Embed Size (px)
Citation preview

Cartel de la Reunión

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
2
Instituciones Patrocinadoras
La VIII Reunión de la Sociedad Mexicana de Astrobiología (SOMA) se ha realizado gracias al apoyo institucional y financiero de las siguientes corporaciones:

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
3
Comité Organizador
Comité organizador (SOMA)
Dra. Sandra Ignacia Ramírez Jiménez, Centro de Investigaciones Químicas (CIQ), UAEM Dra. Antígona Segura Peralta, Instituto de Ciencias Nucleares (ICN), UNAM M. en C. Irma Lozada Chávez, Centro Interdisciplinario de Bioinformática (IZBI), Alemania Dra. María Guadalupe Cordero Tercero, Instituto de Geofísica (IGF), UNAM
Comité local Dra. Sandra I. Ramírez Jiménez, CIQ, UAEM Dr. Rodrigo Morales Cueto, CIQ, UAEM Dr. Thomas Buhse, CIQ, UAEM
Comité científico Dr. Roberto Vázquez Meza, Instituto de Astronomía, UNAM (Campus Ensenada) Dr. Thomas Buhse, CIQ, UAEM Dra. María Colín García, Instituto de Geología, UNAM
Página web Dr. Víctor de la Luz, Instituto Nacional de Astrofísica, Óptica y Electrónica (INAOE) Mat. Enrique Palacios Boneta, ICN, UNAM
Diseño gráfico Ana Ruiz, IGF, UNAM
Fotografía M. en E. Miguel Ángel Reza Urueta, Facultad de Arquitectura, UAEM
Maestra de Ceremonias Lic. Susana Ballesteros Carpintero, UFM Alterna, Radio UAEM
Apoyo logístico Cristina Bojórquez Espinosa Lilia Montoya Lorenzana Eduardo Piña Mendoza Dení Zenteno Gómez Rocío Elizabeth Avendaño Serrano Jonathan Valdez Camacho Sandra Hidalgo Neri Gabriel Iván Martínez Solís Yuritzi Judith Quiroz Cortés Roselena Arroyo Basave Ricardo González Lizardi
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
4
Estadísticas de la asistencia a la Reunión
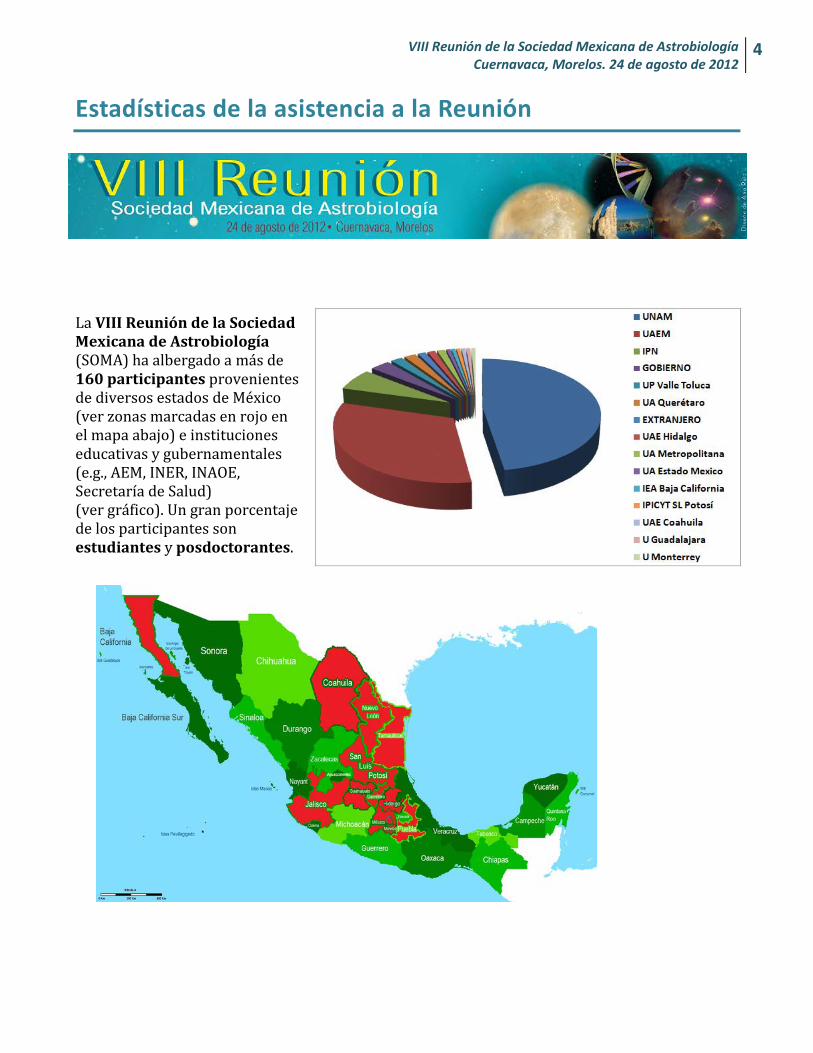
La VIII Reunión de la Sociedad Mexicana de Astrobiología (SOMA) ha albergado a más de 160 participantes provenientes de diversos estados de México (ver zonas marcadas en rojo en el mapa abajo) e instituciones educativas y gubernamentales (e.g., AEM, INER, INAOE, Secretaría de Salud) (ver gráfico). Un gran porcentaje de los participantes son estudiantes y posdoctorantes.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
5
Programa de la Reunión
Horario Título de la Plática Conferencista Institución
09:30 - 10:10 Entrega de paquete de participación: Auditorio "Emiliano Zapata" de la UAEM
10:10 - 10:30 Inauguración y palabras de bienvenida
Presídium: Dr. Mario Fernández Zertuche, Director del CIQ Biol. Juan Carlos Sandoval Manrique, Director de la FCB Dr. Gustavo Urquiza, Secretario de Investigación e Innovación UAEM Dra. Leticia Carigi, Coordinadora del Posgrado en Astrofísica, UNAM Dra. Antígona Segura Peralta, Presidente de SOMA Dra. Sandra I. Ramírez Jiménez, Representante del CA Química y Física del Ambiente y Coordinadora del Comité Organizador
10:30 - 10:45 C1. Parámetros del sistema exoplanetario que transita HAT-P-23b
Dr. Pedro Valdés-Sada Universidad de Monterrey
10:45 - 11:00 C2. Clasificación planetaria obtenida a partir de leyes de potencia empíricas
Dr. Héctor Durand-Manterola
Instituto de Geofísica, UNAM
11:00 - 12:00 P1. ¿Qué es Cuatro Ciénegas y por qué se parece a Marte?
Dra. Valeria Souza Saldívar Instituto de Ecología, UNAM
12:00 - 12:30 DESCANSO PARA CAFÉ
12:30 - 12:45 C3. Origen de la nube progenitora del Sistema Solar
Dra. Leticia Carigi Delgado Instituto de Astronomía, UNAM
12:45 - 13:00 C4. Generación experimental de fundidos tipo condros para determinar los mecanismos de su formación
Dra. Antígona Segura Peralta Instituto de Ciencias Nucleares, UNAM
13:00 - 13:15 C5. El corazón en el ámbito espacial Dr. Ramiro Iglesias-Leal Secretaría de Salud de Tamaulipas
13:15 - 13:30 C6. Perspectivas de la investigación científica en la Agencia Espacial Mexicana
Dr. Celso Gutiérrez Agencia Espacial Mexicana
13:30 - 15:00 COMIDA: Jardín del Centro de Investigaciones Químicas
15:00 - 15:15 C7. La halotolerancia en el contexto de la Astrobiología
Dra. Sandra I. Ramírez-Jiménez
Centro de Investigaciones Químicas, UAEM
15:15 - 15:30 C8. El registro geológico del Hadeano y la emergencia posible de la primera vida en el sistema solar
Dr. Fernando Ortega-Gutiérrez
Instituto de Geología, UNAM
15:30 - 16:30 P2. Ciclos autocatalíticos, corrección de pruebas y el origen de la homoquiralidad
Dr. Thomas Buhse Centro de Investigaciones Químicas, UAEM
16:30 - 17:00 DESCANSO PARA CAFÉ
17:00 - 17:15 C9. Roll-ups: estructuras sedimentarias inducidas por microbios y la búsqueda de vida en Marte
Dr. Hugo Beraldi-Campesi Instituto de Geología, UNAM
17:15 - 17:30 C10. ¿Cuál es el papel de la selección natural en el origen de nuevas funciones proteicas? Estudio teórico de la evolución molecular de proteínas
LAQB. Zurisadai Miguel Muñoz-González
CINVESTAV, Irapuato
17:30 - 18:30 SESIÓN DE CARTELES: Vestíbulo del Centro de Investigaciones Químicas

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
6
Índice de resúmenes (vínculos activos)
CCOONNFFEERREENNCCIIAASS PPLLEENNAARRIIAASS
Cuatro Ciénegas, una máquina del tiempo que nos puede llevar al pasado del planeta Tierra y ayudarnos a entender si hay o hubo vida en Marte.
Valeria Souza Saldivar, Luis E. Eguiarte Fruns y Germán Bonilla Rosso
10
Ciclos autocatalíticos, corrección de pruebas y el origen de la homoquiralidad Thomas Buhse, José-Manuel Cruz, Jean-Claude Micheau y Christophe Coudret
13
PPRREESSEENNTTAACCIIOONNEESS OORRAALLEESS
Parámetros del sistema exoplanetario que transita HAT-P-23b Pedro Valdés-Sada y Felipe G. Ramón Fox
16
Clasificación planetaria obtenida a partir de leyes de potencia empíricas Héctor Durand-Manterola
20
Origen de la nube progenitora del Sistema Solar Leticia Carigi y Manuel Peimbert
23
Generación experimental de fundidos tipo condros para determinar los mecanismos de su formación
Antígona Segura y Karina Cervantes
26
Perspectivas de la investigación científica en la Agencia Espacial Mexicana Celso Gutiérrez
29
El corazón en el ámbito espacial Ramiro Iglesias-Leal
31
La halotolerancia en el contexto de la Astrobiología Sandra I. Ramírez-Jiménez
34
El registro geológico del Hadeano y la emergencia posible de la primera vida en el Sistema Solar Fernando Ortega-Gutiérrez
36
Roll-ups: estructuras sedimentarias inducidas por microbios y la búsqueda de vida en Marte Hugo Beraldi-Campesi
38
¿Cuál es el papel de la selección natural en el origen de nuevas funciones proteicas? Estudio teórico de la evolución molecular de proteínas
Zurisadai Miguel Muñoz-González, Luis Jośe Delaye Arredondo y Alexander de Luna Fors
40
PPRREESSEENNTTAACCIIOONNEESS EENN CCAARRTTEELLEESS
AASSTTRROOBBIIOOLLOOGGÍÍAA EENN MMEEXXIICCOO
Propuesta de la Sociedad Mexicana de Astrobiología para convertirse en afiliado internacional del Instituto de Astrobiología de la NASA
Antígona Segura, Sandra Ramírez e Irma Lozada-Chávez
44
AASSTTRROOFFÍÍSSIICCAA
Estudio molecular de CO en galaxias cercanas Ilhuiyolitzin Villicaña Pedraza y J. Martin-Pintado
47
GGEEOOFFÍÍSSIICCAA
Detección de meteoros y estudio de las propiedades físicas de los meteoroides asociados María Guadalupe Cordero Tercero
49
Meteoroide del 18 de mayo del 2012: estimación del área de caída José Luis García Martínez
50

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
7
Cometas y el origen del agua en la Tierra Saúl Alberto Villafañe Barajas
54
Río Tinto y la posible preservación de moléculas orgánicas en condiciones extremas María Colín-García, Basem Kanawati, Mourad Harir, Schmitt-Kopplin, Ricardo Amils, et al.
57
Bioformas microscópicas en pirita sedimentaria y sus implicaciones astrobiológicas Susana A. Ángeles Trigueros, Alberto Blanco-Piñón, Juan Hernández-Ávila y F. Javier Zavala-Díaz
60
Avances sobre panspermia inversa Roberto Vázquez Meza, Patricia G. Núñez, Mauricio Reyes-Ruiz, Carlos E. Chávez, Stephania Hernández y Héctor Aceves
64
Biochar: una estrategia de mitigación de calentamiento global en la Tierra, para después lograr una ecopoiesis en Marte
Ramón Agustín Bacre González
67
QQUUÍÍMMIICCAA PPLLAANNEETTAARRIIAA
La importancia de la detección de litio en la superficie de Marte María Colín-García, Alejandro Heredia, Julio Valdivia Silva, Hugo Beraldi, et al.
69
Cantidad de H2O y CO2 desgasados en Marte desde su formación y sus implicaciones para la vida Alexia Nailee Medina-Amayo y Héctor J. Durand-Manterola
72
Producción abiótica de metano en planetas terrestres Andrés Guzmán Marmolejo, Antígona Segura y Elva Escobar Briones
75
Influencia de las superficies minerales en el desarrollo y estabilización de moléculas orgánicas en condiciones prebióticas
Ellen Yvette Aguilar Ovando y Alicia Negrón-Mendoza
78
Simulación de relámpagos a baja temperatura en la atmósfera de Titán Dení Tanibé Zenteno Gómez y Sandra Ignacia Ramírez Jiménez
81
BBIIOOLLOOGGÍÍAA YY EEVVOOLLUUCCIIÓÓNN
Endosimbiontes como modelos de células mínimas Luis José Delaye y David José Martínez Cano
84
Bacillus pumilus: bacteria modelo en estudios de potencial de habitabilidad Rocio Elizabeth Avendaño Serrano y Sandra I. Ramírez Jiménez
87
Ubicación de posibles ecosistemas de procariontes reductores de sulfato (PSR) en Europa, satélite de Júpiter
Guadalupe Vaneza Yazmín, Lilia Montoya-Lorenzana y Héctor Durand-Manterola
90
Microorganismos degradadores de acetato, objeto de estudio en Astrobiología Lilia Montoya, Lourdes B. Celis, Elías Razo-Flores y Ángel G. Alpuche-Solís
93
Extremófilos: una revisión Ricardo Gonzalez Lizardi, Roselena Arroyo Basave y Sandra I. Ramírez Jiménez
95
Extremófilos anaerobios en el ciclo del carbono, una revisión Lilia Montoya, Lourdes B. Celis, Elías Razo-Flores y Ángel G. Alpuche-Solís
98
Evolución molecular de los efectores secretados por Trichoderma spp. y su participacion en la comunicación con sus hospederos
Mario Iván Alemán Duarte, Luis José Delaye Arredondo y Alfredo Herrera-Estrella
101
Análisis evolutivo de los patrones de selección en las cápsides de tres virus icosaédricos: virus del mosaico del frijol del sur, virus del tomate y virus satélite de la necrosis del tabaco
Karina Espinoza Muñoz y Luis José Delaye Arredondo
104
Estandarización de una técnica de extracción de ADN microbiano en muestras de agua del lago-cráter Rincón de Parangueo, Guanajuato, México
Christian Emmanuel Robles Rivera, Mayra Alejandra Campos Hernández y Fausto Arellano Carbajal
106
MMEEDDIICCIINNAA EESSPPAACCIIAALL
Efecto de la posicion supina en la descalcificacion María Elena Aguilar Mena
110

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
8
CONFERENCIAS PLENARIAS

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
9
Conferencia Plenaria
CUATRO CIÉNEGAS, UNA MÁQUINA DEL TIEMPO QUE NOS PUEDE LLEVAR AL PASADO DEL PLANETA TIERRA Y AYUDARNOS A ENTENDER SI HAY O HUBO VIDA
EN MARTE.
Valeria Souza Saldivar, Instituto de Ecología, Universidad Nacional Autónoma de México [email protected]
Luis E. Eguiarte Fruns, Instituto de Ecología, Universidad Nacional Autónoma de México Germán Bonilla Rosso, Instituto de Ecología, Universidad Nacional Autónoma de México
Introducción
Sabemos que desde el principio, la vida estuvo formada de los átomos más abundantes del universo, aquellos elementos que fueran solubles en agua y que por lo tanto fueran abundantes en la sopa primigenia. Sin embargo, la vida también necesita fósforo (P) y el P puro es tan reactivo que cuando entra en contacto con el oxígeno del aire explota generando una fuerte luz blanca formando fosfatos, una molécula rica en energía que se volvió, por razones termodinámicas, en un elemento esencial de los bloques que construyen la vida. Sin embargo, en la tierra primitiva las rocas de apatita que guardan este elemento estabilizándolo con calcio y no estaban expuestas al oxígeno salvo en sitios muy particulares donde el agua liberaba O2 de manera local y solo en esos sitios había P soluble. Por lo tanto, la mayor parte de la historia de la vida en la tierra, el Precámbrico, se caracterizó por ser un ambiente pobre en fósforo y rico en azufre donde las comunidades microbianas formaban tapetes microbianos y en condiciones ricas en calcio, estos tapetes se volvían arrecifes llamados estromatolitos. Es por ésto que el funcionamiento fisiológico de los tapetes microbianos a gran escala ha sido estudiado desde hace más de tres décadas por los ecólogos microbianos, revelando un marcado microgradiente en las concentraciones de oxígeno, pH y ácido sulfídrico. Resultados y Discusión
En Cuatro Ciénegas los tapetes microbianos y los estromatolitos son muy abundantes y diversos, ya que en este sitio no hay suficiente fósforo (P) para que las algas puedan crecer [1]. Iniciamos el trabajo microbiológico molecular para describir el sitio y descubrimos que 50% de las bacterias eran de afiliación marina utilizando como marcador el gen 16S rDNA [2, 3], secuenciamos el genoma de varias de estas bacterias, en particular del género Bacillus y Exiguobacterium [4, 5, 6] y encontramos que esta filiación marina es consistente en todo el genoma y que los organismos secuenciados presentan múltiples adaptaciones al ambiente pobre en nutrientes. Recordemos que este valle está ahora en el centro del Norte de México, en el estado de Coahuila a 700 m sobre el nivel del mar ¿por qué hay organismos marinos en el desierto? ¿Por qué hay tantos tapetes microbianos y estromatolitos en este lugar tan particular?
Una hipótesis posible es que los tapetes microbianos que vivían en las costas Oeste de Laurentia por al menos mil millones de años, encontraron un lugar nuevo donde crecer hace 238 millones de años cuando Pangea se rompe en dos y Laurentia, junto con el hemisferio norte, migran, desde el ecuador hasta donde se encuentran ahora. El inicio de ese enorme evento ocurre precisamente en Coahuila en lo que ahora es la Sierra de la Fragua que limita por el sur al valle de Cuatro Ciénegas, por lo que los

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
10
tapetes microbianos entran a esta zona en el inicio del rompimiento de Pangea y el sedimento marino del jurásico, rico en yeso (sulfato de calcio) domina a la zona. En la actualidad, muchos de los parientes cercanos de las bacterias de Cuatro Ciénegas están ahora en sitios que eran parte del mar de Thetys, incluyendo los lagos de altura de los Andes o en los salares del Himalaya. Hace solo 35 millones de años, al levantarse las sierras se fue el mar y el valle también se levanto aislando así a los tapetes microbianos de Cuatro Ciénegas junto con sus moluscos, crustáceos, peces, tortugas, diatomeas y virus [7]. Al aislarse el valle, las comunidades se adaptaron a las aguas continentales ricas en iones y evolucionaron aisladas del resto del mundo volviéndose endémicas pero manteniendo la señal filogenética de sus ancestros marinos [8, 9].
Para poder explorar esta ecología del pasado secuenciamos 4 comunidades microbianas, 2 formadoras de tapetes microbianos (PR y PG) y 2 estromatolitos (RM y PA) para determinar la diversidad y función de la comunidad por medio de la metagenómica [10, 11]. Sin embargo, cuanto a la diversidad funcional, el conocimiento existente sobre el metabolismo bacteriano es tan sólo una parte del existente en comunidades naturales, y ésto limita nuestra capacidad de realizar análisis comparativos porque se encontrarán sesgados hacia la representación de nuestro conocimiento en las muestras. El tapete PG se desarrolla en una poza permanente fisicoquímicamente estable, y posee una mayor complejidad y diversidad taxonómica y funcional, caracterizada por la abundancia de mecanismos de producción primaria y metabolismos autotróficos aeróbicos. El tapete PR se desarrolla en una poza de desecación fisicoquímicamente variable, y se encuentra fuertemente dominada por varias especies del género Pseudomonas. Su metabolismo es predominantemente heterotrófico, y está caracterizado por la presencia de rutas relacionadas con la detoxificación y tolerancia a altas concentraciones de toxinas, iones y metales. Ésto indica que los ambientes oligotróficos con bajas concentraciones de fósforo y nitrógeno pueden mantener comunidades complejas y diversas, y que la estructura del tapete presenta una solución para la concentración de nutrientes y protección de los miembros de la comunidad per se. Al comparar los tapetes microbianos contra los metagenomas de estromatolitos de la misma región, encontramos una mayor similitud entre los sistemas con regímenes de perturbación similares que entre tipos de sistema o proximidad geográfica. Esto es que la mayor similitud observada no ocurre entre los dos metagenomas de tapetes microbianos (PG y PR) o de estromatolitos (PA y RM), sino entre los dos sistemas desarrollados en los ambientes estables de las pozas permanentes (PG y PA). Éstas dos comunidades son las más diversas taxonómica y funcionalmente, es decir con composiciones más complejas, mayor número de phyla y especies estimadas, un mayor número de proteínas y de funciones y con genomas más pequeños. En contraste, aunque PR y RM no son similares entre sí, presentan un menor número de especies y una mayor dominancia, un menor número de proteínas y funciones, una mayor redundancia funcional y sus genomas son más grandes. Éstos resultados sugieren que los ambientes más estables permiten la diversificación de organismos con genomas pequeños especializados en la explotación de micro nichos diversos, al tiempo que los ambientes más variables promueven la diversificación de organismos generalistas con genomas grandes que les permite una mayor plasticidad y versatilidad metabólica.
No hay duda que la máquina del tiempo de Cuatro Ciénegas hace de este sitio extraordinario un lugar único en el planeta actual, un parque precámbrico. Sin embargo, la historia geológica de Cuatro Ciénegas y sus minerales ricos en azufre lo hacen el mejor análogo del Cráter Gale en Marte, esto es muy importante, ya que en agosto de 2012, el laboratorio Curiosity llegará a Marte al Cráter Gale y empezará a buscar señales de vida tanto en el pasado como en el presente de Marte. El Cráter Gale tiene evidencias de un mar ancestral donde cayó un gran meteorito generando sedimentos ricos en yeso y es probable que, a pesar de que ahora se ve seco, todavía tenga agua profunda donde la vida se pudo haber refugiado [12, 13].

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
11
Referencias [1] V. Souza, L. E. Eguiarte, Nature Review Microbiology 6: 559-564 (2008). [2] V. Souza, L. Espinosa, et al., PNAS 103: 6566-6570 (2006). [3] V. Souza, J. Siefert, et al., en prensa Astrobiology. [4] L. D. Alcaraz, G. Olmedo, et al., PNAS 105:5803-5808 (2008). [5] L. D. Alcaraz, G. Moreno-Hagelsieb, et al., BMC Genomics 11:332 (2010). [6] E. A. Rebollar, M. Avitia, et al., en prensa Microbial Ecology [7] Ch. G. Desnues, B. Rodriguez-Brito et al., Nature. 452:340-342 (2008) [8] Moreno-Letelier, G. Olmedo Gabriela, et al., International Journal of Evolutionary Biology.
doi: 10.4061/2011/781642 (2011). [9] Moreno-Letelier, G. Olmedo et al., en prensa Astrobiology
[10] G. Bonilla-Rosso, M. Peimbert, et al., en prensa en Astrobiology [11] M. Peimbert L. D. Alcaraz, et al., en prensa Astrobiology. [12] N. E. López-Lozano, G. Bonilla, et al., en prensa en Astrobiology. [13] J. L. Siefert, V. Souza et al., en prensa Astrobiology

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
12
Conferencia Plenaria
CICLOS AUTOCATALÍTICOS, CORRECCIÓN DE PRUEBAS Y EL ORIGEN DE LA HOMOQUIRALIDAD
Thomas Buhse y José-Manuel Cruz, Centro de Investigaciones Químicas, UAEM Jean-Claude Micheau y Christophe Coudret, Université Paul Sabatier, Toulouse
[email protected] Introducción
La búsqueda del origen de la homoquiralidad biomolecular se mantiene entre los objetivos más prominentes por resolverse en la ciencia [1]. Los posibles entornos para alcanzar la pureza enantiomérica en un escenario prebiótico han sido sugeridos por varios modelos matemáticos, la mayor parte de ellos inspirados en el esquema seminal de Frank [2]. Estas investigaciones teóricas se vieron apoyadas con observaciones experimentales de estados ópticamente activos emergentes a partir de precursores aquirales que ocurrieron en algunos procesos de cristalización [3] y por la influencia de fuerzas quirales externas tales como la luz circularmente polarizada [4] o por movimiento de vórtice [5]. Otro fenómeno es la denominada “síntesis asimétrica absoluta” [6] donde – al igual que en las moléculas de la vida – la quiralidad se sitúa dentro de un carbono asimétrico. Muchos de estos modelos u observaciones experimentales incluyen procesos cinéticos que toman lugar en sistemas lejos de equilibrio y que requieren de procesos autocatalíticos [7].
Seguramente el ejemplo más importante de síntesis asimétrica absoluta es la adición autocatalítica de diisopropilzinc en pirimidilcarbaldehídos descubierto por Soai [8]. La reacción exhibe una amplificación de pequeños excesos enantioméricos iniciales y la generación sistemática de altos excesos enantioméricos a partir de mezclas iniciales aquirales, un fenómeno conocido como el rompimiento de la simetría especular. La reacción de Soai se mantiene como un ejemplo exclusivo de un sistema de reacción quiralmente autocatalítico en química orgánica y que ha atraído la atención desde diversos puntos de vista: como una explicación para el origen de la homoquiralidad biomolecular, como innovación potencial en la síntesis enantioselectiva o como una notable manifestación de la dinámica no lineal en sistemas químicos. Resultados
Nuestras simulaciones cinéticas de la reacción de Soai revelaron que los procesos de autocatálisis enantioselectiva e inhibición mutua son suficientes para reproducir una extremadamente larga amplificación de excesos enantioméricos y para modelar la síntesis asimétrica absoluta [9-12]. En esta plática se presentan los resultados actuales de un análisis cinético de tres prototípicos modelos cíclicos autocatalíticos basados en la reacción de Soai [13]. Los tres modelos varían por el tamaño que asumen por sus oligómeros catalíticos. Mostrando que el incremento en el tamaño del oligómero está asociado a una mejor tolerancia a un débil reconocimiento quiral entre los diastereómeros en el proceso de la inhibición mutua para lograr el rompimiento de la simetría especular. Esta robustez puede ser entendida como un caso particular de la llamada “corrección de prueba cinética” [14], desarrollada para entender el sorprendente mínimo error generado en procesos bioquímicos. El concepto de corrección de pruebas

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
13
se presenta ahora como una nueva faceta en el creciente campo de la química de sistemas y como un ejemplo para la posible eficaz de procesos prebióticos en el marco de la evolución prebiótica [15]. Referencias
[1] N. Jones, Nature, 2012, 481, 14-17. [2] F. C. Frank, Biochim. Biophys. Acta, 1953, 11, 459-463. [3] D. K. Kondepudi, R. J. Kaufman and N. Singh, Science, 1990, 250, 975–976. [4] M. Avalos, R. Babiano, P. Cintas, J. L. Jiménez, J. C. Palacios and L. D. Barron, Chem. Rev.,
1998, 98, 2391-2404. [5] J. M. Ribó, J. Crusats, F. Sagués, J. Claret and R. Rubires, Science, 2001, 292, 2063-2066. [6] K. Mislow, Collect. Czech. Chem. Commun., 2003, 68, 849-864. [7] D. K. Kondepudi and K. Asakura, Acc. Chem. Res., 2001, 43, 946-954. [8] K. Soai, T. Shibata, H. Morioka and K. Choji, Nature, 1995, 378, 767-768. [9] T. Buhse, Tetrahedron: Asymmetry, 2003, 14, 1055-1061. [10] J. Rivera Islas, D. Lavabre, J. M. Grevy, R. Hernández Lamoneda, H. Rojas Cabrera, J. C.
Micheau and T. Buhse, Proc. Natl. Acad. Sci. USA, 2005, 102, 13743-13748. [11] D. Lavabre, J. C. Micheau, J. Rivera Islas and T. Buhse, J. Phys. Chem. A, 2007, 111, 281-286. [12] J. C. Micheau, J. M. Cruz, C. Coudret and T. Buhse, ChemPhysChem, 2010, 11, 3417-3419. [13] J. C. Micheau, C. Coudret, J. M. Cruz and T. Buhse, Phys. Chem. Chem. Phys., 2012,
DOI:10.1039/C2CP42041D. [14] J. J. Hopfield, Proc. Natl. Acad. Sci. USA, 1974, 71, 4135-4139. [15] J. C. Micheau, C. Coudret and T. Buhse, In: The Soai Reaction and Related Topic. (G. Pályi, C.
Zucchi and L. Caglioti; Eds.), Collana di studi dell'Accademia Nazionale di Scienze Lettere e Arti (Modena), Vol. 37, 2012, pp. 169-196.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
14
PRESENTACIONES ORALES

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
15
Presentación Oral
PARÁMETROS DEL SISTEMA EXOPLANETARIO QUE TRANSITA HAT-P-23b
Pedro A. Valdés Sada, Universidad de Monterrey [email protected]
Felipe G. Ramón Fox, I.T.E.S.M.–Campus Monterrey Introducción
El descubrimiento de planetas orbitando alrededor de otras estrellas, llamados planetas extrasolares o exoplanetas, ha creado una nueva vertiente de vigorosa investigación científica. Muchos planetas orbitan muy cerca de sus estrellas y son gigantes gaseosos. Es común que ocurran tránsitos del planeta frente al disco estelar detectables desde la Tierra. Esta geometría permite medir directamente el tamaño del planeta y, estimando la masa con mediciones de velocidad radial, comprender su estructura interna. Los planetas de período corto, los más cercanos a la estrella, son de particular interés pues la intensa irradiación puede afectar el tamaño del planeta.
HAT-P-23b es uno de estos “júpiteres calientes inflados”. Es un planeta relativamente masivo (~2MJUP) orbitando a una estrella enana G0 con un período de ~1.2129 días. Bakos et al. [1] concluyen que tiene uno de los tiempos de decaimiento orbital más cortos. Se estima que en ~7.5+2.9
-1.8 millones de años sea absorbido por la estrella. En esta presentación se analizan algunos tránsitos recientes del planeta, registrados con un telescopio relativamente pequeño. La intención es de tratar de mejorar los valores de los parámetros esenciales obtenidos de las curvas de luz; como lo son la inclinación orbital (i), el semi-eje mayor normalizado (a/R*) y el tamaño relativo del planeta con respecto a la estrella (Rp/R*). Métodos Observaciones Las observaciones fueron realizadas en el Observatorio de la Universidad de Monterrey (MPC 720) con un telescopio de 0.36m de diámetro y una cámara CCD comercial (SBIG STL-1301E) con un filtro fotométrico estándar Rc (630 nm). La escala de ~1 arcsec/píxel resulta en un campo de visión de ~21.3’×17.1’ y suficientes estrellas de comparación (5). El telescopio fue ligeramente desafocado para mejorar la precisión fotométrica y aumentar los tiempos de exposición (60s). Se registraron cuatro tránsitos entre Junio 4 y Agosto 21 del 2011, con suficiente tiempo de observación para cubrir completamente el tránsito y por lo menos una hora a cada extremo del evento. Reducción y Modelaje
Las imágenes se procesaron de manera estándar con darks y flats, y se realizó fotometría relativa utilizando las mismas estrellas de comparación. Las curvas de luz inicialmente mostraban una pendiente probablemente causada por la extinción diferencial entre las estrellas de comparación y la de interés debido a los diferentes colores de las mismas. Esto se corrigió con una función del tipo δm=c(1-X)+b, donde δm es la corrección aplicada en magnitudes, X es la masa de aire, c y b son constantes de mejor ajuste a las porciones de las curvas fuera del tránsito. Las curvas resultantes se muestran en la parte superior de la Fig. 1.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
16
Curva de Luz Combinada y Efemérides
Las curvas de luz originales poseen una relativamente alta cantidad de ruido causado por utilizar un telescopio de diámetro pequeño. Se decidió combinarlas en una sola y reducir la cantidad de puntos promediando los valores obtenidos dentro de un intervalo de tiempo determinado (2 minutos). El tiempo central (Tc) se obtuvo ajustando modelos de curvas de luz generados con el software Binary Maker 3.0 [2] y los parámetros del sistema derivados por Bakos et al. [1]. Se utilizó un modelo lineal para el ensombrecimiento al limbo y el coeficiente (u) se obtuvo de Claret [3]. El mejor valor del tiempo central se obtuvo alterando las curvas modelo ligeramente en profundidad y duración hasta obtener el mejor ajuste (minimizando χ2) a las curvas de luz observadas. La curva de luz combinada se obtuvo de los cuatro tiempos centrales y fue ajustada para obtener valores generales del modelo. Éstos fueron aplicados iterativamente a las curvas de luz individuales para refinar los tiempos centrales. La curva combinada se muestra en la parte inferior de la Figura 1.
Se derivó un período orbital mejorado utilizando los cuatro tiempos centrales y las efemérides de Bakos et al. [1], ponderando cada valor por el inverso de su incertidumbre. Se obtuvo un período 1.2128868±0.0000004 días (To=2,454,852.26542±0.00018 BJDTDB).
Modelos
Los parámetros esenciales de las curvas se obtuvieron asumiendo los parámetros orbitales e y ω de Bakos et al [1]. En un sentido estricto, es necesario iterar por todos los parámetros (P, a/R*, Rp/R*, i, e, ω, u, Tc de cada tránsito, etc.) simultáneamente, utilizando las curvas de luz individuales y de velocidad radial disponibles. Dicho proceso es mucho más costoso computacionalmente y requiere una gran cantidad de información para converger a un conjunto de soluciones consistentes con bajas incertidumbres. El trabajo se simplifica sustancialmente acotando valores que pueden obtenerse con mayor precisión por métodos alternos.
Una vez fijados los mejores valores para P, e, ω, u, Tc1, Tc2, Tc3 y Tc4 se analizaron los restantes (a/R*, Rp/R* e i) con el código Transit Analysis Package (TAP) [4], que representa las curvas de luz con los modelos de Mandel & Agol [5] e implementa un algoritmo de Monte Carlo de cadenas de Markov para encontrar los parámetros de mejor ajuste. Se corrieron 10 cadenas con 106 muestras por cadena. Los resultados se muestran en la Tabla 1 con los parámetros originales de Bakos et al. [1], y se incluyen los resultados de un modelaje donde se dejan libres los tiempos centrales. Se consideró que los parámetros (P, e, ω y u) están suficientemente bien establecidos por otros métodos y afectan poco a los resultados de los demás.
Resultados Los resultados (Tabla 1) en general coinciden con los de Bakos et al. [1]. En algunos casos, se redujeron las incertidumbres. Particularmente, la inclinación (i) y semi-eje mayor normalizado (a/R*) concuerdan con los de Bakos et al. [1] a 1σ. Aunque se consideró que la incertidumbre de nuestra inclinación es mayor, la del semi-eje mayor normalizado es menor. Se obtiene un tránsito más centrado y una distancia planeta-estrella ligeramente mayor. En general, existe una relación entre estas dos variables ya que un parámetro de impacto menor resulta en una duración mayor del tránsito. Dicho efecto también puede ser consecuencia un semi-eje mayor, donde el planeta viaja más lentamente. Observaciones en otras bandas podrían ayudar a solucionar este problema al esclarecer los efectos del ensombrecimiento al limbo.
El tamaño relativo del planeta (Rp/R*) nos resulta menor y con incertidumbres similares a las de Bakos et al. [1]. Este parámetro depende esencialmente de la forma de las curvas de luz y depende poco de otros parámetros. Dado un radio fijo para la estrella, el radio del planeta resulta ser ~5.5% menor.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
17
Esto concuerda con los modelos de Fortney et al. [6] que predicen un tamaño menor del planeta (~8.4%) comparado con el modelo de Bakos et al. [1]. Cabe aclarar que estos modelos tienen relativamente gran margen de error.
Los resultados de los parámetros esenciales tienen mayores incertidumbres cuando no se acotan los tiempos centrales, aunque el valor central de los parámetros aún concuerda con los anteriores. Esto demuestra la importancia de ajustar lo mejor posible los tiempos centrales, aprovechando un método que ocupa menor tiempo computacional.
Conclusiones Se ha demostrado que varias curvas de luz de tránsitos de exoplaetas pueden combinarse para obtener una de mejor calidad y puede modelarse con éxito, con un menor tiempo de cómputo. Los resultados se comparan favorablemente con los de Bakos et al. [1], con incertidumbres similares y, en cierto caso, menores. Los tiempos centrales han permitido mejorar las efemérides del sistema. En particular, un mejor valor del período es importante para comparar los tiempos de los tránsitos primarios con los de eclipses secundarios (su observación se favorece por la cercanía del planeta a la estrella), lo que puede proporcionar una medida más precisa de la excentricidad orbital. Además, pueden buscarse posibles variaciones en los tiempos centrales, lo que puede llevar a la detección de otros planetas en el sistema por su perturbación gravitacional sobre el que transita. Referencias
[1] Bakos, G. Á., et al. 2011, ApJ, 742, 116. [2] Bradstreet, D. H. 2005, SASS-24, 23 [3] Claret, A. 2000, A&A, 363, 1081 [4] Gazak, J. A., Johnson, J. A., Tonry, J., Eastman, J., Mann, A. W., & Agol, E. 2011, (arXiv:
1102:1036v1) [5] Mandel, K. & Agol, E. 2002, ApJ580, L171 [6] Fortney, J. J., Lodders, K., Marley, M. S., & Freedman, R. S. 2008, ApJ, 678, 1419
Tablas Tabla 1. Parámetros del Modelo de HAT-P-23
Parámetro Curva de luz Combinada Ajuste Global Bakos et al. [1]
i 87.9 +1.5-2.2 87.2 +1.9
-2.0 85.1 ± 1.5
a / R* 4.23 +0.06-0.12
* 4.32 +0.11-0.17
* 4.14 ± 0.23 Rp / R*
0.1105 +0.0015-0.0013 0.1057 +0.0031
-0.0026 0.1169 ± 0.0012
* Este parámetro se extrajo usando los valores orbitales e y ω de Bakos et al. [1] y nuestro periodo P mejorado.
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
18
Figuras
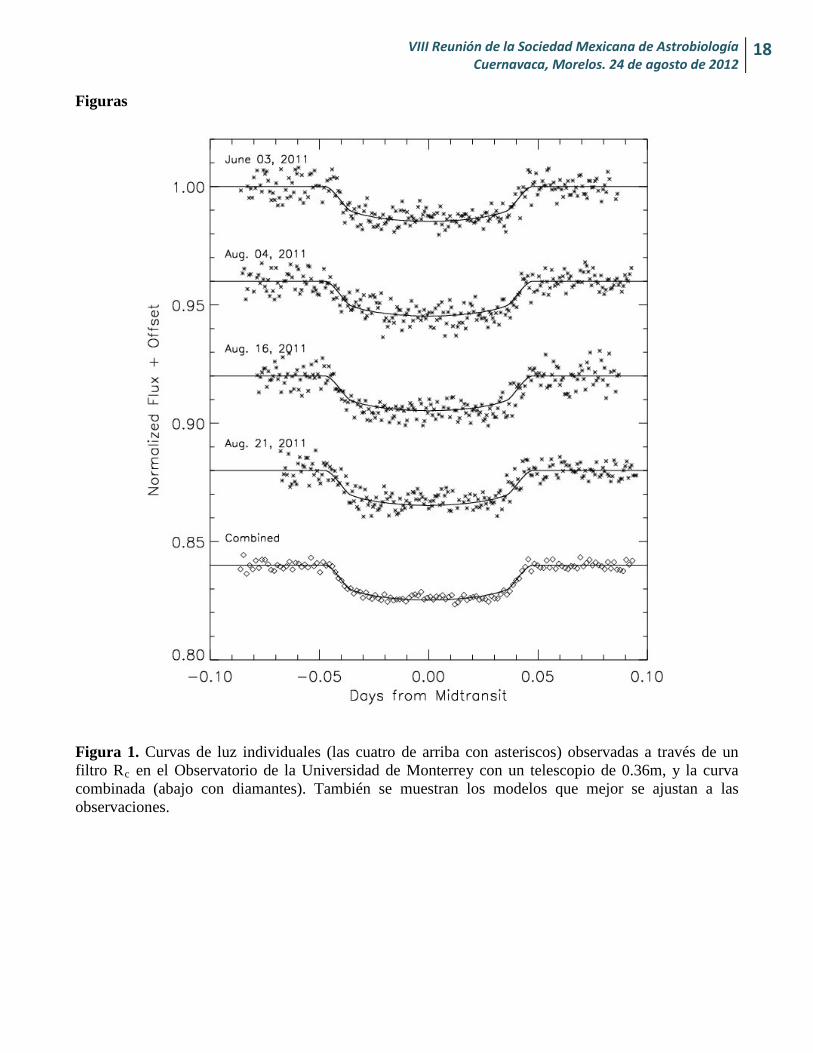
Figura 1. Curvas de luz individuales (las cuatro de arriba con asteriscos) observadas a través de un filtro Rc en el Observatorio de la Universidad de Monterrey con un telescopio de 0.36m, y la curva combinada (abajo con diamantes). También se muestran los modelos que mejor se ajustan a las observaciones.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
19
Presentación Oral
CLASIFICACION PLANETARIA OBTENIDA A PARTIR DE LEYES DE POTENCIA EMPIRICAS
Hector Javier Durand Manterola
Departamento de Ciencias Espaciales, Instituto de Geofísica, UNAM [email protected]
Introducción
Cuando se trata de estudiar las leyes generales que rigen la estructura de cuerpos tales como los exoplanetas, Júpiter, La Tierra, La Luna, Titán, Encelado o Plutón no es importante distinguir si se trata de una luna, un planeta o un planeta enano ya que esas categorías solo se refieren a su mecánica celeste y no a su naturaleza interna. Por eso en este trabajo se tomara como planeta cualquier cuerpo que cumpla con la siguiente definición:
Cuerpo celeste, no importa cómo se haya formado, no importa en qué sitio o vecindario se encuentre, que tenga una masa que esté por debajo de la masa umbral para la fusión termonuclear del deuterio y una masa suficientemente grande como para que su forma (esferoide) este determinada por una relajación viscosa inducida por gravedad.
Desde este punto de vista, para los fines de este trabajo, serán planetas no solo los ocho aceptados por la IAU (International Astronomical Union) sino también La Luna, los cuatro satélites galileanos, Titán, Encelado o cualquiera de los satélites esferoides, así como los planetas enanos Plutón, Eris, Ceres o cualquiera de los cuerpos del cinturón de Kuiper que cumplan con la definición anterior. Igualmente serán incluidos en esta clase los exoplanetas, que aunque no se tienen imágenes de ellos, su gran masa nos inclina a pensar que deben tener formas esferoides determinadas por gravedad.
Uno de los problemas principales al estudiar un planeta es conocer como se distribuye la masa en su interior [1; 2]. Un primer paso hacia comprender esta distribución en los interiores de los planetas en general es ver cómo se comporta el radio, la densidad media y la gravedad superficial al crecer la masa del planeta. Se conocen masa y radio tanto de los planetas del sistema solar como de los planetas extrasolares de transito, y por lo tanto se puede calcular estos parámetros para todos ellos.
En este estudio se trabajó con los datos de 26 planetas del Sistema Solar y 92 exoplanetas de transito y se obtuvieron leyes de potencia empíricas para el radio, la densidad y la gravedad superficial. En base a estas leyes se ve que los planetas se separan, de manera natural, en tres clases. Clases de Planetas
Con datos de 26 planetas del Sistema Solar [3] y de 92 exoplanetas de tránsito [4] obtuve una ley de potencia empírica radio-masa única para todos estos cuerpos (R = 0.0342M0.3428). Los planetas con masa > 3x1025 kg no se ajustan muy bien a esta ley general y parecen formar dos grupos aparte. Estudiando otros parámetros (la densidad y la gravedad superficial) se observa más claramente que todos los planetas se separan de manera natural en tres clases A, B y C. La clase A son todos los planetas con masa < 3x1025 kg, la clase B esta en el intervalo 3x1025kg < M < 1x1027kg y la clase C son planetas con masa > 1027 kg. En nuestro Sistema Solar todos

