para vertebrados.
Figura 19. Proteínas disponibles en la base de datos UniProtKB para decápodos y
para vertebrados. Fuente: UniProtKB, Julio 2009.
En estos casos, se hace necesario recurrir a la secuenciación de novo de los espectros de
fragmentación, obteniendo tags de secuencia que por análisis de homología nos proporcionen
información sobre el posible origen de la proteína.
51


OBJETIVOS


OBJETIVOS
El objetivo general de la presente Tesis Doctoral es la descripción de nuevos procedimientos de
autentificación de las principales especies comerciales de langostinos del orden Decapoda, basados en el
análisis de proteínas y péptidos por electroforesis, cromatografía y espectrometría de masas, los cuales
supongan una mejora en términos de sensibilidad, especificidad, sencillez y rapidez frente a las
metodologías disponibles hasta la fecha.
Este objetivo general puede desglosarse en los siguientes objetivos específicos:
• Identificar patrones de bandas obtenidos por IEF que resulten específicos para las principales
especies comerciales de langostinos.
• Identificar las proteínas cuyas movilidades electroforéticas (Mr y/o pI), obtenidas por
electroforesis bidimensional, sean diferenciales para las principales especies comerciales de
langostinos.
• Identificar mediante espectrometría de masas tipo MALDI-TOF las variaciones en las huellas
peptídicas de las proteínas diferenciales anteriormente identificadas, con la finalidad de determinar
los picos específicos que permitan la diferenciación de las distintas especies de langostinos.
Desarrollar un nuevo método de identificación de las principales especies comerciales de
langostinos, mediante la comparación matemática de los espectros de huella peptídica de las
proteínas diferenciales previamente identificadas.
• Identificar mediante espectrometría de masas tipo ESI-IT variaciones en la secuencia
aminoacídica de los péptidos trípticos de las proteínas diferenciales anteriormente identificadas, y
caracterizar los péptidos diferenciales utilizando espectrometría de masas en tándem, mediante
ESI-IT o nESI-IT. La determinación de la secuencia de los péptidos específicos, tanto por
identificación mediante búsqueda en bases de datos proteicas como por secuenciación de novo,
permitirá la consideración de los mismos como péptidos biomarcadores para la identificación
inequívoca de las distintas especies de langostinos.
• Desarrollar un nuevo procedimiento de identificación rápida de las principales especies
comerciales de langostinos, mediante la monitorización, mediante cromatrografía en fase reversa
acoplada a espectrometría de masas, de los distintos péptidos específicos caracterizados en el
objetivo anterior.
55


MATERIALES Y MÉTODOS

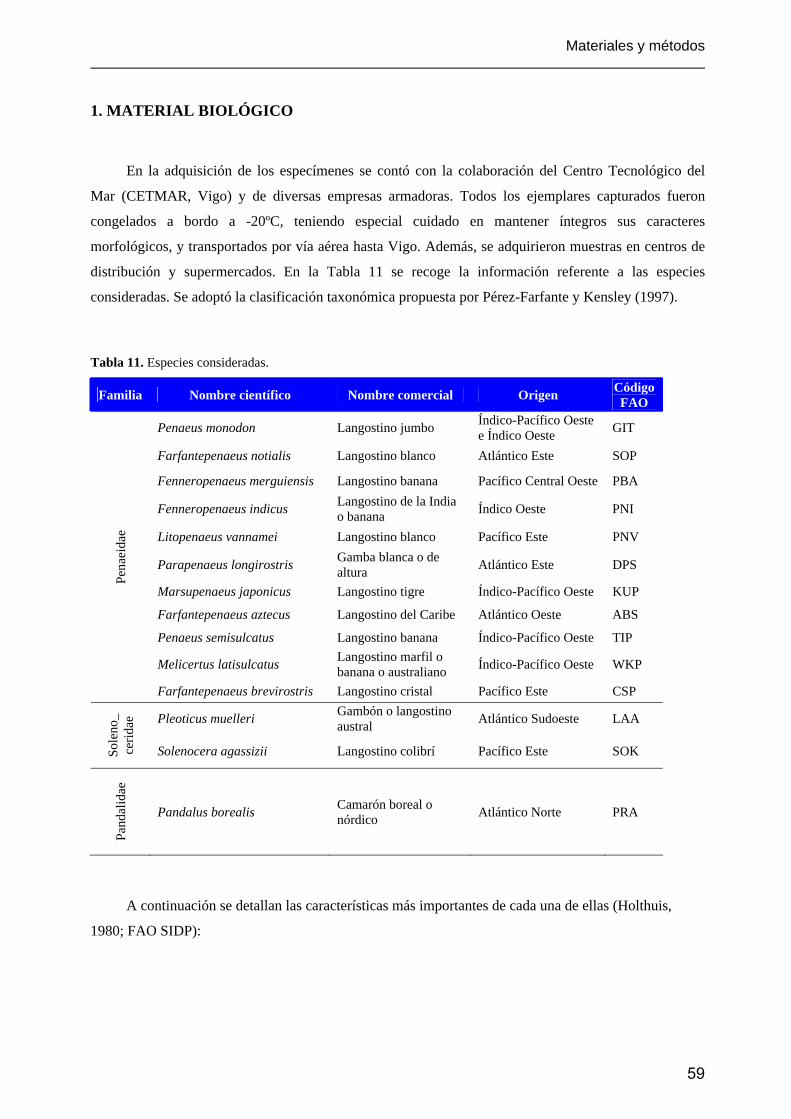
Materiales y métodos
1. MATERIAL BIOLÓGICO
En la adquisición de los especímenes se contó con la colaboración del Centro Tecnológico del
Mar (CETMAR, Vigo) y de diversas empresas armadoras. Todos los ejemplares capturados fueron
congelados a bordo a -20ºC, teniendo especial cuidado en mantener íntegros sus caracteres
morfológicos, y transportados por vía aérea hasta Vigo. Además, se adquirieron muestras en centros de
distribución y supermercados. En la Tabla 11 se recoge la información referente a las especies
consideradas. Se adoptó la clasificación taxonómica propuesta por Pérez-Farfante y Kensley (1997).
Tabla 11. Especies consideradas.
Familia Nombre científico Nombre comercial Origen Código FAO
Penaeus monodon Langostino jumbo Índico-Pacífico Oeste e Índico Oeste GIT
Farfantepenaeus notialis Langostino blanco Atlántico Este SOP
Fenneropenaeus merguiensis Langostino banana Pacífico Central Oeste PBA
Fenneropenaeus indicus Langostino de la India o banana Índico Oeste PNI
Litopenaeus vannamei Langostino blanco Pacífico Este PNV
Parapenaeus longirostris Gamba blanca o de altura Atlántico Este DPS
Marsupenaeus japonicus Langostino tigre Índico-Pacífico Oeste KUP
Farfantepenaeus aztecus Langostino del Caribe Atlántico Oeste ABS
Penaeus semisulcatus Langostino banana Índico-Pacífico Oeste TIP
Melicertus latisulcatus Langostino marfil o banana o australiano Índico-Pacífico Oeste WKP
Pena
eida
e
Farfantepenaeus brevirostris Langostino cristal Pacífico Este CSP
Pleoticus muelleri Gambón o langostino austral Atlántico Sudoeste LAA
Sole
no_
cerid
ae
Solenocera agassizii Langostino colibrí Pacífico Este SOK
Pand
alid
ae
Pandalus borealis Camarón boreal o nórdico Atlántico Norte PRA
A continuación se detallan las características más importantes de cada una de ellas (Holthuis,
1980; FAO SIDP):
59

Materiales y métodos
Farfantepenaeus aztecus (Ives, 1891)
De nombre común camarón café norteño, y nombre comercial langostino del caribe, F. aztecus
está muy restringido a la zona del Atlántico Oeste que va de Estados Unidos a México. Se distribuye en
un rango de profundidad de 4 a 160 m, ocupando los fondos fangosos. Los machos alcanzan los 195 mm
de longitud, y las hembras los 236 mm. Es de gran importancia en Estados Unidos, en los estados de
Texas y Carolina del Norte.
Farfantepenaeus brevirostris (Kingsley, 1878)
La distribución geográfica de esta especie, conocida como langostino cristal, está restringida a la
costa del Pacífico Este, desde México hasta el norte de Perú y las Islas Galápagos. Ocupa los fondos
lodosos y arenosos, en un rango de profundidades entre los 36 y los 120 m. El tamaño máximo que
alcanza es de 150 mm en los machos, y de 170 mm en las hembras. Esta especie tiene una considerable
importancia comercial en México, y también es capturada en Panamá y Ecuador.
Farfantepenaeus notialis (Pérez-Farfante, 1967)
Esta especie, de nombre común camarón rosado sureño, y nombre comercial langostino blanco, se
distribuye en el Atlántico, tanto en las costas africanas (de Mauritania a Angola) como en las costas de
Centro y Sudamérica (de México a Brasil). Habita en los fondos arenosos a profundidades entre 3 y 100
metros. Constituye una pesquería muy importante para Nigeria y Senegal.
Fenneropenaeus indicus (H. Milne Edwards, 1837)
El langostino blanco de la india, de nombre comercial langostino banana, está distribuido en el
Océano Índico y Pacífico Oeste, del este y sudeste de África al sudeste asiático y norte de Australia.
60

Materiales y métodos
Ocupa los fondos fangosos y arenosos de entre 2 y 90 m de profundidad. Alcanza longitudes máximas
de 184 mm (machos) y 228 mm (hembras). Es la especie pesquera de mayor importancia en
Mozambique, Kenia, Somalia y la India, y también Malasia y Tailandia la explotan comercialmente.
Fenneropenaeus merguiensis (De Man, 1888)
Esta especie, de nombre común langostino banana, se distribuye en los fondos fangosos del
Océano Índico y Pacífico Oeste, desde el Golfo Pérsico hasta Tailandia, Indonesia y el norte de
Australia. Vive a profundidades de entre 10 y 45 m. Es muy importante económicamente para Indonesia
y Tailandia, y frecuentemente se confunde con F. indicus.
Litopenaeus vannamei (Boone, 1931)
Esta especie, de nombre común camarón patiblanco y nombre comercial langostino blanco, habita
en el Pacífico Este, de México al norte de Perú. Se encuentra en los fondos fangosos de 0 a 72 m de
profundidad. Alcanza una longitud máxima de 230 mm. Tiene gran importancia económica en Ecuador,
Costa Rica y Colombia, y algo menor en México. Es el langostino de más importancia económica en
acuicultura, con 2.133.000 t de producción en el año 2006.
Marsupenaeus japonicus (Bate, 1888)
Esta especie, de nombre común langostino japonés, y nombre comercial langostino tigre, se
encuentra en el Océano Índico y Pacífico Oeste, desde el Mar Rojo y el sudeste de África hasta Corea,
Japón y el archipiélago malayo. Además se encuentra en el Mediterráneo oriental, alcanzando la costa
sur de Turquía, a donde llegó a través del Canal de Suez. Ocupa fondos arenosos y fangosos, de
profundidades entre 0 y 90 m. El tamaño máximo llega a los 190 mm del macho y los 225 mm de la
hembra. La pesca de esta especie es de gran importancia en Japón, donde tiene gran valor comercial.
También se captura en el este y sudeste de África, aunque aquí tiene una menor importancia.
Actualmente también se obtiene por acuicultura en Japón, Corea y Taiwán.
61

Materiales y métodos
Melicertus latisulcatus (Kishinouye, 1896)
Esta especie, de nombre común camarón real, y nombre comercial langostino marfil, banana o
australiano, se distribuye por el Océano Índico y Pacífico Oeste: desde el Mar Rojo y el Sudeste de
África, hasta Corea, Japón, Malasia y Australia. Ocupa costas arenosas o rocosas, entre los 0 y los 80 m
de profundidad. Alcanza una longitud máxima de 137 mm (macho). Es explotada comercialmente en
Australia, donde tiene gran valor comercial, en el sudeste africano y en la India. Además, se cultiva en
Tailandia.
Pandalus borealis Krøyer, 1838
El camarón boreal o nórdico se distribuye por el Atlántico Norte, desde el Mar del Norte a
Massachussets pasando por Groenlandia. Ocupa fondos arcillosos entre los 20 y los 1.380 m de
profundidad. Alcanza longitudes de 120 mm (machos) y 165 mm (hembras). Es el principal producto de
las pesquerías de decápodos del Atlántico Noroeste, capturándose intensivamente en Groenlandia,
Islandia y Noruega. Es una de las principales especies en cuanto a producción por captura (388.000 t en
el año 2006).
Parapenaeus longirostris (Lucas, 1846)
Conocido como gamba blanca o de altura, P. longirostris se distribuye en el Atlántico Este (de
Portugal a Angola), en el Mar Mediterráneo y en el Atlántico Oeste, del estado de Massachussets a la
Guayana Francesa. Se encuentra a partir de 20 m de profundidad, llegando hasta los 700 m, aunque
normalmente habita entre 150 y 400 m, en los fondos fangosos y arenosos. Normalmente no pasa de 140
mm (macho) y 160 mm (hembra) de longitud. El mayor número de capturas se produce en las costas
Mediterráneas de España, Italia y Francia, países en los que es muy apreciada.
62

Materiales y métodos
Penaeus monodon Fabricius, 1798
Esta especie, conocida como langostino jumbo o langostino tigre, se distribuye por las costas de
los Océanos Índico y Pacífico Oeste, del sudeste de África hasta Malasia, Japón y norte de Australia.
Habita los fondos fangosos y arenosos hasta profundidades de 110 metros. Alcanza longitudes de 336
mm y pesos de hasta 130 g. Debido a su gran tamaño, tiene gran importancia económica, destacando
Indonesia, Malasia e India entre los países que la explotan comercialmente. Es uno de los langostinos
con mayor producción, tanto por acuicultura como por captura.
Penaeus semisulcatus De Haan, 1844
Esta especie, de nombre común camarón tigre verde, pero nombre comercial langostino banana, se
confunde frecuentemente con P. monodon. P. semisulcatus se encuentra en el Océano Índico y Pacífico
Oeste, del Mar Rojo y sudeste de África a Japón, Corea, Malasia y norte de Australia. Además ha
alcanzado el Mediterráneo oriental a través del Canal de Suez, encontrándose en la actualidad en las
costas de Egipto, Israel, Líbano, Siria y sur de Turquía. Ocupa los fondos fangosos y arenosos, en
profundidades de 2 a 130 m. Los machos alcanzan una longitud de 180 mm, y las hembras de 228 mm.
Esta especie tiene una moderada importancia económica en el sudeste africano (Madagascar,
Mozambique y Somalia), Mar Rojo, Golfo Pérsico, Paquistán y Japón. También tiene relevancia en el
sudeste asiático, y su cultivo es importante en el delta del Ganges y en Tailandia.
Pleoticus muelleri (Bate, 1888)
Esta especie, conocida como langostino argentino o austral, se encuentra en el Atlántico Sudoeste,
principalmente en las costas de Argentina, aunque también en Brasil y Uruguay, en los fondos fangosos
63

Materiales y métodos
y arenosos a profundidades entre 2 y 100 m. Alcanza longitudes de hasta 190 mm, aunque la media está
en torno a los 100 mm. Es el crustáceo más importante comercialmente para Argentina.
Solenocera agassizii Faxon, 1893
Se conoce como langostino colibrí o chupaflor, y habita en aguas del Pacífico Este, de Costa Rica
a Ecuador y probablemente también en la costa norte de Perú. Su hábitat se sitúa entre los 280 y los 384
m de profundidad. Alcanza una longitud media de 149 mm (hembras). Esta especie es explotada
comercialmente por Costa Rica, Panamá y Nicaragua.
2. ANÁLISIS TAXONÓMICO DE EJEMPLARES DE REFERENCIA
En este tipo de estudios es imprescindible contar con especímenes de referencia, clasificados
perfectamente, de cada una de las especies objeto de estudio. Por ello fue necesario realizar previamente
un análisis taxonómico a todos los individuos susceptibles de ser material de referencia, para cada una
de las especies comprendidas en esta tesis.
Al no disponer en el momento de inicio de esta tesis de marcadores moleculares de las especies
del orden Decapoda que permitan la autentificación taxonómica de los individuos adquiridos, fue
necesaria una primera etapa de obtención de secuencias de referencia para cada una de las especies, con
las que posteriormente se compararon los especímenes que se estudiarían por técnicas proteómicas.
2.1. IDENTIFICACIÓN MORFOLÓGICA Y OBTENCIÓN DE SECUENCIAS DE
REFERENCIA
A su llegada al laboratorio, al menos 2 individuos pertenecientes a cada una de las potenciales
especies de langostinos fueron examinados y clasificados por taxónomos expertos, atendiendo a diversos
caracteres anatómicos y/o morfológicos. De la misma manera, la localización geográfica del buque en el
momento de la captura fue considerada como una información primordial para el mismo propósito. Una
vez perfectamente clasificados estos individuos, se tomó una muestra de músculo blanco abdominal –
limpio de restos de hemolinfa, vísceras y cutícula- de cada uno de ellos y se envió al Laboratorio de
64

Materiales y métodos
Higiene, Inspección y Control de Alimentos (LHICA) de la Universidad de Santiago de Compostela, a
efectos de establecer secuencias de ADN de referencia para cada especie.
2.2. IDENTIFICACIÓN GENÉTICA
Con el fin de asegurar la identificación taxonómica inequívoca de los especímenes a estudiar, cada
uno de ellos fue sometido a autentificación genética por comparación con las secuencias de ADN de
referencia establecidas previamente. Así, de cada ejemplar a estudiar se tomó una muestra de músculo
blanco limpio y se envió al LHICA para establecer genéticamente la identidad de cada espécimen.
3. EXTRACCIÓN DE LAS PROTEÍNAS SARCOPLÁSMICAS
La extracción de las proteínas sarcoplásmicas de cada uno de los especímenes objeto de estudio se
realizó en base a la característica de este grupo de proteínas de ser solubles en agua y en soluciones
salinas de baja fuerza iónica. El extracto se obtuvo a partir de 1 g de músculo blanco de la parte
comestible (cola o abdomen) de cada individuo. El trozo de tejido, limpio de restos de exoesqueleto,
cutícula, vísceras y hemolinfa, se cortó y trituró con un bisturí y una espátula, y se introdujo en tubos de
ensayo de 10 mL, a los que se añadió 2 mL de agua milliQ con PMSF 5 mM (fluoruro de fenil-metil-
sulfonilo, un inhibidor de proteasas). La muestra se homogenizó con un Ultra-Turrax (IKA-Werke,
Staufen, Alemania) a velocidad media, durante 2 ciclos de 15 s y a 4ºC (para evitar proteolisis).
Posteriormente, el homogenizado se centrifugó a 30.000 g durante 10 min a 4ºC (J221-M centrifugue;
Beckman, 16 Palo Alto, CA, EEUU) y el sobrenadante, conteniendo las proteínas sarcoplásmicas, se
repartió en tubos eppendorf para almacenarlo a -80ºC hasta su utilización.
La concentración de las proteínas solubilizadas se cuantificó mediante el método colorimétrico del
ácido bicinconínico (BCA, Sigma-Aldrich Co. St. Louis, EEUU). Para la elaboración del patrón de
proteínas se utilizaron disoluciones de albúmina de suero bovino (BSA), con un rango de
concentraciones conocidas entre 0,05 mg/mL y 1 mg/mL.
65

Materiales y métodos
4. ISOELECTROENFOQUE NATIVO
El isoelectroenfoque (IEF) se realizó en geles de poliacrilamida comerciales (Ampholine
PAGplate IEF, GE Healthcare, Uppsala, Suecia) en dos rangos de pH diferentes (Tabla 12). Los geles
(5% T, 3% C; dimensiones 250 x 110 x 1 mm) se procesaron a 10ºC en un equipo de electroforesis
horizontal Multiphor II (GE Healthcare) acoplado a un sistema de refrigeración MultiTemp III (GE
Healthcare). Los electrolitos del cátodo y del ánodo (Tabla 12) se aplicaron en dos tiras de papel de
filtro (IEF Electrode Strips, GE Healthcare) en ambos extremos del gel. Las muestras, 15 μL de cada
uno de los extractos de proteínas sarcoplásmicas a una concentración de 4 mg/mL, se aplicaron en
pequeñas piezas de papel de filtro (Sample Application Pieces, GE Healthcare) dispuestas a una
distancia de 1 cm del cátodo.
Tabla 12. Composición de las soluciones electrolíticas para los geles de IEF en función del gradiente
utilizado.
Rango de pH Solución anódica Solución catódica 4,0-6,5 Ácido glutámico 0,1 M en H3PO4 0,5 M β-alanina 0,1 M 4,0-5,0 H3PO4 1 M Glicina 1 M
Como patrones de punto isoeléctrico (pI) se utilizaron para todos los gradientes de pH dos
posibles mezclas de estándares proteicos de rango de pI amplio, Broad pI kit (GE Healthcare) o IEF
Markers 3-10 (Invitrogen, Carlsbad, EEUU), en función de su disponibilidad en el mercado, cuya
composición y pI para cada proteína se muestra en la Tabla 13. Al menos 2 calles de patrones de pI
Tabla 13. Composición de los patrones de punto isoeléctrico utilizados.
pI Broad pI kit pI IEF Markers 3-10 9,30 Tripsinógeno 10,7 Citocromo C 8,65 Lectina 9,5 Ribonucleasa A 8,45 Lectina 8,3 Lectina 8,15 Lectina 8,0 Lectina 7,35 Mioglobina 7,8 Lectina 6,85 Mioglobina 7,4 Mioglobina 6,55 Anhidrasa carbónica B humana 6,9 Mioglobina 5,85 Anhidrasa carbónica B bovina 6,0 Anhidrasa carbónica 5,20 Lactoglobulina A 5,3 β-lactoglobulina 4,55 Inhibidor de tripsina 5,2 β-lactoglobulina 3,50 Amiloglucosidasa 4,5 Inhibidor de tripsina
4,2 Glucosa oxidasa 3,5 Amiloglucosidasa
66

Materiales y métodos
fueron incluidas en cada gel en posiciones distintas para compensar posibles distorsiones entre calles de
un mismo gel.
Las condiciones de desarrollo de los geles para los diferentes rangos se muestran en la Tabla 14.
Tabla 14. Condiciones de desarrollo de los geles de IEF para los diferentes rangos.
Rango de pH Voltaje (V) Intensidad (mA) Potencia (W) 4,0-6,5 2.000 25 25 4,0-5,0 1.400 50 30
La electroforesis se dejó correr hasta alcanzar unos 4.000 Vh, tras lo que los geles se mantuvieron
en solución fijadora con ácido sulfosalicílico al 3,5% (p/v) y ácido tricloroacético al 11,5% (p/v), se
lavaron dos veces durante 5 min en solución desteñidora con etanol:ácido acético:agua (25:8:67) y se
tiñeron a 60ºC durante 5 min con azul brillante de Coomassie (CBB) R-350 (PhastGel Blue R, GE
Healthcare) al 0,04% (p/v) en solución desteñidora. A continuación, los geles fueron decolorados
mediante sucesivos lavados en solución desteñidora hasta conseguir un fondo transparente. Finalmente,
los geles fueron preservados por fijación con glicerol 10% (v/v), empaquetados con una membrana de
celofán y dejados secar a temperatura ambiente.
Los geles fueron escaneados y analizados mediante el programa LabImage 1D L340 versión 4.1
(Kapelan GmbH, Leipzig, Alemania), utilizando un método de ajuste lineal en base a los patrones de pI.
5. ELECTROFORESIS BIDIMENSIONAL
La primera dimensión se realizó en geles de poliacrilamida comerciales de rango medio, pH 4,0-
6,5 (Ampholine PAGplate, GE Healthcare), tal y como se describe en el apartado anterior, con las
siguientes modificaciones. En un mismo gel se cargaron por duplicado las distintas muestras a analizar,
cada una de ellas en cada una de las dos mitades del gel. En una de las mitades, la mitad analítica, se
cargaron 15 μL de cada uno de los extractos, y en la otra mitad, la mitad preparativa, se cargaron 30 μL,
siempre previo ajuste de la concentración proteica de los extractos a 4 mg/mL. Finalizada la
electroforesis, la mitad analítica fue teñida con CBB R-350 (GE Healthcare) y la mitad preparativa fue
recortada en tiras correspondientes a cada una de las muestras, las cuales fueron almacenadas a -80ºC
hasta su posterior utilización.
67

Materiales y métodos
Las tiras de gel provenientes de la primera dimensión fueron equilibradas a temperatura ambiente
durante 10 min con tampón de muestra para electroforesis (Laemmli, 1970) sustituyendo el β–
mercaptoetanol por ditiotreitol (DTT) al 0,75% (p/v), y a continuación durante otros 10 min con un
tampón similar al anterior pero sustituyendo el DTT por iodoacetamida al 4,5% y trazas de azul de
bromofenol. El DTT rompe los puentes disulfuro que existen en las proteínas, y la iodoacetamida
estabiliza los grupos –SH formados en el paso anterior para evitar que se formen de nuevo.
Una vez equilibradas las tiras, se realizó la segunda dimensión utilizando geles verticales (14 x
13,5 x 1,5 cm) de SDS-PAGE (7,5% T y 3% C) en tampón Tris-tricina (Schägger y von Jagow, 1987).
El crosslinker o agente entrecruzante fue N’N’-bis-metilen-acrilamida, y los agentes aceleradores de la
reacción, persulfato amónico (APS) al 10% (p/v) y N’N’N’N’-tetra-metil-etilendiamina (TEMED) al
0,05% (v/v). Los geles se desarrollaron en un sistema vertical (Hoefer SE600, Pharmacia Biotech Inc.,
San Francisco, CA, EEUU) a temperatura controlada de 15ºC por medio de un sistema de refrigeración
MultiTemp III (GE Healthcare). En cada gel se incluyeron patrones proteicos de baja masa molecular
(Low Molecular Weight, GE Healthcare). La electroforesis se desarrolló a una intensidad constante de
40 mA por gel, 100 V y 150 W durante 12-14 horas, utilizando como tampón para el cátodo Tris 0,1 M,
Tricina 0,1 M, SDS 0,1%, pH 8,25 y para el ánodo Tris 0,2 M, pH 8,9.
Finalizada la electroforesis, los geles se revelaron mediante tinción con CBB R-350 (GE
Healthcare) al 0,04 % (p/v) disuelto en disolución desteñidora compuesta por etanol 25%, ácido acético
8% y agua 67%. A continuación, los geles fueron decolorados mediante sucesivos lavados en solución
desteñidora hasta conseguir un fondo transparente. Los geles fueron escaneados y se analizaron
mediante el programa de análisis de imagen de geles bidimensionales PDQuest versión 7.1.0 (Bio-Rad
Laboratories, Hercules, CA, EEUU). Tras ello, los spots de interés fueron recortados para su posterior
análisis mediante espectrometría de masas (apartado 6). Finalmente, los geles fueron preservados por
fijación con glicerol 10% (v/v), empaquetados con una membrana de celofán y dejados secar a
temperatura ambiente.
6. DIGESTIÓN DE LAS PROTEÍNAS EN GEL
Los spots de interés fueron recortados en condiciones estériles de los geles 2-DE, maximizando la
relación proteína/gel, de manera que la zona recortada fue la región más intensa de cada spot. Los spots
fueron digeridos en gel con tripsina (Sequencing Grade; Roche Diagnostics GmbH, Mannheim,
Alemania), siguiendo el protocolo descrito por Jensen y col. (1999) con pequeñas modificaciones. Así,
la pieza de gel, cortada en piezas menores, se lavó 2 veces con 50 μL de agua MilliQ durante 10 min en
68

Materiales y métodos
agitación. Tras ello, se deshidrató 2 veces con 40 μL de acetonitrilo (ACN) durante 10 min con
agitación, y se rehidrató con 30 μL de bicarbonato amónico 25 mM durante 5 min en agitación. Tras la
rehidratación, se adicionaron sobre el bicarbonato amónico 30 μL de ACN y se incubó la muestra
durante 15 min en agitación. Seguidamente, la muestra se secó en un concentrador de vacío speed-vac
RVC-2-18/Alpha 1-2 (Christ, Osterode am Harz, Alemania) y se rehidrató de nuevo con 20 μL de una
disolución de tripsina 12,5 ng/μL en bicarbonato amónico 25 mM pH 8. La muestra se mantuvo en hielo
durante al menos 45 min para que la tripsina penetrase en el gel, tras lo que se retiró la disolución de
tripsina que no fue absorbida. Se adicionaron 25 μL de bicarbonato amónico 12,5 mM pH 8, y se incubó
durante al menos 14 horas a 37ºC.
7. MAPEO PEPTÍDICO MEDIANTE ESPECTROMETRÍA DE MASAS MALDI-TOF
7.1. OBTENCIÓN DE PMF
El análisis mediante espectrometría de masas tipo MALDI-TOF de los péptidos trípticos se llevó a
cabo en un espectrómetro de masas Voyager DE STR (Applied Biosystems, Foster City, CA, EEUU)
operando en modo reflectrón, extracción retardada e ión-positivo. Después de la digestión tríptica de los
spots, la disolución peptídica se secó en un concentrador de vacío y se acidificó mediante 5 μL de ácido
trifluoroacético (TFA) 0,1%. Como soporte para la ionización MALDI se utilizaron placas de acero
inoxidable de 100 pocillos (Applied Biosystems), en las que se depositaron 0,8 μL del extracto peptídico
que se dejaron secar a temperatura ambiente. Una vez seco, se depositaron sobre él 0,8 μL de la solución
de matriz, consistente en una disolución saturada de ácido α–ciano-4-hidroxicinámico (CHCA) en ACN
al 50% (v/v) con TFA al 0,1% (v/v), y también se dejo secar a temperatura ambiente.
Los valores de los parámetros del espectrómetro de masas fueron: low mass gate, m/z 500 Da;
tiempo de retardo, 350 ns; voltaje de aceleración, 20.000 V; voltaje de rejilla (grid voltage), 68,5%. Se
utilizó una intensidad del láser justo sobre el umbral de la generación de iones. Los espectros fueron
adquiridos promediando 150 disparos del láser, en el rango de m/z de 850 a 3500. Los espectros se
calibraron externamente utilizando una mezcla estándar de péptidos compuesta por péptidos sintéticos
(Calibration Mixture 2 del Sequazyme Peptide Mass Standards Kit; Applied Biosystems): angiotensina I
(MH+ 1.296,69), ACTH 1-17 (MH+ 2.093,09), ACTH 18-39 (MH+ 2.465,20), ACTH 7-38 (MH+
3.657,93), Insulina (MH+ 5.730,61). Para asegurar la mayor precisión, estos calibrantes se añadieron a la
placa en posiciones adyacentes a las muestras. Además, los espectros se calibraron internamente
69

Materiales y métodos
utilizando los picos de los péptidos resultantes de la autodigestión de la tripsina (MH+ 1.153,57,
2.163.06, 2.273,16 y 2.289,16).
7.2. PROCESAMIENTO DE LOS ESPECTROS DE MALDI-TOF
Los espectros de masas obtenidos por MALDI-TOF se procesaron con el software del instrumento
(Data Explores, version 4.0.0.0; Applied Biosystems). Se corrigió la línea base, se eliminaron los picos
no monoisotópicos, y se extrajeron los valores de m/z monoisotópicos con intensidades mayores al 5%
en el rango de m/z de 920 a 3500. Las listas de picos obtenidas se analizaron visualmente para eliminar
todas aquellas masas procedentes de la autodigestión de la tripsina, de la matriz, y de otras proteínas
contaminantes, como ciertas queratinas.
Las listas de picos se utilizaron para la identificación de proteínas y péptidos mediante búsqueda
en bases de datos por PMF (Peptide Mass Fingerprinting). Para ello se utilizó el motor de búsqueda
MASCOT (http://www.matrixscience.com) y la base de datos no redundante del NCBI (NCBInr)
(http://www.ncbi.nih.gov; NCBI Resources, NIH, Bethseda, MD, EEUU), utilizando los siguientes
parámetros: masas monoisotópicas y protonadas, hasta 2 sitos de corte de la tripsina fallidos, tolerancia
de masa del péptido de 100 ppm, y tres modificaciones variables: acetilación del extremo N-terminal,
cisteína carbamidometilada, y metionina sulfóxido.
8. ANÁLISIS FILOGENÉTICO DE LOS ESPECTROS PMF
Se compararon los espectros MALDI-TOF de varios individuos mediante análisis cluster. Para
ello, se extrajeron de los espectros MALDI-TOF los valores de m/z monoisotópicos con intensidades
mayores al 5% en el rango de m/z de 920 a 3000. La lista de picos de cada individuo se integró en una
matriz de picos con 2 posibles valores: presencia del pico (valor “1”) o ausencia del pico (valor “0”).
Cuando la diferencia de masa entre el pico mi en la lista de picos del individuo i y el pico mj en la lista
de picos del individuo j era menor de 200 ppm, los dos picos se supusieron el mismo péptido,
permitiendo este error en la medida exacta de las masas. A partir de la matriz de presencia/ausencia de
picos, se generó una matriz de distancias, para lo que se utilizó el software Windist (del paquete
Winboot, disponible en www.irri.org), empleando el coeficiente de similitud de Ochiai (Ochiai, 1957).
Según este coeficiente, la medida de la similitud, Si,j, entre la lista de picos del individuo i y la lista de
picos del individuo j se define como (1):
70

Materiales y métodos
ji
ijji
nnaS =, (1)
donde ni y nj son el número total de picos de cada lista individual de picos, y aij es el número de picos en
común entre las dos listas de picos, i y j. En la matriz de distancias se recogió no el coeficiente de
similitud, sino el coeficiente de distancia, di,j, que se calcula sustrayendo el coeficiente de similitud Si,j
de 1 (2).
di,j = 1 - Si,j (2)
El valor de di,j está por tanto limitado al rango 0 ≤ di,j ≤ 1. Cuando las listas de picos i y j son
indistinguibles, di,j = 0; cuando las listas de picos son completamente distintas, di,j = 1.
La matriz de distancias entre listas de picos se representó como un dendrograma utilizando el
software MEGA (Tamura y col., 2007), empleando el algoritmo neighbor-joining (Saitou y Nei, 1987).
Para evaluar el soporte estadístico de la topología obtenida se realizó un bootstrap con 1000 réplicas.
9. ANÁLISIS DE ADN
9.1. EXTRACCIÓN DEL ADN
La extracción del ADN de las muestras se llevó a cabo mediante un método comercial de
extracción de ADN a partir de tejidos, el DNeasy Tissue kit (Qiagen, Hilden, Alemania). Se
homogenizaron 250 mg de músculo de cada individuo a analizar, utilizando bisturí y pinzas, y se dejó
incubando con tampón de lisis durante toda la noche a 55ºC. Una vez completada la lisis celular, el
ADN se purificó utilizando las columnas incluidas en el DNeasy Tissue kit, siguiendo las indicaciones
del fabricante. Resumidamente, consiste en pasar la muestra por pequeñas columnas, tras lo que se lavan
las columnas con distintos buffers incluidos en el kit, para finalmente eluir el ADN con un buffer de
elución. En cada uno de los pasos –ligado del ADN, 2 etapas de lavado de la columna, y elución del
ADN- se centrifugaron las columnas a 8.000 rpm durante 1 min –excepto en la segunda etapa de lavado,
71

Materiales y métodos
que se centrifugaron a 13.000 rpm durante 3 min- utilizando una microcentrífuga modelo 5415 D
(Eppendorf, Hamburgo, Alemania).
El ADN extraído se cuantificó por el método de Downs y Wilfinger (1983), que consiste en la
determinación de la fluorescencia obtenida al mezclar un volumen conocido del extracto de ADN con el
reactivo Hoechst 33258 (Sigma, St. Louis, MO, EEUU). Este reactivo está compuesto por bis-
benzimida, que se intercala entre las moléculas de ADN y tiene unas longitudes de onda de excitación y
de emisión de 356 nm y 458 nm, respectivamente. Las medidas se llevaron a cabo en un fluorímetro LS
50 (Perkin Elmer, Waltham, MA, EEUU).
9.2. AMPLIFICACIÓN DEL ADN
Una vez purificado el ADN, se procedió a amplificar mediante PCR una zona del ADNmt,
concretamente en la región 16S rRNA-tRNAVal. La amplificación del ADN se llevó a cabo en el
termociclador GeneAmp PCR System 2700 (Applied Biosystems), utilizando los cebadores o primers
16ScruC3 (5′-CGTTGAGAAGTTCGTTGTGCA-3′) y 16ScruC4 (5′-
AATATGGCTGTTTTTAAGCCTAATTCA-3′) (Pascoal y col., 2008a).
La mezcla de la reacción de PCR consistió en:
- 100 ng de ADN molde.
- 25 μL de una mezcla de reacción comercial (BioMix, Bioline, Londres, Reino Unido), que
incluye el buffer de la reacción, los deoxirribonucleótidos trifosfato (dNTPs), cloruro de
magnesio y la Taq DNA polimerasa.
- 25 pmol de cada cebador.
- agua PCR (Genaxis, Montigny le Bretonneaux, Francia) hasta alcanzar un volumen de
reacción de 50 μL.
Las condiciones de la amplificación fueron:
- desnaturalización inicial a 94ºC durante 90 s.
- 35 ciclos de desnaturalización (94ºC durante 20s), anillamiento (51-55ºC durante 20 s) y
extensión (72ºC durante 30 s).
- extensión final a 72ºC durante 15 min.
72

Materiales y métodos
9.3. SECUENCIACIÓN
Previamente a la secuenciación, los productos de PCR amplificados fueron purificados mediante
el kit ExoSAP-IT (GE Healthcare) con el fin de eliminar componentes de la reacción que pudieran
interferir en la reacción de secuenciación. La secuenciación directa de los amplicones purificados se
realizó mediante el método comercial BigDye Terminator v3.1 Cycle Sequencing Kit (Applied
Biosystems). Para llevar a cabo las reacciones de secuenciación, se emplearon los mismos cebadores
utilizados en la PCR, realizándose la secuenciación de las dos hebras de ADN. Las reacciones de
secuenciación se analizaron en un secuenciador capilar ABI 3730XL DNA Analyzer (Applied
Biosystems).
9.4. ANÁLISIS FILOGENÉTICO
Los cromatogramas de las muestras secuenciadas se revisaron visualmente mediante el software
CHROMAS versión 1.45 (Technelysium Pty, Tewantin, Australia). La secuencias en formato FASTA se
alinearon con el programa ClustalX versión 1.83 (Thompson y col., 1997), y a partir de estas secuencias
se realizó el análisis filogenético utilizando el programa MEGA versión 4.0 (Tamura y col., 2007). El
árbol filogenético se obtuvo empleando el algoritmo neighbor-joining, realizando un bootstrap con 1000
réplicas para evaluar el soporte estadístico de la topología obtenida.
10. CARACTERIZACIÓN DE PÉPTIDOS MEDIANTE ESPECTROMETRÍA DE
MASAS EN TÁNDEM
Los digeridos proteicos fueron analizados bien online mediante LC-ESI-IT-MS/MS, o bien offline
mediante nESI-IT-MS/MS, con objeto de identificar los péptidos obteniendo sus espectros de
fragmentación.
10.1. LC-ESI-IT-MS/MS
El análisis online se realizó mediante LC-ESI-IT-MS/MS, utilizando un sistema de HPLC modelo
SpectraSystem P4000 (Thermo Fisher Scientific, San José, CA, EEUU) acoplado a un equipo de MS de
73

Materiales y métodos
trampa iónica tridimensional modelo LCQ Deca XP Plus (Thermo Fisher Scientific), con fuente de
ionización de tipo microspray.
El extracto peptídico obtenido de la digestión se retiró de las piezas de gel y se secó a vacío en un
speed-vac (Christ). Posteriormente, se resuspendió la muestra en 10 μL de ácido acético al 5%, se sonicó
en un baño de ultrasonidos 5 min y se inyectaron 5 μL en el equipo LC-MS. La separación
cromatrográfica fue realizada en una columna de fase reversa tipo BioBasic C18 (0,18 mm x 150 mm)
(Thermo Fisher Scientific), utilizando un gradiente lineal del 5% al 60% del componente B de la fase
móvil durante 90 minutos, a un flujo de trabajo de 1,5-1,8 μL/min (fase móvil A: 0,5% de ácido acético
(v/v); fase móvil B: 80% ACN (v/v) en 0,5% ácido acético (v/v)). Para la electronebulización se
estableció una diferencia de potencial de 3,5 kV entre la salida del capilar y la entrada del espectrómetro
de masas, aplicándose un flujo de N2 de 10 unidades arbitrarias para favorecer la ionización. La
temperatura del capilar se fijó en 200ºC.
Los péptidos fueron detectados y analizados realizando el seguimiento denominado Triple Play
(Figura 20), que consiste en la sucesión de los siguientes tres eventos: 1) un barrido completo (Full
Scan) de 300 a 1300 amu (3 μscans), en el que, mediante el modo de trabajo “Data-dependent”, se
seleccionaron los iones de mayor intensidad a los que se les realizó, 2) un barrido de alta resolución
(Zoom Scan) (5 μscans), y 3) un evento de MS/MS (5 μscans), utilizando una ventana de aislamiento de
3 amu y una energía de colisión normalizada del 35%. Los picos elegidos para ser fragmentados fueron
excluidos de una posible nueva fragmentación durante 3 min, utilizando el procedimiento de exclusión
dinámica. Los iones monocargados fueron descartados para sus posterior análisis mediante MS/MS.
Figura 20. Esquema del modo de trabajo Triple Play.
74

Materiales y métodos
10.2. nESI-IT-MS/MS
El análisis offline se realizó utilizando un espectrómetro de masas con trampa iónica
tridimensional modelo LCQ Deca XP Plus (Thermo Fisher Scientific), equipado con una fuente de
ionización por nanospray. El digerido peptídico fue sometido previamente a un proceso de limpieza,
desalado y concentración, utilizando microcolumnas C18 tipo ZipTip (Millipore Co., Bedford, MA,
EEUU). Los péptidos se eluyeron con 10 μL de 70% MeOH/0,5% CH3COOH. 3 μL de esta disolución
se cargaron en un nanocapilar de borosilicato (PicoTips, New Objective, Woburn, MA, EEUU), con un
diámetro de salida de 1 μm. Los parámetros del instrumento se fueron ajustando durante el análisis,
utilizando un voltaje de spray entre 0,7 y 1,3 kV y una temperatura del capilar de desolvatación de
150ºC. Para la fragmentación de los péptidos, la anchura de la ventana de selección del ión precursor fue
de 3 amu, y la energía de colisión se ajustó entre el 35% y el 45% en función del estado de carga del ión
precursor. Para cada péptido estudiado se obtuvo un barrido de alta resolución (ZoomScan) y otro de
fragmentación (MS/MS), promediando la señal alrededor de 1 min.
10.3. PROCESAMIENTO DE LOS DATOS DE MS/MS
Los espectros de fragmentación obtenidos se analizaron mediante el motor de búsqueda
SEQUEST (Bioworks Browser 3.1 package, Thermo Fisher Scientific), frente a la base de datos no
redundante NCBInr. Los parámetros introducidos para la realización de las búsquedas fueron: masas
monoisotópicas, hasta dos sitios de corte consecutivos de la enzima fallidos, tolerancia de masa del ión
precursor de ±1,5 Da y de los iones fragmento de ±0,8 Da, y dos posibles modificaciones, C* (cisteína
carbamidometilada) y Mox (metionina sulfóxido). Además se indicó que el enzima utilizado fue
tripsina. Los espectros obtenidos fueron confirmados manualmente para evitar falsos positivos.
Cuando no se consiguió una adecuada identificación del péptido, bien porque el espectro era de
calidad insuficiente para ser comparado por SEQUEST, o bien porque el péptido no se encontraba
descrito en las bases de datos, se procedió a su secuenciación de novo mediante interpretación manual de
las series de iones fragmento, con ayuda del software PEAKS Studio 4.2 (Bioinformatics Solutions,
Waterloo, Ontario, Canadá).
Las secuencias obtenidas fueron analizadas mediante el programa BLAST: protein blast,
http://www.ncbi.nlm.nih.gov/BLAST (Altschul y col., 1990), utilizando la base de datos NCBInr, con
objeto de encontrar secuencias homólogas en proteínas descritas previamente.
75

Materiales y métodos
11. DESARROLLO DE UN PROCEDIMIENTO DE ANÁLISIS RÁPIDO POR MS
PARA LA IDENTIFICACIÓN DE LAS PRINCIPALES ESPECIES COMERCIALES
DE LANGOSTINOS
Una vez identificados y caracterizados los péptidos que nos permiten diferenciar entre las distintas
especies de langostinos, se desarrolló un método rápido para detectar la presencia en una muestra de
dichos péptidos. Se procedió a digerir directamente el extracto proteico, con el fin de evitar la
electroforesis bidimensional, que es una etapa que consume mucho tiempo. Se realizaron 2 tipos de
digestiones en solución del extracto proteico sarcoplásmico, bien una digestión tradicional, o bien una
digestión ultrarrápida utilizando ultrasonidos focalizados de alta intensidad (HIFU). Así mismo, para la
monitorización de los péptidos trípticos resultantes de la digestión se utilizó el método de barrido
“monitorización de la fragmentación de un ión seleccionado” (Selected MS/MS Ion Monitoring, SMIM)
en el espectrómetro de masas, previa separación de los péptidos en un HPLC acoplado online.
11.1. DIGESTIÓN EN SOLUCIÓN TRADICIONAL
Se secaron a vacío en un speed-vac 100 μg de proteínas del extracto sarcoplásmico, y se
desnaturalizaron las proteínas por adición de 25 μL de urea 8 M en bicarbonato amónico pH 8,0. Tras
sonicar la muestra en un baño de ultrasonidos, se añadieron 3 μL de DTT, de modo que su concentración
final fuese 10 mM, y se incubó a 37ºC durante 1 h. A continuación se añadió iodoacetamida a una
concentración final de 50 mM, y se incubó en oscuridad a temperatura ambiente durante 1 h.
Transcurrido este tiempo, la mezcla se diluyó 4 veces con bicarbonato amónico 25 mM con el fin de
reducir la concentración de urea presente en la muestra, y se digirió durante al menos 16 horas a 37ºC
con tripsina (Sequencing Grade; Roche Diagnostics GmbH) a una relación de 1:50 tripsina:proteína.
11.2. DIGESTIÓN EN SOLUCIÓN ULTRARRÁPIDA CON HIFU
100 μg del extracto sarcoplásmico se digirieron enzimáticamente con tripsina en un volumen final
de 104 μL, aplicando simultáneamente HIFU (López-Ferrer y col., 2005). Para ello se añadió a la
muestra tripsina (Sequencing Grade; Roche Diagnostics GmbH) en una relación 1:25 tripsina:proteína, y
se introdujo una sonda de ultrasonidos (calibre de 2 mm, intensidad 50%, Sonics & Materials modelo
Vibra Cell CV 18, Newtown, CT, EEUU) durante 1 min.
76

Materiales y métodos
11.3. SEPARACIÓN Y MONITORIZACIÓN DE PÉPTIDOS MEDIANTE SMIM
Como etapa recomendable, pero opcional, los péptidos resultantes de la digestión se purificaron y
desalaron a través de columnas de fase reversa C18 tipo ZipTip (Millipore Co.), eluyendo los péptidos
en 10 μL de 0,1% ácido fórmico/70% ACN. Una vez eliminado el ACN mediante evaporación, se
inyectaron 5 μL del eluído conteniendo los péptidos trípticos en un sistema HPLC (Thermo Fisher
Scientific), acoplado online a un espectrómetro de masas con una fuente de ionización microspray y con
una trampa iónica como analizador (LCQ Deca XP, Thermo Fisher Scientific). La separación se realizó
a través de una columna tipo fase reversa (0,18 mm x 150 mm) (Thermo Fisher Scientific), a un flujo de
1,5-2 µL/min. Los péptidos fueron eluidos en un gradiente de 65 min del 5% al 45% de solvente B
(Solvente A: 0,5% ácido acético (v/v); solvente B: 0,5% ácido acético (v/v), 80% ACN (v/v)). A medida
que los péptidos iban eluyendo de la columna cromatográfica, el espectrómetro de masas configurado en
modo SMIM realizó una continua monitorización de la fragmentación de los iones doblemente cargados
de cada uno de los péptidos específicos descritos en el texto (capítulo 5). Para la electronebulización, se
estableció una diferencia de potencial de 3,5 kV y un flujo de N2 de 10 unidades arbitrarias. La
temperatura del capilar se fijó en 200ºC y los eventos de fragmentación se realizaron con una energía de
colisión normalizada del 35% y una apertura de la ventana de aislamiento del ión precursor de 3 amu.
Para cada ión precursor se representó el cromatrograma “virtual” de uno de los iones fragmento
(masa del ión precursor→masa del ión fragmento), utilizando el software QualBrowser (Thermo Fisher
Scientific).
77


RESULTADOS Y DISCUSIÓN


CAPÍTULO 1 IDENTIFICACIÓN DE ESPECIES DE LANGOSTINOS Y GAMBAS DE INTERÉS ALIMENTARIO POR ISOELECTROENFOQUE


Capítulo 1: Identificación de especies de langostinos y gambas de interés
alimentario por isoelectroenfoque.
Comentario
Aunque los métodos moleculares o las técnicas proteómicas son cada vez más frecuentemente
utilizados en los laboratorios de control de productos marinos, los métodos electroforéticos clásicos
como el IEF, que separa proteínas en función de su punto isoeléctrico, han demostrado ser una técnica
barata, rápida y fácil de aplicar. Es más, la identificación de especies utilizando IEF de la fracción
proteica sarcoplásmica es el único método oficial validado. Sin embargo, no existen trabajos que
apliquen esta técnica a la identificación de las principales especies comerciales de langostinos. Por ello,
es conveniente el desarrollo de un estudio que incluya un número importante de estas especies.
En este trabajo se describen los patrones de bandas que resultan de someter a IEF a las fracciones
proteicas sarcoplásmicas de 14 especies de langostinos y camarones de interés comercial. Estos perfiles
electroforéticos resultaron ser especie-específicos, permitiendo la inequívoca diferenciación de cada una
de las especies consideradas. Las bandas más características fueron analizadas por espectrometría de
masas, identificándose como isoformas de una proteína conocida como Sarcoplasmic Calcium-binding
Protein (SCP), perteneciente a la misma familia que las parvalbúminas, proteínas presentes en pescado y
descritas como especie-específicas y termoestables.
Estos resultados demuestran que los perfiles electroforéticos diferenciales, debidos a la presencia
de isoformas de SCPs, permiten una rápida y efectiva identificación de las especies de decápodos
consideradas.
83


PUBLICACIÓN
85


Elsevier Editorial System(tm) for Food Chemistry
Manuscript Draft
Manuscript Number: FOODCHEM-D-09-00690
Title: Identification of commercial prawn and shrimp species of food interest by native isoelectric
focusing
Article Type: Research Article
Keywords: IEF; shrimps; species identification; SCP; MS/MS
Corresponding Author: Mr Ignacio Ortea,
Corresponding Author's Institution: Instituto de Investigaciones Marinas
First Author: Ignacio Ortea
Order of Authors: Ignacio Ortea; Benito Cañas; Pilar Calo-Mata; Jorge Barros-Velázquez; José M
Gallardo
Abstract: Native isoelectric focusing (IEF) of water-soluble sarcoplasmic proteins was applied to the
identification of 14 shrimp species of food interest belonging to the order Decapoda. These species
have different commercial values, but due to their phenotypic similarities and carapace removal in
their industrial processing, incorrect food labelling and deliberate or inadvertent adulteration can
occur. Each of the 14 tested species showed species-specific protein band profiles and intra-
specific polymorphism was low, not preventing the correct identification of the species. Therefore,
IEF of water-soluble sarcoplasmic proteins allowed the differentiation of the 14 species considered.
In addition, sarcoplasmic calcium-binding proteins (SCPs) were identified by tandem mass
spectrometry (MS/MS) as the major species-specific proteins in these species, opening the way to
further studies focusing on their potential use as specific biomarkers.
87

Identification of commercial prawn and shrimp
species of food interest by native isoelectric
focusing
Ignacio Orteaa,*, Benito Cañasb, Pilar Calo-Matac, Jorge Barros-
Velázquezc and José M. Gallardoa
a Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Científicas
(CSIC), Eduardo Cabello 6, E-36208 Vigo, Spain
b Department of Analytical Chemistry; School of Chemistry, Complutense University of
Madrid. E-28040 Madrid, Spain
c Department of Analytical Chemistry, Nutrition and Food Science; School of
Veterinary Sciences, University of Santiago de Compostela. E-27002 Lugo, Spain
* Corresponding author. Tel.: +34-986-231-930; fax: +34-986-292-762.
E-mail address: [email protected] (I. Ortea).
Running title: Shrimp species identification by IEF.
88

Abstract
Native isoelectric focusing (IEF) of water-soluble sarcoplasmic proteins was applied to
the identification of 14 shrimp species of food interest belonging to the order Decapoda.
These species have different commercial values, but due to their phenotypic similarities
and carapace removal in their industrial processing, incorrect food labeling and
deliberate or inadvertent adulteration can occur. Each of the 14 tested species showed
species-specific protein band profiles and intra-specific polymorphism was low, not
preventing the correct identification of the species. Therefore, IEF of water-soluble
sarcoplasmic proteins allowed the differentiation of the 14 species considered. In
addition, sarcoplasmic calcium-binding proteins (SCPs) were identified by tandem mass
spectrometry (MS/MS) as the major species-specific proteins in these species, opening
the way to further studies focusing on their potential use as specific biomarkers.
Keywords: IEF, shrimps, species identification, SCP, MS/MS.
1. Introduction
Seafood products include a wide variety of species with a significant importance for
food industry. Among them, crustaceans belonging to the order Decapoda, comprising
prawns, shrimps, lobsters, crayfish, crabs and hermit crabs, are of remarkable
commercial interest. This order includes the superfamily Penaeoidea, the penaeid
shrimps, which are the most important economic resource in the world’s crustacean
fishery and aquafarming industry (Dall, Hill, Rothlisberg & Staples, 1990; Holthius,
1980; Pérez-Farfante & Kensley, 1997).
89

Food authentication is a major concern not only for the prevention of commercial
fraud, but also for the assessment of safety risks derived from the inadvertent
introduction of any food ingredient that might be harmful to human health (Brzezinski,
2005; Civera, 2003; Lockley & Bardsley, 2000). Thus, identification of marine species
is an issue of capital relevance for the seafood industry, because of commercial
requirements concerning labelling and traceability. In response to consumer concern,
global regulations have been implemented to assure complete and correct information,
guaranteeing market transparency and avoiding substitution of certain fish species by
others of less commercial value (Council Regulation, 2000; Royal Decree, 2004).
Anatomical characters are particularly difficult to use for penaeid shrimps species
differentiation due to their phenotypic similarities and to the fact that in their processing
the external carapace is often removed. This may lead to mislabelled and/or adulterated
products, so unintentional fraud may occur. Therefore, the increasing demand of
crustaceans in general, and of high quality penaeid shrimps in particular, may be
compromised by deliberate or inadvertent adulteration along the food chain by the
substitution of higher quality species by others of lower commercial quality. Therefore,
it is highly recommendable the development of the necessary analytical tools to make
possible distinguishing among closely related species.
Methods based on protein analysis, including sodium dodecyl sulphate-
polyacrylamide gel electrophoresis (SDS-PAGE), isoelectric focusing electrophoresis
(IEF) (Berrini, Tepedino, Borromeo & Secchi, 2006; Chen, Shiau, Noguchi, Wei &
Hwang, 2003; Etienne et al., 2001; Mackie et al., 2000; Piñeiro et al., 1999a) and two-
dimensional gel electrophoresis (2-DE) (Piñeiro, Barros-Velázquez, Sotelo & Gallardo,
1999b) have been extensively used for fish species identification. Immunoassay
(Asensio, González, García & Martín, 2008) and, more recently, mass spectrometry
90

(MS) methodologies (Carrera, Cañas, Piñeiro, Vázquez & Gallardo, 2006; Mazzeo et
al., 2008) have also been used for species identification.
Molecular biological methods as mitochondrial DNA (mtDNA) analysis has been
used in polymerase chain reaction (PCR) based studies for fish species identification
(Rasmussen & Morrissey, 2008). Among the mtDNA targets, the 16S rRNA gene and
the cytochrome oxidase I gene have been reported to serve as good interspecific
markers in some crustacean species, although most of these studies were focused not on
species identification, but on population structures, phylogeography and phylogenetic
relationships (Baldwin, Bass, Bowen & Clark, 1998; Lavery, Chan, Tam & Chu, 2004;
Maggioni, Rogers, Maclean & Incao, 2001; Quan, Zhuang, Deng, Dai & Zhang, 2004;
Voloch, Freire & Russo, 2005). More recently, two PCR-restriction fragment length
polymorphism (RFLP) methods for the detection of crustaceans (Brzezinski, 2005) and
penaeid shrimps (Pascoal, Barros-Velázquez, Cepeda, Gallardo & Calo-Mata, 2008)
have been proposed.
Although the molecular methods or the proteomic technologies will be the future in
the seafood control laboratories, the classic electrophoretic methods such as IEF, which
separates proteins based on their differences in isoelectric point (pI), has revealed as a
cheaper, faster, easy to apply by the control laboratories and less sophisticated
technology than the DNA-based techniques for species identification purposes (Piñeiro
et al., 1999a). In this context, little effort has been made to elucidate differences among
closely related shrimp species using these protein-based methodologies. Related
previous reports described a SDS-PAGE study which was not able to discriminate
among close related species (Civera & Parisi, 1991), and two urea IEF studies including
only 3 shrimp species (An, Marshall, Otwell & Wei, 1989; Wei, Chen & Marshall,
1990).
91

