Embed Size (px)
Citation preview



iii
DEDICATORIA
A la vida porque me demuestra todos los días que todo lo que vale la pena cuesta,
por todas esas maravillosas personas que pone en mi camino.
A mis padres por ser personas ejemplares, por todo el amor y comprensión que
me brindan, por haber puesto mis estudios antes de todo, por trabajar
arduamente todos los días de sol a sol para salir adelante, por haberme regalado
el tesoro más preciado que el propio oro, mis estudios; Por todos esos consejos y
regaños; Por tomar a sus hijos como el motor para seguir adelante, por ser mi
vida.
A toda mi familia por su confianza y apoyo, por haber sembrado en mí sus
ilusiones.
José Luis Elorza Tejada

iv
AGRADECIMIENTOS
Agradezco infinitamente de todo corazón a la Universidad Autónoma Chapingo por
haberme cobijado, por haber hecho realidad mi más grande anhelo – una carrera, por
haberme regalado tantos momentos de felicidad, de tristezas, ilusiones, desilusiones, por
haber confiado en mí y por haberme dado una segunda familia.
Agradezco a mi mamá Carmen Tejada Castro por ser una mujer maravillosa, una mujer
que si es ignorante descubrió los secretos de mi vida con más acierto que un sabio, una
mujer que siendo pobre se satisface con la felicidad de los que ama, mi madre, una
mujer estricta que se estremece con mi llanto, una mujer débil que se revistió tantas
veces con la bravura de un león para protegerme, gracias madre porque a tu lado todos
los dolores se olvidan. Por saber perdonar, por nunca haber mirado atrás y siempre
tener la frente en alto y caminar hacia delante por más pesado que sea el camino.
Agradezco a mi papá Jovito Elorza Romero por toda su dedicación para sacar adelante
a su familia, por haberme enseñado que debes luchar y dar todo en la batalla para
obtener eso que quieres, que es mejor intentar arriesgándolo todo a sentarse
tranquilamente a mirar el desfile de los triunfadores; que si haces algo debes hacerlo con
amor, porque solo así las cosas salen bien, agradezco de todo corazón haberme
demostrado que incluso los héroes se equivocan, lloran, se doblegan, que nadie es
perfecto y que todos tenemos algo de egoístas, por demostrarme que todo es bueno sin
exceso.
Agradezco a mi hermano Gabriel Elorza Tejada por todas las locuras que hicimos de
niños, todas las peleas que tuvimos, por esas lagrimas que lloramos juntos cuando nos
regañaban, por ser mi cómplice y ser siempre un amigo fiel, porque después de las
peleas reíamos y confiábamos otra vez el uno del otro.
Agradezco a mi tía Silvia por ser una segunda madre, a mi tía Vicky por ser mi
confidente, a mi tía Anguie por ser mi amiga y hermana, por todas las peleas que

v
tuvimos, por su protección cuando era niño, a mi tía Alis y Gloria por ser mis
psicólogas, agradezco a todas por el amor que me brindan, por su protección y
consejos. A todas esas personas que su sueño era verme convertido en un hombre de
bien.
Agradezco al Ph. Dr. Ignacio Tovar Luna por toda su dedicación, apoyo y confianza
para realizar este trabajo de tesis, por sus enseñanzas durante la carrera.
Agradezco a todas esas personas que no sólo fueron mis profesores sino amigos,
aquellos que me brindaron su amistad y apoyo, que cada día me daban ánimos para ser
mejor: al Medico José Luis Ortega, al profesor Ramón, al profesor Chalo, al Dr. Ruiz
Torres a todas esas personas que laboran en URUZA y a mis compañeros de grupo que
me brindaron su amistad y que sin duda alguna hicieron de mi estadía en Chapingo un
periodo inolvidable.
Agradezco a Abigail Ramírez Pérez, Keila A. Hernández Mariscal, Gerardo Espinosa
(Gera), Carlos Arnoldo Romero, Carlos Mendoza (Carlitos), Rodrigo Domínguez
(Cachorro), Arturo Beltrán (Woody) y a Perusi por haber sido mi segunda familia, por
todos esos buenos y malos momentos que vivimos juntos, por todo su apoyo y cariño.
Agradezco a Adriana Juárez por su apoyo en la escritura de este trabajo, por confiar en
mí y compartir su historia, por todos los momentos juntos –unos buenos, otros malos
pero maravillosos, por ser diferente y estar conmigo aun cuando no todo va bien, por
soportar todo y brindarme su cariño y amor, porque a pesar de ser infantil conoce de la
vida más de lo que debería. Atte. José Luis Elorza Tejada.
La vida es una obra de teatro que no permite ensayos por eso, canta,
ríe, baila, llora y vive intensamente cada momento de tu vida antes que
el telón baje y la obra termine sin aplausos (Charles Chaplin).

vi
INDICE DE CONTENIDO
ÍNDICE DE CUADROS ................................................................................. viii
INDICE DE FIGURAS ..................................................................................... ix
ABREVIATURAS ............................................................................................ x
RESUMEN ...................................................................................................... xi
ABSTRACT .................................................................................................... xii
INTRODUCCIÓN ............................................................................................ 1
II. OBJETIVOS E HIPÓTESIS ........................................................................ 3
2.1. Objetivos .............................................................................................. 3
2.1.1. Generales ...................................................................................... 3
2.1.2. Particulares ................................................................................... 3
2.2. Hipótesis .............................................................................................. 3
III. REVISIÓN DE LITERATURA ..................................................................... 4
3.1 Población y distribución caprina ............................................................ 4
3.2 Producción caprina ............................................................................... 6
3.3 Sistema de producción caprina en México. .......................................... 8
3.3.1 Sistema extensivo .......................................................................... 8
3.3.2 Sistema intensivo ........................................................................... 8
3.3.3 Sistema semi-intensivo ................................................................... 8
3.5. Necesidades nutritivas ......................................................................... 9
3.5.1. Energía .......................................................................................... 9
3.5.2 Proteína ........................................................................................ 10
3.6. Digestión de las proteínas ................................................................. 10
3.7. Producción de proteína microbiana ................................................... 13
3.8. Proteína de paso ............................................................................... 13
3.8.1. Factores que incrementan la proteína de paso ........................... 15
3.9. Utilización del nitrógeno no proteico .................................................. 18
3.10 Harina de pescado. ........................................................................... 19

vii
3.11. SOY70 ® (Nutrilag, S. A. de C. V.). ................................................. 22
3.12. Efecto del nivel de proteína sobre el comportamiento de cabritos... 24
IV. MATERIALES Y METODOS ................................................................... 26
4.1. Localización del área experimental .................................................... 26
4.2. Formación de grupos experimentales, manejo de animales y
mediciones durante el periodo experimental. ............................................ 26
4.2.1. Etapa1: Prueba de alimentación en corral. .................................. 26
4.2.2 Etapa 2: Medición de la digestibilidad en jaulas metabólicas. ...... 29
4.2.3 Cálculos y análisis estadísticos .................................................... 31
V. RESULTADOS Y DISCUSION ................................................................ 33
VI. CONCLUSIONES y RECOMENDACIONES .......................................... 41
6.1. Conclusiones: .................................................................................... 41
6.2. Recomendaciones: ............................................................................ 41
VII. LITERATURA CITADA ........................................................................... 42

viii
ÍNDICE DE CUADROS
Cuadro 1. Población de caprinos en México ................................................... 5
Cuadro 2. Producción caprina y su valor en la producción animal total en
México. ........................................................................................ 7
Cuadro 3. Nutrientes de la harina de pescado. ............................................. 20
Cuadro 4. Composición del Prolak (as fed1)................................................ 22
Cuadro 5. Análisis bromatológico (% base seca) de soy 70 y de la harina de
soya ........................................................................................... 24
Cuadro 6. Contenido de aminoácidos en Soy 70 (%). ................................. 24
Cuadro 7. Distribución de los animales en las dos fuentes de proteína. ....... 27
Cuadro 8. Composición de las dietas (% base seca). .................................. 29
Cuadro 9. Consumo y digestibilidad por cabritos en crecimiento consumiendo
dietas con dos fuentes de proteína en jaulas metabólicas. .......... 36
Cuadro 10. Consumo de materia seca (CMS) por cabritos en crecimiento
alimentados con dos fuentes de proteína durante 35 días. .......... 37
Cuadro 11. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en crecimiento
consumiendo dietas con dos fuentes de proteína en los primeros
21 días del estudio. ...................................................................... 38
Cuadro 12. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en crecimiento
consumiendo dietas con dos fuentes de proteína en los últimos 14
días del estudio. ........................................................................... 39
Cuadro 13. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en crecimiento
consumiendo dietas con dos fuentes de proteína durante 35 días.
..................................................................................................... 40

ix
INDICE DE FIGURAS
Figura. 1 Población caprina en la Comarca Lagunera, SIAP (2013). ............. 6

x
ABREVIATURAS
°C ............ Grados centígrados
AA ........... Aminoácidos
AAE ......... Aminoácidos esenciales
CO2 .............. Dióxido de Carbono
CEA ........ Centro de Estudios Agropecuarios
CNSP ...... Comité Nacional Sistema Producto Caprinos
CNE ........ Carbohidratos no estructurales
DM .......... Ingestión de Materia seca
EM .......... Energía Metabolizable
ENm .............. Energía Neta de Mantenimiento
FAD ......... Fibra detergente acido
FND ........ Fibra Detergente Neutro
DNDE ...... Fibra Detergente Neutro Efectivo
Grs .......... Gramos
GDP ........ Ganancia diaria de peso
Kg ........... Kilogramos
Mm .......... Milímetros
Ms ........... Materia seca
Msnm Metros sobre el nivel del mar
N ............. Nitrógeno
NH2 ............... Amoniaco
NNP ........ Nitrógeno no proteico
NRC ........ National Reserch Council
PC ........... Proteína Cruda
PB ........... Proteína Bruta
PE ........... Proteína de escape
pH ........... Potencial de hidrogeno
PND ........ Proteína no degradable
PP ........... Proteína de paso
RD ........... Degradabilidad en el rumen
REP Relación de Eficiencia Proteica
SIAP ........ Servicio de Información Agroalimentaria y Pesquera
Ton .......... Toneladas
UIP .......... Consumo de proteína no degradable en rumen
INP .......... Utilización Neta de la Proteína
W0.75 ............ Peso Metabólico

xi
RESUMEN
El objetivo fue determinar el consumo, digestibilidad y la eficiencia de
utilización del alimento en cabritos en crecimiento consumiendo dietas con
dos fuentes de proteína. Se utilizaron 36 cabritos de alrededor de 7 meses
de edad, (24.2±5.8 kg PV), los cuales fueron asignados al azar a uno de dos
fuentes de proteína (PROLAK y SOY70). Los animales fueron alimentados
en corrales individuales durante 38 días con las dietas experimentales (12%
PC y 2.8 Mcal EM/kg MS). Además ocho cabritos de cada dieta se colocaron
en jaulas metabólicas para determinar la digestibilidad de la materia seca y
materia orgánica. Los datos fueron analizados como un diseño
completamente al azar. En la fase de jaulas, el consumo y digestibilidad fue
mayor (P < .05) en la dieta con PROLAK. Sin embargo, en la fase de
alimentación en corral, el consumo fue ligeramente mayor en la dieta con
SOY70 pero sin diferencias significativas (P > .05). La ganancia diaria de
peso, ganancia de peso por cada kg de alimento, y eficiencia de utilización
de la energía metabolizable para ganancia de peso también fueron similares
(P > .05) en ambas ditas. En conclusión, los resultados del estudio indican
que el comportamiento de cabritos en crecimiento es similar consumiendo
dietas con PROLAK y SOY70, por lo que se asume que el valor nutritivo de
ambos productos sea similar.
Palabras Clave: Cabritos, Consumo, Digestibilidad, Eficiencia.

xii
ABSTRACT
The objective was to evaluate feed efficiency in growing kids feeding diets
with two crude protein sources, PROLAK and SOY70. Thirty six 7 months of
age kids (24.2±5.8 kg BW) were randomly assigned to be fed one of two
diets, containing PROLAK or SOY70. The kids were allocated in individual
pens, and fed the experimental diets (12% CP and 2.8 Mcal ME/kg DM) over
a 38 days period. Following this phase, eight kids from each diet were placed
in metabolic cages to measure DM and OM digestibility. Data was analyzed
as completely randomized design. During the cages phase, dry matter intake
and digestibility were greater (P < .01) for the PROLAK diet. However, during
the pen feeding, DMI was slightly greater in the SOY70 diet, but the
differences were not significant (P > .05). Daily gain and gain per kg of feed,
and efficiency of utilization of ME for body weight gain were similar (P > .05)
in both diets. In conclusion, results of this study indicate that kid’s
performance is similar consuming diets containing PROLAK or SOY70.
Key Words: Kids, Intake, Digestibility, Feed Efficiency.

