Embed Size (px)
Citation preview

Capítulo VII
INGENIERÍA DE PROTEÍNAS Y EVOLUCIÓN DIRIGIDA
X. SOBERÓN MAINERO
G.M. MONTERO MORÁN
LAS PROTEÍNAS COMO HERRAMIENTAS MOLECULARES
A partir del concepto básico que establece la correspondencia entre ungene y una proteína específica, se pudo anticipar que las primeras apli-caciones, conceptual y metodológicamente más simples de la ingenieríagenética consistirían en la expresión de proteínas heterólogas (es decir,codificadas por genes provenientes de organismos distintos) en célulasbacterianas. En los albores de la ingeniería genética, el paso de obtenerun gene cuyo producto fuera interesante representaba un reto en símismo, pero hoy en día esta situación ha cambiado radicalmente. Porotra parte, el simple hecho de contar con un gene y la capacidad deintroducirlo a una célula capaz de expresarlo, no implica que se dispongade un sistema de producción eficiente de la proteína respectiva, ya queexiste una multitud de variables que pueden afectar los niveles de expre-sión de una proteína dada. De cualquier manera como se ha señalado encapítulos anteriores, la clonación y expresión de la mayoría de los genesespecíficos, de cualquier origen, natural o sintético, es una operaciónrazonablemente rutinaria con las herramientas disponibles hoy en día.
Las proteínas no son solamente el producto primario de los genes,sino que son moléculas que intervienen de manera directa en la realiza-ción de la mayoría de las reacciones biológicas. Las proteínas puedentener funciones estructurales, de síntesis y degradación de compuestos(enzimas), señalización (hormonas, proteínas de superficie), defensa(anticuerpos), realización de trabajo mecánico, etc. En este sentido, lacapacidad de estudiar proteínas expresadas de manera controlada en sis-temas manipulados a través de DNA recombinante ha constituido, desdehace cerca de 30 años, un área efervescente de la investigación biológica
195

experimental. Por ejemplo, a través de una de las herramientas funda-mentales de las metodologías de la ingeniería genética, se tiene la capa-cidad de modificar de manera precisa la secuencia nucleotídica de losgenes, utilizando DNA sintetizado químicamente en el laboratorio. Así,aunque las proteínas han sido estudiadas desde hace mucho tiempo,como productos naturales, la era de la ingeniería genética ha permitidomaterializar una etapa verdaderamente “experimental” en el estudio delas proteínas. Esto es, ha permitido enfocar este estudio al modificarel nivel básico de identidad en una proteína, que es su secuencia de ami-noácidos, en lo que constituye el enfoque denominado “ingeniería deproteínas”.
Debe señalarse, como se ha presentado en el capítulo I, que una pro-teína puede ser definida de manera simple e inequívoca por su estruc-tura primaria, o secuencia de aminoácidos, pero sus propiedades quími-cas y su función biológica resultan de un nivel de organización que vamas allá de esta secuencia, a través del plegamiento espontáneo en tresdimensiones (estructura terciaria) (figura I.15). El proceso por el que elsistema molecular que constituye una proteína típica, es decir una cadenade cientos de aminoácidos (y, por lo tanto, miles de átomos), se pliegaen el espacio es un problema aún no resuelto por la ciencia contempo-ránea. En este sentido, la capacidad de modificar a voluntad la secuenciade aminoácidos de una proteína no significa que tengamos la capacidad depredecir las implicaciones estructurales y funcionales de dichos cambios.
Más de 25 años después del establecimiento de las técnicas básicas dela ingeniería genética, el estudio fundamental de la relación entre lasecuencia, la estructura y la función de las proteínas continúa siendo uncampo muy activo de investigación. El concepto de ingeniería de pro-teínas constituye un enfoque experimental cuya utilidad se manifiestadesde este nivel de generación de conocimiento fundamental. En esen-cia, un investigador analiza la información de que dispone acerca de unaproteína de interés, desde su secuencia de aminoácidos y la secuencia deotras proteínas relacionadas evolutivamente, hasta la función descritapara la misma, las condiciones en que funciona, la estructura tridimen-sional de la misma, la identidad y posición en la secuencia de los amino-ácidos que juegan un papel especialmente importante en la función dela misma. Con esta información se generan hipótesis específicas sobre elpapel que puede jugar un cambio de algún aminoácido en la proteína;se introduce el cambio diseñado en el gene que codifica para la proteína
196

197
Figura VII.1COMPARACIÓN ENTRE EL DISEÑO RACIONAL DE PROTEÍNAS
Y LA EVOLUCIÓN DIRIGIDADurante el diseño racional de proteínas, las mutaciones se planean con base en su estruc-tura y son generadas por mutagénesis sitio dirigida. Después de la transformación en elorganismo huésped, la variante es expresada, purificada y analizada para las propiedadesdeseadas.
La evolución dirigida empieza con la preparación de las librerías de los genesmutantes por mutagénesis al azar, las cuales son, entonces, expresadas en el organismohuésped. Las librerías de proteínas son usualmente evaluadas en placas de microtitu-lación, usando ciertos parámetros de selección. La caracterización de la proteína y elanálisis del producto permite seleccionar las mutaciones deseadas y no deseadas. Larecombinación in vitro por “DNA shuffling”, por ejemplo, puede ser usada para mejorarlas propiedades. Los dos enfoques de ingeniería de proteínas pueden ser repetidosy combinados hasta que el biocatalizador con las características deseadas sea generado.