In this study, we used native IEF for the unequivocal identification of 14 shrimp
species of commercial interest for the food industry worldwide.
2. Materials and methods
2.1. Fish material
14 different prawn and shrimp species of commercial interest were considered (Table
1), 13 species belonging to the superfamily Penaeoidea and one species –the northern
shrimp Pandalus borealis- to the superfamily Pandaloidea. Specimens were collected
using extractive fishing practices or from aquaculture facilities in different continents
worldwide. Shrimps were frozen on board and shipped to our laboratory for the
analyses. Six individuals from each species were analyzed. When required, specimens
were classified in their respective taxons according to their anatomical external features
with the help of a marine biologist from the Marine Sciences Institute (Mediterranean
Centre for Marine and Environmental Research, Higher Council for Scientific Research,
CMIMA-CSIC, Barcelona, Spain) with expertise in penaeid shrimp taxonomy.
2.2. Extraction of sarcoplasmic proteins
Extraction of the sarcoplasmic proteins was performed by homogenizing 1 g of raw
white muscle from each individual in two volumes of milliQ water, using an Ultra-
Turrax blender for 3x15 s–with interruptions of 45 s to avoid warming the samples. The
extracts were then centrifuged at 30 000g for 15 min at 4ºC (J25 centrifuge; Beckman,
Palo Alto, USA), and the supernatants were maintained at -80ºC until the
electrophoretic analysis. Protein concentration in the extracts was determined by the
bicinchoninic acid method (Sigma-Aldrich, St. Louis, USA).
2.3. IEF
92

Native IEF was conducted by using Ampholine PAGplate precast polyacrylamide
gels in two different pH ranges (pH 4.0-6.5, pH 4.0-5.0; GE Healthcare, Uppsala,
Sweden), and was performed at 10ºC in a Multiphor II electrophoresis unit (Amersham
Biosciences, Sweden) with a thermostatic circulator. Most of the specific bands
described for other fish species are in the pH range 4.0-7.0 (Piñeiro et al., 1999b), so
narrow acidic pH ranges were chosen. IEF electrode strips were soaked in 0.1 M
glutamic acid in 0.5 M H3PO4 for anode and 0.1 M β–alanine for cathode (4.0-6.5 pH
range); and 1 M H3PO4 for anode and 1 M glycine for cathode (4.0-5.0 pH range). 40 μg
of total proteins were loaded near the cathode using sample application pieces (GE
Healthcare). Two lanes of protein standards in the 3.5-10.7 pH range (IEF Markers 3-
10, Invitrogen, Carlsbad, USA) were included in each gel. The running conditions were
1500 V – 50 mA – 30 W, until 4000 V·h was reached, with 30 min prefocusing. 60
minutes after the initiation of electrophoresis, the application pieces were removed to
avoid any smearing of proteins on the gel. Following electrophoresis, proteins were
fixed for 30 minutes and gels were stained with Coomassie Brilliant Blue (GE
Healthcare) equilibrated in 23% ethanol and 8% acetic acid, and dried in air between
sheets of cellophane.
2.4. Analysis of patterns
IEF gels were scanned and the acquired images were analyzed using the LabImage
1D software (version 4.1, Kapelan GmbH, Leipzig, Germany). Intra-gel distortions
between lines were compensated by aligning the band position of the two lines of
standards loaded at different positions on the gel. Different gels were normalized for
differences in running conditions by comparing the pattern of pI standard proteins
separated in the gel to be analyzed with the pI standard pattern chosen as a database
standard.
93

2.5. Protein identification of characteristic bands
Major characteristic bands were separated in a closer pH range (pH 4.0-5.0) for
identification by means of MS analysis. They were excised from gel and subjected to in-
gel digestion with trypsin (Roche Diagnostics GmbH, Mannheim, Germany), performed
overnight at 37 ºC as described elsewhere (Jensen, Wilm, Shevchenko & Mann, 1999).
Final digestion solution was dried under vacuum and resuspended in 5 % acetic acid.
The produced peptides were subjected to tandem mass spectrometry (MS/MS) analysis,
using an ion trap mass spectrometer model LCQ Deca XP Plus (Thermo-Finnigan, San
Jose, USA). Peptides were fragmented, obtaining MS/MS spectra; these spectra were
then used for database searching against the nr.fasta database (NCBI Resources, NiH,
Bethseda, USA) using SEQUEST (Bioworks 3.1 package, Thermo-Finnigan).
3. Results and discussion
3.1. Native IEF
Native IEF profiles of sarcoplasmic proteins, extracted from the muscle of 14 shrimp
species, were obtained and the patterns analyzed for species identification. Most of the
specific bands described for fish species are in the pH range 4.0-7.0 (Piñeiro et al.,
1999b), so a narrow acidic pH range (4.0-6.5) was chosen for the analysis. Six
specimens of each of the species were analyzed, in either the same or different gels.
Figure 1 represents the fourteen different protein patterns from the shrimp species. Each
protein fingerprint is composed of a series of protein bands, which are characteristic for
each species, allowing an easy discrimination. IEF patterns exhibited many species-
specific bands over a pI range of 4.0-6.5, with most of the bands in the 4.2-5.0 range.
Band patterns were scanned, the characteristic bands identified and their pI calculated.
Table 2 reports a schematic diagram of the patterns showing the average pI (±RSD)
94

values of characteristic bands determined using the LabImage 1D software according to
the procedure described in the section 2.4. Firstly, the pIs were determined for each gel,
and then the average pI values and RSD for each band were calculated. The RSD values
varied between 0.0 and 2.3, indicating the good precision for this method. The band
patterns were consistent for the six specimens analyzed for each of the species under
study. Low intra-specific polymorphism was found, only in 3 of the species: a sample of
F. notialis showed a significant band at pI 5.22 which was not visible in the other five;
two samples of P. semisulcatus showed a substitution of a band at pI 4.74 by a band at
pI 5.07; and two samples of P. monodon showed a triplet where other four samples had
a unique band (pI 4.95). However, the presence of these changes did not prevent correct
identification of the species. It might be necessary to test larger number of individuals
from each species to detect intra-specific differences. However, in the analytical
conditions used in this work, the inter-specific band variation is much greater than the
intra-specific variation and good enough for correct species identification.
3.2. Protein identification of major characteristic bands
Major characteristic bands were in-gel digested with trypsin and subjected to MS/MS
analysis. To achieve an unequivocal identification of the proteins presented in the
specific bands, an optimal separation of the possible marker proteins was needed, so
preparative native IEF was performed using an even closer pH range (pH 4.0-5.0). The
following bands were selected for mass spectrometry characterization (Figure 2): band
1, pI 4.96; band 2, pI 4.76; band 3, pI 4.93; band 4, pI 4.53; band 5, pI 4.49; and band 6,
pI 4.86. Several peptides (between 4 and 10) were produced after tryptic digestion of
each of these bands, and all of them matched with some of the five Decapoda
Sarcoplasmic Calcium-binding Proteins (SCPs) described in databases (Table 3): SCP α
chain from Penaeus sp. (accession P02636), SCP β chain from Penaeus sp. (P02635),
95

SCP from Procambarus clarkii (ABB58783), SCP I from Pontastacus leptodactylus
(P05946), and SCP αB from Fenneropenaeus orientalis (1006232A). In Figure 3,
several fragmentation spectra as examples of the SCP peptides found are shown,
together with the mass-to-charge ratio (m/z) of the precursor ions. Therefore, all the
bands studied were identified by MS/MS as isoforms of the SCP, a soluble protein
found in invertebrate muscle, and constituent, like parvalbumins, of the EF-hand-type
family of the calcium-binding proteins. These proteins, previously described as species-
specific proteins (Piñeiro et al., 1999b; Rehbein, 1995), belong to a group of acidic and
heat-stable proteins, which accounts for the majority of the water extractable proteins
from fish sarcoplasma. Recently, SCPs have also been established as crustacean
allergens (Shiomi, Sato, Hamamoto, Mita & Shimakura, 2008), and interspecific
variability in their amino acid sequences has been described as the cause of differences
in IgE reactivity to SCP in sensitized individuals.
4. Conclusions
To the best of our knowledge, the results reported here are representative of the most
complete electrophoretic method described to date for the identification of penaeid
shrimp species of commercial interest. The specific patterns allowed the unambiguous
differentiation of all the 14 tested species. Therefore, this simple and fast technique
could be usable by fisheries and related industries as a good tool for preventing
mislabelling and fraudulent practices, providing a rapid and effective identification and
guaranteeing market transparency. SCPs exhibited extensive polymorphism between
species, a very useful property for electrophoretic species identification, as described
previously (Rehbein, 1995). Inter-specific variability in the pI of SCPs isomers suggest
96

the potential presence of aminoacid substitutions in their sequences, opening the way to
further studies focusing on their potential use as specific biomarkers.
Acknowledgements
We thank Ms Lorena Barros for her excellent technical assistance. We also
acknowledge members from CETMAR for their helpful collaboration in the collection
of specimens for this study. This work was supported by the National Food Program of
the INIA (Spanish Ministry for Education and Science) (Project CAL-03-030-C2-2) and
by the PGIDIT Research Program in Marine Resources (Project
PGIDIT04RMA261004PR) of the Xunta de Galicia (Galician Council for Industry,
Commerce and Innovation). I.O. is supported by the FPU program (AP-2004-5826) and
B.C. is supported by the RyC program, under the auspices of the Spanish MICINN.
Abbreviations used
2-DE, two-dimensional gel electrophoresis; IEF, isoelectric focusing; MS, mass
spectrometry; MS/MS, tandem mass spectrometry; mtDNA, mitochondrial DNA; m/z,
mass-to-charge ratio; PCR, polymerase chain reaction; pI, isoelectric point; RFLP,
restriction fragment length polymorphism; RSD, relative standard deviation; SCP,
sarcoplasmic calcium-binding protein; SDS-PAGE, sodium dodecyl sulphate-
polyacrylamide gel electrophoresis.
References
An, H., Marshall, M. R., Otwell, W. S., & Wei, C. I. (1989). Species identification of
raw and boiled shrimp by a urea gel isoelectric focusing technique. Journal of Food
Science, 54(2), 233-236.
97

Asensio, L., González, I., García, T., & Martín, R. (2008). Determination of food
authenticity by enzyme-linked immunosorbent assay (ELISA). Food Control, 19(1),
1-8.
Baldwin, J. D., Bass, A. L., Bowen, B. W., & Clark, W. H. (1998). Molecular
phylogeny and biogeography of the marine shrimp Penaeus. Molecular
Phylogenetics and Evolution, 10, 399-407.
Berrini, A., Tepedino, V., Borromeo, V., & Secchi, C. (2006). Identification of
freshwater fish commercially labelled "perch" by isoelectric focusing and two-
dimensional electrophoresis. Food chemistry, 96(1), 163-168.
Brzezinski, J. L. (2005). Detection of crustacean DNA and species identification using a
PCR-restriction fragment length polymorphism method. Journal of Food
Protection, 68, 1866-1873.
Carrera, M., Cañas, B., Piñeiro, C., Vázquez, J., & Gallardo, J. M. (2006). Identification
of commercial hake and grenadier species by proteomic analysis of the parvalbumin
fraction. Proteomics, 6(19), 5278-5287.
Civera, T. (2003). Species identification and safety of fish products. Veterinary
Research Communications, 27, 481-489.
Civera, T., & Parisi, E. (1991). The use of sodium dodecyl sulfate polyacrylamide gel
electrophoresis for the identification of raw and cooked shrimps and crabs. Italian
Journal of Food Science, 3(2), 9.
Council Regulation (2000). Council Regulation (EC) No. 104/2000 of 17 December
1999 on the common organization of the markets in fishery and aquaculture
products. Official Journal of the European Communities L 17/22.
98

Chen, T.-Y., Shiau, C.-Y., Noguchi, T., Wei, C.-i., & Hwang, D.-F. (2003).
Identification of puffer fish species by native isoelectric focusing technique. Food
chemistry, 83(3), 475-479.
Dall, W., Hill, B. J., Rothlisberg, P. C., & Staples, D. J. (1990). The biology of
Penaeidae. Advances in Marine Biology, 27, 1-484.
Etienne, M., Jérôme, M., Fleurence, J., Rehbein, H., Kündiger, R., Mendes, R., Costa,
H., & Martínez, I. (2001). Species identification of formed fishery products and high
pressure-treated fish by electrophoresis: a collaborative study. Food chemistry,
72(1), 105-112.
Holthius, L. B. (1980). FAO species catalogue. Volume 1. Shrimps and prawns of the
world. An annotated catalogue of species of interest to fisheries. FAO Fisheries
Synopsis, 125(1), 1-271.
Jensen, O. N., Wilm, M., Shevchenko, A., & Mann, M. (1999). Sample preparation
methods for mass spectrometric peptide mapping directly from 2-DE gels. In: A. J.
Link, 2-D proteome analysis protocols (pp. 513-530). Totowa, NJ: Humana Press.
Lavery, S., Chan, T. Y., Tam, Y. K., & Chu, K. H. (2004). Phylogenetic relationships
and evolutionary history of the shrimp genus Penaeus s.l. derived from
mitochondrial DNA. Molecular Phylogenetics and Evolution, 31(1), 39-49.
Lockley, A. K., & Bardsley, R. G. (2000). DNA-based methods for food authentication.
Trends in Food Science & Technology, 11(2), 67-77.
Mackie, I., Craig, A., Etienne, M., Jérôme, M., Fleurence, J., Jessen, F., Smelt, A.,
Kruijt, A., Malmheden Yman, I., Ferm, M., Martinez, I., Pérez-Martín, R., Piñeiro,
C., Rehbein, H., & Kündiger, R. (2000). Species identification of smoked and
gravad fish products by sodium dodecylsulphate polyacrylamide gel electrophoresis,
99

urea isoelectric focusing and native isoelectric focusing: a collaborative study. Food
chemistry, 71(1), 1-7.
Maggioni, R., Rogers, A. D., Maclean, N., & Incao, F. (2001). Molecular phylogeny of
western Atlantic Farfantepenaeus and Litopenaeus shrimp based on mitochondrial
16S partial sequences. Molecular Phylogenetics and Evolution, 18, 66-73.
Mazzeo, M. F., Giulio, B. D., Guerriero, G., Ciarcia, G., Malorni, A., Russo, G. L., &
Siciliano, R. A. (2008). Fish authentication by MALDI-TOF mass spectrometry.
Journal of Agricultural and Food Chemistry, 56(23), 11071-11076.
Pascoal, A., Barros-Velázquez, J., Cepeda, A., Gallardo, J. M., & Calo-Mata, P. (2008).
A polymerase chain reaction-restriction fragment length polymorphism method
based on the analysis of a 16S rRNA/tRNAVal mitochondrial region for species
identification of commercial penaeid shrimps (Crustacea: Decapoda: Penaeoidea)
of food interest. Electrophoresis, 29(2), 499-509.
Pérez-Farfante, I., & Kensley, B. (1997). Penaeoid and sergestoid shrimps and prawns
of the world. Keys and diagnoses for the families and genera. Paris: Éditions du
Muséum National d’Histoire Naturelle.
Piñeiro, C., Barros-Velázquez, J., Pérez-Martín, R. I., Martínez, I., Jacobsen, T.,
Rehbein, H., Kündiger, R., Mendes, R., Etienne, M., Jerome, M., Craig, A., Mackie,
I. M., & Jessen, F. (1999a). Development of a sodium dodecyl sulfate-
polyacrylamide gel electrophoresis reference method for the analysis and
identification of fish species in raw and heat-processed samples: A collaborative
study. Electrophoresis, 20(7), 1425-1432.
Piñeiro, C., Barros-Velázquez, J., Sotelo, C. G., & Gallardo, J. M. (1999b). The use of
two-dimensional electrophoresis in the characterization of the water-soluble protein
100

fraction of commercial flat fish species. Zeitschrift für Lebensmitteluntersuchung
und -Forschung A, 208(5), 342-348.
Quan, J., Zhuang, Z., Deng, J., Dai, J., & Zhang, Y.-p. (2004). Phylogenetic
relationships of 12 Penaeoidea shrimp species deduced from mitochondrial DNA
sequences. Biochemical Genetics, 42(9), 331-345.
Rasmussen, R. S., & Morrissey, M. T. (2008). DNA-based methods for the
identification of commercial fish and seafood species. Comprehensive Reviews in
Food Science and Food Safety, 7(3), 280-295.
Rehbein, H. (1995). Food control by applied biochemistry of marine organisms:
Comparison of proteins and metabolites from fish and invertebrate muscle.
Helgoland Marine Research, 49(1), 747-757.
Royal Decree (2004). Royal Decree 121/2004 of 23 January 2004 related to the
identification of live, fresh or refrigerated fish and fishery products. Ministry of
Agriculture, Fisheries and Food, Spain. BOE number 31, of 5 February 2004.
Shiomi, K., Sato, Y., Hamamoto, S., Mita, H., & Shimakura, K. (2008). Sarcoplasmic
calcium-binding protein: identification as a new allergen of the black tiger shrimp
Penaeus monodon. International Archives of Allergy and Immunology, 146(2), 91-
98.
Voloch, C. M., Freire, P. R., & Russo, C. A. M. (2005). Molecular phylogeny of
penaeid shrimps inferred from two mitochondrial markers. Genetics and Molecular
Research, 4(4), 668-674.
Wei, C.-I., Chen, H. A. J.-S., & Marshall, M. R. (1990). Use of a modified urea gel
isoelectric focusing method for species identification of raw or boiled white, pink,
and rock shrimp. Journal of Food Biochemistry, 14(2), 91-102.
101

Figure captions
Fig. 1. Native IEF patterns (pH range 4.0-6.5) of water-soluble sarcoplasmic proteins
representative of the 14 shrimp species tested.
Fig. 2. IEF in polyacrylamide gel (pH range 4.0-5.0) of water-soluble sarcoplasmic
proteins of four shrimp species, showing bands selected for MS/MS analysis.
Fig. 3. Ion-trap MS/MS spectrum of the SCPs peptides a) DGEVTVDEFK, present in
all bands analyzed; b) VFIANQFK, present in bands 1, 2 ,3, 4 and 6; and c)
YM*YDIDNDGFLDK, present in bands 1, 2, 4, 5 and 6. M*, methionine sulfoxide.
102
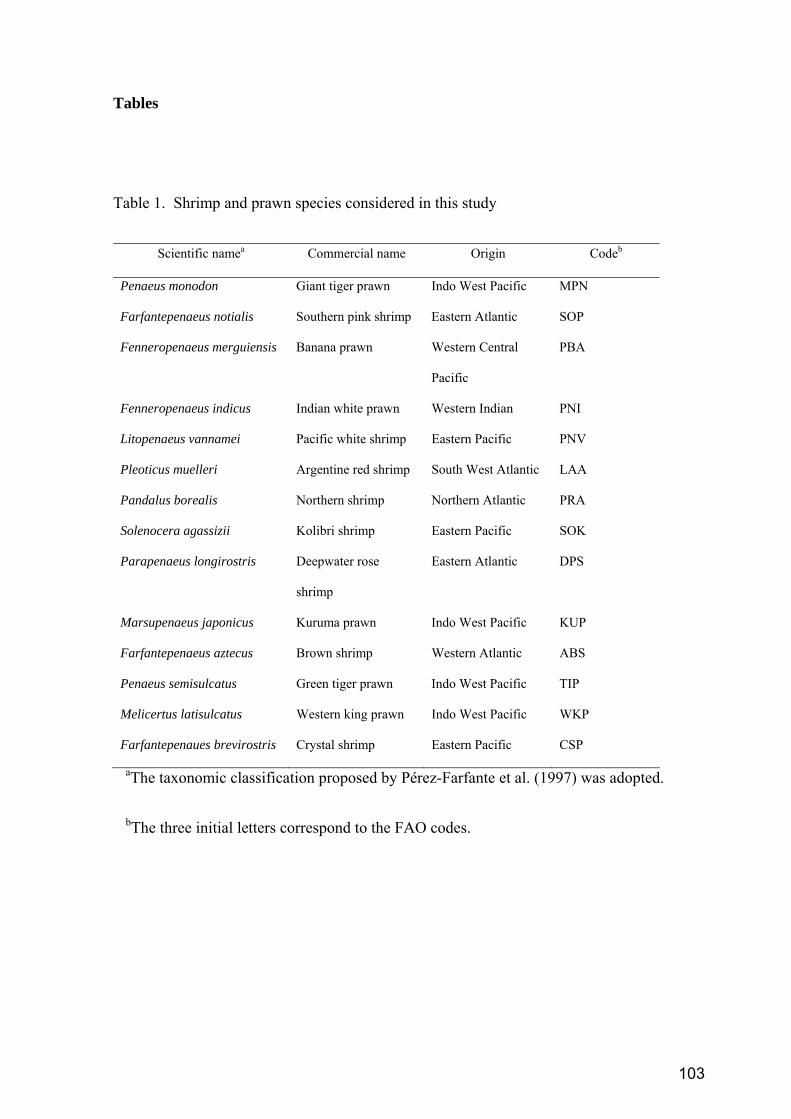
Tables
Table 1. Shrimp and prawn species considered in this study
Scientific namea Commercial name Origin Codeb
Penaeus monodon Giant tiger prawn Indo West Pacific MPN
Farfantepenaeus notialis Southern pink shrimp Eastern Atlantic SOP
Fenneropenaeus merguiensis Banana prawn Western Central
Pacific
PBA
Fenneropenaeus indicus Indian white prawn Western Indian PNI
Litopenaeus vannamei Pacific white shrimp Eastern Pacific PNV
Pleoticus muelleri Argentine red shrimp South West Atlantic LAA
Pandalus borealis Northern shrimp Northern Atlantic PRA
Solenocera agassizii Kolibri shrimp Eastern Pacific SOK
Parapenaeus longirostris Deepwater rose
shrimp
Eastern Atlantic DPS
Marsupenaeus japonicus Kuruma prawn Indo West Pacific KUP
Farfantepenaeus aztecus Brown shrimp Western Atlantic ABS
Penaeus semisulcatus Green tiger prawn Indo West Pacific TIP
Melicertus latisulcatus Western king prawn Indo West Pacific WKP
Farfantepenaues brevirostris Crystal shrimp Eastern Pacific CSP
aThe taxonomic classification proposed by Pérez-Farfante et al. (1997) was adopted.
bThe three initial letters correspond to the FAO codes.
103

Table 2. Species-specific band patterns of water-soluble shrimp proteins. Mean pI
±RSD (%) were calculated from the patterns of the six shrimps of each species
analyzed. The RSD values varied between 0.0 and 2.3.
MPN SOP PBA PNI PNV LAA PRA SOK DPS KUP ABS TIP WKP CSP
6.13
6.05
5.21
5.10
5.04
4.95
4.86
4.75
4.61
4.54
5.65
5.60
5.52
5.00
4.95
4.87
4.80
4.74
4.72
4.60
4.48
5.83
5.17
5.07
5.00
4.93
4.88
4.84
4.74
4.57
4.49
5.79
5.10
5.04
4.96
4.91
4.86
4.76
4.73
4.64
4.59
4.52
6.10
6.04
5.60
5.56
5.52
5.34
5.03
4.93
4.87
4.82
4.74
4.72
4.57
4.51
4.53
4.49
4.89
4.84
4.74
4.62
4.60
4.58
4.60
4.57
4.50
4.86
4.64
4.60
4.58
4.51
5.00
4.91
4.86
4.81
4.74
4.72
4.70
4.59
4.56
4.48
4.42
5.68
5.63
5.57
4.97
4.88
4.80
4.66
4.60
4.55
6.28
6.22
5.65
5.14
5.09
5.07
4.99
4.94
4.89
4.82
4.74
4.59
4.51
5.88
5.19
5.08
5.02
4.94
4.86
4.77
4.61
4.53
4.22
6.11
5.57
5.50
5.23
4.93
4.88
4.85
4.79
4.75
4.62
4.53
4.44
104

Table 3. Sequences found that matched with Decapoda Sarcoplasmic Calcium-binding
Protein peptides.
sequence 100% matched with proteina Found in band no.
NLWNEIAELADFNK P02636, P02635, ABB58783, P05946,
1006232A
1, 2, 6
DGEVTVDEFK P02636, P02635, 1006232A 1, 2, 3, 4, 5, 6
VFIANQFK P02636, P05946, 1006232A 1, 2, 3, 4, 6
NDFECLAVR P02636, P02635, 1006232A 1, 2
VGLDEYR P02636, 1006232A 1, 2, 3, 6
EIDDAYNK P02636, 1006232A 1, 2
AGGLTLER P02636, 1006232A 2, 3
GEFSAADYANNQK P02636, 1006232A 3, 6
YMYDIDNDGFLDK P02635 1, 2, 4, 6
YM*YDIDNDGFLDK P02635 1, 2, 4, 5, 6
SAFADVK P02635 1, 2, 6
AGGINLAR P02635 5
VTLIEGR P02635 5
YM*YDIDNNGFLDK ABB58783, P05946 3, 4, 6
aP02636, SCP alpha chain; P02635, SCP beta chain; ABB58783, SCP (Procambarus
clarkii); P05946, SCP I; 1006232A, SCP αB.
105
FIGURES.
106

107

108

CAPÍTULO 2 IDENTIFICACIÓN DE ESPECIES DE LANGOSTINOS PRÓXIMAS POR MALDI-TOF MS


Capítulo 2: Identificación de especies de langostinos próximas por MALDI-
TOF MS.
Comentario
Una de las metodologías proteómicas más frecuentemente utilizada es la caracterización de
proteínas mediante huella peptídica, peptide mass fingerprinting (PMF) en la terminología inglesa. A
partir de la digestión con una proteasa de una proteína previamente aislada, se obtiene una mezcla de
péptidos que se analiza mediante MALDI-TOF. El espectro de masas obtenido, o mapa peptídico, es
específico para esa proteína.
En este trabajo se demostró la capacidad de esta metodología para diferenciar entre dos especies
de langostinos muy próximos, tanto filogenética y morfológicamente, como en distribución geográfica.
Las especies consideradas fueron dos de las de mayor importancia económica, Penaeus monodon y
Fenneropenaeus indicus, que algunos autores incluyen en el mismo género Penaeus, y que, como se ha
indicado anteriormente, comparten distribución geográfica y hábitat, además de tener caracteres
morfológicos muy similares.
Los resultados obtenidos demuestran que los espectros de masas de los péptidos provenientes de la
digestión tríptica de la proteína arginina quinasa nos permiten diferenciar de forma precisa entre las dos
especies consideradas.
111


PUBLICACIÓN
113


146
Journal of Aquatic Food Product Technology, 18:146–155, 2009Copyright © Taylor & Francis Group, LLC ISSN: 1049-8850 print/1547-0636 onlineDOI: 10.1080/10498850802584280
WAFP1049-88501547-0636Journal of Aquatic Food Product Technology, Vol. 18, No. 1-2, February 2009: pp. 1–18Journal of Aquatic Food Product Technology
Closely Related Shrimp Species Identification by MALDI-ToF Mass Spectrometry
Shrimp Species Identification by MALDI-ToF MSI. Ortea et al.
IGNACIO ORTEA, LORENA BARROS, and JOSÉ M. GALLARDODepartment of Marine Product Chemistry, Marine Research Institute, CSIC, Vigo, Spain
Tiger prawn, Penaeus monodon, and banana white prawn,
Fenneropenaeus indicus—two of the most important commercial
shrimp species—share world distribution, habitat, and similar
morphometric characters. Because of commercial regulations
about labeling and traceability requirements, species authentica-
tion methods are of great interest to fisheries and seafood indus-
tries. In this study, peptide mass fingerprints of both species are
compared, using Matrix Assisted Laser Desorption Ionization–
Time of Flight Mass Spectrometry (MALDI-TOF MS) after trypsin
digestion of a protein resolved by two-dimensional gel electro-
phoresis. Though low variability among these species is observed,
two peptides of mass 1077.6 and 1205.7 Da are found to be useful
to differentiate them.
KEYWORDS shrimp, species authentication, peptide mass finger-
print, MALDI-TOF MS
INTRODUCTION
Identification of marine species is a matter of capital importance in theseafood industry because of commercial requirements about labeling and
This work was supported by Instituto Nacional de Investigaciones Agrarias project No.CAL03-030-C2-2 “Aplicación de técnicas de genómica y proteómica a la identificación de
especies de crustáceos comerciales pertenecientes al Orden Decapoda” and by PGIDITResearch Program in Marine Resources Project No. PGIDIT04RMA261004PR of the Xunta deGalicia. I. O. is supported by the FPU program (AP-2004-5826) under the auspices of theSpanish Ministry of Education and Science.
Address correspondence to Ignacio Ortea, Department of Marine Product Chemistry,Marine Research Institute, Consejo Superior de Investigaciones Científicas, Eduardo Cabello6, 36208 Vigo, Spain. E-mail: [email protected]
115

Shrimp Species Identification by MALDI-ToF MS 147
traceability. Global regulations have been implemented to assure completeand correct information, guarantee market transparency, and avoid substitu-tion of certain fish species by others of less commercial value (MAPA, 2004).The handling of fillets or minced fish instead of whole specimens as rawmaterial is gaining importance in the manufacturing industry. This compli-cates the identification of fish species, which has been traditionally basedon the analysis of external anatomical and morphological features.
Tiger prawn, Penaeus monodon (Fabricius, 1798), and Indian whiteprawn, Fenneropenaeus indicus (H. Milne Edwards, 1837), are two of themost economically important crustacean species (Lavery et al., 2004). Bothof them are distributed in the Indo-West Pacific region, from East Africa toSoutheast Asia and Australia, and they share habitat and similar morphometriccharacters (Pérez Farfante and Kensley, 1997). This, added to the increasinguse of filleted or minced material mentioned above, makes species authenti-cation methods of great interest to fisheries and seafood industries.
The amplification of selected DNA sequences by polymerase chainreaction (PCR) has been applied to the identification of certain groups of fishspecies (Rasmussen and Morrissey, 2008). Among DNA targets, mitochondrialDNA (mtDNA), and more specifically the 16S rRNA gene and the cytochromeoxidase I (COI) gene, have been reported to serve as good interspecific mark-ers in some crustacean species, although most of these studies were focusedon population structures, phylogeography, and phylogenetic relationships(Lavery et al., 2004; Baldwin et al., 1998; Quan et al., 2004; Maggioni et al.,2001; Voloch et al., 2005), not on species identification. RFLP (RestrictionFragment Length Polymorphism) analyses of PCR products have been exten-sively used for species discrimination, but there are only a few studies dealingwith RFLP-based species identification and crustaceans. A method for thedetection of crustacean DNA based on a PCR-RFLP approach has been pro-posed, but it did not allow shrimp species identification, although it did per-mit their generic detection and differentiation with respect to crab, lobster,and crawfish species (Brzezinski, 2005). More recently, a PCR-RFLP protocolhas been proposed (Pascoal et al., 2008) that does allow the differentiation ofseveral penaeid shrimp species, including P. monodon and F. indicus,although RFLP patterns described for these two species are similar and maylead to ambiguous identification. RFLP method has the drawback that incom-plete digestion may occasionally occur, and intraspecific variation coulddelete or create additional restriction sites (Lockley and Bardsley, 2000) dueto the high degree of intraspecific variability of mtDNA. In addition, the needfor specific probe sequences and the associated DNA sequencing when thespecies under study are not present in databases makes genotypic methods forspecies identification technically demanding, time consuming, and expensive.
Protein-based methodologies (Mackie, 1997; Gallardo et al., 1995;Armstrong et al., 1992) have been used for authentication purposes in seafoodproducts. Recently, proteomic tools have been applied to the identification of
116

148 I. Ortea et al.
species (Martínez and Friis, 2004; Piñeiro et al., 1998), but little effort has beenmade to elucidate differences among closely related seafood species usingmass spectrometry (MS; Carrera et al., 2006; Piñeiro et al., 2001).
With the advent of proteomics, one of the important areas of interest isthe characterization of proteins through peptide mass fingerprinting (PMF).The Matrix Assisted Laser Desorption Ionization–Time of Flight MassSpectrometry (MALDI-TOF MS) is an effective method for the analysis ofproteins and peptides. This approach is highly advantageous due to its rela-tively low cost, simplicity in design, and sensitivity. In this work, we appliedMALDI-TOF MS to the identification of species-specific peptides within thesarcoplasmic protein fraction. Peptides detected allow a highly specificidentification of individuals belonging to the P. monodon taxonomic form,when compared to those belonging to F. indicus.
MATERIALS AND METHODS
Materials
Black tiger prawns (Penaeus monodon) and banana white prawns (Fenner-
openaeus indicus) were acquired by the Centro Tecnológico del Mar (Vigo,Spain). Shrimps were frozen onboard and shipped to our laboratory for theanalyses. Three specimens of P. monodon (samples mon1, mon2, andmon3), corresponding to three different populations (one from the coasts ofMalaysia and two from Mozambique); and three specimens of F. indicus
(samples indi1, indi2, and indi3), corresponding to another three popula-tions (all from the coasts of Mozambique), were evaluated in the study. Tailmuscle of these specimens was subjected to two-dimensional gel electro-phoresis (2-DE) and MS analysis.
Extraction of Sarcoplasmic Proteins
Extraction of the sarcoplasmic proteins was performed by homogenizing 1 gof raw white muscle from each individual in two volumes of milliQ water,in an Ultra-Turrax blender for 3 × 15 s—with interruptions of 45 s to avoidwarming the samples. The extracts were then centrifuged at 30,000 g for 15 minat 4ºC (J25 centrifuge; Beckman, Palo Alto, CA), and the supernatants weremaintained at −80ºC until the electrophoretic analysis. Protein concentrationin the extracts was determined by the bicinchoninic acid (BCA) method(Sigma Chemical Co., USA).
Two-Dimension Electrophoresis
First-dimension native IEF was performed at 10ºC in a Multiphor II electro-phoresis unit (Amersham Biosciences, Sweden) as described elsewhere
117

Shrimp Species Identification by MALDI-ToF MS 149
(Carrera et al., 2006). Briefly, protein extracts (4 mg/mL) were loaded onIEF precast polyacrylamide gels (Ampholine PAGplate pH 4.0–6.5, GEHealthcare, Uppsala, Sweden) and run under 1500 V–50 mA–30 W condi-tions, until 4000 V·h were reached. IEF strips corresponding to individuallanes were cut after the run was completed and kept at −80ºC until second-dimension electrophoresis analysis.
Equilibration of pH 4–6.5 IEF gel strips was carried out at room temper-ature as described previously (Piñeiro et al., 1998).
Preparative second dimension electrophoresis for MS analysis wereperformed using vertical sodium dodecyl sulphate (SDS) polyacrylamide gelelectrophoresis (PAGE; 7.5% T and 3% C) gels with Tris-tricine buffer andstained with Coomassie Brilliant Blue (CBB; GE Healthcare) according tothe manufacturer’s instructions.
In-Gel Digestion
One major spot, presented in all gels, was selected. This spot was excisedfrom each gel, and subjected to in-gel digestion with trypsin (RocheDiagnostics GmbH, Mannheim, Germany), performed overnight at 37ºC,according to Jensen et al. (1999). Final digestion solution was dried undervacuum and resuspended in 0.1% trifluoroacetic acid.
Spectra Acquisition and Processing
A 1 µL aliquot of the final sample solution was manually deposited onto astainless steel MALDI probe and allowed to dry at room temperature. Then,0.8 µL of matrix solution (saturated a-cyano-4 hydroxycinnamic acid in 50%aqueous acetonitrile and 0.1% trifluoroacetic acid) was added and againallowed to dry at room temperature.
All mass spectra were acquired with a Voyager DE STR MALDI-TOF MS(Applied Biosystems, Foster City, USA) operating in the reflector, delayextraction, positive-ion mode. The laser intensity was set just above the iongeneration threshold. The low mass gate was set to m/z 500 Da, the delaytime was 350 ns, the accelerating voltage was 20,000 V, and the grid voltagewas set to 68.5%. Mass spectra were acquired by accumulating 150 laser shotsin the m/z range from 850 to 3500. External close calibration with the Calibra-tion Mixture 2 of the Sequazyme Peptide Mass Standards Kit (Applied Biosys-tems) was used. Mass spectra were baseline corrected, and data listscontaining monoisotopic m/z values were extracted from mass spectral datawith the specific software of the instrument (Data Explorer, version 4.0.0.0).Signals with relative intensities greater than 4% were included in the lists.Species-specific biomarkers were extracted by visual comparison of the peaklists obtained. Mass lists were also used for protein and peptide identificationusing MASCOT Peptide Mass Fingerprint (Perkins et al., 1999). Search
118
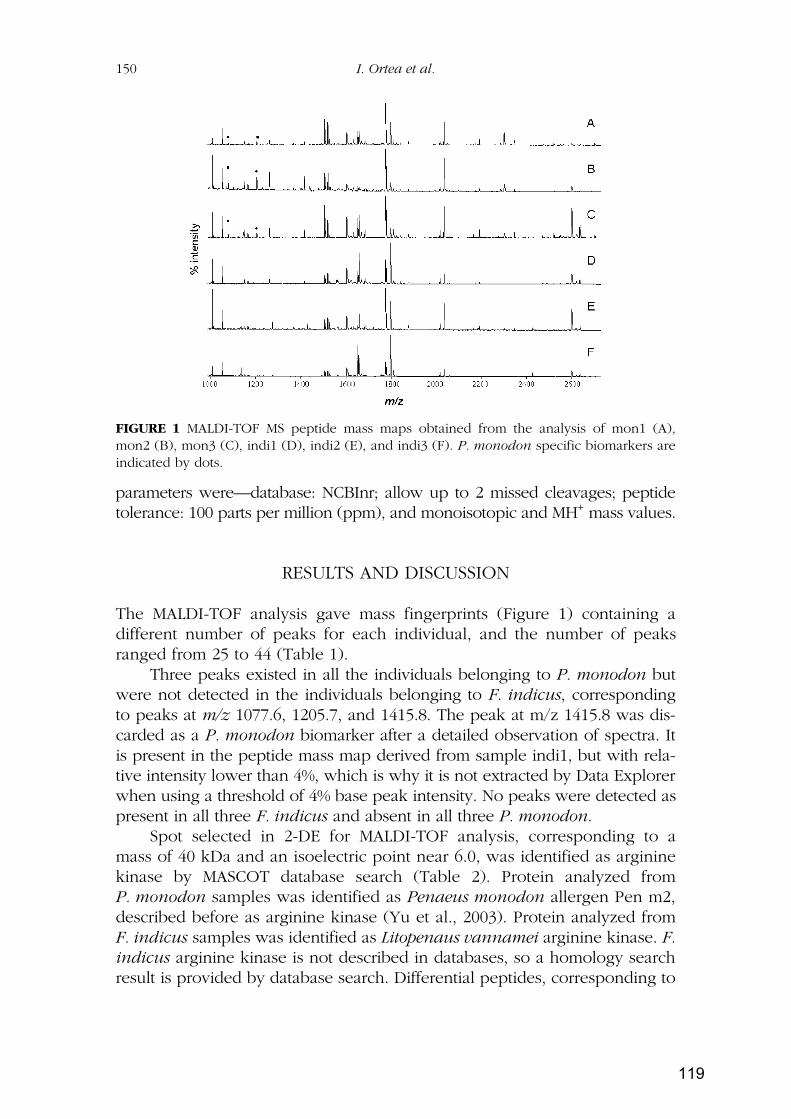
150 I. Ortea et al.
parameters were—database: NCBInr; allow up to 2 missed cleavages; peptidetolerance: 100 parts per million (ppm), and monoisotopic and MH+ mass values.
RESULTS AND DISCUSSION
The MALDI-TOF analysis gave mass fingerprints (Figure 1) containing adifferent number of peaks for each individual, and the number of peaksranged from 25 to 44 (Table 1).
Three peaks existed in all the individuals belonging to P. monodon butwere not detected in the individuals belonging to F. indicus, correspondingto peaks at m/z 1077.6, 1205.7, and 1415.8. The peak at m/z 1415.8 was dis-carded as a P. monodon biomarker after a detailed observation of spectra. Itis present in the peptide mass map derived from sample indi1, but with rela-tive intensity lower than 4%, which is why it is not extracted by Data Explorerwhen using a threshold of 4% base peak intensity. No peaks were detected aspresent in all three F. indicus and absent in all three P. monodon.
Spot selected in 2-DE for MALDI-TOF analysis, corresponding to amass of 40 kDa and an isoelectric point near 6.0, was identified as argininekinase by MASCOT database search (Table 2). Protein analyzed fromP. monodon samples was identified as Penaeus monodon allergen Pen m2,described before as arginine kinase (Yu et al., 2003). Protein analyzed fromF. indicus samples was identified as Litopenaus vannamei arginine kinase. F.
indicus arginine kinase is not described in databases, so a homology searchresult is provided by database search. Differential peptides, corresponding to
FIGURE 1 MALDI-TOF MS peptide mass maps obtained from the analysis of mon1 (A),mon2 (B), mon3 (C), indi1 (D), indi2 (E), and indi3 (F). P. monodon specific biomarkers areindicated by dots.
119
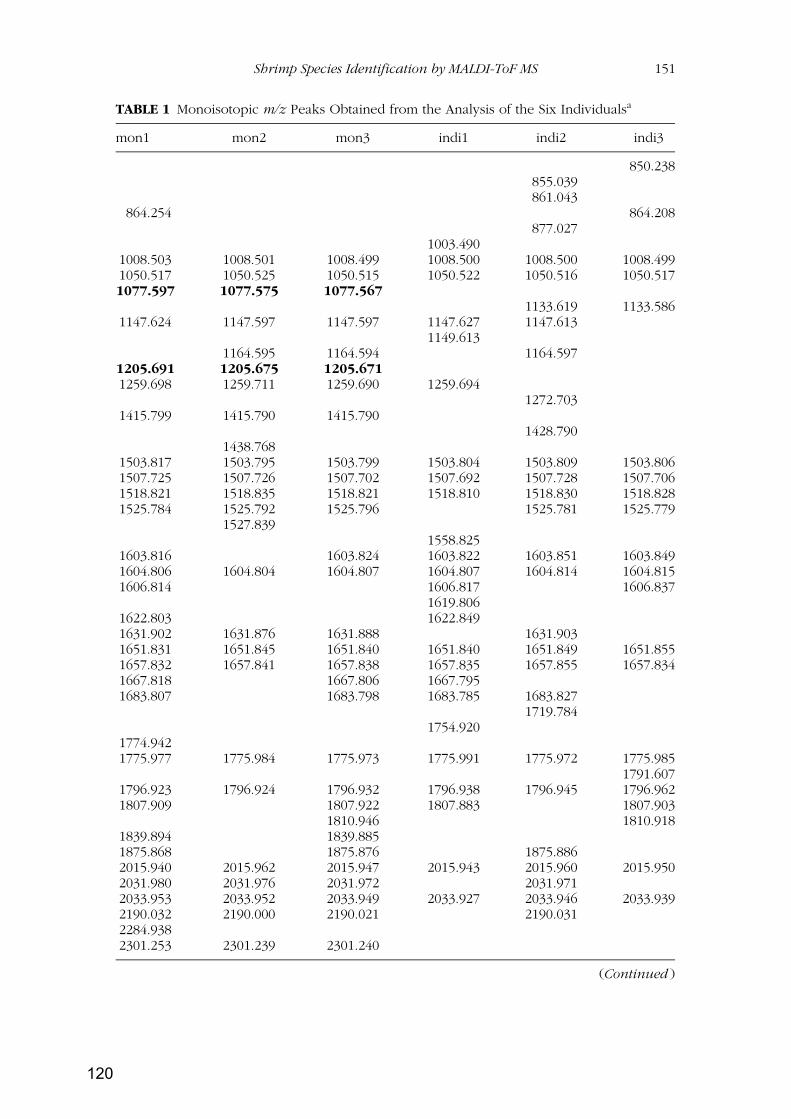
Shrimp Species Identification by MALDI-ToF MS 151
TABLE 1 Monoisotopic m/z Peaks Obtained from the Analysis of the Six Individualsa
mon1 mon2 mon3 indi1 indi2 indi3
850.238855.039861.043
864.254 864.208877.027
1003.4901008.503 1008.501 1008.499 1008.500 1008.500 1008.4991050.517 1050.525 1050.515 1050.522 1050.516 1050.5171077.597 1077.575 1077.567
1133.619 1133.5861147.624 1147.597 1147.597 1147.627 1147.613
1149.6131164.595 1164.594 1164.597
1205.691 1205.675 1205.6711259.698 1259.711 1259.690 1259.694
1272.7031415.799 1415.790 1415.790
1428.7901438.768
1503.817 1503.795 1503.799 1503.804 1503.809 1503.8061507.725 1507.726 1507.702 1507.692 1507.728 1507.7061518.821 1518.835 1518.821 1518.810 1518.830 1518.8281525.784 1525.792 1525.796 1525.781 1525.779
1527.8391558.825
1603.816 1603.824 1603.822 1603.851 1603.8491604.806 1604.804 1604.807 1604.807 1604.814 1604.8151606.814 1606.817 1606.837
1619.8061622.803 1622.8491631.902 1631.876 1631.888 1631.9031651.831 1651.845 1651.840 1651.840 1651.849 1651.8551657.832 1657.841 1657.838 1657.835 1657.855 1657.8341667.818 1667.806 1667.7951683.807 1683.798 1683.785 1683.827
1719.7841754.920
1774.9421775.977 1775.984 1775.973 1775.991 1775.972 1775.985
1791.6071796.923 1796.924 1796.932 1796.938 1796.945 1796.9621807.909 1807.922 1807.883 1807.903
1810.946 1810.9181839.894 1839.8851875.868 1875.876 1875.8862015.940 2015.962 2015.947 2015.943 2015.960 2015.9502031.980 2031.976 2031.972 2031.9712033.953 2033.952 2033.949 2033.927 2033.946 2033.9392190.032 2190.000 2190.021 2190.0312284.9382301.253 2301.239 2301.240
(Continued )
120

152 I. Ortea et al.
m/z 1077.6 and 1205.7, were identified as AVFDQLKEK and AVFDQLKEKK,respectively (Table 3).
CONCLUSIONS
Proteomics methodology, using peptide mass fingerprint generation and fastprotein identification by MALDI-TOF MS analysis, after 2-DE and trypsindigestion, was used to find differences within a sarcoplasmic protein fromtwo different species. The capacity of this methodology to discriminatebetween seafood species was ascertained by analyzing the closely relateddecapod species, P. monodon and F. indicus. Some authors include bothspecies in the same genus, Penaeus (Lavery et al., 2004), indicating that thetwo species selected for this study are very closely related to each other.
This approach is highly advantageous due to its relatively low cost,simplicity in design and sensitivity, which avoids technically demanding and
TABLE 1 (Continued )
mon1 mon2 mon3 indi1 indi2 indi3
2346.120 2346.1142426.150 2426.123
2472.1722520.265 2520.259
2548.2402580.1982597.146
2599.230 2599.260 2599.206 2599.250 2599.2432600.314
2619.000 2619.0492619.9892623.1152636.263 2636.295 2636.246 2636.2492701.219
3290.474 3290.5823403.593
3420.544 3420.587
aSpecific biomarkers are indicated by boldface type.
TABLE 2 Mascot Top Score Protein Matches for each Sample
Sample Protein Accession No. Mascot Score
mon1 allergen Pen m2 [Penaeus monodon] 27463265 196mon2 allergen Pen m2 [Penaeus monodon] 27463265 248mon3 allergen Pen m2 [Penaeus monodon] 27463265 274indi1 arginine kinase [Litopenaeus vannamei] 115492980 159indi2 arginine kinase [Litopenaeus vannamei] 115492980 226indi3 arginine kinase [Litopenaeus vannamei] 115492980 152
121
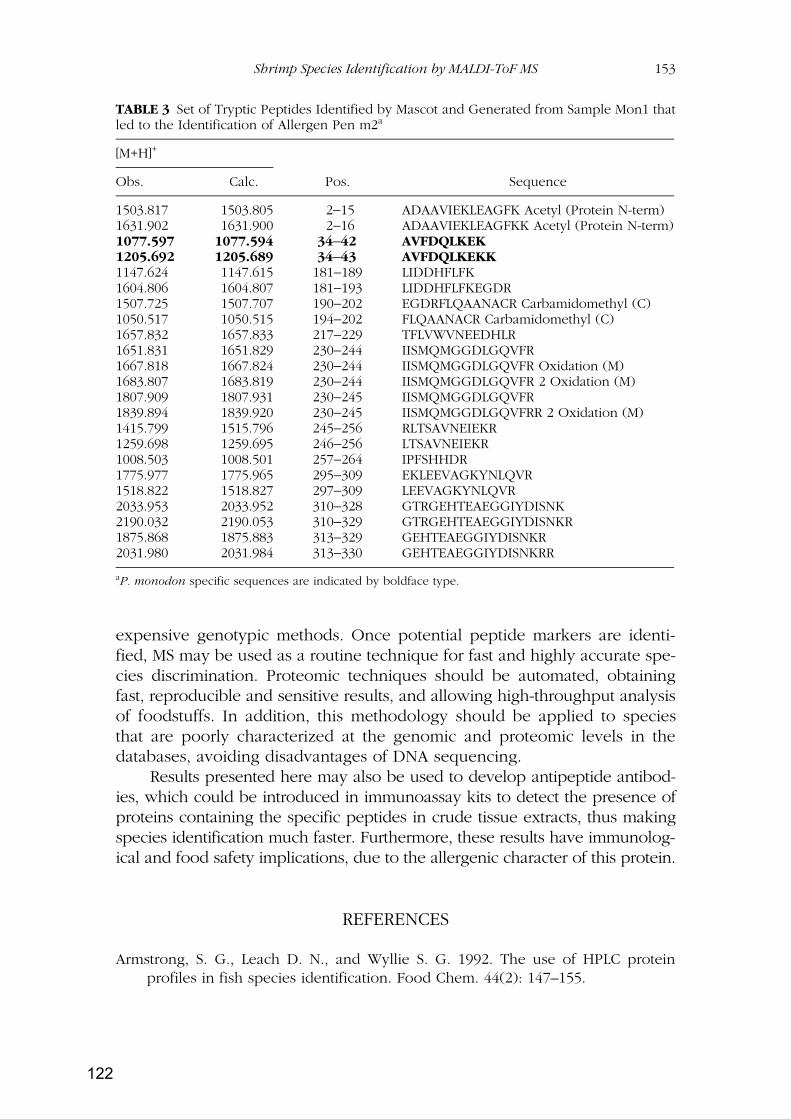
Shrimp Species Identification by MALDI-ToF MS 153
expensive genotypic methods. Once potential peptide markers are identi-fied, MS may be used as a routine technique for fast and highly accurate spe-cies discrimination. Proteomic techniques should be automated, obtainingfast, reproducible and sensitive results, and allowing high-throughput analysisof foodstuffs. In addition, this methodology should be applied to speciesthat are poorly characterized at the genomic and proteomic levels in thedatabases, avoiding disadvantages of DNA sequencing.
Results presented here may also be used to develop antipeptide antibod-ies, which could be introduced in immunoassay kits to detect the presence ofproteins containing the specific peptides in crude tissue extracts, thus makingspecies identification much faster. Furthermore, these results have immunolog-ical and food safety implications, due to the allergenic character of this protein.
REFERENCES
Armstrong, S. G., Leach D. N., and Wyllie S. G. 1992. The use of HPLC proteinprofiles in fish species identification. Food Chem. 44(2): 147–155.
TABLE 3 Set of Tryptic Peptides Identified by Mascot and Generated from Sample Mon1 thatled to the Identification of Allergen Pen m2a
[M+H]+
Pos. SequenceObs. Calc.
1503.817 1503.805 2−15 ADAAVIEKLEAGFK Acetyl (Protein N-term)1631.902 1631.900 2−16 ADAAVIEKLEAGFKK Acetyl (Protein N-term)1077.597 1077.594 34-42 AVFDQLKEK1205.692 1205.689 34-43 AVFDQLKEKK1147.624 1147.615 181−189 LIDDHFLFK1604.806 1604.807 181−193 LIDDHFLFKEGDR1507.725 1507.707 190−202 EGDRFLQAANACR Carbamidomethyl (C)1050.517 1050.515 194−202 FLQAANACR Carbamidomethyl (C)1657.832 1657.833 217−229 TFLVWVNEEDHLR1651.831 1651.829 230−244 IISMQMGGDLGQVFR1667.818 1667.824 230−244 IISMQMGGDLGQVFR Oxidation (M)1683.807 1683.819 230−244 IISMQMGGDLGQVFR 2 Oxidation (M)1807.909 1807.931 230−245 IISMQMGGDLGQVFR1839.894 1839.920 230−245 IISMQMGGDLGQVFRR 2 Oxidation (M)1415.799 1515.796 245−256 RLTSAVNEIEKR1259.698 1259.695 246−256 LTSAVNEIEKR1008.503 1008.501 257−264 IPFSHHDR1775.977 1775.965 295−309 EKLEEVAGKYNLQVR1518.822 1518.827 297−309 LEEVAGKYNLQVR2033.953 2033.952 310−328 GTRGEHTEAEGGIYDISNK2190.032 2190.053 310−329 GTRGEHTEAEGGIYDISNKR1875.868 1875.883 313−329 GEHTEAEGGIYDISNKR2031.980 2031.984 313−330 GEHTEAEGGIYDISNKRR
aP. monodon specific sequences are indicated by boldface type.
122

154 I. Ortea et al.
Baldwin, J. D., Bass, A. L., Bowen, B. W., and Clark, W. H. 1998. Molecular phylog-eny and biogeography of the marine shrimp Penaeus. Mol. Phylogenet. Evol.10(3): 399–407.
Brzezinski, J. L. 2005. Detection of crustacean DNA and species identification usinga PCR–restriction fragment length polymorphism method. J. Food Prot. 68(9):1866–1873.
Carrera, M., Cañas, B., Piñeiro, C., Vázquez, J., and Gallardo, J. M. 2006. Identifica-tion of commercial hake and grenadier species by proteomic analysis of theparvalbumin fraction. Proteomics 6(19): 5278–5287.
Gallardo, J. M., Sotelo, C. G., Piñeiro, C., and Pérez-Martín, R. I. 1995. Use of capil-lary zone electrophoresis for fish species identification. Differentiation of flatfishspecies. J. Agric. Food Chem. 43: 1238–1244.
Jensen, O. N., Wilm, M., Shevchenko, A., and Mann, M. 1999. Sample preparationmethods for mass spectrometric peptide mapping directly from 2-DE gels. In:2-D Proteome Analysis Protocols. Link, A. J. (Ed.). Totowa, NJ: Humana Press.pp. 513–530.
Lavery, S., Chan, T. Y., Tam, Y. K., and Chu, K. H. 2004. Phylogenetic relationshipsand evolutionary history of the shrimp genus Penaeus s.l. derived frommitochondrial DNA. Mol. Phylogenet. Evol. 31: 39–49.
Lockley, A. K., and Bardsley, R. G. 2000. DNA-based methods for food authentica-tion. Trends Food Sci. Tech. 11: 67–77.
Mackie, I. M. 1997. Methods of identifying species of raw and processed fish. In:Fish Processing Technology. Hall, G. M. (Ed.). London: Chapman & Hall.pp. 160–199.
Maggioni, R., Rogers, A. D., Maclean, N., and D’Incao, F. 2001. Molecular phylog-eny of Western Atlantic Farfantepenaeus and Litopenaeus shrimp based onmitochondrial 16S partial sequences. Mol. Phylogenet. Evol. 18: 66–73.
MAPA. 2004. Real Decreto 121/2004, de 23 de enero, sobre la identificación de losproductos de la pesca, de la acuicultura y del marisqueo vivos, frescos, refrig-erados o cocidos. (Royal Decree 121/2004, 23 January, About identification ofproducts from fisheries, aquaculture and shellfish gathering.) Ministerio deAgricultura, Pesca y Alimentación, Spain.
Martínez, I., and Friis, T. J. 2004. Application of proteome analysis to seafoodauthentication. Proteomics 4: 347–354.
Pascoal, A., Barros-Velázquez, J., Cepeda, A., Gallardo, J. M., and Calo-Mata, P.2008. A polymerase chain reaction-restriction fragment length polymorphismmethod based on the analysis of a 16S rRNA/tRNAVal mitochondrial region forspecies identification of commercial penaeid shrimps (Crustacea: Decapoda:
Penaeoidea) of food interest. Electrophoresis 29: 499–509.Perez Farfante, I., and Kensley, B. 1997. Penaeoid and Sergestoid Shrimps and
Prawns of the World: Keys and Diagnoses for the Families and Genera. Paris,France: Éditions du Muséum.
Perkins, D. N., Pappin, D. J. C., Creasy, D. M., and Cottrell, J. S. 1999. Probability-based protein identification by searching sequence databases using mass spec-trometry data. Electrophoresis 20: 3551–3567.
Piñeiro, C., Barros-Velazquez, J., Sotelo, C. G., Perez-Martin, R. I., and Gallardo, J.M.1998. Two-dimensional electrophoretic study of the water-soluble protein
123

Shrimp Species Identification by MALDI-ToF MS 155
fraction in white muscle of gadoid fish species. J. Agric. Food Chem. 46(10):3991–3997.
Piñeiro, C., Vázquez, J., Marina, A. I., Barros-Velázquez, J., and Gallardo J. M. 2001.Characterization and partial sequencing of species-specific sarcoplasmicpolypeptides from commercial hake species by mass spectrometry followingtwo-dimensional electrophoresis. Electrophoresis 22(8): 1545–1552.
Quan, J., Zhuang, Z., Deng, J., Dai, J., and Zang, Y. P. 2004. Phylogenetic relation-ships of 12 Penaeoidea shrimp species deduced from mitochondrial DNAsequences. Biochem. Genet. 42: 331–345.
Rasmussen, R. S., and Morrissey, M. T. 2008. DNA-based methods for the identifica-tion of commercial fish and seafood species. Compr. Rev. Food Sci. Food Saf.7: 280–295.
Voloch, C. M., Freire, P. R., and Russo, C. A. M. 2005. Molecular phylogeny ofpenaeid shrimps inferred from two mitochondrial markers. Genet. Mol. Res. 4:668–674.
Yu, C. J., Lin,Y. F., Chiang, B. L., and Chow, L. P. 2003. Proteomics and immunolog-ical analysis of a novel shrimp allergen, Pen m 2. J. Immunol. 170(1): 445–453.
124

CAPÍTULO 3 HUELLA PEPTÍDICA DE ARGININA QUINASA, ENFOQUE PROTEÓMICO PARA LA IDENTIFICACIÓN Y ANÁLISIS TAXONÓMICO DE ESPECIES DE LANGOSTINOS DE INTERÉS COMERCIAL


