Embed Size (px)
Citation preview

INSTITUTO POLITÉCNICO NACIONAL
UNIDAD PROFESIONAL INTERDISCIPLINARIA DE BIOTECNOLOGÍA
OBTENCIÓN DE FRACCIONES PEPTÍDICAS DE
Phaseolus vulgaris L. Y EVALUACIÓN DE SU
ACTIVIDAD BIOLÓGICA
TESIS
Que para obtener el Grado de:
Doctora en Ciencias en Bioprocesos
Presenta:
M. en C. Teresita de Jesús Ariza Ortega
Directores:
Dr. Jorge Yáñez Fernández
Dra. María del Carmen Oliver Salvador
México D. F., julio 2014

Declaración de originalidad
2
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica

Autorización de uso de obra
3
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica

Acta de revisión de tesis
4
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica

Resumen
5
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Resumen
En México, el cultivo de frijol es el segundo en importancia después del maíz, ya que
además de proveer al ser humano de proteína de alta calidad, se le han atribuido diversas
actividades biológicas que pueden favorecer la salud humana. Por lo que, en este trabajo se
evaluó la actividad biológica in vitro (antioxidante, antimicrobiana e inhibidora de la enzima
convertidora de la angiotensina I) e in vivo (antihipertensiva) de péptidos obtenidos por hidrólisis
enzimática de tres variedades de frijol (Phaseolus vulgaris L.): frijol negro plus, frijol azufrado
higuera y frijol pinto Saltillo.
Los hidrolizados totales de proteínas de frijol presentaron actividad antibacteriana sobre
cepas indicadoras mostrando su capacidad de inhibición en amplio espectro. Los péptidos de la f<1
kDa inhibieron la cepa Shigella dysenteriae con concentración mínima inhibitoria de 10 a 40 µg de
ésta fracción. Y la mayor inhibición del crecimiento bacteriano fue observada con la variedad FAH.
Los hidrolizados totales de frijol mostraron actividad antioxidante, así como sus fracciones
peptídicas; y se observó la tendencia a incrementar ésta actividad biológica al disminuir la masa
molecular de los péptidos. La actividad antioxidante se determinó por cuatro métodos y los
valores máximos obtenidos por métodos espectrofotométricos fueron de: DPPH+ = 40.19 ± 3.83 %
(FNP, f1-3), ABTS+ = 99.17 ± 0.91 % (FNP, f<1) y DMPD+ = 89.08 ± 2.46 % (FPS, f<1); y por el método
de quimioluminiscencia de 94.02 ± 1.09 % (FAH, f1-3).
Todos los hidrolizados totales y las fracciones peptídicas de frijol presentaron actividad
inhibidora de la enzima convertidora de la angiotensina. El mayor porcentaje de actividad fue del
95 % con la fracción f1-3 kDa de las variedades FAH y FPS. Y se comprobó la actividad
antihipertensiva de los péptidos de FAH (f3-10) in vivo y se observó una reducción de la presión
arterial sistólica (27.13 mmHg) de ratas hipertensas similar a la del Captopril (29.67 mmHg) a las 2
h de haberse administrado.
Adicionalmente, se purificaron dos péptidos con actividad antioxidante de FAH (f1-3 kDa),
por espectrometría de masas se determinó su peso molecular (656.219 y 879.242 Da) y por
análisis bioinformático se determinaron las secuencias de aminoácidos posibles.
Las actividades biológicas estudiadas, antimicrobianas, antioxidantes e hipertensivas de las
fracciones peptídicas de hidrolizados de proteína de frijol tienen un considerable potencial en el
desarrollo de alimentos nutracéuticos para el mejoramiento de la salud humana.

Abstract
6
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Abstract
In Mexico, the bean crop is the second in importance after corn because of its quality
protein contain, furthermore, it has recently been studied their human health enhance. Therefore,
in this study the in vitro (antioxidant, antimicrobial and inhibitory of angiotensin converting
enzyme) and in vivo (antihypertensive) biological activities of peptides obtained by enzymatic
hydrolysis of proteins from three common bean (Phaseolus vulgaris L.) varieties (negro plus,
azufrado higuera and pinto Saltillo) were evaluated.
Antimicrobial activity was observed with the total hydrolysate of bean proteins in a wide
broad. The peptidic fraction f<1 kDa inhibited the growth of Shigella dysenteriae with a minimal
inhibitory concentration of 10 to 40 µg of peptide.
Antioxidant activity was observed in the total hydrolysates and peptidic fractions tested. It
was observed that the trend in the antioxidant activity was increased while the group of peptides
decreased in molecular mass. The maximum antioxidant activity observed in spectrophotometric
methods was: DPPH+ = 40.19 ± 3.83 % (FNP, f1-3), ABTS+ = 99.17 ± 0.91 % (FNP, f<1), and DMPD+ =
89.08 ± 2.46 % (FPS, f<1); and by chemiluminescence of 94.02 ± 1.09 % (FAH, f1-3),
Inhibitory angiotensin converting enzyme activity was observed in the total hydrolysates
and peptidic fractions tested. The maximum activity was 95 % with f1-3 kDa from FAH and FPS
beans. Also, the antihypertensive activity was probed with the f3-10 from FAH in vivo over Wistar
spontaneously hypertense rats resulting in a similar systolic pressure values (27.13 mmHg) to
positive control (Captopril, 29.67 mmHg) at 2 h of administration.
The molecular mass of two isolated peptides with antioxidant activity from FAH (f1-3 kDa)
was determined (656.219 y 879.242 Da) by mass spectrometry, and the possible amino acid
sequences were determined by a bioinformatics analysis.
The biological activities studied, antimicrobial, antioxidant, and antihypertensive of the
peptidic fractions from common bean hydrolysates (from the three bean varieties of this work)
support their potential use in the pharmaceutics and food fields to develop a nutraceutic product.

Apoyos para la realización de éste trabajo
7
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Apoyos para la realización de éste trabajo
Este trabajo fue realizado en los laboratorios de Biotecnología Alimentaria y Biología
Molecular de la sección de estudios de posgrado e investigación de la Unidad Profesional
Interdisciplinaria de Biotecnología del Instituto Politécnico Nacional.
El programa de posgrado de Doctorado en Ciencias en
Bioprocesos de la Unidad Profesional Interdisciplinaria de
Biotecnología del Instituto Politécnico Nacional está
incluido en el Padrón Nacional de Posgrado con el número
de referencia 000653.
La presente Tesis fue realizada con el apoyo de la Beca
350021 del Consejo Nacional de Ciencia y Tecnología en el
periodo 2012-2013.
El Instituto Politécnico Nacional a través de la Secretaría de
Investigación y Posgrado y por conducto de la Dirección de
Posgrado, otorgo apoyo por parte del Programa de Beca
Institucional de Posgrado en el periodo 2010-2012 y 2013-
2014.

Índice
8
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Índice general
Resumen…………………………………………………………………………… ..............................................................5
Abstract……………………………………………………………………………………………………………………………………...….6
Índice...................................................................................................................................................8
1. Introducción ............................................................................................................................16
1.1 Particularidades e importancia del frijol ...........................................................................16
1.1.1 El frijol común (Phaseolus vulgaris L.).......................................................................16
1.1.2 El cultivo de frijol en México .....................................................................................17
1.1.3 Importancia de las semillas de frijol en la alimentación ...........................................18
1.2 Las proteínas y su función en el organismo ......................................................................19
1.2.1 Concepto general de proteínas ................................................................................19
1.2.2 Importancia de las proteínas ....................................................................................19
1.2.3 Proteínas de origen animal .......................................................................................20
1.2.4 Proteínas de origen vegetal ......................................................................................20
1.3 Proceso de hidrólisis enzimática .......................................................................................21
1.3.1 Hidrólisis de proteínas ..............................................................................................21
1.3.2 Importancia de los hidrolizados de proteína ............................................................22
1.3.3 Enzimas proteolíticas ................................................................................................22
1.4 Actividad biológica y funcionalidad de péptidos ..............................................................24
1.4.1 Alimentos funcionales ..............................................................................................24
1.4.2 Péptidos con actividad biológica ..............................................................................25
1.5 Aplicación y función de péptidos en la salud ....................................................................26
1.5.1 Problemática actual y riesgos en la salud .................................................................26
1.5.2 Actividad antibacteriana como control de patógenos ..............................................26
1.5.3 Actividad antioxidante como control de la oxidación celular ...................................28
1.5.4 Actividad inhibidora de la enzima convertidora de la angiotensina como control de
la hipertensión ..........................................................................................................................30
1.6 Péptidos identificados como biológicamente activos .......................................................32
2. Justificación .............................................................................................................................35
3. Objetivos .................................................................................................................................36

Índice
9
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
3.1 Objetivo general ...............................................................................................................36
3.2 Objetivos particulares.......................................................................................................36
4. Materiales y Métodos ..............................................................................................................37
4.1 Materiales biológicos y reactivos .....................................................................................37
4.2 Caracterización física de las semillas de frijol ...................................................................37
4.3 Preparación de las harinas de semillas de frijol ................................................................38
4.4 Caracterización químico proximal de las harinas de semillas de frijol ..............................38
4.4.1 Determinación de humedad .....................................................................................39
4.4.2 Determinación de proteína total ..............................................................................39
4.4.3 Determinación del extracto etéreo ..........................................................................40
4.4.4 Determinación de fibra cruda ...................................................................................40
4.4.5 Determinación de cenizas totales .............................................................................41
4.4.6 Carbohidratos totales ...............................................................................................41
4.5 Proceso de desengrasado de las harinas de frijol .............................................................41
4.6 Obtención de los concentrados proteicos de frijol ...........................................................42
4.7 Caracterización de los concentrados proteicos de frijol ...................................................42
4.7.1 Contenido total de proteína en los concentrados de frijol .......................................42
4.7.2 Determinación de proteína soluble en los concentrados proteicos de frijol ............42
4.7.3 Perfil proteico de los concentrados de proteína de frijol .........................................43
4.7.4 Electroforesis bidimensional de los concentrados de proteína de frijol ...................44
4.8 Hidrólisis enzimática de las proteínas de frijol .................................................................44
4.9 Fraccionamiento de los hidrolizados proteicos por ultrafiltración ...................................45
4.10 Caracterización del fraccionamiento por ultrafiltración ...................................................46
4.11 Determinación de la actividad antibacteriana de los hidrolizados totales y fracciones
peptídicas de frijol .......................................................................................................................46
4.11.1 Prueba de susceptibilidad microbiana ......................................................................46
4.11.2 Determinación de la concentración mínima inhibitoria ...........................................47
4.12 Determinación de la actividad antioxidante de los hidrolizados totales y fracciones
peptídicas de frijol .......................................................................................................................48
4.12.1 Actividad antioxidante como captura del radical libre DPPH+ ..................................48
4.12.2 Actividad antioxidante en función de la decoloración del radical libre ABTS+ ..........49