y se analiza su comportamiento en un sistema de expresión (típicamenteen células bacterianas). Con base en los resultados obtenidos se generannuevas hipótesis y se repite el ciclo (figura VII.1).
Traslademos ahora sólo por un momento, nuestro centro de atenciónde las técnicas básicas de la ingeniería genética para considerar el desti-no que depara la biotecnología a uno de los grupos de proteínas másimportantes del mundo: las enzimas. Para estimar la importancia de estasmoléculas es indispensable saber que controlan las innumerables reac-ciones químicas cotidianas de todos los organismos vivos, desde antes dela concepción hasta después de la muerte. Su papel como catalizadoresconsiste en acelerar las reacciones químicas, formándose o destruyéndo-se moléculas en el proceso. Colaboran en la transmisión de los impulsosnerviosos, evitan la pérdida de la sangre que brota de las heridas e inclu-so regulan la fabricación de la materia básica de la vida: el DNA. Trabajanmuy de prisa, tardando menos de una millonésima de segundo en reali-zar su trabajo. Actualmente, su poderosa actividad biológica se ha apro-vechado con fines industriales. Son importantes en las industrias ali-menticia y química, a causa de su especificidad, eficiencia y potenciaa temperaturas y acidez moderadas. Las enzimas se extraen tradicional-mente de plantas y animales, pero su producción a partir de bacterias,levaduras y hongos está aumentando rápidamente a medida que se dis-pone de más organismos de este tipo y que la ingeniería genética mejo-ra los rendimientos y procesos.
Si consideramos el ámbito aplicado de la ingeniería de proteínas, elesquema planteado se implementa tomando en cuenta objetivos funcio-nales específicos, que confieren una mayor utilidad a la proteína o enzi-ma en cuestión, con la finalidad de mejorar el producto o proceso bio-tecnológico en el que interviene. En lo que resta de este capítulo se haráuna descripción de los retos, las limitaciones, los enfoques y el potencialde la ingeniería de proteínas en el ámbito aplicativo.
BIOCATÁLISIS Y BIOTECNOLOGÍA MODERNA
Como se ha señalado, ya avanzada la segunda mitad del siglo XX, labiotecnología se encontraba asociada a una industria productora de bie-nes y servicios basada en el uso de microorganismos o partes de ellos,relacionada fundamentalmente con las industrias farmacéutica y ali-menticia, y sólo en menor grado con las industrias química y del medio
198

ambiente. Esto es lo que podríamos denominar como biotecnología“tradicional o clásica” que ha permitido a la humanidad hacer vinoo cerveza, producir queso o yogur, a partir de leche, fabricar pan o con-servar carnes, así como producir antibióticos o ácidos orgánicos. Conformesurge el conocimiento científico se perfeccionaron estos procesos y seincluyeron otros como la mejora genética de animales y plantas.
La industria química, todavía cimentada en las reservas petroleras delplaneta, ha visto en la biotecnología apenas una alternativa futura parala producción de materias primas, obtención de energéticos y el desa-rrollo de procesos más limpios para hacer frente al problema de ladegradación del medio ambiente y de la destrucción de la biodiversidad.
Pero es claro que existe una tendencia hacia el desarrollo de unabioindustria sustentada en la capacidad de transformación, mediantecélulas vegetales, animales o microbianas, actuando tanto en reactoresindustriales como en el campo y la granja. Como consecuencia de estaintegración, la industria irá sustituyendo paulatinamente los combustiblesfósiles por renovables, provistos por la propia industria biotecnológica.También cabe destacar que a este campo tecnológico lo caracteriza unanecesidad moderada de inversión, la existencia actual de recursos huma-nos capacitados, y la ventaja natural para aquellos países que, comoMéxico, cuentan con regiones de diversas características climáticas y sobretodo con una gran riqueza de recursos genéticos debido a su biodiversidad.
Ahora bien, como se ha señalado en capítulos previos, la biotecnologíamoderna está basada en los avances científicos que se iniciaron en ladécada de los setenta, especialmente en biología molecular, que hanpermitido aislar y modificar información genética de los organismos,mediante las técnicas de DNA recombinante. Así, los alcances de la nuevabiología permiten visualizar un nuevo concepto de industria y un nuevoconcepto de la actividad industrial con el que sin duda culminaremos elsiglo XXI. Se trata de una industria que, abarcando todas las esferas de laactividad económica, estará caracterizada por el conocimiento profundoy la capacidad de modificación y de la reingeniería misma de la célulaviva, la cual tenderá a transformar a la industria química basada en elpetróleo y las transformaciones químicas, a una industria bioquímica,basada en las biomasas, en las transformaciones biológicas y en la biocatá-lisis (procesos que utilizan enzimas para acelerar las reacciones químicas).
Toda esta nueva generación de conocimiento permite definir a labiotecnología moderna tal y como ha sido previamente señalado como“la utilización de organismos vivos, o parte de los mismos, para obtener
199

o modificar productos, mejorar animales o plantas, o desarrollar micro-organismos para objetivos específicos”. En este concepto se incluyen labiotecnología tradicional y la moderna. En la moderna se incluye el usoindustrial de las técnicas de DNA recombinante, la producción de anti-cuerpos monoclonales, los bioprocesos, la ingeniería de proteínas y eluso de enzimas. Este último concepto, que consiste en el uso de enzimasen los procesos de la industria química, es el objeto primordial de estecapítulo.
RETOS A RESOLVER. MITOS Y REALIDADES
La mencionada convergencia entre la maduración de tecnologíasestablecidas y la aparición reciente de tecnologías habilitadoras poten-ciará fuertemente el desarrollo de la biotecnología en el campo indus-trial. Adicionalmente, la batería de herramientas contribuida por el DNA
recombinante ha hecho posible el tránsito hacia el conocimiento de losgenomas completos de microorganismos, plantas y animales. Como con-secuencia, el número de especies y procesos en los que es viable pensaren aplicaciones biotecnológicas crece constantemente.
En este contexto general, se observa un despegue importante delámbito industrial como una tercera fase de aplicaciones de la biotecno-logía, debido a la inserción del potencial de la biocatálisis y las tecnolo-gías habilitadoras que impactan su desarrollo.
En la actualidad, a pesar de representar una proporción muy peque-ña de la actividad industrial, ya se emplean las enzimas en un buennúmero de procesos (tabla VII.1).
Cabe preguntarse en este punto: ¿por qué es hasta ahora que se espe-ra un impacto importante de las tecnologías biológicas en la industriaquímica en general? Parecería obvio que en muchos procesos químicosindustriales, la tecnología biológica ofrecería una alternativa limpia y efi-ciente. Después de todo, una observación superficial de la materia vivaque nos rodea muestra elocuentemente su enorme capacidad de síntesisde toda una diversidad de compuestos químicos. Hasta la más simplebacteria alberga en su interior literalmente miles de compuestos quími-cos distintos. Vale la pena reparar en que los seres humanos seguimosmostrando un aprecio especial por las materias primas naturales, talescomo el algodón, la lana, las pieles y las maderas, aun cuando estamos
200

