Embed Size (px)
Citation preview

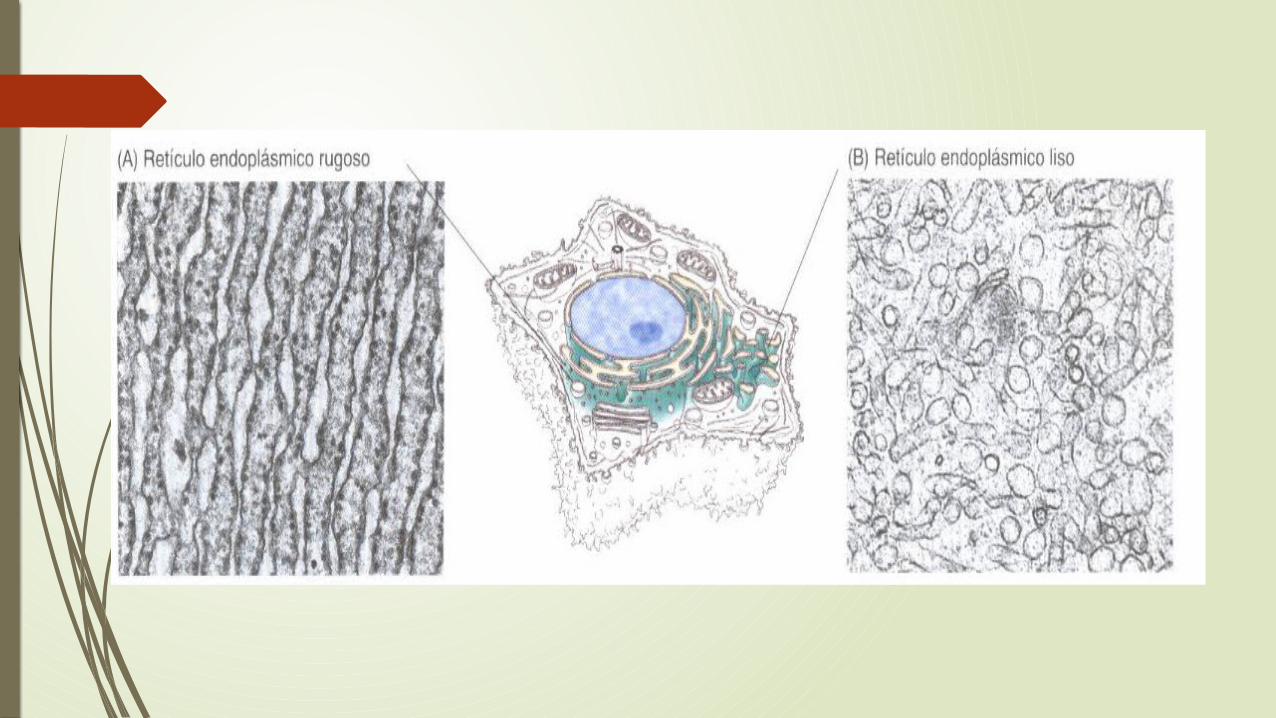
Retículo endoplasmatico rugoso y liso
Integrantes: Nayla Alarcón Camacho
Luis Ángel Campos Báez
Ma. Fernanda Carmona Huesca
Crystel Guevara Duran
Jose Blandino Carballo Landa

Retículo endoplasmatico
Es una red de túbulos y sacos (cisternas) rodeados de membrana que se extiende desde la membrana nuclear por todo el citoplasma.
El retículo endoplasmatico esta rodeado por una membrana continua y es el orgánulo mas grande de la mayoría de las células eucariotas.

Hay 2 tipos de RE los cuales realizan funciones diferentes en la célula: RE rugoso, que esta cubierto por ribosomas en su superficie externa y el RE de transición de donde parten vesículas hacia el aparato de Golgi, estos funcionan en el procesamiento de las proteínas. El RE liso no esta asociado con los ribosomas y esta implicado en el metabolismo de los lípidos, en lugar de las proteínas.