Índice
10
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.12.3 Actividad antioxidante en función de la decoloración del radical libre DMPD+ ........50
4.12.4 Actividad antioxidante determinada por el método de quimioluminiscencia ..........51
4.13 Determinación de la actividad inhibidora de la enzima convertidora de la angiotensina 52
4.14 Purificación de las fracciones peptídicas con actividad antioxidante ...............................54
4.15 Determinación de la masa molecular del péptido con actividad antioxidante .................54
4.16 Determinación de la secuencia de aminoácidos por bioinformática del péptido con
actividad antioxidante .................................................................................................................54
4.17 Análisis estadístico de datos .............................................................................................55
5. Resultados y Discusión ............................................................................................................56
5.1 Evaluación y caracterización física de las semillas de frijol ...............................................56
5.2 Obtención de las harinas de frijol .....................................................................................57
5.3 Caracterización químico proximal de las harinas de frijol ................................................58
5.4 Obtención del concentrado de proteínas de frijol ............................................................60
5.5 Comparación de los perfiles electroforéticos de las harinas de frijol ...............................61
5.5.1 Electroforesis de los concentrados de proteína de frijol ..........................................61
5.5.2 Electroforesis bidimensional de los concentrados de proteína de frijol ...................62
5.6 Hidrólisis enzimática de las proteínas de frijol .................................................................65
5.6.1 Hidrólisis enzimática de las proteínas contenidas en las harinas de frijol ................65
5.6.2 Hidrólisis enzimática de los concentrados proteicos de frijol ...................................66
5.7 Fraccionamiento de péptidos por ultrafiltración ..............................................................67
5.8 Determinación de la actividad antibacteriana ..................................................................69
5.8.1 Controles positivos - Resistencia de las cepas indicadoras a los antibióticos ...........69
5.8.2 Actividad antibacteriana en harinas de frijol hidrolizadas ........................................70
5.8.3 Actividad antibacteriana en harinas desengrasadas e hidrolizadas de frijol .............70
5.8.4 Actividad antibacteriana de los hidrolizados totales obtenidos a partir de la proteína
concentrada de frijol ................................................................................................................71
5.8.5 Actividad antibacteriana de las fracciones peptídicas de los hidrolizados de los
concentrados proteicos de frijol ...............................................................................................72
5.8.6 Determinación de la concentración mínima inhibitoria de las fracciones peptídicas e
hidrolizados de frijol .................................................................................................................73
5.9 Determinación de la actividad antioxidante .....................................................................75

Índice
11
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.9.1 Método de captación del radical libre DPPH+ ...........................................................75
5.9.2 Método de captación del radical libre ABTS+ ............................................................78
5.9.3 Método de captación del radical libre DMPD+ ..........................................................80
5.9.4 Método de quimioluminiscencia ..............................................................................82
5.9.5 Comparación de métodos de determinación de la actividad antioxidante ..............84
5.10 Determinación de la actividad inhibidora de la enzima convertidora de la angiotensina 85
5.11 Purificación de las fracciones con actividad .....................................................................88
5.11.1 Separación de péptidos por grupos y determinación de su actividad antioxidante .88
5.11.2 Separación de péptidos individualmente y determinación de su actividad
antioxidante .............................................................................................................................90
5.11.3 Comparativo de la actividad antioxidante de péptidos purificados ..........................92
5.12 Identificación del péptido con actividad antioxidante ......................................................93
5.12.1 Determinación del peso molecular del péptido ........................................................93
5.12.2 Identificación molecular del péptido ........................................................................95
6. Conclusiones ............................................................................................................................99
7. Perspectivas ...........................................................................................................................100
8. Referencias ............................................................................................................................101
9. Anexos ...................................................................................................................................111
9.1 Productos derivados de ésta Tesis..................................................................................111
9.2 Antioxidantes de referencia utilizados como control .....................................................113
9.3 Lista de aminoácidos ......................................................................................................114

Índice
12
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Índice de Cuadros
Cuadro 1-1. Proteasas comerciales grado alimenticio. ....................................................................23
Cuadro 1-2. Péptidos bioactivos obtenidos de proteínas animales. .................................................33
Cuadro 1-3. Péptidos bioactivos sintetizados químicamente. ..........................................................33
Cuadro 1-4. Péptidos bioactivos obtenidos de proteínas vegetales. ................................................34
Cuadro 5-1. Características físicas de las semillas de las tres variedades de frijol............................56
Cuadro 5-2. Masas y rendimientos de las harinas de frijol después de la molienda y tamizado. .....57
Cuadro 5-3. Análisis químico proximal de las harinas de frijol (%) ...................................................58
Cuadro 5-4. Composición químico proximal en harinas de frijol reportadas previamente (%) ........59
Cuadro 5-5. Contenido de proteína en los concentrados de frijol (%)..............................................60
Cuadro 5-6. Grado de hidrólisis alcanzado por las harinas de frijol durante el tratamiento (%) ......65
Cuadro 5-7. Grado de hidrólisis obtenido en 2 h de reacción con los concentrados de proteína de
frijol (%) ...................................................................................................................................66
Cuadro 5-8. Determinación de la resistencia de las cepas a los antibióticos de referencia (control
positivo). ..................................................................................................................................69
Cuadro 5-9. Actividad antibacteriana del hidrolizado total de los concentrados proteicos de frijol 71
Cuadro 5-10. Actividad antibacteriana de las fracciones del hidrolizado del concentrado de FAH ..73
Cuadro 5-11. CMI determinada para las fracciones de hidrolizados de frijol. ..................................74
Cuadro 5-12. Actividad antioxidante de péptidos de frijol determinada por captación de DPPH+ (%)
.................................................................................................................................................76
Cuadro 5-13. Equivalentes del antioxidante de referencia con respecto de las fracciones de
péptidos de frijol determinada por captación del radical libre DPPH+ .....................................77
Cuadro 5-14. Actividad antioxidante de péptidos de frijol determinada por decoloración del ABTS+
(%) ...........................................................................................................................................78
Cuadro 5-15. Equivalentes del antioxidante de referencia con respecto de las fracciones de
péptidos de frijol determinada por captación del radical libre ABTS+......................................79
Cuadro 5-16. Actividad antioxidante de péptidos de frijol determinada por decoloración del
DMPD+ (%) ...............................................................................................................................80
Cuadro 5-17. Equivalentes del antioxidante de referencia con respecto de las fracciones de
péptidos de frijol determinada por captación del radical libre DMPD+ ...................................81

Índice
13
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Cuadro 5-18. Actividad quimioluminiscente de los controles positivos y blancos de control. .........82
Cuadro 5-19. Actividad antioxidante determinada por quimioluminiscencia de péptidos de frijol
(%) ...........................................................................................................................................83
Cuadro 5-20. Inhibición de la ECA-I por los péptidos de frijol de 1-3 kDa (%) ..................................85
Cuadro 5-21. Efecto en la presión sanguínea (PS) de la administración de la F3-10 del FAH sobre
RWH.........................................................................................................................................87
Cuadro 5-22. Actividad antioxidante (%) de los péptidos individuales colectados en la separación
individual por HPLC de los péptidos de los grupos 40A y 45A del FAH. ...................................91
Cuadro 5-23. Análisis de aminoácidos realizado en el sistema de predicción virtual ExPASY
(UniProt: P02853). ...................................................................................................................95
Cuadro 5-24. Análisis de aminoácidos realizado en el sistema de predicción virtual ExPASY
(UniProt: Q8RVH1). ..................................................................................................................96

Índice
14
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Índice de Figuras
Figura 1-1. Cultivo y vaina del frijol común (Phaseolus vulgaris L.). .................................................16
Figura 1-2. Variedades de frijol utilizadas en este trabajo. ..............................................................18
Figura 1-3. Representación del mecanismo de acción de los péptidos antibacterianos del tipo alfa
hélice. ......................................................................................................................................28
Figura 1-4. Mecanismo de acción de la inhibición de la enzima convertidora de la angiotensina I
(modificado de Keidar y col., 2007). ........................................................................................31
Figura 4-1. Harinas obtenidas de las tres variedades de frijol. .........................................................38
Figura 4-2. Caracterización químico proximal de las harinas de frijol según métodos de la AOAC. .39
Figura 4-3. Equipo de (a) isoelectroenfoque y (b) electroforesis en gel de poliacrilamida dodecil
sulfato de sodio de las proteínas de frijol. ...............................................................................43
Figura 4-4. Montaje experimental utilizado durante el proceso de hidrólisis enzimática. ...............44
Figura 4-5. Determinación de la actividad antibacteriana de los hidrolizados totales y fracciones
peptídicas. ...............................................................................................................................47
Figura 4-6. Estructura química (a) del radical libre difenilpicrilhidrazilo y (b) del compuesto estable
difenilpicrilhidrazina (Molyneux, 2004). ..................................................................................48
Figura 4-7. Reacción de formación del radical libre ABTS+. ..............................................................49
Figura 4-8. Reacción de decoloración del radical libre DMPD+ (Chagas y col., 2009). ......................50
Figura 4-9. Reacción quimioluminiscente llevada a cabo por el radical libre 3-aminoptalatohidrazilo
(Douglas y Garley, 2010). .........................................................................................................51
Figura 4-10. Montaje experimental del equipo de quimioluminiscencia. ........................................52
Figura 5-1. Semilla de frijol de las tres variedades estudiadas en este trabajo. ...............................56
Figura 5-2. SDS-PAGE del concentrado de proteínas de frijol de las variedades FNP, FAH y FPS en
condiciones no reductoras y reductoras. Gel al 15 % de acrilamida con 5 µg de proteína por
pozo. ........................................................................................................................................61
Figura 5-3. Electroforesis bidimensional en gel de poliacrilamida y dodecil-sulfato de sodio del
concentrado de proteínas de frijol. .........................................................................................63
Figura 5-4. SDS-PAGE al 15 % de los concentrados proteicos y los hidrolizados totales de proteínas
de frijol. ...................................................................................................................................67

