Embed Size (px)
Citation preview

165
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
Correspondencia:Jaime Robledo, Unidad de Bacteriología y Micobacterias,Corporación para Investigaciones Biológicas, Carrera 72ªNo.78B-141, Medellín, Colombia.Teléfono: 441 0855; fax: 441 [email protected]
Recibido: 15/08/03; aceptado: 31/01/04
Inactivación de genes de Mycobacterium tuberculosis ysu potencial utilidad en la prevención y el control de
la tuberculosisOfelia Chacón 1,2, Teresa Realpe 1, Raúl Gerardo Barletta 2, Jaime Robledo 1,2,3
1 Unidad de Bacteriología y Micobacterias, Corporación para Investigaciones Biológicas, Medellín, Colombia.2 Department of Veterinary and Biomedical Sciences, University of Nebraska, Lincoln, Nebraska, USA.3 Facultad de Medicina, Universidad Pontificia Bolivariana
El conocimiento derivado del genoma de Mycobacterium tuberculosis, junto con el desarrollode sofisticados sistemas para la manipulación genética del bacilo, ofrece la mayor promesapara el desarrollo de herramientas nuevas y más eficientes para prevenir y controlar latuberculosis. Se han desarrollado métodos más eficientes para la inactivación de genesmicobacterianos que se han convertido en el pilar de la genómica funcional micobacteriana.La generación de mutantes mediante la inactivación génica, apoyada directa o indirectamentepor el desciframiento del genoma micobacteriano, ha permitido la generación de un númerosignificativo de mutantes de M. tuberculosis. En algunos casos, el análisis de estas mutantesha establecido relaciones entre los productos génicos y sus funciones en la fisiología y lapatogenicidad de la micobacteria. En esta revisión se describen los estudios másrepresentativos basados en dichas mutantes.
Palabras claves: Mycobacterium tuberculosis, inactivación de genes, genómica funcional,mutantes atenuadas.
Gene inactivation in Mycobacterium tuberculosis and its use in tuberculosis control andprevention
Availability of the M. tuberculosis genome sequence and the development of sophisticatedsystems for genetic manipulation of bacilli offer the potential for new and effective tools toprevent and control tuberculosis. Efficient methods to inactivate mycobacterial genes havebeen developed. These methods have become the cornerstone for the application anddevelopment of mycobacterial functional genomics. Specific mutants are generated to establishthe role of targetted genes associated with mycobacterial physiology and pathogenesis. Geneinactivation, supported directly or indirectly by the deciphering of the mycobacterial genome,has permitted the generation of large numbers of M. tuberculosis mutants. Analysis of thesemutants has (in some cases) established relationships between gene products and their role inmycobacterial physiology and pathogenesis.
Key words: Mycobacterium tuberculosis, gene inactivation, functional genomic, attenuatedmutants.
La identificación de Mycobacterium tuberculosiscomo agente causal de tuberculosis hecha porRobert Koch en 1882 marcó el primer hito en el
REVISIÓN DE TEMA
estudio de esta enfermedad (1). Estos hallazgos,seguidos por el desarrollo y refinamiento de lastécnicas de coloración y cultivo realizadas porPaul Erhlich, Franz Ziehl y Friedrich K.A. Nielsenproporcionaron las primeras herramientas paracombatir racionalmente la tuberculosis (2). En másde un siglo desde la introducción de estasherramientas, se han desarrollado una vacuna yvarios agentes quimioterapéuticos. Aun así, en laactualidad, la tuberculosis sigue siendo la mayor

166
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
causa de muerte en el mundo debida a un únicoagente infeccioso (3,4).
Se sabe que casi un tercio de la población mundialestá infectada con el bacilo tuberculoso, el cuales la causa de muerte de 2 millones de personas,aproximadamente, cada año. Esta situación haempeorado recientemente por factores como laaparición de cepas de bacilos multirresistentes alos medicamentos antes efectivos en eltratamiento y por la pandemia VIH/sida. Deacuerdo con este panorama, se necesitan nuevasestrategias para prevenir y controlar eficiente-mente esta enfermedad.
Las características propias de M. tuberculosisplantean un verdadero reto para los investigadores.Por ejemplo, M. tuberculosis tiene un tiempo dereplicación lento, sus requerimientos para elcrecimiento in vitro son exigentes y presentadificultades para la manipulación genética.Afortunadamente, el conocimiento reciente delgenoma de la micobacteria y el desarrollo de latecnología genómica han proporcionadoinstrumentos para el desarrollo de nuevastécnicas que permiten la manipulación genéticadel bacilo (5-8). La inactivación de genesespecíficos y el estudio de los efectos que dichasinactivaciones tienen en las funcionesestructurales y fisiológicas de la micobacteria,incluso sus efectos en la virulencia, así como elestudio de la esencialidad de los genes quecodifican posibles blancos para agentesterapéuticos, se han convertido en la piedraangular de la aplicación de la genómica funcionalcomo disciplina para el estudio de la tuberculosis.
Actualmente, la genómica funcional está abriendoun camino totalmente nuevo en el estudio de latuberculosis y su utilización, probablemente,llevará al diseño racional de estrategias para laidentificación de blancos potenciales para laintervención terapéutica, el diagnóstico y lageneración de vacunas.
En esta revisión, que no pretende ser exhaustivapor la imposibilidad de cubrir la totalidad del tema,se abordan los hallazgos recientes suministradospor la manipulación genética y la inactivación dealgunos genes relacionados con la estructura o elmetabolismo de M. tuberculosis. Se describen
brevemente las estrategias que se han usado parainactivar diferentes genes y los posibles papelesde cada gene en la fisiología o en el proceso depatogénesis. Esperamos que esta información seade utilidad en el entendimiento de la genéticamicobacteriana y que sea aplicable en el diseñoy en el desarrollo de nuevas medidas de control yprevención de la tuberculosis.
Para un mejor entendimiento de la informaciónproporcionada, se describirán inicialmente algunasde las metodologías utilizadas para la inactivacióngénica en micobacterias y, posteriormente, variosestudios en los cuales se han generado mutantesde M. tuberculosis con el uso de algunas de estasmetodologías. Esto último se presenta clasificandolos genes en: aquéllos relacionados con labiosíntesis de la pared celular - un componenteestructural que es cuantitativa y cualitativamenteimportante para la fisiología micobacteriana - ysus proteínas asociadas; los relacionados con labiosíntesis, el metabolismo o el transporte demetabolitos y nutrientes como aminoácidos,metales y ácidos grasos, y los genes involucradosen procesos complejos que juegan papelesimportantes en la adaptación micobacteriana adiferentes factores medio-ambientales como laregulación de la transducción de señales, latranscripción y la resistencia a los mecanismosbactericidas de los macrófagos.
Técnicas utilizadas parala inactivación génica de micobacterias
Las micobacterias, especialmente aquéllas decrecimiento lento como M. tuberculosis, sonmicroorganismos difíciles de manipular genética-mente. Los métodos utilizados inicialmente paragenerar mutantes de micobacterias se basaronen el uso de diversos agentes mutagénicos comola luz ultravioleta y los agentes químicos (9). Sinembargo, estos métodos no fueron lo suficiente-mente efectivos como para permitir un desarrollorápido de la genética funcional micobacteriana.Décadas más tarde, se aplicaron exitosamenteotras técnicas como el empleo de vectoresadecuados que incluían plásmidos, fagos,fásmidos y transposones que permiten lainactivación de genes por recombinaciónhomóloga y la generación de mutaciones al azar.
167
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
La inactivación de genes por recombinaciónhomóloga consiste en la introducción de una copiamutada del gen en la micobacteria. Esta copia serecombina luego por mecanismos de recom-binación homóloga con el gen sin mutación", elcual se reemplaza en el cromosoma mico-bacteriano (figura 1). Mientras que algunos pocosensayos han tenido éxito introduciendo el genmutado como un fragmento lineal (10-13), lamayoría de los métodos, como se presenta másadelante, se basan en el uso de plásmidos y fagos.
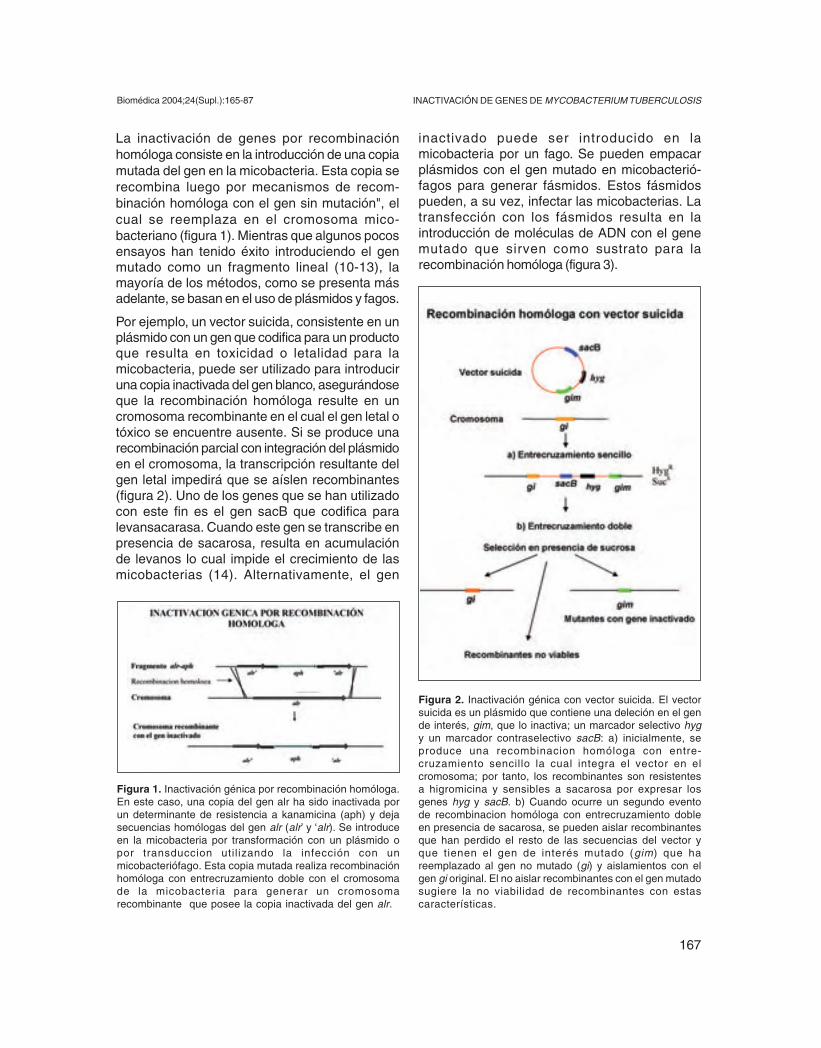
Por ejemplo, un vector suicida, consistente en unplásmido con un gen que codifica para un productoque resulta en toxicidad o letalidad para lamicobacteria, puede ser utilizado para introduciruna copia inactivada del gen blanco, asegurándoseque la recombinación homóloga resulte en uncromosoma recombinante en el cual el gen letal otóxico se encuentre ausente. Si se produce unarecombinación parcial con integración del plásmidoen el cromosoma, la transcripción resultante delgen letal impedirá que se aíslen recombinantes(figura 2). Uno de los genes que se han utilizadocon este fin es el gen sacB que codifica paralevansacarasa. Cuando este gen se transcribe enpresencia de sacarosa, resulta en acumulaciónde levanos lo cual impide el crecimiento de lasmicobacterias (14). Alternativamente, el gen
Figura 1. Inactivación génica por recombinación homóloga.En este caso, una copia del gen alr ha sido inactivada porun determinante de resistencia a kanamicina (aph) y dejasecuencias homólogas del gen alr (alr’ y ‘alr). Se introduceen la micobacteria por transformación con un plásmido opor transduccion util izando la infección con unmicobacteriófago. Esta copia mutada realiza recombinaciónhomóloga con entrecruzamiento doble con el cromosomade la micobacteria para generar un cromosomarecombinante que posee la copia inactivada del gen alr.
Figura 2. Inactivación génica con vector suicida. El vectorsuicida es un plásmido que contiene una deleción en el gende interés, gim, que lo inactiva; un marcador selectivo hygy un marcador contraselectivo sacB: a) inicialmente, seproduce una recombinacion homóloga con entre-cruzamiento sencillo la cual integra el vector en elcromosoma; por tanto, los recombinantes son resistentesa higromicina y sensibles a sacarosa por expresar losgenes hyg y sacB. b) Cuando ocurre un segundo eventode recombinacion homóloga con entrecruzamiento dobleen presencia de sacarosa, se pueden aislar recombinantesque han perdido el resto de las secuencias del vector yque tienen el gen de interés mutado (gim) que hareemplazado al gen no mutado (gi) y aislamientos con elgen gi original. El no aislar recombinantes con el gen mutadosugiere la no viabilidad de recombinantes con estascaracterísticas.
inactivado puede ser introducido en lamicobacteria por un fago. Se pueden empacarplásmidos con el gen mutado en micobacterió-fagos para generar fásmidos. Estos fásmidospueden, a su vez, infectar las micobacterias. Latransfección con los fásmidos resulta en laintroducción de moléculas de ADN con el genemutado que sirven como sustrato para larecombinación homóloga (figura 3).

