Embed Size (px)
Citation preview

Tlamati Sabiduría, Volumen 7 Número Especial 2 (2016)
4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 21 y 23 de septiembre 2016
Memorias
Caracterización parcial de células mesenquimales de pulpa dental expuestas a
dos medios de cultivo.
Luisa Fernanda Franco Pérez
Facultad Académica de Ciencias Químico Biológicas de la UAGro
Programa de Verano Delfín
Área en la que participa: III Medicina y Salud
Dr. Marco Antonio Meraz Ríos
Profesor- Investigador del Centro de Estudios avanzados
(CINVESTAV)
Karla Mariana Suárez Galván
Innovaciones y Desarrollo en Biotecnología Celular S.A. de C. V.
(INDEBIOC, S.A. de C.V.)
Resumen
Las células troncales de la pulpa dental (DPSC, del inglés Dental Pulp Stem Cells), son
tejido fibroso que contiene células troncales mesenquimatosas / estromales (MSC) derivadas de
la cresta neural craneal embrionaria, que por su origen se han pensado como un nuevo sistema de
terapia celular para enfermedades neurodegenerativas, sin embargo los protocolos de aislamiento
implican el uso de suero fetal bovino (SFB) ocasionando riesgo de rechazo si son usados en
terapia celular, por lo que el objetivo principal del trabajo fue comparar la respuesta de las DPSC
cuando son expuestas en un medio estándar con una baja concentración de SFB y cuando se
exponen en medio neurobasal suplementado con B27 y factores de crecimiento (FC) libres de
SFB, que además dirijan a las células hacia un linaje neural desde pases más tempranos,
evaluando morfología, inmunofenotipo y expresión de genes de pluripotencia. Los resultados
obtenidos mostraron que las DPSC expuestas en medio sin SFB requieren del contacto célula –
célula para poder adherirse y proliferar en placa, además sugieren que la disminución en la
expresión de los marcadores CD90 y CD146 parecen actuar como reguladores de la
diferenciación multilinaje, por lo que el aumento en la expresión del gen crítico para dirigir la
diferenciación a progenitores neurales (Sox2) sugiere que las DPSC expuestas en medio
neurobasal mas B27 y factores de crecimiento están dejando el estado mesenquimal y
dirigiéndose hacia un linaje neural.
Palabras Clave: DPSC, SFB, Progenitores neurales, Sox2, CD90 Y 146.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
Introducción
Las células mesenquimales (MSC) fueron descritas por primera vez por Friedenstein en
1976, como células estromales clonales, adherentes en plástico, que pueden dar origen a líneas
celulares osteoblásticas, adipogénicas y condrogénicas (Friedenstein et al. 1976), además, son
capaces de replicarse casi indefinidamente. De acuerdo a su compromiso a convertirse en un tipo
de célula en particular, las células troncales mesenquimales pueden ser definidas como
"multipotentes" o "pluripotentes", la restricción de potencia se produce cuando las células
troncales no son capaces de diferenciarse en tipos de células más específicas (Evans y, Kaufman
1981; Sch€oler. 2007). La Sociedad Internacional de Terapia Celular en 2006 propuso 3 criterios
para identificar las MSC, que comprenden (1) adherencia al plástico en condiciones normales de
cultivo; (2) la expresión de las moléculas de superficie CD73, CD90, CD105 y ausencia de
CD34, CD45, HLA-DR, CD14 o CD11b, CD79, y (3) una capacidad para la diferenciación de
osteoblastos, adipocitos, y condroblastos in vitro (Dominici at al. 2006). Las MSC derivan de
varios tejidos, como, la médula ósea, sangre de cordón umbilical, el tejido adiposo y la sangre
periférica (Friedenstein et al 1968; Pittenger et al 1999; Bianco et al 2013), aunque en la
actualidad las MSC también se han aislado del folículo dental, ligamento periodontal, la papila
apical y pulpa dental (Hilkens et al. 2015; Honda et al. 2010; Sonoyama et al.2008; Gronthos et
al. 2004; Sch€oler 2007), esta última, es un tejido fibroso que contiene células troncales
mesenquimatosas / estromales (MSC) derivadas de la cresta neural craneal embrionaria (Chai Y
et al. 2000), por lo que contiene diferentes tipos de células, como las células endoteliales,
fibroblastos, osteoblastos / osteoclastos, odontoblastos y células de linaje neural (Nanci, A. 2014)
siendo estas últimas las de mayor interés para el uso de MSC derivados de la pulpa dental como
un nuevo sistema de terapia celular para enfermedades neurodegenerativas y desordenes
neurogeneticos (Urraca N et al.2015). Las células troncales de la pulpa dental (DPSC, del inglés
Dental Pulp Stem Cell), también conocidas como células troncales de la pulpa dental postnatal,
primero se aislaron por Gronthos et al, en el 2002 de los terceros molares y se caracterizaron
