Embed Size (px)
Citation preview

Peptidasen aktibitatearen aldaketak heste lodiko neoplasietan
Jakintza-arloa: Medikuntza
Egilea: ITXARO PEREZ URZELAI
Urtea: 2011
Zuzendariak: BEGOÑA SANZ ETXEBARRIA, GORKA LARRINAGA ENBEITA
Unibertsitatea: UPV-EHU
ISBN: 978-84-8438-458-8

Hitzaurrea
Itxaro Perez Urzelai naiz (Beasain,1981), biokimikan lizentziatua duela urte batzuk jada (2004an) eta 2011ko ekainaren 17tik EHUko doktorea.
Medikuntza eta Odontologia fakultateko fisiologia sailean egin nuen Tesi hau 2006-2011 urteetan zehar. 2011ko Ekainaren 17a egun adierazgarria da nire bizitzan orduz geroztik. Duela egun batzuk dagoeneko urte bat pasa dela konturatzeak atzera begira jarri ninduen. Ia 5 urte luze hauetan ikertu eta ikasitako guztiak bizia aldatzen duela ezin da ukatu.
Nire ikerketa taldea sistema peptidikoak ikertzen ari zen ni bertara sartu nintzenean. Lerro ezberdinak ditugu egun, orduan bezala, eta ni minbizi arloan sartu nintzen. Hasiera batean giltzurruneko minbizian egin nituen lehenengo urratsak eta emaitza adierazgarriak ateratzen zirela ikusita, koloneko/heste lodiko neoplasietan ikertzen hasi nintzen, jada tesian pentsatuz.
Errenina-angiotentsina sistemako (RAS) peptidasak heste lodiko neoplasietan ikertzearen ekarpena, giltzurrunekoek ez bezala, lesioaren aurrekaria den adenoma baten ikerketa egiteko aukera-edo eskaintzen duela da, hau da, minbiziaren bilakaera ikusi litekeela nolabait. Beraz, paziente baten ehun osasuntsua, adenoma eta adenokartzinoma ikertzeko aukera ematen du heste lodiak eta bertan gertatzen diren prozesu ezberdinen arteko konparaketa egin liteke. Heste lodiko minbizia, intzidentzia altuenetarikoa duen minbizia izanik, eta fase berantiarrera arte sintoma nabaririk ez agertzean, adenoma ikertzeak abantaila handia ematen du minbiziaren kartzinogenesia ulertzeko. Bestalde, minbiziaren estadio eta gradu desberdina duten pazienteen adenokartzinomak beraien artean konparatuz ere, minbiziaren agresibitateari buruzko informazioa ere jaso da tesi honetan.
Peptidasa hauen funtzioa giza ehunetan ikertzeaz gain pazienteen odol-laginetan, zehazki plasmetan ere ikertu dira eta bertan lortutako ondorioak interesgarriak dira oso. Plasmak ikertzeak gerora klinikara begira, aurrerapenak ekar litzakeelako gaixotasunaren pronostikoa, diagnosia eta gaixoaren jarraipena egiterako orduan.
Ikerketa honetan lortutako ondorioek argi pixka bat eskaintzen dute minbiziaren sorreran eta bilakaeran gertatzen ari diren prozesuak ulertzeko unean. Konparaketak egin diren laginetan, entzimen aktibitateetan aldaketa adierazgarriak aurkitu dira baina, aipatu behar da, egin den ikerketa oinarrizko ikerketa dela. Ikerketa gehiago eta sakonagoak egin behar dira oraindik.
Ikerketa gehiago eta sakonagoak egitea dira gure ondorengo pausuak. Lehenik eta behin, biziraupenak aztertzen hasi gara (eta ondorio interesgarriak lortu ditugu) peptidasa hauen eragina gaixotasunaren pronostikoan ikertzeko asmoz. Bestalde, sistema peptidikoa modu orokor batean ikertzea da hurrengo helburua. Entzima hauen aktibitatea neurtu dugu oraingoz eta RAS sistema peptidikoa osatzen duten gainontzeko partaideak (hartzaileak, peptidoak…) ikertu nahi ditugu. Horretarako, beraien adierazpena eta funtzioa teknika

ezberdinak erabiliaz minbiziaren bilakaeran zehar aldatzen diren ikertu nahi dugu, eta hala bada, zergatik edo zein prozesuren bidez aldatzen diren jakin.
Itxaro Perez Urzelai, 2012

DoktoreDoktoreDoktoreDoktore----TesiaTesiaTesiaTesia
Peptidasen aktibitatearen aldaketak Peptidasen aktibitatearen aldaketak Peptidasen aktibitatearen aldaketak Peptidasen aktibitatearen aldaketak
heste lheste lheste lheste lodikoodikoodikoodiko n n n neoplasietaneoplasietaneoplasietaneoplasietan
Itxaro Perez UrzelaiItxaro Perez UrzelaiItxaro Perez UrzelaiItxaro Perez Urzelai
2011201120112011ko Ekainako Ekainako Ekainako Ekaina
Zuzendariak:
Gorka Larrinaga Enbeita
Begoña Sanz Echevarría
Departamento de FisiologíaDepartamento de FisiologíaDepartamento de FisiologíaDepartamento de Fisiología
Facultad de Medicina y OdontologíaFacultad de Medicina y OdontologíaFacultad de Medicina y OdontologíaFacultad de Medicina y Odontología
FisFisFisFisiologia Sailaiologia Sailaiologia Sailaiologia Saila
Medikuntza eta Odontologia FakultateaMedikuntza eta Odontologia FakultateaMedikuntza eta Odontologia FakultateaMedikuntza eta Odontologia Fakultatea


Eskerrak
Etxe handi honetan denok gara familia bat. Etxearen administrariak, bizilagunak, Etxe handi honetan denok gara familia bat. Etxearen administrariak, bizilagunak, Etxe handi honetan denok gara familia bat. Etxearen administrariak, bizilagunak, Etxe handi honetan denok gara familia bat. Etxearen administrariak, bizilagunak,
lagunak eta etsaiak. Familiaren barnean, gurasoak, anaialagunak eta etsaiak. Familiaren barnean, gurasoak, anaialagunak eta etsaiak. Familiaren barnean, gurasoak, anaialagunak eta etsaiak. Familiaren barnean, gurasoak, anaia----arrebak, izebaarrebak, izebaarrebak, izebaarrebak, izeba----osabak, osabak, osabak, osabak,
lehengusulehengusulehengusulehengusu----lehengusinak eta ulehengusinak eta ulehengusinak eta ulehengusinak eta urruneko lehengusurruneko lehengusurruneko lehengusurruneko lehengusu----lehengusinak. Denon artean egin lehengusinak. Denon artean egin lehengusinak. Denon artean egin lehengusinak. Denon artean egin
dezagun gustura bizitzeko ingurunea.dezagun gustura bizitzeko ingurunea.dezagun gustura bizitzeko ingurunea.dezagun gustura bizitzeko ingurunea.
Esker aunitz guztiEsker aunitz guztiEsker aunitz guztiEsker aunitz guzti----guztioi.guztioi.guztioi.guztioi.
Lehenik eta behin, eskerrak eman nahi nizkieke nire zuzendariei, lan hau ondo atera Lehenik eta behin, eskerrak eman nahi nizkieke nire zuzendariei, lan hau ondo atera Lehenik eta behin, eskerrak eman nahi nizkieke nire zuzendariei, lan hau ondo atera Lehenik eta behin, eskerrak eman nahi nizkieke nire zuzendariei, lan hau ondo atera
dadin egin duten esfortzuagatik. dadin egin duten esfortzuagatik. dadin egin duten esfortzuagatik. dadin egin duten esfortzuagatik.
Bego, eskerrik asko egBego, eskerrik asko egBego, eskerrik asko egBego, eskerrik asko egindako lan izugarriagatik. indako lan izugarriagatik. indako lan izugarriagatik. indako lan izugarriagatik.
Gorka, ez dezazula inoiz galdu ongi ateratako esperimentuetako ilusio hori eta hark Gorka, ez dezazula inoiz galdu ongi ateratako esperimentuetako ilusio hori eta hark Gorka, ez dezazula inoiz galdu ongi ateratako esperimentuetako ilusio hori eta hark Gorka, ez dezazula inoiz galdu ongi ateratako esperimentuetako ilusio hori eta hark
zioen bezala, zioen bezala, zioen bezala, zioen bezala, “Que me perdone el que se sienta desgraciado. Esta es una canción “Que me perdone el que se sienta desgraciado. Esta es una canción “Que me perdone el que se sienta desgraciado. Esta es una canción “Que me perdone el que se sienta desgraciado. Esta es una canción
en honor de la alegría de vivir”. en honor de la alegría de vivir”. en honor de la alegría de vivir”. en honor de la alegría de vivir”.
Gorka eta Adolfo, Adolfo eta Gorka eta Adolfo, Adolfo eta Gorka eta Adolfo, Adolfo eta Gorka eta Adolfo, Adolfo eta Gorka: Gorka: Gorka: Gorka: YinYinYinYin edo edo edo edo YangYangYangYang? Nor da Nor? Inoiz ez dut argi ? Nor da Nor? Inoiz ez dut argi ? Nor da Nor? Inoiz ez dut argi ? Nor da Nor? Inoiz ez dut argi
izan... izan... izan... izan... AAAAdolfo, egunerodolfo, egunerodolfo, egunerodolfo, egunero----egunero gogoratzen naiz zurekin, ezin jakin zenbat botatzen egunero gogoratzen naiz zurekin, ezin jakin zenbat botatzen egunero gogoratzen naiz zurekin, ezin jakin zenbat botatzen egunero gogoratzen naiz zurekin, ezin jakin zenbat botatzen
zaitudan faltan. Berdin jarraitzen dugu, batak A, besteak B eta guk beti C!!!!zaitudan faltan. Berdin jarraitzen dugu, batak A, besteak B eta guk beti C!!!!zaitudan faltan. Berdin jarraitzen dugu, batak A, besteak B eta guk beti C!!!!zaitudan faltan. Berdin jarraitzen dugu, batak A, besteak B eta guk beti C!!!!
Lorena, esker mila, irakatsi didazun guztiLorena, esker mila, irakatsi didazun guztiLorena, esker mila, irakatsi didazun guztiLorena, esker mila, irakatsi didazun guztiagatik, hasierako momentuetan izandako agatik, hasierako momentuetan izandako agatik, hasierako momentuetan izandako agatik, hasierako momentuetan izandako
pazientziagatik eta neure buruan konfiantza izaten eta baloratzen erakusteagatik. pazientziagatik eta neure buruan konfiantza izaten eta baloratzen erakusteagatik. pazientziagatik eta neure buruan konfiantza izaten eta baloratzen erakusteagatik. pazientziagatik eta neure buruan konfiantza izaten eta baloratzen erakusteagatik.
Ez dakit nola eskertu zure laguntza amaigabea urte luze hauetan zehar bizi izan Ez dakit nola eskertu zure laguntza amaigabea urte luze hauetan zehar bizi izan Ez dakit nola eskertu zure laguntza amaigabea urte luze hauetan zehar bizi izan Ez dakit nola eskertu zure laguntza amaigabea urte luze hauetan zehar bizi izan
ditugun arlo ezberdinetan. Espero ser tu ditugun arlo ezberdinetan. Espero ser tu ditugun arlo ezberdinetan. Espero ser tu ditugun arlo ezberdinetan. Espero ser tu yangyangyangyang en esos f en esos f en esos f en esos futuros proyectos que ya uturos proyectos que ya uturos proyectos que ya uturos proyectos que ya
tenemos pensados y hay que pulir.tenemos pensados y hay que pulir.tenemos pensados y hay que pulir.tenemos pensados y hay que pulir.
Eskerrik asko, bost urte luze hauetan zehar, nire ametsa egi bihurtzeko ahaleginak egin Eskerrik asko, bost urte luze hauetan zehar, nire ametsa egi bihurtzeko ahaleginak egin Eskerrik asko, bost urte luze hauetan zehar, nire ametsa egi bihurtzeko ahaleginak egin Eskerrik asko, bost urte luze hauetan zehar, nire ametsa egi bihurtzeko ahaleginak egin
dituzten guztiei. Laborategiko familia, ikaragarri lagundu didazue! Luis, Jon, Javi, dituzten guztiei. Laborategiko familia, ikaragarri lagundu didazue! Luis, Jon, Javi, dituzten guztiei. Laborategiko familia, ikaragarri lagundu didazue! Luis, Jon, Javi, dituzten guztiei. Laborategiko familia, ikaragarri lagundu didazue! Luis, Jon, Javi,
Adolfo, Bego, Gorka, Adolfo, Bego, Gorka, Adolfo, Bego, Gorka, Adolfo, Bego, Gorka, IñakIñakIñakIñaki L., Leire A., Carmen E., Luz, Paco, i L., Leire A., Carmen E., Luz, Paco, i L., Leire A., Carmen E., Luz, Paco, i L., Leire A., Carmen E., Luz, Paco, Fátima, Asier, Nerea, Fátima, Asier, Nerea, Fátima, Asier, Nerea, Fátima, Asier, Nerea,
Usue, Leyre, Lorena,Usue, Leyre, Lorena,Usue, Leyre, Lorena,Usue, Leyre, Lorena, Iñaki A., Iñaki A., Iñaki A., Iñaki A., Jaime (umoretsu!), Naiara (pelirroja), Elena Jaime (umoretsu!), Naiara (pelirroja), Elena Jaime (umoretsu!), Naiara (pelirroja), Elena Jaime (umoretsu!), Naiara (pelirroja), Elena
(detailista), Ekaitz eta Naia ((detailista), Ekaitz eta Naia ((detailista), Ekaitz eta Naia ((detailista), Ekaitz eta Naia (ChapeauChapeauChapeauChapeau, aurrera bolie!!), Itziar, Maider, Amaia, , aurrera bolie!!), Itziar, Maider, Amaia, , aurrera bolie!!), Itziar, Maider, Amaia, , aurrera bolie!!), Itziar, Maider, Amaia,
Clara Clara Clara Clara eta Ana (erizainoi esker milaeta Ana (erizainoi esker milaeta Ana (erizainoi esker milaeta Ana (erizainoi esker mila hor hor hor horren ondo zaintzeagatikren ondo zaintzeagatikren ondo zaintzeagatikren ondo zaintzeagatik). ). ). ).
Baina batez ere, Alex, gracias, por esas charlas de despacho, por escucharme y Baina batez ere, Alex, gracias, por esas charlas de despacho, por escucharme y Baina batez ere, Alex, gracias, por esas charlas de despacho, por escucharme y Baina batez ere, Alex, gracias, por esas charlas de despacho, por escucharme y
animarme a tirar para adelante, aún cuando no he podido. Y mis Blondieanimarme a tirar para adelante, aún cuando no he podido. Y mis Blondieanimarme a tirar para adelante, aún cuando no he podido. Y mis Blondieanimarme a tirar para adelante, aún cuando no he podido. Y mis Blondie----s!! s!! s!! s!!
¿Cómo agradeceros todo lo que habéis hecho por mí? Mis compis de piso, Jon y¿Cómo agradeceros todo lo que habéis hecho por mí? Mis compis de piso, Jon y¿Cómo agradeceros todo lo que habéis hecho por mí? Mis compis de piso, Jon y¿Cómo agradeceros todo lo que habéis hecho por mí? Mis compis de piso, Jon y
Laura, gracias por esos momentos de cenita y tele, risas o locuraLaura, gracias por esos momentos de cenita y tele, risas o locuraLaura, gracias por esos momentos de cenita y tele, risas o locuraLaura, gracias por esos momentos de cenita y tele, risas o locura. Cuá. Cuá. Cuá. Cuánto te echo nto te echo nto te echo nto te echo
de menos, Laura!! Gracias, por ser mi Pon, mi otra mitad. Idoia, my little sister!! de menos, Laura!! Gracias, por ser mi Pon, mi otra mitad. Idoia, my little sister!! de menos, Laura!! Gracias, por ser mi Pon, mi otra mitad. Idoia, my little sister!! de menos, Laura!! Gracias, por ser mi Pon, mi otra mitad. Idoia, my little sister!!
Eskerrik askorik handiena zuretzat, Eskerrik askorik handiena zuretzat, Eskerrik askorik handiena zuretzat, Eskerrik askorik handiena zuretzat, zure alaitasunagatik, positibotasunagatik, zure alaitasunagatik, positibotasunagatik, zure alaitasunagatik, positibotasunagatik, zure alaitasunagatik, positibotasunagatik,
laguntzagalaguntzagalaguntzagalaguntzagatik, adiskidetasunagatik,tik, adiskidetasunagatik,tik, adiskidetasunagatik,tik, adiskidetasunagatik, en fin, en fin, en fin, en fin, denagatik, denagatik, denagatik, denagatik, ez nuke bukatukoez nuke bukatukoez nuke bukatukoez nuke bukatuko eta!! eta!! eta!! eta!!
Mila esker, Arantza, momenturo hor egoteagatik. Dudan euskarako ikaslerik finena zara. Mila esker, Arantza, momenturo hor egoteagatik. Dudan euskarako ikaslerik finena zara. Mila esker, Arantza, momenturo hor egoteagatik. Dudan euskarako ikaslerik finena zara. Mila esker, Arantza, momenturo hor egoteagatik. Dudan euskarako ikaslerik finena zara.
Zeurea ere bada tesiaren zatitxo bat.Zeurea ere bada tesiaren zatitxo bat.Zeurea ere bada tesiaren zatitxo bat.Zeurea ere bada tesiaren zatitxo bat.
Ezin eskertu gabe uEzin eskertu gabe uEzin eskertu gabe uEzin eskertu gabe utzi, gure “bizilagun” maiteak, tzi, gure “bizilagun” maiteak, tzi, gure “bizilagun” maiteak, tzi, gure “bizilagun” maiteak, gugatik balitgugatik balitgugatik balitgugatik balitz, mundua konponduta z, mundua konponduta z, mundua konponduta z, mundua konponduta
legoklegoklegoklegoke=e=e=e=
Ai ama, Janire, zenbat berriketa, negar eta buruhauste lan honen inguruan. Ai ama, Janire, zenbat berriketa, negar eta buruhauste lan honen inguruan. Ai ama, Janire, zenbat berriketa, negar eta buruhauste lan honen inguruan. Ai ama, Janire, zenbat berriketa, negar eta buruhauste lan honen inguruan.
Izugarri lagundu didazu hilabete luze hauetan, nire laneko eta buruko “zuzendari Izugarri lagundu didazu hilabete luze hauetan, nire laneko eta buruko “zuzendari Izugarri lagundu didazu hilabete luze hauetan, nire laneko eta buruko “zuzendari Izugarri lagundu didazu hilabete luze hauetan, nire laneko eta buruko “zuzendari
pertsonala” izan zara. Benepertsonala” izan zara. Benepertsonala” izan zara. Benepertsonala” izan zara. Bene----benetan, eskerrik asko. Zure falta ikaragarbenetan, eskerrik asko. Zure falta ikaragarbenetan, eskerrik asko. Zure falta ikaragarbenetan, eskerrik asko. Zure falta ikaragarri sumatuko ri sumatuko ri sumatuko ri sumatuko
dut. Zorte ona opa dizut!dut. Zorte ona opa dizut!dut. Zorte ona opa dizut!dut. Zorte ona opa dizut!
Aintzane y Marian: Gracias por vuestros muchísimos consejos y ayuda cuando más Aintzane y Marian: Gracias por vuestros muchísimos consejos y ayuda cuando más Aintzane y Marian: Gracias por vuestros muchísimos consejos y ayuda cuando más Aintzane y Marian: Gracias por vuestros muchísimos consejos y ayuda cuando más
lo he necesitado.lo he necesitado.lo he necesitado.lo he necesitado.
Encarni, la alegría de la huerta!!! Gracias por alegrarnos el día cada mañana.Encarni, la alegría de la huerta!!! Gracias por alegrarnos el día cada mañana.Encarni, la alegría de la huerta!!! Gracias por alegrarnos el día cada mañana.Encarni, la alegría de la huerta!!! Gracias por alegrarnos el día cada mañana.
Carlos, eskariak egiten eskainiCarlos, eskariak egiten eskainiCarlos, eskariak egiten eskainiCarlos, eskariak egiten eskaini diguzun d diguzun d diguzun d diguzun denbora eta laguntzaenbora eta laguntzaenbora eta laguntzaenbora eta laguntzagatik, eskerrik asko.gatik, eskerrik asko.gatik, eskerrik asko.gatik, eskerrik asko.

Kuadrilako lagunak, Ainhoa eta ItziarKuadrilako lagunak, Ainhoa eta ItziarKuadrilako lagunak, Ainhoa eta ItziarKuadrilako lagunak, Ainhoa eta Itziar, eskerrik beroenak, benetan,, eskerrik beroenak, benetan,, eskerrik beroenak, benetan,, eskerrik beroenak, benetan, zuen zuen zuen zuen
adiskidetasunagatik eta adiskidetasunagatik eta adiskidetasunagatik eta adiskidetasunagatik eta izan duzuen pazientziagatik! Ea noiz egiten dugun bidaia izan duzuen pazientziagatik! Ea noiz egiten dugun bidaia izan duzuen pazientziagatik! Ea noiz egiten dugun bidaia izan duzuen pazientziagatik! Ea noiz egiten dugun bidaia
hori. Umeak koxkortu ondoren igual?? hori. Umeak koxkortu ondoren igual?? hori. Umeak koxkortu ondoren igual?? hori. Umeak koxkortu ondoren igual?? Aitor,Aitor,Aitor,Aitor, Xabi, Oier eta JuXabi, Oier eta JuXabi, Oier eta JuXabi, Oier eta June, zenbat momentu ne, zenbat momentu ne, zenbat momentu ne, zenbat momentu
on pasa arazi dizkidazuenon pasa arazi dizkidazuenon pasa arazi dizkidazuenon pasa arazi dizkidazuen…………
UrzelaiUrzelaiUrzelaiUrzelai ´́́́tar familia: Eskerrik asko Euskara maitatzen eta beragatik borrokatzen tar familia: Eskerrik asko Euskara maitatzen eta beragatik borrokatzen tar familia: Eskerrik asko Euskara maitatzen eta beragatik borrokatzen tar familia: Eskerrik asko Euskara maitatzen eta beragatik borrokatzen
erakusteagatikerakusteagatikerakusteagatikerakusteagatik.... Gurekin ez dauden Jon, Xanti eta Inaxiorengandik hasi, eta Gurekin ez dauden Jon, Xanti eta Inaxiorengandik hasi, eta Gurekin ez dauden Jon, Xanti eta Inaxiorengandik hasi, eta Gurekin ez dauden Jon, Xanti eta Inaxiorengandik hasi, eta
bukatzeko, Manolo, Maixa, Ion eta Uxue. Goretti, Igor ebukatzeko, Manolo, Maixa, Ion eta Uxue. Goretti, Igor ebukatzeko, Manolo, Maixa, Ion eta Uxue. Goretti, Igor ebukatzeko, Manolo, Maixa, Ion eta Uxue. Goretti, Igor eta Saioa. Angeli, Mariasun, ta Saioa. Angeli, Mariasun, ta Saioa. Angeli, Mariasun, ta Saioa. Angeli, Mariasun,
Txarles, Jon, Xabi eta Amaia. Mila esker benetan, izugarriak zarete!!Txarles, Jon, Xabi eta Amaia. Mila esker benetan, izugarriak zarete!!Txarles, Jon, Xabi eta Amaia. Mila esker benetan, izugarriak zarete!!Txarles, Jon, Xabi eta Amaia. Mila esker benetan, izugarriak zarete!! Bazkari bat Bazkari bat Bazkari bat Bazkari bat
zor dizuet esker modura soziedadean!!!!zor dizuet esker modura soziedadean!!!!zor dizuet esker modura soziedadean!!!!zor dizuet esker modura soziedadean!!!!
Familia Pérez: Esa mitad castellana de mí, que sale más en verano, pero siempre está Familia Pérez: Esa mitad castellana de mí, que sale más en verano, pero siempre está Familia Pérez: Esa mitad castellana de mí, que sale más en verano, pero siempre está Familia Pérez: Esa mitad castellana de mí, que sale más en verano, pero siempre está
presente. Gracias, popresente. Gracias, popresente. Gracias, popresente. Gracias, por hacerme pasar esos veranos increíbles que tanta falta hacen r hacerme pasar esos veranos increíbles que tanta falta hacen r hacerme pasar esos veranos increíbles que tanta falta hacen r hacerme pasar esos veranos increíbles que tanta falta hacen
para olvidarse de todos los problemas. Por esas tardes de piscina o pantano y para olvidarse de todos los problemas. Por esas tardes de piscina o pantano y para olvidarse de todos los problemas. Por esas tardes de piscina o pantano y para olvidarse de todos los problemas. Por esas tardes de piscina o pantano y
pipas en las escaleras de la plaza mayor, helado y verbenas, los paseos de pipas en las escaleras de la plaza mayor, helado y verbenas, los paseos de pipas en las escaleras de la plaza mayor, helado y verbenas, los paseos de pipas en las escaleras de la plaza mayor, helado y verbenas, los paseos de
medianoche, las noches con la fuente de cmedianoche, las noches con la fuente de cmedianoche, las noches con la fuente de cmedianoche, las noches con la fuente de colores, picnic y lluvia de estrellas, olores, picnic y lluvia de estrellas, olores, picnic y lluvia de estrellas, olores, picnic y lluvia de estrellas,
chismorreos de juventud en nuestra querida calle tan familiar. El olor a tierra chismorreos de juventud en nuestra querida calle tan familiar. El olor a tierra chismorreos de juventud en nuestra querida calle tan familiar. El olor a tierra chismorreos de juventud en nuestra querida calle tan familiar. El olor a tierra
húmeda y a pipas tostadas, mmm!. Gracias, tíoshúmeda y a pipas tostadas, mmm!. Gracias, tíoshúmeda y a pipas tostadas, mmm!. Gracias, tíoshúmeda y a pipas tostadas, mmm!. Gracias, tíos----tías, primos y primas. tías, primos y primas. tías, primos y primas. tías, primos y primas.
Y cómo era aquello deY cómo era aquello deY cómo era aquello deY cómo era aquello de………… “Dijo el toro al morir: “Dijo el toro al morir: “Dijo el toro al morir: “Dijo el toro al morir:
Siento dejar este mSiento dejar este mSiento dejar este mSiento dejar este mundo, undo, undo, undo,
sin probar Pipas Facundo”.sin probar Pipas Facundo”.sin probar Pipas Facundo”.sin probar Pipas Facundo”.
EskerrakEskerrakEskerrakEskerrak eman nahi dizkizuet, aipatu gabe geratu eman nahi dizkizuet, aipatu gabe geratu eman nahi dizkizuet, aipatu gabe geratu eman nahi dizkizuet, aipatu gabe geratu arren, familiakotzat zaituztedan arren, familiakotzat zaituztedan arren, familiakotzat zaituztedan arren, familiakotzat zaituztedan
guztioi. Nigatik arduratzeagatik, itxaroteagatik eta batzuetan ez agertzeagatik, zer guztioi. Nigatik arduratzeagatik, itxaroteagatik eta batzuetan ez agertzeagatik, zer guztioi. Nigatik arduratzeagatik, itxaroteagatik eta batzuetan ez agertzeagatik, zer guztioi. Nigatik arduratzeagatik, itxaroteagatik eta batzuetan ez agertzeagatik, zer
moduz doa? galdetzeagatik, animoengatik, emandako lagumoduz doa? galdetzeagatik, animoengatik, emandako lagumoduz doa? galdetzeagatik, animoengatik, emandako lagumoduz doa? galdetzeagatik, animoengatik, emandako laguntzagatik eta abar luze ntzagatik eta abar luze ntzagatik eta abar luze ntzagatik eta abar luze
bategatik.bategatik.bategatik.bategatik.
Gaizki esanak ahaztuGaizki esanak ahaztuGaizki esanak ahaztuGaizki esanak ahaztu
eta ongi esanak gogoan hartu!!eta ongi esanak gogoan hartu!!eta ongi esanak gogoan hartu!!eta ongi esanak gogoan hartu!!
MilaMilaMilaMila----milamilamilamila esker etxekoei. Zer egingo nuen esker etxekoei. Zer egingo nuen esker etxekoei. Zer egingo nuen esker etxekoei. Zer egingo nuen zuek gabe? Ezer ez, seguraski. Nire ametsa zuek gabe? Ezer ez, seguraski. Nire ametsa zuek gabe? Ezer ez, seguraski. Nire ametsa zuek gabe? Ezer ez, seguraski. Nire ametsa
dena lortzen laguntzeagatik eta horiek lortzeko ez dela hetsi behar erakusteagatikdena lortzen laguntzeagatik eta horiek lortzeko ez dela hetsi behar erakusteagatikdena lortzen laguntzeagatik eta horiek lortzeko ez dela hetsi behar erakusteagatikdena lortzen laguntzeagatik eta horiek lortzeko ez dela hetsi behar erakusteagatik, , , ,
eskerrik asko!! Zuengandik ikasi eta jaso dut naizen guztia. Aita, páspas! nuestro eskerrik asko!! Zuengandik ikasi eta jaso dut naizen guztia. Aita, páspas! nuestro eskerrik asko!! Zuengandik ikasi eta jaso dut naizen guztia. Aita, páspas! nuestro eskerrik asko!! Zuengandik ikasi eta jaso dut naizen guztia. Aita, páspas! nuestro
pitufo gruñón, de ti he heredado la tranquilidad que tanta falta me ha hecho para pitufo gruñón, de ti he heredado la tranquilidad que tanta falta me ha hecho para pitufo gruñón, de ti he heredado la tranquilidad que tanta falta me ha hecho para pitufo gruñón, de ti he heredado la tranquilidad que tanta falta me ha hecho para
seguir adelante en muchísimas ocasiones. Ama, zugandik, buruseguir adelante en muchísimas ocasiones. Ama, zugandik, buruseguir adelante en muchísimas ocasiones. Ama, zugandik, buruseguir adelante en muchísimas ocasiones. Ama, zugandik, buru----gogorkeria eta batgogorkeria eta batgogorkeria eta batgogorkeria eta bat----
bateko odbateko odbateko odbateko odolololol----beroa, beste momentu askotan atera behar izan dudana. Biongandik, beroa, beste momentu askotan atera behar izan dudana. Biongandik, beroa, beste momentu askotan atera behar izan dudana. Biongandik, beroa, beste momentu askotan atera behar izan dudana. Biongandik,
jenioa baina baita pazientzia, maitasuna, besteekiko errespetua eta langile izatea. jenioa baina baita pazientzia, maitasuna, besteekiko errespetua eta langile izatea. jenioa baina baita pazientzia, maitasuna, besteekiko errespetua eta langile izatea. jenioa baina baita pazientzia, maitasuna, besteekiko errespetua eta langile izatea.
Añes, ahizpa eta adiskidea, eta orain amatxo!! beti izan zara niretzat eredu. Añes, ahizpa eta adiskidea, eta orain amatxo!! beti izan zara niretzat eredu. Añes, ahizpa eta adiskidea, eta orain amatxo!! beti izan zara niretzat eredu. Añes, ahizpa eta adiskidea, eta orain amatxo!! beti izan zara niretzat eredu.
Imanol eta Adur txikia. ZImanol eta Adur txikia. ZImanol eta Adur txikia. ZImanol eta Adur txikia. Zuengatik eta zuentzat nire Tesia. Pilauengatik eta zuentzat nire Tesia. Pilauengatik eta zuentzat nire Tesia. Pilauengatik eta zuentzat nire Tesia. Pila----pila, ilargiraino pila, ilargiraino pila, ilargiraino pila, ilargiraino
ta bueltan MAITTE ZAITTUZTET!!!ta bueltan MAITTE ZAITTUZTET!!!ta bueltan MAITTE ZAITTUZTET!!!ta bueltan MAITTE ZAITTUZTET!!!
Liburuaren ilustrazioa: Itziar Barrutia Urretabizkaia


Laburdurak
ACE: Angiotentsina eraldatzen duen entzima
ACE2: Angiotentsina eraldatzen duen entzima 2
APA: Aminopeptidasa azidoa
APB: Aminopeptidasa basikoa
APN: Aminopeptidasa neutroa
Asp-AP: Aspartil aminopeptidasa
b/b: Bolumena/bolumena
BSA: Behi-seroalbumina
CAP: Zistil aminopeptidasa
DAGNPG: N-Dansyl-D-Ala-Gly-pNO2-Phe-Gly
DMSO: Dimetil-sulfoxidoa
DNA: Azido desoxiribonukleikoa
DPPIV: Dipeptidil peptidasa IV
DTT: DL-ditioteitrola
EDTA: Etileno diamino tetrazetikoa azidoa
ES: Errore estandarra
FAPGG: 3-(2-furylacryloyl-l-Phe-Ala-Gly-Gly)
MMP: Matrizeko metaloproteasa
mRNA: RNA mezularia
NEP: Endopeptidasa neutroa
OME: Osasunerako Mundu Erakundea
p/b: pisua/bolumena
PEP: Prolil endopeptidasa
PGI: Piroglutamil peptidasa I
PSA: Puromizinarekiko sentikortasuna duen aminopeptidasa
PU: Peptidasa unitatea
RAS: Errenina angiotentsina sistema
SPNPT: Abz-Ser-Pro-3-nitro-Tyr-OH
TNM: Tumorea, Gongoil linfatikoak, Metastasia (Tumour, Node, Metastasis)


Aurkibidea
1. Sarrera_________________________________________ ______________________ 1
1. Heste lodia eta bere egitura.........................................................................................3
2. Heste lodiko neoplasiak...............................................................................................4
2.1. Neoplasia mota ezberdinen sailkapena 4
2.2. Neoplasien ezaugarri orokorrak 7
2.3. Neoplasien sintomatologia, diagnosia eta pronostikoa 9
2.4. Minbizi-markatzaileak 10
3. Peptidasak.................................................................................................................11
3.1. Peptidasen sailkapena eta nomenklatura 11
3.2. Peptidasen funtzio biologikoak 13
3.2.1. Peptidoen eraldaketa metabolikoa 13
3.2.2. Peptidasen ekintza intrakrinoa 14
3.2.3. Peptidasen ekintza batzuk 15
3.3. Peptidasak fisiologian eta patologian 15
3.4. Peptidasen ekintza tumoreen biologian 16
3.4.1. Peptidoak eta peptidasak tumoreen biologian 16
3.4.2. Peptidoen erregulazioa ulertzeko eredua 17
3.4.3. Peptidasen funtzioa hazkuntza prozesuetan 18
3.5. Ikertutako peptidasak 20
3.5.1. Puromizinarekiko sentikortasuna duen aminopeptidasa (PSA) 23
3.5.2. Aminopeptidasa neutroa (APN) 24
3.5.3. Aminopeptidasa basikoa (APB) 25
3.5.4. Aspartil aminopeptidasa (Asp-AP) 25
3.5.5. Zistil aminopeptidasa (CAP) 27
3.5.6. Piroglutamil peptidasa I (PGI) 28
3.5.7. Dipeptidil peptidasa IV (DPPIV) 29
3.5.8. Prolil endopeptidasa (PEP) 30
3.5.9. Aminopeptidasa azidoa (APA) 30
3.5.10. Angiotentsina eraldatzen duen entzima (ACE) 31
3.5.11. Angiotentsina eraldatzen duen entzima 2 (ACE2) 33
3.5.12. Endopeptidasa neutroa (NEP) 34
Helburuak _________________________________________ _______________ 35

2. Materialak eta Metodoak _________________________ ______________________ 39
1. Materiala....................................................................................................................41
2. Erabilitako laginen erdiespena edo lorpena...............................................................43
2.1. Heste-laginak 43
2.2. Plasma-laginak 43
3. Peptidasen entzima-jardueraren azterketa ................................................................43
3.1. Laginen prestaketa 43
3.1.1. Heste-laginen prestaketa 44
3.1.2. Plasma-laginen prestaketa 45
3.2. Lagineko proteina kopuruaren neurketa 45
3.3. Entzima-jarduera 45
3.3.1. Entzima-jardueraren azterketa β-naftilamidak substratu gisa erabiliz 46
3.3.2. Entzima-jardueraren azterketa FAPGG (3-(2-furylacryloyl-l-Phe-
Gly-Gly)) substratu gisa erabiliz 47
3.3.3. Entzima-jardueraren azterketa SPNPT (Abz-Ser-Pro-3-nitro-
Tyr-OH) substratu gisa erabiliz 48
3.3.4. Entzima-jardueraren azterketa DAGNPG (N-Dansyl-D-Ala-Gly-
(pNO2)-Phe-Gly) substratu gisa erabiliz 49
4. Minbiziaren sailkapena ..............................................................................................50
4.1. Gradu histologikoaren sailkapena: OME-ren graduak 50
4.2. Estadioaren sailkapena: TNM sistema 52
5. Datuen analisia..........................................................................................................53
3. Emaitzak ________________________________________ ____________________ 55
1. Peptidasen azterketa: Entzima-jarduera....................................................................57
1.1. Puromizinarekiko sentikortasuna duen aminopeptidasa (PSA) 58
1.2. Aminopeptidasa neutroa (APN) 59
1.3. Aminopeptidasa basikoa (APB) 60
1.4. Aspartil aminopeptidasa (Asp-AP) 61
1.5. Zistil aminopeptidasa (CAP) 62
1.6. Piroglutamil peptidasa I (PGI) 63
1.7. Dipeptidil peptidasa IV (DPPIV) 64
1.8. Prolil endopeptidasa (PEP) 65
1.9. Aminopeptidasa azidoa (APA) 66
1.10. Angiotentsina eraldatzen duen entzima (ACE) 67

1.11. Angiotentsina eraldatzen duen entzima 2 (ACE2) 68
1.12. Endopeptidasa neutroa (NEP) 69
2. Peptidasen jardueraren azterketa: Graduak eta Estadioak .......................................70
2.1. Heste-zeluletako entzimen jarduera 70
2.2. Plasmako entzimen jarduera 70
3. Peptidasen aktibitatearen laburpena .........................................................................72
4. Eztabaida _______________________________________ ____________________ 73
1. Peptidasen entzima-jarduera mukosa normalean eta heste lodiko
neoplasietan 75
2. Plasmako peptidasen ekintza koloneko minbizia zuten gaixoetan eta
kontroletan 84
3. Peptidasen aktibitatea minbiziaren gradu eta estadio ezberdinetan 89
4. Peptidasen asaldurak eta erregulazio peptidikoa heste lodiko kartzinogenesian 95
5. Ondorioak _______________________________________ ___________________ 101
6. Bibliografia ____________________________________ _____________________ 105
7. Eranskinak ______________________________________ ___________________ 125


Sarrera

Sarrera
3
1. SARRERA
1. Heste lodia eta bere egitura
Heste lodia, heste mehearen amaieratik uzkiraino doan hodi muskulutsua da eta
digestio aparatuaren azken zatia da. Bere funtzio nagusiak, hondakinen gordetzea,
prozesamendua eta gorozkien kanporatzea dira. Prozesamendu horren parte da ur eta
elektrolitoen zurgapena (De Gandarias et al., 2001).
Hesteak sabel aldean daude kokatuta, gibelaren eta urdailaren azpian. Ahoratzen
dugun elikagaien prozesamenduaren azken fasea hemen gertatzen da. Heste mehean
gertatzen den nutrienteen zurgapenaren ondoren, mugimendu peristaltikoen bidez kimoa
heste lodira mugitzen da. Heste lodian kimoari ur kopuru handiena kentzen zaio gorozki
lehorra sortuz. Hau, ondestean pilatuko da sabel-hustutzera arte. Prozesu honetan zehar,
heste lodiak eginkizun fisiologikoak eta biokimikoak betetzen ditu, egokiro gertatu behar
direnak (Garro, 2000).
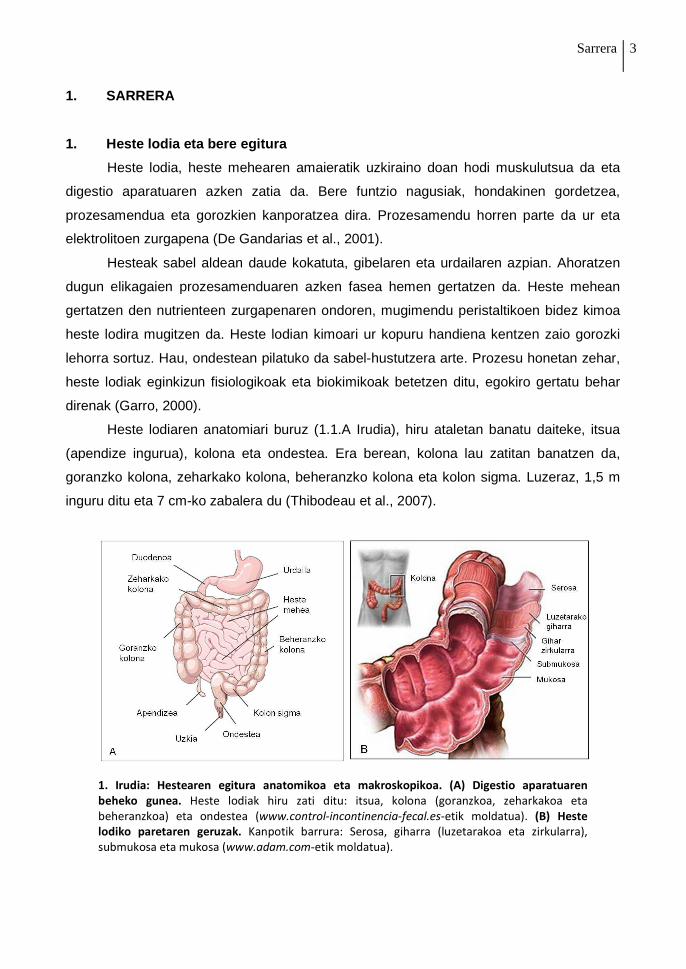
Heste lodiaren anatomiari buruz (1.1.A Irudia), hiru ataletan banatu daiteke, itsua
(apendize ingurua), kolona eta ondestea. Era berean, kolona lau zatitan banatzen da,
goranzko kolona, zeharkako kolona, beheranzko kolona eta kolon sigma. Luzeraz, 1,5 m
inguru ditu eta 7 cm-ko zabalera du (Thibodeau et al., 2007).
1. Irudia: Hestearen egitura anatomikoa eta makroskopikoa. (A) Digestio aparatuaren
beheko gunea. Heste lodiak hiru zati ditu: itsua, kolona (goranzkoa, zeharkakoa eta beheranzkoa) eta ondestea (www.control-incontinencia-fecal.es-etik moldatua). (B) Heste
lodiko paretaren geruzak. Kanpotik barrura: Serosa, giharra (luzetarakoa eta zirkularra), submukosa eta mukosa (www.adam.com-etik moldatua).

4
Sarrera
Hestearen, hau da, hodi digestibo osoko paretaren egitura makroskopikoari
dagokionez, hainbat geruza dituela aipatu behar da (1.1.B Irudia). Kanpotik barrura,
lehenengo geruza, serosa (mesenterioa) da. Hau, ehun konektiboz eta egiturazko ehunez
osatua dago, bertara heltzen dira digestio hodia elikatzen duten odol-hodiak, kapilar
linfatikoak eta nerbio-inerbazioak. Bigarren geruzan giharra dago, luzetarako giharra
lehenbizi eta zirkularra ondoren. Digestio-mugimenduetarako beharrezko dira muskulu
geruza hauek. Bien artean, nerbio sistema enterikoaren Auerbach plexua kokatzen da,
mugimendu hauen kontrolaren arduraduna. Hirugarren geruzan, azpimukosa dago, ehun
konektiboz osatua, odol-hodiak, guruin exokrinoak eta kapilar linfatikoak dituen geruza da.
Geruza honetan ere, nerbio sistema enterikoaren Meissner plexua kokatzen da, digestio-
jariaketaren kontrolaren arduraduna. Azkenik, mukosa dago, bi azpigeruzetan banatzen
dena. Bata, muskularis mukosa, muskulu leuneko mintz mehea, mugimenduek bertako
mukosa-guruinen jariaketa eragiten dute. Bestea, epitelioa, tolesturak dituena eta hodiaren
argiarekin kontaktuan dagoena (Berne et al., 2002; Stevens et al., 2006).
2. Heste lodiko neoplasiak
Neoplasia bat, normala ez den ehun masa bat da, ehun normalen aldean, neurri
ezkontrolatuan hazten dena, behin hazteko estimulu hori galdu ostean ere. Neoplasiei
tumore ere baderitze eta bi taldetan sailkatzen dira, onberak eta gaiztoak, baldin eta
tumorearen izaera klinikoa zein den (Kumar et al., 2008). Minbizi bezala izendatzen diren
gaixotasunek ondorengo ezaugarriak dituzte, egiturarik eta kontrolik gabeko hazkuntza
zelularra jasaten dutela eta beste ehunetarako hedatze gaitasuna (metastasia) dutela.
Egun, minbizia da gehien ikertzen den gaixotasuna, baita ikuspuntu ezberdin
askotatik ikertzen dena ere. Gainera, arlo molekularrean lortutako informazio ugariari
esker, urte gutxian aurrerapen handiak egin dira gaixotasun honi buruzko ezagutzan.
2.1. Neoplasia mota ezberdinen sailkapena
1958-tik Osasunerako Mundu Erakundeak (OME) hainbat zentru ditu tumore mota
ezberdinen anatomi-, histologi- eta gaiztotze graduaren sailkapena egitearen proiektuan
dihardutenak. Jada, 1976-an OME-k argitaratu zuen, Minbizien Nazioarteko Histologia
Sailkapenaren lehen edizioa eta handik hamar urtera bigarren hau, Hesteetako tumoreen
histologi-sailkapena (Histological typing of intestinal tumours, Jass eta Sobin 1989).Bertan,
agertzen den sailkapenaren moldapen bat da jarraian adierazten dena (1.1. Taula).

Sarrera
5
1.1. Taula: heste lodiko neoplasia garrantzitsuen sailkapena (Kumar et al., 2008).
Heste lodiko neoplasiak
Polipo ez neoplasikoak
Polipo hiperplasikoak Polipo hamartomatosoak Gaztaroko polipoak Peutz-Jeghers-en polipoak Hanturazko polipoak Polipo linfoideak Epitelioko lesio neoplasikoak
Polipo onberak Adenomak
Lesio gaiztoak Adenokartzinoma Uzkiko kartzinoma ezkatatsua
Bestelako tumoreak Gastrohesteetako estromako tumoreak Tumore kartzinoidea Linfoma
Polipoa, hesteko argirantz ateratzen den tumore-masa bat da. Itxurari dagokionez,
polipoak zurtoindunak ala eseriak izaten dira eta oro har, hestearen barneko epitelioan
dute jatorria, hots, mukosan. Bi motatako polipoak sailkatzen dira. Bata, polipo ez-
neoplasikoak deritzena. Hauek, mukosaren heltze edo egitura ezohikoaren edota
hanturaren ondorioz sor daitezke. Gaiztotzeko arriskurik ez dute izaten normalean eta
ezagunenak polipo hiperplasikoak dira. Bestea, polipo neoplasikoak deritzena. Hauen
artean ezagunenak polipo adenomatosoak edo adenomak dira. Kasu honetan,
epitelioaren hazkuntzaren eta displasiaren ondorioz sortzen diren polipoak dira. Tumore
onberak izan arren, azken hauek gerora, kartzinoma sortzeko gai izan daitezke.
Adenomak izateko aurretiko joera familiarra dago, adibidez, Poliposi familiar sindromea
(1FAP), zeinetan gaixoek 500-2000 adenoma izan ditzaketen hestean zehar. Gaixoak
heste lodiko minbizia izango du bere bizitzaren momentu batean, gehienetan bizitzaren
erdialdean (Leslie et al., 2002; Kumar et al., 2008; Meyerhardt et al., 2008).
Adenokartzinoma: ia kasurik gehienetan polipo adenomatoso bat izaten da
minbiziaren jatorria. Adenoma bat gaiztotu eta kartzinoman bilakatzen dela azaltzen duten
bi bide patogeniko deskribatu dira. Bata, adenoma-kartzinoma sekuentzia (edo APC/β-
katenina bidea) deiturikoa eta bestea, DNA zuzentzen duten geneen bidea (edo
mikrosateliteen ezegonkortasuna). Bi kasuetan, ehun osasuntsuko zelulen gene-
mutazioen pilaketen ondorioz gertatzen da gaiztotzea. Baina asaldatuta dauden geneak
1 FAP: Familial adenomatous polyposis

6
Sarrera
ez dira berdinak bi bideetan, ezta mutazioak gertatzeko prozesuak ere (Leslie et al., 2002;
Meyerhardt et al., 2008).
Adenoma-kartzinoma sekuentziak, ezegonkortasun kromosomikoa eta
onkogeneetan zein gene supresoreetan mutazioen pilaketa mailakatua ditu ezaugarritzat
(1.2. Irudia). Hasiera batean, gene supresoreetan mutazio bat jaiotzetik izateak edo gerora
mutatzeak, bilakaeraren lehen pausua eragiten du. Ondorengo metilazioek eta mutazioek
epitelioko zelulen hazkuntza ezkontrolatua eragiten dute, adenoma bat sortuz. Fase
honetan, beste hainbat mutazio eta kromosometan asaldura larriak gertatzen direla
deskribatu da, adenoma progresiboki handituz, displasikoago bihurtuz eta azkenean,
minbizi inbasore bat, hots, heste lodiko adenokartzinoma bat sortuz. Sekuentzia hau,
koloneko minbizi esporadikoen % 80-aren erantzule da (Leslie et al., 2002; Kumar et al.,
2008; Spisak et al., 2010). Bide patogeniko honetan eraldatzen diren geneak, APC eta
p53 gene supresoreak, K-ras protoonkogenea edo kromosomen galerak (SMAD2 eta 4)
direla deskribatu da, eta ikertutako minbizien % 50-80 kasuetan eraldatuta daudela
adierazi da. APC genea eta β-kateninak (zelula arteko lotura-proteinak), elkar
erlazionatuta daude koloneko minbiziaren bilakaeran. Batak bestearen aldaketa eragiten
du, nukleoko geneen transkribapena bultzatuz eta ondorioz, zelulen hazkuntza eragiten
dute koloneko kartzinogenesiari hasiera emanez (Tucker et al., 2000; Hao et al., 2001;
Leslie et al., 2002; Arnold et al., 2005). FAP-dun gaixoek gene honetako mutazioa izaten
dute jaiotzetik, polipo adenomatoso ugari sortzea eragiten duena.
1.2. Irudia: Adenoma-adenokartzinoma sekuentzian gertatzen diren pausuak. (1) gene supresoreetan mutazioak (germinala edo somatikoa), (2) metilazioen asaldurak edo alelo normalen inaktibazioa, (3) protoonkogeneen mutazioak, (4) minbiziko gene supresoreen homozigosi galera, (5) mutazio gehigarriak eta kromosometako asaldura larriak (Davies et al., 2005 eta Kumar et al., 2008-tik moldatua).