168
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
En el caso de mutación por transposones, el gense inactiva debido a la inserción de un fragmentomóvil de ADN (transposón). La inserción de estosfragmentos es al azar y, por tanto, con estesistema no es posible predecir cuál o cuáles genesserán inactivados. Al introducir los transposonespor intermedio de plásmidos o de fagos se puedengenerar librerías de mutantes que deben, luego,ser analizadas para buscar característicasespecíficas de las mutantes de interés (figura 4).
Figura 3. Recombinación homóloga con fásmidos. 1) Elgen de interés es inactivado por un determinante deresistencia a higromicina (gim:hygR) y subclonado en uncósmido. 2) Este se alinea y se liga al genoma del fago. 3)El cósmido es, luego, empacado en un fago termosensiblepara generar un fásmido. 4) Se infecta E. coli portransducción y se recupera el fásmido. 5) Éste seelectropora en M. smegmatis y se incuba a 30oC para inducirel ciclo lítico. 6) Los micobacteriófagos recuperados seutilizan para infectar la micobacteria blanco en la cual ocurrela recombinación homóloga entre el gen original (gi) y elgen mutado (gim). 7) Las colonias recombinantes con lamutación se recuperan a 37oC por ser resistentes a lahigromicina.
Figura 4. Inserción génica con transposones. Untransposón con un gen determinante de resistencia a lahigromicina (hyg), las secuencias invertidas IR y el gen dela transposasa A (tnpA) se introducen en las micobacteriaspor medio de transducción utilizando un micobacteriófago.La integración del transposón ocurre al azar y generamutantes en diferentes genes que se pueden seleccionaren medio con higromicina.
Se han utilizado algunas variaciones para hacermas rápido y efectivo el análisis de librerías demutantes, por ejemplo, la mutagénesis del tiposignature tag, en las cual el transposón lleva unapequeña secuencia que permite identificarlo(15,16), o hibridizaciones con el sitio del transposón(TRASH), en las cuales se utilizan microarreglospara detectar los genes que poseen inserciones(17).
Inactivación de genes involucrados enla biosíntesis de la pared celular yde proteínas asociadas
La pared celular y sus proteínas asociadas estánsituadas estratégicamente en la estructura externade la micobacteria y juegan un papel importanteen la interacción patógeno-hospedero. En estosmicroorganismos, el efecto bactericida deinhibidores de la síntesis de la pared celularconfirma la esencialidad de este componente parala supervivencia de la bacteria. Además, variasproteínas asociadas con la pared celular se hanpropuesto como factores de virulencia (18-20).
La pared celular de la micobacteria tiene unaconformación compleja de glicolípidos y proteínasasociadas con la estructura central micolil-arabinogalactano-peptidoglicano (21,22). Elarabinogalactano, el mayor componente de lapared celular, está unido a los residuos de ácido

169
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
murámico del peptidoglicano (23,24). Estamolécula tiene substituciones de ácidos micólicos,los cuales son ácidos grasos de alto pesomolecular. En M. tuberculosis, los ácidosmicólicos son ciclopropanos y se pueden clasificarde acuerdo con la presencia y la naturaleza delos sustitutos que contienen oxígeno en la porcióndistal de la rama del meromicolato, en ácidosalfa micólicos no oxigenados y ácidos oxigenadosceto y metoximicólicos (25-29). La síntesis de losácidos micólicos involucra varias metiltrans-ferasas necesarias para la formación de los anillosciclopropanos y las ramas metiladas (25-28,30,31).
Aunque la mayoría de los ácidos micólicos en lapared celular de la micobacteria se encuentrancomo ésteres ligados al arabinogalactano, algunostambién son transferidos por las micoliltransferasas a la α−α'-trehalosa para formarglicolípidos extraíbles, incluso α−α'-trehalosamonomicolato (TMM) y α−α'- trehalosa dimicolato(TDM) o factor cordón (18,23). Otros componentesextraíbles que se han identificado en la paredcelular de M. tuberculosis, incluyen ioceroldimicocerosato, sulfolípidos y lipoarabinomanam(18,23).
Se han sugerido papeles importantes para loscomponentes individuales de la pared celularmicobacteriana en la patogenicidad y laresistencia a medicamentos del bacilo tuberculoso.Por ejemplo, las alteraciones en las proporcionesy en las estructuras de los ácidos micólicos sehan relacionado con variaciones en la fluidez ylapermeabilidad de la pared celular. Estasvariaciones pueden, a su vez, estar relacionadascon la resistencia de las micobacterias asustancias bactericidas (32). Algunas de lasfunciones sugeridas para los componentes de lapared celular micobacteriana se han confirmadopor análisis genético recientes en M. tuberculosis(16,28-30,33,34).
Genes involucrados en la síntesis ytransporte de dimicocerosatos
Los dimicoserosatos (DIM), inclusive eldimicoserosato tiocerol (PDIM), parecen jugar unpapel impor tante en la virulencia de lamicobacteria (35). La síntesis de DIM involucravarios genes que incluyen los genes ppsA-E que
codifican para una policétido sintasa requeridapara la síntesis de phtiocerol (36), el gen masque codifica para una sintasa del ácidomicocerósico (37,38) y el gen fadD28 que codificapara una acil-coA sintasa, la cual parece jugarun papel importante en la transferencia acil delácido micocerósico a phtiocerol (39).
En M. tuberculosis, estos genes están localizadosen una región de 50 kb del cromosoma en el quetambién se localizan los genes drrA-C quecodifican para un polipéptido similar al del ligadordel transportador ABC (40), el gen mmpL7 quecodifica para una proteína conservada demembrana de micobacterias (8), el gen fadD26que codifica para una acil-coA sintasa (39,41) yel gen epapA5 que codifica para una policétidosintasa asociada con proteínas de funcióndesconocida (8).
En un aislamiento clínico de M. tuberculosis, cepaMt103, utilizando mutagénesis con transposones,se generaron mutantes de los genes fadD26,fadD28, mmpL7 y drrC. Estos mutantes seaislaron luego de una selección por su atenuaciónpotencial en pulmones de ratones BALB/cinfectados por vía intravenosa (15). Utilizando lamisma estrategia se han generado mutantes dela región promotora en la cepa Erdman de M.tuberculosis que incluyen los genes faD26, ppsA-E y los genes fadD28 y mmpL7. Estos últimos seaislaron luego de seleccionarse por su inhabilidadpara replicarse en el interior de los pulmones deratones C57BL/6 infectados por vía intravenosa(16).
El análisis de los lípidos de células enterasextraídos de la mutante que contiene insercionesen la región que incluye los genes fadD26 y ppsA-E, revelaron que tenían cantidades pequeñas oindetectables de PDIM (15,16). Lo anterior sugiereque la expresión de los genes ppsA-E podría estaracoplada a la expresión del gene fadD26 situadoinmediatamente corriente arriba (15,36). Futurasinvestigaciones deberán dirigirse a desentrañar elpapel individual de las proteínas PpsA-E y FaD26en la virulencia de M. tuberculosis.
Estudios adicionales de las mutantes fadD26- dela cepa Mt103 han demostrado que, en ausenciade DIM, se observa una susceptibilidad

170
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
aumentada a los detergentes como el SDS (sodiododecil sulfato), aunque no se afecta lasusceptibilidad a reactivos intermediarios delnitrógeno (NRI) ni a antibióticos hidrofóbicos ohidrofílicos (40). Estos últimos resultados sugierenque la atenuación observada, probablemente, esel resultado de la susceptibilidad a losmecanismos de muerte en macrófagos diferentesal NRI.
Los análisis de mutantes también han reveladoun papel importante de MmpL7 en el transportede DIM. El estudio de los lípidos de célulasenteras, extraídos de las mutantes mmpL7- de lacepa Erdman de M. tuberculosis, mostró unaacumulación significativa de PDIM maduro (16).Adicionalmente, el análisis de la distribuciónextracelular del DIM en las mutantes mmpL7- ydrrC- del aislamiento clínico Mt103 mostró quesus productos son esenciales para la translocacióndel DIM (40).
Además de lo anterior, Cox y colaboradores hanmostrado en mutantes ppsA-E-, mmpL7- y fadD28-generados en la cepa Erdman de M. tuberculosis,una incapacidad para crecer en pulmones deratones y una velocidad de crecimiento en bazo yen hígado similar a la cepa original (16). Estudiosfuturos,que deben incluir las mutantes y sus cepascomplementarias correspondientes, podríanindicar si estos genes se requieren para unareacción tisular específica.
En la síntesis de DIM también se ha demostradola participación de, al menos, uno de los genessimilares a los que codifican para una sintasa delácido micoserósico, denominados msl. Estosgenes codifican para una serie de dominioscatalíticos requeridos para la síntesis de ácidosgrasos saturados.
Dos de estos genes, el msl6 (pks12) y el msl5(pks8 y pks17), fueron inactivados en la cepaH37Rv mediante recombinación homólogautilizando un sistema de fagos (42,43). Lainactivación del gen msl5 no afectó el crecimientode la cepa en macrófagos alveolares murinos oen pulmones de ratones C57BL6/J infectados pormedio de aerosoles (43). Por el contrario, lainactivación del gen msl6 resultó en una drástica
disminución en la síntesis de DIM, acompañadade reducción en el crecimiento tanto enmacrófagos alveolares murinos como enpulmones de ratones C57BL6/J infectados por víaintranasal (42).
A pesar de los estudios anteriores, aún quedanpor determinarse las bases moleculares quepuedan explicar la asociación de DIM convirulencia. No es claro todavía si PDIM o DIMfuncionan solamente en la superficie celular de lamicobacteria, afectando la permeabilidad celulary, posiblemente, la sensibilidad a los mecanismosbactericidas de los macrófagos o si, además, suexcreción en el hospedero es importante para lavirulencia y la modulación de la respuesta inmune.
Genes involucrados en la síntesis o en latransferencia de los ácidos micólicos y lastrehalosas
El papel de los ácidos micólicos en la patogénesisde la tuberculosis se ha confirmado por análisisgenéticos recientes en los cuales se haninactivado los genes involucrados en la síntesisy la transferencia de estos ácidos (29,30,33,34).En uno de estos estudios, el gen pcaA (umaA2)que codifica para una ciclopropano sintasarequerida para la síntesis del anillo proximalciclopropano de los ácidos micólicos α, fueinactivado por recombinación homóloga en la cepaErdman de M. tuberculosis (30). Las mutantespcaA- exhibieron una morfología alterada del factorcordón y un perfil de ácidos micólicos igualmentealterado. Aunque las tres clases mayores deácidos micólicos estaban presentes, loscetomicolatos eran significantemente másabundantes en la cepa mutante y se presentaronmodificaciones en el alfa micolato, como un ligeroincremento en la cantidad de ácido micólico trans-cis-ciclopropo. El fenotipo alterado de factor cordónde la mutante pcaA- sugiere que el contenidolipídico de la envoltura celular podría relacionarsecon las características morfológicas de la bacteria(30).
Estudios in vivo revelaron que estas mutantes seeliminaron más rápidamente en ratones C57B1/6infectados por vía venosa al compararlas con lacepa original. Los ratones infectados con la