como células de autorrenovación y multipotentes que expresan CD29, CD90, CD105, CD146,
CD166, CD271 y marcadores similares a la células troncales neurales por ejemplo, nestina,
proteína ácida glial fibrilar GFAP, del inglés, Glial Fibrilar (Kawashima 2012). Aunque los
protocolos de extracción y aislamiento de las DPSC siguen representando grandes retos,
diferentes grupos de investigación han diseñado y evaluado métodos cada vez más eficaces y
clínicamente seguros (Kerkis y Caplan. 2012), sin embargo los protocolos para el cultivo de
DPSCs también implican el uso de suero fetal bovino (FBS), composición que es desconocida y
que varía entre lotes (Reinhardt et al. 2011), además de condiciones éticas por utilizar un
producto de origen no humano en células que se plantean para terapia celular humana, en este
sentido, existe el riesgo de que los suplementos de crecimiento no humanos pueden estar
contaminados con agentes patógenos tales como virus, priones micoplasmas, u otros agentes,
tóxicos o inmunogénicos ( Eloit, 1999; Wessman y Levings 1999) , por lo que se han propuesto
varios medios de cultivo libres de suero fetal bovino con suplementos para sustituir a los medios
convencionales.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Por lo que el objetivo principal del presente trabajo es evaluar la morfología, inmunofenotipo y
marcadores de pluripotencia al exponer las DPSC ante dos medios de cultivo, uno con una baja
concentración de SFB (SFBlow)y otro que contiene medio neurobasal, B27 y factores de
crecimiento como EGF (factor de crecimiento epidermal) y FGF(factor de crecimiento
fibroblastico), necesarios para la proliferación celular (Hebert Teddi L. et al 2009) y sin SFB
(SFB-), con el fin de obtener desde pases o etapas más tempranas células dirigidas al linaje neural
y que posteriormente faciliten la diferenciación a neuronas maduras funcionales con posible uso
en terapia celular de enfermedades neurodegenerativas.
Materiales y Métodos
Declaración de ética
La pulpa dental fue obtenida de una donante mexicana de 25 años de edad el día 24 de
Octubre del 2012 quien firmo un consentimiento informado que cumple con la declaración de
Helsinki y avalado por la comisión de ética de la empresa Innovaciones y Desarrollo en
Biotecnología Celular, INDEBIOC S.A.de C.V.
Preparación de medios de cultivo para aislamiento de células mesenquimales
Medio neurobasal con B27 y factores de crecimiento (SFB-)
Se prepararon 50 ml de medio neuroabsal + B27 1X y factores de crecimiento en un tubo
falcón graduado estéril, adicionando 49 ml de neurobasal médium (GIBCO/Invitrogen), 1 ml de
B27 supplement (GIBCO), 500 ul de Anti-Anti (antimicótico-antibiotico), FGF (Factor de
Crecimiento Fibroblastico) y EGF (Factor de Crecimiento Epidermal) ambos a [20 ng/ml]
concentración final.
Medio estándar (SFBlow)
El medio estándar es preparado con el protocolo de la empresa Desarrollo en
Biotecnología Celular, INDEBIOC.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
Aislamiento y cultivo de células mesénquimales de la pulpa dental
Las células mesenquimales de pulpa dental fueron extraídas en condiciones de esterilidad
en campana de flujo laminar, los molares fueron expuestos en solución desinfectante para
posteriormente fragmentar el molar con pinzas estériles, la pulpa dental fue expuesta a
degradación por colagenasa por 20 minutos a 37° C, la inactivación de la colagenasa fue con
medio DMEM (GIBCO) por dilución, la muestra se centrifuga a 1200 rpm por 7 minutos, una
vez obtenido el pelet, las células son expuestas a dos diferentes medios de cultivo, uno estándar
con SFBlow y otro neurobasal mas factores de crecimiento SFB_, en cajas p60 para cultivo
primario, se monitoreo el crecimiento de las células en ambos medios por una semana, sin
embargo las células mesenquimales de la pulpa dental (DPSC) en este cultivo primario no
proliferaron y las cajas fueron desechadas, por lo que se procedió a descongelar un vial de DPSC
de un donante mexicano de sexo femenino de 25 años de edad de pase 2, el vial se descongelo a
temperatura ambiente, posteriormente se realizó una dilución 1:20 con azul de tripano para
conteo celular, y se cultivaron 2X104 células por pozo en una caja de 6 pozos, y se evaluó la
morfología por 7 días, las DPSC en medio neurobasal mas factores SFB_ no lograron adherirse a
la placa, por lo que después de los 7 días se procedió a sembrar 2x105 células por pozo en caja de
6 pozos, y se evaluó la morfología celular, las DPSC sembradas fueron incubadas a 37° C con
5% de CO2, el medio de cultivo fue cambiado cada tercer día.