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
20
los planetas excepto los cuatro gigantes gaseosos pertenecen a la clase A. Saturno, Urano y Neptuno pertenecen a la clase B y únicamente Júpiter pertenece a la clase C. Propiedades de las Tres Clases Planetarias
En la figura 1 vemos que los radios de los planetas clase A siguen una ley de potencias más suave que los de clase B que tienen una pendiente más dura. Esto indica que el volumen de los planetas clase B aumenta más rápidamente con la masa que los de clase A. Los planetas de clase C tienen una pendiente casi nula, lo cual implica que al aumentar su masa su volumen no varía.
Al graficar la densidad media planetaria contra su masa (Figura 2) se observa más claramente la separación de los planetas en tres clases. En general los planetas de las clases B y C son menos densos que los de la clase A mostrando la separación entre planetas sólidos (clase A) y los gigantes gaseosos (B y C). La separación entre la clase B y la clase C es bien clara mostrando que existen dos clases de gigantes gaseosos.
Por su parte la gravedad superficial (figura 3) también muestra una clara diferencia en la pendiente en las tres clases.
En las tres figuras también vemos las leyes de potencia para las tres clases de planetas. Las leyes de potencia se obtuvieron con un ajuste de mínimos cuadrados de los datos de cada clase. Conclusiones
Los más relevantes resultados de este trabajo son: • El radio, la densidad y la gravedad superficial son leyes de potencia de la masa. • Los planetas en el intervalo de 1019 a 1029 kg se separan en tres clases de planetas. • Los planetas de clase A, a la cual pertenece la Tierra y casi la totalidad de los planetas del
Sistema Solar, están en el intervalo de masa de 1019 kg < M < 3x1025 kg, y su radio, su densidad y su gravedad superficial crecen al crecer la masa.
• Los planetas de clase B, a la cual pertenecen Saturno, Urano y Neptuno, están en el intervalo de masa 3x1025 kg < M < 1027 kg, su radio crece con la masa y tienen la aparentemente inesperada propiedad de disminuir su densidad y su gravedad superficial al aumentar su masa.
• Los planetas de clase C, a la cual pertenece Júpiter, están en el intervalo de masa 1027 kg < M < 1029 kg y tienen la propiedad de que su radio se mantiene constante al aumentar la masa. Su densidad y su gravedad superficial crecen con la masa.
Una versión más extensa de este trabajo puede verse en [5]. Referencias
[1] Valencia, D. et al. Detailed Models of super-Earths: How well can we infer bulk properties? Ap. J. 665: 1413-1420 (2007).
[2] Swift, D.C., J. Eggert, D.G. Hicks, S. Hamel, K. Caspersen, E. Schwegler, G.W. Collins, N. Nettelmann and G.J. Ackland.. Mass-radius relationships for exoplanets. arXiv: 1001.4851v2, (2011).
[3] Tholen, D.J., V.G. Tejfel and A.N. Cox. Chapter 12 Planets and Satellites. In Allen’s Astrophysical Quantities. 4th Edition Editor A.N. Cox. AIP Press and Springer. (2000)
[4] Schneider, J. 2010. The Extrasolar Planets Encyclopedia, http://exoplanet.eu/ [5] Durand-Manterola, H.J. Planets: Power Laws and Classification. arXiv: 1111.3986v1, (2011)
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
21
Figuras
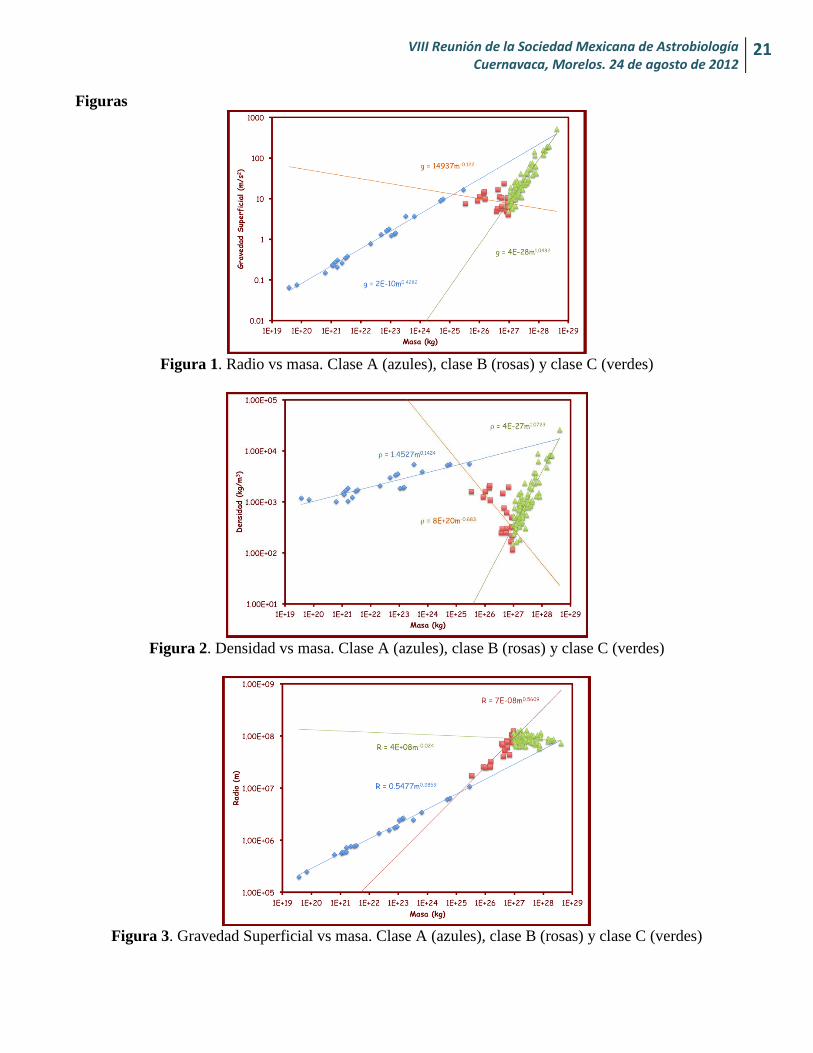
Figura 1. Radio vs masa. Clase A (azules), clase B (rosas) y clase C (verdes)
Figura 2. Densidad vs masa. Clase A (azules), clase B (rosas) y clase C (verdes)
Figura 3. Gravedad Superficial vs masa. Clase A (azules), clase B (rosas) y clase C (verdes)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
22
Presentación Oral
ORIGEN DE LA NUBE PROGENITORA DEL SISTEMA SOLAR
Leticia Carigi, Instituto de Astronomía, Universidad Nacional Autónoma de México [email protected]
Manuel Peimbert, Instituto de Astronomía, Universidad Nacional Autónoma de México
Introducción Hace varias décadas, a partir de la comparación de abundancias químicas del Sol y de sus
estrellas vecinas que se contaban en esa época, se concluyó que el Sol presentaba mayor contenido de metales (elementos más pesados que el He) que sus compañeras. Debido a que el disco de la Vía Láctea tiene un gradiente químico, siendo el contenido de metales mayor en zonas internas del disco que en zonas externas, se propuso que el Sol podría haberse formado a un radio galactocéntrico (r, distancia medida desde el centro de la galaxia a un punto sobre el disco de la misma) menor al que se halla en la actualidad (8 kpc, 1kpc~3,300 años luz). Paralelamente, se propuso que el Sol pudo haberse formado de un gas enriquecido por una supernova cercana, ya que las estrellas masivas (progenitoras de supernovas del tipo II) producen la mayor cantidad de los elementos pesados. Recientemente, la primera propuesta ha tomado auge con el surgimiento de la teoría de migración estelar, a partir de la cual las órbitas estelares alrededor del centro Galáctico podrían ser perturbadas por la barra y los brazos de la Vía Láctea, haciendo que las estrellas migren a través del disco galáctico de r menores a r mayores, y viceversa.
Confrontando modelos detallados de evolución química de galaxias (MEQ) con abundancias químicas, precisas y recientes, en estrellas y en el gas interestelar, es posible probar los modelos y las abundancias en estrellas de diferentes edades y en el medio interestelar actualmente ionizado (regiones HII, ubicadas todas en el disco Galáctico a diferentes r). A fin de obtener un modelo robusto para la Vía Láctea, se utilizó como principal restricción observacional la pendiente y la ordenada en el origen del gradiente de O/H mostrado por las regiones HII. El MEQ fue probado con otras observaciones del disco de nuestra galaxia (con r~25 kpc), y en particular de la Vecindad Solar (a r=8 kpc), logrando responder a las preguntas: dónde se encontraba la nube progenitora del Sistema Solar y qué tipos de estrellas aportaron los elementos químicos presentes en el Sol y en los planetas. Una discusión completa de este trabajo ha sido presentada por [1]. Revisión y adecuación de las restricciones observacionales
Los modelos de evolución química son construidos para reproducir el gradiente de un elemento en particular. Aquí se diseñó el MEQ para ajustar el gradiente de O/H, ya que el oxígeno es el elemento pesado más abundante en el universo y el mejor determinado en las regiones HII.
Los gradientes de O/H publicados en la literatura son obtenidos a partir de diversos métodos, la mayoría de los gradientes muestran pendientes similares, pero con una dispersión considerable en la ordenada en el origen de O/H. Una comparación de muchos de los métodos utilizados ha sido realizada por [2]. Ellas encontraron que las diferencias entre los valores de O/H derivados por los distintos métodos para una determinada región HII puede ser tan grande como 0.7 dex, o 5 (100.7) en escala lineal. La mayor parte de las diferencias entre las diversas calibraciones se debe a la distribución en

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
23
temperatura dentro de las regiones. El método de líneas de recombinación (LR), el cual es casi independiente de la temperatura electrónica, arroja abundancias gaseosas de O y C más altas, entre 0.15 y 0.35 dex, que el método de líneas prohibida (LP), el cual es fuertemente dependiente de la temperatura electrónica. En este trabajo se utilizan las abundancias de H, He, C y O de regiones HII calculadas únicamente con el método de LR. Dado que los cocientes de LR para esos cuatro elementos son prácticamente independientes de la temperatura electrónica, sus abundancias gaseosas relativas son muy fiables. Además, estas abundancias fueron corregidas bajo la suposición de que el 35% de los átomos de O y el 25% de los átomos de C están atrapados en granos de polvo [3], incrementando los valores gaseosos de O/H y C/H por 0.12 y 0.10 dex.
A fin de conocer la historia química del medio interestelar, hemos considerado estrellas no evolucionadas de diferentes edades, ya que al ser relativamente jóvenes, las abundancias químicas observadas en las fotósferas estelares corresponden a las abundancias del material gaseoso donde se formaron las estrellas. Resultados del modelo de evolución química para la Vía Láctea
En las Figuras 1 y 2 se presentan los resultados de nuestro modelo, junto con las mejores restricciones observacionales disponibles (mencionadas en la sección anterior). Para que el modelo reprodujera todas las características del gradiente de O/H actual, a t = 13 Gyr (1 Gyr=1 Gigaño = mil millones de años), se supuso un escenario de formación galáctica dentro-fuera (las zonas centrales de la Vía Láctea se formaron más eficiente y tempranamente que las zonas periféricas) y se consideró que las estrellas masivas de alto contenido de metales pierden masa con una tasa moderada en la etapa de vientos estelares.
Específicamente, nuestro modelo reproduce: (a) Los valores de O/H, C/H y Fe/H, y la relación de Fe/H vs el tiempo derivada por estrellas de diferentes edades ubicadas en el halo y en el disco de la Vecindad Solar (Fig. 1). (b) Las abundancias protosolares (cuando el Sol se formó) de He (Y), C y O (Fig. 2, paneles izquierdos) y de Fe (Fig. 1, panel derecho inferior), correspondiente al contenido de esos elementos en el medio interestelar hace 4.5 Gyr. (c) Los gradientes actuales de O/H, C/H y C/O (Fig. 2, paneles derechos), tanto la pendiente como las ordenadas en el origen.
Sabiendo que el valor protosolar es 12+log(O/H)=8.73±0.05 [4] y que los valores de O/H obtenidos por nuestro MEQ a r=8 y 7 kpc son 8.70 y 8.77 para el medio interestelar cuando el Sol se formó (t=8.5 Gyr), se obtiene que el Sol se originó en r=7.6 ± 0.8 kpc. Conclusiones
1. El acuerdo de He/O, C/O y Fe/O entre el modelo y las abundancias protosolares implica que el Sol se formó a partir de un material gaseoso químicamente bien mezclado y sin contribución mayoritaria de ningún tipo de supernova, ya que la mitad del He generado en la nucleosíntesis estelar (y el C) es producido por estrellas masivas (supernovas del tipo II) y la otra mitad por estrellas de baja masa [5], y una fracción importante del Fe proviene de supernovas del tipo Ia.
2. La concordancia del modelo con las abundancias protosolares al tiempo de formación del Sol apoya la idea de que el Sol se originó a una distancia galactocéntrica de 7.6±0.8 kpc, muy cerca de su posición actual (r=8 kpc).
Referencias
[1] Carigi, L., & Peimbert, M. 2011, RevMexAA, 47, 139 [2] Kewley, L. J., & Ellison, S. L. 2008, ApJ, 681, 1183 [3] Peimbert, A., & Peimbert, M. 2010, ApJ, 724, 791 [4] Asplund, M., Grevesse, N., Sauval, A. J., & Scott, P. 2009, ARA&A, 47, 481
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
24
[5] Carigi, L., & Peimbert, M. 2008, RevMexAA, 44, 311
Figuras
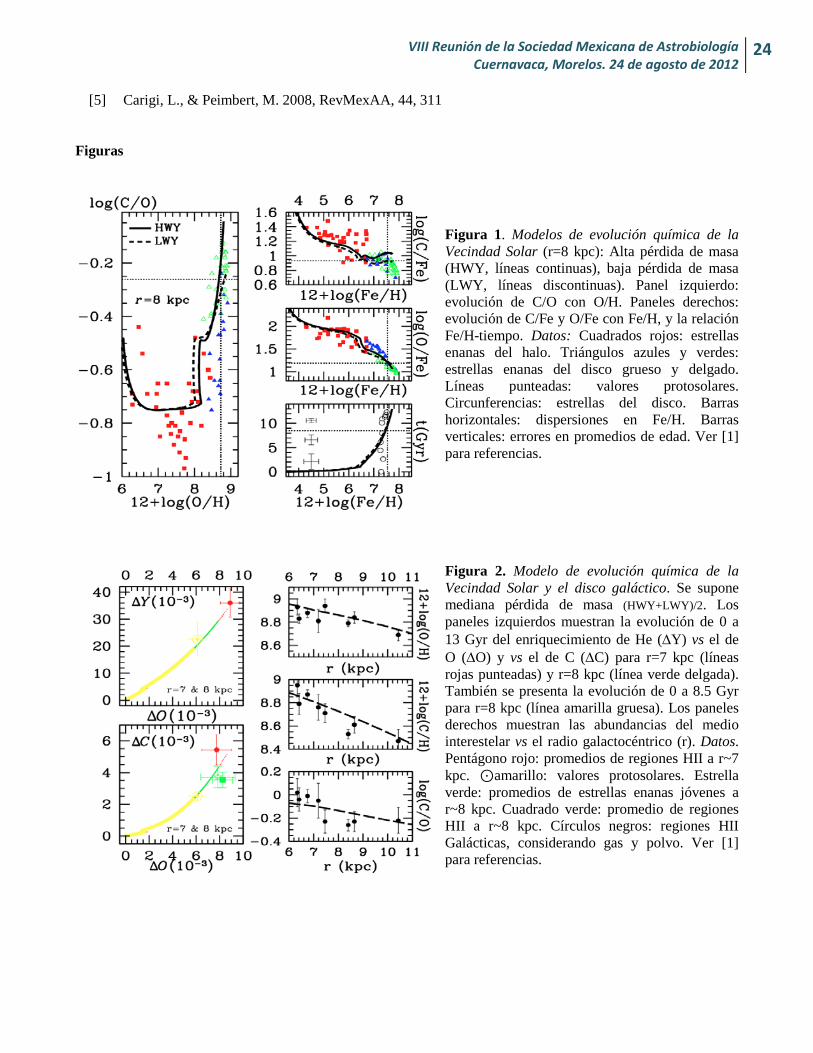
Figura 1. Modelos de evolución química de la Vecindad Solar (r=8 kpc): Alta pérdida de masa (HWY, líneas continuas), baja pérdida de masa (LWY, líneas discontinuas). Panel izquierdo: evolución de C/O con O/H. Paneles derechos: evolución de C/Fe y O/Fe con Fe/H, y la relación Fe/H-tiempo. Datos: Cuadrados rojos: estrellas enanas del halo. Triángulos azules y verdes: estrellas enanas del disco grueso y delgado. Líneas punteadas: valores protosolares. Circunferencias: estrellas del disco. Barras horizontales: dispersiones en Fe/H. Barras verticales: errores en promedios de edad. Ver [1] para referencias.
Figura 2. Modelo de evolución química de la Vecindad Solar y el disco galáctico. Se supone mediana pérdida de masa (HWY+LWY)/2. Los paneles izquierdos muestran la evolución de 0 a 13 Gyr del enriquecimiento de He (∆Y) vs el de O (∆O) y vs el de C (∆C) para r=7 kpc (líneas rojas punteadas) y r=8 kpc (línea verde delgada). También se presenta la evolución de 0 a 8.5 Gyr para r=8 kpc (línea amarilla gruesa). Los paneles derechos muestran las abundancias del medio interestelar vs el radio galactocéntrico (r). Datos. Pentágono rojo: promedios de regiones HII a r~7 kpc. ⊙amarillo: valores protosolares. Estrella verde: promedios de estrellas enanas jóvenes a r~8 kpc. Cuadrado verde: promedio de regiones HII a r~8 kpc. Círculos negros: regiones HII Galácticas, considerando gas y polvo. Ver [1] para referencias.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
25
Presentación Oral
GENERACIÓN EXPERIMENTAL DE FUNDIDOS TIPO CONDROS PARA DETERMINAR LOS MECANISMOS DE SU FORMACIÓN
Antígona Segura, Instituto de Ciencias Nucleares, Universidad Nacional Autónoma de México
[email protected] Karina Cervantes, Centro de Geociencias, Universidad Nacional Autónoma de México
Introducción
Los condros son objetos ígneos y cuasi-esféricos de tamaño milimétrico, con mineralogía y texturas características, que se formaron en el disco de acreción protoplanetario cuando los planetas y asteroides todavía no se agregaban. Por más de 200 años ha existido un debate muy intenso a nivel mundial en torno a los mecanismos que actuaron en la formación de los condros. Los condros se encuentran dentro de las meteoritas de tipo condrítico cuyo estudio ha sido fundamental para interpretar las condiciones fisico-químicas de la formación del Sistema Solar temprano y es la clave para descifrar el material y procesos que formaron a la Tierra y los planetas vecinos. Hipótesis sobre la formación de los condros
El rango temporal de formación de los condros (~4,564 Ma.) va de 1 a 3 Ma después de la formación de las inclusiones de calcio y aluminio (~4,567 Ma.) que fueron los primeros sólidos que se condensaron en el nebulosa solar [1,2]. Las características petrológicas de los condros sugieren historias de formación instantáneas las cuales no se vuelven a repetir dentro del sistema solar después del intervalo de edad mencionado.
A pesar de los avances realizados para comprender la génesis del sistema solar, los astrofísicos aún no han logrado llegar a un consenso acerca del mecanismo preciso del calentamiento de los condros, la hipótesis más favorecida es la de los calentamientos tipo “flash” [3]. Los calentamientos flash derivados de frentes de onda de choque encabezan la lista de los posibles mecanismos formadores de condros [3]. Este modelo se refiere a ondas de choque que se propagan en el disco protoplanetario, de manera tal que pueden tomar agregados precursores y calentarlos hasta el punto de fusión por medio de un proceso de calentamiento, en el cual, tanto el arrastre del gas como la energía térmica, se pierde por las colisiones grano-grano hasta alcanzar la radiación termal del frente de onda y de otros granos de polvo [4,5].
El mecanismo de calentamiento de partículas sólidas por ondas de choque [6] funciona de la siguiente forma: cuando una onda de choque es generada en la nebulosa solar (que es una mezcla de gas y polvo) el gas es frenado por su presión mientras que las partículas de polvo tienden a conservar su velocidad inicial. Debido a este comportamiento dinámico diferente detrás del frente de la onda de choque, se genera una diferencia de velocidad relativa entre el gas y las partículas de polvo, así las partículas de polvo son frenadas por la fuerza de arrastre de gas causada por la velocidad relativa con el gas y son calentadas por la energía de arrastre o fricción. Existen por lo menos cinco propuestas diferentes de mecanismos generadores de ondas de choque: El primer mecanismo es por medio de choques de acreción a la superficie del disco protoplanetario [7,8]. El segundo por la acreción de masas

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
26
de gas en la nebulosa solar [9, 10] y el tercero por ondas de proa generadas por el movimiento de planetésimos de 1000 km de diámetro en órbitas altamente excéntricas. Un cuarto mecanismo son las ondas en espiral [4; 11] generadas por inestabilidades gravitacionales, con tendencia de formación de planetas más allá de la órbita de Júpiter, y cuyas velocidades de choque alcanzan los 10 km/s [12; 13; 3]. Un quinto mecanismo puede ser el de la inducción de choques por ráfagas de rayos X en la parte superior del disco protoplanetario, cuyas velocidades de frente de choque son de 40 km/s [6]. Simulaciones experimentales
La aproximación experimental al estudio de los procesos de la formación de los condros se ha realizado en su mayoría a partir de generar fundidos en estufas. Estos experimentos han sido cuestionados debido a que no toman en cuenta las condiciones reales de formación de los condros en dos aspectos: 1) La retención de materiales volátiles como el FeS, Na y K, los cuales no sobreviven a calentamientos y/o enfriamientos que requieren de varios minutos, horas y hasta días y 2) los granos relictos y bordes ígneos que indican diferentes pulsos de calentamiento y no enfriamientos monótonos posteriores a un evento único de calentamiento [14, 15]. Los experimentos en los que se ha utilizado radiación láser para estudiar la formación de condros permiten en cambio el enfriamiento rápido de los fundidos y el recalentamiento de los mismos a partir de pulsos de radiación subsecuentes.
Los experimentos reportados donde se utiliza un láser para la formación de condros han logrado reproducir las texturas observadas en ellos [16, 17,18]. En la UNAM una serie de experimentos preliminares fueron realizados en el Laboratorio de Geocronología K-Ar del Instituto de Geología con resultados similares a los reportados en la literatura [19]. Los experimentos mencionados anteriormente no fueron realizados en condiciones de vacío como las que se esperan del disco protoplanetario en el cual se formaron los condros y las temperaturas reportadas son derivadas a partir de modelos teóricos y no se miden directamente durante los experimentos [16, 17, 18].
Propuesta para el estudio de mecanismos de formación de condros
En este proyecto se realizará un estudio de las propiedades petrológicas de fundidos tipo condros que se generarán en simulaciones experimentales. La finalidad de los experimentos es reproducir los procesos de calentamiento que pudieron dar origen a los condros. Las simulaciones permitirán determinar a qué temperaturas se formaron las distintas fases y texturas minerales de los condros simulados y éstas se compararán con las propiedades petrológicas observadas en los condros de metoritas condríticas. De esta manera podremos acotar las temperaturas de formación de los condros y por lo tanto los posibles procesos que los originaron. Metodología
Los experimentos inician con la determinación de las propiedades químicas del material precursor que en este caso es tomado de minerales naturales provenientes de xenolitos ultramáficos (peridotitas). Los precursores se colocarán dentro de la cámara de vacío para ser irradiados con el láser del CO2 durante periodos tiempos específicos a diferentes potencias. La temperatura de los fundidos será medida antes, durante y después de la irradiación.
Los fundidos resultantes serán cortados con un disco de diamante para la elaboración de secciones delgadas y su posterior descripción petrográfica. La composición química de los minerales será evaluada mediante una microsonda electrónica por medio de la espectrometría de energía dispersiva de rayos X. También se evaluará la estructura mineral mediante espectroscopía micro-Raman.
La petrología de los fundidos tipo condros será comparada con las características observadas en condros de meteoritas condríticas. Para aquellos fundidos con propiedades petrológicas similares a las

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
27
observadas se analizará su historia térmica durante el proceso experimental de fundido. Estas historias, en particular la temperatura máxima alcanzada y la tasa de enfriamiento, se compararan con las que se proponen para cada uno de los mecanismos propuestos para la formación de condros.
Agradecimientos
Este trabajo es apoyado con los fondos de los proyectos PAPIIT IA101312 y CONACYT No. 128228. Referencias
[1] Y. Amelin, A.N. Krot, I. D. Hutcheon, A.A. Ulyanov, Science 297, 1678-1683 (2002) [2] R.H. Hewins, R.H. Jones, E.R. D. Scott, Chondrules and the ProtoplanetaryDisk (Cambridge
Univ. Press, Cambridge, U.K, 1996). [3] A.P. Boss, R.H. Durisen, Astrophys. J. 621, L137-L140 (2005). [4] L.L. Hood, M. Horanyi, Icarus 93, 259-269 (1991). [5] L.L. Hood, M. Horanyi, Icarus 106, 179-189 (1993). [6] T. Nakamoto, H. Miura, presentado en Lunar and Planetary Science Conference XXXV, 2004. [7] J.A. Wood, Earth Panet. Sci. Lett. 70, 11-26 (1984). [8] T.V Ruzmaikina, W.H. Ip, Icarus 112, 430-447 (1995). [9] A.P. Boss, J.A. Graham, Icarus 106, 168-178 (1993). [10] L.L. Hood, D.A. Kring en Chondrules and the protoplanetary disk, R.H. Hewins, R.H. Jones,
E.R.D. Scott, Eds. (Cambridge Univ. Press., Gran Bretaña, 1996), pp. 265-276. [11] J.A. Wood en Chondrules and the protoplanetary disk, R.H. Hewins, R.H. Jones, E.R.D. Scott,
Eds. (Cambridge Univ. Press., Gran Bretaña, 1996), pp. 55-69. [12] A.P. Boss, Astrophys. J. 576, 462-472 (2002). [13] B.K., Pickett, et.al. Astrophys. J. 590, 1060-1080 (2003). [14] J.T. Wasson, en Chondrules and the protoplanetary disk, R.H. Hewins, R.H. Jones, E.R.D.
Scott, Eds. (Cambridge Univ. Press., Gran Bretaña, 1996), pp. 45-54. [15] S. Desch, M. Morris, H. Connolly, A. Boss, presentado en Lunar and Planetary Science
Institute., id.8008, 2010. [16] L. S. Nelson, M. Blander, S.R. Skaggs, K. Keil, Earth Planet. Sci. Lett. 14, 338 (1972). [17] M. Blander, H. N. Planner, K. Keil, L.S. Nelson, N.L. Richardson, Geochim. Cosmochim.
Acta 40, 889-896 (1976). [18] D.D. Eisenhour, T.L. Daulton, P.R. Buseck, Science 265, 1067-1070 (1994). [19] K. Cervantes de la Cruz, Tesis de doctorado, Universidad Nacional Autónoma de México,
(2009).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
28
Presentación Oral
PERSPECTIVAS DE LA INVESTIGACIÓN CIENTÍFICA EN LA AGENCIA ESPACIAL MEXICANA
Celso Gutiérrez Martínez, Dirección de Investigación Científica
Agencia Espacial Mexicana (AEM) [email protected]
Introducción
La creación formal de la Agencia Espacial Mexicana (AEM), en noviembre de 2011, abre grandes perspectivas para que México avance en el campo espacial, en diferentes aspectos relacionados con desarrollo científico, tecnológico, formación de capital humano e impulso industrial. Estos temas forman parte de la Ley de Creación de la AEM y constituyen los ejes rectores del Programa Nacional de Actividades Espaciales de nuestro país.
En el marco de funciones de la AEM, destaca la facultad de promoción de actividades relacionadas con la investigación científica y tecnológica. Para cumplir con esta función, una de las coordinaciones sustantivas de la AEM es la de Investigación Científica y Desarrollo Tecnológico Espacial, la cual está conformada por las direcciones correspondientes. El impulso a la investigación científica y tecnológica es una actividad estratégica, particularmente porque sus resultados inciden directa o indirectamente en la solución de necesidades sociales, en el desarrollo y crecimiento de la industria nacional y en la generación de innovaciones que aumenten la competitividad. En nuestro país se cuenta con capacidades humanas y de infraestructura vinculadas a la investigación científica y tecnológica del sector aeroespacial. En el marco de la AEM, el área de investigación científica espacial considera dos áreas de competencia. Investigación Astronómica, Astrofísica, y Planetaria.
Con una importante tradición, las ciencias espaciales mexicanas se han desarrollado en diversos escenarios con instituciones de reconocido prestigio nacional e internacional. Las actividades en astronomía, astrofísica, geofísica han tenido un acelerado crecimiento tanto en aspectos teóricos como experimentales, con información de diferentes plataformas terrestres y espaciales; asimismo importantes investigaciones apoyadas en cómputo de alto rendimiento están siendo llevadas a cabo, trabajando en el modelo de redes de procesamiento distribuido; así como la contribución de equipos de vuelo para misiones científicas internacionales en plataformas espaciales que están desarrollando infraestructura crítica para la producción y certificación de tecnología espacial en el país. Ciencias Médicas Espaciales y Telesalud.
México se ha destacado por ser un pionero en los estudios de ciencias médicas aeroespaciales, contando con destacados investigadores en estos campos, quienes han realizado contribuciones importantes en esta materia. También hemos venido siendo pioneros en la implementación de programas de telesalud y cibersalud que consideran la sustentabilidad de los mismos. Los programas desarrollados han permitido a México convertirse el líder de esfuerzos continentales en materia de