Capítulo 3: Huella peptídica de arginina quinasa. Enfoque proteómico para
la identificación y análisis taxonómico de especies de langostinos de interés
comercial.
Comentario
Una vez demostrado que la comparación de los espectros de huella peptídica de la arginina
quinasa nos permite diferenciar entre dos especies de langostinos muy próximas filogenéticamente, en
un segundo estudio se comprobó la idoneidad de la aproximación proteómica, combinando 2-DE y
PMF, para discriminar entre seis de las principales especies comerciales de langostinos y camarones.
Los resultados del análisis de 2-DE mostraron que el spot correspondiente a la proteína arginina
quinasa presentaba variaciones inter-específicas en su punto isoeléctrico, lo que concordaba con los
estudios previos en los que se había descrito variabilidad en la secuencia de esta proteína entre las
especies de Decápodos.
Para efectuar la comparación entre los espectros de huella peptídica de las distintas especies, se
desarrolló un método matemático que nos proporciona un valor numérico objetivo de la similitud entre
los espectros. Además, a partir de estos valores de similitud se elaboró un diagrama de datos en forma de
árbol o dendrograma.
Los resultados obtenidos demuestran la capacidad de la metodología de huella peptídica de la
arginina quinasa para diferenciar entre las seis especies consideradas y su utilidad en la identificación de
especies. Además, el dendrograma elaborado a partir de los valores de similitud entre los espectros PMF
resultó ser consistente con los árboles filogenéticos construidos con datos de secuencias de ADN de las
mismas especies, por lo que este método podría ser utilizado para inferir relaciones filogenéticas.
127


PUBLICACIÓN
129


pubs.acs.org/JAFCPublished on Web 06/03/2009© 2009 American Chemical Society
J. Agric. Food Chem. 2009, 57, 5665–5672 5665
DOI:10.1021/jf900520h
Arginine Kinase Peptide Mass Fingerprinting as a ProteomicApproach for Species Identification andTaxonomicAnalysis of
Commercially Relevant Shrimp Species
IGNACIO ORTEA,*,† BENITO CANAS,‡ PILAR CALO-MATA,§ JORGE BARROS-VELAZQUEZ,§
AND JOSE M. GALLARDO†
†Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Cientıficas (CSIC), Eduardo
Cabello 6, E-36208Vigo, Spain, ‡Analytical ChemistryDepartment, UniversidadComplutense deMadrid,
E-28040 Madrid, Spain, and §Analytical Chemistry, Nutrition and Food Science Department, School of
Veterinary Sciences, University of Santiago de Compostela, E-27002 Lugo, Spain
A proteomic approach aimed at species identification and taxonomic analysis of shrimp species of
commercial interest is presented. Six different species belonging to the order Decapoda were
considered. Preliminary, two-dimensional gel electrophoresis (2-DE) analysis of the sarcoplasmic
proteome revealed interspecific variability in the isoelectric point (pI) of arginine kinase. For this
reason, arginine kinase spot was selected as a potential molecular marker and subjected to tryptic
digestion followed by matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) peptide
mass fingerprinting (PMF) analysis. Arginine kinase PMF allowed the differentiation of the six
species studied. Four samples of commercial origin obtained in local markets were analyzed to
validate the methodology. The PMF cluster analysis also provided information about the phyloge-
netic relationships in these species. The application of this methodology may be of interest for the
differentiation and taxonomic analysis of shrimp species complementing DNA-based phylogenetic
studies.
KEYWORDS: Species identification; shrimp; MALDI-TOF MS; PMF; arginine kinase; proteomics
1. INTRODUCTION
Crustaceans belonging to the order Decapoda are of notablecommercial interest. This order includes the superfamily Penaeoi-dea,which is themost important economic resource in theworld’scrustacean fishery and aquafarming industry (1,2). Identificationof marine species is an issue of capital relevance for the seafoodindustry because of global commercial requirements concerninglabeling and traceability (3, 4), not only for the prevention ofcommercial fraud but also for the assessment of safety risksderived from the inadvertent introduction of any food ingredientthat might be harmful to human health (5). Anatomical char-acters are particularly difficult to use for shrimp species differ-entiation because of their phenotypic similarities and because ofthe fact that in their processing the external carapace is oftenremoved. Therefore, fast and reliable analytical tools are neces-sary for distinguishing between these closely related species.
Methods for species identification are currently based onDNAor protein analysis. Mitochondrial DNA (mtDNA) has beenextensively used in PCR-based studies for fish species identifica-tion (6). Among the mtDNA targets, the 16S rRNA gene andthe cytochrome oxidase I gene have been reported to serve asgood interspecific markers in some crustacean species, althoughmost of these studies were focused on population structures,
phylogeography, and phylogenetic relationships (7-11), and noton species identification. More recently, several PCR-RFLP-based methods for the detection of crustacean (12) and penaeidshrimps (13) DNAhave been proposed. Protein-basedmethodol-ogies have also been used for authentication purposes. Recently,proteomic tools have been applied to the identification of speciesin seafood products (14, 15), but little effort has been made toelucidate differences among closely related seafood species usingmass spectrometry (MS) (16-18), and only electrophoretic andimmunological assays have been reported to date for the detec-tion and differentiation of decapod crustaceans (19-21). Proteo-mics is concerned with identifying and determining the structure,expression, localization, interactions, and cellular roles of pro-teins. Though great advances have been made in this field in thepast decade, there are but only a few papers dealing specificallywith taxonomic studies and proteomics (22). Thus, with theemergence of molecular and bioinformatics analyses of wholegenomes, selection at the molecular level is measured almostexclusively by analysis of DNA sequence variation. However,proteins are responsible for the phenotype, and selection acts onthe structures that proteins build; therefore, analyses based onprotein sequences have capital relevance and complement DNA-based methods (23).
Matrix-assisted laser desorption/ionization-time-of-flight(MALDI-TOF) is one of the MS systems most extensivelyapplied to proteomic studies. This technology, though limited
*Corresponding author. Tel: 0034-986-231-930. Fax: 0034-986-292-762. E-mail: [email protected].
131

5666 J. Agric. Food Chem., Vol. 57, No. 13, 2009 Ortea et al.
due to the need for relatively pure samples, is highly advantageousdue to its relatively low cost, simplicity, and sensitivity. MALDI-TOF MS methodologies have been used in the protein profilefingerprint-based studies for species differentiation and phyloge-netic relationships of microorganisms belonging to differentgenera and species (24-26). Moreover, the term phyloproteomicswas introduced for the development and application of proteo-mics methodology to phylogenetic and evolutionary studies (24).The other main application of MALDI-TOF MS is peptidemass fingerprinting (PMF), for the identification of proteins bycomparing the obtained spectra with a database (27). One of theadvantages over protein profiling is that PMF, because ofprevious tryptic digestion, allows the identificationof both proteinand peptides, directly by database searching of the PMF spectra,and may be followed by peptide MS/MS fragmentation if resultsare conclusive enough. On the contrary, protein profiling does notidentify any of the constituent proteins but simply provides theirmasses. In addition, peptide mass values can be assigned withhigher precision because of the peak isotopic resolution possiblefor the low range (<5000 Da) of mass spectra in commonMALDI-TOF equipment. PMF has revealed itself to be a usefultechnique to differentiate among related marine species (16-18).
The objective of this work was to study the suitability of aproteomic approach, combining 2-DE and PMF, to discriminateamong closely related shrimp species of commercial relevance.Four different samples of commercial specimens were obtained inlocal markets to validate the methodology. On the basis of thecluster analysis of the MALDI-TOF PMF spectra obtained,dendrograms were generated, which were validated with thoseobtained using DNA-based methods.
2. MATERIALS AND METHODS
2.1. Shrimp Species and Populations Considered. Specimens
analyzed are shown inTable 1. Theywere collected using extractive fishing
practices or from aquaculture facilities in different continents worldwide.
We tested PMF’s ability to differentiate among closely related species by
considering six species from the order Decapoda, four of them from the
Penaeidae family, Penaeus monodon, Litopenaeus vannamei, Fennerope-
naeus indicus, and Farfantepenaeus notialis, one from the Solenoceridae
family, Pleoticus muelleri, and one from the Pandalidae family, Pandalus
borealis. Shrimp, whole animals, were frozen on board and shipped to our
laboratory for analyses. Special care was taken in keeping their morpho-
logical characteristics in good shape. Whole frozen shrimps from com-
mercial sources purchased in retail markets in Spain were also considered
(Table 1). At least three individuals of each species were analyzed.
Specimens were classified in their respective taxons according to their
anatomical external features with the help of a marine biologist from the
Marine Sciences Institute (Mediterranean Centre for Marine and Envir-
onmental Research, Higher Council for Scientific Research, CMIMA-
CSIC, Barcelona, Spain) with expertise in penaeid shrimp taxonomy.
2.2. Extraction of Sarcoplasmic Proteins. Extraction of the
sarcoplasmic proteins was performed by homogenizing 1 g of raw white
muscle from each individual in two volumes of Milli-Q water, using an
Ultra-Turrax blender for 3 15 s with interruptions of 45 s to avoid
warming the samples. The extracts were then centrifuged at 30000g for
15 min at 4 �C (J25 centrifuge; Beckman, Palo Alto, CA), and the
supernatants were maintained at -80 �C until electrophoretic analysis.
Protein concentration in the extracts was determined by the bicinchoninic
acid (BCA) method (Sigma Chemical Co., USA).
2.3. 2-DE. Three gels were run per individual, and at least three
individuals of each species were analyzed. First dimension native
isoelectric focusing (IEF) was performed at 10 �C in a Multiphor II
electrophoresis unit (Amersham Biosciences, Sweden) as described else-
where (17). Briefly, protein extracts (4 mg/mL, 100-120 μg of protein)
were loaded on IEF precast polyacrylamide gels (Ampholine PAGplate
pH 4.0-6.5; GE Healthcare, Uppsala, Sweden) in duplicate using sample
applicator paper and run under 1500 V, 50 mA, 30 W conditions, until
4000 V3h was reached. To avoid overlapping of proteins, strips with a
narrow pH range (pH 4.0-6.5) were chosen. IEF strips corresponding to
individual lanes were cut after the run was completed and kept at-80 �C
until second dimension electrophoresis analysis was performed.
Equilibration of pH 4-6.5 IEF gel strips was carried out at room
temperature as described previously (28). Briefly, the strips were placed for
10 min in sample buffer containing 0.75% DTT and then for another
10 min in sample buffer containing 4.5% iodoacetamide.
Preparative second dimension electrophoresis for MS analysis was
performed at 15 �C in the Multiphor II electrophoresis unit (Amersham
Biosciences, Sweden), using vertical SDS-PAGE (7.5%T and 3%C) gels
with Tris-tricine buffer. Running conditions were 100 V, 40 mA per gel,
and 150 W. Gels were stained with Coomassie brilliant blue (CBB)
(GE Healthcare), which is more compatible with sample preparation for
MS analysis, according to themanufacturer’s instructions. Three gels were
run per individual to ensure reproducibility andwere analyzed bymeans of
PDQuest 2-D Analysis Software, version 7.1.0 (Bio-Rad Laboratories,
Hercules, CA).
2.4. Mass Fingerprinting of Tryptic Peptides. 2.4.1. In-Gel
Digestion.Onemajor spot from the sarcoplasmic proteome, present in all
gels, was selected as a potential molecular marker because its pI exhibited
interspecific variability. This spot was excised from each gel and subjected
to in-gel digestion with trypsin (Roche Diagnostics GmbH, Mannheim,
Germany), performed overnight at 37 �C as described elsewhere (29).
The final digestion solution was dried under vacuum and resuspended in
0.1% TFA.
Table 1. Penaeid Shrimp Populations Considered in This Study
scientific namea commercial name originb codec accession number
Penaeus monodon giant tiger prawn IWP (Commercial) MPN-A EF589684
Penaeus monodon giant tiger prawn IWP (Malaysia) MPN-B/C EF589682/EF589685
MPN-D EF589683
Penaeus monodon giant tiger prawn WI (Mozambique) MPN-E/F FJ744153/FJ744154
Litopenaeus vannamei Pacific white shrimp EP (Costa Rica) PNV-A/B/C EF589702/EF589703/-
Fenneropenaeus indicus Indian white prawn WI (Mozambique) PNI-A/B EF589688/EF589689
PNI-C/D EF589690/EF589687
PNI-E EF589686
Farfantepenaeus notialis Southern pink shrimp EA (Senegal) SOP-1/2A EF589698/EF589697
SOP-2C EF589696
Pleoticus muelleri Argentine red shrimp SWA (Argentina) LAA-1/2A EF589718/EF589717
LAA-2B/D/E EF589716/-/-
Pleoticus muelleri Argentine red shrimp SWA (Commercial) LAA-F/G -/-
Pandalus borealis Northern shrimp NA (Greenland) PRA-A/B/C EU548069/EU548070/-
Pandalus borealis Northern shrimp NA (Commercial) PRA-D -
a The taxonomic classification proposed by Perez-Farfante and Kensley (2) was adopted. bOrigin abbreviations: IWP, Indo-West Pacific ocean; WI, Western Indian ocean; EP,Eastern Pacific ocean; WCP, Western Central Pacific ocean; EA, Eastern Atlantic ocean; SWA, Southern West Atlantic ocean; NA, Northern Atlantic ocean. c The three initialletters correspond to the FAO codes. Different letters and numbers after the FAO code indicate different individuals.
132
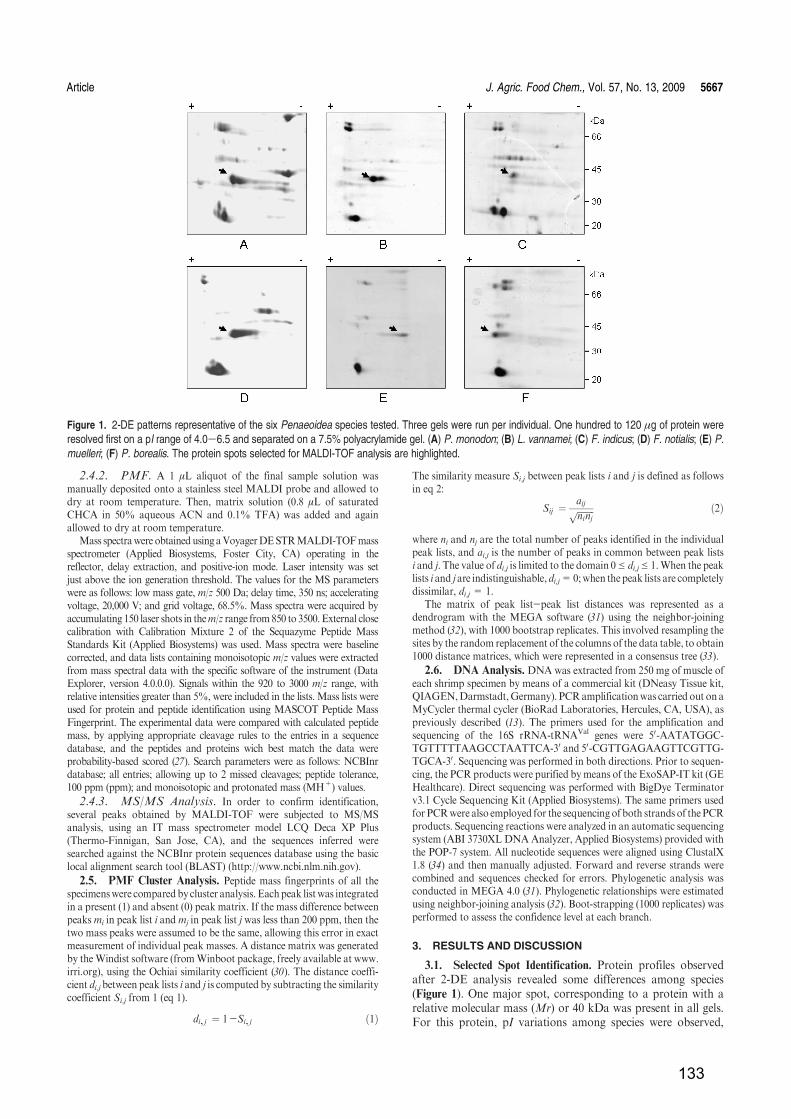
Article J. Agric. Food Chem., Vol. 57, No. 13, 2009 5667
2.4.2. PMF. A 1 μL aliquot of the final sample solution was
manually deposited onto a stainless steel MALDI probe and allowed to
dry at room temperature. Then, matrix solution (0.8 μL of saturated
CHCA in 50% aqueous ACN and 0.1% TFA) was added and again
allowed to dry at room temperature.
Mass spectrawere obtainedusing aVoyagerDESTRMALDI-TOFmass
spectrometer (Applied Biosystems, Foster City, CA) operating in the
reflector, delay extraction, and positive-ion mode. Laser intensity was set
just above the ion generation threshold. The values for the MS parameters
were as follows: low mass gate, m/z 500 Da; delay time, 350 ns; accelerating
voltage, 20,000 V; and grid voltage, 68.5%. Mass spectra were acquired by
accumulating 150 laser shots in them/z range from850 to 3500.External close
calibration with Calibration Mixture 2 of the Sequazyme Peptide Mass
Standards Kit (Applied Biosystems) was used. Mass spectra were baseline
corrected, and data lists containing monoisotopic m/z values were extracted
from mass spectral data with the specific software of the instrument (Data
Explorer, version 4.0.0.0). Signals within the 920 to 3000 m/z range, with
relative intensities greater than 5%, were included in the lists. Mass lists were
used for protein and peptide identification using MASCOT Peptide Mass
Fingerprint. The experimental data were compared with calculated peptide
mass, by applying appropriate cleavage rules to the entries in a sequence
database, and the peptides and proteins wich best match the data were
probability-based scored (27). Search parameters were as follows: NCBInr
database; all entries; allowing up to 2 missed cleavages; peptide tolerance,
100 ppm (ppm); and monoisotopic and protonated mass (MH+) values.
2.4.3. MS/MS Analysis. In order to confirm identification,
several peaks obtained by MALDI-TOF were subjected to MS/MS
analysis, using an IT mass spectrometer model LCQ Deca XP Plus
(Thermo-Finnigan, San Jose, CA), and the sequences inferred were
searched against the NCBInr protein sequences database using the basic
local alignment search tool (BLAST) (http://www.ncbi.nlm.nih.gov).
2.5. PMF Cluster Analysis. Peptide mass fingerprints of all the
specimenswere comparedby cluster analysis. Eachpeak list was integrated
in a present (1) and absent (0) peak matrix. If the mass difference between
peaks mi in peak list i and mj in peak list j was less than 200 ppm, then the
two mass peaks were assumed to be the same, allowing this error in exact
measurement of individual peak masses. A distance matrix was generated
by the Windist software (fromWinboot package, freely available at www.
irri.org), using the Ochiai similarity coefficient (30). The distance coeffi-
cient di,j between peak lists i and j is computed by subtracting the similarity
coefficient Si,j from 1 (eq 1).
di, j ¼ 1-Si, j ð1Þ
The similarity measure Si,j between peak lists i and j is defined as follows
in eq 2:
Sij ¼aijffiffiffiffiffiffiffiffi
ninjp ð2Þ
where ni and nj are the total number of peaks identified in the individual
peak lists, and ai,j is the number of peaks in common between peak lists
i and j. The value of di,j is limited to the domain 0e di,je 1.When the peak
lists i and j are indistinguishable, di,j=0;when the peak lists are completely
dissimilar, di,j = 1.
The matrix of peak list-peak list distances was represented as a
dendrogram with the MEGA software (31) using the neighbor-joining
method (32), with 1000 bootstrap replicates. This involved resampling the
sites by the random replacement of the columns of the data table, to obtain
1000 distance matrices, which were represented in a consensus tree (33).
2.6. DNA Analysis. DNA was extracted from 250 mg of muscle of
each shrimp specimen by means of a commercial kit (DNeasy Tissue kit,
QIAGEN,Darmstadt,Germany). PCRamplificationwas carried out on a
MyCycler thermal cycler (BioRad Laboratories, Hercules, CA, USA), as
previously described (13). The primers used for the amplification and
sequencing of the 16S rRNA-tRNAVal genes were 50-AATATGGC-
TGTTTTTAAGCCTAATTCA-30 and 50-CGTTGAGAAGTTCGTTG-
TGCA-30. Sequencing was performed in both directions. Prior to sequen-
cing, the PCR products were purified bymeans of the ExoSAP-IT kit (GE
Healthcare). Direct sequencing was performed with BigDye Terminator
v3.1 Cycle Sequencing Kit (Applied Biosystems). The same primers used
for PCRwere also employed for the sequencing of both strands of the PCR
products. Sequencing reactions were analyzed in an automatic sequencing
system (ABI 3730XL DNAAnalyzer, Applied Biosystems) provided with
the POP-7 system. All nucleotide sequences were aligned using ClustalX
1.8 (34) and then manually adjusted. Forward and reverse strands were
combined and sequences checked for errors. Phylogenetic analysis was
conducted in MEGA 4.0 (31). Phylogenetic relationships were estimated
using neighbor-joining analysis (32). Boot-strapping (1000 replicates) was
performed to assess the confidence level at each branch.
3. RESULTS AND DISCUSSION
3.1. Selected Spot Identification. Protein profiles observedafter 2-DE analysis revealed some differences among species(Figure 1). One major spot, corresponding to a protein with arelative molecular mass (Mr) or 40 kDa was present in all gels.For this protein, pI variations among species were observed,
Figure 1. 2-DE patterns representative of the six Penaeoidea species tested. Three gels were run per individual. One hundred to 120 μg of protein were
resolved first on a pI range of 4.0-6.5 and separated on a 7.5% polyacrylamide gel. (A) P. monodon; (B) L. vannamei; (C) F. indicus; (D) F. notialis; (E) P.
muelleri; (F) P. borealis. The protein spots selected for MALDI-TOF analysis are highlighted.
133
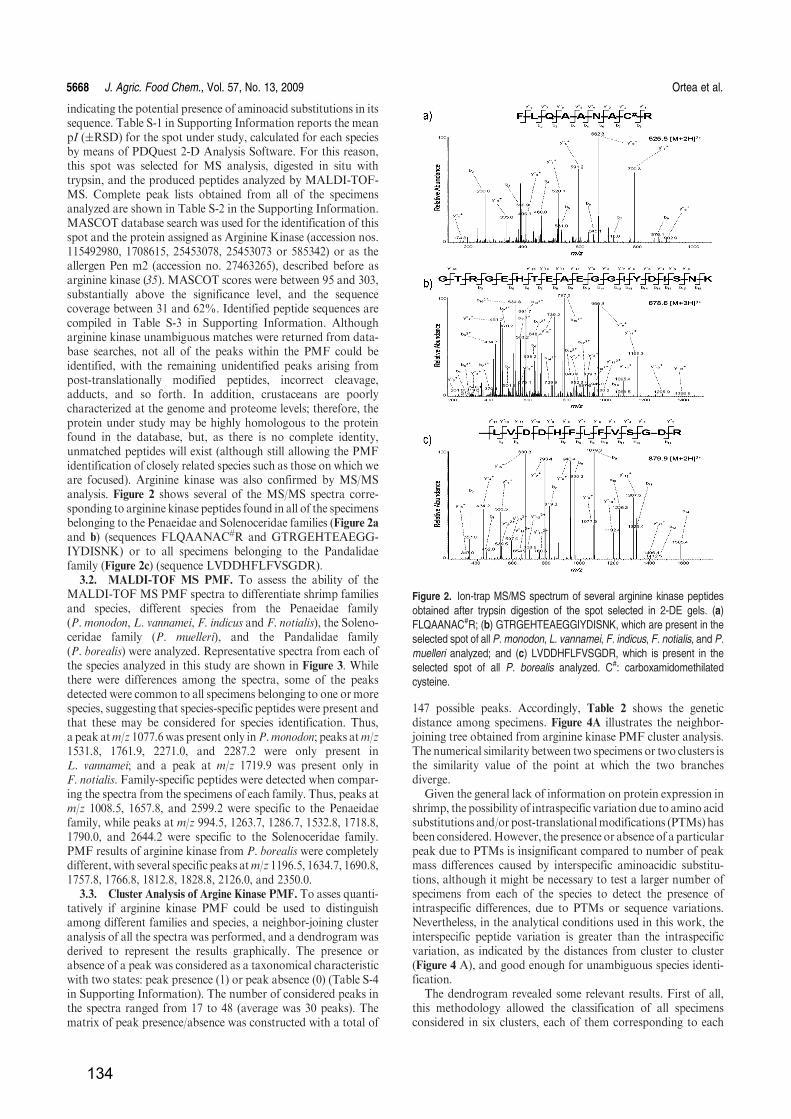
5668 J. Agric. Food Chem., Vol. 57, No. 13, 2009 Ortea et al.
indicating the potential presence of aminoacid substitutions in itssequence. Table S-1 in Supporting Information reports the meanpI ((RSD) for the spot under study, calculated for each speciesby means of PDQuest 2-D Analysis Software. For this reason,this spot was selected for MS analysis, digested in situ withtrypsin, and the produced peptides analyzed by MALDI-TOF-MS. Complete peak lists obtained from all of the specimensanalyzed are shown in Table S-2 in the Supporting Information.MASCOT database search was used for the identification of thisspot and the protein assigned as Arginine Kinase (accession nos.115492980, 1708615, 25453078, 25453073 or 585342) or as theallergen Pen m2 (accession no. 27463265), described before asarginine kinase (35). MASCOT scores were between 95 and 303,substantially above the significance level, and the sequencecoverage between 31 and 62%. Identified peptide sequences arecompiled in Table S-3 in Supporting Information. Althougharginine kinase unambiguous matches were returned from data-base searches, not all of the peaks within the PMF could beidentified, with the remaining unidentified peaks arising frompost-translationally modified peptides, incorrect cleavage,adducts, and so forth. In addition, crustaceans are poorlycharacterized at the genome and proteome levels; therefore, theprotein under study may be highly homologous to the proteinfound in the database, but, as there is no complete identity,unmatched peptides will exist (although still allowing the PMFidentification of closely related species such as those on which weare focused). Arginine kinase was also confirmed by MS/MSanalysis. Figure 2 shows several of the MS/MS spectra corre-sponding to arginine kinase peptides found in all of the specimensbelonging to the Penaeidae and Solenoceridae families (Figure 2aand b) (sequences FLQAANAC#R and GTRGEHTEAEGG-IYDISNK) or to all specimens belonging to the Pandalidaefamily (Figure 2c) (sequence LVDDHFLFVSGDR).
3.2. MALDI-TOF MS PMF. To assess the ability of theMALDI-TOF MS PMF spectra to differentiate shrimp familiesand species, different species from the Penaeidae family(P. monodon, L. vannamei, F. indicus and F. notialis), the Soleno-ceridae family (P. muelleri), and the Pandalidae family(P. borealis) were analyzed. Representative spectra from each ofthe species analyzed in this study are shown in Figure 3. Whilethere were differences among the spectra, some of the peaksdetected were common to all specimens belonging to one ormorespecies, suggesting that species-specific peptides were present andthat these may be considered for species identification. Thus,a peak atm/z 1077.6was present only inP.monodon; peaks atm/z1531.8, 1761.9, 2271.0, and 2287.2 were only present inL. vannamei; and a peak at m/z 1719.9 was present only inF. notialis. Family-specific peptides were detected when compar-ing the spectra from the specimens of each family. Thus, peaks atm/z 1008.5, 1657.8, and 2599.2 were specific to the Penaeidaefamily, while peaks at m/z 994.5, 1263.7, 1286.7, 1532.8, 1718.8,1790.0, and 2644.2 were specific to the Solenoceridae family.PMF results of arginine kinase from P. borealis were completelydifferent, with several specific peaks atm/z 1196.5, 1634.7, 1690.8,1757.8, 1766.8, 1812.8, 1828.8, 2126.0, and 2350.0.
3.3. Cluster Analysis of Argine Kinase PMF. To asses quanti-tatively if arginine kinase PMF could be used to distinguishamong different families and species, a neighbor-joining clusteranalysis of all the spectra was performed, and a dendrogram wasderived to represent the results graphically. The presence orabsence of a peak was considered as a taxonomical characteristicwith two states: peak presence (1) or peak absence (0) (Table S-4in Supporting Information). The number of considered peaks inthe spectra ranged from 17 to 48 (average was 30 peaks). Thematrix of peak presence/absence was constructed with a total of
147 possible peaks. Accordingly, Table 2 shows the geneticdistance among specimens. Figure 4A illustrates the neighbor-joining tree obtained from arginine kinase PMF cluster analysis.The numerical similarity between two specimens or two clusters isthe similarity value of the point at which the two branchesdiverge.
Given the general lack of information on protein expression inshrimp, the possibility of intraspecific variation due to amino acidsubstitutions and/or post-translationalmodifications (PTMs) hasbeen considered.However, the presence or absence of a particularpeak due to PTMs is insignificant compared to number of peakmass differences caused by interspecific aminoacidic substitu-tions, although it might be necessary to test a larger number ofspecimens from each of the species to detect the presence ofintraspecific differences, due to PTMs or sequence variations.Nevertheless, in the analytical conditions used in this work, theinterspecific peptide variation is greater than the intraspecificvariation, as indicated by the distances from cluster to cluster(Figure 4 A), and good enough for unambiguous species identi-fication.
The dendrogram revealed some relevant results. First of all,this methodology allowed the classification of all specimensconsidered in six clusters, each of them corresponding to each
Figure 2. Ion-trap MS/MS spectrum of several arginine kinase peptides
obtained after trypsin digestion of the spot selected in 2-DE gels. (a)
FLQAANAC#R; (b) GTRGEHTEAEGGIYDISNK, which are present in the
selected spot of all P. monodon, L. vannamei, F. indicus, F. notialis, and P.
muelleri analyzed; and (c) LVDDHFLFVSGDR, which is present in the
selected spot of all P. borealis analyzed. C#: carboxamidomethilated
cysteine.
134

Article J. Agric. Food Chem., Vol. 57, No. 13, 2009 5669
species. The first cluster corresponded to P. monodon specimens,with similarity values of ∼0.85 but a bootstrap value of 52(Figure 4 A). All three F. notialis specimens were tightlyclustered, exhibiting similarity values of ∼0.9 and a bootstrapsupport of 82. Another cluster was formed by L. vannamei
specimens, with a similarity value over ∼0.85 and bootstrapvalues over 90. All five F. indicus specimens were clustered withsimilarity values over 0.8, although the bootstrap value wasbelow 50. The fifth cluster was formed by P. muelleri speci-mens, with similarity values over 0.8 and the highest bootstrapvalue of 100. Finally, the sixth cluster included P. borealis
specimens with high similarity and a bootstrap value of 100.The first five clusters could be strongly grouped (indicated bythe highest bootstrap support, 100) into two bigger clusterswith similarity values of ∼0.7, one corresponding to thePenaeidae family (bootstrap value of 91) and the other corre-sponding to the Solenoceridae family (bootstrap value of 100)(Figure 4A). These clustering results showed the reproducibil-ity of PMF spectra formeasurements of different individuals ofa particular species. Commercial samples were also effectivelyidentified after measuring their level of similarity against therest of the specimens.
Topology resulting from the PMF cluster analysis (Figure 4A)was highly concordant with a dendrogram derived from an ana-lysis of 16S rRNA-tRNAValmithocondrial genes (Figure 4B) andwith previously described taxonomical levels in penaeid shrimpdetermined by independent validated methods (8-11, 13). Re-markably, a detailed comparisonwith such previous phylogenetictrees, constructed on the basis of mtDNA data, revealed onlyslight differences. This result highlights the accuracy of theinformation provided by the arginine kinase PMF methoddescribed in this work, as compared to the genetic studiesbased on either 16S rRNA or cytochrome oxidase I mitochon-drial genes.
The efficiency of the arginine kinase PMF methodology todifferentiate between the three families and the six shrimp
species and its usefulness in the identification of four commer-
cial samples has been demonstrated. At first sight, the proteo-
mic approach described in this work seems to be as time-
consuming and complicated as the usual genetic analyses, such
as RFLP, which usually takes two days. However, once we
know the pI and the Mw of the protein of interest and thus its
position in the 2-DE gels, we can obtain the PMF spectra,
avoiding the more complicated steps of this proteomic
approach such as identification and sequencing of the spot.
Proteomic techniques may be automated, producing fast,
reproducible, and sensitive results and allowing high-through-
put analysis of foodstuffs. In addition, this methodology can
be applied to species that are poorly characterized at the
genomic and proteomic levels in the databases, avoiding the
disadvantages of DNA sequencing. Sarcoplasmic proteins are
usually more heat-stable than other myofibrillar proteins;
thus, the selection of this fraction of proteins seems to be an
asset to the applicability of this proteomic approach on
products treated at high temperatures. However, heated sam-
ples were not included in this study. For this purpose, further
studies have to be made.A great advantage of the proteomic approach is that it is the
first step toward designing easy and cheap detection analyses:
the identification of specific peptides, as done by this work, can
be used to identify not only taxons but also proteins of interest.
These diagnostic peptides can be used in fast, cheap, and easy-
to-use kits based on antibodies against the specific peptides.
The selection of arginine kinase is particularly interesting
because in addition to discriminating species, it is also an
allergen (35), and therefore, an analysis targeting this protein
would have a double application: in species identification, if
the peptide selected discriminates at the species level, and in
food safety, if the peptide selected discriminates for common
peptides for several relevant species. This may permit the
detection of the allergen in certain products and help the
Figure 3. Arginine kinase PMF for representative individuals from 6 different shrimp species. (A) P. monodon; (B) L. vannamei; (C) F. indicus; (D) F. notialis;
(E) P. muelleri; (F) P. borealis. Family specific peptides (•) and species-specific peptides (*) are highlighted.
135

5670 J. Agric. Food Chem., Vol. 57, No. 13, 2009 Ortea et al.
manufacturer to identify, in real-time, the existence of acontamination in a production line.
The use of a mathematical approach for the comparison ofPMF spectra, instead of performing the visual spectral evaluationused in other species differentiation studies (16, 17), eliminatessubjectivity bias providing a quantitative measure of how welltwo spectra match. The phylogenetic relationships derived fromthe PMF cluster analysis also provided information for thegrouping of species belonging to the order Decapoda. To ourknowledge, the results reported here represent the first applica-tion of a MALDI-TOF MS PMF method to infer taxonomicrelationships in any type of organism and are consistent with thephylogenies established with mtDNA data. This study opens theway tomore in depth studies with a wider range of shrimp speciesand provides the basis for further investigations on phyloproteo-mics. Since proteins constitute the biological machinery, andselection acts on the structures formed by them, analyses based on
protein sequences should be considered as a complementary toolto mtDNA-based methods.
ABBREVIATIONS USED
ACN, acetonitrile; BCA, bicinchoninic acid; BLAST, basiclocal alignment search tool; CBB, Coomassie brilliant blue;CHCA, R-cyano-4-hydroxycinnamic acid; Da, Dalton (molecu-lar mass); 2-DE, two-dimensional gel electrophoresis; DTT,dithiothreitol; IEF, isoelectric focusing; MALDI-TOF, matrix-assisted laser desorption/ionization-time-of-flight; Mr, relativemolecular mass; MS, mass spectrometry; MS/MS, tandem massspectrometry; m/z, mass-to-charge ratio; PCR, polymerasechain reaction; pI, isoelectric point; PMF, peptide mass finger-printing; ppm, parts per million; PTM, post-translational mod-ifications; RFLP, restriction fragment length polymorphism;RSD, relative standard deviation; SDS-PAGE, sodium dodecyl
Table 2. Distance Matrix between the Arginine Kinase PMF Peak Lists of All Different Specimensa, Calculated by Means of the Ochiai Similarity Coefficient
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1
2 0.277
3 0.158 0.221
4 0.444 0.176 0.332
5 0.958 0.956 0.958 0.956
6 0.957 0.955 0.957 0.955 0.186
7 0.955 0.952 0.955 0.952 0.212 0.235
8 0.960 0.958 0.960 0.958 0.269 0.224 0.249
9 0.963 0.961 0.963 0.961 0.289 0.217 0.268 0.153
10 0.961 0.958 0.961 0.958 0.280 0.268 0.294 0.373 0.332
11 1.000 1.000 1.000 1.000 0.487 0.449 0.449 0.456 0.367 0.545
12 1.000 1.000 1.000 1.000 0.484 0.475 0.376 0.477 0.455 0.545 0.310
13 1.000 1.000 1.000 1.000 0.499 0.437 0.434 0.472 0.368 0.480 0.188 0.337
14 0.956 0.953 0.956 0.953 0.543 0.500 0.509 0.363 0.407 0.505 0.579 0.626 0.583
15 0.947 0.944 0.947 0.944 0.497 0.446 0.370 0.401 0.405 0.489 0.606 0.473 0.570 0.294
16 0.947 0.944 0.947 0.944 0.372 0.318 0.235 0.281 0.330 0.449 0.498 0.432 0.470 0.382
17 0.953 0.950 0.953 0.950 0.441 0.356 0.359 0.360 0.338 0.510 0.490 0.350 0.440 0.450
18 0.955 0.952 0.955 0.952 0.427 0.308 0.308 0.317 0.268 0.495 0.449 0.445 0.406 0.434
19 1.000 1.000 1.000 1.000 0.452 0.350 0.412 0.362 0.243 0.428 0.401 0.470 0.399 0.487
20 1.000 1.000 1.000 1.000 0.430 0.359 0.387 0.342 0.200 0.464 0.332 0.419 0.312 0.494
21 1.000 1.000 1.000 1.000 0.408 0.337 0.364 0.322 0.263 0.444 0.417 0.426 0.415 0.438
22 1.000 1.000 1.000 1.000 0.704 0.639 0.682 0.548 0.500 0.666 0.569 0.685 0.579 0.500
23 1.000 1.000 1.000 1.000 0.567 0.559 0.606 0.587 0.585 0.624 0.658 0.677 0.631 0.649
24 1.000 1.000 1.000 1.000 0.673 0.585 0.620 0.533 0.492 0.591 0.581 0.657 0.570 0.512
25 1.000 1.000 1.000 1.000 0.629 0.536 0.571 0.511 0.468 0.625 0.537 0.641 0.549 0.579
26 1.000 1.000 1.000 1.000 0.627 0.586 0.636 0.580 0.608 0.713 0.652 0.639 0.625 0.714
27 1.000 1.000 1.000 1.000 0.755 0.626 0.693 0.689 0.637 0.732 0.686 0.684 0.677 0.613
28 1.000 1.000 1.000 1.000 0.707 0.574 0.640 0.641 0.628 0.725 0.678 0.635 0.669 0.691
15 16 17 18 19 20 21 22 23 24 25 26 27 28
15
16 0.211
17 0.298 0.204
18 0.325 0.190 0.119
19 0.465 0.426 0.422 0.346
20 0.472 0.397 0.362 0.291 0.151
21 0.405 0.367 0.371 0.332 0.243 0.227
22 0.591 0.591 0.603 0.586 0.513 0.467 0.526
23 0.623 0.581 0.590 0.535 0.574 0.580 0.556 0.437
24 0.555 0.624 0.574 0.591 0.503 0.510 0.444 0.250 0.347
25 0.606 0.534 0.554 0.479 0.479 0.435 0.443 0.291 0.259 0.185
26 0.702 0.574 0.545 0.490 0.629 0.542 0.608 0.488 0.186 0.446 0.304
27 0.590 0.641 0.544 0.518 0.627 0.596 0.637 0.456 0.428 0.400 0.371 0.377
28 0.632 0.632 0.579 0.550 0.618 0.585 0.628 0.479 0.372 0.384 0.319 0.318 0.179
aCodes: 1, PRA A; 2, PRA C; 3, PRA B; 4, PRA D; 5, MPN B; 6, MPN D; 7, MPN C; 8, MPN E; 9, MPN F; 10, MPN A; 11, PNV A; 12, PNV B; 13, PNV C; 14,PNI E; 15, PNI D; 16, PNI C; 17, PNI B; 18, PNI A; 19, SOP 1, 20, SOP 2A; 21, SOP 2C; 22, LAA 2B; 23, LAA 2A; 24, LAA 1; 25, LAA D; 26, LAA E; 27, LAA F; 28,LAA G.
136

Article J. Agric. Food Chem., Vol. 57, No. 13, 2009 5671
sulfate-polyacrylamide gel electrophoresis; TFA, trifluoroaceticacid; Tris, tris(hydroxymethyl)aminomethane.
ACKNOWLEDGMENT
We thank Lorena Barros for her excellent technical assistanceandDr. VanesaDıaz for her helpful assistance.We also acknowl-edge members from CETMAR for their helpful collaboration inthe collection of specimens for this study. The manuscriptbenefited from critical comments by three anonymous reviewers.
Supporting Information Available: pI of the analyzed spot,
calculated from the 2-DE patterns of all the individuals of each
species studied; complete peaks lists obtained fromarginine kinase
PMFofall specimensanalyzed in this study; complete information
of the MALDI-TOF analysis showing Decapoda arginine kinase
matching peptide sequences; and matrix of peaks presence (1=
present; 0=absent) of all different specimens. This material is
available free of charge via the Internet at http://pubs.acs.org.
LITERATURE CITED
(1) Holthuis, L. B. FAO Species Catalogue: Shrimps and Prawns of the
World. An Annotated Catalogue of Species of Interest to Fisheries;
FAO Fish. Synop. (125); FAO: Rome, Italy, 1980; Vol. 1, pp 1-271.
(2) Perez-Farfante, I., Kensley, B. Penaeoid and Sergestoid Shrimps
and Prawns of the World. Keys and Diagnoses for the Families and
Genera; �Editions du Museum National d’Histoire Naturelle: Paris,
France, 1997.
(3) Council Regulation (EC) No. 104/2000 of the European Parliament
of Dec. 17, 1999 on the common organization of the markets in
fishery and aquaculture products.
(4) Royal Decree 121/2004 of Jan. Twenty-three (BOE No. 31, Feb. 5,
2004) from Ministry of Agriculture, Fisheries and Food, Spain.
(5) Civera, T. Species identification and safety of fish products.Vet. Res.
Commun. 2003, 27, 481–489.
(6) Rasmussen, R. S.; Morrissey, M. T. DNA-based methods for the
identification of commercial fish and seafood species. Compr. Rev.
Food Sci. Food Saf. 2008, 7, 280–295.
(7) Maggioni, R.; Rogers, A. D.; Maclean, N.; Incao, F. Molecular
phylogeny of Western Atlantic Farfantepenaeus and Litopenaeus
shrimp based on mitochondrial 16S partial sequences. Mol. Phylo-
genet. Evol. 2001, 18, 66–73.
(8) Baldwin, J. D.; Bass, A. L.; Bowen, B. W.; Clark, W. H. Molecular
phylogeny and biogeography of the marine shrimp Penaeus. Mol.
Phylogenet. Evol. 1998, 10, 399–407.
(9) Quan, J.; Zhuang, Z.; Deng, J.; Dai, J.; Zhang, Y.-p. Phylogenetic
relationships of 12 Penaeoidea shrimp species deduced from mito-
chondrial DNA sequences. Biochem. Genet. 2004, 42, 331–345.
(10) Lavery, S.; Chan, T. Y.; Tam, Y. K.; Chu, K. H. Phylogenetic
relationships and evolutionary history of the shrimp genusPenaeus s.
l. derived frommitochondrial DNA.Mol. Phylogenet. Evol. 2004, 31,
39–49.
(11) Voloch, C. M.; Freire, P. R.; Russo, C. A. M. Molecular phylogeny
of penaeid shrimps inferred from twomitochondrial markers. Genet.
Mol. Res. 2005, 4, 668–674.
(12) Brzezinski, J. L. Detection of crustacean DNA and species identifi-
cation using a PCR-restriction fragment length polymorphism
method. J. Food Prot. 2005, 68, 1866–1873.
(13) Pascoal, A.; Barros-Velazquez, J.; Cepeda, A.; Gallardo, J. M.;
Calo-Mata, P. A polymerase chain reaction-restriction fragment
length polymorphism method based on the analysis of a 16S rRNA/
tRNAVal mitochondrial region for species identification of commer-
cial penaeid shrimps (Crustacea: Decapoda: Penaeoidea) of food
interest. Electrophoresis 2008, 29, 499–509.
(14) Pineiro, C.; Barros-Velazquez, J.; Vazquez, J.; Figueras, A.; Gallardo,
J. M. Proteomics as a tool for the investigation of seafood and other
marine products. J. Proteome Res. 2003, 2, 127–135.
(15) Martınez, I.; Friis, T. J. Application of proteome analysis to seafood
authentication. Proteomics 2004, 4, 347–354.
(16) L�opez, J. L.; Marina, A.; �Alvarez, G.; Vazquez, J. Application of
proteomics for fast identification of species-specific peptides from
marine species. Proteomics 2002, 2, 1658–1665.
Figure 4. Topologies resulting from (A) arginine kinase PMF cluster analysis; (B) phylogenetic analysis of the nucleotide sequences of 16S rRNA-tRNAVal
mithocondrial genes. Both were obtained by means of the neighbor-joining method; bootstrap values >50 are shown. MPN, P. monodon; PNV, L. vannamei;
SOP, F. notialis; PNI, F. indicus; LAA, P. muelleri; PRA, P. borealis; as referred to in Table 1.
137

5672 J. Agric. Food Chem., Vol. 57, No. 13, 2009 Ortea et al.
(17) Carrera, M.; Canas, B.; Pineiro, C.; Vazquez, J.; Gallardo, J. M.
Identification of commercial hake and grenadier species by proteomic
analysis of the parvalbumin fraction. Proteomics 2006, 6, 5278–5287.
(18) Carrera, M.; Canas, B.; Pineiro, C.; Vazquez, J.; Gallardo, J. M. De
novo mass spectrometry sequencing and characterization of species-
specific peptides from nucleoside diphosphate kinase B for the
classification of commercial fish species belonging to the family
Merlucciidae. J. Proteome Res. 2007, 6, 3070–3080.
(19) An, H.; Wei, C. I.; Zhao, J.; Marshall, M. R.; Lee, C. M. Electro-
phoretic identification of fish species used in surimi products. J. Food
Sci. 1989, 54, 253–257.
(20) Civera, T.; Parisi, E. The use of sodium dodecyl sulphate polyacry-
lamide gel electrophoresis for the identification of raw and cooked
shrimps and crabs. Ital. J. Food Sci. 1991, 3, 149–157.
(21) Shanti, K. N.; Martin, B. M.; Nagpal, S.; Metcalfe, D. D.;
Subba Rao, P. V. Identification of tropomyosin as the major shrimp
allergen and characterization of its IgE-binding epitopes. J. Immu-
nol. 1993, 151, 5354–5363.
(22) Navas, A.; Albar, J. P. Application of proteomics in phylogenetic
and evolutionary studies. Proteomics 2004, 4, 299–302.
(23) Karr, T. L. Application of proteomics to ecology and population
biology. Heredity 2008, 100, 200–206.
(24) Conway, G. C.; Smole, S. C.; Sarracino, D. A.; Arbeit, R. D.;
Leopold, P. E. Phyloproteomics: species identification of Entero-
bacteriaceae using matrix-assisted laser desorption/ionization time-
of-flight mass spectrometry. J. Mol. Microbiol. Biotechnol. 2001, 3,
103–112.
(25) Dickinson, D. N.; La Duc, M. T.; Haskins, W. E.; Gornushkin, I.;
Winefordner, J. D.; Powell, D. H.; Venkateswaran, K. Species
differentiation of a diverse suite of Bacillus spores by mass spectro-
metry-based protein profiling. Appl. Environ. Microbiol. 2004, 70,
475–482.
(26) Mazzeo,M. F.; Sorrentino, A.; Gaita,M.; Cacace, G.; Di Stasio,M.;
Facchiano, A.; Comi, G.; Malorni, A.; Siciliano, R. A. Matrix-
assisted laser desorption ionization-time of flight mass spectrometry
for the discrimination of food-borne microorganisms.Appl. Environ.
Microbiol. 2006, 72, 1180–1189.
(27) Perkins, D. N.; Pappin, D. J. C.; Creasy, D. M.; Cottrell, J. S.
Probability-based protein identification by searching sequence
databases using mass spectrometry data. Electrophoresis 1999, 20,
3551–3567.
(28) Pineiro, C.; Barros-Velazquez, J.; Sotelo, C. G.; Perez-Martin, R. I.;
Gallardo, J. M. Two-dimensional electrophoretic study of the water-
soluble protein fraction in white muscle of Gadoid fish species.
J. Agric. Food Chem. 1998, 46, 3991–3997.
(29) Jensen, O. N.; Wilm, M.; Shevchenko, A.; Mann, M. Sample
Preparation Methods for Mass Spectrometric Peptide Mapping
Directly from 2-DE Gels. In 2-D Proteome Analysis Protocols; Link,
A. J., Ed.; Humana Press: Totowa, NJ, 1999; pp 513-530.
(30) Ochiai, A. Zoogeographical studies on the Soleoid fishes found in
Japan and its neighbouring regions. Bull. Jpn. Soc. Sci. Fish. 1957,
22, 526–530.
(31) Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0. Mol.
Biol. Evol. 2007, 24, 1596–1599.
(32) Saitou, N.; Nei, M. The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425.
(33) Efron, B. The Jackknife, the Bootstrap and Other Resampling Plans.
CBMS-NSF Regional Conference Series in Applied Mathematics,
Monograph 38, SIAM: Philadelphia, PA, 1982.
(34) Thompson, J. D.; Higgins, D. G.; Gibson, T. J. CLUSTAL W:
improving the sensitivity of progressive multiple sequence alignment
through sequence weighting, positions-specific gap penalties and
weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680.
(35) Yu, C.-J.; Lin, Y.-F.; Chiang, B.-L.; Chow, L.-P. Proteomics and
immunological analysis of a novel shrimp allergen, Pen m 2.
J. Immunol. 2003, 170, 445–453.
Received February 13, 2009.Revisedmanuscript receivedMay12, 2009.
AcceptedMay 15, 2009. This work was supported by the National Food
Program of the INIA (Spanish Ministry for Education and Science)
(Project CAL-03-030-C2-2) and by the PGIDIT Research Program in
Marine Resources (Project PGIDIT04RMA261004PR) of the Xunta
de Galicia (Galician Council for Industry, Commerce and Innovation).
I.O. is supported by the FPU program (AP-2004-5826) and B.C. is
supported by the RyC program, under the auspices of the Spanish
MICINN.
138

1
SUPPORTING INFORMATION:
RESULTS AND DISCUSSION
2-DE
Table S-1. Mean pI of the analyzed spot, calculated from the 2-DE patterns of all
individuals analysed of each species.
species pI ± RSD
Penaeus monodon 4.99 ± 0.6
Litopenaeus vannamei 4.98 ± 0.8
Fenneropenaeus indicus 5.18 ± 0.3
Farfantepenaeus notialis 5.22 ± 0.6
Pleoticus muelleri 5.25 ± 1.1
Pandalus borealis 4.68 ± 1.6
139

2
MALDI-TOF MS analysis
Table S-2. Complete peak lists obtained from arginine kinase PMF of 28 specimens.
MPN A MPN B MPN C MPN D MPN E MPN F PNV A PNV B PNV C PNI A PNI B PNI E PNI C
920.53 977.56 1008.50 1008.50 1008.49 1008.50 1008.48 948.50 1008.49 1008.50 1008.50 1050.47 1008.50
977.56 1008.50 1050.52 1050.51 1050.51 1050.51 1050.50 958.52 1050.51 1050.52 1050.52 1133.58 1050.52
1008.50 1050.52 1077.59 1077.59 1077.58 1077.57 1147.59 1003.50 1147.61 1147.61 1147.63 1476.66 1259.69
1050.521 1070.43 1147.62 1147.61 1147.60 1147.60 1149.59 1008.50 1149.61 1259.70 1259.69 1503.80 1503.81
1077.58 1077.57 1149.60 1205.69 1149.63 1164.59 1164.59 1050.51 1164.59 1503.81 1503.80 1518.83 1507.70
1147.62 1147.60 1164.59 1259.70 1259.69 1205.67 1259.69 1147.61 1259.70 1507.71 1507.69 1603.82 1518.82
1149.63 1164.59 1259.69 1296.69 1503.80 1259.69 1369.75 1149.58 1296.64 1518.84 1518.81 1604.81 1525.77
1205.69 1205.67 1438.74 1415.81 1507.70 1415.79 1415.79 1164.59 1366.69 1525.83 1558.82 1651.83 1603.82
1259.70 1259.71 1503.82 1503.81 1518.82 1503.80 1438.76 1259.67 1369.76 1603.83 1603.82 1657.83 1604.81
1296.64 1415.79 1507.72 1507.73 1525.78 1507.70 1507.70 1415.74 1409.72 1604.82 1604.81 1683.83 1651.84
1399.77 1438.77 1518.81 1518.84 1604.80 1518.82 1513.84 1438.72 1415.79 1619.81 1606.82 1775.97 1657.83
1415.80 1503.79 1525.76 1525.79 1631.88 1603.82 1531.82 1507.69 1438.76 1622.89 1619.81 1791.58 1775.98
1476.68 1507.73 1603.81 1604.82 1651.84 1604.81 1553.81 1531.83 1476.67 1651.83 1622.85 1796.94 1796.94
1503.81 1518.83 1604.81 1631.90 1657.84 1631.89 1604.81 1553.75 1507.70 1657.83 1651.84 1807.92 1807.91
1518.83 1525.79 1631.90 1651.86 1683.79 1651.84 1651.84 1558.88 1513.83 1667.78 1657.83 1810.91 2015.94
1527.85 1527.84 1651.84 1657.85 1775.98 1657.84 1657.84 1603.77 1518.83 1683.77 1667.79 2033.95 2033.94
1541.77 1604.80 1657.83 1683.79 1796.93 1667.80 1667.83 1604.80 1531.83 1775.99 1683.78 2107.06 2599.24
1603.84 1631.88 1776.01 1775.98 1807.91 1683.80 1683.82 1606.83 1576.79 1796.92 1775.99 2284.92 2619.93
1604.81 1651.84 1796.93 1796.92 1810.93 1775.97 1761.95 1622.85 1603.83 1810.91 1796.94 2301.27 2636.26
1631.89 1657.84 1807.90 1875.87 2015.94 1796.93 1791.86 1651.83 1604.81 1839.86 2015.94 2346.15
1651.84 1775.98 1875.88 2015.95 2031.97 1807.92 1796.93 1657.83 1622.78 1875.89 2033.93 2426.18
1657.84 1796.92 2015.97 2031.97 2033.94 1810.94 1807.93 1683.84 1651.84 2015.95 2599.20 2520.30
1689.82 2015.96 2033.99 2033.94 2190.01 1839.88 1823.89 1761.96 1657.84 2033.96 2619.99 2599.28
1775.97 2031.98 2190.10 2093.04 2284.97 1875.87 1839.89 1796.95 1667.82 2599.21 2636.29 2620.00
1796.92 2033.95 2301.33 2190.00 2301.24 2015.95 1875.87 1807.91 1683.81 2619.97 2636.29
1807.91 2190.00 2599.54 2301.22 2346.11 2031.97 2015.94 2015.99 1761.95 2636.19 2770.01
1875.87 2285.02 2346.08 2472.15 2033.95 2031.98 2033.99 1778.88
2786.42
2031.98 2301.24 2599.19 2520.26 2190.02 2033.95 2190.15 1796.93
2033.95 2346.07 2636.17 2599.30 2301.24 2135.10 2270.98 1807.92
2190.03 2599.23
2619.95 2346.11 2190.03 2287.38 1823.91
2301.24
2623.14 2472.17 2247.08 2599.56 1839.89
2346.12
2636.28 2520.26 2270.97 2636.53 1875.87
2599.22
2701.25 2548.24 2287.23
2015.94
2631.24
2599.26 2329.82 2031.97
3420.59
2619.00 2346.12 2033.95
2623.11 2431.13 2112.00
2636.26 2548.28 2114.02
2701.22 2599.24 2190.03
2620.04 2270.92
2636.25 2287.23
2786.38 2329.85
2346.12
2431.12
2548.33
2559.19
2599.24
2619.96
2636.27
140