1
INTRODUCCIÓN
La cabra es un animal muy rustico que se adapta fácilmente a
diferentes situaciones del ambiente. Fueron domesticados antes que los
bovinos y ovinos y han servido a la humanidad desde hace largos periodos
de tiempo. Estos animales se desarrollan en ambientes áridos, semi-
tropicales y lugares montañosos. En todo el mundo la cabra vive
principalmente en regiones muy secas; en ellas están 55% de las cabras en
comparación con 39% de los bovinos y 25% de los ovinos existentes
(Attfield et al., 2004). El ganado caprino es una de las especies domesticas
que se destacaron en la ganadería por su gran capacidad para producir
leche, carne, piel, pelo y estiércol, donde otros animales domésticos ni
siquiera podían subsistir (Carrera, 1984).
Las necesidades nutricionales de las cabras varían según el peso, edad
y etapa de crecimiento y/o estado fisiológico. Estas necesidades pueden ser
cubiertas con una variedad de ingredientes alimenticios y el adecuado
programa de alimentación ya que, estas son incapaces de sintetizar todos los
aminoácidos que requieren para formar las proteínas de su cuerpo, por lo
que estas tienen que estar presentes en la dieta en cantidades y
proporciones adecuadas (Castro, 2014), el ingrediente más caro dentro de
una dieta es la proteína por lo que es necesario conocerlas a fondo para
invertir en mejores productos que nos den los resultados más favorables
posibles siempre buscando abaratar la ración sin perder la calidad del
alimento balanceado.
Los costos de alimentación en todo establecimiento ganadero
representan el 60-70 % de los costos totales de la producción. Así, la

2
alimentación es sin duda el factor de mayor relevancia en el éxito del sistema
productivo. El costo de la alimentación de las cabras puede variar
fuertemente según la fuente de suministros de los ingredientes y la
composición de las raciones.
Aunque es importante tomar en consideración en una recomendación
técnica todos los nutrientes requeridos por los animales, la proteína debido al
alto costo de los ingredientes que se usan como fuente de esta, requiere
atención especial (Meda y Maldonado, 2009).
Entre todos los alimentos proteicos vegetales para rumiantes, la pasta
de soya contiene el porcentaje más elevado de aminoácidos esenciales (47.6
%) expresados como porcentaje de la proteína cruda, por otro lado la harina
de pescado tiene una proporción ideal de aminoácidos esenciales altamente
digestibles por lo que resulta muy adecuada en las raciones de animales
jóvenes, la harina de pescado se considera una buena fuente de proteína,
lisina y metionina bypass en rumiantes
En el mercado actual existen diferentes ingredientes que nos
proporcionan proteína, sin embargo, muchas veces compramos ingredientes
caros que nos dan el mismo resultado que otro más accesible
económicamente, con lo anterior, el presente trabajo fue hecho con el
propósito de evaluar dos fuentes de proteína una de origen animal (harina de
pescado) y otra de origen vegetal (pasta de soya), con un mismo nivel de
inclusión en una dieta para cabritos en la fase de crecimiento y de esta
manera evaluar las ganancias de peso y dar a conocer que ingrediente es
más conveniente usar.

3
II. OBJETIVOS E HIPÓTESIS 2.1. Objetivos
2.1.1. Generales
2.1.1.1. Determinar el efecto de dos fuentes de proteína sobre la ganancia
de peso de cabritos en la fase de crecimiento.
2.1.1.2. Generar información que permita hacer recomendaciones a los
productores de la región sobre que ingrediente usar como fuente de
proteína y de esta manera abaratar la ración sin perder la calidad
de la misma.
2.1.2. Particulares
2.1.2.1 Determinar el consumo voluntario del alimento, ganancia de peso,
conversión alimenticia y eficiencia en la utilización del alimento por
cabritos, alimentados con dos diferentes fuentes de proteína.
2.2. Hipótesis
El comportamiento de cabritos en crecimiento será similar
consumiendo dietas con las dos fuentes de proteína, PROLAK y
SOY70.

4
III. REVISIÓN DE LITERATURA
3.1 Población y distribución caprina
Se estima que la población mundial de cabras es de 996, 120, 851
cabezas las cuales se encuentran distribuidas de la siguiente manera: 59.7
% en Asia, 34.6 % en África, el continente americano tiene el 3.6 % de la
población mundial y Oceanía es el continente que menos cabras tiene (0.4
%) (CNSP, 2014).
Se encuentran en todos los países donde hay algún tipo de pastos,
plantas, matorrales o arbustos que ramonear; su capacidad de supervivencia
con casi cualquier tipo de alimento y su rusticidad la han convertido en la
especie doméstica más difundida. De las cabras de nuestro continente más
de la mitad están en Sudamérica, y del resto México contribuye con un hato
de 8.6 millones de cabezas. Es sólo 1.33% del total, contra el 20.61% de
China y el 17.08% de la India, pero éstos son países que poseen 14 y 10
veces más población humana, respectivamente (ANCGCR., 2013). El
inventario nacional de caprinos corresponde a un total de 8,664,613 cabezas
para el año 2013. En el año 2011 el inventario nacional correspondió a 9,004,
377 de cabezas lo que demuestra una reducción en estos últimos dos años
de 339 mil cabezas que representan una pérdida de 3.7 % en el periodo y
1.8 % anual (SIAP, 2013).
La Conafupro (Coordinadora Nacional de las fundaciones produce,
2013) menciona que las 8.6 millones de cabras existentes son explotadas en
194 mil unidades de producción por un millón y medio de productores
5
dedicados a esa actividad en zonas áridas y semiáridas del país, esta
actividad se ha convertido en fuente generadora de ingresos para miles de
mexicanos que han encontrado en la explotación de esta especie un empleo
remunerado en regiones donde la agricultura no tiene posibilidades de
desarrollo.
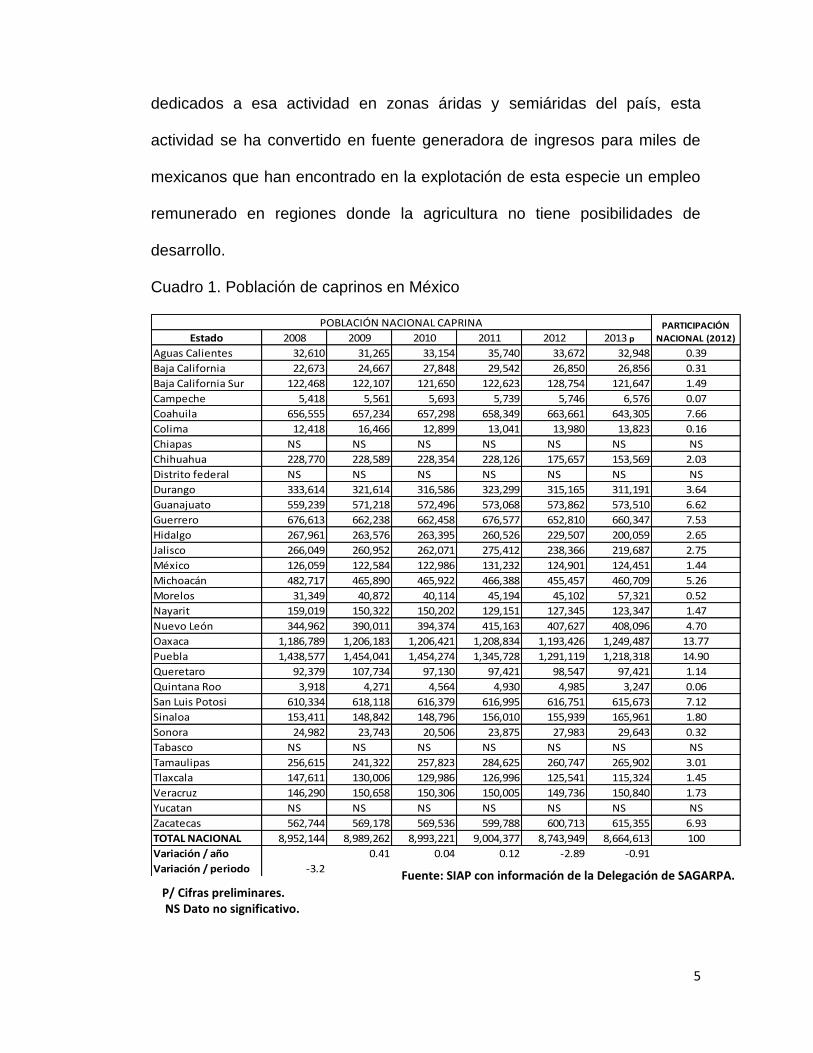
Cuadro 1. Población de caprinos en México
Estado 2008 2009 2010 2011 2012 2013 p
Aguas Calientes 32,610 31,265 33,154 35,740 33,672 32,948 0.39
Baja California 22,673 24,667 27,848 29,542 26,850 26,856 0.31
Baja California Sur 122,468 122,107 121,650 122,623 128,754 121,647 1.49
Campeche 5,418 5,561 5,693 5,739 5,746 6,576 0.07
Coahuila 656,555 657,234 657,298 658,349 663,661 643,305 7.66
Colima 12,418 16,466 12,899 13,041 13,980 13,823 0.16
Chiapas NS NS NS NS NS NS NS
Chihuahua 228,770 228,589 228,354 228,126 175,657 153,569 2.03
Distrito federal NS NS NS NS NS NS NS
Durango 333,614 321,614 316,586 323,299 315,165 311,191 3.64
Guanajuato 559,239 571,218 572,496 573,068 573,862 573,510 6.62
Guerrero 676,613 662,238 662,458 676,577 652,810 660,347 7.53
Hidalgo 267,961 263,576 263,395 260,526 229,507 200,059 2.65
Jalisco 266,049 260,952 262,071 275,412 238,366 219,687 2.75
México 126,059 122,584 122,986 131,232 124,901 124,451 1.44
Michoacán 482,717 465,890 465,922 466,388 455,457 460,709 5.26
Morelos 31,349 40,872 40,114 45,194 45,102 57,321 0.52
Nayarit 159,019 150,322 150,202 129,151 127,345 123,347 1.47
Nuevo León 344,962 390,011 394,374 415,163 407,627 408,096 4.70
Oaxaca 1,186,789 1,206,183 1,206,421 1,208,834 1,193,426 1,249,487 13.77
Puebla 1,438,577 1,454,041 1,454,274 1,345,728 1,291,119 1,218,318 14.90
Queretaro 92,379 107,734 97,130 97,421 98,547 97,421 1.14
Quintana Roo 3,918 4,271 4,564 4,930 4,985 3,247 0.06
San Luis Potosi 610,334 618,118 616,379 616,995 616,751 615,673 7.12
Sinaloa 153,411 148,842 148,796 156,010 155,939 165,961 1.80
Sonora 24,982 23,743 20,506 23,875 27,983 29,643 0.32
Tabasco NS NS NS NS NS NS NS
Tamaulipas 256,615 241,322 257,823 284,625 260,747 265,902 3.01
Tlaxcala 147,611 130,006 129,986 126,996 125,541 115,324 1.45
Veracruz 146,290 150,658 150,306 150,005 149,736 150,840 1.73
Yucatan NS NS NS NS NS NS NS
Zacatecas 562,744 569,178 569,536 599,788 600,713 615,355 6.93
TOTAL NACIONAL 8,952,144 8,989,262 8,993,221 9,004,377 8,743,949 8,664,613 100
Variación / año 0.41 0.04 0.12 -2.89 -0.91
Variación / periodo -3.2
POBLACIÓN NACIONAL CAPRINA PARTICIPACIÓN
NACIONAL (2012)
Fuente: SIAP con información de la Delegación de SAGARPA.
P/ Cifras preliminares. NS Dato no significativo.
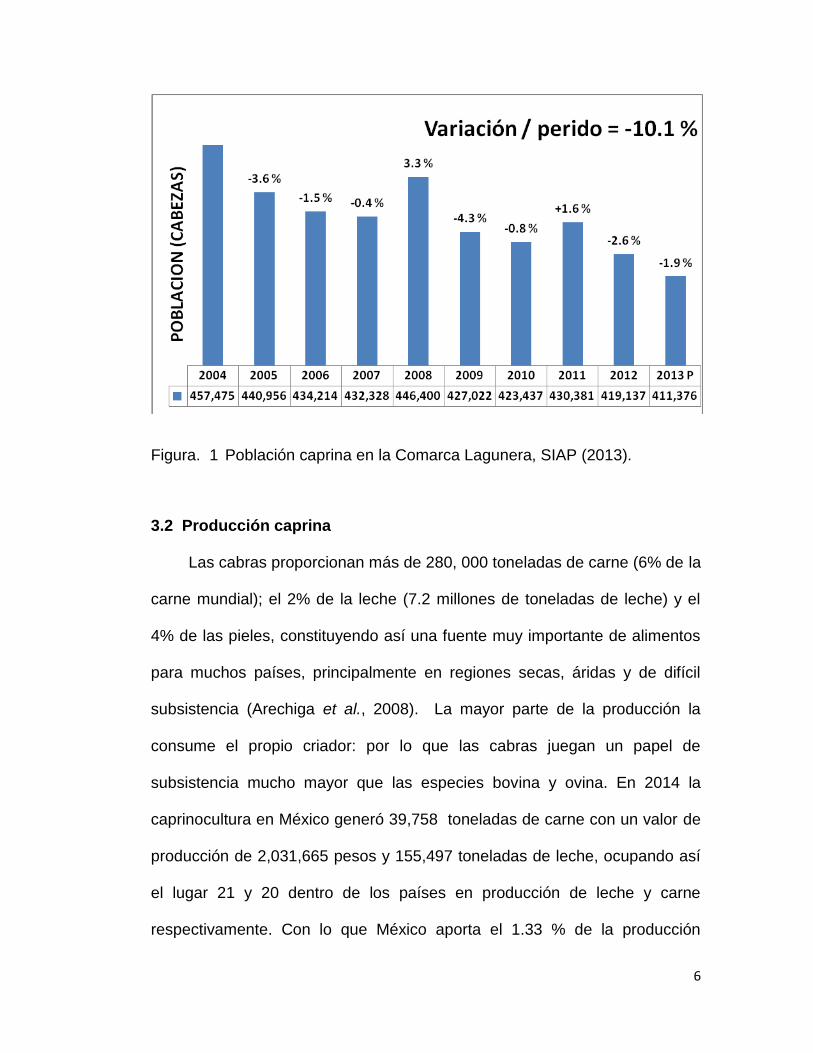
6
Figura. 1 Población caprina en la Comarca Lagunera, SIAP (2013).
3.2 Producción caprina
Las cabras proporcionan más de 280, 000 toneladas de carne (6% de la
carne mundial); el 2% de la leche (7.2 millones de toneladas de leche) y el
4% de las pieles, constituyendo así una fuente muy importante de alimentos
para muchos países, principalmente en regiones secas, áridas y de difícil
subsistencia (Arechiga et al., 2008). La mayor parte de la producción la
consume el propio criador: por lo que las cabras juegan un papel de
subsistencia mucho mayor que las especies bovina y ovina. En 2014 la
caprinocultura en México generó 39,758 toneladas de carne con un valor de
producción de 2,031,665 pesos y 155,497 toneladas de leche, ocupando así
el lugar 21 y 20 dentro de los países en producción de leche y carne
respectivamente. Con lo que México aporta el 1.33 % de la producción