Índice
15
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-5. SDS-PAGE al 18%, de los péptidos obtenidos de la ultrafiltración de los hidrolizados de
frijol a) negro plus, b) azufrado higuera y c) pinto Saltillo. ......................................................68
Figura 5-6. Comparación de la actividad antioxidante de los hidrolizados totales y las fracciones
peptídicas de hidrolizados de proteína de frijol determinada por los cuatro métodos. ..........84
Figura 5-7. Inhibición de la enzima convertidora de la angiotensina con el hidrolizado total y las
fracciones peptídicas de proteínas de frijol. ............................................................................86
Figura 5-8. Cromatografía de liquídos de alta eficiencia (HPLC) del perfil de separación de péptidos
la fracción f1-3 de: a) FNP, b) FAH y c) FPS. .............................................................................88
Figura 5-9. Actividad antioxidante de los trece grupos de péptidos colectados por HPLC del perfil
de separación de péptidos la fracción f1-3 de: a) FNP, b) FAH y c) FPS; determinada por el
método de quimioluminiscencia. ............................................................................................89
Figura 5-10. Cromatografía de la separación individual de péptidos de los grupos a) 40A y b) 45A
del FAH. ...................................................................................................................................91
Figura 5-11. Comparativo de la actividad antioxidante de la fracciones (FAH f<3), grupos de
péptidos (40A y 45A), picos (4A y 10A) y picos puros colectados por HPLC.............................92
Figura 5-12. Resultados del análisis de espectrometría de masas de la muestra 4A. .......................93
Figura 5-13. Resultados del análisis de espectrometría de masas de la muestra 10A. .....................94
Figura 5-14. Posición de péptidos con actividad antioxidante en la faseolina. ................................96
Figura 5-15. Posición de péptidos con actividad antioxidante en la lectina. ....................................97

Introducción
16
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
1. Introducción
1.1 Particularidades e importancia del frijol
1.1.1 El frijol común (Phaseolus vulgaris L.)
El frijol es una especie perteneciente a las leguminosas que es cultivada y consumida como
parte de la dieta del ser humano por su alto contenido de proteínas (Boye y col., 2010)
principalmente en países en desarrollo como América latina (Camacho-Espinoza y col., 2010), y
algunas regiones de Europa, Asia y África (Makri y Doxastakis, 2006).
En México los estados más ricos en especies de esta familia son Oaxaca y Chiapas (Lutz y Edel-
León, 2009). Además, en nuestro país se han encontrado vestigios de domesticación de cinco
especies de leguminosas comestibles: Phaseolus vulgaris (frijol común), P. coccineus (ayocote), P.
acutifolius (alubia), Phaseolus lunatus (lima o furuna) y P. polyanthus (botil) (Fraile y col., 2007). En su
composición se encuentran grupos importantes de aminoácidos, aunque su valor nutricional es
limitado por su deficiencia en aminoácidos sulfurados (metionina y cisteína) y triptófano (Carrasco-
Castilla y col., 2012), en la dieta de los mexicanos dicha carencia es compensada con el consumo de
maíz (Barrón-Contreras, 2006).
La familia Leguminosae comprende alrededor de 650 géneros y 18,000 especies en el mundo,
ésta es una de las seis familias de angiospermas más diversas que existen (Fraile y col., 2007). El frijol
es una especie perteneciente a las leguminosas y su información taxonómica se presenta a
continuación (SIOVM, 2011). Reino: Plantae, División: Magnoliophyta, Clase: Magnoliopsida, Orden:
Fabales, Familia: Fabaceae, Género: Phaseolus L. y Especie: P. vulgaris L.
Figura 1-1. Cultivo y vaina del frijol común (Phaseolus vulgaris L.).

Introducción
17
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
El fruto de las leguminosas es la legumbre, nombre con que se designan las semillas
encerradas en una vaina (Figura 1-1), también nos encontramos con lomentos, es una falsa legumbre,
los frutos de las leguminosas pratenses tienen muy diferentes formas, siendo los más característicos
las legumbres esféricas, aovadas, alargadas o espiraladas, recubiertas de costillas, espinas, verrugas,
articulados (White y González, 1990).
1.1.2 El cultivo de frijol en México
En México los cuatro cultivos principales por superficie sembrada son: maíz, frijol, sorgo y trigo.
Después del maíz (52 %), el 14 % de la superficie de total de cultivo es utilizada para la siembra de las
cinco especies ancestralmente domesticadas de frijol, el 95 % es para el cultivo de frijol, debido a que
en éste existe una amplia variación de color, tamaño, forma de grano, así como hábito de crecimiento
y precocida, en intervalo de adaptación y potencial de producción, en calidad comercial y nutritiva
(SIAP, 2013).
Ya que el frijol ha sido cultivado a nivel mundial desde épocas ancestrales, éste ha tomado gran
importancia en varios países; en 2001 Nigeria fue el principal productor de fríjol en el mundo, desde
2002 hasta 2009 Brasil ha ocupado ese lugar, y de manera invariable México ha ocupado el quinto
lugar en este grupo de productores (FAO, 2013). En nuestro país los estados productores de frijol son:
Zacatecas, Sinaloa, Durango, Chihuahua y Nayarit (COVECA, 2012). India es el principal país
importador de frijol seguido de Brasil y Estados Unidos, México ocupa el octavo lugar (FAO, 2013).
En México después del maíz, el frijol es la semilla a la que más familias recurren para
complementar su alimentación. El consumo per cápita de frijol en México de 1994 a 2005 oscila entre
10 y 15 kg anuales, curiosamente el incremento en la producción anual coincide con el incremento en
el consumo (SIAP, 2013). Por lo que, se ha reportado que la producción nacional no es suficiente para
cubrir la demanda por lo que se ha recurrido a importar de países como Estados Unidos y Canadá (96
% y 4 % de las importaciones, respectivamente). Durante este periodo las variedades importadas
fueron en su mayoría de frijol negra, seguidas de las variedades de frijol pinto y otras variedades con
testa de colores claros (Figura 1-2).
Este asunto representa un problema, ya que depender de las importaciones significa perder
autosuficiencia alimentaria, situación que no es recomendable ya que más de 100 millones de
mexicanos consumen frijol (Borja-Bravo y García-Salazar, 2008).

Introducción
18
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 1-2. Variedades de frijol utilizadas en este trabajo.
a) negro plus, b) azufrado higuera y c) pinto Saltillo.
1.1.3 Importancia de las semillas de frijol en la alimentación
La mayoría de las leguminosas han sido explotadas a gran escala lo son por sus semillas, varias
de las cuales han sido alimentos básicos de la dieta desde tiempos ancestrales en ciertas regiones del
mundo, como es el caso del frijol común, en el sur de México y Centroamérica (Drago-Serrano y col.,
2006). Varios frijoles (género Phaseolus) se domesticaron desde épocas muy antiguas y dieron pie al
desarrollo de civilizaciones en nuestro país y otros lugares de América. Son desde entonces las
leguminosas base en la dieta mexicana, especialmente el frijol común, que fue importante
componente de la dieta en Mesoamérica y hoy es, sólo después del maíz, la planta más cultivada en
México e importante fuente de proteínas, sobre todo para las poblaciones de bajos recursos (Borja-
Bravo y García-Salazar, 2008).
El frijol es probablemente la leguminosa más importante en Latinoamérica y África, ya que en
estas zonas geográficas las proteínas de origen animal no son ampliamente consumidas y por lo
regular son sustituidas con frijoles para satisfacer los requerimientos proteínicos (Delgado-Sánchez y
col., 2006), se considera además que el frijol común es una fuente importante de aminoácidos ya que
contiene 19 de ellos en variadas proporciones.
La composición de frijol varía de acuerdo con las condiciones de cultivo y la localidad de
siembra, pero el perfil de antocianinas quienes determinan el color del frijol se conserva. México,
como parte de Mesoamérica, es considerado uno de los centros de origen más importantes de varios
tipos de frijol, entre los que destaca por su valor comercial Phaseolus vulgaris L., especie donde se
encuentra el frijol negro que es el de mayor consumo nacional (Sánchez y col., 2001).
a) b) c)

Introducción
19
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
1.2 Las proteínas y su función en el organismo
1.2.1 Concepto general de proteínas
Las proteínas son moléculas formadas por cadenas lineales de aminoácidos. El término
proteína significa “de primera calidad”, debido a su importancia estructural y nutricional en los seres
vivos. Las proteínas pueden ser clasificadas de diferentes formas; por sus propiedades físico-químicas
las proteínas se clasifican en: proteínas simples, que por hidrólisis dan solo aminoácidos o sus
derivados; proteínas conjugadas, que por hidrólisis dan aminoácidos acompañados de sustancias
diversas, y proteínas derivadas, sustancias formadas por desnaturalización y desdoblamiento de las
anteriores.
1.2.2 Importancia de las proteínas
Las proteínas desempeñan un papel fundamental para la vida y son las biomoléculas más
versátiles y diversas. Son imprescindibles para el crecimiento del organismo y realizan una enorme
cantidad de funciones diferentes.
Las proteínas son necesarias para la vida, sobre todo por su función plástica, pero también por
sus funciones biorreguladoras y de defensa. Las proteínas son el principal componente estructural y
funcional de las células y tienen numerosas e importantes funciones dentro del organismo que van
desde su papel catalítico (enzimas) hasta su función en la motilidad corporal (actina, miosina),
pasando por su papel mecánico (elastina, colágeno), de transporte y almacén (hemoglobina,
mioglobina, citocromos), protección (anticuerpos) y reguladora (hormonas), (Sánchez-Contreras,
2007). Su característica más importante es que contienen nitrógeno, siendo el contenido medio de
este elemento de un 16 %. Las proteínas son macromoléculas formadas por cadenas de unidades
estructurales, los aminoácidos. Estos aminoácidos se unen por medio de enlaces peptídicos entre los
grupos carboxilo y el grupo α-amino (imino), con pérdida de agua. La secuencia de aminoácidos que
componen una proteína constituye su estructura primaria, de vital importancia desde el punto de
vista nutricional. También tienen importancia nutricional, aunque en menor medida, la estructura
secundaria y terciaria (Sánchez-Contreras, 2007).
Las proteínas se clasifican atendiendo a distintos puntos de vista como son: solubilidad,
composición, forma, propiedades físicas, función y estructura tridimensional. La proteína supone
aproximadamente el 17 % de la masa corporal de un individuo. A pesar de su diversidad funcional