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
29
telesalud, así como establecer cátedras y estar en posición de contribuir con propuestas de normatividad internacional.
En el marco de estas dos líneas, algunas atribuciones de la Dirección de Investigación Científica Espacial incluyen, aspectos de promoción y organización y conducción de actividades de investigación en el sector espacial del país, que propicien la generación de conocimiento científico y que se traduzcan en aplicaciones que respondan a demandas económicas y sociales y que propicien un desarrollo sustentable. En estas perspectivas se consideran proyectos científicos de investigación espacial, relativos a las ciencias astronómicas, astrofísicas y planetarias, desde plataformas terrestres ó desde el espacio, proyectos de telesalud y cibersalud para apoyar las necesidades del sector salud y de la población mexicana en esta materia, proyectos para la generación e intercambio de conocimientos y experiencias científicas entre expertos mediante la realización de congresos, conferencias, talleres, entre otros.
La AEM fungirá como ente coordinador y de enlace entre instituciones académicas, dependencias gubernamentales, empresas y asociaciones públicas y privadas con objeto de promover la realización de proyectos de investigación científica y su potencial desarrollo tecnológico, así como todas las actividades relativas al intercambio de conocimientos y experiencias en materia espacial.
A la fecha, la AEM, está trabajando con objeto de ir avanzando en el cumplimiento de sus atribuciones como organismo rector de la política nacional en materia espacial. La AEM está en proceso de concretar convenios de colaboración con instancias públicas y privadas en todo el país. En materia de investigación científica espacial y de desarrollo tecnológico, la AEM está colaborando estrechamente con la Red de Ciencia y Tecnología Espaciales (RedCyTE), dependiente del CONACYT y con la Sociedad Mexicana de Ciencia y Tecnología Aeroespacial, A. C. (SOMECYTA), así como con los gobiernos de los estados de Puebla, Querétaro, Baja California y Jalisco, en la perspectiva de proyectos espaciales conjuntos.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
30
Presentación Oral
EL CORAZÓN EN EL ÁMBITO ESPACIAL
Dr. Ramiro Iglesias Leal, Secretaria de Salud de Tamaulipas [email protected]
Introducción
El ámbito espacial remodela la anatomía y fisiología humana, introduce cambios en prácticamente todos los aparatos y sistemas, de tal manera que un astronauta al regresar a la Tierra parece un ser distinto al que vimos partir. Esos cambios obedecen a un proceso de adaptación al medio ambiente espacial. (Fig.1) Las condiciones del espacio exterior son incompatibles con la vida: la ausencia de atmosfera, la radiación cósmica, las temperaturas extremas, algunos componentes del espectro electromagnético (luz infrarroja, luz ultravioleta, rayos X, rayos gamma), la ruptura del siclo día/noche, y la presencia de micrometeoroides requieren ser contrarrestados con sistemas ecológicos sellados como trajes, naves y estaciones espaciales. La ausencia de gravedad es el único factor que no ha requerido de control especial, porque el organismo humano lo tolera por tiempo prolongado, quizás indefinidamente, pero es el que impone casi todos los cambios que el cuerpo humano experimenta. (Fig.2)
El sistema cardiovascular es uno de los componentes del organismo que cambia con mayor intensidad en el espacio, y la ausencia de gravedad es el único factor responsable de esa transformación. (Fig.3)
El propósito de este trabajo es significar un hecho excepcional en la medicina y es que cada una de las numerosas modificaciones que experimenta el corazón y la circulación en el estado de ingravidez son normales en el ámbito espacial, pero si se valoran con criterios de cardiología “terrestre”, son indicativas de patología severa.
Métodos
Este trabajo se sustenta en la experiencia del autor adquirida en la especialidad de medicina aeroespacial realizada en la Fuerza Aérea y la NASA de los Estados Unidos de Norteamérica; en su participación como invitado y observador en diferentes misiones espaciales; en una revisión general de la literatura médica mundial sobre el comportamiento del sistema cardiovascular en el espacio; y en la elaboración del libro “Cardiología Aeroespacial” de su autoría. (Fig.4).
Resultados
Los estudios cardiovasculares practicados a los astronautas en el espacio exterior muestran los siguientes cambios:
* Edema de la cara. (Fig.5) * Venas del rostro y el cuello dilatadas. * Presión arterial similar en todas las regiones del cuerpo. * Corazón horizontalizado (choque del ápex en cuarto espacio intercostal izq.) * El diafragma se desplaza hacia arriba cinco a seis centímetros. * El tórax se vuelve más corto, más amplio y redondeado.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
31
* El hígado se eleva de cinco a seis centímetros. * Las venas de los miembros inferiores se colapsan. * El pulso arterial en miembros inferiores disminuye. * El corazón se vuelve más pequeño (15 a 20%). * El tórax aloja mayor cantidad de sangre (± 800 cc). * La presión venosa central desciende (se sitúa alrededor de 2 mm Hg). * La cantidad total de sangre disminuye (± un litro). (Fig.6) * Bradicardia sinusal acentuada durante el sueño (hasta 40 latidos por minuto).
Conclusiones 1) Los cambios del sistema cardiovascular que se observan en el espacio obedecen a un proceso
de adaptación a ese medio, específicamente a la ausencia de gravedad; 2) todos y cada uno de esos cambios, si se observan en tierra, tienen un significado patológico severo; 3) un medico no familiarizado con la fisiología humana en el espacio, estaría haciendo diagnósticos muy graves y, por supuestos equivocados; 4) por estas y otras razones que no se cubren en el presente trabajo, debe incluirse la enseñanza formal de la medicina espacial en la formación del médico del futuro. Referencias
[1] Atkov O., Bednenko VS., AND Fomina GA., Ultrasound techniques in space medicine. Avitat Space Environ Med, 1987; suppl58:69-71
[2] Buckey JC., Gaffney FA., Lane LD., Levine BD., Watenpaugh DE., Wright SJ., Yancy CW Jr. and Blomqvist CG. Central venous pressure in space. J. Appl Physiol, 1996; 81:19-25.
[3] Bungo M. W., Goldwater D.J., Propp, R.L. and Sandler, H., Echocardiographic evaluation of Space Shuttle crewmembers. J. Appl Physiol. 62: 278-283, 1987.
[1] Churchill SE., Bungo MW., Responses of the cardiovascular system to spaceflight. In: Churchill SE, ed. Fundamentals of space life science vol. 1. Malabar: Krieger; 1997: 41-64.
[2] Hoffler G.W., Cardiovascular studies of U.S. space crew: An overview and perspective. In: Cardiovascular Flow Dynamics and Measurements. Edited by Hwang, N.H. C., and Normann, N.A. Baltimore, MD, University Park Press, 1887.
[3] Iglesias R. Cardiología aeroespacial. México: Limusa, 2012. [4] Nicogossian A. Hoffler G.W., Johnson R.L., and Gowen R.J., Determination of cardiac size
following space missions of different durations: The second manned Skylab mission. Aviat Space Environ. Med. 47(4): 362-365,1976.
[5] Watenpaugh DE., Hargens AR., The cardiovascular system in microgravity. In: Fregly MJ., Blatteis CMIII, eds. Handbook of Physiology: environmental physiology. New York: Oxford University Press, 1996: 631-674.
Figuras
Radiación cósmica
Fig. 1. El ámbito especial remodela la Fig. 2. Las condiciones del ámbito Fig. 3. La gravedad cero produce la anatomía y fisiología humana. espacial son incompatibles con la casi totalidad de los cambios anatómicos vida, excepto la ingravidez. y funcionales.
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
32
Fig. 4. Esta obra resume el Fig. 5. El rostro de un astronauta se edemematiza conocimiento del comportamiento en gravedad cero, en tierra ese signo clínico significa del corazón en el ámbito espacial. patología severa cardiaca o renal.
Fig. 6. Esta grafica de cateterismo cardiaco derecho revela un dato inesperado en la dinámica cardiaca en ingravidez: la presión venosa central disminuye en lugar de aumentar como se esperaba.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
33
Presentación Oral
LA HALOTOLERANCIA EN EL CONTEXTO DE LA ASTROBIOLOGIA
Sandra I. Ramírez Jiménez, Centro de Investigaciones Químicas, UAEM [email protected]
Introducción
La halotolerancia es la capacidad de adaptación de los seres vivos a condiciones de alta salinidad. Los procariontes halotolerantes y los halófilos requieren concentraciones salinas comparables o superiores a las existentes en los océanos terrestres en donde la sal predominante es el cloruro de sodio (NaCl).
Evaluar la halotolerancia en términos de sales inorgánicas diferentes al NaCl es difícil porque la información necesaria es escasa o inexistente. Adicionalmente, la existencia de cuerpos de agua salados debajo de la superficie de algunos de los denominados satélites helados, fueron motivos que nos impulsaron a desarrollar un protocolo experimental para evaluar la adecuación de Halomonas halodurans y Halomonas magadiensis, bacterias halófilas, junto con Bacillus pumillus como organismo control, en medios enriquecidos con MgSO4, Mg(NO3)2, (NH4)SO4 o MgCl2 en concentraciones equivalentes al valor de actividad de agua (aw) reportado como óptimo en términos de NaCl.
Los resultados obtenidos pueden explicarse en términos de la serie de Hofmeister y pueden ser utilizados para extrapolar los límites de actividad biológica terrestre hacia las condiciones del océano de los satélites Europa o Encélado o del subsuelo del planeta Marte, sitios de interés astrobiológico.
Métodos
El crecimiento de cada bacteria, en cada condición experimental, se monitoreó a través de cinéticas de crecimiento, midiendo el valor de densidad óptica (D. O.) a 630 nm en intervalos de tiempo regulares hasta observar la fase estacionaria. Se aplicó un ajuste exponencial a cada una de estas gráficas para obtener la velocidad específica de crecimiento (µ).
Con la información recolectada se elaborarón graficas que evidencian la variación en la velocidad específica de crecimiento (µ) de cada bacteria para las diferentes concentraciones y sales inorgánicas bajo estudio. Debido a que la naturaleza química de cada sal es diferente, es necesario contar con un parámetro que permita, de manera imparcial, evaluar su influencia en el crecimiento bacteriano. Dicho parámetro es la actividad de agua (aw), la cual está directamente relacionada con la concentración salina.
Una vez determinados estos valores se elaboraron gráficos que relacionan el valor de aw correspondiente a cada condición de salinidad, con las respectivas velocidades específicas de crecimiento. Resultados
Se obtuvieron gráficas de correspondencia entre los valores experimentales de actividad de agua (aw) del medio modificado y la concentración de una sal específica.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
34
Se realizaron también las gráficas de velocidad específica de crecimiento de cada bacteria en función de la aw del medio modificado. En estas gráficas se pueden comparar las velocidades específicas de crecimiento de cada bacteria para un mismo valor de aw aunque éste provenga de diferentes sales.
Conclusiones
Las bacterias bajo estudio lograron todas crecer en los medios modificados con las sales de interés astrobiológico. El crecimiento en cada caso es distinto. En algunos casos es superior a lo reportado cómo óptimo de crecimiento en NaCl y en otros casos se observan crecimientos por debajo de ese óptimo. Estas diferencias se pueden explicar en función de la serie de Hofmeister, la cual permite ubicar a algunos aniones y cationes como iones cosmotrópicos si favorecen la persistencia de estructuras celulares, o como caotrópicos si por el contrario, desestabilizan a estas estructuras. El ion sulfato por ejemplo, manifiesta claramente un carácter cosmotrópico en los ensayos realizados. Estos resultados son relevantes en el contexto de la exploración de los océanos de los satélites helados debido a la existencia de sales como el sulfato de magnesio (MgSO4) en el satélite Europa o el bicarbonato de sodio (NaHCO3) en Encélado. El hecho de que bacterias halófilas y halotolerantes terrestres tengan la capacidad de desarrollarse en entornos distintos a los enriquecidos con NaCl amplía las posibilidades del estudio de la habitabilidad de esos satélites helados.
Agradecimientos
Apoyo económico recibido de PROMEP (Proyecto Consolidación 2011)
Referencias [1] Duckworth, A. W., Grant, W. D., Jones, B. E., Meijer, D., Márquez, C., & Ventosa, A.
Halomonas magadii sp. nov., a new member of the genus Halomonas,. Extremophiles , 53–60. (2000).
[2] González, J. C., & Peña, A. Estrategias de adaptación de microorganismos halófilos y Debaryomyces hansenii (Levadura halófila). Revista Latinoamericana de Microbiologìa , 137-156. (2002).
[3] Grant, W. D. Life at low water activity. The Royal Society , 1249-1267. (2004). [4] Krämer, R. BActerial Stimulus Perception and Signal Transduction: Response to Osmotic
Stress. The Chemical Record , 217-229. [5] Oren, A. Microbial life at high salt concentrations: phylogenetic and metabolic diversity. Saline
Systems. (2008). [6] Sleator, R. D., & Hill, C. Bacterial Osmoadaptation: the role of osmolytes in bacterial stress and
virulence. FEMS (Microbiology Reviews) , 49-71. (2001). [7] Ventosa, A., Nieto, J. J., & Oren, A. Biology of Moderately Halophilic Aerobic Bacteria.
Microbiology and Molecular Biology Reviews , 504-544. (1998).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
35
Presentación Oral
EL REGISTRO GEOLÓGICO DEL HADEANO Y LA EMERGENCIA POSIBLE DE LA PRIMERA VIDA EN EL SISTEMA SOLAR
Fernando Ortega Gutiérrez, Instituto de Geología, UNAM; SIOV, UNAM
Los 400 millones de años transcurridos entre la primera evidencia en la Tierra de océanos de agua líquida [1] hace 4,300 millones de años (Ma) y el final del gran bombardeo tardío (GBT), están prácticamente excluidos del registro geológico conocido. Sin embargo, diversas consideraciones teóricas indican que ese periodo pudo ser lo suficientemente tranquilo como para permitir el enfriamiento del planeta después de que fue fundido y devolatizado por el evento que formó la Luna hace 4,500 Ma. ¿Pero qué tan firme es la evidencia de la presencia de tales océanos?, ¿y permitieron las condiciones físicas de ese tiempo que el agua permaneciera líquida por millones de años donde la vida pudiera surgir? En caso de que así hubiese sido, ¿pudo la vida trascender los afectos catastróficos del GBT, el cual se extendió [2] entre 4,000 y 3,900 Ma?
Justo después del impacto que formó la Luna depositando una energía de ~5 x 1030 calorías (suficiente para fundir todo el planeta), los elementos volátiles adquiridos con anterioridad (gases principalmente) habrían sido por completo erradicados de las capas principales de la Tierra, y probablemente se necesitaron millones de años para refrigerar su superficie por debajo de los 100º C. La atmósfera perdida tendría entonces que haber sido renovada a partir de cuatro fuentes posibles: cometas, asteroides, volcanismo y recondensación. Las diferencias entre las relaciones deuterio/hidrógeno (D/H) de la mayoría de los cometas estudiados hasta ahora (3 x10-4) y el agua de la Tierra (Vienna Standard Mean Ocean Water, 1.50 x 10-4) no apoyan una fuente en esos cuerpos, pero si una fuente asteroidal, cuyas relaciones D/H (meteoritas), son iguales a las del agua terrestre. Sin embargo, si esta agua hubiese sido reintroducida por impacto de cuerpos exteriores durante la época post lunar y previa al GBT, ese tiempo (400 Ma) no habría sido en realidad tan tranquilo, comprometiendo la gestación de la primera vida durante el Hadeano (4500-4000 Ma). Los materiales fechados más antiguos en la Tierra (4,404 ± 8 Ma) [3] son zircones (ZrSiO4) detríticos del conglomerado Arqueano Jack Hills en el cratón Pilbara del oeste de Australia, los cuales aparentemente cristalizaron en magmas saturados de agua a temperaturas relativamente bajas y derivados de la fusión de un manto primitivo, indicando que nada o muy poca corteza continental (granítica) se había formado entonces. Si la vida surgió durante esa época, ésta se habría asociado más fácilmente con procesos hidrotermales en los fondos basálticos de los océanos más primitivos. El Gneiss Acasta del NW de Canadá contiene rocas originalmente ígneas (metatonalitas) hasta de 4,031 Ma de edad [4], y rocas metavolcánicas mas jóvenes en la misma secuencia contiene zircones heredados de hasta 4,200 Ma, aunque hasta a la fecha no se han encontrado vestigios de sedimentos en el registro geológico del Hadeano. El GBT depositó en la Tierra 3-5 x 1020 kg de masa y una energía de 1028-29 J, lo cual fue dos veces mayor por unidad de superficie que la depositada en la Luna y probablemente el fenómeno duró más tiempo [5]. Se ha propuesto que toda la superficie terrestre se cubrió de cráteres y cuencas de impacto, 40 de los cuales tuvieron diámetros superiores a los 1,000 km y varios impactos del GBT

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
36
produjeron cuencas con diámetros mayores que los 5,000 km, implicando eventos esterilizantes de toda aquella forma de vida que se hubiese podido desarrollar antes del GBT. Sin embargo, el mismo fenómeno poco después de cada catástrofe pudo haber creado las condiciones idóneas (energía, nutrientes y movilidad química) para el resurgimiento o aparición de la primera vida en la Tierra al terminar el Hadeano. La coincidencia temporal extraordinaria que hay entre los 3,850 Ma cuando se han propuesto los primeros vestigios (isotópicos) de vida en la Tierra (6) y el final o pico del GBT, puede interpretarse de dos maneras igualmente relevantes: 1. El nacimiento o renacimiento de la vida en nuestro planeta fue retardado hasta que la intensidad del GBT declinó hace 3,850 Ma, o bien el GBT fue de hecho el factor crítico que hizo posible el surgimiento de la primera vida en la Tierra y probablemente en todo el Sistema Solar. Referencias
[1] S.J. Mojzsis, T.M., Harrison and R.T. Pidgeon, Nature 249, 178-181 (2001). [2] D.A. Kring, , Jour. Geophys. Res. 107, 10.10.29/2001JE001529 (2002) [3] S.A. Wilde, J.W. Valley, W.H. Peck, and C. M. Graham., Nature 249, 175-178 (2001) [4] S.A. Bowring, and I.S., Williams, 1999, Contrib. Miner. Petrol. 134, 3–16 (1999). [5] U. G. Jørgensen, et al., Icarus 204, 368-380 (2009) [6] S. J. Mojzsis et al., Nature 384, 55-59 (1996).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
37
Presentación Oral
ROLL-UPS: ESTRUCTURAS SEDIMENTARIAS INDUCIDAS POR MICROBIOS Y LA BÚSQUEDA DE VIDA EN MARTE
Hugo Beraldi, SIOV1, Instituto de Geología, UNAM
Una manera que tienen los microbios de dejar su huella en el registro geológico es en la forma de estructuras sedimentarias de origen químico y detrítico, como los estromatolitos y las MISS2 respectivamente. Sin embargo, algunos procesos abióticos pueden formar estructuras muy similares a las biogénicas. Por ello, es necesario caracterizar a las estructuras sedimentarias, más allá de su sola morfología, para encontrar caracteres diagnósticos que puedan diferenciar lo biótico de lo abiótico. Estas características diagnósticas son llamadas ‘biofirmas’ y son necesarias para probar la existencia, presente o pasada, de vida basada en carbono y agua, en un ambiente determinado, en la Tierra o fuera de ella. Si conocemos los procesos microbianos involucrados en la formación de una estructura sedimentaria, entonces la expresión macroscópica de tal estructura puede ser reconocida como biofirma. Las estructuras sedimentarias más valiosas como biofirmas para la exploración planetaria, son aquellas que pueden reconocerse en la escala macroscópica (de centímetros a metros), sobre todo porque operadores de rovers en misiones exploratorias distantes, pueden tomar decisiones sobre dónde acercarse a hacer análisis, lo cual implica grandes inversiones de tiempo, dinero, esfuerzo, y riesgos. Pocas estructuras sedimentarias pueden ser reconocidas como verdaderas biofirmas3.
Los ‘roll-ups’ terrestres [2] son estructuras sedimentarias macroscópicas que se forman en sedimentos superficiales infestados de microbios, únicamente a través de procesos de humedad-desecación. De no estar presentes los microbios, pueden formarse ‘rollos de lodo’ (mud curls), pero éstos no se enrollan totalmente y no resisten el transporte por agua. Sólo los roll-ups biogénicos, que son sostenidos internamente por la cohesión de polisacáridos cementantes y un entramado de filamentos que mantienen a las partículas minerales en su sitio, pueden resistir la erosión y ser sepultados (implica potencial fosilización). Roll-ups fósiles se han encontrado en depósitos de inter-duna, en rocas siliciclásticas de hasta 1800 Ma de antigüedad.
Los roll-ups son entonces buenos candidatos para la búsqueda de biofirmas en Marte o cualquier otro planeta, dado que los microbios son esenciales en su formación. Sin embargo, los roll-ups requieren de un mínimo de condiciones para formarse: a) condiciones habitables en el planeta, b) depositación de sedimento fino, c) condiciones favorables para la colonización de los sedimentos, d) microbios filamentosos como agentes bióticos (los microbios no-filamentosos difícilmente pueden crear un entramado), y e) ciclos de humedad-desecación que afectan la reología del sedimento. En la Tierra, microbios filamentosos pueden rastrearse en rocas de 3500 Ma, sugiriendo que morfologías filamentosas pueden ser una forma exitosa y estable de vida (por si acaso surgió la vida en Marte). Por otro lado, la depositación de sedimento fino puede ocurrir en una gran variedad de ambientes 1 Seminario Interdisciplinario sobre Origen de la Vida 2 Microbially Induced Sedimentary Structures [1]. 3 Por ejemplo, estromatolitos dendrolíticos (muy ramificados) o roll-ups.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
38
sedimentarios, y ciclos de humedad-desecación pueden operar mientras el agua líquida sea estable en la superficie.
Especulando que el Marte temprano tuvo condiciones favorables para la vida, las condiciones necesarias para la formación de roll-ups se vislumbran plausibles. Los roll-ups deberían buscarse en aquellos ambientes marcianos, donde tales condiciones puedan ser reconocidas. Referencias
[1] Noffke N. 2010, Microbial mats in sandy deposits from the Archean era to today, en Microbially Induced Sedimentary Structures, Springer-Verlag Berlin. 200p.
[2] Beraldi-Campesi y Garcia-Pichel, 2011. Biogenicity of roll-up structures and their potential as biosignatures of ancient life on land. Geobiology. 9: 10-23

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
39
Presentación Oral
¿CUÁL ES EL PAPEL DE LA SELECCIÓN NATURAL EN EL ORIGEN DE NUEVAS FUNCIONES PROTEICAS?
ESTUDIO TEÓRICO DE LA EVOLUCIÓN MOLECULAR DE LAS PROTEÍNAS
L.A.Q.B. Zurisadai Miguel Muñoz González, Centro de Investigación y de Estudios Avanzados (CINVESTAV) Unidad Irapuato, Guanajuato.
[email protected] Dr. Luis Jośe Delaye Arredondo, Departamento de Ingeniería Genética
Dr. Alexander de Luna Fors, Laboratorio Nacional de Genómica para la Biodiversidad (LANGEBIO).
Introducción
La mutación es la fuente última de variación genética, materia prima para la evolución, sobre la cual actúan diversos mecanismos evolutivos fijadores de caracteres hereditarios en las poblaciones naturales. El proceso de las duplicaciones génicas seguidas de sustituciones nucleotídicas sinónimas y no-sinónimas en secuencias codificantes representa uno de los principales motores de las innovaciones bioquímicas funcionales en los genomas eucariontes. En este contexto nos interesa conocer el papel que ha tenido el mecanismo darwinista de la selección natural en el origen, evolución molecular y destino funcional de proteínas parálogas codificadas en el genoma del organismo modelo Saccharomyces cerevisiae.
La selección natural es el mecanismo evolutivo responsable de las adaptaciones. A nivel poblacional puede cambiar las frecuencias alélicas a través de las generaciones.1 Para detectar la acción de la selección natural a nivel ecológico, se tendría que cuantificar intrapoblacionalmente la relación existente entre determinados caracteres fenotípicos y su impacto en el éxito reproductivo y de supervivencia de los organismos que portan dicha característica.2 Sin embargo, para detectar la selección natural pretérita es necesario buscar su huella en el actual material genético. Es decir, la cantidad de diferencias entre genomas y las frecuencias de las distintas variantes alélicas. De esta forma es posible detectar ejemplos genómicos de evolución adaptativa incluso desconociendo la adaptación específica que ha evolucionado. Haciendo uso de herramientas bioinformáticas y bioestadísticas en la investigación genómica, es ahora posible conocer las bases moleculares de la selección natural darwinista.3
Detección bioinformática de la selección natural a nivel de linajes
El análisis de selección natural por ramas en las filogenias correspondientes de los genes parálogos se realizó utilizando el programa PAML4, aplicando el diseño de una serie de pruebas de rechazo de hipótesis estadísticas (LRT, prueba de razón de verosimilitud) para detectar diferencias significativas en los valores de verosimilitud asociados con las siguientes hipótesis:
H0: Ω0 = Ωa = Ωb H1: Ω0 ≠ Ωa = Ωb H2: Ω0 ≠ Ωa ≠ Ωb

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
40
Donde Ωa y Ωb corresponden al cociente entre tasas de sustitución sinónimas con respecto a no-sinónimas (dN/dS) de las dos ramas emergentes a partir del evento evolutivo de duplicación genómica ancestral en los ascomicetos (levaduras), y Ω0 al dN/dS del resto de la filogenia. Las hipótesis: H0, H1 y H2, y sus respectivas pruebas de contraste, implementan estadísticamente la hipótesis evolutiva según la cual ocurre un cambio en las tasas de evolución molecular de los genes parálogos resultantes de una duplicación génica, inmediatamente posterior al mismo, debido a una relajación de la presión de selección negativa y/o a la acción de verdadera selección positiva, dando como resultado procesos de divergencia funcional (neofuncionalización y subfuncionalización).
Para la realización de este estudio bioinformático se utilizó la información contenida en la base de datos “Fungal Othogroups Repository”5, que contiene las secuencias genómicas de 23 hongos ascomicetos. Además, la selección específica de los 7 pares de parálogos y ortogrupos correspondientes se hizo con base a los criterios de divergencia fenotípica en las secuencias codificantes que se encontraron experimentalmente por los investigadores del Laboratorio de Biología de Sistemas Genéticos (Ascencio, en preparación).
Resultados
Los resultados de las estimaciones por máxima verosimilitud y sus pruebas estadísticas de contraste de hipótesis se pueden consultar en las tablas 1 y 2. Conclusiones Conforme a los resultados obtenidos se concluye que la presión de selección natural se modifica como consecuencia del evento de duplicación genómica ancestral en 5 de los 7 pares de genes parálogos analizados. El incremento relativo en el índice omega de las ramas posteriores a la duplicación génica puede estar asociado con relajación de la selección negativa o selección positiva genuina, posiblemente relacionado con la divergencia funcional de estos genes y sus proteínas codificadas. Agradecimientos Agradecemos a los miembros del Laboratorio de Biología de Sistemas Genéticos, en especial a la M. en C. Diana Itzel Ascencio Sánchez por su valiosa colaboración científica, y del Grupo de Genómica Evolutiva por todo su apoyo, así como a la Química María Cristina Ortega Moo por su asesoría y ayuda en la redacción del presente escrito. Referencias
[1] A. Castillo, in Ecología Molecular, E. L. Eguiarte, V. Souza, X. Aguirre, Eds. (SEMARNAT, 2007), chap. 1.
[2] F. Perfectti, F.X. Picó, J.M. Gómez, Ecosistemas, 18(1),10-16 (2009). [3] J. Bertranpetit, SEBBM, 160, 12 (2009). [4] Z. Yang, Mol. Biol. Evol. 24(8),1586-1592 (2007). [5] I. Wapinski, A. Pfeffer, N. Friedman, A. Regev, Nat. 449(7158),54-61 (2007).
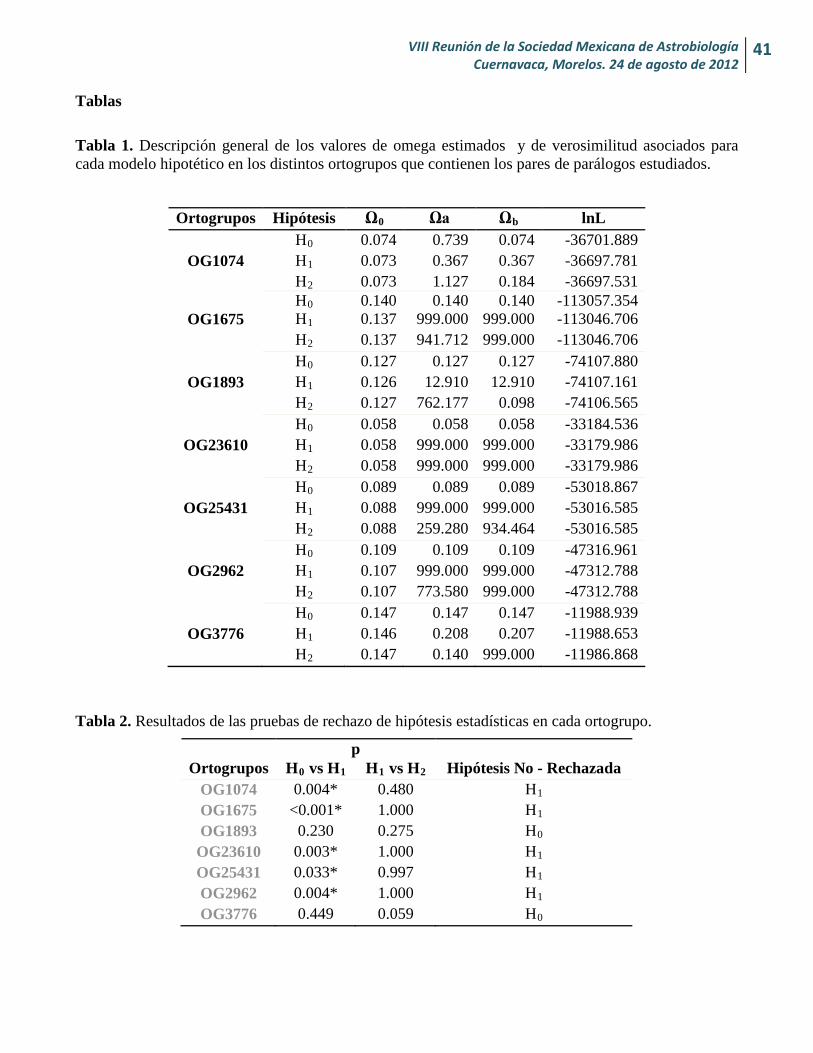
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
41
Tablas Tabla 1. Descripción general de los valores de omega estimados y de verosimilitud asociados para cada modelo hipotético en los distintos ortogrupos que contienen los pares de parálogos estudiados.
Ortogrupos Hipótesis Ω0 Ωa Ωb lnL H0 0.074 0.739 0.074 -36701.889
OG1074 H1 0.073 0.367 0.367 -36697.781 H2 0.073 1.127 0.184 -36697.531 H0 0.140 0.140 0.140 -113057.354
OG1675 H1 0.137 999.000 999.000 -113046.706 H2 0.137 941.712 999.000 -113046.706 H0 0.127 0.127 0.127 -74107.880
OG1893 H1 0.126 12.910 12.910 -74107.161 H2 0.127 762.177 0.098 -74106.565 H0 0.058 0.058 0.058 -33184.536
OG23610 H1 0.058 999.000 999.000 -33179.986 H2 0.058 999.000 999.000 -33179.986 H0 0.089 0.089 0.089 -53018.867
OG25431 H1 0.088 999.000 999.000 -53016.585 H2 0.088 259.280 934.464 -53016.585 H0 0.109 0.109 0.109 -47316.961
OG2962 H1 0.107 999.000 999.000 -47312.788 H2 0.107 773.580 999.000 -47312.788 H0 0.147 0.147 0.147 -11988.939
OG3776 H1 0.146 0.208 0.207 -11988.653 H2 0.147 0.140 999.000 -11986.868
Tabla 2. Resultados de las pruebas de rechazo de hipótesis estadísticas en cada ortogrupo.
p Ortogrupos H0 vs H1 H1 vs H2 Hipótesis No - Rechazada
OG1074 0.004* 0.480 H1 OG1675 <0.001* 1.000 H1 OG1893 0.230 0.275 H0 OG23610 0.003* 1.000 H1 OG25431 0.033* 0.997 H1 OG2962 0.004* 1.000 H1 OG3776 0.449 0.059 H0

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
42
PRESENTACIONES EN CARTEL