7
mundial (SIAP, 2014). Más de 70% es producido en los sistemas extensivos
de las zonas áridas y semiáridas y aproximadamente 25% en los sistemas
intensivos (ANCGCR, 2013). México se ubica en el lugar 19 dentro de los
países exportadores de carne caprina con 59 toneladas, en los primeros tres
lugares se encuentran Australia (25,149 ton), le sigue Etiopia (12,306 ton) y
China continental con 4,614 toneladas de carne (FAO 2012 citado por CNSP,
2014).
Cuadro 2. Producción caprina y su valor en la producción animal total en
México.
Año Producción
en canal Precio Valor de la producción
Animales sacrificados
Peso Variación de la producción en canal (%)
(toneladas) pesos/kg
(miles de pesos)
(cabezas) (kg)
2004 42,029 34.33 1,442,784 2,546,004 17
2005 42,389 35.97 1,524,691 2,468,920 17 0.86
2006 42,728 37.05 1,582,888 2,486,452 17 0.80
2007 42,873 38.19 1,637,297 2,550,262 17 0.34
2008 43,128 39.18 1,689,931 2,550,863 17 0.59
2009 43,242 39.73 1,717,936 2,530,438 17 0.26
2010 43,867 40.73 1,786,650 2,584,057 17 1.45
2011 43,839 42.14 1,847,235 2,511,108 17 -0.06
2012 41,492 45.00 1,867,178 2,383,778 17 -5.35
2013 39,656 46.75 1,854,062 2,262,579 18 -4.42
2014 39,758 51.10 2,031,665 2,315,639 17 0.26
variación/periodo -5.40
Fuente: SIAP, 2015

8
3.3 Sistema de producción caprina en México.
3.3.1 Sistema extensivo
El sistema extensivo se caracteriza por el pastoreo en extensas áreas
de tierras con pobre productividad, debido a la baja fertilidad del suelo, alto
grado de aridez, escasa precipitación y bajas temperaturas, propiciando baja
producción y calidad del forraje, causas principales de los bajos parámetros
productivos y reproductivos que se observan en la explotación pecuaria en
estos sistemas (Ponce, 2004). Este sistema requiere de grandes extensiones
de terreno ya que las cabras se alimentan pastoreando a voluntad en forma
semi–nómada o sedentaria. Presenta la ventaja de abaratar costos en
alimentación e instalaciones.
3.3.2 Sistema intensivo
En el sistema intensivo los animales se mantienen en praderas
cultivadas con elevada producción de forraje y de excelente calidad nutritiva;
o bien en total confinamiento (Ponce, 2004). Este sistema presenta la
desventaja de requerir mayores costos pero facilita el manejo de los animales
y se obtienen mejores índices productivos en la producción (Arechiga et al.,
2008).
3.3.3 Sistema semi-intensivo
Los animales pastorean y ramonean y en la tarde-noche los animales
se estabulan y se les proporciona un suplemento alimenticio. Requiere de
inversión en instalaciones y alimentos concentrados de gran valor proteico y
energético. Generalmente presenta mejores rendimientos productivos que en
el sistema extensivo (CEA, 2001).

9
3.5. Necesidades nutritivas
3.5.1. Energía
La energía se puede definir como la capacidad de efectuar trabajo, y
trabajo se define como el producto de una fuerza dada que actúa a lo largo
de una determinada distancia. La energía química se puede medir en
términos de calor y se expresa como calorías, la cual se define como: la
cantidad de calor que se necesita para elevar la temperatura de un gramo de
agua de 14.5 grados a 15.5 grados a una atmosfera de presión (Church,
1982).
No toda la energía contenida en el alimento consumido por el animal es
utilizado, una parte de esa energía se pierde a través de las heces después
del proceso de digestión, en forma de alimento sin digerir, microbios
entéricos y sus productos, excreciones al aparto digestivo, y en forma de
restos celulares provenientes del aparato digestivo; otro poco se pierde en
forma de gases durante la fermentación y por medio de la orina, también hay
perdidas en forma de calor durante las reacciones químicas en el proceso de
digestión, absorción y metabolismo de los nutrientes (McDonald et al., 2002).
La fuente primaria de energía para los animales es el sol. Las plantas
verdes convierten la energía luminosa en energía química de modo muy
eficaz, y esta energía, cuando se ingiere por el animal en forma de alimento
se libera en el metabolismo para permitir el trabajo osmótico, el transporte
molecular, el trabajo mecánico, la síntesis de nuevas moléculas o la
formación de energía eléctrica como en el caso de la transmisión nerviosa
(Blaxter, 1964).

10
Los animales obtienen energía de los alimentos. Al consumir los
animales los productos vegetales los componentes se degradan , liberando
la energía, que es empleada por los animales para realizar trabajo
,mecánico, transporte, mantenimiento de la integridad de las membranas
celulares, para procesos de síntesis y para proporcionar calor si las
condiciones ambientales son frías (McDonald, 2002).
3.5.2 Proteína
Las proteínas son muy importantes en la nutrición del rumiante, las
utilizan las partes del cuerpo (sangre, músculos, etc.), sistemas enzimáticos,
sistemas de producción de proteína bacteriana. Están compuestas por
cadenas nitrogenadas de aminoácidos, los cuales tiene la estructura CH
(NH2) COOH. La proteína dietaría es degradada en el rumen a amoniaco y
compuestos carbonados, el amoniaco (grupo amino) es usado por las
bacterias para sintetizar sus propias proteínas (Mues y Walz, 2005).
Los rumiantes obtienen una parte de éstas bacterias que resbalan o son
empujadas al tracto digestivo bajo (estomago e intestino). El sistema
digestivo del rumiante es poco complejo y depende de microorganismos para
digerir alimentos con altos contenidos de celulosa y lignina (elementos
presentes en forrajes y harinas – granos y concentrados), por lo tanto cuando
alimentamos a los rumiantes, debemos abastecer también los requerimientos
proteicos de la flora microbiana (Mues y Walz, 2005).
3.6. Digestión de las proteínas
Los microorganismos del rumen hidrolizan las proteínas, los péptidos y
los aminoácidos, algunos de los cuales posteriormente se degradan a ácidos

11
orgánicos, amoniaco y CO2. Un ejemplo de desaminación es la valina, es
convertida en acido isobutirico. Entonces, los ácidos grasos volátiles de
cadena ramificada que se encuentran en el rumen proceden de los
aminoácidos. El amoniaco producido, junto con los péptidos de cadena corta
y los aminoácidos libres, son utilizados por los microorganismos del rumen
para sintetizar sus proteínas, que posteriormente se digieren en el abomaso
e intestino delgado. Un aspecto importante de la síntesis de proteína
microbiana es que las bacterias pueden utilizar tanto aminoácidos
indispensables como no indispensables, lo que asegura al animal hospedero
un aporte de los primeros, independientemente de su contenido en la dieta
(Van Soest, 1994).
En la mayoría de las dietas, la mayor parte (y en ocasiones el total) de
la proteína que llega al intestino delgado es proteína de origen microbiano,
cuya composición es bastante constante. Una cantidad menor
correspondería a la proteína del alimento que no se ha degradado, la cual
tendrá una composición en aminoácidos variable, dependiendo de la
naturaleza de la dieta (Haresign y Cole, 1988).
El contenido de amoniaco del líquido ruminal refleja el resultado de los
procesos de degradación y de síntesis proteínica. Una baja en el contenido
ruminal de amoniaco puede suceder con una dieta deficiente en proteína o si
ésta es resistente a la degradación. Por otro lado, si la degradación de las
proteínas es más rápida que la síntesis, habrá un aumento de amoniaco en
el liquido ruminal, sobrepasándose la concentración óptima. Cuando esto
llega a suceder, el amoniaco pasa a la sangre y se transporta al hígado,

12
donde se transforma en urea. Una pequeña cantidad de esta urea vuelve al
rumen con la saliva o directamente a través de la pared ruminal, pero la
mayor parte se excreta en la orina y por tanto se desperdicia (Church y Pond,
1976).
Si el alimento es pobre en proteína y la concentración de amoniaco del
líquido ruminal es baja, la cantidad de nitrógeno que retorna al rumen en
forma de urea puede exceder a la absorbida en el rumen en forma de
amoniaco. Esta ganancia neta de nitrógeno reciclado se transforma en
proteína microbiana, por lo que la cantidad de proteína que llega al intestino
puede superar a la aportada con el alimento. En esta forma el rumiante,
mediante el retorno de la urea, puede conservar nitrógeno que de otra
manera se excretaría en la orina (Orskov, 1982).
Los microorganismos del rumen tienen por tanto un efecto nivelador en
el suministro de proteínas, complementan cuantitativa y cualitativamente
alimentos gruesos o toscos, tanto de baja cantidad como calidad, pero tiene
un efecto perjudicial en los concentrados ricos en proteínas. En la actualidad
es frecuente aprovechar la capacidad de síntesis de las bacterias del rumen,
complementando con urea la dieta de los rumiantes. Una innovación mas
reciente consiste en proteger de la degradación en el rumen a las proteínas
de buena calidad, ya sea tratándolas químicamente (con formalina) para
reducir su solubilidad, o bien administrándolas en suspensiones liquidas que
puedan atravesar el rumen mediante el conducto esofágico (McDonald,
1998).