Introducción
20
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
(enzimática, de transporte y almacén, mecánica, motilidad, protección y reguladora) un 25 % es
proteína estructural y hemoglobina (Martínez-Agustín y Martínez de Victoria-Muñoz, 2006).
Las proteínas están conformadas por cadenas mayores a 70 aminoácidos, éstos son
fundamentales para el buen funcionamiento del organismo. Para una persona adulta son ocho los
aminoácidos esenciales (fenilalanina, leucina, isoleucina, lisina, metionina, treonina, triptófano y
valina), mientras que durante el crecimiento se precisan dos más (arginina e histidina, durante la
infancia y adolescencia); y los aminoácidos no esenciales (alanina, cisteína, cistina, glicina,
hidroxiprolina, prolina, serina, tirosina, ácido aspártico, y glutámico), (Sánchez-Contreras, 2007).
1.2.3 Proteínas de origen animal
Las proteínas de origen animal poseen un alto valor biológico desde el punto de vista
nutricional, ya que de ellas se obtiene la gran mayoría de nitrógeno y aminoácidos necesarios para el
buen funcionamiento del organismo humano. Se han realizado estudios en los cuales se ha
determinado que los alimentos de origen animal no presentaron aminoácidos limitantes y las cifras
de digestibilidad y disponibilidad se encontraron en un intervalo que varió de 94 % en las carnes a 97
% en el huevo, situación que no es posible observar en las proteínas de fuentes vegetales. La soya, el
garbanzo, el pistacho y la remolacha son los únicos alimentos vegetales que no contienen
aminoácidos limitantes, sin embargo, presentaron un intervalo del 48 % al 78 % de digestibilidad y
disponibilidad (Suárez-López y col., 2006). Algunas fuentes de proteínas animales son: huevo, leche y
sus derivados, hemoglobina, todo tipo de carne, y músculo de productos marinos (Vrese y
Schrezenmeir, 2008).
1.2.4 Proteínas de origen vegetal
La búsqueda de fuentes alternativas de proteínas se ha convertido, a lo largo de las últimas
décadas, en una importante tendencia de investigación, no solo para encarar la creciente demanda
de proteínas sino también para conseguir cultivos alternativos que sustituyan a las proteínas de
origen animal, capaces de suplementar proteína de alta calidad y prevenir la malnutrición. Las
proteínas vegetales en general, gozan de aceptación mundial y son valoradas por sus atributos
sensoriales y nutricionales (Montaño-Pérez y Vargas-Albores, 2002). Algunas fuentes de proteínas
vegetales son: la semillas de girasol, lupinos, frijol, haba, alverjón, garbanzo, chícharos y otras semillas
(Vrese y Schrezenmeir, 2008).

Introducción
21
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
En su forma natural las proteínas vegetales no tienen las mismas propiedades que después de
un proceso de fraccionamiento, es por ello que la hidrólisis de proteínas de fuentes alimenticias tiene
gran importancia porque permite explicar la contribución de cada fracción a la calidad nutricional y
sus propiedades funcionales. La calidad de una proteína es importante, seguida de la digestibilidad,
pues no todas las proteínas son digeridas, absorbidas y utilizadas en la misma medida por el
organismo humano (Padilla y col., 2010; Möller y col., 2008).
Una ventaja adicional al consumo de proteínas de origen vegetal es que se ha reportado que la
inclusión de leguminosas en la dieta permite la prevención de varias enfermedades metabólicas como
diabetes mellitus, enfermedades coronarias cardiacas y cáncer de colón, debido a su importante
contenido de fibra, fenoles y otros compuestos como son los péptidos derivados del proceso de
hidrólisis (Siddiq y col., 2010).
1.3 Proceso de hidrólisis enzimática
1.3.1 Hidrólisis de proteínas
La hidrólisis proteica se realiza para cortar la proteína original en fragmentos y modificar sus
funciones. Normalmente su proceso se lleva a cabo en un reactor, con control de agitación, pH,
temperatura y tiempo del proceso. El sustrato se disuelve en agua hasta que el pH y la temperatura
se estabilizan; a continuación se agrega la proteasa dando inicio a la hidrólisis. A medida que ésta
progresa se produce una disminución del pH debido a la rotura de los enlaces peptídicos. En los casos
de hidrólisis enzimática el pH debe ser mantenido en el óptimo de la enzima mediante la adición de
base diluida. Para finalizar la hidrólisis proteica la enzima puede ser inactivada con calor, mediante
una disminución del pH o con una combinación de ambos. O también puede ser retirada del medio
mediante filtración y la proteína finalmente precipitada (Alvarado-Carrasco y Guerra, 2010).
En un proceso de hidrólisis se deben tomar en consideración varios factores como: las
características del reactor, volumen de operación, control de agitación, pH, temperatura, tiempo de
reacción, tipo y concentración de sustrato, especificidad y concentración de la enzima, que son
importantes para la obtención de hidrolizados con características específicas de grado de hidrólisis,
tamaño de fracciones de proteína, aminoácido donde se ubica el sitio de acción y otros factores
(Benítez y col., 2008).

Introducción
22
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
1.3.2 Importancia de los hidrolizados de proteína
El término propiedades funcionales se aplica a los ingredientes alimenticios y se define como
toda propiedad no nutricional que influencia la utilidad de un ingrediente en un alimento. La mayor
parte de las propiedades funcionales influyen sobre el carácter sensorial del alimento, pero también
pueden tener un papel decisivo en el comportamiento físico de los alimentos o de los ingredientes
alimenticios durante su preparación, transformación o almacenamiento (Belén-Camacho y col.,
2007); desde el punto de vista técnico-funcional de las proteínas, estas propiedades tienden a
mejorar al ser sometidas a un proceso de hidrólisis (Acuña-Goycolea, 2001). La hidrólisis proteica se
realiza para modificar las funciones propias de la proteína. En los hidrolizados de proteína se
potencian diversas características funcionales, tales como viscosidad baja, mayor capacidad de
agitación, dispersión y alta solubilidad, que les conceden ventajas para el uso en muchos productos
alimenticios, respecto a las proteínas originales (Chung y col., 2008; Guadix y col., 2000).
Recientemente se ha incrementado el interés del estudio de péptidos debido a la potencial
actividad biológica (como: antihipertensivos, antibacterianos, anticariogénicos, antiulceratívos,
anticarcinogénicos, anticoagulantes, antitrombóticos y antioxidantes) que han mostrado diversos
fragmentos proteicos obtenidos vía hidrólisis enzimática con variación de tipo de enzima y fuente de
obtención. Dichos péptidos se han obtenido de proteínas animales como: leche, huevo, plasma
sanguíneo, músculo de pescado; o fuentes vegetales como la soya, garbanzo, girasol, colza, lupino,
amaranto y otras semillas (Torruco-Uco y col., 2009). En el caso del presente trabajo se utilizó una
fuente de proteína vegetal, el frijol común, quien contiene entre 13 - 27 % de proteína (Zubirán,
1996) y que puede llegar a concentrarse hasta el 80 % según los reportes previos (Hernández-Álvarez
y col., 2010).
1.3.3 Enzimas proteolíticas
Actualmente se encuentran disponibles comercialmente muchas proteasas grado alimenticio.
Estas proteasas pueden ser clasificadas, por su origen, (animal, vegetal, bacteriano o fúngico), por su
modo de acción catalítica (endo- o exo-actividad) o con base en su sitio catalítico. Las endo-proteasas
hidrolizan enlaces amídicos dentro de la cadena de la proteína. Las exoproteasas, por el contrario,
eliminan aminoácidos terminales de las proteínas o péptidos (Villanueva y col., 1999). La naturaleza
del centro catalítico de las proteasas difiere de acuerdo con los aminoácidos y otros ligandos que
intervienen en la formación del complejo enzima-sustrato. El centro activo contiene aminoácidos o
Introducción
23
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
bien cationes metálicos que promueven la catálisis, denominándose proteasas serinicas, proteasas
cisteínicas, proteasas aspárticas, según intervengan los aminoácidos serina, cisteína o ácido aspártico
(Benítez y col., 2008).
Las primeras enzimas proteolíticas utilizadas en la industria alimentaria fueron proteasas
pancreáticas de origen animal, si bien cada vez están adquiriendo mayor importancia las de origen
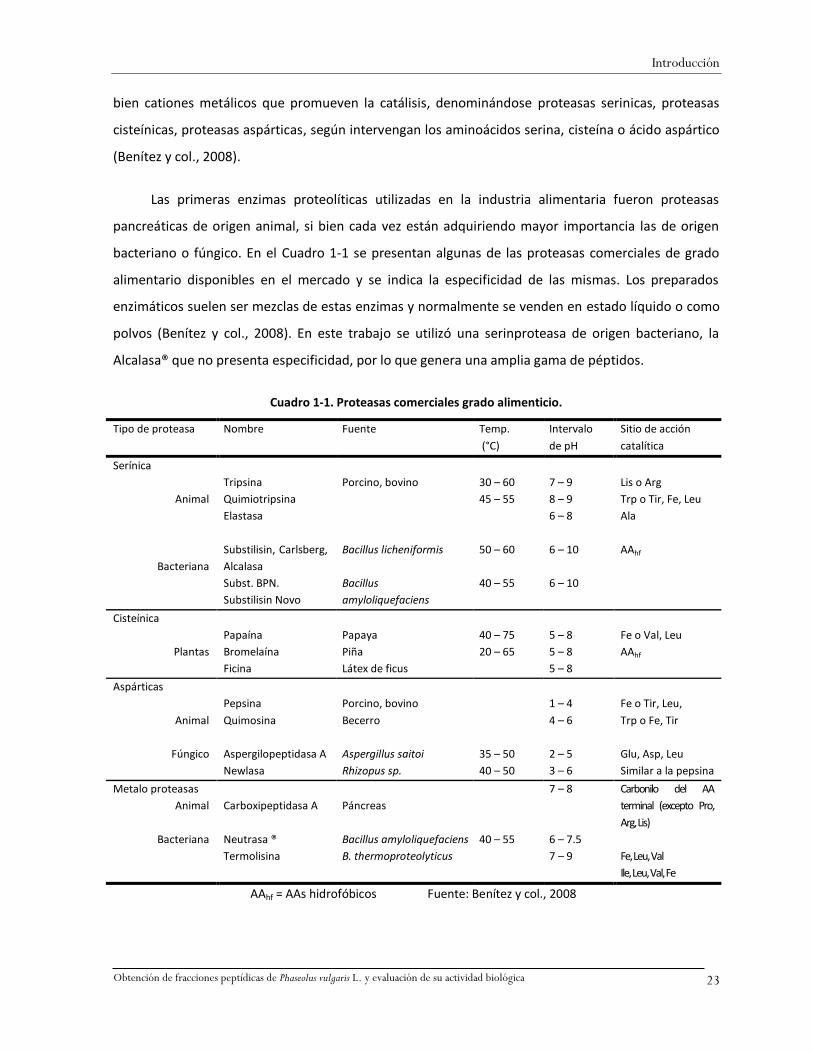
bacteriano o fúngico. En el Cuadro 1-1 se presentan algunas de las proteasas comerciales de grado
alimentario disponibles en el mercado y se indica la especificidad de las mismas. Los preparados
enzimáticos suelen ser mezclas de estas enzimas y normalmente se venden en estado líquido o como
polvos (Benítez y col., 2008). En este trabajo se utilizó una serinproteasa de origen bacteriano, la
Alcalasa® que no presenta especificidad, por lo que genera una amplia gama de péptidos.
Cuadro 1-1. Proteasas comerciales grado alimenticio.
Tipo de proteasa Nombre Fuente Temp.
(°C)
Intervalo
de pH
Sitio de acción
catalítica
Serínica
Animal
Bacteriana
Tripsina
Quimiotripsina
Elastasa
Substilisin, Carlsberg,
Alcalasa
Subst. BPN.
Substilisin Novo
Porcino, bovino
Bacillus licheniformis
Bacillus
amyloliquefaciens
30 – 60
45 – 55
50 – 60
40 – 55
7 – 9
8 – 9
6 – 8
6 – 10
6 – 10
Lis o Arg
Trp o Tir, Fe, Leu
Ala
AAhf
Cisteínica
Plantas
Papaína
Bromelaína
Ficina
Papaya
Piña
Látex de ficus
40 – 75
20 – 65
5 – 8
5 – 8
5 – 8
Fe o Val, Leu
AAhf
Aspárticas
Animal
Fúngico
Pepsina
Quimosina
Aspergilopeptidasa A
Newlasa
Porcino, bovino
Becerro
Aspergillus saitoi
Rhizopus sp.
35 – 50
40 – 50
1 – 4
4 – 6
2 – 5
3 – 6
Fe o Tir, Leu,
Trp o Fe, Tir
Glu, Asp, Leu
Similar a la pepsina
Metalo proteasas
Animal
Bacteriana
Carboxipeptidasa A
Neutrasa ®
Termolisina
Páncreas
Bacillus amyloliquefaciens
B. thermoproteolyticus
40 – 55
7 – 8
6 – 7.5
7 – 9
Carbonilo del AA
terminal (excepto Pro,
Arg, Lis)
Fe, Leu, Val
Ile, Leu, Val, Fe
AAhf = AAs hidrofóbicos Fuente: Benítez y col., 2008