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
43
Presentación en cartel: ASTROBIOLOGÍA EN MEXICO
PROPUESTA DE LA SOCIEDAD MEXICANA DE ASTROBIOLOGÍA PARA CONVERTIRSE EN AFILIADO INTERNACIONAL DEL INSTITUTO DE
ASTROBIOLOGÍA DE LA NASA
Mesa Directiva, Sociedad Mexicana de Astrobiología (SOMA) [email protected]
Antígona Segura, Presidente de SOMA, Instituto de Ciencias Nucleares, UNAM Sandra I. Ramírez Jiménez, Vicepresidente SOMA, Centro de Investigaciones Químicas, UAEM
Irma Lozada-Chávez, Secretaria de difusión de SOMA, Universidad de Leipzig, Alemania
Introducción El próximo 13 de septiembre del año en curso se presentará ante el Comité Ejecutivo del Instituto
de Astrobiología de la NASA (NAI por sus siglas en inglés) la propuesta para que SOMA se convierta en un afiliado internacional de dicho instituto. El Instituto de Astrobiología de la NASA (http://astrobiology.nasa.gov/nai/) es una institución virtual que apoya económicamente a grupos de investigación (equipos) para realizar proyectos específicos. Estos equipos deben presentar una propuesta de investigación para pertenecer al NAI durante 5 años. Además, el NAI cuenta con miembros y afiliados internacionales. Los miembros internacionales se establecen a partir de convenios con el gobierno de un país, este es el caso del Centro de Astrobiología (España). Los afiliados internacionales son organizaciones que pueden ser gubernamentales, académicas, privadas o no lucrativas. Estas organizaciones establecen proyectos a largo plazo y deben presentar una propuesta ante el Comité Ejecutivo del NAI para convertirse en afiliados internacionales. El Comité Ejecutivo del NAI está conformado por el director del Programa de Astrobiología de la NASA, el director del NAI, los investigadores principales de cada equipo de investigación y un representante por cada institución u organización internacional.
Siendo SOMA una organización no lucrativa, la propuesta pretende que ésta sea incluida como Afiliado Internacional del NAI. De ser aceptada, la propuesta le permitirá a la Sociedad coordinar esfuerzos con el NAI para incidir en tres aspectos que generarán beneficios para ambos países, pero principalmente para México:
• Investigación en astrobiología. • Formación de recursos humanos para la astrobiología. • Educación y divulgación de la astrobiología.
La propuesta
En la propuesta se presenta un resumen de lo que es la Sociedad Mexicana de Astrobiología y las actividades que la Sociedad ha realizado en los últimos años, las investigaciones que actualmente se llevan a cabo por parte de sus miembros y las colaboraciones que algunos de ellos tienen con grupos de investigación que forman parte del NAI. La propuesta se divide en tres secciones principales: actividades a corto plazo, recursos y actividades a largo plazo.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
44
A. Actividades a corto plazo Investigaciones en astrobiología
Se propone generar una base de datos que sirva como referencia a los investigadores nacionales e internacionales para conocer las investigaciones relacionadas con la astrobiología que se realizan en México. La base de datos incluirá oportunidades de colaboración en diversas áreas de investigación relacionadas a la astrobiología y en aquellos sitios que son actualmente de interés para la astrobiología en México. En un apéndice de la propuesta se presenta brevemente la descripción de algunos de estos sitios, tales como: la cuenca de Cuatro Ciénagas en Coahuila, la cuenca de Guaymas en Baja California, el lago de Alchichica en Puebla, los estanques salinos de Guerrero Negro en Baja California, el cenote “El Zacatón” en Tamaulipas, yacimiento geotérmico “Los azufres” en Michoacán, las cuevas de Villa Luz y Luna Azufre en Tabasco, la laguna de Bacalar en Quintana Roo, el lago en el cráter en el Rincón de Parangueo en Guanajuato, y la laguna de Figueroa en Baja California.
Formación de recursos humanos para la astrobiología
Esta parte de la propuesta está dirigida a apoyar a los estudiantes de posgrado e investigadores posdoctorales para que participen en actividades académicas como escuelas, talleres y estancias que sean organizadas por instituciones internacionales. Este apoyo se dará a partir de recursos económicos que se gestionarán con CONACyT. Por lo que se establecerá un mayor contacto con investigadores que pertenecen al NAI.
Además, SOMA colaborará en la organización de un seminario hispanoamericano de astrobiología y la escuela hispanoamericana de astrobiología. Estas actividades se realizarán en conjunto con los afiliados internacionales del NAI de habla hispana: el Centro de Astrobiología (España), el Instituto de Astrobiología (Colombia) y la Unidad de Investigación en Astrobiología de Sao Paulo (Brasil). Educación y divulgación de la astrobiología
SOMA ha generado diversos materiales para la divulgación de la astrobiología entre diversos públicos. Estos materiales serán compartidos con el NAI para ser difundidos entre los hispano parlantes en los Estados Unidos y en los sitios web en español generados por el NAI.
Se propone crear nuevos materiales, en español e inglés, con temas relacionados con la investigación en astrobiología que se realiza en México. También se creará un sitio web en español dirigido a jóvenes entre 12 y 18 años con temas clave para la astrobiología. Este esfuerzo se realizará en colaboración con caricaturistas, comunicadores de la ciencia e investigadores.
Como parte de las actividades del grupo del NAI de la Arizona State University, se han creado una serie de “viajes de campo virtuales”. Estos “viajes” muestran fotos panorámicas e interactivas de un sitio de interés astrobiológico y contienen información y actividades dirigidas a desarrollar un tema específico para estudiantes de licenciaturas relacionadas con la astrobiología (biología, química, física, ciencias de la Tierra). En http://vft.asu.edu/ se pueden observar ejemplos de “viajes de campo virtuales” realizados para Australia. Como parte de la propuesta, SOMA plantea generar un viaje virtual a las pozas de Cuatro Ciénegas (Coahuila, México) en colaboración con la Dra. Valeria Souza (Instituto de Ecología, UNAM) y la Dra. Janet Siefert (Rice University, Texas, Virtual Planetary Laboratory y Arizona State University). B. Recursos
Para llevar a cabo las actividades propuestas SOMA cuenta con: • Sitios web albergados en el Instituto de Ciencias Nucleares, UNAM. • Recursos humanos para la organización de eventos académicos.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
45
• Sitios relevantes para la astrobiología. • Materiales educativos y de divulgación. • Recursos para la realización de eventos académicos (p.e. auditorios, salas de
videoconferencia). Se establecerán lazos con el CONACyT para disponer de recursos económicos en la organización
de las siguientes actividades: • Organización de la Reunión Bianual de SOMA. • Organización de la Escuela Bianual Mexicana de Astrobiología. • Apoyar a los estudiantes de posgrado e investigadores posdoctorales para asistir a
actividades internacionales como escuelas, talleres y congresos. • Apoyar la generación de viajes virtuales para Cuatro Ciénegas y otros sitios de interés
astrobiológico. C. Actividades a largo plazo Investigación
• Fortalecer y promover la colaboración entre investigadores que trabajan en México y miembros del NAI.
• Obtener recursos para un programa regular de viajes de campo a sitios de interés astrobiológico en México.
Formación de recursos humanos • Establecer programas para la realización de estancias de investigación para trabajar con los
equipos del NAI y en instituciones mexicanas. • Colaborar con el NAI e instituciones académicas en México para crear un programa de
posgrado en astrobiología Educación y divulgación de la ciencia
• Crear “viajes de campo virtuales” para los sitios en México de interés para la astrobiología. • Traducir al español materiales educativos y de divulgación generados por el NAI para ser
usados en países de habla hispana y para la comunidad hispano parlante de los EU. • Colaborar con el NAI para generar materiales impresos y recursos electrónicos para la
divulgación de la astrobiología entre diferentes púbicos. Agradecimientos
La Mesa Directiva agradece a los investigadores que colaboraron en la conformación de la propuesta (por orden alfabético): Hugo Beraldi-Campesi (Instituto de Geología, UNAM), Thomas Buhse (Centro de Investigaciones Químicas, UAEM), Elizabeth Chacón-Baca (Facultad de Ciencias de la Tierra, UANL), Luis Delaye Arredondo (CINVESTAV, Irapuato), Carlos del Burgo (Instituto Nacional de Óptica, Electrónica y Astrofísica, INAOE), Eduardo de la Fuente-Acosta (Universidad de Guadalajara), Elva Escobar-Briones (Instituto de Ciencias del Mar y Limnología, UNAM), Dolores Maravilla-Meza (Instituto de Geofísica, UNAM), Fernando Ortega-Gutiérrez (Instituto de Geología, UNAM), Valeria F. Souza Saldívar (Instituto de Ecología, UNAM), Pedro Valdés-Sada (Universidad de Monterrey), Roberto Vázquez-Meza (Instituto de Astronomía, Ensenada, UNAM).
Agradecemos especialmente a Cristina Bojórquez Espinosa (Vancouver Community College, Canada) y Shawn Domagal-Goldman (NASA Planetary Science Division) por su ayuda con la revisión del documento en su versión inglesa.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
46
Presentación en cartel: ASTROFÍSICA
ESTUDIO MOLECULAR DE CO EN GALAXIAS CERCANAS
Villicaña Pedraza I., C. de Astrobiología-NASA Asociated, Instituto de Técnica Aeroespacial. [email protected]
Martin-Pintado J., C. de Astrobiología-NASA Asociated, Instituto de Técnica Aeroespacial
Introducción En este trabajo se estudia el medio interestelar y la posibilidad de que ciertas regiones den origen a algún tipo de vida incluyendo organismos extremófilos. Usamos la molécula de CO como herramienta para el estudio de nubes de H molecular que ayuda a la formación de estrellas y planetas. Antecedentes
En un primer estudio (1) de líneas moleculares en una fuente extragaláctica – NGC 253 cubriendo de 129.1 a 175.2 Ghz (2mm) y han obtenido la densidad de columna de diferentes moléculas. Ellos obtuvieron 111 características espectrales como transiciones de 25 especies moleculares diferentes, 8 de las cuales fueron detectadas por primera vez en el MI. Por otro lado, un estudio milimétrico (2) de NGC 4945 en el rango de 85-354 Ghz y han incluido 80 transiciones de 19 moléculas, además de incluir estudio en 1.3mm. Finalmente, en 2010 un grupo estudio ARP 220 y presentaron un estudio de líneas espectrales cubriendo un rango de 40GHz entre 202 y 242 GHz (3). Adicionalmente se han realizado estudios del comportamiento del gas en el centro galáctico usando cocientes isotópicos (4).
La observación de varias líneas moleculares con resolución angular similar nos permite describir la complejidad química de la fuente en estudio, la cual nos da información fundamental de los procesos químicos de calentamiento del medio de regiones muy oscurecidas.
Descripción de la muestra estudiada
En este trabajo se presenta el análisis molecular de CO y sus isótopos C13O, CO17, CO18, C13O17, C13O18 derivado de los espectros obtenidos para tres galaxias cercanas que son NGC 253, NGC 4945 y ARP 220 observados con el telescopio APEX (Atacama Pathfinder Experiment telescope) que se encuentra a una elevación de 5100m en Chajnantor plateau en la región de Atacama Chile. APEX es un telescopio de 12m de diámetro que opera en longitudes de onda milimétrica y submilimétrica (entre IR y radio) y es el telescopio más grande submilimétrico que opera en el hemisferio sur, APEX es un telescopio prototipo para ALMA. La galaxia NGC 253
NGC 253 es una galaxia SAB(s)cd con núcleo Seyfert, es una de las fuentes más brillantes con mayor número de líneas moleculares extragalácticas, se encuentra en la constelación de Sculptor. Se encuentra a una distancia de 3Mpc. La región nuclear de NGC 253 contiene una de las fuentes IRAS extragalácticas (IR) más brillantes con un flujo de 1860 Jy.
La luminosidad de esta galaxia es LIR =2.1x1010 L⊙ y la mayoría de esta es luminosidad provienente de regiones de formación de estrellas masivas intensa en su centro (cientos de pársecs)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
47
teniendo una SFRIR =3.6 M⊙/yr. Formación violenta de estrellas es revelada por la alta tasa de supernovas de 0.05-0.3 /yr en la región nuclear de esta galaxia. La alta actividad de formación de estrellas nuclear es manejada por la gran cantidad de gas molecular en su centro (1.3-2.6 x109 M⊙) .
La galaxia NGC 4945
NGC 4945 Es una galaxia SB(s)cd o SAB(s)cd con núcleo Seyfert 2, es una de las tres fuentes más brillantes en IR (IRAS) con 686 Jy, se encuentra en la constelación de Centauro, a una distancia de 3.9Mpc. Fue en esta galaxia donde se encontró el primer mega máser detectado en la línea de vapor de agua a 22GHz. Se encuentra localizada en AR=13h05m27.279s y DEC= –49º28′04.4″ y un z=0.001908.
La galaxia ARP 220
Arp 220 es la galaxia ultra luminosa en IR (ULIRG) más cercana. Esta galaxia es un sistema de colisión galáctica en la que se ha observado el doble núcleo en longitudes de onda en submilimétrico e IR cercano. Esos núcleos están rodeados por dos discos de gas. Estudios en IR medio de ULIRGs con ISO sugieren que del 70%-80% de la población es alimentada por formación de estrellas mientras del 20%-30% son por AGN. Se encuentra localizada en AR=15h34m57.272s, DEC= +23º30′10.48″, z=0.018116. Métodos
Se han obtenido 54 espectros para la galaxia NGC 253, 46 espectros para NGC 4945 y 34 espectros para ARP 220 de las observaciones realizadas con APEX en la banda de radio con rangos que van de 280-370 GHz para NGC 253, 270-375 GHz para NGC 4945 y 275-370 GHz para ARP 220 GHz.
A partir de los espectros que se han obtenido se ha empleado un programa llamado MADCUBA para identificación de las líneas moleculares, se ha sacado un espectro promedio para cada galaxia, se realizaron ajustes gausianos para CO y sus isótopos obteniendo el área, ancho, velocidad, intensidad y energía mínima. Para el análisis de los isótopos dobles de CO se ha empleado el límite superior del área
3 (3 ) / ( / )A V V vσ σ δ= ∆ ∆ . Donde δv es la separación entre canales y ΔV es el ancho de línea. Una vez obtenidos los parámetros básicos se ha calculado la densidad de columna para CO y sus isótopos para lo cual se debió sacar la función de partición para el caso especifico de temperatura e isótopo y con lo obtenido de la densidad de columna se obtuvieron cocientes de las líneas moleculares. Referencias
[1] S. Martín et al. ApJS. 164, 450 (2006). [2] M. Wang et al. A&A. 422, 883-905 (2004) [3] S. Martín et al. A&A., enviado (disponible en http://arxiv.org/abs/1012.3753v2). [4] D. Riquelme et al. A&A., enviado (disponible en http://arxiv.org/abs/1008.1873v1)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
48
Presentación en cartel: GEOFÍSICA
DETECCIÓN DE METEOROS Y ESTUDIO DE LAS PROPIEDADES FÍSICAS DE LOS METEOROIDES ASOCIADOS
María Guadalupe Cordero Tercero, Depto. Cs. Espaciales, Instituto de Geofísica, UNAM [email protected]
Todos los años cae a la Tierra del orden de 40,000 toneladas de material interplanetario. La mayoría de este material, cuyo origen es principalmente asteroidal y cometario, tiene un tamaño de entre 5 μm y unos cuantos metros aunque ocasionalmente caen objetos de dimensiones mayores. Los objetos mayores a 50 μm suelen producir meteoros (estrellas fugaces) y bólidos cuando los cuerpos tienen dimensiones de metros. La forma en que los meteoroides se comportan cuando atraviesan la atmósfera de la Tierra o de otro planeta depende de distintas variables, entre ellas, las más importantes son su masa, la forma de su sección transversal, la velocidad de entrada, el ángulo de entrada y su densidad. De acuerdo con algunas observaciones y simulaciones, meteoroides de más de dos metros pueden llegar a romperse súbitamente y sus fragmentos quedar esparcidos en un área muy grande tal como ocurrió con el meteorito Allende. Estudiar el movimiento de los meteoroides a través de su paso por la atmósfera proporciona información importante sobre su composición, resistencia, lugar de origen y área donde podría ser recuperado el meteorito, sobreviviente del paso del meteoroide por la atmósfera. En México, en los últimos 3 años se han observado al menos 3 bólidos, asociados presumiblemente a objetos de unos cuantos metros, que han causado temor en la población. Tanto por su importancia científica como por sus implicaciones sociales, es necesario monitorear la entrada de estos objetos a nuestra atmósfera. Para esto se propuso la creación de una red de cámaras de video que se encargarán de vigilar el cielo constantemente para detectar y estudiar a los meteoroides y posiblemente recuperar meteoritos. Consideramos que está red estará funcionando en la zona central de México de manera continua en un par de años.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
49
Presentación en cartel: GEOFÍSICA
METEOROIDE DEL 18 DE MAYO DEL 2012: ESTIMACIÓN DEL ÁREA DE CAÍDA
José Luis García Martínez, Seminario Interdisciplinario del Origen de la Vida, SIOV [email protected]
Introducción
Cantidades significativas de materia orgánica traída por meteoritos se han agregado a la superficie terrestre a través de su historia, y esta materia orgánica indudablemente fue relevante en la síntesis de moléculas cada vez más complejas en el ambiente de la tierra primitiva hasta el surgimiento de la vida, pues la síntesis de orgánicos en abundancia suficiente en la superficie terrestre de la época pudo haber sido inhibida por una atmósfera fundamentalmente neutra [1]. Lo anterior justifica el estudio de los meteoritos como potenciales proveedores de orgánicos a la Tierra primitiva [2], lo que a su vez justifica el esfuerzo por recuperar fragmentos de ellos durante las caídas, sobre todo cuando las circunstancias son favorables a dicho propósito, como es el caso del meteoroide del 18 de mayo pasado que ahora nos ocupa, y que es motivo del presente trabajo. Fuentes de Información Para estimar el área de caída del meteorito con la mejor precisión posible se utilizaron dos videos de dominio público, el primero tomado en Potrero del Llano, Veracruz, viendo al sureste, y el segundo, en Atizapán de Zaragoza, Estado de México, viendo hacia el noreste (Figura 1); la geometría correspondiente se muestra en la Figura 2. Información complementaria fue obtenida a partir de los testimonio de los habitantes de Martínez de la Torre, Veracruz, y de las autoridades de Protección Civil de la misma población, quienes previamente realizaron dos expediciones a el área donde los habitantes reportaron el fenómeno con mayor insistencia. Observaciones Tomando como referencia la imagen tomada en Atizapán y la configuración geométrica de ambas observaciones (Figura 2), la imagen de Potrero indica que la trayectoria seguida por el meteoroide antes del impacto no está en el plano de la imagen de Atizapán, sino que está inclinada ligeramente hacia el noreste en un ángulo Β respecto al plano de esta misma imagen, como se ilustra en la Figura 3. Cálculo del ángulo B Para calcular el ángulo B basta observar que si el desplazamiento angular A del observador en Potrero (Pt) fuera de 0o, el ángulo medido Bm de la imagen de Potrero (Figura 1a) con respecto a la vertical sería igual a B. Por otro lado, si el ángulo A del observador Pt fuera 90o, el ángulo medido Bm sería igual a 0o. Lo anterior conduce a la relación
entre los ángulos A, Bm y B. De
las Figuras 1a y 2, los valores medidos de A y Bm son 6.7º y 22.86º, respectivamente. Consecuentemente B = 24.7º.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
50
Angulo de incidencia y punto de impacto Una vez conocido el ángulo B, estimar el ángulo de incidencia C y el punto de impacto es posible bajo la siguiente observación: debido a la diferencia de altura entre Atizapán y la zona de impacto, cuando el meteoroide se pone en el horizonte de Atizapán, éste se encuentra a una altura h = 2220 m previo al sitio de impacto, y a una distancia r del punto de impacto. Así, de acuerdo a la Figura 3 se debe cumplir que , donde Co = 42.38º según la Figura 1b. De esta forma, despejando r y sustituyendo los demás valores tenemos que r = 3626 m. Usando el teorema de Pitágoras, la proyección de r sobre el piso es d = 2867 m. Esta es la longitud de la línea d de búsqueda, y está medida a partir del cruce de las líneas de visión de Potrero y de Atizapán, y se ubica cerca de la comunidad Belisario Domínguez y orientada hacia la comunidad vecina de Pabanco, ambas dentro del municipio de Papantla, Veracruz. Usando los valores de h y d y la definición de la tangente tenemos que el ángulo de incidencia C = 37.75º. La Figura 4 ilustra estos resultados. Observaciones
Debido a la incompletez del video de Potrero fue necesario hacer la aproximación “razonable” de que el meteoroide pasó cerca del cenit de Tlapcoyan, suposición fundada en la información proporcionada por el director de Protección Civil y Bomberos de Martínez de la Torre en el sentido de que la onda de choque se habría sentido con mayor intensidad en esa población que en Martínez de la Torre. Resultados
Los resultados de este análisis definen una línea de búsqueda de 2.87 km, limitada por los puntos definidos por las coordenadas (20º12’23.4”N, 97º18’07”W) y (20º13’32”N, 97º19’14”W), con máxima probabilidad en este último punto. Esta línea está limitada por las comunidades de Belisario Domínguez y Pabanco, sur del municipio de Papantla, Veracruz. Conclusiones
Si como lo indica la evidencia el objeto era un meteoroide, la mayor parte del material caído como meteoritos estará en las inmediaciones de la línea teórica de búsqueda, con los fragmentos más grandes hacia el extremo NW, y los más pequeños hacia el extremo SE de dicha línea, e incluso antes de la misma. Agradecimientos Agradezco a las siguientes personas que amablemente me proporcionaron o me facilitaron acceso a información vital para la realización del presente trabajo:
• Lic. Patricio Guevara A., Director de Protección Civil y Bomberos de Martínez de la Torre, V. • Lic. Antonio Rosas D., Coordinador de Protección Civil Regional en Martínez de la Torre, V. • Javier García Pérez., Protección Civil y Bomberos de Martínez de la Torre, V. • Lic. Guillermo Iván Murueta M., Dirección de Protección Civil y Bomberos de Atizapán, EM • Ing. José Luis Rueda, Centro de Investigación Rueda Cósmica A. C. • Misael Mata E., estudiante del Instituto Tecnológico Superior de Alamo-Temapache • Garvin Andrés H., estudiante del Instituto Tecnológico Superior de Alamo-Temapache
Referencias
[1] C. Sagan and C. Chyba, Sicence 276, 1217-1221 (1997) [2] E. Anders, Nature 342, 255–257 (1989)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
51
Figuras
Figura 1. a) video del meteoroide del 18 de mayo tomado en Potrero del Llano, Veracruz;
b) video del mismo objeto tomado en Atizapán de Zaragoza, Estado de México.
Figura 2. Configuración geométrica definida por las dos observaciones del meteoroide del 18 de mayo del presente año.
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
52
Figura 3. Esquema tridimensional que muestra la trayectoria r del meteoroide, desplazada del plano de la imagen de Atizapán en un ángulo B.
Figura 4. Línea de búsqueda de los fragmentos del meteoroide del 18 de mayo del 2012, cerca de los poblados de Pabanco, Joloapan y Vista Hermosa de Juárez, en el municipio de Papantla, Veracruz. El extremo noroccidental de la línea es el sitio más probable de impacto.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
53
Presentación en cartel: GEOFÍSICA
COMETAS Y EL ORIGEN DEL AGUA EN LA TIERRA
Saúl Alberto Villafañe Barajas, Ciencias de la Tierra, Facultad de Ciencias, UNAM [email protected]
Introducción
La vida en la Tierra, forzosamente necesita de agua, y actualmente nuestro planeta tiene cerca del 70% de su superficie cubierta por este líquido, sin embargo, no siempre fue así. La Tierra se formó hace aproximadamente 4.550 Ma en los principios del Sistema Solar [1], sin embargo, no era la única que iniciaba su formación, alrededor de ella había una inmensa congregación de fragmentos sólidos de materia, y más aun, cuerpos compuestos de gases congelados como metano, fragmentos de roca y/o metales, dióxido de carbono y sobre todo agua, que estaban continuamente impactándose con la Tierra primitiva [1]. Los cometas son sobrevivientes de una amplia gama de cuerpos formados en regiones frías de la nebulosa solar y representan el estadio más primitivo del desarrollo del sistema planetario, de ahí que sean muy valiosos e interesantes. Dado que son cuerpos formados en diferentes regiones, esto hace que difieran en la temperatura, presión, condiciones de acreción tales como la velocidad de impacto [1].
Actualmente existen básicamente tres teorías sobre el origen del agua en la Tierra [2,3,4] (Teoría Volcánica, Teoría Rocosa y Teoría Extraterrestre), sin embargo, no son absolutamente satisfactorias. Particularmente la Teoría del origen extraterrestre del agua no era bien aceptada por la comunidad científica debido principalmente a las relaciones Deuterio/Hidrogeno medidas en al menos seis cometas [4,5] (doblan la cantidad de lo que está presente en los mares terrestres). Sin embargo, la teoría volvió a tomar vida debido a los estudios de la NASA y ESA [4], durante la visita de cometas al Sistema Solar interno en la última década. Esto es sumamente importante, ya que abre la posibilidad de mejorar y complementar las teorías actuales. Cometas: reservorios de agua y procedencia
Es necesario diferenciar entre ciertos cuerpos celestes que principio podrían ser similares, como lo son los asteroides y cometas. En esencia, la principal diferencia entre los cometas y los asteroides, es que los primeros tienen actividad gaseosa debido a la presencia de material volátil cerca de su superficie que sublima por la acción de la radiación solar, mientras que los asteroides son cuerpos rocosos inactivos [1]. Los cometas son cuerpos de forma irregular que orbitan alrededor del Sol normalmente con gran excentricidad formados principalmente por agua en forma de hielo y partículas de polvo [4]. Normalmente un cometa tiene menos de 10 km de diámetro, y la mayor parte de sus vidas son cuerpos sólidos congelados [4], hasta que logran aproximarse lo suficiente al Sol y eventualmente este empieza a evaporar sus capas más externas, formando así, colas, formadas esencialmente por gas ionizado y polvo, alcanzando longitudes de miles de km [1].
Es importante conocer donde se originan los cometas, ya que su composición química debería reflejar las condiciones de presión y temperatura del medio en que se formaron [5], principalmente. Básicamente los cometas que nos interesan se forman en dos regiones [6], una llamada Cinturón de Kuiper en donde los cometas se mueven en orbitas casi circulares, análogamente al movimiento de los

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
54
planetas interiores. Esta zona se encuentra más alejada de la órbita de Neptuno, de tal manera que también se le suele llamar TNOs (objetos tras-neptunianos [6]). Debido a la lejanía a la que se encuentran (30-100 UA), la luz del sol tarda mucho en llegar y por tanto la radiación que reciben no sea suficiente para calentarlos, presentando así, temperaturas superficiales de -220°C aprox [6]. La Nube de Oort, presenta cometas y asteroides de largo periodo y resalta que sus orbitas no están en el plano de la elíptica. Podemos imaginarla como una gran nube (billones de cuerpos) que rodea el Sistema Solar, con cuerpos continuamente haciendo colisión a velocidades muy altas, a una distancia de 1.5 años luz del Sol [6]. El hecho de que en ocasiones podamos observarlos, se debe a colisiones entre ellos o con otros cuerpos y la influencia gravitacional de otros cuerpos, que alterarían su órbita lo bastante como la enviarlos al sistema solar interno.
El problema de una teoría
Si bien sabemos que los cometas tienen considerables cantidades de agua [4,5], no es tan sencillo decir que los impactos primitivos de estos cuerpos con la Tierra originaron los océanos, esto debido a las posibles características químicas que podría presentar las moléculas de agua terrestres y la de los cometa. Para relacionar el agua de los cometas con el agua terrestre, se utiliza la relación deuterio/ hidrogeno (VSMOW*4). La composición y estructura química del agua existente en los océanos debe estar ligada a la de estos cuerpos, si consideramos que esta procede de ellos [5]. Lo que conocemos como agua pesada (D2O) se encuentra en toda el agua independientemente del cuerpo o profundidad en la que este en una relación siempre constante (99.85% de H y .15 % D) [2]. Esto es el principal problema de la teoría, particularmente la relación D/H en los cometas, muestra que los cometas tienen cerca del doble de deuterio (hidrogeno más un neutrón) que el agua del océano, como indican las mediciones que se han hecho en cometas como el Hale Bopp (1998), Hyakutaque (1998), Halley (1995), todos procedentes de la Nube de Oort [5]. En un momento dado la posibilidad de que el agua viniera de cuerpos extra planetarios, hacia énfasis en los asteroides particularmente en las condritas carbonoceas ya que presentaban una relación D/H de (1.4±.1)×10-4 [5]. Pero como se mencionó los cometas, dados sus relaciones isotópicas prácticamente eran deslindados de cualquier relación con el origen del agua. Además de acuerdo a las observaciones realizadas, los cometas típicos no son tan grandes, como para proveer toda el agua del planeta [2].
En contra parte la teoría volcánica (plantea que el agua se formo en el centro de la tierra por reacciones a alta temperatura entre átomos de H y O y una consecuente desgasificación), era un tanto más aceptada aunque también tiene sus desventajas, por ejemplo, no es suficiente para explicar la vasta cantidad de agua que tenemos [2], además asumía que la Tierra se formó a partir del material del Sol, y hoy se sabe que se formaron independientemente y al mismo tiempo, además, dada la intensidad de la gravedad en ese entonces, la mayor parte del agua no se retuvo [7].
Nuevas expectativas
Recientes investigaciones sobre el Cometa 1999/S4 (linear) y 103 P/Hartley 2 [5] han abierto las posibilidades de que una fuerte cantidad de agua presente en nuestro planeta, realmente tenga un origen extraterrestre, debido a que muestran una similitud en composición y estructura química con el agua que existe actualmente en los océanos, así como presencia de deuterio (característico de estos cometas) [5].
Como se menciono, el deuterio es importante para describir el origen e historia de una masa de agua. El cometa 103P/ Hartley 2, de la familia de Júpiter [5] presenta una relación D/H de (1.61±.24)×10-4, muy similar al del agua en los océanos (1.558±.001)×10-4 [5], resultado que pone 4 Vienna Standard Mean Ocean Water(VSMOW) es un estándar que define la composición isotópica del agua.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
55
de manifiesto la posibilidad del origen extraterrestre del agua, además también mostro que el interior del mismo cometa se encontraba millones de toneladas de agua, lo cual refuerza la teoría planteada [5]. Además de que tenga relaciones isotópicas semejantes y una vasta cantidad de agua, el hecho de que pertenezca a la familia de Júpiter, en esencia, originado en el cinturón de Kuiper [5] a diferencia de los cometas anteriormente estudiados, que provenían de la nube de Oort, también tendría una gran influencia de considerar debido a las diferentes condiciones y por tanto características que presentarían los cometas [5].
Como hacen énfasis en el artículo de Hartogh et al. [5] el hecho de encontrar una similitud entre las relaciones isotópicas del cometa Hartley y el agua en la Tierra, más que resolver una pregunta genera muchas más, dado que ahora será necesario diseñar modelos que consideren distintas zonas donde se podrían generar estos cuerpos, características y composición de esas zonas, además de la influencia de otros cuerpos en el Sistemas Solar primitivo, entre muchas otras a considerar, pero en primera instancia resucita la Teoría del origen extraterrestre del agua. Conclusión
Si bien los resultados del Cometa Hartley 2 muestran una gran similitud con nuestros océanos, sería muy atrevido concluir que la procedencia del agua viene a partir de ellos, ya que hay que contestar muchas más preguntas y considerar también las otras hipótesis [2,3,4,5]. Yo creo que el agua no viene de una sola fuente, sino más bien de una serie de procesos que poco a poco lograron llenar de agua este planeta. Ahora hace falta investigar mucho más, y esperar en su momento la oportunidad de estudiar más cometas, ya que un problema importante es la estadística, pues se han analizado pocos cometas y se necesita corroborar su relación isotópica. También es necesario generar modelos, estudiar el manto de Tierra, rocas, meteoritos y todo lo que nos provea de información para poder ir respondiendo a las incógnitas que rigen este apasionante tema. Referencias
[1] A. Fernandez, Comets: Nature, dynamics, origin and cosmological relevance. (Springer-Verlag New York, 2005.).
[2] C. Tobias. Scientific American. Ask the experts, ( 21 Octubre 1999. Disponible en web: http://www.scientificamerican.com
[3] F. Fontúrbel R, Ciencia y cultura, marzo-mayo, 11, 3-9 (2004). [4] A. Surroca,. Origen de los mares, como apareció agua en la tierra. XXVII Semana de estudios del
mar, 2008. Disponible en web: http://www.asesmar.org. [5] P. Hartogh, D. Lis, D. Bockelée-Morvan, et al. Letter Research Nature, 10519, .218-220 (2011). [6] Meteorites,comets and planets . edited Andrew.M.Davis. Elsevier, 2005. Capq.25 vol 1 Comets,
D.E Brownlee university of Washington WA, USA,2005 [7] W. Waller, American Scientist, enviado (disponible en
http://www.sigmaxi.org/programs/issues/Waller.pdf)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
56
Presentación en cartel: GEOFÍSICA
RÍO TINTO Y LA POSIBLE PRESERVACIÓN DE MOLÉCULAS ORGÁNICAS EN CONDICIONES EXTREMAS
María Colín-García, Instituto de Geología, UNAM, (México)
[email protected] Basem Kanawati, Departmento de BioGeoQuímica y Analítica, Instituto de Química Ecológica Centro
Alemán de Investigación para la Salud Ambiental; Munich (Alemania) Mourad Harir, Departmento de BioGeoQuímica y Analítica, Instituto de Química Ecológica Centro
Alemán de Investigación para la Salud Ambiental; Munich (Alemania) Schmitt-Kopplin, Departmento de BioGeoQuímica y Analítica, Instituto de Química Ecológica Centro
Alemán de Investigación para la Salud Ambiental; Munich (Alemania) Ricardo Amils, Centro de Astrobiología-CSIC, Madrid (España) Víctor Parro, Centro de Astrobiología-CSIC, Madrid (España)
David Fernández-Remolar, Centro de Astrobiología-CSIC, Madrid (España)
Introducción Los biomarcadores son moléculas producidas por o relacionadas con actividades biológicas.
Mackenzie [1] los definió como los compuestos orgánicos detectados en la geosfera, cuya estructura básica sugiere una relación con un producto natural conocido. Por eso, los biomarcadores se pueden utilizar para interpretar el pasado, las condiciones geológicas, físico-químicas y biológicas que imperaron en un ambiente. La posibilidad de encontrar vida en otros lugares del Sistema Solar, o incluso el Universo se ha expandido la aplicación de biomarcadores que ahora se utilizan para los propósitos astrobiológicas [2]. Teóricamente, todas las moléculas derivadas de un organismo se podrían utilizarse como biomarcadores. Sin embargo, para los compuestos orgánicos simples, es muy difícil, y en algunos casos imposible determinar su origen biológico. Este es el caso de los monómeros como los aminoácidos y los azúcares, que han sido encontrados en los meteoritos [3, 4, 5] o se pueden sintetizar en forma abiótica. Una de las características que no puede ser reproducida abióticamente es la complejidad molecular; los biopolímeros (proteínas, polisacáridos, polinucleótidos, etc) son ejemplos de estructuras complejas [2]. En este trabajo se muestra la preservación de grandes biomoléculas portadoras de información biológica como péptidos, en los sedimentos antiguos de un ambiente extremo. Rio Tinto es considerado como un análogo marciano por tener un bajo pH y condiciones altamente oxidantes [6]. A pesar de las condiciones extremas posee una alta diversidad microbiana [7].
Método
Se recogieron muestras de los depósitos modernos y antiguos de tres terrazas diferentes, definidas como terraza joven, intermedia y antigua [6] cuyas edades van desde 2000 años a 2.1 Ma. Las muestras fueron molidas y dos diferentes fracciones fueron separadas y se analizaron como sigue: (1) una pequeña fracción (0.5 g aproximadamente) se separó para ser analizados por FTIR, y (2) 50 g se trató con una solución de PBS-EDTA solución (salina tamponada) para la extracción de la fracción de proteína.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
57
Posteriormente, los extractos se analizaron mediante técnicas espectrofotométricas y por una de las técnicas más sensibles, la Espectrometría de Masas acoplada a un ciclotrón y analizada por Transformada de Fourier (FTICR-MS) para corroborar la presencia de péptidos. Para esta última técnica (FTICR-MS), se usó un equipo Bruker (Bremen, Alemania) APEX 12 Qe equipado con un imán superconductor de 12 Tesla.
Resultados y Discusión
Las muestras fueron analizadas sin preparación previa mediante FT-IR con el fin de identificar los enlaces típicos de grupos amida: alrededor de 1690-1630 cm-1 para el estiramiento del enlace C = O, y alrededor de 3700-3500 cm-1 para el estiramiento del NH en una amida. En todas las tres muestras, el C = O estiramiento banda fue identificado. Sin embargo, la región de 3000 a 3700 cm-1 fue enmascarada por una banda muy amplia, por lo que no permitió la correcta identificación de la banda del estiramiento NH (Fig. 1).
Los análisis por métodos colorimétricos, dieron una respuesta positiva para la determinación de péptidos (método de Bradford); sin embargo, las cantidades eran muy pequeñas. Por lo tanto, es difícil considerarlos como los válidos, porque estaban muy cerca del límite de detección de la técnica.
En el caso del análisis de espectrometría de masa FTICR, los resultados mostraron una determinación positiva de péptidos. De hecho, hemos detectado en la terraza antigua la presencia tentativa de cinco diferentes péptidos con las masas: 1053.140 y 1065.466 y 1083.105 y 1086.114, y 1097.096 Da. Las intensidades decrecientes de péptidos en el tiempo sugieren que las señales no son el resultado de contaminación externa, sino las cadenas de péptidos originales conservados en los sedimentos.
De acuerdo con estos resultados, la conservación de fragmentos de proteína es factible bajo condiciones extremas (pH bajo y las condiciones de oxidación). Aunque las proteínas apenas sean capaces de sobrevivir en ambientes naturales, hay varios factores, incluyendo la naturaleza de la molécula y las condiciones ambientales, que determinan la velocidad de degradación. Cuando un organismo muere su cuerpo comienza un largo proceso de descomposición, promovido por sus propias enzimas o por otros organismos. Según Ambler y Daniel [8] las vías para la alteración de proteínas son: la hidrólisis de enlaces peptídicos, la modificación de las cadenas de aminoácidos, y racemización de los centros quirales. Se ha informado que algunos aminoácidos y péptidos podrían ser aislados de depósitos antiguos y de huesos, pero se produjeron cambios con el tiempo. De hecho, se ha sugerido también, que la asociación de proteínas con minerales puede retrasar la descomposición [9, 10].
La persistencia de un registro molecular en este hábitat ácido abre nuevas posibilidades para la búsqueda de rastros de vida a lo largo de los extensos depósitos de ácidos de Marte. Referencias
[1] A. S. Mackenzie, Applications of biological markers in petroleum geochemistry. J, D Books Ed., Adv. in Petr. Geochemistry Vol 1. (Ac Press, London. Pp. 115-214, 1984).
[2] P. Fernández Calvo, C. Näke, L.A. Rivas, M. García Villadangos, J, Gómez-Elvira, V. Parro, Planet Space Sci 54, 1612-1621(2006).
[3] J. Oró, Nature 190, 389-390(1961). [4] M.A. Sephton, G.D. Love, J.S. Watson, A.B. Verchovsky, I.P. Wright, C.E. Snape, I. Gilmour,
Geochim. Cosmochim. Ac. 68, 1385-1393 (2004). [5] S. Pizzarrello, Chem Biodivers 4, 680-693 (2007). [6] D. Fernández-Remolar, R. V. Morris, J.E. Gruener, R. Amils, A.H. Knoll. Earth and Planet. Sci.
Lett. 240, 149–167 (2005). [7] LA Amaral-Zettler, F Gómez, E Zettler, BG Keenan, et al., Nature 417, 137 (2002).
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
58
[8] R.P. Ambler, M. Daniel, Philos Trans Biol Sci 333, 381-389 (1991). [9] L.A. Pinck, F.E. Allison Science 114:130-131 (1951). [10] B. Waggoner, Enc Life Sci 12, 227-232. (2002).
Figuras
Fig. 1. Espectro IR de la muestra de la terraza intermedia. La muestra molida se analizó para la detección de enlaces tipo amida sin tratamiento previo. La banda de estiramiento C = O se aprecia cerca de 1600 cm-1. La banda alrededor de 3500-3700cm-1, correspondiente al grupo amida NH no se aprecia.
0
4
8
12
16
500150025003500Wavenumber (cm-1)
Refle
ctan
ce (%
)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
59
Presentación en cartel: GEOFÍSICA
BIOFORMAS MICROSCÓPICAS EN PIRITA SEDIMENTARIA Y SUS IMPLICACIONES ASTROBIOLÓGICAS
Susana Abigail Ángeles-Trigueros. Área Académica en Ciencias de la Tierra y Materiales.
Universidad Autónoma del Estado de Hidalgo. [email protected]
Alberto Blanco-Piñón. Área Académica en Ciencias de la Tierra y Materiales, UAEH Juan Hernández-Ávila. Área Académica en Ciencias de la Tierra y Materiales, UAEH
Fco. Javier Zavala-Díaz de la Serna. Fac. de Cs. Químicas, Universidad Aut. de Chihuahua Introducción
Microbios fósiles en pirita sedimentaria han sido reportados por varios autores [1], [2], [3] en varias partes del mundo. La pirita sedimentaria se forma durante un entierro poco profundo por la reacción de minerales de Fe detríticos con H2S, que a su vez, es producido por la reacción de sulfato intersticial disuelto por bacterias usando materia orgánica sedimentaria. Es por esto, que los hallazgos de microorganismos fósiles, especialmente en pirita sedimentaria en registro estratigráfico desde el Arqueozoico hasta el Reciente, han sido una clave importante tanto para entender condiciones físico-químicas en ambientes sedimentarios anóxicos, como para la búsqueda de vida en otros planetas donde pudieron existir condiciones similares para la formación de este mineral, por ejemplo, Marte.
En este trabajo se reportan microestructuras biomórficas en pirita sedimentaria de la Fm. Agua Nueva (Cretácico Superior) en Xilitla, S. L. P. (México), con el propósito de aumentar los reportes que existen sobre estas bioestructuras y discutir sus posibles implicaciones paleoambientales y astrobiológicas. Métodos
Muestras de pirita sedimentaria fueron colectadas en capas de la Fm. Agua Nueva en Xilitla, S. L. P. y transportadas al Área Académica de Ciencias de la Tierra y Materiales de la Universidad Autónoma del Estado de Hidalgo. En el laboratorio, se obtuvieron fragmentos de 1cc de la parte más interna de la muestra para evitar contaminación. Posteriormente, las muestras fueron cubiertas por oro durante 3 minutos y observadas en el Microscopio Electrónico de Barrido (MEB) para identificación de bioestructuras. Las estructuras de interés fueron sometidas a Espectroscopía de Energía Dispersiva (EED) para determinar su composición elemental. Resultados
En la pirita sedimentaria analizada se encontraron estructuras microscópicas que, de acuerdo a su origen, se dividieron en dos grupos: a) Estructuras biomórficas y b) Estructuras no biomórficas asociadas a actividad orgánica. Estructuras Biomórficas
Elementos cocoides (Fig. 1A): Consisten en estructuras ovales a esféricas con superficies lisas preservadas en tres dimensiones. Sus tamaños varían de entre 0.5µm a 2µm y se encuentran preservadas como elementos aislados y en agrupaciones similares a colonias de bacterias. Presentan

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
60
morfologías muy similares a las reportadas como restos mineralizados de bacterias sulfato- reductoras en rocas del Ordovícico de Canadá [4].
Bastones (Fig. 1B): consisten en estructuras alargadas y estrechas con tamaños aproximados de 1µm a 8µm y que coinciden con la descripción para morfologías microscópicas tipo bacilus [5]. Estructuras similares e interpretadas como restos de microbios han sido descritas en rocas del Proterozoico Medio de India [3].
Diatomeas (Fig.1C): consisten en bioformas con simetría bilateral con exoesqueleto o frústula alargada con una longitud aproximada de 2 µm -14 µm y diámetro aparente que varía entre 0.8 µm y 4 µm. Esta morfología es consistente con la descripción de diatomeas del orden Pennales [6], y coincide con las diatomeas reportadas en la localidad de Frying Pan Lake del Reciente de Nueva Zelanda [7].
Esferas no determinadas (Fig. 1D): En las muestras analizadas estas esferas consisten en formas redondeadas aisladas cuyos diámetros varían entre 5µm y 9µm. Al igual que las estructuras cocoides, presentan superficies lisas no ornamentadas. No biomórficas asociadas a actividad orgánica
Superficies lisas (medio extracelular) y estructuras “teeth & sockets” (Fig. 1E): Entre las muestras de pirita, se observaron extensiones compuestas por superficies totalmente lisas que en algunas ocasiones son interrumpidas por pequeños cristales o bien, por huecos cuya forma y tamaño variados sugieren que estuvieron ocupados por cristales de pirita. Estas estructuras son conocidas como “teeth & sockets” y son el resultado de la formación de fluido extracelular por la acción de microorganismos, lo que permite la formación de estas superficies lisas [8].
Tubos (Fig. 1F): En las muestras analizadas se encontraron fragmentos de estructuras cilíndricas cuya longitud varía de 5µm a 20µm y un diámetro de menos de una micra a 5µm. Estas estructuras son huecas con un diámetro interno de aproximadamente una micra. Estas estructuras son comparables con aquellas descritas como “fundas” de filamentos algales/bacteriales reemplazadas por pirita sedimentaria bajo condiciones de laboratorio [9]. Análisis de EED
Los análisis EED (Fig. 2) fueron realizados a estructuras como cocoides y “teeth & sockets” mostrando valores de carbono (C) de 76.39%Wt y 39.76%Wt respectivamente, mientras que en los cristales de pirita no asociados a estructuras biomórficas el C estuvo ausente. Esto sugiere que el C está asociado a las bioestructuras lo que permite sugerir un origen biogénico de las mismas; sin embargo, análisis elementales más precisos deben ser realizados para confirmar esta hipótesis. Conclusiones
Dentro de los lentes de pirita analizados mediante el MEB, se reportan 6 tipos de estructuras divididas en dos grupos: a) biogénicas (cocoides, bastones, diatomeas y esferas no determinadas) y b) no biogénicas asociadas a actividad orgánica (superficies lisas y teeth & sockets, y tubos. Morfológicamente, las bioestructuras son similares a estructuras microscópicas biogénicas reportadas por diversos autores en pirita sedimentaria de diferentes localidades y tiempos geológicos del mundo.
Los análisis EED de estas estructuras muestran valores de C medios a altos en las bioformas, mientras que en los cristales de pirita no asociados a estructuras biomórficas el C estuvo ausente. Esto indica que el C está asociado a las bioestructuras lo que permite sugerir un origen biogénico de las mismas. El estudio de bioestructuras en pirita sedimentaria, tal como es el caso de las formas encontradas en los lentes de pirita sedimentaria de la localidad de Xilitla, S. L. P., son clave para la prospección de vida en otros cuerpos del Sistema Solar, como por ejemplo Marte, en donde se han encontrado evidencias indirectas (presencia de óxidos de hierro cristalinos [10] así como sulfatos [11]) de la existencia de pirita en rocas sedimentarias. Esto, sugiere la presencia de sistemas acuáticos