Morfología celular
La morfología celular fue observada en microscopio Leica de contraste de fases, a los días
3, 5 y 7 después de la siembra en ambos medios de cultivo, las fotos fueron tomadas a una escala
de 200 micras al objetivo 10X después del cambio de medio.
Citometria de flujo
Las DPSC fueron fijadas con BD Cytofix™ Fixation Buffer y siguiendo las indicaciones
del fabricante fueron marcadas con los anticuerpos anti-humano acoplados a fluoroforos CD105-
FITC, CD90-FITC, CD73-PE, CD146-PE , CD271-APC, como marcadores positivos de DPSC, y
como marcadores negativos Anti-HLA-DR-PE y CD31-FITC, todos de la casa comercial
Miltenyi Biotec. Para la muestra sin marcaje se utilizaron 2X105 células para adquirir la
población de interés. Las muestras fueron procesadas en el clitómetro de flujo departamental
FORTESA y analizado con el software Cyflogic versión 2.1.2.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Marcador (CD) Procesos biológicos en los que está involucrado
CD90 Molécula de adhesión celular involucrada en interacción célula –célula y célula -
matriz extracelular.
CD73 Molécula de adhesión celular.
CD105 Componente de factor de crecimiento transformante beta (TGFb), involucrado en
el mantenimiento del citoesqueleto, morfología y migración celular.
CD271 Receptor de baja afinidad para neurotrofinas, involucrada en la supervivencia de
neuronas maduras.
CD146 Molécula de adhesión de melanoma presente en todas las células mesenquimales
CD31 Molécula de adhesión endotelial plaquetaria, involucrado en la angiogénesis y la
progresión de tumores
HLA-DR Complejo principal de histocompatibilidad, involucrada en la supresión de la
respuesta inmune por linfocitos T y potencia la respuesta por linfocitos B, y en el
rechazo por trasplantes de órganos y tejidos.
Cultivo de línea celular de teratocarcinoma pluripotente.
Se cultivó la línea celular NCCIT (teratocarcinoma pluripotente) en medio DEMEM-F12
con 10% de SFB para su posterior extracción de RNA como control positivo para determinar la
expresión de genes de pluripotencia.
Extracción de RNA de las DPSC
El RNA fue extraído de las DPSC y la línea celular NCCIT por el método de
TRIZOL®LS Reagent para células en monocapa de la empresa comercial Life Technologies Cat.
No. 10296
Cuantificación de RNA
El RNA extraído fue cuantificado en un espectrofotómetro llamado NanoDrop de Thermo
scientific, la cuantificación de RNA fue expresada en ng/ul.
RNA ng/ul Relación 260/280
DPSC sin SFB 29.5 2.22
DPSC con SFB 899.9 2.4
NCCIT 448.6 2.45
Tabla 1. CDs o cúmulos de diferenciación positivos y negativos utilizados para la caracterización del inmunofenotipo de
células mesenquimales de la pulpa dental medidos en los dos diferentes medios de cultivo.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
DNAsa
El protocolo de DNAsa para la degradación del DNA que pudiese seguir presente en la
muestra se realizó de acuerdo al kit DNase I, Amplification Grade de la casa comercial
Invitrogen, Código de catálogo: 18068015, que requiere de 1 ug de RNA para el tratamiento con
DNAsa para el posterior uso del RNA en la transcripatasa reversa-PCR. El RNA tratado con la
DNAsa fue cuantificado por segunda vez y se realizaron los cálculos para llevarlo a una
concentración de 10ng/ul para el uso en la RT-PCR.
Transcriptasa reversa-PCR
La RT-PCR o transcriptasa reversa fue realizada mediante el kit para RT-PCR QuantiFast
Probe de Qiagen, el kit realiza la síntesis del cDNA y posteriormente la amplificación del
fragmento de interés mediante PCR convencional, se utilizaron Oligos para genes de
pluripotencia, Oct 4 (sentido 5’ CGTGAAGCTGGAGAAGGAGAAGCTG 3´ y antisentido 5´
AAGGGCCGCAGCTTACACATGTTC 3´), Sox 2 (sentido 5´ ACACCAATCCCATCCACACT
3´y antisentido 5´ GCAAACTTCCTGCAAAGCTC 3´), Nanog (sentido 5´
TGCAAATGTCTTCTGCTGAGAT 3´y antisentido 5´ GTTCAGGATGTTGGAGAGTTC 3´) y
DNAMT3 (sentido 5´ TTGAATATGAAGCCCCCAAG 3´y antisentido 5´
GGTTCCAACAGCAATGGACT 3´) el control de carga fue Actina, la RT-PCR fue corrida en
un termociclador con un programa de temperaturas en gradiente a 30 ciclos, y reveleda en un
fotodocumentador de luz UV con bromuro de etidio.