Retículo endóplasmatico y secreción de proteínas
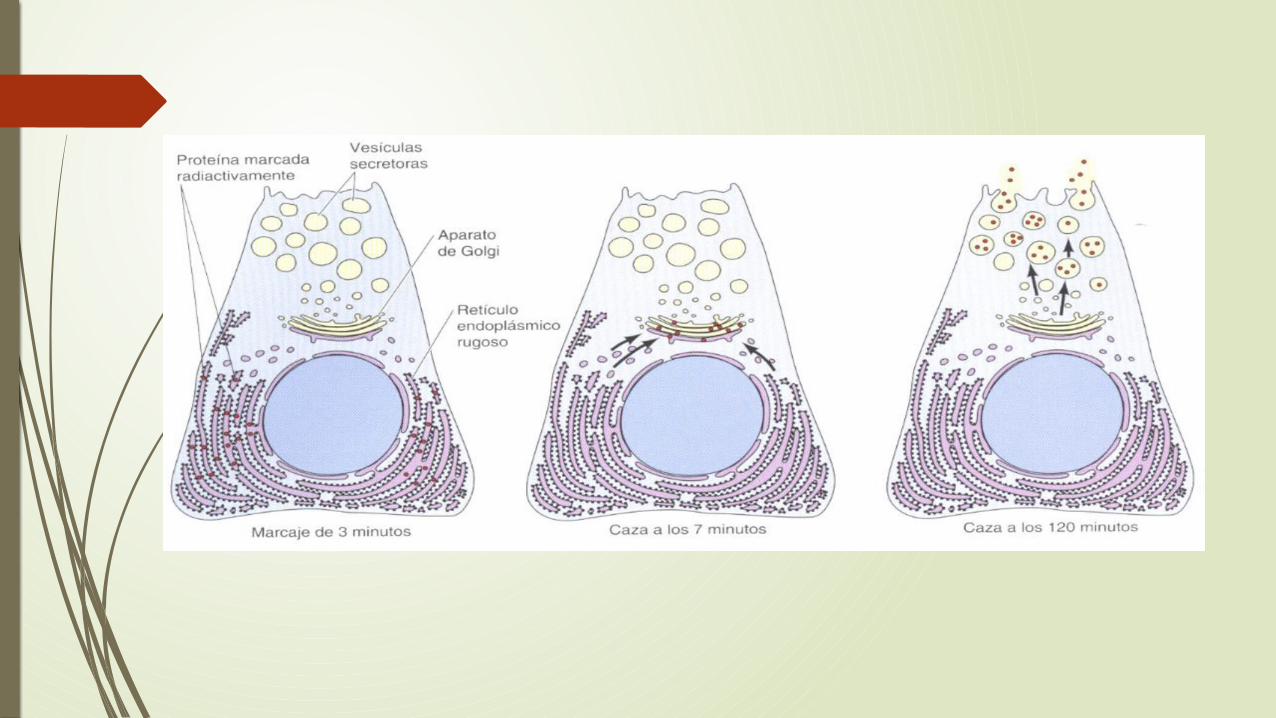
El papel del RE en el procesamiento y distribución de las proteínas fue demostrado por primera vez por George Palade y sus colaboradores en los años 60.
Ellos estudiaron el destino de las proteínas recién sintetizadas en unas células especializadas en el páncreas.
Palade y colaboradores fueron capaces de estudiar la ruta tomada por las proteínas secretadas simplemente mediante el marcaje con aminoácidos radiactivos de las proteínas recién sintetizadas. La localización en la células de las proteínas marcadas radiactivamente se le llama autorradiografia.

Experimentos definieron una vía tomada por las proteínas secretadas de la vía secretora: RE rugoso Golgi vesículas de secreción–exterior de la célula. Estudios adicionales demostraron y ampliaron los resultados que esta vía no esta restringida a las proteínas destinadas a ser secretadas. Las proteínas de la membrana plasmática y las lisosomas también migran desde el RE rugoso hasta el Golgi y posteriormente a sus destinos finales.
Otras proteínas pasan por la etapa inicial de las vía secretora pero posteriormente son retenidas y su actividad tiene lugar en el RE o en el aparato de Golgi .