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
61
análogos a aquellos existentes en la Tierra, en donde la pirita sedimentaria está presente y la vida microbiana prospera.
Agradecimientos
Este trabajo fue parcialmente financiado por el Fondo de Investigación para la Educación SEP-CONACYT Proyecto No. 83849.
Referencias
[1] J. M. Schopf, E. G. Ehlers, D.V. Stiles, J. D. Birle, Proc. Amer. Philos. Soc. 109, 228-308 (1965).
[2] R. L. Folk, J. Sediment. Petrol. 63, 990-999 (1993). [3] J. Schieber, Geology. 30(6), 531-534 (2002). [4] J. Schieber, PALAIOS. 17, 104-109 (2002). [5] T. D. Brock et al., Microbiología (Prentice-Hall Hispanoamericana, México, ed. 4, 1987). [6] H. Des Abbayes et al., Botánica: vegetales inferiores (Reverté, Barcelona, 1989). [7] B. Jones, R. W. Renaut, K. O. Konhauser, Sedimentology. 52, 1229-1252 (2005). [8] S. Sur, J. Schieber, S. Banerjee, J. Earth Syst. Sci. 115(1), 61-66 (2006). [9] J. Schieber, M. Glamoclija, K. Thaisen, LPSC. XXXVIII. 1626.pdf. (2007). [10] C. M. Egglestone, B. A. Parkinson, E. S. Bramlett, Goldsmith Conf. Abst., A261 (2010). [11] L. Lefticariu, L. M. Pratt, J. A. Laverne, LPSC XXXVIII. 1953.pdf. (2006).
Figuras
Fig. 1. Estructuras microscópicas en pirita sedimentaria. A) Cocoides. B) Bacilos. C) Diatomeas. D) Esfera lisa no determinada. E) Teeth & Sockets. F) Estructura tubular.
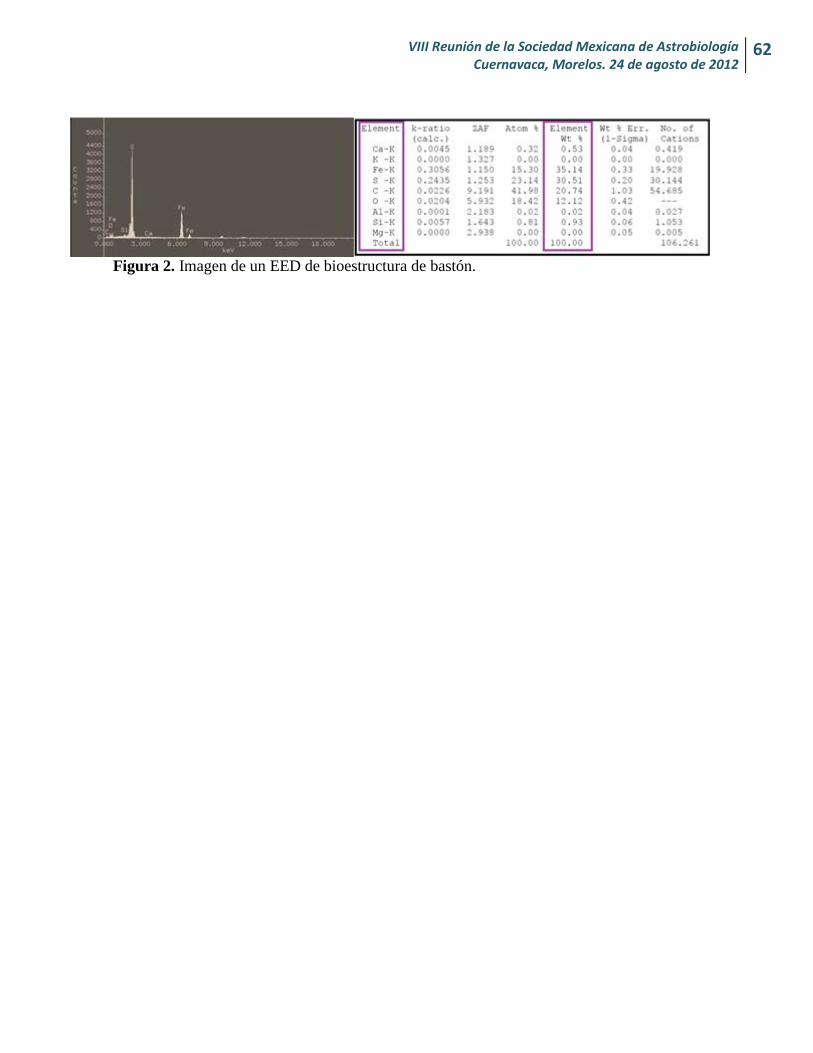
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
62
Figura 2. Imagen de un EED de bioestructura de bastón.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
63
Presentación en cartel: GEOFÍSICA
AVANCES SOBRE PANSPERMIA INVERSA
Roberto Vázquez, Instituto de Astronomía, Universidad Nacional Autónoma de México
(Ensenada) [email protected] Patricia G. Núñez, Instituto de Estudios Avanzados de Baja California, A. C.
Mauricio Reyes-Ruiz, Instituto de Astronomía, Universidad Nacional Autónoma de México Carlos E. Chávez, FIME, Universidad Autónoma de Nuevo León
Stephania Hernández, Instituto de Astronomía, Universidad Nacional Autónoma de México Héctor Aceves, Instituto de Astronomía, Universidad Nacional Autónoma de México
Introducción
Los meteoritos marcianos son fragmentos de corteza arrojados hacia el espacio exterior como resultado de grandes impactos de asteroides y cometas con Marte. Luego de salir del planeta, los meteoritos viajaron a través del espacio para posteriormente llegar a la superficie de la Tierra [1]. Del mismo modo, es válido pensar que material de la corteza terrestre, acelerado por un gran impacto, pueda resultar en la exportación de material biológico hacia otros cuerpos del Sistema Solar. Esto se conoce como “Panspermia inversa” o “Litopanspermia” [2,3]. De hecho, ya se ha hecho trabajo, tanto teórico como experimental acerca de la viabilidad de que organismos terrestres resistan tales procesos [4].
El presente proyecto [5], el cual consta de tres fases, tiene como objetivo fundamental estudiar la posibilidad de que granos de polen terrestre puedan haber sido transportados a otros planetas por medio del mecanismo de “Panspermia inversa”
Objetivos Fase I
Analizar el destino del material residual eyectado desde la Tierra después de un impacto meteorítico. Para ello realizamos simulaciones numéricas de la dinámica de una gran colección de partículas de prueba lo que nos permitió determinar la probabilidad y las condiciones necesarias para que el material eyectado pudiera alcanzar a otros planetas del Sistema Solar. Tanto la sección de RESULTADOS como la de CONCLUSIONES se refieren a esta fase del proyecto. Fase II
Estudiar los granos de polen de especies resistentes para conocer su viabilidad bajo condiciones extremas, tanto en el medio interplanetario como en la superficie de otros planetas. El grano de polen forma un polvo que puede ser fino o grueso, el cual contiene los microgametofitos de la semilla de las plantas, las cuales producen los gametos masculinos (células espermáticas). Una pared protectora (la exina) hecha de un material llamado esporopolenina, convierte al polen en una estructura muy resistente a las condiciones ambientales adversas. Se sabe que el grano de polen muestra una alta

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
64
resistencia a técnicas de laboratorio muy severas, algunas de las cuales incluyen ácidos y otras sustancias para limpiar la exina y estudiar mejor sus componentes.
A la fecha se han desarrollado modelos para la estratificación de la exina que indican que cuanto más compleja es su estructura, mayor es su resistencia a las condiciones extremas [6]. Aunque existen varios estudios referentes a la arquitectura del grano de polen y su viabilidad bajo diversos ambientes, no existen estudios astrobiológicos acerca de la viabilidad del polen en el espacio exterior ni en las atmósferas de otros planetas. Hasta este momento, nuestro equipo ha obtenido muestras de granos de polen de algunas especies resistentes las cuales se encuentran en proceso de caracterización y análisis (por ejemplo, en la Fig. 1 se muestra una fotografía de polen de Opuntia littorallis). Fase III
Estudiar las condiciones físicas relacionadas con la entrada a otro planeta del “transportador de polen”. Esta etapa del viaje interplanetario del material biológico es la menos estudiada de todas. Esto se debe a que en su ingreso de alta velocidad a la atmósfera de un planeta, el calentamiento por fricción derrite la superficie del meteorito y el flujo de viento remueve la roca líquida enfriando el remanente. Debido a que el tiempo en que el meteorito cruza la atmósfera es muy corto (unos cuantas decenas de segundos) solamente la capa superficial del mismo (del orden de 1 mm) se funde, y el interior del meteorito no se calienta significativamente [7]. De manera que esta fase del trayecto interplanetario no representa un gran peligro para el material biológico contenido en el interior del meteorito.
Para meteoritos de tamaños moderados (de centímetros a metros) como los mencionados aquí, generalmente se considera que el impacto con la superficie del planeta blanco ocurre a la velocidad terminal en la atmósfera de dicho planeta (300 m/s para Marte y 10 m/s para Venus en la actualidad). En este caso, las fuerzas generadas durante el impacto final son mucho menores que aquellas a las que se sometió el material al ser eyectado. De tal manera que el efecto del impacto a baja velocidad contribuye a mezclar el contenido del meteorito con el material en el sitio de impacto [8]. Esta fase del proyecto ha sufrido cierto retraso en su inicio, pero esperamos que en el transcurso del segundo año del proyecto se pueda iniciar sin mayor problema. Resultados
Hemos calculado la trayectoria de un conjunto de partículas que representan el material eyectado de la Tierra, como resultado de un gran impacto de un cometa o asteroide, para determinar su probabilidad de colisión con distintos cuerpos del Sistema Solar. Se han explorado varias velocidades de eyección, así como diferentes configuraciones planetarias iniciales, basados en simulaciones de 163,842 partículas sobre un periodo de 30,000 años. En concordancia con trabajos previos [3], encontramos que el material terrestre puede llegar a la Luna, Venus, e incluso regresar de vuelta a la Tierra después de un periodo de varios miles de años en el espacio. Por primera vez se predice que el material de la Tierra también puede alcanzar, así como a Júpiter e incluso a Saturno. Conclusiones
1. En colisiones con velocidades de impacto U > 2Vesc, una cantidad significativa de material (del orden de unos cuantos puntos porcentuales) regresará a la Tierra después de permanecer menos de 10,000 años en el espacio interplanetario.
2. El transporte de material hacia Venus y la Luna puede ocurrir mientras U ≳ 2Vesc. 3. La transferencia de material a Marte requiere una velocidad de eyección sólo ligeramente
mayor a la velocidad de escape de la Tierra: menos de un 5% adicional.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
65
4. El transporte de material de la corteza terrestre hacia la vecindad de Júpiter y Saturno requiere velocidades de impacto U ≈ 3 Vesc, y puede ocurrir sólo si el impacto ocurre en la cara del planeta que se encuentra viendo hacia la dirección de su movimiento orbital.
5. Una fracción importante, del orden del 40%, de todo el material que llega a otros planetas dentro del tiempo considerado en nuestras simulaciones, lo hacen en 10,000 años o menos.
Estos resultados se publicaron recientemente en la revista Icarus [9], aunque ya muchos medios hablaban sobre sus implicaciones [http://www.bbc.co.uk/news/science-environment-14637109], [http://news.discovery.com/space/has-earth-brand-life-seeded-the-galaxy-110822.html]. Agradecimientos
Este es un proyecto apoyado por el CONACYT 128563. Referencias
[1] H. J. Melosh, Geology, 13, 144-148 (1985) [2] C. Mileikowsky, F. A. Cucinotta, J. W. Wilson, B. Gladman, G. Horneck, L. Lindegren, J.
Melosh, H. Rickman, M. Valtonen, J. Q. Zheng, Icarus, 145, 391-427 (2000).
[3] B. Gladman, L. Dones, H. F. Levison, J. A. Burns, Astrobiology, 5, 483-496 (2005). [4] G. Horneck, D. Stöffler, S. Ott, U. Hornemann, C. S. Cockell, R. Moeller, C. Meyer, J. P.
P. de Vera, J. Fritz, S. Schade, N. A. Artemieva. Astrobiology, 8, 17-14 (2008). [5] R. Vázquez, P. G. Núñez, M. reyes-Ruiz, C. E. Chávez, zS. Hernández, Y. Oceguera, S. Gil,
P. F. Guillén, S. Rodríguez, A. Segura, F. J. Martín-Torres, L. Olguín, trabajo presentado en ORIGINS 2011: ISSOL and Bioastronomy Joint International Conference, Montpellier, Francia, 3-8 de Julio de 2011.
[6] J. R. Rowley, A. O. Dahl, S. Sengupta, J. S. Rowley, Palynology, 5, 107-152 (1981). [7] P. Fajardo-Cavazos, A. C. Schuerger, W. L. Nicholson, Acta Astronautica, 60, 534-540
(2007). [8] R. M. E. Mastrapa, H. Glanzberg, J. N., Head, H. J. Melosh, W. L. Nicholson, trabajo
presentado en XXXI Lunar and Planetary Science Conference, Houston, Texas, EEUU, 13-17 de marzo de 2000.
[9] M. Reyes-Ruiz, C. E. Chávez, H. Aceves, M. S. Hernández, R. Vázquez, P. G. Núñez, Icarus, 220, 777-786 (2012).
Figuras
Fig. 1. Grano de polen de Opuntia littorallis.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
66
Presentación en cartel: GEOFÍSICA
BIOCHAR: UNA ESTRATEGIA DE MITIGACIÓN DE CALENTAMIENTO GLOBAL EN LA
TIERRA, PARA DESPUÉS LOGRAR UNA ECOPOIESIS EN MARTE
Ramón A. Bacre González, Instituto de Geología, U.N.A.M. [email protected]
Introducción
Robert Hall Haynes concibió el término ecopoiesis (del griego: “hacer una casa para la vida”) para describir el proceso de establecimiento de un ecosistema, o biosfera, en un planeta inerte [1]. En la actual misión del Mars Science Laboratory (MSL), de la National Aeronautics and Space Administration (NASA) al planeta Marte, se analiza si existen o existieron las condiciones adecuadas para hospedar vida en el planeta rojo [1**]. En caso de no encontrar evidencias de un posible desarrollo de vida marciana, la información derivada de los análisis de suelo, rocas, atmósfera, radiación y agua de la misión en el planeta rojo, permitirá el planteamiento de la introducción de la vida terrestre en Marte [2].
En un contexto terrestre, el Panel Intergubernamental de Cambio Climático (IPCC) presenta en su cuarto reporte del año 2007, como un objetivo de mitigación, el alcanzar y mantener una concentración atmosférica de alrededor de 350 ppmv de CO2 en la atmósfera [1*]. Para lograr esa concentración, se proponen mecanismos económicos, políticas de gobierno, desarrollo tecnológico e inclusión social, entre otros. Dentro del objetivo principal de este trabajo, se presenta una opción de mitigación eficiente del calentamiento global en el planeta Tierra, haciendo énfasis en la necesidad de adecuar las condiciones terrestres a nuestros requerimientos, antes de emprender un intento por hacerlo en otro planeta.
La opción que se presenta es la estabilización de residuos de materia orgánica por medio de pirólisis de baja temperatura, 350-500 °C, para producir carbón vegetal. Éste material producto de la pirólisis, es conocido como en la literatura como biochar [3]. Es un material rico en carbono orgánico recalcitrante que puede ser introducido en la matriz del suelo para la captura de carbono y a su vez enriquecer la fracción orgánica necesaria para el desarrollo de vida vegetal y microbiana en el suelo. De acuerdo con Woodrow et al, para alcanzar la concentración de 350 ppmv de CO2 atmosférico, es necesario retirar alrededor de 35 Gt de CO2 para el año 2030 y la aplicación del biochar en el suelo de manera global, podría ser una solución muy efectiva [4].
El campus de Ciudad Universitaria de la UNAM, se ha tomado como objeto de estudio en la presente investigación, en vista de que se manejan aproximadamente 1500 t/año de residuos provenientes de sus áreas verdes, que tienen como destino final la planta de composteo de la misma UNAM. De este material orgánico se desconoce tanto la proporción de carbono orgánico recalcitrante (COR) en la composta producida, como la cantidad de carbono orgánico mineralizado y liberado a la atmósfera en forma de CO2.
Los residuos resultantes de la jardinería urbana y los derivados de las podas y residuos de cosecha de un número considerable de cultivos en el medio rural, tradicionalmente han sido la materia prima para la generación de compostas; sin embargo, para los residuos de naturaleza leñosa derivados

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
67
de estas mismas actividades y su transformación a través de la pirólisis, puede resultar ambiental y económicamente más productiva. Metodología Por lo antes expuesto, es necesario conocer el rendimiento de carbono orgánico recalcitrante en forma de biochar, evaluando previamente la composición, a fin de que este tipo de procesos puedan ser utilizados como alternativas para el de manejo de residuos a nivel global. De forma paralela se puede conocer la cantidad de carbono orgánico mineralizado que escapa a la atmósfera en forma de CO2 durante el proceso.
Para este propósito, se utilizó un cilindrico metálico diseñado para la pirólisis de baja temperatura, con capacidad de 12 Kg/carga, con intervalo de temperatura de operación de 250-400°C, durante 180 min para la producción de biochar.
El contenido de carbono antes y después del proceso, es evaluado por medio de análisis elemental en el laboratorio de fisicoquímica de suelos en el Instituto de Geología, UNAM. Resultados
Al momento de realizar éste resumen en extenso, los resultados están siendo procesados y se espera el contar con ellos en la presentación del poster con las tablas y figuras correspondientes. Referencias
[1] Haynes, R. (1990). Ecce Ecopoiesis: Playing God on Mars. MoruI Expertise , 161-183. [2] Haynes, R. H., & McKay, C. P. (1992). The implantation of life on Mars: Feasibility and
motivation. Advanced Space Research , 12 (4), (4)133-140. [3] Lehman, J. (2006). Biochar sequestration in Terrestrial Ecosystems: A review. Mitigation and
Adaptation Strategies for Global Change (11), 403-427. [4] Woodrow, W. II, & Chrisman, R. (2009). Challenges in Global Warming: Reduced emissions
and sequestration. Milken Institute; Atodyne Technologies and Affiliate Faculty, Forest Resources, UW, Seattle, Washington, USA.
[5] Rogner, H.-H., Zhou, D., Bradley, R., Crabbé, P., Edenhofer, O., Hare, B., y otros. (2007). Introduction. In Climate Change 2007: Mitigation. Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA.: Cambridge University Press.
[6] NASA. (2012). Mars Science Laboratory. Recuperado el 07 de Agosto de 2012, de Overview: http://mars.jpl.nasa.gov/msl/mission/overview/

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
68
Presentación en cartel: QUÍMICA PLANETARIA
LA IMPORTANCIA DE LA DETECCIÓN DE LITIO EN LA SUPERFICIE DE MARTE
María Colín-García, Instituto de Geología, UNAM [email protected]
Grupo SIOV Alejandro Heredia, Instituto de Ciencias Nucleares, UNAM; Centro de Tecnología Mecánica y
Automatización, TEMA, Universidad de Aveiro, Portugal, Julio Valdivia Silva, NASA, USA
Hugo Beraldi, Instituto de Geología, UNAM Alicia Negrón-Mendoza, Instituto de Ciencias Nucleares, UNAM
Héctor Durand-Manterola, Instituto de Geofísica, UNAM José Luis García-Martínez, Instituto de Ciencias Nucleares, UNAM
Sergio Ramos Bernal, Instituto de Ciencias Nucleares, UNAM Fernando Ortega Gutiérrez, Instituto de Geología, UNAM
Introducción
El origen de la vida tal como la conocemos se basa en la presencia de agua líquida, un compuesto químico simple formado por hidrógeno y oxígeno, el primer y tercer elemento más abundantes del Universo, respectivamente. El litio (Li), por su parte, es el tercer elemento de la tabla periódica y fue sintetizado durante el Big Bang junto con el hidrógeno y el helio. Debido a su solubilidad extrema, forma parte de la composición de algunas arcillas, y puede formar también algunas sales poco comunes que precipitan al evaporarse los cuerpos de agua. El pequeño tamaño iónico del Li, similar al del magnesio, favorece su incorporación en la estructura del olivino y el piroxeno, las principales fases que forman el manto de los planetas terrestres. Sin embargo, en conjunto el Li es un elemento escaso en la naturaleza (alrededor de 2 ppm en el manto de la Tierra), pero su contenido se puede incrementar hasta en un orden de magnitud en basaltos alterados y en espilitas que datan del Arcaico. En este trabajo se sugiere que la detección de Litio en rocas ígneas o sedimentarias en las proporciones de varias decenas de ppm en la superficie de Marte, implicaría la acción de sistemas hidrotermales sub-acuosos y sub-aéreos, y por lo tanto, la presencia en el pasado de lagos u océanos, con volúmenes y duración considerables, lo que aumentaría las posibilidades para que se diera el origen de la vida en ese planeta.
Propiedades generales del Li
El litio tiene el calor específico más alto entre los sólidos, y por lo tanto presenta una capacidad de transferencia de calor elevado. En su forma iónica tiene un número de coordinación de entre de 4 a 8, y la relación carga/tamaño es muy alta, lo que lo hace un ión pequeño altamente cargado; todo ello resulta en propiedades considerablemente diferente de otros iones similares como el sodio y el potasio. En solución acuosa, el Li exhibe propiedades únicas como son: presión de vapor muy baja y el abatimiento del punto de congelación del agua, además de otras propiedades coligativas, lo que amplía el intervalo de disponibilidad de agua líquida. El litio muestra características que son consistentes con el grupo de metales alcalinos (Grupo IA de la tabla periódica) al cual pertenece. Sin embargo, presenta algunas singularidades, tales como su reactividad, similar al magnesio (Grupo 2A). Por ejemplo, en una

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
69
reacción importante, el litio puede combinarse con el nitrógeno (N2) dando lugar al nitruro de litio, una reacción parecida sólo a la que presenta el Mg: 6Li(s) + N2(g) →2Li3N(s)
Algunas sales comunes de litio son el borato, el carbonato, el nitrato y el borohidruro (LiBH4). Con grafito, se forma el carburo de litio (Li2C2), que forma parte de una amplia gama de compuestos de carbono-litio. Además, con compuestos orgánicos se forman compuestos de organo-litio caracterizados por tener un enlace covalente directo entre los átomos de carbono y el de litio, lo cual genera bases extremadamente reactivas.
Li fuentes e isótopos
Hay dos procesos por los que los planetas pueden adquirir litio. El primero es el litio original, el primordial, que estaba contenido en la nebulosa y que fue añadido al planeta durante su formación. El segundo es el resultado de una fisión nuclear por el impacto de los rayos cósmicos de alta energía. Los isótopos del Li son escasos en el universo, sin embargo se ven enriquecidos por la acción de los rayos cósmicos en el medio interestelar; cuando los protones golpean los núcleos de átomos pesados (como el oxígeno y el carbono) los fragmentan, produciendo elementos más ligeros como el litio [1].
El bombardeo de rayos cósmicos en la superficie de un planeta durante miles de millones de años es capaz de producir cantidades sustanciales de Li cosmogénico, esto es particularmente importante en atmósferas ricas en CO2 o en superficies que contienen agua. Después de la desaparición de la atmósfera de CO2 en Marte, los rayos cósmicos debieron interactuar básicamente con el suelo, pero en el permafrost, rico en CO2, y en los hielos polares, este bombardeo continuo podría haber producido grandes cantidades de litio.
Otra singularidad importante es la abundancia relativa de los dos isótopos 7Li/6Li. La abundancia ha cambiado durante los últimos 4.6 Gy y algunos autores sugieren que el 7Li es entre 14 y 12 veces más abundante que el 6Li. Sin embargo, la producción de 6Li es un proceso continuo, promovido por la acción de los rayos cósmicos, por lo que dicha proporción seguirá disminuyendo. Minerales que contienen Litio y su papel en el origen de la vida
Muchos minerales contienen litio, este elemento puede ser encontrado abundantemente en salmueras naturales, salmueras asociadas a pozos petroleros, campos geotérmicos y el agua de mar. Por ejemplo, la hectorita, es un mineral raro (Na0.3 (Mg, Li) 3Si4O10 (OH) 2), una arcilla perteneciente al grupo de las esmectitas, capaz de formar complejos orgánicos-arcilla y podría haber contribuido al ensamblaje de algunos aminoácidos y otras moléculas de importancia prebiótica.
Aunque la química del litio ha sido ampliamente estudiada, se sabe aún poco sobre el rol que pudo haber jugado en los procesos de evolución química. Desde el punto de vista astrobiológico, el Li pudo haber jugado un papel principal alterando las propiedades fisicoquímicas del agua y participando en el ensamblaje y coordinación de moléculas orgánicas [2]. Nuestra investigación actual está centrada en comprender las propiedades potenciales relacionadas con superficies minerales que contienen litio en el ensamblaje de biomoléculas importantes o sus precursores. Por ejemplo, la adsorción del ácido cianhídrico en hectorita (Fig. 1), muestra que la adsorción se da en un porcentaje bajo, aproximadamente 16% comparado con la adsorción en montmorillonita de sodio, en donde la adsorción es del 100%. En este sentido, podría suceder que el mineral que contiene Li esté estabilizando moléculas de agua y previniendo la adsorción de moléculas de HCN. Además, de favorecer el autoensamblaje, los sólidos que contienen Li podrían aumentar la reactividad de moléculas orgánicas y favorecer el ensamblaje de moléculas más grandes. Estas propiedades son parte de nuestras investigaciones actuales.
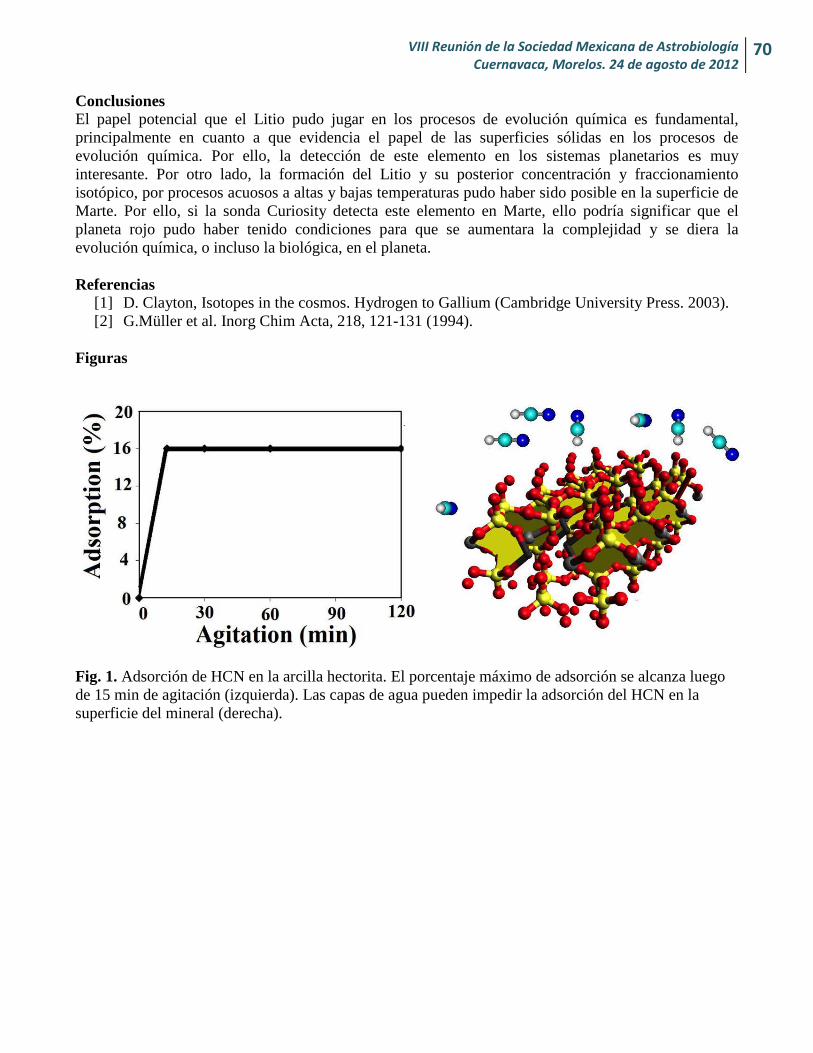
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
70
Conclusiones El papel potencial que el Litio pudo jugar en los procesos de evolución química es fundamental, principalmente en cuanto a que evidencia el papel de las superficies sólidas en los procesos de evolución química. Por ello, la detección de este elemento en los sistemas planetarios es muy interesante. Por otro lado, la formación del Litio y su posterior concentración y fraccionamiento isotópico, por procesos acuosos a altas y bajas temperaturas pudo haber sido posible en la superficie de Marte. Por ello, si la sonda Curiosity detecta este elemento en Marte, ello podría significar que el planeta rojo pudo haber tenido condiciones para que se aumentara la complejidad y se diera la evolución química, o incluso la biológica, en el planeta. Referencias
[1] D. Clayton, Isotopes in the cosmos. Hydrogen to Gallium (Cambridge University Press. 2003). [2] G.Müller et al. Inorg Chim Acta, 218, 121-131 (1994).
Figuras
Fig. 1. Adsorción de HCN en la arcilla hectorita. El porcentaje máximo de adsorción se alcanza luego de 15 min de agitación (izquierda). Las capas de agua pueden impedir la adsorción del HCN en la superficie del mineral (derecha).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
71
Presentación en cartel: QUÍMICA PLANETARIA
CANTIDAD DE H2O Y CO2 DESGASADOS EN MARTE DESDE SU FORMACIÓN Y SUS IMPLICACIONES PARA LA VIDA.
A. N. Medina-Amayo, Facultad de Ciencias, UNAM [email protected]
H.J. Durand-Manterola, Departamento de Ciencias Espaciales, Instituto de Geofísica, UNAM., [email protected]
Introducción
Durand-Manterola [1] estimó la cantidad de volátiles desgasados en la historia de Marte (Figura 1). En base a los valores publicados por este autor, y suponiendo que los únicos volátiles emitidos fueron H2O y CO2, obtuvimos la cantidad de estos desgasados a lo largo del tiempo. Esta suposición no es tan mala ya que estos dos son los principales volátiles que se desgasan por vulcanismo.
Cantidad de CO2 y de H2O en el tiempo geológico
Los modelos de acreción favorecen la proporción de volátiles en la zona de formación de Marte respecto a la de la Tierra [2, 3, 4, 5, 6]. Según estos modelos la razón de volátiles de Marte a la Tierra es de 8:3, es decir, si Marte tuviera la misma masa de la Tierra tendría 2.666 veces la cantidad de volátiles de esta. Pero como la masa de Marte es 0.107 veces la masa de la Tierra entonces la cantidad de volátiles en Marte al momento de su acreción seria 2.666x0.107 = 0.285 veces los volátiles acretados por la Tierra [1]. De esta fracción acretada hay que tomar la fracción desgasada que Durand-Manterola [1] estima en 0.381, por lo tanto la cantidad de volátiles desgasados en la historia de Marte debe ser del orden de 0.285x0.381 = 0.108 veces los volátiles desgasados en la Tierra. En la Tierra se desgasaron del orden de 661 Masas Atmosféricas Terrestres (MAT) (1 MAT = 5.28x1018 kg) de volátiles de los cuales 600 MAT son de agua [7]. Por otro lado Pollack [7] estimó que el inventario actual de volátiles en la Tierra es 10 veces más agua que CO2, es decir, que de todos los volátiles desgasados 1/11 es CO2 y 10/11 es H2O. Si estas proporciones son las mismas en Marte entonces debieron desgasarse del orden de 71 MAT de volátiles de los cuales serian 64.5 MAT de agua y 6.5 MAT de CO2. En la figura 2 podemos ver la cantidad total de volátiles, la cantidad de agua y la cantidad de bióxido de carbono que se fue desgasando a lo largo del tiempo geológico.
Desde hace 4600 Ma hasta 3400 Ma el desgasamiento que tuvo Marte fue muy intenso (Figura 3). Después de esa época el desgasamiento disminuyó drásticamente cambiando claramente su tendencia.
Respecto al agua es muy posible que todavía se encuentre ahí en forma de hielo, pero en cuanto al CO2 hay un problema ya que si toda la masa desgasada de CO2 estuviera ahí habría 8.7 bars de presión y no los 6 mb que hay actualmente. Es posible que este bióxido de carbono faltante parte de él haya sido arrastrado por el viento solar. Otra parte pudo perderse con los impactos en los primeros días del planeta. Otra parte está en los polos congelada como hielo seco. Y otra posibilidad es que exista enterrada como clatratos de CO2.
Las conclusiones de este trabajo son que: 1. Marte desgasó suficiente agua como para, si la temperatura era alta en el pasado, tener
océanos que podrían ayudar al desarrollo de la vida.
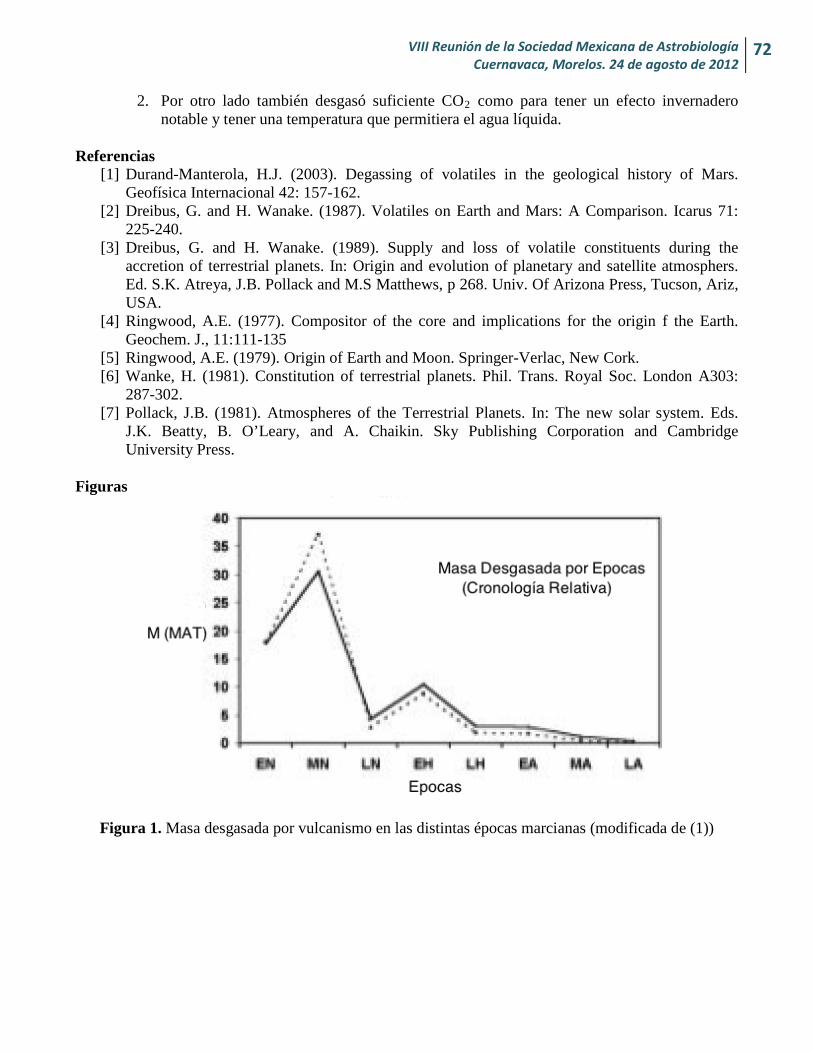
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
72
2. Por otro lado también desgasó suficiente CO2 como para tener un efecto invernadero notable y tener una temperatura que permitiera el agua líquida.
Referencias
[1] Durand-Manterola, H.J. (2003). Degassing of volatiles in the geological history of Mars. Geofísica Internacional 42: 157-162.
[2] Dreibus, G. and H. Wanake. (1987). Volatiles on Earth and Mars: A Comparison. Icarus 71: 225-240.
[3] Dreibus, G. and H. Wanake. (1989). Supply and loss of volatile constituents during the accretion of terrestrial planets. In: Origin and evolution of planetary and satellite atmosphers. Ed. S.K. Atreya, J.B. Pollack and M.S Matthews, p 268. Univ. Of Arizona Press, Tucson, Ariz, USA.
[4] Ringwood, A.E. (1977). Compositor of the core and implications for the origin f the Earth. Geochem. J., 11:111-135
[5] Ringwood, A.E. (1979). Origin of Earth and Moon. Springer-Verlac, New Cork. [6] Wanke, H. (1981). Constitution of terrestrial planets. Phil. Trans. Royal Soc. London A303:
287-302. [7] Pollack, J.B. (1981). Atmospheres of the Terrestrial Planets. In: The new solar system. Eds.
J.K. Beatty, B. O’Leary, and A. Chaikin. Sky Publishing Corporation and Cambridge University Press.
Figuras
Figura 1. Masa desgasada por vulcanismo en las distintas épocas marcianas (modificada de (1))
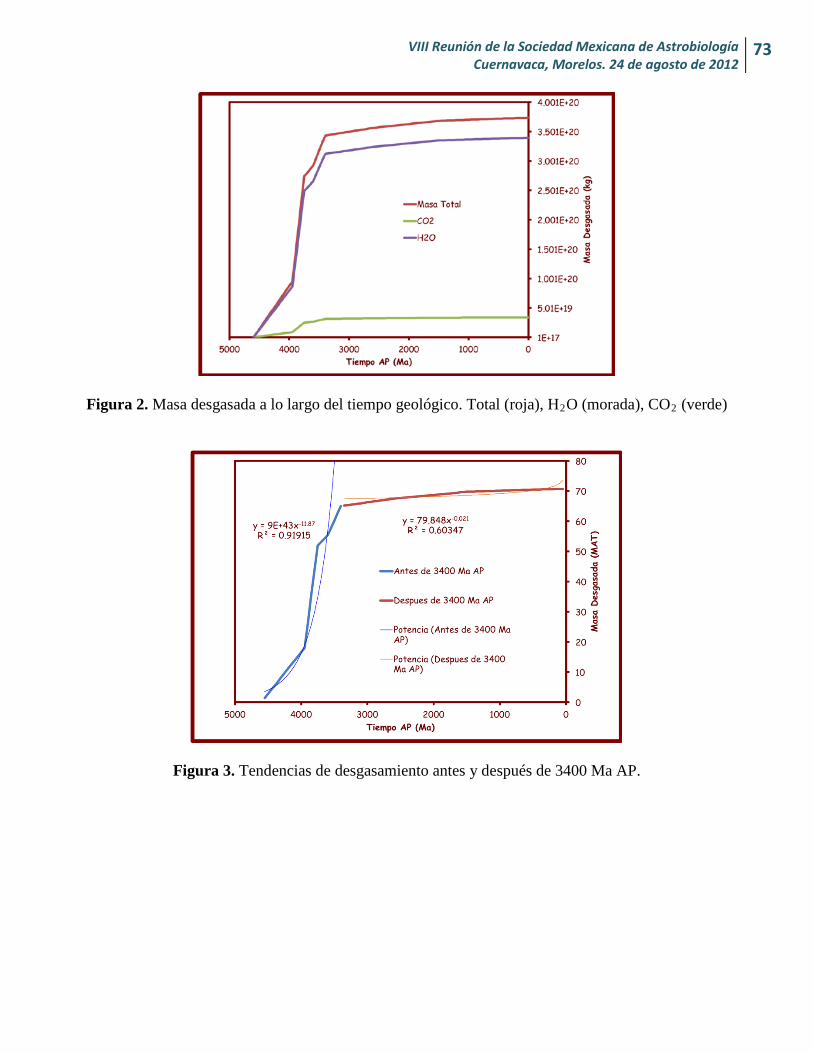
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
73
Figura 2. Masa desgasada a lo largo del tiempo geológico. Total (roja), H2O (morada), CO2 (verde)
Figura 3. Tendencias de desgasamiento antes y después de 3400 Ma AP.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
74
Presentación en cartel: QUÍMICA PLANETARIA
PRODUCCIÓN ABIÓTICA DE METANO EN PLANETAS TERRESTRES
Andrés Guzmán Marmolejo, Posgrado en Ciencias de la Tierra, UNAM. Instituto de Geofísica. Ciudad Universitaria, Coyoacán. C.P. 04510, México D.F. México.
[email protected] Antígona Segura, Instituto de Ciencias Nucleares, UNAM
Elva Escobar Briones, Instituto de Ciencias del Mar y Limnología, UNAM Introducción
El metano (CH4) ha sido propuesto como una bioseñal; no obstante, puede provenir de fuentes biológicas y geológicas [1]; tal y como sucede en la Tierra, donde procesos de serpentinización en la corteza son la principal fuente geológica que contribuye a su abundancia atmosférica [2].
Existe poca información relacionada con este tema para planetas fuera del sistema solar [3]. No obstante, determinar los límites máximos que el CH4 puede alcanzar por fuentes geológicas en planetas terrestres, será de utilidad para futuras misiones con la capacidad de analizar las atmósferas planetarias. Estas misiones podrían diferenciar a un planeta habitable de uno habitado, si encuentran abundancias de metano superiores a las formadas por sus límites máximos de producción geológica, lo que indicaría la existencia de fuentes biológicas.
Formación de metano geológico
Geológicamente el CH4 se forma por la reducción de Dióxido de Carbono (CO2) disuelto en agua (CO2 + 4H2 → CH4 + 2H2O, reacción 1). El H2 necesario proviene de la reacción de oxidación del Fe2+ durante la serpentinización (3Fe2SiO4 + 2H2O ↔ 2Fe3O4 + 3SiO2 + 2H2, reacción 2).
Basados en el modelo de Rudge et al. [4], inferimos una ecuación de primer orden para la velocidad de formación de H2 (vH2) de la reacción 2, vH2 = 2κ[Fe2SiO4], donde κ es la constante de
velocidad y [Fe2SiO4] es la concentración de fayalita; ( )20
20
0TTαe
aa
κ=κ −−⋅
, donde a es el tamaño
del grano de fayalita, T es la temperatura de reacción, κ0 = 10−6 s−1, a0 = 70 μm, α = 2.09 × 10−4 ºC−2 y T0 = 260 ºC [5].
A partir de un análisis teórico de variación de concentraciones de la reacción 1, expresamos el
rendimiento de la formación de metano como , donde Keq es la constante de
equilibrio de la reacción 1, [H2] es la concentración de H2 y x es el rendimiento de reacción. Keq puede estimarse a partir de la energía libre de Gibbs estándar de reacción usando el programa SUPCRT92 (http://geopig.asu.edu/) [6].
Descripción de los modelos
Se empleó un modelo fotoquímico unidimensional desarrollado por Pavlov y Kasting [7] y modificado por Kharecha et al. [8] y Segura et al. [9], que determina las abundancias al equilibrio termodinámico de 73 especies químicas, relacionadas en 359 reacciones atmosféricas. El modelo