ID GEN SECUENCIA PB TM CICLOS
NM_002701.5 Oct 4 F:CGTGAAGCTGGAGAAGGAGAAGCTG
R: AAGGGCCGCAGCTTACACATGTTC
245 68° 30
NM_003106.3 Sox 2 F: ACACCAATCCCATCCACACT
R: GCAAACTTCCTGCAAAGCTC
224 60° 30
XM_011520850.1 Nanog F: TGCAAATGTCTTCTGCTGAGAT
R: GTTCAGGATGTTGGAGAGTTC
286 58° 30
XR_936512.2 DNMT3 F: TTGAATATGAAGCCCCCAAG
R: GGTTCCAACAGCAATGGACT
179 58° 30
Resultados
La morfología celular de las DPSC al transcurso de los 7 días expuestos en medio SFB_ se
mostró diferente en comparación con el medio estándar, ya que en estas últimas al sembrarse
2x104 células por pozo en placas de pozos de 6 se observó una mayor capacidad de adherencia y
proliferación con morfología alargada fibroblastoide (Fig.1A), apariencia que coincide con la
reportada en la literatura para células mesenquimales, por otro lado las DPSC expuestas en medio
Tabla 2. Oligos utilizados para medir la expresión de genes de pluripotencia, con sus respectivas temperaturas, ciclos y
pares de bases donde amplifican

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
SFB_ (neurobasal con B27 y factores de crecimiento) mostraron dificultad para adherirse en placa
de cultivo cuando se sembraron a una densidad de 2x104, por lo que se realizó una resiembra en
placa de 6 pozos a una densidad de 2x105 por pozo, demostrando que a una densidad mayor sí
logran adherirse parcialmente a la placa y mostrando morfología esférica y menos fibroblastoide
con una gran cantidad de células viables despegadas (Fig. 1B), morfología distinta a la reportada
en la literatura para mesenquimales pero similar a la reportada para células progenitoras de linaje
neural.
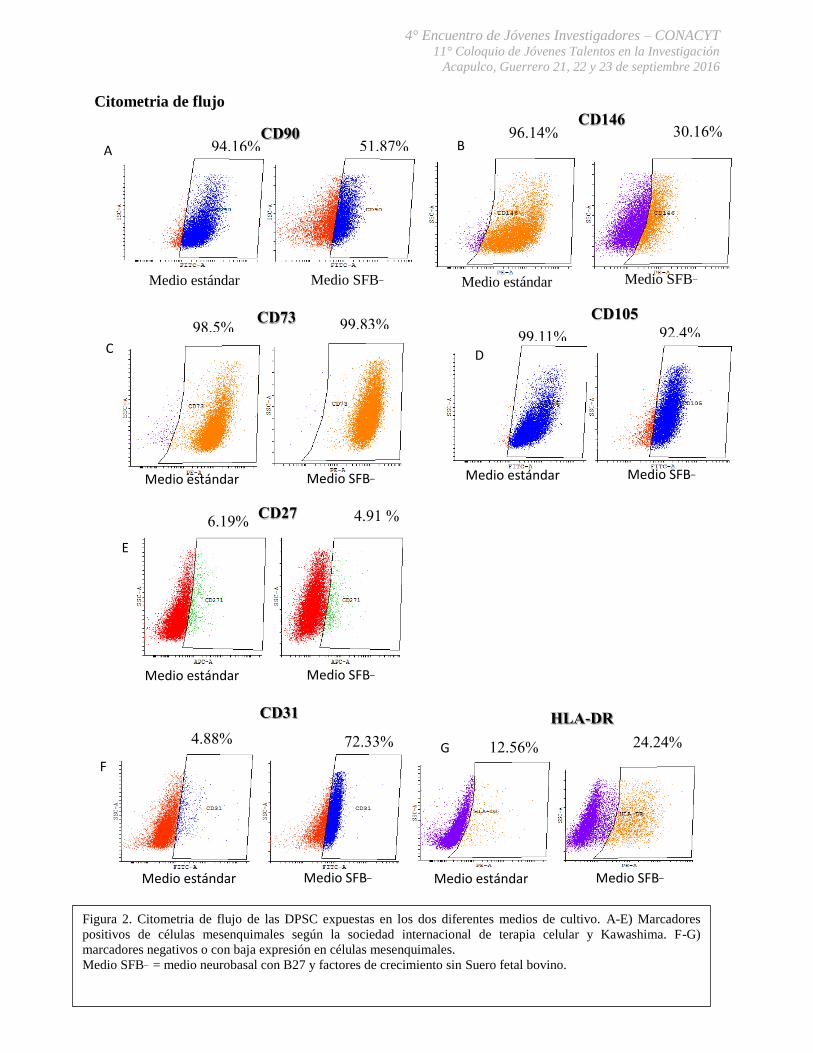
El inmunofenotipo de las DPSC se observó claramente alterado ya que las DPSC en
medio SFB_ mostraron una menor expresión de los marcadores CD90 y de CD146 (ambas
moléculas involucradas con la adhesión celular) en comparación con las DPSC en medio
estándar (Figura. 2A y 2B), por otra parte las DPSC SFB_ a su vez mostraron un aumento en la
expresión de los marcadores HLA-DR o complejo principal de histocompatibilidad y CD31 en
comparación con las DPSC en medio estándar (Fig. 2F y 2G), estos últimos datos no han sido
reportados en la literatura en células mesenquimales por lo que deberán ser discutidos más
adelante, estos resultados fueron complementados con RT-PCR para la expresión de genes de
pluripotencia, donde se midió la expresión de los genes Oct4, Sox 2, DNMT3 Y Nanog (Figura.