171
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
mutante también exhibieron un daño pulmonarmenos importante. Los resultados anterioressugieren que la ciclopropano sintasa proximal esnecesaria para la supervivencia a largo plazo y lavirulencia de M. tuberculosis in vivo (30). Futurasinvestigaciones podrían revelar si las alteracionesen el contenido de ciclopropano de los lípidosasociados con la pared celular en la mutantepcaA-, incluso TDM, están relacionadas conmodulaciones diferentes de la respuesta inmuneen el hospedero.
Se generaron por recombinación homóloga cepasErdman de M. tuberculosis con mutaciones enlos genes cmaA2 y mmaA2. Estos genes codificanpara una sintasa de ácidos micólicos trans-ciclopropano y una ciclopropano sintasa distal,respectivamente (28,44). Los análisis de lamutante cmaA2- han indicado que el cmaA2 serequiere para la síntesis de los anillos trans-ciclopropano de los ácidos micólicos oxigenados.En el caso de la mutante mmaA2-, se detectarondefectos en la cis-ciclopropanación de los αmicolatos (28,44). Estudios futuros podríaninvolucrar análisis adicionales de estas mutantestanto in vivo como in vitro.
La inactivación del gen hma (cmaA, mmaA) quecodifica para una enzima que se requiere para lasíntesis de ácidos micólicos oxigenados, hasugerido la forma como este tipo de ácidosmicólicos participa en la patogénesis de lasmicobacterias. Dubnau et al. inactivaron el genhma en la cepa H37Rv de M. tuberculosis porrecombinación homóloga. La comparación de lacomposición de los lípidos de la cepa original yde la mutante hma- reveló que esta última esincapaz de sintetizar metoximicolatos ocetomicolatos. Estos cambios parecen estaracompañados de modificaciones en lapermeabilidad de la pared celular en la cepamutante, como lo sugieren una captacióndisminuida de moléculas hidrofílicas ehidrofóbicas, su acumulación y una granresistencia al peróxido de hidrógeno (H
2O
2) (29).
Estudios in vitro de esta mutante revelaron queaunque el crecimiento en la línea celular THP-1no se afectó, hubo una disminución significativaen su crecimiento en ratones C57BL/6 infectados
mediante aerosoles. La mutante, además,persistió luego de varias semanas de infección,lo cual sugiere que su atenuación para elcrecimiento probablemente no se debió a unincremento en la susceptibilidad a los mecanismosbactericidas del hospedero, sino a un efecto en elestado nutricional de la cepa. Estudios futuros eneste campo podrían confirmar si la incapacidadde la mutante hma- para adquirir nutrientesespecíficos puede explicar su comportamientodurante el crecimiento en tejidos de ratón o en elinterior de macrófagos humanos (29).
Entre los genes involucrados en la transferenciade ácidos micólicos están los que codifican parael complejo del antígeno 85 (Ag85) (33,45). Elcomplejo está constituido por los antígenos 85A,85B, 85C, codificados por los genes fbpA, fbpB yfbpC2, respectivamente. Estos antígenos sonproteínas ligadoras de fibronectina que catalizanla transferencia de micolatos a trehalosa, un pasonecesario para la síntesis de α,α'-trehalosamonomicolato (TMM) y el factor cordón (18). Seha identificado un cuarto miembro de esta familia,codificada por el gen fbpC1, pero su actividadligadora micoliltransferasa no ha sido aúndemostrada (8,46). Un posible papel del complejoAg85 en la virulencia de M. tuberculosis podríaexplicarse por su función en la biosíntesis de lapared celular. Las proteínas del complejo Ag85también podrían jugar un papel en la virulencia delas micobacterias debido a su habilidad para ligarfibronectina, característica que se ha relacionadocon la virulencia en otros microorganismospatógenos (47). Además, debido a suspropiedades ligadoras de fibronectina, se hapropuesto un papel importante para el complejoAg85 en la entrada de micobacterias patógenas ala célula (48,49).
Armitige y colaboradores obtuvieron mutantes delos genes fbpA y fbpB en la cepa de M.tuberculosis H37Rv por medio de recombinaciónhomóloga (33). Mientras que la inactivación delgen fbpA resultó en crecimiento disminuido enmedio de cultivo mínimo en la línea celular humanaTHP-1 o en la línea celular de macrófagos murinosJ774.A1 (33), la inactivación del gen fbpB no afectóel crecimiento de la mutante bajo las mismas

172
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
condiciones. El análisis de las cepascomplementadas confirmaron estos resultados.
Estudios futuros podrían demostrar si lasalteraciones observadas en la mutante fbpA- sedeben al requerimiento de lípidos o de otroscompuestos que se presentan usualmente en losmedios enriquecidos o a una susceptibilidadincrementada a lípidos tóxicos que de otra maneraserían ligados por la albúmina presente en mediosenriquecidos (33). Además, una alteración en elprocesamiento de fagosomas o en la activaciónde los macrófagos por esta mutante tambiénpodría resultar en una supervivencia intracelulardeteriorada. En relación con lo anterior, estudiosprevios han mostrado resultados promisorios enratones inmunizados con vectores de ADN quellevan el gen del Ag 85A (50), lo cual sugiere unposible papel de este antígeno en la respuestainmunológica a la infección por M. tuberculosis.
La inactivación del gen fbpC2 en un aislamientoclínico de M. tuberculosis, obtenida pormutagénesis con transposones, no afectósignificativamente el crecimiento de la cepa enmedio mínimo o en macrófagos derivados demédula ósea de ratones BALB/c (34). Estoshallazgos fueron similares a los descritos para lainactivación del gen fbpB en la cepa de M.tuberculosis H37Rv. Como la capacidad deligamiento a fibronectina es común para FbpA,FbpB y FbpC2, estos resultados sugieren que elefecto de la inactivación del gen fbpA en lospatrones de crecimiento podría no estardirectamente relacionado con la capacidad deadhesión a la fibronectina.
El análisis de lípidos de la mutante fbpC2-MYC1554 no mostró diferencias significativas enla naturaleza y en las cantidades relativas demicolatos entre la cepa original y la mutante. Sinembargo, la cepa mutante fbpC2- contenía 40%menos micolatos ligados a la pared celular que lacepa original, lo cual sugiere que el Ag 85C podríaestar involucrado en la transferencia de micolatosa arabinogalactano. Además, los estudios depermeabilidad celular de esta mutante sugierenque la capa de micolatos puede servir comobarrera para la difusión de pequeñas moléculashidrofílicas e hidrofóbicas. Este fenómeno fue
independiente de las subsecuentes víasmetabólicas del substrato y no pareció afectar ladifusión de isoniacida y otros antibióticos a travésde la capa de micolatos (34).
Estudios futuros centrados en el papel específicodel complejo del antígeno 85 y los antígenosrelacionados codificados por el gen fbpC1 en lavirulencia e inmunogenicidad micobacteriana,podrían ser de importancia tanto para el diseñoracional de agentes antimicobacterianos dirigidoscontra estos antígenos como para la generaciónde potenciales mutantes inmunogénicasatenuadas.
Los ácidos micolipanoico, micolipénico ymicolipodienoico constituyen los mayorescomponentes de las trehalosas poliaciladas (51).La inactivación del gen msl3 (pks3 y pks4) en lacepa H37Rv de M. tuberculosis por transducciónespecializada generó una mutante que no producíaácido micolipanoico ni ácido micolipénico y,además, no producía trehalosas poliaciladas (51).Se necesitan estudios adicionales de estamutante para demostrar el posible papel del genmsl3 en la virulencia de M. tuberculosis.
Genes involucrados enla síntesis de sulfolípidos
Los sulfolípidos son ácidos grasos metilados yesterificados a trehalosa sulfato (52-54). Se hasugerido que juegan un papel importante en lapatogénesis de las infecciones por micobacterias,posiblemente, a través de la interacción con lascélulas efectoras del sistema inmune delhospedero. Estas interacciones incluyen lainhibición de la activación de los macrófagos(55,56) y la inducción de cambios en la funcióncelular fagocítica, relacionados con procesosinflamatorios exagerados (57).
Se ha descrito la inactivación del gen pks2 (msl2)en la cepa H37Rv de M. tuberculosis porrecombinación homóloga. Este gen codifica parauna policétido sintasa involucrada posiblementeen la síntesis de ácidos hidroxiptioseránicos, elmayor componente acil de los sulfolípidos. Lacaracterización bioquímica de los lípidos celularesreveló que estos mutantes son deficientes ensulfolípidos (58).

173
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
Más recientemente, se ha descrito el gen mmpl8,un miembro de los 13 integrantes de la familiammpl ya mencionados como importantes en eltransporte de lípidos. Este gen (mmp18) juega unpapel importante en la síntesis de sulfolípido-1(SL-1) al igual que el gen de la policétido sintasa2 (pks2). La generación de mutantes mmpl8- ypks2- por recombinación homóloga con un sistemade fagos demostró un defecto acentuado en lasíntesis de SL-1 en ambas mutantes. El análisisde estas cepas en ratones C57/B6 infectados porvía venosa sugieren que SL-1 no es absolutamenteindispensable para la replicación de M. tuberculo-sis durante la infección y que MmpL8 es importantepara la persistencia y el crecimiento bacterianosostenido (59).
Los hallazgos anteriores cuestionan el papeldirecto de SL-1 en la patogénesis de la fase agudade la infección, pero sugieren su importancia enlas formas crónicas y persistentes lo que señalasu potencial como blancos para el diseño denuevos medicamentos antituberculosos quetengan utilidad en el tratamiento de las formaslatentes de la enfermedad.
Genes que codifican para proteínasasociadas con la pared celular
Las proteínas asociadas con la pared celularincluyen las proteínas exportadas que se asociancon la envoltura celular después de sertranslocadas a través de la membranacitoplasmática (60). Entre estas proteínas que seasocian con la pared celular de las micobacterias,están el complejo del antígeno 85, descritoanteriormente (18,61), y la proteína Erp (20). Laproteína Erp contiene una secuencia señal N-terminal y se sospecha que sea translocada porla vía dependiente de la proteína Sec (60).
Se ha descrito la inactivación del gen erp (pirG),que codifica para la proteína Erp, en la cepaH37Rv de M. tuberculosis por medio derecombinación homóloga. La mutante que seobtuvo exhibió disminución en su replicación,efectos citopáticos reducidos en los macrófagosderivados de la médula ósea de ratones BALB/cy disminución en la supervivencia de los ratonesBALB/c infectados por vía intravenosa (20). Estosresultados, junto con el análisis de microscopía
inmunoelectrónica de macrófagos J774, indicanque la proteína Erp se produce en los fagosomasde M. tuberculosis y, posiblemente, tiene tráficointracelular (20). Esto sugiere un importante papelpara esta proteína durante el desarrollo delfagosoma y en la multiplicación del patógeno enel interior de la célula hospedera. Los estudiosfuturos deben enfocarse en entender el papelespecífico que juega la proteína Erp en losprocesos relacionados con el estado intracelulary la patogénesis del microorganismo. En estecontexto, estudios recientes han demostrado quela naturaleza del gen erp afecta significativamenteel número y el tamaño de las lesiones pulmonaresen ratones BALB/c infectados por vía venosa; seobservaron lesiones individuales de mayor tamañoy mayor superficie total acumulada de las lesionesen la mutante erp- complementada con el gen erpproveniente de cepas de micobacterias virulentas,comparadas con la mutante complementada congenes provenientes de cepas no virulentas (62).
Inactivación de genes relacionados con labiosíntesis, el metabolismo y el transportede metabolitos y nutrientes
Genes involucrados en la biosíntesis deaminoácidos. Las micobacterias son capaces desintetizar todos los aminoácidos necesarios parasu crecimiento; se han descrito fenotiposatenuados auxotrópicos de aminoácidos en estosmicroorganismos (63-65). La biosíntesis deaminoácidos en las micobacterias parece tenerlas vías usuales descritas en otros micro-organismos (66,67). No obstante, existendiferencias que parecen encontrarse en losmecanismos reguladores para estas vías (66,67),tal como lo ha mostrado la vía de la familia delaspartato en M. smegmatis (68). El estudio demutantes auxotróficas para aminoácidos es unaherramienta para entender las característicasambientales que confronta la micobacteria quereside en el interior del macrófago y losmecanismos que el microorganismo hadesarrollado para sobrevivir en este tipo deambiente. Además, las mutantes auxotróficasatenuadas también se pueden considerar comopotenciales candidatas a vacunas.
Entre los genes involucrados en la biosíntesis yla captación de aminoácidos en los cuales se ha