Sarrera
7
Bigarren kartzinogenesi bidea, minbizi esporadikoen % 10-15 kasuen eragile da.
Kasu honetan, mutazioen pilaketa jasaten da ere, baina DNA-ren gene zuzentzaileetan
gertatzen da. Ondorioz, DNA kaltetua duten zelulak kontrolik gabe hazi, ezberdintzatu eta
hedatzen dira. Adenokartzinoma ugari izateko probabilitate altua izaten dute, Lynch-en
sindromea edo heste lodiko minbizi ez-poliposi hereditarioaren sindromea (2HNPCC)
duten gaixoek, DNA-ren gene zuzentzaile diren MLH1 eta MLH2-an mutazioak dituztelako.
Mutazio hauek, mikrosateliteen ezegonkortasuna (3MSI) eragiten dute DNA
konpontzerako unean eta asaldurak gertatzen dira erreplikazioan. Honegatik, izen hau ere
erabiltzen da bide hau izendatzeko (Leslie et al., 2002; Arnold et al., 2005; Kumar et al.,
2008; Noffsinger et al., 2009).
2.2. Neoplasien ezaugarri orokorrak
Heste lodiko kartzinogenesiaren ikerketak, minbiziaren bilakaeraren oinarrizko
mekanismoei buruzko ezagutza zabala eskaini du.
Ohar orokor batzuk deskribatu dira adenoma-kartzinoma bilakaeraren inguruan,
hala nola lehenik, adenoma ugari izateko prebalentzia altua duten lurraldeetan
adenokartzinomak izateko ere prebalentzia altua dutela atzeman dute. Bigarrenik,
adenomek kolon eta ondestean duten banaketa, heste lodiko minbizien banaketarekin
alderatu daiteke ere. Hirugarrenik, polipo adenomatosoak izateko probabilitate gorena,
koloneko adenokartzinoma baino urte batzuk lehenago gertatzen da. Laugarrenik, estadio
goiztiarrean diagnostikatzen denean kartzinoma inbaditzaile bat, normalean gaixoek ehun
adenomatoso bat ere izaten dute oso hurbil. Bosgarrenik, heste lodiko kartzinoma izateko
arriskuak erlazio zuzena du adenomen kopuruarekin, horregatik dute FAP-dun gaixoek
kartzinoma izateko probabilitate altua. Azkenik, osasun programei esker, adenomaren
aldizkako jarraipenak egiten badira eta agertutako adenomak erauzten badira, asko jaisten
da minbizia izateko intzidentzia (Gómez-Domínguez et al., 2006; Kumar et al., 2008).
Heste lodiko minbizien artean gehienak adenokartzinomak izaten dira, % 98-ra
iristeraino. Nahiz eta arrisku faktore ugari egon, familia aurrekariak, dieta eta bizi-jarduerak
zein gaixotasun zehatz batzuk oso esanguratsuak dira minbiziaren genesi unean.
Ezaugarri genetikoek, lehen aipatu bezala, garrantzi handia dute, baina ingurumeneko
ezaugarriek ere eragina dute minbizien garapenean. Osasun eta dieta ohiturak dira
ingurumeneko ezaugarrien artean aipagarrienak. Heste lodiko minbizian, beste minbizi
2 HNPCC: Hereditary nonpolyposis colorectal cancer syndrome 3 MSI: Microsatellite instability

8
Sarrera
askotan gertatzen den bezala, alkohola, tabakoa eta kirol gutxi egitea kaltegarriak direla
deskribatu da. Barazki-zuntzean txiroak eta animali gantzean aberatsak diren elikagaiek,
gorozkien kopurua txikitzen eta sabel-hustutze denbora luzatzen dute. Ondorioz,
hesteetako flora asaldatu egiten da heste mukosa kaltetuz (Meyerhardt et al., 2008;
Kumar et al., 2008).
Hesteek, mehea eta lodia kontuan harturik, ia 8 m luze dute. Hodi digestiboaren %
75-a heste mehea izan arren, bertan agertzen diren neoplasiak ez dira % 5-era iristen.
Gainera, neoplasia hauen artean ugariagoak izan ohi dira potentzial onbera dutenak.
Heste lodian aldiz, luzeraz motzagoa izanagatik ere, hodi digestiboko neoplasien
gehiengoa gune honetan agertzen da. Adibidez, gastrohesteetan azaltzen diren neoplasia
guztietatik, % 70 inguru adenokartzinoma kolorrektalak izaten direla deskribatu da (Kumar
et al., 2008).
Neoplasien agerpenaren edo intzidentziaren adin tarteari dagokionez, estadistikek
diote, bai polipoak zein kartzinomak 50-70 adinen artean direla sarrienak, horregatik,
jarraipeneko programetan ere tarte hau hartzen dute azterketa tarte gisa. 50 urtetik
beherakoen intzidentzia ez da % 20-ra iristen. Betiere, gizonengan emakumeengan baino
ohikoagoa dela deskribatu da (Meyerhardt et al., 2008; Izarzugaza et al., 2010).
Eusko Jaurlaritzak berriki argitaratu du (Izarzugaza et al., 2010) Euskal Autonomi
Erkidegoan (EAE) 1986. urtez geroztik, minbizien intzidentziak (2002-2006) eta
hilkortasunak (2004-2008) izan duen bilakaera, baita minbizien biziraupena bost urteetan
zehar (2000-2004). Ondorio gisa, minbiziaren ondoriozko heriotza-tasak murriztu egin
direla azaldu dute, bai gizonengan, bai emakumeengan. Urtean, batez beste, 11.229 kasu
berri diagnostikatu dira aldi horretan EAEn. Gizonen artean prostatan (% 21,3) eta birikan
(% 14,7) ageri dira kasu gehien; emakumeen artean, berriz, bularrean (% 27,2) eta
kolonean (% 9,4). Aztertutako aldian, tumore gaiztoek, batez beste, 3.490 heriotza eragin
dituzte urtean gizonen artean eta 1.962 heriotza emakumeen artean, hurrenez hurren.
Koloneko minbiziaren intzidentzia-tasak, eta hein txikiagoan, hilkortasun-tasak
etengabe handitu dira 25 urtetan (ia % 3-ra eta % 2-ra bi sexuak kontuan izanik hurrenez-
hurren). Koloneko minbiziaren biziraupenak gora egin du etengabe sexu bakoitzean,
biziraupen erlatiboak ikertutako aldian % 56-ra iritsi arte. Ondestea eta uzkiaren
minbiziaren intzidentzia-tasa estandarizatuak nabarmen hazi dira bi sexuetan. Aldi osoan,
gizonen hilkortasun-tasetan ez da aldaketa estatistikoki esanguratsurik gertatu. Aldiz,
emakumeen artean hilkortasun-tasak modu esanguratsuan jaitsi dira, urtean % 1,6
murriztuz azterlan guztian. Tumoreen biziraupenak gora egin du bi sexuetan, baina

Sarrera
9
hazkundea handiagoa da gizonetan emakumeetan baino. Bost urteren buruan bi sexuak
kontuan izanik batazbesteko biziraupen erlatiboa ikertutako aldian % 57-ra iritsi arte
(Bujanda et al., 2010; Izarzugaza et al., 2010).
Izarzugaza et al.,-ek lortutako informazio hori funtsezkoa da herritarrek minbiziari
buruzko informazio zabala izan dezaten eta Espainiako nahiz nazioarteko beste eremu
batzuetako erregistroekin konparazioak egiteko, baita diagnostikoaren eta tratamenduaren
eraginkortasuna hobetu eta arriskuak murrizteko ezinbestekoak diren ekintza eta politika
sanitarioak zehazten jarraitzeko ere.
2.3. Neoplasien sintomatologia, diagnosia eta prono stikoa
Heste lodiko neoplasiak asintomatiko mantentzen dira urteetan zehar. Sintomak
isilpean garatzen dira eta hilabete edota urteak egon daitezke diagnostikatu arte. Oro har,
minbiziaren fase berantiarrera arte ez da agertzen sintomarik eta kokatzen den gunearen
arabera sintomak ezberdinak izan daitezke, hala nola, ahulezia, burdin anemia, odoljarioa,
beherakoa, pisu galera, sabel-hustutzearen ohitura aldaketak, sukarra edo sabeleko mina.
Tumore guztiak inguruko egituretara hedatzen dira zabaltze zuzenez eta beste
organoetara metastasiz, odol eta hodi linfatikoetatik. Kasu honetan, heste lodiko minbiziak
metastatizatzerakoan gertuko gongoil linfatikoak, gibela, birika eta hezurra ditu
lehentasuneko organo. Gainera, % 25-30 kasutan kirurgia sendagarriaren mugatik at
hedatzen da (Kumar et al., 2008).
Neoplasiek sintomarik ez izateak diagnosi goiztiarra zailtzen dute. Diagnosirako
erabiltzen diren metodo ohikoenak ondeste-ukipena eta gorozkietako odol ezkutuen
analisia dira. Sigmoidoskopia, kolonoskopia eta bariozko enema ere egiten dira, baina
diagnosi hauen emaitzak ziurtatzeko edo baieztatzeko biopsia bat behar izaten da.
Konputerizatutako tomografiak eta erradiografiak normalean hedapen metastatikoa
neurtzeko erabiltzen dira (Ahlquist et al., 2002).
Heste lodiko minbiziaren pronostikoa zein izango den jakitea ez da erraza izaten.
Egun, ez da ezagutzen zein diren minbiziaren bilakaeraren faktore pronostikoak.
Esaterako, ulertezina da oraindik, heste lodiko minbizia jasan duten gaixoetan, behin
kirurgia bidez minbizia erauzi eta urte askotara, metastasien agerpena (Meyerhardt et al.,
2008; Saha et al., 2010). Ezagutzen diren adierazlerik garrantzitsuenak, diagnosia
egiterako unean minbiziaren hedapena eta izaera adierazten duten 4TNM sailkapen-
sistema eta minbiziaren histologi-gradua dira, geroago azalduko den bezala. Ezaugarri
4 TNM: Tumour, Node, Metastasis

10
Sarrera
morfologikoak, molekularrak edo markatzaile immunohistokimikoak ere lagungarriak izan
daitezkeela diruditen arren, oraindik ez dira klinikan erabilgarriak, ikertze fasean
daudelako eta oraingoz ez dutelako esperotako emaitzik eskaini (Selves, 2009).
2.4. Minbizi-markatzaileak
Minbizi-markatzaileak, minbiziaren agerpenarekin zerikusia duten substantziak dira,
hots, minbiziaren ausentziaren edo presentziaren adierazle dira. Substantzia hauek zelula
mailan agertzen dira edota gorputz likidoetan. Oro har, izaera heterogeneoa dute, baina
hormona edo entzima funtzioa izan ohi dute eta peptidasak, azken talde honen kide dira.
Azken urteotan, aipatu berri diren markatzaile hauen ikerketan asko sakontzen ari
da zientzialarien lana. Molekula egokien bilaketan dihardute, ez soilik pronostikorako
erabilgarriak izan daitezkeelako, baizik, minbiziaren ezberdintzapenerako eta diagnosirako
ere lagungarri izan daitezkeelako.
Minbizi-markatzaileak gaixotasunarekiko espezifizitatea eta sentsibilitatea izan
behar du. Bestalde, ahal bada erraztasunaz neurtu ahal izateko aukera eman behar du,
beste metodo erasokor edota garestiagoak ekidinez. Minbiziaren prebentzio
saiakeretarako, zeharkako irizpideak ebaluatzeko molekula fidagarri gisa erabil liteke
(Goessl, 2003). Minbizi-markatzaile bakarraren neurketa ez da nahikoa gaixotasunen
estadio goiztiarrak aurkitzeko, sentsibilitate gutxi baitute. Beharrezkoa da beraz, hainbat
biomarkatzaileren neurketa aldi berean egitea (Cordero et al., 2008).
Minbizi Markatzaileen Talde Europarrak (5EGTM) arau berrituak edo ildo nagusiak
deskribatzen ditu biomarkatzaileen inguruan, ondoren klinikan aplikagarri izan
daitezkeelarik. Kasu honetan, heste lodiko minbizian erabiltzen diren ehuneko, sueroko
edota gorotzetako markatzaileen erabilera eta efizientzia adierazten dira (Duffy et al.,
2007). Ehuneko markatzaileei buruz, gaixotasunaren pronostikorako eta minbiziaren
norakoaren iragarle gisa erabili ahal izateko helburuarekin ikertu dira hainbat molekula.
Euren artean, timidilato sintetasa (TS), mikrosateliteen ezegonkortasuna, p53 eta K-ras.
Kasu guztietan EGTM-ak ez du klinikarako erabilera gomendatzen, orain arteko lanetan
neurketa metodo ezberdinen erabilera eta kontraesanak aurkitu baitituzte. Gorotzetako
markatzaileak, minbiziaren estadio goiztiarraren screening-ak edo behaketak egiteko oso
erabilgarriak dira, neurterrazak dira, ez-erasokorrak eta emaitza ugari lortzeko modu
azkarra da. Hauen artean daude, gorotzetako odol ezkutuaren azterketa (6FOBT) eta
5 EGTM: European Group on Tumour Markers 6 FOBT: Faecal Occult Blood Test

Sarrera
11
gorotzetako zelulen DNA azterketa. Lehena, 50 urtetik gorako gizakietan gomendagarria
dela esan arren, bigarren metodoa oraingoz ez dute erabilgarri irizten. Azkenik, sueroko
edo plasmako markatzaileak, kirurgia ondorengo zaintzarako edo jarraipenerako
erabiltzen diren molekulak izan ohi dira. Euren artean heste lodiko minbizian erabilienak
edo ezagunenak sueroko antigeno kartzinoenbrionarioa (CEA), 19.9 karbohidrato
antigenoa (CA19.9) eta metaloproteinasen inhibitzailea (TIMP-1) dira. Talde europar
honen iritziz, CEA da gomendagarri bakarra, beste metodo pronostikoen lagungarri gisa,
II. eta III. estadioa duten gaixoetan eta kirurgia ondorengo terapian 2-3 hilabetero neurtu
daitekeen markatzailea da (Duffy et al., 2007). Beste autore batzuen iritziz ordea, CEA
bezalako markatzaileen balio adierazgarriak lortzean minbizia dagoeneko tamaina aldetik
handia dela diote, eta ziurrenik dagoeneko hedatua egongo da. Bestalde markatzaile
hauek beste hainbat minbizirengatik ere igotzen dira, adibidez, birika, bular, obario,
maskuri edo prostata, zein gaixotasun ez-neoplasikoak diren alkoholaren ondoriozko
zirrosian edo pankreatitisan eta kolitis ulzerosoan (Kumar et al., 2008).
3. Peptidasak
Peptidasek entzima talde zabal eta heterogeneoa osatzen dute. XVIII. mendetik
dira ezagunak eta urte luze hauetako beraien funtzioari buruzko ikerketek minbizi-
markatzaile eta terapia-itu bezala erabiltzeko bidea zabaldu dute.
3.1. Peptidasen sailkapena eta nomenklatura
Transkribatzen diren geneen % 2-a peptidasak dira, ondorioz proteina funtzionalen
talde handienetariko bat da. Proteina oso heterogeneoek osatzen dute talde hau, mintzari
loturiko proteinak zein proteina solugarriak dira, eta 25-300 kDa arteko tamaina dute
(Taylor, 1993). Sailkapena eta nomenklatura aldatuz joan da urteetan zehar, peptidasei
buruzko informazioa zabaltzen joan den heinean. Terminologiari dagokionez ere nahasi
xamarra izan da, entzima berberari hainbat izen eman baitzaizkio kasu askotan.
Historikoki, esanahi ezberdinak izan dituzte proteasa, proteinasa eta peptidasa
hitzek. Egun, idazle batzuek entzima proteolitiko erabiltzea gomendatzen duten arren,
ulerterraza eta egokiena delako, 7NC-IUBMB Komiteak peptidasa hitza erabiltzea
gomendatzen du peptidoen arteko loturak apurtzen dituen entzimez ari garenean. Talde
7 NC-IUBMB: Nomenclature Committe of the International Union of Biochemistry and Molecular Biology

12
Sarrera
honetako partaide diren proteinak exopeptidasa eta endopeptidasa bezala sailkatzen dira,
apurtzen duten muturreko edo kate barneko loturaren arabera.
Beraz, exopeptidasek azken 1-3 aminoazidoekiko lotura peptidikoa hidrolizatzen
dute. Gertaera hau amino ertzean ematen bada, aminopeptidasa, dipeptidil-peptidasa edo
tripeptidil-peptidasa izena hartzen dute entzimek. Karboxilo ertzean gertatzen bada aldiz,
karboxipeptidasa edo peptidil-peptidasa izendatzen dira. Dipeptidasa hitza bi animoazidoz
osatutako dipeptidoak hidrolizatzen dituen entzimari deitzeko erabiltzen da espezifikoki.
Azkenik, omega-peptidasa deituriko entzimek ordezkatuak, ziklatuak edo lotura
isopeptidikoez (α-aminoa eta α-karboxiloa ez diren taldeekiko lotura peptidikoak) lotutako
aminoazidoak apurtuko dituzte.
Endopeptidasek, kate peptidikoaren barneko lotura peptidikoak hidrolizatuko dituzte
eta egiten duten ekintza katalitikoaren arabera sailkatzen dira. Hots, katalisiaren erantzule
den taldearen jatorri kimikoaren arabera, hala nola, aspartil-endopeptidasak, zisteinil-
endopeptidasak, metaloendopeptidasak, seril-endopeptidasak, treonil-endopeptidasak eta
azkenik, oraindik sailkatu gabe dauden endopeptidasak.
Proposamen posible bat aurkezten dute Marks et al.-ek (1968a, 1968b, 1975)
entzima hauen sailkapenerako lanetan, substratu kromogeniko batzuekiko peptidasek
duten afinitatea oinarri bezala hartuz. Sailkapen honen arabera aminopeptidasak hiru
taldetan banatzen dira: azidoak, basikoak eta neutroak. Hauek, substratu azido, basiko eta
neutroak hidrolizatuko dituzte hurrenez hurren. Bestalde, eraldatzen duten peptidoaren
izena ere erabili ohi da entzima hauek izendatzeko, hala nola, entzefalinak eraldatzen
dituzten peptidasak, TRH (tirotropina askatzen duen hormona), angiotentsinak, etab.
eraldatzen dituzten peptidasak.
90. hamarkadan ezarri ziren sailkapen sistema baten oinarriak, peptidasen egitura
eta sekuentzia kontuan izanik. Hemendik sortu zen MEROPS datu-basea, 1996. urtetik
Interneten erreferentzia-iturri nagusitzat bihurtu dena, bertan aurkitu daiteke peptidasa
guztiei buruzko edozein datu. Bertan, sailkapen hierarkiko bat azaltzen da (1.2. Taula),
Katalisi mota, Klan, Familia eta Peptidasa kontzeptuak kontuan hartzen dira (Barrett et al.,
2004).
Hasteko, peptidasak Katalisi motaren arabera taldekatzen dira, lehen aipatu bezala,
katalisiaren erantzule den taldearen izate kimikoaren arabera. Hauek, taldearen lehen
letra larria hartuko lukete sailkapenean (A, C, M, S, T eta U). Ondoren, peptidasak Klan
edo Familiatan banatzen dira, Peptidasa homogeneoen taldeetan.

Sarrera
13
Klan bakoitza, jatorri komuna duten Familia bat edo batzuez osatuta dago,
Klanaren izena bigarren letra larri batez adierazten da eta ezingo da berrerabili Klan hau
ondorengo sailkapen antolaketetan galduz gero. Badaude Familia batzuk Klanen barnean
sartu ezin direnak ere.
Familiak talde horretako peptidasa ordezkagarrienarekin dituzten antzekotasunen
arabera sailkatzen dira, peptidasaren unitatea kontuan izanik. Sekuentzia zenbaki bat
ezartzen dio nomenklaturari, zenbaki hau ere berrerabili ezin delarik. Bukatzeko Peptidasa
bakoitzak izendatzaile bakar bat du, puntu batez banatutako bi zatirekin.
Azaldu bezala, sailkapen ezberdinak erabiltzen dira baina egun onartuenak
Entzimen Komisioak (8EC) aurkezten dituenak dira. Sailkapen honetan, peptidasak EC
3.4. taldean daude, lotura peptidikoak apurtzen dituzten hidrolasen taldea alegia.
Ondorengo azpisailkapena Rawlings eta Barrett-ek 1995 eta 1999 urteetan orain arte
azaldutako adierazpen berberak erabiliz izendatuko litzateke.
3.2. Peptidasen funtzio biologikoak
3.2.1. Peptidoen eraldaketa metabolikoa
Edozein organismotan, honek koordinatuta funtzionatu dezan, zelulen arteko
komunikazioa ezinbesteko baldintza da. Komunikazio hau hiru sistema nagusiren bitartez
egiten da: bitartekarien sistema, sistema hormonala eta nerbio sistema. Hiru sistemek
erabilitako molekula mezularien artean peptido bioaktiboak oso garrantzitsuak dira, eta
hauekin batera sistema peptidiko ezberdinak osatzen dituzten entzima eta hartzaileak
(Turner, 1986, 1987).
Sistema peptidikoen kontrola oso konplexua da, ekintza anitza eta ezberdina da,
baina oso moldagarria bai funtzio fisiologikoaren arabera baita zelularen arabera ere.
Beraz, entzima hidrolizatzaile espezifikoek peptido bioaktiboak erregulatzen dituzte
(Hallberg et al., 2005).
Peptidasen ekintza biologikorik ezagunena peptido bioaktiboen eraldaketa
metabolikoa da. Euren aktibitate hidrolitikoari esker peptidasek mezulari peptidiko hauen
ekintzak modulatu ditzakete, bai forma aktiboak ekoiztuz zein hauek inaktibatuz (Antczak
et al., 2001). Zeregin honetan arduradun nagusiak mintz zelularrean kokatzen diren
peptidasak direla adierazi izan da (Turner, 1986, 1987). Gune katalitikoa zelularen
kanpoaldera zuzenduta dutelarik, ektopeptidasa hauek peptidoa eta hartzailearen arteko
8 EC: Enzyme Commission

14
Sarrera
elkarrekintza eta azken finean mezulariaren funtzioa erregulatzen dute (Turner, 1986,
1987; Taylor, 1993).
3.2.2. Peptidasen ekintza intrakrinoa
Hala ere, entzima hauek bai zelula barneko organuluen mintzei lotuta eta bai forma
solugarri bezala, zitoplasman, gune extrazelularrean, odolean eta gorputzeko beste
jariakin ezberdinetan aurki daitezke (Mantle, 1992; O’Cuinn, 1998; Gil et al., 2001; Irazusta
et al., 2004; Valdivia et al., 2004; Larrinaga et al., 2005). Kokagune hauetan peptidasek
izan dezaketen funtzioaren inguruan, hipotesi ugari daude (1.3. Irudia).
1.3. Irudia: Peptidasen kokagune ezberdinak eta beraien funtzioa gune hauetan. MP: mintzeko peptidasa; PS: peptidasa solugarria, zitosolekoa zein kanpo gunera askatua; N: nukleoa eta M: mintza.
Alde batetik, ektopeptidasekin gertatzen den bezala, zelula barneko forma
solugarriak gune extrazelularrera jariatu daitezkeela deskribatu da eta bertan peptidoen
andeakuntzan parte har dezaketela (Balogh et al., 1998; Shrimpton et al., 2002). Bestetik,
duela gutxi arte peptidasa hauen funtzioa zelula barnean proteinen katabolismoaren
azkeneko etapetara mugatzen dela uste da (Mantle, 1992; O’Cuinn, 1998). Azken finean,
peptido bioaktiboen ekintza zelula kanpoaldean gertatuz gero, ez luke zentzu handirik
zelula barneko peptidasek hauen erregulazioan parte hartzea. Baina azkeneko
hamarkadan, funtzio endokrino, parakrino eta autokrinoaz gain, mezulari peptidiko hauek

Sarrera
15
zelula barnean ekintza izan dezaketela deskribatu da. Ekoiztutako zelulan zuzenean
eraginez edota kanpotik barneratuta, peptido hauek zelula barneko hartzaileei lotu eta
funtzio “intrakrino” ezberdinak betetzen dituzte (zelula hazkundea, adibidez) (Re, 2002;
2003; Re et al., 2006). Hau honela izanik, zelula barneko peptidasek proteinen
birziklaketaz gain mezulari intrakrino hauen erregulazioan parte har lezaketela proposatu
da (Ferro et al., 2004).
3.2.3. Peptidasen ekintza batzuk
Euren aktibitate katalitikoari esker baita ere, peptido bioaktiboak andeatzeaz gain
peptidasek zelulaz kanpoko matrizearen degradazioan parte hartzen dute, beste proteasa
askok egiten duten bezala. Honela, ehunen garapenean, hantura prozesuetan eta prozesu
neoplasikoetan garrantzia dutela ikusi da (Carl-McGrath et al., 2006).
Baina peptidasen ekintza biologikoak ere bere aktibitate entzimatikoaz
independenteak izan daitezke (Antczak et al., 2001; Carl-McGrath et al., 2006). Alde
batetik, zelulen migrazioan beharrezkoa den molekula ezberdinen arteko ezagutzarako,
peptidasa batzuk atxikitze molekula bezala joka dezaketela frogatu da, metastasia
bezalako prozesuetan parte hartuz (Riemann et al., 1999; Carl-McGrath et al., 2006; Pro
et al., 2006). Bestetik, peptidasa batzuk hartzaile funtzioa ere egiten dutela deskribatu da.
Hain zuzen ere, tesi honetan ikertu diren ektopeptidasa askoren egituran aminoazido
gutxiko zitoplasma barneko domeinua deskribatu izan da. Dirudienez, peptidasa motaren
arabera molekula ezberdinekin elkarrekinez entzima hauek zelula barneko seinale-
transdukzioa eragin dezakete, zelula hazkundea eta migrazioa bezalako funtzioak
baldintzatuz (Antczak et al., 2001; Nanus, 2003; Sumitomo et al., 2005).
3.3. Peptidasak fisiologian eta patologian
Beraz, peptidasek funtsezko zeregina betetzen dute zelula arteko komunikazioan,
funtzio fisiologiko ezberdinak baldintzatuz eta prozesu patologiko ezberdinen genesian eta
eboluzioan eraginez. Nerbio sistema zentralean adibidez, neuropeptidoak eraldatuz,
oroimena, portaera eta nozizepzioa bezalako funtzioetan parte hartzen dutela deskribatu
da (Hui, 2007). Oso ezaguna da baita ere sistema kardiobaskularrean eta giltzurrun
funtzioan duten garrantzia, errenina-angiotentsina sistemaren bide metabolikoak
peptidasek erregulatzen baitute (Fyhrquist et al., 2008). Era berean, gogo aldartearen eta
oroimenaren asalduretan, bihotzeko eta giltzurruneko gaixotasunak, gaixotasun

16
Sarrera
endokrino-metabolikoak eta beste hainbat prozesu patologikotan peptidasen asaldurak
deskribatu dira (Wiedeman et al., 2003; Hui, 2007; Leung, 2007).
Are gehiago, gaur egun terapeutika arloan peptidasen inhibitzaile ezberdinak
erabiltzen dira. ACE-ren inhibitzaileak adibidez (captopril, enalapril, lisinopril), oso erabiliak
dira gaixotasun kardiobaskularren tratamendurako (Wilcox et al., 1988). Duela urte gutxitik
ere DPPIV/CD26-aren inhibitzaileak (gliptinak) erabilgarri daude 2. motako diabetearen
tratamendurako (Wiedeman et al., 2003; Drucker et al., 2006). Hauez gain, beste
batzuekin ikerketa nahiko aurreratuak daude, adibidez, PEP-aren inhibitzaileak garuneko
endekapenezko gaixotasunen tratamenduan, NEP eta APN-aren inhibitzaileak analgesian,
etab. (García-Horsman et al., 2007; Noble et al., 2007).
Honen guztiaren inguruan, gure ikerketa taldearen esperientzia urteetakoa da.
Adibidez, neuropeptidoen metabolismoa aztertuz, peptidasek gizakia eta beste ugaztunen
ehun ezberdinen garapenarekin eta ugalketa funtzioarekin erlazioa daukatela frogatu da
(De Gandarias et al., 1999; 2000; Irazusta et al., 2002, 2004; Agirregoitia et al., 2003a,
2003b; 2007; Valdivia et al., 2004). Ildo honi eutsiz, proliferaziozko asalduretan peptidasa
ezberdinen aktibitateak eta adierazpena ikertzen hasi zen eta hurrengo ataletan sakonago
aztertuko den bezala, tumore mota ezberdinetan peptidasen aldaketak ikertzen ari gara
(Varona et al., 2007; Larrinaga et al., 2007; Blanco et al., 2008; Perez et al., 2009;
Larrinaga et al., 2010a, 2010b, 2011; Varona et al., 2010; Blanco et al., 2010).
3.4. Peptidasen ekintza tumoreen biologian
3.4.1. Peptidoak eta peptidasak tumoreen biologian
Tumorea edo neoplasia ezohizko ehun masa bat da, zeina, inguruko ehun
normalarekin koordinatu gabe hazten den, honek baino hazkunde handiagoa duelarik.
Asaldura molekular ezberdinek, mutazioen pilaketa dela medio, tumorearen sorrera,
garapena eta hedaketa baldintzatuko duten asaldurak eragiten dituzte. Kartzinogenesi
prozesuan, hazkunde zelularraren erregulazio bide normaletatik ihes egiteko gaitasuna eta
gaitasun angiogenikoa erakusten dute zelula neoplasikoek. Alegia, minbizi-zelulen izaera
autonomoa da beraien ezaugarrietako bat. Honez gain, ehunak lokalki edo distantzian
inbaditzeko gaitasuna ere lortzen dute (Hanahan et al., 2000, 2011; Carl-McGrath et al.,
2006).
Zelula neoplasikoek bere izaera autonomoa bermatuko duten gaitasun ezberdinak
eskuratzen dituzte. Alde batetik, euren hazkunde seinaleak ekoizteko gai dira eta baita
inguruko zelulek bidalitako hazkunde seinaleei era berezian erantzuteko gai ere. Bestetik,

Sarrera
17
programatutako heriotza zelularretik (apoptosia) ihes egiteko gaitasuna eskuratzen dute.
Hauekin erlazionatuta, zelulak hilezkor bihurtzen dira eta mugarik gabeko hazkundean
hasten dira. Honi guztiari, odol-hodi berrien eraketa sustatzeko gaitasuna gehitzen zaio
(angiogenesia), eta honela, era autonomoan hazi eta biziraun daitekeen minbizi-zelulez
eraturiko masak ager daitezke edozein ehun motetan (Hanahan et al., 2000).
Nahiz eta minbiziaren ikerketan lan ugarienak zelula barneko seinaleztapen
zirkuituen ulermenean oinarritu diren, guzti honek zentzua galduko luke minbizi-zelulen
mikroingurumeneko gertakariak ulertzen ez badira (Carl-McGrath et al., 2006). Azken
finean, kanpoko zein barneko gertakariak elkarrekin erlazionatuta daude eta zitokina,
hormona eta hazkunde faktore asko funtsezko seinale molekularrak dira goian azaldutako
prozesuetan. Hauen artean era parakrino, autokrino edo intrakrinoan jokatu dezaketen
molekula peptidiko askok zelula hazkundean, apoptosiaren inhibizioan eta angiogenesian
eragin zuzena dutela deskribatu da (Heasley, 2001; Re et al., 2006). Minbizi-zelulek
beraien hazkundea bermatuko duten hazkunde faktore peptidiko berriak ekoiztu, jariapena
handitu, edota hauekiko sentikortasuna handitu dezakete, ezagunak direlarik tumore
askotan honelako kiribilen sorrera. Honez gain, hazkundearen aurkako peptidoekiko
erresistentzia edo sentikortasun eza ere garatzen dute (Hanahan et al., 2000, 2011).
Gehien ezagutzen diren hazkunde faktoreez gain, adibidez hazkunde faktore
epidermikoak (EGF), beste peptido askok ere kartzinogenesian garrantzia dutela frogatu
da. Euren artean tesi honetarako aukeratutako peptidasen substratu natural asko,
adibidez, angiotentsinak, zininak, opioideak, oxitozina, basopresina, endotelina-1,
bonbesinak, etab. (Carl-McGrath et al., 2006).
Peptido bioaktibo hauen erregulazioari dagokionez, mikroingurumen honetan
peptidasek eraginen bat izan behar dutela argi dago, peptidoen metabolismoaren
arduradunak baitira.
3.4.2. Peptidoen erregulazioa ulertzeko eredua
Autore batzuk oinarrizko eskema bat proposatu dute peptidoen erregulazioaren
eredu moduan (Antczak et al., 2001; Nanus, 2003; Carl-McGrath, 2006).
Tumore motaren arabera, peptidasaren funtzioa galdua ala gehitua topa genezake.
Paradoxikoa dirudien arren, egoera biek fenomeno neoplasikoak eragin ditzakete. Alde
batetik, peptidasa zehatz baten adierazpena edo funtzioa galdua badago, eta bere
substratu peptidikoak(-ek) zelularen biziraupena sustatu edo zatiketa zelularra, zein
angiogenesia estimulatzen badu, kartzinogenesia eragingo du. Gerta liteke baita ere,

18
Sarrera
substratua hazkunde zelularra inhibitzen duen propeptidoa izatea, alegia peptidasa hori
beharrezkoa izatea peptidoa aktibo bilakatzeko. Ondorioz, hazkunde inhibizioko seinalerik
gabe, tumorea hazi egingo da. Edo alderantzizkoa gerta liteke, minbizi-zelulek peptidasa
bat de novo adierazten dutenean edo adierazpena handitzean, beraien substratua
hazkunde inhibizio seinalea bada, hazkundeak ez du gelditzerik izango. Era berean,
substratua hazkunde faktore propeptidoa bada, sortutako peptidoak kartzinogenesia
eragingo du (Antczak et al., 2001; Nanus, 2003; Carl-McGrath, 2006).
Eredu honen adierazle diren ikerlan ezberdinak daude. Prostatan adibidez,
endotelina-1 (ET-1) eta bonbesinak minbiziaren aldi ezberdinetan eragiten dutela
deskribatu da, tumore zelulen garapena, apoptosi aurkako ekintza eta zelula migrazioa
eragin baitezakete. Peptido hauek andeatzen dituen NEP-aren adierazpena asko
murrizten da minbizi mota honetan, galera honek peptido hauen mailak emendatzea
dakarrelarik (Sumitomo et al., 2005). Era berean, prostatako minbizian baita ere, ET-1
ekoizten duen peptidasaren (endotelina eraldatzen duen entzima 1 edo ECE-1)
adierazpena gehituta dago, gaitasun kartzinogenikoa duen peptido honen mailak handituz
(Dawson et al., 2006). Beste adibide adierazgarri bat areako minbizian ikusi da duela
gutxi. Agresibitate nabarmeneko tumore honetan angiotentsina II-a (Ang II) eta hau
ekoizten duen entzima eraldatzailearen (ACE) adierazpena handitua dago. Berriz, peptido
hau Ang 1-7-an eraldatzen duen ACE2-arena murriztua dago (Zhou et al., 2009). Egoera
honek, gaitasun proliferatiboa eta angiogenikoa duen Ang II-aren eta propietate
antiproliferatiboa eta apoptotikoa omen duen Ang 1-7-aren arteko desoreka eragiten du
(Deshayes et al., 2005; Ruiz-Ortega et al., 2007; Zhou et al., 2009).
Hala ere, peptidoak eta hauek hidrolizatzen dituzten peptidasak prozesu
neoplasikoekin erlazionatzen dituen eredu hau lagungarria izan daitekeen arren,
peptidasek kartzinogenesian dituzten eginkizunak ulertzeko faktore asko kontuan hartu
behar dira. Alde batetik, peptidasen adierazpena tumore bakoitzean ezberdina izan
daiteke. Adibidez, prostatako minbizian adierazpena murriztua duen NEP eta handitua
azaltzen duen ECE-1, justu alderantziz azaltzen dira urdailekoan eta biriketakoan
hurrenez hurren (Ahmed et al., 2000; Antzcak et al., 2001; Carl-McGrath et al, 2006).
Bestetik, peptidasa gehienak substratu natural bat baino gehiago eraldatzeko gai dira. Era
berean, tumore bakoitzaren mikroingurumena ezberdina da, bertan ekoiztutako peptido
guztiak behar bada, ez dira jariatuko, ez dira proportzio berdinetan agertuko ere, eta agian
ez du efektu berbera eragingo, hartzaile mota, kopurua eta funtzioa ere aldagarria izan

Sarrera
19
daitekeelako. Beraz, tumore mota bakoitzak (edo talde bakoitzak) peptidoen erregulazioari
dagokionez bere ezaugarri espezifikoak izango ditu (Nanus, 2003).
3.4.3. Peptidasen funtzioa hazkuntza prozesuetan
Peptido bioaktiboen erregulazioaz gain, kontuan hartu behar da peptidasek euren
aktibitate katalitikoari esker tumorearentzako beharrezkoak diren beste zeregin batzuk
betetzen dituztela. Minbizia sortu den organoan zehar hazi, inguruko ehunak infiltratu eta
distantzian hedatzeko, mintz basalak eta zelulaz kanpoko matrizearen endekapena
beharrezkoa da. Zeregin honetan entzima proteolitiko ezberdinen arteko koordinazioa
ematen da, euren artean peptidasak daudelarik (Carl-McGrath et al., 2006). Melanoma
adibidez, oso pronostiko txarreko minbizia da, nahiko erraz migratu eta metastasia
eragiten duelako. Melanozito normalek peptidasa hau azaltzen ez duten arren, melanoma
zelula batzuk APN adierazten dutela frogatu da. Matrizeko metaloproteasekin (MMP)
koordinatuz, APN-ak kolagenoa hidrolizatzen du (Petrovic et al., 2004). Gainera,
matrizearen suntsiketa ezinbestekoa da bertatik endotelioko zelulak migratu eta odol-hodi
berriak sortu ahal izateko, minbizien neoangiogenesi prozesuan (Hanahan et al., 2000),
zeinetan APN-aren eginkizuna oso garrantzitsua dela deskribatu den (Pasqualini et al.,
2000; Petrovic et al., 2004). Beste peptidasa batzuetan deskribatu den bezala, APN-aren
adierazpena positiboki korrelazionatzen da tumore askoren pronostiko txarrarekin, alegia
tumoreen hazkuntzarekin, ehunak inbaditzearekin eta migratzeko gaitasunarekin (Petrovic
et al., 2004; Carl-McGrath et al., 2006).
Orain aipatutako mekanismoez gain, edo hauekin batera, peptidasek euren
aktibitate katalitikoaz independenteak diren ekintzen bitartez eragiten dute hazkuntza
prozesuetan. Berriro ere prostata eredutzat hartuz, Sumitomo et al.-ek (2005) hazkuntza
zeluletan NEP-ak zelulen hazkundea eta migrazioa inhibitzen duela ere frogatu zuten,
entzimaren domeinu intrazitoplasmikoak beste molekula batzuekin izandako
elkarrekintzaren ondorioz (PTEN proteina tumore supresorea adibidez). Prostatako
minbizian NEP-aren adierazpena galtzean, ez da bakarrik peptidoen erregulazioa
asaldatzen, baizik eta hartzaile bezala duen funtzioa galdu eta zelula proliferazioa eta
hedapena kontrolpetik kanpo gelditzen dira. Bestalde, minbizi zelulek euren migrazioan,
bai lokalki zein distantzian, matrizeko proteinak, odol-hodietako zeluletako proteinak, etab.
ezagutzen dituzte eta hauei lotzen zaizkie. Honetarako, atxikitze molekula funtzioa izango
luketen peptidasa ezberdinak erabilgarriak direla deskribatu da. Adibidez, DPPIV eta APN,
minbizi zeluletan adierazi eta matrizeko kolagenoari lotzea posible da. Eta alderantziz,

20
Sarrera
minbizi zelulen gainazaleko proteina batzuk beste ehunetako basoetako endotelioan
dauden peptidasa hauei lotzen zaizkie, honela, minbizi zelulak itsatsi eta metastasia
gertatu daiteke (Riemann et al., 1999; Carl-McGrath et al., 2006; Pro et al., 2004).
Peptidasek prozesu tumoralen aldi ezberdinetan eginkizun anitzak izan ditzaketela
deskribatu da. Alegia, hurrengo ataletan ikusiko den bezala, hodi digestiboko minbizietan
ere proteasa ezberdinen aldaketa nabarmenak azaldu dira eta hauen artean peptidasek
funtzio garrantzitsua izan dezaketela deskribatu da.
3.5. Ikertutako peptidasak
Sarrera honen aurreko azpiataletan, aipatu dira peptidasa funtzioa duten hainbat
entzima, baita beraien eragina edo erlazioa prozesu patologikoetan ere, adibidez,
minbizian. Deskribaturik daude entzima batzuen erabilera minbizi-markatzaile gisa, hala
nola, NEP/CD10, DPPIV/CD26, ACE/CD143, APA/gp160 eta APN/CD13 (Murakami et al.,
2005; Lui et al., 2007; Havre et al., 2008). Arrazoi honengatik peptidasa hauek ikertu izan
dira, koloneko minbiziaren bilakaeran izan dezaketen erlazioaren bila.
Aipatutako bost peptidasak, entzima mota honetarako proposatu den sailkapen-
sistemako Familia ezberdinetako partaide dira, eta entzimen talde anitza izanik, substratu
ezberdinak erabiltzen dituzte haien artean. Baliteke beraz, beste peptidasa batzuk ere,
lehen aipatu diren prozesuen eragile izan arren, egun ez ezagutzea, eta gerora
markatzaile edo itu modura erabiltzea diagnosian edo beharbada koloneko minbiziaren
tratamenduan orain daudenen lagungarri gisa.
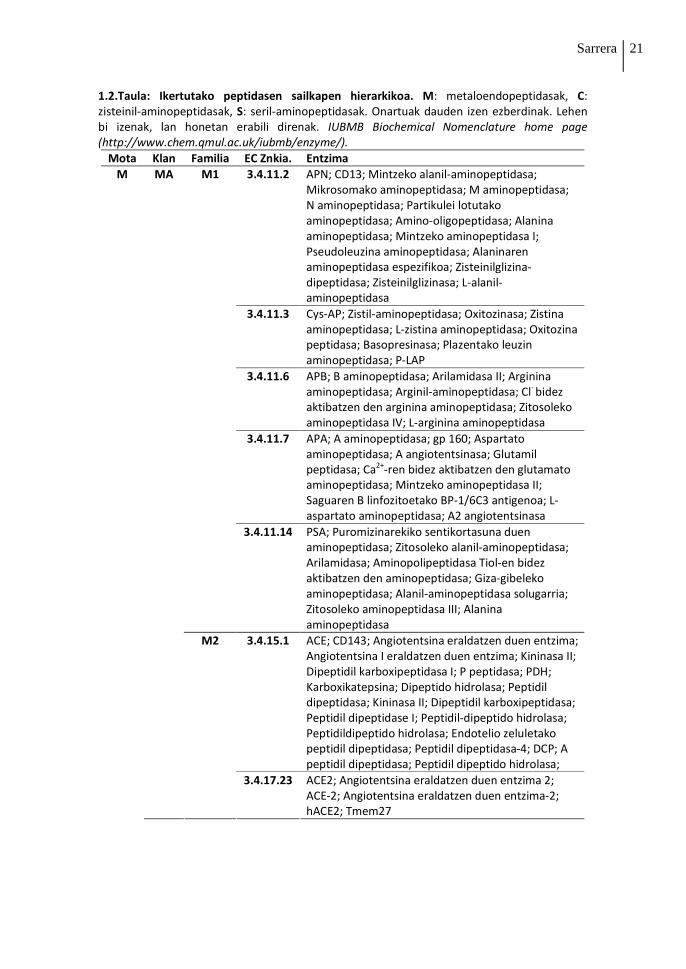
1.2. Taulan agertzen dira bost entzima markatzaileak zein prozesu patologikoetan
garrantzitsuak izan daitezkeen peptidasa gehiagoren sailkapena lehen aipatutako IUBMB-
etik hartua. Bestalde, 1.4. Irudian agertzen da peptidasa bakoitzaren kokapena zelulan eta
zein kromosometan kodetzen den beraien genea eta genearen izena.
Sarrera
21
1.2.Taula: Ikertutako peptidasen sailkapen hierarkikoa. M: metaloendopeptidasak, C: zisteinil-aminopeptidasak, S: seril-aminopeptidasak. Onartuak dauden izen ezberdinak. Lehen bi izenak, lan honetan erabili direnak. IUBMB Biochemical Nomenclature home page
(http://www.chem.qmul.ac.uk/iubmb/enzyme/).
Mota Klan Familia EC Znkia. Entzima
3.4.11.2 APN; CD13; Mintzeko alanil-aminopeptidasa; Mikrosomako aminopeptidasa; M aminopeptidasa; N aminopeptidasa; Partikulei lotutako aminopeptidasa; Amino-oligopeptidasa; Alanina aminopeptidasa; Mintzeko aminopeptidasa I; Pseudoleuzina aminopeptidasa; Alaninaren aminopeptidasa espezifikoa; Zisteinilglizina-dipeptidasa; Zisteinilglizinasa; L-alanil-aminopeptidasa
3.4.11.3 Cys-AP; Zistil-aminopeptidasa; Oxitozinasa; Zistina aminopeptidasa; L-zistina aminopeptidasa; Oxitozina peptidasa; Basopresinasa; Plazentako leuzin aminopeptidasa; P-LAP
3.4.11.6 APB; B aminopeptidasa; Arilamidasa II; Arginina aminopeptidasa; Arginil-aminopeptidasa; Cl- bidez aktibatzen den arginina aminopeptidasa; Zitosoleko aminopeptidasa IV; L-arginina aminopeptidasa
3.4.11.7 APA; A aminopeptidasa; gp 160; Aspartato aminopeptidasa; A angiotentsinasa; Glutamil peptidasa; Ca2+-ren bidez aktibatzen den glutamato aminopeptidasa; Mintzeko aminopeptidasa II; Saguaren B linfozitoetako BP-1/6C3 antigenoa; L-aspartato aminopeptidasa; A2 angiotentsinasa
M1
3.4.11.14 PSA; Puromizinarekiko sentikortasuna duen aminopeptidasa; Zitosoleko alanil-aminopeptidasa; Arilamidasa; Aminopolipeptidasa Tiol-en bidez aktibatzen den aminopeptidasa; Giza-gibeleko aminopeptidasa; Alanil-aminopeptidasa solugarria; Zitosoleko aminopeptidasa III; Alanina aminopeptidasa
M2 3.4.15.1 ACE; CD143; Angiotentsina eraldatzen duen entzima; Angiotentsina I eraldatzen duen entzima; Kininasa II; Dipeptidil karboxipeptidasa I; P peptidasa; PDH; Karboxikatepsina; Dipeptido hidrolasa; Peptidil dipeptidasa; Kininasa II; Dipeptidil karboxipeptidasa; Peptidil dipeptidase I; Peptidil-dipeptido hidrolasa; Peptidildipeptido hidrolasa; Endotelio zeluletako peptidil dipeptidasa; Peptidil dipeptidasa-4; DCP; A peptidil dipeptidasa; Peptidil dipeptido hidrolasa;
M MA
3.4.17.23 ACE2; Angiotentsina eraldatzen duen entzima 2; ACE-2; Angiotentsina eraldatzen duen entzima-2; hACE2; Tmem27

22
Sarrera
M13 3.4.24.11 NEP; CD10; Endopeptidasa neutroa; Neprilisina; Endopeptidasa 24.11; Giltzurruneko ertz eskuilakarako peptidasa neutroa; Endopeptidasa 2; CALLA antigenoa; Endopeptidasa; Mintzeko metaloendopeptidasa; Giltzurruneko ertz eskuilakarako proteinasa neutroa; Endopeptidasa-2; CALLA Glikoproteína; CALLA; Metaloendopeptidasa neutroa; Mintzeko metaloendopeptidasa; Endopeptidasa neutroa 24.11
MH M18 3.4.11.21 Asp-AP; Aspartil aminopeptidasa
C CF C15 3.4.19.3 PG I; Piroglutamil-peptidasa I; 5-oxoprolil-peptidasa; Pirasa; Piroglutamato aminopeptidasa; Piroglutamil aminopeptidasa; L-piroglutamil peptido hidrolasa; Pirrolidona-karboxil-peptidasa; Pirrolidona-karboxil peptidasa; Pirrolidonil peptidasa; Piroglutamidasa; Pirrolidonekarboxilil peptidasa; L-pirrolidonakarboxil peptidasa;
3.4.14.5 DPPIV; CD26; Dipeptidil-peptidasa IV; Dipeptidil aminopeptidasa IV; Xaa-Pro-dipeptidil-aminopeptidasa; Gly-Pro naftilamidasa; Postprolina dipeptidil aminopeptidasa IV; Linfozitoetako CD26 antigenoa, Glikoprotena GP110; Pep X; Dipeptidil dipeptidasa IV; Glizilprolina aminopeptidasa; X-prolil dipetidil aminopeptidasa; Leukozitoetako CD26 antigenoa; X-PDAP; Glizilprolil dipeptidil aminopeptidasa; Dipeptidil-peptido hidrolasa; Glizilprolil aminopeptidasa; Dipeptidil-aminopeptidasa IV; Amino azil-prolil dipeptidil aminopeptidas; T zelulen aktibatzaile den Tp103 molekula
S SC S9
3.4.21.26 PEP; Prolil oligopeptidasa; Post-prolina ondorengo entzima; Prolinaren endopeptidasa espezifikoa; Post-prolina endopeptidasa; Endoprolilpeptidasa; Prolil endopeptidasa

Sarrera
23
3.5.1. Puromizinarekiko sentikortasuna duen aminope ptidasa (PSA)
Metalopeptidasen M1 familiakoa da entzima hau. Zn2+ menpeko sialoglikoproteina
bat da. Co2+ bidez aktibatzen den arren, kasu honetan, sortzen den konplexua ez da hain
egonkorra.
PSA aminopeptidasaren ekintza nahiko zabala da. Oligopeptido ugariren N-
muturreko alanina hondarrak hidrolizatuko ditu lehenik, 13 aminoazido baino gutxiago
baditu bederen. Baina, hidrolizatu ditzake ere afinitate handiz leuzina, arginina, tirosina,
fenilalanina eta metionina, beste batzuen artean (Taylor, 1993). Prolina aminoazidoa
baldin bada azkena edo azkenaurrena dagoena, PSA entzimak ez du eraginik izango
oligopeptido horretan (Dando et al., 1997). Bere substratuen artean, entzefalinak,
dinorfinak eta prodinorfina eratorriak, etab. daude (Gros et al., 1985b; McLellan et al.,
1988).
Burmuinean deskribatu da gehien bere agerpena. Hala ere, gibelean, giltzurrunean,
muskulu eskeletikoan, barrabiletan eta heste meharrean ere adierazten da (De Souza et
al., 1991; Hiroi et al., 1992; Tobler et al., 1997).
Peptidasen funtzio klasikoa peptido bioaktiboen andeaketa izan arren, entzima hau
nagusiki zitosolekoa izanik, ez dirudi funtzio hau denik garrantzitsuena (Tobler et al.,
1997). Alabaina, autore batzuek zelularen hazkuntzarako garrantzitsu diren oinarrizko
proteolisi prozesuetan parte hartzen duela proposatu dute (Constam et al., 1995). Honen
harira, Stoltze et al.-ek (2000) iradokitzen dutenez, exopeptidasa eta oligopeptidasa
solugarriek, turnover proteikoan gertatzen diren prozesuetan, hainbat ekintza dituzte
oligopeptidoen andeaketan. Ugal-prozesuetan ere parte har lezakeela deskribatu da
(Osada et al., 2001a, 2001b).
PSA-ren kode-genea, NPEPPS, 17. kromosoman kokatzen da. 40 kb ditu, 23
exonez eta 22 intronez osatua (Thompson et al., 1999). 100 kDa inguruko kate bakarreko
metaloproteina bat da. PSA-ren aktibitate gorena pH 7,5-an gertatzen da. Kaltzio ioia
edota tiol konposatuak entzimaren aktibatzaileak dira (Haeggström, 2004), aldiz, EDTA,
1,10-fenantrolina, amastatina, bestatina eta puromizina inhibitzaileak dira (Mantle et al.,
1983; Dando et al., 1997). Tiol konposatuengatiko aktibazioa eta puromizinarekiko duen
sentsibilitatea dira entzima honen ezaugarri ezberdintzaileak, honengatik ere entzimaren
izena. Ugaztunen zelulatako entzima zitosolikorik ugariena dela esan liteke (Mantle,
1992).

