Embed Size (px)
Citation preview

B o l e t í n
C i e n t í f i c o
Laboratorio de Microbiología
UNIVERSIDAD RICARDO PALMA
Diciembre, 2014
Volumen 1, Nº1
http://www.urp.edu.pe/biologia/index.php?urp=Laboratorios
VOLUMEN 1, Número 2

Boletín Científico
Página 1 Laborator io de Microbio logía Volumen 1, n°2
Comité Editor
Tania Churasacari Vinces
Cindy Cajachagua Pucuhuaranga
Diego Santiago Bravo
Carla Saldaña Serrano
Luis Pabón Rodríguez
Miguel Gonzáles Olivos
Mario Aguayo Cerna
Pavel Pastrana Sumari
Miguel Morales Campos
Claudia Monge Pimentel
Miguel Montero Orozco
Roma Ballena Palomino
Decano
Dr. Tomás Agurto Saenz
Director
Blgo. Alcides Guerra Santa Cruz
Editor General
Santiago Justo Arévalo
Editor de Estilo
Paola Nunja Huamán
Editor Científico
Marlon Morales Moisela
1° ed., 1° impresión, diciembre 2014
Hecho el Deposito Legal en la Biblioteca Nacional del Perú N° 2014-19353
Editado por:
Santiago Justo Arévalo
Av. Malecon Asent. H. Nuevo Luriin Mz. A5 Lt. 01, Lurin—Lima
Impreso en:
Impreso en la Universidad Ricardo Palma,
Av. Benavides 5440, Santiago de Surco
Diciembre, 2013

Contenido
Página 2 Laborator io de Microbio logía Volumen 1, n°2
Editorial
Presentación…………………………………………………………………….03 Editorial……...…………………………………………….……………………04
Actividades del Laboratorio
Difusión y Exposición de Trabajos de Investigación URP presente en el IV CONCIMAR ...…………………………….….…..05 Encuentro Científico de invierno internacional 2014 (ECI) ...……………..05 Curso-Taller de Redacción Científica ……......…….….……………………….....06 Ciclo de conferencias: Inmunidad y Cáncer.………………..…………….……....07 Proyección Social con alumnos del Colegio EUROAMERICANO….…..….....…08 Reunión de Integración con los Padres de Familia...…...…………...………….....09 Primera capacitación de Interciclo-2014 ...………………………………….…....10 Integrantes Demuestran lo Aprendido en Importantes Laboratorios.…………....11 Visita de estudios a Petramás: Empresa 100% Peruana….….…………….…..….12
Artículos de Divulgación Virus contra Bacterias………………... ...………………………………….…....13 Proteínas Recombinantes…………….. ...………………………………….…....14
Artículos de Investigación Actividad antimicrobiana del extracto acetona-agua de Macrocystis pyrifera (C. Agardh 1820) en bacterias de importancia clínica ………………………...…….15 Determinación de Géneros Bacterianos en el Mar de la Playa Cantolao - La Punta - Callao …………………………………………………………………………...20 Efecto antibiótico de Piocianina de Pseudomonas aureginosa sobre Escherichia coli y Staphylococcus aureus …………….……..………...…………………………….......24
MiniReview Aspectos generales de la crianza de Cuy y su agente infeccioso: Salmonella……....28 Fundamentos básicos de PCR convencional y sus variantes..……………….…....34 Integración de la Biotecnología Bacteriana y la Ingeniería Genética de Plantas......43

Presentación Les presentamos el segundo número del Boletín Científico del Laboratorio de
Microbiología de la Facultad de Ciencias Biológicas de la Universidad Ricardo
Palma. En esta nueva edición tenemos el placer de informarles acerca de las
actividades realizadas en el presente año, donde se incluyen cursos,
capacitaciones, artículos de divulgación, trabajos de investigación y artículos de
revisión; todo esto realizado por integrantes del laboratorio con el único deseo
de hacer la mejor ciencia posible. Actualmente, la intedisciplinariedad de la
ciencia hace necesario que trabajemos en conjunto con otras áreas, es por esto
que el divulgar e informar sobre las actividades que se realicen en un
determinado lugar no solo sirve para dar a conocer las labores de un equipo,
sino pretende además ampliar horizontes a través de la formación de nuevos
nexos con otras instituciones y equipos para así mejorar e integrar los
conocimientos adquiridos; así, si cada uno aporta de un punto de vista diferente,
los logros se hacen más completos y útiles. Les reiteramos nuestros saludos y
les deseamos mucho éxito en todas las áreas que se desempeñen y trabajos que
realicen, recuerden que todo esfuerzo tiene su recompensa.
Editorial
Página 3 Laborator io de Microbio logía Volumen 1, n°2
Equipo de Investigación del Laboratorio de Microbiología

El Microbiólogo
Dr. Tomás Agurto Sáenz
Página 4 Laborator io de Microbio logía Volumen 1, n°2
Editorial
En un libro titulado “los cazadores de microbios” se
relata de los trabajos de científicos que querrán
saber y demostrar el porqué de las enfermedades del
humano, en animales y plantas. Robert Koch,
postulaba que una infección es producida por un
agente a quien hay que encontrarlo y este agente
debe reproducir el mismo mal al ser inoculado en un
animal, eso es lo que se hace ahora, se mira cara a
cara a las colonias crecidas en una placa Petri y bajo
el microscopio se puede definir su forma,
coloración; así como mediante pruebas bioquímicas
y moleculares se puede determinar su composición
química y su metabolismo, logrando una mejor
identificación de bacterias, hongos, y virus que
pueblan el mundo, en el suelo, agua, aire. Existen
especies no patógenas, en las que se explora sus
actividades metabólicas para transformar y elaborar
una serie de productos que se industrializan, los
antibióticos que ocupan un alto lugar en producción
industrial; los alimentos, cada vez más
transformados, conservados, para que sean idóneos
e inocuos. En los ciclos biológicos intervienen las
bacterias y en la cadena alimenticia son las
iniciadoras de los procesos metabólicos, así, se
puede decir que los microrganismos son los
verdaderos responsables para que la vida exista y
este en desarrollo continuo y aquí en el laboratorio
de microbiología un grupo entusiasta, activo,
guardan por ser maestros en dar acción a los
microorganismo con sendos trabajos de
investigación ya al paso de crear el curso y la
especialización en genética bacteriana.
Jóvenes sigan adelante, el éxito depende de ustedes.
Dr. Tomás Agurto Sáenz Decano de la Facultad de Ciencias Biológicas Profesor Principal en la Cátedra de Microbiología

Difusión y Exposición de Trabajos de Investigación
En este invierno 2014 la Facultad de Ciencias Biológicas
tuvo el honor de ser la sede de la sesión de Biología del ECI
2014. Esta se llevó a cabo en el auditorio Biotempo con la
dirección del Prof. Alcides Guerra. Nuestro laboratorio
manifestó su presencia a través de la presentación de 3
trabajos de investigación: “Determinación de Géneros
Bacterianos en el Mar de la Playa Cantolao – La Punta –
Callao”, “Extracción y Actividad Antimicrobiana del
Pigmento Piocianina de 8 cepas de Pseudomonas
aeruginosa” y “Actividad Antibacteriana del Extracto
Acetona-Agua de Macrocystis pyrifera (C. Agardh 1820) en
Bacterias de Importancia Clínica”
La jornada se realizó con éxito con la presencia de
estudiantes de varias universidades del país, además en toda
nuestra universidad se expusieron trabajos de investigación
relacionados a otras áreas de la ciencia.
En el año 2015 se realizará el ECI de Verano en las fechas 2
al 6 de Enero donde seguiremos participando con
investigaciones que venimos realizando gracias al esfuerzo de
todos los integrantes abocados a la ciencia en la Facultad de
Ciencias Biológicas.
Con la finalidad de difundir y discutir los resultados de las
últimas investigaciones relacionadas a los diversos aspectos de
las ciencias del Mar realizadas por investigadores y estudiantes
universitarios en nuestro país, se llevó cabo el IV Congreso de
Ciencias del Mar (IV CONCIMAR). Este importante evento,
realizado en el mes de Junio en la Universidad Peruana
Cayetano Heredia, estuvo dirigido a la comunidad académica,
científica y empresarial nacional e internacional, así como al
sector pesquero y a sus representantes.
El equipo de investigación del Laboratorio de Microbiología se
hizo presente con el tema Determinación de Géneros
Bacterianos en el Mar de la Playa Cantolao – La Punta –
Callao, el cual tuvo el principal objetivo de expandir y motivar
los trabajos en el área de microbiología marina para sus
aplicaciones ecológicas y biotecnológicas.
ENCUENTRO CIENTIFICO DE INVIERNO
INTERNACIONAL 2014 (ECI)
URP presente en el IV CONCIMAR
Santiago Justo Arévalo y Luis David Pabón presentando el panel
“Determinación de géneros bacterianos en el Mar de la Playa Cantolao”
Marlon Morales Moisela en su exposición del
tema: “Actividad antibacteriana del Extracto
Acetona-Agua de Macrocystis pyrifera (C. Agardh
1820) en Bacterias de Importancia Clínica”
Página 5 Laborator io de Microbio logía Volumen 1, n°2
Actividades

Curso-Taller de Redacción Científica
Cierre del curso Taller de Redacción
Página 6 Laborator io de Microbio logía Volumen 1, n°2
El 30 de mayo, 6 y 13 de
Junio del presente año se
organizó el Curso-Taller de
Redacción Científica, el cual
contó con la participación de
ponentes de la Dirección
Científica Académica de
ONCOSALUD – AUNA:
Joseph Pinto, Claudio Flores
y Priscila Valdivieso,
quienes expusieron y dieron
importantes pautas sobre
cómo redactar de manera
ordenada y organizada un
artículo de investigación
científica, cómo presentarlo
a una revista para que evalúe
su publicación, además de
dar a conocer aspectos
generales para tomar en
cuenta en la edición de una
revista científica, el proceso
editorial para la evaluación
de trabajos y cómo mejorar
la calidad y contenidos de esta.
Este evento se realizó con el
fin de promover la publicación
de artículos científicos en
estudiantes y profesionales de
nuestra facultad,
contribuyendo en gran medida
al desarrollo científico.
Cabe agregar que en el evento
no sólo participaron
estudiantes y profesionales de
nuestra casa de estudios, sino
también de otras universidades
como la Universidad Nacional
de Ingeniería, Universidad
Nacional Agraria, Universidad
Nacional Federico Villareal y
la Universidad Peruana
Cayetano Heredia.
Actividades
Afiche de Presentación del Curso-taller

Ciclo de Conferencias: Inmunidad y Cáncer
En los últimos años las
ciencias biológicas se han
desarrollado rápidamente, lo
que ha permitido el estudio
de mecanismos moleculares
de defensa de nuestro
sistema inmune frente a
señales de advertencia de un
mal funcionamiento celular.
El ciclo de conferencias
“Inmunidad y Cáncer” tuvo
como objetivos actualizar a
los asistentes en las últimas
investigaciones sobre la
inmunidad del cáncer,
promover nuevas
investigaciones, además de
crear un foro de debate en
aspectos relacionados a los
avances en el tratamiento del
cáncer.
Dicho evento fue realizado
con éxito gracias a la acción
conjunta del Laboratorio de
Microbiología – URP y la
Dirección Científico
Académica de ONCOSALUD
– AUNA, el día 21 de Junio
del 2014 en el Auditorio Ccori
Wasi – URP, el ciclo de
conferencias conto con la
participación de destacados
ponentes en el área tales como
el Dr. Alfredo Aguilar
Cartagena y representando a
nuestra casa de estudios el
Blgo. Alcides Guerra Santa
Cruz.
Página 7 Laborator io de Microbio logía Volumen 1, n°2
Cierre del Ciclo de Conferencias: Inmunidad y Cáncer
Actividades
Afiche de Presentación del Ciclo de Conferencias

Proyección Social con los alumnos de Colegio EUROAMERICANO
Alumnos del Colegio
Euroamericano en la
capacitación de técnicas
experimentales básicas
de microbiología
Página 8 Laborator io de Microbio logía Volumen 1, n°2
Actividades
El Colegio Euroamericano es una escuela situada
en el valle de Pachacámac, esta resaltante
institución maneja un programa poco común en
los centros educativos del país, el IB o
Bachillerato Internacional, lo que permite que sus
alumnos egresen con un Diploma adicional que
les permite postular universidades de alto
prestigio internacional una vez terminados sus
estudios secundarios. Para contribuir a la
formación de los alumnos de esta institución, la
Universidad Ricardo Palma a través del
Laboratorio de Microbiología de la Facultad de
Ciencias Biológicas y como parte de su programa
de Proyección Social, ha capacitado a alumnos
del último año del Bachillerato de Biología en
técnicas experimentales básicas de la
microbiología como Coloración Gram, ELISA y
electroforesis.
Además ha permitido que el alumno Rodrigo
Alcorta del Bachillerato de Biología investigue el
efecto antimicrobiano de plantas del Perú,
realizando su trabajo de graduación del IB
asesorado por el Prof. Jefe de Laboratorio, Alcides
Guerra y los ayudantes de laboratorio.
Es así como se integran partes fundamentales de
la educación y el progreso del país: Colegios y
Universidades.
Alumna Tania Churasacari capacitando alumnos del
EUROAMERICANO

Reunión de Integración con los Padres de Familia
Palabras del Decano a los padres
Visita de los padres a las
diferentes ambientes del
Laboratorio de
Microbiología
Página 9 Laborator io de Microbio logía Volumen 1, n°2
Actividades
El Laboratorio de Microbiología, celebró en el
interciclo 2014 una reunión en la que el Dr. Tomás
Agurto Sáenz, Decano de la Facultad de Ciencias
Biológicas; el Blgo. Alcides Guerra, Jefe del
laboratorio e integrantes del mismo se reunieron con
los padres de familia, en una muy amena asamblea
donde los padres eran los invitados.
La bienvenida fue en el Auditorio Biotempo y la dio
el Dr. Agurto, con su siempre jovial prosa,
adentrándolos en el panorama de nuestra carrera y la
microbiología en la actualidad; luego tomó la
palabra el Blgo. Alcides Guerra, Jefe de nuestro
laboratorio, enterándolos de todas nuestras
actividades realizadas hasta el momento así como
también de los proyectos que tenemos en curso; para
finalizar esta primera etapa de la reunión, tomó la
palabra el ayudante principal, Santiago Justo, quien
a nombre de los demás integrantes, emitió una muy
emotiva y sincera perorata, donde agradeció la
asistencia de todos los presentes. Seguidamente los
padres fueron guiados al laboratorio y vieron
directamente el lugar de trabajo de sus hijos,
entonces pudieron intercambiar impresiones.
No nos queda más que agradecer a nuestros padres,
por haberse dado un momento para compartir con
nosotros y siempre estar ahí, apoyándonos en cada
etapa de nuestras vidas, por ser nuestros guías,
consejeros y amigos.

Primera Capacitación De Interciclo-2014
Página 10 Laborator io de Microbio logía Volumen 1, n°2
“Dime y lo olvido, enséñame y lo recuerdo,
involúcrame y lo aprendo” Benjamin Franklin
“El aprendizaje es experiencia, todo lo demás
es información” Albert Einstein
Actividades
El Laboratorio de Microbiología de la Universidad
Ricardo Palma conoce el entusiasmo y ganas de
aprender que tenemos los alumnos de la Facultad de
Ciencias Biológícas, además del espíritu científico
que nos caracteriza, ante esta situación se realizó
una capacitación de un poco más de un mes dirigida
a todas las personas que deseen compartir
conocimientos con los ayudantes del laboratorio
Microbiología, esta incluía clases teóricas, prácticas
y discusión de papers en temas desde los básicos
hasta los más actuales: medios de cultivo, pruebas
bioquímicas, antibiograma, extracción de DNA
bacteriano, identificación molecular de una bacteria,
transformación bacteriana, purificación de proteínas
y electroforesis. La finalidad era que los asistentes
se sientan involucrados e identificados como
partícipes directos de cada tema desarrollado con la
intención de motivarlos en el campo de la
investigación, que conozcan que en cada laboratorio
de la facultad hay personas que les gusta la ciencia
tanto como a ellos. Pavel Pastrana, uno de los
participantes de la capacitación nos comenta que:
“Participar en el interciclo, del laboratorio de
microbiología, ha sido una muy buena experiencia
tanto para mi vida profesional como personal; ya
que aparte de haber adquirido nuevos conocimientos
en el área de la microbiología también me ayudó a
ser más meticuloso con las técnicas empleadas en un
laboratorio y ser más organizado en el día a día. Por
otro lado me siento muy contento de haber
participado, pues lo aprendido y el material
complementario, me ha ayudado bastante en los
cursos que estoy actualmente llevando. Me siento
muy agradecido y cómodo en este ambiente, y
agradezco a los muchachos del laboratorio por
permitirme llevar este curso con ellos y brindarme
su conocimiento y su confianza.”