174
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
descrito inactivación en M. tuberculosis, están:lysA, leuD, proC, metB, hisD, trpD, argF y glnA1.Estos genes codifican respectivamente paramesodiaminopimelato decarboxilasa, isopropil-malatoisomerasa, pirroline-5-carboxilato-reductasa, cistatión-gamma sintasa, histidinoldehidrogenasa, antranilato fosforibosiltransferasa,ornitina carbamoiltransferasa y glutamina sintasa(68-72). Las anteriores enzimas están involucradasen el metabolismo o en la biosíntesis de losaminoácidos: lisina, leucina, metionina, histidina,prolina, tr iptófano, arginina y glutamina,respectivamente (67,69-78).
En la cepa H37Rv de M. tuberculosis se hadescrito la inactivación de los genes lysA, leuD,proC, metB, hisD, trpD y argF medianterecombinación homóloga (68-71). El gen glnA1 hasido inactivado en M. tuberculosis cepa Erdmanpor intercambio alélico utilizando un vectortermosensible (72).
Aunque la inmunización única de ratones C57BL/6 con la mutante de lysA no confirió proteccióncontra un reto con M. tuberculosis virulentaadministrada por aerosol, el protocolo con dosdosis de la mutante resultó en niveles deprotección similares a los conferidos por la BCG.Además, el auxótrofo lysA- no persistió en ratonesinmunocompetentes ni en ratones inmuno-comprometidos, probablemente debido a laincapacidad de esta mutante de sobrevivir enausencia de lisina. Se ha sugerido el uso potencialde esta mutante auxotrófica como una vacuna derefuerzo independiente de replicación (79).
Los patrones de crecimiento intracelular hanmostrado que la mutante leuD- es incapaz dereplicarse en el interior de macrófagos derivadosde la médula ósea de ratones BALB/cJ, encontraste con la cepa original y con las mismascepas complementadas (69). Esto sugiere que laleucina del hospedero no es utilizada por lamicobacteria intracelularmente. Esta mutante nofue superior a la BCG en términos de protecciónen ratones BALB/cJ retados con M. tuberculosis(69). Los estudios futuros con este modelo animalpodrían dirigirse a definir si este comportamientopuede explicarse por un bajo nivel de persistenciade las mutantes auxotróficas leu-.
En contraste con las características descritaspara las mutantes leu-, la inactivación del genemetB- generó una cepa no auxotrófica que exhibióuna tasa de supervivencia similar a la de la cepanativa en macrófagos derivados de la médula óseade ratones BALB/c. Esta cepa fue virulenta enratones SCID infectados por vía intravenosa, loscuales han mostrado ser altamente susceptiblesa la infección con M. tuberculosis. La mutantemetB- tuvo una menor virulencia en ratonesinmunocompetentes DBA infectados por víaintravenosa (80). Las futuras investigaciones sonnecesarias para indicar si la actividad MetBresidual estaba presente en la mutante metB- osi el producto de otro gen puede compensar en lamutante la pérdida de la actividad MetB. Los genescandidatos para estas funciones substitutasincluyen el gen metZ el cual se ha identificado enla secuencia genómica de M. tuberculosis (8), quetambién puede codificar para una cistationina γ-sintasa (70).
En relación con las características observadasen las mutantes proC- y trpD-, como son lareducción significativa en la habilidad paramultiplicarse en el interior de los macrófagosderivados de la médula ósea de ratones BALB/cy menor virulencia en ratones SCID y DBA, se hasugerido un papel importante de estos genes enla virulencia de M. tuberculosis (80). La vacunaciónen modelos animales utilizando estos mutantesy retados con M. tuberculosis confirió niveles deprotección similares (mutante proC-) o más altos(trpD) que los conferidos por BCG, lo cual indicaun potencial interesante de estas cepasatenuadas como candidatas a vacunas (80).Similarmente, el fenotipo atenuado de la mutanteglnA1- en macrófagos humanos THP-1 y encobayos infectados por aerosoles sugiere queeste gen juega un papel importante en la virulenciade M. tuberculosis (72).
Genes involucrados en la captación de hierroy en la regulación dependiente de hierro. Lasmicobacterias, como la mayoría de losorganismos vivos, requieren hierro para unapropiado crecimiento y supervivencia (66,81).Evidencias previas sugieren que la captación dehierro y su regulación pueden jugar un importantepapel en la patogénesis de las infecciones

175
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
causadas por micobacterias, tal como se hadescrito en otros patógenos (66,81). En algunosde estos organismos, el hierro juega un papelimportante en la regulación de genes y, porejemplo, la no disponibilidad de este nutrientepuede producir represión de genes de virulencia(67,81,82).
El bacilo tuberculoso es capaz de obtener hierroextracelular a través de la micobactina T, unsideróforo hidrofílico que transfiere el hierro amicobactinas solubles en lípidos y asociadas conla célula (83,84). Varias enzimas involucradas enla biosíntesis de micobactinas son codificadaspor el grupo mbt, el cual incluye los genes mbtAa mbtH (8, 67,85).
La expresión de éstos y otros genes relacionadoscon el transporte de hierro parecen estar bajo elcontrol de genes reguladores. Los genesreguladores responsables del hierro en otrasbacterias codifican para las proteínas Fur y DtxR,las cuales son capaces de inhibir la expresiónbajo concentraciones altas de hierro mediante launión a secuencias palindrómicas conservadasen los promotores blanco (86-88). Se haidentificado un homólogo del gen dtxR en M.tuberculosis. Este gen denominado ideR codificapara una metaloproteína dependiente de hierro quereconoce una secuencia promotora específica('caja de hierro') en otros genes regulados por estemetal (8, 89).
Una mutante ideR- de M. tuberculosis H37Rv fuegenerada por recombinación homóloga utilizandoun vector suicida. Debido a la esencialidad delgen ideR, esta mutante solamente se pudo aislaren la presencia de una segunda copia funcionaldel mismo gen. La mutante generada exhibiómayor sensibilidad al estrés oxidativo comparadacon las cepas original y complementada. Elanálisis posterior de la mutante permitió laidentificación de varios genes regulados por hierroy por IdeR, inclusive endogenes codificadores yproteínas involucradas en la síntesis desideróforos y en el almacenamiento de hierro (89).
El gen mbtB codifica una enzima que, aparente-mente, cataliza algunos de los primeros pasosen la biosíntesis de la micobactina T, incluso lacondensación del ácido salicílico con serina y la
formación de la estructura hidroxifeniltiazolina deeste sideróforo (90). Este gen se inactivó en lacepa de M. tuberculosis H37Rv a través derecombinación homóloga (91). El análisisestructural de la mutante demostró crecimientoen presencia del ácido salicílico [7-14C] y revelóque esta cepa no parecía incorporar el marcadoren material extraíble de células enteras. Unaporción del marcador se encontró incorporado enun metabolito liberado en el medio de cultivo, perono se le demostró capacidad de ligar hierro. Losensayos funcionales revelaron, además, que lamutante no exhibió la capacidad de quelar hierro.Estos resultados, junto con la observación decrecimiento deficiente de esta mutante en mediopobre en hierro, sugieren un importante papel paralas moléculas queladoras de hierro en elcrecimiento de M. tuberculosis (91).
El papel de los sideróforos en la replicación de lamicobacteria en el interior de macrófagoshumanos está adicionalmente soportado por elcrecimiento defectuoso de la mutante mbtB- encélulas THP-1 comparado con la cepa nativa. Elcrecimiento defectuoso no se modificó por laincubación previa de los microorganismos enmedio enriquecido con hierro (91). Lasinvestigaciones futuras podrían enfocarse en eldetalle estructural y en la caracterización funcionalde las micobactinas de M. tuberculosis, laidentificación de los papeles específicos quejuegan otros genes mbt y los genes relacionadoscon mbt en la biosíntesis y la virulencia de lasmicobacterias. Además, debe abordarse elestudio y la identificación del papel que jueganlos genes reguladores en la expresión de genesdependientes del hierro en M. tuberculosis.
El gen mramp, un homólogo micobacteriano delgen que codifica para la proteína de macrófagos1, asociada con la resistencia natural (Nramp1),parece jugar un papel importante en la captaciónde varios cationes, incluso Fe2+ y Mn2+. Unamutante mramp- de M. tuberculosis MT103generada por recombinación homóloga exhibiócrecimiento alterado bajo condiciones limitantesde hierro. El crecimiento y la supervivencia de lamutante en líneas celulares de macrófagosmurinos y su supervivencia en ratones BALB/cinfectados por vía venosa no se alteraron, lo cual

176
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
indica que Mramp no juega un papel realmenteimportante en la virulencia de M. tuberculosis enel modelo murino (92).
Genes involucrados en el transporte demagnesio. La habilidad para transportar cationescomo el magnesio parece ser un mecanismo deadaptación bacteriano que le permite a lospatógenos intracelulares sobrevivir dentro delfagosoma (11,93). Se ha sugerido que en M.tuberculosis este mecanismo puede estarinvolucrado en la supervivencia del micro-organismo dentro de un fagosoma ligeramenteacídico y limitado en Mg2+ (11).
El gen mgtC, que codifica para una proteína Ctransportadora de magnesio, fue identificado enM. tuberculosis basado en la identidad de suproducto con el del gen mgtC de Salmonellaenterica (8,11). Este gene se inactivó en la cepaErdman de M. tuberculosis por recombinaciónhomóloga con un fragmento lineal que conteníael gen inactivado (11).
El análisis de los patrones de crecimiento de lamutante en diferentes medios indicó que el genmgtC se requiere para el crecimiento de M.tuberculosis bajo condiciones de bajaconcentración de Mg+. Este efecto fue másevidente bajo condiciones levemente acídicas. Lamutante mgtC- también exhibió crecimientoalterado en macrófagos derivados de monocitoshumanos y menor supervivencia en ratonesBALB/c infectados por vía venosa, lo cual indicaque el gen mgtC juega un papel importante en elcrecimiento de M. tuberculosis en macrófagoshumanos y en su virulencia en ratones (11).
Genes involucrados en el metabolismo deácidos grasos. Los ácidos grasos son la mayorfuente de carbono y energía que puedencatabolizarse o incorporarse en complejos lipídicosy se ha sugerido que juegan un importante papelen el metabolismo de M. tuberculosis y de otrasbacterias (94,95). El catabolismo de los ácidosgrasos incluye el ciclo de la β-oxidación, la cualproduce varias moléculas de acetil-CoA y la víaacortada del glioxilato, la cual está involucradaen la utilización de las moléculas de acetil-CoAgeneradas por la vía de la β-oxidación (95).
La expresión de la isocitrato liasa (Icl), una enzimaderivada del glioxilato, está aumentada enmicobacterias durante la infección de macrófagosy durante la adaptación para sobrevivir bajocondiciones de anaerobiosis (96-99). El gen quecodifica para esta enzima, icl, ha sido inactivadoen la cepa Erdman de M. tuberculosis porrecombinación homóloga (94).
La mutante icl- obtenida por recombinaciónhomóloga mostró persistencia y virulenciadisminuida en ratones BALB/c infectados por víaintravenosa, comparada con las cepas nativa ycomplementada. La virulencia de esta mutante enratones IFN-γ-/- infectados por vía intravenosa,sugiere la posible relación entre el estado inmunedel hospedero y el estado metabólico de M.tuberculosis in vivo (94).
La evaluación de la expresión Icl-Gfp (isocitratoliasa-green fluorescent protein) en cepastransformadas con un plásmido que contiene lafusión icl-gfp, en macrófagos murinos de médulaósea inactivados y activados con INFγ y con LPS,ha mostrado que la expresión de Icl permanecióelevada luego de 24 horas posinfección sólo enlos macrófagos infectados. Además, lasupervivencia de la mutante icl- fuesignificativamente más reducida en macrófagosactivados, comparados con las cepas nativa ycomplementada. En el resto de los macrófagos,la supervivencia de la mutante fue ligeramentemás reducida comparada con las mismas cepas.Estos resultados sugieren que la Icl juega un papelmuy importante en la supervivencia bacterianaen el interior de los macrófagos activados (94).Las investigaciones futuras podrían indicar si esposible desarrollar inhibidores de Icl con actividadpreferencial contra micobacterias en estado depersistencia.
El ácido pantoténico se requiere para la síntesisde la coenzima A y de la proteína transportadorade acilos (PTA), los cuales juegan un papelimportante en el metabolismo de los ácidosgrasos, en el ciclo del ácido tricarboxílico y enotras reacciones, inclusive la biosíntesis depolicétidos (100). Una mutante doble de los genespanC y panD, que están involucrados en la síntesisde novo del pantotenato, fue generada en M.