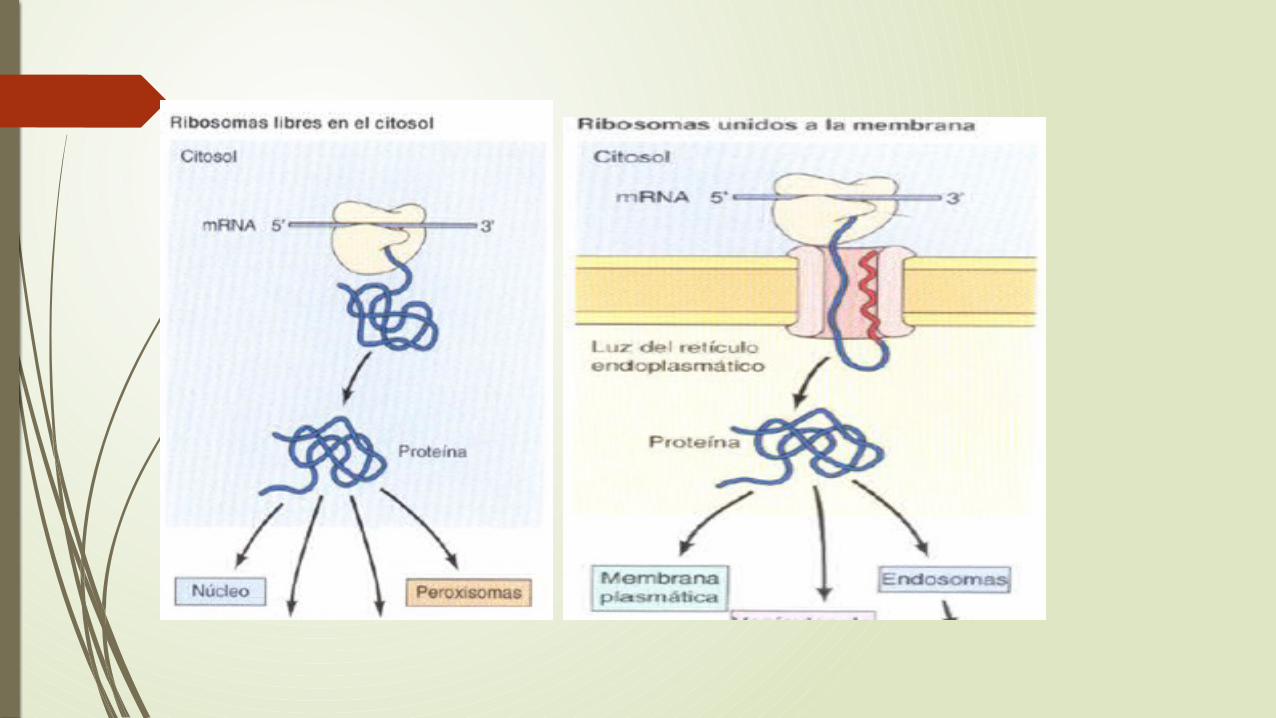
La entrada de las proteínas en el RE representa un cruce de caminos muy importante en el trafico de proteínas en las células eucariotas.
En las células de los mamíferos, la mayoría de las proteínas son transferidas al RE mientras están siendo traducidas por los ribosomas unidos a la membrana por el contrario , las proteínas destinadas a permanecer en el citosol o que se van a incorporar al núcleo, a las mitocondrias, cloroplastos o peroxisomas son sintetizadas en los ribosomas libres y liberadas al citosol cuando finaliza su traducción.

Marcaje de las proteínas para dirigirse al retículo endoplasmatico
Las proteínas pueden ser translocadas al RE durante su síntesis en los ribosomas unidos a la membrana (translocación contraduccional) o una vez que la traducción se ha completado en los ribosomas libres de citosol (translocación postraduccional).
En las células de mamíferos, la mayoría de las proteínas entran en el RE de manera contraduccional , mientras que en las levaduras se utilizan la vía contraduccional y postraduccional.