3), donde se mostró una mayor expresión del gen Sox 2 (marcador de células progenitoras
neurales) en las DPSC SFB_ que en aquellas cultivadas en medio estándar.
Morfología celular de las DPCS
A)
d03 d05 d07
B)
Figura 1. Morfología de las DPSC cultivadas en dos medios de cultivo. A) Morfología fibroblastoide y en adhesión al
plato de cultivo de las DPSC en medio estándar observadas a los 3, 5 y 7 días. B) Morfología circular y pequeña de
las DPSC cultivadas en medio neurobasal + B27 y factores de crecimiento SFB_ al día 3, 5 y 7. Escala=200um.
C/
SF
B
S/
SF
B
4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
Citometria de flujo
94.16% 51.87% CD90
CD146 96.14% 30.16%
Medio estándar Medio estándar Medio SFB_
99.11% 92.4%
CD105 CD73 98.5% 99.83%
CD27
16.19% 4.91 %
4.88% 72.33%
CD31 HLA-DR
12.56% 24.24%
A B
C D
Figura 2. Citometria de flujo de las DPSC expuestas en los dos diferentes medios de cultivo. A-E) Marcadores
positivos de células mesenquimales según la sociedad internacional de terapia celular y Kawashima. F-G)
marcadores negativos o con baja expresión en células mesenquimales.
Medio SFB_ = medio neurobasal con B27 y factores de crecimiento sin Suero fetal bovino.
E
F
G
Medio SFB_
Medio estándar Medio SFB_ Medio estándar Medio SFB_
Medio estándar Medio SFB_
Medio estándar Medio SFB_ Medio estándar Medio SFB_

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
RT-PCR para genes de pluripotencia
Discusión
Las células mesenquimales varían en cuanto a su expresión de marcadores o cluster de
membrana (CDs) de acuerdo a su origen (Moraes D. et al. 2016), el estudio de la expresión de los
marcadores de células mesenquimales ha sido muy extenso e implica el uso de interferentes para
determinar el nivel de participación que tienen en la conservación de su troncalidad, la expresión
de algunos marcadores se observa disminuido cuando las células mesenquimales son expuestas
en medio de cultivo libre se suero fetal bovino (Peter Mark et al. 2013), sin embargo de acuerdo a
los resultados obtenidos solo los marcadores CD90 y CD146 presentan cambios considerables en
la expresión, en medio libre de suero fetal bovino en comparación con el medio estándar, por lo
que esto podría sugerir que la reducción en la expresión de algunos marcadores no solo varía
dependiendo del origen de las células mesenquimales si no también del medio al que son
expuestas, recientemente la expresión del marcador CD90 o Thy-1 la cual es una glicoproteína de
la superficie celular que juega un papel en las interacciones célula-célula y célula - matriz
Oct4
Sox2
Nanog
DNMT3
ACTB
NCCI
T
SFB S/ SFB
Figura. 3. RT-PCR de genes de pluripotencia. Se puede observar la expresión de todos los genes de pluripotencia en
la línea celular teratocarcinoma pluripotente (NCCIT) utilizada como control positivo, en el siguiente carril se puede
observar la expresión relativa de los genes del circuito de pluripotencia en las DPSC con medio estándar, y en el
carril siguiente un comportamiento similar de las DPSC en medio SFB_, sin embargo las células expuestas en este
medio tienen una mayor expresión del marcador de pluripotencia Sox2, gen presente en las células progenitoras
neurales.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
extracelular, así como la motilidad celular (Rege TA & Hagood JS. 2006) se ha encontrado
asociada con una mayor potencialidad de diferenciación hacia otros linajes celulares sin afectar la
morfología de las células expuestas en medio con suero fetal bovino (Moraes D. et al. 2016),
aunado a esto, también existen reportes donde indican que la reducción en la expresión del
marcador CD146 o M-CAM cuya función es mediar las interacciones célula-célula, la migración
en las células endoteliales, y la remodelación del citoesqueleto (Ouhtit A et al. 2009) está
directamente relacionada con la capacidad de diferenciación multilinaje en células