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
75
simula la influencia del Sol sobre la atmósfera con un ángulo de cenit de 50º, dividiendo la atmósfera en 70 capas de 1 km de ancho para los planetas de 1 M⊕ y 0.5 km para los de 5 M⊕. Una ventaja de éste modelo es su capacidad para mantener el balance redox, mediante el balance de moléculas de H2 consumidas o formadas en las diferentes reacciones [10]. El modelo fue modificado para incluir el degasamiento de CH4 geológico.
Acoplado al modelo fotoquímico se empleó un modelo climático (radiativo-convectivo) que calcula los perfiles de temperatura a partir de la radiación solar adsorbida por la atmósfera y su composición química. El modelo considera una presión total de 1 bar para los planetas de 1M⊕ y 2 bares para los de 5 M⊕. Las nuevas temperaturas calculadas por el modelo climático, sirven como datos de entrada para que el modelo fotoquímico determine las abundancias al equilibrio de las especies químicas; éstas son devueltas al modelo climático para recalcular la temperatura. El proceso se repite hasta que se llega a una convergencia.
Resultados
Usando las mejores condiciones para maximizar los rendimientos, estimamos que el CH4 puede alcanzar flujos de hasta 2.25×1011 moléculas cm-2 s-1. Estos se logran a temperaturas de 533 K que pueden existir en el interior de los sistemas hidrotermales. En la Tierra se han medido concentraciones de CH4 de alrededor de 2.5 mmol/kg de agua en este tipo de ambientes [11]. Análisis de isotopos de H2 sugieren que el CH4 encontrado en las ventilas hidrotermales en The Lost City es de origen geológico [12], aunque la discusión sobre su origen continua aún [13]; no obstante, diferentes estudios teóricos y experimentales sugieren que, tanto el H2, como el CH4 puede ser formados en esos ambientes por procesos de serpentinización [2,14].
A partir de los flujos superficiales de CH4 calculamos su abundancia en atmósferas de planetas terrestres de 1 M⊕ y 5 M⊕ alrededor de una estrella similar al Sol. Los planetas fueron colocados a una distancia de su estrella de 1.13 UA y 1.16 UA para planetas de 1 M⊕ y 5 M⊕ respectivamente, donde la temperatura mínima obtenidas están por encima del punto de congelamiento del agua (Tabla 1). En todos los casos las atmósferas estuvieron libres de oxígeno y contenían presiones parciales de CO2 de 0.01, 0.03, 0.1 y 0.3 con la cantidad necesaria de N2 para alcanzar una presión total de 1 bar para los planetas de 1 M⊕ y 2 bares para los de 5 M⊕.
La tabla 2 muestra las abundancias de CH4 superficial obtenidas para todos los sistemas ensayados, donde podemos observar que la tasa de mezclado de CH4 atmosférico aumenta con su flujo superficial y la figura 1 muestra sus perfiles de abundancia a diferentes alturas Comentarios finales
La capacidad de producción de CH4 en planetas terrestres fue estimada en base a las propiedades cinéticas y termodinámicas de reacciones que ocurren en el interior de los sistemas hidrotermales, ligados a la actividad de las placas tectónicas. Valencia et al. [15] sugiere que el tectonismo es inevitable en planetas rocosos de 1 M⊕ a 10 M⊕; aunque no es posible saber cuánta superficie de un planeta extrasolar estaría cubierta por sistemas hidrotermales. Esto no supone un problema pues nuestros cálculos están enfocados a estimar un máximo posible de producción.
Referencias
[1] J. D. Des Marais et al, Astrobiology 2, 153–81 (2002). [2] C. Marcaillou et al. Earth Planet. Sci. Lett. 303, 281-290 (2011). [3] G. Etiope, D. Z. Oehler, C. C. Allen. Planet Space Sci 59, 182–195 (2011). [4] J. F. Rudge, P.B. Kelemen, M. Spiegelman, M. Earth Planet. Sci. Lett., 291, 215-227 (2010).
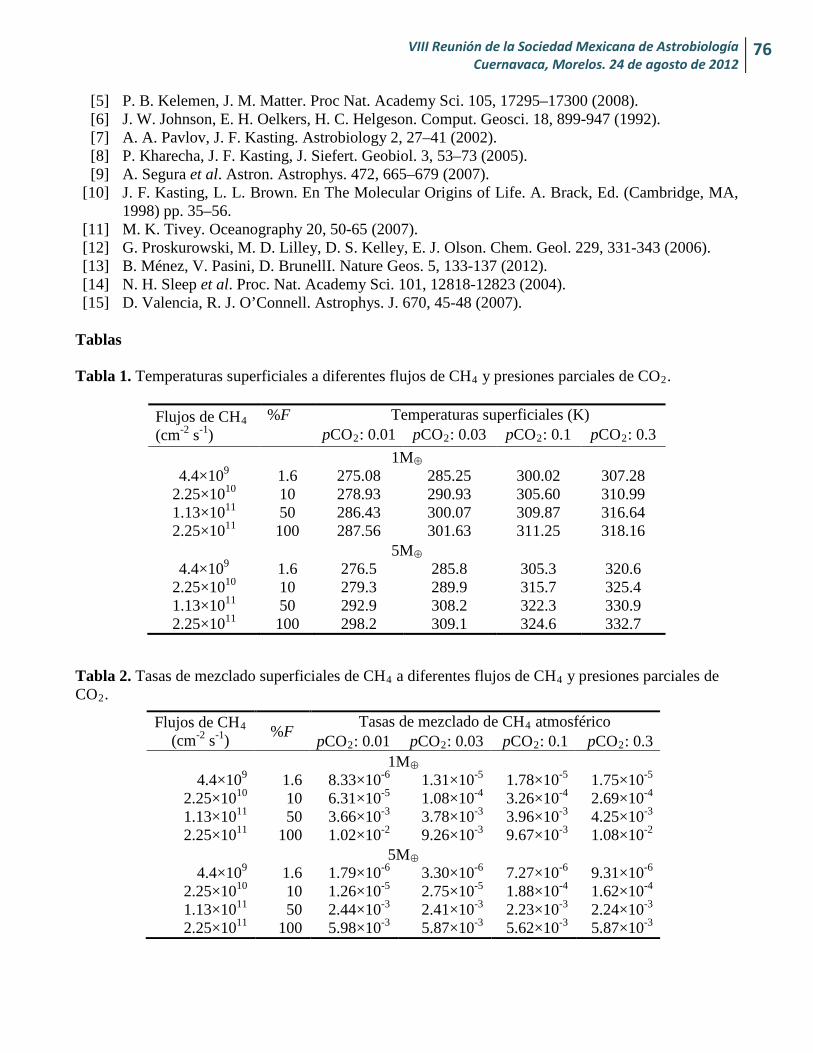
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
76
[5] P. B. Kelemen, J. M. Matter. Proc Nat. Academy Sci. 105, 17295–17300 (2008). [6] J. W. Johnson, E. H. Oelkers, H. C. Helgeson. Comput. Geosci. 18, 899-947 (1992). [7] A. A. Pavlov, J. F. Kasting. Astrobiology 2, 27–41 (2002). [8] P. Kharecha, J. F. Kasting, J. Siefert. Geobiol. 3, 53–73 (2005). [9] A. Segura et al. Astron. Astrophys. 472, 665–679 (2007).
[10] J. F. Kasting, L. L. Brown. En The Molecular Origins of Life. A. Brack, Ed. (Cambridge, MA, 1998) pp. 35–56.
[11] M. K. Tivey. Oceanography 20, 50-65 (2007). [12] G. Proskurowski, M. D. Lilley, D. S. Kelley, E. J. Olson. Chem. Geol. 229, 331-343 (2006). [13] B. Ménez, V. Pasini, D. BrunellI. Nature Geos. 5, 133-137 (2012). [14] N. H. Sleep et al. Proc. Nat. Academy Sci. 101, 12818-12823 (2004). [15] D. Valencia, R. J. O’Connell. Astrophys. J. 670, 45-48 (2007).
Tablas Tabla 1. Temperaturas superficiales a diferentes flujos de CH4 y presiones parciales de CO2.
Flujos de CH4 (cm-2 s-1)
%F Temperaturas superficiales (K) pCO2: 0.01 pCO2: 0.03 pCO2: 0.1 pCO2: 0.3
1M⊕ 4.4×109 1.6 275.08 285.25 300.02 307.28
2.25×1010 10 278.93 290.93 305.60 310.99 1.13×1011 50 286.43 300.07 309.87 316.64 2.25×1011 100 287.56 301.63 311.25 318.16
5M⊕ 4.4×109 1.6 276.5 285.8 305.3 320.6
2.25×1010 10 279.3 289.9 315.7 325.4 1.13×1011 50 292.9 308.2 322.3 330.9 2.25×1011 100 298.2 309.1 324.6 332.7
Tabla 2. Tasas de mezclado superficiales de CH4 a diferentes flujos de CH4 y presiones parciales de CO2.
Flujos de CH4 (cm-2 s-1) %F Tasas de mezclado de CH4 atmosférico
pCO2: 0.01 pCO2: 0.03 pCO2: 0.1 pCO2: 0.3 1M⊕
4.4×109 1.6 8.33×10-6 1.31×10-5 1.78×10-5 1.75×10-5 2.25×1010 10 6.31×10-5 1.08×10-4 3.26×10-4 2.69×10-4 1.13×1011 50 3.66×10-3 3.78×10-3 3.96×10-3 4.25×10-3 2.25×1011 100 1.02×10-2 9.26×10-3 9.67×10-3 1.08×10-2
5M⊕ 4.4×109 1.6 1.79×10-6 3.30×10-6 7.27×10-6 9.31×10-6
2.25×1010 10 1.26×10-5 2.75×10-5 1.88×10-4 1.62×10-4 1.13×1011 50 2.44×10-3 2.41×10-3 2.23×10-3 2.24×10-3 2.25×1011 100 5.98×10-3 5.87×10-3 5.62×10-3 5.87×10-3

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
77
Presentación en cartel: QUÍMICA PLANETARIA
INFLUENCIA DE LAS SUPERFICIES MINERALES EN EL DESARROLLO Y ESTABILIZACIÓN DE MOLÉCULAS ORGÁNICAS EN CONDICIONES PREBIÓTICAS
Ellen Yvette Aguilar-Ovando, Instituto de Ciencias Nucleares, UNAM
[email protected] Alicia Negrón-Mendoza, Instituto de Ciencias Nucleares, UNAM
Introducción
La condición homoquiral de ciertas moléculas orgánicas fundamentales presentes en los sistemas biológicos es considerada base en la organización de la materia orgánica capaz de manifestar y soportar la vida. Todos los sistemas biológicos conocidos, incluyendo virus y priones, presentan homoquiralidad en sus bloques de construcción, siendo aminoácidos de la serie L los que constituyen las proteínas, y azúcares de la serie D los que intervienen en las rutas metabólicas y forman parte de los ácidos nucleicos.
El hecho de que esta característica sea común a todos los organismos vivos ha hecho que la búsqueda de su origen sea un tema fundamental en estudios de origen de la vida y evolución.
Las primeras investigaciones efectuadas en este campo datan de hace aproximadamente 80 años, y han dado múltiples enfoques al problema de la aparición y mantenimiento de un sistema homoquiral. Por ejemplo, se han hecho consideraciones sobre la posibilidad de procesos autocatalíticos de amplificación de las variaciones estadísticas naturales de un sistema racémico [1], se ha investigado sobre la fotodestrucción quirioselectiva en medios interestelares [2,3,4], y se han hecho algunos experimentos sobre la adsorción preferente de las especies en las superficies cristalinas de minerales presentes en épocas primitivas, como calcitas (CaCO3), olivino ((Mg,Fe2+)2SiO4), arcillas, etc., que pudieran haber ocasionado la concentración de un enantiómero particular, estabilizando el exceso de la especie, o por lo menos retrasando su racemización. Estudios previos como los de Lahav (1999), Hazen y Sholl (2003), y Yoreo y Dove (2004) [5], han destacado la viabilidad de esta última hipótesis, pero aún se requiere de más pruebas.
El presente estudio propone aportar nueva evidencia al respecto de esta última hipótesis, investigando si la presencia de ciertas superficies minerales bajo condiciones específicas de pH, radiación gamma y condiciones anóxicas que simulen un ambiente terrestre primitivo, puede promover la acumulación de un exceso enantiomérico en disoluciones de cuatro aminoácidos (lisina, ácido aspártico, alanina y serina), por procesos combinados de adsorción preferente.
De esta forma, el proyecto planteado permitiría, en primer lugar, apoyar con más datos experimentales los estudios previos, permitiendo discernir estadísticamente la validez de dicha hipótesis. En segundo lugar, añade la radiación ionizante como una variable más a considerar en los estudios, con lo que podría establecerse la capacidad de las superficies minerales para proteger (y no sólo concentrar) enantioselectivamente los aminoácidos. En tercer lugar, el análisis de aminoácidos pertenecientes a los cuatro grupos químicos existentes (ácidos, básicos, polares y apolares) permitiría correlacionar su capacidad de ser adsorbidos y sobrevivir determinadas dosis de radiación gamma dentro de superficies minerales, con la concentración en que son hallados en estudios prebióticos,

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
78
según los cuales, los de características ácidas y apolares predominan en concentración frente a los polares y básicos. Métodos
El trabajo experimental se dividió en grupos de experimentos: a) Pruebas de adsorción, b) Determinación de sitios de adsorción, c) Pruebas de desorción, y d) Pruebas de irradiación.
Las pruebas de adsorción y desorción (a y c) se llevan a cabo mediante cambios de pH y temperatura a las disoluciones de aminoácidos, en presencia y ausencia de los minerales finamente divididos, verificándose la adsorción y desorción mediante cromatografías de líquidos de alta resolución acoplada a un detector de luz dispersa (HPLC/ELSD). Los sitios de adsorción (b) se determinan mediante análisis de difracción de rayos X de polvos y bloqueo químico de sitios activos de las estructuras minerales.
Las pruebas de irradiación (d) se efectúan en disolución en ausencia de oxígeno, exponiendo las muestras a distintas dosis de radiación gamma proveniente de una fuente de cobalto-60.
Toda la cristalería empleada se lava de acuerdo al protocolo de Draganič & Draganič (1971) para eliminación de materia orgánica [6]. Resultados
Hasta el momento del proyecto se han obtenido experimentalmente las condiciones cromatográficas y analíticas para la separación e identificación de cada aminoácido en sus dos formas enantioméricas y se ha preparado la materia prima de los experimentos de adsorción, triturándose manualmente los minerales y lavándoles con una solución oxidante para eliminar materia orgánica.
Posteriormente se han llevado a cabo experimentos de adsorción de alanina sobre arcilla, meteorito y olivino a pH=2. Una vez adsorbido el aminoácido, se trataron las muestras para permitir su análisis mediante la técnica seleccionada (ajustando su pH y retirando el sodio con una resina de intercambio catiónico, Dowex 50®). Utilizando los datos de las áreas de pico en cada muestra se calcularon los porcentajes adsorbidos de cada enantiómero sobre cada mineral y se calcularon porcentajes de adsorción para cada enantiómero libre y en disolución racémica. Los resultados señalan un mayor porcentaje de adsorción sobre la superficie del mineral de meteorito, frente a la observada en arcilla al mismo valor de pH. Sobre el olivino no se observó adsorción alguna. Así mismo se observa un mayor porcentaje de adsorción del enantiómero L en ambas superficies minerales, cuando éste se encuentra puro en la disolución, comparado con el que se adsorbe cuando se presenta la mezcla racémica.
Tras efectuar la dosimetría de las posiciones a emplear en la etapa de irradiación de muestras mediante el Método de Fricke Modificado, se ensayó la irradiación de disoluciones de alanina racémica y en sus formas enantioméricas puras, en concentraciones 0,01M, a pH=2 y sin acidular. Se probaron diferentes dosis y se llevó a cabo el experimento 10 veces para comprobar la repetibilidad de los resultados. La tendencia general observada muestra a la L-Alanina como más resistente a la radiación ionizante, en especial cuando es irradiada en disoluciones enantioméricamente puras y en ausencia de ácido (véase Figura 1). Así mismo, se observó la destrucción casi total del aminoácido sobre los 91 KGy, y que el proceso de descomposición una vez irradiadas las muestras continúa conforme pasa el tiempo, acelerándose al incrementarse la temperatura.
Se efectuó un ensayo de irradiación de alanina en presencia de arcilla, aplicando una dosis de 91 KGy, para determinar la capacidad de protección que le brinda el mineral. Se observó que la presencia del mineral disminuye drásticamente el daño que sufre el aminoácido por acción de la radiación gamma, manteniéndose el mayor porcentaje de L-Alanina (véase Tabla 1).
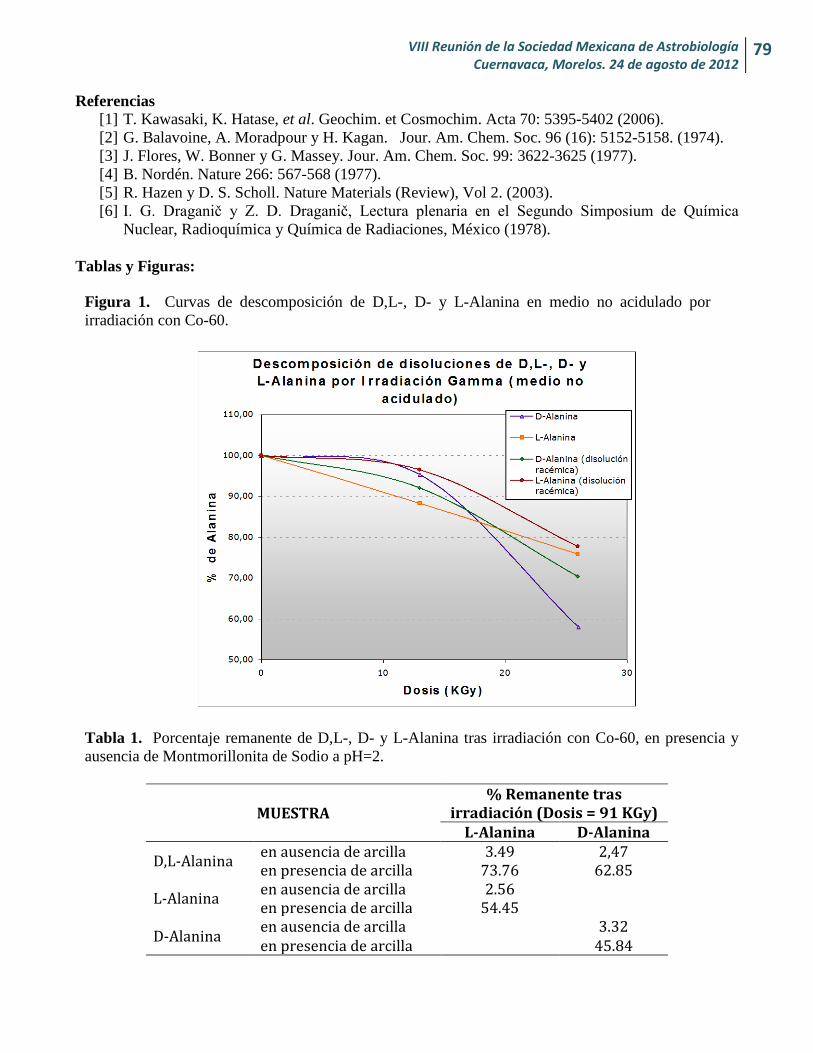
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
79
Referencias [1] T. Kawasaki, K. Hatase, et al. Geochim. et Cosmochim. Acta 70: 5395-5402 (2006). [2] G. Balavoine, A. Moradpour y H. Kagan. Jour. Am. Chem. Soc. 96 (16): 5152-5158. (1974). [3] J. Flores, W. Bonner y G. Massey. Jour. Am. Chem. Soc. 99: 3622-3625 (1977). [4] B. Nordén. Nature 266: 567-568 (1977). [5] R. Hazen y D. S. Scholl. Nature Materials (Review), Vol 2. (2003). [6] I. G. Draganič y Z. D. Draganič, Lectura plenaria en el Segundo Simposium de Química
Nuclear, Radioquímica y Química de Radiaciones, México (1978). Tablas y Figuras:
Figura 1. Curvas de descomposición de D,L-, D- y L-Alanina en medio no acidulado por irradiación con Co-60.
Tabla 1. Porcentaje remanente de D,L-, D- y L-Alanina tras irradiación con Co-60, en presencia y ausencia de Montmorillonita de Sodio a pH=2.
MUESTRA % Remanente tras
irradiación (Dosis = 91 KGy) L-Alanina D-Alanina
D,L-Alanina en ausencia de arcilla 3.49 2,47 en presencia de arcilla 73.76 62.85
L-Alanina en ausencia de arcilla 2.56 en presencia de arcilla 54.45
D-Alanina en ausencia de arcilla 3.32 en presencia de arcilla 45.84

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
80
Presentación en cartel: QUÍMICA PLANETARIA
SIMULACIÓN DE RELÁMPAGOS A BAJA TEMPERATURA EN LA ATMÓSFERA DE TITÁN
Dení Tanibé Zenteno Gómez, Centro de Investigaciones Químicas, UAEM
[email protected] Sandra Ignacia Ramírez Jiménez, Centro de Investigaciones Químicas, UAEM
Introducción
Titán es un satélite natural del planeta Saturno que tiene la peculiaridad de poseer una atmósfera formada principalmente por nitrógeno molecular (N2), como sucede con la atmósfera de la Tierra, aunque con una densidad ligeramente mayor. Otro de los componentes atmosféricos importantes de esta luna es el metano (CH4). La proporción de ambos gases varía dependiendo de la altura a la que se encuentren por encima de la superficie [1].
Dentro de la atmósfera de Titán se han encontrado compuestos de tipo hidrocarburo y nitrilo, que si bien no se sabe con certeza cómo se originan, parecen formarse debido a la posible ocurrencia de relámpagos cuya energía permite la ruptura de enlaces de las moléculas de los constituyentes principales y en consecuencia la combinación de los iones o radicales originados para formar nuevos compuestos químicos [2]. Aunque no se han detectado descargas eléctricas de tipo relámpago en Titán, o de alguna otra forma de descarga eléctrica atmosférica, existe evidencia de su existencia gracias a las observaciones realizadas por el instrumental de la misión Cassini-Huygens [1, 2].
En este trabajo se determinó el rendimiento energético de los compuestos químicos resultantes de una simulación de la ocurrencia de descargas eléctricas en la atmósfera de Titán, en un régimen de temperatura de -180 °C. Simulación a baja temperatura de la atmósfera de Titán
Se llenó un reactor con 600 mbar de una mezcla gaseosa compuesta por 98% de N2 y 2% de CH4, previa evacuación de cualquier otro material dentro del reactor por medio de bombas de vacío. Posteriormente se enfrió el reactor con nitrógeno líquido para simular la temperatura de la atmósfera de Titán (aproximadamente -180ºC) y, con ayuda de una fuente de poder, se generó una descarga eléctrica utilizando como electrodos dos filamentos de tungsteno, de tal forma que la mezcla gaseosa contenida en el reactor fuese irradiada. Este procedimiento se realizó para distintos tiempos de irradiación, específicamente para 10, 20, 30, 40, 50, 75 y 100 minutos.
Al finalizar la irradiación, los compuestos químicos sintetizados se introdujeron a un cromatógrafo de gases para separarlos y fueron caracterizados con la ayuda de un espectrómetro de masas.
Además, para cada tiempo de irradiación se calculó la energía eléctrica asociada a la producción de cada uno de los compuestos obtenidos utilizando los valores de voltaje e intensidad de corriente proporcionados por la fuente de poder, integrándolos respecto al tiempo de irradiación. Graficando el número de moléculas obtenido para cada compuesto vs la energía de la descarga, se obtuvo el rendimiento energético para cada uno de los compuestos mediante el cálculo de la pendiente de cada curva resultante.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
81
Resultados Para todas las irradiaciones se obtuvieron diferentes abundancias en cada uno de los compuestos
producidos. Además, no todos los compuestos se forman para todas las irradiaciones. En la Figura 1 se muestra un cromatograma a modo de ejemplo, en donde están indicados todos
los compuestos que se obtuvieron para una irradiación de 100 minutos a una presión de 600 mbar. Siete de los principales compuestos obtenidos en los experimentos se enlistan en la Tabla 1,
donde también se indica el tiempo de retención y el rendimiento energético de cada uno de ellos. La Figura 2 corresponde a una gráfica que muestra el número de moléculas vs la energía de la
descarga para los siete compuestos de la Tabla 1. Es a partir de gráficas como ésta de donde se obtiene la pendiente (rendimiento energético) para cada uno de los compuestos sintetizados. Conclusiones
Para cualquier energía, el compuesto que tiene la mayor producción es el cianuro de hidrógeno (HCN). El etano (CH3-CH3), el etino (CH≡CH) y el etanonitrilo (CH3-CN) son los compuestos que se sintetizan con mayor abundancia para cualquier valor de energía. Esto quiere decir que estos compuestos, junto con el HCN, no requieren de una gran cantidad de energía para formarse en comparación con el resto de los compuestos. Para energías mayores a los 20,000 J, la abundancia de los compuestos mencionados decae. Esto es debido seguramente a que hay mayor energía disponible que es utilizada para formar otros compuestos de mayor peso molecular.
De manera general, puede decirse que las abundancias de todos los compuestos siguen el mismo patrón de comportamiento conforme aumenta la energía de la descarga eléctrica.
Referencias
[1] M. Fulchignoni, et. al., Nature 438, 785-791 (2005). [2] G. Contreras, tesis, Universidad Autónoma del Estado de Morelos (2010).
Figuras y Tablas Tabla 1. Rendimiento energético de siete de los compuestos obtenidos tras la irradiación de una muestra gaseosa de nitrógeno (98%) y metano (2%).
Compuesto Fórmula molecular Rendimiento energético (moléculas/J)
Eteno C2H4 1.84×1013 Etano C2H6 2.84×1014 Etino C2H2 4.23×1014
Propano C3H8 1.58×1014 Etanodinitrilo C2N2 7.79×1013 Etanonitrilo C2H3N 3.46×1014
Propanonitrilo C3H5N 1.2×1014
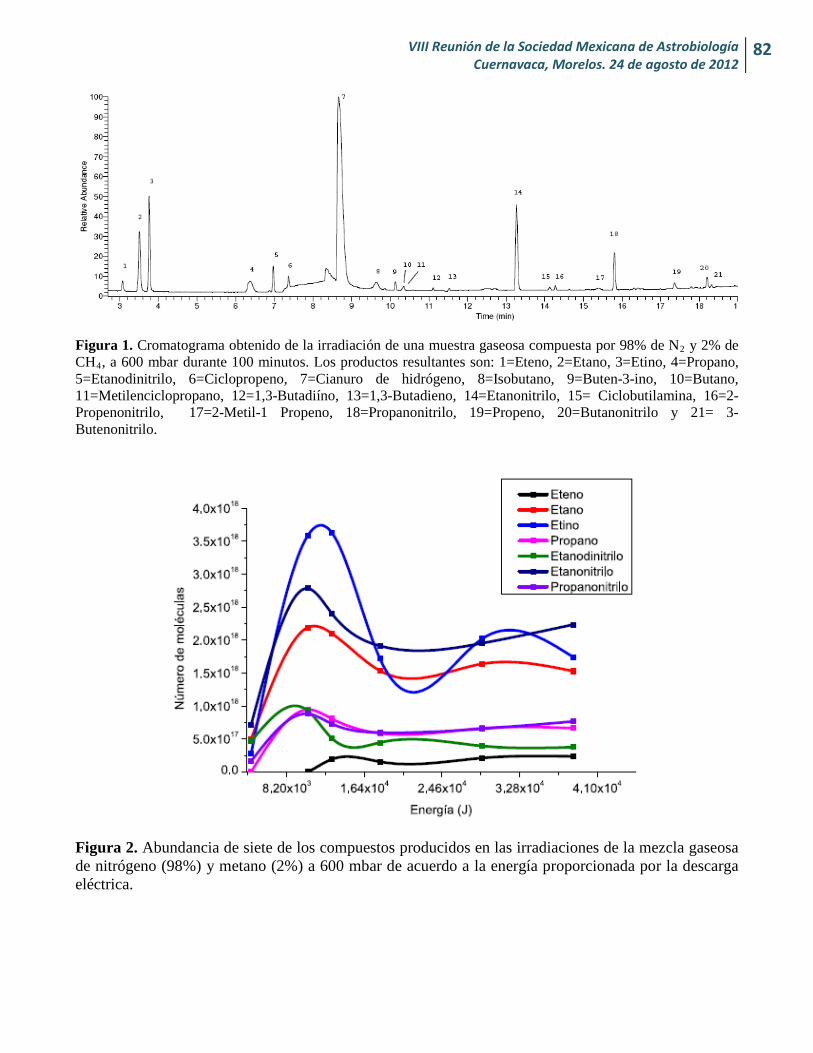
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
82
Figura 1. Cromatograma obtenido de la irradiación de una muestra gaseosa compuesta por 98% de N2 y 2% de CH4, a 600 mbar durante 100 minutos. Los productos resultantes son: 1=Eteno, 2=Etano, 3=Etino, 4=Propano, 5=Etanodinitrilo, 6=Ciclopropeno, 7=Cianuro de hidrógeno, 8=Isobutano, 9=Buten-3-ino, 10=Butano, 11=Metilenciclopropano, 12=1,3-Butadiíno, 13=1,3-Butadieno, 14=Etanonitrilo, 15= Ciclobutilamina, 16=2-Propenonitrilo, 17=2-Metil-1 Propeno, 18=Propanonitrilo, 19=Propeno, 20=Butanonitrilo y 21= 3-Butenonitrilo.
Figura 2. Abundancia de siete de los compuestos producidos en las irradiaciones de la mezcla gaseosa de nitrógeno (98%) y metano (2%) a 600 mbar de acuerdo a la energía proporcionada por la descarga eléctrica.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
83
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
ENDOSIMBIONTES COMO MODELOS DE CÉLULAS MÍNIMAS
Luis José Delaye Arredondo, CINVESTAV Irapuato [email protected]
David José Martínez Cano, CINVESTAV Irapuato Introducción
¿Cuál es el número mínimo de funciones necesarias para sustentar la vida? Esta pregunta, con una larga tradición en biología teórica, ha sido abordada bioinformática y experimentalmente por diversos grupos de investigación en fechas recientes. El concepto de célula mínima presupone que debe de existir un número mínimo de genes, y por lo tanto de funciones, necesarias para mantener a la unidad fundamental de la vida, que es la célula.
La búsqueda de la célula mínima, en su aproximación más platónica, es también la búsqueda de leyes, principios o procesos comunes a todo ser vivo. En este sentido, la búsqueda de la célula mínima es similar al estudio de las leyes últimas de la física [1]. Sin embargo, la realidad biológica, a diferencia de lo que ocurre en física, nos impone una aproximación aristotélica al problema. Dado que el conjunto mínimo de genes necesarios para mantener una célula viva depende en gran medida del ambiente, en la práctica existe una infinidad de sistemas celulares mínimos, cada uno adaptado a su ambiente particular [2]. De hecho, la definición de célula mínima incluye la relación que existe entre un ambiente determinado y el contenido mínimo indispensable de genes para sobrevivir en ese ambiente [3]. Esta dependencia del ambiente es clara si se compara el número de genes necesarios para mantener una célula en un ambiente químicamente complejo con el número de genes necesarios, en un ambiente químicamente simple. Por ejemplo Mycoplasma genitalium requiere alrededor de 400 genes para vivir en un ambiente intracelular, mientras que la cianobacteria Prochlorococcus marinus requiere alrededor de 1,400 para vivir en un ambiente marino [4].
El hecho de que no exista un único genoma mínimo, hace muy interesante el preguntarse sobre las similitudes y diferencias, en términos genéticos y bioquímicos, existentes entre los diversos procariontes con genomas pequeños y como la evolución biológica ha resuelto por diversos caminos, el problema de vivir con un número reducido de genes.
En este trabajo proponemos que el estudio comparativo de los genomas reducidos de bacterias endosimbiontes y parásitas pueden ser utilizado para abordar las preguntas anteriores.
Métodos Las aproximaciones para identificar el número mínimo de genes necesarios para mantener una célula viva se pueden dividir en dos clases. Por un lado, la aproximación bottom-up pretende construir sistemas artificiales químicos capaces de replicarse y evolucionar a partir de componentes moleculares más simples. Por otro lado, la aproximación top-down parte de los organismos actuales con el objetivo de simplificar su genoma para obtener una versión mínima de una célula. En este trabajo utilizaremos la aproximación top-down para identificar genes conservados entre distintas especies de procariontes cuyo genoma ha evolucionado hacia un contenido bajo de genes. Haremos un estudio de la biodiversidad de microorganismos con genoma reducido y discutiremos las implicaciones de esta