La vía contraduccional empieza por la asociación de los ribosomas con el RE. La marca que determina que los ribosomas se unan con la membrana del RE es la secuencia de aminoácidos de la cadena polipeptidica que esta siendo sintetizada, toda la síntesis de proteica se inicia en los ribosomas que están libres en el citosol.
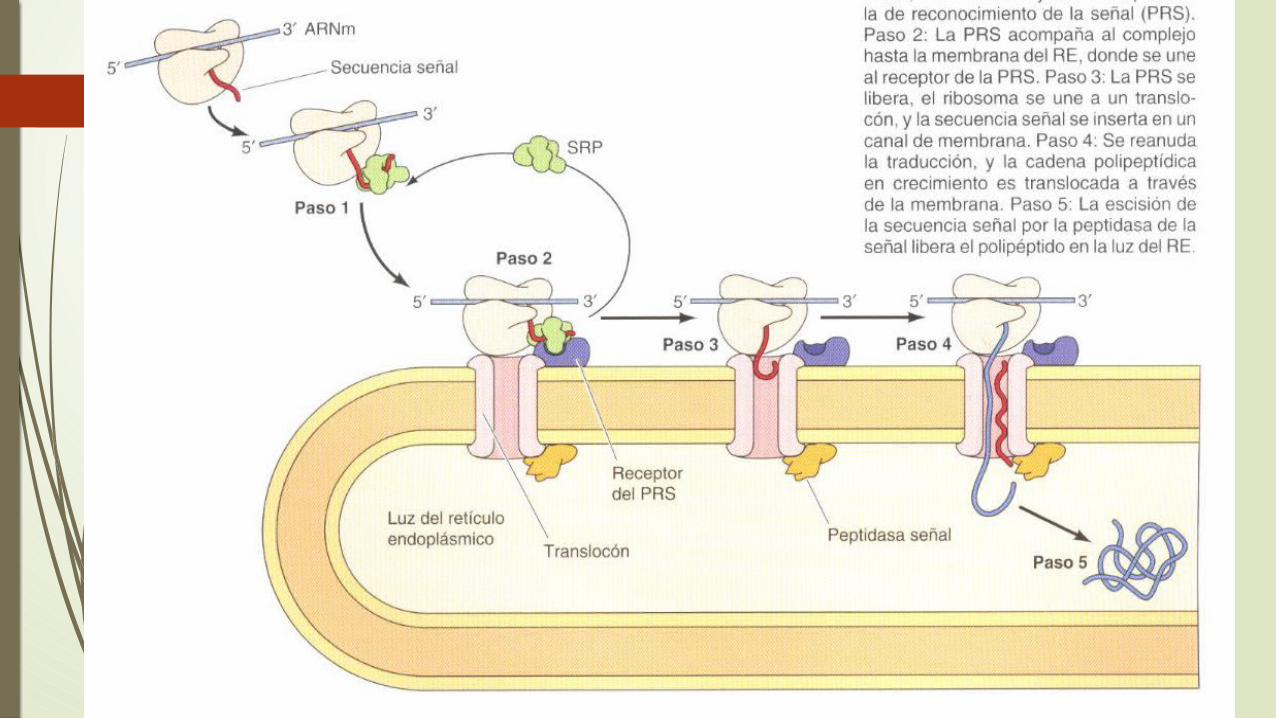
Los ribosomas implicados en la síntesis de proteínas destinadas a ser secretadas están marcados para dirigirse al RE mediante una secuencia de señal localizada en el extremo amino-terminal de la cadena polipeptidica en crecimiento. Estos son pequeños segmentos de aminoácidos hidrófobos que son escindidos de la cadena polipeptidica durante su transferencia a la luz del RE.