mesenquimales (Paduano F. et al. 2016), estos datos sugieren que las DPSC en medio Neurobasal
sin suero fetal bovino podría facilitar la diferenciación hacia células progenitoras neurales y que
las células mesenquimales están dejando el estado mesenquimal siendo más susceptibles a
diferenciarse probablemente hacia linaje progenitor neural, esto puede verse reflejado en la
disminución de los marcadores CD90 y CD146, por otro lado la disminución en la expresión de
estos marcadores individualmente no se han reportado como moléculas que afecten la morfología
de las DPSC (Moraes D. et al. 2016; Paduano F. et al. 2016; Masoud Maleki et al. 2014) por lo
que los datos obtenidos podrían sugerir que los cambios morfológicos observados en las DPSC se
deben únicamente al estímulo del medio neurobasal + B27 y factores de crecimiento como
consecuencia de su inducción para diferenciarse hacia un linaje neural, sin embargo la gran
cantidad de células despegadas con reducida capacidad de adherencia a bajas densidad en placa
de plástico sugiere que las DPSC en este medio requieren de la interacción célula – célula para
lograr su proliferación, adherencia y supervivencia.
Por otra parte, se ha reportado que los niveles del Antígeno principal de
histocompatibilidad (HLA-DR) tienden a incrementar ligeramente su expresión y su capacidad
parcial de estimular una respuesta inmune en células mesenquimales cuando estas son
diferenciadas hacia otro linaje celular (Le Blanc et al. 2003), lo cual podría apoyar los datos
sugeridos por los marcadores CD90 y CD146, es decir la salida del estado mesenquimal hacia
otro linaje celular. Aunque no existen reportes acerca del aumento en la expresión del marcador
CD31 en células mesenquimales, este marcador se encuentra asociado a células altamente
angiogénicas y vasculogénicas (Hyongbum Kim et al. 2010), lo cual podría destinar estas células
hacia un nuevo modelo de terapia celular (Sung-Whan et al. 2011).
Además la expresión de genes del circuito de pluripotencia obtenidos mediante RT-PCR,
muestran la disminución en la expresión de dos de estos genes Oct4 y Nanog pero una remarcada
expresión del gen Sox 2, dicho gen regula la diferenciación a linaje neuroectodermal mientras
que Oct4 y Nanog promueven la diferenciación a linaje mesoendodermico, (Thomson et al.
2011), además estudios recientes reportan a Sox2 como un factor de la transcripción crítico para
dirigir la diferenciación a progenitores neurales y para el mantenimiento de las propiedades de las
células madre progenitoras neurales (Shuchen Zhang & Wei Cui. 2014), dichos datos sugieren
que las DPSC expuestas en medio de cultivo sin SFB no solo están perdiendo pluripotencialidad,
la presencia de Sox 2 y la perdida de la expresión de Oct 4 sugiere que las DPSC también están
iniciando su diferenciación hacia un linaje celular neural.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
Conclusiones
Las DPSC expuestas en medio de cultivo sin SFB con factores de crecimiento son menos
adherentes en placa de plástico, y se requiere cultivar una cantidad mayor de DPSC, es decir
requieren del contacto célula – célula para que puedan proliferar, por otra parte el
inmunofenotipo de las DPSC cambia al ser cultivadas en este medio, siendo los marcadores
CD90 y CD146 los más significativos, y estos parecen actuar como reguladores de potencial
multilinaje, los marcadores CD31 y HLA-DR reportados como negativos para células
mesenquimales en la literatura, en las DPSC expuestas a medio sin SFB muestran una expresión
positiva, el cambio en su expresión parece estar influenciada por el probable cambio de estado
mesenquimal hacia otro linaje celular, por otra parte la presencia del gen Sox 2 indica que el
estímulo del medio de cultivo neurobasal con B27 y factores de crecimiento (SFB_) está
favoreciendo la diferenciación de las células mesenquimales hacia un linaje progenitor neural.