Introducción
24
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
1.4 Actividad biológica y funcionalidad de péptidos
1.4.1 Alimentos funcionales
En la actualidad, los péptidos obtenidos tanto de fuentes animales como vegetales han sido
identificados y algunos sintetizados químicamente, si éstos presentan alguna actividad biológica. Se
han considerado como “alimentos funcionales” a aquellos en los cuales se ha modificado su
composición y en ellos se encuentran presentes por adición o enriquecimiento algunos compuestos
modificando los procesos usuales de manufactura. Los alimentos funcionales ejercen su actividad en
múltiples sistemas, especialmente el gastrointestinal, cardiovascular e inmunológico. Se comportan
como potenciadores del desarrollo y la diferenciación, moduladores del metabolismo de nutrientes,
la expresión génica y el estrés oxidativo (Silveira y col., 2003). Algunas actividades biológicas que
pueden presentar las proteínas alimenticias y sus péptidos son: la actividad antioxidante, la
antitumoral, la antitrombótica, la antibacteriana y la inhibidora de la enzima convertidora de la
angiotensina (Möller y col., 2008).
Algunos alimentos funcionales pueden contener péptidos bioactivos obtenidos vía hidrólisis
enzimática de proteínas, estos péptidos tienen la capacidad de mejorar la bio-disponibilidad de las
proteínas propiciando un efecto benéfico al consumirlos, ya sea a través de los mismos productos o
“creando” nuevos a través de la adición o fortificación de péptidos. Éste tipo de procedimientos se ha
reportado para productos de origen animal, en especial para el grupo de los lácteos (Dziuba y
Darewicz, 2007; Figueroa-Hernández, 2007; Norton y col., 2006; Drago-Serrano y col., 2006; Torres-
Llanez y col., 2005; Meisel y Bockelmann, 1999; Knorr, 1998; Vrese y Schrezenmeir, 2008).
También, se ha reportado que algunos compuestos responsables de la funcionalidad de
alimentos son los inhibidores de proteasas que se encuentran en todas las legumbres, sobre todo en
la soya y se encargan de que las semillas no hagan uso de sus reservas proteicas hasta su
germinación. Estas sustancias pueden inhibir también enzimas proteolíticas por lo que, hasta hace
poco, se las consideraban sustancias nocivas. Sin embargo, en la actualidad se ponderan sus efectos:
como protectores evitando la proliferación de células cancerígenas, actuando como antioxidantes,
reguladores de la glucemia y antiinflamatorios (Palencia-Mendoza, 1999; Hsieh y col., 2010).
En menor grado, pero no por ello dejan de ser importantes son las antocianinas que
pertenecen al grupo de los compuestos fenólicos (flavonoides) y se caracterizan por su solubilidad en
agua y por sus colores brillantes. Se encuentran en las variedades de frijol con testa de color rojo,
rosa y negro, y además de contribuir en su identificación debido a sus diferentes coloraciones; las

Introducción
25
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
antocianinas tienen una gran actividad antioxidante que inhibe los radicales libres de las células,
previniendo enfermedades como el cáncer, arterosclerosis e inflamaciones. La presencia de
antocianinas en el grano de frijol negro lo hace un producto potencial para el suministro de
colorantes y antioxidantes naturales. Por ello el estudio de los pigmentos del frijol negro (Phaseolus
vulgaris L.) ha despertado gran interés (Salinas-Moreno y col., 2005).
1.4.2 Péptidos con actividad biológica
En los últimos años se han realizado hidrolizados de proteínas provenientes de diversas fuentes
con la intención de obtener péptidos con potencial bioactividad ya que se ha estudiado que derivados
peptídicos pueden ser biológicamente activos (Benítez y col., 2008); los péptidos consisten en
cadenas cortas de 2 a 15 residuos aminoácidos (Torruco-Uco y col., 2009). Y se sabe que el consumo
diario de este nutriente por medio de las leguminosas, en los países en vías de desarrollo, éstas
contribuyen con el 10 % de las proteínas diarias y el 5 % del aporte energético en la dieta.
Recientes investigaciones han demostrado que los péptidos de origen animal poseen una o
varias actividades biológicas (Möller y col., 2008), incluso ha sido posible determinar la secuencia de
aminoácidos responsable de dichas actividades (Srihongthong y col., 2012; Gómez-Ruiz y col., 2008).
En este sentido la bioactividad de los péptidos de proteína de las diferentes fuentes han presentado
su funcionalidad como: antihipertensivos (Megías y col., 2009; Torruco-Uco y col., 2009; Valdéz-Ortíz
y col., 2012), antibacterianos (Srihongthong y col., 2012), anticariogénicos (Aimutis, 2004),
antiulceratívos (Tavares y col., 2011), anticarcinogénicos (Cardador-Martínez y col., 2002),
anticoagulante (Xiong y col., 2009), antitrombóticos (Fraga-Silva y col., 2010; Xiong y col., 2009) y
antioxidantes (Alemán y col., 2011; Valdés y col., 2011; Valdéz-Ortíz y col., 2012) por mencionar
algunas que incluso han sido identificados y sintetizados químicamente (Yamada y col., 2008).
En cuanto a los alimentos de origen vegetal, se han estudiado una gran variedad de
hidrolizados de proteínas (garbanzo, soya, chícharo, lenteja, frijol, entre otras) con la finalidad de
determinar si además de su alto contenido nutricional, poseen alguna actividad biológica, la cual
resultaría en un beneficio para la salud del ser humano que lo consume. Dentro de las propiedades
farmacológicas más reportadas de las diversas leguminosas destacan la actividad antioxidante y la
inhibidora de la enzima convertidora de la angiotensina (Möller y col., 2008).

Introducción
26
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
1.5 Aplicación y función de péptidos en la salud
1.5.1 Problemática actual y riesgos en la salud
En la actualidad la población se encuentra propensa a diversos padecimientos desencadenados
por procesos oxidativos celulares, la presencia de agentes patógenos en el ambiente o desequilibrio
en la regulación de funciones básicas como la presión sanguínea. Por lo tanto, la administración de
diversos agentes terapéuticos para el control de estos padecimientos es una práctica frecuente. Sin
embargo, la ingesta de estos está relacionada con efectos adversos derivados del uso individual o
como resultado de su interacción farmacológica durante su uso. En este sentido, la capacidad de
diversos péptidos de regular estos procesos representa una alternativa segura y sencilla para el
tratamiento de los padecimientos asociados al envejecimiento y estrés (Gómez-Ruiz y col., 2008).
1.5.2 Actividad antibacteriana como control de patógenos
El ser humano es susceptible a enfermedades causadas por microorganismos patógenos que
pueden resultar desde inofensivos hasta letales dependiendo del número y la naturaleza de los
mismos (Haque y Chand, 2008). Los compuestos antibacterianos en su mayoría son de origen
sintético, sin embargo, en la actualidad las personas han demandado por compuestos
antibacterianos de origen natural (aceites esenciales, bacteriocinas, etc.) con la finalidad de evitar las
consecuencias a largo plazo que pudieran contener los productos químicos (Tessera y col., 2012).
Existen pocos trabajos referentes al potencial antibacteriano de proteínas e hidrolizados de
proteínas, incluso no se ha establecido el mecanismo de acción que ejercen, aunque se han
planteado algunas propuestas como la interferencia en la síntesis de enzimas metabólicas y la
alteración de la membrana celular ya sea alterando la permeabilidad o lisándola mediante la
formación de canales o poros (Montaño-Pérez y Vargas-Albores, 2002; Wong y Ng, 2005).
Es posible que la actividad antibacteriana se encuentre presente en los hidrolizados de
fuentes de origen vegetal ya que se ha demostrado que los péptidos de origen animal pueden ser
usados como agentes antibacterianos y antifúngicos (Chan y col., 2012). Para esto, las propuestas del
mecanismo de acción ejercida por los péptidos antimicrobianos son las siguientes: a) interferir en la
síntesis de enzimas metabólicas o del DNA, como en el caso de algunos péptidos de origen
microbiano, cuyo proceso aún no es bien conocido; o b) actuar directamente a nivel de la membrana
celular ya sea alterando la permeabilidad o lisándola mediante la formación de canales o poros,
como en el caso de los péptidos α-hélice (Nissen-Meyer y Nes, 1997; Shai, 1998). Aunque se