por ingresar al tercer milenio. Asimismo, casi la totalidad de los medica-mentos que hoy utilizamos y la mayoría de los que se están desarrollan-do tienen su origen en productos naturales, notablemente de plantasmedicinales. Por otra parte, existe un sinnúmero de compuestos sintéti-cos cuyas propiedades y bajo costo de fabricación los hacen difícilmentesustituibles por productos naturales. Dentro de este juego de fuerzas,aparentemente encontradas, debemos ubicar el potencial de la tecnolo-gía biológica en el ámbito de la industria química.
Podemos hacer un recuento esquemático de las ventajas y desventajasde los procesos enzimáticos y de las percepciones que de ellas tienen losactores principales en este campo. En el lado de las ventajas (algunas delas cuales ya se han esbozado arriba) se citan frecuentemente las siguien-tes (1).
— Se pueden lograr niveles de especificidad muy altos, especialmenteútiles cuando se trabaja con materias primas complejas.
— Típicamente operan en condiciones relativamente suaves de tem-peratura y presión. Las instalaciones y equipos utilizados en los pro-cesos fermentativos y biocatalíticos son independientes del pro-ducto particular de que se trate. Como consecuencia, la inversiónrealizada en estos equipos puede ser, en principio, reutilizable ennuevos procesos conforme surgen demandas y oportunidades.
— En algunos casos, los bioprocesos son la única opción para la sín-tesis de compuestos complejos. Esto es particularmente cierto paracompuestos asimétricos (quirales), cada vez más necesarios en esta-do puro en el sector farmacéutico y alimentario, por razones regu-latorias o nutrimentales.
— Los bioprocesos son inherentemente benignos con el ambiente y dehecho son parte integral de los ciclos de asimilación de la materiaorgánica en el planeta.
En el lado de las desventajas, podemos contar las siguientes (algunasde las cuales tienen que ver más con la percepción que actualmente semanifiesta en gran parte de la industria química que con desventajasinherentes o insalvables):
— Las enzimas son caras.— Las enzimas son inestables.
201
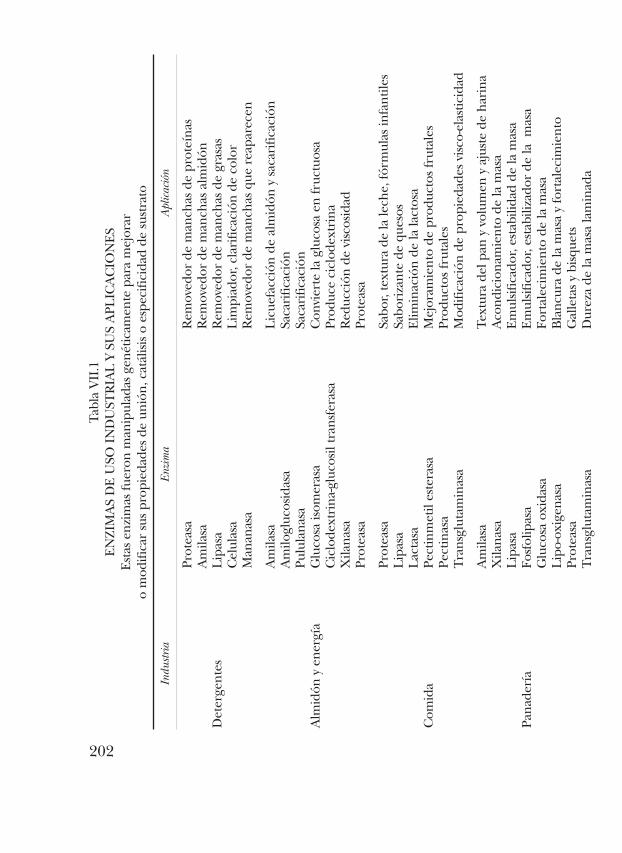
202
Tabl
a V
II.1
EN
ZIM
AS
DE
USO
IN
DU
STR
IAL
Y S
US
APL
ICA
CIO
NE
SE
stas
enz
imas
fuer
on m
anip
ulad
as g
enét
icam
ente
par
a m
ejor
aro
mod
ifica
r su
s pr
opie
dade
s de
uni
ón, c
atál
isis
o e
spec
ifici
dad
de s
ustr
ato
Indu
stria
Enzi
ma
Apl
icac
ión
Prot
easa
Rem
oved
or d
e m
anch
as d
e pr
oteí
nas
Am
ilasa
Rem
oved
or d
e m
anch
as a
lmid
ónD
eter
gent
esL
ipas
aR
emov
edor
de
man
chas
de
gras
asC
elul
asa
Lim
piad
or, c
lari
ficac
ión
de c
olor
Man
anas
aR
emov
edor
de
man
chas
que
rea
pare
cen
Am
ilasa
Lic
uefa
cció
n de
alm
idón
y s
acar
ifica
ción
Am
ilogl
ucos
idas
aSa
cari
ficac
ión
Pulu
lana
saSa
cari
ficac
ión
Alm
idón
y e
nerg
íaG
luco
sa is
omer
asa
Con
vier
te la
glu
cosa
en
fruc
tuos
a C
iclo
dext
rina
-glu
cosi
l tra
nsfe
rasa
Prod
uce
cicl
odex
trin
aX
ilana
saR
educ
ción
de
visc
osid
adPr
otea
saPr
otea
sa
Prot
easa
Sabo
r, te
xtur
a de
la le
che,
fórm
ulas
infa
ntile
sL
ipas
aSa
bori
zant
e de
que
sos
Lac
tasa
Elim
inac
ión
de la
lact
osa
Com
ida
Pect
inm
etil
este
rasa
Mej
oram
ient
o de
pro
duct
os fr
utal
esPe
ctin
asa
Prod
ucto
s fr
utal
es
Tra
nsgl
utam
inas
aM
odifi
caci
ón d
e pr
opie
dade
s vi
sco-
elas
ticid
ad
Am
ilasa
Text
ura
del p
an y
vol
umen
y a
just
e de
har
ina
Xila
nasa
Aco
ndic
iona
mie
nto
de la
mas
aL
ipas
aE
mul
sific
ador
, est
abili
dad
de la
mas
aPa
nade
ría
Fosf
olip
asa
Em
ulsi
ficad
or, e
stab
iliza
dor
de la
mas
aG
luco
sa o
xida
saFo
rtal
ecim
ient
o de
la m
asa
Lip
o-ox
igen
asa
Bla
ncur
a de
la m
asa
y fo
rtal
ecim
ient
oPr
otea
saG
alle
tas
y bi
sque
tsT
rans
glut
amin
asa
Dur
eza
de la
mas
a la
min
ada
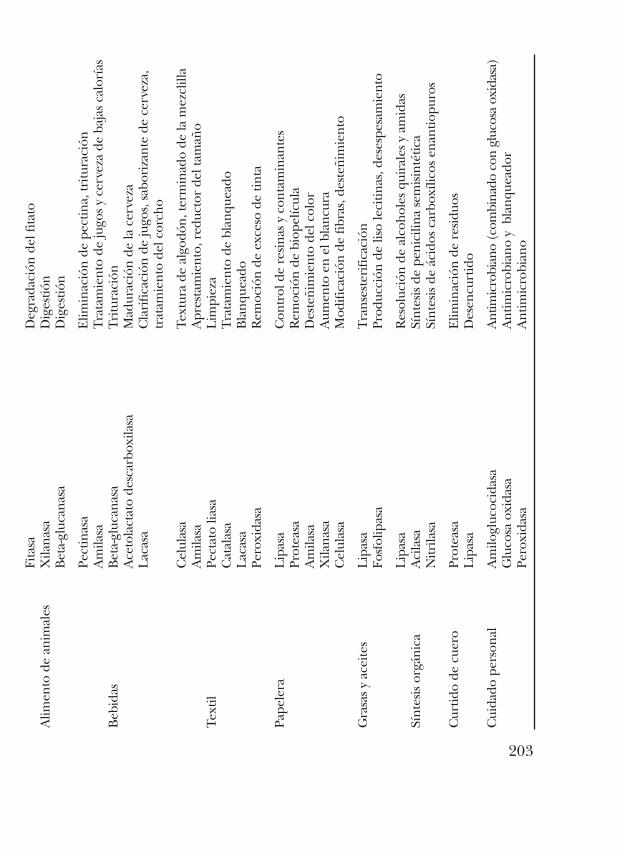
203
Fita
saD
egra
daci
ón d
el fi
tato
Alim
ento
de
anim
ales
Xila
nasa
Dig
estió
nB
eta-
gluc
anas
aD
iges
tión
Pect
inas
aE
limin
ació
n de
pec
tina,
tritu
raci
ónA
mila
saT
rata
mie
nto
de ju
gos
y ce
rvez
a de
baj
as c
alor
ías
Beb
idas
Bet
a-gl
ucan
asa
Tri
tura
ción
Ace
tola
ctat
o de
scar
boxi
lasa
Mad
urac
ión
de la
cer
veza
Lac
asa
Cla
rific
ació
n de
jugo
s, sa
bori
zant
e de
cer
veza
,tr
atam
ient
o de
l cor
cho
Cel
ulas
aTe
xtur
a de
alg
odón
, ter
min
ado
de la
mez
clill
aA
mila
saA
pres
tam
ient
o, r
educ
tor
del t
amañ
oTe
xtil
Pect
ato
liasa
Lim
piez
aC
atal
asa
Tra
tam
ient
o de
bla
nque
ado
Lac
asa
Bla
nque
ado
Pero
xida
saR
emoc
ión
de e
xces
o de
tint
a
Pape
lera
Lip
asa
Con
trol
de
resi
nas
y co
ntam
inan
tes
Prot
easa
Rem
oció
n de
bio
pelíc
ula
Am
ilasa
Des
teñi
mie
nto
del c
olor
Xila
nasa
Aum
ento
en
el b
lanc
ura
Cel
ulas
aM
odifi
caci
ón d
e fib
ras,
dest
eñim
ient
o
Gra
sas
y ac
eite
sL
ipas
aT
rans
este
rific
ació
nFo
sfol
ipas
aPr
oduc
ción
de
liso
leci
tinas
, des
espe
sam
ient
o
Lip
asa
Res
oluc
ión
de a
lcoh
oles
qui
rale
s y
amid
asSí
ntes
is o
rgán
ica
Aci
lasa
Sínt
esis
de
peni
cilin
a se
mis
inté
tica
Nitr
ilasa
Sínt
esis
de
ácid
os c
arbo
xílic
os e
nant
iopu
ros
Cur
tido
de c
uero
Prot
easa
Elim
inac
ión
de r
esid
uos
Lip
asa
Des
encu
rtid
o
Cui
dado
per
sona
lA
milo
gluc
ocid
asa
Ant
imic
robi
ano
(com
bina
do c
on g
luco
sa o
xida
sa)
Glu
cosa
oxi
dasa
Ant
imic
robi
ano
y b
lanq
uead
orPe
roxi
dasa
Ant
imic
robi
ano