13
3.7. Producción de proteína microbiana
Las necesidades de mantenimiento de los microorganismos, que se han
estimado son de 0.022 y 0.187 g de carbohidratos por g de bacterias por
hora. Si la fermentación es lenta, como ocurre con las raciones ricas en
carbohidratos estructurales, los gastos de mantenimiento pueden ser
importantes y las estimaciones de la producción microbiana resultar
exageradas. El medio ambiente ruminal, Se ha comprobado que el descenso
del pH en el rumen de 6.7 a 5.7 reduce a la mitad la producción de proteína
microbiana, lo cual puede ser importante al administrar raciones ricas en
carbohidratos solubles y de bajo contenido en fibra, con la siguiente
producción de acido láctico y acidosis ruminal (McDonald et al., 2002).
Las variaciones en las formas del nitrógeno necesario para los distintos
tipos de microorganismos. Por ejemplo, los microorganismos que inciden los
CNE pueden utilizar nitrógeno peptídico y amoniacal, en tanto que los que
inciden los carbohidratos estructurales no pueden usar el nitrógeno de los
aminoácidos y han de depender del amoniaco como fuente de nitrógeno
(McDonald et al., 2002).
3.8. Proteína de paso
A la proteína de la dieta que escapa a la fermentación ruminal se
transporta al tracto digestivo bajo se le llama proteína de paso (PP) o
proteína de escape (PE), para diferenciarla de la proteína sintetizada por
microbios ruminales y de las secreciones endógenas (NRC, 1985).
La proteína de la dieta que pasa al abomaso consiste en dos
fracciones: 1) La proteína que evade al ataque de microorganismos en el

14
rumen y que a través de la ranura esofágica pasa al abomaso sin mezclarse
completamente con el contenido ruminal, al la cual se le denomina proteína
bypass; esto pasa en los rumiantes jóvenes (Orskov, 1972), y 2) la proteína
que resiste el ataque microbiano en el rumen (NRC, 1985). La cantidad de
proteína no degradable en rumen de la dieta puede ser una cantidad
significativa que genere una respuesta de eficiencia en los rumiantes (Van
Soest, 1994). Del nitrógeno total de los microorganismos ruminales, solo del
60 al 70 % está en forma de proteína verdadera, el remanente está en ácidos
nucléicos y peptidoglucanos (Chalupa, 1972). Por tanto, si la proteína de
calidad se fermenta, una mayor porción se convierte en productos no
disponibles. La proteína de escape puede ser alterada por la manipulación
de las tasas de digestión y la tasa de pasaje. La cantidad de proteína de
escape del rumen es variable y depende del tipo de proteína, su tasa de
degradación, su nivel de consumo, su tasa de pasaje y de otros factores. En
contraste, los factores como tasa de pasaje y tasa de degradación no tienen
efecto sobre la proteína de paso, ya que esta nunca entra al rumen para
sujetarse a degradación ruminal (Van Soest, 1994).
Hill (1991) estimó la degradabilidad ruminal de la harina de semillas, y
obtuvo que fluctuarón entre 59 y 77%; la variación de estos valores de
degradabilidad podría atribuirse a diferentes factores, entre ellos, altas
temperaturas durante las fases de tostado que pueden provocar una baja
degradabilidad ruminal, y tener un mayor valor de proteína de escape.
Titgemeyer et al. (1989) reportaron que 92 % del N en la harina de sangre
escapa a la degradación ruminal, en comparación con un 21 % de la harina

15
de soya. Sin embargo, algunas fuentes de proteínas resistentes a la
degradación ruminal, como productos destilados, proteínas tratadas
químicamente y materiales dañados por el calor, pueden tener un balance
inferior de AA o contener altas cantidades de N indigestible (Owens y Berger,
1983). Por otra parte, cuando se alimenta con proteínas de alto pasaje o se
produce una sobreprotección de la proteína por diversos tratamientos, se
puede generar proteína totalmente indigestible (Amos, 1980), lo cual forzaría
a los microbios ruminales a que dependieran de la urea reciclada, la que es
inadecuada como única fuente para apoyar la fermentación; el crecimiento
de éstos seria bajo, y se podría afectar el consumo de alimentos así como su
digestibilidad (Van Soest, 1994).
Los forrajes frescos contienen casi la mitad de proteína verdadera en
forma hidrosoluble, rápidamente fermentable. Es probable que mucha de
esta proteína se degrade en el rumen. La alta humedad de los ensilajes
origina una situación similar, aunque mucho de ese N hidrosoluble es NNP.
En estos casos, el escape de N es de 10 - 30 % de la cantidad total, pero
considerando que cerca de 5 - 15 % del N del forraje forma un complejo de
N-Lignina, totalmente indigestible, entonces la proteína de escape disponible
es la diferencia, por lo que puede ser solo de 0-25 % del N consumido de
forrajes frescos y ensilados (Van Soest, 1994)
3.8.1. Factores que incrementan la proteína de paso
Naturaleza física de la proteína: es de importancia fundamental conocer
si la proteína es soluble y se mueve en líquidos o es insoluble y se mueve en
el material solido. La materia líquida pasa por el rumen mucho más rápido

16
que el material solido, debido a la presión osmótica y el lavado hacia el
exterior. El paso del material solido es una función del consumo de la pared
celular, la ruminación y el tamaño de la partícula (Van Soest, 1994). El
tamaño de la partícula es importante. Si el forraje es deshidratado y molido,
la alteración física tendría un gran efecto sobre el escape de rumen.
Concentración de proteína en la dieta, también tiene influencia en el
escape de rumen, ya que el pasaje de cualquier ingrediente depende de su
concentración en el rumen por esta razón se puede esperar que alimentos
con altos contenidos de proteína provoquen el escape de grandes cantidades
de la proteína de la dieta, y que las proteínas menos degradables puedan
mostrar un gran escape (Van Soest, 1994).
Consumo elevado de alimentos provoca un pasaje rápido y por lo tanto
incrementa marcadamente la proteína de escape en el ganado lechero
(Tamminga, 1979) y novillos (Zinn y Owens, 1983); las mismas aplicaciones
se consideran para forrajes molidos y peletizados (Van Soest, 1994). La
cantidad de PE en vacas lactantes que comían 8.2 ó 12.9 kg de MS
diariamente fue de 29 y 45 %, respectivamente (Tamminga, 1979). Los
rumiantes con alta producción consumen altas cantidades de alimento y
comúnmente tienen un gran porcentaje de PE, aquellos animales que comen
bajas o moderadas cantidades de alimento. El pH ruminal puede afectar la
degradación proteínica por alteración de la actividad de las enzimas
proteolíticas y desaminasas, así como modificar la solubilidad de las
proteínas. El pH optimo para la actividad de estas enzimas es entre 6 y 7
(Tamminga, 1979); un cambio en este intervalo podría alterar la solubilidad

17
de la proteína con punto isoeléctrico, lo que alteraría la degradabilidad
ruminal (NRC, 1982). La fibra también puede limitar el acceso microbial a la
proteína del forraje, y la digestión reducida de la fibra a un pH bajo podría
también estar involucrado (Ganev et al., 1979).
Tipo de dieta también influye en la proteína de sobrepaso. El ganado
que se alimenta con dietas de forraje, en comparación con el que se alimenta
a base de dietas altas en proteína in vitro e in vivo (Zinn y Owens, 1983 b).
Tiempo de retención en rumen es otro factor que puede influir en la
degradación de la proteína. Las proteínas retenidas por un corto tiempo se
degradan en menor grado que las que tienen gran tiempo de retención (NRC,
1985). El tiempo de retención de los ingredientes de la dieta varía entre
animales (Balch y Campling, 1965), entre especies y entre los ingredientes
de la dieta (Church, 1982). El tiempo de retención es también influido por el
tamaño de la partícula del alimento, así como por el nivel de consumo (NRC,
1985), aunque según Varga y Prigge (1982), el nivel de consumo tiene poco
efecto sobre el tiempo de retención, el impacto es sobre la degradación de la
proteína menor o no hay efecto (McAllan y Smith, 1983).
La tasa de dilución del fluido ruminal aumentada eleva la tasa de
dilución del fluido ruminal, se incrementa el flujo de proteína hacia el
abomaso en borregos y novillos (Prigge et al., 1978). Parte de este aumento
probablemente se debe a un incremento neto en la PCB y otra parte debido a
un incremento en la cantidad de proteína de escape (Hemsley, 1975).
El tiempo de resistencia ruminal puede verse afectado por la
temperatura ambiental. Kennedy et al. (1976) demostraron que borregas en

18
un ambiente frío, tuvieron una elevada tasa de pasaje de la ingesta. Lo
anterior incrementa la PCB y la cantidad de PE. Así mismo, Kennedy et al.
(1982) encontraron que el porcentaje de PE en el rumen se incrementó de
20-24% para el heno de alfalfa y de 40-49 % para el heno de bromegrass,
cuando los borregos se expusieron a temperaturas frías. Por tanto, el
incremento en la proteína de escape puede deberse a la disminución del
tiempo de permanencia de los alimentos en el rumen y al cambio de las
características de fermentación ruminal.
3.9. Utilización del nitrógeno no proteico
No solo la proteína degradable de origen alimenticio contribuye con el
amoniaco del rumen. En los alimentos utilizados por los rumiantes, hasta
30% del N puede encontrarse en forma de compuestos orgánicos simples
como aminoácidos, amidas y aminas, o de compuestos inorgánicos como los
nitratos. La mayoría de estos se degrada rápidamente a portando su N al
pool de amoniaco. En la práctica, es posible aprovechar la capacidad de los
microorganismos del rumen para transformar los compuestos nitrogenados
no proteínicos (NNP) en proteínas, mediante la adición de aquellos en la
dieta. El compuesto más usado es la urea pero pueden utilizarse varios
derivados de ésta incluso el amoniaco (McDonald, 1998).
La urea que entra al rumen se hidroliza rápidamente y se convierte en
amoniaco gracias a la ureasa de origen bacteriana, por lo que la
concentración de amoniaco en el rumen puede aumentar considerablemente.
Para que este amoniaco se incorpore en forma eficiente a la proteína
microbial se han de cumplir dos condiciones: en primer lugar, la

19
concentración inicial de amoniaco debe ser inferior al nivel optimo (de lo
contrario, el amoniaco producido en exceso será absorbido y eliminado por el
animal), y en segundo lugar, los microorganismos deben disponer para la
síntesis proteínica de una fuente de energía rápidamente utilizable. Con el
propósito de que estas condiciones se cumplan, se suele administrar en la
práctica la urea mezclada con otros alimentos (para prolongar el periodo en
que es ingerida y desaminada). Tales alimentos deben tener concentración
baja en proteína degradable y alta en carbohidratos rápidamente
fermentables (Church y Pond, 1976).
Es importante evitar el sobre consumo accidental de la urea ya que la
rápida absorción siguiente de amoniaco en el rumen o pudiera sobrepasar la
capacidad del hígado para transformarlo en urea, provocando una
concentración de amoniaco en sangre periférica que alcance niveles tóxicos.
Otro compuesto nitrogenado no proteínico que puede ser utilizado por las
bacterias del rumen y, por tanto, por el rumiante es el acido úrico. Las heces
de las aves tienen gran cantidad, y a veces se utilizan deshidratadas para
complementar las dietas de los rumiantes (Church, 1976).
3.10 Harina de pescado.
La utilización de la harina de pescado en la formulación de alimentos
para aves, ganadería, cultivos de camarones o langostinos y peces, ha sido
cuestionada por las autoridades sanitarias de muchos países importadores,
juzgándola como fuente probable de agentes causales de diversas
patologías enfermedades o de estar contaminada por dioxinas, micotoxinas u
otros agentes patógenos (Grau et al., 2007).