Página 11 Laborator io de Microbio logía Volumen 1, n°2
Actividades
Integrantes Demuestran lo Aprendido en Importantes Laboratorios
El laboratorio de Microbiología viene esforzándose
por ampliar las barreras del conocimiento. Es así que
dos más de nuestros integrantes han obtenido la
oportunidad de realizar prácticas en laboratorios de
prestigio a nivel nacional e internacional.
Uno de ellos es Marlon Morales, quien terminó
exitosamente su ciclo de prácticas en el Laboratorio
Mixto Internacional (LMI) del Institut de Recherche
pour le développement (IRD), un instituto de
investigación francés que se encuentra asociado a la
Universidad Peruana Cayetano Heredia que viene
dedicándose a la investigación del aislamiento y
fraccionamiento de productos naturales de planta
nativas del Perú con el fin de probar la capacidad de
estas contra bacterias, enfermedades como
Leishmania y Malaria, y hasta contra el mismo
cáncer. Marlon expresó que su formación en el
laboratorio de Microbiología le sirvió de base, pues
le permitió aportar con ideas claras sobre técnicas
empleadas en pruebas de sensibilidad bacteriana y el
mecanismo de acción de los antibióticos conocidos,
además de formarse con un concepto de
interdisciplinariedad en la ciencia, ya que el LMI
trabaja en conjunto con químicos y farmacéuticos en
busca de nuevas opciones para generar antibióticos.
Por su parte, Santiago Justo, integrante veterano del
Laboratorio de Microbiología, ha conseguido la
oportunidad de practicar en el Instituto de Química
de la Universidad de Sao Paulo, en el área de
Bioquímica. Durante seis meses, Santiago podrá
acompañar al equipo de investigación que se
encuentra liderado por el Dr. Shaker Chuck Farah,
además tendrá la oportunidad de afianzar sus
conocimientos en técnicas moleculares como
clonación, secuenciamiento de ADN, expresión y
purificación de proteínas recombinantes; y técnicas
espectroscópicas como cristalografías, dicroísmo
circular y microscopias. Santiago opina que estar
encargado del laboratorio de Microbiología, le ha
dado la capacidad de desenvolverse de manera
activa y constante en el área y en las investigaciones
científicas
De esta manera, el Laboratorio de Microbiología
abre la puerta a sus integrantes en diferentes
instituciones de prestigio a nivel nacional e
internacional.
Marlon Morales
trabajando con
extractos de
Pseudomonas
aeruginosa
Santiago Justo
exponiendo en
Auditorio
Biotempo

Página 12 Laborator io de Microbio logía Volumen 1, n°2
Actividades
Visita de Estudios a Petramás:
Empresa 100% Peruana
Alumnos en Planta procesadora de Petramás
El día 14 de Julio del 2014, los integrantes del
Laboratorio de Microbiología de la Facultad de
Ciencias Biológicas realizamos una visita a la
primera empresa 100% peruana, productora de
energía eléctrica a partir de residuos. La visita fue al
Relleno Sanitario Huaycoloro, siendo esta una de
las tres plantas procesadoras de residuos sólidos de
la empresa Petramás ubicada en el distrito de San
Antonio, provincia de Huarochirí. Durante el
recorrido se observó grandes extensiones de terreno,
las cuales están destinadas a diferentes procesos en
el tratamiento de residuos, ya sea residuos sólidos
orgánicos hasta residuos de alta peligrosidad. La
planta trabaja con el modelo de Desarrollo Limpio
de Residuos que consiste en la captura y combustión
del biogás generado en el relleno, la captura del gas
se dan en grandes pozos que están conectados a un
gaseoducto que llevan el biogás a la Central
Térmica, la cual es una moderna estación
automatizada de succión y quemado del gas
conectada a una moderna central generadora de
electricidad que finalmente se interconecta a redes
que abastecerán de energía eléctrica a diferentes
lugares de Lima.
La gran importancia que tiene esta planta
generadora de energía eléctrica a partir de desechos,
es que reduce la cantidad de emisiones de gases
contaminantes al medio. Petramás gestiona uno de
los mecanismos más limpios en el tratamiento de
residuos sólidos contribuyendo a mitigar los efectos
que el calentamiento global causa al Perú y a todo el
mundo.

Virus contra Bacterias Por Claudia Monge.
Página 13 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Divulgación
En 1917 el microbiólogo Félix d’Herelle descubrió
que existían virus que eran capaces de parasitar
bacterias a los que llamo “bacteriófagos” o fagos. Los
siguientes años realizó los primeros estudios de
empleo de fagos para tratar la disentería. No obstante
recién en 1921 se estandarizaron tratamientos por
Richard Bruyboghe, al usar fagos para tratar
infecciones de estafilococos en la piel. Durante esa
época los bacteriófagos o también llamados fagos
fueron comercializados para combatir diversas
infecciones. Especialmente durante la primera guerra
mundial, donde la fagoterapia fue utilizada para
combatir la disentería.
Con el paso de los años debido a la aparición de los
antibióticos las investigaciones en tratamientos de
fagoterapia fueron dejadas de lado. Sin embargo, la
aparición de cepas bacterianas resistentes a un elevado
número de antibióticos obligaron a los científicos a
encontrar nuevas formas de combatir las bacterias.
La especificidad del ataque de los bacteriófagos
constituye por un lado una ventaja para la fagoterapia
frente a los antibióticos, ya que los fagos atacan
directamente a una determinada cepa de bacterias,
mientras que los antibióticos arrasan con todas las
bacterias: las patógenas que causan alguna
infección pero también con las beneficiosas como
las de la flora bacteriana natural del cuerpo.
Además dicha especificidad de los fagos es a su
vez una desventaja frente a los antibióticos puesto
que para combatir una infección de bacterias se
necesita conocer la cepa específica mientras que
con los antibióticos no es necesario este
conocimiento por su amplio rango de ataque contra
bacterias.
Otra ventaja frente a los antibióticos es la
capacidad de los bacteriófagos de tener mayor
alcance en los tejidos (puede incluso atravesar la
barrera hematoencefálica, a la cual los antibióticos
no tienen alcance)
Por último una gran desventaja de la fagoterapia es
que algunos fagos pueden ser portadores de genes
de resistencia a antibióticos o de genes que
codifican toxinas y por lo tanto, pueden introducir
estos genes a las bacterias otorgándoles
características no deseadas.
El laboratorio de microbiología está en proceso de
estandarización de metodologías para el
aislamiento de bacteriófagos con miras a
posteriores investigaciones en fagoterapia.
Figura 1. Representación gráfica de
posicionamiento de un fago sobre la
superficie bacteriana
Fuente:
http://bacteriasactuaciencia.blogspot.com/2014/02/e
l-redescubrimiento-de-la-terapia-por.html

Página 14 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Divulgación
Proteínas Recombinantes Por Miguel Morales.
Durante el siglo xx se lograron importantes avances,
uno de ellos fue determinar la estructura de doble
hélice del ADN. Esto les valió a Watson J y Crick F
el premio nobel en 1962, luego permitió conocer
como el ADN se transmite de una generación a otra.
Además se reconoció que un organismo desde el
más simple al más complejo tenía el mismo código
génetico. Esto quiere decir que el ADN puede ser
interpretado y traducido por cualquier otro
organismo del planeta. También se descubrió que
esta información se traduce a proteínas, moléculas
imprescindibles para todo organismo. Como dijo el
químico holandés Mulder: “sin proteínas no hay
vida posible en nuestro planeta”
En el año 1975 Nathans D y Smith H descubrieron
un tipo de enzimas (proteínas que aceleran
reacciones químicas) de virus y bacterias, las cuales
actualmente son de gran utilidad para el desarrollo
de las proteínas recombinantes, dando origen a la
ingeniería genética.
Una proteína recombinante se genera al colocar un
fragmento de ADN en un organismo distinto al
original, ese fragmento se denomina ADN
recombinante y las proteínas producidas a partir de
ese ADN: proteínas recombinantes.
De manera básica la metodología de producción de
proteínas recombinantes es la siguiente: primero se
aísla el ARNm que codifica la proteína de interés,
por acción de las transcriptasas inversas se sintetiza
ADN a partir de ARN (normalmente la transferencia
de información, es de ADN a ARN y luego, a una
proteína), el ADN obtenido se inserta en un vector
(transporte del gen de interés) “cortado” por unas
enzimas de restricción denominadas endonucleasas
(“tijeras moleculares” que cortan los enlaces fosfato
de la cadena de ADN sin dañar la secuencia de
nucleótidos); así quedaran bases sin aparear que
estarán disponibles para aparearse con el fragmento
de ADN que contenga las bases complementarias
(en este caso el gen de interés) y por último se
ligaran con ayuda de la enzima, ADN ligasa, en el
vector, obteniendo ADN recombinante.
Para la producción de proteínas recombinantes se
siembra bacterias conteniendo el ADN
recombinante, luego se separa la proteína
recombinante de las otras proteínas bacterianas en
un proceso de purificación (normalmente
cromatografías), en algunos casos es necesario tratar
a la proteína para conseguir una forma estable que
pueda cumplir su función de manera óptima.
En el laboratorio de microbiología se viene
trabajando en el diseño y construcción de plásmidos,
con el objetivo de producir proteínas de interés
biotecnológico.
Figura 1. Metodología general de generación de proteínas
recombinantes.
Fuente:
http://www.dfarmacia.com/farma/ctl_servlet?_f=37&id=13128906

Actividad antimicrobiana del extracto acetona-agua de Macrocystis pyrifera (C. Agardh 1820) en bacterias de importancia clínica Morales M*, Nunja P, Burga L, Justo S, Ávila J, Guerra A. Laboratorio de Microbiología, Facultad de Ciencias Biológicas de la Universidad Ricardo Palma. Laboratorio de Biología Marina y Continental, Facultad de Ciencias Biológicas de la Universidad Ricardo Palma. *Autor para la correspondencia: Morales Moisela, Marlon. [email protected]
Palabras claves: Macrocystis
pyrifera, Discos de Difusión.
Extracto acetona-agua.
R E S U M E N
Se evaluó la actividad antibacteriana del extracto acetona-agua de
Macrocystis pyrifera. Se seleccionaron cuatro cepas de importancia clínica:
Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus y
Salmonella enterica del Laboratorio de Microbiología. Los extractos fueron
obtenidos de los filoides, esporófilos y del alga entera de una muestra seca
de la especie; y, a su vez, de una muestra fresca. La prueba de sensibilidad
se realizó mediante discos de difusión cargados con 30 µl de cada extracto;
se utilizaron discos con 10 mg de Gentamicina como control. Ningún
extracto presentó actividad antibacteriana en ninguna de las cepas
evaluadas; por otro lado, todas las cepas se mostraron sensibles ante la
acción de la Gentamicina. Se concluye que Macrocystis pyrifera no posee
actividad antimicrobiana ante las cepas de importancia clínica evaluadas.
1. Introducción
Las bacterias se han adaptado a un sinfín de hábitats
con diferentes mecanismos de sobrevivencia.
Actualmente la resistencia bacteriana a antibióticos
es un problema de gran importancia en el área
clínica. A raíz de este fenómeno es que cada cierto
tiempo se necesita sintetizar nuevos antibióticos para
combatir a estos microorganismos.
Las algas marinas son consideradas una parte
importante en los ambientes acuáticos, ya que
cumplen un nicho ecológico fundamental
equilibrando los niveles de oxígeno en el agua, por
medio de fotosíntesis; participando en las redes
tróficas; y produciendo metabolitos secundarios que
poseen diferentes propiedades, entre ellas, la
actividad antimicrobiana. Macrocystis pyrifera es
una macroalga de importancia económica a nivel
mundial. Pertenece a la división de las Phaeophytas,
conocidas como algas pardas, las cuales son
conocidas por producir sustancias como diterpenos,
florotaninos, fucoxantinas, acetogeninas, entre otros
metabolitos. De estos, los florotaninos son los que
han recibido mayor atención en los últimos 5 años
por las diversas propiedades de interés biomédico
que poseen. No obstante, los estudios relacionados a
la actividad antibacteriana de extractos de algas
pardas no abundan en la literatura científica, y
menos aún en Macrocystis pyrifera.
Partiendo del concepto que ciertas algas producen
sustancias bioactivas que impiden el crecimiento de
organismos patógenos en su hábitat natural resulta
interesante el uso de metabolitos obtenidos de dichos
organismos como antibióticos alternativos. En este
sentido, el presente trabajo busca evaluar la
actividad antibacteriana de dos extractos obtenidos
de diferentes partes anatómicas de Macrocystis
Página 15 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
INFORMACIÓN DEL
ARTÍCULO:
Morales M, Nunja P, Burga L,
Justo S, Ávila J, Guerra A.
Actividad antimicrobiana del
extracto acetona-agua de
Macrocystis pyrifera (C.
Agardh 1820) en bacterias de
importancia clínica. Boletín
Científico. 2014. 1(2): 15-19.

Artículo de Investigación
pyrifiera en bacterias patógenas de importancia
clínica
2. Materiales y Métodos
Se colectaron ejemplares juveniles flotantes de
Macrocystis pyrifera (DMR<20 cm) de la bahía de
Pucusana (12º28’38”LS 76º47’53”LO) en los meses
de enero y julio del 2014. Los ejemplares fueron
transportados en bolsas negras sin agua de mar al
Laboratorio de Biología Marina y Continental de la
Universidad Ricardo Palma, donde se limpiaron de
epífitos y sales con agua de caño. Previo a la
extracción, las muestras (filoides, esporófilos y toda
el alga) se secaron en una estufa a 60°C hasta
obtener un peso constante, 1g se maceró en
oscuridad por 1 hora en 10 ml de acetona: agua
(7:3) a temperatura ambiente con agitación cada 15
minutos. Se realizó un tratamiento más utilizando
10 g de alga fresca siguiendo el procedimiento antes
mencionado. El extracto se filtró en papel filtro
(Rundfilter 125 mm), luego se evaporó el solvente
en estufa, se resuspendió en agua destilada y,
finalmente, se filtró a 0.22 µm (Merck Millipore).
Para relacionar la actividad antibacteriana con el
contenido de polifenoles de los extractos, estos se
cuantificaron usando el método de Folin-Ciocalteu
usando como estándar floroglucinol. El contenido
de fenoles totales (CFT) fueron expresados en mg
equivalentes de floroglucinol/100 g de alga seca
(mg PGE/100 g).
La actividad antimicrobiana se evaluó empleando la
técnica del antibiograma por el método de discos de
difusión en agar (INS, 2002). Se usaron discos de
papel de filtro Whatman N° 42 y se cargaron
asépticamente con 30 µl del extracto algal. Se
cargaron discos con 10 mg de Gentamicina que se
usó como control y se dejó en absorción durante 2-3
horas.
Se usaron 4 cepas bacterianas aisladas de diferentes
hospitales de Lima: Staphylococcus aureus,
Escherichia coli, Salmonella enterica y
Pseudomonas aeruginosa. Cada cepa fue reactivada
en Caldo CASO; posteriormente, se procedió con la
preparación del estándar para el inóculo. El inóculo
se ajustó hasta una turbidez de 0.08 OD a 0.1 OD en
625 nm, usando cubetas esterilizadas con radiación
U.V. Luego del ajuste del estándar las cepas fueron
sembradas en placas con Agar Mueller-Hinton e
incubadas a 37°C por 24 horas.
Los halos de inhibición obtenidos fueron medidos
con vernier. La actividad antibacteriana se clasificó
como Resistente (R), Sensible (S) e Intermedia (I)
(Tabla 1). Este procedimiento se realizó con tres
repeticiones para cada uno de los extractos.
3. Resultados
El extracto realizado a partir de toda el alga fresca
(AF) tuvo el mayor CTF (contenido total de
fenoles), lo cual fue estadísticamente mayor en
comparación al resto de extractos (Tabla 2).
De los extractos evaluados, ninguno presentó una
actividad antimicrobiana. El control presentó un
diámetro de inhibición esperado lo que permite
validar la prueba, además según dicho diámetro, las
cepas fueron clasificadas como sensibles a
gentamicina (Tabla 3).
4. Discusión
La actividad antibacteriana de distintas algas
pertenecientes al grupo de las Phaeophytas se ha
evaluado constantemente en diferentes laboratorios
alrededor del mundo. Las sustancias bioactivas de
estas algas les permiten defenderse en su medio
natural, contra organismos microbianos y epífitos;
entre estas sustancias se encuentran los florotaninos,
los cuales son los responsables de la alta actividad
Clasificación Diámetro del halo de
inhibición formado por
Control
Resistente (R) 12mm ≥ DHI
Intermedia (I) 12mm ≤ DHI < 14mm
Sensible (S) DHI ≥ 15mm
Página 16 Laborator io de Microbio logía Volumen 1, n°2
Tabla 1. Clasificación de la actividad antibacteriana
con Gentaminica.
DHI: diámetro de inhibición de la sustancia evaluar.