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
84
diversidad para entender cuál podría ser el número mínimo de genes necesarios para mantener una célula viva. Resultados En la actualidad, existe una amplia diversidad de procariontes cuyo genoma se ha secuenciado completamente. De ellos, los procariontes que son parásitos obligados o que son endosimbiontes tienen los genomas con el menor número de genes (Figura 1), siendo estos últimos los campeones de la reducción genómica. Se ha sugerido que el concepto de célula mínima puede ser utilizado como una herramienta para comprender mejor el funcionamiento celular [1]. Siguiendo esta lógica, hace pocos años se publicó un intento por diseñar una célula mínima utilizando la información proveniente de genes conservados entre distintas especies de bacterias simbiontes junto con datos experimentales de esencialidad génica [5]. El resultado es un sistema celular con apenas 206 genes. Este sistema mínimo codifica para todos los genes necesarios para: a) replicar, reparar y transcribir el DNA; b) traducir mRNA en proteíans; c) plegar y secretar proteínas; d) dividir la membrana celular; e) transportar algunos metabolitos a través de la membrana celular; f) un metabolismo básico. Si bien este sistema teórico cuenta con tan solo la mitad de los genes que tiene la bacteria Mycoplasma genitalium, entre la diversidad de bacterias endosimbiontes existen algunas con un menor número de genes, tal como es el caso de las bacterias que se muestran en la Tabla 1. Candidatus Zinderia insectícola (202), Candidatus Carsonella ruddii (182), Candidatus Hodgkinia cicadicola y Candidatus Tremblaya princeps (121 genes). Cómo estos organismos se las arreglan para llevar a cabo las funciones celulares básicas sigue siendo un misterio. Conclusiones Durante las etapas tempranas de la evolución de la vida en la Tierra, debieron haber existido sistemas celulares o pre-celulares mucho más simples que las células que conocemos actualmente. Sin embargo, los procariontes descritos anteriormente son organismos modernos que pertenecen a la biología actual y que seguramente evolucionaron por reducción genómica de bacterias con genomas celulares con genomas mayores. Debido a que no es posible manipular genéticamente a dichas bacterias, es probable que la dilucidación de su funcionamiento celular siga siendo un misterio por algunos años más. Sin embargo, dichos organismos nos muestran que la vida celular es posible aun sin la presencia de genes considerados como esenciales. Agradecimientos Agradecemos al apoyo proporcionado por CONACYT a través del proyecto 157220. Referencias
[1] S.N. Peterson, C.M. Fraser. Genome Biology. 2(2): comment2002.1–comment2002.8 (2001). [2] M. Huynen. Trends in Genetics. 16: 116 (2000). [3] C. Henry, R. Overbeek, R.L. Stevens. Biothecnhology Journal.5: 695-704 (2010). [4] H. Morowitz, Beginnings of Cellular Life (Yale Universtity Press, New Haven and London,
1992). [5] R. Gil, F.J. Silva, J. Peretó, A. Moya. Microbiology and Molecular Biology Reviews. 68:518-
537 (2004).
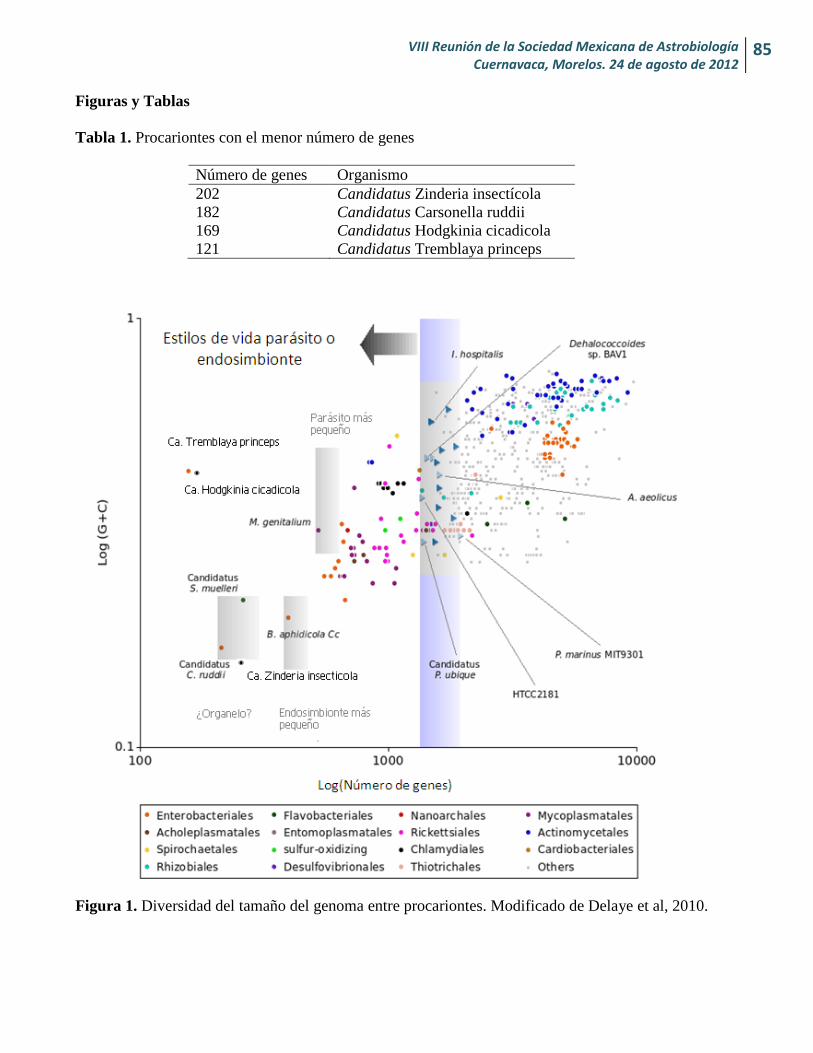
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
85
Figuras y Tablas Tabla 1. Procariontes con el menor número de genes
Número de genes Organismo 202 Candidatus Zinderia insectícola 182 Candidatus Carsonella ruddii 169 Candidatus Hodgkinia cicadicola 121 Candidatus Tremblaya princeps
Figura 1. Diversidad del tamaño del genoma entre procariontes. Modificado de Delaye et al, 2010.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
86
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
BACILLUS PUMILUS: BACTERIA MODELO EN ESTUDIOS DE POTENCIAL DE HABITABILIDAD
Rocío E. Avendaño Serrano, Universidad Autónoma del Estado de Morelos
[email protected] Sandra I. Ramírez Jiménez, Universidad Autónoma del Estado de Morelos
Introducción
Uno de los asuntos de mayor interés en Astrobiología es la posibilidad de que otros objetos planetarios, además de la Tierra, sean habitables. Una manera de evaluar la habitabilidad es recreando en el laboratorio las características del ambiente extraterrestre de interés y estudiar en esas condiciones, las posibilidades de adecuación de algún organismo. Los objetos planetarios que actualmente se identifican con las mayores probabilidades de habitabilidad en el Sistema Solar son una variedad de satélites naturales entre los que se encuentran Europa y Ganímedes, satélites del planeta Júpiter y Encélado, satélite del planeta Saturno. La principal característica de estos cuerpos planetarios es que albergan océanos de agua líquida salada debajo de una gruesa corteza de hielo1, lo que podría sustentar la presencia de algún tipo de vida.
Es importante hacer notar que las sales predominantes en esos océanos son diferentes al cloruro de sodio (NaCl), la sal mayoritaria en los océanos terrestres. La concentración e identidad química de la o las sales presentes en estos océanos se conocerá con exactitud cuando sea posible analizar una muestra obtenida directamente. Existen sin embargo, algunas propuestas realizadas por diferentes grupos de investigación a partir de determinaciones espectroscópicas de la superficie de los satélites. Los espectros sugieren la presencia de sulfato de magnesio (MgSO4) y sulfato de sodio (Na2SO4),3,4,5,6 para los océanos de los satélites Galileanos y para Encélado una salmuera alcalina de sodio, cloruro y bicarbonato (Na, Cl, HCO3
-)7, 8, 9.
La presencia de agua líquida y de sales disueltas en los océanos de estos satélites abre las posibilidades de explorarlos como posibles nichos para el desarrollo de la vida como la conocemos. En este sentido, los organismos extremófilos tienen un papel central. En este estudio nos interesa particularmente, evaluar la posibilidad de que bacterias halófilas o halotolerantes puedan desarrollarse en presencia de sales distintas al NaCl y comprender el mecanismo que utilizan para realizar esa adecuación.
Las bacterias halófilas necesariamente requieren ciertas concentraciones de NaCl para subsistir, mientras que las bacterias halotolerantes son capaces de crecer y desarrollarse en medios que contengan o no NaCl10. Buscamos determinar el potencial de adecuación de Bacillus pumilus, una bacteria halotolerante, en medios modificados con sales diferentes a NaCl, como Mg(NO3)2, MgSO4, (NH4)2SO4 y K3PO4, sales de interés astrobiológico.
Metodología experimental Se usó caldo nutritivo como medio para crecer a Bacilllus pumilus. Este medio de cultivo se
modificó agregando distintas concentraciones de NaCl, K3PO4, Mg(NO3)2, Mg(SO)4 y (NH4)2SO4

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
87
ajustándolas a un valor específico de actividad de agua (aw). El valor de aw de los medios modificados se determinó con un medidor de humedad de punto de rocío (Aqua Lab). Se construyeron cinéticas de crecimiento monitoreando, en intervalos de una hora, la concentración de proteína expresada por la bacteria en cada condición experimental.
Resultados Se realizaron las cinéticas de crecimiento para B. pumilus en NaCl y Mg(SO4)2. Para conocer el
efecto de la concentración e identidad de la sal en el crecimiento de la bacteria se calculó la velocidad específica de crecimiento (µ) en cada uno de los medios modificados.
Una vez calculadas las velocidades específicas de crecimiento se elaboró una gráfica para visualizar el efecto del incremento en la sal adicionada al medio (expresada como aw) sobre la velocidad específica de crecimiento de la bacteria (Fig. 1). Esta gráfica muestra que B. pumilus presenta una mayor velocidad específica de crecimiento en caldo nutritivo libre de sal, como era de esperarse, al ser B. pumilus una bacteria halotolerante. Además nuestros resultados evidencian que a pesar de que B. pumilus presenta menores velocidades específicas de crecimiento en medios que contienen MgSO4, es capaz de desarrollarse en medios modificados con dicha sal. También observamos que la velocidad específica de crecimiento ya sea en NaCl o MgSO4 presenta la tendencia de disminuir conforme disminuye la actividad de agua del medio.
Evaluar la halotolerancia o halofilia de organismos terrestres cuando son expuestos a sales de naturaleza química distinta al NaCl es difícil ya que la mayor parte de los reportes de halotolerancia se expresan en términos de NaCl. En este sentido, este trabajo es un aporte experimental que evalúa la influencia de sales de interés astrobiológico (K3PO4, Mg(NO3)2, Mg(SO)4 y (NH4)2SO4) sobre el crecimiento de bacterias halófilas y halotolereantes con la intención de determinar si favorecen o no su desarrollo. Estos resultados podrían sustentar hipótesis de habitabilidad en las condiciones que prevalecen en los océanos de Europa, Ganímedes y Encélado.
Conclusiones
Bacillus pumilus es capaz de desarrollarse en medios modificados con diferentes concentraciones de NaCl y MgSO4, lo que permite catalogarla como una bacteria halotolerante. Cuando se evalúa la velocidad específica de crecimiento de B. pumilus en medios modificados con NaCl, se observan valores mayores que cuando se evalúa en medios que contienen MgSO4. Por lo tanto puede argumentarse que el desarrollo de la bacteria se ve menos afectado por la presencia de NaCl.
Al incrementar la concentración de NaCl o de MgSO4 en el medio de cultivo, las velocidades específicas de crecimiento son menores. B. pumilus como bacteria halotolerante es capaz de crecer en medios que contienen una sal inorgánica. Sin embargo, la disminución de las velocidades de crecimiento observada cuando se incrementa el contenido de sal en el medio, representa una situación de estrés para la bacteria dificultando su crecimiento.
Referencias [1] Gormly, S., y Adams, V.D., Physical Simulation for Low- Energy Astrobiology Environmental
Scenarios. Astrobiology, 3. (2003). [2] McCord, T., G. Hansen, et al., "Salts on Europa's Surface Detected by Galileo's Near Infrared
Mapping Spectrometer." Science 280: 1242-1245. (1998). [3] Showman, A. and R. Malhotra., "The galilean satellites." Science 286: 77-84. (1999). [4] Kargel, J., J. Kaye, et al., "Europa’s Crust and Ocean: Origin, Composition, and the Prospects for
Life." Icarus 148: 226 –265. (2000).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
88
[5] McCord, T., G. Hansen, et al., "Hydrated salt minerals on Ganymede´s surface: Evidence of an ocean below." Science 292: 1523-1525. (2001).
[6] Orlando, T., T. McCord, et al, "The chemical nature of Europa surface material and the relation to a subsurface ocean." Icarus 177: 528-533. (2005).
[7] Zolotov, M., "An oceanic composition on early and today´s Enceladus." Geophysical Research Letters 34: 1-5. (2007).
[8] Postberg, F., S. Kempf, et al., "Sodium salts in E-ring ice grains from an ocean below the surface of Enceladus." Nature 459: 1098-1101. . (2009).
[9] Postberg, F., J. Schmidt, et al., "A salt-water reservoir as the source of a compositionally stratified plume on Enceladus." Nature. (2011).
[10] Gonzalez -Hernández, J. C. y Peña, A., Estrategias de adaptación de microorganismos halófilos y Debaryomyces hansenii. Revista Latioamericana de Microbiologia. 44, 137-156.(2002).
Figuras
Figura 1. Velocidad específica de crecimiento (µ) para B. pumilus a diferentes valores de actividad de agua debidas a la presencia de NaCl (rombos) y de MgSO4 (cuadros).

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
89
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
UBICACIÓN DE POSIBLES ECOSISTEMAS DE PROCARIONTES REDUCTORES DE SULFATO (PSR) EN EUROPA, SATÉLITE DE JÚPITER
Gpe. Vaneza Y. Peña-Cabrera, Instituto de Geofísica, UNAM
[email protected] Montoya-Lorenzana, L, Instituto Potosino de Investigación Científica y Tecnológica
Durand-Manterola, H.J. Instituto de Geofísica, UNAM
Introducción Morfología de Europa Mediciones hechas por la nave Galileo sugieren que Europa está completamente diferenciada, incluyendo un núcleo rico en hierro, un manto de silicatos y una capa de agua [1]. A diferencia de los otros satélites Galileanos, Europa presenta características tectónicas como domos, bloques interrumpidos, terrenos caóticos, y fracturas [2,3,4]. Los pocos cráteres de impacto sugieren que Europa has estado sometida a algún tipo de repavimentación por procesos tectónicos y/o críovolcánicos. Es difícil de estimar la edad de la superficie de Europa, sin embargo, algunos modelos sugieren una edad entre 10 Ma y 1 billón de años [1].
Europa está sujeta a un calentamiento por fuerzas de marea, ya que la cantidad de hielo es muy baja, el océano puede estar en contacto con el núcleo de silicatos, lo cual hace a Europa bastante interesante desde el punto de vista de la astrobiología.
Información aportada por la misión Galileo, indica la presencia de un campo magnético consistente con una conducción eléctrica interior y con un radio cercanamente igual al radio de la Luna. Este campo requiere de una capa global cerca de la superficie, para lo cual la más probable explicación es un océano salado con una conducción similar a la terrestre, el cual tendría un grosor de 10 km [5,6]. El calentamiento por mareas en la capa explica la presencia de un océano en Europa aún sin involucrar la presencia de un material anticongelante como el amoniaco, sin implicar esto que no haya dicho material. La inducción del campo magnético indica la presencia de iones, como conductores eléctricos y por lo tanto, se puede esperar una reducción en el punto de fusión [1].
Debido a la resonancia de Europa, su excentricidad se mantiene por escalas de tiempo geológico. Por lo que el calor no cesa, lo cual es esencial para que la evolución biológica haya surgido en algún momento de la historia de Europa. Dependiendo de la interacción de marea con Ío y con Júpiter, el calentamiento por mareas puede actuar en Europa por Giga-años evitando que el océano se enfríe y aportando calor al interior de Europa. Por otra lado, la capa de agua está probablemente en contacto con el núcleo de silicatos, permitiendo que se lleven a cabo procesos químicos complejos [7], por lo que en la interfase agua/silicatos las condiciones pueden ser similares a las del piso oceánico terrestre, específicamente como las de las chimeneas hidrotermales donde la vida se desarrolla sin la luz del Sol y donde viven bacterias específicas. Aunque aún no hay evidencia de que estos procesos se lleven a cabo en Europa [1].

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
90
Elementos biogénicos en Europa La abundancia de elementos biogénicos en Europa es desconocida, pero se asume que la composición del cuerpo es como la de los meteoritos condritas carbonáceas, por lo que los elementos biogénicos podrían ser abundantes [6]. Evidencia espectral ha revelado ciertos grupos funcionales (C-N, C≡N) en Ganímedes y Calisto, otras lunas de Júpiter y se ha propuesto su presencia en Europa [8]. Organismos Modelo de Posibles Habitantes en Europa Estados de oxidación del Azufre Aunque son posibles varios estados de oxidación del azufre, solamente tres se forman en cantidades significativas del azufre en la naturaleza, -2 (sulfhidrilo, R-SH y sulfuros, HS), O (azufre elemental, S0) y +6 (sulfato) [9].
La utilización de sulfato para la síntesis de constituyentes celulares que contienen azufre y la subsiguiente descomposición de estos compuestos da como resultado la reducción total de sulfato a H2S, el cual se forma a través de las actividades de diversas bacterias que en conjunto reciben el nombre de bacterias reductoras de sulfato (BSR) [9].
Estas bacterias anaerobias estrictas oxidan compuestos orgánicos e hidrógeno molecular utilizando sulfato como agente oxidante. Se encuentran ampliamente distribuidas en ambientes anóxicos tanto acuáticos como terrestres. La actividad de estas bacterias es patente en el cieno del fondo de los pantanos y arroyos, en turberas y a lo largo de las costas. Como el agua de mar contiene una elevada concentración de sulfatos, su reducción es un importante factor en la mineralización de la materia orgánica de los suelos de las plataformas oceánicas [9].
Las BSR morfológicamente son muy diversas. Se conocen trece géneros de bacterias reductoras de sulfato y se han dividido en dos grupos. Los géneros que se encuentran en el grupo I, las bacterias reductoras de sulfato que no oxidan el acetato, conociéndose también como degradadoras incompletas, utilizan diversos substratos como fuentes de carbono y energía como el lactato, piruvato, etanol o ácidos grasos, oxidan su fuente de energía al nivel de acetato y excretan este ácido graso como un producto final. En el grupo II, las bacterias reductoras de sulfato que oxidan el acetato, conociéndose también como degradadotas completas, oxidan ácidos grasos como el acetato hasta CO2 como producto final [10].
Muchas de las bacterias reductoras de sulfato crecen sobre acetato como única fuente de energía, estos organismos oxidan totalmente el acetato a CO2 y reducen el sulfato a sulfuro por medio de la reacción siguiente [9].
Acetato + SO42-+ 3 H + → 2 CO + H2S + 2 H2O
Desulfobacter psychrotolerans sp. nov. Un grupo restringido nutricionalmente de sulfato-reductores, que usan preferencialmente acetato como su donador de electrones y fuente de carbono, es el género Desulfobacter. Actualmente seis especies han sido descritas, las cuales son mesófilas [11].
Tarpgaard, et al [11] aislaron una bacteria psicrotolerante y acetooxidante de la parte norte del Mar del Norte. La cepa akvbT fue identificada como perteneciente al género Desulforbacter por medio de análisis filogenéticos del gen 16S rRNA. La cepa creció exclusivamente por oxidación de acetato acoplada a la reducción de sulfato. Las células crecieron en un intervalo de temperaturas de -3.6 a 26.3°C, con una temperatura óptima de crecimiento de 20.1°C. Dicha especie fue nombrada como Desulfobacter psychrotolerans sp. nov. Son células móviles que miden 1-3 μm de ancho y 2-5 μm de largo, crecen como agregados, los cuales a menudo se adjuntan a precipitados del medio de cultivo [11].

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
91
En la Tierra un ambiente anóxico y sulfatado con la presencia de hidrógeno o material orgánico es habitable por PRS, si la temperatura del océano de Europa tiene la temperatura en el rango de tolerancia de las PRS psicrófilas, las PRS psicrófilas son un objeto de interés astrobiológico. Existen datos que describen el crecimiento y la actividad de las PRS en bajas temperaturas que ayudarían a inferir la cantidad de ácido sulfhídrico resultante de su metabolismo en condiciones europanas para comprender si la actividad biológica de las PRS sería detectable. Referencias
[1] H. Hussman, J. I. Lunine, 2009. Interiors and Evolution of Ice Satellites. In: Treatise on Geophysics PLANETS AND MOONS. Editores: Schubert G. and Spohn T. Ed. Elservier. Amsterdam. 509-539 pp.
[2] M.H Carr, Belton M.J.S., Chapman C.R. 1998. Evidence for a subsurface ocean on Europa. Nature. 391. 363-365.
[3] R.Greeley, Sullivan R, Klemaszewki J. 1998. Europa: Initial Galileo geological observations. Icarus. 135. 4-24.
[4] R.T. Pappalardo, Head J., Greeley W.R., 1998. Geologic evidence for solid-state convection in Europa´s ice shell. Nature. 391. 365-368.
[5] M.G. Kivelson, Khurana K.K. Krishan K. 2000. Galileo Magnetometer measurements: A Stronger case for a subsurface ocean at Europa. Science. 289. 1340-1343.
[6] C.F. Chyba and C.B Phillips, 2001. Possible ecosystems and the search for life on Europa. PANS. 98 (3). 801-804.
[7] J. S. Kargel, Kaye J.Z. Head J.W. 2000. Europa´s crust and ocean: Origin, composition, and the prospect for life. Icarus. 148. 226-265.
[8] T. McCord, G.B. Hansen, R.N. Clark, P.D. Martin, , C.A. Hibbitts, F.P. Fanale, J.C. Granahan, M. Segura, D.L. Matson, T.V Johnson, W. D. Smythe, J. K. Crowley, P. D. Martin, A. Ocampo, C. A. Hibbitts, J. C. Granahan and the NIMS Team 1998. Salts on Europa's Surface Detected by Galileo's Near Infrared Mapping Spectrometer. J.Gheophys. Res. 103.8603-8626.
[9] R.B.A. Pulido, 2006. Actividad sulfato reductora y desulfurante de cultivos de BSR. Licenciatura en Biología Experimental. Ciencias Biológicas y de la Salud. UAM, Iztapalapa. pp 38; http://148.206.53.231/UAMI13342.pdf
[10] T. D. Brock, Smith D. W. y Madigan M. T. (1993). "Microbiología". Prentice Hall. México. 906 pp.
[11] I.H Tarpgaard; Boetius A;Finster K; 2006: Desulfobacter psychrotolerans sp. nov., a new psychrotolerant sulfate-reducing bacterium and descriptions of its physiological response to temperature changes. Antonie van Leeuwenhoek 89: 109-124; http://www.dnabanknetwork.eu/Query.php?sqlType=Detail&UnitID=DSM%20217155&CollCode=DNA%20Bank&InstCode=DSMZ&UnitIDS=DSM%2017155&CollCodeS=Prokarya&InstCodeS=DSMZ&ID_Cache=14653

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
92
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
MICROORGANISMOS DEGRADADORES DE ACETATO, OBJETO DE ESTUDIO EN ASTROBIOLOGÍA
Lilia Montoya, Instituto Potosino de Investigación Científica y Tecnológica
[email protected] Lourdes B. Celis, Instituto Potosino de Investigación Científica y Tecnológica Elías Razo-Flores, Instituto Potosino de Investigación Científica y Tecnológica
Ángel G. Alpuche-Solís, Instituto Potosino de Investigación Científica y Tecnológica Introducción
De ocurrir la vida fuera de la Tierra, ya sea en Marte o los satélites Europa o Encélado, es probable que sea anaerobia porque a excepción de Europa no se ha propuesto un proceso químico que provea de oxígeno a estos cuerpos del Sistema Solar [1]. Bajo esta condición, los ecosistemas anaerobios conocidos en la Tierra sean extremos o no son objeto de interés para la Astrobiología. Los ecosistemas anaerobios conocidos son: digestores anaerobios, rumen y ambientes hidrotermales [2].
La comprensión del ciclo del carbono presente en los ecosistemas anaerobios es útil para la Astrobiología porque permite el estudio de bioseñales, un ejemplo son las comunidades metanogénicas donde el metano es la molécula terminal de mineralización del carbono. Sin embargo, en Marte y Europa está presente el sulfato [3, 4] y en los ecosistemas anaerobios sulfatados la molécula terminal del carbono es el dióxido de carbono y no el metano.
La mineralización en forma de CO2 la realizan microorganismos acetoclásticos o acetotróficos. Los microorganismos que oxidan la materia orgánica requieren de sulfatos, hierro III, manganeso, entre otros. Cuando estos organismos no están activos por la ausencia del aceptor de electrones la materia orgánica no se degrada completamente y se acumula en forma de acetato [5]. De hecho, en los ecosistemas extremos (por ejemplo, halófilos, acidófilos y psicrófilos) el acetato se acumula. Es por esta razón que el estudio de los microorganismos acetotróficos es de interés astrobiológico.
En el presente trabajo reportamos la riqueza bacteriana presente en una biopelícula proveniente de un digestor anaerobio que exhibe actividad sulfato reductora acetotrófica en condiciones de pH de neutras a ligeramente ácidas. Referencias
[1] Hand, K. P., Carlson, R. W., Chyba, C. F., Energy, chemical disequilibrium, and geological constraints on Europa. Astrobiology 2007, 7, 1006-1022.
[2] Garcia, J. L., Patel, B. K., Ollivier, B., Taxonomic, phylogenetic, and ecological diversity of methanogenic Archaea. Anaerobe 2000, 6, 205-226.
[3] Kargel, J. S., Kaye, J. Z., Head III, J., Marion, G. M., et al., Europa’s Crust and Ocean: Origin, Composition, and the Prospects for Life. Icarus 2000, 148, 226 -265.
[4] Tosca, N. J., Knoll, A. H., McLennan, S. M., Water activity and the challenge for life on early Mars. Science 2008, 320, 1204-1207.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
93
[5] Canfield, D. E., Kristensen, E., Thamdrup, B., Aquatic geomicrobiology. Adv Mar Biol 2005, 48, 1-599.
[6] Garrity, G. M., Bell, J. A., Lilburn, T. G., Taxonomic outline of the Prokaryotes, Springer-Verlag, New York 2004.
Figuras
Figura 1. Árbol filogenético bayesiano inferido con secuencias del gen 16S rRNA de biopelículas acetotróficas obtenidas del presente estudio (en color). Los filotipos de los organismos cultivados más próximos fueron nombrados de acuerdo al manual Bergey’s [6]. El número de filotipos colapsados asignados a cada OTU por RFLP y secuenciación se indica después del número de acceso para cada período operacional del bioreactor (intervalos A, B, C; 22-75, 76-89, 90-110, días, respectivamente). Thermotoga thermarum DSM 9912 (AB039769) se usó como grupo externo. La probabiblidad posterior se indica entre paréntesis después de cada nodo.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
94
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
EXTREMÓFILOS: UNA REVISIÓN
Ricardo A. González Lizardi, Esc. de Téc. Laboratoristas, Universidad del Estado de Morelos [email protected]
Roselena Arroyo Basave, Escuela de Técnicos Laboratoristas, Universidad del Estado de Morelos Sandra I. Ramírez Jiménez, Centro de Investigaciones Químicas, UAEM
Introducción
Los extremófilos son organismos que no solamente sobreviven sino que se desarrollan exitosamente en las condiciones más inhóspitas e impensables para la vida. Son organismos que no sólo toleran y sobreviven a ciertas condiciones naturales extremas, sino que las requieren para poder crecer y multiplicarse.
Estos organismos cuentan con enzimas (catalizadores biológicos) que les ayudan a funcionar en las circunstancias extremas de su entorno. Estos catalizadores sintéticos o enzimas, que son proteínas y que aceleran las reacciones químicas sin alterarlas, permanecen activos cuando otras fallan ante condiciones extremas, eliminando la necesidad potencial de ciertas precauciones, aumentando así su eficacia.
Los primeros organismos extremófilos se descubrieron a mediados de los años setenta. Su estudio ha impulsado el fortalecimiento de proyectos orientados hacia la búsqueda de vida fuera de nuestro planeta; ha abonado en la teoría de la panspermia como una posible explicación del origen de la vida en la Tierra; ha impulsado el desarrollo de la industria biotecnológica, y ha permitido un mejor entendimiento de los factores que sustentan la vida tal como la conocemos.
Resultados
Según la condición más extrema en la que se encuentren, los extremófilos se pueden clasificar en: Termófilos: A medida que la temperatura ambiente aumenta, la velocidad a que se desarrollan
las reacciones enzimáticas aumenta de forma proporcional, lo que implica un desarrollo más rápido del organismo. Esto sucede hasta llegar a una temperatura máxima, por encima de la cual ciertos componentes de la célula comienzan a degradarse de forma irreversible, cesando el crecimiento y sobreviniendo la muerte celular.
Los hipertermófilos como el Sulfolobus acidocaldarius se desarrollan en temperaturas por encima de los 85 °C. El más resistente al calor de estos microorganismos es el Pyrolobus fumarii que crece en las paredes de las fumarolas hidrotermales submarinas.
Psicrófilos: Se desarrollan en ambientes con temperaturas cercanas a los 0 °C, siempre que el agua se encuentre en estado líquido, gracias a que evitan la formación de cristales de hielo en su interior.
Chlamydomonas nivalis es un alga microscópica que aparece en zonas de nieve dándole a ésta un intenso color verde o rojo. Esto es así porque vive en el interior de las capas de nieve en estado vegetativo (verde) y cuando las condiciones se vuelven imposibles, esporula en grandes cantidades y sus esporas son de color rojo. La bacteria Polaromonas vacuolata tiene una temperatura óptima de crecimiento de 4 °C.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
95
Barófilos: Son seres vivos que se desarrollan en ambientes con una presión muy alta. Un ejemplo son las bacterias que habitan las fumarolas o brechas de la corteza terrestre en el suelo marino, a miles de metros de profundidad.
Alcalófilos: Se desarrollan en ambientes tan alcalinos (básicos) que parecen un detergente concentrado y suelen estar asociados a sedimentos con una alta concentración de metano, recordando a lugares como Marte, los cometas, o las lunas heladas de Júpiter o Saturno.
Radiófilos: Soportan gran cantidad de radiación, como un hongo negro que vive en las paredes de los reactores de Chernóbil, o como la bacteria Deinococcus radiodurans que fue descubierta a 3.2 kilómetros de profundidad, asentada en rocas de uranio, un elemento radiactivo, además de estar entre elementos químicos similares a la lejía.
Metanógenos: Son organismos capaces de generar metano (CH4), un gas combustible, en condiciones anaeróbicas (sin oxígeno). Estos organismos viven en el fondo de depósitos acuáticos y en el rumen de algunos mamíferos herbívoros.
Halófilos: Se desarrollan en ambientes hipersalinos, como las bacterias del género Halobacterium que viven en lugares como el Mar Muerto y tienen forma cuadrada. En organismos normales, la sal hace que mueran por deshidratación debido a la fuerte presión osmótica exterior a la que están expuestos. Es decir, se deshidratan y mueren. Los halófilos cuentan con mecanismos de adaptación como el albergar en el interior de sus células, algún soluto compatible, un compuesto orgánico, en concentraciones iguales o mayores a la de las sales del exterior. Así evitan la deshidratación.
Xerófilos: Estos organismos se desarrollan en ambientes con muy baja humedad. En el desierto de Atacama, a una altura de 5,800 metros, se ha localizado en el volcán Socompa una comunidad de microorganismos que viven gracias al dióxido de carbono (CO2), escasa agua y metano que les proporcionan las fumarolas volcánicas.
Acidófilos: Se desarrollan en ambientes de alta acidez, como el Picrophilus. En California, se han descubierto microorganismos increíblemente diminutos que viven cómodamente en el drenaje de las minas a un valor de pH tan bajo como 0.5, equivalente al ácido de una batería de automóvil. Conclusiones
Los extremófilos son organismos sorprendentes debido a que se han adaptado a vivir en condiciones no imaginadas, ya sea en un ambiente salino, o con muy poca humedad, o sometido a mucha presión, o de baja o alta temperatura, entre otros. Para cualquier otro organismo sería imposible vivir en esas condiciones.
Otra cosa muy importante es que los extremófilos no solo se adaptan a esas condiciones sino que las necesitan para poder desarrollarse.
Los extremófilos pueden utilizarse como un modelo de estudio para proponer que podrían existir organismos como ellos en otros planetas o satélites del sistema solar o de otros sistemas planetarios. Por ejemplo en satélites como Titán, una luna de Saturno, que se caracteriza por tener océanos de hidrocarburos, se puede proponer que organismos como los metanógenos tendrían alguna posibilidad de adecuación. Europa es una luna de Júpiter con una corteza hecha principalmente de hielo de agua de un mínimo de 75 a 100 km de grosor, por lo que se puede proponer que organismos psicrófilos puedan tener alguna posibilidad de adaptarse. En esta misma luna se postula que debajo de la capa de hielo existe un océano, posiblemente de agua salada, donde se sugiere que organismos halófilos podrían existir.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
96
Referencias [1] Ramírez, Sandra I. Las fronteras de la vida desde la perspectiva de los extremófilos. Inventio
11, 59-62(2010) [2] Ramírez, Sandra I. Terrazas, Horacio. Astrobiología, una nueva disciplina científica. Inventio 3,
52-55(2006). [3] http://www.cienciapopular.com/n/Biologia_y_Fosiles/Extremofilos/Extremofilos.php
Consultada el 6 agosto de 2012 [4] http://www.earthfacts.com/nature/extremophiles/
Consultada el 5 agosto de 2012 [5] http://www.bacteriamuseum.org/cms/Evolution/extremophiles.html
Consultada el 5 de agosto de 2012 [6] http://serc.carleton.edu/microbelife/index.html
Consultada el 5 de agosto de 2012 [7] http://www.cosmonoticias.org/la-sorpresa-salada-de-encelado/
Consultada el 10 de agosto de 2012 [8] http://ciencia.nasa.gov/ciencias-especiales/27mar_enceladus
Consultada el 10 de agosto de 2012 [9] http://www.astroseti.org/noticia/3632/conceptos-de-astrobiologia-psicrofilos
Consultada el 10 de agosto de 2012

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
97
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
EXTREMÓFILOS ANAEROBIOS EN EL CICLO DEL CARBONO, UNA REVISIÓN
Lilia Montoya, Instituto Potosino de Investigación Científica y Tecnológica [email protected]
Lourdes B. Celis, Instituto Potosino de Investigación Científica y Tecnológica Elías Razo-Flores, Instituto Potosino de Investigación Científica y Tecnológica
Ángel G. Alpuche-Solís, Instituto Potosino de Investigación Científica y Tecnológica Introducción
Los biotopos extremos se refieren a aquellos ambientes que están lejos de las condiciones normales de pH, temperatura, presión, radiación o salinidad, entre otras, siendo las condiciones extremas constantes. A los microorganismos que crecen y realizan su metabolismo bajo tales condiciones extremas son conocidos como “extremófilos”. Sus mecanismos de adaptación van desde estructurales a metabólicos [1,2]. Tales mecanismos de adaptación implican una inversión de energía, por lo que los mecanismos de conservación de energía difieren entre los extremófilos y neutrófilos [3].
El modo de vida anaerobio, añade una restricción en términos bioenergéticos: la energía libre de Gibbs (∆Gº’) proporcionada por estos metabolismos, es menor que por la vía aerobia utilizando el mismo sustrato [4]. Esta diferencia en ∆Gº’ se puede ejemplificar con diversos sustratos como el acetato y explica la localización de los microorganismos en horizontes de una forma jerárquica [5]. Además, la energía disponible para el metabolismo en muchos ambientes anaerobios es tan baja que los microorganismos anaerobios pueden operar cerca del equilibrio termodinámico [6]. Esta aproximación basada en las diferencias de la energía libre de Gibbs es conocida como “ecología termodinámica” [7].
Asímismo, los extremófilos anaerobios constituyen un grupo de interés para analizar aspectos como los límites para la vida y aunque existen revisiones acerca de las adaptaciones biológicas y diversidad de los extremófilos anaerobios [8,9], su papel en el ciclo del carbono tanto en la fijación como en la degradación, no ha sido abordada a pesar de su papel esencial en los ecosistemas anóxicos [10]. Con estos antecedentes, el presente trabajo se enfoca al estudio de los microorganismos extremófilos en su condición de la asimilación del carbono inorgánico y la degradación del carbono orgánico. Inventario de los microorganismos Una síntesis del inventario de los extremófilos autotróficos y degradadores anaerobios en cada biotopo se presenta en la Tabla 1.
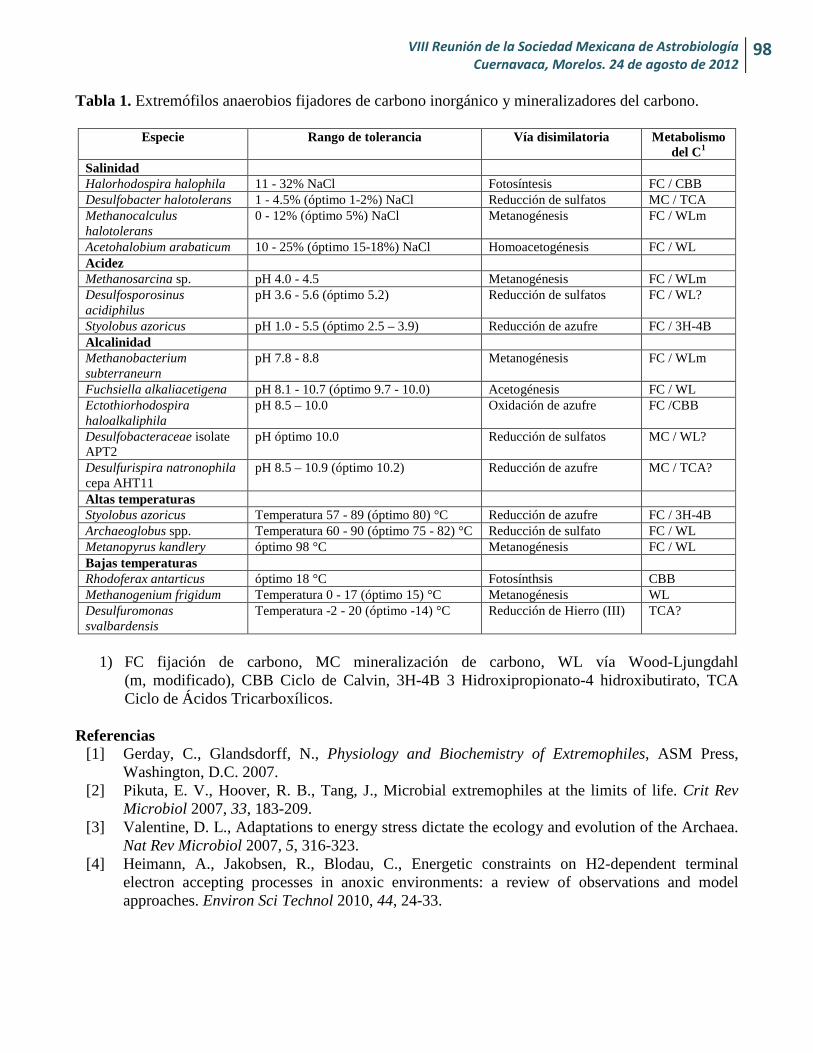
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
98
Tabla 1. Extremófilos anaerobios fijadores de carbono inorgánico y mineralizadores del carbono.
1) FC fijación de carbono, MC mineralización de carbono, WL vía Wood-Ljungdahl
(m, modificado), CBB Ciclo de Calvin, 3H-4B 3 Hidroxipropionato-4 hidroxibutirato, TCA Ciclo de Ácidos Tricarboxílicos.
Referencias
[1] Gerday, C., Glandsdorff, N., Physiology and Biochemistry of Extremophiles, ASM Press, Washington, D.C. 2007.
[2] Pikuta, E. V., Hoover, R. B., Tang, J., Microbial extremophiles at the limits of life. Crit Rev Microbiol 2007, 33, 183-209.
[3] Valentine, D. L., Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nat Rev Microbiol 2007, 5, 316-323.
[4] Heimann, A., Jakobsen, R., Blodau, C., Energetic constraints on H2-dependent terminal electron accepting processes in anoxic environments: a review of observations and model approaches. Environ Sci Technol 2010, 44, 24-33.
Especie Rango de tolerancia Vía disimilatoria Metabolismo del C1
Salinidad Halorhodospira halophila 11 - 32% NaCl Fotosíntesis FC / CBB Desulfobacter halotolerans 1 - 4.5% (óptimo 1-2%) NaCl Reducción de sulfatos MC / TCA Methanocalculus halotolerans
0 - 12% (óptimo 5%) NaCl Metanogénesis FC / WLm
Acetohalobium arabaticum 10 - 25% (óptimo 15-18%) NaCl Homoacetogénesis FC / WL Acidez Methanosarcina sp. pH 4.0 - 4.5 Metanogénesis FC / WLm Desulfosporosinus acidiphilus
pH 3.6 - 5.6 (óptimo 5.2) Reducción de sulfatos FC / WL?
Styolobus azoricus pH 1.0 - 5.5 (óptimo 2.5 – 3.9) Reducción de azufre FC / 3H-4B Alcalinidad Methanobacterium subterraneurn
pH 7.8 - 8.8 Metanogénesis FC / WLm
Fuchsiella alkaliacetigena pH 8.1 - 10.7 (óptimo 9.7 - 10.0) Acetogénesis FC / WL Ectothiorhodospira haloalkaliphila
pH 8.5 – 10.0 Oxidación de azufre FC /CBB
Desulfobacteraceae isolate APT2
pH óptimo 10.0 Reducción de sulfatos MC / WL?
Desulfurispira natronophila cepa AHT11
pH 8.5 – 10.9 (óptimo 10.2) Reducción de azufre MC / TCA?
Altas temperaturas Styolobus azoricus Temperatura 57 - 89 (óptimo 80) °C Reducción de azufre FC / 3H-4B Archaeoglobus spp. Temperatura 60 - 90 (óptimo 75 - 82) °C Reducción de sulfato FC / WL Metanopyrus kandlery óptimo 98 °C Metanogénesis FC / WL Bajas temperaturas Rhodoferax antarticus óptimo 18 °C Fotosínthsis CBB Methanogenium frigidum Temperatura 0 - 17 (óptimo 15) °C Metanogénesis WL Desulfuromonas svalbardensis
Temperatura -2 - 20 (óptimo -14) °C Reducción de Hierro (III) TCA?