24
Sarrera
3.5.2. Aminopeptidasa neutroa (APN)
PSA bezala, APN entzima M1 familiakoa da, metalopeptidasa bat. II. motako
mintzarteko peptidasa bat da (McLellan et al., 1988; Riemann et al., 1999). Glikosilatua
agertzen da kasu askotan, eta gune-aktiboa zelularen kanpo aldera begira izaten du, Zn2+-
a bertan kokatzen den metal bakarra izanik. Gainera, ektoentzima honen bi azpiunitate ez-
kobalenteki lotzen dira homodimeroak osatuz, eta 150 kDa-eko pisu molekularreko
molekula bat osatzen du (Riemann et al., 1999). Klasikoki mintzeko peptidasa bezala
deskribatu arren, azken urteetan APN solugarria plasman ere deskribatu dute (Erbeznik et
al., 1997; van Hensbergen et al., 2002; Murakami et al., 2005).
Entzima hau ugaria da heste mehearen, plazentaren eta giltzurrunaren ertz
eskuilakarako zelulen mintzean. Gibelean eta biriketan ere agertzen da, nahiz eta APN-
arekiko interesa burmuineko entzefalinen inaktibazio eta hidrolisiarengatik sortu zen.
Entzefalinaz gain, angiotentsinak, bradikininak, zitokinak eta kimiokinak dira aktibitateari
dagokionez erregulatzen dituen peptidoak (Gros et al., 1985a; Matsas et al., 1985).
Entzimaren ekintzaren ondorioz, 3 eta 16 aminoazidotako peptidoaren amino
muturreko azken aminoazido neutroa askatzen da. Azken aminoazido hau alanina bat
bada, jarduera azkarrago joango da prolina bat bada baino, hala ere, biak hidrolizatzeko
gai da. Prolinaren aurreko animoazidoa hidrofoboa bada dipeptidoa apurtuko da
(McDonald et al., 1986). Gainera, dipeptido ugari hidrolizatzeko gai da APN-a, baina ez
aspartil, glutamil, histidil, glizil edo balil hondarrak badaude N-muturrean (O´Cuinn, 1998).
ANPEP da entzima hau kodetzen duen genearen izena, 15. kromosomaren beso
luzean kokatzen da eta 35 kb ditu. APN-aren adierazpena aldakorra da hazkuntzan zehar
zein zelularen arabera. Ezaugarri hau, bere ekintzaren “plastizidadearen” isla da (Riemann
et al., 1999). Minbizi mota batzuen histologi-markatzaile gisa erabiltzen da (APN/CD13),
baita pronostikorako ere (Bogenrieder et al., 1997; Ishii et al., 2001; Hashida et al., 2002;
Murakami et al., 2005). Gainera, sistema immunean eta angiogenesian duen funtzioa
sakonki ikertu da (Riemann et al., 1999; Bhagwat et al., 2001).
APN-aren gehiengo aktibitatea pH 7-an gertatzen dela deskribatu da. DTT-a
aktibatzailea da, eta kelatzaileak, tiol-erreaktiboak, bestatina eta batez ere amastatinak
entzima inhibitzen duten molekulak dira (O´Cuinn, 1998; Riemann et al., 1999; Sjöstrom et
al., 2000; Turner, 2004a).

Sarrera
25
3.5.3. Aminopeptidasa basikoa (APB)
APB entzima ere metalopeptidasen M1 familiako kide da. Gainontzeko kideek
bezala, peptidasak Zn2+-ari lotzeko gunea du (Suda et al., 1976; Ocain et al., 1987). Ioi
honi dion loturagatik, kelatzaileak inhibitzaileak izango dira, baita, bestatina, tiol-
erreaktiboak, arfamenina A eta B ere (Hopsu et al., 1966; Cadel et al., 1995; O´Cuinn,
1998; Foulon et al., 2004). 72 kDa dituen entzima honen aktibatzaileak ioi kloruroak (Cl-)
izango dira.
Aminopeptidasa basikoa ere baderitzo, izan ere, peptidoen amino muturreko azken
aminoazido basikoak hidrolizatzen ditu. Lisina edo arginina aminoazidoak askatuko ditu,
betiere aurrekoa prolina ez bada (Gainer et al., 1984; Cadel et al., 1995). Substratu
naturalen artean, bradikininak, entzefalinak, somatostatina, angiotentsina III eta kalidinak
daude (McLellan et al., 1988; Foulon et al., 2004).
APB-aren adierazpena ugaztunetan oso ugaria da eta ehun mota anitzetan
deskribatu da (Foulon et al., 1996). Zelulako kokapenari dagokionez, hasiera batean
zitoplasmako entzima gisa deskribatu zen, gerora erretikulu endoplasmatikoan aurkitu da
eta zelularen kanpo matrizera jariatzen dela ere jakina da (Lauffart et al., 1988; Tanioka et
al., 2003; Cadel et al., 2004). Bestalde, bere aktibitatea mintz plasmatikoan ere neurtu
izan da. Ezaugarri hauek guztiak kontuan hartuta, esan daiteke oso molekula ubikuoa dela
(Varona et al., 2007).
pH tarte zabalean lan egiteko gai da, hala ere, pH optimoa 7 da. Ezaugarri hau eta
bere adierazpen zabala kontuan izanik, prozesu fisiologiko ezberdinetan parte har
dezakeela uste da. Adibidez, hanturazko prozesuetan, bere substratuak diren kalidinak,
entzefalinak eta somatostatinak funtzio garrantzitsua dute. Autore batzuen iritziz, proteinen
itzulpen osteko heltzean eta mintz plasmatikoko hainbat prozesuren erregulazioan parte
hartzen du (Foulon et al., 2004).
APB-aren kode-geneak, RNPEP, exon ugari ditu. 24kb ditu, eta 1. kromosomako
beso luzean kokatzen da. 1997an identifikatu zuen Aurich-Costa et al.-ek. Gen honen
azken produktua, proteina monomeriko bat da (Fukasawa et al., 1996; Cadel et al., 1997).
3.5.4. Aspartil aminopeptidasa (Asp-AP)
M18 familiako metalopeptidasa bat da Asp-AP. Entzima honen kopurua, zitosoleko
proteina solugarrien % 0,1-a da hainbat ehunetan, honegatik, zelula barneko peptidasa
nagusi bezala kontsideratu beharko litzateke. Entzima solugarria izan arren, mintzari
loturiko Asp-AP aktibitatea deskribatu izan da (Ramírez-Expósito et al., 2000; Varona et

26
Sarrera
al., 2007; Blanco et al., 2008). Pisu molekular handia du beste peptidasa solugarrien
antzera, oligomero bat osatuz ageri da normalean. Oligomeroa da eta 53 kDa-eko 8
azpiunitate berdinek osatzen dute.
Kopuru handian agertzen den peptidasa bat da eta gainera, ehun mota gehienetan
deskribatu da. Hau honela, proteinen eta peptidoen azken prozesu metabolikoan funtzio
garrantzitsuren bat bete dezakeela pentsa daiteke (Wilk et al., 1998, 2002). Alabaina,
beste funtzio espezializatuagoak edo zehatzagoak bete ditzakeela ere uste da,
angiotentsina II, kolezistokinina-8 edota K neuropeptidoa bezalako peptido bioaktiboen
metabolismoan parte hartzen baitu (Kelly et al., 1983; Wilk et al., 1998). Aminopeptidasa
bat izanik, N-muturreko aminoazido azidoetan du eragina, hots, azido glutamikoa edo
aspartikoa (hobetsiena) hidrolizatzen ditu, lehentasuna izanik aurreko aminoazidoak
neutroak edo hidrofobikoak badira.
DNPEP genea da Asp-AP entzima kodetzen duena. 2. kromosoman kokatzen da
gene hau, bere sekuentzia erabat kontserbatua dago, ugaztunetan deskribatu den M18
familiako peptidasa bakarra izateraino. Metalopeptidasa izanik, EDTA, 1,10-fenantrolina
eta DTT-az inhibitzen da (O`Cuinn, 1998; Sanderink et al., 1988) eta Ca2+ aktibatzailea
izango da. Entzimaren pH optimoa 7,5-ekoa da (Wilk, 2004).
1.4.Irudia: Ikertutako peptidasen azalpen grafikoa. Zelulan duten kokapena (mintzekoa edo solugarria: zitoplaman edo zelulaz kanpokoa). Genearen izena eta zein kromosometan kodetzen den genea.

Sarrera
27
3.5.5. Zistil aminopeptidasa (CAP)
Sialoglikoproteina bat da, hau ere, M1 familiako metalopeptidasa da. Bere
ekintzaren ondorioz, amino muturreko zisteina bat askatzen da substratu zabal bateko
peptidoetan, hala nola, oxitozina, basopresina, somatostatina eta angiotentsina III
(Sjöholm et al., 1967; Mitsui et al., 2004). CAP entzima, oxitozina andeatzen duen
mintzeko aminopeptidasa bakar bezala deskribatu da. Zisteinaz gain, beste aminoazido
neutro edo basiko batzuk hidrolizatzeko gai ere bada, baina jarraian duen aminoazidoaren
izaeraren arabera zein peptidoaren luzeraren arabera (Nomura et al., 2005).
50 urte baino gehiago badira haurdun dauden emakumeen sueroan CAP entzima
solugarria neurtzen dela, nahiz eta geroago II. motako mintzarteko proteina deskribatu
den. Ez soilik plazentan, baizik, beste ehun batzuetan ere deskribatu da, hala nola,
bihotzean, muskulu eskeletikoan eta burmuinean (Keller et al., 1995; Rogi et al., 1996;
Laustsen et al., 1997).
Mintzarteko egiturak, gune zitoplasmatiko bat agertzen du, mintzarteko gune txiki
bat eta tamaina handiagoko gune extrazelularra (Rogi et al., 1996; Laustsen et al., 1997).
Egitura honetan, AT4 hartzaile bezala ere identifikatu da. Hartzaileari lotzen zaion
angiotentsina IV-ak proteinaren entzima-jarduera inhibitzen du. Bestalde, CAP entzima
solugarriaren askapena, funtzio katalitikoa duten metaloproteasen ekintzaren ondorio dela
uste da. Azken hauek, karboxilo muturraren hidrolizazioa eragiten dute zirkulaziora
askatuz (Nomura et al., 2005).
Haurdunaldiko homeostasia erregulatzeaz gain, intsulinaren metabolismoaz
arduratzen den proteinetako bat dela ere jakina da (Rogi et al., 1996) eta prozesu
fisiopatologiko ugaritan erlazioa duela deskribatu da, euren artean minbizi-prozesuekin
zerikusia daukaten batzuk (Ito et al., 2001; Suzuki et al., 2003). Intsulinari dagokionez,
homona honek ektoentzimaren translokazioak nolabait erregulatzen dituela iradoki izan da
(Sumitani et al., 1997).
Beste hainbat metalopeptidasa bezala, CAP entzimaren aktibitatea agente
kelatzaileen bidez inhibi daiteke, baita 1,10-fenantrolina, katioi dibaleak, amastatina,
bestatina eta puromizina ere, nahiz eta azken bi hauek neurri txikiagoan eragin (Kandror
et al., 1994; Matsumoto et al., 2000; Nomura et al., 2004a, 2004b). Entzimaren aktibitate
gorena pH 7,4-an lortzen da eta 160 kDa-eko pisu molekularra du. 5. kromosoman
kokatzen da LNPEP genea, eta 18 exonez eta 17 intronez osatua dago.

28
Sarrera
3.5.6. Piroglutamil peptidasa I (PGI)
Piroglutamil peptidasa, omega peptidasa bat da. C15 familiako kide da entzima
solugarri hau. C Klan-ekoa izanik, zisteina baten agerpena adierazten du gune aktiboan.
Autore gehienek zitosoleko entzima gisa deskribatu arren, batzuek mintzeko aktibitatea
neurtu dute (Alba et al., 1995; Blanco et al., 2010). Piroglutamil peptidasa II-arekin batera,
2-30 aminoazido arteko peptidoen amino muturreko piroglutamil hondarrak hidrolizatzen
dituen entzima taldean sartzen da PGI-a. Nahiz eta antzeko ekintzak izan eta entzima
bera zela pentsatu hasiera batean, bi entzima hauek Klan ezberdinekoak dira. Hiru
azpimota banatu daitezke. PG I motakoa, solugarria dena, PG II motakoa, mintzekoa dena
eta tiroliberinasa izendatu duten plasmako piroglutamil peptidasa. Tiroliberinasa, PGII-
aren hidrolisiaren ondorioz askatzen den entzima dela adostu da, egitura molekular
antzekoak baitituzte (Bauer, 1995; Cummins et al., 1998).
PGI-aren ekintzak peptido bioaktiboen inaktibazioa eragiten du. Peptidasa honen
substratu naturalak lirateke, hala nola, neurotentsina, hormona luteinizatzailearen
hormona askatzailea (LHRH), tirotropinaren hormona askatzailea (TRH), bonbesina eta
leukopirokinesina (Awade et al., 1994; Mantle et al., 1992; Schmitmeier et al., 2002).
Bestalde, piroglutamina askeak berak, badu ekintza biologikoa, eta erlazioa izan dezake
zahartzapenarekin lotutako hainbat gaixotasunetan edo memoriaren galeran. Autore
batzuen ustez, PGI-aren funtzioen artean piroglutamina hauen kopurua erregulatzen
zerikusia izan dezake, peptidoen inaktibazioaz gain (Grioli et al., 1990; Cummins et al.,
1998).
Entzima monomerikoa da, 19. kromosoman kokatzen da bere genea, PGPEP1.
Pisu molekular eskasekoa, 24 kDa, eta aktibitate gorena 8,5 pH-an du. PGI-aren
adierazpena gorputz osoan hedatua dago. Hala ere, burmuin eta digestio aparatuan
nabariagoa da (Szewczuk et al., 1970), garunaren eta nerbio sistema zentralaren garapen
egokian garrantzi handiko funtzioa izanik, behintzat arratoietan (De Gandarias et al.,
2000). Arratoietan ere, PGI-aren jarduera aldaketak deskribatu dituzte burmuin-kortexean
garapenaren etapa ezberdinetan egin dituzten neurketetan (Agirregoitia et al., 2007).
PGI-ren aktibatzaileak PGII-ren inhibitzaileak dira (Cummins et al., 1998), hala nola,
entzimaren jarduera egokia izan dadin, DTT-a bezalako tiol-erreduzitzaileak, 2-
merkaptoetanola edo tris(karboxietil)fosfina behar da. Hala ere, bere hidrolisirako N-
muturreko piroglutaminaren aurreko aminoazidoaren izaera ere kontuan hartu behar da.

Sarrera
29
3.5.7. Dipeptidil peptidasa IV (DPPIV)
DPPIV/CD26-a minbizi-markatzaile gisa erabiltzen den exopeptidasa bat da. S9
familiako kide da, seril-aminopeptidasen talde anitzaren barnean. Entzimaren ekintzaren
ondorioz, dipeptido bat askatzen da oligopeptido eta polipeptidoen amino muturretik,
betiere, azkenaurreko aminoazidoa prolina bada (edo alanina, baina afinitate gutxiagoz),
eta honen aurrekoa prolina edo hidroxiprolina ez bada (O´Cuinn, 1998; Pro et al., 2004).
Azken aminoazidoaren izaera ez da garrantzi handikoa, baldin eta amino talde bat aske
badu (Yoshimoto et al., 1978).
Mintzeko glikoproteina bat da, 6 aminoazidotako amino muturra du zitoplasmarantz
begira eta hidrofobikoa den zatia mintzarteko gunean. Zelularen kanpoaldera begira
dagoena proteinaren zatirik handiena da eta bertan dago gune aktiboa. Bere amino
muturraren galerak, peptidasen ekintzaren ondorioz, suerora askatzen den entzima
solugarria bihurtzen du (Iwaki-Egawa et al., 1998). Izan ere, entzima hau zitokina eta
kimokina ugariren aktibazioaz eta inaktibazioaz arduratzen dela deskribatu da (Havre et
al., 2008), eta peptido aktiboetan duen funtzioak, zein sistema immunean duen
garrantziak, tripeptido eta dipeptidoen garraioan zerikusia izan dezakeela iradoki da
(Tiruppathi et al., 1990).
DPPIV entzima ehun mota anitzetan deskribatu da, hala ere, bere kontzentrazioa
aldakorra da ehunaren arabera. Giltzurrunean, heste mehean eta plazentan ugaria da,
mihian eta gibelean ez da horren nabaria eta bihotzean, arean eta burmuinean urria da.
Adierazpenean aurkitu daitezkeen ezberdintasun hauek, seril-proteasak kodetzen dituzten
geneen ezaugarri batengatik izan daitezke (Rawlings et al., 1991), izan ere, DPP4 geneak,
exon ugari ditu (Abbott et al., 1994). Gune aktiboa adierazten duten aminoazidoak bi exon
ezberdinetan aurkitzen dira. Beraz, deskribatu izan da, aminoazido hauetako batek
mutaziorik jasaten badu, proteina erretikulu endoplasmatikoan sortu ondoren, ez dela
mintz plasmatikora garraiatuko eta andeatua izango da (Tsuji et al., 1992).
Biokimikari dagokionez, DPPIV-aren aktibitate hobetsia pH 7,8-an gertatuko da eta
bere egitura heldu glikosilatuaren pisu molekularra 110 kDa da. Entzimaren inhibitzaileak,
PMSF (fenilmetilsufoniloaren fluoruroa), diprotina A, bazitrazina, puromizina eta agente
tiol-erreaktiboak: p-kloromerkuribenzoatoa eta N-etilmaleimida dira (O´Cuinn, 1998;
Kikkawa et al., 2005).

30
Sarrera
3.5.8. Prolil endopeptidasa (PEP)
PEP entzima, DPPIV bezala, S9 familiako kide da S Klan-ean. Entzima solugarria
izan arren, mintzeko aktibitatea ere badu. Odola eta likido zefalorakideoan deskribatu da
eta plasman sueroan baino kopuru handiagoak aztertu dituzte (Gotoh et al., 1988; O´Leary
et al., 1996; Momeni et al., 2003; Szeltner et al., 2008).
Endopeptidasa hau, peptidoen karboxilo muturreko azkenaurreko prolina hondakina
hidrolizatzeko gai da (Gass et al., 2007). Hasiera batean, umetokian deskribatu zen,
oxitozinaren entzima degradatzaile gisa (Walter et al., 1971). Egun, jakina da, hainbat
peptidoren prolina-lotura hidrolizatzen duela. Adibidez, P substantzia, basopresina,
neurotentsinak, angiotentsina I eta II, dinorfinak, bradikininak, TRH, LHRH eta alfa-
melanozitoen hormona kitzikatzailea daude bere substratu ezagunen artean (Welches et
al., 1991; Ito et al., 2004a; Ruiz-Ortega et al., 2007; Myohanen et al. 2010). Hala ere, ez
da gai tamaina handiagoa duten molekulak hidrolizatzeko. Bere espezifikotasunean,
prolina azkenaurreko gunean egoteaz gain, molekularen pisu molekularra kontuan izan
behar da.
PEP-en adierazpena gorputz osoan dago hedatua, batez ere muskuluan,
barrabiletan, giltzurrunetan eta gongoil submaxilarrean aurkitzen da (Goossens et al.,
1996b; Gass et al., 2007). Funtzio nagusiei dagokionez, aipatutako peptidoen erregulazio
mekanismoan parte hartzen duela uste da, bere ekintza, memoria eta prozesu
neurologikoekin erlazionatua dagoela ere pentsatzen da (Li et al., 1996; Agirregoitia et al.,
2003).
Peptidasa monomerikoa da, 75 kDa inguruko pisu molekularrekoa eta 7,5 pH-an du
aktibitate gorena. NaCl, potasioa, fosfatoa eta nitrato ioiak aktibatzaileak dira entzima
honentzat. Aldiz, Z-tio-prolina-tioprolinala entzima-jardueraren inhibitzailea da (Gass et al.,
2007).
6. kromosomaren beso luzean kokatzen da PEP entzimaren gene kodetzailea,
PRPEP izenekoa, Goossens et al.-ek deskribaturikoa (1996a).
3.5.9. Aminopeptidasa azidoa (APA)
M1 familiako metalopeptidasa bat da APA. Ektopeptidasa honek amino muturreko
aminoazido azidoak askatzen ditu, honegatik bere izena. Izan ere, glutamil hondarrak
hidrolizatzen ditu nagusiki eta aspartil hondarrak ere apurtzeko gai da, baina espezifizitate
baxuagoaz (Glenner et al., 1962; Wang et al., 2004). Hasiera batean Aspartil
aminopeptidasa (Asp-AP) eta APA, entzima bera zirela pentsatzen zen arren, haiei

Sarrera
31
buruzko informazio gehiago jakin zen heinean ezaugarri propioak zituztela deskribatu
zuten, sailkatzerako garaian familia ezberdinetan banatu direlarik (Wilk et al., 1990).
Aipatzekoa da, saguen B linfozitoetako APA adierazpena, ezberdintzapen antigeno gisa
erabiltzen dela, BP-1/6C3-aren berdina da.
APA, II. motako mintzarteko proteina da eta beste hainbat metalopeptidasek
bezala, Zn2+-ari lotzeko gunea du zelularen kanpo alderantz. Ugaztunetan, entzima
homodimerikoa dela deskribatu da, zeina, 140 kDa inguruko bi azpiunitatez osatua
dagoen (Wang et al., 2004). Mintzeko entzima gisa deskribatu den arren, gune solugarrian
ere aktibitatea aurkitu izan da, baita sueroan ere, mintzeko proteinaren hidrolisiaren
ondorioz kanpo gunera askatzen dena (Carrera et al., 2004; Goto et al., 2006). Bere
aktibitaterako pH-rik egokiena 7,2 ingurukoa da. Ca2+ ioiaz entzimaren aktibitatea
areagotu egiten da eta inhibitzaile nagusiak kelatzaileak dira (EDTA, EGTA eta 1,10-
fenantrolina), baita metal dibalenteak ere (Zini et al., 1996).
Ehun ugaritan adierazten da APA aminopeptidasa. Hala ere, hesteko eta
giltzurruneko ertz eskuilakaran eta plazentan nabariagoa da. Odol-hodiko zeluletan ere,
entzimaren adierazpenak garrantzi handia duela deskribatu izan da (Marchió et al., 2004;
Goto et al., 2006). Entzimaren substratu ezaguna da aspaldidanik angiotentsina II-a, odol
presioaren erregulatzailea dena (Marchió et al., 2004; Ruiz-Ortega et al., 2007). Honez
gain, hainbat neoplasia motekin erlazionatu izan da, hauen artean, prostatako, umetokiko,
giltzurruneko minbizietan eta linfoman (Fujimura et al., 2000; Ino et al., 2003; Blanco et al.,
2008; Perez et al., 2009).
APA kodetzen duen genea, ENPEP, gizakien 4. kromosomako beso luzean
kokatzen da, 100 kb inguruko genea da eta 20 exon eta hainbat intron dituena (Vazeux et
al., 1996; Wang et al., 1996).
3.5.10. Angiotentsina eraldatzen duen entzima (ACE)
Zink metalopeptidasen taldean sailkatzen da, M2 familian zehazki. Mintz
plasmatikoari loturiko I. motako ektoentzima bat da. N-muturra da masarik gehien duena
eta zelularen kanpo gunerantz du norabidetua. Mintzarteko gune bat du eta C-muturra,
zitoplasmara begira dago (Turner et al., 2002a; Carl-McGrath et al., 2006).
Bi isoforma deskribatu dira, bata somatikoa, 150-180 kDa ingurukoa dena, eta
bestea, zelula germinaletan, tamainaz txikiagoa dena. Somatikoa dena, zelula
endotelialetan, epitelialetan eta neuronetan aurkitu daiteke. Baina batez ere, odol-
hodietako zelula endotelialetan eta birikan adierazten da gehien. Giltzurruneko eta digestio

32
Sarrera
hodiko zein hesteko ertz eskuilakaran eta burmuinean ACE-ren adierazpena ere oso
nabaria da (Oparil et al., 1974b; Hirasawa et al., 2002; Larrinaga et al., 2010a). Plasmako
isoforma bat ere deskribatu da, gorputz likidoetan aurkitu dena (sueroa, likido seminala,
likido amniotikoa edo likido zefalorrakideoa). Isoforma hau, mintzeko peptidasaren
hidrolisiaz lortzen dela deskribatu da (Oparil et al., 1974a; Hooper et al., 1991, 2003;
Antczak et al., 2001; Allinson et al., 2004). Honez gain, ACE/CD143-a markatzaile gisa
erabiltzen da klinikan hainbat gaixotasunen diagnosirako (Beneteau-Burnat et al., 1991;
Baudin, 2005).
In vivo, ACE entzimaren substratu nagusiak angiotentsina I eta bradikinina dira.
Karboxipeptidasaren ekintzaren ondorioz, substratu hauen C-muturreko azken dipeptidoak
askatuko dira. Bi substratu hauek, odol presioa eta ura eta gatzen metabolismoa
erregulatzen dute. Hala ere, in vitro beste substratu ugari erabil ditzakeela deskribatu da.
Adibidez, neurotentsina, entzefalinak, endorfinak eta dinorfinak, kolezistokinina-8, LHRH,
P substantzia, Ac-Ser-Pro-Asp-Lys-Pro baita insulina ere. Hematopoiesian, immunitate
sisteman, ugalketna eta neuropeptidoen erregulazioan ere parte hartzen du. ACE
entzimaren ezaugarri azpimarragarria da, nagusiki ektoentzima bat izan arren,
endoentzima bezala ere jardun dezakeela. Beraz, bi gune katalitiko dituela esan daiteke
eta aipatu berri diren substratu batzuen N-muturreko zein C-muturreko tripeptido eta
tetrapeptidoak ere apurtu ditzakeela (Skidgel et al., 1985; Azizi et al., 1999; Carl-McGrath
et al., 2006).
ACE-ak, beste metaloproteasek baino pH alkalinoagoan du aktibitaterik gorena, 8
inguruan (Fernandez et al., 2001). Aktibatzaile gisa, Cl- ioia behar izaten duela deskribatu
da, substratuaren eta gune aktiboaren arabera, N-domeinuak kloroarekiko menpekotasun
gutxiago duelarik (Corvol et al., 2004). Captopril, lisinopril, trandolaprilat eta enalaprilat
dira inhibitzaile garrantzitsuenak ACE entzimarentzat. Hala ere, inhibitzaile bakoitzak
espezifizitate ezberdina erakusten du substratuaren arabera eta baita bi doimeinu
aktiboen arabera ere (Wei et al., 1992; Cotton et al., 2002).
ACE geneak kodetzen du metalopeptidasa hau. 17. kromosomako beso luzean
kokatzen da gizakian, 21 kb eta 26 exon ditu. Proteina somatikoa 1-26 exonera
transkribatzen den bitartean, proteina germinala txikiagoa da eta 13-26 exonera
trasnskribatuko da soilik (Hubert et al., 1991). Gene honetan gertatzen diren
polimorfismoen ondorioz, plasma eta ehuneko ACE kopuruan aldaketak gertatzen direla
deskribatu izan da. Kontuan izateko ezaugarria da hau, gaixotasun batzuek ere ACE
entzimaren kopuru aldaketak eragiten baitituzte (Usmani et al., 2000; Röcken et al., 2007).

Sarrera
33
3.5.11. Angiotentsina eraldatzen duen entzima 2 (AC E2)
ACE2 entzima, hasiera batean ACE-ren homologo bezala deskribatu zen. Gerora,
aldiz, ezaugarri ezberdinak dituztela aurkitu zenez, identitate propioa eman zitzaion
(Donoghue et al., 2000).
Metalopeptidasen taldean sailkatzen da ACE2-a, M2 familiako kide baita. Kasu
honetan karboxipeptidasa bat izanik, oligopeptidoen C-muturreko dipeptido hondarrak
hidrolizatzen ditu. Oligopeptidoen azken aminoazidoa hibrofobikoa edo basikoa bada eta
1, 3 edo 5 gunean prolina bat badago hidrolisia errazagoa izango da. I. motako
mintzarteko peptidasa bat da, N-muturra mintzaren kanpo alderantz du norabidetua,
mintzarteko gune hidrofobikoa eta fosforilazioa jasaten duen C-muturra zitoplasmarantz
begira. Zink-ari lotzeko gunea du beste metalopeptidasen antzera (Turner et al., 2004b).
Substratuei dagokionez, ACE entzimak baino espezifizitate handiagoa azaltzen du
ACE2-ak, angiotentsina I eta angiotentsina II-ari lotzen zaiola deskribatu da, baina afinitate
handiagoa du angiotentsina II-arekiko. Honela, errenina angiotentsina sisteman (RAS)
garrantzitsuena den peptidoaren (Ang II) oreka lortzen dute bi entzima hauek. Bestalde,
apelina-13, kaspasa-1 eta A dinorfina ere ACE2-aren substratuak direla azaldu da
(Vickers et al., 2002; Turner et al., 2002b; Warner et al., 2004; Guy et al., 2005).
ACE2-aren adierazpena ez dago bere homologoarena bezain hedatua. Kopuru
handian agertzen da ugaztunen biriketan. Bihotzeko gaixotasun batzuetan duen eragina
ikusita, bertan funtzio garrantzitsuren bat duela uste da (Ferrario et al., 1997; Crackower et
al., 2002). Giltzurrunean eta digestio aparatuan, hots, area eta hestean ere agertzen da
ACE2 entzima, organo bakoitzaren guneko RAS sistemaren kide garrantzitsu bezala
(Zhang et al., 2001; Zhou et al., 2009; Larrinaga et al., 2010a). Aipatu berri den sistema
honetan, ACE-ak eta ACE2-ak eragin kontrajarriak dituztela deskribatu da. Lehenak,
zelularen hazkuntza eragiten duen bitartean, besteak efektu antiproliferatiboak ditu, eta
bata, basodilatadorea den bitartean, bestea baso-ertsatzailea da. Honela, hainbat
prozesuren oreka lortzen da bi entzimen aktibitateen artean (Zhou et al., 2009; Larrinaga
et al., 2010a).
Entzimaren jarduerari dagokionez, pH optimoa 6,5 ingurukoa da. Pisu molekularra,
90 kDa-koa dela deskribatu da, ACE-a baina txikiagoa. Ioi kloridoek (Cl-), kasu honetan
ere aktibatzaile funtzioa betetzen dute, baina inhibitzaileen kasuan, ACE-a inhibitzen
duten molekulak ez dute ACE2-a inaktibatzen. ACE2-a inhibitzeko, ordea, metal-kelatzaile
eragileak erabiltzen dira (Vickers et al., 2002).

34
Sarrera
Izen berbera du peptidasa hau kodetzen duen geneak, ACE2, X. kromosomako
beso luzean kokatzen da zehazki eta 18 exon ditu.
3.5.12. Endopeptidasa neutroa (NEP)
M13 familiako kide da endopeptidasa hau. Metaloproteasaren ekintza, 40-50
aminoazidoko luzera duten peptidoetara mugatzen da eta espezifizitate handia azaltzen
du aminoazido hidrofobikoen N-muturrarekiko, batez ere, leuzina, metionina eta
fenilalanina badira (Kerr et al., 1974a). Bere substratu nagusiak, entzefalinak, takikininak,
P substantzia, endotelinak, bonbesinak, bradikininak eta angiotentsina I dira (McLellan et
al., 1988; Roques et al., 1990; Ruiz-Ortega et al., 2007).
NEP, entzima ugaria da hesteko ertz eskuilakaran, APN entzimaren antzera.
Giltzurrun, plazenta, hezur-muin eta immune-sistemako zelula erretikularretan ere oso
nabaria da kopuru aldetik (Howell et al., 1991).
Peptidasaren egitura purifikatuan, kate bakarreko polipeptidoa dela deskribatu da
eta bere pisu molekularra 90 kDa-koa izan arren, aldagarria da glikosilazioen arabera
(Malfroy et al., 1988). II. motako mintzarteko proteina bezala agertzen da lehen aipaturiko
ehun guztietan, amino muturra zelularen barnealdera izaten du, gune hidrofobiko bat
mintzean zehar eta gune aktiboa handiena den zatian, hots, zelularen kanpo-matrizera
zuzendua.
Honez gain, NEP-ak Zn2+-ari lotzeko gunea agertzen du zelularen kanpoaldeko
espaziora begira, alabaina, beste ioi dibalente batzuek ere peptidasa aktiba dezakete,
ordea, neurri baxuagoan. Entzimaren aktibitatea inhibitzen duten molekulak ugariak dira,
hala nola, EDTA, beta-mercaptoetanola eta 1,10-fenantrolina bezalako kelatzaileak, DTT-
a bezalako tiolak, tiorfana eta bere deribatuak. Bere pH-rik egokiena 6 ingurukoa da (Kerr
et al., 1974b; Roques et al., 1980). 3. kromosoman dago MME genea, alegia, NEP
entzima kodetzen duena (Barker et al., 1989).
Minbizi askoren bilakaerarekin erlazionatu izan da endopeptidasa neutroa, hala
nola, prostatako, endometrioko, giltzurruneko eta birikako minbizietan (Cohen et al., 1996;
Papandreou et al., 1998; Göhring et al., 1998; Suzuki et al., 2001). Markatzaile histologiko
edota pronostiko gisa ere deskribatu da NEP/CD10 entzimaren erabilera hainbat
neoplasia mota ezberdinetarako (Göhring et al., 1998; Liu et al., 2007).

Helburuak


Helburuak 37
HELBURUAK
Lan hau peptidasen ikerketan 20 urteko esperientzia duen ikerlan taldean egin da.
Bere hastapenetan, talde honek neuropeptidoen metabolismoak nerbio sistemako prozesu
fisiologiko eta patologiko anitzetan duen eragina aztertu zuen. Gerora, ikerketa beste
organo eta sistema batzuetara zabaldu zen, bi lan-lerro sortu zirelarik. Bata, ugalketako
sistema peptidikoaren ikerketan aritzen dena, eta bestea, garapenean zentratu dena.
Animalia-eredu eta giza ehunekin lanean urte batzuk aritu ondoren, lortutako emaitzek
peptidasak ugalkortasunarekin, garapenarekin eta hainbat ehun mota ezberdinen
zahartzapenarekin erlazionatuta daudela adierazi zuten (De Gandarias et al., 1993, 1996,
1999, 2000; Irazusta et al., 2002, 2004; Agirregoitia et al., 2003a, 2003b, 2007; Valdivia et
al., 2004).
Behin puntu honetan, ikerketa arlo berri bat ireki zen, peptidasek garapen
neoplasikoan izan dezaketen erlazioan oinarritzen zena. Hasieran, giltzurruna izan zen
aztergai, peptidasetan aberatsa dena. Emaitza adierazgarriak izan ondoren (Larrinaga et
al., 2007; Varona et al., 2007; Blanco et al., 2008; Larrinaga et al., 2010a, 2010b; Varona
et al., 2010; Blanco et al., 2010), peptidasetan aberatsa den beste ehun bat aztertu zen:
heste lodiko mukosa eta bertan jatorria duten neoplasiak (adenoma eta adenokartzinoma).
Neoplasia hauek, minbiziaren sorrera, bilakaera eta agresibitatea ikasteko eredu oso
erabilgarrian bilakatu dira (Leslie et al., 2002; Kumar et al., 2008; Meyerhardt et al., 2008).
Bertan lortutako emaitzek gainera, peptidasek minbiziaren prozesu hauetan duten
garrantzia zehazten lagundu lezakete, neoplasia hauen diagnosiaren, pronostikoaren eta
tratamenduaren hobekuntzan lagungarri izan litezkeelarik.

38
Helburuak
Beraz, ikerlan honen helburuak ondorengoak izan ziren:
1. Garrantzi biologikoa eta fisiopatologikoa duten peptidasen entzima-jarduera
neurtzea heste lodiko mukosa osasuntsu-adenoma-adenokartzinoma progresioan,
heste-zelulen mintzeko gunean eta gune solugarrian.
2. Koloneko adenokartzinoma duten gaixoen plasman aukeratutako peptidasen
aktibitatea aztertzea eta plasma-kontrolekin alderatzea.
3. Peptidasen ekintzaren azterketa egitea koloneko minbiziaren gradu histologikoaren
eta estadioaren arabera, bai heste-zelula laginetan bai plasma-laginetan.

Materialak eta Metodoak


Materialak eta Metodoak
41
2. MATERIALAK ETA METODOAK
1. Materiala
Ikerketa lan honetan erabili diren aparatuak, soluzioak, erreaktiboak eta produktuak
2.1. Taulan adierazten dira.
2.1. Taula: Aparatuak, soluzioak eta produktuak zein erreaktiboak. (I) Aparatuak. Aparatu-modeloa eta aparatu-marka (II) Soluzioak eta horiek prestatzeko erabili diren osagaiak, (b/b) bolumena/bolumena, (p/b) pisu/bolumena. (III) Erreaktiboen eta produktuen hornitzaileak ageri dira, hala nola, jatorrizko hiria eta lurraldea.
(I) Aparatuak Modeloa. Marka
Espektrofotometroa UV-2401 PC. Shimadzu Espektrofotometroa Hitachi-911. Hitachi Roche Espektrofluorimetroa RF-540. Shimadzu Inkubagailu lehorra Hotcold S, 0-60ºC. Selecta pH-metroa Micro-pH 2002. Crison Termoegonkortutako irabiagailu magnetikoa. Agimatic Ultraizozgailua MDF-U5 186 S. Sanyo Ultraturrax homogeneizagailua Miccra D1. ART Ultrazentrifuga Centrikon T2070. Kontron Angelu finkoko errotorea TFT 45.6. Kontron Zehaztasunezko balantza R180D. Sartorius Zentrifuga Labofuge 200. Heraeus
(II) Soluzioak Osagaiak
Tris-HCl-Indargetzailea 0,2 M (pH 7,4) 1000 ml H2O 24,22 g TRIZMA-base
Homogeneizazio soluzioa, Tris-HCl 10 mM % 5 Tris-HCl 0,2 M
Fosfato-Indargetzailea 0,1 M (pH 7,4) 18,7 ml A + 81,3 ml B
A Fosfatoa 1000 ml H2O 27,6 g NaH2PO4 •H2O 0,2 M
B Fosfatoa 1000 ml H2O 28,4 g Na2HPO4 0,2 M
Azetato-Indargetzailea 0,1 M (pH 4,2) 36,8 ml A + 13,2 ml B + 50 H2O
A Azetatoa 1000 ml arte H2O 11,55 ml CH3COOH 0,2 M
B Azetatoa 1000 ml H2O 27,22 g CH3COONa•3H2O 0,2 M
Bradford erreaktiboa
0,117 mM Coomasie % 5 (b/b) Etanola % 15 (p/b) H3PO4

42
Materialak eta Metodoak
(III) Erreaktiboak eta Produktuak Hornitzailea
Hiria, Lurraldea
Abz-Ser-Pro-3-nitro-Tyr-OH (SPNPT) Cys-β-naftilamida H-Glu-β-naftilamida H-Gly-Pro-β-naftilamida Z-Gly-Pro-β-naftilamida
Bachem
Bubendorf, Suitza
Azido ortofosforikoa (H3PO4) EDTA Etanol absolutua (CH3CH2OH) Manganeso kloruroa (MnCl2) Sodio-azetato trihidratatua (CH3COONa•3H2O )
Sodio fosfato dibasikoa (NaH2PO4)
Sodio fosfato monobasiko monohidratatua (Na2HPO4) Sodio kloruroa (NaCl)
Merck Darmstadt, Alemania
Ala-β-naftilamida Arg-β-naftilamida Asp-AP-β-naftilamida Azido piroglutamiko- β-naftilamida Behi-seroalbumina (BSA) Captopril Coomasie urdin distiratsua G-250 Dimetil-sulfoxidoa (DMSO) DL-ditioteitrola (DTT) Kaltzio kloruroa (CaCl2) N-Dansyl-D-Ala-Gly-pNO2-Phe-Gly (DAGNPG) Puromizina Tris (hidroximetil) aminometazola (Tris-base)
Sigma-Aldrich
Steinheim, Alemania
3-(2-furylacryloyl-l-Phe-Ala-Gly-Gly) (FAPGG) Trinity Biotech
Bray, Irlanda
Azido azetikoa (CH3COOH) Probus
Badalona, Espainia
Azido klorhidrikoa (HCl) Panreac
Bartzelona, Espainia

Materialak eta Metodoak
43
2. Erabilitako laginen erdiespena edo lorpena
2.1. Heste-laginak
Ikerlan honetan aztertu ziren laginak koloneko adenokartzinoma zuten gaixoenak
ziren, kirurgia bidez erauzitako heste-ehun zatia edo biopsia bat zehatzago esanda.
Basurtuko Ospitaleko (Bilbao, Bizkaia) Anatomia Patologikoko Sailean diagnostikatuak eta
aukeratuak izan ziren denak. Ondorengo ikerlan honek Osakidetzaren Ikerkuntza
Klinikoaren eta Unibertsitatearen Batzorde Etikoaren adostasuna jaso du.
Heste-lagin hauek Basurtuko Ospitaleko Anatomia Patologikoko biltegi batean
gordeta eduki zituzten -80 ºC-tan gurera iritsi arte.
Aztertutako laginak, paziente berberaren koloneko hiru gune izan ziren: bi
neoplasia eta kontrola (n=30). Neoplasietako bat, adenoma edo tumore onbera (polipoa)
zen eta bestea, tumore gaiztoa (koloneko adenokartzinoma) zuen ehun zatia. Kontrol
modura, minbiziaren aldamenean baina gaixorik ez zegoen heste-ehun zatia erabili zen.
Sarreran aipaturiko heste lodiko bilakaera kartzinogenetikoaren teoria (osasuntsu-
adenoma-adenokartzinoma) aztertu nahian aukeratu ziren hiru ehun zati hauek.
2.2. Plasma-laginak
Ikerlan honetan, gaixoen plasma, eta ez minbizirik ezta urdail-hesteetako
gaixotasunik ez zutenen plasma, konparatu ziren (n=45). Azken hau kontrol modura erabili
zen pazienteen plasmarekin konparaketa egiteko.
Paziente bakoitzari 3 ml odol atera zitzaizkion eta bertatik plasma banatu zuten
zentrifugazio labur baten bidez (2600 xg, 3 min). Zentrifugazio honen ondorioz, jalkinean
geratuko dira zelula guztiak eta gainjalkinean geratzen den likidoa plasma izango da.
Plasma-lagin hauek izan ziren ikerlan honen ikerketako lanean erabili zirenak.
Heste-laginekin egin zen bezala, plasma-laginak -80 ºC-tan gorde zituzten
Basurtuko Ospitalean gurera iritsi arte. Era berean, ultraizozgailu (Sanyo) batean gorde
ziren -80 ºC-tan entseguko egunera arte.
Kasu bakoitzari zegozkion hiru ehun mota hauek, zein bi plasma laginek protokolo
berbera jarraitu zuten behin laginaren prestaketa egin ostean.
3. Peptidasen entzima-jardueraren azterketa
3.1. Laginen prestaketa
Aipatu berri diren lagin mota guztietan 12 entzimen aktibitatea neurtu zen. Jarduera

44
Materialak eta Metodoak
hauek metodo fluorimetrikoak erabiliz neurtu ziren, baina horretarako laginek prestaketa
prozesu bat behar zuten. Prozesu osoan zehar, laginak 4 ºC-tan mantendu ziren.
3.1.1. Heste-laginen prestaketa
Aukeratutako laginak desizoztu ondoren, 100 mg inguru pisatu ziren ehun fresko
mota bakoitzetik, 1,5 ml Tris-HCl 10 mM (pH7,4) indargetzaile gehitu zitzaien eta ondoren
ultraturrax-a (ART) erabiliz, homogeneizatu egin ziren. Lorturiko nahasketa
ultrazentrifugatu egin zen ultrazentrifuga (Kontron) erabiliz, TFT 45.6 errotorean, 100.000
xg-tara, 35 min-ko epean, 4 ºC-tan. Zentrifugatu ondoren, bi fase ezberdindu zitezkeen,
gainjalkina eta jalkina. Gainjalkina eppendorf saihodietan gorde zen, hau baitzen Gune
Solugarria (2.1. Irudia). Gelditzen zen hauspeakina edo jalkina lehen aipatutako
indargetzaile berbera erabiliz, bi aldiz garbitu zen gelditu zitezkeen ezpurutasunak
kentzeko asmoarekin. Jalkin honi 1 ml Tris-HCl 10 mM (pH 7,4) gehitu zitzaion eta lehen
aipatutako homogenizazio prozesua errepikatu zen. Ondoren, zentrifuga (Heraeus)
erabiliz, zentrifugazio labur bat egin zen, 800 xg-tan 3 min-z. Kasu honetan ere,
gainjalkineko fasea jaso genuen, hau baitzen Mintzeko Gunea.
2.1. Irudia: Heste-lagin erauzi berritik zeluletako mintzeko gunea zein gune solugarria lortzeko prozedura eskematikoa.