antibacteriana de varias algas pardas, tal como lo
reportó Jeong L (2014) en su trabajo con Eisenia
byciclis. La mayoría de algas pardas a las que se les
atribuye actividad antibacteriana lo realizan
mediante los florotaninos. En nuestro trabajo se usó
Macrocystis pyrifera, la cual posee
aproximadamente 34,4 mg/g de taninos en épocas de
verano y 0.547 mg/g en épocas de invierno del total
de polifenoles que existen en el alga, lo cual influye
en su capacidad antibacteriana (Castro M.1993).
Por otro lado, la acción de los solventes en los
extractos de algas también influye de alguna manera
en los resultados que se obtienen. Es así que la
extracción acuosa de algas como Sargassum
wigthiiy Padina tetrastromatica presentaron
actividad antimicrobiana ante cepas patógenas tales
como Staphylococcus aureus, Escherichia coli y
Pseudomonas aeruginosa y en cepas patógenas de
peces como Vibrio fischeri, Vibrio alginolithycus.
(Johnsi G. 2011).
El método por discos de difusión es uno de los más
usuales cuando se realiza un antibiograma y se
encuentra actualmente estandarizado por el Instituto
Nacional de Salud del Perú (2002), en la cual se
recomienda el uso de antibióticos obligatorios según
las cepas que se evalúan. Entre estos, la gentamicina,
es uno de los antibióticos de uso oftálmico que se
usan para todas las cepas que se evaluaron en este
trabajo. Además se ha usado esta sustancia como
control en cultivos de Escherichia coli a una
concentración de 2 mg/ml, tal como se realiza en
nuestro trabajo.
Los discos de difusión fueron cargados con 30 µl del
extracto algal y colocados en el medio de cultivo. En
otros estudios, se usaron 20 µl de extracto acuoso en
los discos, los cuales presentaron un resultado
positivo de actividad antimicrobiana ante cepas
clínicas (Johnsi G. 2011). Sin embargo, también
existe el método de pocillos de 6 mm en el medio de
cultivo, los cuales fueron cargados con 50 µl del
extracto algal, permitiendo también la obtención de
resultados positivos (Magallanes C. 2003). El uso de
ambos métodos permite la difusión de los extractos.
Finalmente, varias especies emparentadas con
Macrocystis pyrifera presentan una actividad
antibacteriana tratadas con diferentes métodos de
extracción y de antibiograma. No obstante, nuestra
especie no presentó ningún resultado positivo
durante la investigación, tal como un trabajo
realizado en nuestro país donde trabajaron con
extractos etanólicos de M. pyrifera evaluado
mediante el método de pocillos, sin encontrar
actividad antibacteriana en cepas clínicas y no
clínicas. Estos resultados los atribuyeron a que
posiblemente existe una concentración mínima de
sustancias bioactivas en estas algas las cuales son
Página 17 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
Tabla 2. Contenido Total de Fenoles (CTF) de los
diferentes extractos evaluados.
A: Extracto de alga entera seca. E: Extracto de los esporófilos
de una muestra seca. F: Extracto de Filoides de una muestra
seca. AF: Extracto de una alga fresca. Las letras indican
diferencias significativas (p<0.01).
Tabla 3. Promedio del diámetro de los halos de
inhibición formados en el ensayo con los extractos
acetona-agua de Macrocystis pyrifera
*C: Control (Gentamicina). A: Extracto de alga entera seca.
E: Extracto de los esporófilos de una muestra seca. F:
Extracto de Filoides de una muestra seca. AF: Extracto de una
alga fresca.
Extracto CTF (mg PGE/100 g alga
seca)
AF 385.12±8.13a
A 201.87±20.87b
E 373.87±4.93c
F 327.57±0.94d

solubilizadas en el ambiente acuático (Magallanes
C. 2003).
5. Conclusiones
El extracto acetona-agua de los filoides, esporófilos,
y del alga entera seca y fresca de Macrocystis
pyrifera no posee actividad antibacteriana ante las
cepas de importancia clínica evaluadas.
6. Referencias Bibliográficas
Cardozo K, Guaratini T, Barros M, Falcao V, Tonon
A, Lopes N, Campos S, Torres M, Souza A,
Colepicolo P, Pinto E. 2007. Metabolites from algae
with economical impact. Comparative
Biomchemistry and Physiology. Parte C, 146: 60-
78.
Castro M, Carrillo S, Pérez-Gil F. 1994.
Composición química de Macrocystis pyrifera
(Sargazo Gigante) recolectada en verano e invierno
y su posible empleo en la alimentación animal.
Ciencias Marinas. 20(1): 33-40.
Dae-Sung L, Min-Seung K, Hye-Jin H, Sung-Hwan
E, Ji-Young Y, Myung-Suk L, Won-Jae L, You-Jin
J, Yae-Sue C, Young-Mog K. 2008. Synergistic
effect between Dieckol from Ecklonia stolonifera
and β-lactams against Methicillin-resistant
Staphylococcus aureus. Biotechnology and
Bioprocesses Engineering 13: 758-764.
García A, Gutiérrez M, Acedo E, Burgos A, López
M, Valdés M, Burboa M. 2013. Las algas y otros
organismos marinos como fuente de moléculas
bioactivas. Revista Biotecnia 15(1): 25-32.
Instituto Nacional de Salud del Perú. 2002. Manual
de procedimientos para la prueba de sensibilidad
antimicrobiana por el método de discos de difusión.
Serie de Normas 30: 30 – 36.
Jeong L, Sung E, Eun L, Yeoun J, Hyo K, Mi J,
Kwang S, Hee L, Ji K, Myung L, Young K. 2014. In
vitro antibacterial and synergistic effect of
phlorotanins isolated from edible brown seaweed
Eisenia bicyclis against acne-related bacteria. Algae.
29(1): 47-55.
Jhonsi G, Lipton A, Aishwarya M, Sakira A,
Udayakumar A. 2011. Antibacterial activity of
aqueous extract from selected macroalgae of
southwest coast of India. Seaweed Research and
Utilization. 33(1-2): 67-75.
Kayalvizhi K, Vasuki S, Anantharaman P,
Kathiresan K. 2013. Antimicriobial activity of
seaweeds from the gulf of Mannar. International
Journal of Pharmaceutical Applications. 3(2): 306-
314.
Lara-Issasi G. 1991. Propiedades antibióticas de
algunas algas marinas bentónicas. Hidrobiológica. 1
(2): 21-28.
Lara-Issasi G, Álvarez J, Lozano C, Hernández N.
1999. Nuevas adiciones al conocimiento de la
actividad antibiótica de macroalgas marinas
mexicanas. Hidrobiológica 9(2): 159-169.
Lee D, Kang M, Hwang H, Eom S, Yang J, Lee M,
Lee W, Jeon Y, Choi J, Kim Y. 2008. Synergistic
Effect between Dieckol from Ecklonia stolonifera y
β-Lactams against Methicillin-resistant
Staphylococcus aureus. Biotechnology and
Bioprocess Engineering 13: 758-764.
Magallanes C, Córdova C, Orozco R. 2003.
Actividad antibacteriana de extractos etanólicos de
macroalgas marinas de la costa central del Perú.
Revista Peruana de Biología. 10(2): 125-132.
Manivannan K, Karthikai G, Anantharaman P,
Balasubramanian T. 2011. Antimicrobial potential
of selected brown seaweeds from Vedalai coastal
waters, Gulf of Mannar. Asian Pacific Journal of
Tropical Biomedicine. 10(11): 114-120.
Página 18 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación

Nagayama K, Iwamura Y, Shibata T, Hirayama I,
Nakamura T. 2002. Bactericidal activity of
phlorotanins from the brown alga Ecklonia kurome.
Journal of Antimicrobial Chemotherapy 50: 889-
893.
Ríos N, Medina G, Jiménez J, Yáñez C, García M,
Di, Bernardo M, Gualtieri M. 2009. Actividad
antibacteriana y antifúngica de extractos algales de
algas marinas venezolanas. Revista Peruana de
Biología. 16(1): 97 -100.
Seenivasan R, Rekha M, Indu H, Geetha S. 2012.
Antibacterial activity and phytochemical analysis of
selected seaweeds from Mandapam Coast, India.
Journal of Applied Pharmaceutical Science. 2(10):
159-169.
Srivastava N, Saurav K., Mohanasrinivasan V,
Kannabiran K., Singh M. 2010. Antibacterial
potential of macroalgae collected from Madappam
coast, India. British Journal of Pharmacology and
Toxicology. 1(2): 72-76.
Sung E, Min K, Young K. 2008. Antibacterial
activity of the Phaeophyta Ecklonia stoloniferaon
Methicillin resistant Staphylococcus aureus. Journal
of Fisheries Science and Technology. 11(1): 1-6.
Wisespongpand P, Kuniyoshi M. 2003. Bioactive
phloroglucinols from the brown alga Zonaria
diesingiana. Journal of Applied Phycology. 15: 225-
228.
Zbakh H, Chiheb H, Bouziane H, Motilva V, Riadi
H. 2012. Antibacterial activity of benthic marine
algae extracts from the Mediterranean coast of
Morocco. Journal of Microbiology, Biotechnology
and Food Sciences. 2(1): 219-228.
Página 19 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación

Determinación de Géneros Bacterianos en el Mar de la Playa Cantolao – La Punta – Callao Justo S*, Churasacari T, Saldaña C, Cajachagua C, Pabón L, Gónzales M, Santiago D, Guerra A. Laboratorio de Microbiología. Facultad de Ciencias Biológicas. Universidad Ricardo Palma. *Autor para la correspondencia: Justo Arévalo, Santiago. [email protected]
1. Introducción
Los océanos comprenden aproximadamente el 70%
de la superficie terrestre; la diversidad de sus
ambientes permite que se desarrolle gran cantidad de
formas de vida; además tienen importancia como
regulador de la temperatura global y como sumidero
de carbono
De esta primera idea se desprenden dos principales:
la diversidad de especies y la importancia ecológica
de los ecosistemas marinos.
En primer lugar, la diversidad biológica marina
comprende todo tipo de organismos que habiten en
los mares, un grupo importante con poca
consideración en nuestro país son las bacterias;
estudios recientes en otras partes del mundo han
hallado bacterias marinas con propiedades que van
desde la producción de pigmentos utilizables (Yada
S et al. 2008) hasta bacterias que sintetizan
nanopartículas de oro a partir de otros compuestos
(Shama N et al. 2012), además de bacterias
ampliamente estudiadas como Marinobacter
hydrocarbonoclasticus capaz de producir enzimas
que degradan hidrocarburos aromáticos, Vibrio
fischeri que tiene en su genoma el operon LUX
(biolumniscencia) o Pseudomonas
pseudoalcaligenes productora de enzima que
degrada el cianuro.
En segundo lugar, el bacterioplancton tiene un
INFORMACIÓN DEL
ARTÍCULO:
Justo S, Churasacari T,
Saldaña C, Cajachagua C,
Pabón L, Gonzáles M,
Santiago D, Guerra A.
Determinación de Géneros
Bacterianos en el Mar de la
Playa –La Punta-Callao.
Boletín Científico. 2014.
1(2): 22-25
R E S U M E N
Los océanos albergan una gran diversidad biológica, hasta el momento
relativamente poco explorada por el humano. Con el objetivo de ampliar los
conocimientos en la biodiversidad bacteriana del mar peruano, se realizaron
muestreos aleatorios en el mar de Playa Cantolao-La Punta-Callao
(12º04´06´´ S 77º10´10´´O), en total se tomaron 5 muestras en frascos
estériles a una profundidad de 3m utilizando una vara de madera
acondicionada a una jeringa de 20ml. Para el aislamiento se utilizó el método
de dispersión en placa en Agar Marino, se trabajó con diluciones sucesivas
para obtener colonias aisladas. Para la identificación se realizaron pruebas
bioquímicas que incluían determinación de citocromo oxidasa, motilidad,
producción de H2S, producción de indol, metabolismo de glucosa, producción
de pigmento, luminiscencia, catalasa y desarrollo en anaerobiosis; además de
las coloraciones Gram y de espora. Los esquemas propuestos por Das S et al.
(2007), Oliver J (1982) y el Bergey´s Manual 2da y 7ma edición fueron
utilizados para la determinación de géneros. Se logró el aislamiento de 20
cepas bacterianas. Los géneros encontrados fueron: Moraxella, Bacillus,
Acinetobacter, Vibrio, Aeromonas, Photobacterium, Alteromonas,
Alcaligenes, Streptococcus y Chromobacterium.
Palabras Claves: Bacterias
Marinas, Esquemas de Das
S et al (2007) y Oliver J
(1982), Pruebas bioquímicas
Página 20 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación

papel significativo como principal contribuyente en
los procesos biogeoquímicos de los ambientes
marinos, y representa un componente fundamental
en las redes alimentarias . En los últimos años, el
concepto de red trófica clásica ha cambiado a un
modelo alternativo más complejo denominado
"bucle microbiano", basado en la capacidad de las
bacterias heterótrofas de reciclar la materia orgánica
disuelta y de hacerla circular mediante diversas
interacciones ecológicas a través de los otros
componentes del plancton y, en forma indirecta,
también a través de los niveles tróficos superiores
del ecosistema marino (Schauer M et al. 2011).
En los océanos, los microorganismos son los
responsables del 98% de la producción primaria e
intervienen en todos los ciclos de la materia, con un
papel fundamental, a su vez, en el destino de los
contaminantes en zonas altamente antropizadas.
Además los organismos heterótrofos como las
bacterias consumen entre el 18% y 77% de carbono
orgánico disuelto en el mar (Azam F et al. 1983).
Por lo tanto el estudio de las bacterias en los
ambientes marinos de nuestro país nos permitirá
aumentar nuestros conocimientos en la ecología y
buscar nuevos recursos aprovechables. El presente
estudio tiene como objetivo identificar los géneros
bacterianos presentes en el mar de Playa Cantolao-
Callao como base para futuros estudios ecológicos y
biotecnológicos.
2. Materiales y Métodos
Recolección de muestras: Se realizaron muestreos
aleatorios en el mar de la playa Cantolao-La Punta-
Callao (12º04´06´´ S 77º10´10´´O). Se tomaron 5
muestras, a una profundidad de aproximadamente 3
metros utilizando una vara de madera acondicionada
con jeringas estériles de 20ml. Además se colectaron
3 litros de agua de mar para la preparación de los
medios de cultivo. Las muestras de agua fueron
transportadas al Laboratorio Microbiología de la
Universidad Ricardo Palma en un cooler refrigerado
a 4ºC.
Aislamiento de bacterias:
Se utilizó el método de dispersión en placa con
medio marino (fig. 1).Para esto se tomó 1ml de la
muestra y se colocó en un tubo con 9ml de agua de
mar esterilizada y filtrada obteniendo la dilución 10-
1, se repitió este procedimiento obteniendo 3
diluciones (hasta 10-3). Las placas fueron incubadas
a Tº ambiente (20-25°C) durante 48h. Al cabo de
este tiempo se seleccionaron las colonias aisladas y
fueron reaisladas en tubos con Agar Marino. Estas
fueron incubadas nuevamente a Tº ambiente hasta
observar crecimiento visible y luego fueron
almacenadas a 4ºC hasta el momento de la
caracterización bioquímica.
Determinación Bioquímica:
Se realizaron las siguientes pruebas bioquímicas:
coloración gram, citocromo oxidasa, motilidad,
producción de H2S, producción de indol,
metabolismo de glucosa, producción de pigmento,
luminiscencia, catalasa, desarrollo en anaerobiosis,
coloración de esporas. (fig.2)
Los esquemas propuestos por Das S et al. (2007),
Oliver J (1982) y el Bergey´s Manual 2da y 7ma
edición fueron utilizados para la determinación de
géneros
3. Resultados
En total se obtuvieron 20 cepas, de las cuales 17
lograron ser identificadas hasta la categoría de
Figura 1 Aislamiento por diluciones sucesivas en medio
Marino.
Página 21 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación

género; 3 no tuvieron crecimiento suficiente para su
identificación.
Se encontraron 10 géneros. De los cuales, 8 fueron
bacilos gramnegativos y sólo 2 grampositivos, un
bacilo y un coco (tabla 1).
4. Discusiones
Pellón et al. (2001) en su trabajó con muestras de 2
moluscos bivalvos cultivados en Pucusana y el
Callao: Argopecten purpuratus “concha de abanico”
y Crassostrea gigas “ostra” logran identificar 6
géneros bacterianos: Vibrio (40%), Aeromonas
(15%), Flavobacterium (10%), Pseudomonas (5%),
Moraxella (5%), Flexibacter (5%) y no identificados
(20%), León et al (2010) por su parte en su trabajó
con muestras de 5 invertebrados intermareales
recolectados en la Bahía de Ancón: Phymactis
clematis “anémona”, Tetrapygus niger “erizo de
mar”, Heliaster helianthus “estrella de mar”, peje
sapo y Tegula atra “caracol” logra identificar 4
géneros: Vibrio (50%), Pseudomonas (10%),
Flavobacterium (20%) y Actinomycete (20%), el
presente trabajo reporta la presencia en el ambiente
marino de 10 géneros bacterianos entre ellos
Aeromonas, Moraxella y Vibrio sin embargo no se
encontró cepas de Flavobacterium, Flexibacter ni
Actynomicetes lo que sugiere que estos géneros
podrián estar más relacionados a organismos que
géneros como Bacillus, Photobacterium y
Alteromonas que no fueron encontrados en los
trabajos mencionados anteriormente pero sí en este.
Pellón et al. (2001) y León et al. (2010) determinan
que la cepa más abundante es la correspondiente a
Vibrio sin embargo en este trabajo la cepa más
abundante fue la de Moraxella, esto puede ser a
Figura 2: A; Prueba de catalasa, B; Citocromo oxidasa,
C; SIM, D; Agar P (pigmentación), E; Agar Marino
(Anaerobiosis), F; HughLeifson (metabolismo de
glucosa).
Página 22 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
CAT=Catalasa; OXID=Oxidación; MOT=Motilidad; PIG= Pigmentación; ANAEROB=Anaerobiosis; H.F= HughLeifson;
Ab=Aerobio, Cer=Anaerobio; NP=. No hay presencia de espora; N= No se realizó la prueba.
Tabla 1. Resultados de las Pruebas Bioquímicas realizadas a las cepas aisladas
A B C D
E F

Página 23 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
causa de que hay mayor asociación de Vibrio con
invertebrados que el género Moraxella. Además en
un estudio realizado en Argentina, en la zona
costanera de Comodoro se reporta en muestras de
sedimentos y agua marina 6 de los géneros hallados
en este trabajo (Acinetobacter, Aeromonas, Vibrio,
Moraxella, Bacillus, Photobacterium),
adicionalmente determinan que la mayor cantidad
de cepas halladas fueron grampositivas a diferencia
del presente trabajo que encuentra que el 80% de
los géneros y el 90% de las cepas aisladas fueron
gramnegativas, esto sugiere y afirma que las
grampositivas están mayormente asociados a los
suelos y que las gramnegativas más bien habitan
asociadas a la columna de agua.
5. Conclusiones
Se determinó la presencia de 10 géneros
bacterianos, estos fueron: Moraxella (23,5%),
Bacillus (5,88%), Acinetobacter (5,88%), Vibrio
(11,76%), Aeromonas (11,76%), Photobacterium
(5,88%), Alteromonas (11,76%), Alcaligenes
(5,88%), Streptococcus (5,88%) y
Chromobacterium (11,76%).
6. Referencias Bibliográficas
Azam F, Fenchel T, Field J, Gray J, Meyer-Reil A,
Thingstad F. 1983. The ecological role of water-
column microbes in the sea. Marine Ecology
Progress Series 10: 257-263.
Das S, Lyla P, Ajmal S. 2007. A simple scheme for
the identification of marine heterotrophic bacteria.
An international journal of marine sciences
Thalassas. 23(2): 17-21.
Kaysner C, De Paola A. 2004. Bacteriological
Analytical Manual. FDA (internet). (citado el 09 de
Febrero del 2014). Disponible en: http://
www.fda.gov/Food/FoodScienceResearch/
LaboratoryMethods/ucm070830.htm
Oliver J. 1982. Taxonomic scheme for the
identification of marine bacteria. Deep Sea
Research. 29(6A): 795-798.
Pucci G, Acuña A, Llanes M, Tiedemann M, Pucci,
O. 2009. Identificación de bacterias marinas
cultivables de la ciudad costera Comodoro
Rivadavia, Argentina. Revista de biología marina y
oceanografía. 44(1), 49-58.
Schauer M, Balagué V, Pedros-Alio C, Massana R.
2003. Seasonal changes in the taxonomic
composition of bacterioplankton in a coastal
oligotrophic system. Aquatic Microbial Ecology. 31:
163-74.
Shama N, Pinnaka A, Raje M, Fnu A, Sharkar M,
Roy A. 2012. Exploitation of marine bacteria for
production of gold nanoparticles. Microbial cell
factories. 11: 86-92
Shikongo M, Chimwamurombe P, Venter S. 2012.
Identification of putative Vibrio species isolated
from processed marine fish using thiosulphate-
citrate-bile-sucrose (TCBS) agar. British
biotechnology journal. 2(4): 229-246.
Yada S, Wang Y, Zou Y, Nagasaki K., Hosokawa
K, Osaka I, Arakawa R, Enomoto K. 2008. Isolation
and characterization of two groups of novel marine
bacteria producing violcain. Marine biotechnology.
10: 128-132.
Pellón F, Orozco R, León J. 2001. Bacterias marinas
con capacidad antimicrobiana aisladas de moluscos
bivalvos en cultivos. Revista Peruana de Biología. 8
(2): 1-3.
León J, Liza L, Soto I, Torres M, Orosco A. 2010.
Bacterias marinas productoras de compuestos
antibacterianos aisladas a partir de invertebrados
intermareales. Revista Peruana de Medicina
Experimental. 27(2): 215-21.
Ruíz C. 2013. Respuesta antagónica de
microorganismos aislados de los cultivos de
microalgas en el laboratorio de cultivos marinos.
DELOS. 6(18):1-5.