3
Table S-2 (Continued).
PNI D SOP 1 SOP 2A SOP 2C LAA 1 LAA 2A LAA 2B LAA D LAA E LAA F LAA G PRA A PARA B
1008.50 1008.50 1008.50 1008.50 946.25 977.54 1050.53 994.48 948.49 994.48 994.48 1196.49 1141.53
1050.52 1050.53 1050.51 1014.62 994.49 994.48 1263.67 1050.51 959.51 1050.51 1050.50 1228.47 1158.56
1133.59 1147.61 1147.60 1050.52 1050.53 1050.53 1424.75 1133.62 994.48 1263.70 1263.62 1634.74 1194.56
1503.80 1259.72 1164.60 1133.66 1133.64 1133.61 1476.66 1263.69 1050.52 1286.72 1286.70 1638.74 1196.58
1518.83 1415.80 1259.69 1147.60 1147.61 1147.58 1503.77 1286.70 1147.62 1503.80 1442.76 1650.74 1634.82
1603.85 1476.81 1415.79 1164.60 1263.70 1150.56 1532.85 1424.74 1150.58 1532.85 1503.74 1666.73 1666.81
1604.81 1503.81 1503.79 1169.60 1286.71 1263.66 1560.82 1442.81 1263.69 1604.82 1532.84 1690.81 1690.88
1606.84 1507.74 1507.70 1259.70 1357.60 1286.70 1603.85 1503.80 1286.70 1622.92 1604.80 1757.78 1757.85
1651.85 1603.82 1603.83 1415.79 1366.69 1366.68 1604.81 1507.71 1424.76 1651.84 1623.14 1758.79 1766.91
1657.83 1604.81 1604.81 1471.82 1442.80 1424.74 1651.83 1525.79 1442.80 1667.82 1683.76 1766.85 1775.87
1775.98 1631.90 1622.83 1476.80 1476.67 1442.78 1667.79 1532.84 1503.80 1683.81 1718.78 1775.81 1812.83
1796.96 1651.83 1631.89 1503.79 1503.80 1503.77 1683.78 1541.85 1507.72 1718.80 1789.99 1812.76 1828.81
1807.90 1657.83 1651.84 1507.71 1532.83 1507.71 1718.79 1603.82 1525.79 1789.99 1796.92 1828.76 2080.85
1810.92 1667.81 1657.83 1525.78 1541.78 1525.80 1789.97 1604.81 1532.84 1796.94 1875.89 1844.76 2126.04
2015.95 1673.86 1667.80 1603.83 1560.87 1532.84 1796.92 1622.80 1541.83 1857.88 2015.95 2080.94 2142.02
2033.93 1683.81 1683.79 1604.81 1603.81 1541.85 1807.90 1631.89 1604.81 1875.87 2033.95 2126.01 2350.26
2426.12 1719.93 1719.93 1606.81 1604.80 1603.83 1823.87 1651.83 1622.79 2033.94 2190.07 2142.00 2531.28
2599.24 1796.93 1778.93 1631.88 1606.81 1604.80 1839.85 1667.83 1651.82 2426.15 2636.36 2350.26
2636.25 1807.92 1796.94 1651.83 1631.89 1651.82 1841.86 1683.82 1683.79 2636.25 2644.29 2563.28
1810.95 1807.92 1657.83 1651.82 1683.78 2015.91 1718.79 1718.78 2644.17
1839.89 1810.93 1667.80 1667.81 1718.80 2031.97 1789.98 1778.85
1875.88 1823.89 1683.81 1683.81 1790.01 2033.95 1796.93 1790.01
2015.95 1839.88 1719.94 1718.79 1796.96 2064.02 1807.92 1796.93
2031.98 1875.87 1741.87 1789.98 1839.86 2107.05 1839.91 1839.85
2033.95 2015.96 1796.94 1796.92 2015.94 2135.06 1841.90 2015.94
2190.03 2031.98 1807.91 1807.90 2031.99 2190.02 1875.87 2031.98
2346.11 2033.95 1810.94 1841.88 2033.95 2238.07 2015.94 2033.95
2442.16 2190.03 1875.87 1875.88 2315.28 2315.25 2031.98 2620.04
2520.26 2346.12 2015.95 2015.94 2346.07 2346.10 2033.95 2644.31
2548.23 2442.14 2031.98 2031.99 2644.24 2426.13 2135.08
2564.23 2520.23 2033.95 2033.95 2520.23 2190.04
2599.23 2548.19 2055.87 2107.05 2620.06 2298.96
2636.26 2599.25 2190.02 2135.06 2623.08 2315.28
2655.08 2620.00 2346.11 2190.02 2636.25 2329.78
2670.19 2636.24 2520.26 2298.93 2640.15 2346.15
2687.21 2640.15 2599.25 2315.29 2644.16 2520.28
2687.17 2619.97 2329.79 2671.13 2619.99
2636.26 2346.13 2772.29 2636.37
2426.13
2644.24
2520.26
2756.09
2619.99
2772.40
2623.19
2636.34
.
2644.20
2772.36
141

4
Table S-2 (Continued).
PRA C PRA D
1141.59 1141.69
1196.64 1158.71
1634.85 1194.71
1638.84 1196.73
1666.84 1382.87
1690.90 1634.88
1757.88 1690.94
1758.87 1757.91
1766.94 1766.96
1775.89 1775.91
1812.84 1812.86
1828.82 1828.86
1966.89 1966.90
1987.89 2126.05
2080.93 2350.24
2126.03 2531.23
2142.02 2616.22
2350.24
2563.22
142
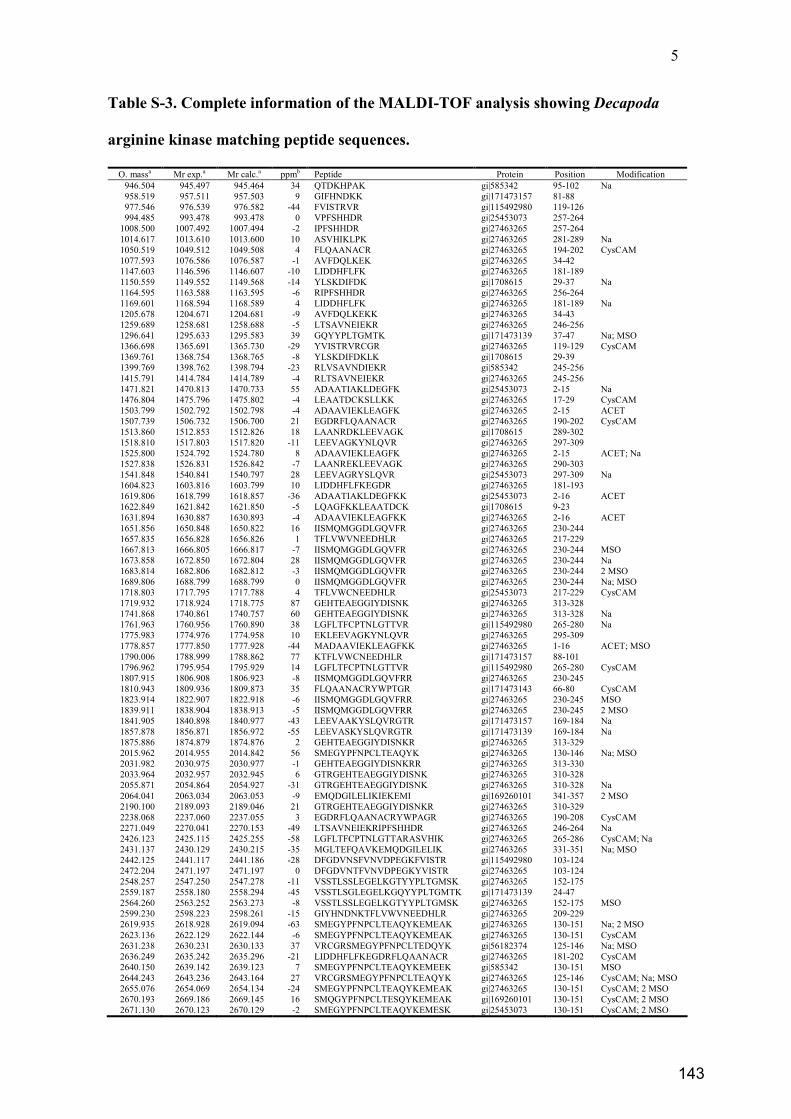
5
Table S-3. Complete information of the MALDI-TOF analysis showing Decapoda
arginine kinase matching peptide sequences.
O. massa Mr exp.a Mr calc.a ppmb Peptide Protein Position Modification
946.504 945.497 945.464 34 QTDKHPAK gi|585342 95-102 Na
958.519 957.511 957.503 9 GIFHNDKK gi|171473157 81-88
977.546 976.539 976.582 -44 FVISTRVR gi|115492980 119-126
994.485 993.478 993.478 0 VPFSHHDR gi|25453073 257-264
1008.500 1007.492 1007.494 -2 IPFSHHDR gi|27463265 257-264
1014.617 1013.610 1013.600 10 ASVHIKLPK gi|27463265 281-289 Na
1050.519 1049.512 1049.508 4 FLQAANACR gi|27463265 194-202 CysCAM
1077.593 1076.586 1076.587 -1 AVFDQLKEK gi|27463265 34-42
1147.603 1146.596 1146.607 -10 LIDDHFLFK gi|27463265 181-189
1150.559 1149.552 1149.568 -14 YLSKDIFDK gi|1708615 29-37 Na
1164.595 1163.588 1163.595 -6 RIPFSHHDR gi|27463265 256-264
1169.601 1168.594 1168.589 4 LIDDHFLFK gi|27463265 181-189 Na
1205.678 1204.671 1204.681 -9 AVFDQLKEKK gi|27463265 34-43
1259.689 1258.681 1258.688 -5 LTSAVNEIEKR gi|27463265 246-256
1296.641 1295.633 1295.583 39 GQYYPLTGMTK gi|171473139 37-47 Na; MSO
1366.698 1365.691 1365.730 -29 YVISTRVRCGR gi|27463265 119-129 CysCAM
1369.761 1368.754 1368.765 -8 YLSKDIFDKLK gi|1708615 29-39
1399.769 1398.762 1398.794 -23 RLVSAVNDIEKR gi|585342 245-256
1415.791 1414.784 1414.789 -4 RLTSAVNEIEKR gi|27463265 245-256
1471.821 1470.813 1470.733 55 ADAATIAKLDEGFK gi|25453073 2-15 Na
1476.804 1475.796 1475.802 -4 LEAATDCKSLLKK gi|27463265 17-29 CysCAM
1503.799 1502.792 1502.798 -4 ADAAVIEKLEAGFK gi|27463265 2-15 ACET
1507.739 1506.732 1506.700 21 EGDRFLQAANACR gi|27463265 190-202 CysCAM
1513.860 1512.853 1512.826 18 LAANRDKLEEVAGK gi|1708615 289-302
1518.810 1517.803 1517.820 -11 LEEVAGKYNLQVR gi|27463265 297-309
1525.800 1524.792 1524.780 8 ADAAVIEKLEAGFK gi|27463265 2-15 ACET; Na
1527.838 1526.831 1526.842 -7 LAANREKLEEVAGK gi|27463265 290-303
1541.848 1540.841 1540.797 28 LEEVAGRYSLQVR gi|25453073 297-309 Na
1604.823 1603.816 1603.799 10 LIDDHFLFKEGDR gi|27463265 181-193
1619.806 1618.799 1618.857 -36 ADAATIAKLDEGFKK gi|25453073 2-16 ACET
1622.849 1621.842 1621.850 -5 LQAGFKKLEAATDCK gi|1708615 9-23
1631.894 1630.887 1630.893 -4 ADAAVIEKLEAGFKK gi|27463265 2-16 ACET
1651.856 1650.848 1650.822 16 IISMQMGGDLGQVFR gi|27463265 230-244
1657.835 1656.828 1656.826 1 TFLVWVNEEDHLR gi|27463265 217-229
1667.813 1666.805 1666.817 -7 IISMQMGGDLGQVFR gi|27463265 230-244 MSO
1673.858 1672.850 1672.804 28 IISMQMGGDLGQVFR gi|27463265 230-244 Na
1683.814 1682.806 1682.812 -3 IISMQMGGDLGQVFR gi|27463265 230-244 2 MSO
1689.806 1688.799 1688.799 0 IISMQMGGDLGQVFR gi|27463265 230-244 Na; MSO
1718.803 1717.795 1717.788 4 TFLVWCNEEDHLR gi|25453073 217-229 CysCAM
1719.932 1718.924 1718.775 87 GEHTEAEGGIYDISNK gi|27463265 313-328
1741.868 1740.861 1740.757 60 GEHTEAEGGIYDISNK gi|27463265 313-328 Na
1761.963 1760.956 1760.890 38 LGFLTFCPTNLGTTVR gi|115492980 265-280 Na
1775.983 1774.976 1774.958 10 EKLEEVAGKYNLQVR gi|27463265 295-309
1778.857 1777.850 1777.928 -44 MADAAVIEKLEAGFKK gi|27463265 1-16 ACET; MSO
1790.006 1788.999 1788.862 77 KTFLVWCNEEDHLR gi|171473157 88-101
1796.962 1795.954 1795.929 14 LGFLTFCPTNLGTTVR gi|115492980 265-280 CysCAM
1807.915 1806.908 1806.923 -8 IISMQMGGDLGQVFRR gi|27463265 230-245
1810.943 1809.936 1809.873 35 FLQAANACRYWPTGR gi|171473143 66-80 CysCAM
1823.914 1822.907 1822.918 -6 IISMQMGGDLGQVFRR gi|27463265 230-245 MSO
1839.911 1838.904 1838.913 -5 IISMQMGGDLGQVFRR gi|27463265 230-245 2 MSO
1841.905 1840.898 1840.977 -43 LEEVAAKYSLQVRGTR gi|171473157 169-184 Na
1857.878 1856.871 1856.972 -55 LEEVASKYSLQVRGTR gi|171473139 169-184 Na
1875.886 1874.879 1874.876 2 GEHTEAEGGIYDISNKR gi|27463265 313-329
2015.962 2014.955 2014.842 56 SMEGYPFNPCLTEAQYK gi|27463265 130-146 Na; MSO
2031.982 2030.975 2030.977 -1 GEHTEAEGGIYDISNKRR gi|27463265 313-330
2033.964 2032.957 2032.945 6 GTRGEHTEAEGGIYDISNK gi|27463265 310-328
2055.871 2054.864 2054.927 -31 GTRGEHTEAEGGIYDISNK gi|27463265 310-328 Na
2064.041 2063.034 2063.053 -9 EMQDGILELIKIEKEMI gi|169260101 341-357 2 MSO
2190.100 2189.093 2189.046 21 GTRGEHTEAEGGIYDISNKR gi|27463265 310-329
2238.068 2237.060 2237.055 3 EGDRFLQAANACRYWPAGR gi|27463265 190-208 CysCAM
2271.049 2270.041 2270.153 -49 LTSAVNEIEKRIPFSHHDR gi|27463265 246-264 Na
2426.123 2425.115 2425.255 -58 LGFLTFCPTNLGTTARASVHIK gi|27463265 265-286 CysCAM; Na
2431.137 2430.129 2430.215 -35 MGLTEFQAVKEMQDGILELIK gi|27463265 331-351 Na; MSO
2442.125 2441.117 2441.186 -28 DFGDVNSFVNVDPEGKFVISTR gi|115492980 103-124
2472.204 2471.197 2471.197 0 DFGDVNTFVNVDPEGKYVISTR gi|27463265 103-124
2548.257 2547.250 2547.278 -11 VSSTLSSLEGELKGTYYPLTGMSK gi|27463265 152-175
2559.187 2558.180 2558.294 -45 VSSTLSGLEGELKGQYYPLTGMTK gi|171473139 24-47
2564.260 2563.252 2563.273 -8 VSSTLSSLEGELKGTYYPLTGMSK gi|27463265 152-175 MSO
2599.230 2598.223 2598.261 -15 GIYHNDNKTFLVWVNEEDHLR gi|27463265 209-229
2619.935 2618.928 2619.094 -63 SMEGYPFNPCLTEAQYKEMEAK gi|27463265 130-151 Na; 2 MSO
2623.136 2622.129 2622.144 -6 SMEGYPFNPCLTEAQYKEMEAK gi|27463265 130-151 CysCAM
2631.238 2630.231 2630.133 37 VRCGRSMEGYPFNPCLTEDQYK gi|56182374 125-146 Na; MSO
2636.249 2635.242 2635.296 -21 LIDDHFLFKEGDRFLQAANACR gi|27463265 181-202 CysCAM
2640.150 2639.142 2639.123 7 SMEGYPFNPCLTEAQYKEMEEK gi|585342 130-151 MSO
2644.243 2643.236 2643.164 27 VRCGRSMEGYPFNPCLTEAQYK gi|27463265 125-146 CysCAM; Na; MSO
2655.076 2654.069 2654.134 -24 SMEGYPFNPCLTEAQYKEMEAK gi|27463265 130-151 CysCAM; 2 MSO
2670.193 2669.186 2669.145 16 SMQGYPFNPCLTESQYKEMEAK gi|169260101 130-151 CysCAM; 2 MSO
2671.130 2670.123 2670.129 -2 SMEGYPFNPCLTEAQYKEMESK gi|25453073 130-151 CysCAM; 2 MSO
143

6
2687.269 2686.262 2686.259 1 FLQAANACRYWPAGRGIYHNDNK gi|27463265 194-216 Na
2701.352 2700.345 2700.359 -5 IPFSHHDRLGFLTFCPTNLGTTAR gi|27463265 257-280
2770.178 2769.171 2769.208 -13 RSMEGYPFNPCLTEAQYKEMESK gi|171473143 1-23 2 MSO
2772.403 2771.395 2771.397 0 VPFSHHDRLGFLTFCPTNLGTTVR gi|25453073 257-280 CysCAM
O.mass (observed mass, MH+), Mr exp. (neutral observed mass) and Mr calc. (neutral calculated mass);
a:
peaks represent monoisotopic mass in Da. b: Mr exp-Mr calc in Da. Na: adduct sodium; MSO: methionine
sulphoxide; ACET: acetyl N-terminal; CysCAM: carboxamidomethylated cysteine.
Table S-4. Matrix of peaks presence (1 = Present; 0 = Absent) of all different
specimens
1 2 3 4
0 0 0 0
PRA A 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA C 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA D 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
MPN A 0 0 0 0 0 1 0 0 1 0 1 1 1 0 0 1 0 0 0 1 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 1 1 0 1 1 1 0 0
MPN B 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 1 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 1 1 0 0 0
MPN C 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 1 1 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 1 1 0 0 0
MPN D 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0
MPN E 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 1 0 0 0 1 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 1 0 0 0 0
MPN F 1 0 0 0 0 1 0 0 1 0 1 0 1 0 0 1 1 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 1 0 1 0 0 0 0 1 1 0 0 1 0 1 0 0
PNV A 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 1 0 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 1 0 1 0 0 0 0 1 1 0 0 0 1 0
PNV B 0 0 1 1 0 0 0 1 1 0 1 0 0 0 0 1 1 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 0 0 0 0 1 0
PNV C 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 1 0 0 1 0 0 0 0 0 1 0 0 1 0 1 1 0 0 1 1 0 1 0 0 1 0 1 1 1 0 0 1 0
PNI A 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 1 0 0 0 0
PNI B 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0
PNI C 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0
PNI D 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0
PNI E 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0
SOP A 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0
SOP B 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 1 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 0 0 0 0
SOP C 0 0 0 0 0 0 0 0 1 1 1 0 0 1 0 1 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 1 1 0 0 1 0 0 0
LAA A 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 1
LAA B 0 0 0 0 0 1 1 0 0 0 1 0 0 1 0 1 0 1 0 0 0 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 1 0 1 0 0 1 1 0 0 1 0 0 1
LAA C 0 1 0 0 0 0 1 0 0 0 1 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 1 0 1 1 0 0 0 0 0 0 1
LAA D 0 0 0 0 0 0 1 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0 1 0 0 1
LAA E 0 0 1 0 1 0 1 0 0 0 1 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 1 0 0 1 1 0 0 1 0 0 1
LAA F 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1
LAA G 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1
144

7
Table S-4 (Continued).
5 6 7 8 9
0 0 0 0 0
PRA A 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 1 0 0 0 0 1 0 0 0 1 1 0 1 1 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0
PRA B 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 1 0 0 0 1 0 0 1 1 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0
PRA C 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 0 0 1 0 0 0 1 1 0 1 1 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 1 0 0 0
PRA D 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 1 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 0 0 0 0
MPN A 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1
MPN B 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 1 1
MPN C 0 0 0 0 0 1 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 1 0 1
MPN D 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 1 1 1
MPN E 0 0 0 0 0 1 1 0 0 0 1 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 1 0 0 0 1 0 0 0 1 0 0 1 1 1
MPN F 1 0 0 0 0 1 1 0 0 0 1 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 0 1 1
PNV A 0 1 0 0 0 0 1 0 0 0 0 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1
PNV B 0 1 1 0 0 1 1 1 0 1 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 1
PNV C 0 0 0 0 1 1 1 0 0 1 0 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 0 1 0 1 0 0 0 1 0 0 1 1 1
PNI A 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1
PNI B 0 0 0 0 0 1 1 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1
PNI C 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 1
PNI D 0 0 1 0 0 1 1 1 1 1 0 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1
PNI E 0 0 0 0 0 1 1 0 1 1 0 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 1 0 1
SOP A 0 0 0 0 0 1 1 0 0 0 1 0 0 0 1 1 0 1 1 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 1 0 0 0 1 0 0 1 1 1
SOP B 0 0 0 0 0 1 1 0 0 1 1 0 0 0 1 1 0 1 0 1 0 0 0 1 0 0 0 0 0 0 1 0 0 1 1 1 0 1 0 1 0 0 0 1 0 0 1 1 1
SOP C 0 0 0 0 0 1 1 1 0 0 1 0 0 0 1 1 0 1 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 1 0 0 1 1 1
LAA A 0 0 0 1 0 1 1 0 0 0 0 0 0 0 1 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 1 0 0 1 0 1 1 0 0 0 0 0 1 1 1
LAA B 1 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 0 0 0 0 0 0 1 1 1
LAA C 1 0 0 1 0 1 1 1 0 0 1 0 0 0 1 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 1 0 0 0 0 0 1 0 0 1 0 0 1 1 1
LAA D 1 0 0 0 0 1 1 0 0 1 1 0 0 0 1 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 1 0 0 0 0 1 1 0 0 1 0 0 1 1 1
LAA E 1 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 1 0 0 0 0 0 0 1 1 1
LAA F 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 1
LAA G 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 1
145

8
Table S-4 (Continued).
1 1 1 1 1
0 1 2 3 4
0 0 0 0 0
PRA A 0 0 1 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA B 0 0 1 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA C 0 0 1 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
PRA D 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
MPN A 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
MPN B 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
MPN C 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
MPN D 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 0 0 1 0 0 0 0 1 1 0 0 0 0 0 1 0 1 1 0 1 0 0 0 0 0 0 1 0 0 0 0
MPN E 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 1 0 1 0 0 0 1 0 1 1 0 1 0 0 0 0 0 0 1 0 0 0 0
MPN F 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0
PNV A 0 0 0 0 0 0 0 0 1 0 1 0 1 1 0 1 0 0 0 1 1 0 0 1 0 0 0 0 1 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 1
PNV B 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
PNV C 0 0 0 0 0 1 1 0 0 0 1 0 0 1 0 1 0 0 0 1 1 0 0 1 0 0 0 0 1 1 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0
PNI A 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1 0 1 0 0 0 1 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 1
PNI B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
PNI C 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0
PNI D 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0
PNI E 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0
SOP A 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 1 0 0 1 1 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0
SOP B 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 1 0 0 0 1 0 1 0 0 1 1 0 0 0 0 1 0 0 0 0 0
SOP C 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0
LAA A 0 1 0 0 1 0 0 0 1 0 1 1 0 0 0 0 0 0 1 0 1 0 1 0 0 0 1 0 0 0 0 0 0 0 1 1 0 1 1 1 0 0 1 0 0 0 0 1 0
LAA B 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0
LAA C 0 0 0 0 1 0 0 0 1 0 1 0 0 0 0 0 1 0 1 1 1 0 1 0 0 0 1 0 0 0 0 0 0 0 1 1 0 1 0 1 0 0 0 0 0 0 0 1 0
LAA D 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 0 1 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 1 0 1 0 0 0 0 0 1 0 1 0
LAA E 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0
LAA F 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
LAA G 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
146

CAPÍTULO 4
CARACTERIZACIÓN POR ESPECTROMETRÍA DE MASAS DE PÉPTIDOS DE ARGININA QUINASA ESPECÍFICOS PARA ESPECIES DE LANGOSTINOS COMERCIALES


Capítulo 4: Caracterización por espectrometría de masas de péptidos de
arginina quinasa específicos para especies de langostinos comerciales.
Comentario
Tras demostrarse la posibilidad de diferenciar las especies de langostinos estudiadas utilizando la
huella peptídica de la arginina quinasa, el siguiente paso consistió en identificar y caracterizar los
péptidos diferenciadores.
Este trabajo consistió en una aproximación proteómica combinando 2-DE, digestión tríptica en
gel, MALDI-TOF MS y análisis ESI-MS/MS (Figura 21) a partir de los extractos proteicos
sarcoplásmicos de siete especies de langostinos de interés comercial. En una primera etapa se realizó un
estudio exhaustivo de los mapas peptídicos de la arginina quinasa, estableciendo los picos diferenciales.
Figura 21. Etapas consideradas y herramientas proteómicas utilizadas para la identificación y
caracterización de péptidos específicos en langostinos.
149

Capítulo 4: Caracterización por espectrometría de masas de péptidos (…)
En una segunda etapa, se obtuvieron los espectros de fragmentación del mayor número posible de
péptidos de la arginina quinasa, mediante ESI-IT-MS, con objeto de caracterizar sus secuencias,
prestando especial atención a aquellos péptidos especie-específicos o diferenciales que pudieran ser
utilizados para autentificación de especies. Al disponer de pocas secuencias en las bases de datos
genómicas y proteómicas de los miembros del orden Decapoda, fue necesario recurrir a la
secuenciación de novo de muchos de los espectros.
Los resultados obtenidos demuestran que, aunque se encontró un alto grado de homología entre
las secuencias de arginina quinasa, las diferencias descritas nos permiten la inequívoca diferenciación de
estas especies.
150

PUBLICACIÓN
151


���������� ���������������������������������������������� ���������������������������������������
�� �������������������� ���������
�� ����!� ������������������ ��������
��� ������"!� ���#$$%�$$&&'��
��� �����(���!� ������
"���� )�����)�� ��� ��!�
#'�� ��#$$%�
��������*������� ���!� +���,�������-��.�.�.�.,����� ��������������������������������,�/����-������ �����0������������������,����������� �������1�������,�����-��.�.�.�.,����� ��������������������������-��.�.�.�.,����� ���������������������������
ACS Paragon Plus Environment
Journal of Proteome Research
153

Mass Spectrometry Characterization of Species-
Specific Peptides from Arginine Kinase for the
Identification of Commercially Relevant
Shrimp Species
Ignacio Ortea1*, Benito Cañas2, and José M. Gallardo1
AUTHORS ADDRESS
1 Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Científicas
(CSIC), Eduardo Cabello 6, E-36208 Vigo, Spain.
2 Analytical Chemistry Department, Universidad Complutense de Madrid. E-28040
Madrid, Spain.
TITLE RUNNING HEAD. Shrimp species identification by MS/MS.
* CORRESPONDING AUTHOR (telephone: 0034-986-231-930); fax: 0034-986-292-
762; e-mail: [email protected]).
ACS Paragon Plus Environment
Journal of Proteome Research
154

ABSTRACT
The identification of commercial shrimp species is a relevant issue to ensure correct
labelling, maintain consumer confidence and enhance the knowledge of the captured
species, benefiting both, fisheries and manufacturers. A proteomic approach, based on
2DE, tryptic in-gel digestion, MALDI-TOF MS, and ESI-MS/MS analyses, is proposed
for the identification of shrimp species with commercial interest. MALDI-TOF peptide
mass fingerprint from arginine kinase tryptic digests were used for the identification of
seven commercial, closely related species of Decapoda shrimps. Further identification
and characterization of these peptides was performed by CID on an ESI-IT instrument,
database search and de novo sequence interpretation, paying special attention to
differential, species-specific, peptides. Fisheries and manufacturers may take advantage
of this methodology as a tool for a rapid and effective seafood product identification and
authentication, providing and guaranteeing the quality and safety of the foodstuffs to
consumers.
KEYWORDS. Arginine kinase, MS/MS, proteomics, shrimps, species identification.
ACS Paragon Plus Environment
Journal of Proteome Research
155

Introduction
Sea food products include an extense variety of species widely used for human
nutrition, having a significant impact in food industry. Among them, crustaceans
belonging to the order of Decapoda are of remarkable commercial interest. This order
includes the Superfamily Penaeoidea, which is one of the most important economic
resources in fishery and aquafarming industry.1,2 The identification of marine species is
of primary relevance for the seafood industry, due to the global commercial demands
concerning labelling and traceability3,4 for the prevention of commercial fraud, and to
avoid the safety risks derived from the inadvertent introduction of food ingredients
which might be harmful for human health.5 Anatomical characters are particularly
difficult to be used in penaeid shrimp species differentiation due to their phenotypic
similarities and to removal of the external carapace, which is very frequent during the
manufacturing process. Therefore, it is highly recommendable to develop the analytical
tools necessary to make possible the distinction between these closely related species,
preventing the inadvertent or deliberate mislabelling and adulteration of these products.
Methods for species identification are currently based on DNA or protein analysis.
Among the DNA targets, the mitochondrial genes 16S rRNA and cytochrome oxidase I
have been used as interspecific markers for several crustacean species, although most of
the studies focused on phylogeography and phylogenetic relationships,6-10 but not on
species identification. More recently, two PCR-RFLP-based methods for the detection
of crustacean 11 and penaeid shrimps 12 DNA have been proposed. Protein-based
methodologies have also been used with authentication purposes. Proteomics tools have
been applied to the identification of species in seafood products, 13, 14 but little effort has
been made to elucidate differences among closely related species using mass
spectrometry (MS).15-17 Some electrophoretic and immunological assays have been
ACS Paragon Plus Environment
Journal of Proteome Research
156

reported for the detection and differentiation of decapod crustaceans, 18-20 but only one
MS-based method has been used for the authentication of shrimp species.21
Arginine kinase (EC 2.7.3.3) is a monomeric phosphagen-ATP phosphotransferase,
widely distributed in invertebrates22 and functionally analogous to the, more thoroughly
studied, creatine kinase from vertebrates.23 Arginine kinase (AK) catalyzes the
reversible phosphorylation of arginine using ATP:
Mg ATP + L-arginine � MgADP + L-arginine phosphate + H+
A comparison of AK amino acid content in two types of shrimp, Penaeus aztecus and
Penaeus japonicus, revealed that, although nearly identical, some slight differences
appeared, even in these closely-related species.24 Inter-specific variability was also
reported for the AK derived from four Decapoda species.25 In a previous work, AK
proteins proved to be a potential molecular marker for shrimp species identification,
showing marked differences in the MALDI-TOF MS spectra from tryptic digests.21
Now, using a combination of preparative 2-DE, MALDI-TOF MS, and sequencing by
ESI-IT MS/MS, we have characterized the AK proteins from seven closely related
species of commercial relevance, belonging to the order Decapoda. Efforts were made
in peptide de novo sequencing, as genomes from members belonging to the order
Decapoda still await sequencing. Particular attention was focused on the identification
and characterization of several species-specific peptides for each of the species
analyzed, which may be used as a tool for seafood product authentication.
Materials and Methods
Shrimp Species and Populations Considered. Specimens analyzed are shown in
Table 1. They were collected using either, extractive fishing practices, or from
aquaculture facilities in different continents worldwide. Seven different shrimp species
from the order Decapoda were considered in this study, being five of them from the
ACS Paragon Plus Environment
Journal of Proteome Research
157

Penaeidae family –Penaeus monodon, Litopenaeus vannamei, Fenneropenaeus indicus,
Fenneropenaeus merguiensis, Farfantepenaeus notialis-, one from the Solenoceridae
family –Pleoticus muelleri-, and one from the Pandalidae family –Pandalus borealis.
Penaeidae and Solenoceridae families belong to the superfamily Penaeoidea, and
Pandalidae family belongs to the superfamily Pandaloidea. Shrimps, whole animals,
were frozen on board and shipped to our laboratory for the analyses. Special care was
taken in keeping their morphological characteristics in good shape. At least six
individuals of each species were analyzed. Specimens were classified in their respective
taxons according to their anatomical external features with the help of a marine
biologist from the Marine Sciences Institute (Mediterranean Centre for Marine and
Environmental Research, Higher Council for Scientific Research, CMIMA-CSIC,
Barcelona, Spain) with expertise in penaeid shrimp taxonomy.
Extraction of Sarcoplasmic Proteins. Approximately 1 g of raw white muscle from
each of the individuals studied were homogenized in two volumes of milliQ water using
an Ultra-Turrax blender for 3x15 s with interruptions of 45 s to avoid warming the
samples. Sarcoplasmic protein extracts were then centrifuged at 30000g for 15 min at
4ºC (J25 centrifuge; Beckman, Palo Alto, CA), and the supernatants were frozen at -
80ºC until the electrophoretic analysis. Protein concentration in the extracts was
determined by the bicinchoninic acid method (Sigma Chemical Co., USA).
2-DE. Isoelectric focusing (IEF) was performed at 10 ºC in a Multiphor II
electrophoresis unit (Amersham Biosciences, Sweden) as described elsewhere.16
Briefly, protein extracts (4 mg/mL, 100-120 μg of protein) were loaded in duplicate on
IEF precast polyacrylamide gels with a narrow range of pH (Ampholine PAGplate pH
4.0-6.5; GE Healthcare, Uppsala, Sweden), using a sample applicator paper, and run at
1500 V, 50 mA, 30 W until 4000 Vh were reached. To avoid overlapping of proteins,
ACS Paragon Plus Environment
Journal of Proteome Research
158

strips with a narrow pH range (pH 4.0-6.5) were chosen. IEF strips corresponding to
individual lanes were cut after the run was completed and kept at -80 ºC until second
dimension electrophoresis analysis was performed.
Equilibration of pH 4-6.5 IEF gel strips was carried out at room temperature as
described previously.26 Briefly, the strips were placed for 10 min in sample buffer
containing 0.75 % DTT followed by 10 min incubation in sample buffer containing 4.5
% iodoacetamide.
Preparative second dimension SDS-PAGE for further MS analysis were performed at
15 °C in the Multiphor II electrophoresis unit (Amersham Biosciences, Sweden), using
vertical gels (7.5%T and 3%C) with Tris-tricine buffer. Running conditions were: 100
V, 40 mA per gel, and 150 W. Gels were stained with Coomassie Brilliant Blue (CBB)
(GE Healthcare), which is very compatible with sample preparation for MS analysis,
according to the manufacturer’s instructions. PDQuest 2-D Analysis Software Version
7.1.0 (Bio-Rad Laboratories, Hercules, CA) was used for the image analysis.
Peptide Sample Preparation for MS and MS/MS Analysis. AK spots were
localized based on the data (pI and Mr) obtained by previous work, 21 and excised from
the gel taking care in maximizing the protein-to-gel ratio. Only the most intensely
stained region at the centre of the spot was excised to avoid extracting an excess of gel
matrix. Excised pieces were subjected to in-gel digestion with trypsin (Roche
Diagnostics GmbH, Mannheim, Germany) as described elsewhere.27 Prior to nESI-IT
MS analysis, the tryptic peptide samples were desalted and concentrated using in-tip
reverse-phase resins (ZipTip C18, Millipore, Bedford, MA), according to the
manufacturer’s recommendations.
Peptide Mapping. Peptide digests were acidified with TFA and 0.8 μL aliquots were
manually deposited onto the stainless steel MALDI probe and allowed to dry at room
ACS Paragon Plus Environment
Journal of Proteome Research
159

temperature. Matrix solution (0.8 μL of saturated CHCA in 50% aqueous ACN and 0.1
% TFA) was added and again allowed to dry at room temperature.
Mass spectra were obtained using a Voyager DE STR MALDI-TOF mass
spectrometer (Applied Biosystems, Foster City, CA) operating in the reflector, delay
extraction, and positive-ion mode. Laser intensity was set just above the ion generation
threshold. The values for the MS parameters were: low mass gate, m/z 500 Da; delay
time, 350 ns; accelerating voltage, 20,000 V; and grid voltage, 68.5 %. Mass spectra
were acquired by accumulating 150 laser shots in the m/z range from 850 to 3500.
External close calibration with the Calibration Mixture 2 of the Sequazyme Peptide
Mass Standards Kit (Applied Biosystems) was used. Mass spectra were baseline
corrected and data lists containing monoisotopic m/z values were extracted from mass
spectral data with the specific software of the instrument (Data Explorer, version
4.0.0.0). Signals within the 920 to 3500 m/z range, with relative intensities greater than
5% were included in the lists. Database searches were performed against the NCBI non-
redundant protein sequence database (http://www.ncbi.nih.gov) using the MASCOT
Peptide Mass Fingerprint search program (http://matrixscience.com). Search parameters
were: monoisotopic and protonated masses, two missed cleavages allowed, 100 ppm
mass tolerance, and three potential variable modifications: ACET sites (acetyl N-
terminal), CysCAM (carbamidomethylcysteine) and MSO (methionine sulfoxide).
Peptide Sequencing by ESI-IT MS/MS. Peptide digests were analyzed online by
LC-ESI-IT-MS/MS using a LC system model SpectraSystem P4000 (Thermo-Finnigan,
San Jose, CA) coupled to an IT mass spectrometer model LCQ Deca XP Plus (Thermo-
Finnigan). The separation was performed on a 0.18 mm × 150 mm BioBasic-18 RP
column (ThermoHypersil-Keystone) using 0.5% acetic acid in water and 0.5% acetic
acid in ACN as mobile phases A and B, respectively. A 90 min linear gradient from 5 to
ACS Paragon Plus Environment
Journal of Proteome Research
160

60% B, at a flow rate of 1.4-1.7 μL /min, was used. Peptides were detected using survey
scans from 400 to 1600 amu (3 μscans), followed by a data-dependent ZoomScan (5
μscans) and MS/MS scan (5 μscans), using an isolation width of 3 amu and a
normalized collision energy of 35%. Fragmented masses were set in dynamic exclusion
for 3 min after the fragmentation event and singly charged ions were excluded from
MS/MS analysis.
Off-line analyses by nESI-IT were performed using an ion trap mass spectrometer,
model LCQ Deca XP Plus (Thermo-Finnigan) equipped with a nanospray interface.
Peptides were previously desalted by micro-columns ZipTip C18 eluted with 10 μL of
70% MeOH/ 0.5%CH3COOH. PicoTips borosilicate glass needles with 1 μm orifice
(New Objective, Woburn, MA) were filled with 3-5 μL of sample and used as emitters.
MS/MS spectra were searched using SEQUEST (Bioworks 3.1 package, thermo-
Finnigan) against the nr.fasta database (NCBI Resources, NIH, Bethseda MD). The
following constraints were used for the searches: tryptic cleavage after Arg and Lys, up
to two missed cleavage sites, and tolerances ±1.5 Da for precursor ions and ±0.8 Da for
MS/MS fragment ions. The variable modifications allowed were methionine oxidation,
carbamidomethylation of Cys, and acetylation of the N-terminus.
De novo sequencing was performed by manual interpretation of the ion series in the
spectra. BLAST program was used for homology searches between de novo obtained
sequences and those in the nr-NCBI database.
Results and Discussion
2-DE. Sarcoplasmic protein extracts from 6 individuals for each of the 7 species under
study were subjected to 2-DE. Gels were Coomassie stained and analyzed with the
image software. To ensure reproducibility of the experiments, 3 gels were run per
individual. In accordance with previously reported data, 21 arginine kinase isoforms run
ACS Paragon Plus Environment
Journal of Proteome Research
161

at 40 kDa with an isoelectric point (pI) between 4.68 and 5.25. The 2-DE patterns for
the different AKs are shown in Figure S-1 in the Supporting Information and pIs for all
the AK spots studied are compiled in Table S-1 in the Supporting Information.
Differences in the AK spots pIs pointed to the presence of amino acid substitutions in
their sequences; consequently, characterization of each of the AK isoforms was
performed by mass spectrometry.
MALDI-TOF MS Analysis of AK Spots. After preparative 2-DE, AK spots were in
gel digested with trypsin, and the peptides analyzed by MALDI-TOF MS. The complete
list of the peaks detected can be found in Table S-2 in the Supporting Information. Mass
coincidences between experimental data and in silico produced tryptic digests from the
AKs present in protein databases were apparent: 24 peak masses were coincident with
theoretical peptides from the P. monodon allergen Pen m 2 (gi|27463265), seven with
peptides from an AK found in L. vannamei (gi|115492980), another seven with peptides
from Marsupenaeus japonicus AK (gi|1708615); four more peaks matched peptides
from Galapaganus conwayensis (gi|209402936); two peaks were present in the AK
from Solenopsis invicta (gi|192337578); other two were present in the AK from
Chasmagnathus granulata (gi|7243761) and one was contained in the AK sequences
from Periplaneta americana (gi|50428904), Cimberis pilosa (gi|228014712), Apis
mellifera (gi|58585146), Homarus gammarus (gi|585342), Lepidophthalmus
louisianensis (gi|171473157), Artemia franciscana (gi|25453072), Pterapion sp.
(gi|228014764), Cancer antennarius (gi|171473177), and Hemigrapsus oregonensis
(gi|171473159).
MALDI-TOF MS peptide maps containing specific peaks are shown in figure 1.
Highlighted are those used for the definition and differentiation of the species studied.
According to the results presented in table S-2, eleven peaks with an m/z of 1050.51
ACS Paragon Plus Environment
Journal of Proteome Research
162

(Figure 1a), 1507.73, 1604.81, 1683.82, 1796.95, 1807.92, 2015.98, 2031.99, 2033.98,
2190.05, and 2636.32 were present in all the species of the superfamily Penaeoidea.
Several differences were also obtained among the Penaeoidea species, which may allow
the classification of shrimps into two groups (Figure 1b, c, d): the family Penaeidae,
presenting four specific peaks at m/z 1008.49, 1259.63, 1657.85, and 2599.31 Da, and
the family Solenoceridae, with other twelve specific peaks at 994.45, 1263.62, 1286.63,
1424.68, 1442.73, 1532.78, 1541.89, 1718.85, 1789.89, 2315.25, 2644.36, and 2772.53
Da. Within the Penaeidae family, species-specific peaks were found (Figure 1e,f): one
peak with an m/z of 1077.54 was specific to P. monodon; other peak with an m/z of
1463.75 was specific to F. notialis; while two peaks at 2285.06 and 2472.38 Da were
specific for F. merguiensis. 17 specific peaks were found for P. borealis, making the
pattern very different from those found in the other species, and therefore to the family
Pandalidae (e.g. peak at 1634.82 Da in Figure 1d). In addition, intra-species variations,
depending on the origin of the population considered, were found in L. vannamei and F.
indicus: peaks with m/z 1148.53, 1369.71, 1513.78, 1531.75 (Figure 1g), 1761.92,
2271.18, 2287.23, 2431.23, 2548.35, and 3379.73 were found in L. vannamei from
Costa Rica, while peaks with m/z 1503.71, 1631.90, 1775.89, 2301.21, and 3390.74
were found L. vannamei from Argentina, being the last one an specific marker found
only in this L. vannamei population. Differential peaks were found as well for two
populations of F. indicus: 1133.58 (Figure 1h), 2301.21 and 2426.31 Da in the
specimens coming from Mozambique, while 1622.80, and 2619.97 in those from
Madagascar. The information provided by these specific peaks and the combination of
the presence or absence of other peaks in the different groups of these species (Table S-
2) is enough to clearly differentiate between the two superfamilies, the three families
and the seven species studied. Furthermore, intra-specific differences described for L.
ACS Paragon Plus Environment
Journal of Proteome Research
163

vannamei and F. indicus may be used to differentiate populations from these two
species.
MS/MS Characterization of Peptides from AK. Tryptic digests were further
analyzed by MS/MS using a combination of LC-ESI-IT and nESI-IT. The spectra
produced were used for database search and for manual de novo amino acid sequence
interpretation. Results of the sequencing efforts are shown in tables 2 and S-3; in the
most extended, presented in the supplementary information, a summary of the 131
peptide sequences obtained is presented, being table 2 a concise summary with the
masses and sequences used for shrimp species identification. Peaks with coincident
molecular weight were previously found by MALDI-TOF MS analysis for 54 peptides
and 35 were previously assigned by PMF.
Several peptides were found to be specific for each of the superfamilies. Those with
Mr 1147.60, 1050.52, 1667.82, 1683.82, 1796.94, 792.43, 2033.94, 1719.77, 1139.58,
and 1304.68 Da were specific to the superfamily Penaeoidea. Specific peptides were
found for each of the families included in this superfamily: peptides at 2599.26 and
1657.82 Da for the Penaeidae family, and peptides at 1397.76, 1403.67, 1718.80,
1442.80, 1130.59, 1286.70, 2772.40, 1016.55, 1789.97, and 1532.83 Da for the
Solenoceridae family.
Within the Penaeidae family, species-specific peptides were also found. Examples are
peptides at 3057.50, 3073.50, 3089.49, 3105.49, and 1984.93 Da for P. monodon;
1205.68 Da for F. merguiensis; 2639.15 Da for F. notialis.
As previously revealed using MALDI-TOF spectra, intra-species differences were
found for L. vannamei and F. indicus: peptides at 1531.83, 1659.92, 1201.58, 1513.82
and 1761.94 Da, were specific for L. vannamei from Costa Rica; 1383.78 and 3390.66
Da for L. vannamei from Argentina; 1215.60 and 1231.60 Da, for F. indicus from
ACS Paragon Plus Environment
Journal of Proteome Research
164

Mozambique; and 1063.57 Da, was specific for F. indicus from Madagascar. Another
pair of isobaric peptides, with a Mr of 1369.76 Da and sequences YLSKDIFDKLK and
YLSKEVFDQLK were found specific for L. vannamei from Costa Rica, and to F.
indicus from Mozambique, respectively.
Several examples of the fragmentation spectra from species-specific peptides,
together with the m/z of the precursor ions, are shown in Figures 2, 3, and 4. The peak
with m/z 525.59 (charge 2+) was chosen because it corresponds to one of the ten
peptides specific for the Penaeoidea superfamily (Figure 2). A pair of homologous
peptides serves to differentiate the Penaeidae and Solenoceridae families: the peak at
m/z 829.26 (2+), which is only present in Penaeidae, corresponds to the sequence
TFLVWVNEEDHLR (Figure 3a), whereas the peak at 860.09 (2+), only present in
Solenoceridae, corresponds to TFLC*WVNEEDHLR (Figure 3b). Both are absent in
the Pandalidae family. A substitution of Val for Cys in the sequence is responsible for
the difference in the mass of the precursors and for the presence of different
fragmentation diagnostic peaks at m/z 1011.3, 1197.4 (for the 829.26 precursor) and
1072.3, 1258.4 (for the 860.09 precursor).
Only one P. borealis specific peptide could be completely characterized by manual de
novo sequencing: SFLVWVNEEDQLR, with a Mr of 1634.81 Da. This peak was
chosen because it was only found in the Pandalidae family species (Figure 3c). This
sequence has two substitutions with respect to that present in the Penaeidae species: a
Thr for a Ser, and a His for a Gln.
The peak with a Mr of 1077.58 Da was found in P. monodon and F. merguiensis
samples, but it happened to be a mixture of two isobaric peptides, with specific
sequences for each of the species: AVFDQLKEK and ALFDQLKDK, respectively.
There are a Val and a Glu for P. monodon and a Leu and an Asp for F. merguiensis, in
ACS Paragon Plus Environment
Journal of Proteome Research
165

positions 2 and 8. Fragmentation spectra are shown in Figure 4; as expected, these two
substitutions are responsible for the different diagnostic fragmentation peaks at m/z
454.3, 760.3, 907.3 for P. monodon (Figure 4a) and at m/z 447.1, 746.1, 893.1 for F.
indicus (Figure 4b). As the MS/MS fragmentation patterns are different for each
sequence, these peptides can be used in a multiple reaction monitoring (MRM)
experiment to differentiate both shrimp species.
In addition to species-specific peptides, there are others which are always present in
several species, being always absent in others. Combining the presence or absence in
the samples of carefully selected peptides among those shown in Table 2, unequivocal
shrimp species differentiation is clearly possible. Figure 5 presents one of the possible
schematic flow diagrams which can be used for the systematic discrimination among
the seven species studied and even, among the different geographical populations for L.
vannamei and F. indicus.
The AK amino acid sequences from the different shrimp considered, as deduced from
MS/MS data, were aligned together with the sequence Pen m 2 from P. monodon,25
accessible in the protein databases, which presents the highest homology with those
found in the present work (Figure 6). As AKs are highly conserved, only small
differences are observed, but these differences may be used to distribute the species into
different groups.
Conclusions
Proteomics methodology was used to find sequence similarities and differences
among the arginine kinases from seven different shrimp species belonging to the order
Decapoda. A high degree of homology was found among the AK sequences, but the
sequence variations found are very useful to distinguish the different members of these
species, providing (i) a selective differentiation between the superfamilies Penaeoidea
ACS Paragon Plus Environment
Journal of Proteome Research
166

and Pandaloidea and between the families Penaeidae, Solenoceridae, and Pandalidae;
and (ii) an unequivocal identification of several shrimp species. Due to AK inter-
specific variability and the high concentration at which it is present in the muscle from
shrimp, we can forecast that they could be used as a good biomarker for species
identification. The identification and characterization of specific peptides, as done by
this work, is the first step toward the designing of fast and cheap detection analyses.
Identified species-specific peptides can be used to prepare antibody-based, easy-to-use
kits for the sensitive detection of each of the species.
Acknowledgement. We thank Lorena Barros and Eva María Rodríguez for their
excellent technical assistance. We also acknowledge members from CETMAR for their
helpful collaboration in the collection of specimens for this study. This work was
supported by the INIA National Food Program; Spanish Ministry for Education and
Science (Project CAL-03-030-C2-2) and by the PGIDIT Research Program in Marine
Resources (Project PGIDIT04RMA261004PR), Xunta de Galicia.
Supporting Information Available. Results and Discussion, including Figure S-1,
and Tables S-1, S-2 and S-3. This material is available free at http://pubs.acs.org.
Conflict of interest statement All authors declare there are no financial/commercial
conflicts of interest.
ACS Paragon Plus Environment
Journal of Proteome Research
167

Figure captions
Figure 1. Regions of the MALDI-TOF MS peptide maps from AK trypsin digested
spots, showing peaks specific to (a) superfamily Penaeoidea (1050.51 Da); (b, c, d)
family Penaeidae (1008.49, 1259.63, and 1657.85 Da), family Solenoceridae (994.45,
1263.62, and 1286.63 Da) or family Pandalidae (1634.82 Da); (e) P. monodon (1077.54
Da); (f) F. notialis (1463.75 Da); (g) L. vannamei from Costa Rica (1531.75 Da); (h) F.
indicus from Mozambique (1133.58 Da). Populations from: (1) Costa Rica; (2)
Argentina; (3) Mozambique and (4) Madagascar.
Figure 2. MS/MS spectrum of the AK peptide FLQAANAC*R. This peptide is present
in all the species belonging to the superfamily Penaeoidea. C* carboxymethylated
cysteine.
Figure 3. MS/MS spectra of the AK differential peptides: (a) TFLVWVNEEDHLR, (b)
TFLVWVC*NEEDHLR, and (c) SFLVWVNEEDQLR, which are present in the
Penaeidae, Solenoceridae and Pandalidae families, respectively. C* carboxymethylated
cysteine.
Figure 4. MS/MS spectra of the isobaric peptides (a) AVFDQLKEK and (b)
ALFDQLKDK, which are present in AK from P. monodon and F. merguiensis,
respectively.
Figure 5. Flow diagram showing the systematic identification of shrimp species using
nine differential peptides from AK. m/z of the doubly charged ions were: 525.6, 817.8,
829.3, 759.8, 675.2, 539.2, 766.2, 603.3, and 616.1. “Y” denotes the presence and “N”
the absence of a particular peptide.
Figure 6. Alignment of the AK amino acid sequences, as obtained by MS/MS analysis,
for all the shrimp species studied. The sequences were aligned with the allergen Pen m
2 sequence from P. monodon (accession number gi|27463265), which presents the
ACS Paragon Plus Environment
Journal of Proteome Research
168

highest degree of homology with them. (�) not determined amino acid. (1) Costa Rica;
(2) Argentina; (3) Mozambique and (4) Madagascar populations.
ACS Paragon Plus Environment
Journal of Proteome Research
169
Table 1. Penaeid shrimp species considered in the study
Scientific namea Commercial name Originb
Penaeus monodon Giant tiger prawn IWP and WI
Litopenaeus vannamei Pacific white shrimp EP
Fenneropenaeus indicus Indian white prawn WI
Fenneropenaeus merguiensis Banana prawn WCP
Farfantepenaeus notialis Southern pink shrimp EA
Pleoticus muelleri Argentine red shrimp SWA
Pandalus borealils Northern shrimp NA
aThe taxonomic classification proposed by Pérez-Farfante and Kensley [2]) was adopted.bOrigin
abbreviations: IWP, Indo-West Pacific Ocean; WI, Western Indian Ocean; EP, Eastern Pacific
Ocean; WCP, Western Central Pacific Ocean; EA, Eastern Atlantic Ocean; SWA, Southern
West Atlantic Ocean.; NA, Northern Atlantic Ocean.
ACS Paragon Plus Environment
Journal of Proteome Research
170

Tab
le 2
. Con
cise
sum
mar
y of
pep
tide
sequ
ence
dat
a ob
tain
ed b
y M
S/M
S fo
r A
Ks
from
the
shri
mp
spec
ies
stud
ied
usef
ul f
or s
peci
es
iden
tific
atio
n.
m/z
(z)
ob
serv
ed
(M+
H)+
mat
ched
P
eptid
e S
eque
nce
Mod
ifica
tion
Pos
ition
M
ALD
I ob
serv
ed
SO
PP
RA
G
IT
PN
V
(1)
PN
V
(2)
PN
I (3
) P
NI
(4)
PB
A
LAA
752.
30 (
2+)
1503
.798
A
DA
AV
IEK
LEA
GF
K
N-A
cyl
2-15
Y
(P
MF
)
766.
22 (
2+)
1531
.83
VD
AA
VIE
KLE
AG
FK
N
-Acy
l 2-
15
Y
660.
20 (
2+)
1319
.69
VS
ST
LSG
LEG
ELK
152-
164
675.
22 (
2+)
1349
.71
VS
ST
LSS
LEG
ELK
152-
164
685.
20 (
2+);
457
.07
(3+
); 3
43.2
3 (4
+)
1369
.76
YLS
KD
IFD
KLK
30-4
0 Y
(P
MF
)
685.
25 (
2+);
457
.14
(3+
) 13
69.7
3 Y
LSK
EV
FD
QLK
30-4
0 Y
692.
33 (
2+)
1383
.78
YLT
KE
VF
DK
LK
30
-40
699.
20 (
2+);
466
.43
(3+
) 13
97.7
6 Y
LTK
EIF
DQ
LK
30
-40
532.
18 (
2+)
1063
.57
AV
FD
QLK
DK
34-4
2
539.
16(2
+)
1077
.58
AV
FD
QLK
EK
34-4
2 Y
(P
MF
)
539.
14 (
2+)
1077
.58
ALF
DQ
LKD
K
34
-42
Y
603.
26 (
2+)
1205
.68
ALF
DQ
LKD
KK
34-4
3 Y
601.
18 (
2+)
1217
.59
GT
YF
PLT
GM
SK
M
SO
16
5-17
5
616.
09 (
2+)
1231
.60
GT
YF
PLT
GM
TK
M
SO
16
5-17
5
617.
11 (
2+)
1233
.58
GT
YY
PLT
GM
SK
M
SO
16
5-17
5
525.
59 (
2+)
1050
.52
FLQ
AA
NA
CR
C
ysC
AM
19
4-20
2 Y
(P
MF
)
817.
76 (
2+)
1634
.81
SF
LVW
VN
EE
DQ
LR
21
7-22
9 Y
829.
26 (
2+);
553
.14
(3+
) 16
57.8
2 T
FLV
WV
NE
ED
HLR
217-
229
Y (
PM
F)
860.
09 (
2+)/
573
.71
(3+
) 17
18.8
0 T
FLV
WC
NE
ED
HLR
C
ysC
AM
21
7-22
9 Y
(P
MF
)
759.
85 (
2+);
506
.73
(3+
) 15
18.8
2 LE
EV
AG
KY
NLQ
VR
297-
309
Y (
PM
F)
766.
80 (
2+)
1532
.83
LEE
IAG
KY
NLQ
VR
297-
309
Y
m/z
: mas
s/ch
arge
. MA
LD
I ob
serv
ed; (
Y):
pep
tide
foun
d by
MA
LD
I-T
OF
MS;
(P
MF)
: pep
tide
mat
ched
by
pept
ide
mas
s fi
nger
prin
t wit
h an
AK
ent
ry in
pro
tein
data
base
s. (�)
den
otes
the
pres
ence
of
a pe
ptid
e, a
nd (�
) th
e ab
senc
e. C
ysC
AM
, car
boxy
met
hyla
ted
cyst
eine
; MSO
, met
hion
ine
sufo
xide
; N-A
cyl,
acet
yl N
-
term
inal
. SO
P, F
. not
iali
s; P
RA
, Pan
dalu
s bo
real
is, G
IT, P
enae
us m
onod
on; P
NV
, Lit
open
aeus
van
nam
ei; P
NI,
Fen
nero
pena
eus
indi
cus;
PB
A, F
enne
rope
naeu
s
mer
guie
nsis
; LA
A, P
leot
icus
mue
ller
i. (1
) C
osta
Ric
a po
pula
tion;
(2)
Arg
enti
na p
opul
atio
n; (
3) M
ozam
biqu
e po
pula
tion;
(4)
Mad
agas
car
popu
latio
n.
AC
S P
arag
on
Plu
s E
nvi
ron
men
t
Jou
rnal
of
Pro
teo
me
Res
earc
h
171