— Su productividad es baja.— Las reacciones redox requieren cofactores difíciles de reciclar.— Las enzimas no catalizan reacciones interesantes para la industria.
Desde nuestro punto de vista, más que hablar de desventajas podría-mos enfocarnos en los retos a vencer para la utilización más generaliza-da de las enzimas y los elementos tecnológicos que pueden ir contribu-yendo a resolverlos.
Hasta épocas recientes la disponibilidad de enzimas había sido bas-tante limitada. Sin embargo, en los últimos veinte años, esta situaciónestá siendo rápidamente revertida por las dos manifestaciones de las téc-nicas del DNA recombinante mencionadas anteriormente. Por una parte,la posibilidad de clonar los genes e introducirlos en sistemas que permi-ten su expresión abundante y controlada ha permitido ir dejando atrásla limitación de materia prima inherente a los sistemas naturales (porejemplo, cuando la fuente de la enzima útil era un tejido poco abun-dante de una especie rara). Por otra parte, el desarrollo de los proyectosgenómicos, aunado a la capacidad de aislar genes directamente de mues-tras ambientales como aguas y suelos (sin necesidad de cultivar los micro-organismos ahí presentes), ha multiplicado, literalmente por miles, elnúmero de enzimas que pueden potencialmente utilizarse.
Otro obstáculo importante se deriva de la historia natural de los siste-mas vivientes. Los atributos de los catalizadores biológicos han sido mol-deados por la selección natural. En consecuencia, la función, estabilidady respuesta a factores ambientales de una enzima están directamenterelacionados con su función y ambiente biológicos. De ninguna manerase puede esperar que sean óptimos para funcionar en procesos indus-triales, con miras a la obtención de productos provenientes de la deman-da del mundo moderno. Es entonces evidente que se necesita, virtual-mente en todos los casos, modificar las propiedades de las enzimas paraadaptarlas, inclusive en grados bastante drásticos para los requisitos deestabilidad, especificidad, etc., planteados en función del proceso indus-trial en cuestión (figura VII.2) (2). En este ámbito, el impacto del cono-cimiento científico se ha mostrado de manera más lenta. Como se expli-cará más adelante, la modalidad de ingeniería de proteínas por evolucióndirigida está contribuyendo de manera notable a vencer esta limitación.
Finalmente, es crucial tomar en cuenta que, independientemente delos horizontes de aplicación que se perciben o pronostican desde el sec-tor científico/académico, la asimilación de los mismos por parte de la
204