Materialak eta Metodoak
45
3.1.2. Plasma-laginen prestaketa
Plasma-laginak ultraizozgailutik atera ziren desizoztu zitezen. Ondoren,
ezpurutasun posibleak kentzeko asmoarekin, zentrifugazio labur bat (2000 xg, 3 min) egin
zen. Gainjalkinetik 1,5 ml plasma-lagin hartu ziren entsegu fluorimetrikoen bidezko
analisietarako.
3.2. Lagineko proteina kopuruaren neurketa
Lagin mota ezberdinetan zegoen guztizko proteinaren kopurua zein zen neurtzeko,
Bradford et al.-en teknika erabili zen (1976). Horretarako, 2 ml Bradford erreaktibo gehitu
zitzaizkien 10 µl laginei. Ongi irabiatu ziren eta 10 min erreakziona zezan itxaron ostean,
absorbantzia neurtu zen 595 nm-tara espektrofotometroan (Shimadzu).
Gure laginak, BSA (1 mg/ml) kopuru ezaguneko patroi zuzenaren erdigunean
gelditzen zirela ziurtatu genuen bi arrazoiengatik, bata, Bradfordaren absorbantzia eta
lagin kontzentrazioa proportzionalak direlako gune honetan eta bestea, ondorengo
entzima-jarduerarako kontzentrazioa egokiena delako. Behar izatekotan, Tris-HCl 0,2 M
(pH 7,4) indargetzailea erabili zen laginak diluitzeko.
3.3. Entzima-jarduera
Entzimen ekintzak heste-zelulen mintzeko gunean zein gune solugarrian aztertu
ziren. Beti ere, heste-ehuneko hiru motetan: koloneko adenokartzinoman, polipoan eta
hauen aldamenean baina gaixorik ez zegoen ehun zati osasuntsuan. Baita paziente
gaixoen plasman eta kontrol modura erabili zirenen plasman ere.
Erreakzioa betetzeko, entzima bakoitza neurtzeko beharrezko zen lagin kopuruari
(30-100 µl) artifiziala den substratu fluorogenikotik 1 ml bota zitzaion (2.2. Taula).
Nahastura ondo irabiatu ondoren, 37 ºC-tan mantendu zen 30 min-ko epean. Erreakzioa
gelditzeko, 4,2-ko pH duen 0,1 M azetato indargetzailearen 1 ml erabili zen. Peptidasen
entzima-jarduera, entsegu fluorimetrikoen bidezko metodoak erabiliz aztertu zen
espektrofluorimetroan (Shimadzu).
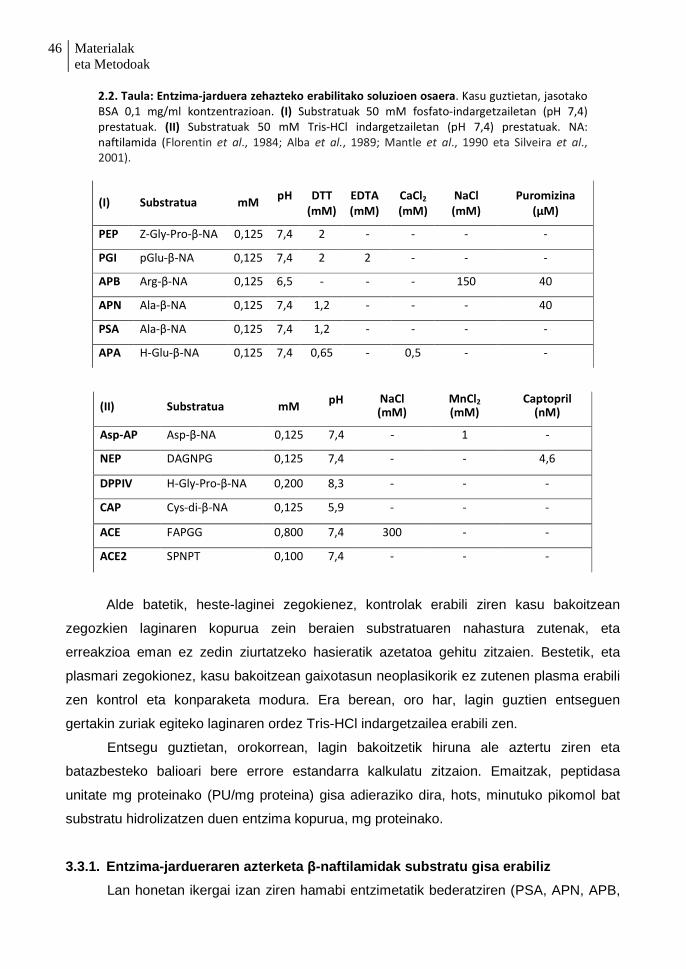
46
Materialak eta Metodoak
2.2. Taula: Entzima-jarduera zehazteko erabilitako soluzioen osaera. Kasu guztietan, jasotako BSA 0,1 mg/ml kontzentrazioan. (I) Substratuak 50 mM fosfato-indargetzailetan (pH 7,4) prestatuak. (II) Substratuak 50 mM Tris-HCl indargetzailetan (pH 7,4) prestatuak. NA: naftilamida (Florentin et al., 1984; Alba et al., 1989; Mantle et al., 1990 eta Silveira et al., 2001).
(I) Substratua mM pH DTT
(mM)
EDTA
(mM)
CaCl2
(mM)
NaCl
(mM)
Puromizina
(µM)
PEP Z-Gly-Pro-β-NA 0,125 7,4 2 - - - -
PGI pGlu-β-NA 0,125 7,4 2 2 - - -
APB Arg-β-NA 0,125 6,5 - - - 150 40
APN Ala-β-NA 0,125 7,4 1,2 - - - 40
PSA Ala-β-NA 0,125 7,4 1,2 - - - -
APA H-Glu-β-NA 0,125 7,4 0,65 - 0,5 - -
(II) Substratua mM pH NaCl
(mM) MnCl2
(mM)
Captopril (nM)
Asp-AP Asp-β-NA 0,125 7,4 - 1 -
NEP DAGNPG 0,125 7,4 - - 4,6
DPPIV H-Gly-Pro-β-NA 0,200 8,3 - - -
CAP Cys-di-β-NA 0,125 5,9 - - -
ACE FAPGG 0,800 7,4 300 - -
ACE2 SPNPT 0,100 7,4 - - -
Alde batetik, heste-laginei zegokienez, kontrolak erabili ziren kasu bakoitzean
zegozkien laginaren kopurua zein beraien substratuaren nahastura zutenak, eta
erreakzioa eman ez zedin ziurtatzeko hasieratik azetatoa gehitu zitzaien. Bestetik, eta
plasmari zegokionez, kasu bakoitzean gaixotasun neoplasikorik ez zutenen plasma erabili
zen kontrol eta konparaketa modura. Era berean, oro har, lagin guztien entseguen
gertakin zuriak egiteko laginaren ordez Tris-HCl indargetzailea erabili zen.
Entsegu guztietan, orokorrean, lagin bakoitzetik hiruna ale aztertu ziren eta
batazbesteko balioari bere errore estandarra kalkulatu zitzaion. Emaitzak, peptidasa
unitate mg proteinako (PU/mg proteina) gisa adieraziko dira, hots, minutuko pikomol bat
substratu hidrolizatzen duen entzima kopurua, mg proteinako.
3.3.1. Entzima-jardueraren azterketa β-naftilamidak substratu gisa erabiliz
Lan honetan ikergai izan ziren hamabi entzimetatik bederatziren (PSA, APN, APB,

Materialak eta Metodoak
47
Asp-AP, CAP, PGI, DPPIV, PEP eta APA) jarduera aztertzeko peptidil-β-naftilamida edo
aminoazil-β-naftilamida motako substratuak erabili ziren.
PSA, APN eta APB peptidasen ekintza aztertzeko 30 µl lagin erabili ziren 3.1.
azpiatalean azaldu den bezala prestatutako hiru heste-laginetarako (gune solugarrikoa
zein mintzeko gunekoa) eta bi plasma-laginetarako. 50 µl erabili ziren ordea, Asp-AP,
CAP, PGI, DPPIV, PEP eta APA entzimen jarduera neurtzeko.
Laginean aurkitzen diren aminopeptidasek peptidil-β-naftilamida edo aminoazil-β-
naftilamida substratua bitan hidrolizatuko dute, peptidiloa edo aminoaziloa eta β-
naftilamina loturatik, 2.2. Irudian azaltzen den bezala. Azken molekula honek,
fluoreszentea denak, 345 nm-ko uhin-luzera duen argi izpi batez kitzikatzen bada, beste
argi izpi bat jaurtiko du, energia txikiagokoa dena eta 412 nm-ra jasoko dena. Ezaugarri
honi esker, espektrofluorimetroaren bidez, erraz asko neur daiteke askatu den β-
naftilamina kopurua. Produktuak askatutako argi kopuru hau neurtuz, laginean dagoen
entzimaren jarduera zenbatekoa den jakitea baimenduko du.
2.2. Irudia: Peptidil-β-naftilamida gisako substratuen hidrolisia eskematikoki azaldua.
3.3.2. Entzima-jardueraren azterketa FAPGG (3-(2-fu rylacryloyl-l-Phe-Gly-Gly))
substratu gisa erabiliz
ACE entzimaren aktibitatea neurtzeko FAPGG (3-(2-furylacryloyl-l-Phe-Gly-Gly))
gisako substratu artifiziala erabili zen. Entzimak Phe-Gly lotura hidrolizatuko du, FAP (3-
(2-furylacryloyl-l-Phe)) eta Gly-Gly banatuz. Lehen hau, FAP, izan zen metodo
fotometrikoen bidez neurtu zena eta aurreko kasuan bezala, estrapolatze kurba bat egin
ezkero entzimaren aktibitatea zein den jakin daiteke (2.3. Irudia).

48
Materialak eta Metodoak
Heste-laginen mintzeko gunean aztertu zen ACE entzimaren ekintza eta baita
plasma-laginetan ere. Kasu honetan, bina ale aztertu ziren soilik eta zuriak egiteko
laginaren ordez indargetzailea zuten hodiak erabili ziren gainontzeko entseguetan bezala.
Honela, laginari ez dagokion argia ezeztatuko da.
Substratuaren 900 µl nahastu ziren 100 µl lagin prestakinekin eta jarraian (t0),
espektrofotometroaren (Hitachi Roche) bidez, absorbantzia neurtu zen 340 nm-ko uhin-
luzeran, 25 ºC-tan. 15 min pasa ostean (t15) berriro neurtu zen absorbantzia uhin-luzera
berberean. Denbora tarte horretako absorbantziaren espektru aldaketa horrek adierazten
du entzimak substratuaren gainean egin duen ekintza. Denboran zeharreko substratuaren
desagerpena neurtu zen kasu honetan, ondorioz, ∆A340-ko datuak negatiboak ziren. ACE
entzimaren aktibitatea U/mg proteinako adierazi zen, entzima unitate bat (U), minutuko,
340 nm-tan gertatzen diren -0,0001-eko absorbantzia aldaketa kopurua (∆A340) izanik.
2.3. Irudia: FAPGG gisako substratuen hidrolisia eskematikoki azaldua.
3.3.3. Entzima-jardueraren azterketa SPNPT (Abz-Ser -Pro-3-nitro-Tyr-OH) substratu
gisa erabiliz
ACE2 entzimaren aktibitatea neurtzeko SPNPT (Abz-Ser-Pro-3-nitro-Tyr-OH)
gisako substratu artifizial eta fluorogenikoa erabili zen. Peptidasa honen kasuan, bina ale
aztertu ziren eta heste-zeluletako mintzeko gunean zein plasma-laginetan aztertu zen.
Laginetik 100 µl hartu ziren alikuota bakoitzerako eta 1 ml substratu gehitu zitzaion
bakoitzari. Erreakzioa eman zedin 37 ºC-tan 2 h-z inkubatu ziren. Gelditzeko aldiz, 1 ml
azetato 0,1 M (pH 4,2) erabili zen eta jarraian espektrofluorimetroan (Shimadzu) 410 nm-
ko uhin-luzeran jaso zen entzimaren aktibitatea 360 nm-ko uhin-luzerarekin kitzikatu

Materialak eta Metodoak
49
ostean (2.4. Irudia).
Entzimak Abz-Ser-Pro-3-nitro-Tyr-OH substratua hidrolizatu ostean, Abz-Ser-Pro
eta 3-nitro-Tyr-OH-an banatuko da. Abz (2-Aminobenzoil) gisako substratuek berezko
fluoreszentzia dute, baina 3-nitro-Tyr-OH-ak pantaila modura jokatzen du argi hori
ezkutatuz. Behin entzimak bere funtzioa betetzen duenean Abz-Ser-Pro aske geratuko da
eta askatzen duen fluoreszentzia entzimaren aktibitatearekiko zuzenki proportzionala da.
Ondorioz, erregresio kurba bat egin ezkero errez asko jakin daiteke zein izan den ACE2-
aren ekintza substratu honen gainean.
2.4. Irudia: SPNPT gisako substratuen hidrolisia eskematikoki azaldua.
3.3.4. Entzima-jardueraren azterketa DAGNPG (N-Dans yl-D-Ala-Gly-(p-NO 2)-Phe-Gly)
substratu gisa erabiliz
NEP entzimaren kasuan, DAGNPG (N-Dansyl-D-Ala-Gly-(p-NO2)-Phe-Gly)
substratu artifiziala erabili zen entseguetarako, hau ere fluorogenikoa da eta
fluorimetroaren bidez neurtzen da entzimaren ekintza.
Beste entzimen antzera, heste-zelulen mintzeko guneko prestakinetik 100 µl eta 1
ml substratu artifizial inkubatu ziren 37 ºC-tan, 30 min-z.
NEP entzima zelulen mintzeko endopeptidasa bat da eta peptidoen erdiko guneko
loturak apurtzen ditu, substratuaren Gly-(pNO2)-Phe lotura moztuko du orduan. Dansyl-ak
(5-(Dimetilamino)naftaleno-1-sulfonil) duen berezko fluoreszentzia, pNO2-Phe taldeak
ezkutatzen du baina behin NEP-ak lotura hau moztuta, N-Dansyl-D-Ala-Gly (DAG) aske
geratuko da eta Dansyl-aren argia neur daiteke 342 nm-ko uhin-luzerako argi izpi batez
kitzikatuz, eta 562 nm-tara jasoaz Shimadzu espektrofluorimetroan (2.5. Irudia).

50
Materialak eta Metodoak
Beste entzimekin gertatzen den bezala, hemen ere DAG produktuaren argi kopurua
laginean dagoen NEP entzimaren ekintzaren menpekoa da eta estrapolatze bat egin
daiteke.
2.5. Irudia: DAGNPG gisako substratuen hidrolisia eskematikoki azaldua.
4. Minbiziaren sailkapena
Basurtuko Ospitaleko Anatomia Patologikoko Sailean diagnostikatu eta sailkatu
zituzten laginak. Bertan, kirurgia bidez erauzitako heste lodiko lagin erauzi berriak,
Osasunerako Mundu Erakundeak (OME) adierazi bezala, eta heste lodiko minbizietarako
espresuki moldatutako TNM (Tumour, Node, Metastasis) sisteman oinarritutako protokoloa
jarraituz sailkatzen dira. Ospitaleek jarraitzen duten protokolo honetan azterketa zabal bat
egiten da. Minbiziaren egitura makroskopikoa zein mikroskopikoa aztertzen da I.
Eranskinean agertzen den adibidean ikus daitekeen bezala.
4.1. Gradu histologikoaren sailkapena: OME-ren grad uak
Koloneko Tumorearen gradu histologikoa zehazteko OME-ren gradu sailkapena
erabili zen. Honen arabera, bi sailkapen mota egiten dira, hiru gradu sistemakoa eta bi
gradu sistemakoa. Lehenengo kasuan, graduak batetik hirura (G1, G2 eta G3) izendatzen
dira zelulen nukleoaren ezberdintzapenaren arabera. Bigarren sailkapenean, bi talde
sortuko dira, gradu baxua eta gradu altua. Bi sailkapen hauek ondorengo taulan agertzen
den bezala erlaziona daitezke (2.3. Taula).

Materialak eta Metodoak
51
2.3. Taula: Heste lodiko minbiziko histologi-graduaren sailkapena OME-ren irizpideak
jarraituz.
Gradu Baxua
G1: nukleo ongi ezberdintzatuak
G2: nukleo erdi ezberdintzatuak
Gradu Altua
G3: nukleo gutxi ezberdintzatuak
Mikroskopi-azterketen bidez, tumore-zelulek duten agresibitatea edo gaiztotze
maila aztertzen da. Hala nola, nukleoen ezaugarri mikroskopikoak, egitura,
pleomorfismoak eta mitosi-jarduera ikus daitezke. Honen arabera gradu bat edo beste
egokituko zaie. Nukleo gutxi ezberdintzatuak dituzten tumore-zelulek joera agresiboagoa
dute (Kumar et al., 2008). 2.6. Irudian agertzen diren argazkietan heste-zelulen nukleoen
ezberdintzapen mailak adierazten dira.
2.6. Irudia: Heste lodiko histologi-gradua (G1-
G3) adierazten duten hiru mikroargazki, OME-
ren sailkapena, 100x.
(I) Ongi ezberdintzatutako nukleoak, (II) nukleo erdi ezberdintzatuak eta (III) nukleo gutxi ezberdintzatuak.
I
II
III

52 Materialak eta Metodoak
4.2. Estadioaren sailkapena: TNM sistema
Hesteari egokitutako TNM sistema erabili zen koloneko minbiziko estadioaren
sailkapena egiteko. Sistema honek irizpide modura dauzka adibidez, lehen mailako
minbiziaren kokapena eta gehiengo diametroa, inguruko egituren inbasioa, gongoil
linfatikoen ezaugarriak eta metastasien agerpena, beste hainbat ezaugarriren artean
(Greene et al., 2002; Kumar et al., 2008). 2.4. Taulan zabalago adierazten dira TNM
sistemaren oinarriak.
2.4. Taula: TNM sistema bidezko heste lodiko minbiziaren estadio-sailkapena.
LEHEN MAILAKO TUMOREA (PRIMARIOA) (pT)
-pTX : Ezin da neurtu (ebaluatu) -pT0 : Ez dago lehen mailako tumore nabarmenik -pTis : In situ Kartzinoma, intraepiteliala (inbasiorik gabe) -pTis : In situ Kartzinoma, Lamina propiaren inbasioarekin -pT1 : Azpimukosaren inbasioa -pT2 : Muskulu-propia geruzaren inbasioa -pT3 : Muskulu geruzan barna, serosaren inbasioa, baita heste inguruko gantzarena
eta peritonealizatu gabeko zuzen inguruko ehunarena ere *pT3a : Gutxieneko Inbasioa: Muskulu-propia geruzaren mugatik <1 mm *pT3b : Inbasio arina: Muskulu-propia geruzaren mugatik 1-5 mm *pT3c : Inbasio moderatua: Muskulu-propia geruzaren mugatik 5-15 mm *pT3d : Inbasio hedatua: Muskulu-propia geruzaren mugatik > 15 mm
-pT4a : Beste organoen eta egituren inbasio zuzena -pT4b : Errai-peritoneoa zulatu GUNEKO GONGOIL LINFATIKOAK (pN)
-pNX : Ezin da neurtu -pN0 : Ez dago guneko gongoil linfatikoetan metastasia nabarmenik -pN1 : Metastasia 1-3 guneko gongoil linfatikoetan -pN2 : Metastasia >4 guneko gongoil linfatikoetan
URRUNEKO METASTASIA (pM)
-pMX : Ezin da neurtu -pM1 :Urruneko metastasia
Heste lodiko adenokartzinomaren estadio-sailkapena egiteko TNM sistema erabili
zen ikerlan honetarako, baina badaude beste hainbat sistema oso erabilgarriak eta
ezagunak. Hala nola, Dukes Estadioa da ezagunena eta honen moldapen bat ere badago
Aster-Coller izenekoa (2.5. Taula). Dukes-en estadio sisteman hiru talde ezberdintzen dira
(A-C) eta sistemarik trinkoena dela esan daiteke. Aster-Coller-en estadio sistema Dukes

Materialak eta Metodoak
53
sistemaren moldapen bat dela esaten da, lau taldetan (A-D) banatzen dituelako estadioak
eta azpitaldeak ere ezartzen dizkiolako Dukes-en sistemari.
2.5. Taula: Estadio-sailkapen ezberdinen erlazioa. (T) lehen mailako tumorea, (N) gongoil linfatikoak, (M) metastasia, (A-D) talde banaketa (American Joint Committee on Cancer: http://www.cancerstaging.org/ ).
Estadioa TNM sistema Dukes Estadioa Aster-Coller (Dukes Moldatua)
0 Estadioa Tis N0 M0 - Super A
I Estadioa T1 N0 M0 A A
T2 N0 M0 A B1
IIA Estadioa T3 N0 M0 B B2
IIB Estadioa T4 N0 M0 B B3
IIIA Estadioa T1,T2 N1 M0 C C1
IIIB Estadioa T3,T4 N1 M0 C C2,C3
IIIC Estadioa Edozein T N2 M0 C C1,C2,C3
IV Estadioa Edozein T Edozein T M1 - D
5. Datuen analisia
Heste laginen kasuan, minbizia, polipoa eta normalaren arteko konparaketa
koloneko adenokartzinoma zuen gaixo berberarenak izan ziren. Laginen datuei
dagokionez, heste lodiko minbizia zuten gizon-emakumeen (2:1 ratioa hurrenez-hurren)
biopsiak aztertu ziren. Pazienteen batazbesteko adina 68,81±1,54-koa izan zen (47-84
urte bitartean), n=30 izanik. Plasmaren konparaketa egiteko unean, pazienteen sexua eta
adina kontuan izan ziren bi lagin bikoteak parekatu ahal izateko (n=45).
Fotometroaren eta fluorimetroaren bidez lortutako datuak askatutako metabolitoen
pikomol kopuruetara pasa ziren. Eraldaketa hau patroi-zuzen baten erregresio-ekuazioan
estrapolatze ez-zuzen bat eginez lortzen da. Datu hauek ondoren peptidasaren aktibitate
unitateetara pasa ziren (PU), honela azalduko da unitate bat: minutuko pikomol bat
substratu hidrolizatzen duen entzima kopurua. Azken emaitzak aktibitate zehatz modura
azaldu ziren, peptidasaren aktibitate unitatea, lagineko proteina miligramoko (PU/mg
proteina). Plasmen kasuan, Nazioarteko Unitate Sisteman, U (µmol/min-etan), neurtu zen
aktibitatea eta lagin litroko egin zen: entzimaren aktibitate unitatea, litro bat plasman
dagoen proteinako (U/l).
Behin entzima bakoitzaren aktibitatea zein zen jakinda, bi plasma laginak beraien
artean konparatzerakoan Student-en T testa egin zen, baita minbizien gradu histologikoen

54 Materialak eta Metodoak
eta estadioen arteko konparaketa egiteko ere. Ehun gaixoan, polipoan eta osasuntsuan
ezberdintasunik ba ote zegoen edo ez ikusteko ANOVA egin zen. SPSS 16.0 programa
estatistikoa erabili zen eragiketa hauek egiteko. Adierazgarritasun estatistikoa adierazteko
(*) p<0,05, (**) p<0,01 eta (***) p<0,001 erabili zen. Bi ehunen arteko konparaketa egitean
adierazgarritasun estatistikoa adierazteko (a) Normala vs. Polipoa, (b) Minbizia vs. Polipoa
eta (c) Normala vs. Minbizia erabili da.

Emaitzak


Emaitzak 57
3. EMAITZAK
1. Peptidasen azterketa: Entzima-jarduera
Ikerlan honetan aztertutako peptidasa guztien entzima-jarduera neurtu ahal izateko
entsegu fluorometrikoak egin ziren.
Alde batetik, tumoreak jarraitzen duen progresio berbera ikertzeko asmoarekin,
entseguak koloneko minbizia zuten pazienteen hiru heste-ehun ezberdinetan aztertu ziren:
gaixorik ez zegoen ehunean, polipoan eta minbizi-ehunean. Gehiago zehaztearren, hiru
ehunetako heste-zelulen mintzari lotutako eta zatiki solugarriari lotutako aktibitateak
aztertu ziren (Materialak eta Metodoak, 3.1.1. Azpiatala).
Terminologia errazagoa izan dadin, ondoren agertuko diren emaitzetan normala
hitza erabiliko da neoplasiarik ez zuen ehuna adierazteko, polipoa, tumore ona
zenarentzako (adenoma) eta minbizia hitza adenokartzinoma ehuna adierazteko.
Bestetik, plasman ere egin ziren entsegu fluorometrikoak. Kasu honetan,
konparaketa koloneko minbizia zuten pazienteen plasmaren eta minbizirik zein
gastrohesteetako patologiarik ez zuten pazienteen plasmaren artekoa izan zen.
Plasmaren kasuan, gaixoa koloneko minbizia zuten pazienteen plasma adierazteko
eta kontrola paziente osasuntsuen plasma adierazteko erabili da.
Atal honen amaieran estatistikoki adierazgarriak izan ziren emaitzen laburpen bat
agertuko da (3. Azpiatala).
Jarraian aipatuko diren emaitzetan, grafikoen bidez azalduko da entzima bakoitzak
gune ezberdinetan duen entzima-jarduera eta eranskin gisa, tauletan jarri dira, zenbakizko
datu guztiak (II. eta III. Eranskinak).
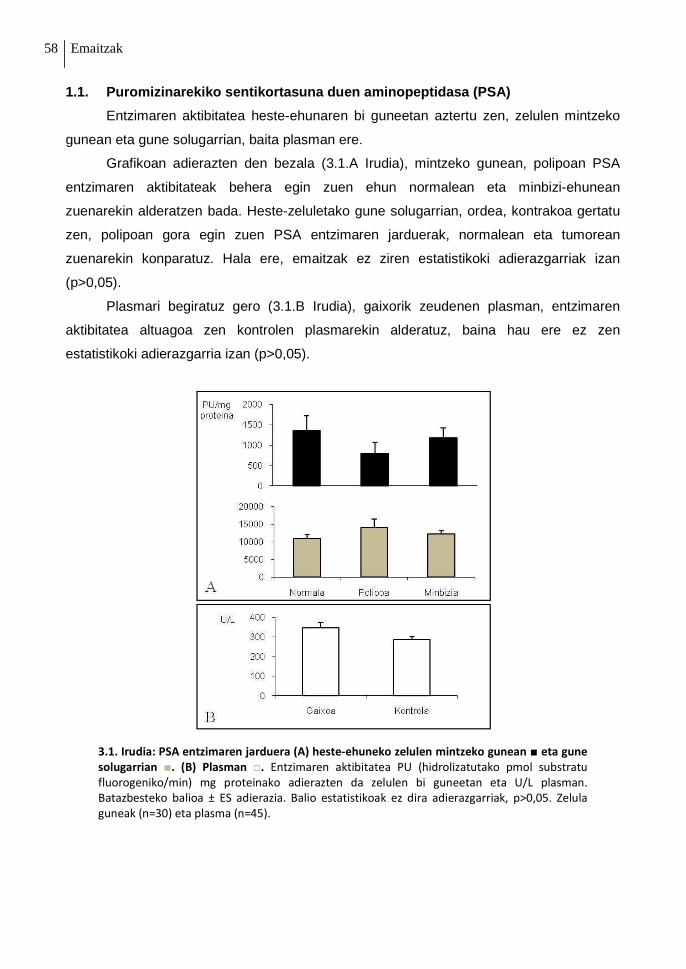
58 Emaitzak
1.1. Puromizinarekiko sentikortasuna duen aminopept idasa (PSA)
Entzimaren aktibitatea heste-ehunaren bi guneetan aztertu zen, zelulen mintzeko
gunean eta gune solugarrian, baita plasman ere.
Grafikoan adierazten den bezala (3.1.A Irudia), mintzeko gunean, polipoan PSA
entzimaren aktibitateak behera egin zuen ehun normalean eta minbizi-ehunean
zuenarekin alderatzen bada. Heste-zeluletako gune solugarrian, ordea, kontrakoa gertatu
zen, polipoan gora egin zuen PSA entzimaren jarduerak, normalean eta tumorean
zuenarekin konparatuz. Hala ere, emaitzak ez ziren estatistikoki adierazgarriak izan
(p>0,05).
Plasmari begiratuz gero (3.1.B Irudia), gaixorik zeudenen plasman, entzimaren
aktibitatea altuagoa zen kontrolen plasmarekin alderatuz, baina hau ere ez zen
estatistikoki adierazgarria izan (p>0,05).
3.1. Irudia: PSA entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistikoak ez dira adierazgarriak, p>0,05. Zelula guneak (n=30) eta plasma (n=45).
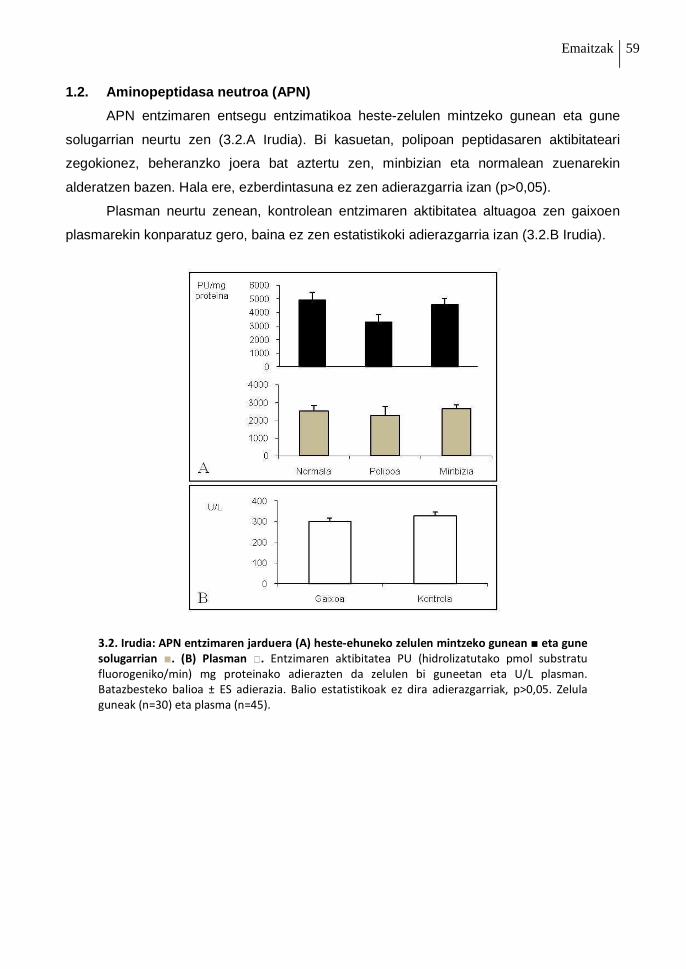
Emaitzak 59
1.2. Aminopeptidasa neutroa (APN)
APN entzimaren entsegu entzimatikoa heste-zelulen mintzeko gunean eta gune
solugarrian neurtu zen (3.2.A Irudia). Bi kasuetan, polipoan peptidasaren aktibitateari
zegokionez, beheranzko joera bat aztertu zen, minbizian eta normalean zuenarekin
alderatzen bazen. Hala ere, ezberdintasuna ez zen adierazgarria izan (p>0,05).
Plasman neurtu zenean, kontrolean entzimaren aktibitatea altuagoa zen gaixoen
plasmarekin konparatuz gero, baina ez zen estatistikoki adierazgarria izan (3.2.B Irudia).
3.2. Irudia: APN entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistikoak ez dira adierazgarriak, p>0,05. Zelula guneak (n=30) eta plasma (n=45).
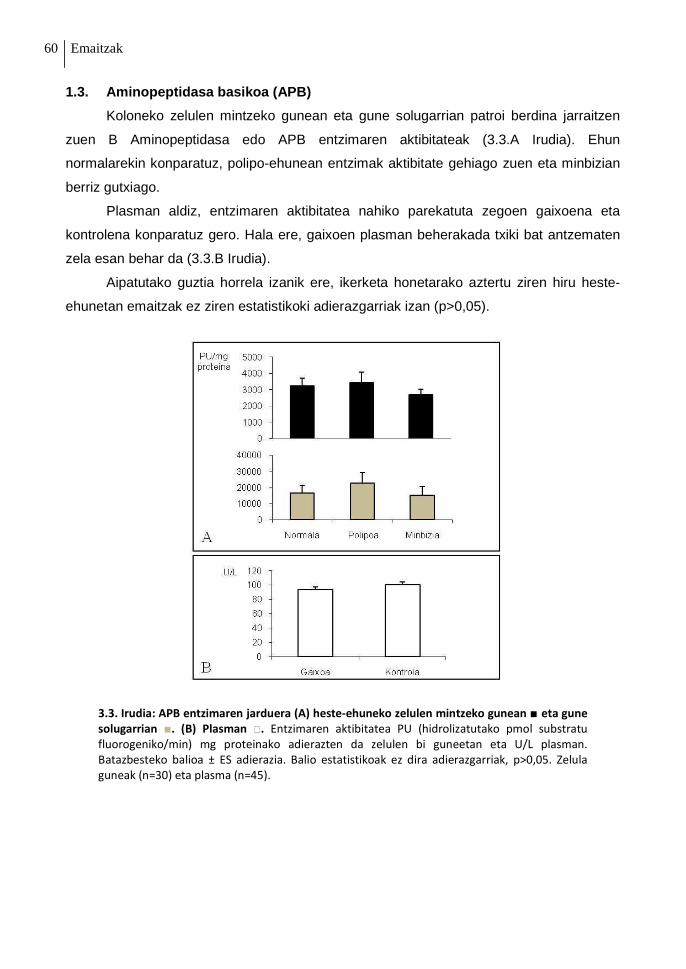
60 Emaitzak
1.3. Aminopeptidasa basikoa (APB)
Koloneko zelulen mintzeko gunean eta gune solugarrian patroi berdina jarraitzen
zuen B Aminopeptidasa edo APB entzimaren aktibitateak (3.3.A Irudia). Ehun
normalarekin konparatuz, polipo-ehunean entzimak aktibitate gehiago zuen eta minbizian
berriz gutxiago.
Plasman aldiz, entzimaren aktibitatea nahiko parekatuta zegoen gaixoena eta
kontrolena konparatuz gero. Hala ere, gaixoen plasman beherakada txiki bat antzematen
zela esan behar da (3.3.B Irudia).
Aipatutako guztia horrela izanik ere, ikerketa honetarako aztertu ziren hiru heste-
ehunetan emaitzak ez ziren estatistikoki adierazgarriak izan (p>0,05).
3.3. Irudia: APB entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistikoak ez dira adierazgarriak, p>0,05. Zelula guneak (n=30) eta plasma (n=45).
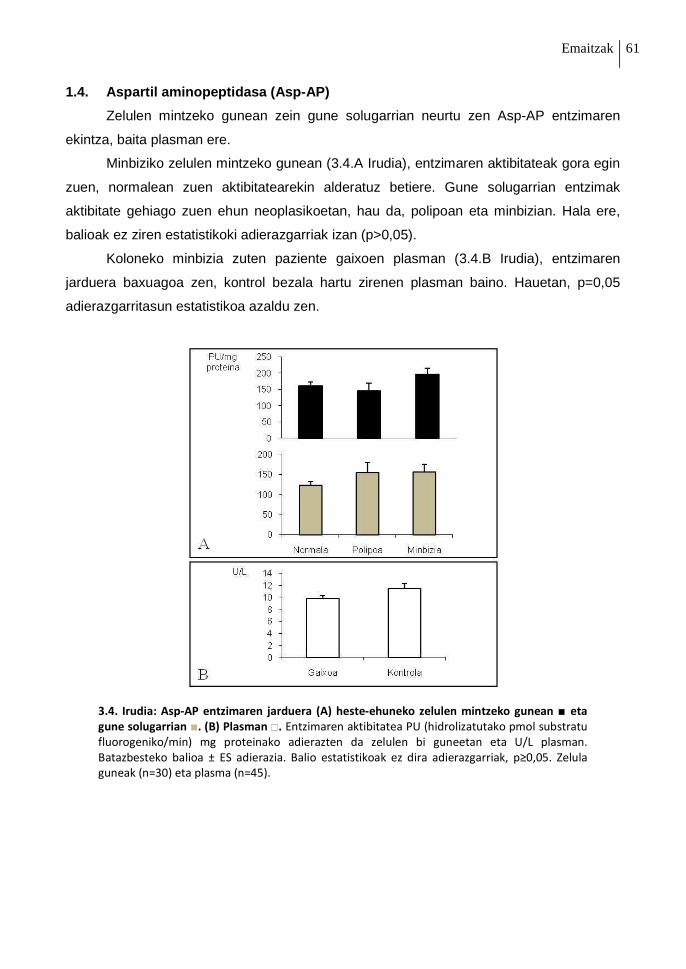
Emaitzak 61
1.4. Aspartil aminopeptidasa (Asp-AP)
Zelulen mintzeko gunean zein gune solugarrian neurtu zen Asp-AP entzimaren
ekintza, baita plasman ere.
Minbiziko zelulen mintzeko gunean (3.4.A Irudia), entzimaren aktibitateak gora egin
zuen, normalean zuen aktibitatearekin alderatuz betiere. Gune solugarrian entzimak
aktibitate gehiago zuen ehun neoplasikoetan, hau da, polipoan eta minbizian. Hala ere,
balioak ez ziren estatistikoki adierazgarriak izan (p>0,05).
Koloneko minbizia zuten paziente gaixoen plasman (3.4.B Irudia), entzimaren
jarduera baxuagoa zen, kontrol bezala hartu zirenen plasman baino. Hauetan, p=0,05
adierazgarritasun estatistikoa azaldu zen.
3.4. Irudia: Asp-AP entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta
gune solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistikoak ez dira adierazgarriak, p≥0,05. Zelula guneak (n=30) eta plasma (n=45).
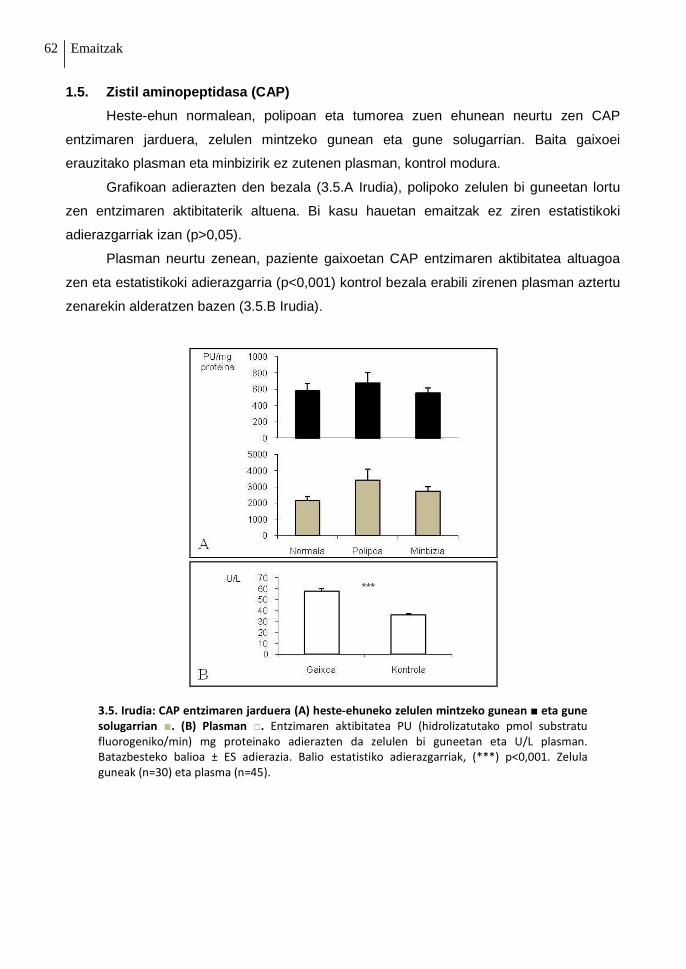
62 Emaitzak
1.5. Zistil aminopeptidasa (CAP)
Heste-ehun normalean, polipoan eta tumorea zuen ehunean neurtu zen CAP
entzimaren jarduera, zelulen mintzeko gunean eta gune solugarrian. Baita gaixoei
erauzitako plasman eta minbizirik ez zutenen plasman, kontrol modura.
Grafikoan adierazten den bezala (3.5.A Irudia), polipoko zelulen bi guneetan lortu
zen entzimaren aktibitaterik altuena. Bi kasu hauetan emaitzak ez ziren estatistikoki
adierazgarriak izan (p>0,05).
Plasman neurtu zenean, paziente gaixoetan CAP entzimaren aktibitatea altuagoa
zen eta estatistikoki adierazgarria (p<0,001) kontrol bezala erabili zirenen plasman aztertu
zenarekin alderatzen bazen (3.5.B Irudia).
3.5. Irudia: CAP entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (***) p<0,001. Zelula guneak (n=30) eta plasma (n=45).
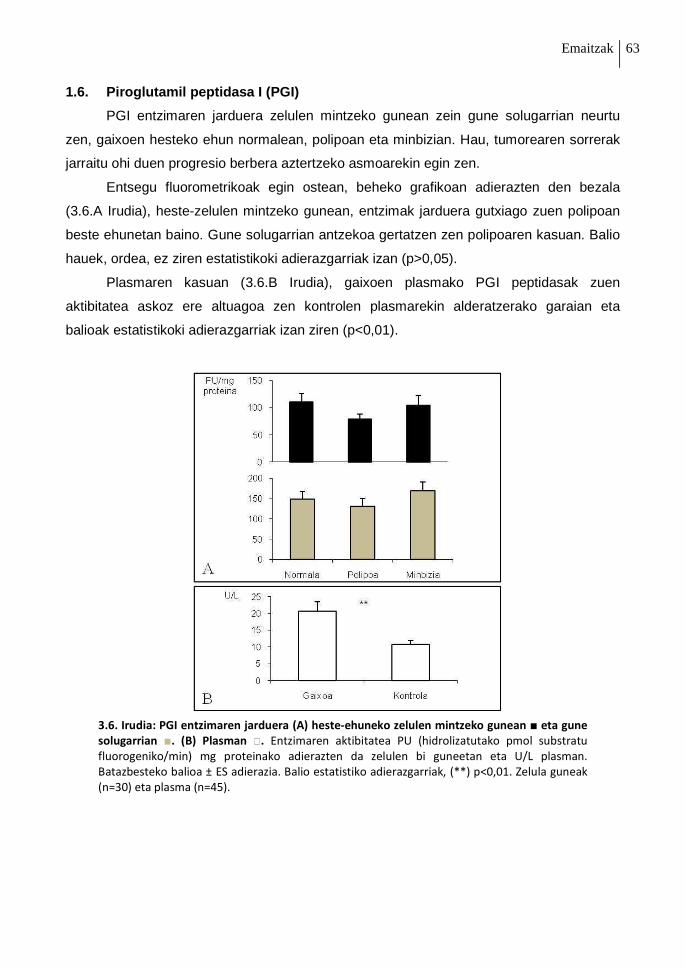
Emaitzak 63
1.6. Piroglutamil peptidasa I (PGI)
PGI entzimaren jarduera zelulen mintzeko gunean zein gune solugarrian neurtu
zen, gaixoen hesteko ehun normalean, polipoan eta minbizian. Hau, tumorearen sorrerak
jarraitu ohi duen progresio berbera aztertzeko asmoarekin egin zen.
Entsegu fluorometrikoak egin ostean, beheko grafikoan adierazten den bezala
(3.6.A Irudia), heste-zelulen mintzeko gunean, entzimak jarduera gutxiago zuen polipoan
beste ehunetan baino. Gune solugarrian antzekoa gertatzen zen polipoaren kasuan. Balio
hauek, ordea, ez ziren estatistikoki adierazgarriak izan (p>0,05).
Plasmaren kasuan (3.6.B Irudia), gaixoen plasmako PGI peptidasak zuen
aktibitatea askoz ere altuagoa zen kontrolen plasmarekin alderatzerako garaian eta
balioak estatistikoki adierazgarriak izan ziren (p<0,01).
3.6. Irudia: PGI entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (**) p<0,01. Zelula guneak (n=30) eta plasma (n=45).
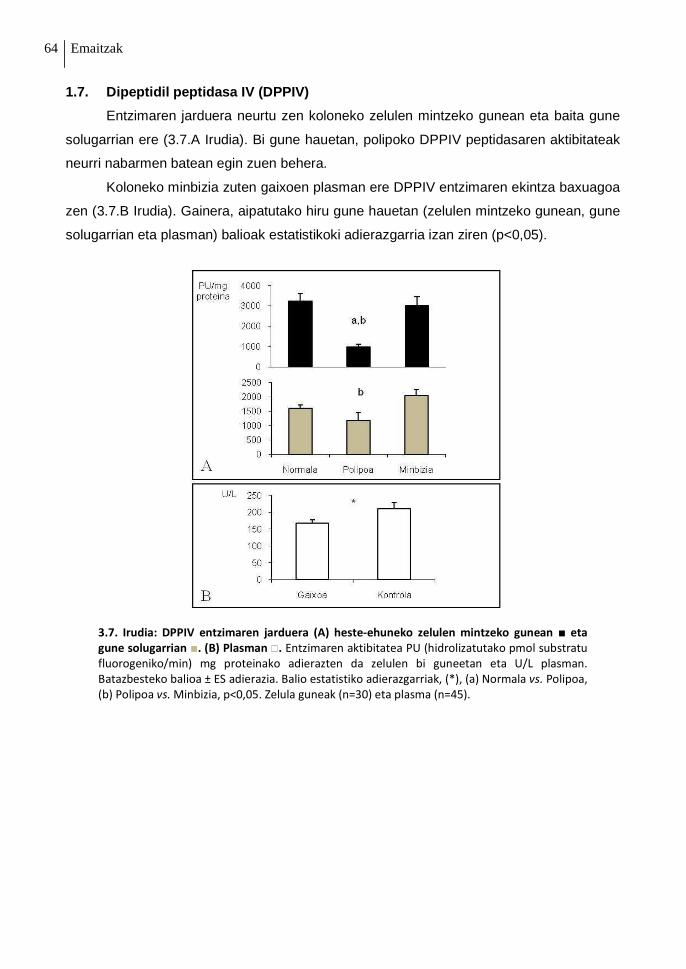
64 Emaitzak
1.7. Dipeptidil peptidasa IV (DPPIV)
Entzimaren jarduera neurtu zen koloneko zelulen mintzeko gunean eta baita gune
solugarrian ere (3.7.A Irudia). Bi gune hauetan, polipoko DPPIV peptidasaren aktibitateak
neurri nabarmen batean egin zuen behera.
Koloneko minbizia zuten gaixoen plasman ere DPPIV entzimaren ekintza baxuagoa
zen (3.7.B Irudia). Gainera, aipatutako hiru gune hauetan (zelulen mintzeko gunean, gune
solugarrian eta plasman) balioak estatistikoki adierazgarria izan ziren (p<0,05).
3.7. Irudia: DPPIV entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta
gune solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (*), (a) Normala vs. Polipoa, (b) Polipoa vs. Minbizia, p<0,05. Zelula guneak (n=30) eta plasma (n=45).
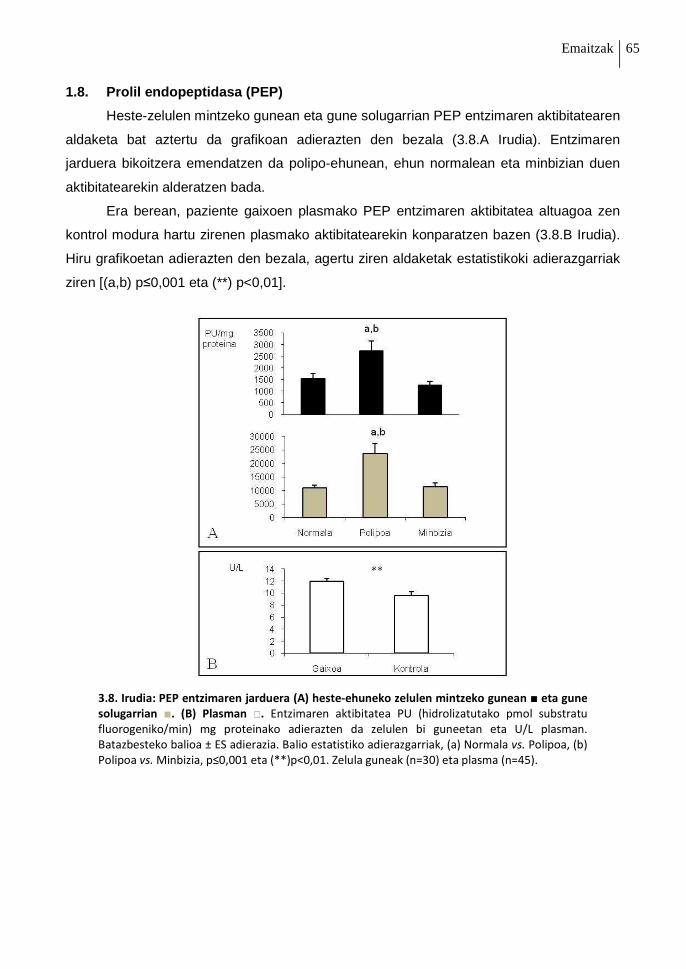
Emaitzak 65
1.8. Prolil endopeptidasa (PEP)
Heste-zelulen mintzeko gunean eta gune solugarrian PEP entzimaren aktibitatearen
aldaketa bat aztertu da grafikoan adierazten den bezala (3.8.A Irudia). Entzimaren
jarduera bikoitzera emendatzen da polipo-ehunean, ehun normalean eta minbizian duen
aktibitatearekin alderatzen bada.
Era berean, paziente gaixoen plasmako PEP entzimaren aktibitatea altuagoa zen
kontrol modura hartu zirenen plasmako aktibitatearekin konparatzen bazen (3.8.B Irudia).
Hiru grafikoetan adierazten den bezala, agertu ziren aldaketak estatistikoki adierazgarriak
ziren [(a,b) p≤0,001 eta (**) p<0,01].
3.8. Irudia: PEP entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (a) Normala vs. Polipoa, (b) Polipoa vs. Minbizia, p≤0,001 eta (**)p<0,01. Zelula guneak (n=30) eta plasma (n=45).
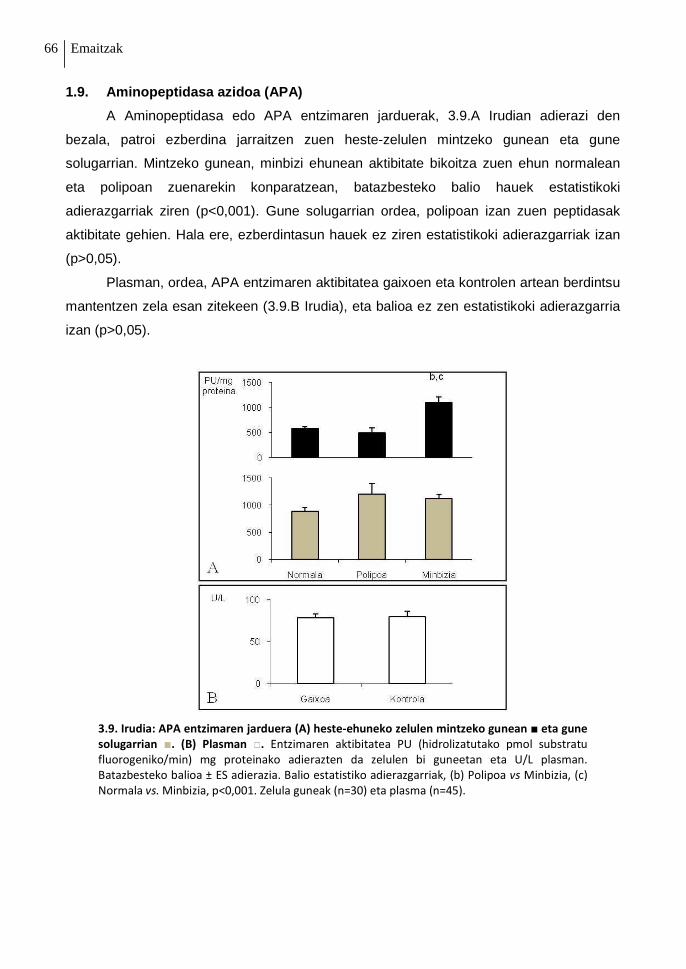
66 Emaitzak
1.9. Aminopeptidasa azidoa (APA)
A Aminopeptidasa edo APA entzimaren jarduerak, 3.9.A Irudian adierazi den
bezala, patroi ezberdina jarraitzen zuen heste-zelulen mintzeko gunean eta gune
solugarrian. Mintzeko gunean, minbizi ehunean aktibitate bikoitza zuen ehun normalean
eta polipoan zuenarekin konparatzean, batazbesteko balio hauek estatistikoki
adierazgarriak ziren (p<0,001). Gune solugarrian ordea, polipoan izan zuen peptidasak
aktibitate gehien. Hala ere, ezberdintasun hauek ez ziren estatistikoki adierazgarriak izan
(p>0,05).
Plasman, ordea, APA entzimaren aktibitatea gaixoen eta kontrolen artean berdintsu
mantentzen zela esan zitekeen (3.9.B Irudia), eta balioa ez zen estatistikoki adierazgarria
izan (p>0,05).
3.9. Irudia: APA entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■ eta gune
solugarrian ■. (B) Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen bi guneetan eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (b) Polipoa vs Minbizia, (c) Normala vs. Minbizia, p<0,001. Zelula guneak (n=30) eta plasma (n=45).
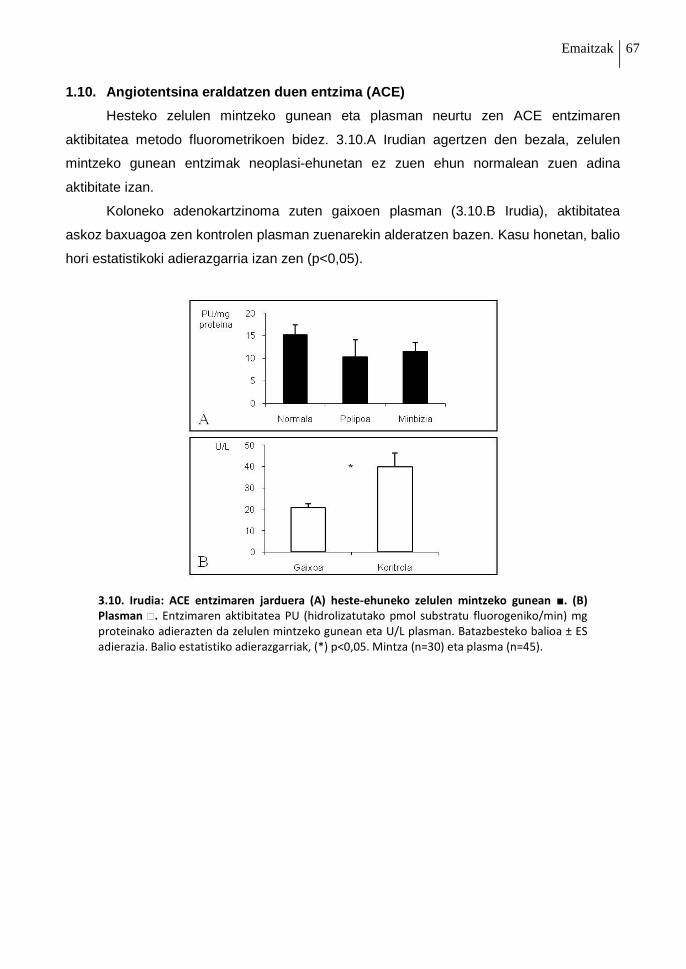
Emaitzak 67
1.10. Angiotentsina eraldatzen duen entzima (ACE)
Hesteko zelulen mintzeko gunean eta plasman neurtu zen ACE entzimaren
aktibitatea metodo fluorometrikoen bidez. 3.10.A Irudian agertzen den bezala, zelulen
mintzeko gunean entzimak neoplasi-ehunetan ez zuen ehun normalean zuen adina
aktibitate izan.
Koloneko adenokartzinoma zuten gaixoen plasman (3.10.B Irudia), aktibitatea
askoz baxuagoa zen kontrolen plasman zuenarekin alderatzen bazen. Kasu honetan, balio
hori estatistikoki adierazgarria izan zen (p<0,05).
3.10. Irudia: ACE entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■. (B)
Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen mintzeko gunean eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (*) p<0,05. Mintza (n=30) eta plasma (n=45).
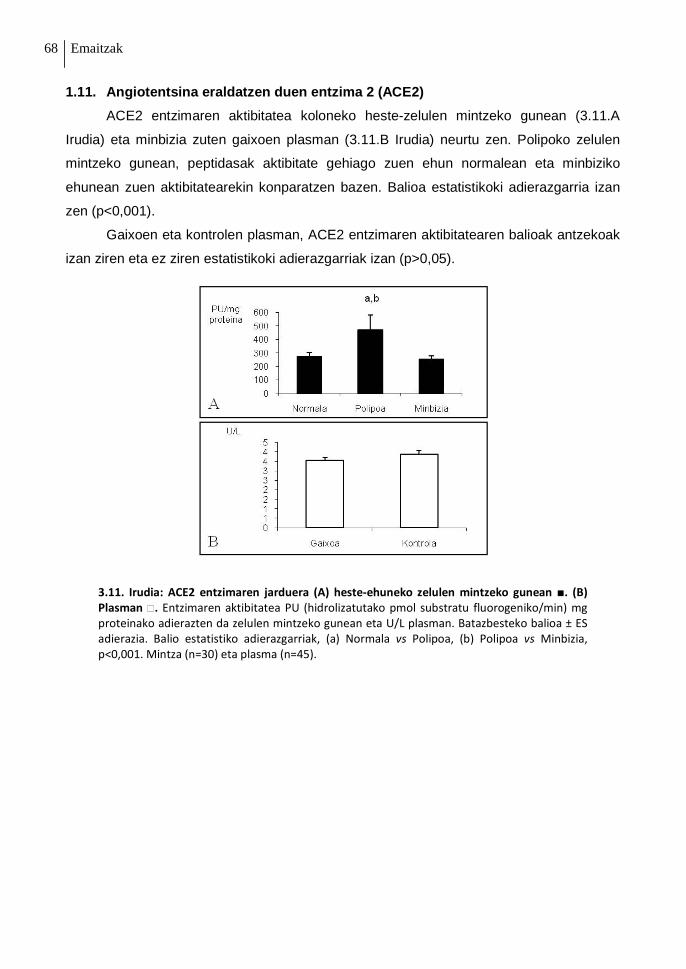
68 Emaitzak
1.11. Angiotentsina eraldatzen duen entzima 2 (ACE2 )
ACE2 entzimaren aktibitatea koloneko heste-zelulen mintzeko gunean (3.11.A
Irudia) eta minbizia zuten gaixoen plasman (3.11.B Irudia) neurtu zen. Polipoko zelulen
mintzeko gunean, peptidasak aktibitate gehiago zuen ehun normalean eta minbiziko
ehunean zuen aktibitatearekin konparatzen bazen. Balioa estatistikoki adierazgarria izan
zen (p<0,001).
Gaixoen eta kontrolen plasman, ACE2 entzimaren aktibitatearen balioak antzekoak
izan ziren eta ez ziren estatistikoki adierazgarriak izan (p>0,05).
3.11. Irudia: ACE2 entzimaren jarduera (A) heste-ehuneko zelulen mintzeko gunean ■. (B)
Plasman . Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen mintzeko gunean eta U/L plasman. Batazbesteko balioa ± ES adierazia. Balio estatistiko adierazgarriak, (a) Normala vs Polipoa, (b) Polipoa vs Minbizia, p<0,001. Mintza (n=30) eta plasma (n=45).
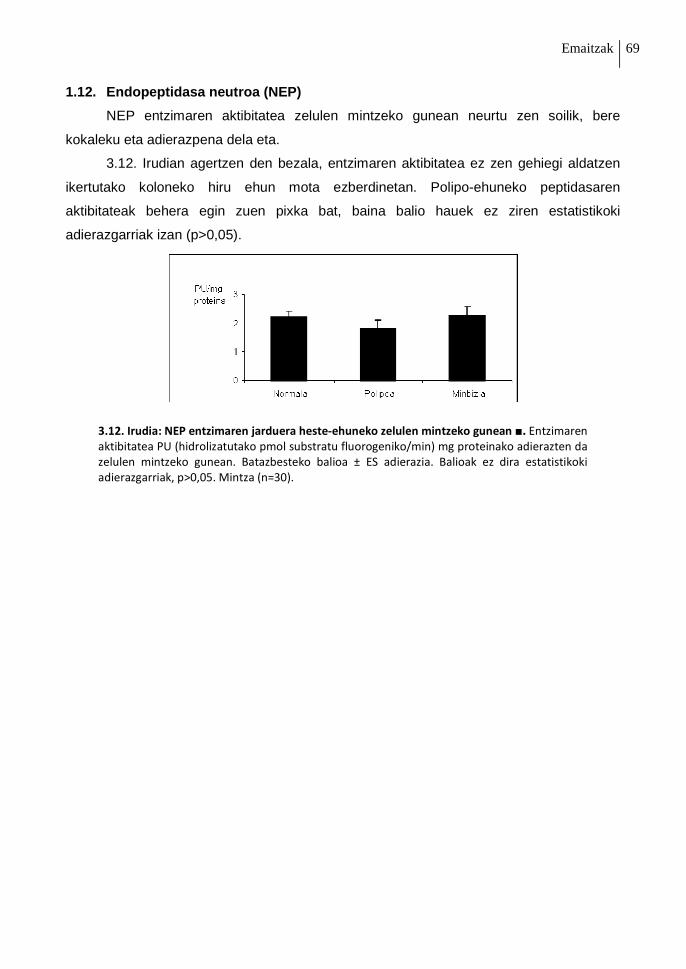
Emaitzak 69
1.12. Endopeptidasa neutroa (NEP)
NEP entzimaren aktibitatea zelulen mintzeko gunean neurtu zen soilik, bere
kokaleku eta adierazpena dela eta.
3.12. Irudian agertzen den bezala, entzimaren aktibitatea ez zen gehiegi aldatzen
ikertutako koloneko hiru ehun mota ezberdinetan. Polipo-ehuneko peptidasaren
aktibitateak behera egin zuen pixka bat, baina balio hauek ez ziren estatistikoki
adierazgarriak izan (p>0,05).
3.12. Irudia: NEP entzimaren jarduera heste-ehuneko zelulen mintzeko gunean ■. Entzimaren aktibitatea PU (hidrolizatutako pmol substratu fluorogeniko/min) mg proteinako adierazten da zelulen mintzeko gunean. Batazbesteko balioa ± ES adierazia. Balioak ez dira estatistikoki adierazgarriak, p>0,05. Mintza (n=30).