References
(1) Holthius, L. B., FAO Species Catalogue: Shrimps and Prawns of the World. An
Annotated Catalogue of Species of Interest to Fisheries; FAO Fish. Synop. (125);
FAO: Rome, Italy, 1980; Vol. 1, pp 1-271.
(2) Pérez-Farfante, I.; Kensley, B., Penaeoid and Sergestoid Shrimps and Prawns of
the World. Keys and Diagnoses for the Families and Genera. Éditions du Muséum
National d’Histoire Naturelle: Paris, France, 1997.
(3) Council Regulation (EC) No. 104/2000 of the European Parliament of Dec. 17,
1999 on the common organization of the markets in fishery and aquaculture
products.
(4) Royal Decree 121/2004 of Jan. Twenty-three (BOE No. 31, Feb. 5, 2004) from
Ministry of Agriculture, Fisheries and Food, Spain.
(5) Civera, T., Species identification and safety of fish products. Vet. Res. Commun.
2003, 27, 481-489.
(6) Maggioni, R.; Rogers, A. D.; Maclean, N.; Incao, F., Molecular phylogeny of
Western Atlantic Farfantepenaeus and Litopenaeus shrimp based on
mitochondrial 16S partial sequences. Mol. Phylogenet. Evol. 2001, 18, 66-73.
(7) Baldwin, J. D.; Bass, A. L.; Bowen, B. W.; Clark, W. H., Molecular phylogeny
and biogeography of the marine shrimp Penaeus. Mol. Phylogenet. Evol. 1998, 10,
399-407.
(8) Quan, J.; Zhuang, Z.; Deng, J.; Dai, J.; Zhang, Y. -p., Phylogenetic relationships
of 12 Penaeoidea shrimp species deduced from mitochondrial DNA sequences.
Biochem. Genet. 2004, 42, 331-345.
(9) Lavery, S.; Chan, T. Y.; Tam, Y. K.; Chu, K. H., Phylogenetic relationships and
evolutionary history of the shrimp genus Penaeus s.l. derived from mitochondrial
DNA. Mol. Phylogenet. Evol. 2004, 31, 39-49.
(10) Voloch, C. M.; Freire, P. R.; Russo, C. A. M., Molecular phylogeny of penaeid
shrimps inferred from two mitochondrial markers. Genet. Mol. Res. 2005, 4, 668-
674.
(11) Brzezinski, J. L., Detection of crustacean DNA and species identification using a
PCR-restriction fragment length polymorphism method. J. Food Prot. 2005, 68,
1866-1873.
(12) Pascoal, A.; Barros-Velázquez, J.; Cepeda, A.; Gallardo, J. M.; Calo-Mata, P., A
polymerase chain reaction-restriction fragment length polymorphism method
ACS Paragon Plus Environment
Journal of Proteome Research
172

based on the analysis of a 16S rRNA/tRNAVal mitochondrial region for species
identification of commercial penaeid shrimps (Crustacea: Decapoda:
Penaeoidea) of food interest. Electrophoresis 2008, 29, 499-509.
(13) Piñeiro, C.; Barros-Velázquez, J.; Vázquez, J.; Figueras, A.; Gallardo, M.,
Proteomics as a tool for the investigation of seafood and other marine products. J.
Proteome Res. 2003, 2, 127-135.
(14) Martinez, I.; Friis, T. J., Application of proteome analysis to seafood
authentication. Proteomics 2004, 4, 347-354.
(15) López, J. L.; Marina, A.; Álvarez, G.; Vázquez, J., Application of proteomics for
fast identification of species-specific peptides from marine species. Proteomics
2002, 2, 1658-1665.
(16) Carrera, M.; Cañas, B.; Piñeiro, C.; Vázquez, J.; Gallardo, J. M., Identification of
commercial hake and grenadier species by proteomic analysis of the parvalbumin
fraction. Proteomics 2006, 6, 5278-5287.
(17) Carrera, M.; Canas, B.; Pineiro, C.; Vazquez, J.; Gallardo, J. M., De novo mass
spectrometry sequencing and characterization of species-specific peptides from
nucleoside diphosphate kinase B for the classification of commercial fish species
belonging to the family Merlucciidae. J. Proteome Res. 2007, 6, 3070-3080.
(18) An, H.; Marshall, M. R.; Otwell, W. S.; Wei, C. I., Species identification of raw
and boiled shrimp by a urea gel isoelectric focusing technique. J. Food Sci. 1989,
54, 233-236.
(19) Civera, T.; Parisi, E., The use of sodium dodecyl sulfate polyacrylamide gel
electrophoresis for the identification of raw and cooked shrimps and crabs. Ital. J.
Food Sci. 1991, 3, 149-157.
(20) Shanti, K. N.; Martin, B. M.; Nagpal, S.; Metcalfe, D. D.; Rao, P. V.,
Identification of tropomyosin as the major shrimp allergen and characterization of
its IgE-binding epitopes. J. Immunol. 1993, 151, 5354-5363.
(21) Ortea, I.; Cañas, B.; Calo-Mata, P.; Barros-Velázquez, J.; Gallardo, J. M.,
Arginine kinase peptide mass fingerprinting as a proteomic approach for species
identification and taxonomic analysis of commercially relevant shrimp species. J.
Agric. Food Chem. 2009, 57, 5665-5672.
(22) Morrison, J. F., Arginine kinase and other invertebrate guanidine kinases. In The
Enzymes, 3rd ed.; Boyer, P. D., Ed.; Academic Press: New York, 1973; Vol. 8, pp
457-486.
ACS Paragon Plus Environment
Journal of Proteome Research
173

(23) Watts, D. C., Creatine kinase (adenosine 5'-triphosphate creatine
phosphotransferase. In The Enzymes, 3rd ed.; Boyer, P. D., Ed.; Academic Press:
New York, 1973; Vol. 8, pp 383-455.
(24) France, R. M.; Sellers, D. S.; Grossman, S. H., Purification, characterization, and
hydrodynamic properties of arginine kinase from Gulf shrimp (Penaeus aztecus).
Arch. Biochem. Biophys. 1997, 345, 73-78.
(25) Yu, C.-J.; Lin, Y.-F.; Chiang, B.-L.; Chow, L.-P., Proteomics and immunological
analysis of a novel shrimp allergen, Pen m 2. J. Immunol. 2003, 170, 445-453.
(26) Piñeiro, C.; Barros-Velázquez, J.; Sotelo, C. G.; Pérez-Martín, R. I.; Gallardo, J.
M., Two-dimensional electrophoretic study of the water-soluble protein fraction in
white muscle of gadoid fish species. J. Agric. Food Chem. 1998, 46, 3991-3997.
(27) Jensen, O. N.; Wilm, M.; Shevchenko, A.; Mann, M., Sample preparation methods
for mass spectrometric peptide mapping directly from 2-DE gels. In 2-D Proteome
Analysis Protocols, Link, A. J., Ed.; Humana Press: Totowa, NJ, 1999; pp 513-
530.
Synopsis TOC.
The identification of commercial shrimp species is a relevant issue to ensure correct labelling, maintain consumer confidence and enhance the knowledge of the captured species, benefiting both, fisheries and manufacturers. A proteomic approach, based on 2DE, tryptic in-gel digestion, MALDI-TOF MS, and ESI-MS/MS analyses, paying special attention to de novo sequence interpretation of differential species-specific peptides, is proposed for the identification of shrimp species with commercial interest.
ACS Paragon Plus Environment
Journal of Proteome Research
174

����������� ����*"��(+2�������������������������������������������,�� �3�������4�����������5�6�� ���������������������57$8$.87�"�6-�5),�,��6������������������57$$9.:%,�7#8%.&',�����7&8;.98��"�6,���������������������5%%:.:8,�7#&'.&#,�����7#9&.&'�"�6����������������������
57&':.9#�"�6-�5�6��.���������57$;;.8:�"�6-�5�6�2.���������57:&'.;8�"�6-�5�6�*.������������������������578'7.;8�"�6-�5 6�2.����� ������������)�< ��577''.89�"�6.���� �����������!�576�
��������-�5#6���������-�5'6������)�< ������5:6����������.��#8:=7%$���5%&�=�%&�"��6��
ACS Paragon Plus Environment
Journal of Proteome Research
175

��
�
�
��>������� ����� ������������2*?��@��A�.�( ��������������������������� ���������)����������� ��� ��������������������.��A���)�=��� ������������.��
#8:=7%$���5%&�=�%&�"��6����
ACS Paragon Plus Environment
Journal of Proteome Research
176
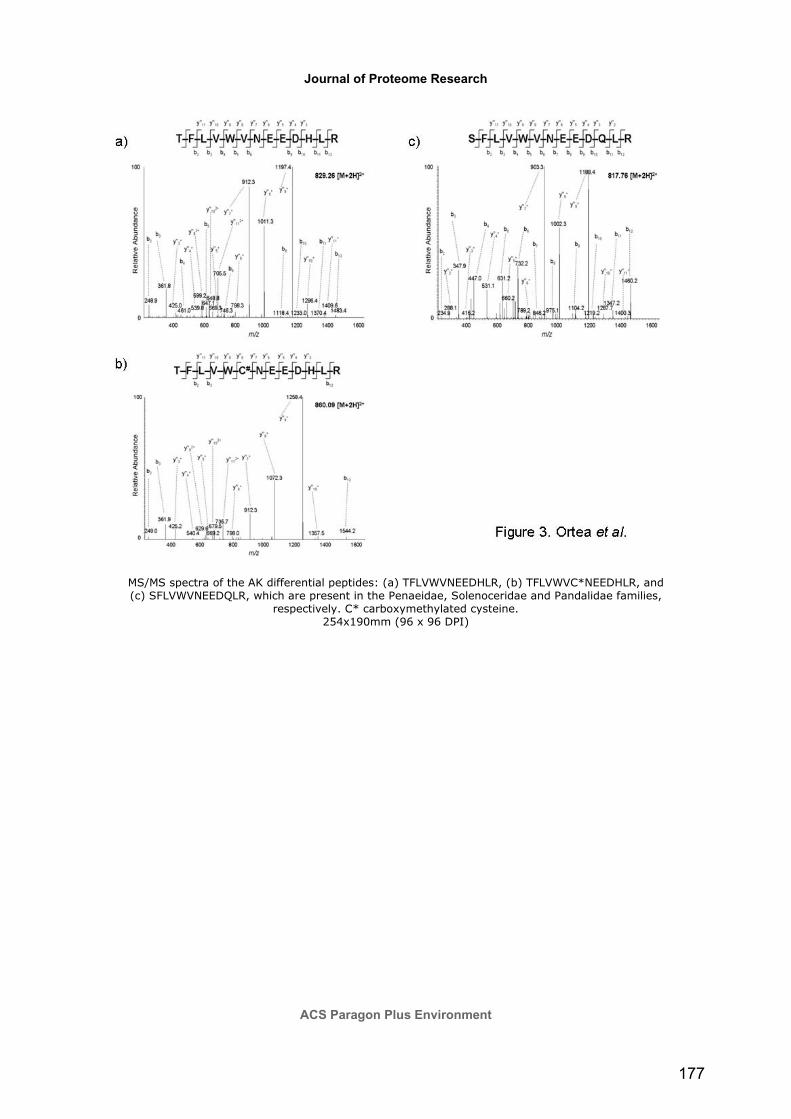
��>������������ ������������������������!�5�6�(2*BCB@DD"E*�,�5)6�(2*BCB�A@DD"E*�,�����56��2*BCB@DD"?*�,�3 � ��������������� �����������,�������������������������������������,�
����������.��A���)�=��� ������������.��#8:=7%$���5%&�=�%&�"��6��
ACS Paragon Plus Environment
Journal of Proteome Research
177
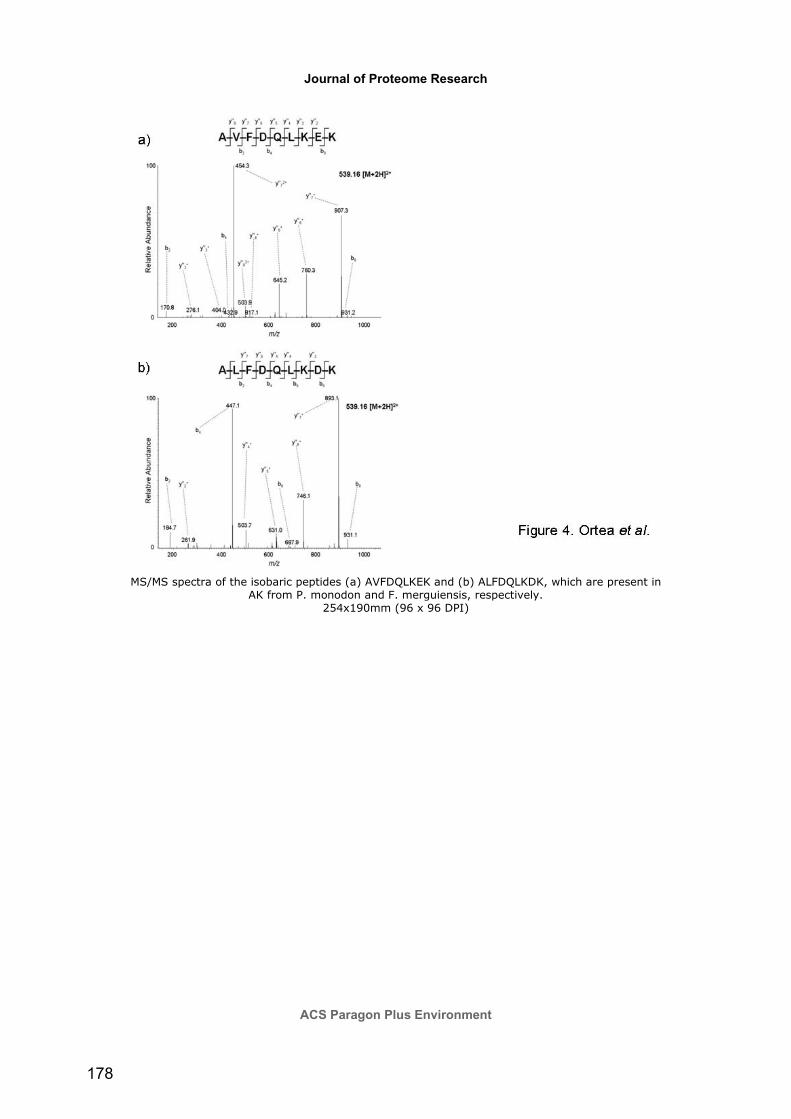
��>������������ �����)������������5�6��B2"?*�D������5)6��*2"?*�"�,�3 � ������������������������.�������������2.����� ������,�����������.��
#8:=7%$���5%&�=�%&�"��6��
ACS Paragon Plus Environment
Journal of Proteome Research
178

2��3���������� �3���� �������������������������� ������������ �������������������������������������.��>����� ���� )��� �����������3���!�8#8.&,�97;.9,�9#%.',�;8%.9,�&;8.#,�8'%.#,�;&&.#,�
&$'.',�����&7&.7.�FGH�������� ��������������F@H� ���)�������������� ����������.��#8:=7%$���5%&�=�%&�"��6��
ACS Paragon Plus Environment
Journal of Proteome Research
179

������������ �����������������< ����,�����)������)����>�����������,��������� ��� ������������� ����.�( ����< �����3������������3� � �����������������#���< ����������.���������5��������� �)�����I#;:&'#&86,�3 � ��������� �� �� ������������� ��������3� � ��.�5J6�����������������������.�576���������-�5#6���������-�5'6������)�< ������5:6�����������
��� ������.��#8:=7%$���5%&�=�%&�"��6��
ACS Paragon Plus Environment
Journal of Proteome Research
180

1
SUPPORTING INFORMATION: RESULTS AND DISCUSSION
2-DE
Figure S-1. Cropped regions of representative 2-DE gels
showing the separated arginine kinase from all shrimps studied.
Pen, Penaeus; Far, Farfantepenaeus, Lit, Litopenaeus; Fen,
Fenneropenaeus; Ple, Pleoticus; Pan, Pandalus.
Table S-1. Mean pI of the analyzed spot, calculated from the 2-DE patterns of all
individuals analysed of each species.
species pI ± RSD Penaeus monodon 4.99 ± 0.6 Farfantepenaeus notialis 5.22 ± 0.6 Litopenaeus vannamei 4.98 ± 0.8 Fenneropenaeus indicus 5.18 ± 0.3 Fenneropenaeus merguiensis 5.16 ± 1.0 Pleoticus muelleri 5.25 ± 1.1 Pandalus borealis 4.68 ± 1.6
MALDI-TOF MS analysis
181

2
Tab
le S
-2. C
ompl
ete
info
rmat
ion
of th
e M
AL
DI-
TO
F M
S a
naly
sis
of a
rgin
ine
kina
se s
pots
in th
e se
ven
shri
mp
spec
ies
cons
ider
ed.
exp
ma
ss
exp
mr
ca
lc m
r d
elta
p
ep
tid
e
mo
dific
atio
n
Org
anis
m
Po
sitio
n
SO
P
PR
A
GIT
P
NV
P
NI
PB
A
LA
A
99
4.4
52
3
99
3.4
45
9
93
.47
81
-0
.03
V
PF
SH
HD
R
G
alap
agan
us c
onw
ayen
sis
18
9-1
96
10
08
.49
1
00
7.4
82
1
00
7.4
94
-0
.01
IP
FS
HH
DR
Pen
aeus
mon
odon
2
57
-26
4
10
50
.50
6
10
49
.49
9
10
49
.50
8
-0.0
1
FL
QA
AN
AC
R
CysC
AM
P
enae
us m
onod
on
19
4-2
02
10
77
.54
4
10
76
.53
7
10
76
.58
7
-0.0
5
AV
FD
QL
KE
K
P
enae
us m
onod
on
34
-42
11
33
.57
6
(3
)
11
41
.56
1
11
47
.59
7
11
46
.58
9
11
46
.60
7
-0.0
2
LID
DH
FL
FK
Pen
aeus
mon
odon
1
81
-18
9
11
48
.53
4
(1)
11
49
.56
8
11
48
.56
1
11
48
.61
9
-0.0
6
EIF
DQ
LK
TR
Sol
enop
sis
invi
cta
33
-41
(1)
11
58
.58
3
11
64
.65
1
11
63
.64
4
11
63
.59
5
0.0
4
RIP
FS
HH
DR
Lito
pena
eus
vann
amei
2
56
-26
4
11
96
.63
4
12
05
.66
7
12
04
.66
1
20
4.6
82
-0
.02
A
VF
DQ
LK
EK
K
P
enae
us m
onod
on
34
-43
12
59
.63
1
25
8.6
23
1
25
8.6
88
-0
.07
L
TS
AV
NE
IEK
R
P
enae
us m
onod
on
24
6-2
56
12
63
.61
8
12
86
.63
4
13
69
.71
1
36
8.7
02
1
36
8.7
65
-0
.06
Y
LS
KD
IFD
KL
K
M
arsu
pena
eus
japo
nicu
s 2
9-3
9
(1
)
14
15
.77
7
14
14
.76
9
14
14
.78
9
-0.0
2
RL
TS
AV
NE
IEK
R
P
enae
us m
onod
on
24
5-2
56
14
24
.67
9
14
38
.70
8
14
37
.70
1
14
37
.74
-0
.04
R
RM
GL
TE
YD
AV
K
P
erip
lane
ta a
mer
ican
a 3
29
-34
0
14
42
.72
7
14
41
.71
9
14
41
.76
4
-0.0
4
LK
GT
FY
PL
TG
MS
K
C
imbe
ris p
ilosa
1
13
-12
5
14
63
.75
1
14
62
.74
3
14
62
.83
7
-0.0
9
RL
VH
AV
NE
IEK
R
A
pis
mel
lifer
a 2
44
-25
5
15
03
.71
5
15
02
.70
8
15
02
.79
8
-0.0
9
AD
AA
VIE
KL
EA
GF
K
N-A
cyl
Pen
aeus
mon
odon
2
-15
(2)
15
07
.73
1
15
06
.72
3
15
06
.7
0.0
2
EG
DR
FL
QA
AN
AC
R
CysC
AM
P
enae
us m
onod
on
19
0-2
02
15
13
.78
1
15
12
.77
4
15
12
.82
6
-0.0
5
LA
AN
RD
KL
EE
VA
GK
Mar
supe
naeu
s ja
poni
cus
28
9-3
02
(1)
15
18
.73
2
15
17
.72
4
15
17
.82
-0
.10
L
EE
VA
GK
YN
LQ
VR
Pen
aeus
mon
odon
2
97
-30
9
15
25
.77
9
15
24
.77
2
15
24
.85
5
-0.0
8
KY
LT
KE
IFD
QL
K
S
olen
opsi
s in
vict
a 2
8-3
9
15
31
.75
3
(1)
15
32
.78
2
15
31
.77
4
15
31
.83
6
-0.0
6
LE
EV
AA
KY
NL
QV
R
G
alap
agan
us c
onw
ayen
sis
22
9-2
41
15
41
.89
1
15
40
.88
4
15
40
.85
7
0.0
3
LA
AN
RE
KL
EE
VA
AK
Hom
arus
gam
mar
us
29
0-3
03
16
04
.81
4
16
03
.80
6
16
03
.8
0.0
1
LID
DH
FL
FK
EG
DR
Pen
aeus
mon
odon
1
81
-19
3
16
06
.77
6
16
22
.79
9
16
21
.79
2
16
21
.85
-0
.06
L
QA
GF
KK
LE
AA
TD
CK
Mar
supe
naeu
s ja
poni
cus
9-2
3
(4)
16
31
.89
9
16
30
.89
2
16
30
.89
3
-0.0
1
AD
AA
VIE
KL
EA
GF
KK
N
-Acyl
Pen
aeus
mon
odon
2
-16
(2)
16
34
.82
4
16
38
.82
6
16
51
.83
9
16
50
.83
2
16
50
.82
2
0.0
1
IIS
MQ
MG
GD
LG
QV
FR
Pen
aeus
mon
odon
2
30
-24
4
16
57
.85
1
16
56
.84
3
16
56
.82
6
0.0
2
TF
LV
WV
NE
ED
HL
R
P
enae
us m
onod
on
21
7-2
29
16
66
.81
5
16
67
.81
1
66
6.8
02
1
66
6.8
17
-0
.01
II
SM
QM
GG
DL
GQ
VF
R
MS
O
Lito
pena
eus
vann
amei
2
30
-24
4
(1
)
16
83
.81
6
16
82
.80
9
16
82
.81
2
-0.0
1
IIS
MQ
MG
GD
LG
QV
FR
2
MS
O
Pen
aeus
mon
odon
2
30
-24
4
16
90
.87
2
182
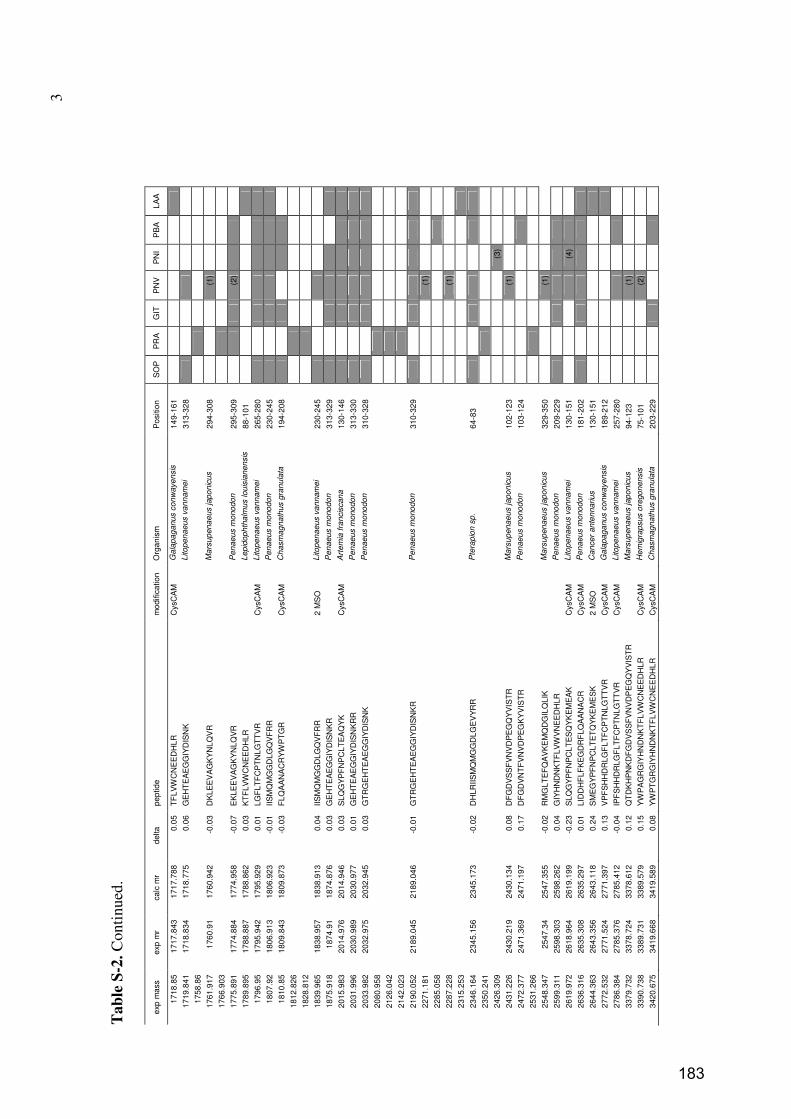
3
Tab
le S
-2. C
onti
nued
.
exp
ma
ss
exp
mr
ca
lc m
r d
elta
p
ep
tid
e
mo
dific
atio
n
Org
anis
m
Po
sitio
n
SO
P
PR
A
GIT
P
NV
P
NI
PB
A
LA
A
17
18
.85
1
71
7.8
43
1
71
7.7
88
0
.05
T
FL
VW
CN
EE
DH
LR
C
ysC
AM
G
alap
agan
us c
onw
ayen
sis
14
9-1
61
17
19
.84
1
17
18
.83
4
17
18
.77
5
0.0
6
GE
HT
EA
EG
GIY
DIS
NK
Lito
pena
eus
vann
amei
3
13
-32
8
17
58
.86
17
61
.91
7
17
60
.91
1
76
0.9
42
-0
.03
D
KL
EE
VA
GK
YN
LQ
VR
Mar
supe
naeu
s ja
poni
cus
29
4-3
08
(1)
17
66
.90
3
17
75
.89
1
17
74
.88
4
17
74
.95
8
-0.0
7
EK
LE
EV
AG
KY
NL
QV
R
P
enae
us m
onod
on
29
5-3
09
(2)
17
89
.89
5
17
88
.88
7
17
88
.86
2
0.0
3
KT
FL
VW
CN
EE
DH
LR
Lepi
doph
thal
mus
loui
sian
ensi
s 8
8-1
01
17
96
.95
1
79
5.9
42
1
79
5.9
29
0
.01
L
GF
LT
FC
PT
NL
GT
TV
R
CysC
AM
Li
tope
naeu
s va
nnam
ei
26
5-2
80
18
07
.92
1
80
6.9
13
1
80
6.9
23
-0
.01
II
SM
QM
GG
DL
GQ
VF
RR
Pen
aeus
mon
odon
2
30
-24
5
18
10
.85
1
80
9.8
43
1
80
9.8
73
-0
.03
F
LQ
AA
NA
CR
YW
PT
GR
C
ysC
AM
C
hasm
agna
thus
gra
nula
ta
19
4-2
08
18
12
.82
6
18
28
.81
2
18
39
.96
5
18
38
.95
7
18
38
.91
3
0.0
4
IIS
MQ
MG
GD
LG
QV
FR
R
2 M
SO
Li
tope
naeu
s va
nnam
ei
23
0-2
45
18
75
.91
8
18
74
.91
1
87
4.8
76
0
.03
G
EH
TE
AE
GG
IYD
ISN
KR
Pen
aeus
mon
odon
3
13
-32
9
20
15
.98
3
20
14
.97
6
20
14
.94
6
0.0
3
SL
QG
YP
FN
PC
LT
EA
QY
K
CysC
AM
A
rtem
ia fr
anci
scan
a 1
30
-14
6
20
31
.99
6
20
30
.98
9
20
30
.97
7
0.0
1
GE
HT
EA
EG
GIY
DIS
NK
RR
Pen
aeus
mon
odon
3
13
-33
0
20
33
.98
2
20
32
.97
5
20
32
.94
5
0.0
3
GT
RG
EH
TE
AE
GG
IYD
ISN
K
P
enae
us m
onod
on
31
0-3
28
20
80
.95
8
21
26
.04
2
21
42
.02
3
21
90
.05
2
21
89
.04
5
21
89
.04
6
-0.0
1
GT
RG
EH
TE
AE
GG
IYD
ISN
KR
Pen
aeus
mon
odon
3
10
-32
9
22
71
.18
1
(1)
22
85
.05
8
22
87
.22
8
(1)
23
15
.25
3
23
46
.16
4
23
45
.15
6
23
45
.17
3
-0.0
2
DH
LR
IIS
MQ
MG
GD
LG
EV
YR
R
P
tera
pion
sp.
6
4-8
3
23
50
.24
1
24
26
.30
9
(3
)
24
31
.22
6
24
30
.21
9
24
30
.13
4
0.0
8
DF
GD
VS
SF
VN
VD
PE
GQ
YV
IST
R
M
arsu
pena
eus
japo
nicu
s 1
02
-12
3
(1
)
24
72
.37
7
24
71
.36
9
24
71
.19
7
0.1
7
DF
GD
VN
TF
VN
VD
PE
GK
YV
IST
R
P
enae
us m
onod
on
10
3-1
24
25
31
.26
6
25
48
.34
7
25
47
.34
2
54
7.3
55
-0
.02
R
MG
LT
EF
QA
VK
EM
QD
GIL
QL
IK
M
arsu
pena
eus
japo
nicu
s 3
29
-35
0
(1
)
25
99
.31
1
25
98
.30
3
25
98
.26
2
0.0
4
GIY
HN
DN
KT
FL
VW
VN
EE
DH
LR
Pen
aeus
mon
odon
2
09
-22
9
26
19
.97
2
26
18
.96
4
26
19
.19
9
-0.2
3
SL
QG
YP
FN
PC
LT
ES
QY
KE
ME
AK
C
ysC
AM
Li
tope
naeu
s va
nnam
ei
13
0-1
51
(4
)
26
36
.31
6
26
35
.30
8
26
35
.29
7
0.0
1
LID
DH
FL
FK
EG
DR
FL
QA
AN
AC
R
CysC
AM
P
enae
us m
onod
on
18
1-2
02
26
44
.36
3
26
43
.35
6
26
43
.11
8
0.2
4
SM
EG
YP
FN
PC
LT
ET
QY
KE
ME
SK
2
MS
O
Can
cer
ante
nnar
ius
13
0-1
51
27
72
.53
2
27
71
.52
4
27
71
.39
7
0.1
3
VP
FS
HH
DR
LG
FL
TF
CP
TN
LG
TT
VR
C
ysC
AM
G
alap
agan
us c
onw
ayen
sis
18
9-2
12
27
86
.38
4
27
85
.37
6
27
85
.41
2
-0.0
4
IPF
SH
HD
RL
GF
LT
FC
PT
NL
GT
TV
R
CysC
AM
Li
tope
naeu
s va
nnam
ei
25
7-2
80
33
79
.73
2
33
78
.72
4
33
78
.61
2
0.1
2
QT
DK
HP
NK
DF
GD
VS
SF
VN
VD
PE
GQ
YV
IST
R
M
arsu
pena
eus
japo
nicu
s 9
4-1
23
(1)
33
90
.73
8
33
89
.73
1
33
89
.57
9
0.1
5
YW
PA
GR
GIY
HN
DN
KT
FL
VW
CN
EE
DH
LR
C
ysC
AM
H
emig
raps
us o
rego
nens
is
75
-10
1
(2
)
34
20
.67
5
34
19
.66
8
34
19
.58
9
0.0
8
YW
PT
GR
GIY
HN
DN
KT
FL
VW
CN
EE
DH
LR
C
ysC
AM
C
hasm
agna
thus
gra
nula
ta
20
3-2
29
183

4
exp
mas
s, o
bser
ved
prot
onat
ed m
onoi
soto
pic
mas
ses
in D
a; e
xp M
r, o
bser
ved
neut
ral m
asse
s in
Da;
cal
c M
r, c
alcu
late
d ne
utra
l mas
ses
in D
a; d
elta
, exp
Mr-
calc
Mr;
; (■
) de
note
s th
e pr
esen
ce o
f a
peak
, an
d (�
) th
e ab
senc
e. C
ysC
AM
, ca
rbox
ymet
hyla
ted
cyst
eine
; M
SO
, m
ethi
onin
e su
foxi
de;
N-A
cyl,
acet
yl N
-
term
inal
. SO
P,
F.
noti
alis
; P
RA
, P
anda
lus
bore
alis
, G
IT,
Pen
aeus
m
onod
on;
PN
V,
Lit
open
aeus
va
nnam
ei;
PN
I,
Fen
nero
pena
eus
indi
cus;
P
BA
,
Fen
nero
pena
eus
mer
guie
nsis
; L
AA
, Ple
otic
us m
uell
eri.
(1)
Cos
ta R
ica
popu
latio
n; (
2) A
rgen
tina
pop
ulat
ion;
(3)
Moz
ambi
que
popu
latio
n; (
4)
Mad
agas
car
popu
latio
n.
MS/
MS
Cha
ract
eriz
atio
n of
Pep
tide
s fr
om A
K.
184

5
Tab
le S
-3. C
ompl
ete
info
rmat
ion
of p
epti
de s
eque
nce
data
obt
aine
d by
MS
/MS
for
AK
s fr
om th
e sh
rim
p sp
ecie
s st
udie
d
m/z
(z)
ob
se
rve
d
(M+
H)+
m
atc
he
d
Pe
ptid
e S
eq
ue
nce
M
od
ific
atio
n
Po
sitio
n
MA
LD
I o
bse
rve
d
SO
P
PR
A
GIT
P
NV
(1
) P
NV
(2
) P
NI
(3)
PN
I (4
) P
BA
L
AA
54
6.0
8 (
3+
) 1
63
6.8
6
MV
DA
AV
LE
KL
EA
GF
K
MS
O
1-1
5
42
9.4
9 (
2+
) 8
58
.45
A
DA
AV
IEK
N
-Acyl
2-9
44
3.3
8 (
2+
) 8
86
.48
V
DA
AV
IEK
N
-Acyl
2-9
75
2.3
0 (
2+
) 1
50
3.7
98
A
DA
AV
IEK
LE
AG
FK
N
-Acyl
2-1
5
Y (
PM
F)
76
6.2
2 (
2+
) 1
53
1.8
3
VD
AA
VIE
KL
EA
GF
K
N-A
cyl
2-1
5
Y
81
6.3
7 (
2+
); 5
44
.49
(3
+)
16
31
.89
A
DA
AV
IEK
LE
AG
FK
K
N-A
cyl
2-1
6
Y (
PM
F)
83
0.3
9 (
2+
) 1
65
9.9
2
VD
AA
VIE
KL
EA
GF
KK
N
-Acyl
2-1
6
84
0.7
0 (
3+
) 2
52
0.2
9
AD
AA
VIE
KL
EA
GF
KK
LE
AA
TD
CK
N
-Acyl;C
ysC
AM
2
-24
84
0.7
2 (
2+
) 1
68
0.8
6
LE
AG
FK
KL
EA
AT
DC
K
CysC
AM
1
0-2
4
51
8.1
0 (
2+
) 1
03
5.5
1
KL
EA
AT
DC
K
CysC
AM
1
6-2
4
68
5.2
0 (
2+
); 4
57
.07
(3
+);
34
3.2
3 (
4+
) 1
36
9.7
6
YL
SK
DIF
DK
LK
30
-40
Y
(P
MF
)
68
5.2
5 (
2+
); 4
57
.14
(3
+)
13
69
.73
Y
LS
KE
VF
DQ
LK
30
-40
Y
69
2.3
3 (
2+
) 1
38
3.7
8
YL
TK
EV
FD
KL
K
3
0-4
0
69
9.2
0 (
2+
); 4
66
.43
(3
+)
13
97
.76
Y
LT
KE
IFD
QL
K
3
0-4
0
41
0.4
8 (
2+
) 8
20
.45
A
VF
DQ
IK
3
4-4
0
41
7.5
2 (
2+
) 8
34
.46
A
LF
DQ
LK
34
-40
53
2.1
8 (
2+
) 1
06
3.5
7
AV
FD
QL
KD
K
3
4-4
2
53
9.1
6(2
+)
10
77
.58
A
VF
DQ
LK
EK
34
-42
Y
(P
MF
)
53
9.1
4 (
2+
) 1
07
7.5
8
AL
FD
QL
KD
K
3
4-4
2
Y
59
6.1
4 (
2+
) 1
19
1.6
6
DIF
DK
LK
GQ
K
3
4-4
3
60
3.2
6 (
2+
) 1
20
5.6
8
AL
FD
QL
KD
KK
34
-43
Y
61
5.9
9 (
2+
) 1
23
0.7
2
TS
LG
AT
LL
DV
IK
4
4-5
5
90
1.0
2 (
3+
); 6
76
.12
(4
+)
27
01
.27
Q
TD
KH
PN
KD
FG
DV
NT
FV
NV
DP
EG
K
9
5-1
18
11
27
.09
(3
+)
33
79
.61
Q
TD
KH
PN
KD
FG
DV
SS
FV
NV
DP
EG
QY
VIS
TR
95
-12
4
Y (
PM
F)
84
8.3
3 (
4+
) 3
39
0.6
6
QT
DK
HP
NK
DF
GD
VN
SF
VN
VD
PE
GK
FV
IST
R
9
5-1
24
Y
85
5.9
0 (
4+
) 3
42
0.6
7
QT
DK
HP
NK
DF
GD
VN
TF
VN
VD
PE
GK
YV
IST
R
9
5-1
24
Y
86
9.7
0 (
2+
) 1
73
8.7
8
DF
GD
VN
SF
VN
VD
PE
GK
10
3-1
18
87
6.7
7 (
2+
) 1
75
2.8
0
DF
GD
VN
TF
VN
VD
PE
GK
10
3-1
18
12
16
.22
(2
+);
81
1.1
4 (
3+
) 2
43
1.1
3
DF
GD
VS
SF
VN
VD
PE
GQ
YV
IST
R
1
03
-12
4
Y (
PM
F)
81
4.7
5 (
3+
) 2
44
2.1
8
DF
GD
VN
SF
VN
VD
PE
GK
FV
IST
R
1
03
-12
4
82
4.7
1 (
3+
) 2
47
2.1
9
DF
GD
VN
TF
VN
VD
PE
GK
YV
IST
R
1
03
-12
4
Y (
PM
F)
70
2.4
8 (
2+
) 1
40
3.6
7
DV
NA
FV
NV
DP
EG
K
1
06
-11
8
80
8.7
8 (
3+
) 2
42
4.0
4
CG
RS
ME
GY
PF
NP
CL
TE
AQ
YK
C
ysC
AM
12
7;M
SO
;CysC
AM
139
12
7-1
46
10
16
.43
(2
+)
20
31
.95
S
LQ
GY
PF
NP
CL
TE
SQ
YK
C
ysC
AM
1
30
-14
6
Y
10
17
.30
(2
+);
67
8.4
7 (
3+
) 2
03
3.9
1
SM
QG
YP
FN
PC
LT
EA
QY
K
CysC
AM
1
30
-14
6
Y
10
17
.77
(2
+)
20
34
.89
S
ME
GY
PF
NP
CL
TE
AQ
YK
C
ysC
AM
1
30
-14
6
10
25
.25
(2
+);
68
3.9
6 (
3+
) 2
04
9.9
0
SM
QG
YP
FN
PC
LT
EA
QY
K
MS
O;C
ysC
AM
1
30
-14
6
10
25
.85
(2
+);
68
4.3
0 (
3+
) 2
05
0.8
9
SM
EG
YP
FN
PC
LT
EA
QY
K
MS
O;C
ysC
AM
1
30
-14
6
88
0.3
9 (
3+
) 2
63
9.1
5
SM
EG
YP
FN
PC
LT
EA
QY
KE
ME
AK
C
ysC
AM
;MS
O14
8
13
0-1
51
88
5.6
9 (
3+
) 2
65
5.1
4
SM
EG
YP
FN
PC
LT
EA
QY
KE
ME
SK
M
SO
13
1;C
ysC
AM
1
30
-15
1
185
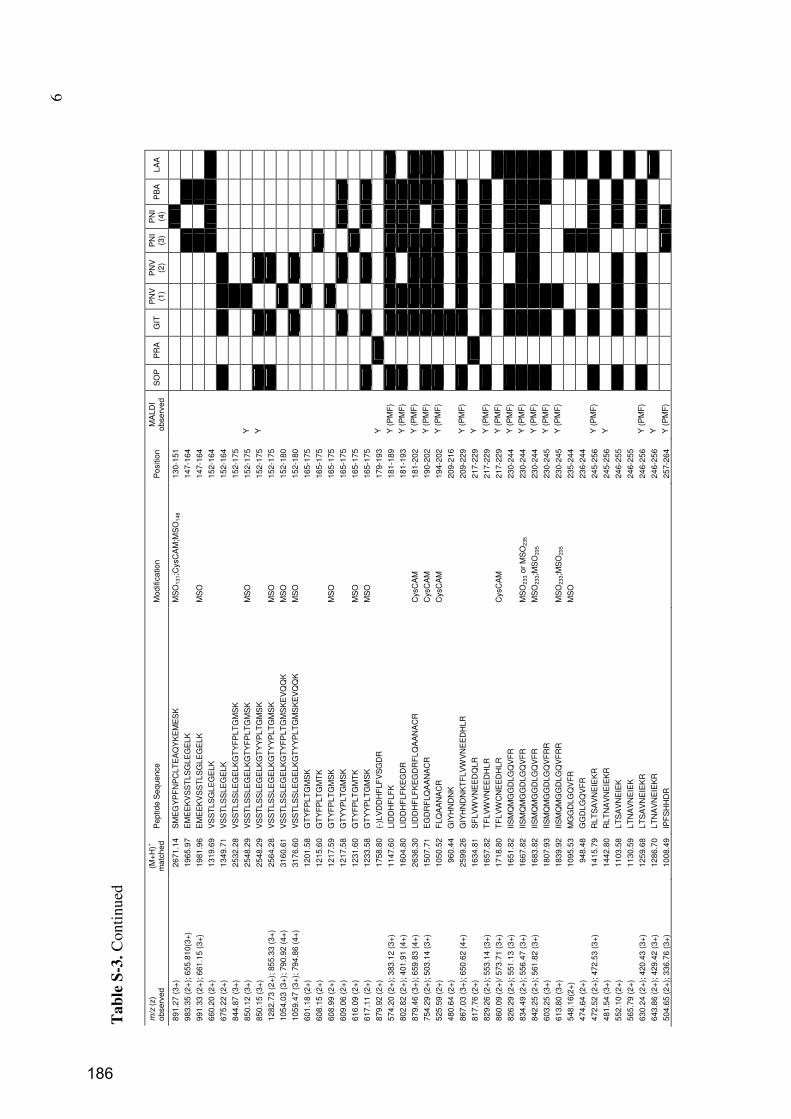
6
Tab
le S
-3. C
onti
nued
m/z
(z)
ob
se
rve
d
(M+
H)+
m
atc
he
d
Pe
ptid
e S
eq
ue
nce
M
od
ific
atio
n
Po
sitio
n
MA
LD
I o
bse
rve
d
SO
P
PR
A
GIT
P
NV
(1
) P
NV
(2
) P
NI
(3)
PN
I (4
) P
BA
L
AA
89
1.2
7 (
3+
) 2
67
1.1
4
SM
EG
YP
FN
PC
LT
EA
QY
KE
ME
SK
M
SO
13
1;C
ysC
AM
;MS
O1
48
13
0-1
51
98
3.3
5 (
2+
); 6
55
.81
0(3
+)
19
65
.97
E
ME
EK
VS
ST
LS
GL
EG
EL
K
1
47
-16
4
99
1.3
3 (
2+
); 6
61
.15
(3
+)
19
81
.96
E
ME
EK
VS
ST
LS
GL
EG
EL
K
MS
O
14
7-1
64
66
0.2
0 (
2+
) 1
31
9.6
9
VS
ST
LS
GL
EG
EL
K
1
52
-16
4
67
5.2
2 (
2+
) 1
34
9.7
1
VS
ST
LS
SL
EG
EL
K
1
52
-16
4
84
4.8
7 (
3+
) 2
53
2.2
8
VS
ST
LS
SL
EG
EL
KG
TY
FP
LT
GM
SK
15
2-1
75
85
0.1
2 (
3+
) 2
54
8.2
9
VS
ST
LS
SL
EG
EL
KG
TY
FP
LT
GM
SK
M
SO
1
52
-17
5
Y
85
0.1
5 (
3+
) 2
54
8.2
9
VS
ST
LS
SL
EG
EL
KG
TY
YP
LT
GM
SK
15
2-1
75
Y
12
82
.73
(2
+);
85
5.3
3 (
3+
) 2
56
4.2
8
VS
ST
LS
SL
EG
EL
KG
TY
YP
LT
GM
SK
M
SO
1
52
-17
5
10
54
.03
(3
+);
79
0.9
2 (
4+
) 3
16
0.6
1
VS
ST
LS
SL
EG
EL
KG
TY
FP
LT
GM
SK
EV
K
MS
O
15
2-1
80
10
59
.47
(3
+);
79
4.8
6 (
4+
) 3
17
6.6
0
VS
ST
LS
SL
EG
EL
KG
TY
YP
LT
GM
SK
EV
K
MS
O
15
2-1
80
60
1.1
8 (
2+
) 1
20
1.5
8
GT
YF
PL
TG
MS
K
1
65
-17
5
60
8.1
5 (
2+
) 1
21
5.6
0
GT
YF
PL
TG
MT
K
1
65
-17
5
60
8.9
9 (
2+
) 1
21
7.5
9
GT
YF
PL
TG
MS
K
MS
O
16
5-1
75
60
9.0
6 (
2+
) 1
21
7.5
8
GT
YY
PL
TG
MS
K
1
65
-17
5
61
6.0
9 (
2+
) 1
23
1.6
0
GT
YF
PL
TG
MT
K
MS
O
16
5-1
75
61
7.1
1 (
2+
) 1
23
3.5
8
GT
YY
PL
TG
MS
K
MS
O
16
5-1
75
87
9.9
2 (
2+
) 1
75
8.8
0
(-)L
VD
DH
FLF
VS
GD
R
1
79
-19
3
Y
57
4.2
0 (
2+
); 3
83
.12
(3
+)
11
47
.60
L
IDD
HF
LF
K
1
81
-18
9
Y (
PM
F)
80
2.8
2 (
2+
); 4
01
.91
(4
+)
16
04
.80
L
IDD
HF
LF
KE
GD
R
1
81
-19
3
Y (
PM
F)
87
9.4
6 (
3+
); 6
59
.83
(4
+)
26
36
.30
L
IDD
HF
LF
KE
GD
RF
LQ
AA
NA
CR
C
ysC
AM
1
81
-20
2
Y (
PM
F)
75
4.2
9 (
2+
); 5
03
.14
(3
+)
15
07
.71
E
GD
RF
LQ
AA
NA
CR
C
ysC
AM
1
90
-20
2
Y (
PM
F)
52
5.5
9 (
2+
) 1
05
0.5
2
FL
QA
AN
AC
R
CysC
AM
1
94
-20
2
Y (
PM
F)
48
0.6
4 (
2+
) 9
60
.44
G
IYH
ND
NK
20
9-2
16
86
7.0
3 (
3+
); 6
50
.62
(4
+)
25
99
.26
G
IYH
ND
NK
TF
LV
WV
NE
ED
HL
R
2
09
-22
9
Y (
PM
F)
81
7.7
6 (
2+
) 1
63
4.8
1
SF
LV
WV
NE
ED
QL
R
2
17
-22
9
Y
82
9.2
6 (
2+
); 5
53
.14
(3
+)
16
57
.82
T
FL
VW
VN
EE
DH
LR
21
7-2
29
Y
(P
MF
)
86
0.0
9 (
2+
)/ 5
73
.71
(3
+)
17
18
.80
T
FL
VW
CN
EE
DH
LR
C
ysC
AM
2
17
-22
9
Y (
PM
F)
82
6.2
9 (
2+
); 5
51
.13
(3
+)
16
51
.82
II
SM
QM
GG
DL
GQ
VF
R
2
30
-24
4
Y (
PM
F)
83
4.4
9 (
2+
); 5
56
.47
(3
+)
16
67
.82
II
SM
QM
GG
DL
GQ
VF
R
MS
O2
33 o
r M
SO
235
23
0-2
44
Y
(P
MF
)
84
2.2
5 (
2+
); 5
61
.82
(3
+)
16
83
.82
II
SM
QM
GG
DL
GQ
VF
R
MS
O2
33;M
SO
23
5
23
0-2
44
Y
(P
MF
)
60
3.2
5 (
3+
) 1
80
7.9
3
IIS
MQ
MG
GD
LG
QV
FR
R
2
30
-24
5
Y (
PM
F)
61
3.8
0 (
3+
) 1
83
9.9
2
IIS
MQ
MG
GD
LG
QV
FR
R
MS
O2
33;M
SO
23
5
23
0-2
45
Y
(P
MF
)
54
8.1
6(2
+)
10
95
.53
M
GG
DL
GQ
VF
R
MS
O
23
5-2
44
47
4.6
4 (
2+
) 9
48
.48
G
GD
LG
QV
FR
23
6-2
44
47
2.5
2 (
2+
); 4
72
.53
(3
+)
14
15
.79
R
LT
SA
VN
EIE
KR
24
5-2
56
Y
(P
MF
)
48
1.5
4 (
3+
) 1
44
2.8
0
RL
TN
AV
NE
IEK
R
2
45
-25
6
Y
55
2.1
0 (
2+
) 1
10
3.5
8
LT
SA
VN
EIE
K
2
46
-25
5
56
5.7
9 (
2+
) 1
13
0.5
9
LT
NA
VN
EIE
K
2
46
-25
5
63
0.2
4 (
2+
); 4
20
.43
(3
+)
12
59
.68
L
TS
AV
NE
IEK
R
2
46
-25
6
Y (
PM
F)
64
3.8
6 (
2+
); 4
29
.42
(3
+)
12
86
.70
L
TN
AV
NE
IEK
R
2
46
-25
6
Y
50
4.6
5 (
2+
); 3
36
.76
(3
+)
10
08
.49
IP
FS
HH
DR
25
7-2
64
Y
(P
MF
)
186

7
Tab
le S
-3. C
onti
nued
m/z
(z)
ob
se
rve
d
(M+
H)+
m
atc
he
d
Pe
ptid
e S
eq
ue
nce
M
od
ific
atio
n
Po
sitio
n
MA
LD
I o
bse
rve
d
SO
P
PR
A
GIT
P
NV
(1
) P
NV
(2
) P
NI
(3)
PN
I (4
) P
BA
L
AA
92
4.7
7 (
3+
) 2
77
2.4
0
VP
FS
HH
DR
LG
FL
TF
CP
TN
LG
TT
VR
C
ysC
AM
2
57
-28
0
Y (
PM
F)
92
9.4
5 (
3+
); 6
97
.37
(4
+)
27
86
.42
IP
FS
HH
DR
LG
FL
TF
CP
TN
LG
TT
VR
C
ysC
AM
2
57
-28
0
Y (
PM
F)
72
0.9
6 (
2+
) 1
44
0.7
2
LG
FL
TF
CP
TN
LG
T
CysC
AM
2
65
-27
7
89
8.8
5 (
2+
) 1
79
6.9
4
LG
FL
TF
CP
TN
LG
TT
VR
C
ysC
AM
2
65
-28
0
Y (
PM
F)
49
6.7
9 (
2+
); 3
31
.38
(3
+)
99
2.6
1
AS
VH
IKL
PK
28
1-2
89
50
5.0
7 (
3+
) 1
51
3.8
2
LA
AN
RD
KL
EE
VA
GK
29
0-3
03
Y
(P
MF
)
50
9.9
7(3
+)
15
27
.84
L
AA
NR
EK
LE
EV
AG
K
2
90
-30
3
57
2.6
0 (
3+
); 5
72
.39
(4
+)
22
87
.24
L
AA
NR
DK
LE
EV
AG
KY
NL
QV
R
2
90
-30
9
Y
76
7.7
0 (
3+
); 5
76
.04
(4
+)
23
01
.26
L
AA
NR
EK
LE
EV
AG
KY
NL
QV
R
2
90
-30
9
Y
50
1.6
5 (
2+
); 3
34
.89
(3
+)
10
02
.54
E
KL
EE
VA
GK
29
5-3
03
49
4.5
8 (
2+
) 9
88
.52
D
KL
EE
VA
GK
29
5-3
03
50
8.6
6 (
2+
) 1
01
6.5
5
EK
LE
EIA
GK
29
5-3
03
58
7.8
0 (
3+
) 1
76
1.9
4
DK
LE
EV
AG
KY
NL
QV
R
2
95
-30
9
Y (
PM
F)
88
8.4
2 (
2+
); 5
92
.62
(3
+)
17
75
.95
E
KL
EE
VA
GK
YN
LQ
VR
29
5-3
09
Y
(P
MF
)
59
7.3
6 (
3+
) 1
78
9.9
7
EK
LE
EIA
GK
YN
LQ
VR
29
5-3
09
Y
75
9.8
5 (
2+
); 5
06
.73
(3
+)
15
18
.82
L
EE
VA
GK
YN
LQ
VR
29
7-3
09
Y
(P
MF
)
76
6.8
0 (
2+
) 1
53
2.8
3
LE
EIA
GK
YN
LQ
VR
29
7-3
09
Y
39
6.5
3 (
2+
) 7
92
.43
Y
NL
QV
R
3
04
-30
9
10
17
.19
(2
+);
67
8.4
9 (
3+
); 5
09
.24
(4
+)
20
33
.94
G
TR
GE
HT
EA
EG
GIY
DIS
NK
31
0-3
28
Y
(P
MF
)
54
8.1
8 (
4+
) 2
19
0.0
4
GT
RG
EH
TE
AE
GG
IYD
ISN
KR
31
0-3
29
Y
(P
MF
)
58
7.3
4 (
4+
); 4
69
.91
(5
+)
23
46
.14
G
TR
GE
HT
EA
EG
GIY
DIS
NK
RR
31
0-3
30
Y
86
0.3
2 (
2+
); 5
73
.94
(3
+)
17
19
.77
G
EH
TE
AE
GG
IYD
ISN
K
3
13
-32
8
Y (
PM
F)
62
5.9
2 (
3+
); 4
69
.66
(4
+)
18
75
.87
G
EH
TE
AE
GG
IYD
ISN
KR
31
3-3
29
Y
(P
MF
)
50
8.7
7 (
4+
) 2
03
1.9
8
GE
HT
EA
EG
GIY
DIS
NK
RR
31
3-3
30
Y
(P
MF
)
42
7.0
0 (
3+
) 1
27
9.6
7
RM
GL
TE
FQ
AV
K
3
30
-34
0
43
2.5
5 (
3+
) 1
29
5.6
8
RM
GL
TE
FQ
AV
K
MS
O
33
0-3
40
64
5.9
8 (
4+
) 2
58
1.3
4
RM
GL
TE
FQ
AV
KE
MQ
DG
ILE
LIK
M
SO
33
1;M
SO
34
2
33
0-3
51
56
2.1
5 (
2+
) 1
12
3.5
7
MG
LT
EF
QA
VK
33
1-3
40
57
0.1
6 (
2+
) 1
13
9.5
8
MG
LT
EF
QA
VK
M
SO
3
31
-34
0
11
97
.10
(2
+);
79
8.4
2 (
3+
) 2
39
3.2
4
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IK
3
31
-35
1
12
05
.19
(2
+);
80
3.6
4 (
3+
) 2
40
9.2
3
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IK
MS
O3
31 o
r M
SO
342
33
1-3
51
12
13
.06
(2
+);
80
9.0
1 (
3+
) 2
42
5.2
4
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IK
MS
O3
31;M
SO
34
2
33
1-3
51
69
1.7
5 (
4+
) 2
76
3.4
6
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
K
3
31
-35
4
69
5.4
5 (
4+
) 2
77
9.4
5
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
K
MS
O3
31
33
1-3
54
69
9.5
1 (
4+
) 2
79
5.4
6
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
K
MS
O3
31;M
SO
34
2
33
1-3
54
10
08
.49
(3
+);
75
6.6
6 (
4+
) 3
02
3.5
5
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
KE
M
3
31
-35
6
10
13
.82
(3
+);
76
0.6
1 (
4+
) 3
03
9.5
5
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
KE
M
MS
O3
31 o
r M
SO
342 o
r M
SO
356
33
1-3
56
10
19
.05
(3
+);
76
4.5
0 (
4+
) 3
05
5.5
4
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
KE
M
two
of
MS
O33
1 o
r M
SO
34
2 o
r M
SO
35
6
33
1-3
56
76
5.1
4 (
4+
) 3
05
7.5
0
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKM
EK
EM
M
SO
33
1 o
r M
SO
342 o
r M
SO
356
33
1-3
56
10
24
.50
(3
+);
76
8.7
9 (
4+
) 3
07
1.5
4
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKIE
KE
M
MS
O3
31;M
SO
34
2;M
SO
356
33
1-3
56
10
25
.14
(3
+);
76
9.0
9 (
4+
) 3
07
3.5
0
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKM
EK
EM
M
SO
33
1 o
r M
SO
34
2 o
r M
SO
35
2;M
SO
35
6
33
1-3
56
187

8
Tab
le S
-3. C
onti
nued
m/z
(z)
ob
se
rve
d
(M+
H)+
m
atc
he
d
Pe
ptid
e S
eq
ue
nce
M
od
ific
atio
n
Po
sitio
n
MA
LD
I o
bse
rve
d
SO
P
PR
A
GIT
P
NV
(1
) P
NV
(2
) P
NI
(3)
PN
I (4
) P
BA
L
AA
10
30
.56
(3
+)
30
89
.49
M
GL
TE
FQ
AV
KE
MQ
DG
ILE
LIK
ME
KE
M
MS
O3
31 o
r M
SO
34
2;M
SO
35
2;M
SO
356
33
1-3
56
10
35
.39
(3
+);
77
7.0
3 (
4+
) 3
10
5.4
9
MG
LT
EF
QA
VK
EM
QD
GIL
EL
IKM
EK
EM
M
SO
33
1;M
SO
34
2;M
SO
352;M
SO
35
6
33
1-3
56
64
4.6
6 (
2+
) 1
28
8.6
7
EM
QD
GIL
EL
IK
3
41
-35
1
65
2.7
1 (
2+
) 1
30
4.6
8
EM
QD
GIL
EL
IK
MS
O
34
1-3
51
55
8.8
2 (
3+
) 1
67
4.9
0
EM
QD
GIL
EL
IKIE
K
MS
O
34
1-3
54
64
0.2
2 (
3+
) 1
91
8.9
8
EM
QD
GIL
EL
IKIE
KE
M
3
41
-35
6
97
5.8
6 (
2+
); 6
50
.88
(3
+)
19
50
.98
E
MQ
DG
ILE
LIK
IEK
EM
M
SO
34
2;M
SO
35
6
34
1-3
56
66
2.2
2 (
3+
) 1
98
4.9
3
EM
QD
GIL
EL
IKM
EK
EM
M
SO
34
2;M
SO
35
2;M
SO
356
34
1-3
56
m/z
: mas
s/ch
arge
. MA
LD
I ob
serv
ed; (
Y):
pep
tide
foun
d by
MA
LD
I-T
OF
MS;
(P
MF)
: pep
tide
mat
ched
by
pept
ide
mas
s fi
nger
prin
t wit
h an
AK
ent
ry in
pro
tein
dat
abas
es. (■
)
deno
tes
the
pres
ence
of
a pe
ptid
e, a
nd (�
) th
e ab
senc
e. C
ysC
AM
, car
boxy
met
hyla
ted
cyst
eine
; MSO
, met
hion
ine
sufo
xide
; N-A
cyl,
acet
yl N
-ter
min
al. S
OP
, F. n
otia
lis;
PR
A,
Pan
dalu
s bo
real
is,
GIT
, P
enae
us m
onod
on;
PN
V,
Lit
open
aeus
van
nam
ei;
PN
I, F
enne
rope
naeu
s in
dicu
s; P
BA
, F
enne
rope
naeu
s m
ergu
iens
is;
LA
A,
Ple
otic
us m
uell
eri.
(1)
Cos
ta R
ica
popu
latio
n; (
2) A
rgen
tina
pop
ulat
ion;
(3)
Moz
ambi
que
popu
latio
n; (
4) M
adag
asca
r po
pula
tion.
188