205
Figura VII.2ESTRATEGIAS PARA EL DESARROLLO DE ENZIMAS DE ALTA EFICIENCIA
Los puntos claves para que la enzima cumpla con los requisitos del desarrollo del procesoson cubiertos por métodos moleculares, entre ellos, mutagénesis al azar y evolucióndirigida de proteínas.

industria está dominada por los aspectos comerciales y las fuerzas delmercado. En otras palabras, los procesos comerciales que se vayan plan-teando tienen que resultar económicamente viables. En este ámbito, valela pena observar que las empresas productoras de productos químicoshan hecho cuantiosas inversiones en sus plantas actuales, mismas quedeben ser amortizadas en periodos de tiempo largos. Por ello, cualquierproceso nuevo, biológico o no, que pretenda sustituir a uno ya estableci-do, se enfrenta no sólo a la necesidad de presentar viabilidad técnica,sino que tiene que remontar a las inercias e inversiones previamente esta-blecidas (figura VII.3).
ENFOQUES
La ingeniería de proteínas “clásica”
Como ya se mencionó, es factible emitir hipótesis concretas sobre losefectos que se esperan de algún cambio específico y discreto en lasecuencia de aminoácidos de una proteína. Es también relativamentesencillo implementar estos cambios, a través de ingeniería genética, demanera que se produzca la proteína alterada y se observe la concordan-cia entre la predicción y la realidad.
Un prerrequisito para el estudio de la ingeniería de proteínas es quela enzima ya haya sido clonada y expresada (figuras II.15 y IV.2).
La metodología ideal para confirmar el papel de un residuo de ami-noácido en particular, previamente identificado por otros métodos(modificación química, predicción, cristalografía, resonancia magnéticanuclear [RMN]), ya sea en la estructura o en la función, es la mutagéne-sis dirigida. Ésta se realiza mediante la técnica conocida como mutagé-nesis por oligonucleótido, desarrollada inicialmente por Hutchinson ycolaboradores y modificada por Kunkel, en la que un oligonucleótidosintético se emplea como mutágeno altamente específico (3). La muta-génesis dirigida consiste en cuatro pasos fundamentales (figura VII.4) (4).
Debe notarse que el enfoque clásico de la ingeniería de proteínas quese muestra en la figura VII.1 se describe como un ciclo, es decir, se destacaque es poco probable que en un solo intento se logre el resultado deseado,debido a que la capacidad predictiva de la que disponemos actualmentees muy limitada.
De cualquier manera, existen un buen número de casos exitosos demejoras en las propiedades de proteínas útiles valiéndose de este enfoque.
206

207
Figura VII.3ENFOQUE DE BIOPROCESOS
El primer paso en el desarrollo de un proceso es identificar la reacción. Con base en ésta,se define el biocatalizador ideal, se caracteriza y modifica el proceso. Subsecuentementeel biocatalizador es producido, se aplica en el bioproceso y el producto se recupera.

Podemos considerar que hay dos tipos generales de información en lasque se basan los diseños empleados en ingeniería de proteínas. Por unaparte, se encuentran los que provienen del conocimiento de las propie-dades fisicoquímicas de los sistemas proteicos. Por otra parte, se puedenincorporar en el análisis los datos aportados por las secuencias de ami-noácidos de varias proteínas similares a la que es objeto de la experi-mentación, es decir, inferir posibles alteraciones a partir de las solucio-nes que la naturaleza ya ha ensayado. Por ejemplo, a partir de conceptosfisicoquímicos, se han logrado estabilizar proteínas implementando puen-tes disulfuro (enlaces covalentes que fortalecen la estructura tridimen-sional), puentes salinos o tratando de optimizar el empaquetamiento delos residuos internos. En la proteasa subtilisina se identificó mediantemutagénesis al azar, que la modificación de los residuos de los aminoá-cidos Lys211 y Arg212 aumentan su termoestabilidad (5,6).
Por otro lado, Liebeton y col. 2000 (7), aumentaron la enantioselecti-vidad de una lipasa de Pseudomonas aeruginosa, sugiriendo que el cambioen esta propiedad se debe a un aumento en la flexibilidad de las asas leja-nas al sitio de unión del sustrato (8). Otro caso exitoso en la modificaciónde las propiedades enantioselectivas se realizó en una enzima hidantoi-nasa, donde la sustitución de un solo residuo de aminoácido de D-metio-nina a L-metionina fue suficiente para invertir la enantioselectividad dela enzima (9). En la industria de los detergentes se han modificado ami-lasas para aumentar su actividad a bajas temperaturas y a pH alcaninossin comprometer su estabilidad. Existen muchos ejemplos de enzimasque son utilizadas a nivel industrial que han sido modificadas; algunosejemplos de las enzimas más utilizadas en diferentes campos de la indus-tria y sus usos se muestran en la tabla VII.1 (10). Similarmente, se hanlogrado implementar sitios de ligación a metales en proteínas que origi-nalmente no lo hacían. En cuanto al uso de secuencias similares (infor-mación filogenética), hay ejemplos notables en los que secuencias ine-xistentes en la naturaleza (por ejemplo secuencias consenso) resultantener propiedades útiles distintas a cualquiera de las proteínas naturales,sin pérdida de la especificidad y actividad catalítica. También ha sido útilel análisis de proteínas homólogas para diseñar cambios de especificidada cofactores a través de un número pequeño de cambios de ciertos resi-duos de aminoácidos (11).
Incluso varias enzimas ya han sido manipuladas genéticamente paramejorar la catálisis en los procesos industriales (tabla VII.2). Éstos inclu-yen proteinasas, celulasas, lipasas, α-amilasas y glucoamilasas. Uno de los
208