Efecto antibiótico de Piocianina de Pseudomonas aureginosa sobre Escherichia coli y Staphylococcus aureus. Justo S, Churasacari T, Elías C, Guerra A. Laboratorio de Microbiología. Facultad de Ciencias Biológicas. Universidad Ricardo Palma. *Autor para la correspondencia: Justo Arévalo, Santiago. [email protected]
R E S U M E N
Pseudomonas aeruginosa es una bacteria gramnegativa, patógeno oportunista
de importancia clínica pues causa gran cantidad de enfermedades
intrahospitalarias. Además este es un organismo ubicuo debido
principalmente a su alta capacidad para metabolizar variedad de sustratos.
Esta bacteria tiene la capacidad de producir varios pigmentos extracelulares,
uno de ellos, la piocianina es una exotoxina perteneciente al grupo de las
fenazinas que cuenta con la capacidad de inhibir el desarrollo de organismos
competidores. Se utilizaron 8 cepas de Pseudomonas aeruginosa, 2
ambientales, 5 nosocomiales y 1 cepa de referencia (ATCC 27853). Para
estimular la producción de piocianina las cepas fueron incubadas por 7 días a
37ºC en caldo P. La extracción de Piocianina se realizó por el método
propuesto por Knight et al. (1978) que constó de una extracción
clorofórmica, seguida de una extracción con HCl. No se observó diferencias
en el efecto de la piocianina de cepas nosocomiales y ambientales frente a
Staphylococcus aureus, sin embargo sólo la piocianina de la cepa de muestra
respiratorio mostró un leve efecto sobre Escherichia coli.
1. Introducción
Psedudomonas sp. es un género cosmopolita, este
grupo es capaz de utilizar y metabolizar un amplio
rango de sustratos, ya sean orgánicos o inorgánicos,
por ello puede vivir bajo muy diversas condiciones,
poseen también gran plasticidad génica (Ruiz L,
2007); encontrándose en suelos y aguas, tanto
continentales como marinas (Baumann L et al.
1971), Pseudomonas aureginosa es el patógeno más
importante de este género, debido a que es un
patógeno oportunista (Francis K y Brown P, 2012)
siendo el más importante en humanos debido al
número, tipo de infecciones y mortalidad asociadas
(Ruiz L, 2007), así mismo es también conocido por
su capacidad de sintetizar diferentes pigmentos
hidrosolubles extracelulares entre los que se destaca
un pigmento verde fluorescente, pioverdina (Merino
L, 2007) que actúa como sideróforo en condiciones
mínimas de Fe (III) y actúan como inhibidores del
crecimiento de microorganismos patógenos en
plantas (Bello D et al, 2006) y un pigmento azul,
piocianina (Young G, 1947), perteneciente a las
fenazinas, soluble en cloroformo.
Se ha demostrado que este pigmento posee actividad
antibacteriana (Merino L, 2007), y que su
producción es activada principalmente bajo efecto
de quorum sensing (Williams P y Cámara M, 2009).
Página 24 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
INFORMACIÓN DEL
ARTÍCULO:
Justo S, Churasacari T,
Elias C. Efecto Antibiótico
de Piocianina de
Pseudomonas aeruginosa
sobre Escherichia coli y
Staphylococcus aureus.
Boletín Científico. 2014.
1(2): 26-29.
Palabras Claves:
Pseudomonas aeruginosa,
Piocianina, antibiótico

El objetivo del presente trabajo es determinar la
actividad antibacteriana del pigmento piocianina
sobre una cepa de Escherichia coli y una cepa de
Staphylococcus aureus.
2. Materiales y Métodos
Obtención de Cepas de Pseudomonas
Se trabajó con 8 cepas de Pseudomonas aeruginosa
(Tabla 1); 5 fueron aislamientos nosocomiales, 1
aislada a partir de eyecciones de paloma, 1 de un
manantial ubicado en la costa verde, además de la
cepa ATCC 27853 de Pseudomonas aeruginosa.
Para la confirmación de las cepas se utilizaron
pruebas bioquímicas que consistían en metabolismo
de glucosa, degradación de citrato, motilidad,
producción de Indol y producción de pigmentos.
Obtención de Pigmentos
Las ocho cepas aisladas fueron sembradas en 5ml de
caldo CASO por 18 horas a 37 º C, luego se ajustó la
turbidez del caldo hasta una OD de entre 0.10 y 0.08
a 625nm; por último se tomó una alícuota de 1ml y
se sembró en 50 ml de caldo P los cuales fueron
incubados por 7 días a 37ºC, agitándose
manualmente dos veces al día para favorecer la
oxigenación del medio.
Extracción de Piocianina
Transcurridos los 7 días se tomaron muestras de 10
ml de Caldo P, se centrifugaron a 3000 rpm por 10
minutos y se transfirió el sobrenadante a un tubo
limpio; se realizó una primera extracción con
cloroformo seguido de una extracción con HCl
0.2N. La fase ácida fue neutralizada con NaOH 0.2N
hasta observar el cambio de coloración. Una vez más
se realizó la extracción clorofórmica y a partir de
esta se cargaron discos de 6 mm de papel filtro
Whatman Nº 42 con 10ul del extracto.
Prueba de Sensibilidad
Para esta prueba se utilizó una cepa de Escherichia
coli ATCC 25922 y una cepa de Staphylococcus
aureus ATCC 25923. Se ajustó la turbidez del caldo
CASO, donde fueron previamente inoculadas, entre
0.10 y 0.08 de OD a 625nm. Cada extracto fue
probado por triplicado en placas con agar Muller
Hinton, como control positivo se utilizó discos de
gentamicina.
Análisis estadísticos
Se realizó la prueba de ANOVA para determinar
diferencias significativas entre los efectos
inhibitorios de los extractos de piocianina de las
diferentes cepas utilizadas.
3. Resultados
No se observó diferencias significativas entre los
halos de inhibición del extracto de piocianina frente
a la cepa de Staphylococcus aureus (Tabla 2).
Sólo la piocianina de la cepa aislada de muestra
respiratorio (Tabla 1) mostró un leve efecto
inhibitoria sobre Escherichia coli.
Código Origen Descripción
A A Aislamiento de
eyecciones de
palomas
B A Manantial de la Costa
Verde en Playa La
Estrella
C N Aislamiento del
Hospital Grau
D N Heridas epiteliales
aislada en el HNGAI
E N Heridas epiteliales
aislada en el HNGAI
F N Heridas epiteliales
aislada en el HNGAI
G N Muestra respiratoria
aislada en el HNGAI
ATCC
27853
----- Donación del aisladas
en el HNGAI
Página 25 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
Tabla 1. Cepas de Pseudomonas aeruginosa
utilizadas.
N= Nosocomial, A=Ambiental. HNGAI= Hospital Nacional
Guillermo Almenara Irigoyen

Código Escherichia
coli
Staphylococcus
aureus
A - 15.79
B - 16.02
C - 18.00
D - .1513
E - 14.37
F - 16.00
G 8.61 16,70
ATCC 27853 - 14.07
Gentamicina 21.30 21.30
4. Discusión
Stephen et al (1981) realizan un ensayo similar
con piocianina donde una cantidad de 2.5 ug por
disco resulta en un halo de inhibición de 17 mm
para Staphylococcus aureus. Un resultado similar
se evidencia en la cepa G la cual también mostró
halo de inhibición frente a Staphylococcus aureus
esto sugiere que las concentraciones obtenidas
pueden haber sido inferiores en los otros casos a
los 2.5ug por disco.
Baron S y Rowe J (1981) encuentran que la
actividad antibacteriana de Pseudomonasa
eruginosa no se manifiesta frente a cepas como
Proteus vulgaris, Enterobacter aerogenes otras
Pseudomonas ni frente a Escehrichia coli; sin
embargo otros autores como Young G (1947) y
Nowroozi J et al (2012) han determinado que la
Piocianina sí tiene efecto contra Escherichia coli e
incluso Nowroozi et al (2011) menciona que las
nanopartículas de plata tienen un efecto sinérgico
inhibitorio tanto en Escherichia coli como en
Staphylococcus aureus. En este trabajo se utilizan
discos cargados con 2 ug, nuestro trabajo también
reportó efecto inhibitorio sobre E. coli, esto
demuestra que al parecer el efecto de
antibacteriano de la piocinana es dependiente de la
concentración y se necesita mayores concentraciones
para inhibir el crecimiento de bacterias
gramnegativas.
Esto se confirma con el trabajo de Knight et al.
(1978) que utilizando el método de pocillos en agar,
una metodología conocida por ser más directa que
los discos de difusión, reportan efecto inhibitorio de
la piocianina sobre Proteus morganii sugiriendo que
altas concentraciones del pigmento no permiten el
desarrollo bacteriano.
Por último, de manera muy interesante ninguno de
los trabajos mencionados anteriormente reportaron
un efecto inhibitorio de la piocianina sobre cepas de
Pseudomonas sugiriendo la existencia de un
mecanismo que contrarreste la acción de la
piocianina.
5. Conclusiones
La cepa de Staphylococcus auereus se mostró
sensible frente al extracto de piocianina de todas las
cepas obtenidas de Pseudomonas aureginosa, por el
contrario Escherichia coli se mostró sensible solo
frente al extracto de piocianina producido por la
muestra G, aislada de HNGAI.
Estudios bibliográficos no muestran efecto
inhibitorio de la Piocianina sobre Pseudomonas
aeruginosa lo que sugiere un mecanismo de
protección frente al efecto de este pigmento.
6. Referencias Bibliográficas
Baron S y Rowe J. 1981. Antibiotic action of
Pyocyanin. Antimicrobial Agents and Chemoterapy.
20(6): 814-820.
Baumann L, Baumann P, Mandel, M, Allen R. 1972.
Taxonomy of aerobic marine eubacteria. Journal of
Bacteriology. 110(1): 402-429.
Bello D, Santo J, Diaz M, Bell A, Torres E, Villa P.
2006. Purificación parcial del pioverdina a partir de
caldos de fermentación de Pseudomonas aureginosa.
ICIDCA. 11(1): 36-39.
Página 26 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación
Tabla 2. Mediciones de Halos de Inhibición de
Piocianina frente a Escherichia coli y
Staphylococcus aureus
*Los resultados mostrados son la media de tres ensayos expresados
en mm.

Francis K y Brown P. 2012. Diversity of
antimicrobial resistance and virulence determinants
in Pseudomonas aeruginosa associated with fresh
vegetables. International Journal of Food
Microbiology. 426241.
Knight M, Hartman P, Hartman, Z, Young V. 1978.
A new Method of Preparation of Pyocyanin and
Demonstration of an Unusual Bacterial Sensivity.
Analitical Biochemistry. 95(1): 19-23.
Merino L. 2007. Pseudomonas aureginosa: Una
bacteria con personalidades múltiples. Revista
Argentina de Microbiología. 39:143
Nowroozi J, Sepahi A, Rashnonejad A. 2012.
Pyocyanine Biosinthetic genes in Clinical and
Environmental Isolates of Pseudomonas aeruginosa
and Detection of Pyocyanine’s Antimicrobial Effects
with or without Colloidal Silver Nanoparticles. Cell
Journal. 14(1): 7-18.
Ruiz L. 2007. Pseudomonas aureginosa: Aportación
al conocimiento de su estructura y al de los
mecanismos que contribuyen a su resistencia a los
antimicrobianos. Tesis Doctoral. Barcelona –
España. Universidad de Barcelona.180 pp
Williams P y Cámara M. 2009. Quorum sensing and
environmental adaptation in Pseudomonas
aeruginosa: a tale of regulatory networks and
multifunctional signal molecules. Current Opinion in
Microbiology. 12(2): 182-191.
Young G. 1947. Pigment production and antibiotic
activity in cultures of Pseudomonas aeruginosa.
Universidad de Boston, Massachusetts. 54: 109-117.
Página 27 Laborator io de Microbio logía Volumen 1, n°2
Artículo de Investigación

Aspectos generales de la crianza del Cuy y
su agente infeccioso: Salmonella Justo, S.
Correo Electrónico: [email protected] Laboratorio de Microbiología. Facultad de Ciencias Biológicas. Universidad Ricardo Palma.
Introducción
Cavia porcellus denominado comúnmente como
cuy, cobayo, curi, conejillo de indias y en inglés
Guinea pig es un mamífero roedor originario de la
región andina, se ha estimado que su domesticación
en el Perú se inició hace 2500 a 3600 años por restos
de abundantes excretas de cuy encontrados en el
templo del Cerro Sechín (Departamento de
Ancash); en el período de Paracas Cavernas entre
700 a 500 a.C se han encontrado indicios de
alimentación con carne de cuy en los pobladores,
además en el Período de Paracas Necrópolis casi
todas las casas tenían un espacio de crianza de cuyes
(500 a.C – 200 d.C.)(Moreno A, 1989). El cuy ha
constituido desde esas épocas una importante fuente
proteica, se ha estimado que en Lima Metropolitana
se consume anualmente 0.1 kg de carne de cuy por
persona anualmente y en la Sierra del Perú 1.8 kg
(INEI, 2009), alcanzando las 116500 toneladas
consumidas anualmente en todo el Perú (Chauca L,
1995). Actualmente viene aumentando la demanda
del cuy para la exportación es así que en el año 2000
se exportaron solo 145,00Kg de carne de Cuy pero
para el año 2006 las cifras se habían incrementado
hasta 7762,73Kg con un valor total de 56794,66
dólares americanos siendo el principal país de
destino Estados Unidos (MINAG, 2014).
Aunque la distribución del cuy no está restringida al
Perú, este presenta la mayor población de cuyes
alcanzando los 23 240 846 animales, estando el 92%
de la población en la sierra y solo el 6% en la costa
R E S U M E N
Actualmente la demanda de cuy para consumo, tanto al interior como al
exterior del país, viene en aumento. La distribución y crianza de esta especie
con fines de comercio está restringido a 4 países Sudamericanos: Ecuador,
Colombia, Bolivia y Perú; siendo Perú el país con la mayor población. Los
sistemas de crianza del cuy se clasifican según su finalidad y la cantidad de
individuos criados. Debido principalmente a deficiencias en la técnicas de
crianza y falta de investigación; la producción del cuy atraviesa graves
problemas por enfermedades infecciosas las cuales diezman a gran parte de la
población en los criaderos. Salmonella se ha identificado como el agente
infeccioso bacteriano más importante por ocasionar altos índices de
mortalidad y morbilidad; los sistemas de control empleados actualmente no
han sido suficientes para atenuar este mal. A nivel mundial se han desarrollado
vacunas contra Salmonella pero estas están diseñadas para su principal uso en
aves de corral; los estudios existentes en vacunas contra Salmonella para
cuyes son escasos y muy pocos enfocados realmente a proteger al cuy de la
Salmonella. En conclusión, se necesita de mayor cantidad y rigurosa
investigación científica tanto en el agente infeccioso como en el cuy y los
sistemas de crianza para mejorar la producción de cuy en el país.
Página 28 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
INFORMACIÓN DEL
ARTÍCULO:
Justo S. Aspectos Generales
de la Crianza del Cuy y Su
Agente Infeccioso:
Salmonella. Boletín
científico. 2014. 1(2): 30-
35.
Palabras Claves: Cuy,
Salmonella, Producción.
(MINAG, 2003). Tanto en Perú como en Ecuador la
distribución del cuy es amplia, encontrándose casi
en la totalidad del territorio, a diferencia de otros
países como Colombia y Bolivia donde la
distribución está restringida a algunos
departamentos y por lo tanto presentan poblaciones
menores. (Chauca L, 1997).
Producción del cuy según los sistemas de crianza
La producción del cuy se puede clasificar en tres
niveles de acuerdo al sistema de crianza (Chauca L,
1997): el sistema familiar, el familiar-comercial y el
comercial. El sistema familiar se caracteriza por
desarrollarse fundamentalmente como sustento para
la alimentación de la familia, los cuyes son criados
por los miembros de la familia y los alimentan con
malezas, residuos de cosecha y residuos de cocina,
se crían entre 20 a 50 cuyes por lugar de crianza
(Zaldivar M et al, 1990); este sistema se encuentra
en departamentos como Cajamarca, Junín y Lima
(Perú), Nariño (Colombia), La Paz y Cochabamba
(Bolivia) y el 90% de la crianza en Ecuador (Chauca
L, 1997; Caycedo, 1981; Chauca L, 1991; INIA,
1999; Moncayo R, 2000). El sistema familiar-
comercial nace siempre de los sistemas familiares en
áreas rurales cercanas a la ciudad, donde el criador
puede fácilmente comercializar su producto; los
productores invierten recursos monetarios para
infraestructura, tierra para la siembra de forrajes y
mano de obra para el manejo de la crianza (Chauca,
L 1995), en este sistema se mantienen entre 50 y
1000 cuyes, y se encuentra en departamentos como
Cajamarca, Junín y Lima (INIA, 1999). Por último
el sistema comercial se presenta mayormente en
empresas agropecuarias, se mantienen áreas de
cultivo para la siembra del forraje, se utiliza además
alimento balanceado; este sistema mantiene por lo
menos 1000 cuyes, Lima es el principal
departamento con este sistema, además de Cuzco y
Arequipa (Peña S, 2007).
Salmonella en cuyes
En la crianza del cuy se han reportado diversas
enfermedades infecciosas ocasionados por bacterias
como Bortedella bronchiseptica, Diplococcus
pneumoniae, Yersinia pseudotuberculosis,
Streptococcus pyogenes, Pasteurella spp.,
Escherichia coli, Clostridium sp., Klebsiella
pneumoniae, Diplococcus pneumoniae,
Corynebacterium pyogenes, Shigella sp.,
Streptococcus zooepidermicus, y especialmente el
género Salmonella (Morales S, 2012; Correa R,
2000).
Salmonella es un género de bacterias gramnegativas
perteneciente a la familia Enterobacteriaceae, este
género está dividido en dos especies Salmonella
enterica y Salmonella bongori, S. enterica
comprende más de 2500 serotipos que están
ordenados en 7 grupos: I, II, IIIa, IIIb, IV, VI y VII.
Existe una gran diversidad en este género.
Actualmente los serotipos de Salmonella pueden
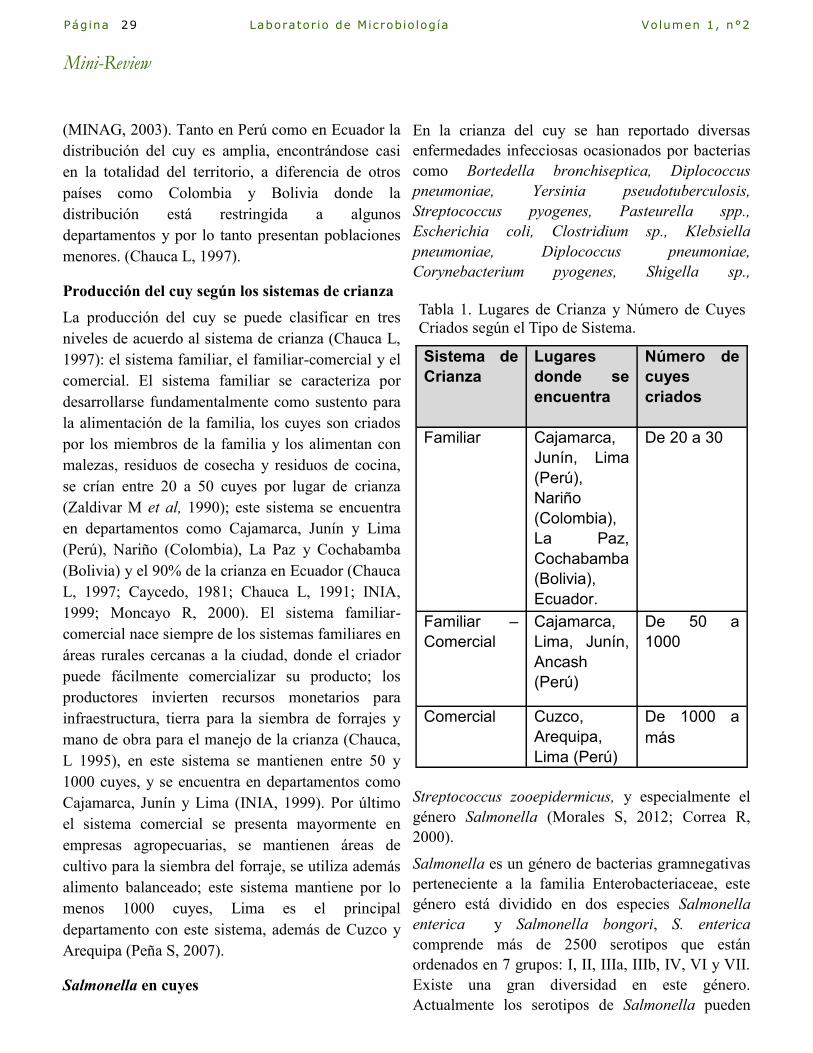
Tabla 1. Lugares de Crianza y Número de Cuyes
Criados según el Tipo de Sistema.
Página 29 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Sistema de
Crianza
Lugares
donde se
encuentra
Número de
cuyes
criados
Familiar Cajamarca,
Junín, Lima
(Perú),
Nariño
(Colombia),
La Paz,
Cochabamba
(Bolivia),
Ecuador.
De 20 a 30
Familiar –
Comercial
Cajamarca,
Lima, Junín,
Ancash
(Perú)
De 50 a
1000
Comercial Cuzco,
Arequipa,
Lima (Perú)
De 1000 a
más