CAPÍTULO 5 PROCEDIMIENTO Y KIT PARA LA IDENTIFICACIÓN RÁPIDA DE LAS PRINCIPALES ESPECIES COMERCIALES DE LANGOSTINOS Y CAMARONES


Capítulo 5: Procedimiento y kit para la identificación rápida de las
principales especies comerciales de langostinos y camarones.
Comentario
Una vez caracterizados los péptidos diferenciales, la siguiente etapa consistió en aplicar todo el
conocimiento adquirido al diseño de un método que permitiera realizar un análisis de identificación de
especies robusto, sencillo y de gran rapidez.
En este trabajo se desarrolló una metodología de monitorización de los péptidos diferenciadores
diana, utilizando un espectrómetro de masas de trampa iónica trabajando en modo SMIM, acoplado on-
line a una separación por RP-HPLC. Para acelerar el procedimiento, la digestión tríptica clásica se
realizó mediante la aplicación de ultrasonidos focalizados de alta intensidad, con lo que el tiempo
necesario para la digestión se redujo de varias horas a un minuto.
Los resultados obtenidos demuestran que, combinando la digestión ultrarrápida con la
monitorización de péptidos diana utilizando la técnica SMIM, se consigue la identificación inequívoca
de las especies consideradas en un tiempo total de 91 minutos, lo que supone un gran avance respecto a
cualquier otro método disponible en la actualidad para la autentificación de especies de langostinos y
camarones, ya sean métodos electroforéticos o genéticos.
En el presente capitulo se describe la patente desarrollada en este estudio. La invención se refiere
a un método de identificación de las principales especies extractivas y de acuicultura de langostinos
comerciales. La metodología desarrollada (Figura 22), basada en la monitorización por espectrometría
de masas de los péptidos diferenciales, permite identificar siete especies de relevancia comercial en
productos y subproductos frescos, refrigerados, congelados y/o elaborados.
191

Capítulo 5: Procedimiento y kit para la identificación rápida de las principales especies (…)
Figura 22. Representación esquemática de las etapas del procedimiento de identificación desarrollado.
192

PUBLICACIÓN
193


���������� �� �������� ��������� �� ��������� �� ����
���� ������� �� � ��� � ����� �� �� �� �� ��� � �� �� ��� � �� �������� �� ������� ��� ���
�������� ��� � � ����� �� ����� �� ����� �� �� ���� �!� "� ��� �� �� �� �� ����� �� ����
���#� �� � �$��� �� ��� � �� % �� ����� �� ������ ��� ������� �� ������� &'�( ��� )��������
���� �� ����� �� �� �� *�% &&+&,-.� �� /0 �� ����� �� �������� *� ����� �� ��������� �� �� ��
��� � �� �� ������ ��� �� ���� // �� �� *�% �� �������� �� ���# ��� ���� ������ �������
1$��� �� ��� � ��2 /00,(0'/'
3���� �� ������ ��2 04 �� � /00,� &(25- 67�89:
�� � �� ���������2 �� ! !��� �
8 �������� �2 �8&.'&�',4
8�� � �����2 7�18�;� 8< �)=�) >� =1?�89=@"7=�1�8 7=�19A3=7"8 678=7:
1$��� �� ��� � ������2 &
���2 �8
9����2 )�7�>=!=�19� B C=9 ")" *" =>�19=3=7"7=D1 >� *"8
)=17= "*�8 �8 �7=�8 7�!�)7="*�8 >� *"1@�89=1�8 B
7"!")�1�8
>������� ��� ����2 >���� �� ������ 6(0 ��:
)� � �� ��� ����E&���� 6. ��:
)����E&���� 6& ��:
> ����E&���� 6&/ ��:
3��)7 9E&���� 6& ��:
8�F* >3���� 6/ ��:
8�F*9G9����
���H���E�������
��E���������
���� ��� ��E���%���
��E���E��������
������������
����������
��� ���� ���2 71I�19=>"> �18 7�18<*9�)�8 >� )� =�>"> =1><89)="*
8" E 7=3 "/-450-,& E 1�!J)� �18 ")=K� "1@�* E 1=3
505('/4,;��<I40(0&5('5��<I��� ����� / ����I31!9�7I��
3���� % ���� ��
������ ��2
04 �� � /00,� &(25, 67�89:
7�� � ��� �� ��� �����2 002"'2,,2-027J24�2J42'52(�2�527�2''2/,2'(2>'25/2"72-�2-&2.3
+!��� �� �� � �� )��������+
195

(1) MODALIDAD:PATENTE DE INVENCIÓN
MODELO DE UTILIDAD[✓][ ]
(2) TIPO DE SOLICITUD:PRIMERA PRESENTACIÓN
ADICIÓN A LA PATENTE EUROPEAADICIÓN A LA PATENTE ESPAÑOLA
SOLICITUD DIVISIONALCAMBIO DE MODALIDAD
TRANSFORMACIÓN SOLICITUD PATENTE EUROPEAPCT: ENTRADA FASE NACIONAL
[✓][ ][ ][ ][ ][ ][ ]
(3) EXP. PRINCIPAL O DE ORIGEN:MODALIDAD:
N.OSOLICITUD:FECHA SOLICITUD:
(4) LUGAR DE PRESENTACIÓN:LUGAR
OEPM, PresentaciónElectrónica
(5-1) SOLICITANTE 1:DENOMINACIÓN SOCIAL: CONSEJO SUPERIOR DE
INVESTIGACIONESCIENTÍFICAS (CSIC)
NACIONALIDAD: EspañaCÓDIGO PAÍS: ES
DNI/CIF/PASAPORTE: Q2818002DCNAE:PYME:
DOMICILIO: C/ Serrano, 117LOCALIDAD: MadridPROVINCIA: 28 Madrid
CÓDIGO POSTAL: 28006PAÍS RESIDENCIA: España
CÓDIGO PAÍS: ESTELÉFONO:
FAX:CORREO ELECTRÓNICO:
PERSONA DE CONTACTO:
MODO DE OBTENCIÓN DEL DERECHO:INVENCIÓN LABORAL: [✓]
CONTRATO: [ ]SUCESIÓN: [ ]
(6-1) INVENTOR 1:APELLIDOS: ORTEA GARCÍA
NOMBRE: IGNACIONACIONALIDAD: España
CÓDIGO PAÍS: ESDNI/PASAPORTE: 0-T.
(6-2) INVENTOR 2:APELLIDOS: GALLARDO ABUÍN
NOMBRE: JOSÉ MANUELNACIONALIDAD: España
CÓDIGO PAÍS: ESDNI/PASAPORTE: 0-T.
(6-3) INVENTOR 3:APELLIDOS: MEDINA MÉNDEZ
NOMBRE: ISABELNACIONALIDAD: España
CÓDIGO PAÍS: ESDNI/PASAPORTE: 0-T.
196

(6-4) INVENTOR 4:APELLIDOS: BARROS TAJES
NOMBRE: LORENANACIONALIDAD: España
CÓDIGO PAÍS: ESDNI/PASAPORTE: 0-T.
(7) TÍTULO DE LA INVENCIÓN:PROCEDIMIENTO Y KITPARA LA IDENTIFICACIÓNDE LAS PRINCIPALESESPECIES COMERCIALESDE LANGOSTINOS YCAMARONES
(8) PETICIÓN DE INFORME SOBRE EL ESTADO DE LA TÉCNICA:SI
NO[ ][✓]
(9) SOLICITA LA INCLUSIÓN EN EL PROCEDIMIENTO ACELERADO DECONCESIÓN
SINO
[ ][✓]
(10) EFECTUADO DEPÓSITO DE MATERÍA BIOLÓGICA:SI
NO[ ][✓]
(11) DEPÓSITO:REFERENCIA DE IDENTIFICACIÓN:
INSTITUCIÓN DE DEPÓSITO:NÚMERO DE DEPÓSITO:
ACCESIBILIDAD RESTRINGIDA A UN EXPERTO (ART. 45.1. B):
(12) DECLARACIONES RELATIVAS A LA LISTA DE SECUENCIAS:
LA LISTA DE SECUENCIAS NO VA MÁS ALLÁ DEL CONTENIDO DE LA SOLICITUDLA LISTA DE SECUENCIAS EN FORMATO PDF Y ASCII SON IDENTICOS
[✓][✓]
(13) EXPOSICIONES OFICIALES:LUGAR:FECHA:
(14) DECLARACIONES DE PRIORIDAD:PAÍS DE ORIGEN:
CÓDIGO PAÍS:NÚMERO:
FECHA:
(15) AGENTE/REPRESENTANTE:APELLIDOS: PONS ARIÑO
NOMBRE: ÁNGEL
NACIONALIDAD: EspañaCÓDIGO PAÍS: ES
DNI/CIF/PASAPORTE: 50534279-J
DOMICILIO: Glorieta Rubén Darío, 4LOCALIDAD: MadridPROVINCIA: 28 Madrid
CÓDIGO POSTAL: 28010PAÍS RESIDENCIA: España
CÓDIGO PAÍS: ESTELÉFONO:
FAX:CORREO ELECTRÓNICO:
NÚMERO DE PODER: 20081765
(16) RELACIÓN DE DOCUMENTOS QUE SE ACOMPAÑAN:
DESCRIPCIÓN: [✓] N.º de páginas: 30REIVINDICACIONES: [✓] N.º de reivindicaciones:
32
197

DIBUJOS: [✓] N.º de dibujos: 5RESUMEN: [✓] N.º de páginas: 1
FIGURA(S) A PUBLICAR CON EL RESUMEN: [ ] N.º de figura(s):ARCHIVO DE PRECONVERSION: [ ]
DOCUMENTO DE REPRESENTACIÓN: [ ] N.º de páginas:JUSTIFICANTE DE PAGO (1): [✓] N.º de páginas: 1LISTA DE SECUENCIAS PDF: [✓] N.º de páginas: 2
ARCHIVO PARA LA BUSQUEDA DE LS: [✓]OTROS (Aparecerán detallados):
(17) EL SOLICITANTE SE ACOGE AL APLAZAMIENTO DE PAGO DE TASAPREVISTO EN EL ART. 162 DE LA LEY 11/1986 DE PATENTES, DECLARA: BAJOJURAMIENTO O PROMESA SER CIERTOS TODOS LOS DATOS QUE FIGURANEN LA DOCUMENTACIÓN ADJUNTA:
[ ]
DOC COPIA DNI: [ ] N.º de páginas:DOC COPIA DECLARACIÓN DE CARENCIA DE MEDIOS: [ ] N.º de páginas:
DOC COPIA CERTIFICACIÓN DE HABERES: [ ] N.º de páginas:DOC COPIA ÚLTIMA DECLARACIÓN DE LA RENTA: [ ] N.º de páginas:
DOC COPIA LIBRO DE FAMILIA: [ ] N.º de páginas:DOC COPIA OTROS: [ ] N.º de páginas:
(18) NOTAS:
(19) FIRMA DIGITAL:
FIRMA DEL SOLICITANTE O REPRESENTANTE: ENTIDAD PONSCONSULTORES DEPROPIEDAD INDUSTRIALSA - CIF A28750891 -NOMBRE PONS ARIÑOANGEL - NIF 50534279J
LUGAR DE FIRMA: MadridFECHA DE FIRMA: 07 Julio 2009
198

2
Procedimiento y kit para la identificación de las principales especies
comerciales de langostinos y camarones.
La presente invención se encuentra dentro del campo de la bioquímica y de la
taxonomía, y se refiere a un procedimiento rápido mediante espectrometría de 5
masas, para la identificación de las principales especies comerciales de
langostinos o camarones (Orden Decapoda) pertenecientes a las familias
Penaeidae (Penaeus monodon, Farfantepenaeus notialis, Fenneropenaeus
indicus, Fenneropenaeus merguiensis, Litopenaeus vannamei) Solenoceridae
(Pleoticus muelleri) y Pandalidae (Pandalus borealis). Este procedimiento 10
permite la identificación inequívoca de la especie concreta en productos y
subproductos frescos, refrigerados, congelados y/o elaborados, de indiscutible
aplicación en los sectores alimentario, importador, sanitario y de control de
calidad.
15
ESTADO DE LA TÉCNICA ANTERIOR
En la actualidad, dentro del mercado de los productos pesqueros, los
langostinos y camarones son productos de gran valor. Así, en 2004
representaban el 16.5% del total del mercado internacional de productos 20
pesqueros y su producción mundial en 2006 fue de unos 6.5 millones de
toneladas, equivalentes a un valor de más de 24.000 millones de dólares
(FAO), producción que no ha dejado de crecer en los últimos años. Destacan
las especies L. vannamei, con 2.300.000 toneladas, P. monodon, con 833.000
toneladas, y P. borealis, con 421.000 toneladas de producción (FIGIS -25
Fisheries Global Information System- FAO; http://www.fao.org/fishery/figis).
Diversos países, entre ellos los integrantes de la Unión Europea, destacan
entre sus objetivos prioritarios lograr la transparencia en los mercados
pesqueros y ofrecer al consumidor información fiable sobre los productos que 30
van a consumir. Este hecho resulta fundamental en un mercado cada vez más
globalizado, en el que se pueden encontrar varias especies distintas bajo una
199

3
misma denominación. Este es el caso concreto de las especies de langostinos
y camarones pertenecientes al Orden Decapoda, del que las especies más
comerciales se engloban en la superfamilia Penaeoidea (familias Penaeidae y
Solenoceridae) y la familia Pandalidae. La incorrecta catalogación, entre otras
razones, debida a que especies de origen y características muy diversas no 5
son adecuadamente identificadas, puede ocasionar problemas de sustituciones
que afectarían tanto al etiquetado como a la actividad comercial del sector y al
propio consumidor. Como el valor comercial varía según las especies y los
productos derivados, las entidades administrativas deben proteger a los
consumidores de casos de sustitución de especies, mediante medidas de 10
control que aseguren un adecuado etiquetado del producto.
El control de origen y autenticidad de especies se hace más necesario cuanto
mayor es la diversidad de especies susceptibles de ser comercializadas bajo el
mismo nombre. En el mercado existe una notable heterogeneidad de las 15
especies y géneros que constituyen las principales denominaciones
comerciales de langostinos. Cabe destacar que bajo los términos “langostino
banana”, “langostino blanco” o “langostino tigre” se encuadran distintas
especies de langostinos e incluso distintos géneros. Respecto a la captura,
cuanto mayor es el grado de solapamiento geográfico entre los caladeros de 20
las distintas especies, mayor es la frecuencia con la que se pueden pescar dos
especies distintas y difícilmente distinguibles en una misma área. Así, por
ejemplo, en el océano Índico coexisten tres de las especies de langostinos más
comerciales, P. monodon, F. indicus y .F. merguiensis, las cuales son
habitualmente capturadas en una misma pesquería. Y por último, cuanto mayor 25
es el grado de procesado o manipulación del crustáceo previo a la venta,
mediante procesos tales como el pelado, eviscerado, descabezado, etc., mayor
es la dificultad de la identificación de los especimenes utilizados en dicho
producto. A estas circunstancias hay que añadir la creciente exigencia del
consumidor por estar adecuadamente informado de la composición de los 30
alimentos en un mercado cada vez más liberalizado y global, y, por tanto, más
expuesto a riesgos sanitarios imprevisibles.
200

4
Por todo lo anteriormente expuesto, es imprescindible lograr una información
contrastada y veraz sobre la naturaleza y el origen de los alimentos, con lo cual
a su vez se protegerán a los productos de calidad frente a casos de sustitución
de especies, que dan lugar a fraudes al consumidor y a competencia desleal
entre productores. 5
En materia de etiquetado a nivel internacional el Codex Alimentarius (una
acción conjunta de la FAO y la OMS) es punto de referencia para armonizar
las normas alimentarias a nivel mundial. Por otro lado, en la Unión Europea, el
Reglamento (CE) nº 104/2000 establece la Organización Común de Mercados 10
en el sector de los productos de la pesca y de la acuicultura, y dispone que “los
Estados miembros establecerán y publicarán el Listado de denominaciones
comerciales de especies pesqueras y de acuicultura”. En España, el Real
Decreto 1380/2002 y el Real Decreto 121/2004 - teniendo en cuenta la Ley
3/2001 de Pesca Marítima del Estado - regulan la identificación y el etiquetado 15
de los productos de la pesca, la acuicultura y el marisqueo, congelados o
ultracongelados el primero, y vivos, frescos, refrigerados o cocidos el segundo.
Un requisito fundamental del etiquetado de productos pesqueros es la
identificación clara del nombre de la especie utilizada.
20
La existencia de toda esta serie de normativas necesita herramientas fiables
que permitan la identificación certera del origen y naturaleza del producto.
Estas herramientas han de basarse en metodologías analíticas que identifiquen
la especie a partir de la cual se ha elaborado el producto a analizar, aunque ya
no existan indicios morfológicos o sensoriales. De hecho, solamente los 25
expertos son capaces de identificar las especies mediante una serie de
caracteres tanto morfológicos como anatómicos cuando el crustáceo se
presenta intacto, ya sea fresco o congelado (Mafra et al., 2008. European Food
Research and Technology, 227(3), 649-665). En cambio una vez procesado
(pelado, congelado, descabezado, etc.), la mayoría de los caracteres 30
morfológicos desaparecen por lo que la identificación se hace prácticamente
imposible.
201

5
Como una primera alternativa a la identificación morfológica para las especies
de langostinos del Orden Decapoda, se abordaron herramientas analíticas
encaminadas al análisis de las proteínas procedentes de sus tejidos. Los
primeros trabajos de investigación fueron dos estudios de An y colaboradores
(An et al., 1988. Journal of Food Science, 53(2), 313-318; An et al., 1989. 5
Journal of Food Science, 54(2), 233-236), utilizando SDS-PAGE el primero e
isoelectroenfoque el segundo. En estos trabajos sólo se incluyeron tres
especies minoritarias (Penaeus duorarum, Penaeus setiferus y Sicyonia
brevirostris), consiguiendo distinguir el segundo estudio entre las tres especies,
pero no cuando se presentaba una mezcla con dos de las especies. 10
Más recientemente, se han propuesto dos métodos basados en el análisis del
ADN para la identificación de especies de langostinos y camarones. En el
primero (Khamnamtong et al., 2005. Journal of Biochemistry and Molecular
Biology, 38(4), 491-499) se cita un método para la identificación de especies de 15
langostinos peneideos en base al polimorfismo del gen 16S rRNA, utilizando
polimorfismos de longitud de los fragmentos de restricción (RFLP) y
polimorfismos de conformación de cadena sencilla (SSCP), previa amplificación
con la reacción en cadena de la polimerasa (PCR). En este estudio se pudo
identificar distintivamente las especies P. monodon, L. vannamei y F. 20
merguiensis, pero no la Penaeus semisulcatus y la Marsupenaeus japonicus.
En el segundo de los estudios basados en ADN (Pascoal et al., 2008.
Electrophoresis, 29(2), 499-509) se recoge un método, que ha sido patentado
(PCT/ES2007/070212), para la identificación de especies de langostinos de la
superfamilia Penaeoidea utilizando la combinación de PCR y RFLP, en el que 25
es necesario el corte con 3 enzimas de restricción distintos para conseguir una
correcta identificación de las especies. Estos mismos autores describen otro
método, también utilizando PCR y RFLP, para un número menor de especies
(Pascoal et al., 2008. Electrophoresis, 29 (15), 3220-3228).
30
Las técnicas de Biología Molecular no se han aplicado a la especie Pandalus
borealis, conocida como camarón nórdico o boreal, de gran interés comercial,
202

6
al ser la especie de camarón con una mayor cantidad de capturas (FIGIS,
FAO). Por otro lado, las técnicas genéticas disponibles hasta el momento para
la identificación de especies de langostinos, proporcionan una metodología
muy laboriosa, con numerosos etapas necesarias para la determinación final de
la especie, incluyendo la amplificación del ADN mediante PCR, el corte con 5
varios enzimas de restricción y la resolución de los fragmentos de restricción
resultantes en geles de electroforesis, al mismo tiempo que son técnicas que
exigen largos tiempos de análisis, ya que el resultado final suelen obtenerse
tras un periodo de tiempo de unas 18 -48 horas. Por tanto, existe la necesidad
de un método alternativo y/o complementario a las técnicas existentes que 10
resuelvan los inconvenientes anteriormente citados, mejorando en este sentido
la rapidez en el diagnóstico y la sencillez en el análisis, para las principales
especies de langostinos y camarones pertenecientes al Orden Decapoda,
incluyendo la especie P. borealis. Metodologías sencillas, rápidas y eficientes
que logren la identificación de la especie en cualquier etapa de la cadena 15
alimentaria y en cualquier producto ya sea fresco, refrigerado, congelado y/o
procesado.
DESCRIPCIÓN DE LA INVENCIÓN
20
La presente invención proporciona un procedimiento para la identificación
rápida, sencilla, sensible e inequívoca de las principales especies comerciales
de langostinos y camarones pertenecientes al orden Decapoda en cualquier
punto de la cadena alimentaria y en cualquier producto fresco, refrigerado,
congelado y/o procesado. La invención se basa en la identificación de una serie 25
de péptidos específicos resultantes de la digestión del extracto proteico
sarcoplásmico.
Por tanto, un primer aspecto de la invención se refiere a un procedimiento para
la obtención de datos útiles en la identificación de especies langostinos y 30
camarones pertenecientes al Orden Decapoda, de ahora en adelante primer
procedimiento de la invención, que comprende:
203

7
a) extracción de las proteínas sarcoplásmicas del músculo blanco de una
muestra biológica aislada
b) digerir las proteínas del extracto sarcoplásmico del paso (a), e
c) identificar la presencia o ausencia, en el extracto digerido del paso (b),
de los péptidos que se seleccionan de la lista que comprende: S-PEN829 (SEQ 5
ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-
PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO:
6), S-PEN603 (SEQ ID NO: 7).
Todos estos péptidos aislados están comprendidos en las distintas secuencias 10
de la proteína arginina kinasa de cada una de las especies de langostinos o
camarones.
En esta memoria se entiende por “muestra biológica” cualquier material
biológico de origen animal, preferiblemente de animales invertebrados, del 15
Phylum Arthropoda, del Subphylum Crustacea, y del Orden Decapoda, y que
pueda albergar información sobre la dotación genética característica de dicho
animal, como tejidos, células y proteínas. Así, podría ser, pero sin limitarse,
cualquier material del que se sospeche la presencia de material biológico de
origen animal, y cuya procedencia quiera ser determinada, como alimentos 20
frescos, refrigerados, congelados y/o procesados.
Las especies identificables con el procedimiento de la invención son las
siguientes: i) Familia Penaeidae: Penaeus monodon, Litopenaeus vannamei,
Farfantepenaeus notialis, Fenneropenaeus indicus, Fenneropenaeus 25
merguiensis; ii) Familia Solenoceridae: Pleoticus muelleri; y iii) Familia
Pandalidae: Pandalus borealis.
Estas especies son las principales especies comerciales de langostinos y
camarones, constituyendo dos de ellas (Litopenaeus vannamei y Penaeus 30
monodon) el 80% a nivel mundial (Pascoal et al., 2008. Electrophoresis, 29
(15), 3220-3228).
204

8
Así, la identificación de los péptidos específicos resumidos en la Tabla 1,
permite de forma escalonada discriminar si en una muestra biológica, por
ejemplo de alimento, existen trazas de langostino o camarón, si éstas
pertenecen a una u otra familia y la especie/s concreta/s utilizadas en dicho
producto. La determinación de la presencia o ausencia en la muestra problema 5
de la Familia Penaeidae, de la Familia Solenoceridae, y/o de la Familia
Pandalidae, se lleva a cabo valorando la presencia/ausencia de un total de 3
péptidos específicos, uno para cada familia:
- Péptido específico para la Familia Penaeidae: S-PEN829 (13
aminoácidos, SEQ ID NO: 1). 10
- Péptido específico para la Familia Solenoceridae: S-MUE643 (11
aminoácidos, SEQ ID NO: 2).
- Péptido específico para la Familia Pandalidae: S-BOR817 (13
aminoácidos, SEQ ID NO: 3).
15
Si la muestra no presenta ninguno de los péptidos específicos de Familia, el
procedimiento termina en esta etapa, y se puede concluir que en la muestra no
existe langostino o camarón. Si la muestra es positiva para la Familia
Solenoceridae, se identificará la especie Pleoticus muelleri. Si la muestra es
positiva para la Familia Pandalidae, se identificará la especie Pandalus 20
borealis.
Por tanto, en una realización preferida de este aspecto de la invención, el
procedimiento para la identificación de especies langostinos y camarones, de
ahora en adelante segundo procedimiento de la invención, que comprende los 25
pasos (a)-(c) del primer procedimiento de la invención y, adicionalmente:
d1) asignar la muestra biológica del paso (a) como perteneciente a la
especie Pleoticus muelleri de la Familia Solenoceridae, si el péptido identificado
en el paso (c) es S-MUE643 (SEQ ID NO: 2).
30
205

9
En otra realización preferida, el segundo procedimiento de la invención
comprende, adicionalmente a los pasos (a)-(c) del primer procedimiento de la
invención:
d2) asignar la muestra biológica del paso (a) como perteneciente a la
especie Pandalus borealis de la Familia Pandalidae, si el péptido identificado 5
en el paso (c) es S-BOR817 (SEQ ID NO: 3).
En otra realización preferida, el procedimiento de la invención comprende,
adicionalmente a los pasos (a)-(c) del primer procedimiento de la invención, la
etapa: 10
d3) asignar la muestra biológica del paso (a) como perteneciente a la
Familia Penaeidae, si el péptido identificado en el paso (c) es S-PEN829 (SEQ
ID NO: 1).
Si el análisis ha dado positivo para la Familia Penaeidae, la combinación de 15
presencia/ausencia de una serie de otros péptidos permitirá la identificación
concreta de la especie. El péptido S-PEN759 (13 aminoácidos, SEQ ID NO: 4)
permitirá la identificación del origen geográfico de los langostinos. Su presencia
revela la existencia en la muestra de alguna de las especies de langostino de
los océanos Índico y Pacífico (P. monodon, L. vannamei, F. indicus y F. 20
merguiensis); por el contrario, su ausencia es indicativa de la presencia de una
especie del océano Atlántico (F. notialis). Así, en una realización aún más
preferida, el segundo procedimiento de la invención comprende,
adicionalmente a la etapa d3, la etapa:
e1) asignar la muestra biológica del paso (a) como perteneciente a la 25
especie Farfantepenaeus notialis, si en el paso (c) no se ha identificado el
péptido S-PEN759 (SEQ ID NO: 4)
Conocida ya la procedencia, una serie de péptidos permitirán diferenciar las
especies. Los péptidos: 30
- S-PEN675 (13 aminoácidos, SEQ ID NO: 5)
- S-PEN539 (9 aminoácidos, SEQ ID NO: 6)
206

10
- S-PEN603 (10 aminoácidos, SEQ ID NO: 7)
permitirán la identificación inequívoca en la muestra de P. monodon, L.
vannamei, F. indicus y F. merguiensis. De esta manera, por ejemplo P.
monodon se identifica de forma precisa con la determinación en la muestra de
los péptidos S-PEN675 y S-PEN539; L. vannamei se identifica de forma precisa 5
con la presencia en la muestra del péptido S-PEN675 y la ausencia del péptido
S-PEN539; F. merguiensis se identifica de forma precisa con la presencia en la
muestra del péptido S-PEN603; F. indicus se identifica de forma clara con la
ausencia de todos los anteriores (Tabla 1).
10
Así, en otra realización más preferida, el segundo procedimiento de la
invención comprende, adicionalmente, y además de la etapa d3, la etapa:
e2) asignar la muestra biológica del paso (a) como perteneciente a la
especie P. monodon, si en el paso (c) se identifican los péptidos S-PEN675
(SEQ ID NO: 5) y S-PEN539 (SEQ ID NO: 6). 15
En otra realización más preferida, el segundo procedimiento de la invención
comprende, adicionalmente a la etapa d3, la etapa:
e3) asignar la muestra biológica del paso (a) como perteneciente a la
especie Litopenaeus vannamei, si en el paso (c) se identifica la presencia del 20
péptido S-PEN675 (SEQ ID NO: 5) y la ausencia del péptido S-PEN539 (SEQ
ID NO: 6).
En otra realización más preferida, el segundo procedimiento de la invención
comprende, adicionalmente a la etapa d3, la etapa: 25
e4) asignar la muestra biológica del paso (a) como perteneciente a la
especie Fenneropenaeus merguiensis, si en el paso (c) se identifica la
presencia del péptido S-PEN603 (SEQ ID NO: 7) y la ausencia de los péptidos
S-PEN675 (SEQ ID NO: 5) y S-PEN539 (SEQ ID NO: 6).
30
En otra realización más preferida, el segundo procedimiento de la invención
comprende, adicionalmente a la etapa d3, la etapa:
207

11
e5) asignar la muestra biológica del paso (a) como perteneciente a la
especie Fenneropenaeus indicus, si en el paso (c) se identifica la presencia del
péptido S-PEN759 (SEQ ID NO: 4) y la ausencia de los péptidos S-PEN675
(SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7)
5
Los pasos (a), (b) y/o (c) del procedimiento descrito anteriormente pueden ser
total o parcialmente automatizados, por ejemplo, por medio de un equipo
robótico sensor para la identificación de los péptidos S-PEN829 (SEQ ID NO:
1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759
(SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-10
PEN603 (SEQ ID NO: 7) en el paso (c). Además de los pasos especificados
anteriormente puede comprender otros pasos adicionales, por ejemplo
relacionados con el pre-tratamiento de la muestra o la evaluación de los
resultados obtenidos mediante este procedimiento.
15
Las técnicas y procedimientos necesarios para extraer las proteínas
sarcoplásmicas del músculo blanco a partir de una muestra biológica son
ampliamente conocidos por los técnicos y expertos en el campo de la técnica, y
pueden ser llevados a cabo igualmente por técnicas similares o ligeramente
modificadas. Así, en una realización preferida, la extracción a partir del músculo 20
blanco se realiza por homogeneización de la muestra, por ejemplo, pero sin
limitarnos, con agua o con un extractante de baja fuerza iónica, centrifugando
posteriormente, preferiblemente a 30.000 x g 10 min.
La digestión de las proteínas puede llevarse a cabo por varios métodos, como 25
por ejemplo mediante una incubación tradicional con una enzima, como la
tripsina. Además, se puede sustituir la tripsina por otras proteasas como la
quimiotripsina, endoproteinasa Glu-C, endoproteinasa Lys-C, etc., o
alternativamente se puede realizar la digestión mediante tratamientos de
hidrólisis química tales como el bromuro de cianógeno, hidroxilamina, etc. 30
Además, la digestión puede realizarse de forma simultánea o secuencial
mediante hidrólisis química y enzimática. En este sentido, la aplicación de
208

12
ultrasonidos a la muestra permite reducir el tiempo de realización de la etapa
de digestión. Con estos métodos alternativos se podrían generar otros péptidos
distintos a los descritos como específicos en la presente invención, si bien la
presencia de éstos últimos en la muestra permitiría reproducir de igual manera
el procedimiento particular. 5
Por tanto, en una realización preferida de este aspecto de la invención, la
digestión de las proteínas del paso (b) se realiza con una enzima proteasa,
más preferiblemente con la enzima tripsina. Aún más preferiblemente, la
digestión de las proteínas del paso (b) es ultrarrápida, y se realiza mediante la 10
aplicación de ultrasonidos focalizados de alta intensidad.
En otra realización preferida, la digestión de las proteínas del paso (b) se
realiza mediante hidrólisis química, y más preferiblemente, la digestión de las
proteínas del paso (b) es ultrarrápida, y se realiza mediante la aplicación de 15
ultrasonidos focalizados de alta intensidad
En la presente invención la separación y/o identificación de los distintos
péptidos se puede llevar a cabo por distintos métodos. Así, por ejemplo,
directamente mediante un espectrómetro de masas sin separación previa, o 20
también por LC-MS, mediante cromatografía HPLC acoplada a un
espectrómetro de masas con ionización por electrospray y con una trampa
iónica tridimensional o lineal, triple cuadrupolo o QqTof como analizador,
configurado en modo SIRM (selected ion reaction monitoring, monitorización de
la reacción de un ión seleccionado), SIM (selected ion monitoring, 25
monitorización de un ión seleccionado) monitorización de cada uno de los iones
específicos) o SMIM (selected MS/MS ion monitoring, monitorización de la
fragmentación de un ión seleccionado) (Jorge et al., 2007. Journal of Mass
Spectrometry, 42: 1391-1403). Para todas estas estrategias los espectros de
fragmentación de cada uno de los péptidos específicos se muestran en la Fig. 30
5.
209

13
Por tanto, en otra realización preferida de este aspecto de la invención, la
identificación de los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID
NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675
(SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7) se
realiza mediante espectrometría de masas. En otra realización más preferida, 5
se realiza mediante espectrometría de masas acoplada a cromatografía líquida
de alta resolución (HPLC) previa. En otra realización aún más preferida, el
espectrómetro de masas acoplado a un sistema HPLC previo trabajan en
cualquiera de los siguientes modos Data Dependent, SIM (selected ion
monitoring), SIRM (selected ion reaction monitoring) y SMIM (selected MS/MS 10
ion monitoring).
En otra realización preferida, la detección de los péptidos S-PEN829 (SEQ ID
NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759
(SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-15
PEN603 (SEQ ID NO: 7), se realiza mediante un anticuerpo. El término
“inmunoensayo”, tal y como se utiliza en la presente descripción se refiere a
cualquier técnica analítica que se basa en la reacción de la conjugación de una
anticuerpo con la muestra obtenida. Ejemplos de inmunoensayos conocidos en
el estado de la técnica son, por ejemplo, pero sin limitarse: hibridación de las 20
proteínas en membrana y posterior detección mediante anticuerpo, ya sea tras
electroforesis previa de las proteínas (western-blot) o tras su aplicación directa
(dot-blot), ensayo inmunoabsorbente ligado a enzimas (ELISA),
radioinmunoensayo (RIA), inmunohistoquímica o chips de proteína
(“microarrays”). 25
El término “anticuerpo” tal como se emplea en esta memoria, se refiere a
moléculas de inmunoglobulinas y porciones inmunológicamente activas de
moléculas de inmunoglobulinas, es decir, moléculas que contienen un sitio de
fijación de antígeno que se une específicamente (inmunorreacciona) con 30
alguno/s de los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO:
2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ
210

14
ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7). Ejemplos de
porciones de moléculas de inmunoglobulinas inmunológicamente activas,
incluyen fragmentos F(ab) y F(ab’)2 que pueden ser generados tratando el
anticuerpo con una enzima tal como la pepsina. Los anticuerpos pueden ser
policlonales (incluyen típicamente anticuerpos distintos dirigidos contra 5
determinantes o epítopos distintos) o monoclonales (dirigidos contra un único
determinante en el antígeno). El anticuerpo monoclonal puede ser alterado
bioquímicamente, por manipulación genética, o puede ser sintético, careciendo,
posiblemente, el anticuerpo en su totalidad o en partes, de porciones que no
son necesarias para el reconocimiento de alguno/s de los péptidos S-PEN829 10
(SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-
PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO:
6), S-PEN603 (SEQ ID NO: 7). y estando sustituidas por otras que comunican
al anticuerpo propiedades ventajosas adicionales. El anticuerpo puede ser
también recombinante, quimérico, sintético o una combinación de cualquiera de 15
los anteriores. Un “anticuerpo recombinante” es un anticuerpo que ha sido
producido en una célula hospedadora que ha sido transformada o transfectada
con el ácido nucleico codificante del anticuerpo, o que lo produce como
resultado de la recombinación homóloga.
20
De la misma manera, una vez conocida la secuencia aminoacídica de un
péptido, pueden diseñarse sondas para la identificación en una muestra de
dicho péptido a nivel de los ácidos nucleicos. Puesto que una misma secuencia
polipeptídica puede estar codificada por distintos oligonucleótidos, sin embargo,
se pueden localizar las secuencias de interés mediante un simple 25
procedimiento de ensayo y error. Así, por ejemplo pero sin limitarnos, todas las
secuencias de ADN que codifican para la secuencia polipeptídica dada (cada
uno de los péptidos de la invención o sus secuencias complementarias) se
sintetizan (por ejemplo mediante un método en fase sólida) y se marcan
radiactivamente mediante la fosforilación en sus extremos 5´ con 32P. Se coloca 30
una membrana de nitrocelulosa que contengan clones con los péptidos de la
invención en presencia de una mezcla que contenga todas estas sondas y se
211

15
somete a autorradiografía para determinar cuales contienen una secuencia que
coincida con la del péptido de interés. Por tanto, en otra realización preferida de
este aspecto de la invención, la detección de los péptidos S-PEN829 (SEQ ID
NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759
(SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-5
PEN603 (SEQ ID NO: 7), se realiza mediante un oligonucleótido.
Otro aspecto de la invención se refiere a un péptido denominado S-PEN829, de
ahora en adelante primer péptido de la invención, cuya secuencia aminoacídica
consiste en la SEQ ID NO: 1. 10
En esta memoria, el péptido S-PEN829 se define también por una secuencia
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-PEN829, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que 15
comprende la secuencia aminoacídica de la SEQ ID NO: 1,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b) debido
a la degeneración del código genético, 20
d) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%,
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 1,
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
actividad y las características estructurales del péptido S-PEN829. Incluye, por 25
tanto, diversas variantes del péptido S-PEN829, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
glicosilación, fosforilación o metilación. El término “variante” se define más
adelante en esta memoria.
30
212

16
Otro aspecto de la invención se refiere a un péptido denominado S-MUE643,
de ahora en adelante segundo péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 2
En esta memoria, el péptido S-MUE643 se define también por una secuencia 5
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-MUE643, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica de la SEQ ID NO: 2,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con 10
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b) debido
a la degeneración del código genético,
d) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%, 15
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 2.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
actividad y las características estructurales del péptido S-MUE643. Incluye, por
tanto, diversas variantes del péptido S-MUE643, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse, 20
glicosilación, fosforilación o metilación.
Otro aspecto de la invención se refiere a un péptido denominado S-BOR643,
de ahora en adelante tercer péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 3. 25
En esta memoria, el péptido S-BOR643 se define también por una secuencia
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-BOR643, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que 30
comprende la secuencia aminoacídica de la SEQ ID NO: 3,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con
213

17
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b) debido
a la degeneración del código genético,
d) moléculas de ácido nucleico que codifican un polipétptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%, 5
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 3.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
actividad y las características estructurales del péptido S-BOR643. Incluye, por
tanto, diversas variantes del péptido S-BOR643, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse, 10
glicosilación, fosforilación o metilación.
Otro aspecto de la invención se refiere a un péptido denominado S-PEN759,
de ahora en adelante cuarto péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 4. 15
En esta memoria, el péptido S-PEN759 se define también por una secuencia
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-BOR643, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que 20
comprende la secuencia aminoacídica de la SEQ ID NO: 4,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b)
debido a la degeneración del código genético, 25
d) moléculas de ácido nucleico que codifican un polipétptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%,
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 4.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
actividad y las características estructurales del péptido S-PEN759. Incluye, por 30
tanto, diversas variantes del péptido S-PEN759, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
214

18
glicosilación, fosforilación o metilación.
Otro aspecto de la invención se refiere a un péptido denominado S-PEN675,
de ahora en adelante quinto péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 5. 5
En esta memoria, el péptido S-PEN675 se define también por una secuencia
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-BOR643, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que 10
comprende la secuencia aminoacídica de la SEQ ID NO: 5,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b)
debido a la degeneración del código genético, 15
d) moléculas de ácido nucleico que codifican un polipétptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%,
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 5.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
actividad y las características estructurales del péptido S-PEN675. Incluye, por 20
tanto, diversas variantes del péptido S-PEN675, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
glicosilación, fosforilación o metilación.
Otro aspecto de la invención se refiere a un péptido denominado S-PEN539, 25
de ahora en adelante sexto péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 6, y su masa para el ión doblemente
cargado de 539,3 Da.
En esta memoria, el péptido S-PEN539 se define también por una secuencia 30
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-BOR643, y que comprendería diversas variantes procedentes de:
215

19
a) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica de la SEQ ID NO: 6,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b) 5
debido a la degeneración del código genético,
d) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%,
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 6.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la 10
actividad y las características estructurales del péptido S-PEN539. Incluye, por
tanto, diversas variantes del péptido S-PEN539, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
glicosilación, fosforilación o metilación.
15
Otro aspecto de la invención se refiere a un péptido denominado S-PEN603, de
ahora en adelante séptimo péptido de la invención, cuya secuencia
aminoacídica consiste en la SEQ ID NO: 7.
En esta memoria, el péptido S-PEN603 se define también por una secuencia 20
de nucleótidos o polinucleótido, que constituye la secuencia codificante del
péptido S-PEN603, y que comprendería diversas variantes procedentes de:
a) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica de la SEQ ID NO: 7,
b) moléculas de ácido nucleico cuya cadena complementaria hibrida con 25
la secuencia polinucleotídica de a),
c) moléculas de ácido nucleico cuya secuencia difiere de a) y/o b) debido
a la degeneración del código genético,
d) moléculas de ácido nucleico que codifican un polipéptido que
comprende la secuencia aminoacídica con una identidad de al menos un 80%, 30
un 90%, un 95%, un 98% o un 99% con la SEQ ID NO: 7.
en las que el polipéptido codificado por dichos ácidos nucleicos posee la
216

20
actividad y las características estructurales del péptido S-PEN603. Incluye, por
tanto, diversas variantes del péptido S-PEN603, esto es, péptidos resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
glicosilación, fosforilación o metilación.
5
En el sentido utilizado en esta descripción, el término "variante" se refiere a un
péptido sustancialmente homólogo a cualquiera de los péptidos que se
seleccionan de la lista que comprende: S-PEN829 (SEQ ID NO: 1), S-MUE643
(SEQ ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-
PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 10
7). En general, una variante incluye adiciones, deleciones o sustituciones de
aminoácidos. El término “variante” incluye también a las proteínas resultantes
de modificaciones postraducción como, por ejemplo, pero sin limitarse,
glicosilación, fosforilación o metilación.
15
Tal como aquí se utiliza esta expresión, un péptido es "sustancialmente
homólogo” a cualquiera de los péptidos que se seleccionan de la lista que
comprende: S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-
BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO:
5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7), cuando su 20
secuencia de aminoácidos presenta un buen alineamiento con la secuencia de
aminoácidos de cualquiera de los péptidos que se seleccionan de la lista que
comprende: S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-
BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO:
5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7), es decir, cuando su 25
secuencia de aminoácidos tiene un grado de identidad respecto a la secuencia
de aminoácidos de SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO:
4, SEQ ID NO: 5, SEQ ID NO: 6 o SEQ ID NO: 7 de, al menos, un 60%,
típicamente de, al menos, un 80%, ventajosamente de, al menos, un 85%,
preferiblemente de, al menos un 90%, más preferiblemente de, al menos, un 30
95%, y, aún más preferiblemente de, al menos, un 99%.
217

21
El término “identidad”, tal y como se utiliza en esta memoria, hace referencia a
la proporción de aminoácidos idénticos entre dos secuencias aminoacídicas
que se comparan. El tanto por ciento de identidad existente entre dos
secuencias puede ser identificado fácilmente por un experto en la materia, por
ejemplo, con la ayuda de un programa informático apropiado para comparar 5
secuencias, que incluye, aunque sin limitarse a ellos, el programa BLASTP o
BLASTN, y FASTA (Altschul et al., 1990, Journal of Molecular Biology 215: 403-
410).
El término "fragmento", tal y como se utiliza en la presente descripción se 10
refiere a una porción de cualquiera de los péptidos que se seleccionan de la
lista que comprende: S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2)
y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ
ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7) o de una sus
variantes. 15
La expresión "funcionalmente equivalente", tal como aquí se utiliza, significa
que el péptido o el fragmento del péptido en cuestión mantienen esencialmente
las propiedades inmunológicas descritas más alante en este documento.
Dichas propiedades inmunológicas se pueden determinar mediante métodos 20
convencionales en el estado de la técnica.
Los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-
BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO:
5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7) de la presente 25
invención pueden emplearse para generar anticuerpos, que son herramientas
que permiten la identificación de los péptidos de la presente invención.
Técnicas para la generación de anticuerpos son conocidas en el estado de la
técnica por los expertos en la materia. Así, por ejemplo, pero sin limitarnos,
inyectando un antígeno en un mamífero, como un ratón, rata o conejo (en el 30
caso de que se requiera poca cantidad) o una cabra, oveja o caballo (si se
requiere grandes cantidades) se puede generar un antisuero a partir de estos
218

22
animales, que está constituido por anticuerpos policlonales. O por ejemplo, si
se desea obtener anticuerpos específicos para un único epítopo de un
antígeno, se aíslan linfocitos secretores de anticuerpos de un animal y se
inmortalizan fusionándolos con una línea celular cancerosa. Las células
fusionadas se denominan hibridomas y continuarán creciendo y secretando 5
anticuerpo en el cultivo. Se aíslan las células de hibridoma individuales
mediante clonado por dilución para generar clones que produzcan todos el
mismo anticuerpo. A estos anticuerpos se les denomina anticuerpos
monoclonales.
10
Por tanto, otro aspecto de la invención se refiere al uso de los péptidos S-
PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-BOR817 (SEQ ID
NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID NO: 5), S-PEN539
(SEQ ID NO: 6), y/o S-PEN603 (SEQ ID NO: 7) para la generación de
anticuerpos. Preferiblemente, los anticuerpos se emplean para la identificación 15
de especies de langostinos y camarones pertenecientes al Orden Decapoda
según el procedimiento de la invención.
Otro aspecto de la invención se refiere a un kit para la identificación de
especies de langostinos y camarones pertenecientes al Orden Decapoda, de 20
ahora en adelante kit de la invención, que comprende los medios necesarios
para la identificación de los péptidos de la invención. Así, puede contener, por
ejemplo pero sin limitarse, tampones, agentes para prevenir la contaminación,
concentraciones adecuadas de tripsina y/o otras enzimas, en el caso de la
detección de los péptidos mediante anticuerpos, podría contener anticuerpos 25
capaces de reconocer específicamente a dichos péptidos, o en el caso de la
detección mediante oligonucleótidos, podrá comprender cebadores, sondas,
anticuerpos poli o monoclonales, y todos aquellos reactivos necesarios para
realizar los procedimientos descritos anteriormente en este documento y/o
conocidos en el estado de la técnica. El kit además puede incluir, sin ningún 30
tipo de limitación, el uso de tampones, enzimas, enzimas polimerasas,
cofactores para obtener una actividad óptima de éstas, agentes para prevenir la
219

23
contaminación, etc. Por otro lado el kit puede incluir todos los soportes y
recipientes necesarios para su puesta en marcha y optimización. El kit puede
contener además otras proteínas o sondas de interés, que sirvan como
controles positivos y negativos. Preferiblemente, el Kit comprende además
todos los medios necesarios para llevar a cabo los procedimientos de la 5
invención, y más preferiblemente, puede incluir las instrucciones.
Los términos “oligonucleótido”, “polinucleótido” y “ácido nucleico” se usan aquí
de manera intercambiable, refiriéndose a formas poliméricas de nucleótidos de
cualquier longitud, tanto ribonucleótidos (ARN) como desoxiribonucleótidos 10
(ADN).
Los términos “secuencia aminoacídica”, “péptido”, “oligopéptido”, “polipéptido” y
“proteína” se usan aquí de manera intercambiable, y se refieren a una forma
polimérica de aminoácidos de cualquier longitud, que pueden estar, o no, 15
química o bioquímicamente modificados.
A lo largo de la descripción y las reivindicaciones, la palabra "comprende" y sus
variantes no pretenden excluir otras características técnicas, aditivos,
componentes o pasos. Para los expertos en la materia, otros objetos, ventajas 20
y características de la invención se desprenderán en parte de la descripción y
en parte de la práctica de la invención. Los siguientes ejemplos y dibujos se
proporcionan a modo de ilustración, y no se pretende que sean limitativos de la
presente invención.
25
DESCRIPCIÓN DE LAS FIGURAS
Fig. 1. Esquema general del procedimiento de la invención.
Fig. 2. Resultado de la monitorización de los péptidos específicos del 30
ejemplo de realización 2. A) Cromatogramas para identificar la familia. B)
220

24
Cromatograma para identificar el origen geográfico. C) Cromatogramas para la
determinación de la especie.
Fig. 3. Resultado de la monitorización de los péptidos específicos del
ejemplo de realización 3. A) Cromatogramas para identificar la familia. B) 5
Cromatogramas para identificar el origen geográfico. C) Cromatogramas para
la determinación de la especie.
Fig. 4. Resultado de la monitorización de los péptidos específicos del
ejemplo de realización 4. A) Cromatogramas para identificar la familia. B) 10
Cromatograma para identificar el origen geográfico. C) Cromatogramas para la
determinación de la especie.
Fig. 5. Espectros de fragmentación de los iones doblemente cargados
para los distintos péptidos de la invención. A) Espectro de fragmentación 15
del péptido S-PEN829 (SEQ ID NO: 1). B) Espectro de fragmentación del
péptido S-MUE643 (SEQ ID NO: 2). C) Espectro de fragmentación del péptido
S-BOR817 (SEQ ID NO: 3). D) Espectro de fragmentación del péptido S-
PEN759 (SEQ ID NO: 4). E) Espectro de fragmentación del péptido S-PEN675
(SEQ ID NO: 5). F) Espectro de fragmentación del péptido S-PEN539 (SEQ ID 20
NO: 6). G) Espectro de fragmentación del péptido S-PEN603 (SEQ ID NO: 7).
EJEMPLOS DE REALIZACIÓN
A continuación se ilustrará la invención mediante unos ensayos realizados por 25
los inventores, que pone de manifiesto la especificidad y efectividad del
procedimiento de la invención. La identificación de los péptidos en el ejemplo
de la invención se ha realizado mediante un sistema HPLC (Surveyor-LC
System, Thermo-Electron, San José, CA, USA), acoplado a un espectrómetro
de masas, lo que unido a una digestión ultrarrápida de las proteínas, permitió la 30
identificación de las especies de langostinos y camarones en unos 90 minutos.
221

25
Ejemplo 1.- Identificación de las distintas especies del Orden Decapoda.
Con el objeto de identificar el patrón de péptidos específicos de cada una de
las principales especies de langostinos o camarones pertenecientes al Orden
Decapoda, se procedió tal y como se describe en los siguientes apartados. 5
Todas las soluciones y el material empleado para el tratamiento de la muestra
deben estar libres de proteínas contaminantes. Se requiere desde el inicio: i)
una manipulación cuidadosa; ii) instrumental libre de proteínas y con baja
capacidad de adsorción proteica, y iii) reactivos de calidad extra (grado HPLC). 10
1.- Extracción de las proteínas sarcoplásmicas.
La extracción de las proteínas sarcoplásmicas de cada especie se realizó a
partir de la homogenización de 1 g de músculo blanco obtenido de la cola de 15
cada individuo analizado. Los trozos de tejido previamente triturados con una
espátula y un bisturí, se introdujeron en tubos de ensayo y se les añadió 2 mL
de agua milliQ con 5 mM de PMSF (este último de manera opcional). La
muestra se homogeneizó con un Ultra-Turrax (IKA-Werke, Staufen, Alemania),
durante 2 ciclos de aproximadamente 30 s. Posteriormente, el homogenizado 20
fue centrifugado a 30.000 g durante 10 minutos (y se recogió el sobrenadante
conteniendo las proteínas sarcoplásmicas. La concentración de proteína en el
mismo fue cuantificada mediante el método del ácido bicinconínico (Smith et
al., 1985. Anal Biochem. 150(1):76-85. Erratum in: Anal Biochem 1987;
163(1):279). 25
2.- Digestión de las proteínas sarcoplásmicas con tripsina.
Unos 100 µg del extracto de proteínas sarcoplásmicas de cada una de las
muestras fueron digeridos con 4 µg de tripsina, aplicando simultáneamente 30
ultrasonidos focalizados de alta intensidad (López-Ferrer et al., 2005. Journal of
Proteome Research 4: 1569-1574) durante 1 min.
222

26
3.- Separación y monitorización de péptidos de las muestras mediante sistema
LC-MS y SMIM.
Como etapa recomendable, pero opcional, los péptidos resultantes de la 5
digestión fueron acidificados y purificados a través de pequeñas columnas de
fase reversa. Del extracto peptídico de cada una de las muestras se inyectaron
5 µL en un sistema HPLC (Surveyor-LC System, Thermo-Electron, San José,
CA, USA), acoplado a un espectrómetro de masas con una fuente de
ionización electrospray y con una trampa iónica como analizador (LCQ-MS 10
Thermo-Finnigan, San Jose, CA, USA). La separación se realizó a través de
una columna de fase reversa (Thermo Hypersil-Keystone, San Jose, CA, USA),
a un flujo de 1,5-2 µL/min. Los péptidos fueron eluidos en un gradiente de 65
min del 5% al 45% de solvente B (Solvente A: 0,5% ácido acético. Solvente B:
0,5% ácido acético, 80% acetonitrilo). A medida que los péptidos iban 15
eluyendo, el espectrómetro de masas configurado en modo SMIM realizó una
monitorización continua de la fragmentación de los iones doblemente cargados
de los 7 péptidos específicos recogidos en la Tabla 1.
A partir de la fragmentación de los 7 péptidos o iones seleccionados, 20
contenidos en la Tabla 1, se pudo discriminar de forma escalonada si la
muestra pertenecía a un langostino o camarón, si éste pertenecía a una u otra
familia, su procedencia geográfica y determinar la especie concreta.
Estos péptidos específicos se seleccionaron teniendo en cuenta la 25
especificidad de los mismos, en comparación con las secuencias de proteínas
disponibles en las bases de datos (Swiss-Prot, UNIPROT). Los espectros de
fragmentación para cada uno de los péptidos específicos se muestran en la
Fig. 5.
30
223

27
Los siguientes ejemplos permiten ilustrar casos en los que la utilización del
procedimiento de la invención con o sin las etapas adicionales descritas
anteriormente permite un mayor control de productos de la cadena alimentaria.
Ejemplo 2.- Utilización del procedimiento de invención para el control de 5
fraude en importaciones.
Se plantea el problema de identificar la especie de langostino de una partida de
langostinos congelados, provenientes del mismo caladero (caladero de
Mozambique) e importadas a España para el consumo humano etiquetadas 10
como Penaeus spp. Para la descripción completa del procedimiento véase
etapas consecutivas (1-3) del Ejemplo 1.
Tras la monitorización SMIM en el sistema LC-MS (etapa 3), se llevó a cabo,
como etapa 4, un procedimiento escalonado para la identificación de cada 15
especie en particular, iniciando la discriminación de la existencia de langostino
o camarón o no en la muestra, la determinación de la familia de langostino o
camarón si esta existiese en la muestra, la procedencia geográfica y finalmente
la determinación particular de la especie concreta.
20
4.- Identificación de las especies capturadas.
A partir de la fragmentación de los péptidos o iones seleccionados se realizó la
identificación de la especie de las cuatro muestras de trabajo en dos etapas
consecutivas: 25
4.1.- Determinación de la presencia/ausencia en la muestra de la Familia
Penaeidae, la Familia Solenoceridae o la familia Pandalidae.
En esta primera etapa se comprobó la presencia o ausencia para cada una de 30
las muestras de tres posibles combinaciones de péptidos específicos: Familia
Penaeidae, S-PEN829 (SEQ ID NO: 1); Familia Solenoceridae S-MUE643
224

28
(SEQ ID NO: 2); Familia Pandalidae S-BOR817 (SEQ ID NO: 3). Todos los
especímenes fueron identificados como pertenecientes a la Familia Penaeidae
(Fig. 2A).
4.2.- Identificación del origen geográfico y de la especie de langostino presente 5
en la muestra.
En una primera aproximación la presencia del péptido S-PEN759 (SEQ ID NO:
4) permitió confirmar que se trataba de langostinos del Índico-Pacífico (Fig. 2B).
10
Finalmente la combinación de una serie de péptidos específicos recogidos en
la Tabla 1 determinó inequívocamente la especie (Fig. 2C). Así, las cuatro
muestras fueron identificadas como Penaeus monodon (langostino tigre).
De esta forma los organismos oficiales encargados del control de 15
importaciones habrían verificado la especie presente en esta partida comercial,
ya que la información proporcionada por el suministrador era insuficiente,
además de que se han importado langostinos de una zona donde se solapan
geográficamente dos especies distintas, lo que puede inducir a errores en el
etiquetado. 20
Ejemplo 3.- Utilización del procedimiento de la invención para la
autentificación de producto final y control de fraude al consumidor.
Se plantea el problema de identificar un lote de 4 muestras congeladas 25
etiquetadas como “colas de langostino vannamei congeladas”.
Para la descripción completa del procedimiento véase etapas consecutivas (1-
3) del Ejemplo 1. Tras la monitorización SMIM en el sistema LC-MS (etapa 3),
se llevó a cabo la etapa 4 30
4.- Identificación de las especies capturadas.
225