mejores ejemplos es una enzima xilanasa, la cual necesita ser estable aaltas temperaturas y ser activa a temperaturas y pH fisiológicos cuando esutilizada como un aditivo de alimentos y en condiciones alcalinas cuan-do es usada en el blanqueo en la industria del papel.
Uno de los organismos en que se produce la enzima xilanasa a nivelindustrial es el hongo del género Trichoderma. Ésta ha sido purificaday cristalizada. Su estabilidad térmica ha sido incrementada cerca de 2 000veces a 70 ºC, y su pH óptimo fue desplazado una unidad de pH hacia laalcalinidad por mutagénesis racional. La estrategia consistió en la modi-ficación de una región alfa hélice y del amino terminal.
La inspección de los logros obtenidos hasta ahora nos indica que lacapacidad de diseñar verdaderamente cambios útiles, a partir de losconocimientos disponibles, es todavía limitada. Especialmente tomandoen cuenta que, con toda certeza, existe un gran número de intentos falli-dos que ni siquiera aparecen en la literatura científica. Sin duda, en lospróximos años el número de ejemplos exitosos deberá ir aumentando,como resultado del correspondiente aumento en nuestro conocimientosobre los sistemas proteicos. Es notable que la capacidad de cálculo delos sistemas computacionales siga aumentando de manera geométrica,dando la posibilidad de implementar algoritmos cada vez más deman-dantes y precisos para la predicción de la relación entre secuencia,estructura y función en las proteínas, sugiriendo un futuro brillante.
209
Tabla VII.2Se muestran algunas de las enzimas más utilizadas a nivel industrial y las carac-
terísticas enzimáticas que han sido modificadas por ingeniería de proteínas.
Enzima Industria Característica deseada
Xilanasa alimentos Estabilidad térmica,papel y su pulpa actividad a pH ácidos
Estabilidad térmicay a pH alcalinos
Glucoamilasa almidón Aumento en el pH óptimo
Glucosa isomerasa fructosa Especificidad de sustratoEstabilidad a pH ácido
Termoestabilidad
Proteinasa detergentes TermoestabilidadEstabilidad a pH alcalino
Estabilidad oxidativa

La capacidad de comprender los sistemas proteicos al grado de diseñar-los es, claramente, una meta fundamental de gran mérito científico. Peroes claro que los resultados más recientes siguen siendo de interés paraquienes desarrollan los métodos y enfoques, sin tener aún un efectonotable en el sector aplicativo, dada la pobre capacidad predictiva. Comose ha señalado, en la figura VII.1, panel izquierdo, se muestra un diagra-ma del proceso básico de la ingeniería de proteínas, y se le compara conlos elementos del enfoque alternativo, llamado evolución dirigida.
Evolución dirigida
En vista del enorme reto que significa el diseño racional de proteínas,basado en el conocimiento estructural y mecánistico, y de lo distante queaún se ve el logro de esta meta en un nivel tecnológicamente útil, ¿quéalternativa queda hoy para avanzar?
En los últimos 10 o 15 años, se ha desarrollado notablemente la expe-rimentación basada en un concepto intuitivamente sencillo, que implicaimitar al proceso natural que, para empezar, ha dado origen a las enzi-mas existentes hoy en día. Es decir, se trata de imitar el proceso evoluti-vo que ocurre día a día en la naturaleza. Para comprender los enfoquesque se han adoptado debe tenerse en cuenta que el proceso natural seha dado en el transcurso de miles de millones de años, operando sobrecantidades vastas de materia viva. Así, aun cuando desde hace muchosaños se han llevado a cabo experimentos de evolución en condiciones delaboratorio, los alcances de estos procedimientos dependen del grado enque el proceso evolutivo se pueda compactar y acelerar. El surgimientode las técnicas del DNA recombinante estableció las bases para avanzardramáticamente en esta dirección, en particular cuando el objeto de evo-lución está en el nivel molecular, como es el caso de las enzimas. La apli-cación de los procesos evolutivos para mejorar las características de bio-moléculas se ha llamado evolución dirigida (también evolución forzadao, simplemente, evolución in vitro).
Se puede decir que la evolución dirigida de proteínas in vitro se iniciódesde el trabajo pionero de Spiegelman en 1967 cuando demostró quese podía reproducir in vitro el proceso darwiniano propuesto como hipó-tesis del origen de la evolución (12). Esta idea, que originalmente naceen un esquema de investigación básica para tratar de entender el origendel proceso evolutivo, fue posteriormente explotada para generar, demanera acelerada, proteínas con propiedades modificadas. El grupo
210

de Arnold, uno de los pioneros en este campo, demostró que llevandoa cabo ciclos iterativos de mutagénesis y selección se podía evolucionaruna proteína en cuestión de meses o incluso semanas (6, 13). La evolucióndirigida, como área, tiene dos retos por afrontar: el primero es la genera-ción de diversidad y el segundo es la selección de las mutantes. En amboscasos, los procesos implementados en el laboratorio se inspiran en los pro-cesos naturales, pero difieren de ellos en razón de las posibilidades tec-nológicas y de la necesidad de lograr la compactación y aceleración refe-ridos anteriormente.
En el proceso natural de evolución, el material genético sufre muta-ciones (figura I.13), inherentemente aleatorias, a través de diversosmecanismos. Los más sencillos serían los errores cometidos al replicar elDNA y los causados por las lesiones en esta macromolécula por efecto deagentes físicos (por ejemplo radiación) o químicos (compuestos muta-génicos, como nitritos entre otros muchos).
Existen desde luego otros muchos fenómenos que contribuyen a lageneración de diversidad genética, notablemente los procesos de recom-binación, que juegan un papel preponderante en la variabilidad obser-vada en la reproducción sexual. En el caso de la evolución in vitro, es fac-tible emplear agentes físicos y químicos, pero la principal adaptación quese hace es concentrar la acción de estos agentes en segmentos específi-cos de DNA, concretamente en genes clonados. En la actualidad, estosmétodos han sido virtualmente suplantados por los que se sustentan enla acción de enzimas replicativas de DNA, pero en condiciones en las quela tasa de error se ha logrado aumentar en gran medida (hasta cerca de1%, comparado con 0.04% en condiciones biológicas típicas). La mayorversatilidad y eficiencia la da la utilización de DNA sintético, que permitela introducción de variabilidad en las secuencias genéticas de maneramuy controlada, tanto en la tasa de mutación (que puede llegar a ser de100%) como en la zona o región del gene de interés a la que se quieremodificar (figura VII.4).
Adicionalmente, en los últimos años se han establecido métodos quepermiten incluir el elemento de recombinación, de importancia funda-mental en la reproducción sexual, a los procesos de generación de varia-bilidad in vitro (figura VII.5).
El entremezclado de genes o “DNA shuffling”, consiste en digerir alazar una colección de genes hasta obtener pequeños fragmentos de DNA
de aproximadamente 100 pares de nucleótidos para reconstruir a partir deellos el gene mediante la técnica de PCR (figura II.12). En esta técnica
211