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
99
[5] Thullner, M., Regnier, P., Van Cappellen, P., Modeling Microbially Induced Carbon Degradation in Redox-Stratified Subsurface Environments: Concepts and Open Questions. Geomicrobiology Journal 2007, 24, 139-155.
[6] Jackson, B. E., McInerney, M. J., Anaerobic microbial metabolism can proceed close to thermodynamic limits. Nature 2002, 415, 454-456.
[7] Valentine, D. L., Thermodynamic Ecology of Hydrogen-Based Syntrophy, in: Seckbach, J. (Ed.), Symbiosis: Mechanisms and Model Systems, Kluwer Acadaemic Publishers, Dordrecht 2004, pp. 149-161.
[8] Lowe, S. E., Jain, M. K., Zeikus, J. G., Biology, ecology, and biotechnological applications of anaerobic bacteria adapted to environmental stresses in temperature, pH, salinity, or substrates. Microbiol Rev 1993, 57, 451-509.
[9] Ollivier, B., Patel, B. K., Garcia, J. L., in: Seckbach, J. (Ed.), Journey to Diverse Microbial Worlds, Kluwer Academic Publishers, Netherlands 2000, pp. 75-90.
[10] Hugler, M., Sievert, S. M., Beyond the Calvin cycle: autotrophic carbon fixation in the ocean. Ann Rev Mar Sci 2011, 3, 261-289.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
100
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
EVOLUCIÓN MOLECULAR DE LOS EFECTORES SECRETADOS POR Trichoderma spp. Y SU PARTICIPACION EN LA COMUNICACIÓN CON SUS HOSPEDEROS
Mario Iván Alemán Duarte, Laboratorio Nacional de Genómica para la Biodiversidad y Centro de
Investigación y de Estudios Avanzados sede Irapuato [email protected]
Luis José Delaye Arredondo, Centro de Investigación y de Estudios Avanzados sede Irapuato Alfredo Herrera-Estrella, Laboratorio Nacional de Genómica para la Biodiversidad
Introducción
Durante miles de millones de años en la Tierra, la mayor parte de las comunicaciones entre seres vivos ha tenido lugar mediante la transmisión y recepción de moléculas [1]. En patógenos por ejemplo, existen pequeñas proteínas que son capaces de internalizarse hasta el núcleo de la célula huésped y evitar la respuesta de defensa para favorecer la invasión del patógeno [2]. Durante muchos años este mecanismo se creía muy particular de especies patógenas desde bacterias hasta insectos [3], sin embargo descubrimientos recientes demuestran que especies simbióticas como Laccaria bicolo r [4] y Glomus intradarices [5] también utilizan este mecanismo para modular la respuesta de su huésped y establecer una comunicación molecular. Estas proteínas son llamadas efectores [6].
Los miembros del género Trichoderma son hongos capaces de interaccionar con una amplia gama de microorganismos gracias a su habilidad como parásitos, pero también pueden establecer relaciones benéficas con plantas actuando como un simbionte mutualista [7]. Esta amplia variedad de interacciones implica una gran diversidad de efectores con los que establecer comunicación con sus huéspedes.
En la actualidad son pocos los efectores involucrados en la interacción Trichoderma-huésped que han sido identificados y que presentan evidencia experimental. Gracias a que actualmente están secuenciados los genomas de tres especies del género Trichoderma (T. atroviride, T. virens, y T. reesei) es posible realizar un análisis genómico que nos permita identificar la diversidad de proteínas tipo efectores y su posible relación durante la interacción con sus huéspedes.
En un contexto evolutivo, nos preguntamos ¿Cuál es el modelo de evolución que describe mejor la evolución molecular de los efectores de Trichoderma utilizados durante la interacción con sus huéspedes? Métodos Predicción y anotación de proteínas extracelulares
El conjunto de proteínas extracelulares fue predicho como se describe a continuación: los proteomas predichos para Trichoderma virens, T. reesei and T. atroviride se descargaron del sitio web JGI. El conjunto de proteínas extracelulares fue definido en una tuberia automatizada utilizando PERL. Primero usamos SignalP4.0 para identificar aquellas proteínas con péptido señal y sin dominios transmembranales. En este conjunto de secuencias se buscó la presencia de motivos de retención en Retículo Endoplasmático (ERrs [PS00014]) utilizando ScanProsite, y señales de retención en membrana tipo GPI utilizando GPI-Anchored Protein Prediction Tandem System (NN+HMM) .

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
101
Todas las secuencias que tuvieron ERrs y GPI fueron removidas del set inicial. El resto de proteínas fueron anotadas con Blast2GO con los parámetros preestablecidos. A este conjunto nos referiremos como proteínas extracelulares. Identificación de proteínas tipo efector en Trichoderma spp.
Primero integramos una base de datos con 80 efectores reportados en la literatura con evidencia experimental directa utilizando minería de texto con el programa Text-Presso. Identificamos después secuencias homologas en nuestro conjunto de proteínas extracelulares utilizando BLASTp (punto de corte 10E-5). Utilizando PERL nosotros seleccionamos los homólogos si: 1) el punto de corte es igual o menos a 10E-5; 2) el alineamiento es igual o mayor a 50 a.a.; 3) la secuencias de referencia esta representada en más del 50% en el alineamiento con la secuencia blanco. Un análisis similar con INTERPRO fue realizado para identificar aquellas proteínas con una estructura de dominios similar a los efectores con evidencia experimental.
Después identificamos en las proteínas extracelulares motivos de translocación con evidencia experimental, tales como: RxLR, RxFLAK y algunas variaciones utilizando combinaciones de la siguiente firma [RKH]x[LYMFYW][RKH]. Este motivo debería aparecer restringido entre los aminoácidos 15 a 75 en las secuencias.
Todas las secuencias que fueron identificadas como extracelulares y que tienen una o más de las características típicas de los efectores validados fueron consideradas efectores en las tres especias de Trichoderma analizadas. Distribución filogenética y evolución molecular de los efectores en Trichoderma spp
Una vez identificados el conjunto de efectores predichos en Trichoderma sp buscamos los ortólogos en los tres Trichoderma usados y en Giberella zeae and Chaetomium globosum utilizando BranchClust. Los ortológos fueron alineados con muscle e identificamos en mejor modelo usando ProtTest. Después reconstruimos las filogenias para cada ortogrupo utilizando Phyml (100 bootstrap). Estas filogenias fueron usadas para realizar análisis de dn/dS tanto para el modelo por ramas como para el modelo de sitios con el programa codeml del paquete PAML.
Resultados Identificación de proteínas extracelulares
El análisis para identificar proteínas extracelulares, nos permitió identificar 2,625 secuencias en las tres especies, lo cual representa alrededor del 7.86% del proteoma total. La distribución de las proteínas extracelulares entre las tres especies es relativamente similar (Ta=8.16%, Tv=7.76%, Tr=7.69%), interesantemente T. atroviride presenta un ligero incremento en el porcentaje total, este incremento podría estar relacionado a su habilidad como micoparásito.
Identificación de proteínas efectoras
Las búsquedas de homólogos cercanos nos permitió identificar 24 secuencias, que se distribuyendo la siguiente manera: 3 homólogos a repetidos de LysM, 4 a serin-proteasas, 12 a cerato-platanin, 3 a repetidos de Thioredoxina, 1 a Nep1 y 1 a RSP1. El análisis de estructura de dominios con Pfam permitió confirmar las secuencias previamente identificadas e incremento el número de secuencias de la misma familia, además permitió identificar la familia CFEM. Sumando en total 97 secuencias identificadas.
La búsqueda de motivos de translocación mostró ser positivo solo para la firma RxLR y algunas posibles variantes. T. atroviride presenta 86 secuencias, 104 para T. virens y 76 para T. Reesei. Interesantemente algunos de los dominios Pfam presentes en este grupo de proteínas son:

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
102
Glucosilhidrolasas, Peptidasas, LysM y Metaloproteasas, CFEM, entre otros; todos ellos relacionados anteriormente con efectores en otros hongos.
En total se han identificado y categorizado un catálogo con 316 posibles efectores. De estos se seleccionamos 28 genes que pertenecen a 5 familias (repetidos de LysM, de CFEM, de Tioredoxina, homologos a Serin proteasas e Hidrofobinas) que poseen todas las cualidades buscadas y que están presentes en los efectores con evidencia experimental.
Hasta el momento se han analizado 3 genes durante la confrontación con su huésped Rhizoctonia solani en tres tiempos s: antes del contacto (BC), durante el contacto (DC) y sobre crecimiento (OG), los resultados previos muestran un incremento en la expresión de los genes durante la confrontación con su huésped con un máximo en la fase de Contacto (DC).
Por otro lado el análisis de 16 genes seleccionados de nuestro conjunto de efectores fue analizado durante la interacción con Arabidopsis thaliana, de los cuáles 8 presentan inducción durante la colonización de las raíces [7]. La reconstrucción filogenética de los ortogrupos obtenidos y el subsecuente análisis de dN/dS para adentrarnos en su evolución molecular están en proceso. Conclusiones
Nuestro análisis permitió la construcción de un catalogo consistente de posibles efectores en Trichoderma spa que permitirá el análisis funcional de estos candidatos. Los resultados previos de la participación de estos efectores en la comunicación con sus huéspedes arrojan resultados interesantes que nos ayudarán a validar nuestro método de predicción. Sin lugar a dudas adentrarnos en la evolución molecular de estos efectores y como se ven afectados por el estilo de vida de la especie nos permitirá comprender un poco más sobre el complejo dialogo molecular de Trichoderma con sus huéspedes. Referencias
[1] Kadloor, S.; Adve, R.S.; Eckford, A.W.; , Molecular Communication Using Brownian Motion With Drift, NanoBioscience, IEEE Transactions on ,11(2), p.89-99, (2012).
[2] Kanneganti, T.-D., Bai, X., Tsai, C.-W., Win, J., Meulia, T., Goodin, M., Kamoun, S. and Hogenhout, S. A. A functional genetic assay for nuclear trafficking in plants. The Plant Journal, 50 149–158, (2007).
[3] Stergiopoulos I., Harrold A., Ökmen B., Henriek G. B., van Liere S., Kema G., and de Wit P.Tomato Cf resistance proteins mediate recognition of cognate homologous effectors from fungi pathogenic on dicots and monocots. Proc Natl Acad Sci U S A, 107(16): p. 7610-5, (2010).
[4] Plett J.M., Kemppainen M., Kale S.D., Kohler A., Legue V., Brun A., Tyler B.M., Martin F.A secreted effector protein of laccaria bicolor is required for symbiosis development. (2011)Current Biology, 21 (14), pp. 1197-1203
[5] Kloppholz S., Kuhn H., Requena N., A Secreted Fungal Effector of Glomus intraradices Promotes Symbiotic Biotrophy Current Biology 21(14), p. 1204-1209, (2011).
[6] Kamoun, S., Groovy times: filamentous pathogen effectors revealed. Curr Opin Plant Biol, 2007. 10(4): p. 358-65.
[7] Harman, G.E., et al., Trichoderma species-opportunistic, avirulent plant symbionts. Nat Rev Microbiol, 2004. 2(1): p. 43-56.
[8] Guzman P., Olmedo-MOnfil V., Herrera-Estrella A., comunicación personal.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
103
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
ANÁLISIS EVOLUTIVO DE LOS PATRONES DE SELECCIÓN EN LAS CÁPSIDES DE TRES VIRUS ICOSAÉDRICOS: VIRUS DEL MOSAICO DEL FRIJOL DEL SUR, VIRUS
DEL TOMATE Y VIRUS SATÉLITE DE LA NECROSIS DEL TABACO
Karina Espinoza Muñoz, CINVESTAV IPN-ITESI [email protected]
Luis José Delaye Arredondo, CINVESTAV IPN Introducción
Debido al proceso de descendencia con modificación la evolución es un proceso aproximadamente gradual [1]. Al nivel de la genética de poblaciones, los dos mecanismos principales de la evolución son la deriva génica y la selección natural. Si bien la selección natural es el mecanismo que explica las adaptaciones morfológicas, a nivel molecular la mayor parte de la variación es mantenida por un balance entre la deriva génica y las mutaciones neutras o casi-neutras [2]. Con base a múltiples discusiones que se han generado, se han desarrollado numerosos métodos para la detección de selección a nivel molecular, por lo que el objetivo de este trabajo es determinar qué presiones selectivas están actuando en el Virus del Mosaico del frijol del Sur, Virus del Tomate y Virus satélite de la Necrosis del tabaco. Ello debido a que se ha sugerido que en las proteínas que conforman la cápside de estos tres virus icosaédricos existen residuos sujetos a selección positiva que son importantes para la interacción proteína-proteína y por lo tanto para la replicación de los mismos. Métodos Secuencias de Virus y Homólogos
Se utilizo la secuencia de la cápside del Virus del Mosaico del Frijol del Sur (SBMV); Virus del Tomate, (TBSV); Y Virus satélite de la Necrosis del Tabaco, (STNV). Se realizó un BLAST en NCBI para la obtención de homólogos. Reconstrucciones filogenéticas
Las alineaciones se realizaron en el programa MEGAv5 y mediante el programa PHYLIP se obtuvieron las filogenias por el método de Neighbor-Joining (NJ). Traducción de la secuencia de aminoácidos a codones
Para el Análisis filogenético de los patrones de selección por máxima verosimilitud se obtuvieron las correspondientes secuencias codificantes y se alinearon respetando los codones con el programa PAL2NAL. Análisis de Selección
El análisis de selección se realizará en PAML donde se requiere de los siguientes archivos: Secuencia de codones, la reconstrucción filogenética y el archivo control.
En el archivo control se especificarán ciertos parámetros que nos permitirán poner a prueba las hipótesis de evolución: por selección natural o por deriva genética y mutación neutral.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
104
Contraste de hipótesis por el Método LRT (likelihood ratio test)
Utilizando el valor de lnL obtenido con PAML, se realizará una prueba para poder determinar cuál es el modelo que mejor se ajusta a nuestros datos (evolución por selección natural o por mutación y deriva genética).
Análisis de Resultados
El archivo que arroja PAML para el análisis de los resultados es el archivo rst el cual contiene los diferentes valores de omega (número de sustituciones no sinónimas/número de sustituciones sinónimas) para cada sitio. A partir de ahí se van a visualizar estos resultados en graficas en Excel y en la estructura tridimensional en RASMOL. Para identificar los sitios que se sugiere se encuentran bajo selección positiva y que son importantes en el ensamblaje de la cápside, posteriormente se mapearan estos sitios la base de datos Viper [3]. Resultados Hasta el momento sólo se cuenta con las reconstrucciones filogenéticas, donde se observa que a partir de nuestro virus de interés y sus homólogos, se definió una topología del Árbol que le va a permitir a PAML calcular la verosimilitud para los dos modelos de evolución (Figura 1). Referencias
[1] Sosa V. 2009. El Árbol de la Vida. 1-10pp [2] Castillo C. A. 2007. Ecología Molecular: Capitulo 1 La selección natural a nivel molecular. 25-
64 pp [3] Carrillo-Tripp M., Shepherd C.M., Borelli I.A., Venkataraman S., Lander G., Natarajan P.,
Johnson J.E., Brooks III C.L., Reddy V.S. 2008. “VIPERdb2: an enhanced and web API enabled relational database for structural virology”, Department of Molecular Biology, Accepted
Figuras Fig. 1 Muestra la reconstrucción filogenética de la proteína de la cápside del Virus del mosaico del frijol de Sao Paulo Brasil (SBMVSPa) el cual representa el virus sujeto al análisis de selección. En la topología del árbol se observa que este virus está emparentada con el SBMV de Japón.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
105
Presentación en cartel: BIOLOGÍA Y EVOLUCIÓN
ESTANDARIZACIÓN DE UNA TÉCNICA DE EXTRACCIÓN DE ADN MICROBIANO EN MUESTRAS DE AGUA DEL LAGO-CRÁTER RINCÓN DE PARANGUEO, GUANAJUATO,
MÉXICO.
Christian Emmanuel Robles Rivera, Fac. de Ciencias Naturales, Universidad Aut. de Querétaro [email protected]
Mayra Alejandra Campos Hernández, Fac. de Ciencias Naturales, Universidad Aut. de Querétaro Fausto Arellano Carbajal, Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro
Introducción
Rincón de Parangueo (RP) es un volcán tipo mar perteneciente al Eje Neovolcánico Transversal, se localiza dentro del municipio de Valle de Santiago en el estado de Guanajuato [1, 2], RP se ubica a 20°25’de latitud norte, 101°15’ de longitud oeste y una altitud de 1700 msnm [1, 2].
En el volcán RP existe un cráter de aproximadamente 2 kilómetros de diámetro, dentro de este cráter se forma un lago salobre poco profundo rodeado principalmente por una vegetación tropical caducifolia [3] (ver Figura 1). Alrededor de lo que era la costa del lago se forma un anillo conformado por estromatolitos, éstos viven en condiciones altamente salinas y alcalinas y se cree están compuestas principalmente por algas y cianobacterias además de otros tipos de microorganismos. Sin embargo, estos estromatolitos están a punto de desaparecer debido a un proceso de desecación progresiva [2, 4].
Debido a la desecación progresiva el contenido de álcalis ha alcanzado valores muy altos [2]. También las concentraciones de salinidad son altas, desde 1986 se han registrado concentraciones con valores de 53-56 g/l y en el 2002 mayor de 120 g/l, esto se cree que es ocasionado en gran medida por los serios problemas de desecación progresiva que sufre el lago de RP [2].
Ambientes parecidos a los de RP albergan una gran variedad de microorganismos que desempeñan un papel fundamental en el funcionamiento del ecosistema, además estos microorganismos contienen en su genoma un registro de su historia evolutiva [5, 6, 7, 8]. Por desgracia este tipo de ecosistemas cada vez son más vulnerables debido a las sequias y presiones antropogénicas.
El presente proyecto busca caracterizar molecularmente la microbiota mediante el empleo de técnicas en biología molecular, ya que se cree que el volcán funciona como un ambiente altamente conservado y podría albergar una gran diversidad microbiana y altos niveles de endemismo. Debido a las condiciones extremas del hábitat se asume que pueden existir organismos análogos a los que existieron en la Tierra hace millones de años o a los que podrían existir en otros planetas [6]. También es importante hacer muchos otros estudios interdisciplinarios como geológicos, químicos y biológicos para entender con más detalle el funcionamiento de dichos ambientes, además de implementar estrategias de conservación del lugar ya que sufre un serio problema de desecación y esta propenso a la extinción [2]. Métodos
En el mes de noviembre del 2011, se tomaron muestras de agua del lago: 3 en frascos (NALGENE) de plástico de 500 ml previamente estériles de tres distintas charcas donde se presentaba

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
106
un mayor volumen de agua, también se tomaron 10 muestras de agua y sedimento húmedo en frascos (FALCON) de plástico de 50 ml. Las muestras se transportaron selladas herméticamente en una hielera hasta el Laboratorio de Genética Molecular y Ecología Evolutiva de la Facultad de Ciencias Naturales en la Universidad Autónoma de Querétaro. En el laboratorio se almacenaron a 4°C hasta que fueron procesadas las muestras. Se midieron datos fisicoquímicos del agua en el lugar de muestreo como pH, salinidad, temperatura, entre otros, solo se midieron los datos fisicoquímicos de dos charcas, una que presentaba el mayor volumen de agua y la otra donde las charcas estaban en un severo proceso de desecación (ver Tabla 1).
De cada una de las 13 muestras se extrajo el ADN total. El protocolo de extracción de ADN se modifico con base al utilizado por Mwirichia et al. [7], en el cual se usó una solución de lisis (10 mM Tris pH 8.5 + 5 mM EDTA pH 8.0 + 1% SDS (10%) + 10 µl Proteinasa K 20 mg/ml) y una solución concentrada de sales (6 M NaCl). El protocolo consiste en tomar 250 µl de cada muestra en tubos para microcentrífuga de 1.5 µl y agregarle 750 µl de la solución de lisis, se incuban las muestras durante 1 hora a 58 °C y después se agregan 300 µl de la solución concentrada de sales y se centrifugan a 13,000 rpm durante 30 minutos, el sobrenadante se transfiere a tubos nuevos y se le agregan 750 µl de isopropanol, después se incuba a -20 °C por 1 hora y enseguida se centrifuga a 13,000 rpm por 20 minutos, el sobrenadante se retira cuidadosamente para no extraer el pellet de ADN, posteriormente al tubo con el pellet de ADN se le agregan 200 µl de etanol al 70% y se centrifuga a 13,000 rpm durante 2 minutos, por último se retira el sobrenadante y se resuspende el pellet en 50-100 µl de agua destilada estéril y se refrigera a 4°C.
El material genético obtenido se amplificó mediante una PCR (Reacción en Cadena de la Polimerasa) utilizando genes universales ribosomales (ver Tabla 2). Se siguió el protocolo para PCR de HotStarTaq Plus Master Mix (Qiagen). Por último los productos de PCR se visualizarán en electroforesis en gel de agarosa al 1% utilizando como marcador fluorescente bromuro de etidio.
Resultados
Se obtuvieron resultados positivos en la prueba de electroforesis en gel de agarosa al 1% de las trece muestras para Bacteria, Arquea y Eucaria (ver Fig. 2).
Perspectivas
1. Caracterizar molecularmente la diversidad microbiana del lago y de los estromatolitos. 2. Establecer relaciones evolutivas entre los principales grupos taxonómicos de
microorganismos. 3. Establecer las bases sobre la importancia del estudio de ambientes similares a los de Rincón
de Parangueo para entender la evolución temprana de la vida en la Tierra y la posible existencia de vida en otros planetas con condiciones parecidas a las de Rincón de Parangueo.
Referencias
[1] Jiménez B., Marín L., Morán D., Escolero Ó., Alcocer J., Martínez V. El Agua en México vista desde la Academia. (Edición Digital, Academia Mexicana de Ciencias, 2005), 405 pp.
[2] Aranda-Gómez J., Levresse G., Pacheco J., Ramos-Leal J., Carrasco-Núñez G., Chacón-Baca E., González-Naranjo G., Chávez-Cabello G., Vega-González M., Origel G. y Noyola-Medrano C. Eighth International Symposium on Land Subsidence. Querétaro, México, 2010.
[3] Aguilera, L. I. Colegio de Postgraduados (1991). [4] Chacón-Baca E., Berrendero E., Montejo G., Malda J., Sánchez-Ramos M. Are cyanobacterial
mats precursors of stromatolites? 2009. 26 pp.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
107
[5] Souza V., Espinosa-Asuar L., Escalante A., Eguiarte L., Farmer J., Forney L., Lloret L., Rodríguez-Martínez J., Soberó X., Dirzo R., Elser J. PNAS. Vol. 103, no 17. 6565–6570 (2006).
[6] Mormile R., Hong B., Benison K. ASTROBIOLOGY. Vol. 9, no. 10, 919-930 (2009). [7] Mwirichia R., Coustin S., muigai A., Boga H., Stackebrandt E. Curr Microbiol. 1-13 (2010). [8] Gugliandolo C., Lentini V., Maugeri T. Curr Microbiol. 1-8 (2010). [9] Zeng J., Yang H. y Lou K. Journal of Basic Microbiology, 50, 484-493 (2010) [10] Dawson S. C. y Pace N. R. PNAS. Vol. 99, no. 12, 8324-8329 (2002).
Figuras
Figura 1. En las fotografías de la izquierda y el centro se aprecian las charcas en proceso de desecación, y en la de la derecha una vista panorámica del lago en el cráter. Fotografías del autor.
Figura 2. Resultados de electroforesis en gel de agarosa al 1% para cada uno de los 4 pares de oligonucleótidos. Dentro de cada imagen la primera banda muestra la escalera control de 1 kb y las siguientes bandas muestran los resultados correspondientes para cada una de las 13 muestras donde se extrajo ADN. De izquierda a derecha: bacterias, arqueas y las dos últimas eucarias.
Tabla 1. Datos fisicoquímicos del lugar de muestreo y un registro de agua potable de la llave.
pH T amb °C T agua °C mbar mS/cm tds ppt sal. ORP DO% Domg/l Lago 9.4 29 25.91 833.6 124.5 62.23 70 -150 69.3 2.34
Charco 9.93 29 26.09 832.8 140.4 70.13 70 -124 8.3 0.3 Agua llave 7 24 23.85 810.3 967 484 0.52 159.1 26.2 1.76
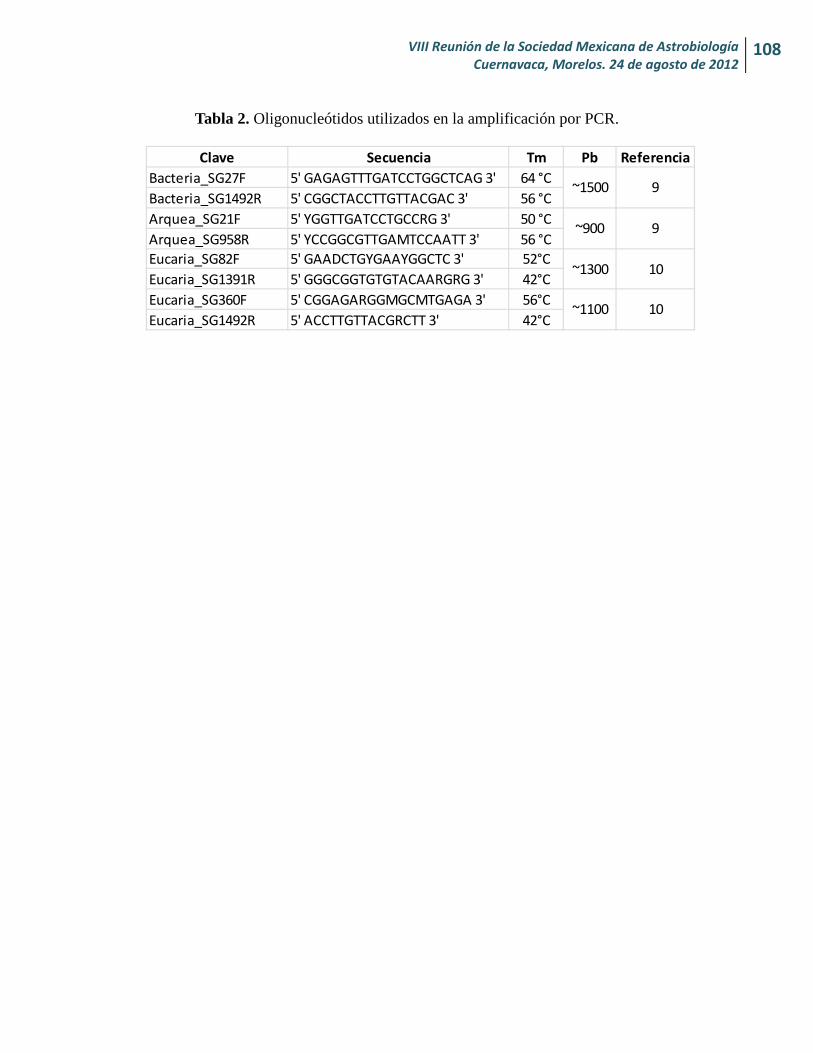
VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
108
Tabla 2. Oligonucleótidos utilizados en la amplificación por PCR.
Clave Secuencia Tm Pb ReferenciaBacteria_SG27F 5' GAGAGTTTGATCCTGGCTCAG 3' 64 °CBacteria_SG1492R 5' CGGCTACCTTGTTACGAC 3' 56 °CArquea_SG21F 5' YGGTTGATCCTGCCRG 3' 50 °CArquea_SG958R 5' YCCGGCGTTGAMTCCAATT 3' 56 °CEucaria_SG82F 5' GAADCTGYGAAYGGCTC 3' 52°CEucaria_SG1391R 5' GGGCGGTGTGTACAARGRG 3' 42°CEucaria_SG360F 5' CGGAGARGGMGCMTGAGA 3' 56°CEucaria_SG1492R 5' ACCTTGTTACGRCTT 3' 42°C
~1100 10
9
9
10
~1500
~900
~1300

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
109
Presentación en cartel: MEDICINA ESPACIAL
EFECTO DE LA POSICION SUPINA EN LA DESCALCIFICACION
María Elena Aguilar Mena, Instituto Nacional de Enfermedades Respiratorias [email protected]
Introducción Uno de los grandes retos en viajes tripulados al espacio de larga duración es evitar la descalcificación que tiene lugar a microgravedad. La calcificación comienza desde el desarrollo intrauterino (1) y se incrementa con la bipedestación. En dicho proceso intervienen la hormona paratiroidea y la calcitonina secretada por la tiroides, además de un buen aporte de calcio vitamina D, la integridad del intestino para su absorción y la integridad renal, así como la fuerza gravitatoria y la luz ultravioleta (2). Sin embargo, existen condiciones en Tierra en las cuales estamos expuestos a gravedad cero. Estas son: la posición supina prolongada (acostado boca arriba en la cama) como la que guardan algunos pacientes crónicamente enfermos y la natación sobre la superficie del agua, donde por principio de Arquímedes nuestro cuerpo flota eliminando la fuerza gravitatoria. El hueso se origina de la capa germinal embrionaria llamada mesodermo (3) Tiene una parte central o medular formadora células sanguíneas y una parte cortical formada por precipitados de apatita de calcio. Los fémures son los huesos más largos y fuertes, dan soporte a todo el cuerpo, teniendo una epífisis radial, una metáfisis donde se encuentra el cartílago hialino de crecimiento y una diáfisis o parte central (4). La arquitectura del fémur le permite dar no sólo soporte al segmento superior con el inferior sino articularlos y permitir la marcha y la carrera. Su primer punto de osificación primitivo aparece en el segundo mes del embarazo. Otros cuatro puntos de osificación complementarios para la cabeza de fémur, el trocánter mayor, el trocánter menor y el de la epífisis distal se forman 15 días antes del nacimiento. La osificación se completa a los 19 años de edad. Se han realizado estudios con personas en posición de decúbito supino por 60 días para verificar los cambios en la masa muscular de los glúteos en reposo total y con carga ponderal así como ejercicio vibratorio (5) corroborando los investigadores que el reposo total en cama por más de 60 días ocasiona atrofia muscular. Por otra parte esta atrofia muscular y la propia descalcificación ósea por falta de uso conduce al paciente a la mayor propensión a fracturas impacto mínimo (6) lo cual prolonga los periodos de rehabilitación y el costo intrahospitalario. Material Se presenta el caso de un paciente crónicamente enfermo que ingresó a los dos meses de edad con problema pulmonar secundario a infección y prematurez, estando internado en terapia intensiva pediátrica bajo ventilación mecánica, hasta su total resolución, posteriormente pasó a terapia intermedia. Su dependencia al oxígeno, la desnutrición GIII y el hecho de haber presentado complicaciones propias de su prematurez a otros niveles prolongó hasta más de un año su estancia, bajo el influjo lumínico artificial y la posición supina. Después de un año de vivir bajo esta condición el paciente presentó una fractura de fémur, al ser únicamente abrazado por la mamá.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
110
Método Además de tratar al paciente multidisciplinariamente. Se revisó literatura sobre el efecto de la
posición supina a largo plazo. Las revistas de 2005 al 2012 con impacto 4 a 5.
Resultados En 2007 los autores Benjamin y Boudignon realizaron una publicación de los resultados de
experimentar con ratas sometidas a microgravedad a las cuales les administraron IGF-I 2.5 mg Kg día por dos semanas contra un grupo control al que no le fue administrado el factor. Ellos observaron aumento en el número de osteoblastos, de hueso esponjoso y densidad ósea (7). Smith y Zwart y colaboradores en 2008 publicaron sus resultados de un estudio realizado en 15 varones voluntarios que por 21 días estuvieron a en posición supina con cabeza baja a 6 grados simulando condiciones de vuelo, a 8 de ellos se les administró una gravedad artificial sometiendo a una Gz por una hora diaria la región cercana al corazón y a 2Gz la región podálica, los voluntarios que no recibieron el beneficio de la microgravedad tuvieron mayor excreción urinaria de calcio, mayor fosfatasa alcalina y la densidad mineral ósea de todo el cuerpo disminuyó (8). Bradley fue uno de los primeros investigadores que a nivel molecular trató de dilucidar el por qué ocurren dichos cambios, con 10 mujeres voluntarias a las que les aplicó ejercicios aeróbicos, de resistencia y mixtos contradijo el conocimiento en voga de que la masa muscular y ósea dependía de proteínas fijadoras de IGF-I (9). En los dos últimos años se han publicado las investigaciones sobre los señaladores celulares en la síntesis o degradación muscular, usando modelos animales en posición antigravitatoria y con personas en reposo total en cama se ha concluido que el señalador AKT de la síntesis proteica muscular aún no está clara, pues varía de un momento a otro, ya sea en reposo, comiendo o durante el ejercicio de rehabilitación, lo que sí está claro es que los niveles de AKT se encuentran muy disminuidos en el decúbito prolongado lo que ocasiona hipotrofia muscular generalizada (10). La repercusión en el músculo cardiaco en la posición supina prolongada ocasiona el adelgazamiento e hipotrofia de la masa muscular cardiaca con la disminución de su función, que se haya compensada por la disminución de la resistencia vascular sistémica y el flujo preferencial hacia el segmento superior durante el vuelo, no así en el reposo en cama por largo tiempo (11).
Conclusiones El decúbito supino ocasiona hipotrofia muscular generalizada, así como la pronta descalcificación
ósea, lo que favorece las fracturas llamadas de “cama” en aquellos pacientes que por su condición se ven confinados a situaciones de reposo prolongado. Aunque estas fracturas son comentadas en el ámbito médico y a pesar de que las ciencias espaciales nos han aportado un caudal creciente de conocimientos, estas lesiones siguen ocurriendo. No se trata sólo de los cuidados y la nutrición sino de los esfuerzos en los ejercicios de rehabilitación temprana, es decir, en la misma cama del paciente, ya sea con vibradores de alta frecuencia como electroestimuladores de grupos musculares, la aplicación de pesas acordes a la edad y la condición, para el entrenamiento muscular y aumentar de esta forma la resistencia muscular y por consiguiente la calcificación ósea y la aplicación de la helioterapia que usaron los médicos de la antigüedad, para evitar en lo posible daños secundarios y mayor permanencia, costo y dolor.
Agradecimiento
Al personal de la Biblioteca del Instituto Nacional de Enfermedades Respiratorias que me ayuda con la obtención del material y a mis compañeros de trabajo que me impulsan con sus críticas sanas y sus buenos deseos.

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
111
Referencias [1] L. Cortés, D. Garzón, R. Cárdenas, Rev Cub Orth Traum 23(2), 1-25. (2009). [2] W. Gannong. Fisiología Medica ( Manual Moderno ed.20ª, 2006 ), pp.359-371. [3] WJ. Larsen. Human Embriology ( Elsevier Science ed 3th, 2002), pp.315-347. [4] F. Netter. Atlas de Anatomía Humana ( Elsevier Masson ed.4ª. 2007 ), Secc.7. [5] T. Miokovic, G. Armbrecht, D. Felsenberg, J Appl Physiol 110, 926-934 (2011) [6] K. Marimuthu, AJ. Murton, P. Greenhaff, J Appl Physiol 110, 555-560 (2011) [7] M. Benjamin, D.Boudignon et al, J Appl Physiol 103,125-131 (2007) [8] SM. Smith, S.R.Zwart et al, J Appl Physiol 107, 47-53 (2009) [9] C.Bradley, J.Nindl et al, J Appl Physiol 109,112-120 (2010) [10] K. Marimuthu, AJ. Murton, et al, J Appl Physiol 110, 555-560 (2011) [11] R.L. Hughson, J.K. Shoemaker, et al, J Appl Physiol 112, 719-727 (2012)

VIII Reunión de la Sociedad Mexicana de Astrobiología Cuernavaca, Morelos. 24 de agosto de 2012
112
Directorios
Sociedad Mexicana de Astrobiología
Dra. Antígona Segura Peralta Presidente
Dra. Sandra I. Ramírez Jiménez Vicepresidente
Dra. Leticia Carigi Delgado Secretaria General
Dr. Luis Delaye Arredondo Secretario Administrativo
Dra. Javiera Cervini Silva Secretaria de Vinculación
M. en C. Irma Lozada Chávez Secretaria de Difusión
Dra. Guadalupe Cordero Tercero M. en A. Eduardo Alberto Piña Mendoza
Secretarios de Finanzas Manet Estefanía Peña Salinas
Representante de los Estudiantes
Universidad Autónoma del Estado de Morelos
Dr. Alejandro Vera Jiménez Rector
Dra. Patricia Castillo España Secretaria Académica
Dr. Gustavo Urquiza Beltrán Secretario de Investigación e Innovación
Dr. Mario Fernández Zertuche Director del Centro de Investigaciones Químicas
Biol. Juan Carlos Sandoval Manrique Director de la Facultad de Ciencias Biológicas
Dra. Sandra I. Ramírez Jiménez Representante del Cuerpo Académico de Química y Física del Ambiente





