Perspectivas
Para corroborar que los marcadores CD90 y CD146 están actuando como marcadores
reguladores de la diferenciación multilinaje, en este caso por el estímulo del medio neurobasal
con B27 y factores de crecimiento, es necesario medir la su expresión aunado a la expresión de
genes de linaje neural mediante RT-PCR o medir la expresión de los mismos a nivel proteína por
Western Blot, la acción permisiva de diferenciación multilinaje de estos marcadores también
podría determinarse iniciando no solo la diferenciación a linaje neural, también hacia otros linajes
celulares como osteoblastos, adipocitos y condrocitos, por otra parte es necesario determinar si
los marcadores CD90 y CD146 (ambas moléculas de adhesión celular) son capaces de afectar la
morfología celular cuando la disminución en su expresión se da de manera conjunta y no
individual como se encuentra reportado en la literatura, por lo que para comprobarlo es necesario
exponer DPSC en las mismas condiciones (ambos medios de cultivo) interfiriendo la expresión
de los marcadores CD90 y CD146 y observar que ocurre con las morfología de las DPSC.
Es necesario repetir el estudio con un mayor número de muestras, para que los resultados
puedan ser considerados como significativamente estadísticos.
Agradecimientos
Al doctor Marco Antonio Meraz Ríos, Profesor- Investigador del Centro de Estudios
avanzados (CINVESTAV) jefe del laboratorio, por permitirme realizar estancia en el
departamento de Biomedicina molecular, por otro lado agradezco a la tesista de Doctorado Karla
Mariana Suárez Galván quien fue mi asesora directa a cargo y estuvo pendiente de la realización
del proyecto en todo momento, a todas las tesistas de doctorado en el laboratorio, así como los
técnicos que laboran en el mismo, por facilitar mi estancia y apoyarme con las dudas que surgían
en el transcurso del proyecto.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
Referencias
1. Bianco P, Cao X, Frenette PS, Mao JJ, Robey PG, Simmons PJ, Wang CY. 2013. The
meaning, the sense and the significance: Translating the science of mesenchymal stem
cell into medicine. Nat Med 19:35–42.
2. Chai Y, Jiang X, Ito Y. 2000. et al. Fate of the mammalian cranial neural crest during
tooth and mandibular morphogenesis. Development 2000;127:1671–1679.
3. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R,
Keating A, Prockop D, Horwitz E. 2006. Minimal criteria for defining multipotent
mesenchymal stromal cells: the International Society
4. Evans MJ, Kaufman MH. 1981. Establishment in culture of pluripotential cells from
mouse embryos. Nature 292:154–156.
5. Friedenstein AJ, Gorskaja U, Kalugina NN. 1976. Fibroblast precursors in normal and
irradiated mouse hematopoietic organs. Exp Hematol 4: 267-274
6. Friedenstein AJ, Petrakova KV, Kurolesovam AI, Frolova GP. 1968. Heterotopic of bone
marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues.
Transplantation 6:230–247.
7. Hilkens P, Meschi N, Lambrechts P, Bronckaers A, Lambrichts I. 2015. Dental stem cells
in pulp regeneration: Near future or long road ahead? Stem Cells Dev 24:1610–1622.
8. Honda MJ, Imaizumi M, Tsuchiya S et al. 2010. Dental follicle stem cells and tissue
engineering. 2010. J Oral Sci;52:541–552.
9. Hyongbum Kim, Hyun-Jai Cho, Sung-Whan Kim, Bianling Liu, Yong Jin Choi, JiYoon
Lee, Young-Doug Sohn, Min-Young Lee, Mackenzie A. Houge, Young-sup Yoon. 2010.
CD31+ Cells Represent Highly Angiogenic and Vasculogenic Cells in Bone Marrow
Circulation Research.Vol. 30. 107: 602-614.
10. Kerkis and A. I. Caplan. 2012. “Stem cells in dental pulp of deciduous teeth,”Tissue
Engineering—PartB: Reviews,vol.18,no.2, pp.129–138.
11. J. Reinhardt, A. St¨uhler, and J. Blumel, 2011. “Safety of bovine sera for production of
mesenchymal stem cells for therapeutic use,” Human Gene Therapy, vol. 22, no.6,p.775.
12. Katarina Le Blanc, Charlotte Tammik, Kerstin Rosendahl, Eva Zetterberg & Olle
Ringde. 2003. HLA expression and immunologic properties of differentiated and
undifferentiated mesenchymal stem cells. Elsevier. Experimental Hematology. Vol. 31
890–896.
13. Kawashima N. 2012. Characterization of dental pulp stem cells: A new horizon for tissue
regeneration? Arch Oral Biol: 2012;57:1439– 1458.
14. M. Eloit, 1999 “Risks of virus transmission associated with animal sera or substitutes and
methods of control,” Developments in Biological Standardization, vol.99,pp.9–16.