212
Figura VII.4REPRESENTACIÓN ESQUEMÁTICA DE LA TÉCNICA
DE LA MUTAGÉNESIS DIRIGIDAEl plásmido con una hebra de DNA del gene que va a ser mutado, es apareado con unfragmento de DNA complementario que contiene la mutación que será incorporada (. ).Esta mutación puede ser el reemplazo, eliminación o inserción de uno o más nucleótidos;el único requerimiento es que los segmentos complementarios flanqueantes de los oligo-nucleótidos sean suficientes para permitir que se apareen específicamente al plásmido. En este punto se introduce variabilidad que puede ser desde un cambio sencillo que ge-nera un solo cambio a nivel de codón, o bien, a saturación logrando producir hasta 1 013variantes. La variabilidad puede crearse con técnicas como: PCR mutagénica, entremez-clado de genes (DNA-shuffling), Itchy y química ortogonal. Ésta ultima consiste en unnuevo método de mutagénesis ejecutado a nivel de codón que permite combinarcodones silvestres y mutantes a lo largo de la ventana que se quiere modificar, lo cual esimposible con los métodos convencionales de oligonucleótidos degenerados (NNGC),además tiene la ventaja de que no hay redundancia como en el código genético. Por otrolado, los métodos enzimáticos y químicos están limitados a generar principalmente muta-ciones puntuales y cada aminoácido silvestre puede cambiar a sólo una fracción de losotros 19 diferentes aminoácidos.
Una segunda cadena de DNA se sintetiza enzimáticamente a partir de la primera cade-na, usando el oligonucleótido como iniciador para incorporar la mutación. El plásmidohíbrido resultante es utilizado para la transformación de las células y generar DNA dedoble cadena. Las moléculas de DNA que contienen la mutación en ambas cadenaspueden ser seleccionadas o identificadas por la secuenciación de su DNA.
Clonación y seleccióndel plásmido con las
mutaciones

213
Figura VII.5 RECOMBINACIÓN DE SECUENCIAS DE GENES Y GENERACIÓN
DE VARIABILIDAD Representación esquemática de algunas estrategias in vitro: A) “DNA shuffling” o entre-mezclado de genes para la recombinación de miembros de una familia de genes consecuencias de alta homología. El primer paso de este método consiste en fragmentarcon DNAsaI una población de genes relacionados. Los fragmentos de DNA generados dediferentes longitudes se reensamblan, es decir, se unen los extremos 5’ de un fragmentocon los extremos 3’ de otro. Los fragmentos con extremos 5’ pueden ser extendidosmediante PCR con la enzima Taq DNA polimerasa dejando los fragmentos con extremos 3’intactos. Como una consecuencia de esta extensión, la longitud promedio de cada frag-mento se incrementa durante cada ciclo (figura II.15). La recombinación ocurre, enton-ces, cuando un fragmento derivado de un templado sirve como oligonucleótido iniciadoren un templado con una diferente secuencia.
B) Estrategia “ITCHY” para la recombinación de dos genes con poca o ninguna secuen-cia homóloga. Itchy o método de truncamiento cíclico o repetido se utiliza para la crea-ción de enzimas híbridas. Se basa en la generación de librerías de fragmentos aminoo carboxilos terminales de dos genes por eliminación progresiva de las secuencias codi-ficantes por la enzima exonucleasa III seguida por la ligación de los productos paragenerar una librería híbrida.
A B

de entrelazado de genes, se intercambian fragmentos de genes homólo-gos generando un camino eficiente para crear proteínas con nuevas carac-terísticas (figura VII.5).
Una variante de este método es el llamado truncamiento por pruebay error para la creación de enzimas híbridas (ITCHY), el cual está basadoen la generación de librerías de fragmentos de DNA que codifican paralas regiones aminos o carboxilo terminales de dos genes por trunca-miento progresivo de las secuencias nucleotídicas codificantes pormedio de la acción de la enzima exonucleasa III seguido por ligación delos productos (14, 15).
Por medio de estos sistemas es factible, y relativamente sencillo, gene-rar en un pequeño tubo de ensayo, millones de millones (1012) de genesdistintos. El reto fundamental consiste en diseñar el tipo de variabilidadque se quiere explorar, que siempre será una fracción muy pequeña deltotal de variantes posibles en los sistemas genéticos (baste decir que ungene de tamaño típico consiste en unos 1 000 pares de nucleótidos, y queel número de combinaciones posibles para una secuencia de esta longi-tud es mayor al número de átomos del universo conocido). Adicional-mente, la variabilidad que hoy día es factible generar con los sistemasin vitro no permite aún implementar eficientemente algunos de losesquemas de intercambio de segmentos que han estado presentes enlos sistemas naturales.
En lo que se refiere a la selección de las mutantes obtenidas por estasmetodologías, ésta opera en la naturaleza a nivel del éxito reproductivode la población. La mayoría de las mutaciones sencillas son, para estosefectos, ya sea neutrales o perjudiciales. Ocasionalmente aparece algunamodificación con efecto positivo, la cual tiende a prosperar en la pobla-ción. Los sistemas usados en evolución dirigida pueden ser similares, porejemplo creando células microbianas que sólo puedan sobrevivir cuandoexpresan un gene (proveniente de una colección de mutantes), con lacaracterística deseada. Desafortunadamente, esto sería factible solamenteen una mínima proporción de los casos para los que se requerirán enzi-mas con atributos modificados. Por esta razón, en evolución moleculardirigida se recurre no sólo a la selección verdadera, sino también a diver-sos sistemas de tamizaje, los más eficientes posibles. Evidentemente, si setuviese que inspeccionar la actividad catalítica de las enzimas variantesuna por una, no estaríamos hablando de evolución dirigida, sino de inge-niería de proteínas clásica (y sería totalmente inútil generar 1012 varian-tes). Así, en el ámbito de la selección, se ha tenido que recurrir también
214