Introducción
27
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
desconocen muchos detalles sobre los mecanismos de acción de los péptidos antimicrobianos α-
hélice, se considera que actúan directamente en la membrana celular y que su secuencia de
aminoácidos es importante (Matsuzaki, 1998), ya que si se sustituyen aminoácidos que alteren la
polaridad del péptido, su actividad disminuye. Aparentemente no requieren receptores específicos,
como se ha demostrado al sustituir aminoácidos por sus correspondientes D-enantiómeros sin
alterar su actividad, este comportamiento molecular y su carácter catiónico hacen que los péptidos
presenten una mayor afinidad por los fosfolípidos ácidos de las bacterias Gram(-) y polisacáridos de
las bacterias Gram(+), que por los fosfolípidos anfipáticos de las membranas de células de
mamíferos; lo anterior permite explicar la falta de citotoxicidad para las células animales y permite
proponer a los péptidos como agentes terapéuticos altamente específicos (Matsuzaki, 1998; Nissen-
Meyer y Nes, 1997; Shai, 1998; Vaucher y col., 2010).
En general, se ha propuesto que el mecanismo de acción de los péptidos antibacterianos está
dado por los siguientes pasos (Figura 1-3): la reacción inicia con la unión entre algunos péptidos y los
fosfolípidos ácidos de las membranas microbianas mediante fuerzas electrostáticas. En seguida, los
monómeros de los péptidos anfipáticos se acomodan en la superficie de la membrana de manera
que la carga positiva de los aminoácidos básicos concuerde con las cabezas de fosfolípidos con carga
negativa; y cuando la concentración relativa entre el péptido y el lípido localmente es baja el péptido
tiende a orientarse paralelamente en la membrana y es inactivo. Cuando la concentración local
relativa del péptido se incrementa, éste tiende a tomar una posición perpendicular a la membrana y
se inserta, orientando los residuos hidrofóbicos hacia la región hidrofóbica de la membrana. La
capacidad de “aceptación” de la membrana por una mayor cantidad de péptido dependerá de la
composición lipídica y propiedades fisicoquímicas de la misma. Esto explica los diferentes grados de
susceptibilidad de una célula por un determinado péptido o grupo de péptidos, y la especificidad
hacia ciertas bacterias. Finalmente, la inserción de los péptidos en la bicapa lipídica alteran la
permeabilidad de la membrana o bien ocurre la lisis celular por la presencia de los poros (Matsuzaki,
1998; Nissen-Meyer y Nes, 1997; Shai, 1998; Huang, 2000).

Introducción
28
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 1-3. Representación del mecanismo de acción de los péptidos antibacterianos del tipo alfa hélice. Dónde: a) Conformación lineal del péptido en solución acuosa donde se polarizan los aminoácidos
hidrofóbicos y los aminoácidos, b) La interacción electrostática entre los lipopolisacáridos de la membrana microbiana y los péptidos, y c) Inserción de péptidos en la membrana que dan lugar a la formación de canales
por los que fluyen elementos celulares (Fuente: Montaño-Pérez y Vargas-Albores, 2002).
1.5.3 Actividad antioxidante como control de la oxidación celular
Cada día aumentan las enfermedades crónicas y neurodegenerativas como la arteriosclerosis,
diabetes, cáncer, Alzheimer, envejecimiento prematuro, problemas cardiovasculares y el mal de
Parkinson asociadas a la presencia o producción de radicales libres, en la actualidad para
contrarrestar el efecto nocivo de los radicales libres se emplean sustancias antioxidantes de origen
natural o sintético (Jiménez-Arellanes y col., 2011). La oxidación celular es un proceso natural de
envejecimiento, debido a los radicales libres generados por una mala alimentación, la contaminación
del medio ambiente, la radiación solar y otros factores, sin embargo, se puede retrasar este proceso
con la presencia de compuestos antioxidantes en el organismo los cuales poseen la cualidad de
estabilizar a los radicales libres mediante la cesión de electrones (Medina-Godoy y col., 2012). La
caracterización de la funcionalidad antioxidante de los péptidos provenientes de diferentes
variedades de frijol permitiría su utilización como ingredientes en la elaboración de alimentos
funcionales y con ello obtener un efecto beneficioso para el ser humano produciendo una mejora en
su salud o reduciendo el riesgo de padecer alguna enfermedad asociada a los radicales libres.
Los antioxidantes son compuestos que al retardar o inhibir la degradación oxidativa de las
moléculas orgánicas, ayudan a prevenir la formación de colores y olores desagradables; estos pueden

Introducción
29
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
ser de origen natural o sintético, pero la mayoría de los utilizados comercialmente son de origen
sintético. Debido a que algunos de los antioxidantes sintéticos, son altamente inestables bajo las
condiciones de trabajo y en ciertos casos ocasionan efectos adversos sobre la salud de animales de
experimentación, los investigadores han intentado encontrar sustancias más estables, eficaces,
versátiles y/o menos tóxicas (Vásquez-Cardeño y col., 2007).
El daño oxidativo también tiene una gran importancia en los alimentos. Una consecuencia
habitual es la peroxidación lipídica que produce rancidez, aparición de sabores indeseables para el
consumidor y disminución de la vida comercial del consumidor (Liu y col., 2005). Para evitar estos
problemas la industria de alimentos utiliza antioxidantes de origen sintético, sin embargo, no se
descarta el empleo de antioxidantes de origen natural (carotenoides, compuestos fenólicos, vitamina
E) aún con la desventaja de ser de baja capacidad antioxidante y en su mayoría insolubles en sistemas
acuosos.
Se ha descrito el mecanismo de acción de los péptidos antioxidantes basado en su capacidad de
secuestrar radicales libres o su capacidad de formar complejos con los iones metálicos (Hernández-
Ledesma y col., 2005). Éstos péptidos actúan impidiendo que otras moléculas se unan a especies
reactivas del oxígeno, al interactuar más rápido con los radicales libres que éstos con el resto de las
moléculas presentes, es decir, el péptido actúa cediéndole un electrón al radical libre una vez que se
colisionan en un determinado microambiente de la membrana plasmática, citosol, núcleo o líquido
extracelular (Ruiz-Ruiz y col., 2013b).
Algunos trabajos han demostrado que los hidrolizados totales (HT) de frijol negro Jamapa
pueden tener una función antioxidante in vitro, incluso existen reportes donde se ha trabajado con
fracciones peptídicas de otro tipo de vegetales como chícharos, garbanzos y semillas de girasol (Ruiz-
Ruiz y col., 2013a). Además, en recientes estudios se ha demostrado que los hidrolizados
provenientes de fuentes animales como: huevo, leche y proteínas de pescado, poseen alto potencial
antioxidante (Srihongthong y col., 2012) y hay un menor número de trabajos reportados con fuentes
vegetales (Fritz y col., 2011; Yust y col., 2012), en particular con fracciones peptídicas de frijol. Ruiz-
Ruiz y col. (2013a) demostraron que al disminuir el peso molecular (PM) de los péptidos contenidos
de las fracciones proteicas, la actividad antioxidante se incrementó respecto a los hidrolizados
totales.
Se ha demostrado que las leguminosas pueden ser una fuente importante de péptidos
antioxidantes, además se ha propuesto que si tienen actividad biológica y resisten la digestión y
absorción in vivo, entonces podrían ser usados en la elaboración de alimentos funcionales para la

Introducción
30
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
prevención de distintas enfermedades y reducir el daño oxidativo de productos alimenticios
aumentando su vida útil (Vioque y col., 2000).
La cuantificación de la actividad antioxidante de un compuesto o de una mezcla se puede
realizar por los métodos de decoloración del radical libre 2,2-difenil-1- picril hidracilo (Marxen y col.,
2007; Mohamad y col., 2004), o por el atrapamiento de los radicales libres ABTS, DMPD o luminol (Ré
y col., 1999; Fogliano y col., 1999; Georgetti y col., 2003).
1.5.4 Actividad inhibidora de la enzima convertidora de la angiotensina como control de la
hipertensión
La hipertensión arterial o presión arterial alta afecta alrededor de mil millones personas en
todo el mundo (Nakahara y col., 2010). La hipertensión es un padecimiento multifactorial y uno de
los mecanismo más estudiados para su regulación y blanco de acción farmacológica es el sistema
renina-angiotensina, conformado por la renina, la enzima convertidora de angiotensina (ECA), la
aldosterona y las angiotensinas I (AGI) y II (AGII) (Keidar y col., 2007).
Y se ha demostrado que péptidos derivados de la hidrólisis enzimática de diferentes alimentos
en particular de origen animal tienen la capacidad de inhibir de forma selectiva a la ECA in vitro e in
vivo (Yamada y col., 2008). En algunos casos la cadena de aminoácidos de los péptidos que han
demostrado inhibir la ECA han sido elucidadas, resultando ser una alternativa segura y potencial para
el control de la hipertensión arterial (Rui y col., 2013). Se ha reportado que los hidrolizados totales y
las fracciones peptídicas de frijol negro Jamapa tienen capacidad de inhibir la ECA-I (Medina-Godoy y
col., 2012), y ésta actividad no ha sido estudiada con fracciones de péptidos de tamaño conocido de
otras variedades de P. vulgaris. Por lo que el estudio de péptidos de tres variedades de frijol (negro
plus, azufrado higuera y pinto Saltillo) podría abrir nuevas fuentes de péptidos con ésta actividad
biológica.
El mecanismo del sistema renina-angiotensina-aldosterona es el mayor regulador de la
fisiología humana, éste tiene el control de la presión sanguínea, volumen y electrolitos que afectan el
corazón, vasculatura y riñones. El proceso de la angiotensina comienza con las hidrólisis del
angiotensinogen por efecto de la renina para formar el decapéptido angiotensina I (Ang I). La
remoción de dos aminoácidos del carboxilo terminal por efecto de la enzima convertidora de la
angiotensina resulta en la formación de un péptido biológicamente activo, angiotensina II (Ang II). El
incremento en el nivel de Ang II en condiciones patológicas ha sido reportado en diversos sistemas
celulares, es por ello que la reducción de la Ang II es un tema de vital importancia clínica.