70 Emaitzak
2. Peptidasen jardueraren azterketa: Graduak eta Es tadioak
Memoria honen 3. helburua aztertu ahal izateko peptidasen entzima-jarduera
konparatu zen minbizi-ehunaren gradu eta estadio ezberdinetan. Orain arte bezala, bai
heste-zeluletako mintzeko gunean zein gune solugarrian neurtu zen.
Plasmaren kasuan, hesteko minbizia zuten paziente gaixoen plasman aztertu ziren
ikerketako entzimen jarduerak. Pairatzen zuten minbiziaren gradu eta estadio
ezberdinetan konparatu ziren aldaketak.
Graduak, OME-ren sailkapen sistema erabiliz, taldekatuta aztertu ziren, gradu
baxuan (G1+G2) eta gradu altuan (G3).
Minbiziaren estadio-sailkapena egiteko heste laginei egokitutako TNM estadio-
sistema erabili zen (Materialak eta Metodoak, 4.2. Azpiatala). Hemen ere taldekatuta
aztertu ziren, estadio baxuan (T1+T2) eta estadio altuan (T3+T4).
2.1. Heste-zeluletako entzimen jarduera
Ikerketa lan hau egiteko aukeratu ziren peptidasen aktibitateak minbizia zuten
ehunean aztertu ziren. Lagin hauek era berean, graduaren eta estadioaren arabera banatu
ziren. Kasu hauetan guztietan, ordea, ez zen emaitza estatistikoki adierazgarririk azaldu
eta dokumentu idatziaren amaieran aurkezten dira tauletan (III. Eranskina).
2.2. Plasmako entzimen jarduera
Plasma-laginak, pazienteek pairatzen zuten hesteko minbiziaren graduaren arabera
banatu ziren ikerketaren atal honetan (3.13.A Irudia).
Gradu baxuko eta gradu altuko laginetan egin zen plasmako entzimen jardueraren
konparaketa. Peptidasa-jardueraren aldaketa adierazgarria izan zuten entzimak PSA,
DPPIV, APA eta PEP ziren. Entzima hauetan guztietan, hesteko minbiziaren gradu
baxuetan plasmako peptidasen aktibitatea altuagoa izan zen minbiziaren gradu altuetan
baino (p<0,05).
Era berean, minbiziaren estadioaren arabera banatu ziren laginak bigarren
grafikoan agertzen den bezala (3.13.B Irudia). Hemen, plasmako PGI entzimaren jarduera
baxuagoa izan zen hesteko minbiziaren estadio baxuetako laginetan, tumorearen estadio
altuetako laginetan baino. Gainera, balio hau estatistikoki adierazgarria izan zen (p<0,05).
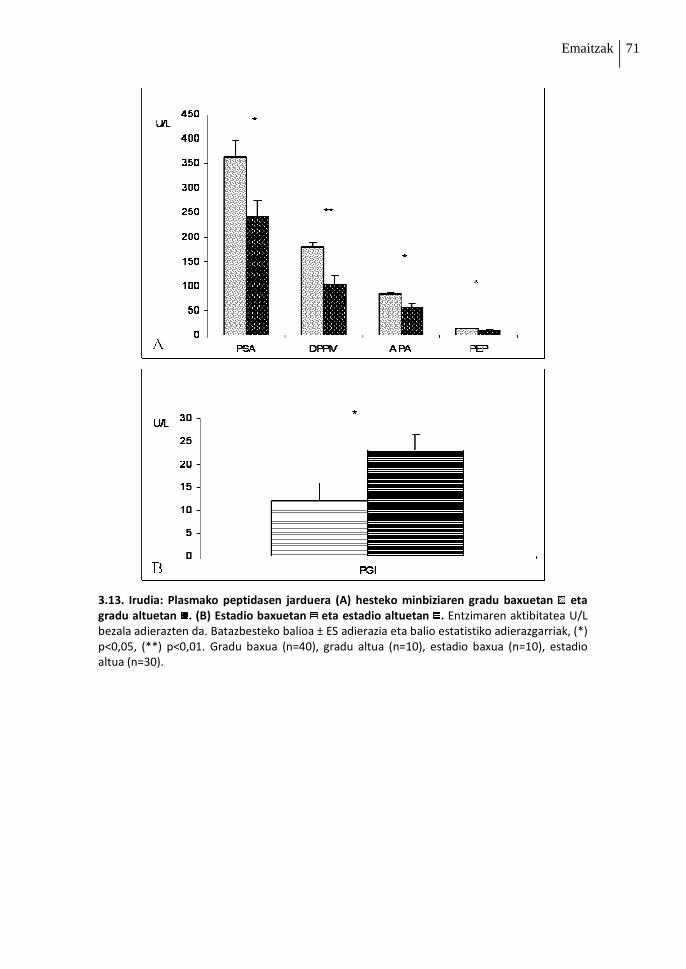
Emaitzak 71
3.13. Irudia: Plasmako peptidasen jarduera (A) hesteko minbiziaren gradu baxuetan eta
gradu altuetan . (B) Estadio baxuetan eta estadio altuetan . Entzimaren aktibitatea U/L bezala adierazten da. Batazbesteko balioa ± ES adierazia eta balio estatistiko adierazgarriak, (*) p<0,05, (**) p<0,01. Gradu baxua (n=40), gradu altua (n=10), estadio baxua (n=10), estadio altua (n=30).

72 Emaitzak
3. Peptidasen aktibitatearen laburpena
Emaitzen atal hau ulerterrazagoa izan dadin eta pixka bat laburtzeko asmoarekin,
eskema bat azaltzen da ondoren.
Hesteko zeluletan (normala, polipoa eta minbizia), bai mintzeko gunean edota gune
solugarrian, estatistikoki adierazgarriak (p<0,05) izan ziren peptidasa-jardueraren
aldaketak ondorengo taulan adierazten dira:
3.1. Taula: Heste-zeluletako peptidasa-jardueraren aldaketa estatistikoki adierazgarriak.
DPPIV PEP ACE2 APA
Mintza Polipoa ↓ Polipoa ↑ Polipoa ↑ Minbizia ↑
Solugarria Minbizia ↑ Polipoa ↑
Gaixoen eta kontrolen plasmako peptidasak aztertu zirenean ere, emaitza
estatistikoki adierazgarriak (p<0,05) atzeman ziren ikertutako ondorengo entzima hauen
jardueretan:
3.2. Taula: Plasma-lagineko peptidasa-jardueraren aldaketa estatistikoki adierazgarriak.
CAP PGI PEP DPPIV ACE
Gaixoa ↑ ↑ ↑ ↓ ↓
Heste lodiko minbizia zuten gaixoen plasmako peptidasen aktibitatea minbiziaren
histologi-graduaren eta estadioaren araberako banaketa egitean, emaitza estatistikoki
adierazgarriak (p<0,05) atzeman ziren ondorengo entzima hauetan:
3.3. Taula: Plasmako peptidasa-jardueraren aldaketa adierazgarriak histologi-graduaren eta
estadioaren arabera. %: peptidasen aktibitate aldaketak gradu altua gradu baxuarekiko, eta estadio altua estadio baxuarekiko.
DPPIV PSA APA PEP
Gradu altua %42,21↓ %33,52 ↓ %32,78↓ %17,15↓
PGI
Estadio altua %47,26↑

Eztabaida


Eztabaida 75
4. EZTABAIDA
1. Peptidasen entzima-jarduera mukosa normalean eta heste lodiko
neoplasietan
Zelularen funtzio fisiologikoetako batzuk ezberdintzapena, hazkuntza, zahartzapena
eta apoptosia dira. Patologietan, baita minbizian ere, funtzio garrantzitsu hauek asaldatuta
egon ohi dira. Aldaketa hauek gertatzeko arrazoiak ezberdinak izan daitezke, baina
gehienetan mutazioak gertatzen dira DNA mailan. Ondorioz, mRNA-ren adierazpenean,
proteinaren adierazpenean edota proteina beraren funtzionaltasunean arazoak egon
daitezke. Horregatik entzima baten aktibitatea da bere funtzioan arazoak dauden jakin
ahal izateko parametrorik egokiena (Wang et al., 2009; Larrinaga et al., 2010a). Ikerlan
honen helburua da beraz, proteina pool zabal baten entzima-aktibitatearen aldaketak
aztertuz, patologiaren zati bat ulertzen saiatzea.
Bibliografian aurkitu ditzakegun lan gehienek minbizia dagoen ehuneko kartzinoma
eta bere aldamenean baina osasuntsua den gunea alderatzen dituzte. Garrantzi handikoa
da honelako konparaketak egitea, gaixoaren ehun berbera izanik, kartzinogenesiaren
ondorioz bertan zein aldaketa gertatu diren ikusita, errealitatean gertatutako prozesua
interpretatzen saia gaitezkeelako.
Lan honetan ikertu den ehuna heste lodia izan da. Heste lodiko minbizien %98a
adenokartzinomak dira eta koloneko kartzinogenesiari buruz hitz egiten denean, jakina da
mukosa osasuntsua - polipo adenomatsua - adenokartzinoma progresioa betetzen dutela
(Jass, 2001; Leslie et al., 2002). Sekuentzia honi esker, beste minbizi mota batzuetan
gertatzen ez den bezala, ehun osasuntsua eta kartzinomaren erdibideko prozesu batean,
kontrolik gabeko hazkuntza jasan duten (baina beti ere onbera izanik) zelulen fisiologia
aztertzeko aukera ere eskaintzen du. Minbiziaren gaiztotzeko bilakaera eta agresibitatea
aztertzeko eredu egokia da beraz. Koloneko neoplasiak ez du sintomarik adierazten fase
berantiarrera arte. Honek, diagnosia zailtzen du eta minbizia zabaldu eta hedatu egiten da
kirurgia bidezko sendaketa nekezagoa eginez. Klinikaren helburu nagusietako bat da,
ikerketen ondorioak aintzat hartuz, neoplasia hauek oraindik sendagarriak diren fase
goiztiarretan diagnostikatzea, hots, polipo adenomatsu erauzgarrietan aurkitzea.
Minbiziaren gaiztotze bilakaera, hein handi batean, geneetan gertatzen diren
mutazioen pilaketen ondorioa da, aipatu bezala. Onkogeneetan, tumoreen gene
erreprimitzaileetan eta DNA zuzentzen duten geneetan gertatzen diren mutazioei egotzi
zaie bilakaera honen arrazoia. Bibliografian, gene ugariren adierazpena asaldatuta egon

76 Eztabaida
ohi dela deskribatu da eta heste mukosa - adenoma - kartzinoma bilakaeran ezinbesteko
diren geneak aurkitzeko asmoz egin diren lanetan islatzen dira aipatutako aldaketak (Lee
et al., 2006; Sillars-Hardebol et al., 2010). Bestalde, DNA-mikrosateliteen
ezegonkortasunak, DNA-metilazioak eta kromosometako zati batzuen galerak ere
zerikusia dutela deskribatu da (Jass, 2001; Leslie et al., 2002; Spisák et al., 2010). Izan
ere, de novo sortzen diren adenokartzinomen portzentajea zenbatekoa den ez jakin arren,
baxua dela uste da (Jass, 2001; Kumar et al., 2008).
DNA-ren adierazpena ikertzea ezinbestekoa da, baina proteinen adierazpena eta
funtzionaltasuna aztertzea ere garrantzi handikoa da minbiziaren garapenean. Minbiziak
zabaldu eta hedatu ahal izateko molekula ugariren beharra dauka, horien artean entzima
proteolitikoak esaterako. Proteasa hauek ezinbesteko funtzioa betetzen dute
kartzinogenesiaren hasierako faseetan, inbasioan, angiogenesian eta metastasian
adibidez. Proteasen substratuen artean, hazkuntza faktoreak, kimiokinak eta zitokinak
daude, baina baita hazkuntza inhibitzen duten beste hainbat substratu ere (Flores-
Reséndiz et al., 2009). Talde oso zabala eta heterogeneoa da proteasena, familia ugari
barneratzen dituena, hala nola, katepsinak, kalikreinak, mintzeko metaloproteasak…
Hauetako batzuk, minbizi ezberdinen markatzaile gisa erabiltzen dira egun klinikan.
Badaude, katepsina eta kalikreina familiako proteasa ugariren funtzioa adenoma-
adenokartzinoma sekuentziarekin alderatzen duten hainbat lan. Kopuru altuagoak aurkitu
dituzte animali modelo eta in vitro esperimentuetako ehun neoplasikoetan (Shuja et al.,
1991; Iacobuzio-Donahue et al., 1997; Kuester et al., 2008; Talieri et al., 2004, 2009; Kim
et al., 2010). Matrizeko metaloproteasen (MMP) familiako kide askoren adierazpena eta
kokapena ikertzen duten beste horrenbeste lanetan ere adierazpen aldaketak
esperimentatu dituzte bilakaeran zehar (Zucker et al., 2004; Herszényi et al., 2008;
Groblewska et al., 2010).
Proteasek bezala, peptidasek funtzio garrantzitsua betetzen dute prozesu
fisiologikoetan, baina baita patologikoetan ere, minbiziarekin erlazionatu izan direlarik
(Hashida et al., 2002; Röcken et al., 2007; Varona et al., 2007; Blanco et al., 2008; Perez
et al., 2009; Larrinaga et al., 2010a, 2010b). Peptidasak, sarreran aipatu gisa, peptido
bioaktiboak eraldatzen dituzten entzimak dira (Antczak et al., 2001). Gainera, MMP-ak eta
peptidasak elkarrekin lan egiten dute, hala nola, APN, DPPIV eta PEP gaixotasun
neoplasikoen eta ez-neoplasikoen bilakaeran (Antczak et al., 2001; Hashida et al., 2002;
Petrovic et al., 2004; Selvakumar et al., 2004; Gaggar et al., 2008; Liu et al., 2008). Beraz,

Eztabaida 77
proteasen aktibitatea minbizietan aldatua dagoela azaldu den bezala peptidasen
aktibitatea ere aldatua egon litekeela pentsa daiteke.
NEP eta ACE, endopeptidasa eta karboxipeptidasa hurrenez-hurren, tumore-
markatzaileak dira. Hauen adierazpena, bai mRNA mailan zein proteina mailan, ikertu da
heste lodiko minbiziaren progresioarekin loturik (Ogawa et al., 2002; Sumitomo et al.,
2005; Röcken et al., 2007; Koga et al., 2008; Del Rio et al., 2010; Neo et al., 2010). Baina
oro har, heste lodiko minbiziaren bilakaera peptidasen familiako kideen adierazpenarekin
eta aktibitatearekin erlazionatzen dituzten ikerlanak ez dira horrenbeste ezagutzen. Esan
bezala, adierazpena ikertzea garrantzitsua da, baina entzima baten funtzioak asaldaturik
dauden edo ez ikertzeko, aktibitatea da hobekien adierazten duen parametroa.
Ikerlan honetan aztergai izan ziren entzimen jarduerak heste lodiaren
kartzinogenesian zehar aldaketarik jasan zuten edo ez aztertzean, gehienetan
aktibitatearen gora-beherak gertatzen zirela esperimentatu zen: bai fase goiztiarrean
(adenoman) zein fase berantiarrean, bai zelularen mintzeko gunean edota gune
solugarrian, zein plasman. Heste-zeluletan lortutako emaitzak azaltzeko intentzioarekin,
aipatzekoa da aztertutako entzima hauetatik, APA, DPPIV, PEP eta ACE2 peptidasek
jasan zituztela beraien aktibitatean aldaketa estatistikoki adierazgarriak (p<0,05). Mintzeko
DPPIV izan ezik, beste hiru entzimen aktibitateek igoera bat izan zuten, APA eta DPPIV
solugarriak heste lodiko adenokartzinoma ehunean eta PEP, ACE2 eta mintzeko DPPIV
entzimak polipo adenomatosoan, beti ere, ehun osasuntsuarekin konparatzean. DPPIV
eta PEP entzimen kasuan, bai heste-zelulen mintzeko gunean zein gune solugarrian
esperimentatu ziren aldaketak. Aldiz, APA eta ACE2 entzimetan, soilik mintzeko gunean
gertatu ziren aldaketak (Emaitzen ataleko 3.1. Taula).
Beraz, entzimen aktibitatearen aldaketak modu ezberdin batean eman ziren. Hots,
ekintzak ez ziren zentzu berberean aldatu entzima guztietan, ez minbiziaren bilakaeraren
arabera, ezta zelularen banaketa guneen arabera ere. Hau dela eta, honekin pentsa
daiteke, gertatzen diren asaldurak ezin direla proteolisi orokorrean gertatzen diren
aldaketa orokorrengatik izan, baizik eta aldaketa selektiboen ondorio liratekeela eta
entzima hauek eragina izango luketela heste lodiko kartzinogenesian. Edo posible da ere
prozesua alderantziz izatea, baliteke kartzinogenesiaren ondorio gisa entzima hauen
aktibitatea aldatzea (Varona et al., 2007; Blanco et al., 2008, 2010; Perez et al., 2009).
Aipatu bezala, hesteko polipo fasean aztertutako entzima batzuen ekintzaren
aldaketa bat nabaritu zen ehun osasuntsuarekin eta minbiziarekin alderatzerakoan.
Aldaketa hau ordea, ez zen neurri berberekoa izan entzima ezberdinetan, ezta norabide

78 Eztabaida
berdinekoa ere. Entzima batzuen aktibitateak igoera bat jasan zuen eta besteetan
beherakada. Hala ere, aldaketa adierazgarriak izan zituzten entzimak mintzeko guneko
PEP, ACE2 eta DPPIV, zein gune solugarriko PEP izan ziren.
Heste lodiko neoplasiaren fase goiztiar honetan geroago garrantzitsuak diren
prozesuak gertatzen dira. Hasiera batean, kontrolik gabeko hazkuntza gertatzen da,
onbera dena, baina gero gaiztotzera egin dezakeena (Leslie et al., 2002; Kumar et al.,
2008). Gialeli et al.-ek berriki argitaratu duten (2011) lanean diotenez, MMP-ak dira,
minbiziaren bilakaeran zehar, mikroingurumenean gertatzen diren aldaketen bitartekari
nagusiak. Heste-zelulen hazkuntzarekin erlazionatu dituzten MMP eta peptidasa ugari
daude. Hauek, minbiziaren progresiorako ezinbesteko garrantzia duten funtzioak dituzte,
hala nola, matrize extrazelularreko molekulei lotzea, angiogenesia, zelularen hazkuntza,
hedapena eta metastasia (Heslin et al., 2001; Herszényi et al., 2008; Roy et al., 2009).
Adierazgarriak izan diren peptidasa-jarduera hauek ere erlazionatu izan dira
minbiziarekin harremana duten honako hainbat prozesutan. Mintzeko DPPIV-ak matrizeko
kolagenoari eta fibronektinari lotutzeko guneak dituela deskribatu da, ondorioz, entzima
honek ere matrizea hidrolizatzeko gaitasuna duela iradoki da (Tan et al., 2004; Havre et
al., 2008). PEP eta ACE2 entzimen aktibitate aldaketak hanturazko gaixotasunekin
erlazionatu dira, aireztapen-hodietan eta digestio-hodietan hurrenez-hurren (Gaggar et al.,
2008; Byrnes et al., 2009). Entzima hauek ere errenina angiotentsina sistemaren (RAS)
partaide izanik, odol-hodi berrien sorreran eta zelulen hazkuntzan eragina dute (Carrera et
al., 2006; Ruiz-Ortega et al., 2007). Peptidasetan ikertu den bezala, beste proteasa
batzuetan ere aurkitu dira aldaketak, adibidez matrizeko metaloproteasetan. MMP-1-a,
adenoma fase goiztiarrean gain-adierazia dagoela deskribatu da, gero adenokartzinoma
fasean nabaritzen ez dena (Tahara et al., 2010). MMP-en emendapen berbera azaldu
dute gaztaroko polipoen ehun zein gernuetan (Manfredi et al., 2009). Gure ikerketako PEP
eta ACE2 peptidasen aktibitateak ere jokaera berbera adierazi zuten. Ondorioz, peptidasa
hauen aktibitate aldaketek adenomaren fase goiztiarretan eraginen bat izan lezaketela
pentsa daiteke.
Gure ikerlaneko emaitzekin jarraituz, ikerturiko beste peptidasa batzuen aktibitatea
heste lodiko minbizian aldatu zen estatistikoki modu adierazgarrian, polipoarekin eta ehun
osasuntsuarekin konparatzean. Hau da esaterako, heste-zelulen mintzeko APA-rekin eta
gune solugarriko DPPIV-arekin atzeman zena. Bi entzimen kasuan, heste lodiko
adenokartzinoman beraien ekintzaren emendapen nabaria aztertu zen. Antzeko gertaerak
deskribatu dituzte ere beste peptidasa batzuekin, metionin aminopeptidasa-2 adibidez

Eztabaida 79
(Selvakumar et al., 2004). KLK6 kalikreina (Kim et al., 2010) zein MMP-2 eta MMP-9-aren
kasuan ere, igoera bat deskribatu dute heste lodiko adenokartzinoman, aitzitik, adenoma
goiztiarretan esperimentatzen ez dena (Heslin et al., 2001; Herszényi et al., 2008;
Groblewska et al., 2008). Heste-zelulen mintzeko guneko eta gune solugarriko DPPIV
aktibitatearen ezberdintasunari dagokionez, deskribatu izan da, MMP familiako kide
batzuek kontrako ekintzak izan ditzaketela. Ekintza hauek ezberdinak izan daitezke
minbiziaren bilakaeran zehar edo baita tumorearen estadioaren arabera, entzimen
kokagunearen arabera edo substratuaren arabera ere (Gialeli et al., 2011). Antzeko
emaitza esperimentatu dugu gure ikertaldean ere urte hauetan zehar eginiko lanetan.
Giltzurruna edo aho barrunbea bezalako organo ezberdinetan izandako emaitzetan
peptidasen aktibitatean aldaketak esperimentatu dira minbizia ehun osasuntsuarekin
alderatzean (Irazusta et al., 2002; Larrinaga et al., 2005; Varona et al., 2007; Perez et al.,
2009; Larrinaga et al., 2011). Ondorioz, peptidasa batzuk adenokartzinoman edo
bilakaeraren fase berantiarrean izan lezakete eraginen bat, edo alderantziz,
kartzinogenesiko fase berantiarraren ondorio gisa, aldaketak gertatzen dira peptidasa
hauetan.
Gure emaitzak bibliografian agertzen direnekin alderatzen denean, aldeko eta
kontrako erantzunak jaso dira. Badaude, gutxi diren arren, heste lodiko adenokartzinoman
aktibitatea aztertzen dutenak, baita beste minbizietan aztertzen duten batzuk ere.
Aldaketak jasan dituzten entzimak banan-banan aztertuko dira jarraian.
DPPIV entzima serina proteasa bat da. Heste lodiko neoplasian duen funtzioa
deskribatu izan da bibliografian, baina gehien, plasmako peptidasari buruz hitz egin da eta
idazki honen hurrengo atalean aipatuko da zabalago horri buruz (Cordero et al., 2000,
2009; De Chiara et al., 2009, 2010). Peptidasa hau hainbat minbizietan adierazten da eta
giltzarri da tumorogenesian zein metastasian (Antczak et al., 2001; Nanus, 2003; Pro et
al., 2004; Carl-McGrath et al., 2006; Havre et al., 2008). Informazio asko dago
ektopeptidasa honen inguruan, funtzio ugari ditu eta oso ezberdinak beraien artean. Ehun
mota gehienetan agertzen da eta ekintza selektiboa duela esaten da, hau da, zelularen
araberako funtzioa duela (Pro et al., 2004; Carl-McGrath et al., 2006; Havre et al., 2008).
Gure emaitzetan lortutako aldagarritasuna, hots, zelularen gunearen araberako emaitza,
peptidasaren funtzio anitzari egotzi diezaiokegu.
DPPIV entzimak jakina da kimiokinen aktibitatea eraldatzen duela, zelularen kanpo-
matrizeko kolagenoa eta fibronektina bezalako proteinei ere lotzen da, zelularen
metabolismoa erregulatuz (Tan et al., 2004; Havre et al., 2008). Hala nola, adenosina

80 Eztabaida
deaminasarekin duen elkarrekintzaz HT-29 koloneko minbizi zeluletan, DPPIV-aren
jarduera jaitsi egiten dela argitaratu da (Tan et al., 2004). Minbizietan, zelularen kanpo
likidoko adenosinaren kontzentrazioa igo egiten dela deskribatu dute, honek tirosin
fosfatasaren aktibitatearen igoera bat dakar eta ondorioz, ERK1/2 MAP kinasaren
fosforilazio gutxitzea eta DPPIV-aren mRNA zein proteina adierazpenaren jaistea ere (Tan
et al., 2006). Yoshioka et al.-ek (1991) aipatzen duten moduan, mintzeko DPPIV-aren
aktibitateak gorantz egiten du gizakiaren koloneko minbiziko Caco-2 zelula
ezberdintzatuetan, baina sintesi ugariagoaren ondorio izan litekeela aipatzen du.
Horregatik, proteinaren sintesia eta adierazpena zein aktibitatea alderatuz, ikerketa
sakonagoa egin beharko litzateke hau argitzeko.
Entzima honen multifuntzioaren adibide gisa, neoplasia ezberdinetan duen
aktibitatea eta adierazpena deskribatu da ikerlan askotan: handiagoa da T-zelulen linfoma
linfoblastikoan, tiroideko minbizian, biriketako adenokartzinoman eta zelula basaletako
kartzinoman. Txikiagoa da, ordea, melanoma gaiztoetan, buruko eta lepoko zelula
ezkatatsuetako kartzinoman eta giltzurruneko neoplasia ohikoenetan (Pro et al., 2004;
Carl-McGrath et al., 2006; Perez et al., 2009; Varona et al., 2010).
A Aminopeptidasa (APA) mintzartekoa izan arren, solugarria den isoforma ere
deskribatu da (Nanus, 2003). Beste hainbat ektopeptidasa bezala, entzima hau
kartzinogenesiarekin erlazionatu izan da bibliografian. Hain zuzen, angiogenesia, zelula-
hazkuntza, minbiziaren hedapena eta gaixotasunen bilakaerarekin (Nanus, 2003; Ino et
al., 2004; Marchiò et al., 2004; Perez et al., 2009). Entzimaren adierazpena bularreko zein
heste lodiko ehun normaletan, eta neoplasikoetan (onbera eta gaiztoa) ikertu duen idazki
bat dago, non, kartzinogenesian zehar perizito aktibatuetako APA adierazpena gero eta
indartsuagoa dela deskribatzen duen. Perizitoak angiogenesiko odol-hodi berriak
inguratzen dituzten zelulak dira, uzkurtzeko gaitasuna dutenak eta endotelio-zelulen
hazkuntza erregulatzen dutela deskribatu da (Schlingemann et al., 1996; Marchiò et al.,
2004). Angiotentsina II (Ang II), odol-hodien hertsatzailea dena, A aminopeptidasaren
substratu garrantzitsuenetakoa da. Peptidasaren adierazpenak eta aktibitateak
minbizietako odol-hodietan gora egiten duela deskribatua izan da saguetan, odol-hodi
normalekin alderatzerakoan. Tumorearen hodi-sistemako funtzio-adierazle bezala erabili
daitekeela adierazi da (Marchiò et al., 2004).
Memoria honetako emaitzak bat datozela esan dezakegu, gure kasuan, entzimaren
jarduera aktiboagoa da heste lodiko kartzinoman, polipoarekin eta ehun normalarekin
konparatuz gero. Emaitza berberak aurkitu dira ere, umetoki-zerbixeko kartzinomaren

Eztabaida 81
progresioan (Ino et al., 2004) eta gure taldeak buruko eta lepoko zelula ezkatatsuetako
kartzinoman egindako ikerlanean (Perez et al., 2009). APA entzimaren jarduera
angiogenesiari lotuta aurkitzen dugu bibliografian kasurik gehienetan. Minbiziak zabaldu
eta hedatu ahal izateko odol-hodi berrien beharra izaten du. Berriki aipatu bezala, Ang II,
RAS-eko protagonista nagusia da eta proteina honen adierazpena hertsiki lotua dago odol
presioarekin, baina baita zelularen hazkuntza edo apoptosiarekin ere (Carrera et al., 2006;
Ruiz-Ortega et al., 2007). Honi lotuta, azken urteetan angiogenesia asko ikertzen den
prozesua da eta baita honek tumorogenesiarekin izan dezakeen erlazioa ere. Honela
adierazten du Raica et al.-ek (2009), neoplasia gaiztotu aurreko baldintzetan egin duten
ikerketan. Angiogenesiak minbiziaren inbasioa adierazten ez duen arren,
tumorogenesiaren hasierako faseetako gertakaria dela diote.
ACE2 entzimari dagokionez, heste lodiarekin erlazionatzen duten oso lan gutxi
daude, 9SARS-koronabirusaren hartzaile bezala deskribatzen dutenak batzuk (Li et al.,
2003; Ren et al., 2006) eta saguetan, hesteetako hanturazko gaixotasunekin erlazionatzen
dutenak (Byrnes et al., 2009). ACE2 entzimaren adierazpena eta aktibitatea sagu eta
arratoietan aztertu da, mRNA-adierazpena saguetan handiagoa izanik arratoietan baino,
aktibitatea ordea bi espezieetan antzekoa da (Gembardt et al., 2005). Gure emaitzetan,
mintzeko ACE2 entzimaren aktibitateak emendapen bat izan zuen polipoan, hesteko
mukosa eta minbiziarekin konparatuz gero. Bibliografian, ordea, ez da heste lodian
aktibitatea ikertzen duen lanik aurkitu emaitzak alderatu ahal izateko.
Bestalde, gure ikertaldeak ACE2 eta ACE proteinen eta mRNA-ren adierazpena eta
aktibitateak aztertu ditu giltzurruneko lau neoplasia mota ohikoenetan. Aktibitateek jaitsiera
nabaria jasan zuten lau neoplasietan ehun osasuntsuarekin alderatzerakoan (Blanco et
al., 2010; Larrinaga et al., 2010b). Beste ikertalde batzuek ere adierazpenaren gutxitzea
esperimentatu dute area-hodiko adenokartzinometan (PDAC). Ang II-aren pilaketak ACE2
entzimaren erregulazio jaitsieran eragiten duela diote, areako minbizi zelulen hazkuntza
ahalbidetuz kultur-ingurunean (Zhou et al., 2009). Feng et al.-ek (2010) antzeko emaitzak
lortu dituzte ere, biriketako zelula ez-txikien minbizian (NSCLC). Aitzitik, gibeleko lesioan
ACE2-aren adierazpena emendatuta dagoela argitaratu dute. Honen produktua den Ang
(1-7) molekulak, Ang II-aren kontrako eraginak dituela adierazi da, hau da, zelulen
hazkuntzaren inhibizioa eragiten duela (Neo et al., 2010). Talde honek berak, saguaren
heste lodiko kartzinomaren ondoriozko gibeleko metastasian, RAS-aren oztopoak
9 SARS: Severe acute respiratory syndrome-coronavirus

82 Eztabaida
tumorearen hazkuntza ekiditen duela esperimentatu du. Beraz, badirudi ACE2 entzimak
RAS sistemaren bidez heste lodiko minbiziaren hazkuntzan eragina izan dezakeela.
Prolil endopeptidasa (PEP) nagusiki gune solugarriko entzima bezala deskribatu
den arren, mintzeko isoforma ere purifikatu zuten O´Leary et al.,-ek (1996) behien
burmuinetik. Gerora, gure ikertaldeak ere deskribatu du arratoien burmuineko gune
ezberdinetan eta baita ehunetan ere (Irazusta, et al., 2002; Agirregoitia et al., 2003a,
2003b). Zelularen hazkuntza, zahartzapena eta minbizien bilakaerarekin erlazionatuta
dauden substratu natural ugari ditu PEP-ak, hala nola, bradikinina, angiotentsinak,
basopresina, TRH, eta timosin-β4 (Salers et al., 1992; Schally et al., 2006; Stewart et al.,
2006). Arratoien ehunetan egindako lanetan deskribatu da PEP entzimaren aktibitatean
aldaketak gertatzen direla zahartzapenean zehar, emendapen bat dagoelarik kasu
gehienetan jaioberrietan helduekin alderatuz (Agirregoitia et al., 2003a, 2003b, 2007).
Badirudi, PEP aktibitatearen igoera bat izaten duela minbizi mota ezberdinetan. Datu
hauekin, pentsa daiteke, minbizien kartzinogenesian komuna den sistemaren batean parte
garrantzitsua beteko duela entzima honek, eta kontuan hartzeko datu azpimarragarria da.
Gure emaitzetan, PEP entzimaren jardueran hesteko mukosa eta minbiziaren
artean ezberdintasunik lortu ez arren, adenoma ehuna aztertzean igoera adierazgarri bat
aztertu zen bertan, aktibitate bikoitza izatera iritsi arte, bai heste zelulen mintzeko gunean
zein gune solugarrian. Heste lodiko zeluletan, PEP-aren aktibitatea aztertu duen lanik ez
da aurkitu bibliografian, beraz, konparaketa bat egitea ezinezkoa izan da.
Beste minbizi mota batzuetan ere, PEP-aren aktibitatea ikertu da. Urrutira joan
gabe, gure ikertaldeak berriki argitaratutako lan batean ikertu du PEP-aren ekintza, ez
heste lodian bakarrik, baita beste sei neoplasietan ere (Larrinaga et al., 2010a). Lan
honetan, PEP entzimaren aktibitatea neoplasia ezberdinetan aztertzean antzeko emaitzak
lortu dira kasu denetan, hots, aktibitateak igoera nabaria izan du giltzurruneko zelula
argietako kartzinoman, giltzurrun-pelbiseko kartzinoma urotelialean eta lepoko eta buruko
zelula ezkatatsuetako kartzinoman. Beste ikertalde batek tiroideko kartzinoman ere
emendapena esperimentatu zuen (Sedo et al., 1991). Liu et al.-ek (2008) PEP entzimaren
aktibitatearen igoera bat aztertu zuten ere minbizi hauetan. Goossens et al.-en arabera
(1996b), PEP entzimaren igoerak aztertzen dira, birika, prostata eta hesteko sigma
minbizietan, mukosa normalarekin alderatuz.
Gure emaitzen arabera, ACE eta NEP entzimen jarduera aldaketak estatistikoki
adierazgarriak ez izan arren, beste ikertalde batzuek emaitza interesgarriak lortu dituzte
heste lodiko minbiziaren bilakaeran zehar peptidasa hauen adierazpenak aztertu

Eztabaida 83
dituztenean. Bi entzimen kasuan, adierazpena nabariagoa zen minbiziaren progresioa
aurrera zihoan heinean (Ogawa et al., 2002; Sumitomo et al., 2005; Röcken et al., 2007;
Koga et al., 2008; Del Rio et al., 2010; Neo et al., 2010). Emaitza hauek ez datoz bat gure
ikerketa honetan esperimentatutakoarekin. Hala ere, desberdintasun hauek ez dute zertan
kontraesan bat izan behar. Gure ikertaldeak, giltzurrunean egindako lanetan deskribatzen
duen gisara, gerta daiteke, entzimaren adierazpen aldaketa jarduerarekin bat ez etortzea
(Wang et al., 2009; Larrinaga et al., 2010a). Honen azalpena, zelularen metabolismoaren
erregulazio maila desberdinean aurkitu daiteke, izan ere, DNAren itzulpen mailan gerta
daiteke erregulazioa edo baita proteinaren adierazpenean.
Azpiatal honen azken ideia bezala, aipatu, ikerlan honetan aztertu diren entzima
batzuen aktibitateek ez dutela aldaketarik jasan heste lodiko minbiziaren bilakaeran zehar.
Aldaketak izan dituzte, ordea, beste minbizi mota batzuetan, hala nola, Martinez et al.-ek
(1999), bularreko minbizietan Asp-AP-ren ekintzan jaitsiera bat deskribatu zuten, eta APN,
PSA eta APB entzimen aktibitate igoera. Gure ikertaldeak aho eta lepoko zelula
ezkatatsuetako kartzinoman egindako lanean, APN-ren aktibitateak ere igoera jasan zuen.
Aldiz, Asp-AP entzimaren aktibitate emendapena deskribatu zen (Perez et al., 2009),
aurreko lanarekin bat ez datorrena, baina bai obarioko kartzinometan Carrera et al.-ek
(2009) deskribatutakoarekin ados dagoena. Azken lan honen arabera, Asp-AP
aktibitatearen igoera, eta APN zein APB-ren aktibitate normalak deskribatu dituzte. Kasu
honetan, APN-ren eta APB-ren aktibitate konstanteak heste lodiko minbizian
esperimentatutako gure emaitzekin bat datoz.
Peptidasen emaitzetako aldagarritasun handi honek adierazten duena da, autore
batzuen iritziz, peptidasen aktibitatean gertatzen diren aldaketa hauek tumorearen
araberakoak direla eta neoplasia bakoitzean egon litekeen mikroingurumeneko aldaketen
erantzunen ondorio direla (Nanus, 2003; Ino et al., 2004; Carl-McGrath et al., 2006).
Honen guzitaren salbuespen gisa, PEP-aren kasuan soilik adostu dira datuak. Are
gehiago, guk dakigula, PEP entzimaren aktibitateak igoera nabarmena jasaten du
neoplasia mota ezberdinetan. Izan ere, ez da peptidasen aktibitatean gertatzen soilik,
geneen adierazpenean ere aldagarritasun hau aurkitu izan da, minbiziaren bilakaeran
existitzen den konplexutasun hori ziurtatuz (Lee et al., 2006).