infectar un amplio rango de hospederos, desde
reptiles y aves hasta mamíferos incluyendo humanos
(Gamazo C e Irache J, 2007)
Salmonella es el principal agente infeccioso en los
cuyes, durante largo tiempo se ha observado una
gran susceptibilidad a la salmonelosis en los cuyes,
sin embargo aún son escasos los reportes con datos
exactos que permitan tener una idea clara de la
realidad de la salmonelosis en cuyes, algunos de
estos reportes son el de Leguía P (1993) que señala
que hasta el 95% de muertes de la morbilidad
general es generada por este agente causal variando
los efectos según el estado de desarrollo del cuy
(52.70% en adultos y 19.83% en lactantes), Morales
S et al (2007) quien menciona que Salmonella
provoca brotes de mortalidad y abortos de hasta el
65%, Matsuura A et al (2010) quién reporta en un
criadero de Ancash una prevalencia de 61.5% y
Ramírez A (1972) una prevalencia de 65.5%.
Aunque no existe referencias exactas de cuál es el
serotipo causante de la Salmonelosis en los cuyes, se
ha reportado mayormente presencia de S. enteritidis
(Correa R, 2000; Matsuura A et al, 2010) y S.
thiphymurium (Correa R, 2000; Guamán M, 2014),
además de S. typhi (Dima V et al, 1989) y S. dublin
(Wagner J y Manning P, 1976)
La principal vía de infección es la oral a través de
alimentos contaminados, esto ocasionado en gran
medida por las malas prácticas de manejo, deficiente
nivel de bioseguridad que ocasiona presencia de
roedores y aves contaminados, ingreso no
controlado de personal, estrés en los cuyes (Figueroa
I y Verdugo A, 2005). Además se ha determinado
que las variaciones de temperatura y humedad
predisponen a la aparición de infecciones (Ramírez
A, 1972).
La salmonelosis en cuyes se puede manifestar de
dos formas: crónica o aguda, siendo esta última la
que mayor mortalidad causa presentándose como un
cuadro septicémico agudo que genera muertes en 24
a 48 horas, los signos más frecuentes son
decaimiento, postración, anorexia, adelgazamiento,
pelos ásperos y erizados, y parálisis de los miembros
posteriores; aunque en muchos casos los animales
mueren sin mostrar síntoma alguno (Layme A,
2010). En los casos crónicos se presenta un
adelgazamiento paulatino, pelaje deslucido y
aumento del volumen abdominal (Evans A, 2005).
Control de Salmonella en cuyes
Actualmente, la mayoría de criaderos realiza el
control de la Salmonelosis utilizando antibióticos
como Sulfatrimetoprim, enrofloxacina,
estreptomicina, amoxicilina, cloranfenicol,
fosfomicina. Matsuura A et al (2010) determinaron
que de 40 cepas de Salmonella aisladas a partir de
cuyes el 100% fueron sensibles los 4 primeros
antibióticos mencionados, el 97,5% a gentamicina y
cloranfenicol, y el 92,5% a fosfomicina. Estos
antibióticos son administrados por vía oral a través
del alimento balanceado. Además como ya se
comentó líneas arriba, las buenas prácticas de
manejo como: el control de roedores y aves, el
ingreso controlado del personal, el aislamiento de
cuyes enfermos, el control de sanidad de los
alimentos, la limpieza constante del criadero, entre
otros permitirián prevenir hasta cierto punto el brote
de enfermedades. Sin embargo se sigue requiriendo
de métodos de prevención más efectivos que
permitan desarrollar un sistema de crianza con
menores índices de morbilidad y mortalidad.
Una forma bastante estudiada para el control de la
Salmonella son las vacunas, sin embargo la mayoría
de vacunas producidas actualmente están destinadas
a la industria de aves de corral (Desin T et al, 2013)
y hasta la fecha son pocos los estudios realizados en
cuyes, además resulta interesante recalcar que gran
parte de la información acerca de vacunas contra
Salmonella probadas en cuyes se ha realizado con el
objetivo de utilizarlas en otros animales y no en el
cuy (el cuy es utilizado como animal modelo). Esto,
por lo tanto, no ha permitido que se desarrollen
trabajos continuos enfocados a desarrollar una
vacuna ideal para cuyes, en cambio se han probado
vacunas contra cepas y serotipos de Salmonella de
Página 30 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
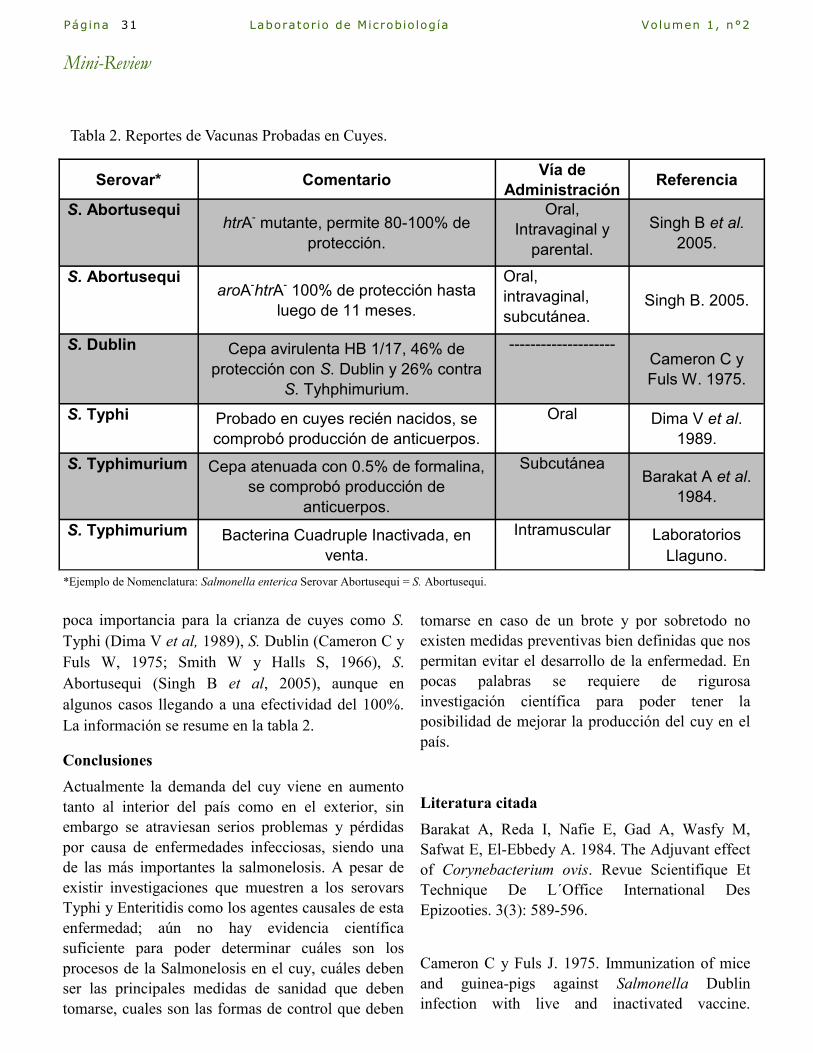
poca importancia para la crianza de cuyes como S.
Typhi (Dima V et al, 1989), S. Dublin (Cameron C y
Fuls W, 1975; Smith W y Halls S, 1966), S.
Abortusequi (Singh B et al, 2005), aunque en
algunos casos llegando a una efectividad del 100%.
La información se resume en la tabla 2.
Conclusiones
Actualmente la demanda del cuy viene en aumento
tanto al interior del país como en el exterior, sin
embargo se atraviesan serios problemas y pérdidas
por causa de enfermedades infecciosas, siendo una
de las más importantes la salmonelosis. A pesar de
existir investigaciones que muestren a los serovars
Typhi y Enteritidis como los agentes causales de esta
enfermedad; aún no hay evidencia científica
suficiente para poder determinar cuáles son los
procesos de la Salmonelosis en el cuy, cuáles deben
ser las principales medidas de sanidad que deben
tomarse, cuales son las formas de control que deben
tomarse en caso de un brote y por sobretodo no
existen medidas preventivas bien definidas que nos
permitan evitar el desarrollo de la enfermedad. En
pocas palabras se requiere de rigurosa
investigación científica para poder tener la
posibilidad de mejorar la producción del cuy en el
país.
Literatura citada
Barakat A, Reda I, Nafie E, Gad A, Wasfy M,
Safwat E, El-Ebbedy A. 1984. The Adjuvant effect
of Corynebacterium ovis. Revue Scientifique Et
Technique De L´Office International Des
Epizooties. 3(3): 589-596.
Cameron C y Fuls J. 1975. Immunization of mice
and guinea-pigs against Salmonella Dublin
infection with live and inactivated vaccine.
Serovar* Comentario Vía de
Administración Referencia
S. Abortusequi htrA
- mutante, permite 80-100% de
protección.
Oral,
Intravaginal y
parental.
Singh B et al.
2005.
S. Abortusequi aroA
-htrA
- 100% de protección hasta
luego de 11 meses.
Oral,
intravaginal,
subcutánea. Singh B. 2005.
S. Dublin Cepa avirulenta HB 1/17, 46% de
protección con S. Dublin y 26% contra
S. Tyhphimurium.
-------------------- Cameron C y
Fuls W. 1975.
S. Typhi Probado en cuyes recién nacidos, se
comprobó producción de anticuerpos.
Oral Dima V et al.
1989.
S. Typhimurium Cepa atenuada con 0.5% de formalina,
se comprobó producción de
anticuerpos.
Subcutánea Barakat A et al.
1984.
S. Typhimurium Bacterina Cuadruple Inactivada, en
venta.
Intramuscular Laboratorios
Llaguno.
Tabla 2. Reportes de Vacunas Probadas en Cuyes.
Página 31 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
*Ejemplo de Nomenclatura: Salmonella enterica Serovar Abortusequi = S. Abortusequi.

Onderstepoort Journal of Veterinary Research. 42
(2): 63-69.
Caycedo V. 1981. Situación de la industria de
Cuyes en Colombia. Memoria Primer Seminario
Andino de Cuyecultura, Colombia, Ecuador y Perú.
Universidad de Nariño. 7-15.
Chauca L. 1997. Producción de Cuyes (Cavia
porcellus). Estudios FAO Producción y Sanidad
Animal. Citado el 02/09/14. Disponible en: http://
www.fao.org/docrep/w6562s/w6562s00.HTM
Chauca L. 1995. Producción de cuyes (Cavia
porcellus) en los países andinos. World Animal
Review. 83(1).
Chauca L. 1991. Caracterización de la Crianza de
cuyes en los Departamentos de Cochabamba, La
Paz y Oruro. Instituto Boliviano de Tecnología
Agropecuaria. 1-65.
Correa R. 2000. Sanidad en Cuyes. V Curso
Latinoamericano de Cuyicultura. Puerto Ayacucho
– Venezuela.
Desin T, Koster W, Potter A. 2013. Salmonella
vaccines in poultry: past, present and future. 2013.
Expert Reviews Vaccines. 12(1): 87-96.
Dima V, Petrovici M, Lacky D. 1989. Reaction and
response of newborn guinea pigs to experimental
Salmonella typhi infection. Archives Roumaines de
Pathologie Experimentale et de Microbiologie. 48
(4): 299-321.
Evans A. 2005. Import risk Analysis: Domestic
Guinea Pig, Cavia porcellus, imported from
Australia. Draft Reports. Citado el 02/09/14.
Disponible en: http://piggyville.com/guinea_pigs/
NZriskanalysis.pdf
Figueroa I. y Verdugo A. 2005. Mecanismos
Moleculares de Patogenicidad de Salmonella sp.
Revista Latinoamericana de Microbiología. 47: 25-
42.
Gamazo C. e Irache J. 2007. Salmonella Vaccines.
Trends in applied Microbiology. SN: 518-524.
Guamán M. 2014. Determinación del Género y
Especie de Salmonella en Cuyes Mestizos en
Diferentes Sistemas de Crianza en la Comunidad de
Oñacapac del Cantón Saraguro. Tesis para obtener
el Título de Médico Veterinario Zootecnista.
Universidad Politécnica Salesiana. Cuenca –
Ecuador.
INEI. 2009. Perú: Consumo per cápita de los
principales alimentos 2008 – 2009. Dirección
Técnica de Demografía e Indicadores Sociales.
Lima – Perú. 117pág.
INIA. 1999. Proyecto: Sistemas de Producción de
Cuyes. Centro Internacional de Investigación para
el Desarrollo. Lima – Perú. 85pág.
Layme A. 2010. Frecuencia de Lesiones
Anatomopatológicas en Cobayos con Diagnóstico
Bacteriológico de Salmonella sp. Remitidos al
Laboratorio de Histología, Embriología y Patología
Veterinaria de la FMV-UNMSM durante el período
2001-2007. Tesis para obtener el Título de Médico
Veterinario. Universidad Nacional Mayor de San
Marcos. Lima – Perú.
Leguía P. 1993. Enfermedades infecciosas y
Parasitarias en Cuyes. Primer Curso Regional de
Reproducción en Cuyes. La Molina – Perú.
Página 32 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review