Introducción
31
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 1-4. Mecanismo de acción de la inhibición de la enzima convertidora de la angiotensina I (modificado de Keidar y col., 2007).
Keidar y col. (2007) propusieron un posible mecanismo de acción en el cual la Ang II y la ECA
tomaron los roles principales para la inhibición de la Ang I; y en el cual se involucran otras
angiotensinas bioactivas, la Ang (1–7) y ACE2 en un sistema de acción más complejo (Figura 1-4). En
el cual se muestra el siguiente fenómeno: primero los Inhibidores de la enzima convertidora de la
angiotensina (IECA) actúan produciendo un bloqueo competitivo de la ECA. Éstos inhiben también la
endopeptidasa neutra que degrada la bradicinina, por lo que su administración aumenta la
producción de cininas vasodilatadoras. Finalmente, los IECA disminuyen el tono simpático y los
valores plasmáticos de ADN estimulados por la angiotensina II.
La acción antihipertensiva se ejerce primero por la vasodilatación mediada por la bradicinina,
que relaja la musculatura lisa arteriolar, unos 15 días después comienza a actuar el mecanismo
vasodilatador y antiadrenérgico causado por la inhibición de la ECA, de 3 a 6 meses más tarde se
manifiesta la acción antitrófica que poco a poco revierte el crecimiento concéntrico de la media
arteriolar, a medida que esto ocurre se reduce la resistencia vascular periférica, lo que a su vez
impide la esclerosis vascular renal y coronaria (Cachofeiro y col., 1991).
Existen inhibidores de la ECA conocidos, sin embargo, éstos tienden a ser de origen químico y
al controlar la hipertensión llegan a afectar otros órganos. Los IECA conocidos se encuentran

Introducción
32
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
clasificados según su grupo químico que interactúa con el zinc en: compuestos sulfidrílicos
(captopril), compuestos carboxílicos (enalapril), y compuestos fosfóricos (fosinopril / ceranapril). La
efectividad de los IECA dependerá de la dosis y el organismo a quien le sea administrado. Los IECA
que tienen bien documentado sus beneficios sobre el corazón y a largo plazo son: captopril, enalapril,
lisinopril, ramipril y trandolapril (González-García y col., 2002). Siendo el Captopril® comercial el
medicamento utilizado como control para las pruebas de I-ECA de este trabajo.
1.6 Péptidos identificados como biológicamente activos
Está reportado que los veinte aminoácidos presentes en las proteínas pueden reaccionar con
los radicales libres si la energía de éstos es alta como aquellos que contienen a los radicales hidroxilo.
Entre los aminoácidos más reactivos se incluyen los azufrados (Met y Cys), los aromáticos (Trp, Tyr,
Phe) y los que contienen anillo imidazol como la His (Samaranayaka y Li-Chan, 2011). La mayoría de
los péptidos antioxidantes derivados de fuentes alimentarias presentan intervalos de peso molecular
de 500 a 1800 Da, y con frecuencia presentan restos de aminoácidos hidrofóbicos como Val o Leu en
el amino terminal, así como Pro, His, Tyr, Met y Cys en sus secuencias (Sarmadi e Ismail, 2010).
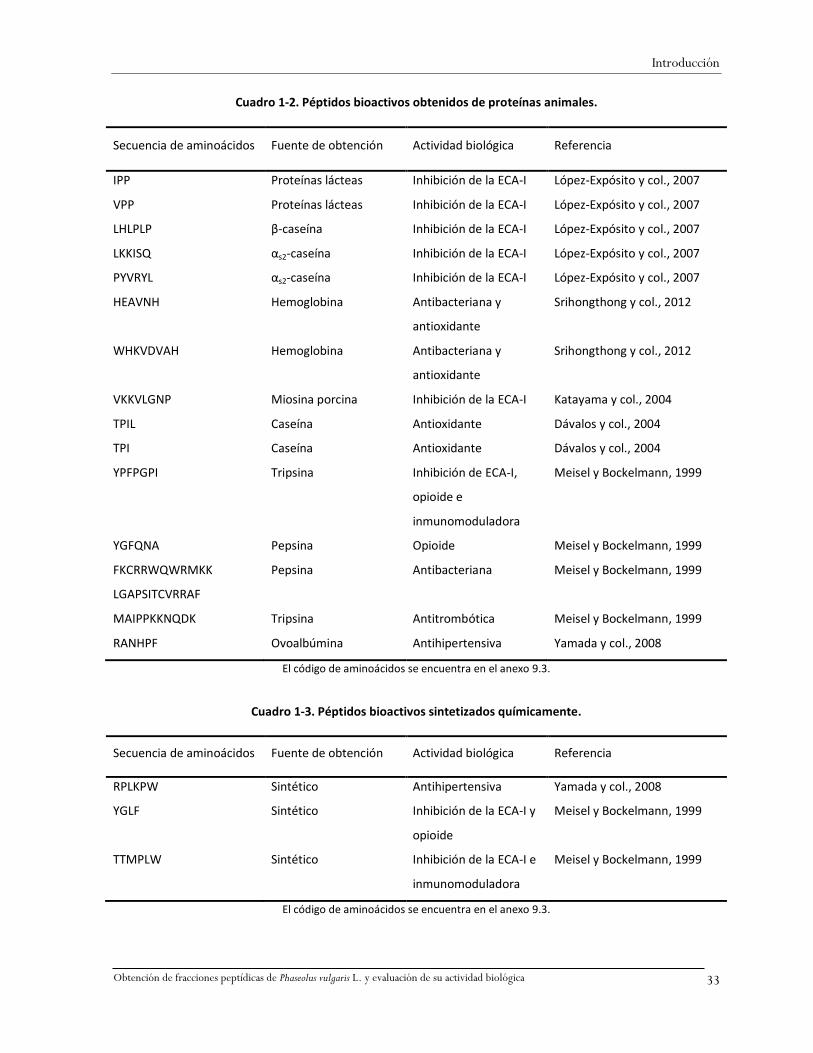
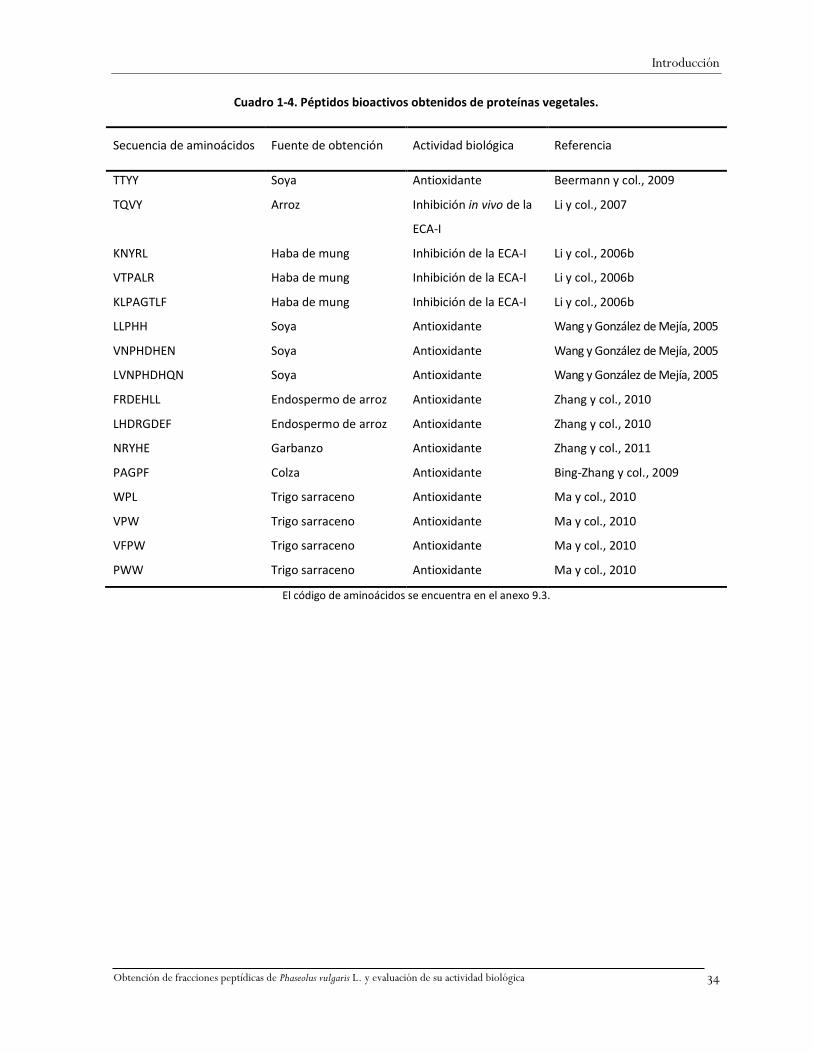
La secuencia de aminoácidos de algunos péptidos de origen animal identificados con actividad
comprobada se presenta en el Cuadro 1-2, de péptidos obtenidos por vía química en el Cuadro 1-3, y
de péptidos obtenidos de fuentes vegetales en el Cuadro 1-4. En éstos cuadros se observa que la
mayoría de los trabajos existentes son con hidrolizados de proteínas animales además de ser los que
se han investigado desde hace más tiempo, principalmente de proteínas lácteas; seguido de trabajos
con péptidos obtenidos sintéticamente y que han sido comparados con péptidos de origen animal; y
finalmente algunos péptidos obtenidos de fuentes vegetales.
Introducción
33
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Cuadro 1-2. Péptidos bioactivos obtenidos de proteínas animales.
Secuencia de aminoácidos Fuente de obtención Actividad biológica Referencia
IPP Proteínas lácteas Inhibición de la ECA-I López-Expósito y col., 2007
VPP Proteínas lácteas Inhibición de la ECA-I López-Expósito y col., 2007
LHLPLP β-caseína Inhibición de la ECA-I López-Expósito y col., 2007
LKKISQ αs2-caseína Inhibición de la ECA-I López-Expósito y col., 2007
PYVRYL αs2-caseína Inhibición de la ECA-I López-Expósito y col., 2007
HEAVNH Hemoglobina Antibacteriana y
antioxidante
Srihongthong y col., 2012
WHKVDVAH Hemoglobina Antibacteriana y
antioxidante
Srihongthong y col., 2012
VKKVLGNP Miosina porcina Inhibición de la ECA-I Katayama y col., 2004
TPIL Caseína Antioxidante Dávalos y col., 2004
TPI Caseína Antioxidante Dávalos y col., 2004
YPFPGPI Tripsina Inhibición de ECA-I,
opioide e
inmunomoduladora
Meisel y Bockelmann, 1999
YGFQNA Pepsina Opioide Meisel y Bockelmann, 1999
FKCRRWQWRMKK
LGAPSITCVRRAF
Pepsina Antibacteriana Meisel y Bockelmann, 1999
MAIPPKKNQDK Tripsina Antitrombótica Meisel y Bockelmann, 1999
RANHPF Ovoalbúmina Antihipertensiva Yamada y col., 2008
El código de aminoácidos se encuentra en el anexo 9.3.
Cuadro 1-3. Péptidos bioactivos sintetizados químicamente.
Secuencia de aminoácidos Fuente de obtención Actividad biológica Referencia
RPLKPW Sintético Antihipertensiva Yamada y col., 2008
YGLF Sintético Inhibición de la ECA-I y
opioide
Meisel y Bockelmann, 1999
TTMPLW Sintético Inhibición de la ECA-I e
inmunomoduladora
Meisel y Bockelmann, 1999
El código de aminoácidos se encuentra en el anexo 9.3.
Introducción
34
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Cuadro 1-4. Péptidos bioactivos obtenidos de proteínas vegetales.
Secuencia de aminoácidos Fuente de obtención Actividad biológica Referencia
TTYY Soya Antioxidante Beermann y col., 2009
TQVY Arroz Inhibición in vivo de la
ECA-I
Li y col., 2007
KNYRL Haba de mung Inhibición de la ECA-I Li y col., 2006b
VTPALR Haba de mung Inhibición de la ECA-I Li y col., 2006b
KLPAGTLF Haba de mung Inhibición de la ECA-I Li y col., 2006b
LLPHH Soya Antioxidante Wang y González de Mejía, 2005
VNPHDHEN Soya Antioxidante Wang y González de Mejía, 2005
LVNPHDHQN Soya Antioxidante Wang y González de Mejía, 2005
FRDEHLL Endospermo de arroz Antioxidante Zhang y col., 2010
LHDRGDEF Endospermo de arroz Antioxidante Zhang y col., 2010
NRYHE Garbanzo Antioxidante Zhang y col., 2011
PAGPF Colza Antioxidante Bing-Zhang y col., 2009
WPL Trigo sarraceno Antioxidante Ma y col., 2010
VPW Trigo sarraceno Antioxidante Ma y col., 2010
VFPW Trigo sarraceno Antioxidante Ma y col., 2010
PWW Trigo sarraceno Antioxidante Ma y col., 2010
El código de aminoácidos se encuentra en el anexo 9.3.