84 Eztabaida
2. Plasmako peptidasen ekintza koloneko minbizia zu ten gaixoetan eta
kontroletan
Peptidasak zelularen mintzetan edota zitosolean daudela ondo ezarrita dago. Era
berean, odolean eta jariakin ezberdinetan peptidasa aktibitateak neur daitezke. Autore
askoren iritziz, entzima hauek mintz zitoplasmatikotik edo zelula barnetik jariatu zein
askatzen dira odolera. Askapen honen zergatia argi ez dagoen arren, peptidoen
andeaketan parte hartzen dutela deskribatu da (Balogh et al., 1998; Shrimpton et al.,
2002). Entzima berdina izanagatik, aurkitzen deneko gunearen arabera, hots, ehuna edo
gorputz likidoa, eragina duteneko prozesuak eta sortzen dituzten ondorioak ez dute zertan
berdinak izan behar. Entzimaren funtzioa ezberdina izan daiteke eta ondorioz, neurtutako
aktibitateak ere ezberdinak izan daitezke (Cordero et al., 2000). Peptidasa batzuen
plasmako aktibitateak prozesu fisiologiko eta patologiko ezberdinetan aldatu egiten direla
deskribatu da, euren artean minbizian (Carrera et al., 2006).
Odola lagin eskuragarria da eta bere osagaiak teknika errazen bidez neur daitezke
klinikan. Bertan, plasmako peptidasen aktibitate neurketak garrantzi handikoak lirateke eta
badira dagoeneko. Lanabes baliagarria da minbiziaren diagnosirako batez ere, baina
pronostiko egokia egin ahal izateko ere erabil liteke. Paziente gaixoen jarraipena egiteko
tresna lagungarria litzateke peptidasa hauen aktibitate aldaketek minbizi-markatzaile gisa
jokatuko balute.
Aztergai izan diren peptidasetako batzuk dagoeneko gaixotasun-markatzaileak dira,
plasmako ACE konbertasa esaterako. Adibidez, geroago sakonago azalduko den bezala,
klinikan erabili ohi da sarkoidosia gaixotasunaren adierazle modura (Bénéteau-Burnat et
al., 1991; Baudin, 2005), eta plasmako CAP entzimaren neurketak ere haurdunaldi
egokiaren adierazletzat erabiltzen dira (Ino et al., 2004). Heste lodiko minbiziari
dagokionez, jatorri ezberdineko molekulak proposatu dira pronostikorako, diagnostikorako
markatzaile edo terapia-itu gisa. Honen harira, entzima proteolitikoak gaixoen gorputzeko
likidoetan, horien artean odolean, ikertu izan dira molekula erabilgarriak izan
daitezkeelako. Gernuko MMP-en emendapena gastro-hesteetako gazte-polipoen detekzio-
markatzaile egokitzat iradoki dute berriki (Manfredi et al., 2009). Plasmako zein ehuneko
KLK6-aren gain-erregulazioa minbizian ere, terapia-itu gisa planteatu dute (Kim et al.,
2010). Eta, plasmako DPPIV entzima heste lodiko adenoma berantiarraren markatzaile
gisa proposatu dute, zuzenki erlazionatua baitago (De Chiara et al., 2009).
Plasmako peptidasen asaldurak neoplasikoak ez diren gaixotasunetan ere
deskribatu dira. Alde batetik, kolonarekin zerikusirik ez duten gaixotasunetan azaldu dira

Eztabaida 85
aldaketak, hots, depresioa, anorexia edo bulimia, gaixo alkoholiko-abstinenteetan edo
trauma baten ondorengoko estresean (Maes et al., 1999a, 1999b, 2001). Eta bestetik,
heste lodiarekin erlazioa duten gaixotasunetan ere deskribatu dira, adibidez, kolitis
ulzerosoa bezalako hanturazko gaixotasunetan (Sturiale et al., 1999; Byrnes et al., 2009).
Gaixotasun neoplasikoak aztertuz, eta heste lodiko minbizia zehazkiago aztertzen
bada, minbizia duten gaixoen odolean MMP familiako kideen eragina aztertu da. Hauetako
batzuen parte hartzea garrantzitsua dela irizten dute autore askok. Plasmako gelatinasek
(MMP-2 eta MMP-9), MMP-1 eta MMP-7-ak igoera jasaten dute polipo adenomatosoetan
zein minbizian (Mook et al., 2004; Hurst et al., 2007; Roy et al., 2009; Tahara et al., 2010).
Minbiziaren erauzketaren ondoren, MMP-2 eta MMP-9-ren kopurua jaitsi egiten dela
aztertu dute, non, terapeutikarako markatzaile bezala egoki egiten dituen.
Lisosomako aspartil eta zisteina entzimak (D eta B katepsinak), matrize
extrazelularreko molekulen hidrolisian parte hartzen duten proteasa solugarri
garrantzitsuenetakoak dira (Skrzydlewska et al., 2005; Kuester et al., 2008). Heste lodi eta
ondesteko minbizia duten pazienteen sueroan, talde kontrolarekin alderatzean, B
katepsinak esaterako aktibitate baxuagoa duela deskribatu da. D katepsinaren aktibitateak
aldiz, igoera bat jasaten du (Skrzydlewska et al., 2005).
Argi dago plasma lagineko molekulen ikerketak garrantzi handia izan dezakeela
klinikako erabileran gaixotasun neoplasikoen osaerarako. Proteasen aktibitatean
aldaketak gertatzen direla ikusita, peptidasen aktibitatean ere berdina gertatu litekeela
pentsatzekoa da. Honela, ikerlan honetarako, peptidasa batzuen aktibitatea aztertu zen
heste lodiko minbizia zuten gaixoen plasman.
Memoria honetako emaitzetan erakusten denez, heste lodiko minbizia zuten
gaixoen plasmako CAP, PGI eta PEP entzimen aktibitateak goranzko joera zuten.
Bitartean, DPPIV eta ACE-ren aktibitateak jaitsi egin ziren, betiere talde kontrolarekin
konparatzerakoan. Adierazgarriak izan ziren aldaketa hauek eta heste-zelulen kasuan
lortutako emaitzak, ildo berean kokatu daitezke. Kasu honetan ere, entzimaren arabera
aldaketak ezberdinak eta selektiboak izateak, gaitz honen konplexutasuna handia dela
ziurtatzen du. Ikerlanaren helmuga, terapia-itu izango diren molekulak aurkitzea bada,
konplexutasun honek komunean dituzten elementuak aurkitzea zailtzen du.
Adierazgarri izan diren plasmako entzimei buruzko lanak bibliografian ugariak dira,
beraien funtzio anitza dela eta, bai prozesu fisiologikoetan zein prozesu patologikoetan.
Prozesu patologikoei buruz hitz egiten denean, gaixotasun neoplasikoekin erlazionatzen
dituztenak badaude, baina baita gaixotasun ez-neoplasikoekin erlazionatzen dituztenak

86 Eztabaida
ere, aipatu bezala. Jarraian, koloneko minbizia zuten gaixoen plasman aldaketak jasan
zituzten peptidasak banan-banan aztertuko dira.
Dipeptidil peptidasa IV (DPPIV) entzimaren plasmako aktibitateari buruz emaitza
kontrajarriak deskribatu dira. Biopeptidoak erregulatzeko ahalmena duela aipatu da, eta
hori horrela izanik, tumore supresore edo aktibatzaile bezala jokatu dezake, tumore-
espezifikoa den mikroingurumeneko substratuen arabera. Aho barrunbeko minbizietan
aktibitate jaitsiera deskribatu da. Aldiz, hepatoma zuten arratoien sueroan, DPPIV-aren
aktibitatea askoz handiagoa da (Havre et al., 2008). DPPIV-ak, heste lodiko minbizidun
gaixoen plasman duen aktibitatea aztertzean ere kontraesanak aurkitzen dira. De la Haba-
Rodriguez et al.-ek (2002) koloneko adenokartzinoma zuten gaixoen sueroan, entsegu
entzimatikoaren teknika erabiliz, DPPIV-ren aktibitate altuagoak aztertu zituzten pertsona
osasuntsuekin alderatzean. Beste zientzialari batzuk ordea, ELISA teknika erabiliz,
kontrakoa aztertu zuten heste lodiko minbizietan egin zuten sueroko DPPIV entzimari
buruzko ikerketa lan zabalean (Cordero et al., 2000, 2009; De Chiara et al., 2009, 2010).
Izan ere, emaitza kontrajarrien arrazoi posiblea, Havre et al.-ek (2008) argitaratutako
errebisioan, bi lan hauetan erabilitako metodologia ezberdina izan litekeela argudiatu dute.
Gure lanean, gaixoen plasmako DPPIV entzimaren aktibitateak jaitsiera bat izan
zuen. Emaitza hauek bat datoz Cordero et al.-ek deskribatutakoarekin (2000). Haien
emaitzek, %90eko espezifizitatea eta sentsibilitatea ematen zuten, pronostiko metodo
eraginkorra dela adieraziz, batez ere fase goiztiarreko kartzinomadun gaixoekin. Duela
urte gutxi, talde honek berak errebisio bat argitaratu zuen serina peptidasa honen
aldaketen inguruan. Likido biologikoetan entzima honen kontzentrazioa oso altua da, nahiz
eta ezezaguna den oraindik, zein mekanismoren bidez askatzen den entzima hau
ingurune likidora eta erregulaturik dagoen ala ez. Euren ustez, garrantzitsua litzateke
DPPIV/CD26-aren organo edo ehun sortzaile nagusia zein den jakitea, eta zerk eragin
ditzakeen minbizia bezalako gaitzetan gertatzen diren kopuru aldaketa eta
presentzia/ausentzia horiek (Cordero et al., 2009). Ikertalde berberak, ikerketa zabala egin
zuen urte berean bi mila pertsonatik gorako talde batekin. DPPIV solugarriaren
kontzentrazioa gaixoek zuten adina eta sexua bezalako parametroen arabera aldatzen
zen aztertu zuten. Edo minbiziarekin erlazioa zuten beste hainbat parametrorekin
konparatu zuten, hots, hemorroideak, uzki odol-jarioa zein hanturazko gaixotasun onbera.
DPPIV kopuruaren aldaketak hainbat gaixotasun onbera zuten gaixoetan gertatzen zirela
aztertu zuten (De Chiara et al., 2009). Berriki argitaratu duten lan batean, sueroko DPPIV-
a proposatzen dute klinikarako teknika lagungarri modura, euren iritziz erlazio zuzena

Eztabaida 87
duelako koloneko adenoma berantiarrarekin edo minbizi goiztiarrarekin (De Chiara et al.,
2010).
ACE entzima, angiotentsinak eraldatzen dituen entzima, RAS-aren entzima nagusia
izan da klasikoki, Ang I eraldatzen baitu Ang II lortu ahal izateko. Alegia, Ang II da sistema
honetako molekula protagonista, bere hartzaileei lotuz zelularen hazkuntza,
ezberdintzapena edo bestalde, apoptosia eragin dezakeelarik (Hunyady et al., 2006; Ruiz-
Ortega et al., 2007). ACE entzimaren aktibitatea hainbat gaixotasunekin erlazionatu izan
da berriki aipatu den bezala. Gaixotasun ez-neoplasikoetan ere deskribatu da entzima
honen eragina, esaterako, ACE entzimaren aktibitateak sarkoidosia bezalako hanturazko
gaixotasuna pairatzen dutenen plasman neurtu da, eta baita gaixotasun baskularretan
zein minbizien aurka erradioterapiak edo kimioterapiak eragiten duten endotelioetako
disfuntzioetan ere. Lehenengoan, ACE-ren aktibitateak igoera bat izaten du, eta beste
kasuetan, ordea, aktibitatea jaitsi egiten da (Bénéteau-Burnat et al., 1991). Lehen aipatu
bezala honegatik guztiagatik, odoleko ACE entzima klinikan ohikoa den markatzailea da.
Neurketa errazekoa da eta odolaz gain, beste gorputz-likidoetan ere azter daiteke.
Adibidez, bronko-albeoloetako garbitze-likidoan (BAL) eta likido zefalorrakideoan (CSF)
ere aztertzen da entzima hau (Baudin, 2005). Beraz, argi dago hainbat gaixotasunen
diagnosian eta pronostikoan lagungarri bezala erabil daitekeela.
Bestalde, konbertasa hau kodetzen duen ACE genean gertatzen diren
inserzio/delezio polimorfismoak plasmako ACE kopuruan eragina dutela deskribatu da
(Rigat et al., 1990). Delezioak, ACE entzimaren emendapena eragiten du, giltzurrun eta
bihotzeko gaixotasunetarako arrisku izanik (Usmani et al., 2000; Antczak et al., 2001).
Polimorfismo mota hauek, tumorearen tamainarekin edo gaixoaren pronostikoarekin
erlazionatu dira, bai bularreko minbizian eta baita heste lodiko adenokartzinoman ere.
Horrela, delezioarekiko homozigosia zuten emakumeek pronostiko hobea zutela
deskribatu dute, hormonen bidezko ACE-ren erregulazioaren seinale. Euren iritziz, ACE
entzimak, heste lodiko adenomatik adenokartzinomarako progresioan eragina dauka,
guneko minbiziaren hazkuntzan eta angiogenesian parte hartzen duelako (Röcken et al.,
2007). Honela, ACE-ren inhibitzaileek minbiziaren aurreko babesa eskaintzen dutela
deskribatu da. Minbiziaren hilkortasuna eta arriskua jaisten dute eta inhibitzaile hauen
bidezko tratamendu luzeek polipo adenomatosoen intzidentzia jaisten dute (Röcken et al.,
2007; Kedika et al., 2011). Gure Memoria honetako emaitzetan, ACE aktibitatearen
jaitsiera aztertu genuen heste lodiko minbizia zuten gaixoen plasman. Heste lodiko
minbizian plasmako ACE entzimaren eragina aztertzen duten lan gutxi aurkitu dira eta

88 Eztabaida
zaila da konparaketa bat egitea. Baina ACE entzima RAS-ari oso loturik agertzen da beste
minbizietan eta heste lodiko minbizian ere sistema honetan eragin dezakeela pentsatzea
posible da.
Zistil aminopeptidasa (CAP), oxitozinasa, basopresinasa edo intsulina
erregulatzailea den aminopeptidasa (IRAP) bezala ere ezagutzen da. Honek, oxitozina,
basopresina eta Ang III bezalako hormonak dituelako substratu natural gisa (Ino et al.,
2004). Klasikoki, mintzeko peptidasa bezala izendatu den arren, gune solugarrian eta
gorputz likidoetan ere aurkitu da entzima honen aktibitatea (Rogi et al., 1996; Martinez et
al., 1999; Matsumoto et al., 2000, 2001; Silveira et al., 2001; Larrinaga et al., 2007).
Oxitozinasa edo P-LAP (plazentako leuzil aminopeptidasa) izenetik, esan behar da,
emakumeen ugal-aparatuan izan dela gehien aztertu den arloa. Haurdunaldian, sueroko
CAP peptidasaren kopuruaren igoerak esperimentatu dituzte, fetuaren ongizatea
ziurtatzen duena. Gorputz likidoen homeostasian ere eragina du, hortik basopresinasa
izena (Rogi et al., 1996; Matsumoto et al., 2001; Ino et al., 2004). Gaixotasun
ginekologikoetan CAP entzimaren aktibitatea nahiko ikertu da eta baita minbizietan ere,
neoplasiaren progresioarekin erlazionatu da endometrioko minbizietan, oxitozina
hidrolizatzen duelako (Ino et al., 2004).
Intsulina erregulatzailea den aminopeptidasa (IRAP) bezala jokatzen du CAP-ek.
RAS-ean, IRAP/CAP-aren substratua Ang IV da, ikerlan honetan aztertu diren APA eta
APN zein APB entzimen bidez Ang II-tik sortzen dena (Ruiz-Ortega et al., 2007; Stragier et
al., 2008). Honen harira, plasmako IRAP/CAP eta APB entzimen aktibitatea neurtu izan da
N-metil nitrosaureak arratoietan eragindako bularreko minbizietan. Bi entzimen aktibitateak
igoera bat jasan zuten, oxitozina, basopresina edo RAS-ak minbizian eraginen bat izango
dutenaren bidea irekita utziz (Carrera et al., 2006). Heste lodiko minbizidun gaixoen
plasmako CAP-aren aktibitateak ere gora egin zuen gure ikerketen arabera eta aipatutako
sistema hauek neoplasia honetan zerikusiren bat izan dezaketela pentsa daiteke.
PEP eta PGI peptidasek TRH (hormona tirotropina askatzailea) neuropeptidoa dute
substratu modura, besteak beste. Hormona honek, sistema neuroendokrinoan eta
gizakiaren portaeran eragina du eta arlo honetako hainbat gaixotasunekin erlazionatu da.
Sueroko PGI entzima, Tiroliberinasa modura izendatu dute ere. Honek eta gorputz pisuak
erlazio zuzena dutela deskribatu da, oraindik argi ez dagoen arren bietatik zein den
erregulatzailea eta zein haren menpekoa (Friedman et al., 1995). Hala ere, ez da ikerketa
sakonagorik egin Tiroliberinasari buruz, eta zaila da adierazten guk lortutako plasmako
PGI aktibitate aldaketak zein prozesuren ondorio diren. Bestalde, sueroko PEP entzimaren

Eztabaida 89
aktibitate baxuagoak esperimentatu dira, anorexia eta bulimia nerbiosoan, depresioetan
eta alkoholiko-abstinenteetan. Baina aktibitate altuagoak deskribatu dira trauma baten
osteko estresean (Maes et al., 1999a, 1999b, 2001). Bi entzima hauen ehuneko
aktibitateak arratoien garapenean eta zahartzapenean ikertu dira bai garunean zein
organo ezberdinetan. Aktibitate ezberdinak esperimentatu dira organoaren arabera eta
baita adinaren arabera ere (Agirregoitia et al., 2007).
Prolil endopeptidasa (PEP), solugarria da nagusiki. Odolean eta gorputz likidoetan
PEP entzimaren aktibitate baxuak deskribatu dira aspalditik (Goossens et al., 1996b).
Angiotentsinak dira PEP-aren beste substratu garrantzitsu batzuk, Ang I eta Ang II
zehatzago esateko. Hipertentsioa zuten gaixoetan, ACE eta PEP entzimaren artean
korrelazio positiboa aurkitu zuten, RAS-ean eraginen bat izango duela iradokiz eta
ondorioz gaixotasun hipertentsiboetan (Goossens et al., 1996b). Prostatako minbizia edo
prostatako hipertrofia duten gaixoen plasmako PEP-aren aktibitate jaitsiera deskribatu da
(Goossens et al., 1996b). Gure lanean, datu hauekin bat ez datorren aktibitatearen igoera
bat aztertu genuen. Heste lodiko minbizia duten gaixoen plasman ez da azken entzima
hauei buruzko ikerketarik egin guk dakigula eta bide honetatik jarraitzea interesgarria
litzateke ondorengo ikerketetan.
Hala ere, aipatu behar da, bibliografian plasmako peptidasei buruzko informazio
asko egon ez arren, hauetan gertatzen diren aktibitate aldaketak garrantzia handikoak
direla eta kontuan hartu beharrekoak. Lehen esan bezala, gaixotasunaren pronostiko eta
baita jarraipenerako ere markatzaile egokiak lirateke. Markatzaile hauek, klinikarako
praktikan, oso erabilgarriak dira, emaitza ugari lortzeko aukera eskaintzen dutenak era
erraz eta azkar batean. Gainera, gaixoarentzat, oso erosoa eta ez-erasokorra den
metodoaren bidez neurtuko lirateke markatzaile hauek plasman.
3. Peptidasen aktibitatea minbiziaren gradu eta est adio ezberdinetan
Heste lodiko minbiziak bilakaera ezjakina du eta badirudi bere agresibitate
biologikoaren araberakoa dela. Minbizien agresibitatea, hedapena eta barreiadura
neurtzeko metodoak beharrezkoak dira, gaixoen pronostiko egokiak eta ezarritako
tratamenduen emaitzen konparaketa egin ahal izateko. Metodo hauen helburu nagusia,
minbiziaren agresibitatea baxua den artean aurkitzea da eta honela, pazienteen bizi-
itxaropena hobetzea. Horretarako erabiltzen diren metodoak “histologi-gradua” eta
“estadioa” neurtzea dira. Klinikan, kirurgia bidez lagina erauzi ostean, histologi-analisia

90 Eztabaida
egitea da ohiko metodologia. Ondoren, neoplasiaren gradu zein estadio ezberdinen
sailkapena egiten da. Oro har, gradu eta estadio altuko neoplasiek agresibitate handiagoa
eta pronostiko okerragoa adierazten dute (Kumar et al., 2008).
Pronostikorako, tumore-markatzaileak diren molekulen bilaketa ezinbestekoa da.
Proteina hauek behin-behineko diagnosia egiteko erabil daitezke. Bestalde,
gaixotasunaren birsorreraren aurkikuntzan erabil daitezke, baita tratamenduaren
eraginkortasuna zehazteko ere. Markatzaile hauen aktibitatea, tumorearen
agresibitatearen arabera aldatzen bada, minbizia fase goiztiarrean aurkitzeko aukera
posibleak emendatzen dira. Tumorearen agresibitatea definitzeko unean, hainbat
ezaugarri izan behar dira kontuan. Minbiziko zelulek, denbora pasa ahala,
tratamenduarekiko erresistentzia sortzen dute. Gainera, inguruko zelulak infiltratzeko
ahalmena izaten dute eta baita beste organoak metastatizatzeko gaitasuna ere (Kumar et
al., 2008). Erresistentzia eta inbasioa ikertu beharreko prozesuak dira, beraien eraginak
momentu egokian ekidin ahal izateko. Peptidasak molekula aproposak lirateke tumore-
markatzaile gisa, aintzat hartu beharreko prozesu hauetan funtzio anitzak baitituzte. Hau
kontuan izanik, minbiziaren agresibitatearen arabera peptidasen aktibitateek ea aldaketak
jasaten dituzten aztertu da.
Gure ikerketak, entzima jarduera eta heste lodiko minbiziaren klinika erlazionatzen
ditu. Adenokartzinomak dira heste lodiko eta ondesteko minbizirik ugarienak (%98) eta
ehun hauetan neurtzeaz gain, gaixo hauen plasman ere neurtu da peptidasen aktibitatea.
Aktibitatearen aldaketak tumorearen pronostikorako giltzarri diren gradua (Toll et al., 2010)
zein estadioa (Greene et al., 2002; Lee et al., 2010) bezalako parametro
histopatologikoekin alderatu dira (Selves, 2009).
Proteasek dituzten ezaugarri molekularrengatik, klinikan zein terapia arloan oso
erabilgarriak diren molekulak dira. Terapeutikari begira, proteasen entzima-jarduera
erregulatzea posible da inhibitzaile naturalak (TIMP10) edota sintetikoak (MMPI11) erabiliz.
MMP-ak adibidez, luzaroan ikertu izan dira terapeutikarako, baita beraien inhibitzaile
endogenoak ere (Mook et al., 2004; Roy et al., 2009). MMP-ak zimogeno (inaktibo)
modura askatzen dira, eta zelulaz kanpo aktibatzen dira. Honela, erregulatu ahal izateko
kontrolpean dauden molekulan bihurtzen dira (Tahara et al., 2010). Adibidez, MMPI
deritzen hauen erabileran zientzialariek ikerketa ugari egin dituzte. Proteasek
kartzinogenesian dituzten funtzio anitz eta kontrajarriengatik, egun ez dute esperotako
10 TIMP: Tissue inhibitors of metalloproteinases 11 MMPI: Matrix metalloproteinase inhibitors

Eztabaida 91
emaitzik lortu, baina MMPI eraginkorren sorreren ikerketak aurrera darrai (Zucker et al.,
2004; Gialeli et al., 2011).
Agresibitate maila ezberdinetako minbizi-ehunetan, proteasen entzima-jardueren
aldaketak gertatzen direla aspaldi deskribatu da. MMP familiakoek minbizi mota
ezberdinetan (bularra, birika, area, heste lodia, maskuria, obarioa, prostata eta burua) edo
minbizien progresioan duten funtzioa ikertu duten hainbat errebisio daude bibliografian.
Lan hauek mintzeko proteasen aktibitate aldaketak deskribatzen dituzte, eta
gaixotasunaren prognosi-biomarkatzaile posibleak diren molekulak aztertzen dituzte (Roy
et al., 2009; Gialeli et al., 2011).
MMP-ek funtzio ezberdina erakutsi dute estadio ezberdinetan. Adibidez, MMP-8
matrizeko proteasaren aktibitatearen igoerak bularreko minbizietan, metastasiaren aurreko
babesle funtzioa duela aztertu dute. Aldiz, mingaineko zelula ezkatatsuetako minbizietan
kontrako efektua du. Antzeko jokaera aztertu dute MMP-9-aren portaeran,
kartzinogenesiaren eragilea izan daiteke edo minbiziaren aurkako entzima gaixotasunaren
fase berantiarretako egoera zehatzetan (Gialeli et al., 2011). Heste lodiko kartzinoman
zehazki, MMP hauen ikerketa zabala egin da. MMP-2 zein MMP-9-aren gain-adierazpena
deskribatu dute kartzinometan. MMP-2-aren eta gradu zein Dukes-en estadioen arteko
erlazio zuzena ere azaldu dute (Papadopoulou et al., 2001; Heslin et al., 2001). Beste
ikerlari batzuek, MMP-1 eta MMP-13-ak ere asaldatuta daudela diote (Roy et al., 2009).
Edonola, ikertalde ezberdinak bat datoz MMP-9-aren gain-adierazpenaz, eta honen erlazio
zuzenaz heste lodiko minbiziaren hainbat ezaugarrirekin: displasia graduarekin,
minbiziaren estadioarekin zein inbasio gaitasunarekin. Gainera, heste lodiko minbizia
duten gaixoen biziraupen baxuarekin erlazionatu dute (Herszényi et al., 2008; Roy et al.,
2009; Groblewska et al., 2010). Matrizeko serina proteasa (MSP) den tripsinaren gain-
adierazpenaz antzeko emaitzak lortu dituzte koloneko minbizian. Inbasioa, metastasia,
estadioa zein minbiziaren birsorrerarekin erlazionatu dute. Tripsinaren gain-adierazpenak,
pronostiko baxuko kasuetan heste lodiko minbiziaren birsorreraren adierazle ona dela
iradokitzen dute (Yamamoto et al., 2003).
Katepsinak eta kalikreinak bezalako beste proteasa batzuk ere erlazionatu dira
minbizien agresibitatearekin. B, D eta L katepsinen aktibitateek, igoera nabaria jasaten
dute koloneko adenokartzinoman, baina minbiziaren agresibitatearekin duten erlazioa ez
dago batere argi. B eta L katepsinak Dukes-en estadio goiztiarretan emendatzen diren
bitartean, D katepsina estadio berantiarretan emendatzen dela diote (Kuester et al., 2008).
Beste batzuek emendapen progresiboa aztertu dute B katepsinan, D katepsinan aurkitu ez

92 Eztabaida
dutena (Talieri et al., 2004). Aldiz, beste batzuek ez dute inolako erlaziorik aurkitu
katepsinen aktibitatearen eta agresibitatearen artean (Adenis et al., 1995). Azkenik,
kalikreinen familiako kide batzuen (KLK5, KLK7 eta KLK14) aktibitateak adinaren,
estadioaren eta graduaren arabera aldatu egiten direla deskribatu da. Honela, pazienteen
bizi-itxaropena neurtzerako unean informazio gehigarri bezala erabil daitezke (Talieri et
al., 2009).
Gure ikerketa lanean, peptidoak hidrolizatzen dituzten peptidasak aztertu ziren.
Badaude lanak, non, minbiziaren agresibotasun maila ezberdinetan, peptidasen
adierazpen edo aktibitate aldaketak aztertu dituzten, eta ondorioz, gaixotasunaren
pronostikoaren hobekuntza edo okertzea eragiten dute (Carl-McGrath et al., 2006). NEP,
APN, DPPIV eta CAP entzimak proposatu dira neoplasia mota ugariren pronostikorako
lanabes bezala. Adibidez, NEP-aren adierazpena prostatako minbizi agresiboekin
erlazionatu da eta gaiztotze-potentzial altuagoarekin (Dall´Era et al., 2007). DPPIV-ren
adierazpena ere obarioko kartzinoma duten gaixoen biziraupen-tasa okerragoekin
erlazionatu dute (Zhang et al., 2008). Honen harira, antzeko emaitzak deskribatzen
dituzten hainbat ikerketa daude. Endometrioko kartzinomako gradu altuetan, DPPIV
entzimaren aktibitate baxuagoak esperimentatu dituzte (Pro et al., 2004; Carl-McGrath et
al., 2006; Cordero et al., 2009). Aipatzekoa da ere, Shibata et al.-ek (2004) endometrioko
kartzinoman egin zuten CAP entzimaren ikerketa zabal batean, adierazi zuten peptidasa
honen proteina-adierazpena histologi-graduekin, gaixoen biziraupenekin, miometrioaren
eta gongoil linfatikoen inbasioarekin zein odoleko infiltrazioarekin positiboki erlazionatua
zegoela. Gure lan taldeak ere giltzurruneko zelula argietako kartzinoman eginiko lanetan,
CAP, DPPIV, PSA, PGI, ACE eta APB entzimen aktibitate aldaketak atzeman zituen
minbiziaren graduaren eta estadioaren arabera. Minbiziaren agresibitate maila altuetan,
peptidasa hauek aktibitate altuagoa zutela deskribatu zuten, pronostikorako markatzaile
egokiak direla iradokiz (Larrinaga et al., 2007; Blanco et al., 2008; Varona et al., 2010;
Larrinaga et al., 2010).
Peptidasen ekintzaren aldaketak eta heste lodiko minbiziaren agresibitatea
erlazionatzen dituzten lan gutxi daude. Ehuneko peptidasei dagokionez, PEP entzimaren
aktibitate emendapena fenotipo okerragoarekin erlazionatu da minbizi mota ezberdinetan,
euren artean kolonean (Liu et al., 2008). Koloneko minbizian ere, APN aktibitatearen
igoera, pronostiko okerragoarekin erlazionatua dagoela deskribatu dute (Hashida et al.,
2002). Gure datuek antzeko joera agertzen dute bi entzima hauetan. Gure emaitzetan
ordea, peptidasen aldaketak minbiziaren graduaren eta estadioaren arabera

Eztabaida 93
alderatzerakoan ez zen ezberdintasun adierazgarririk egon (III. Eranskina). Beraz,
aktibitateen gorabeherei buruzko joeraz hitz egingo da. Emaitzek ez dute goranzko edo
beheranzko joera orokor bat agertzen, peptidasen aldaketa selektiboaz hitz egin daiteke
aurreko bi ataletan adierazi den bezala. Peptidasen gorabeherak ez ziren graduaren
menpekoak, ezta estadioaren menpekoak ere. Ezberdinak ziren kokapenaren arabera ere,
mintzekoak edo solugarriak izan. Graduak eta estadioak taldekatuta zein taldekatu gabe
adierazi, ehuneko peptidasen aktibitatearen aldaketak eta minbiziaren agresibitatearen
artean lortutako emaitzak ez ziren adierazgarriak izan.
Aldiz, heste lodiko minbizia zuten gaixoen plasmako peptidasen aktibitateak
aztertzerako unean, PSA, DPPIV, PEP, APA eta PGI entzimetan aldaketa adierazgarriak
aurkitu ziren, minbiziaren graduaren eta estadioaren arabera banatzerakoan (3.13. Irudia;
III. Eranskina).
Beste metaloproteasa batzuk ere ikertu dira. Adibidez, matrizeko metaloproteasa
batzuen kasuan (MMP-1 eta MMP-7), gradu eta estadioaren araberako banaketa bat
egitean, peptidasekin gertatzen den antzeko emaitzak lortu dituzte. Plasmako MMP-ek
pronostikorako markatzaile gisa joka dezakete, agresibitatearen arabera aldaketak jasaten
baitituzte, ehunean gertatzen ez zaiena (Tahara et al., 2010). Kontrako emaitzak ere
badaude, MMP-2 eta MMP-9-arekin gertatzen den bezala, heste-zeluletako proteasen
aktibitate aldaketak bat datoz minbiziaren agresibitatearekin, plasman azaltzen ez dena
(Papadopoulou et al., 2001; Mook et al., 2004; Roy et al., 2009).
Emaitzen ataleko 3.3. Taulan adierazi den moduan, gure datuek, heste lodiko
minbiziaren histologi-gradu altua (2.3. Taula; G3) zuten gaixoen plasmako PSA, DPPIV,
PEP eta APA entzimek aktibitate baxuagoa zuten gradu baxuko (G1-G2) minbizia
zutenekin alderatuz gero. Hau da, gradu histologikoko agresibitatea igotzen doan heinean
peptidasen aktibitatea jaitsi egiten da. Estadioaren arabera banaketa egitean, estadio
altuko (2.4. Taula; T3-T4) minbizia zuten gaixoen plasmako PGI entzimak aktibitate
gehiago agertu zuen estadio baxuko (T1-T2) minbizia zutenekin konparatuz gero.
Egun, sueroko antigeno kartzinoenbrionarioa (CEA) eta 19.9 karbohidrato
antigenoa (CA19.9) erabiltzen dira minbizi mota batzuen (heste lodiko minbizia barne)
prognosi-markatzaile gisa (Loizate, 1990; Lee et al., 2010). Hala ere, ez da minbizien
prognosi-markatzaile ziur eta hobetsiena existitzen. Horregatik, metodo klasikoen
lagungarri diren molekula egokiak aurkitzeko lanetan jarraitzen dute ikertaldeak. Izan ere,
proteasek zein peptidasek minbizi mota ezberdinen agresibitatearekin erlazio zuzenak
dituztela deskribatu da eta pronostikorako markatzaile posibleak direla iradokitzen da.

94 Eztabaida
Orain arte aipatutako proteasa ugari pronostiko markatzaile gisa proposatu dira. Adibidez,
plasmako zein ehuneko KLK6-aren minbiziko gain-erregulazioa, pronostiko baxuaren
adierazle dela deskribatu dute (Kim et al., 2010). Sueroko MMP-7 eta MMP-1-ari buruz,
fase berantiarreko kartzinometan biziraupenaren jaitsieraren adierazle dela adierazi dute
(Yamamoto et al., 2003; Roy et al., 2009; Tahara et al., 2010). Odoleko B eta D
katepsinek ere aldaketak jasaten dituzte koloneko adenokartzinoman. Hala nola,
aktibitateok tumorearen tamainarekin erlazioa dutela azaldu dute eta egoera txarraren eta
pronostiko okerragoaren adierazle dira (Skrzydlewska et al., 2005).
Plasmako peptidasen adierazpenaren aldaketak minbizi mota ezberdinetako
minbizi-agresibitatearekin erlazionatzen dituzten lanen kopurua urria da (Carl-McGrath et
al., 2006). Ikertzaile batzuek diotenez, biriketako minbizietako eta gaixo hauen plasmako
APN entzimaren adierazpen aldaketak erlazio zuzena du estadio aurreratu eta biziraupen-
tasa baxuko gaixotasunekin (Murakami et al., 2005; Tokuhara et al., 2006). Heste lodiko
minbiziari dagokionez, aipatzekoak dira Cordero et al.-ek lorturiko emaitzak (2009).
Hauetan, sueroko DPPIV entzimaren aktibitatea ez da aldatzen gaixoaren sexu, adin edo
tumorearen tamaina, gradu, eta kokalekuaren arabera. Aldiz, pronostikorako tresna bezala
erabil daitekeela diote adenoma berantiarra eta kartzinoma goiztiarra pairatzen duten
kasuetarako (De Chiara et al., 2009). Gure datuek emaitza hauekin duten antzekotasuna
kontutan izanik, DPPIV-aren joera azpimarragarria dela iruditzen zaigu. DPPIV-aren
aktibitatea adierazgarriki jaitsi zen, ia erdira, histologi-gradu altuko minbizia zuten gaixoen
plasman neurtzean.
Konparaketa gradu baxuko eta gradu altuko minbizia zuten gaixoen plasmako
entzimen aktibitateen artean egin zen. Aktibitateen aldaketa adierazgarriak izan zituzten
gainontzeko entzimetan, hots, PSA, APA eta PEP-ean. Aldaketa hauetan, hain
nabarmenak ez izan arren, antzeko jaitsierak aztertu ziren. PGI-aren kasuan,
aktibitatearen igoera bat aztertu zen minbiziaren estadio altuetan. Gaixotasunaren
agresibitate altuagoaren eta pronostiko baxuaren adierazle dira peptidasen aktibitate
aldaketa hauek.
Plasmako peptidasak izanik, teknika errazen bidez neurtzen dira, eta egungo
metodoen lagungarri gisa erabili ahal izateko egokiagoak dira. Lehen aipatu bezala,
klinikan plasma-lagina lortzea ohikoa eta eskuragarri dagoen lagina da, ez da teknika
erasokorra gaixoentzat eta gaixotasunaren jarraipena egiteko ezin hobea da. Gainera,
peptidasen aktibitate aldaketek minbizien agresibitatearekin erlazioa dutela deskribatu da,
eta ondorioz, pronostikorako markatzaile egokiak direla adierazi daiteke.

Eztabaida 95
4. Peptidasen asaldurak eta erregulazio peptidikoa heste lodiko
kartzinogenesian
Kartzinogenesian zehar, zelularen fisiologian aldaketa ugari gertatzen dira.
Aldaketa hauek maila extrazelularrean eta zelula barnean gertatutako prozesuen
elkarrekintzaren ondorioa dira. Horregatik dute garrantzia seinalearen bide osoan parte
hartzen duten molekulek, bai zelularen mintzekoek zein zelularen zitosolean daudenek.
Zelularen kanpo-mintzean kokatzen diren peptidasen (ektopeptidasak) funtzio
nagusia peptido bioaktiboen eraldaketa da, peptido hauek aktibatuz edo inhibituz (Turner,
1986, 1987). Entzima hauek, zelula neoplasikoetan eragina duten faktore aktibatzaile edo
inhibitzaile jardueradun substratu peptidikoak eraldatuz, tumorearen sorrera, hazkuntza
eta hedapena erregulatu dezakete (Nanus, 2003; Ino et al., 2004; Carl-McGrath et al.,
2006). Ondorioz, mintzeko peptidasa hauen adierazpenaren edota aktibitatearen jaitsierak
edo galerak mintzeko hainbat peptidoen pilaketa areagotu dezake, neoplasiaren
hedapena eraginez (Nanus, 2003; Sumitomo et al., 2005).
Egun, entzima hauek zelularen barne-prozesuetan duten eraginari buruzko
ezjakintasun asko dago eta eztabaidagai da oraindik. Urteetan pentsatu izan da soilik
proteina-katalisiaren azken fasean zutela zerikusia (Mantle, 1992; O´Cuinn, 1998). Ordea,
azken hamarkadan, ekintza endokrinoa, parakrinoa eta autokrinoa izateaz gain, mezulari
peptidikoek zelularen barnean ere ekintzarik izan lezaketela adierazi da, hazkuntza,
zelula-ezberdintzapena eta angiogenesia bezalako prozesu garrantzitsuetan parte hartuz
(Re, 2002; Zagon et al., 2002; Re et al., 2006; 2008). Bide honetan ere, autore batzuen
iritziz, ekintza intrakino hau, zelula barneko peptidasez erregulatua egon liteke, aipatutako
prozesuotan partaide izango liratekeelarik (Shrimpton et al., 2002; Re, 2003; Re eta Cook,
2006).
Lan honetan, zelularen banaketa egin zen, mintzari lotutako entzima jarduera zatiki
solugarriarengandik banatzeko. Banaketa honen helburua zatiki bietako entzimen
aktibitatea ezagutzea izan zen, gainazal zelularrean eta zitosolean gertatutakoaren
adierazlea baita. Honela, heste lodiko neoplasietan peptidasen aktibitatearen aldaketa
selektiboak agertu ziren zelula-gunearen arabera. Adibidez, DPPIV-aren eta PEP-aren
aktibitateak ehun neoplasikoen bi zatiki azpizelularretan asaldatuta aurkitzen direla azaldu
genuen. Ondorioz, esan daiteke, heste lodiko neoplasietan, ez soilik ektopeptidasek,
baizik eta gune solugarriko peptidasek ere, eraginen bat izan dezaketela. Ez kanpora
askatzean edo jariatzean duten ekintzarengatik bakarrik (Balogh et al., 1998; Shrimpton et

96 Eztabaida
al., 2002), baizik zelularen barnean ekintza intrakinoaren bidez dituzten jarduerengatik ere,
hala nola, hazkuntza prozesuetan (Re eta Cook, 2006).
Heste lodiko neoplasietan, zein sistema peptidiko dagoen asaldatuta jakiteko beste
ikerketa ugari egin beharko lirateke. Hala ere, gure datuek peptido sistema konkretu
baterantz zuzendu dute ikerketa lerroa, alegia, errenina angiotentsina sistema (RAS).
RAS, presio arterialaz eta oreka hidroelektrikoaz arduratzen den sistema peptidikoa da.
Klasikoki sistema endokrino bezala definitua izan da, angiotentsina II (Ang II), bera
eraldatzen duen entzima (ACE) eta angiotentsinaren hartzailea (AT1) izan direlarik bere
protagonistak. Baina pixkanaka, sistema konplexuagoa dela aurkitu da, hartzaile
gehiagok, peptido bioaktibo ugarik eta peptidasa ezberdinek parte hartzen dutela
deskribatu baita (Hirasawa et al., 2002; Hunyady et al., 2006). Gainera, azken urte
hauetan, ehun askok sistema honetako osagaiak sortzeko beharrezko makinaria
molekularra dutela deskribatu dute. Adibidez, hesteetan eta giltzurrunean guneko-RAS-ak
existitzen direla adierazi dute (Hirasawa et al., 2002; Paul et al., 2006). Guneko RAS
hauek, oinarrizko funtzioa mantentzeaz gain, zelulen hazkuntza, ezberdintzapena,
angiogenesia eta ugaritze prozesuetan ere parte hartzen dute, ekintza autokrinoak,
parakrinoak eta intrakrinoak erabiliz (Hirasawa et al., 2002; Nanus, 2003; Re, 2003;
Deshayes et al., 2005; Re et al., 2006; Paul et al., 2006; Zhou et al., 2009; Neo et al.,
2010).
Hau dela eta, RAS azken urte hauetan kartzinogenesian ikertzen ari den sistema
peptidikoa da (Hirasawa et al., 2002; Nanus, 2003; Re, 2003; Deshayes et al., 2005; Zhou
et al., 2009). Heste lodiko guneko RAS-ari buruzko aipamenak ere aurkitu dira
bibliografian, baita angiogenesiak duen erlazioa koloneko kartzinogenesian zein bere
gaiztotze-progresioan. Sistema honetako peptidasek zein funtzio betetzen duten
angiogenesian ere ikertu izan da (Hirasawa et al., 2002; Hashida et al., 2002; Marchió et
al., 2004; Röcken et al., 2007; Raica et al., 2009; Larrinaga et al., 2010b).
Gure ikerlaneko emaitzek, ACE2-aren eta PEP-aren aktibitatea polipo
adenomatosoetan, eta APA-rena adenokartzinoman, adierazgarriki emendatua dagoela
adierazten dute. Hiru peptidasa hauek angiotentsina II eraldatzen dute, Ang 1-7 (ACE2-k
eta PEP-k) eta Ang III ekoiztuz (APA-k). Aipatutako angiotentsina hauek neoplasia
ezberdinen sorreran eta hedapenean eragiten dutela adierazi da (Deshayes et al., 2005;
Speth et al., 2008; Teranishi et al., 2008; Zhou et al., 2009; Neo et al., 2010, Larrinaga et
al., 2010a, 2010b). Beraz, posible da heste lodiko guneko RAS asaldatuta egotea
adenoma-adenokartzinoma sekuentzian. Emaitza hauek, eta baita aurreko ikerketetan

Eztabaida 97
giltzurruneko neoplasietan lortutakoek (Varona et al., 2007; Larrinaga et al., 2007; Blanco
et al., 2008; Varona et al., 2010; Larrinaga et al., 2010), ikerketa lerro berri bat zabaldu
dute gure lantaldean, RAS-eko osagai guztiak aztertu nahian eta bere funtzioa neoplasia
hauetan zein den zehazteko helburuarekin.
Bibliografian badaude peptido bioaktiboen eta peptidasen ekintza neoplasien
sorreran eta hedapenean aipatzen duten beste adibide batzuk. Adibidez, DPPIV
entzimaren proteina-adierazpen galera deskribatu izan da neoplasia batzuetan, adibidez,
gibeleko, bularreko, biriketako eta endometrioko kartzinoman, melanoman eta
neuroblastoman (Stecca et al., 1997; Havre et al., 2008). Autore batzuen iritziz,
tumorearen kontrako efektua du DPPIV-ak ehun batzuetan eta honen adierazpenaren
jaitsierak minbiziaren hazkuntza, angiogenesia eta hedapena eragiten du. Hainbat
zitokinen gain duen erregulazio ekintza izan liteke horren arrazoia (Pro et al., 2004; Havre
et al., 2008; Arscott et al., 2009). Adibidez, duela gutxi ziurtatu da neuroblastoman, DPPIV
entzimaren adierazpen edota aktibitatearen jaitsierak edo galerak, estromako deribatu
faktorea 1 (SDF1) peptidoaren kopuru igoera eragiten duela, haurtzaroko gaixotasunik
ohikoenetako bat den honen hazkuntza eta agresibitatea areagotuz (Arscott et al., 2009).
Hainbat ikerlanek adierazi dute ere, giltzurruneko minbizian interleukina batzuek
erregulatzen dutela DPPIV-aren gene- zein proteina-adierazpena (Riemann et al., 1999;
Varona et al., 2010).
Memoria honetako emaitzak ezberdinak dira, berriki aipatu bezala, mintzeko
guneko entzimaren ekintza edo gune solugarriko entzimaren ekintza azaltzerakoan.
Polipoan, mintzeko guneko DPPIV entzimaren aktibitatearen jaitsiera bat antzeman
daiteke eta kasu honetan baliteke lehenengo aipatutako prozesuan parte hartzea, hots,
tumorearen hazkundean fase goiztiarrean, entzima honek inaktibatzen dituen zitokina
ezberdinen maila aldaketak direla medio. Minbizian, ordea, gune solugarriko DPPIV-aren
aktibitateak igoera bat jasaten du. Aktibitatearen ezberdintzapen selektibo hau gibeleko
minbizia duten gaixoetan deskribatu da (Kojima et al., 1987; Cordero et al., 2009).
Interpretatzeko emaitza erraza ez den arren, neoplasia hauetan mintzeko eta gune
solugarriko DPPIV-aren funtzioak ezberdinak izan behar dutela pentsatzera garamatza.
Peptidasen ekintza biologiko guztien artean, peptidoen erregulazioa da gehien
ezagutzen dena, baina badira beste batzuk ere, hots, zelularen kanpo-matrizearen
andeakuntza, atxikimendu-molekula gisako ekintza eta seinalizazio intrazelularra
aktibatzen duen hartzaile funtzioa (Antczak et al., 2001; Carl-McGrath et al., 2006).
Adibide esangarrienetarikoa DPPIV-rena da. Entzima honek tumorearen hazkundean

98 Eztabaida
eragiten du, metaloproteasekin modu koordinatuan zelularen kanpo matrizea andeatzen
baitu. Gainera, zelula neoplasikoak DPPIV-a adieraziz edota beste ehunetako zelulen
DPPIV-a ezagutuz, alegia peptidasaren atxikimendu-molekula funtzioa erabiliz, lokalki zein
distantzian hedatzen dira. Honez gain, DPPIV-ak hartzaile funtzioa bete dezakeela ere
proposatu da (Pro eta Dang, 2004; Carl-McGrath et al., 2006; Havre et al., 2008).
Ezaugarri hauek guztiak kontuan hartuz, posible da peptidasen eragina neoplasia
mota ezberdinetan euren jarduera biologiko guztien emaitza netoa izatea. Ondorioz, heste
lodiko neoplasietan jasotako emaitzen analisietatik sorturiko hipotesien artean, posibilitate
guztiak kontuan hartu beharko lirateke etorkizunean egin beharreko ikerketa berriei begira.

Eztabaida 99
Egun, neoplasien bilakaerarekin erlazionatutako ekintzei buruzko ezagutzak
badaude. Hala ere, ezagutza hauek ez dira nahikoak gaixotasuna guztiz ulertzeko
beharrezko diren elementu guztiak ez baititugu. Ondorioz, zail egiten da gaixotasun honi
aurka egitea edo amaiera ematea. Neoplasietan parte hartzen duten proteinen funtzio
zehatzari buruzko jakintza urria da oraindik, eta nahiz eta ikergai izan diren
peptidasetariko batzuk minbizi-markatzaileak izan, pronostikorako markatzaile egokiaren
bilaketak aurrera darrai, baita terapia-itu izan litekeenaren bila ere. Neoplasien sorrera eta
bilakaera konplexuaren ulermenerako, ez da nahikoa elementu bakarraren ezagutza
izatea, elementu ugari eta desberdinen arteko elkarrekintzak parte hartzen baitute
gaixotasun honetan. Izan ere, elkarrekintza hauen ondorioak desberdinak dira neoplasia
motaren arabera, eta neoplasia berberaren barnean, era berean, azpimotaren arabera.
Aurkezten den Memoria honetan, peptidasek ez dutela soilik entzima andeatzaile funtzioa
betetzen deskribatu da, baizik eta, hartzaile eta atxikimendu molekula gisa ere ekintzak
izan ditzaketela. Peptidasa hauek, peptidoekin eta hartzaileekin batera sistema
peptidikoak osatzen dituzte. Honez gain, zelularen kanpoko kinadek barneko seinalizazio
sistemetan edo barneko erreakzio bideetan eragina dutenez, zelularen kanpo mintzeko
molekulek zein barneko molekula solugarriek funtzio garrantzitsua izango dute neoplasien
biopatologian.
Bide metaboliko hauen ezagutzak eta hauek minbiziaren bilakaeran dituzten
asaldurak, gaixotasun honen diagnosirako eta terapia arlorako tresna eta estrategia
berrien sorreran lagungarri izango dira.


Ondorioak


Ondorioak 103
5. ONDORIOAK
1. Heste lodiko minbiziaren bilakaeran zehar, hau da, ehun osasuntsua, adenoma,
adenokartzinoma sekuentzian, peptidasen aktibitate aldaketak gertatzen dira.
Aldaketa hauek selektiboak dira, neoplasiaren arabera, zelularen banaketaren
arabera, eta entzima motaren arabera. DPPIV-k, PEP-k, ACE2-k eta APA-k eragina
izan lezakete bai neoplasia hauen fase goiztiarretan (adenoma) eta bai
berantiarretan (adenokartzinoma).
2. Heste lodiko minbizidunen plasmako peptidasen aktibitateetan aldaketak gertatzen
dira, plasma-kontrolen plasmako aktibitatearekin alderatzean. Aldaketa hauek ere
selektiboak dira, ez baitie entzima guztiei modu berdinean eragiten. Hau honela
izanik, DPPIV, ACE, PEP, CAP eta PGI peptidasen neurketa plasmatikoa tresna
erabilgarria bilakatu liteke heste lodiko minbizien diagnostikorako.
3. Minbiziaren histologi-graduaren eta estadioaren arabera plasmako peptidasen
aktibitate aldaketa selektiboak gertatzen dira. Plasman DPPIV, APA, PEP, PSA eta
PGI neurtzeak heste lodiko minbizien pronostikoa egiten lagundu lezake.


Bibliografia


Bibliografia 107
6. BIBLIOGRAFIA
A
Abbott, C.A., Baker, E., Sutherland, G.R., McCaughan, G.W., Genomic organization, exact localization, and tissue
expression of the human CD26 (dipeptidyl peptidase IV) gene. Immunogenetics, 1994. 40: p. 331-338.
Adenis, A., Huet, G., Zerimech, F., Hecquet, B., Balduyck, M., Peyrat, J. P., Cathepsin B, L, and D activities in colorectal
carcinomas: relationship with clinico-pathological parameters. Cancer Letters, 1995. 96: p. 267-275.
Agirregoitia, N., Casis, L., Gil, J., Ruiz, F., Irazusta, J., Ontogeny of prolyl endopeptidase and pyroglutamyl peptidase I in
rat tissues. Regulatory Peptides, 2007. 139: p. 52-58.
Agirregoitia, N., Gil, J., Ruiz, F., Irazusta, J., Casis, L., Effect of aging on rat tissue peptidase activities. J Gerontol A Biol Sci Med Sci, 2003a. 58: p. 792-797.
Agirregoitia, N., Irazusta, A., Ruiz, F., Irazusta, J., Gil, J., Ontogeny of Soluble and Particulate Prolyl Endopeptidase
Activity in Several Areas of the Rat Brain and in the Pituitary Gland. Dev Neurosci 2003b. 25: p. 316-323.
Ahlquist, D.A., Shuber, A.P., Stool screening for colorectal cancer: evolution from occult blood to molecular markers. Clinica Chimica Acta, 2002. 315: p. 157-168.
Ahmed, S.I., Thompson, J., Coulson, J.M., Woll, P.J., Studies on the Expression of Endothelin, Its Receptor Subtypes, and
Converting Enzymes in Lung Cancer and in Human Bronchial Epithelium. Am. J. Respir. Cell Mol. Biol., 2000. 22: p. 422-431.
Alba, F., Arenas, J.C., Lopez, M.A., Comparison of soluble and membrane-bound pyroglutamyl peptidase I activities in
rat brain tissues in the presence of detergents. Neuropeptides, 1995. 29: p. 103-107.
Alba, F., Iribar, C., Ramirez, M., Arenas, C., A fluorimetric method for the determination of brain aminopeptidases. Arch Neurobiol (Madr), 1989. 52: p. 169-73.
Allinson, T.M.J., Parkin, E.T., Condon, T.P., Schwager, S.L.U., Sturrock, E.D., Turner, A.J., Hooper, N.M., The role of
ADAM10 and ADAM17 in the ectodomain shedding of angiotensin converting enzyme and the amyloid precursor
protein. European Journal of Biochemistry, 2004. 271: p. 2539-2547.
Antczak, C., De Meester, I., Bauvois, B., Ectopeptidases in pathophysiology. BioEssays, 2001. 23: p. 251-260.
Arnold, C.N., Goel, A., Blum, H.E., Boland, C.R., Molecular pathogenesis of colorectal cancer. Cancer, 2005. 104(10): p. 2035-2047.
Arscott, W.T., LaBauve, A. E., May, V., Wesley, U. V., Suppression of neuroblastoma growth by dipeptidyl peptidase IV:
relevance of chemokine regulation and caspase activation. Oncogene, 2009. 28: p. 479-491.
Aurich-Costa, J., Cadel, S., Gouzy, C., Foulon, T., Chérif, D., Cohen, P., Assignment of the aminopeptidase B gene
(RNPEP) to human chromosome 1 band q32 by in situ hybridization. Cytogenet Cell Genet, 1997. 79: p. 143-144.
Awadé, A.C., Cleuziat, P., Gonzalès, T., Robert-Baudouy, J., Pyrrolidone carboxyl peptidase (Pcp): an enzyme that
removes pyroglutamic acid (pGlu) from pGlu-peptides and pGlu-proteins. Proteins, 1994. 20: p. 34-51.
Azizi, M., Ezan, E., Reny, J.L., Wdzieczak-Bakala, J., Gerineau, V., Menard, J., Renal and Metabolic Clearance of N-
Acetyl-Seryl-Aspartyl-Lysyl-Proline (AcSDKP) During Angiotensin-Converting Enzyme Inhibition in Humans. Hypertension, 1999. 33: p. 879-886.
B
Balogh, A., Cadel, S., Foulon, T., Picart, R., Der Garabedian, A., Rousselet, A., Tougard, C., Cohen, P., Aminopeptidase B:
a processing enzyme secreted and associated with the plasma membrane of rat pheochromocytoma (PC12) cells. J Cell Sci, 1998. 111: p. 161-169.
Barker, P.E., Shipp, M.A., D'Adamio, L., Masteller, E.L., Reinherz, E.L., The common acute lymphoblastic leukemia
antigen gene maps to chromosomal region 3 (q21-q27). The Journal of Immunology, 1989. 142: p. 283-287.