Matsuura A, Morales S, Calle S, Ara M. 2010.
Susceptibilidad a Antibacterianos in vitro de
Salmonella enterica aislada de Cuyes de Crianza
Familiar-Comercial en la Provincia de Carhuaz-
Áncash. Revista de Investigación Veterinaria del
Perú. 21(1): 93-99.
MINAG. 2014. Sector Agrario: Cuyes.
Departamento de Agricultura. Citado el
02/09/14.Disponible en:
http://www.minag.gob.pe/portal/sector-
agrario/pecuaria/situacion-de-las-actividades-de-
crianza-y-produccion/cuyes?limitstart=0
MINAG. 2008. Sector Agrario: Cuyes.
Departamento de Agricultura. Citado el 02/09/14.
Disponible en: http://www.minag.gob.pe/situacion-
de-las-actividades-de-crianza-yproduccion/cuyes-
39.html
Moncayo R. 2000. Crianza Comercial de Cuyes y
Costos de Producción. V Curso Latinoamericano de
Cuyicultura. Puerto Ayacucho – Venezuela.
Moreno A. 1989. Producción de Cuyes.
Universidad Nacional Agraria La Molina.
Departamento de Producción Animal. Lima – Perú.
132pág.
Morales S. 2012. Patógenos Oportunistas por
Transmisión Fecal-Oral en Cuyes Reproductores
Introducidos al Distrito de San Marcos. Científica.
9(1): 33-38.
Morales S, Mattos J, Calle, S. 2007. Efecto de la
muña (Satureja parvifolia) en la dinámica de la
infección por Salmonella entérica en cobayos. XXX
Reunión Científica Anual Asociación Peruana de
Producción Animal. Cusco – Perú.
Peña S. 2007. Estudio de la Comercialización de
Cuyes en una Región Andina. Tesis de final de
Carrera. Universidad Politécnica de Madrid.
Madrid – España.
Ramírez A. 1972. Estudio Bacteriológico y
epidemiológico de un brote infeccioso en cobayos
(Cavia porcellus). Tesis para obtener el Título de
Médico Veterinario. Universidad Mayor de San
Marcos. Lima – Perú.
Singh B, Alam J. y Hansda D. 2005. Alopecia
induced by salmonelosis in guinea pigs. Veterinary
Record. 156: 516-518.
Singh B. 2005. Development of double defined
deletion mutant vaccine for control of Salmonella
Abortusequi infections in equines. IVRI. Izatnagar
– India.
Smith W y Halls S. 1966. The immunity produced
by rough Salmonella Dublin variant against
Salmonella typhimurium and Salmonella
choleraesuis infection in guinea-pigs. Journal of
hygiene. 64(3): 357-359.
Wagner J y Manning P. 1976. The Biology of
Guinea Pig. Academic Press. Londres. Edición 1.
Zaldivar A, Gamarra M, Florian A. 1990.
Determinación de la Capacidad de Carga para
Cuyes (Cavia porcellus L.) machos reproductores.
12ava Reunión ALPA. Campinas – Brasil. 177pág.
Página 33 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review

Fundamentos básicos de PCR convencional y sus variantes
Médico, A.
Correo Electrónico: [email protected]
Laboratorio de Microbiología. Facultad de Ciencias Biológicas. Universidad Ricardo Palma .
R E S U M E N
La reacción en cadena de la polimerasa (PCR) es una técnica de biología
molecular, desarrollada por Kary Mullis en 1986, utilizada para amplificar a
partir de una o unas pocas copias una secuencia de ADN, generando miles a
millones de copias de esa secuencia de ADN. Esta técnica involucra tres pasos
principales: desnaturalización, hibridación y extensión, los cuales se repiten en
ciclos para lograr la amplificación; es necesario dar énfasis al diseño de
cebadores y sus consideraciones básicas, ya que esto aporta la especificidad a
la técnica. Por último, en la actualidad el PCR es una técnica común e
indispensable en los laboratorios de investigación biológica debido a su gran
variedad de aplicaciones; la diversidad de aplicaciones que existen han
permitido que se generen variantes de la técnica clásica: Nested PCR, Real
Time PCR, RT-PCR, Inverse PCR. Este artículo es un corto repaso de los
fundamentos básicos de un PCR convencional y algunas de sus variantes.
Introducción
Es indiscutible la importancia de la replicación del
ADN desde niveles tan simples como el celular
hasta niveles ecológicos, ya que con ella la célula es
capaz de continuar su división celular. Por este
mismo motivo es importante en el estudio del
cáncer, que ocasiona casi 15 millones de muertes al
año en todo el mundo, de las cuales solo en el Perú
se presentan aproximadamente 31 mil nuevos casos
por cada año (Ramos W et al, 2013).
En 1957 los biólogos estadounidenses Matthew
Meselson y Franklin Stahl utilizando cultivos
bacterianos de Escherichia coli, demostraron que el
ADN celular se replica de la forma,
semiconservativa, propuesta por Watson y Crick
(Watson J y Crick F, 1953). Estos investigadores
utilizaron el elemento nitrógeno (N) presente en la
molécula de ADN, con el 14N natural y la
incorporación del isótopo de 15N radiactivo,
pudieron demostrar que cuando el ADN se replica,
una de sus cadenas pasa a las células hijas sin
cambiar (14N) y es la que actúa como molde o
patrón, para así formar una segunda hebra
complementaria (15N) y completar las dos cadenas
del ADN (Watson J et al, 2006) (Ochman H et al,
1988). La replicación del ADN consta de tres
etapas: iniciación, elongación y terminación. Otras
características de la replicación son la
secuencialidad y bidireccionalidad desde un punto
fijo llamado origen de replicación. Este proceso se
desarrolla formando horquillas. In vivo necesita
varias proteínas y enzimas tales como la helicasa,
topoisomerasa, proteínas SSB, ADN polimerasa I y
III, ARN primasa y ADN ligasa (Karp G, 2011).
Con la técnica de la reacción en cadena de la
Página 34 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
INFORMACIÓN DEL
ARTÍCULO:
Medico A. Fundamentos
básicos de PCR
convencional y sus
variantes. Boletín
Científico. 2014. 1(2): 36-
44.
Palabras Claves: PCR,
Cebadores, Fundamentos y
aplicaciones

polimerasa (PCR) todas estas enzimas, a excepción
de las ADN polimerasa I y III, son reemplazadas
por variaciones de temperatura (Tabla 1).
PCR: ¿Por qué temperatura?
La reacción en cadena de la polimerasa (PCR) es
una técnica que fue desarrollada en 1986 por Kary
Mullis. Por el desarrollo de esta técnica Mullis
recibió el Premio Nobel de Química en 1993
(Bartlett J y Stirling D, 2003). Sin embargo,
Gobind Khorana en 1971 ya había descrito el
principio básico de la replicación de una cadena de
ADN utilizando dos cebadores complementarios a
los extremos de la secuencia a amplificar.
Lamentablemente el progreso se vio limitado por la
síntesis de cebadores y la purificación de la
polimerasa (Kleppe K et al, 1971).
El objetivo del PCR es amplificar ADN hasta
obtener un gran número de copias de una secuencia
molde. Pero, ¿cuál es el factor que determina que la
temperatura sea una gran herramienta en la
amplificación de secuencias?
Para esto debemos analizar la estructura del ADN
(Fig. 1). El primer objetivo del PCR es
desnaturalizar las cadenas sin que se afecte la
secuencia de nucleótidos. Esto es posible ya que
los enlaces puente de hidrógeno son enlaces débiles,
por lo tanto se rompen cuando se le aplica calor;
mientras que los enlaces fosfodiéster no se ven tan
afectados por este ya que son enlaces covalentes
fuertes. En la década de los 80, un problema que se
tuvo fue conseguir una ADN polimerasa que no
fuera termolábil, porque la ADN polimerasa común
se desnaturalizaba a altas temperaturas, entonces
después de cada ciclo del PCR se debía agregar
nuevamente, haciendo de este proceso muy lento y
costoso (Erlich H et al, 1991). El uso de una ADN
polimerasa termoestable, obtenida de Thermus
aquaticus (Fig. 2), significó un mayor rendimiento
en el proceso (Erlich H et al, 1991) (Saiki R et al,
1988).
Para desarrollar un óptimo PCR se necesitan
excelentes cebadores ya que estos les dan
especificidad al proceso. Pero debemos saber
primero dos cosas: ¿qué son? (Apartado 1) y ¿con
qué criterio se diseñan? (Apartado 2).
Fue muy difícil en los primeros experimentos
determinar las temperaturas idóneas para la
desnaturalización, hibridación y elongación; es más,
los primeros experimentos de PCR se hicieron a
baño María. Hoy en día se realiza este proceso en
un equipo denominado termociclador de manera
Página 35 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Tabla 1. Lista de reactivos necesarios para
realizar cada proceso. Se aprecia que la
variación de temperatura reemplaza a varias
enzimas necesarias in vivo.
Figura 1. Esquema general de tipos de enlaces en el ADN.
(Tomada de: GeneMol: http://genemol.org/ y modificada
por el autor del artículo).

etapa depende del Tm, tamaño de los cebadores y
del contenido de G/C de cada uno de ellos y es de
50°- 60° C generalmente. La temperatura
incorrecta durante la etapa de hibridación puede
resultar en cebadores que no se unen al ADN
molde en lo absoluto, o la unión inespecífica.
3. Extensión (30’’-2 minutos): La ADN
polimerasa se adhiere a la cadena tomando como
punto de referencia al cebador; añade los
desoxinucleótidos trifosfatados (dNTPs) mientras
se abre camino a lo largo de la cadena de ADN. El
tiempo y la temperatura dependen tanto de la
fidelidad y rendimiento de la ADN polimerasa
como de la longitud del fragmento de ADN
blanco. ,,,,,,,,,,,,,,,,,,,,,,,,,,,,,,,
Después de todo el proceso se puede colocar la
muestra a temperaturas de 4° a 15° si es que se
desea conservar a corto plazo (expuesto a acción de
nucleasas pero no a formación de cristales de
hielo); a -20°, si se desea a mediano plazo y a -70°
controlada y programada. En este se pueden realizar
las variaciones secuenciales repetitivas de
temperatura llamadas ciclos con la confianza de que
la temperatura del ADN y el tiempo en cada paso sea
el correcto.
Pasos de un ciclo de PCR
Cada ciclo consta de tres pasos (Fig. 4); todos estos
ciclos son precedidos de un ‘inicio’ donde el ADN es
desnaturalizado por un tiempo de, aproximadamente,
5 minutos a 94°-96° C para asegurarse de que tanto
el ADN molde y los cebadores estén separados
completamente; este proceso es necesario para ADN
polimerasas que necesitan activación por calor.
Y a su vez todos los ciclo son seguidos de un paso de
‘elongación final’, generalmente de 10 minutos a una
temperatura de entre 70° y 74° C, con esto se asegura
que cualquier ADN de cadena simple restante sea
totalmente ampliado (Haras D yAmoros J, 1994).
1. Desnaturalización (1-4 minutos): El ADN molde
de doble cadena debe ser calentado hasta los 94-96°
C, dependiendo de la cantidad de G/C que tenga, con
el fin de separar las hebras. El calor rompe los
enlaces puente de hidrógeno que une a las bases
nitrogenadas.
2. Hibridación (30’’-2 minutos): Después de
separar las hebras de ADN, la temperatura se reduce
de modo que los cebadores pueden adherirse a las
hebras individuales de ADN. La temperatura de esta
Página 36 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Figura 2. Micrografía de Thermus aquaticus, bacteria
Gram-negativa identificada por Thomas Brock (Tomada de
http://www.musee-frappier.qc.ca/images/site/large/thermus-
aquaticus-dianemontpetitagc.jpg y modificada por el autor).
Apartado 1. Cebadores. Los cebadores son cadenas cortas de ADN artificiales
de no más de cincuenta nucleótidos (mayormente de
18 a 25 pares de bases) que son complementarios al
principio y al final del fragmento de ADN blanco
(Rahman M et al 2013). Luego de hibridarse es
donde la ADN polimerasa actúa y comienza la
síntesis de la nueva cadena en dirección 5’ a 3’ (Fig.
3). El fragmento de ADN a ser amplificado se
delimita por los cebadores utilizados.
Figura 3. Representación de cebadores forward y
reverse y su complementariedad de bases con la
cadena molde. (De
http://en.wikipedia.org/wiki/File:Primers_RevComp.s
vg y modificada por el autor del artículo de revisión)
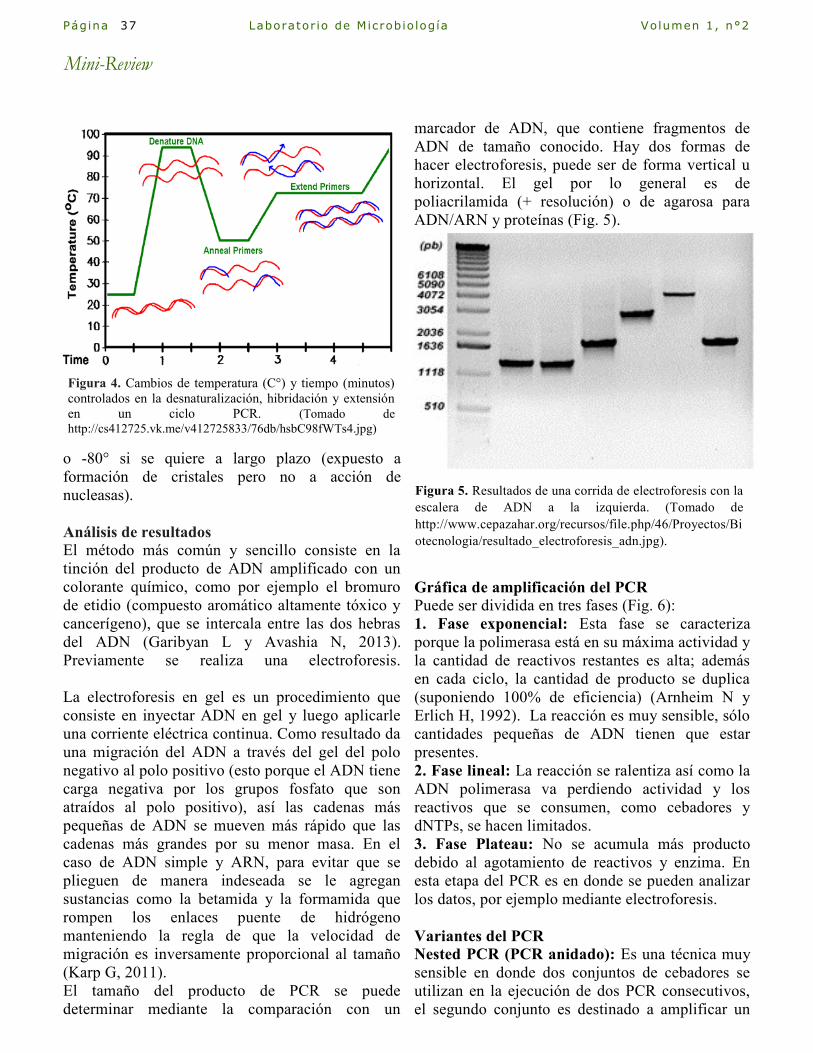
o -80° si se quiere a largo plazo (expuesto a
formación de cristales pero no a acción de
nucleasas).
Análisis de resultados
El método más común y sencillo consiste en la
tinción del producto de ADN amplificado con un
colorante químico, como por ejemplo el bromuro
de etidio (compuesto aromático altamente tóxico y
cancerígeno), que se intercala entre las dos hebras
del ADN (Garibyan L y Avashia N, 2013).
Previamente se realiza una electroforesis.
La electroforesis en gel es un procedimiento que
consiste en inyectar ADN en gel y luego aplicarle
una corriente eléctrica continua. Como resultado da
una migración del ADN a través del gel del polo
negativo al polo positivo (esto porque el ADN tiene
carga negativa por los grupos fosfato que son
atraídos al polo positivo), así las cadenas más
pequeñas de ADN se mueven más rápido que las
cadenas más grandes por su menor masa. En el
caso de ADN simple y ARN, para evitar que se
plieguen de manera indeseada se le agregan
sustancias como la betamida y la formamida que
rompen los enlaces puente de hidrógeno
manteniendo la regla de que la velocidad de
migración es inversamente proporcional al tamaño
(Karp G, 2011).
El tamaño del producto de PCR se puede
determinar mediante la comparación con un
marcador de ADN, que contiene fragmentos de
ADN de tamaño conocido. Hay dos formas de
hacer electroforesis, puede ser de forma vertical u
horizontal. El gel por lo general es de
poliacrilamida (+ resolución) o de agarosa para
ADN/ARN y proteínas (Fig. 5).
Gráfica de amplificación del PCR Puede ser dividida en tres fases (Fig. 6):
1. Fase exponencial: Esta fase se caracteriza
porque la polimerasa está en su máxima actividad y
la cantidad de reactivos restantes es alta; además
en cada ciclo, la cantidad de producto se duplica
(suponiendo 100% de eficiencia) (Arnheim N y
Erlich H, 1992). La reacción es muy sensible, sólo
cantidades pequeñas de ADN tienen que estar
presentes.
2. Fase lineal: La reacción se ralentiza así como la
ADN polimerasa va perdiendo actividad y los
reactivos que se consumen, como cebadores y
dNTPs, se hacen limitados.
3. Fase Plateau: No se acumula más producto
debido al agotamiento de reactivos y enzima. En
esta etapa del PCR es en donde se pueden analizar
los datos, por ejemplo mediante electroforesis.
Variantes del PCR
Nested PCR (PCR anidado): Es una técnica muy
sensible en donde dos conjuntos de cebadores se
utilizan en la ejecución de dos PCR consecutivos,
el segundo conjunto es destinado a amplificar un
Página 37 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Figura 4. Cambios de temperatura (C°) y tiempo (minutos)
controlados en la desnaturalización, hibridación y extensión
en un ciclo PCR. (Tomado de
http://cs412725.vk.me/v412725833/76db/hsbC98fWTs4.jpg)
Figura 5. Resultados de una corrida de electroforesis con la
escalera de ADN a la izquierda. (Tomado de
http://www.cepazahar.org/recursos/file.php/46/Proyectos/Bi
otecnologia/resultado_electroforesis_adn.jpg).