David Sabatini y Guenter Blobel propusieron en 1971 que la señal para la unión del ribosoma al RE era una secuencia de aminoácidos próxima al extremo amino-terminal de la cadena polipeptidica en crecimiento. Esto fue apoyado por los resultados de la traducción in vitro de ARNm que codifican proteínas secretadas como las inmunoglobulinas.
Estos experimentos condujeron a que se formulara de una manera mas detallada la hipótesis de la señal.
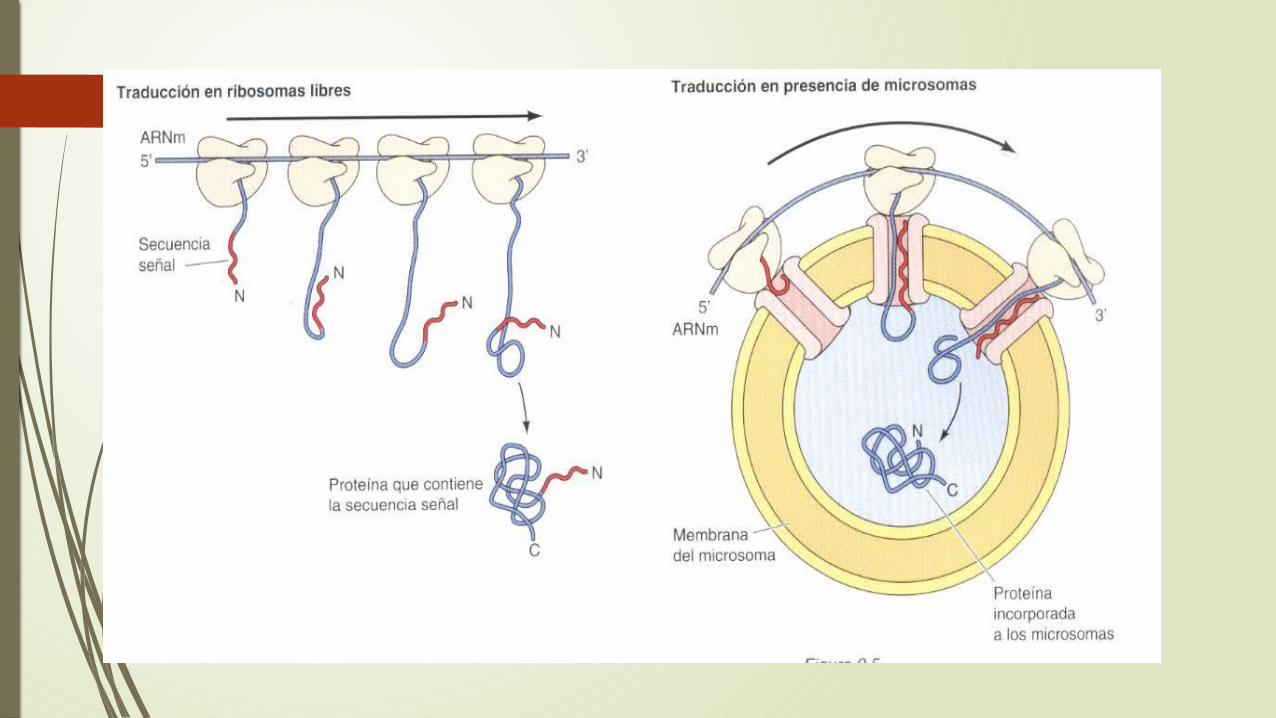

• El mecanismo de las proteínas secretadas son dirigidas al RE durante su traducción. La secuencia de la señal esta constituida aproximadamente 20 aminoácidos incluyendo a un grupo de residuos hidrófobos, habitualmente en el extremo amino-terminal y una cadena polipeptidica.

Salen del ribosoma las secuencias de la señal son reconocidas y unidas a la partícula (PRS) que esta constituida por polipéptidos y un ARN citoplasmático pequeño.

La PRS se une tanto el ribosoma, que la unión del receptor libera a la PRS del ribosoma de la consecuencia de la señal de la cadena polipeptidica en crecimiento.
El ribosomas se une al complejo de translocación de proteínas en la membrana del RE, y la consecuencia de la señal de la cadena polipeptidica en crecimiento.

El ribosoma se une a un complejo de translocación de proteínas en la membrana del RE y la secuencia señal es insertada en un canal de la membrana.
Los translocones que atraviesan la membrana del RE están constituidas por proteínas transmembrana, denominadas proteínas Sec61.
Las proteínas Sec61 de levaduras de mamíferos están relacionadas con las proteínas de la membrana plasmática que translocan los polipéptidos secretados en las bacterias demostrando la conversión maquinaria de las células procariotas y eucariotas.