177
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
tuberculosis H37Rv por recombinación homólogautilizando un sistema de transducciónespecializada. La mutante demostró ser altamenteatenuada en ratones inmunocomprometidos SCIDe inmunocompetentes BALB/c infectados por víavenosa. Además, confirió un nivel de proteccióncomparable con el de la BCG en ratones C57BL/6j inmunizados por vía subcutánea (101).
Inactivación de genes relacionados con latransducción de señales y la transcripción
Sistemas de dos componentes. Los sistemasde regulación de dos componentes son comunesen microorganismos y controlan la transducciónde señales en respuesta al medioambiente y alos estímulos internos en la bacteria. Estossistemas se caracterizan por un dominio histidino-proteín-cinasa, el cual es capaz de catalizar supropia fosforilación y funciona como el sensor dela señal y por un dominio regulador de la respuestaque es capaz de controlar la transcripción de losgenes blanco (102). Se han descrito algunossistemas de dos componentes esenciales,inclusive genes que están involucrados en lapermeabilidad de la membrana (103) y en el controldel ciclo celular (104-106). Se han identificado oncepares de sensores de histidino-cinasas ysecuencias reguladoras de respuestas en M.tuberculosis (8), incluso los genes mtrA-mtrB(107).
La inactivación del gen mtrA que codifica para unactivador transcripcional putativo, se ha descritoen la cepa H37Rv de M. tuberculosis por mediode recombinación homóloga. La esencialidad delgen mtrA se probó por el hecho de que sureemplazo solamente se obtuvo después detransformar con un plásmido que contenía elmismo gen (107). Este gen se expresó durante elcrecimiento de M. tuberculosis en medio 7H9 yen macrófagos de origen humano y murino (107).Las investigaciones futuras, inclusive el análisisdel gen mtrB y de su expresión relativa, podríanrevelar funciones específicas controladas por estesistema de dos componentes en los mecanismosde adaptación y virulencia de M. tuberculosis.
Otro sistema de dos componentes descrito en M.tuberculosis es el denominado prrA-prrB (97). Seobtuvo una mutante en el gen prrA en M.
tuberculosis Mt103 mediante mutagénesis portransposones. La mutante exhibió una velocidadde crecimiento disminuida durante los primerosdías de infección en macrófagos murinosderivados de médula ósea de ratones BALB/c. Lavelocidad de crecimiento de la mutante seequiparó con la cepa original en estadios mástardíos de la infección, lo cual sugiere un posiblepapel de este sistema en las fases tempranas decrecimiento intracelular de M. tuberculosis (108).
El sistema de dos componentes PhoP/PhoR seha asociado con el metabolismo del fosfato en M.tuberculosis (8). Este sistema es similar alsistema PhoP/PhoQ, el cual controla latranscripción de varios genes relacionados convirulencia en diversos patógenos (109). Se generóuna mutante en el gen phoP en M. tuberculosisMt103 por recombinación homóloga utilizando unplásmido con el gen phoP inactivado. Seobservaron alteraciones en la multiplicación de lamutante en macrófagos derivados de médulaósea, a pesar de que la supervivencia total de losmicroorganismos dentro de los macrófagos no sevio afectada. Esta mutante resultó atenuada enratones BALB/c infectados por vía venosa. Losresultados sugieren un posible papel para PhoPen el crecimiento intracelular de M. tuberculosis(109).
Factores sigma. Los factores sigma primarios yalternativos de ARN polimerasa confieren a estaenzima la habilidad de unirse a secuenciaspromotoras específicas e iniciar la transcripción(88). La expresión genética condicional enrespuesta a las condiciones medioambientales,incluso la expresión de genes que codifican parafactores de virulencia, se lleva a cabocomúnmente a través del reemplazo de losfactores sigma primarios por factores sigmaalternativos con cambios resultantes en laespecificidad del promotor de la ARN polimerasa(88,110-115). Ya se han identificado trecesecuencias de factores sigma putativos en M.tuberculosis, incluso los genes del factor sigmasigA y sigB, diez factores sigma con funciónextracitoplasmática y un tipo de factor sigmarelacionado con condiciones de estrés yesporulación, conocido como sigF (8).

178
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
El gen whiB3 codifica una proteína (WhiB3) queinteractúa específicamente con el factor sigmaprincipal RpoV. Una mutante de whiB3 se generóen la cepa H37Rv por recombinación homólogacon un sistema de transducción especializada.No se observaron defectos en la replicación deesta cepa en ratones C57Bl/6 o BALB/c infectadospor vía venosa, o en cobayos infectados por víasubcutánea. Sin embargo, la supervivencia fuemayor en los ratones infectados con la mutantecomparada con la de aquellos infectados con lacepa original. Estos resultados sugieren un papelpara el gen whiB3 en la virulencia de M.tuberculosis (116).
Los miembros de la familia extracitoplasmáticade factores sigma alternativos (ECF) parecenjugar un papel importante en la regulación de laexpresion genética relacionada con las respuestasal estrés oxidativo y térmico. Se ha llegado asugerir que estos factores pueden jugar un papelen la expresión de antígenos de superficie y en lasecreción de sustancias inmunomoduladorasmicobacterianas (97,117,118). En cuanto al factorSigF, se ha demostrado que el gen sigF es reguladopositivamente en la fase de crecimientoestacionario, en condiciones de depleción denitrógeno y choque frío y durante el crecimientoen macrófagos (97,119,120). La exposición amedicamentos antimicobacterianos, inclusiveetambutol, rifampicina, estreptomicina ycicloserina, también conducen a la regulaciónpositiva de este gen (120).
Una mutante sigH de M. tuberculosis H37Rv fuegenerada por recombinación homóloga medianteun sistema de transducción especializada. Comose esperaba, la mutante fue más susceptible alestrés oxidativo que la cepa original y lacomplementada (121). Igualmente, una mutantepara este gen de la cepa de M. tuberculosis H37Rvgenerada por recombinación homóloga con unvector suicida, fue más susceptible al estréstérmico y oxidativo comparado con las cepasnativa y complementada, pero no exhibióalteraciones en su crecimiento dentro demacrófagos murinos. En este caso, el análisis delos cambios en la expresión génica global despuésde la exposición a diamida permitió la identificaciónde 39 genes dependientes de sigH, incluso sigB,
sigE y el mismo sigH (117). El gen sigH fuetambién inactivado en M. tuberculosis CDC1551por recombinación homóloga con un plásmido quellevaba el gen inactivado, lo cual demuestrapersistencia en ratones C57BL/6 infectados poraerosoles. Esta mutante, además, indujo lesionesimmunopatológicas reducidas en estos ratones,al igual que en ratones CH3 infectados por víavenosa. Estos datos sugieren que, aunque el gensigH de M. tuberculosis no es indispensable parael crecimiento y la supervivencia de la bacteriaen el hospedero, sí es indispensable en lageneración de lesiones inmunopatológicascausadas por M. tuberculosis. La mortalidadreducida inducida por la mutante sigH- sugiere unpapel de este gen en la letalidad de M. tuberculosisen ratones (118).
Una mutante en el gen sigE fue generada en M.tuberculosis H37Rv por recombinación homólogacon un vector suicida que contenía el geninactivado. La mutante fue más susceptible alchoque térmico y a SDS que las cepas nativa ycomplementada. Esta mutante también exhibió uncrecimiento defectuoso en macrófagos humanosy murinos, así como una mayor sensibilidad amacrófagos murinos activados con lipopoli-sacárido e interferón gamma. El análisis de estamutante mostró que sigE está involucrado en larespuesta al estrés y, además, permitió laidentificación de varios genes, incluso sigB, queson inducidos de una forma dependiente de sigEdespués del estrés inducido por SDS (122).
El gen sigF ha sido inactivado en la cepa CDC1551de M. tuberculosis mediante recombinaciónhomóloga. La mutante obtenida alcanzó altadensidad en medios ricos durante la faseestacionaria comparada con la cepa nativa.Además, no presentó retraso en la misma faseestacionaria de crecimiento cuando se inoculó enmedio fresco según los procedimientos estándar.Las pruebas de sensibilidad de la mutante y delas cepas complementadas indicaron que lamutación en sigF resultó en una altasusceptibilidad a medicamentos tipo rifamicinas,inclusive la rifampicina (110).
Un modelo de infección in vitro indicó que lasupervivencia a corto plazo y la proliferación de

179
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
la mutante sigF- en monocitos de sangre periféricahumana fueron comparables con los que exhibióla cepa nativa. No se encontraron diferenciassignificativas entre la susceptibilidad de la mutantesigF- y la cepa nativa en la muerte mediada porlinfocitos in vitro (110). Sin embargo, los estudiosen modelos de infección en ratones BALB/cinfectados por vía intravenosa indicaron que lainactivación del gen sigF resultaba en unavirulencia reducida y los animales exhibieron unperfil de citocinas tipo Th2 después de serinfectados (110). Estos resultados sugieren queel SigF juega un importante papel en este modelode infección animal. Las futuras investigacionespodrían indicar si este efecto también es evidenteen el perfil de citocinas tipo Th1 en el modelomurino.
Inactivación de genes relacionados conresistencia a mecanismos bactericidas delos macrófagos
Uno de los aspectos claves en las infeccionescausadas por micobacterias es la interacción conlos macrófagos, las células en las que elmicroorganismo se multiplica en el hospedero. Laactividad bactericida propia de estas células escapaz de destruir más del 90% de los bacilos; sinembargo, los que logran sobrevivir son suficientespara iniciar su replicación intracelular (123). Portanto, la habilidad de las micobacterias patógenaspara sobrevivir dentro de los macrófagos es undeterminante crucial en la patogénesis de estasinfecciones.
Para sobrevivir dentro de las células fagocíticas,las micobacterias deben evitar los mecanismosbactericidas de estas células, como las enzimaslisosómicas y la actividad de los reactivosintermediarios del oxígeno y del nitrógeno(124,125). Para este fin, las micobacteriaspatógenas usan diversas estrategias. Por ejemplo,M. tuberculosis y las especies relacionadassintetizan sustancias que inhiben la acidificacióndel fagosoma (126) y la fusión del fagolisosoma(127). Otros productos génicos que puedencontribuir a la detoxificación de los reactivosintermediarios del oxígeno o del nitrógeno son lasenzimas catalasa/peroxidasa (KatG), alkil-hidroperoxidasa (AhpC) y superóxido dismutasa(SOD) (13,128,129).
La enzima KatG, codificada por el gen katG, esuna catalasa/peroxidasa que parece proteger aM. tuberculosis de la actividad bactericida de losreactivos intermediarios del oxígeno (13,129,130).El gen katG parece estar cotranscrito en unamisma región reguladora con el gen furA, quecodifica para la proteína FurA, la cual es reguladorade la captación de hierro (131). Las mutacionesen katG que codifican para una catalasa/peroxidasa incapaz de convertir la isoniacida (INH)a su forma activa, confieren resistencia a estefármaco (130,132). Se han realizado diversosestudios para definir el efecto que las mutacionesen este gen tienen en la fisiología y la patogénesisde M. tuberculosis (130,131). El análisis de variascepas isogénicas indicaron que la mutación queocurre más frecuentemente, asociada con laresistencia a INH, no afecta la virulencia de lacepa en ratones BALB/c infectados por víaintravenosa (130).
La enzima alkil hidroperoxidasa (AhpC) es capazde detoxificar peróxidos orgánicos y, probable-mente, también peróxido de hidrógeno (133-135).La mayoría de los estudios demuestra bajosniveles de AhpC en M. tuberculosis (128,136-138).Esta enzima se ha inactivado en M. tuberculosisH37Rv por recombinación homóloga utilizando unplásmido con el gen inactivado. La mutanteobtenida fue tan virulenta como la cepa originalen ratones BALB/c infectados por vía intravenosa(138). Estos resultados sugieren que este gen nojuega un papel importante en el inicio de lainfección. Sin embargo, el hallazgo de niveleselevados de expresión de ahpC en cultivosestáticos de M. tuberculosis (138) obligan a definiren el futuro el papel de esta enzima en nivelesposteriores de infección.
Dos genes en M. tuberculosis codifican lasenzimas superóxido dismutasas, sodA y sodC.Estas proteínas parecen jugar un papel importanteen la protección del microorganismo contra laacción de los intermediarios reactivos del oxígeno.El análisis de la mutante secA2- de H37Rvgenerada por recombinación homóloga usando unvector suicida contraselectivo, demostró que SodAes una proteína dependiente de la proteína SecA2para su secreción (139).