108
Bibliografia
Barrett, A.J., Rawling, N.D., Woessner, J.F., Handbook of Proteolytic Enzymes. Barrett AJ, Rawling ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1.
Baudin, B., L'enzyme de conversion de l'angiotensine I (ECA) dans le diagnostic de la sarcoïdose. Pathologie Biologie, 2005. 53: p. 183-188.
Bauer, K., Inactivation of thyrotropin-releasing hormone (TRH) by the hormonally regulated TRH-degrading
ectoenzyme : A potential regulator of TRH signals? Trends in Endocrinology and Metabolism, 1995. 6: p. 101-105.
Bénéteau-Burnat, B., Baudin, B., Angiotensin-Converting Enzyme: Clinical Applications and Laboratory Investigations
on Serum and Other Biological Fluids. Critical Reviews in Clinical Laboratory Sciences, 1991. 28: p. 337-356.
Berne, R.M., Levy, M.N., Fisiologia. Harcourt, Mosby 3. Edizioa, 2002: p. 354-405.
Bhagwat, S.V., Lahdenranta, J., Giordano, R., Arap, W., Pasqualini, R., Shapiro, L.H., CD13/APN is activated by
angiogenic signals and is essential for capillary tube formation. Blood, 2001. 97: p. 652-659.
Blanco, L., Larrinaga, G., Perez, I., Lopez, J.I., Gil, J., Agirregoitia, E., Varona, A., Acid, basic, and neutral peptidases
present different profiles in chromophobe renal cell carcinoma and in oncocytoma. Am J Physiol Renal Physiol, 2008. 294: p. 850-858.
Blanco, L.; Perez, I.; Sanz, B.; Gil, J.; López, J.I.; Ugalde, A.; Varona, A.; Larrinaga, G., Changes in Cell-Surface Peptidase
Activity in Papillary Renal Cell Carcinoma. Anticancer Research, 2010. 30: p. 1137-1141.
Bogenrieder, T., Finstad, C.L., Freeman, R.H., Papandreou, C.N., Scher, H.I., Albino, A.P., Reuter, V.E., Nanus, D.M., Expression and localization of aminopeptidase A, aminopeptidase N, and dipeptidyl peptidase IV in benign and
malignant human prostate tissue. Prostate, 1997. 33: p. 225-232.
Bradford, M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Annal Biochem., 1976. 72: p. 248-254.
Bujanda, L., Sarasqueta, C., Hijona, E., Hijona, L., Cosme, A., Gil, I., Elorza, J.L., Asensio, J.I., Larburu, S., Enríquez-Navascués, J.M. , Jover, R., Balaguer, F., Llor, X., Bessa, X., Andreu, M., Paya, A., Castells, A., Gastrointestinal Oncology Group of the Spanish Gastroenterological Association, Colorectal cancer prognosis twenty years later. World J Gastroenterol, 2010. 16: p. 862–867.
Byrnes, J., Gross, S., Ellard, C., Connolly, K., Donahue, S., Picarella, D., Effects of the ACE2 inhibitor GL1001 on acute
dextran sodium sulfate-induced colitis in mice. Inflammation Research, 2009. 58: p. 819-827.
C
Cadel, S., Foulon, T., Viron, A., Balogh, A., Midol-Monnet, S., Noël, N., Cohen, P., Aminopeptidase B from the rat testis
is a bifunctional enzyme structurally related to leukotriene-A4hydrolase. Proceedings of the National Academy of Sciences, 1997. 94: p. 2963-2968.
Cadel, S., Pierotti, A.R., Foulon, T., Créminon, C., Barré, N., Segrétain, D., Cohen, P., Aminopeptidase-B in the rat testes:
isolation, functional properties and cellular localization in the seminiferous tubules. Mol Cell Endocrinol, 1995. 110: p. 149-160.
Cadel, S., Piesse, C., Gouzy-Darmon, C., Cohen, P., Foulon, T., Arginyl aminopeptidase. Aminopeptidases in biology and disease. Hooper NM, Lendeckel U. Kluwer Academic/Plenum Press, 2004. 2: p. 113-126.
Carl-McGrath, S., Lendeckel, U., Ebert, M., Rocken, C., Ectopeptidases in tumour biology: A review. Histol Histopathol, 2006. 21: p. 1339-1353.
Carrera, M.P., Ramírez-Expósito, M.J., Dueñas, B., Mayas, M.D., Garcia, M.J., De La Chica, S., Cortés, P., Ruíz-San Juan, M., Martínez-Martos, J.M., Insulin-regulated Aminopeptidase/Placental Leucil Aminopeptidase (IRAP/P-LAP) and
Angiotensin IV-forming Activities are Modified in Serum of Rats with Breast Cancer Induced by N-methyl-
nitrosourea. Anticancer Research, 2006. 26: p. 1011-1014.
Carrera, M.P., Ramírez-Expósito, M.J., García, M.J., Mayas, M.D., Martínez-Martos, J.M. , Ovarian Renin-Angiotensin
System-regulating Aminopeptidases Are Involved in Progesterone Overproduction in Rats with Mammary Tumours
Induced by N-Methyl Nitrosourea. Anticancer Research, 2009. 29: p. 4633-4637.

Bibliografia 109
Carrera, M.P., Ramirez-Expósito, M.J., Valenzuela, M.T., García, M.J., Mayas, M.D., Martínez-Martos, J.M., Glutamyl-
but Not Aspartyl- Aminopeptidase Activity is Modified in Serum of N-Methyl Nitrosourea-induced Rat Mammary
Tumours. Anticancer Research, 2004. 24: p. 801-806.
Cohen, A.J., Bunn, P.A., Franklin, W., Magill-Solc, C., Hartmann, C., Helfrich, B., Gilman, L., Folkvord, J., Helm, K., Miller, Y.E., Neutral Endopeptidase: Variable Expression in Human Lung, Inactivation in Lung Cancer, and Modulation of
Peptide-induced Calcium Flux. Cancer Research, 1996. 56: p. 831-839.
Constam, D.B., Tobler, A.R., Rensing-Ehl, A., Kemler, I., Hersh, L.B., Fontana, A., Puromycin-sensitive Aminopeptidase. Journal of Biological Chemistry, 1995. 270: p. 26931-26939.
Cordero, O.J., Ayude, D., Nogueira, M., Rodriguez-Berrocal, F.J., de La Cadena, M.P., Preoperative serum CD26 levels:
diagnostic efficiency and predictive value for colorectal cancer. Br J Cancer, 2000. 83: p. 1139-1146.
Cordero, O.J., De Chiara, L., Lemos-González, Y., De La Cadena, M.P., Rodriguez-Berrocal, F.J., How the Measurements
of a Few Serum Markers Can Be Combined to Enhance their Clinical Values in the Management of Cancer. Anticancer Research, 2008. 28: p. 2333-2341.
Cordero, O.J., Salgado, F., Nogueira, M. On the origin of serum CD26 and its altered concentration in cancer patients. Cancer Immunology, Immunotherapy, 2009. 58: p. 1723-1747.
Corvol, P., Eyries, M., Soubrier, F., Peptidyl-dipeptidase A/angiotensin I-converting enzyme. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 332-346.
Cotton, J., Hayashi, M.A.F., Cuniasse, P., Vazeux, G., Ianzer, D., De Camargo, A.C.M., Dive, V., Selective Inhibition of the
C-Domain of Angiotensin I Converting Enzyme by Bradykinin Potentiating Peptides. Biochemistry, 2002. 41: p. 6065-6071.
Crackower, M.A., Sarao, R., Oudit, G.Y., Yagil, C., Kozieradzki, I., Scanga, S.E., Oliveira-dos-Santos, A.J., da Costa, J., Zhang, L., Pei, Y., Scholey, J., Ferrario, C.M., Manoukian, A.S., Chappell, M.C., Backx, P.H., Yagil, Y., Penninger, J.M., Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature, 2002. 417: p. 822-828.
Cummins, P.M., O'Connor, B., Pyroglutamyl peptidase: an overview of the three known enzymatic forms. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology, 1998. 1429: p. 1-17.
D
Dall'Era, M.A., True, L.D., Siegel, A.F., Porte,r M.P., Sherertz, T.M., Liu, A.Y., Differential expression of CD10 in prostate
cancer and its clinical implication. BMC Urol, 2007. 7: p. 3.
Dando, P.M., Young, N.E., Barrett, A.J., Aminopeptidase PS: a widely distributed cytosolic peptidase. Proteolysis in cell functions. Proceeding of the 11
th International Conference on Proteolysis and Protein Turnover. Hopsu-Havu, V.K.
Edizioa. IOS Press. Amsterdam. , 1997: p. 88-95.
Davies, R.J., Miller, R., Coleman, N., Colorectal cancer screening: prospects for molecular stool analysis. Nat Rev Cancer, 2005. 5: p. 199-209.
Dawson, L.A., Maitland, N.J., Berry, P., Turner, A.J., Usmani, B.A., Expression and Localization of Endothelin-Converting
Enzyme-1 in Human Prostate Cancer. Exp. Biol. Med., 2006. 231: p. 1106-1110.
De Chiara, L., Rodríguez-Piñeiro, A.M., Cordero, O.J., Rodríguez-Berrocal, F.J., Ayude, D., Rivas-Hervada, F.J., de la Cadena M.P., Soluble CD26 levels and its association to epidemiologic parameters in a sample population. Dis Markers. , 2009. 27: p. 311-316.
De Chiara, L., Rodriguez-Pineiro, A., Rodriguez-Berrocal, F., Cordero, O., Martinez-Ares, D., Paez de la Cadena, M., Serum CD26 is related to histopathological polyp traits and behaves as a marker for colorectal cancer and
advanced adenomas. BMC Cancer, 2010. 10: p. 333.
De Gandarias, J.M., Barrallo, G., Digestio aparatuaren fisiologia eta patologia. Euskal Herriko Unibertsitatearen Argitalpen Zerbitzua, 2001: p. 293-359.
De Gandarias, J.M., Gil, J., Valdivia, A., Larrinaga, G., Artola, D., Casis, L., Subcellular distribution of pyroglutamyl-
peptidase I activity in the developing rat cerebellum. Dev Neurosci. 2000. 22: p. 264-273.

110
Bibliografia
De Gandarias, J.M., Irazusta, J., Echevarría, E., Casis, L., Neutral aminopeptidase activity levels during the estrous cycle
and the pregnancy in the hypothalamus and the pituitary of the rat. Life Sci., 1993. 52: p. 1629-1632.
De Gandarias, J.M., Irazusta, J., Echevarría, E., Gil, J., Casis, L., Age-related changes in the soluble and the membrane-
bound Tyr-aminopeptidase activities in several areas of the male and female rat brain. Reprod Fertil Dev., 1996. 8: p. 465-469.
De Gandarias, J.M., Irazusta, J., Gil, J., Fernández, D., Varona, A., Casis, L., Ontogeny of puromycin-sensitive and
insensitive aminopeptidase activities in several subcellular fractions of the rat brain. Brain Research Bulletin, 1999. 50: p. 283-290.
De la Haba-Rodríguez, J., Macho, A., Calzado, M.A., Blázquez, M.V., Gómez, M.A., Muñoz, E.E., Aranda, E., Soluble
dipeptidyl peptidase IV (CD-26) in serum of patients with colorectal carcinoma. Neoplasma, 2002. 49: p. 307-311.
De Souza, A.N., Bruno, J.A., Carvalho, K.M., An enkephalin degrading aminopeptidase of human brain preserved during
the vertebrate phylogeny. Comp Biochem Physiol C, 1991. 99: p. 363-367.
Del Rio, P., Crafa, P., Papadia, C., Dell'Abate, P., Franzè, A., Franzini, G., Campanini, N., Sianesi, M., Evaluation of CD10
positivity in colorectal polyps in neoplastic transformation Ann Ital Chir, 2010. 81:121-127.
Deshayes, F., Nahmias, C., Angiotensin receptors: a new role in cancer? Trends in Endocrinology and Metabolism, 2005. 16: p. 293-299.
Donoghue, M., Hsieh, F., Baronas, E., Godbout, K., Gosselin, M., Stagliano, N., Donovan, M., Woolf, B., Robison, K., Jeyaseelan, R., Breitbart, R.E., Acton, S., A Novel Angiotensin-Converting Enzyme-Related Carboxypeptidase (ACE2)
Converts Angiotensin I to Angiotensin 1-9. Circ Res, 2000. 87: p. 1-9.
Drucker, D.J., Nauck, M.A., The incretin system: glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4
inhibitors in type 2 diabetes. The Lancet, 2006. 368: p. 1696-1705.
Duffy, M.J., van Dalen, A., Haglund, C., Hansson, L., Holinski-Feder, E., Klapdor, R., Lamerz, R., Peltomaki, P., Sturgeon, C., Topolcan, O., Tumour markers in colorectal cancer: European Group on Tumour Markers (EGTM) guidelines for
clinical use. European journal of cancer (Oxford, England : 1990), 2007. 43: p. 1348-1360.
E
Erbeznik, H., Hersh, L.B., A Novel Mammalian High-Molecular-Weight Aminopeptidase. Archives of Biochemistry and Biophysics, 1997. 344: p. 228-234.
F
Feng, Y., Wan, H., Liu J., Zhang, R., Ma, Q., Han, B., Xiang, Y., Che, J., Cao, H., Fei, X., Qiu, W., The angiotensin-
converting enzyme 2 in tumor growth and tumor-associated angiogenesis in non-small cell lung cancer. Oncology reports, 2010. 23: p. 941-948.
Fernandez, M., Liu, X., Wouters, M.A., Heyberger, S., Husain, A., Angiotensin I-converting Enzyme Transition State
Stabilization by His1089. Journal of Biological Chemistry, 2001. 276: p. 4998-5004.
Ferrario, C.M., Chappell, M.C., Tallant, E.A., Brosnihan, K.B., Diz, D.I., Counterregulatory Actions of Angiotensin-(1-7). Hypertension, 1997. 30: p. 535-541.
Ferro, E.S., Hyslop, S., Camargo, A.C.M., Intracellullar peptides as putative natural regulators of protein interactions. Journal of Neurochemistry, 2004. 91: p. 769-777.
Florentin, D., Sassi, A., Roques, B., A highly sensitive fluorimetric assay for "enkephalinase", a neutral
metalloendopeptidase that releases Tyr-Gly-Gly from enkephalins. Annal Biochem, 1984. 141: p. 62-69.
Flores-Reséndiz, D., Castellanos-Juárezb, E., Benítez-Bribiescab, L., Las proteasas en la progresión neoplásica. Gac Méd Méx, 2009. 146: p. 131-142.

Bibliografia 111
Foulon, T., Cadel, S., Chesneau, V., Draoui, M., Prat, A., Cohen, P., Two Novel Metallopeptidases with a Specificity for
Basic Residues Functional Properties, Structure, and Cellular Distribution. Annals of the New York Academy of Sciences, 1996. 780: p. 106-120.
Foulon, T., Cadel, S., Cohen, P., Aminopeptidase B (EC 3.4.11.6). The International Journal of Biochemistry & Cell Biology, 1999. 31: p. 747-750.
Foulon, T., Cadel, S., Piesse, C., Cohen, P., Aminopeptidase B. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 328-332.
Friedman, T.C., Yanovski, J.A., Jayasvasti, V., Yanovski, S.Z., Koenig, R.J., Wilk, S., Pyroglutamyl peptidase-II
("thyroliberinase") activity in human serum: influence of weight and thyroid status. J Clin Endocrinol Metab, 1995. 80: p. 1086-1089.
Fujimura, H., Ino, K., Nagasaka, T., Nakashima, N., Nakazato, H., Kikkawa, F., Mizutani, S., Aminopeptidase A expression
in cervical neoplasia and its relationship to neoplastic transformation and progression. Oncology, 2000. 58: p. 342-352.
Fukasawa, K.M., Fukasawa, K., Kanai, M., Fujii, S., Harada, M., Molecular Cloning and Expression of Rat Liver
Aminopeptidase B. Journal of Biological Chemistry, 1996. 271: p. 30731-30735.
Fyhrquist, F., Saijonmaa, O., Renin-angiotensin system revisited. Journal of Internal Medicine, 2008. 264: p. 224-236.
G
Gaggar, A., Jackson, P.L., Noerager, B.D., O´Reilly, P.J., McQuaid, D.B., Rowe, S.M., Clancy, J.P., Blalock, J.E., A Novel
Proteolytic Cascade Generates an Extracellular Matrix-Derived Chemoattractant in Chronic Neutrophilic
Inflammation. The Journal of Immunology, 2008. 180: p. 5662-5669.
Gainer, H., Russell, J.T., Loh, Y.P., An aminopeptidase activity in bovine pituitary secretory vesicles that cleaves the N-
terminal arginine from [beta]-lipotropin60-65. FEBS Letters, 1984. 175: p. 135-139.
García-Horsman, J.A., Männistö, P.T., Venäläinen, J.I., On the role of prolyl oligopeptidase in health and disease. Neuropeptides, 2007. 41: p. 1-24.
Garro, M.A., Giza Fisiologia. Digestio Aparatua. Elhuyar Fundazioa, 2000: p. 192-220.
Gass, J., Khosla, C., Prolyl endopeptidases. Cellular and Molecular Life Sciences, 2007. 64: p. 345-355.
Gembardt, F., Sterner-Kock, A., Imboden, H., Spalteholz, M., Reibitz, F., Schultheiss, H.P., Siems, W.E., Walther, T., Organ-specific distribution of ACE2 mRNA and correlating peptidase activity in rodents. Peptides, 2005. 26: p. 1270-1277.
Gialeli, C., Theocharis, A.D., Karamanos, N.K., Roles of matrix metalloproteinases in cancer progression and their
pharmacological targeting. FEBS Journal, 2011. 278: p. 16-27.
Gil, J., Larrinaga, G., Meana, J.J., Rodriguez-Puertas, R., Irazusta, J., Casis, L., Regional and subcellular distribution of
soluble aminopeptidase in the human and the rat brain: a comparative study. Neuropeptides, 2001. 35: p. 276-284.
Glenner, G.G., McMillan, P.J., Folk, J.E., A mammalian peptidase specific for the hydrolysis of N-terminal alpha-L-
glutamyl and aspartyl residues. Nature, 1962: p. 194-867.
Goessl, C., Noninvasive molecular detection of cancer - the bench and the bedside. Curr Med Chem, 2003. 10(8): p. 691-706.
Göhring, B., Holzhausen, H.J., Meye, A., Heynemann, H., Rebmann, U., Langner, J., Riemann, D., Endopeptidase
24.11/CD10 is down-regulated in renal cell cancer. Int J Mol Med., 1998. 2: p. 409-414.
Gómez-Domínguez, E., Trapero-Marugán, M., del Pozo, A.J., Cantero, J., Gisbert, J.P., Maté, J., The colorectal
carcinoma prognosis factors. Significance of diagnosis delay. Rev Esp Enferm Dig, 2006. 98: p. 322-329.
Goossens, F., De Meester, I., Vanhoof, G., Scharpé, S., Distribution of Prolyl Oligopeptidase in Human Peripheral
Tissues and Body Fluids. Clinical Chemistry and Laboratory Medicine, 1996b. 34: p. 17-22.

112
Bibliografia
Goossens, F.J., Wauters, J.G., Vanhoof, G.C., Bossuyt, P.J., Schatteman, K.A., Loens, K., Scharpé, S.L., Subregional
mapping of the human lymphocyte prolyl oligopeptidase gene (PREP) to human chromosome 6q22. Cytogenet Cell Genet, 1996a. 74: p. 99-101.
Goto, Y., Hattori, A., Ishii, Y., Mizutani, S., Tsujimoto, M., Enzymatic Properties of Human Aminopeptidase A. Journal of Biological Chemistry, 2006. 281: p. 23503-23513.
Gotoh, H., Hagihara, M., Nagatsu, T., Iwata, H., Miura, T., Activity of dipeptidyl peptidase IV and post-proline cleaving
enzyme in sera from osteoporotic patients. Clin Chem, 1988. 34: p. 2499-2501.
Greene, F.L., Stewart, A.K., Norton, H.J., A New TNM Staging Strategy for Node-Positive (Stage III) Colon Cancer: An
Analysis of 50,042 Patients. Annals of Surgery, 2002. 236: p. 416-421.
Grioli, S., Lomeo, C., Quattropani, M.C., Spignoli, G., Villardita, C., Pyroglutamic acid improves the age associated
memory impairment. Fundam Clin Pharmacol, 1990. 4: p. 169-173.
Groblewska, M., Mroczko, B., Szmitkowski, M., The role of selected matrix metalloproteinases and their inhibitors in
colorectal cancer development. Postepy Hig Med Dosw, 2010. 64: p. 22-30.
Groblewska, M., Mroczko, B., Wereszczyńska-Siemiątkowska, U., Kędra, B., Łukaszewicz, M., Baniukiewicz, A., Szmitkowski, M., Serum interleukin 6 (IL-6) and C-reactive protein (CRP) levels in colorectal adenoma and cancer
patients. Clinical Chemistry and Laboratory Medicine, 2008. 46: p. 1423-1428.
Gros, C., Giros, B., Schwartz, J.C., Identification of aminopeptidase M as an enkephalin-inactivating enzyme in rat
cerebral membranes. Biochemistry, 1985a. 24: p. 2179-85.
Gros, C., Giros, B., Schwartz, J.C., Purification of membrane-bound aminopeptidase from rat brain: Identification of
aminopeptidase M. Neuropeptides, 1985b. 5(4-6): p. 485-488.
Guy, J.L., Lambert, D.W., Warner, F.J., Hooper, N.M., Turner, A.J., Membrane-associated zinc peptidase families:
comparing ACE and ACE2. Biochimica et Biophysica Acta (BBA) - Proteins & Proteomics, 2005. 1751(1): p. 2-8.
H
Haeggström, J.Z., Aminopeptidase PS. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 313-316.
Hallberg, M., Le Grevés, P., Nyberg, F., Neuropeptide processing. Proteases in the Brain. Lendeckel N.M., Hooper U. Edizioa. Springer Science, 2005: p. 203-234.
Hanahan, D., Weinberg, R.A., Hallmarks of Cancer: The Next Generation. Cell, 2011. 144: p. 646-674.
Hanahan, D., Weinberg, R.A., The Hallmarks of Cancer. Cell, 2000. 100: p. 57-70.
Hao, X.P., Pretlow, T.G., Rao, J.S., Pretlow, T.P., β-Catenin Expression Is Altered in Human Colonic Aberrant Crypt Foci. Cancer Research, 2001. 61: p. 8085-8088.
Hashida, H., Takabayashi, A., Kanai, M., Adachi, M., Kondo, K., Kohno, N., Yamaoka, Y., Miyake, M., Aminopeptidase N
is involved in cell motility and angiogenesis: its clinical significance in human colon cancer. Gastroenterology, 2002. 122: 376-386.
Havre, P.A., Abe, M., Urasaki, Y., Ohnuma, K., Morimoto, C., Dang, N.H., The role of CD26/dipeptidyl peptidase IV in
cancer. Frontiers in Bioscience, 2008. 13: p. 1634-1645.
Heasley, L.E., Autocrine and paracrine signaling through neuropeptide receptors in human cancer. Oncogene, 2001. 20: p. 1563-1569.
Herszényi, L., Sipos, F.,Galamb, O., Solymosi, N., Hritz, I., Miheller, P., Berczi, L., Molnár, B., Tulassay, Z., Matrix
Metalloproteinase-9 Expression in the Normal Mucosa–Adenoma–Dysplasia–Adenocarcinoma Sequence of the
Colon. Pathology & amp; Oncology Research, 2008. 14: p. 31-37.
Heslin, M.J., Yan, J., Johnson, M.R., Weiss, H., Diasio, R.B., Urist, M.M., Role of matrix metalloproteinases in colorectal
carcinogenesis. Ann Surg. , 2001. 233: p. 786-792.

Bibliografia 113
Hirasawa, K., Sato, Y., Hosoda, Y., Yamamoto, T., Hanai, H., Immunohistochemical Localization of Angiotensin II
Receptor and Local Renin-Angiotensin System in Human Colonic Mucosa. Journal of Histochemistry & Cytochemistry, 2002. 50: p. 275-282.
Hiroi, Y., Endo, Y., Natori, Y., Purification and properties of an aminopeptidase from rat-liver cytosol. Arch Biochem Biophys, 1992. 294: p. 440-445.
Hooper, N.M., Angiotensin converting enzyme: implications from molecular biology for its physiological functions. Int J Biochem., 1991. 23: p. 641-647.
Hooper, N.M., Turner, A.J., An ACE structure. Nat Struct Mol Biol, 2003. 10(3): p. 155-157.
Hopsu, V.K., Mäkinn, K.K., Glenner, G.G., Characterization of aminopeptidase B: substrate specificity and affector
studies. Arch Biochem Biophys, 1966. 114: p. 567-575.
Howell, S., Murray, H., Scott, C.S., Turner, A.J., Kenny, A.J., A highly sensitive E.L.I.S.A. for endopeptidase-24.11, the
common acute-lymphoblastic-leukaemia antigen (CALLA, CD-10), applicable to material of porcine and human
origin. Biochem J., 1991. 278: p. 417-421.
http://www.control-incontinencia-fecal.es
http://www.adam.com
http://www.chem.qmul.ac.uk/iubmb/enzyme/
http://www.cancerstaging.org/
Hubert, C., Houot, A.M., Corvol, P., Soubrier, F., Structure of the angiotensin I-converting enzyme gene. Two alternate
promoters correspond to evolutionary steps of a duplicated gene. Journal of Biological Chemistry, 1991. 266: p. 15377-15383.
Hui, K.S., Brain-Specific Aminopeptidase: From Enkephalinase to Protector Against Neurodegeneration. Neurochemical Research, 2007. 32: p. 2062-2071.
Hunyady, L., Catt, K.J., Pleiotropic AT1 Receptor Signaling Pathways Mediating Physiological and Pathogenic Actions of
Angiotensin II. Mol Endocrinol, 2006. 20: p. 953-970.
Hurst, N.G., Stocken, D.D., Wilson, S., Keh, C., Wakelam, M.J.O., Ismail, T., Elevated serum matrix metalloproteinase 9
(MMP-9) concentration predicts the presence of colorectal neoplasia in symptomatic patients. Br J Cancer, 2007. 97: p. 971-977.
I
Iacobuzio-Donahue, C.A., Shuja, S., Cai, J., Peng, P., Murnane, M.J., Elevations in Cathepsin B Protein Content and
Enzyme Activity Occur Independently of Glycosylation during Colorectal Tumor Progression. Journal of Biological Chemistry, 1997. 272: p. 29190-29199.
Ino, K., Shibata, K., Kajiyama, H., Kikkawa, F., Mizutani, S., Regulatory role of membrane-bound peptidases in the
progression of gynecologic malignancies. Biological Chemistry, 2004. 385: p. 683-690.
Ino, K., Uehara, C., Kikkawa, F., Kajiyama, H., Shibata, K., Suzuki, T., Khin, E.E., Ito, M., Takeuchi, M., Itakura, A., Mizutani, S., Enhancement of Aminopeptidase A Expression during Angiotensin II-Induced Choriocarcinoma Cell
Proliferation through AT1 Receptor Involving Protein Kinase C- and Mitogen-Activated Protein Kinase-Dependent
Signaling Pathway. J Clin Endocrinol Metab, 2003. 88: p. 3973-3982.
Irazusta, J., Larrinaga, G., Gonzalez-Maeso, J., Gil, J., Meana, J.J., Casis, L., Distribution of prolyl endopeptidase activities
in rat and human brain. Neurochem Int, 2002. 40(4): p. 337-45.
Irazusta, J., Valdivia, A., Fernandez, D., Agirregoitia, E., Ochoa, C., Casis, L., Enkephalin-Degrading Enzymes in Normal
and Subfertile Human Semen. J Androl, 2004. 25: p. 733-739.
Ishii, K., Usui, S., Sugimura, Y., Yoshida, S., Hioki, T., Tatematsu, M., Yamamoto, H., Hirano, K., Aminopeptidase N
regulated by zinc in human prostate participates in tumor cell invasion. Int J Cancer, 2001. 92: p. 49-54.

114
Bibliografia
Ito, N., Nomura, S., Iwase, A., Ito, T., Kikkawa, F., Tsujimoto, M., Ishiura, S., Mizutani, S., ADAMs, a disintegrin and
metalloproteinases, mediate shedding of oxytocinase. Biochemical and Biophysical Research Communications, 2004. 314: p. 1008-1013.
Ito, T., Nomura, S., Okada, M., Katsumata, Y., Iwase, A., Kikkawa, F., Tsujimoto, M., Mizutani, S., Transcriptional
regulation of human placental leucine aminopeptidase/oxytocinase gene. Molecular Human Reproduction, 2001. 7: p. 887-894.
Iwaki-Egawa, S., Watanabe, Y., Kikuya, Y., Fujimoto, Y., Dipeptidyl Peptidase IV from Human Serum: Purification,
Characterization, and N-Terminal Amino Acid Sequence. Journal of Biochemistry, 1998. 124: p. 428-433.
Izarzugaza, M.I., Martínez, R., Audicana, C., Larrañaga, N., Hernández, E., Tobalina, M.C., De la Cruz, M., Laviñeta, M., Hurtado, R., San Sebastián, M.C., De Miguel, A.R., Michelena, M.J., López de la Calle, J., Minbizia EAE. Intzidentzia,
hilkortasuna, biziraupena eta bilakaera denboran zehar. Eusko Jaurlaritza, 2010.
J
Jass, J.R., Serrated route to colorectal cancer: back street or super highway?. J. Pathol, 2001. p. 193-128.
Jass, J.R., Sobin, L.H. , Histological typing of intestinal tumours. World Health Organization, Jass, J.R., Sobin, L.H. In collaboration with pathologists in 9 countries. 2. Edizioa, 1989. p. 8-41.
K
Kandror, K.V., Yu, L., Pilch, P.F., The major protein of GLUT4-containing vesicles, gp160, has aminopeptidase activity. Journal of Biological Chemistry, 1994. 269: p. 30777-30780.
Kedika, R., Patel, M., Pena Sahdala, H.N., Mahgoub, A., Cipher, D., Siddiqui, A.A., Long-term Use of Angiotensin
Converting Enzyme Inhibitors Is Associated With Decreased Incidence of Advanced Adenomatous Colon Polyps. Journal of Clinical Gastroenterology, 2011. 45: p. e12-e16, doi: 10.1097/MCG.0b013e3181ea1044.
Keller, S.R., Scott, H.M., Mastick, C.C., Aebersold, R., Lienhard, G.E., Cloning and Characterization of a Novel Insulin-
regulated Membrane Aminopeptidase from Glut4 Vesicles. Journal of Biological Chemistry, 1995. 270: p. 23612-23618.
Kelly, J.A., Neidle, E.L., Neidle, A., An aminopeptidase from mouse brain cytosol that cleaves N-terminal acidic amino
acid residues. J Neurochem, 1983. 40: p. 1727-1734.
Kerr, M.A., Kenny, A.J., The molecular weight and properties of a neutral metallo-endopeptidase from rabbit kidney
brush border. Biochem J., 1974b. 137: p. 489-495.
Kerr, M.A., Kenny, A.J., The purification and specificity of a neutral endopeptidase from rabbit kidney brush border. Biochem J., 1974a. 137: p. 477-488.
Kikkawa, F., Kajiyama, H., Shibata, K., Ino, K., Nomura, S., Mizutani, S., Dipeptidyl peptidase IV in tumor progression. Biochimica et Biophysica Acta (BBA) - Proteins & Proteomics, 2005. 1751: p. 45-51.
Kim, J.T., Song, E.Y., Chung, K.S., Kang, M.A., Kim, J.W., Kim, S.J., Yeom, Y., Kim, J.H., Kim, K.H., Lee, H.G., Up-regulation
and clinical significance of serine protease kallikrein 6 in colon cancer. Cancer, 2010: doi: 10.1002/cncr.25841.
Koga, Y., Yao, T., Hirahashi, M., Kumashiro, Y., Ohji, Y., Yamada, T., Tanaka, M., Tsuneyoshi, M., Flat adenoma–
carcinoma sequence with high-malignancy potential as demonstrated by CD10 and β-catenin expression: a
different pathway from the polypoid adenoma–carcinoma sequence. Histopathology, 2008. 52: p. 569-577.
Kojima, K., Mihara, R., Sakai T., Togari, A., Matsui, T., Shinpo, K., Fujita, K., Fukasawa, K., Harada, M., Nagatsu, T., Serum activities of dipeptidyl-aminopeptidase II and dipeptidyl-aminopeptidase IV in tumor-bearing animals and in
cancer patients. Biochem Med Metab Biol. , 1987. 37: p. 35-41.
Kuester, D., Lippert, H., Roessner, A., Krueger, S., The cathepsin family and their role in colorectal cancer. Pathology - Research and Practice, 2008. 204: p. 491-500.

Bibliografia 115
Kumar, V., Abbas, A.K., Fausto, N., Mitchell, R.N. Riñón y Vía Urinaria. Robbins Patología Humana. 8. Edizioa. Elsevier Saunders. 2008: p. 587-590.
L
Larrinaga, G., Callado, L.F., Agirregoitia, N., Varona, A., Gil, J., Subcellular distribution of membrane-bound
aminopeptidases in the human and rat brain. Neurosci Lett, 2005. 383: p. 136-140.
Larrinaga, G., López, J.I., Casis, L., Blanco, L., Gil, J., Agirregoitia, E., Varona, A., Cystinyl aminopeptidase activity is
decreased in renal cell carcinomas. Regul Pept, 2007. 144: p. 56-61.
Larrinaga, G., Perez, I., Blanco, L., López, J.I., Andrés, L., Etxezarraga, C., Santaolalla, F., Zabala, A., Varona, A., Irazusta, J., Increased prolyl endopeptidase activity in human neoplasia. Regulatory Peptides, 2010a. 163: p. 102-106.
Larrinaga, G., Perez, I., Sanz, B., Blanco, L., López, J.I., Cándenas, M. L., Pinto, F. M., Gil, J., Irazusta, J., Varona, A., Angiotensin-converting enzymes (ACE and ACE2) are downregulated in renal tumors. Regulatory Peptides, 2010b. 165:218-223.
Larrinaga, G., Perez, I., Sanz, B., Zarrazquin, I., Casis, L., Anta, J.A., Martínez, A., Santaolalla, F., Altered dipeptidyl
peptidase IV and prolyl endopeptidase activities in chronic tonsillitis, tonsillar hyperplasia and adenoid
hyperplasia. Int J of Pediatr Otorhinolaryngol, 2011. 75: p. 347-350
Lauffart, B., Mantle, D., Rationalization of aminopeptidase activities in human skeletal muscle soluble extract. Biochim Biophys Acta, 1988. 956: p. 300-306.
Laustsen, P.G., Rasmussen, T.E., Petersen, K., Pedraza-Diaz, S., Moestrup, S.K., Gliemann, J., Sottrup-Jensen, L., Kristensen, T., The complete amino acid sequence of human placental oxytocinase. Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression, 1997. 1352: p. 1-7.
Lee, S., Bang, S., Song, K., Lee, I., Differential expression in normal-adenoma-carcinoma sequence suggests complex
molecular carcinogenesis in colon. Oncol Rep, 2006. 16: p. 747-754.
Lee, W.S., Baek, J.H., Kim, K.K., Park, Y.H., The prognostic significant of percentage drop in serum CEA post curative
resection for colon cancer. Surgical Oncology, 2010. doi: 10.1016/j.suronc.2010.10.003.
Leslie, A., Carey, F.A., Pratt, N.R., Steele, R.J.C., The colorectal adenoma–carcinoma sequence. British Journal of Surgery, 2002. 89: p. 845-860.
Leung, P.S., Renin-Angiotensin System Proteases and the Cardiometabolic Syndrome: Pathophysiological, Clinical and
Therapeutic Implications. Frontiers in research of the renin-angiotensin system on human disease. Leung P.S. Edizioa. Springer. Holanda., 2007: p. 87-112.
Li, J., Wilk, E., Wilk, S., Inhibition of Prolyl Oligopeptidase by Fmoc-Aminoacylpyrrolidine-2-Nitriles. Journal of Neurochemistry, 1996. 66: p. 2105-2112.
Li, W., Moore, M.J., Vasilieva, N., Sui, J., Wong, S.K., Berne, M.A., Somasundaran, M., Sullivan, J.L., Luzuriaga, K., Greenough, T.C., Choe, H., Farzan, M., Angiotensin-converting enzyme 2 is a functional receptor for the SARS
coronavirus. Nature, 2003. 426: p. 450-454.
Liu, J.M., Kusinski, M., Ilic, V., Bignon, J., Hajem, N., Komorowski, J., Kuzdak, K., Stepien, H., Wdzieczak-Bakala, J., Overexpression of the Angiogenic Tetrapeptide AcSDKP in Human Malignant Tumors. Anticancer Research, 2008. 28: p. 2813-2817.
Liu, L., Qian, J., Singh, H., Meiers, I., Zhou, X., Bostwick, D.G., Immunohistochemical Analysis of Chromophobe Renal
Cell Carcinoma, Renal Oncocytoma, and Clear Cell Carcinoma: An Optimal and Practical Panel for Differential
Diagnosis. Archives of Pathology & Laboratory Medicine, 2007. 131: p. 1290-1297.
Loizate, A., Tumore markatzaileak eta antigorputz monoklonalak. Elhuyar Fundazioa, 1990. 16: p. 61-73.
Luan, Y., Xu, W., The structure and main functions of aminopeptidase N. Curr Med Chem, 2007. 14: p. 639-47.

116
Bibliografia
M
Maes, M., Lin, A., Bonaccorso, S., Vandoolaeghe, E., Song, C., Goossens, F., De Meester, I., Degroote, J., Neels, H., Scharpé, S., Janca, A., Lower Activity of Serum Peptidases in Abstinent Alcohol-Dependent Patients. Alcohol, 1999b. 17: p. 1-6.
Maes, M., Lin, A.H., Bonaccorso, S., Goossens, F., Van Gastel, A., Pioli, R., Delmeire, L., Scharpé, S., Higher serum prolyl
endopeptidase activity in patients with post-traumatic stress disorder. Journal of Affective Disorders, 1999a. 53: p. 27-34.
Maes, M., Monteleone, P., Bencivenga, R., Goossens, F., Maj, M., van West, D., Bosmans, E., Scharpe, S., Lower serum
activity of prolyl endopeptidase in anorexia and bulimia nervosa. Psychoneuroendocrinology, 2001. 26: p. 17-26.
Malfroy, B., Kuang, W.J., Seeburg, P.H., Mason, A.J., Schofield, P.R., Molecular cloning and amino acid sequence of
human enkephalinase (neutral endopeptidase). FEBS Letters, 1988. 229: p. 206-210.
Manfredi, M.A., Fox, V.L., Zurakowski, D., Kutok, J.L., Glickman, J.N., Moses, M.A., Matrix Metalloproteinases in the
Urine and Tissue of Patients With Juvenile Polyps: Potential Biomarkers for the Presence of Polyps. Journal of Pediatric Gastroenterology and Nutrition, 2009. 48: p. 405-411.
Mantle, D., Comparison of soluble aminopeptidases in human cerebral cortex, skeletal muscle and kidney tissues. Clinica Chimica Acta, 1992. 207: p. 107-118.
Mantle, D., Hardy, M.F., Lauffart, B., McDermott, J.R., Smith, A.I., Pennington, R.J., Purification and characterization of
the major aminopeptidase from human skeletal muscle. Biochem J., 1983. 211: p. 567-573.
Mantle, D., Lauffart, B., McDermott, J., Gibson, A., Characterization of aminopeptidases in human kidney soluble
fraction. Clin Chim Acta, 1990. 187: p. 105-113.
Marchiò, S., Lahdenranta, J., Schlingemann, R.O., Valdembri, D., Wesseling, P., Arap, M.A., Hajitou, A., Ozawa, M.G., Trepel, M., Giordano, R.J., Nanus, D.M., Dijkman, H.B.P. M., Oosterwijk, E., Sidman, R.L., Cooper, M.D., Bussolino, F., Pasqualini, R., Arap, W., Aminopeptidase A is a functional target in angiogenic blood vessels. Cancer Cell, 2004. 5: p. 151-162.
Marks, N., Datta, R.K., Lajtha, A., Partial Resolution of Brain Arylamidases and Aminopeptidases. Journal of Biological Chemistry, 1968b. 243: p. 2882-2889.
Marks, N., Datta, R.K., Lajtha, A., The relationship of aminotripeptidase and arylamidase to protein breakdown in the
brain. Macromolecules and Function of the neurone. Lodin Z. Edizioa. Excerpta Medica, 1968a.
Marks, N., Stern, F., Lajtha, A., Changes in proteolytic enzymes and proteins during maturation of the brain. Brain Research, 1975. 86: p. 307-322.
Martinez, J.M., Prieto, I., Ramirez, M.J., Cueva, C., Alba, F., Ramirez, M., Aminopeptidase Activities in Breast Cancer
Tissue. Clin Chem, 1999. 45: p. 1797-1802.
Matsas, R., Stephenson, S.L., Hryszko, J., Kenny, A.J., Turner, A.J., The metabolism of neuropeptides. Phase separation
of synaptic membrane preparations with Triton X-114 reveals the presence of aminopeptidase N. Biochem J. , 1985. 231: p. 445-449.
Matsumoto, H., Nagasaka, T., Hattori, A., Rogi, T., Tsuruoka, N., Mizutani, S., Tsujimoto, M., Expression of placental
leucine aminopeptidase/oxytocinase in neuronal cells and its action on neuronal peptides. European Journal of Biochemistry, 2001. 268: p. 3259-3266.
Matsumoto, H., Rogi, T., Yamashiro, K., Kodama, S., Tsuruoka, N., Hattori, A., Takio, K., Mizutani, S., Tsujimoto, M., Characterization of a recombinant soluble form of human placental leucine aminopeptidase/oxytocinase
expressed in Chinese hamster ovary cells. European Journal of Biochemistry, 2000. 267: p. 46-52.
McDonald, J.K., Barrett, A.J., Mammalian Proteases: A Glossary and Bibliography. Exopeptidases. Academic Press, 1986. 2: p. 59-71.
McLellan, S., Dyer, S.H., Rodriguez, G., Hersh, L.B., Studies on the tissue distribution of the puromycin-sensitive
enkephalin-degrading aminopeptidases. J Neurochem. , 1988. 51: p. 1552-1559.
Meyerhardt, J., Saunders, M., Cáncer Colorrectal. Dana-Farber Cancer Institute, Elsevier Mosby, 2008: p. 23-57.

Bibliografia 117
Mitsui, T., Nomura, S., Itakura, A., Mizutani, S., Role of Aminopeptidases in the Blood Pressure Regulation. Biological & Pharmaceutical Bulletin, 2004. 27: p. 768-771.
Momeni, N., Yoshimoto, T., Ryberg, B., Sandberg-Wollheim, M., Grubb, A., Factors influencing analysis of prolyl
endopeptidase in human blood and cerebrospinal fluid: increase in assay sensitivity. Scand J Clin Lab Invest., 2003. 63: p. 387-395.
Mook, O.R.F., Frederiks, W.M., Van Noorden, C.J.F., The role of gelatinases in colorectal cancer progression and
metastasis. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer, 2004. 1705: p. 69-89.
Murakami, H., Yokoyama, A., Kondo, K., Nakanishi, S., Kohno, N., Miyake, M., Circulating aminopeptidase N/CD13 is an
independent prognostic factor in patients with non-small cell lung cancer. Clin Cancer Res, 2005. 11: p. 8674-8679.
Myöhänen, T.T., Tenorio-Laranga, J., Jokinen, B., Vázquez-Sánchez, R., Moreno-Baylach, M.J., García-Horsman, J.A., Männistö, P.T., Prolyl oligopeptidase induces angiogenesis both in vitro and in vivo in a novel regulatory manner. British Journal of Pharmacology, 2010: doi:10.1111/j.1476-5381.2010.01146.x
N
Nanus, D.M., Of Peptides and Peptidases. Clinical Cancer Research, 2003. 9: p. 6307-6309.
Neo, J., Ager, E., Angus, P., Zhu, J., Herath, C., Christophi, C., Changes in the renin angiotensin system during the
development of colorectal cancer liver metastases. BMC Cancer, 2010. 10: p. 134.
Noble, F., Roques, B.P., Protection of endogenous enkephalin catabolism as natural approach to novel analgesic
and antidepressant drugs. Expert Opinion on Therapeutic Targets, 2007. 11: p. 145-159.
Noffsinger, A.E., Serrated Polyps and Colorectal Cancer: New Pathway to Malignancy. Annual Review of Pathology: Mechanisms of Disease, 2009. 4: p. 343-364.
Nomura, S., Ito, T., Mizutani, S., Placental leucine aminopeptidase. Aminopeptidases in biology and disease. Hooper NM, Lendeckel U. Kluwer Academic/Plenum Press, 2004b: p. 45-59.
Nomura, S., Ito, T., Yamamoto, E., Sumigama, S., Iwase, A., Okada, M., Shibata, K., Ando, H., Ino, K., Kikkawa, F., Mizutani, S., Gene regulation and physiological function of placental leucine aminopeptidase/oxytocinase during
pregnancy. Biochimica et Biophysica Acta (BBA) - Proteins & Proteomics, 2005. 1751: p. 19-25.
Nomura, S., Tsujimoto, M., Mizutani, S., Cystinyl aminopeptidase, oxytocinase and insulin-regulated aminopeptidase. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004a. 1: p. 307-309.
O
O´Cuinn, G., Peptide metabolism in cytoplasm of brain cells. Biochem Soc Trans. , 1998. 26(3): p. 279-92.
Ocain, T.D., Rich, D.H., L-Lysinethiol: A subnanomolar inhibitor of aminopeptidase B. Biochemical and Biophysical Research Communications, 1987. 145: p. 1038-1042.
Ogawa, H., Iwaya, K., Izumi, M., Kuroda, M., Serizawa, H., Koyanagi, Y., Mukai, K., Expression of CD10 by stromal cells
during colorectal tumor development. Human Pathology, 2002. 33: p. 806-811.
O'Leary, R.M., Gallagher, S.P., O'Connor, B., Purification and characterization of a novel membrane-bound form of
prolyl endopeptidase from bovine brain. The International Journal of Biochemistry & Cell Biology, 1996. 28: p. 441-449.
Oparil, S., Koerner, T.J., O'Donoghue, J.K., Structural requirements for substrates and inhibitors of angiotensin I-
converting enzyme in vivo and vitro. Circ Res., 1974b. 40: p. 19-26.
Oparil, S., Koerner, T.J., Haber, E., Effects of pH and Enzyme Inhibitors on Apparent Generation of Angiotensin I in
Human Plasma. J Clin Endocrinol Metab, 1974a. 39: p. 965-968.

118
Bibliografia
Osada, T., Watanabe, G., Kondo, S., Toyoda, M., Sakaki, Y., Takeuchi, T., Male Reproductive Defects Caused by
Puromycin-Sensitive Aminopeptidase Deficiency in Mice. Mol Endocrinol, 2001b. 15: p. 960-971.
Osada, T., Watanabe, G., Sakaki, Y., Takeuchi, T., Puromycin-Sensitive Aminopeptidase Is Essential for the Maternal
Recognition of Pregnancy in Mice. Mol Endocrinol, 2001a. 15: p. 882-893.
Osasunerako Mundu Erakundea (OME) : http://www.who.int/en/ WHO International Histological Classification of Tumours Jass and Sobin: Histological Typing of Intestinal Tumours, 2nd edn, 1989.
Oset, P., Jasińska, A., Szcześniak, P., Orszulak-Michalak, D., Talar-Wojnarowska, R., Małecka-Panas, E., Analysis of
serum gastrin levels in patients with adenomatous polyps of the colon. Pol Merkur Lekarski, 2009. 26: p. 458-461.
P
Papadopoulou, S., Scorilas, A., Arnogianaki, N., Papapanayiotou, B., Tzimogiani, A., Agnantis, N., Talieri, M., Expression
of gelatinase-A (MMP-2) in human colon cancer and normal colon mucosa. Tumour Biol, 2001. 22: p. 383-389.
Papandreou, C.N., Usmani, B., Geng, Y., Bogenrieder, T., Freeman, R., Wilk, S., Finstad, C.L., Reuter, V.E., Powell, C.T., Scheinberg, D., Magill, C., Scher, H.I., Albino, A.P., Nanus, D.M., Neutral endopeptidase 24.11 loss in metastatic
human prostate cancer contributes to androgen-independent progression. Nat Med, 1998. 4: p. 50-57.
Pasqualini, R., Koivunen, E., Kain, R., Lahdenranta, J., Sakamoto, M., Stryhn, A., Ashmun, R.A., Shapiro, L.H., Arap, W., Ruoslahti, E., Aminopeptidase N Is a Receptor for Tumor-homing Peptides and a Target for Inhibiting
Angiogenesis. Cancer Research, 2000. 60: p. 722-727.
Paul, M., Poyan Mehr, A., Kreutz, R., Physiology of Local Renin-Angiotensin Systems. Physiological Reviews, 2006. 86: p. 747-803.
Perez, I., Varona, A., Blanco, L., Gil, J., Santaolalla, F., Zabala, A., Ibarguen, A.M. Irazusta, J., Larrinaga, G., Increased
APN/CD13 and acid aminopeptidase activities in head and neck squamous cell carcinoma. Head & Neck, 2009. 31(10): p. 1335-1340.
Petrovic, N., Schacke, W., Shapiro L.H., CD13/Aminopeptidase N in tumor growth and angiogenesis. Hooper NM, Lendeckel U, edizioa. Aminopeptidases in Biology and Disease. New York: Kluwer Academic/Plenum Press, 2004: p. 179-200.
Pro, B., Dang N.H., CD26/dipeptidyl peptidase IV and its role in cancer. Histol Histopathl, 2004. 19: p. 1345-1351.
R
Raica, M., Cimpean, A.M., Ribatti, D., Angiogenesis in pre-malignant conditions. European Journal of Cancer, 2009. 45: p. 1924-1934.
Ramírez-Expósito, M.J., Martínez, J.M., Prieto, I., Alba, F., Ramírez, M., Comparative distribution of glutamyl and
aspartyl aminopeptidase activities in mouse organs. Horm Metab Res, 2000. 32: p. 161-163.
Rawlings, N.D., Barrett, A.J., Evolutionary families of metallopeptidases. Methods Enzymol, 1995. 248: p. 183-228.
Rawlings, N.D., Barrett, A.J., MEROPS: the peptidase database. Nucleic Acids Research, 1999. 27: p. 325-331.
Rawlings, N.D., Polgar, L., Barrett, A.J., A new family of serine-type peptidases related to prolyl oligopeptidase. Biochem J., 1991. 279: p. 907-908.
Re, R.N., Cook, J.L., An intracrine view of angiogenesis. BioEssays, 2006. 28: p. 943-953.
Re, R.N., Cook, J.L., The Basis of an Intracrine Pharmacology. The Journal of Clinical Pharmacology, 2008. 48: p. 344-350.
Re, R.N., Intracellular Renin and the Nature of Intracrine Enzymes. Hypertension, 2003. 42: p. 117-122.
Re, R.N., The Origins of Intracrine Hormone Action. The American Journal of the Medical Sciences, 2002. 323: p. 43-48.