a una buena dosis de imaginación y a la incorporación de los mayoresavances tecnológicos para ampliar la gama de reacciones que puedenevaluarse de manera rápida y paralela. Específicamente, se ha recurri-do a sistemas de detección muy simplificados basados en el aspecto(color, por ejemplo) de las colonias bacterianas surgidas de una colec-ción numerosa de variantes. En el siguiente nivel, se utilizan sistemasrobóticos para implementar las reacciones enzimáticas en formatos deplacas (por ejemplo, las utilizadas desde hace tiempo en ensayos paraanticuerpos en contextos clínicos). Estos sistemas se han ido perfeccio-nando y sofisticando, hasta lograr lo que se conoce hoy día como HTS
(High Throughput Screening o tamizado de alta productividad).Un importante reto en la evolución dirigida de proteínas es lograr un
ensayo de alta productividad en el que se detecten precisamente los atri-butos deseados, que pueden ser varios simultáneos (por ejemplo estabi-lidad y especificidad) y de cierta complejidad.
ÁMBITO DE OPORTUNIDADES
Retomemos la referencia a la diversidad natural de compuestos quí-micos, mencionada previamente en este capítulo. Es evidente que losseres vivos han logrado fabricar todo tipo de sustancias y materiales conbase en procesos biocatalíticos. Paradójicamente, aunque estos produc-tos naturales se cuentan por millones, y cumplen funciones variadísimas,la actividad humana ha requerido la creación de muchas sustanciasy materiales nuevos, pues el desarrollo de las sociedades tecnológicas haido planteando sus propias necesidades. Evidentemente, muchos de losartefactos usados por la sociedad se basan en productos que no son natu-rales (notablemente aleaciones de metales y plásticos). De toda la varie-dad de compuestos que requieren las sociedades modernas, ¿cuálesserán los más susceptibles de ser producidos mediante procesos biológi-cos? Como se sugiere en la figura VII.6, realmente existe una buenadiversidad de aplicaciones para los procesos biológicos. Podemos apro-ximarnos a la vasta gama de procesos clasificándolos desde el punto devista del valor del producto y el volumen de su mercado. Los productosfarmacéuticos son, usualmente, compuestos cuyos precios son muy altosy de un requerimiento relativamente pequeño. En este campo, es renta-ble una cuantiosa inversión en investigación y la implementación de pro-cesos complejos de síntesis y purificación. Estas aplicaciones se pueden
215

satisfacer típicamente con componentes biológicos purificados, talescomo enzimas aisladas. En un siguiente nivel encontramos los productosde la llamada química fina, es decir productos que se utilizan en canti-dades relativamente pequeñas, con precios intermedios. En estos casos,el volumen de producción puede requerir la intervención de procesosmás simples, por ejemplo los que utilizan microorganismos completos,ya sea como biocatalizadores, para llevar a cabo una sola reacción, o como“biorreactores”, en los que el producto resulta directamente del meta-bolismo celular. A medida que nos movemos hacia los productos de mayorvolumen y menor valor, los llamados “commodities” o productos de altovolumen, se puede requerir la intervención de sistemas mucho más bara-tos, tales como sistemas agronómicos, en los que la biosíntesis del pro-ducto se asocia a procesos tradicionales de gran escala y bajo costo. Existeuna continuidad en cuanto al empleo de estos diversos sistemas, depen-diendo de la aplicación concreta, y en la actualidad existen ejemplospara cada uno de estos enfoques.
216
Figura VII.6LA BIOCATÁLISIS AMPLÍA EL RANGO DEL USO DE ORGANISMOS
Y SUS APLICACIONESSe muestran algunos ejemplos de los diferentes procesos biológicos y sus aplicacionesa nivel industrial.

Un escenario ideal podría consistir en que todos los productos quími-cos y materiales fueran “productos naturales”, en cuanto a que surgierande procesos biológicos limpios. Para acercarse a esta situación, requeri-mos un manejo cada vez más sofisticado de los organismos vivos tratadoscomo sistemas. Se trata de comprender y modificar los sistemas metabó-licos, de manera que el flujo de materia y energía esté adaptado para laproducción del insumo requerido. A este enfoque, que utiliza a laingeniería de vías metabólicas y la ingeniería de proteínas, lo hemosdenominado “ingeniería celular” y se describe en el siguiente capítulo.
Es indudable que la intervención en los procesos vivientes, en la medi-da en que son cercanos a lo que comemos y a aquello de lo que nosotrosmismos estamos hechos, genera una buena dosis de ansiedad y miedo envarios sectores de la sociedad. Es claro, por otro lado, que los procesosbiológicos son casi siempre inherentemente más promisorios en su res-peto al entorno que los procesos tradicionales. Se debería procedertomando las debidas precauciones, al tiempo que, con decisión y visiónrealista, vayamos hacia la consecución de estos fines.
BIBLIOGRAFÍA
1. Rozzell D., Commercial Scale Biocatalysis:Myths y Realities. Bioorg. Med.Chem. 7: 2253-2261 (1999).
2. Kathryn M., M. Koeller, Chi-Huey Wong. Enzymes for chemical synyhesis.Nature Insight 409: 232-239 (2001).
3. Kunkel T. The use of oligonucleotide-directed mutagenesis to probe mole-cular mechanisms of mutagenesis. Nucleic. Acids. Symp. Ser. 19: 43 (1988).
4. Carter, P. Site-directed mutagenesis. Biochem. J. 237: 1-7 (1986).5. Bornscheuer W., M. Pohl. Improved biocatalysts by directed evolution and
rational protein design. Curr. Opin. in Chem. Biol. 5: 137-143 (2001).6. Miyazaki K., F. Arnold. Exploring nonnatural evolutionary pathways by sat-
uration mutagenesis: rapid improvement of protein function. J. Mol. Evol.49: 716-720 (1999).
7. Liebeton K., A. Zonta, K. Schimossek, M. Nardini, D. Lang, B. Dijkstra, M.Reetz, K. Jager. Directed evolution of an enantioselective lipase. Chem. Biol.7: 709-718 (2000).
8. Nardini M., D. Lang, K. Liebeton, K. Jager, B. Dijkstra. Crystal structure ofPseudomonas aeruginosa lipase in the open conformation: the prototype forfamily 1.1 of bacterial lipases. J. Biol. Chem. 275: 31219-25 (2000).
217

9. May O., P. Nguyen, F. Arnold. Inverting enantioselectivity by directed evo-lution of hydantoinase for improved production of L-methionine. NatBiotechnol. 18: 317- 320 (2000).
10. Kirk O., T. Borchert, C. Fuglsang. Industrial enzyme applications. Curr.Opin. Biotechnol. 4: 345-51 (2002).
11. Hart, H., R. Feeney. A specific, highly active malate dehydrogenase byredesign of a lactate dehydrogenase ramework. Science 242: 1541-1544(1988).
12. Mills, D., R. Peterson, S. Spiegelman. An extracellular Darwinian experi-ment with a self-duplicating nucleic acid molecule. Proc. Natl. Acad. Sci. 58:217-24 (1967).
13. Moore, J., H. Jin, O. Kuchner, F. Arnold. Strategies for the in vitro evolutionof protein function: enzyme evolution by random recombination ofimproved sequences. J. Mol. Biol. 272: 336-47 (1997).
14. Stemmer, W.P. Rapid evolution of a protein in vitro by DNA shuffling. Nature370: 389-91 (1994).
15. Michnick, S., F. Arnold. Itching for new strategies in protein engineering.Nature Biotech., 17: 1159-1160, (1999).
218





