180
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
Con respecto a SodC, una mutante sodC- de M.tuberculosis cepa Erdman, obtenida porrecombinación homóloga utilizando un fragmentolineal con el gen sodC inactivado, presentó un in-cremento significativo en la sensibilidad asuperóxido de origen externo. Esta mutante fuetambién más susceptible a los mecanismosbactericidas de macrófagos peritoneales murinosactivados con interferón gamma (INF-γ) ygeneradores de productos de la explosionrespiratoria (13).
Existe también una gran cantidad de genesmicobacterianos con función controladora(housekeeping genes) cuya expresión esnecesaria para que estos microorganismossobrevivan a la acción bactericida de las célulasfagocíticas. Por ejemplo, las mutantes de M.tuberculosis auxotróficas para leucina (69),micobactina (91) o purinas (45) poseen fenotiposatenuados caracterizados por supervivenciadisminuida en macrófagos. Los estudiosrelacionados con algunos de estos genes se handescrito en párrafos anteriores.
En el cuadro 1 que aparece a continuación, sepresenta la bibliografía relacionada con lainactivación de genes de M. tuberculosis con elfin de facilitarle a los lectores la consulta de lamisma.
Consideraciones finales
Desde la secuenciación por primera vez delgenoma de M. tuberculosis H37Rv en 1998, haaparecido una gran cantidad de informaciónrelacionada con las funciones adscritas a losgenes del microorganismo como lo confirma laactual revisión. Esto todavía es una porción muypequeña del total del genoma compuesto por4'411.529 pares de bases que conformanaproximadamente 4.000 genes (8). Aún así,solamente, aproximadamente, al 52% de losgenes del genoma de H37Rv se les ha adscritofunciones precisas o putativas de acuerdo con lahomología con genes de otros microorganismos,mientras que el 48% restante son geneshipotéticos conservados o genes con funcióndesconocida (140).
Las futuras investigaciones probablementecontinuarán enfocándose en la inactivación de
genes para identificar blancos útiles en el diseñode vacunas, agentes terapéuticos y pruebasdiagnósticas. Los estudios comparativos deexpresión génica en diferentes cepas, estadios ymodelos de infección pueden jugar un papelimportante en la identificación de dichos genes.
Especial énfasis se le ha dado a la atenuaciónracional de cepas con el fin de generar candidataspara vacunas. A este respecto, sería de utilidadpara futuras investigaciones recordar que dichascepas deberán poseer inactivaciones en dosgenes, por lo menos, idealmente involucrados endiferentes vías metabólicas del bacilo. Estascepas deben exhibir un grado adecuado deatenuación, de tal modo que sean lo suficiente-mente inmunogénicas y así generen unarespuesta inmunológica protectora que sea útil enel caso eventual de una infección. Idealmente,también la inactivación de genes deberá generarcepas que sean compatibles con pruebasdiagnósticas y que permitan una rápida y eficazdiferenciación entre individuos vacunados einfectados. Además, las cepas atenuadas deberánser productos finales carentes de marcadores,principalmente marcadores que confieranresistencia a agentes quimioterapéuticos, paraevitar la transferencia de estos genes a losaislamientos clínicos (141).
Estudios adicionales podrían eventualmenteemplear la inactivación de genes para identificarposibles candidatos con potencial en otros tiposde vacunas, incluidas vacunas de subunidadesnaturales o sintéticas o vacunas de ADN, o paraidentificar blancos potenciales para pruebasdiagnósticas y nuevos agentes quimio-terapéuticos. Finalmente, vale la pena anotar queaunado al conocimiento del genoma mico-bacteriano, la reciente secuenciación del genomahumano abre aun nuevas puertas para elentendimiento de la patogenicidad en tuberculosis.Las futuras investigaciones basadas en el estudiogenético de las interacciones hospedero-bacteria,a nivel individual y de población, indudablementejugarán un papel primordial en el desentrañamientode los misterios que encierra dicha interacción yproporcionarán nuevos conocimientos útiles parael diagnóstico, la prevención y el tratamiento dela tuberculosis.

181
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
Cuadro 1. Inactivación de genes de Mycobacterium tuberculosis y su relación con funciones estructurales y metabólicas.
Función general Función específica Genes inactivados Referencias
Pared celular y Síntesis y transporte de dimicocerosatos ppsA-E 16proteínas asociadas fadD28 15, 16
drrC 15mmpL7 16, 40fadD26 15, 40msl6 (pks12) 42msl5 (pks8, pks17) 43
Síntesis o transferencia de ácidos pca (umaA2) 30micólicos y trehalosas cmaA2 28
mmaA2 44hma (cmaA, mmaA) 29fbpA 33fbpB 33fbpC2 34msl3 (pks3, pks4) 51
Síntesis de sulfolípidos pks2 (msl2) 58mmpl8 59
Proteínas asociadas erp (pirG) 20, 62
Biosíntesis, metabolismo y Biosíntesis de aminoácidos lysA 68, 79transporte de metabolitos y leuD 69nutrientes metB 70, 80
proC 70, 80hisD 70trpD 70, 80argF 70, 71glnA1 72
Captación de hierro ideR 89
y regulación dependiente de hierro mbtB 91mramp 92
Transporte de magnesio mgtC 11
Metabolismo de ácidos grasos panC 101panD 101icl 94
Transducción de señales Sistemas de dos componentes mtrA 107y transcripción prrA 108
phoP 109
Factores sigma sigF 110whiB3 116sigH 117, 118, 121sigE 122
Resistencia a mecanismos Enzimas detoxificadoras de reactivos katG 130, 131, 132bactericidas de los macrófagos intermediarios del oxígeno o nitrógeno sodA 139
sodC 13ahpC 138
Agradecimientos
El presente manuscrito fue realizado gracias a unconvenio marco Universidad de Nebraska-CIB yal proyecto cofinanciado por Colciencias código2213-04-11899.
Referencias
1. Koch RD. Aetiologie der Tuberculos. Ber KlinWochenschr 1882;19:221. Reprinted as a translationby Pinner B, Pinner M. Am Rev Tuberc 1932;25:285-323.

182
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
2. Daniel TM, Bates JH, Downes KA. History oftuberculosis. En: Bloom B, editor. Tuberculosis:pathogenesis, protection and control. Washington D.C.:American Society for Microbiology Press; 1994. p.13-24.
3. Bloom BR, Murray CJL. Tuberculosis: commentaryon a reemergent killer. Science 1992;257:1055-64.
4. Dye CS, Scheele S, Dolin P, Pathania V, RaviglioneMC. Global burden of tuberculosis: estimated incidence,prevalence, and mor tality by country. JAMA1999;282:677-86.
5. Jacobs WR Jr., Tuckman M, Bloom BR. Introductionof foreign DNA into mycobacteria using a shuttlephasmid. Nature 1987;327:532-5.
6. Bardarov S, Kriakov J, Carriere C, Yu S, VagabondedC, Macadam RA, et al. Conditionally replicatingmycobacteriophages: a system for transposondelivery to Mycobacterium tuberculosis. Proc Natl AcadSci USA 1997;94:10961-6.
7. Pelicic V, Jackson M, Reyrat JM, Jacobs WR Jr.,Gicqel B, Guilhot C. Efficient allelic exchange andtransposon mutagenesis in Mycobacteriumtuberculosis. Proc Natl Acad Sci USA 1997;94:10955-60.
8. Cole ST, Brosch R, Parkhill J, et al. Descipheringthe biology of Mycobacterium tuberculosis from thecomplete genome sequence. Nature 1998;393:537-44.
9. Konickova-Radochova M, Konicek J, Malek L. Thestudy of mutagenesis in Mycobacterium phlei. FoliaMicrobiol 1970;15:88-102.
10. Balasubramanian V, Pavelka MS Jr, Bardarov SS,Martin J, Weisbrod TR, McAdam RA, et al. Allelicexchange in Mycobacterium tuberculosis with longlinear recombination substrates. J Bacteriol1996;178:273-9.
11. Buchmeier N, Blanc-Putard A, Ehrt S, PiddingtonD, Riley L, Goisman EA. A parallel intraphagosomalsurvival strategy shared by Mycobacteriumtuberculosis and Salmonella enterica. Mol Microbiol2000;35:1375-82.
12. Chacon O, Feng Z, Harris NB, Caceres NE, AdamsLG, Barletta RG. Mycobacterium smegmatis D-alanine racemase mutants are not dependent on D-alanine for growth. Antimicrob Agents Chemother2002;46:47-54.
13. Piddington DL, Fang FC, Laessig T, Cooper AM,Orme IM, Buchmeier N. Cu, Zn superoxide dismutaseof Mycobacterium tuberculosis contributes to survivalin activated macrophages that are generating anoxidative burst. Infect Immun 2001;69:4980-7.
14. Pelicic V, Reyrat JM, Gicquel B. Expression of theBacillus subtilis sacB gene confers sucrose sensitivityon mycobacteria. J Bacteriol 1996;178:1197-9.
15. Camacho LR, Ensergueix D, Perez E, Gicquel B,Guilhot C. Identification of a virulence gene cluster ofMycobacterium tuberculosis by signature-taggedtransposon mutagenesis. Mol Microbiol 1999;34:257-67.
16. Cox JS, Chen B, McNell M, Jacobs WR Jr. Complexlipid determines tissue-specific replication ofMycobacterium tuberculosis in mice. Nature1999;402:79-83.
17. Sassetti CM, Boyd DH, Rubin EJ. Genes requiredfor mycobacterial growth defined by high densitymutagenesis. Mol Microbiol 2003;48:77-84.
18. Trias J, Benz R. Permeability of the cell wall ofMycobacterium smegmatis. Mol Microbiol1994;14:283-90.
19. Belisle JT, Vissa VD, Slevert T, Takayama K, BrennanPJ, Besra GS. Role of the major antigen ofMycobacterium tuberculosis in cell wall biogenesis.Science 1997;276:1420-2.
20. Berthet FX, Rasmussen PB, Rosenkrands I,Andersen P, Gicquel B. Attenuation of virulence bydisruption of the Mycobacterium tuberculosis erp gene.Science 1998;282:759-62.
21. McNeil M, Daffe M, Brennan PJ. Location of themycolyl ester substituents in the cell walls ofmycobacteria. J Biol Chem 1991;266:13217-23.
22. Daffe M, Draper P. The envelope layers ofmycobacteria with reference to their pathogenicity.Advances in Microbial Physiology 1998;39:131-203.
23. Brennan PJ, Nikaido H. The envelope of mycobateria.Annu Rev Biochem 1995;64:29-63.
24. Lederer E. Cell walls of mycobacteria and relatedorganisms; chemistry and immunostimulant proper-ties. Molecular and Cellular Biochemistry 1975;7:87-104.
25. Yuan Y, Barry CE 3rd. A common mechanism for thebiosynthesis of methoxy and cyclopropyl mycolic acidsin Mycobacterium tuberculosis. Proc Natl Acad SciUSA 1996;93:12828-33.
26. Yuan Y, Lee RE, Besra GS, Belisle JT, Barry CE 3rd.Identification of a gene involved in the biosynthesis ofcyclopropanated mycolic acids in Mycobacterium tu-berculosis. Proc Natl Acad Sci USA 1995;92:6630-4.
27. George KM, Yuan Y, Sherman DR, Barry CE 3rd.The biosynthesis of cyclopropanated mycolic acids inMycobacterium tuberculosis. Identification andfunctional analysis of CMAS-2. J Biol Chem1995;270:27292-8.
28. Glickman MS, Cahill SM, Jacobs WR Jr. TheMycobacterium tuberculosis cmaA2 gene encodes amycolic acid trans-cyclopropane synthetase. J BiolChem 2001;276:2228-33.
29. Dubnau E, Chan J, Raynaud C, Mohan VP, LanéelleMA, Yu K, et al. Oxygenated mycolic acids are