20
Desde el punto de vista nutritivo, el proceso de desecación es muy
importante puesto que el desecado excesivo puede reducir significativamente
la calidad del producto (Cuadro 3). El componente nutritivo más valioso de la
harina de pescado es la proteína. La degradabilidad media de la proteína es
de alrededor de 40%, pero es altamente variable, dependiendo del grado de
deterioro durante el almacenamiento y de la cantidad de solubles reciclados.
Probablemente, el principal determinante de la calidad proteica guarde
relación con el grado y duración del tratamiento térmico (McDonald et al.,
2002).El contenido en minerales es alto (100 a 220 g/kg), lo que resulta muy
importante desde el punto de vista de la nutrición, ya que aporta buenas
cantidades de calcio y fosforo, así como una serie de micro-elementos como
hierro, yodo y manganeso. A demás, son buena fuente de vitaminas del
complejo B, especialmente colina, B12 y riboflavina (McDonald et al., 2002).
Cuadro 3. Nutrientes de la harina de pescado.
Nutriente %
Materia seca 90
Proteína cruda 66
UIP (% de PC) 60
PD (% de PC) 40
Gras*a 8
Cenizas 10
FC 1
FDN 12
FDA 2
FDN efectiva (% FDN) 10
Calcio) 5.5
Fosforo 3.15
TDN 74
Digestibilidad 90
NRC (2009)

21
La energía de las harinas de pescado se encuentran totalmente en
forma de grasa y proteína, y guarda relación con su contenido en aceite. La
tendencia actual en el mercado de las harinas de pescado se dirige a los
productos especiales, fabricados para ajustarse a las distintas especies. Los
rumiantes adultos pueden obtener aminoácidos y vitaminas del grupo B a
partir de la síntesis microbiana, de modo que la importancia de la harina de
pescado para estos animales radica en su valor como suplemento de
proteína no degradable. Este aspecto, tiene importancia especial en los
animales en crecimiento rápido y las hembras gestantes. Para las vacas
lecheras, la ingestión diaria de harina de pescado debe limitarse a 1 kg como
máximo (McDonald et al., 2002).
3.9.1. PROLAK (H.J. BAKER & BRO., INC)
El PROLAK está hecho a base de productos de origen animal y
marino diseñado para complementar la proteína sintetizada por los
microorganismos del rumen. En el cuadro 4 se muestra la composición típica
del PROLAK.

22
Cuadro 4. Composición del PROLAK (as fed1) Nutriente % Aminoácido %
Humedad 7.69 Lisina 4.46
Proteína 70.3 Leucina 6.31
Grasa 8.02 Isoleucina 2.28
Cenizas 12.5 Valina 4.72
Calcio 3.33 Fenilalanina 3.76
Fósforo 1.77 Treonina 3.01
UIP 68.0 Metionina 1.10
TDN 70.8 Arginina 4.2
ENl (Mcal/kg) 1.61 Triptofano 0.6
Histidina 2.04
1 tal como se ofrece.
3.11. SOY70 ® (Nutrilag, S. A. de C. V.).
Es un suplemento alimenticio de soya con elevado contenido en
proteínas de sobrepaso, altamente digestible para su uso en la alimentación
de rumiantes. Entre todos los alimentos proteicos vegetales para rumiantes,
la pasta de soya contiene el porcentaje más alto de aminoácidos esenciales
(47.6 %) expresados como porcentaje de proteína cruda, y su índice de
aminoácidos esenciales de Bypass es muy próximo al de las proteínas de los
microorganismos del rumen y muy especialmente a la lisina que es el primer
aminoácido limitante para la producción de leche y proteína en las raciones
comunes a base de maíz grano y ensilaje de maíz. sin embargo la pasta de
soya común tiene una reducida eficiencia proteica por la importante
degradación de su proteína en el rumen. El SOY70 es el ingrediente con
mayor contenido en proteína de bypass. Es una proteína 100 % de origen
vegetal, es altamente digestible y de calidad constante, no tiene límite de
inclusión, permite la satisfacción de las necesidades de proteína

23
metabolizable sin exceder la proteína degradable de las raciones, es el
suplemento de elección para la satisfacción de las necesidades mínimas de
lisina metabolizable de las vacas lecheras. La inclusión de SOY70 en las
raciones de vacas lecheras permite obtener un mayor consumo de materia
seca, mayor producción de leche y picos de producción más elevados
(Nutrilag, 2013).
24
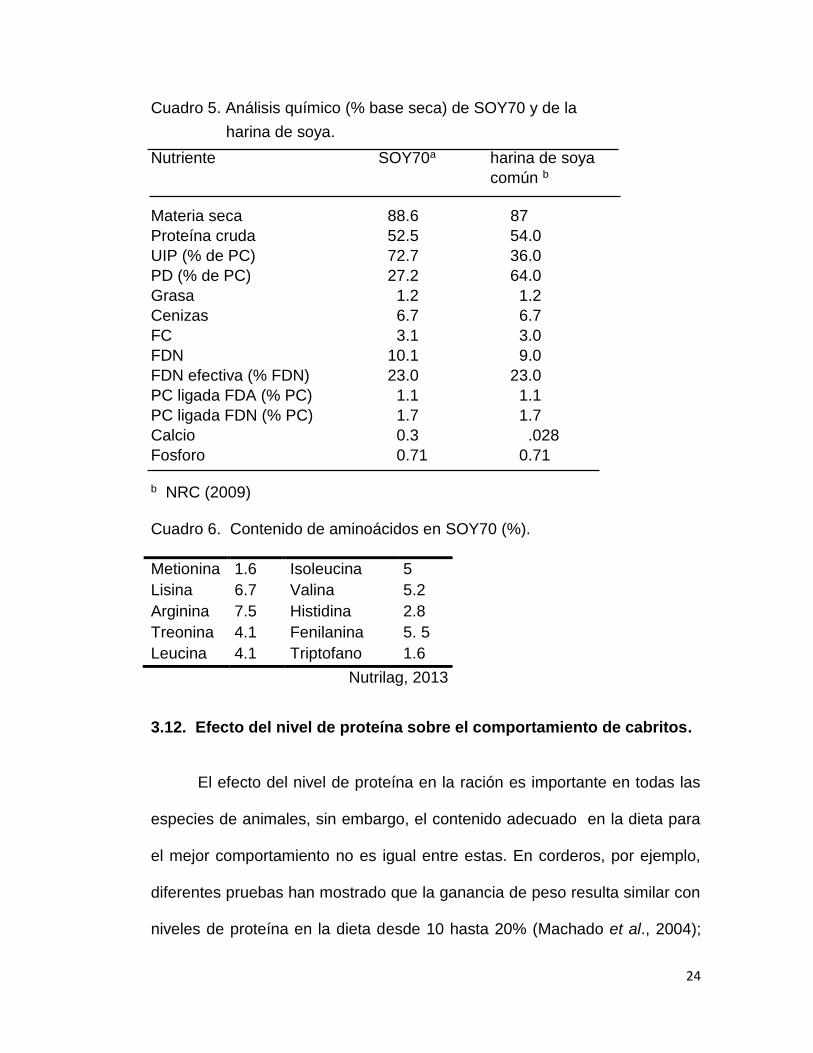
Cuadro 5. Análisis químico (% base seca) de SOY70 y de la
harina de soya.
Nutriente SOY70a harina de soya
común b
Materia seca 88.6 87
Proteína cruda 52.5 54.0
UIP (% de PC) 72.7 36.0
PD (% de PC) 27.2 64.0
Grasa 1.2 1.2
Cenizas 6.7 6.7
FC 3.1 3.0
FDN 10.1 9.0
FDN efectiva (% FDN) 23.0 23.0
PC ligada FDA (% PC) 1.1 1.1
PC ligada FDN (% PC) 1.7 1.7
Calcio 0.3 .028
Fosforo 0.71 0.71 b NRC (2009) Cuadro 6. Contenido de aminoácidos en SOY70 (%).
Metionina 1.6 Isoleucina 5
Lisina 6.7 Valina 5.2
Arginina 7.5 Histidina 2.8
Treonina 4.1 Fenilanina 5. 5
Leucina 4.1 Triptofano 1.6
Nutrilag, 2013
3.12. Efecto del nivel de proteína sobre el comportamiento de cabritos.
El efecto del nivel de proteína en la ración es importante en todas las
especies de animales, sin embargo, el contenido adecuado en la dieta para
el mejor comportamiento no es igual entre estas. En corderos, por ejemplo,
diferentes pruebas han mostrado que la ganancia de peso resulta similar con
niveles de proteína en la dieta desde 10 hasta 20% (Machado et al., 2004);

25
Hadaad et al., 2001; Dabiri et al., 2004). La información disponible referente
al nivel de proteína en la dieta de cabritos es limitada. Las tablas del NRC
recomiendan diferentes cantidades de proteína en la alimentación de cabritos
en crecimiento con diferente peso y tasa de crecimiento. En el estudio
reportado por Galicia y Nevárez (2015) alimentando cabritos de 5 meses de
edad con dietas de 12 y 14% de PC en la ración, las diferencia en GDP
resulto estadísticamente no significativas, aunque la GDP en aquellos
cabritos consumiendo la dieta con 14% de PC fue numéricamente mayor. En
el estudio reportado por Urge et al., (2004) trabajando con cabritos Alpina,
Angora, Boer y españoles, alimentados con dietas de 50% y 75% de
concentrado (17% de PC y 2.7 y 2.37 Mcal/kg de EM, respectivamente), la
GDP entre cabritos Aplina, Angora y Boer fue similar, y los españoles
tuvieron la GDP más baja.

26
IV. MATERIALES Y METODOS
4.1. Localización del área experimental
La presente investigación se realizó en la Unidad Regional
Universitaria de Zonas Áridas, al noreste a 3 km de Bermejillo, Durango, en
el municipio de Tlahualilo, Durango, entre las coordenadas utm 639935 E y
2864331 N (universal transversa mercator), las cuales corresponden a las
coordenadas geográficas 25° 53’ 32’’ longitud norte y 103° 36’ 11’’ longitud
oeste, a una altura de 1117 msnm.
De acuerdo con la clasificación de Köppen modificada por García
(1988), el área presenta un clima muy seco y extremoso semicálido con
lluvias de verano e inviernos fríos, el clima de la región se clasifica como
BW(h´)hw(e). La temperatura anual es de 20.8°C con una mínima promedio
en el invierno de 3.9°C y una máxima promedio en el verano de 36.1°C
(Cornet, 1988), las lluvias son en verano, con un porcentaje de lluvias
invernales que va de 5 a 10 % (García, 1973).
La precipitación es muy escasa en esta región, oscila desde los 200
mm hasta los 250 mm, con un promedio de 201 mm anuales mientras que la
evaporación anual es del orden de los 2600 mm. La máxima precipitación
ocurre entre los meses de agosto y septiembre (CONAGUA, 2009).
4.2. Formación de grupos experimentales, manejo de animales y mediciones
durante el periodo experimental.
4.2.1. Etapa1: Prueba de alimentación en corral.
El estudio se inició el día 23 de Diciembre de 2013. Un total de 36 cabritos
de alrededor de 6 meses de edad (28 Alpina (10 hembras y 18 machos

27
intactos; 5 Toggenburg (3 hembras y 2 machos intactos; y 3 Saanen (1
hembra y 2 machos)) se seleccionaron en la Unidad de Producción Caprina
de la Unidad Regional Universitaria de Zonas Áridas y desparasitados con
Vermectin (BROEL S.A. de C. V., México, D. F.) y dosificados con vitamina
A, D, y E (Adler Pharma S. de R. L. de C. V., Jalisco, México). Los animales
fueron alimentados con las dietas experimentales por dos semanas como
adaptación al manejo y rutina de trabajo diario.
Al final del periodo de adaptación, los treinta y seis cabritos fueron
estratificados de acuerdo a su peso vivo (24.2±5.8 kg de peso) y asignados
al azar a una de dos dietas, o SOY70 (Cuadro 8), teniendo ambos grupos de
animales similar peso vivo promedio y la misma variación. Además, los
animales fueron asignados a los corrales tratando que se tuvieran animales
de ambos tratamientos distribuidos en todas las ubicaciones del área de
trabajo. Los corrales tenían sombra y bebederos automáticos.
Cuadro 7. Distribución de los animales en las dos fuentes de proteína.
Fuente de proteína
Animal SOY70 PROLAK
Machos
Alpina 9 9
Togenburg 1 1
Saanen 0 2
Hembras
Alpina 7 3
Togenburg 1 2
Saanen 0 1
TOTAL 18 18

28
El periodo de alimentación con las dietas experimentales (Cuadro 8)
tuvo una duración de 38 días, durante los cuales los animales fueron
alimentados ad libitum y 5% más de alimento de lo consumido en la última
semana, ofreciendo la mitad del alimento a las 7 am y la otra mitad a las 4
pm. Los animales se pesaron cada semana, en la mañana antes de ser
alimentados con aproximación de 0.1 kg.
Durante el periodo de alimentación se tomaron muestras de alimento
cada semana, y se colectó la totalidad del alimento rechazado diariamente.
Al final del periodo de alimentación, las muestras de alimento ofrecido de
cada ración se mezclaron para formar muestras compuestas, y del alimento
rechazado también se formaron muestra compuesta por animal; tanto del
alimento ofrecido y rechazado se tomaron dos submuestras de cada dieta o
animal, las cuales fueron deshidratadas en estufa de aire forzado a una
temperatura de 55°C por 48 hrs para determinar la materia seca y fueron
molidas en un molino Thomas Willy usando una criba de 1 mm.
Posteriormente submuestras de aproximadamente 2 gramos fueron
deshidratadas a 105°C por un período de 4 hrs para la obtención de la
materia seca total, y las mismas submuestras se incineraron en la mufla a
550 °C por un periodo de 4 hrs. para conocer los porcentajes de materia
orgánica de cada muestra.