29
Realizada en dos etapas consecutivas:
4.1.- Determinación de la presencia en la muestra de la Familia Penaeidae, la
Familia Solenoceridae o la familia Pandalidae.
5
Para cada una de las muestras se comprobó la presencia o ausencia de tres
posibles combinaciones de péptidos específicos: S-PEN829 (SEQ ID NO: 1)
para la Familia Penaeidae; S-MUE643 (SEQ ID NO: 2) para la Familia
Solenoceridae; y S-BOR817 (SEQ ID NO: 3) para la Familia Pandalidae.
10
El resultado obtenido mostró que en todas las muestras existen especimenes
pertenecientes a la Familia Penaeidae (Fig. 3A).
4.2.- Identificación del origen geográfico y de la especie de langostino presente
en la muestra. 15
Una vez identificadas las muestras como langostinos Penaeidae, se procedió a
la determinación del origen geográfico. La presencia del péptido S-PEN759
(SEQ ID NO: 4) en todas las muestras confirmó que se trataba de langostinos
del Índico-Pacífico (Fig. 3B). 20
Posteriormente, la combinación de una serie de péptidos recogidos en la Tabla
1 permitió la determinación de especie (Fig. 3C). Todas las muestras fueron
identificadas como Litopenaeus vannamei (camarón patiblanco).
25
Como resultado, las autoridades competentes, o las empresas responsables de
la inspección de los productos, habrían verificado el correcto etiquetado del
producto por parte de la empresa responsable del envasado, etiquetado o
distribución.
30
226

30
Ejemplo 4.- Utilización del procedimiento de la invención para la
identificación de mezcla de especies.
Se planteó la posibilidad de utilizar esta técnica para la discriminación de
mezclas de especies.
5
Para preparar la mezcla de estudio, se procedió a la homogenización en partes
iguales de músculo de distintas especies de langostinos o camarones. Con
esta mezcla se realizó el procedimiento de extracción, digestión y separación
ya descrito en el Ejemplo 1 (etapas 1-3). Tras la monitorización SMIM en el
sistema LC-MS (etapa 3), se llevó a cabo la etapa 4. 10
4.- Identificación de las especies capturadas.
4.1.- Determinación de la presencia en la muestra de la Familia Penaeidae, la
Familia Solenoceridae o la familia Pandalidae. 15
Se comprobó la presencia o ausencia de las tres posibles combinaciones de
péptidos específicos de familia: S-PEN829 (SEQ ID NO: 1) para la Familia
Penaeidae; S-MUE643 (SEQ ID NO: 2) para la Familia Solenoceridae; y S-
BOR817 (SEQ ID NO3) para la Familia Pandalidae. 20
El resultado obtenido reveló que en la muestra existía una mezcla de
especimenes pertenecientes a las familias Penaeidae y Solenoceridae (Figura
4A).
25
4.2.- Determinación del origen geográfico y de la especie.
Una vez identificada en la muestra la mezcla de familias, se determinó la
procedencia geográfica de los langostinos Penaeidae. La ausencia del péptido
S-PEN759 (SEQ ID NO4) (Figura 4B) indicó que en la mezcla existían 30
langostinos del Atlántico.
227
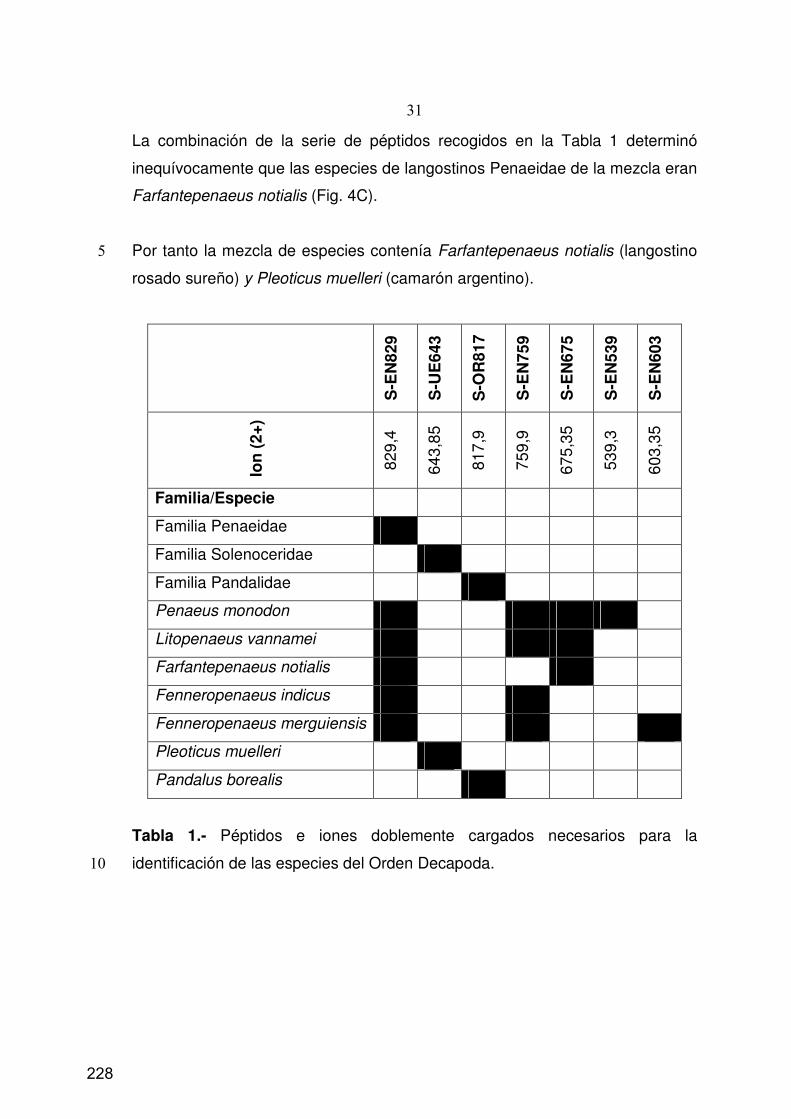
31
La combinación de la serie de péptidos recogidos en la Tabla 1 determinó
inequívocamente que las especies de langostinos Penaeidae de la mezcla eran
Farfantepenaeus notialis (Fig. 4C).
Por tanto la mezcla de especies contenía Farfantepenaeus notialis (langostino 5
rosado sureño) y Pleoticus muelleri (camarón argentino).
S-E
N82
9
S-U
E64
3
S-O
R81
7
S-E
N75
9
S-E
N67
5
S-E
N53
9
S-E
N60
3
Ion
(2+
)
829,
4
643,
85
817,
9
759,
9
675,
35
539,
3
603,
35
Familia/Especie
Familia Penaeidae
Familia Solenoceridae
Familia Pandalidae
Penaeus monodon
Litopenaeus vannamei
Farfantepenaeus notialis
Fenneropenaeus indicus
Fenneropenaeus merguiensis
Pleoticus muelleri
Pandalus borealis
Tabla 1.- Péptidos e iones doblemente cargados necesarios para la
identificación de las especies del Orden Decapoda. 10
228

32
REIVINDICACIONES
1. Procedimiento de obtención de datos útiles para la identificación de
especies de langostinos y camarones que comprende: 5
a. extraer las proteínas sarcoplásmicas del músculo blanco de una
muestra biológica aislada,
b. digerir las proteínas del extracto sarcoplásmico del paso (a), e
c. identificar la presencia o ausencia en el extracto digerido del paso
(b) de los péptidos que se seleccionan de la lista que comprende: 10
S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2), S-
BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675
(SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID
NO: 7).
15
2. Procedimiento para la identificación de especies de langostinos y
camarones que comprende los pasos (a) – (c) según la reivindicación
anterior, y además comprende:
d1. asignar la muestra biológica del paso (a) como perteneciente a la
especie Pleoticus muelleri de la Familia Solenoceridae, si el 20
péptido identificado en el paso c) es S-MUE643 (SEQ ID NO: 2).
3. Procedimiento para la identificación de especies de langostinos y
camarones que comprende los pasos (a) – (c) según la reivindicación 1, y
además comprende: 25
d2. asignar la muestra biológica del paso (a) como perteneciente a la
especie Pandalus borealis de la Familia Pandalidae, si el péptido
identificado en el paso (c) es S-BOR817 (SEQ ID NO: 3).
4. Procedimiento para la identificación de especies de langostinos y 30
camarones que comprende los pasos (a) – (c) según la reivindicación 1, y
además comprende:
229

33
d3. asignar la muestra biológica del paso (a) como perteneciente a la
Familia Penaeidae, si el péptido identificado en el paso (c) es S-
PEN829 (SEQ ID NO: 1).
5. Procedimiento para la identificación de especies de langostinos y 5
camarones según la reivindicación 4, que además comprende:
e1. asignar la muestra biológica del paso (a) como perteneciente a la
especie Farfantepenaeus notialis, si en el paso (c) se identifica la
ausencia del péptido S-PEN759 (SEQ ID NO: 4).
10
6. Procedimiento para la identificación de especies de langostinos y
camarones según la reivindicación 4, que además comprende:
e2. asignar la muestra biológica del paso (a) como perteneciente a la
especie P. monodon, si en el paso (c) se identifican los péptidos
S-PEN675 (SEQ ID NO: 5) y S-PEN539 (SEQ ID NO: 6). 15
7. Procedimiento para la identificación de especies de langostinos y
camarones según la reivindicación 4, que además comprende:
e3. asignar la muestra biológica del paso (a) como perteneciente a la
especie Litopenaeus vannamei, si en el paso (c) se identifica la 20
presencia del péptido S-PEN675 (SEQ ID NO: 5) y la ausencia del
péptido S-PEN539 (SEQ ID NO: 6).
8. Procedimiento para la identificación de especies de langostinos y
camarones según la reivindicación 4, que además comprende: 25
e4. asignar la muestra biológica del paso (a) como perteneciente a la
especie Fenneropenaeus merguiensis, si en el paso (c) se
identifica la presencia del péptido S-PEN603 (SEQ ID NO: 7) y la
ausencia de los péptidos S-PEN675 (SEQ ID NO: 5) y S-PEN539
(SEQ ID NO: 6). 30
230

34
9. Procedimiento para la identificación de especies de langostinos y
camarones según la reivindicación 4, que además comprende:
e5. asignar la muestra biológica del paso (a) como perteneciente a la
especie Fenneropenaeus indicus, si en el paso (c) se identifica la
presencia del péptido S-PEN759 (SEQ ID NO: 4) y la ausencia de 5
los péptidos S-PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO:
6), S-PEN603 (SEQ ID NO: 7).
10. Procedimiento según cualquiera de las reivindicaciones 1-9, donde la
extracción a partir del músculo blanco se realiza por homogeneización y 10
centrifugación de la muestra.
11. Procedimiento según la reivindicación anterior, donde la homogenización de
la muestra se realiza con un extractante de baja fuerza iónica.
15
12. Procedimiento según cualquiera de las reivindicaciones 9-11, donde la
centrifugación se realiza a 30.000 x g 10 min.
13. Procedimiento según cualquiera de las reivindicaciones 1-12, donde la
digestión de las proteínas del paso (b) se realiza con una enzima proteasa. 20
14. Procedimiento según la reivindicación anterior, donde la enzima es la
tripsina.
15. Procedimiento según cualquiera de las reivindicaciones 1-14, donde la 25
digestión de las proteínas del paso (b) se realiza mediante hidrólisis
química.
16. Procedimiento según cualquiera de las reivindicaciones 1-15, donde la
digestión de las proteínas del paso (b) se realiza mediante la aplicación de 30
ultrasonidos focalizados de alta intensidad.
231

35
17. Procedimiento según cualquiera de las reivindicaciones 1-16, donde la
identificación de los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ
ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-
PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID
NO: 7) se realiza mediante espectrometría de masas. 5
18. Procedimiento según la reivindicación anterior, donde la identificación de los
péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ ID NO: 2) y/o S-
BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-PEN675 (SEQ ID
NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID NO: 7) se realiza 10
mediante espectrometría de masas acoplada a cromatografía líquida de alta
resolución (HPLC).
19. Procedimiento según la reivindicación anterior, donde el espectrómetro de
masas acoplado a un sistema de cromatografía líquida de alta resolución 15
trabajan en cualquiera de los siguientes modos Data Dependent, SIM
(selected ion monitoring), SIRM (selected ion reaction monitoring) y SMIM
(selected MS/MS ion monitoring).
20. Procedimiento según cualquiera de las reivindicaciones 1-16, donde la 20
identificación de los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ
ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-
PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID
NO: 7) se realiza mediante un anticuerpo.
25
21. Procedimiento según cualquiera de las reivindicaciones 1-16, donde la
identificación de los péptidos S-PEN829 (SEQ ID NO: 1), S-MUE643 (SEQ
ID NO: 2) y/o S-BOR817 (SEQ ID NO: 3), S-PEN759 (SEQ ID NO: 4), S-
PEN675 (SEQ ID NO: 5), S-PEN539 (SEQ ID NO: 6), S-PEN603 (SEQ ID
NO: 7) se realiza mediante un oligonucleótido. 30
232

36
22. Péptido S-PEN829 cuya secuencia aminoacídica consiste en la SEQ ID NO:
1.
23. Péptido S-MUE643 cuya secuencia aminoacídica consiste en la SEQ ID
NO: 2. 5
24. Péptido S-BOR643 cuya secuencia aminoacídica consiste en la SEQ ID
NO: 3.
25. Péptido S-PEN759 cuya secuencia aminoacídica consiste en la SEQ ID NO: 10
4.
26. Péptido S-PEN675 cuya secuencia aminoacídica consiste en la SEQ ID NO:
5.
15
27. Péptido S-PEN539 cuya secuencia aminoacídica consiste en la SEQ ID NO:
6.
28. Péptido S-PEN603 cuya secuencia aminoacídica consiste en la SEQ ID NO:
7. 20
29. Uso de los péptidos según cualquiera de las reivindicaciones 22-28, para la
generación de anticuerpos.
30. Uso de los anticuerpos según la reivindicación anterior, para la identificación 25
de los péptidos según la reivindicación 20.
31. Kit para la identificación de especies langostinos y camarones que
comprende los medios necesarios para la identificación de los péptidos
según cualquiera de las reivindicaciones 22-28. 30
233

37
32. Kit según la reivindicación anterior que además comprende los medios
necesarios para llevar a cabo un procedimiento según cualquiera de las
reivindicaciones 1-21.
234
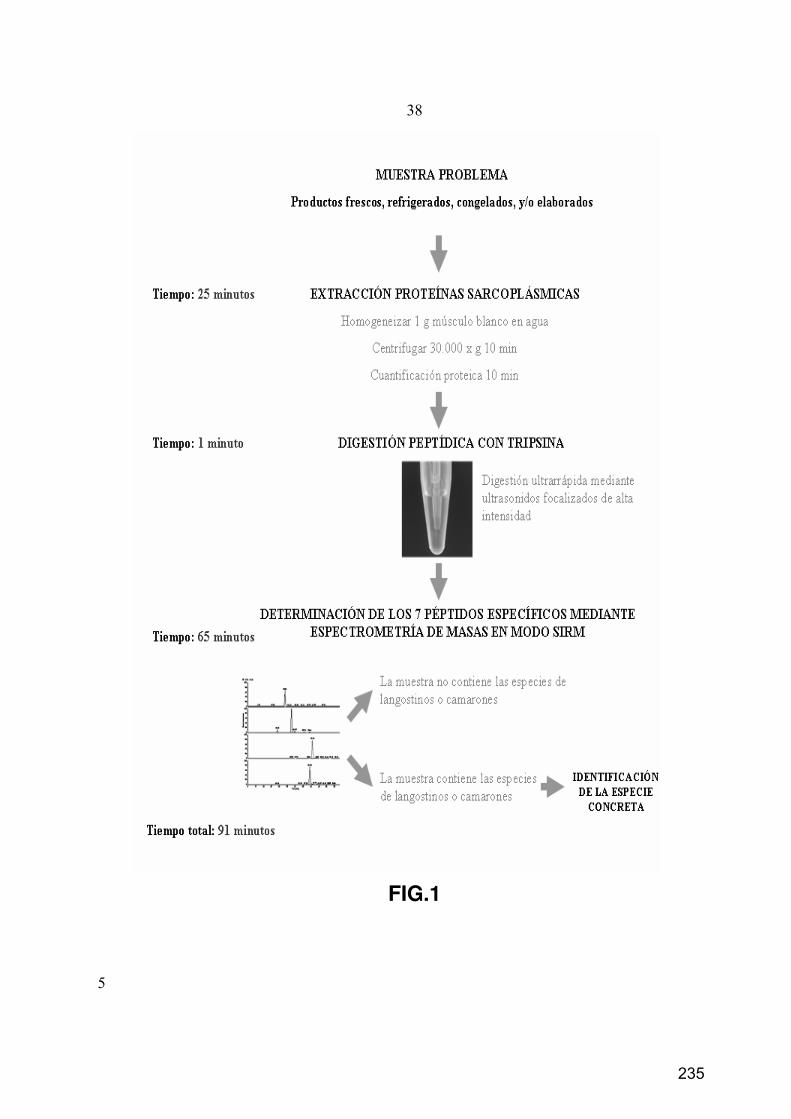
38
FIG.1
5
235
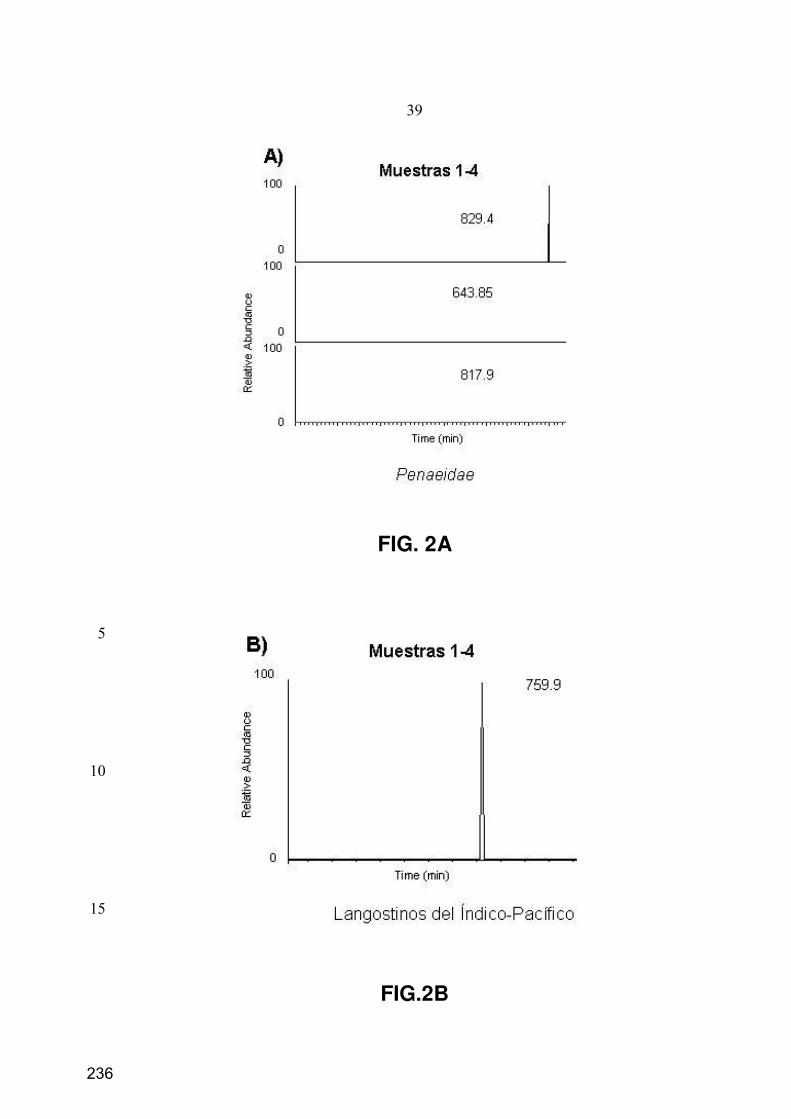
39
FIG. 2A
5
10
15
FIG.2B
236

40
5
10
15
FIG. 2C
20
25
30
237
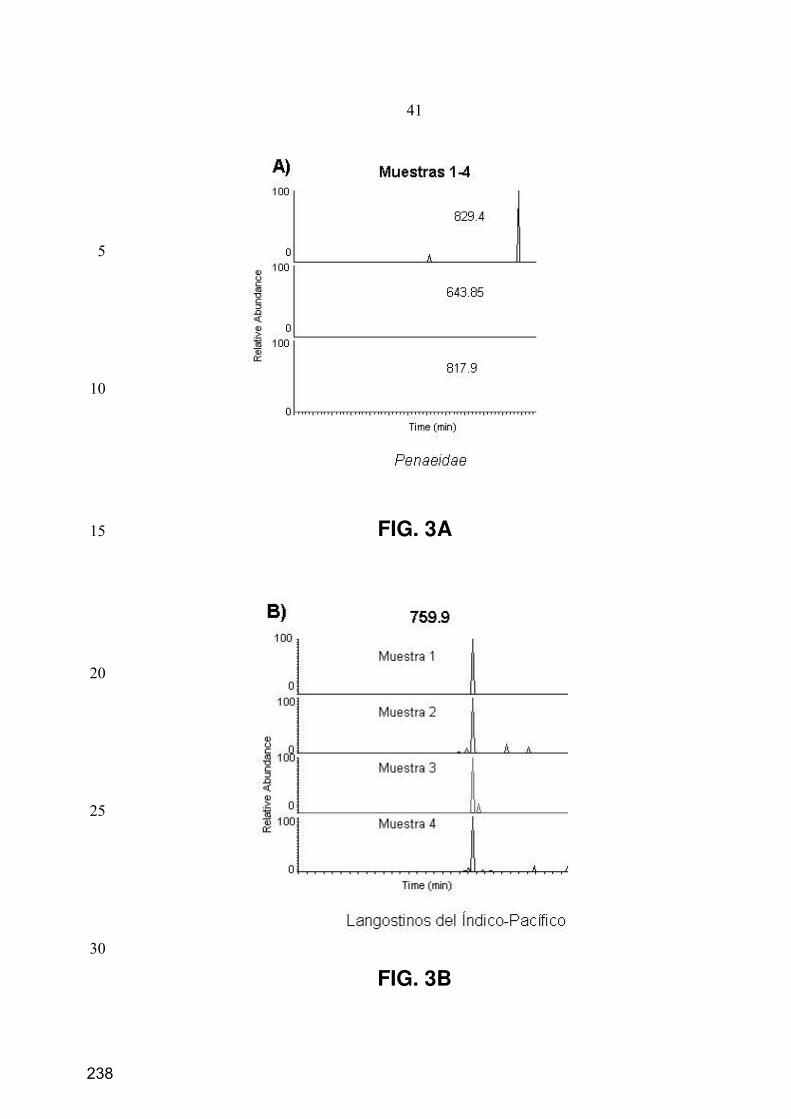
41
5
10
FIG. 3A 15
20
25
30
FIG. 3B
238
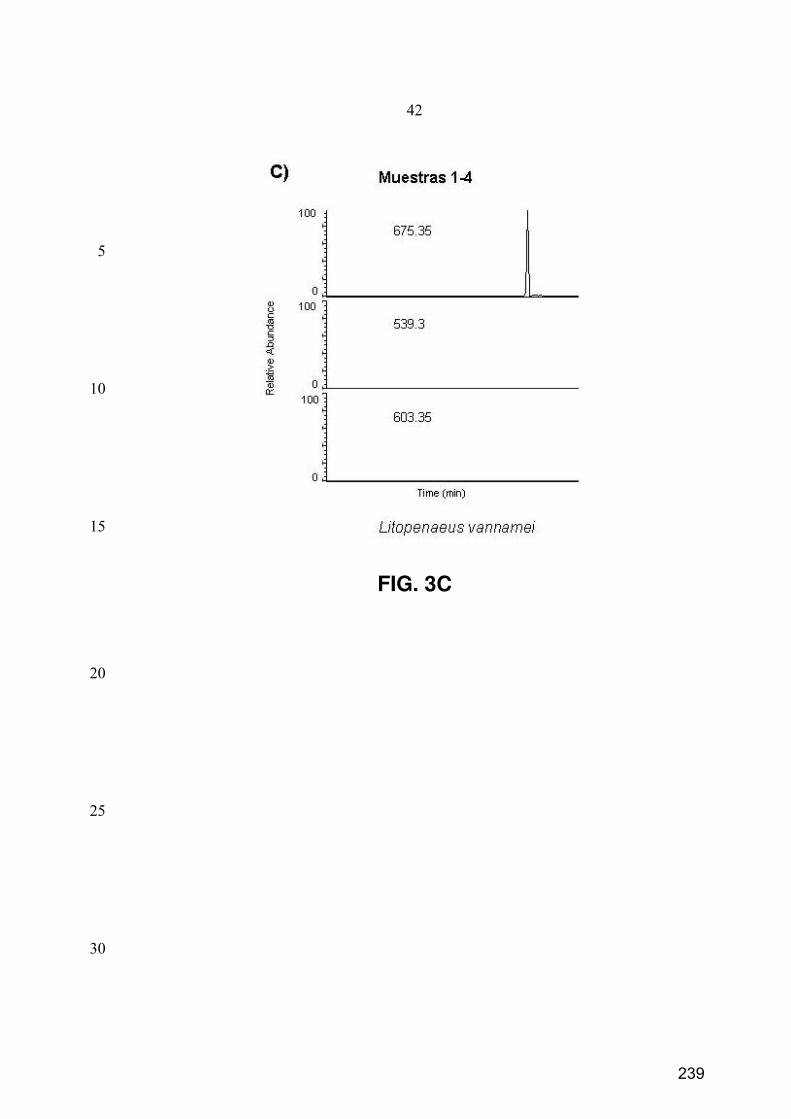
42
5
10
15
FIG. 3C
20
25
30
239
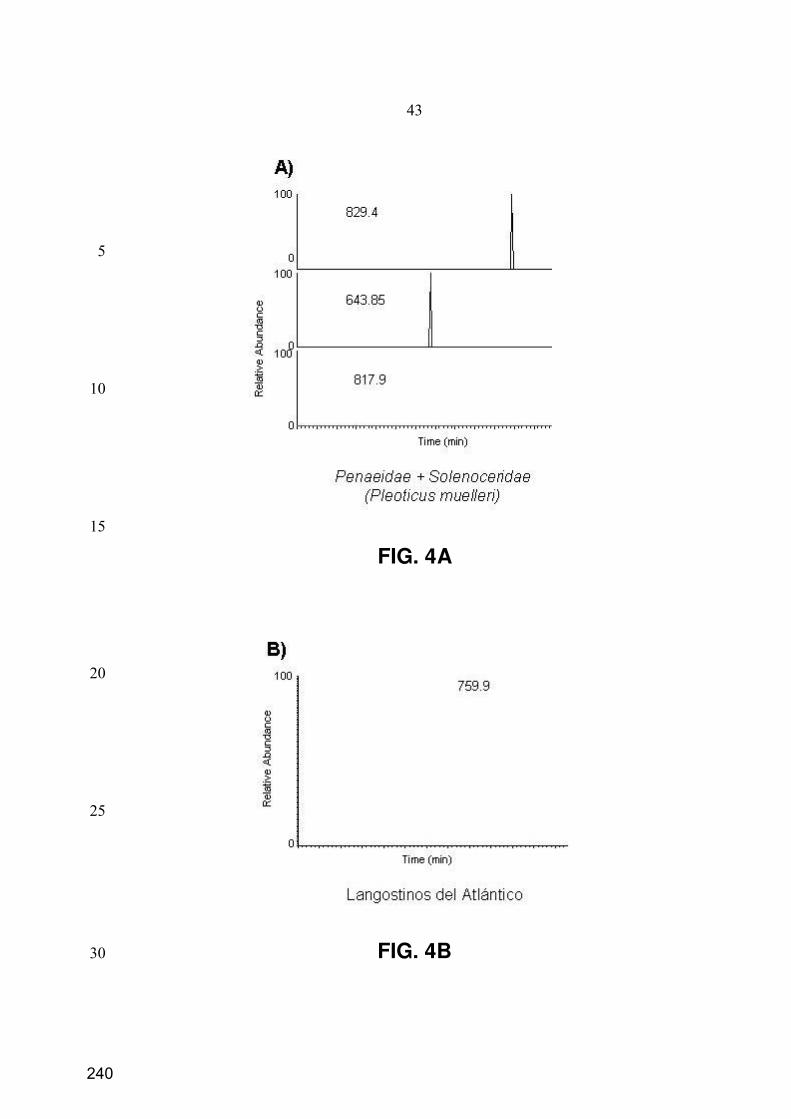
43
5
10
15
FIG. 4A
20
25
FIG. 4B 30
240
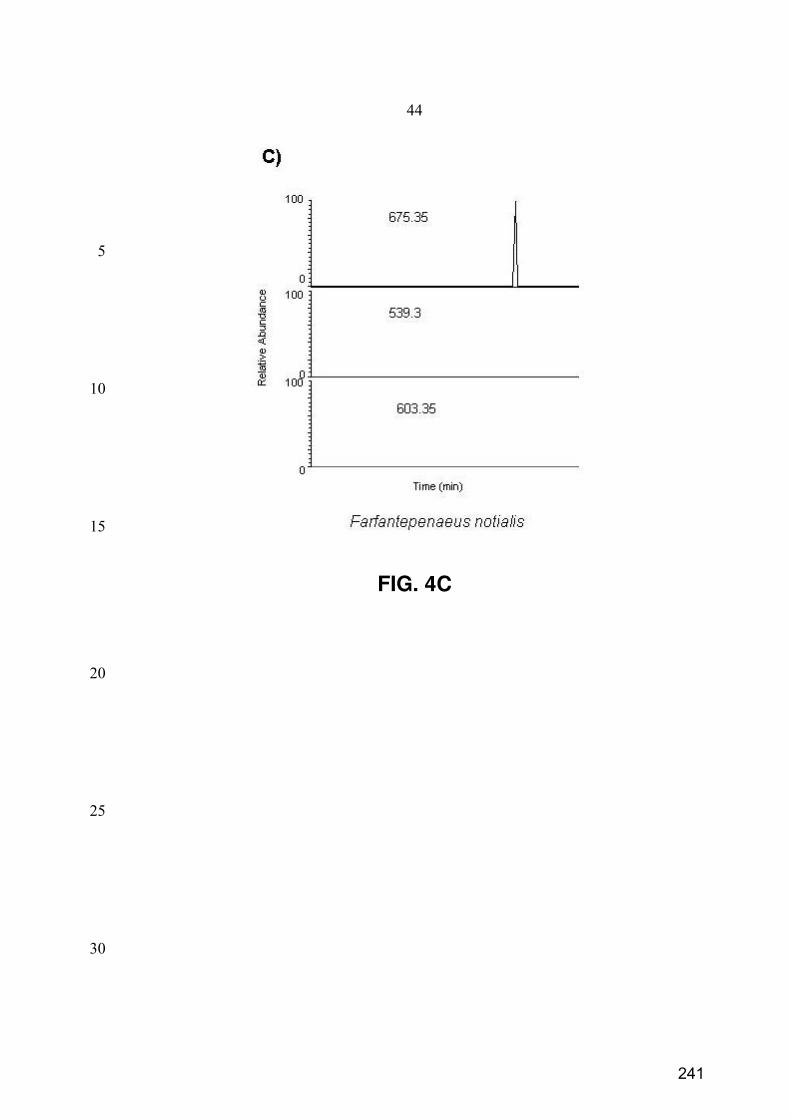
44
5
10
15
FIG. 4C
20
25
30
241
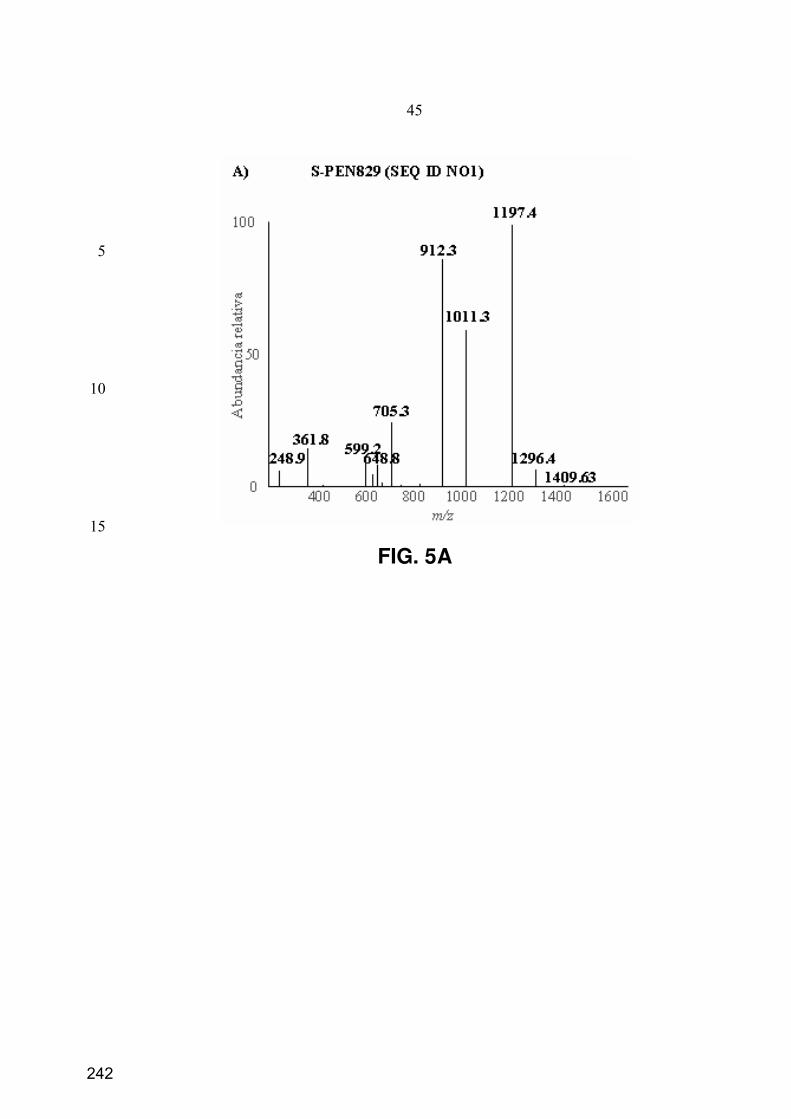
45
5
10
15
FIG. 5A
242

46
FIG. 5B
5
243
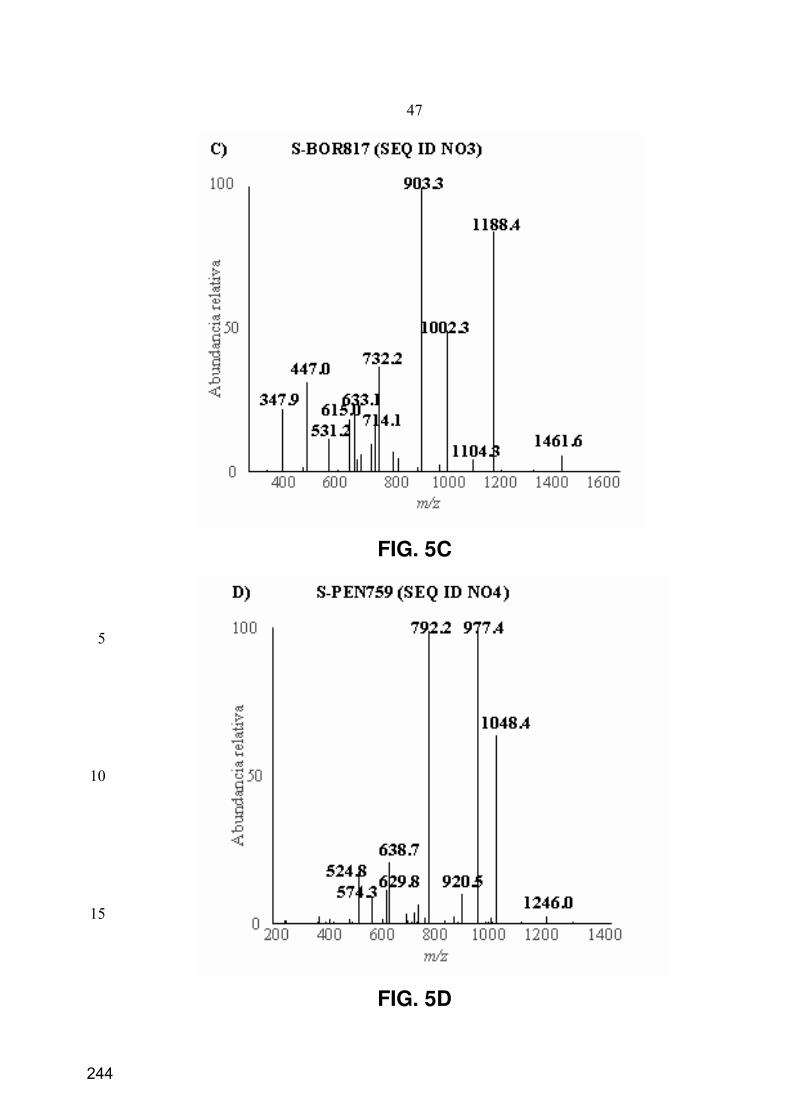
47
FIG. 5C
5
10
15
FIG. 5D
244
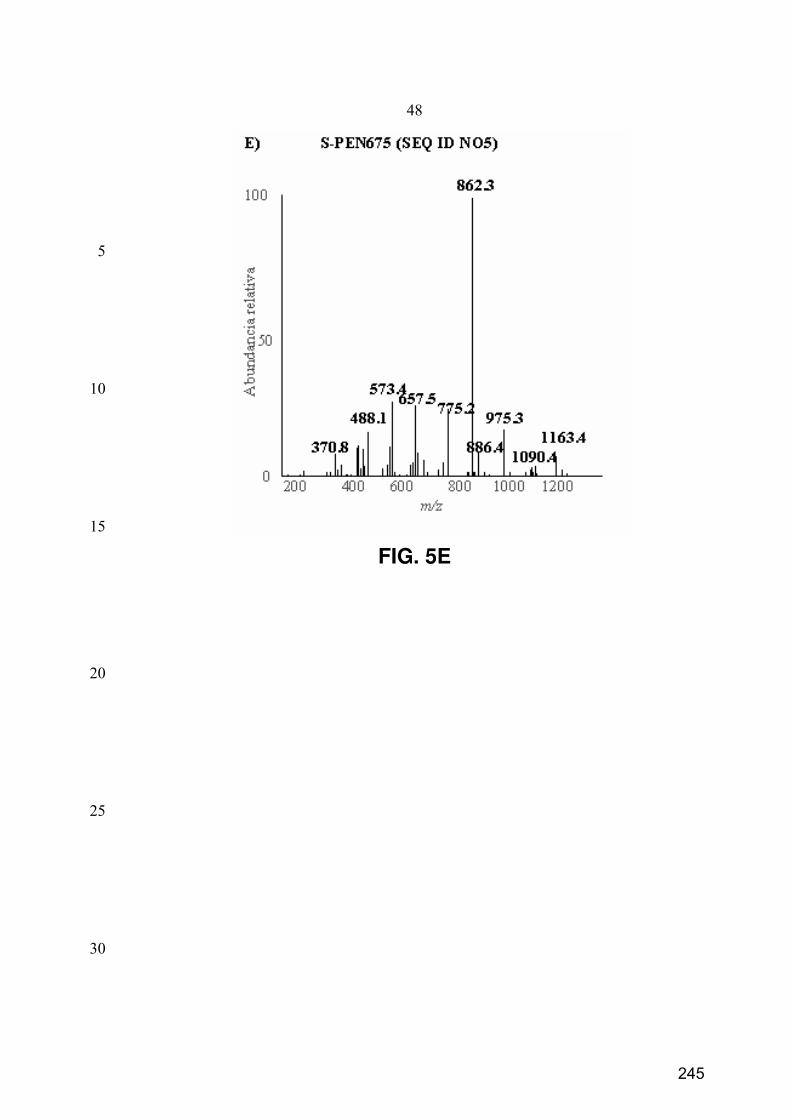
48
5
10
15
FIG. 5E
20
25
30
245
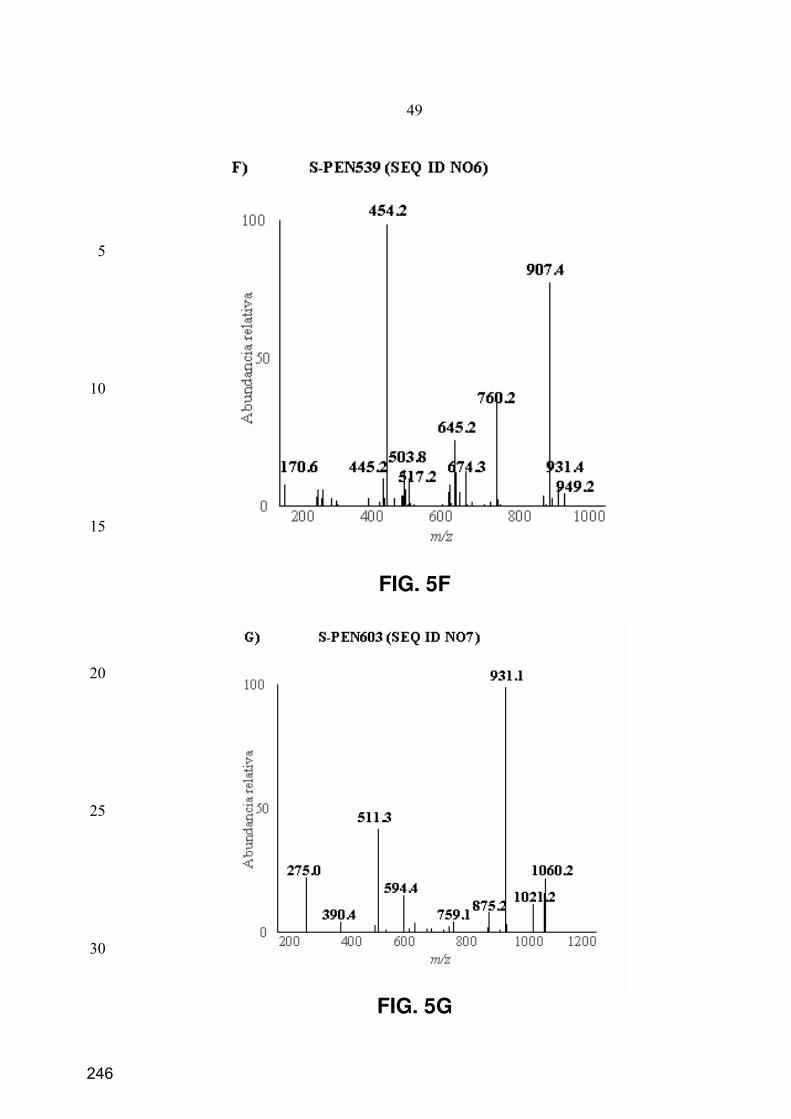
49
5
10
15
FIG. 5F
20
25
30
FIG. 5G
246

50
RESUMEN
Procedimiento y kit para la identificación de las principales especies
comerciales de langostinos y camarones.
5
Procedimiento y kit para la identificación de las principales especies
comerciales de langostinos o camarones (Orden Decapoda) pertenecientes a
las familias Penaeidae (Penaeus monodon, Farfantepenaeus notialis,
Fenneropenaeus indicus, Fenneropenaeus merguiensis, Litopenaeus
vannamei) Solenoceridae (Pleoticus muelleri) y Pandalidae (Pandalus borealis), 10
en productos y subproductos frescos, refrigerados, congelados y/o elaborados.
247

Página 1 LISTADO DE SECUENCIAS
<110> CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS (CSIC) <120> Procedimiento y kit para la identificación de las principales especies comerciales de langostinos y camarones.
<130> PEPT
<160> 7
<170> PatentIn version 3.5
<210> 1<211> 13<212> PRT<213> Artificial
<220><223> Péptido S-PEN829
<400> 1
Thr Phe Leu Val Trp Val Asn Glu Glu Asp His Leu Arg 1 5 10
<210> 2<211> 11<212> PRT<213> Artificial
<220><223> Péptido S-MUE643
<400> 2
Leu Thr Asn Ala Val Asn Glu Ile Glu Lys Arg 1 5 10
<210> 3<211> 13<212> PRT<213> Artificial
<220><223> Péptido S-BOR817
<400> 3
Ser Phe Leu Val Trp Val Asn Glu Glu Asp Gln Leu Arg 1 5 10
<210> 4<211> 13<212> PRT<213> Artificial
<220><223> Péptido S-PEN759
<400> 4
248

Página 2Leu Glu Glu Val Ala Gly Lys Tyr Asn Leu Gln Val Arg 1 5 10
<210> 5<211> 13<212> PRT<213> Artificial
<220><223> Péptido S-PEN675
<400> 5
Val Ser Ser Thr Leu Ser Ser Leu Glu Gly Glu Leu Lys 1 5 10
<210> 6<211> 9<212> PRT<213> Artificial
<220><223> Péptido S-PEN539
<400> 6
Ala Val Phe Asp Gln Leu Lys Glu Lys 1 5
<210> 7<211> 10<212> PRT<213> Artificial
<220><223> Péptido S-PEN603
<400> 7
Ala Leu Phe Asp Gln Leu Lys Asp Lys Lys 1 5 10
249


DISCUSIÓN


DISCUSIÓN INTEGRADORA
Los resultados experimentales recogidos en esta memoria han conducido al cumplimiento de los
objetivos propuestos. Se ha conseguido un avance considerable en el desarrollo de metodologías
robustas y eficaces para la identificación o autentificación de las principales especies comerciales de
langostinos y camarones. Aunque las discusiones de esta memoria han quedado reflejadas en cada uno
de los cinco capítulos anteriores, a efectos prácticos se han reunido dentro de este epígrafe con el
objetivo de integrarlas y reforzar los objetivos de los que partimos.
La autentificación alimentaria tiene en la actualidad gran relevancia en el sector pesquero, debido
a la sustitución, inadvertida o malintencionada, de unas especies por otras, en mayor medida cuando se
trata de productos elaborados en los que las características externas han sido eliminadas. Además, la
correcta identificación de los ingredientes de un producto es importante no sólo por razones económicas,
sino también por razones sanitarias, debido a la introducción de algún componente alergénico o tóxico
para algún sector de la población. Por todo ello, es necesario disponer de metodologías adecuadas para
identificar los componentes de los alimentos.
Aunque el IEF de la fracción proteica sarcoplásmica es el único método oficial validado para la
identificación de especies (AOAC, 1990), hasta el desarrollo de esta tesis se han hecho pocos esfuerzos
para describir un método de identificación de especies de langostinos utilizando esta técnica
electroforética. Por ello, en primer lugar el trabajo se centró en la búsqueda de patrones de bandas
diferenciales utilizando el IEF para la identificación de 14 especies de langostinos y camarones de
interés comercial. Los patrones de bandas que se obtuvieron en este trabajo permiten la diferenciación
inequívoca de cada una de las especies consideradas, debido a las isoformas de Sacoplasmic Calcium-
binding Protein (SCP), unas proteínas pertenecientes, como las parvalbúminas, a la familia de proteínas
de tipo mano-EF, descritas previamente como especie-específicas y termoestables (Rehbein, 1995;
Piñeiro y col., 1999b). Un estudio reciente (Shiomi y col., 2008) ha demostrado la alergenicidad de las
SCPs, por lo que se considera que el estudio de este grupo de proteínas es de gran importancia, no sólo
desde el punto de vista de la autenticidad alimentaria, sino también desde cualquier campo relacionado
con las alergias alimentarias, como pueden ser la detección de proteínas alergénicas en alimentos,
estudios de inmunogenicidad, o estudios de las reacciones cruzadas entre distintas especies o grupos de
especies.
Los protocolos que se han venido utilizando tradicionalmente para la identificación de especies
apenas han evolucionado en los últimos años, consistiendo básicamente en metodologías electroforéticas
y, desde los años 90, en estudios basados en el ADN. Sin embargo, los requisitos en cuanto a calidad,
253

Discusión
trazabilidad y seguridad alimentaria crecen constantemente, debido a la globalización del mercado
alimentario como consecuencia de los avances en los campos de la tecnología alimentaria (como las
mejoras en conservación) y de la distribución. Así, una de las limitaciones de las actuales técnicas
usadas para la detección e identificación de especies en alimentos es el tiempo que requiere el análisis.
Además, conseguir una correcta identificación frecuentemente depende del tratamiento que ha seguido
el producto, y para alimentos sometidos a tratamientos fuertes las mejores metodologías serían aquellas
que utilicen como marcadores moléculas no muy grandes, como pueden ser péptidos (Martínez y Friis,
2004). Por todo ello, nos planteamos el empleo de técnicas proteómicas para definir marcadores
peptídicos que puedan ser utilizados para la detección e identificación de las siete especies de
langostinos y camarones de mayor importancia económica. En los últimos años, las metodologías
proteómicas han sido ampliamente utilizadas en otras áreas, principalmente en biomedicina, pero son
minoritarios los trabajos proteómicos aplicados a tecnología de alimentos, en general, y a identificación
de especies, en particular.
El primer paso hacia la caracterización de marcadores peptídicos especie-específicos consistió en
identificar una proteína con la suficiente variabilidad inter-específica para que permitiera distinguir entre
especies filogenéticamente tan próximas como las consideradas, pero cuya variabilidad intra-específica
no complicara la identificación. Mediante el análisis de las proteínas sarcoplásmicas por 2-DE, se
encontraron diferencias entre las distintas especies estudiadas en el pI del spot correspondiente a la
arginina quinasa (AK). Además, otros autores ya habían descrito la existencia de variabilidad inter-
específica en Decápodos en la secuencia de la AK (France y col., 1997; Yu y col., 2003) y en el gen que
la codifica (Mahon y Neigel, 2008). Por ello, esta proteína fue seleccionada para los análisis por
espectrometría de masas. Un estudio previo demostró la capacidad de la técnica conocida como huella
peptídica (Peptide Mass Figerprinting, PMF), en la que se utiliza MALDI-TOF MS, para diferenciar
entre dos especies de langostinos muy próximas, tanto filogenéticamente como geográficamente, P.
monodon y F. indicus. En un estudio más amplio se consiguió diferenciar entre las siete especies
consideradas, a partir de la comparación de los espectros PMF de las isoformas de AK.
Para efectuar la comparación entre los espectros PMF de las distintas especies, se desarrolló un
método matemático que, por una parte nos proporciona la similitud numérica entre cada par de espectros
(con lo que podemos eliminar la subjetividad que conlleva la evaluación visual de los espectros), y por
otra parte nos proporciona un dendrograma o árbol de los espectros, construido en función de dicha
similitud. Este árbol fue validado por comparación con una filogenia establecida con datos de ADN
mitocondrial y resultó ser consistente con los árboles filogenéticos previos de otros autores, por lo que
se concluye que el método puede ser utilizado, además de para identificación de especies, para realizar
estudios filoproteómicos, complementado los estudios filogenéticos basados en las secuencias de ADN.
254

Discusión
Aunque Conway y col. (2001) introdujeron el término “filoproteómica” en relación al desarrollo y la
aplicación de metodologías proteómicas en estudios de evolución y filogenética, hasta el momento los
estudios realizados en este contexto son escasos (Navas y Albar, 2004). La metodología descrita en el
presente trabajo, en la que se comparan espectros de huella peptídica, es independiente de las bases de
datos, por lo que puede ser aplicada a especies que, como las especies marinas, estén poco
caracterizadas en las bases de datos genómicas y proteómicas, evitando los inconvenientes de tener que
secuenciar el ADN. Alm y col. (2006) desarrollaron un método similar de comparación de espectros
MALDI-TOF de huella peptídica y lo aplicaron a la identificación de isoformas de una misma proteína.
Sin embargo, nuestro estudio representa la primera aplicación de un método de este tipo que utilice
espectros PMF para inferir relaciones filogenéticas en cualquier tipo de organismo.
Una vez que los espectros MALDI-TOF de huella peptídica pusieron de manifiesto las diferencias
en las masas de algunos de los péptidos provenientes de la digestión de la AK, el siguiente paso
consistió en caracterizar dichos péptidos diferenciadores. Para ello se secuenció el mayor número
posible de péptidos de esta proteína, utilizando un espectrómetro de masas con trampa iónica para
obtener los espectros de la fragmentación. A partir de dichos espectros, bien mediante comparación con
los espectros teóricos de los péptidos disponibles en las bases de datos, o bien mediante secuenciación
de novo manual, se estableció la secuencia de un total de 131 péptidos, que permitieron reconstruir las
secuencias parciales de las siete isoformas de AK, correspondientes a cada una de las siete especies
estudiadas, además de dos isoformas más, correspondientes a dos poblaciones distintas dentro de dos de
las especies. Los péptidos diferenciadores caracterizados, muchos de ellos especie-específicos, pueden
ser usados como biomarcadores para la identificación de especies.
Hay que señalar que las técnicas proteómicas pueden ser automatizadas, produciendo resultados
rápidos y reproducibles y permitiendo el análisis de un alto número de muestras. Sin embargo, el gran
avance de esta aproximación proteómica es que constituye el primer paso hacia el diseño de sistemas de
detección baratos y fáciles de usar: los péptidos marcadores caracterizados, así como las secuencias
parciales de las AK obtenidas, podrían usarse para diseñar anticuerpos específicos, que podrían incluirse
en kits de análisis rápidos (como placas de ELISA o chips de proteínas), permitiendo una monitorización
sencilla y barata de un gran número de muestras por personal que no necesitaría un alto nivel de
preparación.
La identificación de péptidos diferenciadores, como se hace en este trabajo, puede usarse no sólo
para identificar taxones (como especies, géneros o familias), sino también para identificar proteínas de
interés. Por ello, la elección de la AK es particularmente interesante porque, además de discriminar
especies debido a su variabilidad, es también un alérgeno (Yu y col., 2003), y por tanto, un análisis
255

Discusión
orientado a esta proteína tendría una doble aplicación: en identificación de especies (si el péptido
seleccionado discrimina a nivel de especies), y en seguridad alimentaria (si el péptido seleccionado es
común para varias especies de interés, por ejemplo, especies que produzcan alergia alimentaria). Esto
permitiría la detección del alérgeno en productos alimentarios, y de esta manera ayudaría al fabricante,
al distribuidor, o a la administración, a identificar, en tiempo real, la existencia de una contaminación en
una línea de producción. Además, aunque la actividad de la AK se elimina al calentar a 60ºC el músculo
del decápodo (Rehbein, 1995), esta desnaturalización térmica no afecta a sus propiedades antigénicas
(Verrez y col., 1992), por lo que los anticuerpos contra esta proteína seguirían detectándola incluso en
muestras sometidas a tratamiento térmico.
La última etapa de este trabajo consistió en aplicar todo el conocimiento obtenido a través de la
secuenciación de las diferentes isoformas de AK al diseño de un método que permitiera realizar un
análisis de identificación o autentificación de especies robusto, sistemático, sencillo y de gran rapidez.
Por esta circunstancia, se desarrolló una metodología de monitorización de los péptidos diana,
previamente caracterizados, utilizando un espectrómetro de masas de trampa iónica trabajando en modo
SMIM. Acoplando on-line la trampa iónica a una separación cromatográfica capilar por fase reversa, es
posible separar y detectar varios péptidos diana en el digerido de una mezcla compleja de proteínas. Una
separación por HPLC de 65 minutos demostró ser suficiente para analizar una muestra de la complejidad
que supone el digerido del extracto sarcoplásmico del músculo abdominal de langostinos y camarones,
de forma que los péptidos diana pudieran ser detectados por la trampa iónica. Se constató que la etapa
limitante en un procedimiento que utilice esta separación y detección de péptidos, es la preparación de la
muestra, es decir, la obtención de los péptidos por digestión del extracto proteico, que supone, con los
actuales protocolos utilizados rutinariamente en proteómica, un mínimo de 12 horas (Kinter y Sherman,
2000). Para acelerar la digestión proteica se aplicaron ultrasonidos focalizados de alta intensidad
(HIFU). López-Ferrer y col. (2005) han desarrollado esta metodología para acelerar la digestión tríptica
de mezclas complejas de proteínas en disolución, demostrando que la eficiencia de la digestión obtenida
aplicando un minuto los ultrasonidos es comparable a la obtenida con los protocolos tradicionales. La
aplicación de los ultrasonidos en el campo alimentario podría ser una aportación clave en los años
venideros, teniendo además una aplicación potencial en otros sectores, como el farmacéutico, que
también hacen uso de enzimas en sus procesos de producción. En el trabajo que aquí se presenta se
demuestra que, combinando esta digestión ultrarrápida con la monitorización de péptidos diana
utilizando la técnica SMIM, se consigue la identificación inequívoca de las especies consideradas en un
tiempo total de 91 minutos. Hay que tener en cuenta que un análisis genético de identificación de
especies, como los que hay descritos para especies de langostinos (Pascoal y col., 2008a), consistente en
extracción del ADN, amplificación por PCR, y estudio de RFLPs, se extiende por término medio
durante uno o dos días, a lo que habría que añadir aún más tiempo si hay que secuenciar el material
256

Discusión
genético. Por tanto el ahorro de tiempo que se consigue con el método que aquí se presenta supone un
gran avance respecto a cualquier otro método disponible en la actualidad para la autentificación de
especies de langostinos y camarones, ya sean métodos electroforéticos o genéticos. El método
desarrollado ha sido patentado para su aplicación en el control de origen y autenticidad de las principales
especies comerciales de langostinos y camarones a nivel internacional.
Próximamente, se plantea reducir aún más el tiempo del análisis de autentificación de especies.
Para ello, se seguirá utilizando la digestión del extracto proteico mediante ultrasonidos focalizados de
alta intensidad, pero se evitará la separación por fase reversa previa a la monitorización por SMIM,
utilizando una fuente de nanoelectrospray (nESI) estático. La eliminación de la separación
cromatográfica permitirá reducir los 65 minutos de la carrera de HPLC a 5 minutos de análisis con nESI,
lo que dejará la duración total del análisis, desde que llega la muestra hasta que se identifica la especie,
en 30 minutos, incluida la preparación de la muestra. Aunque tiene la ventaja de este corto tiempo de
análisis, esta detección con nanoESI presenta inconvenientes: por un lado la posible presencia de
sustancias que interfieran en la ionización y detección de los péptidos marcadores. Además, desde el
punto de vista técnico, es menos automatizable y de manejo más complicado, requiriendo una mayor
preparación técnica.
Para concluir, señalar que los métodos de identificación de especies aquí descritos, así como el
conocimiento generado y recogido en la presente Tesis Doctoral, tienen aplicación, además de en
investigación científica, en los sectores alimentario, importador, sanitario, y de control de calidad:
tecnología de productos pesqueros, control e inspección de la calidad y el origen de materias primas,
control y cumplimiento del etiquetado correcto de productos y subproductos establecido por la
legislación actual, control del fraude al consumidor y/o al importador, gestión de pesquerías, y conflictos
pesqueros en aguas internacionales. El hecho de que el 2% de la población sea alérgica al marisco
(Sampson, 2004), confiere a los estudios sobre la SCP y la AK un interés biosanitario adicional, debido
al carácter alergénico de ambas proteínas.
De cara al futuro, podemos avanzar que la Proteómica, con la utilización de estas metodologías,
será una herramienta clave en los procesos de control de calidad basados en proteínas.
257