183
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
necessary for virulence of Mycobacteriumtuberculosis in mice. Molec Micro 2000;36:630-7.
30. Glickman MS, Cox JS, Jacobs WR Jr. A novel mycolicacid cyclopropane synthetase is required for cording,persistence, and virulence of Mycobacteriumtuberculosis. Molecular Cell 2000;5:717-27.
31. Dubnau E, Laneelle MA, Soares S, Benichou A,Vaz T, Prome D, et al. Mycobacterium bovis BCGgenes involved in the biosynthesis of cyclopropyl keto-and hydroxy-mycolic acids. Mol Microbiol 1997;23:313-22.
32. Liu J, Barry 3rd CE, Besra GS, Nikaido H. Mycolicacid structure determines the fluidity of themycobacterium cell wall. J Biol Chem 1996;271:29545-51.
33. Armitige LY, Jagannath C, Wanger AR, Norris SJ.Disruption of the genes encoding antigen 85A andantigen 85B of Mycobacterium tuberculosis H37Rv:effect on growth in culture and in macrophages. InfectImmun 2000;68:767-78.
34. Jackson M, Raynaud C, Lanéelle MA, Guilhot C,Laurent-Winter C, Ensergueix D, et al. Inactivationof the antigen 85C gene profoundly affects themycolate content and alters the permeability of theMycobacterium tuberculosis cell envelope. Molec Micro1999;31:1573-87.
35. Goren MB, Broki O, Schaefer WB. Lipids of putativerelevance to virulence in Mycobacterium tuberculosis:phthiocerol dimycocerosate and the attenuationindicator lipid. Infect Immun 1974;9:150-8.
36. Azad AK, Sirakova TD, Fernandez ND, KolattukudyPE. Gene knockout reveal a novel gene cluster for thesynthesis of a class of cell wall lipids unique topathogenic mycobacteria. J Biol Chem1997;272:16741-5.
37. Azad AK, Sirakova TD, Rogers LM, KolattudukiPE. Targeted replacement of the mycocerosic acidsynthase gene in Mycobacterium bovis BCG producesa mutant that lacks mycosides. Proc Natl Acad SciUSA 1996;93:4787-92.
38. Mathur M, Kolattuduki PE. Molecular cloning andsequencing of the gene for mycocerosic acid synthase,a novel fatty acid elongating multifunctional enzyme,from Mycobacterium tuberculosis var. bovis bacillusCalmette-Guerin. J Biol Chem 1992;267:19388-95.
39. Fitzmaurice AM, Kolattukudy PE. An acyl-CoA syn-thase (acoas) gene adjacent to the mycocerosic acidsynthase (mas) locus is necessary for mycocerosyllipid synthesis in Mycobacterium tuberculosis var. bovisBCG. J Biol Chem 1998;273:8033-9.
40. Camacho LR, Constant P, Raynaud C, LaneélleMA, Triccas JA, Guicquel B, et al. Analysis of thephthiocerol dimycocerosate locus of Mycobacteriumtuberculosis: evidence that this lipid is involved in cell
wall permeability barrier. J Biol Chem 2001; 276:19845-54.
41. Fitzmaurice AM, Kolattukudy PE. Open readingframe 3, which is adjacent to the mycocerosic acidsynthase gene, is expressed as an acyl coenzyme Asynthase in Mycobacterium bovis BCG. J Bacteriol1997;179:2608-15.
42. Sirakova T, Dubey VS, Kim HJ, Synamon MH,Kolattukudy PE. The largest open reading frame(pks12) in the Mycobacterium tuberculosis genome isenvolved in pathogenesis and dimicocerosil phthiocerolsynthesis. Infect Immun 2003;71:3794-801.
43. Dubey VS, Sirakova TD, Cynamon MH, KolattukudyPE. Biochemical function of msl5 (pks8 plus pks17) inMycobacterium tuberculosis H37Rv: biosynthesis ofmonomethyl branched unsaturated fatty acids. InfecImmun 2003;185:4620-5.
44. Glickman MS. The mmaA2 gen of Mycobacteriumtuberculosis encodes the distal cyclopropane synthaseof the alpha-mycolic acid. J Biol Chem 2003;278:7844-9.
45. Jackson M, Phalen SW, Lagranderie M, EnsergueixD, Chavarot P, Marchal G, et al. Persistence andprotective efficacy of a Mycobacterium tuberculosisauxotroph vaccine. Infect Immun 1999;67:2867-73.
46. Kremer L, Baulard AR, Besra GS. Genetics of mycolicacid biosynthesis. En: Hatfull GF, Jacobs WR Jr, editors.Molecular genetics of Mycobacteria. Washington, D.C.:American Society for Microbiology Press; 2000. p.173-90.
47. Patti JM, Allen BL, McGavin MJ, Hook M.MSCRAMM-mediated adherence of microorganismsto host tissues. Ann Rev Microbiol 1994;48:585-617.
48. Peake P, Gooley A, Britton WJ. Mechanism ofinteraction of the 85B secreted protein ofMycobacterium bovis with fibronectin. Infec Immun1993;61:4828-34.
49. Espitia C, Laclette JP, Mondragon-Palomino M,Amador A, Campuzano J, Martens A, et al. ThePE-PGRS glycine-rich proteins of Mycobacteriumtuberculosis: a new family of fibronectin-bindingproteins? Microbiology 1999;145:3487-95.
50. Huygen K, Content J, Denis O, Montgomery L,Yauman AM, Deck RR, et al. Immunogenicity andprotective efficacy of a tuberculosis DNA vaccine. NatMed 1996;2:893-8.
51. Dubey VS, Sirakova TD, Kolattukudy PE. Disruptionof msl3 abolishes the synthesis of mycolipanoic andmycolipenic acids required for polyacyltrehalosesynthesis in Mycobacterium tuberculosis H37Rv andcauses cell aggregation. Mol Microbiol 2002;45:1451-9.
52. Goren M, Brokl BO, Das BC. Sulfatides ofMycobacterium tuberculosis: the structure of the

184
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
principal sulfatide (SL-I). Biochemistry 1976;15:2728-35.
53. Alugupalli S, Lanelle MA, Larson L, Daffe M.Chemical characterization of the ester-linked 3-hidroxyfatty acyl-containing lipids in Mycobacteriumtuberculosis. J Bacteriol 1995;177:4566-70.
54. Kolattukudy PE, Fernandez ND, Azad AK,Fitzmaurice AM, Sirakova TD. Biochemistry andmolecular genetics of cell-wall lipid biosynthesis inmycobacteria. Mol Microbiol 1997;24:263-70.
55. Pabst MJ, Gross JM, Brozna JP, Goren MB. Inhibitionof macrophage priming by sulfatide fromMycobacterium tuberculosis. J Immunol1988;140:634-40.
56. Brozna JP, Horan M, Rademacher JM, Pabst KM,Pabst MJ. Monocyte responses to sulfatide fromMycobacterium tuberculosis: inhibition of priming forenhanced release of superoxide, associated withincreased secretion of interleukin-1 and tumornecrosis factor alpha, and altered proteinphosphorylation. Infect Immun 1991;59:2542-8.
57. Zhang L, Goren MB, Holzer TJ, Andersen BR. Effectof Mycobacterium tuberculosis derived sulfolipid I onhuman phagocytic cells. Infect Immun 1988;56:2876-83.
58. Sirakova TD, Thirumala AK, Dubey VS, Sprecher H,Kolattukudy PE. The Mycobacterium tuberculosispks2 gene encodes the synthase for the hepta- andoctamethyl branched fatty acids required for sulfolipidsynthesis. J Biol Chem 2001;276:16833-9.
59. Converse SE, Mougous JD, Leavell MD, Leary JA,Bertozzi CR, Cosx JS. Mmpl8 is required forsulpholipid-1 biosynthesis and Mycobacteriumtuberculosis virulence. PNAS 2003;100:6121-6.
60. Braunstein M, Belisle JT. Genetics of proteinssecretion. En: Hatfull GF, Jacobs WR Jr., editors.Molecular genetics of Mycobacteria. Washington, D.C.:American Society for Microbiology Press; 2000. p.203-20.
61. Abou-Zeid C, Ratliff TL, Wiker HG, Harboe M,Bennedsen J, Rook GA. Characterization offibronectin-binding antigens released byMycobacterium tuberculosis and Mycobacterium bovisBCG. Infect Immun 1988;56:3046-51.
62. de Mendonca-Lima L, Bordat Y, Pivert E, Recchi C,Neyrolles O, Maitournam A, et al. The alleleencoding the mycobacterial Erp protein affects lungdisease in mice. Cell Microbiol 2003;5:65-73.
63. McAdam RA, Weisbrod TR, Martin J, Scuderi JD,Brown AM, Cirillo JD, Bloom BR, Jacobs WR. Invivo growth characteristics of leucine and methionineauxotrophic mutants of Mycobacterium bovis BCGgenerated by transposon mutagenesis. Infect Immun1995;63:1004-12.
64. Bange FC, Brown AM, Jacobs WR Jr. Leucineauxotrophy restricts growth of Mycobacterium bovisBCG in macrophages. Infect Immun 1996;64:1794-9.
65. Guleria I, Teitelbaum R, McAdam RA, Kalpana G,Jacobs WR Jr, Bloom BR. Auxotrophic vaccines fortuberculosis. Nat Med 1996;2:334-7.
66. Ratledge C. Nutrition, growth and metabolism. En:Ratledge C, Stanford J, editors. The biology ofmycobacteria. London: United Kingdom AcademicPress; 1982. p.185-271.
67. Pavelka MS Jr. Genetics of Mycobacteriummetabolism. En: Hatful GF, Jacobs WR Jr., editors.Molecular genetics of mycobacteria. Washington,D.C.: American Society for Microbiology Press; 2000.p.221-34.
68. Pavelka MS Jr, Jacobs WR Jr. Comparison of theconstruction of unmarked deletion mutations inMycobacterium smegmatis, Mycobacterium bovisbacillus Calmette-Guérin and Mycobacteriumtuberculosis H37Rv by allelic exchange. J Bacteriol1999;181:4780-9.
69. Hondalus MK, Bardarov S, Russell R, Chan J,Jacobs WR Jr, Bloom BR. Attenuation and protectioninduced by a leucine auxotroph of Mycobacteriumtuberculosis. Infect Immun 2000;68:2888-98.
70. Parish T, Gordhan BG, McAdam RA, Duncan K,Mizrahi V, Stoker NG. Production of mutants in aminoacid biosynthetic genes of Mycobacterium tuberculosisby homologous recombination. Microbiology1999;145:3497-503.
71. Gordhan BG, Smith DA, Alderton H, McAdam RA,Bancroft GJ, Mizrahi V. Construction andcharacterization of an auxotrophic mutant ofMycobacterium tuberculosis defective in l-argininebiosyntesis. Infect Immun 2002;70:3080-4.
72. Tullius M, Harth G, Horwitz MA. Glutamine synthaseGlnA1 is essential for growth of Mycobacterium tu-berculosis in human THP-1 macrophages and Guineapigs. Infec Immun 2003;71:3927-36.
73. Patte JC. Biosynthesis of threonine and lysine. En:Neidhardt FC, editor. Escherichia coli and Salmonellacellular and molecular biology. Washington, D.C.:American Society for Microbiology Press; 1996. p.528-41.
74. Umbarger HE. Biosynthesis of the branched chainaminoacids. En: Neidhardt FC, editor. Escherichia coliand Salmonella cellular and molecular biology.Washington, D.C.: American Society for MicrobiologyPress; 1996. p.442-57.
75. Greene RC. Biosynthesis of methionine. En: NeidhardtFC, editor. Escherichia coli and Salmonella cellularand molecular biology. Washington, D.C.: AmericanSociety for Microbiology Press; 1996. p.542-60.

185
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
76. Winkler ME. Biosynthesis of histidine. En: NeidhardtFC, editor. Escherichia coli and Salmonella cellularand molecular biology. Washington, D.C.: AmericanSociety for Microbiology Press; 1996. p.485-505.
77. Leisinger T. Biosynthesis of proline. En: Neidhardt FC,editor. Escherichia coli and Salmonella cellular andmolecular biology. Washington, D.C.: American Societyfor Microbiology Press; 1996. p.434-41.
78. Pittard AJ. Biosynthesis of the aromatic aminoacids.En: Neidhardt FC, editor. Escherichia coli andSalmonella cellular and molecular biology. Washington,D.C.: American Society for Microbiology Press; 1996.p.458-84.
79. Pavelka MS Jr, Chen B, Kelley CL, Collins FM,Jacobs WR Jr. Vaccine efficacy of a lysine auxotrophof Mycobacterium tuberculosis. Infect Immun2003;71:4190-2.
80. Smith DA, Parish T, Stoker NG, Bancroft GJ.Characterization of auxotrophic mutants ofMycobacterium tuberculosis and their potential asvaccine candidates. Infect Immun 2001;69:1142-50.
81. Litwin CM, Calderwood SB. Role of iron in regulationof virulence genes. Clin Microb Rev 1993;6:137-49.
82. Manabe YC, Saviola BJ, Sun L, Murphy JR, BishaiWR. Attenuation of virulence in Mycobacteriumtuberculosis expressing a constitutively active ironrepressor. Proc Natl Acad Sci USA 1999;96:12844-8.
83. Gobin J, Hortwitz MA. Exochelins of Mycobateriumtuberculosis remove iron from human iron-bindingproteins and donate iron to mycobactins in the M.tuberculosis cell wall. J Exp Med 1996;183:1527-32.
84. Golden CA, Kochan I, Spriggs DR. Role ofmycobactin in the growth and virulence of tuberclebacilli. Infect Immun 1974;9:34-40.
85. Quadri LE, Sello J, Keating TA, Weinreb PH, WalshCT. Identification of a Mycobacterium tuberculosis genecluster encoding the biosynthetic enzymes for as-sembly of the virulence-conferring siderophoremycobactin. Chem Biol 1998;5:631-45.
86. Earhart CF. Uptake and metabolism of iron andmolybdenum. En: Neidhardt FC, editor. Escherichiacoli and Salmonella cellular and molecular biology.Washington, D.C.: American Society for MicrobiologyPress; 1996. p.1075-90.
87. Tao X, Schiering N, Zeng HY, Ringe D, Murphy JR.Iron, DtxR, and the regulation of diphteria toxinexpression. Mol Microbiol 1994;14:191-7.
88. Gómez M, Smith I. Determinants of mycobacterialgene expression. En: Hatfull GF, Jacobs WR Jr, editors.Molecular genetics of Mycobacteria. Washington, D.C.:American Society for Microbiology Press; 2000. p.111-29.
89. Rodríguez GM, Voskuil MI, Gold B, Schollnik GK,Smith I. IdeR, an essential gene in Mycobacterium
tuberculosis: role of IdeR in iron-dependent geneexpression, iron metabolism, and oxidative stressresponse. Infec Immun 2002;70:3371-81.
90. De Voss JJ, Rutter K, Schroeder BG, Barry CE III.Iron acquisition and metabolism in mycobacteria. JBacteriol 1999;181:4443-51.
91. De Voss JJ, Rutter K, Schroeder BG, Su H, Zhu Y,Barry CE 3rd. The salicylate-derived mycobactinsiderophores of Mycobacterium tuberculosis areessential for growth in macrophages. Proc Natl AcadSci USA 2000;97:1252-7.
92. Boechat N, Lagier-Roger B, Petit S, Bordat Y,Rauzier J, Hance AJ et al. Disruption of the genehomologous to mammalian Nramp1 in Mycobacteriumtuberculosis does not affect virulence in mice. InfecImmun 2002;70:4124-31.
93. Blanc-Potard AB, Groisman EA. The Salmonella selClocus contains a pathogenicity island mediating intramacrophage survival. EMBO J 1997;16:5376-85.
94. McKinney JD, Zu Bentrup KH, Muñoz-Elias EJ,Miczak A, Chen B, Chan W-T, et al. Persistence ofMycobacterium tuberculosis in macrophages andmice requires the glyoxylate shunt enzyme isocitratelyase. Nature 2000;406:735-8.
95. Nunn WD. Two-carbon compounds and fatty acidsas carbon sources. En: Neidhar t FC, editor.Escherichia coli and Salmonella: cellular and molecularbiology. Washington, D.C.: American Society forMicrobiology Press; 1987. p.285-301.
96. Zu Bentrup KH, Miczak A, Swenson DL, RussellDG. Characterization of activity and expression ofisocitrate lyase in Mycobacterium avium andMycobacterium tuberculosis. J Bacteriol 1999;181:7161-7.
97. Graham JE, Clark Curtiss JE. Identification of Myco-bacterium tuberculosis RNAs synthesized in responseto phagocytosis by human macrophages by selectivecapture of transcribed sequences (SCOTS). Proc NatlAcad Sci USA 1999;96:11554-9.
98. Wayne LG, Lin KY. Glyoxilate metabolism and adap-tation of Mycobacterium tuberculosis to survival un-der anaerobic conditions. Infect Immun 1982;37:1042-9.
99. Sturgill-Koszycki S, Haddix PL, Russell DG. Theinteraction of the Mycobacterium and the macrophageanalyzed by two-dimensional polyacrylamide gelelectrophoresis. Electrophoresis 1997;18:2558-65.
100. Jackowski S. Biosynthesis of pantothenic acids andcoenzyme A. En: Neidhart FC, editor. Escherichia coliand Salmonella typhimurium: cellular and molecularbiology. Washington, D.C.: American Society forMicrobiology Press; 1996. p.687-94.
101. Sambandamurthy VK, Wang X, Chen B, RussellRG, Derrick S, Collins FM, et al. A pantothenate