Página 38 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Apartado 2. Diseño de Cebadores.
¿Por qué son importantes los cebadores? La principal razón es porque los cebadores le dan la especificidad al PCR.
Por lo que un buen diseño de ellos llevará a un óptimo desempeño del PCR. En este apartado se verán las
consideraciones básicas para el diseño de estos, el diseño de un cebador depende de la variante de PCR.
Ubicación del cebador: Definir la ubicación antes de ser diseñados teniendo en cuenta un posible rango de variación.
Y esta ubicación debe ser flexible para no perder valiosos nucleótidos de la secuencia de interés.
Tamaño del cebador: Influye en el tiempo y temperatura necesarios para la hibridación y en la especificidad (A
mayor tamaño más especificidad ya que eso disminuye el número de lugares posibles de unión).
Temperatura de fusión (Tm): Es una medida de la estabilidad de las secuencias de doble cadena. Es definida como
la temperatura en la cual 50% de los complejos cebador-molde encuentran formados y 50% aún permanecen
disociados o sin formarse. Un alto contenido de G/C incrementa la temperatura de fusión ya que la unión entre estas
bases complementarias tiene 3 puentes de hidrógeno, a diferencia de A/T que solo tiene 2.
Temperatura de hibridación (Th): Es la temperatura utilizada para que los cebadores se hibriden, generalmente es
4° C menos que la temperatura de fusión y varía directamente proporcional a esta. Ambos cebadores deben tener Th
similares porque si no el cebador con Th más alto hibridará inespecíficamente a bajas temperatura mientras que el de
Th más baja puede que no hibride a temperaturas altas.
Evitar homodímeros: Es importante que un cebador no tenga complementariedad entre más de 3 de los nucleótidos
que lo conforman. Ya que las regiones de auto-homología pueden formar estructuras parcialmente de dos cadenas que
puedan interferir con la hibridación al molde. También se recomienda que la energía libre de Gibbs sea mayor de -8.0
kcal/mol de preferencia positivo; de esto depende la cantidad de G/C y A/T (A mayor complementariedad de G/C se
tiene menor energía libre de Gibbs) (Fig. 7).
Evitar heterodímeros: Un heterodímero es la homología entre los dos cebadores. Una homología parcial en las regiones
medias de los cebadores puede que interfiera con la hibridación, en caso la homología ocurra en los extremos 3’ de cada
cebador se dará la formación de dímeros que impedirán la formación del producto por competición.
Contenido en G/C: La composición de bases del cebador en guanina y citosina debe ser de entre el 50% y 60% y se
debe evitar que el cebador contenga zonas poli-G o poli-C que puedan llevar a hibridación inespecífica. También se
debe evitar las zonas ricas en poli-A y poli-T ya que es por dónde puede des-hibridarse el complejo molde-cebador
(Somma M, Querci M 2007).
Secuencia 3’ terminal: La posición terminal 3’ confiere una unión firme entre el cebador y la secuencia molde. Se
debe considerar la existencia de dos residuos de G o C en las últimas bases a modo de grapa (por ejemplo GG, CC,
GCO CG) para que la polimerasa actúe ahí (Figura 6). Se conoce también que los cebadores con secuencias inestables
(Mucha concentración de A/T o ΔG mayor de -8 kcal/mol) suelen ser más específicos ya que la tasa de apareamiento
en lugares incorrectos es menor. (Fig. 8)
Estructuras secundarias del ADN molde: Puede que el cebador se vea obstaculizado si la secuencia blanco tiene
afinidad entre de bases. Se debe evitar que el cebador deba unirse a zonas con apareamiento de base del molde. Para eso
debemos saber que zonas del ADN molde tienen afinidad entre sí (Fig. 9).

Página 39 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Figura 7. Esquema de complementariedad de
bases de un cebador de 20 nucleótidos con ΔG
de 0.22 kcal/mol. (Tomado de The RNA
Institute: College of Arts and Science:
http://mfold.rna.albany.edu/ y modificado por el
autor del artículo de revisión).
Figura 8. Esquema simple de la unión de cebadores
a la secuencia molde, se puede apreciar que en los
extremos hay al menos una guanina a modo de grapa.
(Tomado de Davidson:
http://www.davidson.edu/academics/biology y
modificado por el autor del artículo de revisión).
Figura 9. Presencia de estructuras secundarias en
el ADN molde. (Tomado de The RNA Institute:
College of Arts and Science:
http://mfold.rna.albany.edu/ y modificada por el
autor del artículo de revisión).
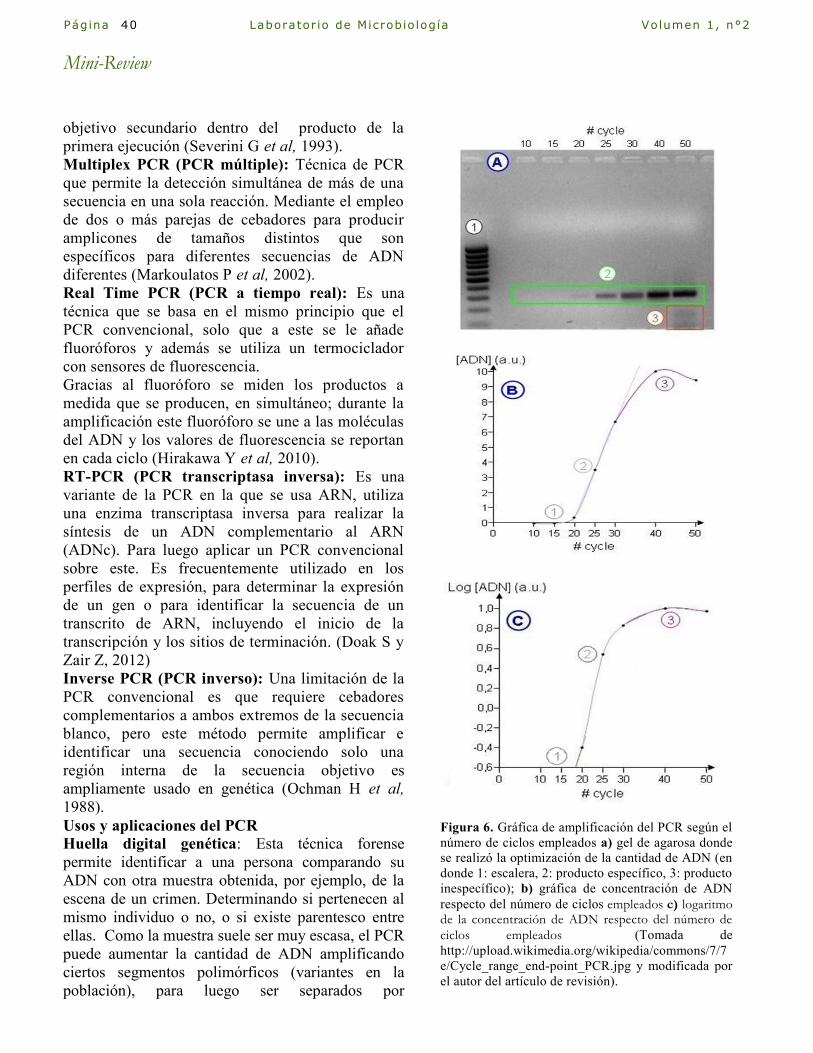
Página 40 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
objetivo secundario dentro del producto de la
primera ejecución (Severini G et al, 1993).
Multiplex PCR (PCR múltiple): Técnica de PCR
que permite la detección simultánea de más de una
secuencia en una sola reacción. Mediante el empleo
de dos o más parejas de cebadores para producir
amplicones de tamaños distintos que son
específicos para diferentes secuencias de ADN
diferentes (Markoulatos P et al, 2002).
Real Time PCR (PCR a tiempo real): Es una
técnica que se basa en el mismo principio que el
PCR convencional, solo que a este se le añade
fluoróforos y además se utiliza un termociclador
con sensores de fluorescencia.
Gracias al fluoróforo se miden los productos a
medida que se producen, en simultáneo; durante la
amplificación este fluoróforo se une a las moléculas
del ADN y los valores de fluorescencia se reportan
en cada ciclo (Hirakawa Y et al, 2010).
RT-PCR (PCR transcriptasa inversa): Es una
variante de la PCR en la que se usa ARN, utiliza
una enzima transcriptasa inversa para realizar la
síntesis de un ADN complementario al ARN
(ADNc). Para luego aplicar un PCR convencional
sobre este. Es frecuentemente utilizado en los
perfiles de expresión, para determinar la expresión
de un gen o para identificar la secuencia de un
transcrito de ARN, incluyendo el inicio de la
transcripción y los sitios de terminación. (Doak S y
Zair Z, 2012)
Inverse PCR (PCR inverso): Una limitación de la
PCR convencional es que requiere cebadores
complementarios a ambos extremos de la secuencia
blanco, pero este método permite amplificar e
identificar una secuencia conociendo solo una
región interna de la secuencia objetivo es
ampliamente usado en genética (Ochman H et al,
1988).
Usos y aplicaciones del PCR
Huella digital genética: Esta técnica forense
permite identificar a una persona comparando su
ADN con otra muestra obtenida, por ejemplo, de la
escena de un crimen. Determinando si pertenecen al
mismo individuo o no, o si existe parentesco entre
ellas. Como la muestra suele ser muy escasa, el PCR
puede aumentar la cantidad de ADN amplificando
ciertos segmentos polimórficos (variantes en la
población), para luego ser separados por
Figura 6. Gráfica de amplificación del PCR según el
número de ciclos empleados a) gel de agarosa donde
se realizó la optimización de la cantidad de ADN (en
donde 1: escalera, 2: producto específico, 3: producto
inespecífico); b) gráfica de concentración de ADN
respecto del número de ciclos empleados c) logaritmo de la concentración de ADN respecto del número de
ciclos empleados (Tomada de
http://upload.wikimedia.org/wikipedia/commons/7/7
e/Cycle_range_end-point_PCR.jpg y modificada por
el autor del artículo de revisión).

Página 41 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
electroforesis lo que se conoce como ADN
fingerprint o huella digital.
Test de paternidad: En esta técnica se amplifica
por PCR fragmentos polimórficos de ADN de la
madre, del niño y de él o los presuntos padres y,
separándolos mediante electroforesis, se puede
visualizar una serie de fragmentos que tiene el
niño, los cuales deben compartir con la madre y
padre biológicos.
Detección de enfermedades Cada gen en estudio
puede ser amplificado por PCR, con los cebadores
adecuados, y luego ser secuenciados; para así
determinar si un individuo porta alguna mutación
que explique la presencia de una enfermedad o su
aparición en el futuro, la que puede ser heredada a
sus progenitores y así se maneje su tratamiento.
Secuenciación y clonación de genes: La PCR es
habitualmente usada para amplificar un
determinado gen o un ARNm (representado por un
ADNc), que puede introducirse en un vector y
luego en un organismo, que al duplicarse también
duplicará el gen que nos interesa. Así podemos
tener una cantidad suficiente de un gen o ADNc,
por ejemplo, para secuenciarlo o incluso para
producir a gran escala la proteína que este gen
codifica. ;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;
Análisis de ADN antiguo: Con el PCR se puede
analizar ADN que tiene miles de años de
antigüedad (siempre y cuando esté en buen estado),
lo que ha permitido estudiar muestras obtenidas
desde momias hasta animales y vegetales ahora
extintos
Conclusiones El conocimiento de los diversos mecanismos
moleculares implicados en la replicación ha
permitido desarrollar la técnica del PCR y
reemplazar el uso de enzimas por variables
controladas como la temperatura en aparatos
automatizados. Además el PCR es altamente
específico y debe esto principalmente a los
cebadores y su diseño, haciendo del PCR una
técnica ampliamente usada en el mundo. A partir
de un PCR convencional, que consta de tres pasos
por ciclo (desnaturalización, hibridación y
extensión), se han desarrollado distintas variantes
de acuerdo al campo y/o finalidad de su uso. Por lo
que el uso del PCR se ha extendido a campos desde
la biología evolutiva hasta la medicina.
Literatura citada
Arnheim N y Erlich H. 1992. Polymerase Chain
Reaction Strategy. Annual review of biochemistry.
61: 131-56.
Bartlett J y Stirling D. 2003. A short history of the
polymerase chain reaction. Methods in Molecular
Biology. 226: 3-6.
Doak S, Zaïr Z. 2012 Real-time reverse-
transcription polymerase chain reaction: technical
considerations for gene expression analysis.
Methods in Molecular Biology. 817: 251-70.
Erlich H, Gelfand D, Sninsky J. 1991. Recent
Advances in the Polymerase Chain Reaction.
Science. 252 (5013): 1643-51.
Garibyan L y Avashia N. 2013. Polymerase chain
reaction. Nature. 133 (3): 1-4.
Haras D y Amoros J. 2004. Polymerase chain
reaction, cold probes and clinical diagnosis. Sante. 4
(1):43-52.
Hirakawa Y, Medh R, Metzenberg S. 2010.
Quantitative polymerase chain reaction analysis by
deconvolution of internal standard. BMC
Molecular Biology. 11:30.
Karp G. 2011. Biología celular y molecular. 6a ed.
México, D.F.: McGraw-Hill
Kleppe K, Ohtsuka E, Kleppe R, Molineux I,
Khorana H. 1971. Studies on polynucleotides.
XCVI. Repair replications of short synthetic
DNA’s as catalyzed by DNA polymerases. Journal
of Molecular Biology. 56: 341-61.

Página 42 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Watson J y Crick F. 1953. Genetical implications
of the structure of deoxyribonucleic acid. Nature
171:964-7.
Markoulatos P, Siafakas N, Moncany M. 2002.
Multiplex polymerase chain reaction: a practical
approach. Journal of Clinical Laboratory Analysis:
16(1): 47-51.
Ochman H, Gerber A, Hartl D. 1998. Genetic
applications of an inverse polymerase chain
reaction. Genetics. 120: 621–623.
Rahman M, Uddin M, Sultana R, Moue A, Setu,
M. 2013. PCR: A short review. Anwer Khan
Modern Medical College Journal 4(1): 30-36.
Ramos W, Venegas D, Medina J, Guerrero P, Cruz
A. 2013. Análisis de la situación del cáncer en el
Perú 2013.[citado 23 Nov 2014]. Disponible en:
http://www.dge.gob.pe/portal/docs/asis_cancer.pdf
Saiki R, Gelfand D, Stoffel S, Scharf S, Higuchi R,
Horn G. 1998. Primer-directed enzymatic
amplification of DNA with a thermostable DNA
polymerase. Science. 239(4839): 487-91.
Severini G, Mestroni L, Falaschi A, Camerini,
Giacca M. 1993. Nested polymerase chain reaction
for high-sensitivity detection of enterovirus RNA in
biological samples. Journal of Clinical
Microbiology. 31(5): 1345.
Somma M, Querci M. 2007 Análisis de la presencia
de organismos genéticamente modificados en
muestras de alimentos. [citado 23 Nov 2014].
Disponible en: http://gmo-crl.jrc.ec.europa.eu/
capacitybuilding/manuals/Manual%20ES/Sesi%
C3%B3n6.pdf
Watson J, Baker T, Bell S, Gann A, Levine M,
Losick R. 2006. Biología molecular del gen. 5a ed.
Madrid: Médica
Panamericana.

R E S U M E N
Los estudios en biotecnología han demostrado que se puede utilizar el
mecanismo de infección de microorganismos fitopatógenos para que las
plantas expresen moléculas cuya producción actual es ineficiente y de alto
costo. Uno de estos organismos utilizados en la ingeniería genética de plantas
es Agrobacterium tumefaciens que tiene la capacidad de transferir una
secuencia del plásmido Ti al genoma vegetal. Esta secuencia se denomina
TDNA y el proceso es regulado, principalmente, por el operón vir del pTi, el
cual está conformado por ocho genes vir (A-H). La transferencia comienza
con la activación de la proteína receptora virA, por interacción con
compuestos fenólicos secretados por la planta; a partir de ello, se fosforila la
proteína VirG, y actúa como factor de transcripción para las demás proteínas
Vir. Las proteínas que se encargan de transportar el TDNA y mantener su
integridad son VirD1 y VirD2, VirB, VirE y VirF. Esta propiedad, ha dado
paso a múltiples investigaciones en donde se reemplazan los genes del TDNA
por un gen de interés y se acompaña de otro vector de A. tumefaciens que no
posee una región de TDNA pero sí el operón vir permitiendo la transferencia
del TDNA modificado, a este sistema se le denomina vector binario. En la
actualidad mediante la aplicación de esta metodología se pueden obtener
plantas transgénicas lo cual permite mejorar la productividad de plantas de
importancia económica u obtener sustancias proteicas que son indispensables
en otras áreas de la ciencia.
Las plantas, fuente de vida
Desde los inicios de la humanidad, las plantas han
sido los principales organismos pluricelulares que
se han usado como alimento, medicina, e inclusive
como ornamento. La distribución de muchas
especies puede llegar a ser global dependiendo del
uso al que esté destinado. Es así que, a medida que
se desarrollaban nuevas técnicas para cultivar y
conservar diferentes tipos de plantas, se adaptaban
otros organismos que, al igual que los humanos,
necesitaban de la planta para sobrevivir, causando
pérdidas en la productividad de cultivos, como el
virus de la papa que afecta al 90% de las plantas
cultivadas (Robles L et al, 2010). Esto inició los
estudios guiados a la fitopatología, área que se ha
desarrollado desde 1729, cuando el botánico
Michelli observó hongos en plantas de melocotón,
dando paso a la determinación de hongos, bacterias
y virus, que producen enfermedades en las plantas
(Agrios G, 2013).
Integración de la Biotecnología Bacteriana
y la Ingeniería Genética de Plantas Morales, M.
Correo electrónico: [email protected]
Página 43 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
INFORMACIÓN DEL
ARTÍCULO:
Morales M. Integración de
la Biotecnología Bacteriana
y la Ingeniería Genética de
Plantas. Boletín Científico.
2014. 1(2): 42-51.
Palabras Claves:
Agrobacterium tumefaciens,
TDNA, operón vir