CONCLUSIONES


CONCLUSIONES
1. La separación de un extracto acuoso de proteínas sarcoplásmicas del músculo abdominal mediante
la técnica, rápida y sencilla, de isoelectroenfoque, permitió detectar bandas diferenciadoras, que se
identificaron como isoformas de SCPs. Estas proteínas son homólogas a las parvalbúminas, que se
utilizan normalmente como marcadores especie-específicos en pescados. Las diferencias en la
movilidad electroforética de las SCPs identificadas permiten la clasificación e identificación de 14
especies comerciales de langostinos y camarones en un solo día de trabajo.
2. El análisis mediante 2-DE de las proteínas sarcoplásmicas del músculo abdominal de siete
especies de langostinos y camarones de gran interés comercial, ha demostrado, a efectos de
identificación de especies, la existencia de una proteína potencialmente diferenciadora, la arginina
quinasa (AK), ya que presenta variaciones en su movilidad electroforética.
3. Los mapas peptídicos de los digeridos trípticos de las AKs, obtenidos mediante MALDI-TOF MS,
presentaron patrones diferenciales, consistentes en una serie de picos comunes y otros específicos
para una o varias especies. Estos patrones pueden ser utilizados para diferenciar claramente entre
las dos superfamilias, las tres familias, y las siete especies estudiadas.
4. Se ha desarrollado un método para la comparación matemática de los mapas peptídicos. Dicho
método se validó para dos tipos de aplicaciones, (i) la identificación de especies, mediante el
análisis de muestras problema, y (ii) el análisis filogenético, mediante la comparación del árbol
filoproteómico obtenido con un árbol filogenético construido a partir del material genético.
5. El análisis por espectrometría de masas en tándem de los digeridos trípticos de las AKs, seguido
por secuenciación manual o por identificación en bases de datos, permitió la caracterización de un
total de 131 secuencias peptídicas susceptibles de ser utilizadas para la clasificación e
identificación de las siete especies de langostinos y camarones estudiadas. En dos de estas
especies, L. vannamei y F. indicus, se encontró variabilidad intra-específica, permitiendo los
péptidos secuenciados distinguir entre dos poblaciones de la misma especie pero de distinto origen
geográfico.
6. Se logró la secuenciación parcial de un total de nueve isoformas de AKs, específicas para cada
una de las siete especies estudiadas –P. monodon, L. vannamei, F. notialis, F. indicus, F.
merguiensis, P. muelleri y P. borealis- y las dos poblaciones diferentes para L. vannamei y F.
indicus.
261

Conclusiones
7. Se ha desarrollado un procedimiento mediante el cual, tras una digestión ultrarrápida con
aplicación de ultrasonidos focalizados de alta intensidad y una separación y monitorización de
siete péptidos diferenciales mediante LC-MS trabajando en modo SMIM, se logra la identificación
inequívoca de las siete especies comerciales principales de langostinos y camarones del orden
Decapoda en 90 minutos, en cualquier producto ya sea fresco, refrigerado, congelado y /o
procesado.
262

BIBLIOGRAFÍA


BIBLIOGRAFÍA
Abercrombie, D., Clarke, S., Shivji, M. (2005). Global-scale genetic identification of hammerhead
sharks: Application to assessment of the international fin trade and law enforcement. Conserv.
Genet., 6, 775-788.
Ahmed, M. M. M., Ali, B. A., Ei-Zaeem, S. Y. (2004). Application of RAPD markers in fish: Part I -
some genera (Tilapia, Sarotherodon and Oreochromis) and species (Oreochromis aureus and
Oreochromis niloticus) of Tilapia. Int. J. Biotechnology, 6, 86-93.
Alm, R., Johansson, P., Hjerno, K., Emanuelsson, C., Ringner, M., Hakkinen, J. (2006). Detection and
identification of protein isoforms using cluster analysis of MALDI-MS Mass Spectra.
J.Proteome Res., 5, 785-792.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., Lipman, D. J. (1990). Basic local alignment search
tool. J. Mol. Biol., 215, 403-410.
An, H., Marshall, M. R., Otwell, W. S., Wei, C. -I. (1988). Electrophoretic identification of raw and
cooked shrimp using various protein extraction systems. J. Food Sci., 53, 313-318.
An, H., Wei, C. I., Zhao, J., Marshall, M. R., & Lee, C. M. (1989a). Electrophoretic identification of fish
species used in surimi products. J. Food Sci., 54, 253-257.
An, H., Marshall, M. R., Otwell, W. S., Wei, C. -I. (1989b). Species identification of raw and boiled
shrimp by a urea gel isoelectric focusing technique. J. Food Sci., 54, 233-236.
An, H., Klein, P. A., Kao, K. J., Marshall, M. R., Otwell, W. S., Wei, C. -I. (1990). Development of
monoclonal antibody for rock shrimp identification using enzyme-linked immunosorbent assay.
J. Agr. Food Chem., 38, 2094-2100.
AOAC, Association of Analytical Communities (1990). En: K. Helrich (ed.), Official Methods of
Analysis, vol. 2. Ed. Arlington.
Appel, R. D., Hochstrasser, D. F., Funk, M., Vargas, J. R., Pellegrini, C., Muller, A. F., Scherrer, J.-R.
(1991). The MELANIE project: from a biopsy to automatic protein map interpretation by
computer. Electrophoresis, 12, 722-735.
Apraiz, I., Mi, J., Cristobal, S. (2006). Identification of proteomic signatures of exposure to marine
pollutants in mussels (Mytilus edulis). Mol. Cell. Proteomics, 5, 1274-1285.
Aranishi, F. (2005). Rapid PCR-RFLP method for discrimination of imported and domestic mackerel.
Mar. Biotechnol., 7, 571-575.
Aranishi, F., Okimoto, T., Izumi, S. (2005a). Identification of gadoid species (Pisces, Gadidae) by PCR-
RFLP analysis. J. Appl. Genet., 46, 69-73.
Aranishi, F., Okimoto, T., Ohkumbo, M., Izumi, S. (2005b). Molecular identification of commercial
spicy pollack roe products by PCR-RFLP analysis. J. Food Sci., 70, C235-C238.
265

Bibliografía
Armstrong, S. G., Leach, D. N. (1992). The use of HPLC protein profiles in fish species indentification.
Food Chem., 44, 147.
Asensio, L., González, I., Fernández, A., Céspedes, A., Rodríguez, M. A., Hernández, P. E., García, T.
(2001a). Identification of Nile perch (Lates niloticus), grouper (Epinephelus guaza), and wreck
fish (Polyprion americanus) fillets by PCR amplification of the 5S rDNA gene. J. AOAC Int.,
84, 777-781.
Asensio, L., Gonzalez, I., Fernandez, A., Rodriguez, M. A., Hernandez, P. E., Garcia, T., Martin, R.
(2001b). PCR-SSCP: a simple method for the authentication of grouper (Epinephelus guaza),
wreck fish (Polyprion americanus), and Nile perch (Lates niloticus) fillets. J. Agr. Food Chem.,
49, 1720-1723.
Asensio, L., González, I., Rodríguez, M. A., Hernández, P. E., García, T., Martín, R. (2003a).
Development of a monoclonal antibody for grouper (Epinephelus marginatus) and wreck fish
(Polyprion americanus) authentication using an indirect ELISA. J. Food Sci., 68, 1900-1903.
Asensio, L., Gonzalez, I., Rodriguez, M. A., Mayoral, B., Lopez-Calleja, I., Hernandez, P. E., Garcia,
T., Martin, R. (2003b). Identification of grouper (Epinephelus guaza), wreck fish (Polyprion
americanus), and Nile perch (Lates niloticus) fillets by polyclonal antibody-based Enzyme-
Linked Immunosorbent Assay. J. Agr. Food Chem., 51, 1169-1172.
Asensio, L. (2008). Application of multiplex PCR for the identification of grouper meals in the
restaurant industry. Food Control, 19, 1096-1099.
Baldwin, J. D., Bass, A. L., Bowen, B. W., Clark, W. H. (1998). Molecular phylogeny and biogeography
of the marine shrimp Penaeus. Mol. Phylogenet. Evol., 10, 399-407.
Barnes, S., Kim, H. (2004). Nutriproteomics: identifying the molecular targets of nutritive and non-
nutritive components of the diet. J. Biochem. Mol. Biol., 37, 59-74.
Bartlett, S. E., Davidson, W. S. (1992). FINS (forensically informative nucleotide sequencing): a
procedure for identifying the animal origin of biological specimens. Biotechniques, 12, 408-411.
Berrini, A., Tepedino, V., Borromeo, V., Secchi, C. (2006). Identification of freshwater fish
commercially labelled "perch" by isoelectric focusing and two-dimensional electrophoresis.
Food Chem., 96, 163-168.
Blanco, M., Pérez-Martín, R. I., Sotelo, C. G. (2008). Identification of shark species in seafood products
by forensically informative nucleotide sequencing (FINS). J. Agr. Food Chem., 56, 9868-9874.
Booy, A. T., Haddow, J. D., Ohlund, L. B., Hardie, D. B., Olafson, R. W. (2005). Application of isotope
coded affinity tag (ICAT) analysis for the identification of differentially expressed proteins
following infection of Atlantic salmon (Salmo salar) with infectious hematopoietic necrosis
virus (IHNV) or Renibacterium salmoninarum (BKD). J. Proteome Res., 4, 325-334.
Bowman, T. E., Abele, L. G. (1982). The Biology of the Crustacea, Vol 1. Systematics, the Fossil
Record and Biogeography. New York: Academic Press.
266

Bibliografía
Brzezinski, J. L. (2005). Detection of crustacean DNA and species identification using a PCR-restriction
fragment length polymorphism method. J. Food Protect., 68, 1866-1873.
Calo-Mata, P., Sotelo, C. G., Pérez-Martín, R. I., Rehbein, H., Hold, G. L., Russell, V. J., Pryde, S.,
Quinteiro, J., Rey-Méndez, M., Rosa, C., Santos, A. T. (2003). Identification of gadoid fish
species using DNA-based techniques. Eur. Food Res. Technol., 217, 259-264.
Calo-Mata, P., Pascoal, A., Prado, M., Cepeda, A., Barros-Velázquez, J. Patente Nº
PCT/ES2007/070212.
Callejas, C., Ochando, M. D. (2001). Molecular identification (RAPD) of the eight species of the genus
Barbus (Cyprinidae) in the Iberian Peninsula. J. Fish Biol., 59, 1589-1599.
Carrera, E., Martín, R., García, T., González, I., Sanz, B., Hernández, P. E. (1996). Development of an
enzyme-linked immunosorbent assay for the identification of smoked salmon (Salmo salar),
trout (Oncorhynchus mykiss) and bream (Brama rail). J. Food Protect., 59, 521-524.
Carrera, E., García, T., Céspedes, A., González, I., Sanz, B., Hernández, P. E., Martín, R. (1997).
Immunostick colorimetric ELISA assay for the identification of smoked salmon, trout and
bream. J. Sci. Food Agr., 74, 547-550.
Carrera, E., Garcia, T., Céspedes, A., González, I., Fernández, A., Hernández, P. E., Martin, R. (1999).
Salmon and trout analysis by PCR-RFLP for identity authentication. J. Food Sci., 64, 410-413.
Carrera, M., Cañas, B., Piñeiro, C., Vázquez, J., Gallardo, J. M. (2006). Identification of commercial
hake and grenadier species by proteomic analysis of the parvalbumin fraction. Proteomics, 6,
5278-5287.
Carrera, M., Canas, B., Piñeiro, C., Vázquez, J., Gallardo, J. M. (2007). De novo mass spectrometry
sequencing and characterization of species-specific peptides from nucleoside diphosphate kinase
B for the classification of commercial fish species belonging to the family Merlucciidae. J.
Proteome Res., 6, 3070-3080.
Céspedes, A., García, T., Carrera, E., González, I., Sanz, B., Hernández, P. E., Martín, R. (1998a).
Polymerase chain reaction-restriction fragment length polymorphism analysis of a short
fragment of the cytochrome b gene for identification of flatfish species. J. Food Protect., 61,
1684-1685.
Céspedes, A., García, T., Carrera, E., González, I., Sanz, B., Hernáz, P. E., Martín, R. (1998b).
Identification of flatfish species using polymerase chain reaction (PCR) amplification and
restriction analysis of the cytochrome b gene. J. Food Sci., 63, 206-209.
Cespedes, A., Garcia, T., Carrera, E., Gonzalez, I., Fernandez, A., Hernandez, P. E., Martin, R. (1999a).
Identification of sole (Solea solea) and Greenland halibut (Reinhardtius hippoglossoides) by
PCR amplification of the 5S rDNA gene. J. Agr. Food Chem., 47, 1046-1050.
267

Bibliografía
Céspedes, A., García, T., Carrera, E., González, I., Fernández, A., Hernández, P. E., Martín, R. (1999b).
Application of polymerase chain reaction-single strand conformational polymorphism (PCR-
SSCP) to identification of flatfish species. J. AOAC Int., 82, 903-907.
Céspedes, A., García, T., Carrera, E., González, I., Fernández, A., Asensio, L., Hernández, P. E., Martín,
R. (1999c). Indirect enzyme-linked immunosorbent assay for the identification of sole (Solea
solea), european plaice (Pleuronectes platessa), flounder (Platichthys flesus), and Greenland
halibut (Reinhardtius hippoglossoides). J. Food Protect., 62(10), 1178-1182.
Céspedes, A., García, T., Carrera, E., González, I., Fernández, A., Asensio, L., Hernández, P. E., Martín,
R. (2000). Genetic differentiation between sole (Solea solea) and Greenland halibut
(Reinhardtius hippoglossoides) by PCR-RFLP analysis of a 12S rRNA gene fragment. J. Sci.
Food Agr., 80, 29-32.
Chakraborty, A., Aranishi, F., Iwatsuki, Y. (2005). Molecular identification of hairtail species (Pisces:
Trichiuridae) based on PCR-RFLP analysis of the mitochondrial 16S rRNA gene. J. Appl.
Genet., 46, 381-385.
Chapela, M. J., Sotelo, C. G., Calo-Mata, P., Pérez-Martín, R. I., Rehbein, H., Hold, G. L., Quinteiro, J.,
Rey-Méndez, M., Rosa, C., Santos, A. T. (2002). Identification of cephalopod species
(Ommastrephidae and Loliginidae) in seafood products by forensically informative nucleotide
sequencing (FINS). J. Food Sci., 67, 1672-1676.
Chapela, M., Sotelo, C., Pérez-Martín, R. (2003). Molecular identification of cephalopod species by
FINS and PCR-RFLP of a cytochrome b gene fragment. Eur. Food Res. Technol., 217, 524-529.
Chapela, M. J., Sanchez, A., Suarez, M. I., Perez-Martin, R. I., Sotelo, C. G. (2007). A rapid
methodology for screening hake species (Merluccius spp.) by single-stranded conformation
polymorphism analysis. J. Agr. Food Chem., 55, 6903-6909.
Chen, I.-C., Chapman, F. A., Wei, C.-l., Portier, K. M., O'Keefe, S. F. (1995). Differentiation of cultured
and wild sturgeon (Acipenser oxyrinchus desotoi) based on fatty acid composition. J. Food Sci.,
60, 631-635.
Chen, T.-Y., Hwang, D. F. (2002). Electrophoretic identification of muscle proteins in 7 puffer species.
J. Food Sci., 67, 936-942.
Chen, T.-Y., Shiau, C.-Y., Noguchi, T., Wei, C.-I., Hwang, D.-F. (2003). Identification of puffer fish
species by native isoelectric focusing technique. Food Chem., 83, 475-479.
Chen, T.-Y., Shiau, C.-Y., Wei, C.-I., Hwang, D.-F. (2004). Preliminary study on puffer fish proteome
species identification of puffer fish by two-dimensional electrophoresis. J. Agr. Food Chem., 52,
2236-2241.
Chou, C.-C., Lin, S.-P., Lee, K.-M., Hsu, C.-T., Vickroy, T. W., Zen, J.-M. (2007). Fast differentiation
of meats from fifteen animal species by liquid chromatography with electrochemical detection
using copper nanoparticle plated electrodes. J. Chromatogr. B, 846, 230-239.
268

Bibliografía
Chow, S., Nohara, K., Tanabe, T., Itoh, T., Tsuji, S., Nishikawa, Y., Uyeyanagi, S., Uchikawa, K.
(2003). Genetic and morphological identification of larval and small juvenile tunas (Pisces:
Scombridae) caught by a mid-water trawl in the western Pacific. Bull. Fish. Res. Agen., 8, 1-14.
Civera, T., Parisi, E. (1991). The use of sodium dodecyl sulfate polyacrylamide gel electrophoresis for
the identification of raw and cooked shrimps and crabs. Ital. J. Food Sci., 3, 149-157.
Clarke, S. C., Magnussen, J. E., Abercrombie, D. L., Mcallister, M. K., Shivji, M. S. (2006).
Identification of shark species composition and proportion in the Hong Kong shark fin market
based on molecular genetics and trade records. Conserv. Biol., 20, 201-211.
Colombo, F., Cerioli, M., Maria Colombo, M., Marchisio, E., Malandra, R., Renon, P. (2002). A simple
polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method for
the differentiation of cephalopod mollusc families Loliginidae from Ommastrephidae, to avoid
substitutions in fishery field. Food Control, 13(3), 185-190.
Colombo, F., Mangiagalli, G., Renon, P. (2005). Identification of tuna species by computer-assisted and
cluster analysis of PCR-SSCP electrophoretic patterns. Food Control, 16, 51-53.
Comesaña, A. S., Abella, P., Sanjuan, A. (2003). Molecular identification of five commercial flatfish
species by PCR-RFLP analysis of a 12S rRNA gene fragment. J. Sci. Food Agr., 83, 752-759.
Comi, G., Iacumin, L., Rantsiou, K., Cantoni, C., Cocolin, L. (2005). Molecular methods for the
differentiation of species used in production of cod-fish can detect commercial frauds. Food
Control, 16, 37-42.
Conway, G. C., Smole, S. C., Sarracino, D. A., Arbeit, R. D., Leopold, P. E. (2001). Phyloproteomics:
species identification of Enterobacteriaceae using matrix-assisted laser desorption/ionization
time-of-flight mass spectrometry. J. Mol. Microbiol. Biotechnol., 3, 103-112.
Cowie, W. P. (1968). Identification of fish species by thin-slab polyacrylamide gel electrophoresis of the
muscle myogens. J. Sci. Food Agr., 19, 226-229.
Devereux, R., Rublee, P., Paul, J., Field, K., Domingo, J. (2006). Development and applications of
microbial ecogenomic indicators for monitoring water quality: report of a workshop assessing
the state of the science, research needs and future directions. Environ. Monit. Assess., 116, 459-
479.
Directiva 2000/13/CE del Parlamento Europeo y del Consejo, de 20 de marzo de 2000, relativa a la
aproximación de las legislaciones de los Estados miembros en materia de etiquetado,
presentación y publicidad de los productos alimenticios.
Directiva 2003/89/CE del Parlamento Europeo y del Consejo de 10 de noviembre de 2003 por la que se
modifica la Directiva 2000/13/CE en lo que respecta a la indicación de los ingredientes
presentes en los productos alimenticios.
269

Bibliografía
Domínguez, E., Perez, M. D., Puyol, P., Calvo, M. (1997). Use of immunological techniques for
detecting species substitution in raw and smoked fish. Z. Lebensm. Unters. Forsch. A, 204, 279-
281.
Dooley, J. J., Sage, H. D., Brown, H. M., Garrett, S. D. (2005a). Improved fish species identification by
use of lab-on-a-chip technology. Food Control, 16, 601-607.
Dooley, J. J., Sage, H. D., Clarke, M.-A. L., Brown, H. M., Garrett, S. D. (2005b). Fish species
identification using PCR-RFLP analysis and lab-on-a-chip capillary electrophoresis: application
to detect white fish species in food products and an interlaboratory study. J. Agr. Food Chem.,
53, 3348-3357.
Downs, T. R., Wilfinger, W. W. (1983). Fluorometric quantification of DNA in cells and tissue. Anal.
Biochem., 131, 538-547.
Emmett, M. R., Caprioli, R. M. (1994). Micro-electrospray mass spectrometry: ultra-high-sensitivity
analysis of peptides and proteins. J. Am. Soc. Mass Spectrom., 5, 605-613.
Eng, J. K., McCormack, A. L., Yates, J. R. (1994). An approach to correlate tandem mass spectral data
of peptides with amino acid sequences in a protein database. J. Am. Soc. Mass Spectrom., 5,
976-989.
Erickson, B. E. (2005). Proteomics reveals effects of aquaculture on fish. J. Proteome Res., 4, 667.
Etienne, M., Jerome, M., Fleurence, J., Rehbein, H., Kundiger, R., Mendes, R., Costa, H., Perez-Martin,
R., Pineiro-Gonzalez, C., Craig, A., Mackie, I., Malmheden Yman, I., Ferm, M., Martinez, I.,
Jessen, F., Smelt, A., Luten, J. (2000). Identification of fish species after cooking by SDS-
PAGE and urea IEF: a collaborative study. J. Agr. Food Chem., 48, 2653-2658.
Evans, F. F., Raftery, M. J., Egan, S., Kjelleberg, S. (2007). Profiling the secretome of the marine
bacterium Pseudoalteromonas tunicata using amine-specific isobaric tagging (iTRAQ). J.
Proteome Res., 6, 967-975.
FAO, 2006. Yearbooks of Fishery Statistics Summary Tables 2006.
ftp://ftp.fao.org/fi/stat/summary/default.htm
FAO SIDP (Species Identification and Data Programme). http://www.fao.org/fishery/sidp/2/en
Fenn, J. B., Mann, M., Meng, C. K., Wong, S. F., Whitehouse, C. M. (1989). Electrospray ionization for
mass spectrometry of large biomolecules. Science, 246, 64-71.
Fernández, A., García, T., González, I., Asensio, L., Rodríguez, M. A., Hernández, P. E., Martín, R.
(2002a). Polymerase chain reaction-restriction fragment length polymorphism analysis of a 16S
rRNA gene fragment for authentication of four clam species. J.Food Protect., 65, 692-695.
Fernández, A., García, T., Asensio, L., Rodríguez, M. A., González, I., Lobo, E., Hernández, P.
E.,Martín, R. (2002b). Genetic differentiation between the clam species Ruditapes decussatus
(grooved carpet shell) and Venerupis pullastra (pullet carpet shell) by PCR-SSCP analysis. J.
Sci. Food Agr., 82, 881-885.
270

Bibliografía
FIGIS FAO, 2006. Fisheries Global Information System. http://www.fao.org/fishery/topic/16140
FISH INFOnetwork. Market Report on Shrimp. http://www.eurofish.dk
France, R. M., Sellers, D. S., Grossman, S. H. (1997). Purification, characterization, and hydrodynamic
properties of arginine kinase from Gulf shrimp (Penaeus aztecus). Arch. Biochem. Biophys.,
345, 73-78.
Frank, A., Pevzner, P. (2005). PepNovo: de novo peptide sequencing via probabilistic network
modeling. Anal. Chem., 77, 964-973.
Gallardo, J. M., Sotelo, C. G., Piñeiro, C., Perez-Martin, R. I. (1995). Use of capillary zone
electrophoresis for fish species identification. Differentiation of flatfish species. J. Agr. Food
Chem., 43, 1238-1244.
Garrels, J. I. (1989). The QUEST system for quantitative analysis of two-dimensional gels. J. Biol.
Chem., 264, 5269-5282.
Goll, D. E., Robson, R. M., Stromer, M. H. (1977). Muscle proteins. En: J. R. Whitaker, S. R.
Tannebaum, Food Proteins (pp. 121-174). Westford, CT: Avi Publishing.
Gomiero, A., Pampanin, D. M., Bjørnstad, A., Larsen, B. K., Provan, F., Lyng, E., Andersen, O. K.
(2006). An ecotoxicoproteomic approach (SELDI-TOF mass spectrometry) to biomarker
discovery in crab exposed to pollutants under laboratory conditions. Aquat. Toxicol., 78
(Supplement 1), S34-S41.
Greig, C., Robertson, J. M., Banks, M. A. (2002). Rapid PCR-based species tests for threatened
sympatric salmonids. Conserv. Genet., 3, 81-84.
Han, K., Ely, B. (2002). Use of AFLP analyses to assess genetic variation in morone and thunnus
species. Mar. Biotechnol., 4, 141-145.
Henzel, W. J., Billeci, T. M., Stults, J. T., Wong, S. C., Grimley, C., Watanabe, C. (1993). Identifying
proteins from two-dimensional gels by molecular mass searching of peptide fragments in protein
sequence databases. P. Natl. Acad. Sci. USA, 90, 5011-5015.
Hird, H. J., Hold, G. L., Chisholm, J., Reece, P., Russell, V. J., Brown, J., Goodier, R., MacArthur, R.
(2005). Development of a method for the quantification of haddock (Melanogrammus
aeglefinus) in commercial products using real-time PCR. Eur. Food Res. Technol., 220, 633-
637.
Holthuis, L. B. (1980). FAO Species Catalogue Vol. 1 – Shrimps and prawns of the World. An
annotated catalogue of species of interest to fisheries. FAO Fisheries Synopsis No. 125, Volume
1.
Horstkotte, B., Rehbein, H. (2003). Fish species identification by means of restriction fragment length
polymorphism and high-performance liquid chromatography. J. Food Sci., 68, 2658-2666.
271

Bibliografía
Hsieh, H. S., Chai, T., Cheng, C. A., Hsieh, Y. W., Hwang, D. F. (2004). Application of DNA technique
for identifying the species of different processed products of swordfish meat. J. Food Sci., 69,
FCT1-FCT6.
Huang, T.-S., Marshall, M. R., Kao, K.-J., Otwell, W. S., Wei, C.-I. (1995). Development of monoclonal
antibodies for red snapper (Lutjanus campechanus) identification using enzyme-linked
immunosorbent assay. J. Agr. Food Chem., 43, 2301-2307.
Jensen, O. N., Wilm, M., Shevchenko, A., Mann, M. (1999). Sample preparation methods for mass
spectrometric peptide mapping directly from 2-DE gels. En: A. J. Link (ed.), 2-D Proteome
Analysis Protocols (pp. 513-530). Totowa, NJ: Humana Press.
Jerome, M., Lemaire, C., Verrez-Bagnis, V., Etienne, M. (2003). Direct sequencing method for species
identification of canned sardine and sardine-type products. J. Agr. Food Chem., 51, 7326-7332.
Jin, L.-G., Cho, J.-Y., Seong, K.-B., Park, J.-Y., Kong, I.-S., Hong, Y.-K. (2006). 18S rRNA gene
sequences and random amplified polymorphic DNA used in discriminating Manchurian trout
from other freshwater salmonids. Fish. Sci., 72, 903-905.
Joensen, H., Grahl-Nielsen, O. (2000). Discrimination of Sebastes viviparus, Sebastes marinus and
Sebastes mentella from Faroe Islands by chemometry of the fatty acid profile in heart and gill
tissues and in the skull oil. Comp. Biochem. Phys. B, 126, 69-79.
Joensen, H., Steingrund, P., Fjallstein, I., Grahl-Nielsen, O. (2000). Discrimination between two reared
stocks of cod (Gadus morhua) from the Faroe Islands by chemometry of the fatty acid
composition in the heart tissue. Mar. Biol., 136, 573-580.
Karas, M., Hillenkamp, F. (1988). Laser desorption ionization of proteins with molecular masses
exceeding 10,000 daltons. Anal. Chem., 60, 2299-2301.
Khamnamtong, B., Klinbunga, S., Menasveta, P. (2005). Species identification of five penaeid shrimps
using PCR-RFLP and SSCP analyses of 16S ribosomal DNA. J. Biochem. Mol. Biol., 38, 491-
499.
Kinter, M., Sherman, N. E. (2000). Protein sequencing and identification using tandem mass
spectrometry (pp. 161-163). New York: Wiley Interscience.
Klose, J. (1975). Protein mapping by combined isoelectric focusing and electrophoresis of mouse
tissues. Hum. Genet., 26, 231-243.
Knigge, T., Monsinjon, T., Andersen, O.-K. (2004). Surface-enhanced laser desorption/ionization-time
of flight-mass spectrometry approach to biomarker discovery in blue mussels (Mytilus edulis)
exposed to polyaromatic hydrocarbons and heavy metals under field conditions. Proteomics, 4,
2722-2727.
Knuutinen, J., Harjula, P. (1998). Identification of fish species by reversed-phase high-performance
liquid chromatography with photodiode-array detection. J. Chromatogr. B, 705, 11-21.
272

Bibliografía
Kvasnicka, F. (2003). Proteomics: general strategies and application to nutritionally relevant proteins. J.
Chromatogr. B, 787, 77-89.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature, 227, 680-685.
Larraín, M. A., Abugoch, L., Quitral, V., Vinagre, J., Segovia, C. (2002). Capillary zone electrophoresis
as a method for identification of golden kinglip (Genypterus blacodes) species during frozen
storage. Food chem., 76, 377-384.
Lavery, S., Chan, T. Y., Tam, Y. K., Chu, K. H. (2004). Phylogenetic relationships and evolutionary
history of the shrimp genus Penaeus s.l. derived from mitochondrial DNA. Mol. Phylogenet.
Evol., 31, 39-49.
LeBlanc, E. L., Singh, S., LeBlanc, R. J. (1994). Capillary zone electrophoresis of fish muscle
sarcoplasmic proteins. J. Food Sci., 59, 1267-1270.
Li, L., Guo, X. (2004). AFLP-based genetic linkage maps of the Pacific oyster Crassostrea gigas
Thunberg. Mar. Biotechnol., 6, 26-36.
Liebler, D. C., ed (2002). Introduction to Proteomics: tools for the new Biology. Totowa, NJ: Humana
Press.
Lin, W.-F., Hwang, D.-F. (2007). Application of PCR-RFLP analysis on species identification of canned
tuna. Food Control, 18, 1050-1057.
Link, A. J., Eng, J., Schieltz, D. M., Carmack, E., Mize, G. J., Morris, D. R., Garvik, B. M., Yates, J. R.
3rd. (1999). Direct analysis of protein complexes using mass spectrometry. Nat. Biotechnol., 17,
676-682.
Liu, Z., Li, P., Argue, B. J., Dunham, R. A. (1998a). Inheritance of RAPD markers in channel catfish
(Ictalurus punctatus), blue catfish (I. furcatus), and their F1, F2 and backcross hybrids. Anim.
Genet., 29, 58-62.
Liu, Z., Nichols, A., Li, P., Dunham, R. A. (1998b). Inheritance and usefulness of AFLP markers in
channel catfish (Ictalurus punctatus), blue catfish (I. furcatus), and their F1, F2, and backcross
hybrids. Mol. Gen. Genet., 258, 260-268.
Lockley, A. K., Bardsley, R. G. (2000a). Novel method for the discrimination of tuna (Thunnus thynnus)
and bonito (Sarda sarda) DNA. J. Agr. Food Chem., 48, 4463-4468.
Lockley, A. K., Bardsley, R. G. (2000b). DNA-based methods for food authentication. Trends Food Sci.
Tech., 11, 67-77.
Lopez, I., Pardo, M. A. (2005). Application of relative quantification TaqMan real-time polymerase
chain reaction technology for the identification and quantification of Thunnus alalunga and
Thunnus albacares. J. Agr. Food Chem., 53, 4554-4560.
López, J. L., Marina, A., Vázquez, J., Álvarez, G. (2002a). A proteomic approach to the study of the
marine mussels Mytilus edulis and M. galloprovincialis. Mar. Biol., 141, 217-223.
273

Bibliografía
López, J. L., Marina, A., Álvarez, G., Vázquez, J. (2002b). Application of proteomics for fast
identification of species-specific peptides from marine species. Proteomics, 2, 1658-1665.
López, J. L. (2007). Two-dimensional electrophoresis in proteome expression analysis. J. Chromatogr.
B, 849, 190-202.
Lopez-Ferrer, D., Capelo, J. L., Vazquez, J. (2005). Ultra fast trypsin digestion of proteins by high
intensity focused ultrasound. J. Proteome Res., 4, 1569-1574.
Lundstrom, R. C. (1979). Fish species identification by thin layer isoelectric focusing. J. Assoc. Off.
Anal. Chem., 62, 624-629.
Ma, B., Zhang, K., Hendrie, C., Liang, C., Li, M., Doherty-Kirby, A., Lajoie, G. (2003). PEAKS:
powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid
Commun. Mass Sp., 17, 2337-2342.
Mackie, I. M., Jones, B. W. (1978). The use of electrophoresis of the water-soluble (sarcoplasmic)
proteins of fish muscle to differentiate the closely related species of hake (Merluccius sp.).
Comp. Biochem. Phys. B, 59, 95-98.
Mackie, I. M. (1997). Methods of identifying species of raw and processed fish. En: G. M. Hall (ed.),
Fish Processing Technology (pp. 160-199). London: Chapman & Hall.
Mackie, I., Craig, A., Etienne, M., Jérôme, M., Fleurence, J., Jessen, F., Smelt, A., Kruijt, A.,
Malmheden Yman, I., Ferm, M., Martinez, I., Pérez-Martín, R., Piñeiro, C., Rehbein, H.,
Kündiger, R. (2000). Species identification of smoked and gravad fish products by sodium
dodecylsulphate polyacrylamide gel electrophoresis, urea isoelectric focusing and native
isoelectric focusing: a collaborative study. Food chem., 71, 1-7.
Mafra, I., Ferreira, I., Oliveira, M. (2008). Food authentication by PCR-based methods. Eur. Food Res.
Technol., 227, 649-665.
Maggioni, R., Rogers, A. D., Maclean, N., Incao, F. (2001). Molecular phylogeny of Western Atlantic
Farfantepenaeus and Litopenaeus shrimp based on mitochondrial 16S partial sequences. Mol.
Phylogenet. Evol., 18, 66-73.
Magnussen, J. E., Pikitch, E. K., Clarke, S. C., Nicholson, C., Hoelzel, A. R., Shivji, M. S. (2007).
Genetic tracking of basking shark products in international trade. Anim. Conserv., 10, 199-207.
Mahon, B. C.,Neigel, J. E. (2008). Utility of arginine kinase for resolution of phylogenetic relationships
among brachyuran genera and families. Mol. Phylogenet. Evol., 48, 718-727.
Mairs, D. F., Sindermann, C. J. (1962). A serological comparison of five species of atlantic clupeoid
fishes. Biol. Bull., 123, 330-343.
Maldini, M., Nonnis Marzano, F., Fortes, G. G., Papa, R., Gandolfi, G. (2006). Fish and seafood
traceability based on AFLP markers: elaboration of a species database. Aquaculture, 261, 487-
494.
274

Bibliografía
Mann, M., Wilm, M. (1994). Error-tolerant identification of peptides in sequence databases by peptide
sequence tags. Anal. Chem., 66, 4390-4399.
MAPA: Ministerio de Agricultura, Pesca y Alimentación. Libro blanco de la pesca. http://www.mapa.es
Marko, P. B., Lee, S. C., Rice, A. M., Gramling, J. M., Fitzhenry, T. M., McAlister, J. S., Harper, G. R.,
Moran, A. L. (2004). Fisheries: mislabelling of a depleted reef fish. Nature, 430, 309-310.
Martin, J. W., Davis, G. E. (2001). An updated classification of the recent Crustacea. Los Angeles, CA
(USA): Natural History Museum of Los Angeles County.
Martinez, I., Ofstad, R., Olsen, R. L. (1990). Electrophoretic study of myosin isoforms in white muscles
of some teleost fishes. Comp. Biochem. Phys. B, 96, 221-227.
Martinez, I., Jakobsen, T., Seppola, F. M. (2001). Requirements for the application of protein sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and randomly amplified polymorphic DNA
analyses to product speciation. Electrophoresis, 22, 1526-1533.
Martinez, I., Friis, T. J. (2004). Application of proteome analysis to seafood authentication. Proteomics,
4, 347-354.
McLafferty, F. W. (1981). Tandem mass spectrometry. Science, 214, 280-287.
McNulty, S. T., Klesius, P. H. (2005). Development of an indirect enzyme-linked immunoabsorbent
assay using a monoclonal antibody to identify Ictalurus sp. fillets. Aquac. Res., 36, 1279-1284.
Medina, I., Aubourg, S., Martín, R. (1995). Composition of phospholipids of white muscle of six tuna
species. Lipids, 30, 1127-1135.
Medina, I., Aubourg, S., Trigo, M., Ares, J. (2003). Fish species identification based on phospholipids
analysis. En: R. Pérez-Martín, & C. G. Sotelo (eds.), Authenticity of Species in Meat and
Seafood Products (pp. 269-279). Vigo: Association "International Congress on Authenticity of
Species in meat and seafood products".
Mi, J., Apraiz, I., Cristobal, S. (2007). Peroxisomal proteomic approach for protein profiling in blue
mussels (Mytilus edulis) exposed to crude oil. Biomarkers, 12, 47 - 60.
Mullis, K., Faloona, F., Scharf, S., Saiki, R., Horn, G., Erlich, H. (1986). Specific enzymatic
amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb. Symp. Quant.
Biol., 51, 263-273.
Navas, A., Albar, J. P. (2004). Application of proteomics in phylogenetic and evolutionary studies.
Proteomics, 4, 299-302.
Ochiai, A. (1957). Zoogeographical studies on the Soleoid fishes found in Japan and its neighbouring
regions. Bull. Jpn. Soc. Sci. Fish., 22, 526-530.
O'Farrell, P. H. (1975). High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem.,
250, 4007-4021.
Osman, M. A., Ashoor, S. H., Marsh, P. C. (1987). Liquid chromatographic identification of common
fish species. J. Assoc. Off. Anal. Chem., 70, 618-625.
275

Bibliografía
Pandey, A., Mann, M. (2000). Proteomics to study genes and genomes. Nature, 405, 837-846.
Pappin, D. J. C., Hojrup, P., Bleasby, A. J. (1993). Rapid identification of proteins by peptide-mass
fingerprinting. Curr. Biol., 3, 327-332.
Pascoal, A., Barros-Velázquez, J., Cepeda, A., Gallardo, J. M., Calo-Mata, P. (2008a). A polymerase
chain reaction-restriction fragment length polymorphism method based on the analysis of a 16S
rRNA/tRNAVal mitochondrial region for species identification of commercial penaeid shrimps
(Crustacea: Decapoda: Penaeoidea) of food interest. Electrophoresis, 29, 499-509.
Pascoal, A., Barros-Velázquez, J., Cepeda, A., Gallardo, J. M., Calo-Mata, P. (2008b). Identification of
shrimp species in raw and processed food products by means of a polymerase chain reaction-
restriction fragment length polymorphism method targeted to cytochrome b mitochondrial
sequences. Electrophoresis, 29, 3220-3228.
Paul, W., Steinwedel, H. (1953). A new mass spectrometer without a magnetic field. Z. Naturforsch. A,
8, 448-450.
Pedrós-Alió, C. (2006). Genomics and marine ecology. Int. Microbiol., 9, 191-197.
Pepe, T., Trotta, M., di Marco, I., Cennamo, P., Anastasio, A., Cortesi, M. (2005). Mitochondrial
cytochrome b DNA sequence variations: an approach to fish species identification in processed
fish products. J. Food Protect., 68, 421-425.
Pérez-Farfante, I., Kensley, B. (1997). Penaeoid and sergestoid shrimps and prawns of the world. Keys
and diagnoses for the families and genera. Paris: Éditions du Muséum national d’Histoire
naturelle.
Perkins, D. N., Pappin, D. J. C., Creasy, D. M., Cottrell, J. S. (1999). Probability-based protein
identification by searching sequence databases using mass spectrometry data. Electrophoresis,
20, 3551-3567.
Piñeiro, C., Sotelo, C. G., Medina, I., Gallardo, J. M., Pérez-Martín, R. I. (1997). Reversed-phase HPLC
as a method for the identification of gadoid fish species. Z. Lebensm. Unters. Forsch. A, 204,
411-416.
Piñeiro, C., Barros-Velázquez, J., Sotelo, C. G., Pérez-Martín, R. I., Gallardo, J. M. (1998). Two-
dimensional electrophoretic study of the water-soluble protein fraction in white muscle of
gadoid fish species. J. Agric. Food Chem., 46, 3991-3997.
Piñeiro, C., Barros-Velázquez, J., Pérez-Martín, R. I., Martínez, I., Jacobsen, T., Rehbein, H., Kündiger,
R., Mendes, R., Etienne, M., Jerome, M., Craig, A., Mackie, I. M., Jessen, F. (1999a).
Development of a sodium dodecyl sulfate-polyacrylamide gel electrophoresis reference method
for the analysis and identification of fish species in raw and heat-processed samples: a
collaborative study. Electrophoresis, 20, 1425-1432.
276

Bibliografía
Piñeiro, C., Barros-Velázquez, J., Sotelo, C. G., Gallardo, J. M. (1999b). The use of two-dimensional
electrophoresis in the characterization of the water-soluble protein fraction of commercial flat
fish species. Z. Lebensm. Unters. Forsch. A, 208, 342-348.
Piñeiro, C., Barros-Velázquez, J., Pérez-Martín, R. I., Gallardo, J. M. (2000). Specific enzyme detection
following isoelectric focusing as a complimentary tool for the differentiation of related gadoid
fish species. Food chem., 70, 241-245.
Piñeiro, C., Vázquez, J., Marina, A. I., Barros-Velázquez, J., Gallardo, J. M. (2001). Characterization
and partial sequencing of species-specific sarcoplasmic polypeptides from commercial hake
species by mass spectrometry following two-dimensional electrophoresis. Electrophoresis, 22,
1545-1552.
Piñeiro, C., Barros-Velázquez, J., Vázquez, J., Figueras, A., Gallardo, J. M. (2003). Proteomics as a tool
for the investigation of seafood and other marine products. J. Proteome Res., 2, 127-135.
Provan, F., Bjørnstad, A., Pampanin, D. M., Lyng, E., Fontanillas, R., Andersen, O. K., Koppe, W.,
Bamber, S. (2006). Mass spectrometric profiling - A diagnostic tool in fish? Mar. Environ. Res.,
62 (Supplement 1), S105-S108.
Quan, J., Zhuang, Z., Deng, J., Dai, J., Zhang, Y.-p. (2004). Phylogenetic relationships of 12 Penaeoidea
shrimp species deduced from mitochondrial DNA sequences. Biochem. Genet., 42, 331-345.
Quinteiro, J., Sotelo, C. G., Rehbein, H., Pryde, S. E., Medina, I., Perez-Martin, R. I., Rey-Mendez, M.,
Mackie, I. M. (1998). Use of mtDNA direct polymerase chain reaction (PCR) sequencing and
PCR-restriction fragment length polymorphism methodologies in species identification of
canned tuna. J. Agr. Food Chem., 46, 1662-1669.
Ram, J. L., Ram, M. L., Baidoun, F. F. (1996). Authentication of canned tuna and bonito by sequence
and restriction site analysis of polymerase chain reaction products of mitochondrial DNA. J.
Agr. Food Chem., 44, 2460-2467.
Ramella, M. S., Kroth, M. A., Tagliari, C., Arisi, A. C. M. (2005). Optimization of random amplified
polymorphic DNA protocol for molecular identification of Lophius gastrophysus. Ciênc.
Tecnol. Aliment., 25, 733-735.
Rasmussen, R. S., Morrissey, M. T. (2008). DNA-based methods for the identification of commercial
fish and seafood species. Compr. Rev. Food Sci. Food Saf., 7, 280-295.
Real Decreto 1380/2002, de 20 de diciembre, de identificación de los productos de la pesca, de la
acuicultura y del marisqueo congelados y ultracongelados. Boletín Oficial del Estado nº 3,
Viernes 3 de enero de 2003.
Real Decreto 121/2004, de 23 de enero, sobre la identificación de los productos de la pesca, de la
acuicultura y del marisqueo vivos, frescos, refrigerados o cocidos. Boletín Oficial del Estado nº
31, Jueves 5 de febrero de 2004.
277

Bibliografía
Real Decreto 1702/2004, de 16 de julio, por el que se modifica el Real Decreto 1380/2002, de 20 de
diciembre, de identificación de los productos de la pesca, de la acuicultura y del marisqueo
congelados y ultracongelados. Boletín Oficial del Estado nº 172, Sábado 17 de julio de 2004.
Redeby, T., Carr, H., Björk, M., Emmer, Å. (2006). A screening procedure for the solubilization of
chloroplast membrane proteins from the marine green macroalga Ulva lactuca using RP-HPLC-
MALDI-MS. Int. J. Biol. Macromol., 39, 29-36.
Rego, I., Martinez, A., Gonzalez-Tizon, A., Vieites, J., Leira, F., Mendez, J. (2002). PCR technique for
identification of mussel species. J. Agr. Food Chem., 50, 1780-1784.
Rehbein, H. (1990). Fish species identification by HPLC of sarcoplasmic proteins. Fresenius J. Anal.
Chem., 337, 106.
Rehbein, H. (1995). Food control by applied biochemistry of marine organisms: comparison of proteins
and metabolites from fish and invertebrate muscle. Helgol. Mar. Res., 49, 747-757.
Rehbein, H., Kress, G., Schmidt, T. (1997). Application of PCR-SSCP to species identification of
fishery products. J. Sci. Food Agr., 74, 35-41.
Rehbein, H., Mackie, I. M., Pryde, S., Gonzales-Sotelo, C., Medina, I., Perez-Martin, R., Quinteiro, J.,
Rey-Mendez, M. (1999a). Fish species identification in canned tuna by PCR-SSCP: validation
by a collaborative study and investigation of intra-species variability of the DNA-patterns. Food
chem., 64, 263-268.
Rehbein, H., Kündiger, R., Yman, I. M., Ferm, M., Etienne, M., Jerome, M., Craig, A., Mackie, I.,
Jessen, F., Martinez, I., Mendes, R., Smelt, A., Luten, J., Pineiro, C., Perez-Martin, R. (1999b).
Species identification of cooked fish by urea isoelectric focusing and sodium dodecylsulfate
polyacrylamide gel electrophoresis: a collaborative study. Food chem., 67, 333-339.
Rehbein, H., Kündiger, R., Piñeiro, C., Perez-Martin, R. I. (2000). Fish muscle parvalbumins as marker
proteins for native and urea isoelectric focusing. Electrophoresis, 21, 1458-1463.
Rehbein, H., Sotelo, C., Perez-Martin, R., Chapela-Garrido, M. J., Hold, G., Russell, V., Pryde, S.,
Santos, A., Rosa, C., Quinteiro, J., Rey-Mendez, M. (2002). Differentiation of raw or processed
eel by PCR-based techniques: restriction fragment length polymorphism analysis (RFLP) and
single strand conformation polymorphism analysis (SSCP). Eur. Food Res. Technol., 214, 171-
177.
Resolución de 4 de marzo de 2009, de la Secretaría General del Mar, por la que se establece y se publica
el listado de denominaciones comerciales de especies pesqueras y de acuicultura admitidas en
España. Boletín Oficial del Estado nº 80, Jueves 2 de abril de 2009.
Roepstorff, P., Fohlman, J. (1984). Proposal for a common nomenclature for sequence ions in mass
spectra of peptides. Biomed. Mass Spectrom., 11, 601.
Rondelet, G. (1554). Libri de Piscibus Marinis, in quibus veræ piscium effigies expressæ sunt. Lugduni
(Bonhomme).
278

Bibliografía
Russell, V. J., Hold, G. L., Pryde, S. E., Rehbein, H., Quinteiro, J., Rey-Mendez, M., Sotelo, C. G.,
Perez-Martin, R. I., Santos, A. T., Rosa, C. (2000). Use of restriction fragment length
polymorphism to distinguish between salmon species. J. Agr. Food Chem., 48, 2184-2188.
Saitou, N., Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic
trees. Mol. Biol. Evol., 4, 406-425.
Sampson, H. A. (2004). Update on food allergy. J. Allergy Clin. Immunol., 113, 805-819.
Santaclara, F., Cabado, A., Vieites, J. (2006). Development of a method for genetic identification of four
species of anchovies: E. encrasicolus, E. anchoita, E. ringens and E. japonicus. Eur. Food Res.
Technol., 223, 609-614.
Sardà, F. (1993). Biología descriptiva: Crustáceos. En: F. Castelló (ed.), Acuicultura marina:
fundamentos biológicos y tecnología de la producción (pp. 69-91). Barcelona: Publicacions i
Edicions Universitat de Barcelona.
Schägger, H., von Jagow, G. (1987). Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis
for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem., 166, 368-379.
Schweder, T., Markert, S., Hecker, M. (2008). Proteomics of marine bacteria. Electrophoresis, 29, 2603-
2616.
Scobbie, A. E., Mackie, I. M. (1988). The use of sodium dodecyl sulphate polyacrylamide gel
electrophoresis in fish species identification-A procedure suitable for cooked and raw fish. J.
Sci. Food Agr., 44, 343-351.
Shiomi, K., Sato, Y., Hamamoto, S., Mita, H., Shimakura, K. (2008). Sarcoplasmic calcium-binding
protein: identification as a new allergen of the black tiger shrimp Penaeus monodon. Int. Arch.
Allergy Imm., 146, 91-98.
Shivji, M., Clarke, S., Pank, M., Natanson, L., Kohler, N., Stanhope, M. (2002). Genetic identification
of pelagic shark body parts for conservation and trade monitoring. Conserv. Biol., 16, 1036-
1047.
Sicherer, S. H., Muñoz-Furlong, A., Sampson, H. A. (2004). Prevalence of seafood allergy in the United
States determined by a random telephone survey. J. Allergy Clin. Immunol., 114, 159-165.
SOFIA (2009). El estado mundial de la pesca y la acuicultura 2008. FAO.
Sotelo, C. G., Piñeiro, C., Gallardo, J. M., Pérez-Martin, R. I. (1992). Identification of fish species in
smoked fish products by electrophoresis and isoelectric focusing. Z. Lebensm. Unters. Forsch.
A, 195, 224-227.
Sotelo, C. G., Calo-Mata, P., Chapela, M. J., Perez-Martin, R. I., Rehbein, H., Hold, G. L., Russell, V.
J., Pryde, S., Quinteiro, J., Izquierdo, M., Rey-Mendez, M., Rosa, C., Santos, A. T. (2001).
Identification of flatfish (Pleuronectiforme) species using DNA-based techniques. J. Agr. Food
Chem., 49, 4562-4569.
279

Bibliografía
Sotelo, C. G., Chapela, M. J., Rey, M., Pérez-Martín, R. (2003). Development of an identification and
quantitation system for cod (Gadus morhua) using taqman assay. En: First Joint Trans-Atlantic
Fisheries Technology Conference (pp. 195-198). Reykjavik, Iceland.
Stafford, G. C., Kelley, P. E., Syka, J. E. P., Reynolds, W. E., Todd, J. F. J. (1984). Recent
improvements in and analytical applications of advanced Ion Trap technology. Int. J. Mass
Spectrom. Ion Processes., 60, 85-98.
Takeyama, H., Chow, S., Tsuzuki, H., Matunaga, T. (2001). Mitochondrial DNA sequence variation
within and between tuna Thunnus species and its application to species identification. J. Fish
Biol., 58, 1646-1657.
Tamura, K., Dudley, J., Nei, M., Kumar, S. (2007). MEGA4: Molecular Evolutionary Genetics Analysis
(MEGA) software version 4.0. Mol. Biol. Evol., 24, 1596-1599.
Tanaka, K., Waki, H., Ido, Y., Akita, S., Yoshida, Y., Yoshida, T., Matsuo, T. (1988). Protein and
polymer analyses up to m/z 100 000 by laser ionization time-of-flight mass spectrometry. Rapid
Commun. Mass Spectrom., 2, 151-153.
Taylor, W. J., Jones, J. L. (1992). An immunoassay for verifying the identity of canned sardines. Food
Agr. Immunol., 4, 169 - 175.
Teletchea, F., Maudet, C., Hänni, C. (2005). Food and forensic molecular identification: update and
challenges. Trends Biotechnol., 23, 359-366.
Tepedino, V., Berrini, A., Borromeo, V., Gaggioli, D., Cantoni, C., Manzoni, P., Secchi, C. (2001).
Identification of commercial fish species belonging to the orders Pleuronectiformes and
Gadiformes: library of isoelectric focusing patterns. J. AOAC Int., 84, 1600-1607.
Thomas, T., Egan, S., Burg, D., Ng, C., Ting, L., Cavicchioli, R. (2007). Integration of genomics and
proteomics into marine microbial ecology. Mar. Ecol. -Prog. Ser., 332, 291-299.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., Higgins, D. G. (1997). The CLUSTALX
windows interface: flexible strategies for multiple sequence alignment aided by quality analysis
tools. Nucleic Acids Res., 25, 4876-4882.
Tritt, K. L., O'Bara, C. J., Wells, M. J. M. (2005). Chemometric discrimination among wild and cultured
age-0 largemouth bass, black crappies, and white crappies based on fatty acid composition. J.
Agr. Food Chem., 53, 5304-5312.
Trotta, M., Schonhuth, S., Pepe, T., Cortesi, M. L., Puyet, A., Bautista, J. M. (2005). Multiplex PCR
method for use in real-time PCR for identification of fish fillets from grouper (Epinephelus and
Mycteroperca species) and common substitute species. J. Agr. Food Chem., 53, 2039-2045.
Tunsjø, H. S., Paulsen, S. M., Mikkelsen, H., L'Abée-Lund, T. M., Skjerve, E., Sørum, H. (2007).
Adaptive response to environmental changes in the fish pathogen Moritella viscosa. Res.
Microbiol., 158, 244-250.
280

Bibliografía
Ukishima, Y., Sakane, Y., Fukuda, A., Wada, S., Okada, S. (1995). Identification of whale species by
liquid chromatographic analysis of sarcoplasmic proteins. Mar. Mamm. Sci., 11, 344-361.
Valenzuela, M. A., Gamarra, N., Gómez, L., Kettlun, A. M., Sardón, M., Pérez, L. M., Vinagre, J.,
Guzman, N. A. (1999). A comparative study of fish species identification by gel
isoelectrofocusing two-dimensional gel electrophoresis, and capillary zone electrophoresis. J.
Capill. Electrophor. Microchip Technol., 6, 85-91.
Verrez, V., Benyamin, Y., Roustan, C. (1992). Detection of marine invertebrates in surimi-based
products. En: H. H. Huss, M. Jakobsen, J. Liston (eds.), Quality Assurance in the Fish Industry
(pp. 441-448). Amsterdam: Elsevier.
Voloch, C. M., Freire, P. R., Russo, C. A. M. (2005). Molecular phylogeny of penaeid shrimps inferred
from two mitochondrial markers. Genet. Mol. Res., 4, 668-674.
Weaver, L. A., Lundstrom, R. C., Colbert, A. A. (1999). Identification of shark species by
polyacrylamide gel isoelectric focusing of sarcoplasmic proteins. J. AOAC Int., 82, 1163-1170.
Weder, J., Rehbein, H., Kaiser, K.-P. (2001). On the specificity of tuna-directed primers in PCR-SSCP
analysis of fish and meat. Eur. Food Res. Technol., 213(2), 139-144.
Wei, C. -I., An, H., Chen, J. -S., Marshall, M. R. (1990). Use of a modified urea gel isoelectric focusing
method for species identification or raw or boiled white, pink, and rock shrimp. J. Food
Biochem., 14, 91-102.
Whitmore, D. H. (1990). Electrophoretic and isoelectric focusing techniques in fisheries management.
Boca Ratón, Florida: CRC Press.
Wilm, M., Mann, M. (1996). Analytical properties of the nanoelectrospray ion source. Anal. Chem., 68,
1-8.
Wu, C. C., MacCoss, M. J. (2002). Shotgun proteomics: tools for the analysis of complex biological
systems. Curr. Opin. Mol. Ther., 4, 242-250.
Yamauchi, M., Miya, M., Machida, R., Nishida, M. (2004). PCR-based approach for sequencing
mitochondrial genomes of decapod crustaceans, with a practical example from kuruma prawn
(Marsupenaeus japonicus). Mar. Biotechnol., 6, 419-429.
Yamazaki, Y., Shimada, N., Tago, Y. (2005). Detection of hybrids between masu salmon Oncorhynchus
masou masou and amago salmon O. m. ishikawae occurred in the Jinzu River using a random
amplified polymorphic DNA technique. Fish. Sci., 71, 320-326.
Young, W. P., Wheeler, P. A., Coryell, V. H., Keim, P., Thorgaard, G. H. (1998). A detailed linkage
map of rainbow trout produced using doubled haploids. Genetics, 148, 839-850.
Yu, Z., Guo, X. (2003). Genetic linkage map of the Eastern oyster Crassostrea virginica Gmelin. Biol.
Bull., 204, 327-338.
Yu, C.-J., Lin, Y.-F., Chiang, B.-L., Chow, L.-P. (2003). Proteomics and immunological analysis of a
novel shrimp allergen, Pen m 2. J. Immunol., 170, 445-453.
281

Bibliografía
Yue, G., Li, Y., Chen, F., Cho, S., Lim, L. C., Orban, L. (2002). Comparison of three DNA marker
systems for assessing genetic diversity in asian arowana (Scleropages formosus).
Electrophoresis, 23, 1025-1032.
Zhang, J., Cai, Z. (2006). Differentiation of the rainbow trout (Oncorhynchus mykiss) from Atlantic
salmon (Salmon salar) by the AFLP-derived SCAR. Eur. Food Res. Technol., 223, 413-417.
282


Recommended

