Bibliografia 119
Ren, X., Glende, J., Al-Falah, M., de Vries, V., Schwegmann-Wessels, C., Qu, X., Tan, L., Tschernig, T., Deng, H., Naim, H.Y., Herrler, G., Analysis of ACE2 in polarized epithelial cells: surface expression and function as receptor for
severe acute respiratory syndrome-associated coronavirus. J Gen Virol, 2006. 87: p. 1691-1695.
Riemann, D., Kehlen, A., Langner, J., CD13-not just a marker in leukemia typing. Immunology Today, 1999. 20: p. 83-88.
Rigat, B., Hubert, C., Alhenc-Gelas, F., Cambien, F., Corvol, P., Soubrier, F., An insertion/deletion polymorphism in the
angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. The Journal of Clinical Investigation, 1990. 86: p. 1343-1346.
Röcken, C., Neumann, K., Carl-McGrath, S., Lage, H., Ebert, M.P.A., Dierkes, J., Jacobi, C.A., Kalmuk, S., Neuhaus, P., Neumann U., The Gene Polymorphism of the Angiotensin I-Converting Enzyme Correlates with Tumor Size and
Patient Survival in Colorectal Cancer Patients. Neoplasia, 2007. 9: p. 716–722.
Rogi, T., Tsujimoto, M., Nakazato, H., Mizutani, S., Tomoda, Y., Human Placental Leucine Aminopeptidase/Oxytocinase. Journal of Biological Chemistry, 1996. 271: p. 56-61.
Roques, B.P., Beaumont, A., Neutral endopeptidase-24.11 inhibitors: from analgesics to antihypertensives? Trends Pharmacol Sci., 1990. 11: p. 245-249.
Roques, B.P., Fournié-Zaluski, M.C., Soroca, E., Lecomte, J.M., Malfroy, B., Llorens, C., Schwartz, J.C., The
enkephalinase inhibitor thiorphan shows antinociceptive activity in mice. Nature, 1980. 288: p. 286-288.
Roy, R., Yang, J., Moses, M.A., Matrix Metalloproteinases As Novel Biomarker s and Potential Therapeutic Targets in
Human Cancer. Journal of Clinical Oncology, 2009. 27: p. 5287-5297.
Ruiz-Ortega, M., Esteban, V., Egido, J., The Regulation of the Inflammatory Response Through Nuclear Factor-[kappa]B
Pathway by Angiotensin IV Extends the Role of the Renin Angiotensin System in Cardiovascular Diseases. Trends in Cardiovascular Medicine, 2007. 17: p. 19-25.
S
Saha, A.K., Smith, K.J.E., Sue-Ling, H., Sagar, P.M., Burke, D., Finan, P.J., Prognostic factors for survival after curative
resection of Dukes’ B colonic cancer. Colorectal Disease, 2010: doi: 10.1111/j.1463-1318.2010.02507.x.
Salers, P., Ouafik, L.H., Giraud, P., Maltese, J.Y., Dutour, A., Oliver, C., Ontogeny of prolyl endopeptidase, pyroglutamyl
peptidase I, TRH, and its metabolites in rat pancreas. American Journal of Physiology - Endocrinology And Metabolism, 1992. 262: p. 845-850.
Sanderink, G.J., Artur, Y., Siest, G., Human aminopeptidases: a review of the literature. J Clin Chem Clin Biochem., 1988. 26: p. 795-807.
Schally, A.V., Engel, J., Analogs of luteinizing hormone-releasing hormone (LHRH) in cancer. Handbook of biologically active peptides. Kastin AJ. Elsevier Academic Press, 2006: p. 421-428.
Schlingemann, R.O., Oosterwijk, E., Wesseling, P., Rietveld, F.J.R., Ruiter, D.J., Aminopeptidase A is a constituent of
activated pericytes in angiogenesis. The Journal of Pathology, 1996. 179: p. 436-442.
Schmitmeier, S., Thole, H., Bader, A., Bauer, K., Purification and characterization of the thyrotropin-releasing hormone
(TRH)-degrading serum enzyme and its identification as a product of liver origin. European Journal of Biochemistry, 2002. 269: p. 1278-1286.
šedo, A., KŘepela, E., Kasafírek, E., Dipeptidyl peptidase IV, prolyl endopeptidase and cathepsin B activities in primary
human lung tumours and lung parenchyma. Journal of Cancer Research and Clinical Oncology, 1991. 117: p. 249-253.
Selvakumar, P., Lakshmikuttyamma, A., Kanthan, R., Kanthan, S.C., Dimmock, J.R., Sharma, R.K., High Expression of
Methionine Aminopeptidase 2 in Human Colorectal Adenocarcinomas. Clinical Cancer Research, 2004. 10: p. 2771-2775.
Selves, J., Histological types and prognostic factors in colorectal cancer. Med Sci (Paris), 2009. 25: p. 9-12.

120
Bibliografia
Shibata, K., Kikkawa, F., Suzuki, Y., Mizokami, Y., Kajiyama, H., Ino, K., Nomura, S., Nagasaka, T., Mizutani, S., Expression of placental leucine aminopeptidase is associated with a poor outcome in endometrial endometrioid
adenocarcinoma. Oncology, 2004.66: p. 288-295.
Shrimpton, C.N., Smith, A.I., Lew, R.A., Soluble Metalloendopeptidases and Neuroendocrine Signaling. Endocr Rev, 2002. 23: p. 647-664.
Shuja, S., Sheahan, K., Murnane, M.J., Cysteine endopeptidase activity levels in normal human tissues, colorectal
adenomas and carcinomas. Int J Cancer, 1991. 49: p. 341-346.
Sillars-Hardebol, A., Carvalho, B., de Wit, M., Postma, C., Delis-van Diemen, P., Mongera, S., Ylstra, B., van de Wiel, M., Meijer, G., Fijneman, R., Identification of key genes for carcinogenic pathways associated with colorectal
adenoma-to-carcinoma progression. Tumor Biology, 2010. 31: p. 89-96.
Silveira, P.F., Irazusta, J., Gil, J., Agirregoitia, N., Casis, L., Interactions among challenges of hydromineral balance,
angiotensin-converting enzyme, and cystine aminopeptidase. Peptides, 2001. 22: p. 2137-2144.
Sjöholm, I., Yman, L., Degradation of oxytocin, lysine-vasopressin, angiotensin II and angiotensin-II-amide by
oxytocinase (cystine aminopeptidase). Acta Pharm Suec, 1967. 4: p. 65-76.
Sjöström, H., Norén, O., Olsen, J., Structure and Function of Aminopeptidase N, in Cellular Peptidases in Immune
Functions and Diseases 2, J. Langner and S. Ansorge, Editors. 2002, Springer Netherlands. p. 25-34.
Skidgel, R.A., Erdös, E.G., Novel activity of human angiotensin I converting enzyme: release of the NH2- and COOH-
terminal tripeptides from the luteinizing hormone-releasing hormone. Proceedings of the National Academy of Sciences, 1985. 82: p. 1025-1029.
Skrzydlewska, E., Sulkowska, M., Wincewicz, A., Koda, M., Sulkowski, S., Evaluation of serum cathepsin B and D in
relation to clinicopathological staging of colorectal cancer. World J Gastroenterol, 2005. 11: p. 4225-4229.
Smith, A.I., Turner, A.J., What´s new in the renin-angiotensin system? Cellular and Molecular Life Sciences, 2004. 61: p. 2475-2676.
Speth, R., Karamyan, V., The significance of brain aminopeptidases in the regulation of the actions of angiotensin
peptides in the brain. Heart Failure Reviews, 2008. 13: p. 299-309.
Spisák, S., Kalmár, A.,Galamb, O., Sipos, F., Wichmann, B., Molnár, B., Tulassay, Z., Identification of methylation related
genes from Lacer Capture Microdissected colon samples during investigation of adenoma–carcinoma sequence. Orvosi Hetilap 2010. 151: p. 805-814.
Stecca, B.A., Nardo, B., Chieco, P., Mazziotti, A., Bolondi, L., Cavallari, A., Aberrant dipeptidyl peptidase IV (DPP
IV/CD26) expression in human hepatocellular carcinoma. Journal of Hepatology, 1997. 27: p. 337-345.
Stevens, A., Lowe, J., Histologia Humana. Tracto digestivo. Stevens A., Lowe J. 3. Edizioa. Elsevier Mosby, 2006: p. 189-229.
Stewart, J.M., Gera, L. , Bradykinin and cancer. Handbook of biologically active peptides. Kastin AJ. Elsevier Academic Press, 2006: p. 443-446.
Stoltze, L., Schirle, M., Schwarz, G., Schroter, C., Thompson, M.W., Hersh, L.B., Kalbacher, H., Stevanovic, S., Rammensee, H.G., Schild, H., Two new proteases in the MHC class I processing pathway. Nat Immunol, 2000. 1: p. 413-418.
Stragier, B., De Bundel, D., Sarre, S., Smolders, I., Vauquelin, G., Dupont, A., Michotte, Y., Vanderheyden, P., Involvement of insulin-regulated aminopeptidase in the effects of the renin–angiotensin fragment angiotensin IV:
a review. Heart Failure Reviews, 2008. 13: p. 321-337.
Sturiale, S., Barbara, G., Qiu, B., Figini, M., Geppetti, P., Gerard, N., Gerard, C., Grady, E.F., Bunnett, N.W., Collins, S.M., Neutral endopeptidase (EC 3.4.24.11) terminates colitis by degrading substance P. Proceedings of the National Academy of Sciences of the United States of America, 1999. 96: p. 11653-11658.
Suda, H., Aoyagi, T., Takeuchi, T., Umezawa, H., Inhibition of aminopeptidase B and leucine aminopeptidase by bestatin
and its stereoisomer. Archives of Biochemistry and Biophysics, 1976. 177: p. 196-200.
Sumitani, S., Ramlal, T., Somwar, R., Keller, S.R., Klip, A., Insulin Regulation and Selective Segregation with Glucose
Transporter-4 of the Membrane Aminopeptidase vp165 in Rat Skeletal Muscle Cells. Endocrinology, 1997. 138: p. 1029-1034.

Bibliografia 121
Sumitomo, M., Shen, R., Nanus, D.M., Involvement of neutral endopeptidase in neoplastic progression. Biochimica et Biophysica Acta (BBA) - Proteins & Proteomics, 2005. 1751: p. 52-59.
Suzuki, T., Kikkawa, F., Ino, K., Nagasaka, T., Tamakoshi, K., Mizutani, S., Imbalance between neutral endopeptidase
24.11 and endothelin-1 expression in human endometrial carcinoma. Oncology, 2001. 60: p. 258-267.
Suzuki, Y., Shibata, K., Kikkawa, F., Kajiyama, H., Ino, K., Nomura, S., Tsujimoto, M., Mizutani, S., Possible Role of
Placental Leucine Aminopeptidase in the Antiproliferative Effect of Oxytocin in Human Endometrial
Adenocarcinoma. Clinical Cancer Research, 2003. 9: p. 1528-1534.
Szeltner, Z., Polgár, L., Structure, function and biological relevance of prolyl oligopeptidase. Curr Protein Pept Sci, 2008. 9: p. 96-107.
Szewczuk, A, Kwiatkowska, J., Pyrrolidonyl peptidase in animal, plant and human tissues. Occurrence and some
properties of the enzyme. Eur J Biochem, 1970. 15(1): p. 92-96.
T
Tahara, K., Mimori, K., Iinuma, H., Iwatsuki, M., Yokobori, T., Ishii, H., Anai, H., Kitano, S., Mori, M., Serum Matrix-
Metalloproteinase-1 is a Bona Fide Prognostic Marker for Colorectal Cancer. Annals of Surgical Oncology, 2010. 17: p. 3362-3369.
Talieri, M., Li, L., Zheng, Y., Alexopoulou, D. K., Soosaipillai, A., Scorilas, A., Xynopoulos, D., Diamandis, E. P., The use of
kallikrein-related peptidases as adjuvant prognostic markers in colorectal cancer. Br J Cancer, 2009. 100: p. 1659-1665.
Talieri, M., Papadopoulou, S., Scorilas, A., Xynopoulos, D., Arnogianaki, N., Plataniotis, G., Yotis, J., Agnanti, N., Cathepsin B and cathepsin D expression in the progression of colorectal adenoma to carcinoma. Cancer Letters, 2004. 205: p. 97-106.
Tan, E.Y., Mujoomdar, M., Blay, J., Adenosine Down-Regulates the Surface Expression of Dipeptidyl Peptidase IV on HT-
29 Human Colorectal Carcinoma Cells: Implications for Cancer Cell Behavior. Am J Pathol, 2004. 165: p. 319-330.
Tan, E.Y., Richard, C.L., Zhang, H., Hoskin, D.W., Blay, J., Adenosine downregulates DPPIV on HT-29 colon cancer cells
by stimulating protein tyrosine phosphatase(s) and reducing ERK1/2 activity via a novel pathway. American Journal of Physiology - Cell Physiology, 2006. 291: p. 433-444.
Tanioka, T., Hattori, A., Masuda, S., Nomura, Y., Nakayama, H., Mizutani, S., Tsujimoto, M., Human Leukocyte-derived
Arginine Aminopeptidase. Journal of Biological Chemistry, 2003. 278: p. 32275-32283.
Taylor, A., Aminopeptidases: structure and function. The FASEB Journal, 1993. 7: p. 290-298.
Teranishi, J., Ishiguro, H., Hoshino, K., Noguchi, K., Kubota, Y., Uemura, H., Evaluation of role of angiotensin III and
aminopeptidases in prostate cancer cells. The Prostate, 2008. 68: p. 1666-1673.
Thibodeau, G.A., Patton, K.T., Anatomia y Fisiologia. Sistema Digestivo. Thibodeau, G.A., Patton, K.T. Elsevier-Mosby Edizioa, 2007: p. 638-641.
Thompson, M.W., Tobler, A., Fontana, A., Hersh, L.B., Cloning and Analysis of the Gene for the Human Puromycin-
Sensitive Aminopeptidase. Biochemical and Biophysical Research Communications, 1999. 258: p. 234-240.
Tiruppathi, C., Ganapathy, V., Leibach, F.H., Evidence for tripeptide-proton symport in renal brush border membrane
vesicles. Studies in a novel rat strain with a genetic absence of dipeptidyl peptidase IV. Journal of Biological Chemistry, 1990. 265: p. 2048-2053.
Tobler, A.R., Constam, D.B., Schmitt-Gräff, A., Malipiero, U., Schlapbach, R., Fontana, A., Cloning of the Human
Puromycin-Sensitive Aminopeptidase and Evidence for Expression in Neurons. Journal of Neurochemistry, 1997. 68: p. 889-897.
Tokuhara, T., Hattori, N., Ishida, H., Hirai, T., Higashiyama, M., Kodama, K., Miyake, M., Clinical significance of
aminopeptidase N in non-small cell lung cancer. Clin Cancer Res, 2006. 12: p. 3971-3978.
Toll, A.D., Fabius, D., Hyslop, T., Pequignot, E., DiMarino, A.J., Infantolino, A., Palazzo, J.P., Prognostic significance of
high grade dysplasia in colorectal adenomas. Colorectal Disease, 2010: doi: 10.1111/j.1463-1318.2010.02385.x.

122
Bibliografia
Tsuji, E., Misumi, Y., Fujiwara, T., Takami, N., Ogata, S., Ikehara, Y., An active-site mutation (Gly633-->Arg) of dipeptidyl
peptidase IV causes its retention and rapid degradation in the endoplasmic reticulum. Biochemistry, 1992. 31: p. 11921-11927.
Tucker, E.L., Pignatelli, M., Catenins and their associated proteins in colorectal cancer. Histol Histopathol, 2000. 15: p. 251-260.
Turner, A.J., Endopeptidase-24.11 and neuropeptide metabolism. Neuropeptides and their peptidases. Ellis Horwood Ltd, 1987.
Turner, A.J., Hooper, N.M., Angiotensin converting enzyme 2. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 349-352.
Turner, A.J., Hooper, N.M., The angiotensin-converting enzyme gene family: genomics and pharmacology. Trends in Pharmacological Sciences, 2002a. 23(4): p. 177-183.
Turner, A.J., Membrane alanyl aminopeptidase. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 289-294.
Turner, A.J., Processing and metabolism of neuropeptides. Essays Biochem, 1986. 22: p. 69-119.
Turner, A.J., Tipnis, S.R., Guy, J.L., Rice, G.I., Hooper, N.M., ACEH/ACE2 is a novel mammalian metallocarboxypeptidase
and a homologue of angiotensin-converting enzyme insensitive to ACE inhibitors. Canadian Journal of Physiology and Pharmacology, 2002b. 80: p. 346-353.
U
Usmani, B.A., Janeczko, M., Shen, R., Mazumdar, M., Papandreou, C.N., Nanus, D.M., Analysis of the insertion/deletion
polymorphism of the human angiotensin converting enzyme (ACE) gene in patients with renal cancer. Br J Cancer, 2000. 82: p. 550-552.
V
Valdivia, A., Irazusta, J., Fernández, D., Múgica, J., Ochoa, C., Casis, L., Pyroglutamyl peptidase I and prolyl
endopeptidase in human semen: increased activity in necrozoospermia. Regulatory Peptides, 2004. 122: p. 79-84.
van Hensbergen, Y., Broxterman, H.J., Hanemaaijer, R., Jorna, A.S., van Lent, N.A., Verheul, H.M.W., Pinedo, H.M., Hoekman, K., Soluble Aminopeptidase N/CD13 in Malignant and Nonmalignant Effusions and Intratumoral Fluid. Clinical Cancer Research, 2002. 8: p. 3747-3754.
Varona, A., Blanco, L., López, J.I., Gil, J., Agirregoitia, E., Irazusta, J., Larrinaga, G., Altered levels of acid, basic, and
neutral peptidase activity and expression in human clear cell renal cell carcinoma. American Journal of Physiology-Renal Physiology, 2007. 292: p. 780-788.
Varona, A., Blanco, L., Perez, I., Gil, J., Irazusta, J., Lopez, J.I., Candenas, M.L., Pinto, F., Larrinaga, G., Expression and
activity profiles of DPP IV/CD26 and NEP/CD10 glycoproteins in the human renal cancer are tumor-type
dependent. BMC Cancer, 2010. 10: p. 193.
Vazeux, G., Wang, J., Corvol, P., Llorens-Cortès, C., Identification of Glutamate Residues Essential for Catalytic Activity
and Zinc Coordination in Aminopeptidase A. Journal of Biological Chemistry, 1996. 271: p. 9069-9074.
Vickers, C., Hales, P., Kaushik, V., Dick, L., Gavin, J., Tang, J., Godbout, K., Parsons, T., Baronas, E., Hsieh, F., Acton, S., Patane, M., Nichols, A., Tummino, P., Hydrolysis of Biological Peptides by Human Angiotensin-converting Enzyme-
related Carboxypeptidase. Journal of Biological Chemistry, 2002. 277: p. 14838-14843.

Bibliografia 123
W
Walter, R., Shlank, H., Glass, J.D., Schwartz, I.L., Kerenyi, T.D., Leucylglycinamide released from oxytocin by human
uterine enzyme. Science, 1971. 173: p. 827-829.
Wang, G., Lai, F.M.M., Lai, K.B., Chow, K.M., Kwan, C.H.B., Li, K.T.P., Szeto, C.C., Discrepancy between Intrarenal
Messenger RNA and Protein Expression of ACE and ACE2 in Human Diabetic Nephropathy. Am J Nephrol, 2009. 29: p. 524-531.
Wang, J., Cooper, M.D., Aminopeptidase A. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 299-303.
Wang, J., Walker, H., Lin, Q., Jenkins, N., Copeland, N.G., Watanabe, T., Burrows, P.D., Cooper, M.D., The MouseBP-
1Gene: Structure, Chromosomal Localization, and Regulation of Expression by Type I Interferons and Interleukin-7. Genomics, 1996. 33: p. 167-176.
Warner, F.J., Smith, A.I., Hooper, N.M., Turner, A.J., Angiotensin-converting enzyme-2: a molecular and cellular
perspective. Cellular and Molecular Life Sciences, 2004. 61: p. 2704-2713.
Wei, L., Clauser, E., Alhenc-Gelas, F., Corvol, P., The two homologous domains of human angiotensin I-converting
enzyme interact differently with competitive inhibitors. Journal of Biological Chemistry, 1992. 267: p. 13398-13405.
Welches, W.R., Santos, R.A., Chappell, M.C., Brosnihan, K.B., Greene, L.J., Ferrario, C.M., Evidence that prolyl
endopeptidase participates in the processing of brain angiotensin. J Hypertens., 1991. 9: p. 631-638.
Wiedeman, P.E., Trevillyan, J.M., Dipeptidyl peptidase IV inhibitors for the treatment of impaired glucose tolerance and
type 2 diabetes. Curr Opin Investig Drugs., 2003. 4: p. 412-420.
Wilcox, C.S., Williams, C.M., Smith, T.B., Frederickson, E.D., Wingo, C., Bucci, C.M., Diagnostic uses of angiotensin-
converting enzyme inhibitors in renovascular hypertension. Am J Hypertens, 1988. 1: p. 344-349.
Wilk, S., Aspartyl aminopeptidase. Handbook of Proteolytic Enzymes. Barrett AJ, Rawlings ND, Woessner JF. 2. Edizioa. Elsevier Academic Press, 2004. 1: p. 937-939.
Wilk, S., Thurston, L.S., Inhibition of angiotensin III formation by thiol derivatives of acidic amino acids. Neuropeptides., 1990. 16: p. 163-168.
Wilk, S., Wilk, E., Magnusson, R.P., Identification of histidine residues important in the catalysis and structure of
aspartyl aminopeptidase. Archives of Biochemistry and Biophysics, 2002. 407: p. 176-183.
Wilk, S., Wilk, E., Magnusson, R.P., Purification, Characterization, and Cloning of a Cytosolic Aspartyl Aminopeptidase. Journal of Biological Chemistry, 1998. 273: p. 15961-15970.
Y
Yamamoto, H., Iku, S., Adachi, Y., Imsumran, A., Taniguchi, H., Nosho, K., Min, Y., Horiuchi, S., Yoshida, M., Itoh, F., Imai, K., Association of trypsin expression with tumour progression and matrilysin expression in human colorectal
cancer. The Journal of Pathology, 2003. 199: p. 176-184.
Yoshimoto, T., Fischl, M., Orlowski, R.C., Walter, R., Post-proline cleaving enzyme and post-proline dipeptidyl
aminopeptidase. Comparison of two peptidases with high specificity for proline residues. Journal of Biological Chemistry, 1978. 253: p. 3708-3716.
Yoshioka, M., Erickson, R.H., Matsumoto, H., Gum, E., Kim, Y.S., Expression of dipeptidyl aminopeptidase IV during
enterocytic differentiation of human colon cancer (Caco-2) cells. Int J Cancer. , 1991. 47: p. 916-921.

124
Bibliografia
Z
Zagon, I.S., Verderame, M.F., McLaughlin, P.J., The biology of the opioid growth factor receptor (OGFr). Brain Research Reviews, 2002. 38: p. 351-376.
Zhang, H., Wada, J., Hida, K., Tsuchiyama, Y., Hiragushi, K., Shikata, K., Wang, H., Lin, S., Kanwar, Y.S., Makino, H., Collectrin, a Collecting Duct-specific Transmembrane Glycoprotein, Is a Novel Homolog of ACE2 and Is
Developmentally Regulated in Embryonic Kidneys. Journal of Biological Chemistry, 2001. 276: p. 17132-17139.
Zhang, M.Z., Qiao, Y.H., Suo, Z.H., Correlation of DPPIV expression with clinicopathological features and prognosis in
epithelial ovarian carcinoma. Zhonghua Zhong Liu Za Zhi, 2008. 30: p. 848-852.
Zhou, L., Zhang, R., Yao, W., Wang, J., Qian, A., Qiao, M., Zhang, Y., Yuan, Y., Decreased Expression of Angiotensin-
Converting Enzyme 2 in Pancreatic Ductal Adenocarcinoma Is Associated with Tumor Progression. Tohoku J. Exp. Med., 2009. 217: p. 123-131.
Zini, S., Fournie-Zaluski, M.C., Chauvel, E., Roques, B.P., Corvol, P., Llorens-Cortes, C., Identification of metabolic
pathways of brain angiotensin II and III using specific aminopeptidase inhibitors: predominant role of angiotensin
III in the control of vasopressin release. Proceedings of the National Academy of Sciences, 1996. 93: p. 11968-11973.
Zucker, S., Vacirca, J., Role of matrix metalloproteinases (MMPs) in colorectal cancer. Cancer and Metastasis Reviews, 2004. 23: p. 101-117.

Eranskinak


Eranskinak 127
I. Eranskina Heste lodiko lagin freskoaren diagnosirako/pronostikorako ospitalean erabiltzen duten protokoloa
_____________________________________________________________________________________ Intraoperatoria SI___ (Referir Nº)___. NO___ Nº Total Bloques: Especificar procedencia (de cada nº) Nº Refs. Previas:_________________________________ RHTs, Ref:____ Patólogo: ____ Información Clínica relevante: CARCINOMA COLORRECTAL:PROTOCOLO DIAGNÓSTICO/PRONÓST ICO (CAP) .[Actualizado 01/2005]. Sólo aplicable en carcinomas invasivos. Basado en el TNM, 6ª ed., 01/2003 EXAMEN MACROSCÓPICO PROCEDIMIENTO QUIRÚRGICO ___ Hemicolectomía Dcha. * Longitud: ___ cm ___ Colectomía Transversa * Longitud: ___ cm ___ Hemicolectomía Izq. * Longitud: ___ cm ___ Sigmoidectomía * Longitud: ___ cm ___ Rectosigmoidectomía (Resección anterior baja) * Longitud: ___ cm ___ Colectomía total abdominal * Longitud: ___ cm ___ Resección Abdominoperineal * Longitud: ___ cm ___ Otro (especificar): ___________________________ * Longitud: ___ cm ___ No especificable LOCALIZACIÓN DEL TUMOR ___ Ciego ___ Colon dcho. (ascendente) ___ Flexura Hepática ___ Colon Transverso ___ Flexura Esplénica ___ Colon izq. (descendente) ___ Colon Sigmoide ___Colon ___ Rectosigmoide ___ Recto ___ No puede determinarse (ver Comentario) * CONFIGURACIÓN DEL TUMOR * ___ Exofítico (Polipoide) * ___ Infiltrativo * ___ Ulcerado * ___ Otro (especificar): ______ TAMAÑO DEL TUMOR Dimensión máxima: en cm ___ * Dimensiones adicionales: en cm ___ x ___ ___ No puede determinarse (ver Comentario) * MESORECTUM * ___ No aplicable * ___ Extirpación completa * ___ Extirpación casi completa * ___ Extirpación incompleta ESTUDIO MICROSCÓPICO TIPO HISTOLÓGICO ___ Adenocarcinoma ___ Adenocarcinoma Mucinoso (> del 50% de componente mucinoso) ___ Carcinoma de celulas "en anillo de sello" (>del 50% de células en "anillo de sello") ___ Carcinoma Medular ___ Carcinoma de Celula Pequeña ___ Carcinoma Indiferenciado ___ Otro (especificar): ________________________ ___ No puede determinarse ## RESUMEN DIAGNOSTICO Y ESTADIAJE FINAL EN PÁGINA 4 ##
Pág. 1/5

128 Eranskinak
GRADO HISTOLÓGICO ___ No aplicable ___ No puede evaluarse ___ Bajo-grado(bien a moderadamente diferenciado) ___ Alto-grado (pobremente diferenciado o indiferenciado) ___ Otro (especificar): ___________________________ ESTADIAJE POSTQUIRÚRGICO TUMOR PRIMARIO (pT) -pTX ___: No puede evaluarse -pT0 ___: No evidencia de tumor primario -pTis ___: Carcinoma "in situ", intraepitelial (sin invasión) -pTis ___: Carcinoma "in situ", con invasión de la lámina propia -pT1____: El tumor invade la submucosa -pT2____: El tumor invade la capa muscular propia -pT3____: El tumor invade, a través de la capa muscular, la subserosa , la grasa pericólica o el tejido perirrectal no peritonealizado
*pT3a___: Invasión mínima: <1 mm desde el límite externo de la capa muscular propia *pT3b___: Invasión ligera: de 1 a 5 mm desde el límite de la capa muscular *pT3c___: Invasión moderada: > de 5 mm y < de 15 mm desde el límite de la capa muscular *pT3d___: Invasión extensa: > de 15 mm desde el límite de la capa muscular
-pT4a____: El tumor invade directamente otros órganos o estructuras -pT4b____: El tumor perfora el peritoneo visceral GANGLIOS LINFATICOS REGIONALES (pN) -pNX ___: No puede evaluarse -pN0 ___: No evidencia de metástasis en ganglios linfáticos regionales -pN1 ___: Metástasis en 1 a 3 ganglios linfáticos regionales -pN2 ___: Metástasis en 4 ó más ganglios linfáticos regionales
Especificar: Nº ganglios examinados: ___ Nº ganglios metastatizados: ___
METÁSTASIS A DISTANCIA (pM) -pMX___: No puede evaluarse -pM1 ___: Metástasis a distancia * Especificar región(es):_____________________________ CATEGORIZACIÓN ESTADIO Estadio Grupos TNM Est. Dukes Modificado (Aster-Coller) Estadio Dukes __________________________________________________________________________________ Estadio 0 Tis N0 M0 Super A - Estadio I T1 N0 M0 A A
T2 N0 M0 B1 A Estadio IIA T3 N0 M0 B2 B Estadio IIB T4 N0 M0 B3 B Estadio IIIA T1,T2 N1 M0 C1 C Estadio IIIB T3,T4 N1 M0 C2,C3 C Estadio IIIC Cualq T N2 M0 C1,C2,C3 C Estadio IV Cualq T Cualq N M1 D - MÁRGENES QUIRÚRGICOS -Margen Proximal ___ No afectado por carcinoma invasivo ___ Afectado por carcinoma invasivo
.Carcinoma in situ/adenoma ausente en margen proximal:___
.Carcinoma in situ/adenoma presente en margen proximal:___ -Margen Distal ___ No afectado por carcinoma invasivo ___ Afectado por carcinoma invasivo
.Carcinoma in situ/adenoma ausente en margen distal:____
.Carcinoma in situ/adenoma presente en margen distal:____ Pág. 2/5

Eranskinak 129
Borde Circunferencial (radial) ___ No aplicable ___ No afectado por carcinoma invasivo ___ Afectado por carcinoma invasivo (tumor presente entre 0-1 mm del borde Radial) -Distancia del carcinoma invasivo al margen más cercano: ____mm. Referir qué margen___________ *Margen mesentérico *___ No puede precisarse *___ No afectado por carcinoma invasivo *___ Afectado por carcinoma invasivo -Distancia del carcinoma invasivo al margen más cercano: ____mm ó ___ cm
Referir qué margen___________
INVASIÓN VASO LINFÁTICO (VASO PEQUEÑO) [L] ___ Ausente ___ Presente: * ___ Intramural * ___ Extramural ___ Indeterminable INVASIÓN VENOSA (VASO GRANDE) [V] ___ Ausente ___ Presente: * ___ Intramural * ___ Extramural ___ Indeterminable *INVASIÓN PERINEURAL * ___ Ausente * ___Presente * PATRÓN DE CRECIMIENTO TUMORAL * ___ Expansivo * ___ Infiltrativo * RESPUESTA LINFOIDE INTRATUMORAL/PERITUMORAL * ___ Nula/muy escasa * ___ Ligera/moderada * ___ Marcada (incluyendo Enf. de Crohn) * DATOS PATOLÓGICOS ADICIONALES * ___ No identificados * ___ Adenoma(s) * ___ Proctocolitis ulcerosa crónica * ___ Enfermedad de Crohn * ___ Displasia * ___ Otro/s pólipo/s (tipo[s]): _______________________ *___ Otros (especificar): _________________________ ESTUDIOS INMUNOHISTOQUÍMICOS COMPLEMENTARIOS Especificar cada uno de los Acs testados y reflejar resultados (intensidad inmunotinción y % de células marcadas
Pág. 3/5

130 Eranskinak
RESUMEN DIAGNÓSTICO Y ESTADIAJE FINAL #COMENTARIO/S:
Pág.4/5

Eranskinak 131
ACLARACIONES SUPLETORIAS.- 1.- Cambios introducidos en relación con la edición previa del TNM # Se realiza una revisión de la anatomía del intestino grueso que establece los límites entre el colon, el recto y el canal anal. - Los Adenocarcinomas del apéndice cecal se clasifican separadamente, pero siguiendo los mismos criterios de la clasificación TNM que los utilizados en el Cáncer Colorrectal - Los cánceres del canal anal se clasifican en un protocolo independiente (segregado del Carcinoma Colorrectal) # Los nódulos metastásicos bien delimitados, presentes en la grasa pericólica o perirrectal se consideran metástasis ganglionares y se valoran en el apartado pN - Por el contrario, los nódulos metastásicos irregularmente contorneados, observados en la grasa peritumoral, pasan a considerarse como invasión vascular y se codificará como una extensión de la categoría pT, en el subapartado V1 si solo se reconoce microscopicamente y en el subapartado V2 si existe evidencia macroscópica grosera . # El estadiaje del Grupo II se subdivide en IIA o en IIB si el tumor primario corresponde a un pT3 o pT4, respectivamente. # El estadiaje del Grupo III se subdivide en IIIA (T1-2N1M0), IIIB (T3-4N1M0) o IIIC (cualquier TN2M0). # El procedimiento quirúrgico consistente en la resección anterior baja ( resección abdominoperineal ) del Cáncer de Recto, se ha agregado al protocolo. ________________________________________________________________ 2.- Cumplimentación de Datos En el Protocolo del Cáncer Colo-Rectal los datos señalados con un asterisco (*) no son requeribles para la cumplimentación del Informe AP acreditado por la Comisión del Cáncer. Estos datos pueden ser clínicamente importantes, pero su utilización aún no está validada o su aplicación no está estandarizada de manera regular. Alternativamente, pueden existir datos necesarios que no estén disponibles o que sean inaplicables por el patólogo en el momento de la valoración anatomopatológica del espécimen. El apartado precedido del signo &&& corresponde a datos complementarios Inmunohistoquímicos que opcional o regladamente venían incluyéndose en los Protocolos inciales confeccionados en el SAP CHT =========================================================================== 3.- Addendum.- Junto con las Indicaciones y Comentarios (sin traducir), anexos a la totalidad de los Protocolos de Cáncer, para la cumplimentación del Protocolo del Carcinoma Colorrectal, se encuentra disponible, además, para consulta y guía, el siguiente documento: Colina F, Ibarrola C. Protocolo e Información sistematizada para los Estudios Histopatológicos relacionados con el Carcinoma Colorrectal Rev Esp Patol 2004;37: 73-90
Pág.5/5
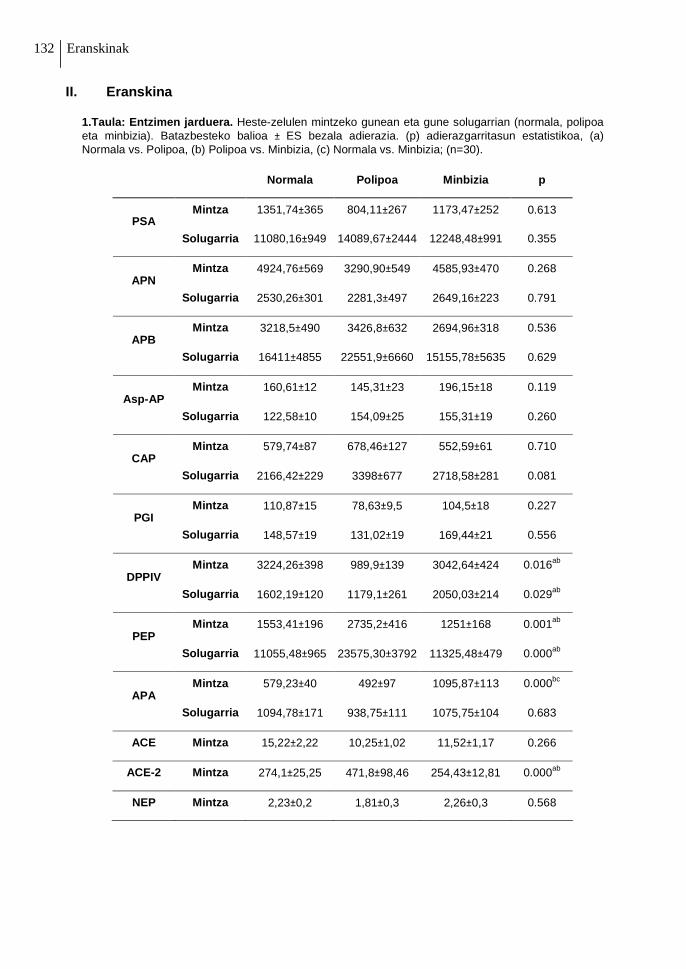
132 Eranskinak
II. Eranskina 1.Taula: Entzimen jarduera. Heste-zelulen mintzeko gunean eta gune solugarrian (normala, polipoa eta minbizia). Batazbesteko balioa ± ES bezala adierazia. (p) adierazgarritasun estatistikoa, (a) Normala vs. Polipoa, (b) Polipoa vs. Minbizia, (c) Normala vs. Minbizia; (n=30).
Normala Polipoa Minbizia p
Mintza 1351,74±365 804,11±267 1173,47±252 0.613 PSA
Solugarria 11080,16±949 14089,67±2444 12248,48±991 0.355
Mintza 4924,76±569 3290,90±549 4585,93±470 0.268 APN
Solugarria 2530,26±301 2281,3±497 2649,16±223 0.791
Mintza 3218,5±490 3426,8±632 2694,96±318 0.536 APB
Solugarria 16411±4855 22551,9±6660 15155,78±5635 0.629
Mintza 160,61±12 145,31±23 196,15±18 0.119 Asp-AP
Solugarria 122,58±10 154,09±25 155,31±19 0.260
Mintza 579,74±87 678,46±127 552,59±61 0.710 CAP
Solugarria 2166,42±229 3398±677 2718,58±281 0.081
Mintza 110,87±15 78,63±9,5 104,5±18 0.227 PGI
Solugarria 148,57±19 131,02±19 169,44±21 0.556
Mintza 3224,26±398 989,9±139 3042,64±424 0.016ab
DPPIV
Solugarria 1602,19±120 1179,1±261 2050,03±214 0.029ab
Mintza 1553,41±196 2735,2±416 1251±168 0.001ab
PEP
Solugarria 11055,48±965 23575,30±3792 11325,48±479 0.000ab
Mintza 579,23±40 492±97 1095,87±113 0.000bc
APA Solugarria 1094,78±171 938,75±111 1075,75±104 0.683
ACE Mintza 15,22±2,22 10,25±1,02 11,52±1,17 0.266
ACE-2 Mintza 274,1±25,25 471,8±98,46 254,43±12,81 0.000ab
NEP Mintza 2,23±0,2 1,81±0,3 2,26±0,3 0.568
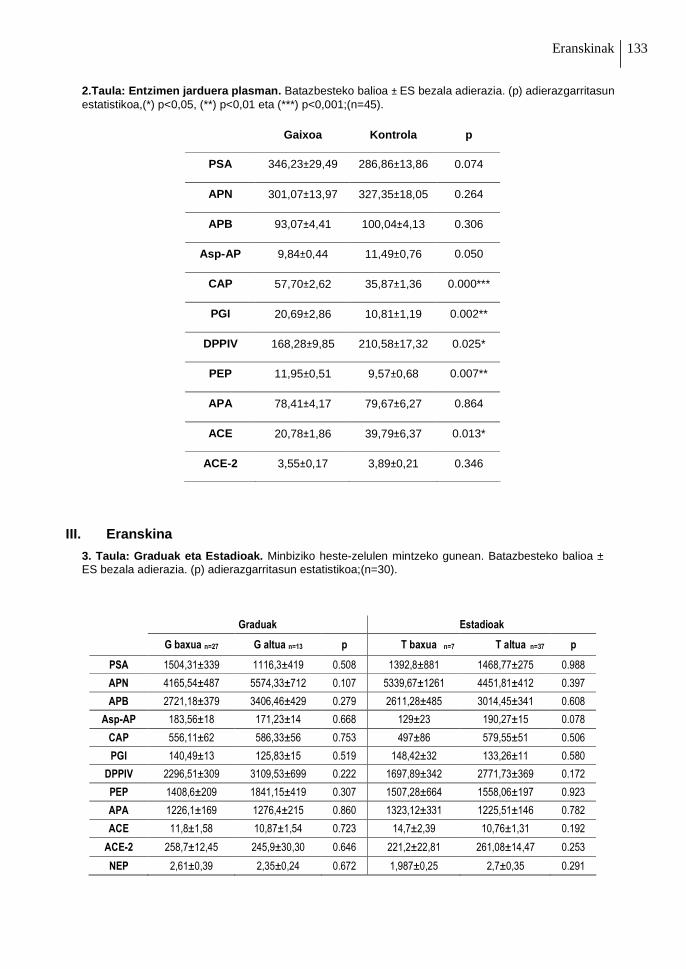
Eranskinak 133
2.Taula: Entzimen jarduera plasman. Batazbesteko balioa ± ES bezala adierazia. (p) adierazgarritasun estatistikoa,(*) p<0,05, (**) p<0,01 eta (***) p<0,001;(n=45).
Gaixoa Kontrola p
PSA 346,23±29,49 286,86±13,86 0.074
APN 301,07±13,97 327,35±18,05 0.264
APB 93,07±4,41 100,04±4,13 0.306
Asp-AP 9,84±0,44 11,49±0,76 0.050
CAP 57,70±2,62 35,87±1,36 0.000***
PGI 20,69±2,86 10,81±1,19 0.002**
DPPIV 168,28±9,85 210,58±17,32 0.025*
PEP 11,95±0,51 9,57±0,68 0.007**
APA 78,41±4,17 79,67±6,27 0.864
ACE 20,78±1,86 39,79±6,37 0.013*
ACE-2 3,55±0,17 3,89±0,21 0.346
III. Eranskina
3. Taula: Graduak eta Estadioak. Minbiziko heste-zelulen mintzeko gunean. Batazbesteko balioa ± ES bezala adierazia. (p) adierazgarritasun estatistikoa;(n=30).
Graduak Estadioak
G baxua n=27 G altua n=13 p T baxua n=7 T altua n=37 p
PSA 1504,31±339 1116,3±419 0.508 1392,8±881 1468,77±275 0.988
APN 4165,54±487 5574,33±712 0.107 5339,67±1261 4451,81±412 0.397
APB 2721,18±379 3406,46±429 0.279 2611,28±485 3014,45±341 0.608
Asp-AP 183,56±18 171,23±14 0.668 129±23 190,27±15 0.078
CAP 556,11±62 586,33±56 0.753 497±86 579,55±51 0.506
PGI 140,49±13 125,83±15 0.519 148,42±32 133,26±11 0.580
DPPIV 2296,51±309 3109,53±699 0.222 1697,89±342 2771,73±369 0.172
PEP 1408,6±209 1841,15±419 0.307 1507,28±664 1558,06±197 0.923
APA 1226,1±169 1276,4±215 0.860 1323,12±331 1225,51±146 0.782
ACE 11,8±1,58 10,87±1,54 0.723 14,7±2,39 10,76±1,31 0.192
ACE-2 258,7±12,45 245,9±30,30 0.646 221,2±22,81 261,08±14,47 0.253
NEP 2,61±0,39 2,35±0,24 0.672 1,987±0,25 2,7±0,35 0.291
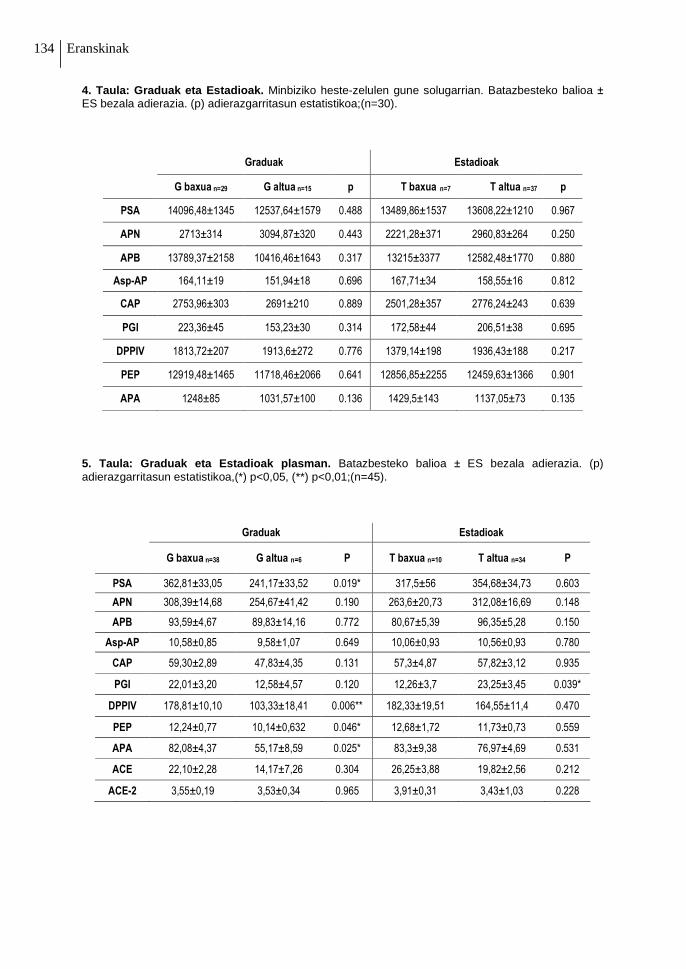
134 Eranskinak
4. Taula: Graduak eta Estadioak. Minbiziko heste-zelulen gune solugarrian. Batazbesteko balioa ±
ES bezala adierazia. (p) adierazgarritasun estatistikoa;(n=30).
Graduak Estadioak
G baxua n=29 G altua n=15 p T baxua n=7 T altua n=37 p
PSA 14096,48±1345 12537,64±1579 0.488 13489,86±1537 13608,22±1210 0.967
APN 2713±314 3094,87±320 0.443 2221,28±371 2960,83±264 0.250
APB 13789,37±2158 10416,46±1643 0.317 13215±3377 12582,48±1770 0.880
Asp-AP 164,11±19 151,94±18 0.696 167,71±34 158,55±16 0.812
CAP 2753,96±303 2691±210 0.889 2501,28±357 2776,24±243 0.639
PGI 223,36±45 153,23±30 0.314 172,58±44 206,51±38 0.695
DPPIV 1813,72±207 1913,6±272 0.776 1379,14±198 1936,43±188 0.217
PEP 12919,48±1465 11718,46±2066 0.641 12856,85±2255 12459,63±1366 0.901
APA 1248±85 1031,57±100 0.136 1429,5±143 1137,05±73 0.135
5. Taula: Graduak eta Estadioak plasman. Batazbesteko balioa ± ES bezala adierazia. (p) adierazgarritasun estatistikoa,(*) p<0,05, (**) p<0,01;(n=45).
Graduak Estadioak
G baxua n=38 G altua n=6 P T baxua n=10 T altua n=34 P
PSA 362,81±33,05 241,17±33,52 0.019* 317,5±56 354,68±34,73 0.603
APN 308,39±14,68 254,67±41,42 0.190 263,6±20,73 312,08±16,69 0.148
APB 93,59±4,67 89,83±14,16 0.772 80,67±5,39 96,35±5,28 0.150
Asp-AP 10,58±0,85 9,58±1,07 0.649 10,06±0,93 10,56±0,93 0.780
CAP 59,30±2,89 47,83±4,35 0.131 57,3±4,87 57,82±3,12 0.935
PGI 22,01±3,20 12,58±4,57 0.120 12,26±3,7 23,25±3,45 0.039*
DPPIV 178,81±10,10 103,33±18,41 0.006** 182,33±19,51 164,55±11,4 0.470
PEP 12,24±0,77 10,14±0,632 0.046* 12,68±1,72 11,73±0,73 0.559
APA 82,08±4,37 55,17±8,59 0.025* 83,3±9,38 76,97±4,69 0.531
ACE 22,10±2,28 14,17±7,26 0.304 26,25±3,88 19,82±2,56 0.212
ACE-2 3,55±0,19 3,53±0,34 0.965 3,91±0,31 3,43±1,03 0.228