29
Cuadro 8. Composición de las dietas (% base seca).
Ingrediente Fuente de proteína
PROLAK SOY70
Heno de alfalfa 17.3 19.6
Avena 14.2 13.1
Maíz rolado 56.6 52
SOY70 0.0 5.5
PROLAK 3.4 0.0
Megalac 0 1.3
Melaza 3 3
Minerales 1 1
Vitaminas .5 0.5
Bicarbonato 2 2
Sal 2 2
Composición química*
PC, % 12 12
DIP, % 7.88 7.75
UIP, % 4.10 4.21
EM, Mcal / kg 2.81 2.81
ED, Mcal / kg 3.27 3.11
Ca, % 0.44 0.3
P, % 0.22 0.15 *Valores estimados en base a valores de las tablas de NRC (1981).
4.2.2 Etapa 2: Medición de la digestibilidad en jaulas metabólicas.
Con el objetivo de determinar la digestibilidad de la materia seca y de
la materia orgánica de las diferentes raciones, al final de la prueba de
alimentación en corral se tomaron ocho cabritos al azar de cada tratamiento,
con peso vivo homogéneos (35±5.5 kg). Los animales fueron pesados antes

30
se subirse a las jaulas metabólicas y el último día de las mediciones. La
rutina de trabajo seguida en el manejo y alimentación de los animales, las
mediciones y toma de muestras para la medición de la digestibilidad fue
similar a la descrita por Cochran y Galyean (1994). El alimento se ofreció
dos veces al día a las 7:00 am y 4:00 pm. Con el objeto que los animales
fueran alimentados ad libitum, la cantidad de alimento ofrecido diariamente
fue un 5% más del consumido el día anterior. Los animales fueron
alimentados en las jaulas metabólicas durante catorce días; los primeros
nueve días se consideraron como adaptación a la jaula, rutina de actividades
de alimentación, limpieza de comeros y jaulas. Los últimos 7 días como
período de recolección de datos o experimental, registró la cantidad de
alimento ofrecido y alimento rechazado, y se registro la cantidad de heces
excretadas y se colectó el 25% de lo excretado. Las muestras de alimento
ofrecido y rechazado se conservaron en congelador a -20°C hasta su
posterior análisis en el laboratorio.
La muestras de heces, muestras de alimento ofrecido y rechazado
fueron deshidratadas en estufa de aire forzado a una temperatura de 55°C
por 48 h. Las muestras secas de ofrecido, rechazo y heces fueron molidas
en un molino Thomas Willy usando una criba de 1 mm. Posteriormente se
determino el contenido de de humedad y cenizas (AOAC, 2002).

31
4.2.3 Cálculos y análisis estadísticos
El consumo de alimento se estimó por diferencia de alimento ofrecido
y alimento rechazado diariamente. El consumo de materia seca se estimó
usando los valores de contenido de humedad tanto del alimento ofrecido y
rechazo (Burns et al., 1994). Los animales fueron pesados cada semana
antes de ofrecer alimento por la mañana. La ganancia diaria de peso se
estimó por diferencia de peso final menos el inicial de cada periodo dividido
entre el número de días que duro el periodo.
Los requerimientos de EMm fueron estimados en base al peso
metabólico (PV0.75) y el requerimiento de EM indicado en las tablas del NRC,
101.38 kcal/ kg PV0.75 (NRC, 1981). El consumo de EM se estimó a partir del
contenido de ED por cada kg de materia orgánica digestible (MOD) (4.62
Mcal ED/kg MOD, NRC, 1981), y asumiendo una eficiencia de utilización de
la ED a EM del 82% (ARC, 1980). La energía retenida en la ganancia de
peso se estimó asumiendo un contenido de energía por cada kg de ganancia
de 4.09 Mcal (NRC, 1981). La eficiencia de utilización de la energía
metabolizable para ganancia de peso se estimó dividiendo la energía
retenida en la ganancia de peso (REg) por la energía metabolizable usada
para ganancia de peso (EMg), la cual se estimó de la diferencia del total de
EM consumida menos la EMm. La eficiencia de utilización del alimento para
ganancia de peso se estimo dividiendo la ganancia diaria de peso dividido
por el consumo diario de alimento, expresando en gramos de ganancia por
cada kg de alimento consumido.

32
Los datos fueron analizados como un diseño completamente al azar.
La comparación de medias fue hecha por la prueba de Diferencia Mínima
significativa (Steel y Torrie, 1980). Todos los análisis estadísticos se
realizaron usando el procedimiento GLM del paquete SAS (2007) de acuerdo
al siguiente modelo:
Yij = µ + FPj + Ɛij
Donde:
i= 1, 2
j= 1…18
Y ij = Respuesta en el i-ésimo nivel de proteína.
μ= Media general, común a todos los animales.
FP j = Efecto del j-ésimo fuente de proteína.
E ij= Error experimental asociado a la ij-ésima unidad experimental.
Ɛijk = Error aleatorio (ɛ ij ~ N (0, σ2)

33
V. RESULTADOS Y DISCUSION
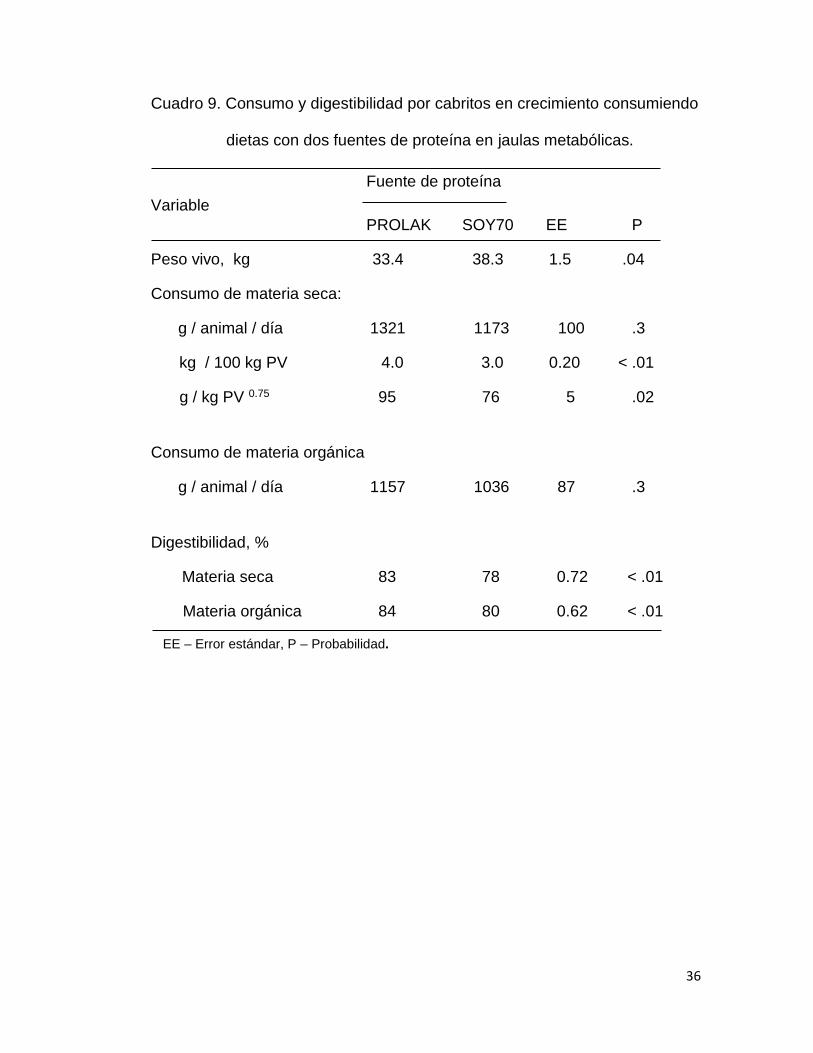
En el cuadro se presenta las medias de consumo y digestibilidad
observados en la fase de evaluación en las jaulas metabólicas. El consumo
de materia seca y materia orgánica en g/día fue similar (P > .05) en los
cabritos consumiendo PROLAK y SOY70; sin embargo al expresar el
consumo en porciento y por peso metabólico el consumo de MS fue más alto
(P < .05) en los cabritos consumiendo la dieta con PROLAK. Así mismo, la
digestibilidad de la materia seca y materia orgánica fue ligeramente mayor (P
< .05) en los cabritos consumiendo PROLAK. El consumo de MS promedio
observado en jaulas metabólicas fue más bajo que en la fase de alimentación
en corral (3.5 vs 3.7 kg/100 kg PV). Estas diferencias también han sido
observadas en estudios previos, en borregos por Meda y Maldonado (2009) y
en cabras lactantes por Domínguez y Pérez (2010).
Los datos de consumo durante la fase de alimentación en corral se
presentan en el cuadro 10. Como se mencionó arriba, el consumo en corral
fue ligeramente más alto que lo registrado en jaulas metabólicas,
posiblemente como resultado de estar en un lugar más confortable para el
animal que estar en la jaula metabólica. No se detectaron diferencias
significativas en el consumo de MS (P > .05) en ninguna de las formas de
expresarlo (g/animal/día, porciento y peso metabólico) entre las dos fuentes
de proteína. Sin embargo, el consumo fue numéricamente mayor en los
cabritos que consumieron la dieta con SOY70, contrario a lo observado en