15. Masoud Maleki, Farideh Ghanbarvand, Mohammad behvarz, Mehri Ejtemaei, y Elham
Ghadirkhomi. 2014. Comparison of Mesenchymal Stem Cell Markers in Multiple Human
Adult Stem Cells. Int J Stem Cells. 7(2): 118–126.

Tlamati Sabiduría Volumen 7 Número Especial 2 (2016)
16. Moraes D, Sibov T, Pavon L, Alvim P, Bonadio R, Da Silva j, et al. 2016. A reduction in
CD90 (THY-1) expression result in increased differentiation of mesenchymal stromal
cells. Stem cell Research & therapy. 7:97.
17. Nanci, A. 2014. Ten Cate’s oral histology: development, structure, and function. Elsevier
Health Sciences. 2014.
18. Ouhtit A, Gaur RL, Abd Elmageed ZY, Fernando A, R Thouta, Trappey AK, Abdraboh
ME, El-Sayyad HI, Rao P, Raj MG. 2009. Hacia la comprensión del modo de acción de
la multifacética receptor de adhesión de células CD146. Biochim Biophys; 1795 :. 130-
136
19. Paduano F, Marrelli M, Palmieri, Tatullo M. 2016. CD146 Expression Influences
Periapical Cyst Mesenchymal Stem Cell Properties.CD146 Expresión influye periapical
Quiste propiedades de células madre mesenquimales. Stem Cell Reviews and Reports pp
1-–12,
20. Peter Mark, Mandy Kleinsorge, Ralf Gaebel, Cornelia A. Lux, Anita Toelk, Erik
Pittermann, Robert David, Gustav Steinhoff, & Nan Ma. 2013. Human Mesenchymal
Stem Cells Display Reduced Expression of CD105 after Culture in Serum-Free Medium.
Stem Cells International. Volume 2013, Article ID 698076.
21. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA,
Simonetti DW, Craig S, Marshak DR. 1999. Multilineage potential of adult human
mesenchymal stem cells. Science 284:143–147.
22. Rege TA, Hagood JS. 2006. Thy-1 as a regulator of cell-cell and cell-matrix interactions
in axon regeneration, apoptosis, adhesion, migration, cancer, and fibrosis. FASEB J
;20:1045–1054.
23. S. Gronthos, J. Brahim, W. Li et al., 2002. “Stem cell properties of human dental pulp
stem cells,”Journal of Dental Research, vol. 81,no.8,pp.531–535.
24. S. J. Wessman &and R. L. Levings, 1999. “Benefits and risks due to animal serum used in
cell culture production, ” Developments in Biological Standardization, vol. 99,pp.3–
8.,1999.
25. Sch€oler HR. 2007. The potential of stem cells: An inventory. In: Knoepffler N,
Schipanski D, Sorgner SL, editors.Humanbiotechnology as social challenge. Farnham,
Surrey, UK: Ashgate Publishing Ltd. p 28
26. Seo BM, Miura M, Gronthos S et al. 2004.Investigation of multipotent postnatal stem
cells from human periodontal ligament. Lancet;364:149–155
27. Shuchen Zhang & Wei Cui. 2014. Sox2, a key factor in the regulation of pluripotency and
neural differentiation. World J Stem Cells. Vol. 6. N°3: 305–311.
28. Sonoyama W, Liu Y, Yamaza T et al. 2008 Characterization of the apical papilla and its
residing stem cells from human immature permanent teeth: A pilot study. J Endod;34:166
171.
29. Sung-Whan Kim, Hyongbum Kim & Young-sup Yoon. 2012. Advances in bone marrow
derived cell therapy: CD31-expressing cells as next generation cardiovascular cell therapy
Regen Med. 2011 May; 6(3): 335–349.

4° Encuentro de Jóvenes Investigadores – CONACYT 11° Coloquio de Jóvenes Talentos en la Investigación
Acapulco, Guerrero 21, 22 y 23 de septiembre 2016
30. Thomson M, Liu SJ, Zou LN, Smith Z, Meissner A & Ramanathan S. 2011. Pluripotency
factors in embryonic stem cells regulate differentiation into germ layers. Cell;145:875–
889.
31. Urraca N, Memon R, El-Iyashi I, Goorha S, Valdez C. et al.2015. Characterization of
neurons from immortalized dental pulp stem cells for the study of neurogenetic disorders.
Elselvier. 722-730.



![Proyecto 09 W. Huanca.ppt [Modo de compatibilidad] · Extracción de RNA total Método TRIzol ( Invitrogen ) RNA total Cuantificación](https://img.pdfslide.es/doc/110x75/5babb31809d3f211798c741d/proyecto-09-w-modo-de-compatibilidad-extraccion-de-rna-total-metodo-trizol.jpg)

