Muchas proteínas en levaduras, a si como algunas proteínas en las células de los mamíferos, son dirigidas al RE tras haber sido traducidas.
Estas proteínas son sintetizadas en ribosomas citosilicos libres, y su incorporación al RE ya que esta no requiere PRS.
La secuencia señal son reconocidas por proteínas receptoras diferentes asociadas con el translocon en la membrana del RE.
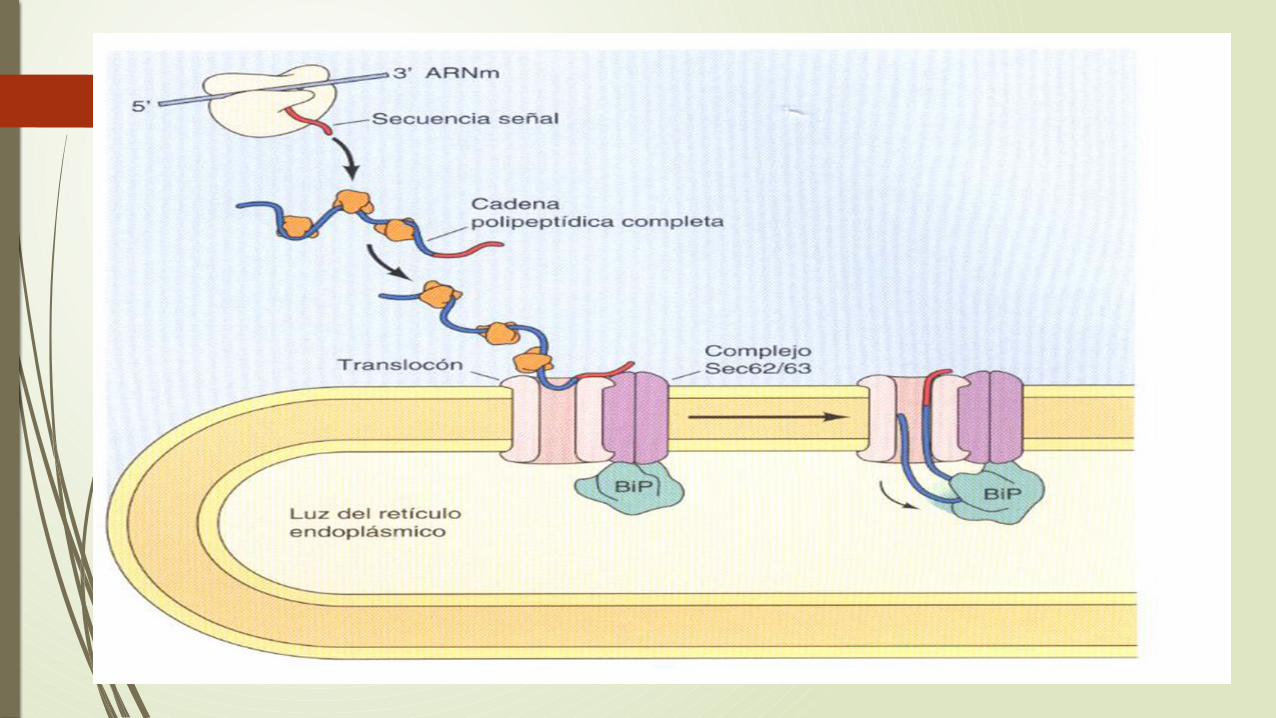

Se adquieren citosilicas Hsp70 para mantener a las cadenas polipeptidicas en su conformación primaria para que puedan penetrar en el translocon y otra Hsp70 en el interior del RE ya que es necesario para que atraviese el canal hasta el interior del RE.
La unión de las cadenas polipeptidicas a BiP es necesaria para dirigir la translocación postraducional de proteínas al RE mientras el crecimiento esta dirigida directamente por el proceso de la síntesis proteica.





