34
jaulas metabólicas donde el consumo fue más alto en los cabritos
consumiendo la dieta con PROLAK. Los valores de consumo observados en
este estudio (en porciento) son ligeramente menores a los reportados por
Galicia y Nevárez (2015) usando una dieta similar (12% de PC y 2.8 Mcal
EM/kg MS), y son mayores que los reportados por Urge et al. (2004) usando
50 y 75% de concentrado en la ración (ambas 17% de PC; con 2.7 y 2.37
Mcal EM/kg MS, respectivamente). Los cabritos usados en este estudio
tenían alrededor de 10 meses de edad y los usados por Galicia y Nevárez
(2015) tenían alrededor de 5 meses de edad, por lo que los animales usados
en ambos estudios muy probablemente fueran fisiológicamente diferentes.
El consumo de MS se incrementó al transcurrir los días de alimentación, lo
cual es esperable que ocurriera ya que los animales aumentaron de tamaño
(Bines, 1979; Martens, 1994).
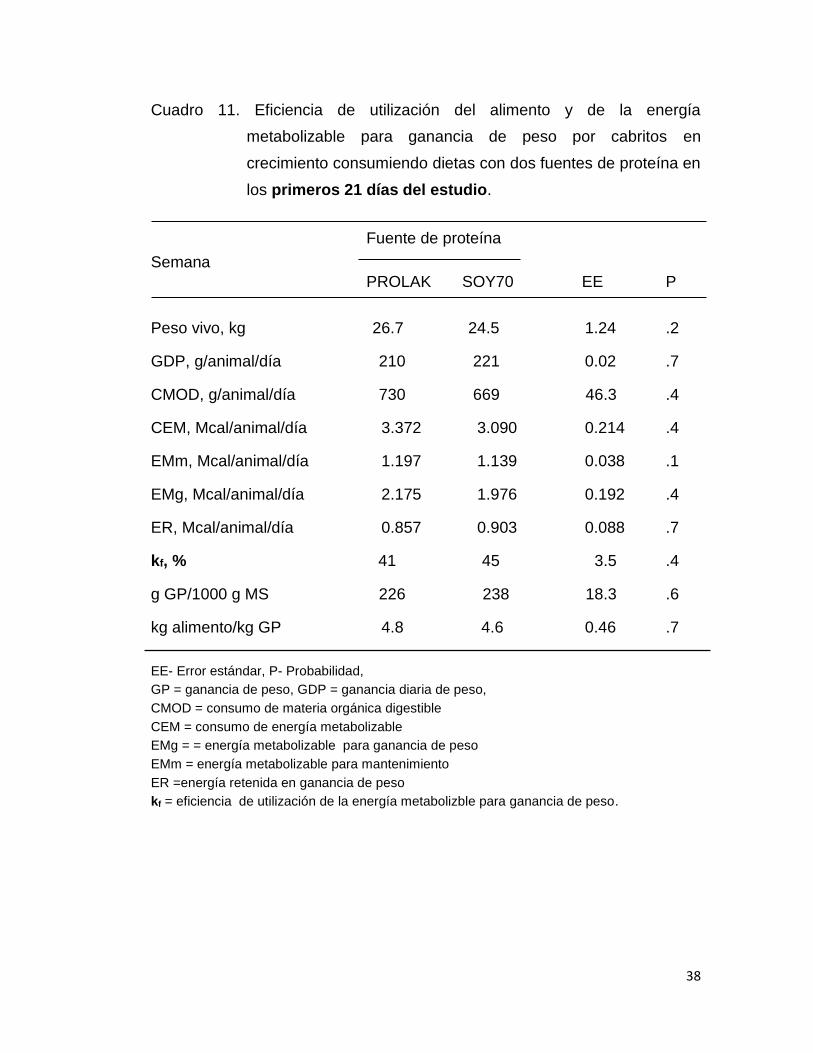
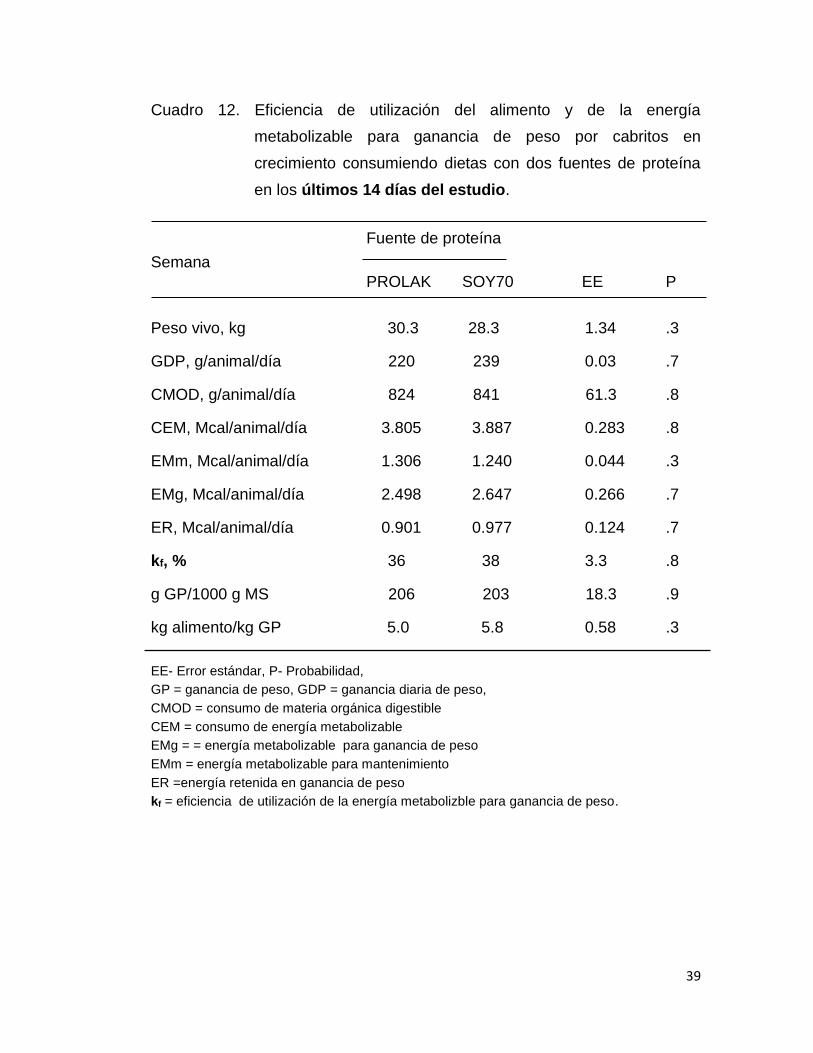
En los cuadros 11, 12 y 13 se presenta los resultados de la eficiencia
de utilización del alimento y de la energía metabolizable. Con el objeto de
evaluar la variación de dichas variables los datos se subdividieron en las
primeras tres semanas (cuadro 11) y las dos últimas semanas (cuadro 12), y
en la Tabla 13 se muestran los resultados analizando los datos de los 35
días de alimentación. En el análisis de los resultados por periodo y todos los
datos no se detectó diferencias significativas en todas las variables
analizadas (PV, GDP, CMOD, CEM, EMg, ER, kf y g GD/kg de alimento), lo
que significa que el valor nutritivo del PROLAK y SOY70 es similar. Los
valores de kf observados en este estudio son similares a los reportados por
Galicia y Nevárez (2015) alimentando cabritos (41%) y corderos (44%) con

35
dietas con contenido de PC y EM igual a los de este estudio. Así mismo, los
valores de kf son similares a los reportados por el NRC en 1984 para ganado
de carne con valores de concentración de EM semejantes a los de este
estudio.
La ganancia de peso por kg de alimento consumido es mayor que lo
reportados por Galicia y Nevárez (2015) y Urge et al. (2004). Los primeros
reportaron ganancias de 153 g/kg de alimento, y los segundos de 80, 121,
104, y 51 g/kg de alimento para cabritos Alpina, Angora, Boer y Españoles,
respectivamente.
36
Cuadro 9. Consumo y digestibilidad por cabritos en crecimiento consumiendo
dietas con dos fuentes de proteína en jaulas metabólicas.
Fuente de proteína
Variable
PROLAK SOY70 EE P
Peso vivo, kg 33.4 38.3 1.5 .04
Consumo de materia seca:
g / animal / día 1321 1173 100 .3
kg / 100 kg PV 4.0 3.0 0.20 < .01
g / kg PV 0.75 95 76 5 .02
Consumo de materia orgánica
g / animal / día 1157 1036 87 .3
Digestibilidad, %
Materia seca 83 78 0.72 < .01
Materia orgánica 84 80 0.62 < .01
EE – Error estándar, P – Probabilidad.

37
Cuadro 10. Consumo de materia seca (CMS) por cabritos en crecimiento
alimentados con dos fuentes de proteína durante 35 días.
Fuente de proteína
Semana
PROLAK SOY70 EE P
CMS, g/animal/día
1 866 800 74 .5
2 966 906 70 .5
3 960 983 65 .8
4 942 1061 77 .3
5 1079 1151 77 .5
Promedio 986 1008 65 .8
CMS, kg/100 kg PV
1 3.6 3.7 0.26 .7
2 3.6 3.8 0.21 .6
3 3.5 4.1 0.29 .2
4 3.4 4.1 0.31 .1
5 3.8 4.4 0.36 .3
Promedio 3.5 3.9 0.22 .2
CMS, g/kg PV 0.75
1 79 80 5.7 .9
2 83 84 4.8 .9
3 81 91 6.1 .2
4 80 94 6.7 .2
5 88 100 7.7 .3
Promedio 80 88 4.7 .3
EE- Error estándar, P- Probabilidad
38
Cuadro 11. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en
crecimiento consumiendo dietas con dos fuentes de proteína en
los primeros 21 días del estudio.
Fuente de proteína
Semana
PROLAK SOY70 EE P
Peso vivo, kg 26.7 24.5 1.24 .2
GDP, g/animal/día 210 221 0.02 .7
CMOD, g/animal/día 730 669 46.3 .4
CEM, Mcal/animal/día 3.372 3.090 0.214 .4
EMm, Mcal/animal/día 1.197 1.139 0.038 .1
EMg, Mcal/animal/día 2.175 1.976 0.192 .4
ER, Mcal/animal/día 0.857 0.903 0.088 .7
kf, % 41 45 3.5 .4
g GP/1000 g MS 226 238 18.3 .6
kg alimento/kg GP 4.8 4.6 0.46 .7
EE- Error estándar, P- Probabilidad,
GP = ganancia de peso, GDP = ganancia diaria de peso,
CMOD = consumo de materia orgánica digestible
CEM = consumo de energía metabolizable
EMg = = energía metabolizable para ganancia de peso
EMm = energía metabolizable para mantenimiento
ER =energía retenida en ganancia de peso
kf = eficiencia de utilización de la energía metabolizble para ganancia de peso.
39
Cuadro 12. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en
crecimiento consumiendo dietas con dos fuentes de proteína
en los últimos 14 días del estudio.
Fuente de proteína
Semana
PROLAK SOY70 EE P
Peso vivo, kg 30.3 28.3 1.34 .3
GDP, g/animal/día 220 239 0.03 .7
CMOD, g/animal/día 824 841 61.3 .8
CEM, Mcal/animal/día 3.805 3.887 0.283 .8
EMm, Mcal/animal/día 1.306 1.240 0.044 .3
EMg, Mcal/animal/día 2.498 2.647 0.266 .7
ER, Mcal/animal/día 0.901 0.977 0.124 .7
kf, % 36 38 3.3 .8
g GP/1000 g MS 206 203 18.3 .9
kg alimento/kg GP 5.0 5.8 0.58 .3
EE- Error estándar, P- Probabilidad,
GP = ganancia de peso, GDP = ganancia diaria de peso,
CMOD = consumo de materia orgánica digestible
CEM = consumo de energía metabolizable
EMg = = energía metabolizable para ganancia de peso
EMm = energía metabolizable para mantenimiento
ER =energía retenida en ganancia de peso
kf = eficiencia de utilización de la energía metabolizble para ganancia de peso.

40
Cuadro 13. Eficiencia de utilización del alimento y de la energía
metabolizable para ganancia de peso por cabritos en
crecimiento consumiendo dietas con dos fuentes de proteína
durante 35 días.
Fuente de proteína
Semana
PROLAK SOY70 EE P
Peso vivo, kg 28.3 26.0 1.22 .2
GDP, g / animal/día 214 228 20 .6
CMOD, g / animal / día 767 738 48 .7
CEM, Mcal / animal / día 3.545 3.409 0.215 .8
EMm, Mcal / animal / día 1.242 1.165 0.040 .2
EMg, Mcal / animal / día 2.304 2.244 0.201 .8
ER, Mcal / animal / día 0.874 0.933 0.080 .6
kf 39 42 2.3 .4
g GP/1000 g MS 217 222 13.4 .8
Kg alimento/kg GP 5.7 4.8 0.29 .9
EE- Error estándar, P- Probabilidad,
GP = ganancia de peso, GDP = ganancia diaria de peso,
CMOD = consumo de materia orgánica digestible
CEM = consumo de energía metabolizable
EMg = = energía metabolizable para ganancia de peso
EMm = energía metabolizable para mantenimiento
ER =energía retenida en ganancia de peso
kf = eficiencia de utilización de la energía metabolizble para ganancia de peso.

41
VI. CONCLUSIONES y RECOMENDACIONES
6.1. Conclusiones:
6.1.1. Los valores de consumo de materia seca fueron más bajos en cabritos
durante la jaula metabólicas que durante la alimentación en corrales con
piso de tierra
6.1.2. La ganancia de peso y la eficiencia de utilización de la energía
metabolizable en cabritos en crecimiento es similar consumiendo dietas
con PROLAK o SOY70.
6.2. Recomendaciones:
6.2.1. En base a los resultados obtenidos en nuestro estudio recomendamos
usar SOY70 como fuente de proteína en la alimentación de cabritos
en crecimiento, ya que es un producto más barato.
6.2.2. Se recomienda realizar evaluaciones en el futuro donde se incluyan
otros niveles de proteína en la ración y conducir las pruebas por
periodos de mínimo tres meses.