De las bacterias a las plantas
Una de estas enfermedades es la “Corona de
Agallas”, denominada así por las agallas de aspecto
tumoroso que aparecen en la parte basal del eje
principal aéreo de la planta, generalmente en la
base del caule, y fue descrita por primera vez por
Fabre F y Dunal F en 1853. Sin embargo, fue en
1997 cuando Chilton M et al, reconocieron
fragmentos de ADN en los tumores axénicos de
plantas de tabaco, procedentes de un bacilo
gramnegativo: Agrobacterium tumefaciens.
Posteriormente, se determinó que estos fragmentos
de ADN contenían genes que inducían la formación
de tumores en las células vegetales, ya que eran
introducidos mediante un mecanismo de
transferencia al genoma de las plantas. Es así que la
investigación causó gran impacto en el área de la
genética de plantas, concibiendo la idea de que una
planta puede expresar un gen que no se encuentre
normalmente en su genoma. A partir de ello se
comenzó a investigar este mecanismo con el fin de
realizar un mejoramiento genético, o inclusive
expresar moléculas difíciles de obtener en otros
sistemas (Valderrama A et al, 2005).
Transferencia de ADN (T-DNA) en la célula
vegetal
El desarrollo de la enfermedad está ligado a la
interacción entre el patógeno, el ambiente y el
hospedero. El patógeno coloniza las heridas e infecta
la planta mediante mecanismos moleculares que
involucran una serie de interacciones genéticas que
contribuyen a que la bacteria sea capaz de adherirse
a la planta y causar la tumoración y transferencia de
ADN; estos procesos se pueden sintetizar en cuatro
pasos fundamentales: la colonización bacteriana; la
inducción del sistema de virulencia; la generación
del complejo de transferencia (T-DNA); y la
integración del T-DNA en el genoma de la planta.
En este proceso resalta la participación de un
plásmido denominado Ti (Tumoral inductor –
“inductor del tumor”) (Fig. 2). (De la Riva G et al,
1998).
El proceso colonización bacteriana incluye la
adherencia del patógeno a la raíz o tallo de la planta;
esto se da debido a que Agrobacterium reconoce
células vegetales que se encuentran dañadas por
factores físicos, químicos o biológicos; ya que dichas
células vegetales secretan sustancias fenólicas de
bajo peso molecular como acetosiringona e
hidroxiacetosiringona que son reconocidos por la
bacteria; por otro lado, las células vegetales dañadas
tiene la característica de no presentar una pared
celular estable lo que conlleva a la alta producción
de lignina y flavonoides que permiten dirigir a la
bacteria hacia las células susceptibles;
posteriormente, la bacteria se une a la célula
mediante la acción de una molécula en la pared
celular sensible a proteasas que es reconocida por la
bacteria; estas moléculas pueden ser dos proteínas de
adhesión en donde se encuentra la vitonectrina y la
ricoadhesina, sin embargo este proceso no ha sido
descrito genéticamente (Llop P, 2003); no obstante,
se han identificado genes que son necesarios para
Figura 1. “Corona de Agallas” causada por A.
tumefaciens (Escobar M y Dadenlar A, 2003).
Página 44 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
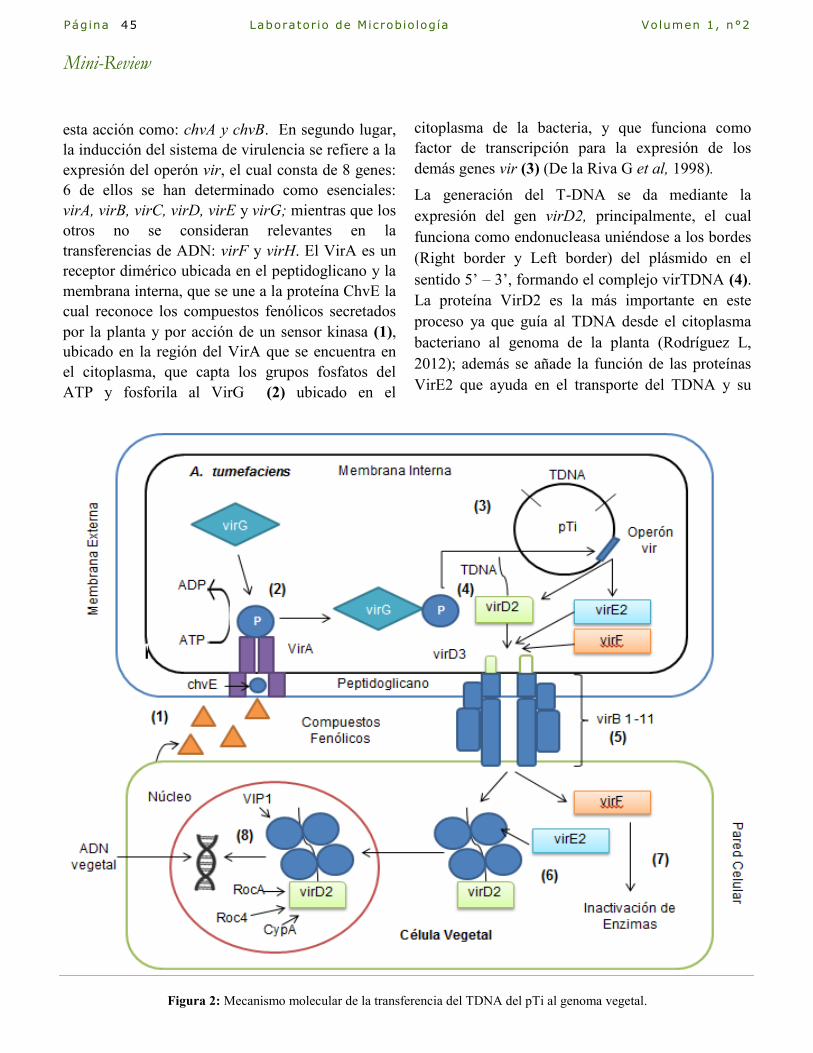
citoplasma de la bacteria, y que funciona como
factor de transcripción para la expresión de los
demás genes vir (3) (De la Riva G et al, 1998).
La generación del T-DNA se da mediante la
expresión del gen virD2, principalmente, el cual
funciona como endonucleasa uniéndose a los bordes
(Right border y Left border) del plásmido en el
sentido 5’ – 3’, formando el complejo virTDNA (4).
La proteína VirD2 es la más importante en este
proceso ya que guía al TDNA desde el citoplasma
bacteriano al genoma de la planta (Rodríguez L,
2012); además se añade la función de las proteínas
VirE2 que ayuda en el transporte del TDNA y su
esta acción como: chvA y chvB. En segundo lugar,
la inducción del sistema de virulencia se refiere a la
expresión del operón vir, el cual consta de 8 genes:
6 de ellos se han determinado como esenciales:
virA, virB, virC, virD, virE y virG; mientras que los
otros no se consideran relevantes en la
transferencias de ADN: virF y virH. El VirA es un
receptor dimérico ubicada en el peptidoglicano y la
membrana interna, que se une a la proteína ChvE la
cual reconoce los compuestos fenólicos secretados
por la planta y por acción de un sensor kinasa (1),
ubicado en la región del VirA que se encuentra en
el citoplasma, que capta los grupos fosfatos del
ATP y fosforila al VirG (2) ubicado en el
Página 45 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Figura 2: Mecanismo molecular de la transferencia del TDNA del pTi al genoma vegetal.

integración (6) (Wroblewski T et al, 2005). Además
actúa la proteína VirF cuya función no está
completamente determinada, pero se cree que actúa
inactivando las enzimas que puedan degradar el
TDNA mediante proteólisis (7) (Schrammeijer B et
al, 2001). Este complejo ingresa en el citoplasma de
la célula hospedera mediante la acción de las
proteínas VirB1-11 y VirD4 los cuales unen la
membrana celular de la bacteria con el citoplasma
de la célula vegetal (5) formando un puente similar a
un proceso de conjugación (McCullen C y Binns A,
2006) (Fig. 2).
Finalmente, el complejo virTDNA es internalizado
en el núcleo de la célula vegetal por la proteína
VirD2 el cual contiene dominios de recombinación
y ligación (Bako L et al, 2003); y con la
participación de histonas como la H2A (Tzfira T y
Citovsky V, 2002); además, existen proteínas de la
planta, como Vip1, que reconocen al virE2 para
permitir la integración del TDNA; por otro lado,
también, es reconocido por las chaperonas de la
planta: RocA, Roc4 y CypA (8) que mantienen la
inserción del TDNA en el genoma de la planta
(Valderrama
A et al, 2005).
La transformación: Base de la Ingeniería
Genética
A partir del mecanismo anterior. La ingeniería
genética ha encontrado un beneficio propio en la
obtención de plantas transgénicas. Es así que se
utiliza un sistema de vector binario, el plásmido Ti
se ha “domesticado” inhibiendo la síntesis de genes
de inducción de tumor y desarrollo de la
enfermedad y manteniendo los genes de
transferencia de ADN: el operón vir (Gutarra B,
2004) (Fig. 3), este plásmido se utiliza con otro
vector que posee el gen de interés con el fin de
obtener varias copias del plásmido, y subclonado
en un plásmido de expresión que posee un
promotor, residuos que facilitan la purificación (Ej.
His-6) y un terminador (Shoji Y et al, 2008).
Además, para facilitar la identificación de una
transferencia exitosa, el vector de expresión
presenta un gen que da resistencia a herbicidas, así
se puede tratar a las plantas con dichas sustancias y
si no resultan afectadas entonces la transferencia
del gen es satisfactoria (Cubero J, 2013). Existen,
actualmente, vectores comerciales con sitios
múltiples de clonación (MCS), como pT7-Blue y
pBluescriptII, en los que se ha insertado el gen del
Página 46 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Figura 3: Sistema de Vector Binario. (izq.) plásmido Ti “desarmado”. (der.) Vector con el gen de interés. Tomado de:
http://concienciadeplantas.blogspot.com/2013/03/tecnicas-que-empleamos-parte-1.html

transgénicos que poseen resistencia a herbicidas y
plagas, producción de colores, y longevidad y sus
usos van desde cultivos, alimentación,
procesamiento, ornamentación e importaciones
(Cuadro 1), además, es posible la venta de semillas
de esta plantas transgénicas (Sebiot, 2000).
En nuestro país, el uso de los transgénicos para las
actividades mencionadas anteriormente no está
permitido debido a la proclamación de la Ley N°
29811 que impide el ingreso, producción y
liberación de OGM en el territorio nacional por un
periodo de 10 años, y simultáneamente, se debe
equipar, adaptar y desarrollar una infraestructura
necesaria para la evaluación de estos organismos y
su aceptación en el país (MINAM, 2012). Para ello,
es necesaria la investigación en las áreas de la
biotecnología bacteriana y vegetal con el fin de
comprobar la viabilidad y uso de los organismos
genéticamente modificados y transformados por
acción de Agrobacterium tumefaciens.
Conclusiones
Es así que a partir de una patología que parecía
tener la capacidad de causar un grave impacto en la
economía de la agricultura y más aún en la
IFNγ controlado por promotores de αAmy y
Ubiquitina, el cual fue subclonado en otro plámsido
comercial de A. tumefaciens: pCAMBIA1390, el
cual está diseñado con el marcador de resistencia a
higromicina; es decir que aquellas plantas que hayan
sido transformadas deberán ser resistente a este
antibiótico y expresar el IFNγ (Chen T et al, 2004).
Aplicaciones de la Transformación de Plantas
mediado por Agrobacterium tumefaciens
La ingeniería genética en microbiología y en
botánica ha permitido la obtención de organismos
genéticamente modificados (OGM), los cuales
pueden expresar características deseables, tales
como colores, resistencia a factores abióticos como
temperatura, potencial hídrico, e inclusive
pesticidas; y bióticos, como plagas principalmente;
además se está evaluando la posibilidad de expresar
proteínas cuya producción es baja y poco efectiva y
que tienen una importancia en el área de salud,
generalmente, tales como las moléculas del sistema
inmunológico o antígenos para la producción de
proteínas recombinantes (NSTA, 2007). Sin
embargo, en la actualidad, Europa es el principal
continente donde se ha permitido el uso de algunos
Extraído de : “Plantas Transgénicas: Preguntas y Respuestas (Sebiot, 2000)
Tabla 1. Plantas Transgénicas permitidas en Europa.
Página 47 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
Organismo Característica Usos
Maíz Bt-176 Resistente a Pyrausta nubilalis“Taladro” Todos los usos
Maíz MON-810 Resistente a Pyrausta nubilalis“Taladro” Todos los usos
Maíz T25 Resistente a glufosinato de amonio (herbicida) Todos los usos
Maíz Bt-11 Resistente al “taladro” y al glufosinato de amonio Importación y
procesamiento
Tabaco Resistente a bromoxinil (herbicida) Cultivo
Soya A-5403 Resistente al glifosato (herbicida) Importación y
procesamiento
Achicoria Tolerante al glufosinato de amonio (herbicida) Cultivo
Claveles Nuevos colores Ornamentación
Claveles Mayor Longevidad Ornamentación

alimentación del hombre, resultó ser una
herramienta para el mejoramiento genético que
actualmente viene solucionando gran cantidad de
problemas de la humanidad. La interacción genética
del plásmido Ti, permite la adherencia de la bacteria
a la planta (chvA, chvB); la inducción del sistema de
virulencia a partir de los genes virA y virG; la
síntesis del complejo virTDNA mediante la
expresión de los genes virB, virC, virD, virE, virF,
virH; y la integración del TDNA en el genoma de la
planta, donde el principal protagonista es el gen
virD2, ya que posee dominios de ligamiento y
recombinación. Este proceso, es aprovechado para
la obtención de transgénicos lo cual permite mejorar
la productividad de plantas de importancia
económica u obtener sustancias proteicas que son
indispensables en otras áreas de ciencia.
Literatura citada
Agrios G. 2013. Fitopatología. 2 ed. Massachusets:
Limusa: p. 800 - 805.
Bako L, Umeda M, Tiburcio A, Schell J, Koncz C.
2003. The VirD2 pilot protein of Agrobacterium –
transferred DNA interacts with the TATA box –
binding protein and a nuclear protein kinase in
plants. PNAS. 100(17): 10108-10113
Chen T, Lin Y, Lee Y, Yang N, Chan M. 2004.
Expression of bioactive human interferon-gamma in
transgenic rice cell suspension cultures. Transgenic
Research. 13: 499 – 510.
Chilton M, Drummond M, Merio D, Sciaky D,
Montoya A, Gordon M, Nester E. 1977. Stable
incorporation of plasmid DNA into higher plant
cells: the molecular basis of crown gall
tumorigenesis. Cell. 11(2): 263-271.
Cubero J. 2013. Introducción a la Mejora Genética
Vegetal. 3 ed. Córdoba: Mundi-Prensa: 375 – 380.
De la Riva G, Gonzáles J, Vásquez R, Ayra C.
1998. Agrobacterium tumefaciens: a natural tool for
plants transformation. Electronic Journal of
Biotechnology. 1(3): 118–133.
Escobar M y Dandelar A. 2003. Agrobacterium
tumefaciens as an agent of disease. Trends in Plant
Science. 8(8): 380-386.
Fabre E y Dunal F. 1853. Observations sur les
maladies regnantes de la vigne. Bulletin de la
Société Centrale d´Agriculture du Département de
l´Hérault. 40:46.
Gutarra B. 2004. Construcción de dos vectores
conteniendo un gen inhibidor de tripsina unido al
promotor de la β-amilasa y agro-transformación de
camote (Ipomoea batatas Lam.). Tesis para obtener
el Título de Biólogo con mención en Genética.
Universidad Nacional Mayor de San Marcos. Lima
— Perú.
Llop P. 2003. Caracterización molecular de la
pérdida del poder patógeno en Agrobacterium
tumefaciens. Tesis para obtener el Título de Doctor.
Universidad de Valencia — España.
McCullen C y Binns A. 2006. Agrobacterium
tumefaciens and Plant Cell interactions and
activities required for Interkingdom
Macromolecular Transfer. Annual Review of Cell
and Developmental Biology. 22: 101-127.
MINAM. 2012. Reglamento de la Ley Nº 29811,
Ley que establece la Moratoria al Ingreso y
Producción de Organismos Vivos Modificados al
Territorio Nacional por un Período de 10 años.
Ministerio del Ambiente, Perú: 1– 19.
Página 48 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review

Página 49 Laborator io de Microbio logía Volumen 1, n°2
Mini-Review
NSTA. 2007. Genetically Modified Crops:
Resources for Environmental Literacy. Ed. NSTA
press. 5 – 36.
Robles L, Gonzáles A, Gill E, Pérez L, López J.
2010. Virus fitopatógenos que afectan al cultivo de
chile en México y análisis de las técnicas de
detección. Revista de Tecnociencia 4(2): 72– 86.
Rodríguez L. 2012. Acercamiento al estudio del
comportamiento biológico de Agrobacterium
tumefaciens. Universidad Nacional Abierta y a
Distancia. Bogotá—Colombia.
Schrammeijer B, Risseeuw E, Pansegrau W,
Regensburg-Tuink T, Crosby W, Hooykaas P.
2001. Interaction of the virulence protein VirF of
Agrobacterium tumefaciens with plant homologs of
the yeast Skp1 protein. Current Biology. 11(4): 258
– 261.
Sebiot. 2000. Plantas Transgénicas: Preguntas y
Respuestas. Sociedad Española de Biotecnología. 1:
4 – 47.
Shoji Y, Chichester J, Bi H, Musiychuk K, De la
Rosa P, Goldschmidt L, Horsey A, Ugulava N,
Palmer G, Mett V, Yusibov V. 2008. Plant –
expressed HA as a seasonal influenza vaccine
candidate. Vaccine 26: 2930 – 2934.
Tzfira T y Citovsky V. 2002. Partners – in –
infection: host proteins involved in the
transformation of plant cells by Agrobacterium.
Trends in Cell Biology. 12(3): 121–129.
Valderrama A, Arango R, Afanador L. 2005.
Transformación de plantas mediada por
Agrobacterium: “Ingeniería Genética Natural
Aplicada”. Revista Facultad Nacional de
Agronomía. 58(1): 2569 – 2585.
Wroblewski T, Tomczak A, Michelmore R. 2005.
Optimization of Agrobacterium-mediated transient
assays of gene expression in lettuce, tomato and
Arabidopsis. Plant Biotechnology Journal. 3: 1 – 13.

Dirección: Facultad de Ciencias Biológicas Av. Benavides 5440, Santiago de Surco, Lima 33—PERU
Contacto:
Laboratorio de Microbiología
999227470





