186
Biomédica 2004;24(Supl.):165-87CHACÓN O., BARLETTA R.G., REALPE T., ROBLEDO J.
auxotroph of Mycobacterium tuberculosis is highlyattenuated and protects mice against tuberculosis.Nat Med 2002;8:1171-4.
102. Ninfa AJ. Regulation of gene transcription byextracellular stimuli. En: Neidhardt FC, editor.Escherichia coli and Salmonella cellular and molecularbiology. Washington, D.C.: American Society forMicrobiology Press; 1996. p.1246-62.
103. Martin PK, Li T, Sun D, Biek DP, Schmid M. Role incell permeability of an essential two-componentsystem in Staphylococcus aureus. J Bacteriol1999;181:3666-73.
104. Quon KC, Marczynski GT, Saphiro L. Cell cyclecontrol by an essential bacterial two-component signaltransduction protein. Cell 1996;84:83-93.
105. Wu J, Ohta N, Newton A. An essential multi-component signal transduction pathway required forcell cycle regulation in Caulobacter. Proc Natl AcadSci USA 1998;95:1443-8.
106. Jacobs C, Domian IJ, Madock JR, Shapiro L. Cellcyle-dependent polar localization of an essentialbacterial histidine kinase that controls DNA replicationand cell division. Cell 1999;97:111-120.
107. Zahrt TC, Deretic V. An essential two-componentsignal transduction system in Mycobacteriumtuberculosis. J Bacteriol 2000;182:3832-8.
108. Graham JE, Clark-Curtiss JE. Identification ofMycobacterium tuberculosis RNAs synthesized inresponse to phagocytosis by human macrophagesby selective capture of transcribed sequences(SCOTS). Proc Natl Acad Sci USA 1999;96:11554-9.
108. Ewann F, Jackson M, Pethe K, Cooper A, MielcarekN, Ensergueix D, Gicquel B, Locht C, Supply P.Transient requirement of the PrrA-PrrB two componentsystem for early intracellular multiplication of Myco-bacterium tuberculosis. Infec Immun 2002;70:2256-63.
109. Pérez E, Samper S, Bordas Y, Guilhot C, GicquelB & Martín C. An essential role for phoP inMycobacterium tuberculosis virulence. Mol Micro2001;41:179-87.
110. Chen P, Ruiz RE, Li Q, Silver RF, Bishai WR.Construction and characterization of a Mycobacte-rium tuberculosis mutant lacking the alternate sigmafactor gene, sigF. Infect Immun 2000;68:5575-80.
111. Deretic V, Schurr MJ, Boucher JC, Martin DW.Conversion of Pseudomonas aeruginosa to mucoidyin cystic fibrosis: environmental stress and regulationof bacterial virulence by alternative sigma factors. JBacteriol 1994;176:2773-80.
112. Fang FC, Libbly SJ, Buchmeier NA, Loewen PC,Switala J, Harwood J, Guiney DG. The alternatives factor KatF (RpoS) regulates Salmonella virulence.Proc Natl Acad Sci USA 1992;89:11978-82.
113. Collins D, Kawakami R, de Lisle G, Pascopella L,Bloom B, Jacobs WR Jr. Mutation of the principal sfactor causes loss of virulence in a strain of theMycobacterium tuberculosis complex. Proc Natl AcadSci USA 1995;92:8036-40.
114. Finlay BB, Falkow S. Common themes in microbialpathogenicity revisited. Microbiol Mol Biol Rev1997;61:136-69.
115. Miller JF, Mekalanos JJ, Falkow S. Coordinateregulation and sensory transduction in the control ofbacterial virulence. Science 1989;243:916-22.
116. Steyn AJC, Collins DM, Hondalus MK, Jacobs WRJr, Kawakami RP, Bloom BR. Mycobacteriumtuberculosis WhiB3 interacts with RpoV to affect hostsurvival but is dispensable for in vivo growth. ProcNatl Acad Sci 2002;99:3147-52.
117. Manganelli R, Voskuil MI, Schoolnik GK, DubnauE, Gomez M, Smith I. Role of the extracytoplasmic-function sigma factor sigma(H) in Mycobacteriumtuberculosis global gene expression. Mol Microbiol2002;45:365-74.
118. Kaushal D, Schroeder BG, Tyagi S, Yoshimatsu T,Scott C, Ko C, et al. Reduced immunopathology andmortality despite tissue persistence in aMycobacterium tuberculosis mutant lacking alternativefactor, SigH. Proc Natl Acad Sci 2002;99:8330-5.
119. DeMaio J, Zhang Y, Ko C, Young D, Bishai WR. Astationary phase stress-response sigma factor fromMycobacterium tuberculosis. Proc Natl Acad Sci USA1996;93:2790-4.
120. Michele T, Ko C, Bishai WR. Antibiotic exposureinduces expression of the Mycobacteriumtuberculosis sigF gene: implications for chemotherapyagainst mycobacterium persistors. Antimicrob AgentsChemother 1999;43:218-25.
121. Raman S, Song T, Puyang X, Bardarov S, JacobsWR Jr, Husson RN. The alternative sigma factor SigHregulates major components of oxidative and heatstress responses in Mycobacterium tuberculosis. JBacteriol 2001;183:6119-25.
122. Manganelli R, Voskuil MI, Schoolnik GK, Smith I.The Mycobacterium tuberculosis ECF sigma factorsigmaE: role in global gene expression and survival inmacrophages. Mol Microbiol 2001;41:423-37.
123. Fenton MJ, Vermeulen MW. Immunopathology oftuberculosis: roles of macrophages and monocytes.Infect Immun 1996;64:683-90.
124. Lowrie DB, Andrwe PW. Macrophage anti-micobacteriamechanisms. BR Med Bull 1988;44:624-34.
125. Nathan C, Shiloh MU. Reactive oxygen and nitrogenintermediates in the relationship between mammalianhost and microbial pathogens. Proc Natl Acad SciUSA 2000;97:8841-8.

187
Biomédica 2004;24(Supl.):165-87 INACTIVACIÓN DE GENES DE MYCOBACTERIUM TUBERCULOSIS
126. Sturgill-Koszycki S, Schlesinger PH, ChakrabortyP, Haddix PL, Collins HL, Fohk AK, et al. Lack ofacidification in Mycobacterium phagosomes producedby exclution of the vesicular proton-ATPase. Science1994;263:678-81.
127. Armstrong JA, Hart PD. Phagosome-lysosomeinteractions in cultured macrophage infected withvirulent tubercle bacilli. Reversal of the usual nonfusionpattern and observation on bacterial survival. J ExpMed 1975;142:1-16.
128. Chen L, Xie QW, Nathan C. Alkyl hydroperoxidereductase subunit C (AhpC) protects bacterial andhuman cells against reactive nitrogen intermediates.Mol Cell 1998;1:795-805.
129. Manca C, Paul S, Barry CE 3rd, Freedman VH,Kaplan G. Mycobacterium tuberculosis catalase andperoxidase activities and resistance to oxidative killingin human monocytes in vitro. Infect Immun 1999;67:74-9.
130. Pym AS, Saint-Joanis B, Cole ST. Effect of katGmutations on the virulence of Mycobacteriumtuberculosis and the implication for transmission inhumans. Infect Immun 2002;70:4955-60.
131. Pym AS, Domenech P , Honore N , Song J, DereticV, Cole ST. Regulation of catalase-peroxidase (KatG)expression, isoniazid sensitivity and virulence by furAof Mycobacterium tuberculosis. Mol Microbiol2001;40:879-89.
132. Johnsson K, Froland WA, Schultz PG. Over-ex-pression, purification, and characterization of thecatalase-peroxidase KatG from Mycobacteriumtuberculosis. J Biol Chem 1997;272:2834-40.
133. Jacobson FS, Morgan RW, Christman MF, AmesBN. An alkyl hydroperoxide reductase from Salmonellatyphimurium involved in the defense of DNA againstoxidative damage. Purification and properties. J BiolChem 1989;264:1488-96.
134. Sherman DR, Mdluli K, Hicky MJ, Arain TN, MorrisSL, Barry CE 3rd, et al. Compensatory ahpCgeneexpression in isoniazid resistance Mycobacteriumtuberculosis. Science 1996;272:1641-3.
135. Niimura Y, Poole LB, Massey V. Amphibacillusxylanus NADH oxidase and Salmonella typhimuriumalkyl-hydroperoxide reductase flavoproteincomponents show extremely high scavenging activityfor both alkyl hydroperoxide and hydrogen peroxidein the presence of S. typhimurium alkyl-hydroperoxidereductase 22-kDa protein component. J Biol Chem1995;270:25645-50.
136. Dhandayuthapani S, Mudd M, Deretic V.Interactions of OxyR with the promoter region of theoxyR and ahpC genes from Mycobacterium lepraeand Mycobacterium tuberculosis. J Bacteriol1997;179:2401-9.
137. Zhang Y, Dhandayuthapani S, Deretic V. Molecularbasis for the exquisite sensitivity of Mycobacteriumtuberculosis to isoniazid. Proc Natl Acad Sci USA1996;93:13212-6.
138. Springer B, Master S, Sander P, Zahrt T, McFaloneM, Song J, et al. Silencing of oxidative stress responsein Mycobacterium tuberculosis: expression patternsof ahpC in virulent and avirulent strains and effect ofahpC inactivation. Infect Immun 2001;69: 5967-73.
139. Braunstein M, Espinosa BJ, Chan J, Belisle JT,Jacobs WR. SecA2 functions in the secretion ofsuperoxide dismutase A and in the virulence ofMycobacterium tuberculosis. Mol Microbiol2003;48:453-64.
140. Cole ST. Comparative and functional genomics ofthe Mycobacterium tuberculosis complex.Microbiology 2002;148:2919-28.
141. Barletta RG, Donis RO, Chacon O, Shams H,Cirillo JD. Vaccines against intracellular pathogens.Subcell Biochem 2000;33:559-99.