42
VII. LITERATURA CITADA
Allen, S.A. and Miller, E.R., 1987. Determination of N requirement for microbial growth from the effect of urea supplementation of a low – N diet on abomasal N – flow and N recycling in wethers and lambs. Brit. J. Nutr. 36: 353 – 368.
Amos H. E., 1980, Treatment of protein to improve utilization by ruminants, Proc. Georgia Nutr. Conf., Pág. 168
AOAC. 1990. Official methods of analysis, 15th ed. Asociation of Analycal Chemists, Washington, D.C.
ARC. 1980, The Nutrient Requeriments of Ruminant Livestok. Commonwealth. Agriculture Bureaux, England.
Arechiga C. F., et al., 2008, Role and perpectives of goat production in a global world, Universidad Autónoma de Zacatecas y Universidad Autónoma Chapingo, recuperado el 18 de Agosto de 2015 de http://www.veterinaria.uady.mx/publicaciones/journal/vol-9-amca/Arechiga1.pdf
Asociación Nacional de Criadores de Ganado Caprino de Registro (ANCGCR) A. C. 2013, recuperado el 12 de Agosto de 2015 de, http://www.conargen.mx/index.php/asociaciones/caprinos
Attfield, H.D., F.W. George, J.Haenlein, E. M. Williams y Moore, 2004, Understanding dairy goat production, volunteers in technical assistance, EUA.
Balch C. C. y Campling R., 1965, Physiology of digestion of the ruminant, R. W. Dougherty (dir), Butterworths, Washington D. C., pág. 108.
Bines, J. A., 1979. Consumo voluntario de alimentos. In: Broster E. H, y Swan, H, 1979. Estrategia de alimentación para vacas lecheras de alta producción; Ed. AGT, S. A; progreso 202, planta alta, 11800 – México, D. F.
Blaxter, K. Lyon. 1964. The Energy Metabolism of Ruminants, Ed. Huitchinson and Co. (Plublishers) Ltd., London, Inglaterra.
Burns, J. C.; Pound, K. R. y Fisher, D. S., 1994, Measurement of Forage Intake. In: Fahey, G. C. Jr., Collins, M., Mertens, D. R., y Moser, L. E. 1994, Forage Quality Evaluation and utilization. American Society of Agronomy. Pp. 497 – 532

43
Carrera, C. 1984. La cabra. Uno de los animales más eficientes Ecológicamente. Productividad Caprina. Fac. Med. Vet. y Zoot. U.N.A.M. México, D.F.
Castro R. A., 2014, La importancia de la energía, la proteína y el agua en la producción caprina, recuperado el 15 de Agosto de 2015 de Ministerio de Agricultura y Ganadería de Costa Rica (MAG) en http://www.mag.go.cr/biblioteca_virtual_animal/cabra_energia.html
Centro de Estudios Agropecuarios. 2001. Crianza de Caprinos. Ed Iberoamérica. México, D.F.
Chalupa, W., 1972. Metabolic aspects of nonprotein nitrogen utilization in ruminant animals, en Fed. Proc., Núm. 31, pág. 1152.
Church D. C. & Pond W.G., 1982. Basic Animal Nutrition And Feeding, 2nd edition, Ed. Jonh Wiley & Sons. Oregon Estados Unidos.
Church, D. C., 1976. Digestive physiology and nutrition of ruminant, Vol. 1, Digestive Physiology, 2nd edition, ed. O&B Books, Corvallis Oregon Estados Unidos.
Cochran, R. C. y Galyean, M.L. 1994, Measurement of in vivo digestion by ruminants. In. Fahey, G. C. Jr., Collins, M., Mertens, D.R., y Moser, L.E. 1994. Forage Quality Evaluation and Utilization. American Society of Agronomy. Pp: 613 – 643.
Comité Nacional Sistema Producto Caprinos, 2014, Entorno Internacional de la Caprinocultura, recuperado el 15 de Agosto de 2015 de http://www.cnsp.caprinos.org.mx/otraspublicaciones/anuario2013.pdf
CONAGUA, 2009, Estadísticas del agua en México. Secretaria de Medio Ambiente y recursos Naturales. Ed. Secretaria de Medio Ambiente y Recursos Naturales, Boulevard Adolfo Ruiz Cortines # 4209, Col. Jardines de la montaña, C.P. 14210, Tlalpan, México, D.F.; pp. 193 – 248
Cornet, A., 1988. "Principales caractéristiques climatiques". En: Estudio integrado de los recursos vegetación, suelo y agua en la Reserva de la Biosfera de Mapimí. C. Montaña, ed., Instituto de Ecología, Publicación 23.
Dabiri, N., y Thonney, M. L. 2004. Source and level of supplemental protein for growing lambs. Journal of Animal Science 2004. 82:3237-3244.
Domínguez, M. P. y Pérez, G. A. 2010. Efecto del nivel de concentrado en la ración y calidad de agua bebida sobre la producción y composición de la leche en cabras. Tesis. Universidad Autónoma Chapingo, Unidad Regional Universitaria de Zonas Áridas. Bermejillo, Dgo. Pág. 14 – 33.

44
Galicia C.M y Nevárez D. A., 2015, Ganancia de peso en corderos y cabritos en crecimiento alimentados con dos niveles de proteína cruda en la ración, Universidad Autónoma Chapingo. Bermejillo, Dgo., México.
Ganev G. E. et al., 1979, the effect of roughage on concentrate feeding and rumen retention time on total degradation of protein in the rumen, J. Agric. Sci. Camb,. núm. 93, pág. 651
García E. 1973. Modificaciones al sistema de clasificación climática de Koopen (para adaptarlo a las condiciones climáticas de la república mexicana); Universidad Autónoma Chapingo; pp. 12 – 13.
García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen. Offset Larios, México.
Grau M. C., Marval H., Marcano A., Z., 2007, Utilización de la harina de pescado en la formulación de alimentos para crecimiento y de engorde animal, Centro de Investigaciones de los Estados Sucre y Nueva Esparta, Argentina.
Haddad, S.G., Nasr, R.E. y Muwalla, M.M. 2001. Optimum dietary crude protein level for finishing Awassi lambs; Small Ruminant Research Volume 39, Issue 1, January 2001, Pages 41-46.
Haresing W. & Cole D.J.A,. 1988. Recend Developments in Ruminant Nutrition 2. Butterworths, Londres.
Hemsley, J. A., 1975, Effect of high intake of sodium chloride on the utilization of a protein concentrate by sheep, en Aust. J. Agric. Res., núm. 26, pág. 709
Hill, R.,1991, Rapeseed meal in the diets of ruminants, en Nutr. Abstr. Rev. (series B), núm. 61, pág. 139
Kennedy P. M. et al., 1976, Effects of cold exposure of sheep on digestion, rumen turnover time and efficiency of microbialsynthesis, en Br. J. Nutr., núm. 47, pág. 231.
Kennedy P. M. et al., 1982, Effects of cold exposure on feed protein degradation, microbial protein synthesis and transfer of plasma urea to the rumen of sheep, en Br. J. Nutr., núm. 47, pág. 521
Machado de Rocha, M. M., Susin, I., Pires, A., Vaz, De Souza, F. J., y Quirino, M. C. 2004. Performance of Santa Ines lambs fed diets of variable Crude Protein levels. Sci. Agric. (Piracicaba, Braz.), v.61, n.2, p.141-145, Mar./Apr. 2004.
McAllan A. B. y Smith R. H. 1983, Estimation of flows of organic matter an nitrogen components in post ruminal digesta and affects of level of

45
dietary intake and physical form of protein supplement on such estimates, en Br. J. Nutr., núm. 49, pág. 119
McDonal, P. et al., 1998, Animal Nutrition, 5 Edition, Logman Scientific Technical, Nueva York.
McDonald P.; Edwars R.A; Greenhalgh J.F.D; Morgan C.A. 2002. Animal Nutrition. 6.a Edition, ed, Pearson. Londres.
Meda A.P y Maldonado J. J., 2009, Ganancia de peso en corderos alimentados con tres niveles de proteína cruda en la ración, Universidad Autónoma Chapingo. Bermejillo, Dgo., México.
Mues Noel y Walz Troy, 2005, Sistema de Proteína Metabólica (MP). Revisión bibliográfica, Instituto de Agricultura y Recursos Naturales de la Universidad de Nebraska. Recuperado el 25 de Agosto de 2015 de http://www.produccion-animal.com.ar/tablas_composicion_alimentos/22-Cap_3_Proteina.pdf
NRC, 1981, Nutrient Requirements of Domestic Animals. No. 15. Nutrient Requirements of Goats: Angora, Dairy, and Meat Goats in temperature and tropical countries. National Research Academic Press. Washington, D.C.
NRC, 1982, United States – Canadian Tables of Feed Composition, National Research Council, National Academy Press, Washington, D.C.
NRC, 1984, Nutrient Requeriments of Animals, Nutrient Requirements of Sheep, 5a. ed., National Academy of Sciences-National Research Council, Washington, D.C.
NRC, 1985, Nutrient Requeriments of Animals, Nutrient Requirements of Sheep, 5a. ed., National Academy of Sciences-National Research Council, Washington, D.C.
NRC, 2009, United States – Canadian Tables of Feed Composition, National Research Council, National Academy Press, Washington, D.C
Nutrilag, 2013, SOY70 ® recuperado el 28 de agosto de 2015 de http://www.nutrilag.com/productos/soy-70
Orskov, E. R. et al., 1972, Digestion and Metabolism in the Ruminant, en Br. J. Ntr., Núm. 28. Pág. 425.
Orskov, E.R., 1982, Protein Nutrition in Ruminants, Academic Press, Londres.
Owens F. N. y Berger W.G., 1983, Nitrogen metabolism of ruminant animals: Historical perspective, current understanding and future implications, en J. Amin. Sci., núm. 57, págs.m 498 – 518.

46
Ponce, C.I. 2004. Sustitución de diferente nivel de cascarilla de soya en sustitución parcial de sorgo y rastrojo de maíz. Tesis profesional. Departamento de zootecnia. Universidad Autónoma Chapingo. Estado de México.
Prigge E. C. et al., 1978, Microbial protein synthesis in steers fed processed corn rations, en J. Anim. Sci., núm. 46, pág. 249
SIAP, 2013, Inventario Caprino, población ganadera 2004 – 2013, recuperado el 12 diciembre de 2014 de http://www.siap.gob.mx/opt/poblagand/Caprinos.pdf
SIAP, 2014, Resumen nacional pecuario, recuperado el 10 de agosto de 2015 de http://www.siap.gob.mx/resumen-nacional-pecuario/http://www.fao.org/
SIAP, 2015, Resumen nacional pecuario preliminar, recuperado el 10 de agosto de 2015 de http://www.siap.gob.mx/resumen-nacional-pecuario/http://www.fao.org/
Steel, D.R.G y Torie, H.J. 1980. Pronciples ad Prodedures of Statistics. 2nd Edition. Ed. McGraw Hill. NY.
Tamminga S., 1979, Protein degradation in the forestomachs of ruminants, en J. Anim., Sci, Núm. 49, pág. 1615
Titgemeyer, E. C. et al., 1982, Evaluation of soybean meal, corn gluten meal, blood meal and fish meal as source of nitrogen and amino acids disappearing from the small intestine of steers, en J. Anim. Sci., núm. 55, pág. 1498
Urge M., Merkel R.C., Sahlu T., Animut G. y Goetsch A. L. 2004. Growth performance by Alpine, Angora, Boer, and Spanish wether goats consuming 50 or 75% concentrate diets. Small Ruminant Reserch, V. 55, Pp. 149 – 158. Langton, OK.
Van Soest, P. J., 1994. Nutritional Ecology Of the Ruminant, 2nd edition, Ed. O & B Books, Corvallis, Oregon.
Varga G. A. y Prigge E.C., 1982, Influence of forage species an level intake on ruminal turnover rates, en J. Anim. Sci., Núm. 55, pág. 1498
Zinn, R. A. y Owens F. N., 1983, Influence of feed intake level on site of digestion in steers fed a high concentrate diet, en J. Anim., Sci., Núm. 56, Pág. 471