Justificación
35
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
2. Justificación
El frijol es una de las leguminosas más consumidas en nuestro país. En México es un cultivo
tradicional que se siembra en todas las regiones agrícolas y representa el segundo producto
agrícola alimenticio después del maíz y uno de los alimentos básicos de la dieta en el medio rural,
del mismo modo está documentado que el frijol posee un alto nivel de proteína comparado con
otros productos de origen vegetal, lo que le permite tener competitividad con respecto a otras
fuentes de proteína.
En los últimos años ha existido un creciente interés por la actividad biológica que han
mostrado diversos péptidos obtenidos de la hidrólisis de proteínas de diferentes fuentes animales
como: leche, huevo, plasma de sangre, músculo de pescado; así como también de fuentes
vegetales como: la soya, garbanzo, girasol, colza, lupino, entre otros. En este sentido la
bioactividad de los péptidos de proteína de diferentes fuentes de frijol no ha sido caracterizada
por completo como: antihipertensivos, antioxidantes, y antibacterianos.
Por lo tanto, la obtención de hidrolizados de proteínas de frijol y la caracterización de los
péptidos obtenidos contribuiría con información para su posible utilización como ingredientes en
la elaboración de alimentos funcionales, los cuales permitan obtener un efecto beneficioso para
mejorar la salud del ser humano o reduciéndole el riesgo de padecer alguna enfermedad.

Objetivos
36
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
3. Objetivos
3.1 Objetivo general
Evaluar la actividad biológica (antioxidante, antibacteriana e inhibidora de la enzima
convertidora de la angiotensina) de péptidos obtenidos por hidrólisis enzimática de tres
variedades de frijol (Phaseolus vulgaris L.): frijol negro plus FRI-064-2109993, frijol azufrado
higuera FRI-001-220995 y frijol pinto Saltillo 1424-FRI-026-120901/C.
3.2 Objetivos particulares
Obtener harinas de frijol de las variedades: negro plus, azufrado higuera y pinto Saltillo; y
determinar su composición químico proximal.
Obtener concentrados proteicos a partir de harinas de frijol de las variedades: negro plus,
azufrado higuera y pinto Saltillo; y observar su perfil proteíco en gel de poliacrilamida -
dodecil sulfato de sodio.
Hidrolizar los concentrados proteicos de frijol utilizando la enzima Alcalasa®.
Separar los péptidos en fracciones peptídicas de: f > 10 kDa, 10 kDa > 3 kDa, 3 kDa > 1 kDa y
f < 1 kDa, por ultrafiltración.
Evaluar la actividad antibacteriana de los hidrolizados y sus fracciones peptídicas.
Evaluar la actividad antioxidante de los hidrolizados y sus fracciones peptídicas.
Evaluar la actividad inhibidora de la enzima convertidora de la angiotensina de las fracciones
peptídicas.
Obtener la secuencia de aminoácidos de los péptidos que presentaron actividad biológica.

Materiales y Métodos
37
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4. Materiales y Métodos
4.1 Materiales biológicos y reactivos
Las semillas de frijol (Phaseolus vulgaris L.) utilizadas fueron proporcionadas por el Instituto
Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP, México); se utilizaron semillas
de las variedades: frijol negro plus (FNP) FRI-064-2109993, frijol azufrado higuera (FAH) FRI-001-
220995 y frijol pinto Saltillo (FPS) 1424-FRI-026-120901/C.
La enzima utilizada para el proceso de hidrólisis de las proteínas de frijol fue Alcalasa®
comercial (EC 3.4.21.62, endoproteinasa de Bacillus licheniformis, 2.4 AU/g) de Novozymes® (Sigma-
Aldrich, USA) conservada a 4 °C.
Las cepas utilizadas para la determinación de la actividad antibacteriana fueron: Bacillus subtilis
(ATCC 6633), Bacillus cereus (ATCC 11778), Escherichia coli, Salmonella typhi, Staphylococcus aureus
nombrada cepa 1 (aislamientos clínicos del CINVESTAV-IPN), Staphylococcus aureus nombrada cepa 2
(ATCC 12398), Listeria monocytogenes (Scott A), Klebsiella pneumoniae (ATCC 13884), Yersinia
enterocolitica, Klebsiella rinhoscleromatis (aislamientos clínicos de la FES Cuautitlán), Shigella
dysenteriae (INDRE B2188) y Enterobacter agglomerans (ATCC 27155, también llamada Pantoea
agglomerans).
Los reactivos empleados en las determinaciones fueron de grado analítico de J. T. Baker
(México), Sigma-Aldrich (Sigma Chemical Co., USA), o Bio-Rad (Bio-Rad Laboratories Inc., USA).
4.2 Caracterización física de las semillas de frijol
Una vez que se eliminaron las impurezas (como hojas y piedras) de las semillas de frijol de las
tres variedades empleadas, éstas fueron caracterizadas físicamente. Se determinó la masa (g) y las
dimensiones largo, alto y ancho (cm) de cien semillas de cada variedad. Y fueron clasificadas de
acuerdo con López-Herrera (2002) quién mencionó que si la masa de 100 semillas es menor de 25 g la
especie se considera de grano pequeño, de 25 -40 g de grano mediano y superior a 40 g de grano
grande. La forma (esfericidad) de las semillas de frijol fue determinada de acuerdo con la ecuación
reportada por Sreerama y col. (2009).
⁄
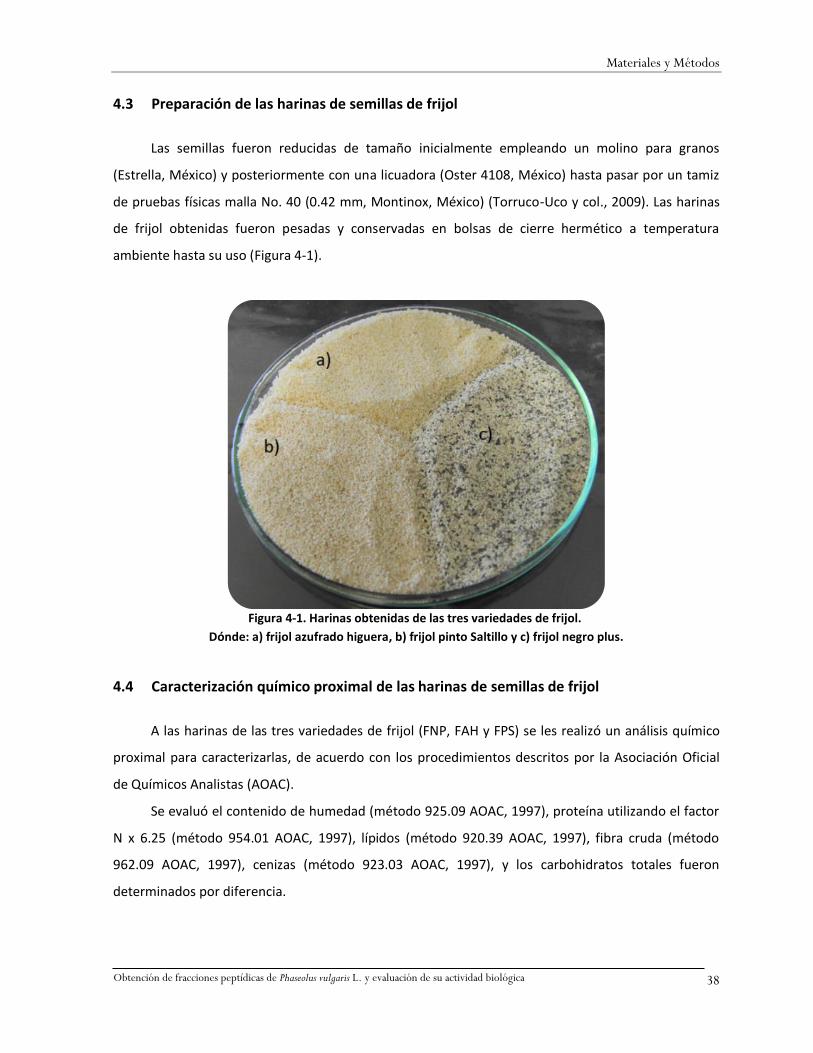
Materiales y Métodos
38
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.3 Preparación de las harinas de semillas de frijol
Las semillas fueron reducidas de tamaño inicialmente empleando un molino para granos
(Estrella, México) y posteriormente con una licuadora (Oster 4108, México) hasta pasar por un tamiz
de pruebas físicas malla No. 40 (0.42 mm, Montinox, México) (Torruco-Uco y col., 2009). Las harinas
de frijol obtenidas fueron pesadas y conservadas en bolsas de cierre hermético a temperatura
ambiente hasta su uso (Figura 4-1).
Figura 4-1. Harinas obtenidas de las tres variedades de frijol.
Dónde: a) frijol azufrado higuera, b) frijol pinto Saltillo y c) frijol negro plus.
4.4 Caracterización químico proximal de las harinas de semillas de frijol
A las harinas de las tres variedades de frijol (FNP, FAH y FPS) se les realizó un análisis químico
proximal para caracterizarlas, de acuerdo con los procedimientos descritos por la Asociación Oficial
de Químicos Analistas (AOAC).
Se evaluó el contenido de humedad (método 925.09 AOAC, 1997), proteína utilizando el factor
N x 6.25 (método 954.01 AOAC, 1997), lípidos (método 920.39 AOAC, 1997), fibra cruda (método
962.09 AOAC, 1997), cenizas (método 923.03 AOAC, 1997), y los carbohidratos totales fueron
determinados por diferencia.

Materiales y Métodos
39
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 4-2. Caracterización químico proximal de las harinas de frijol según métodos de la AOAC. Dónde: a) determinación de humedad, b) determinación de cenizas, c) determinación de grasa,
d) determinación de proteína y e) determinación de fibra.
4.4.1 Determinación de humedad
Se utilizó el método de calentamiento directo para el cual se pesaron 3 g de harina de frijol de
cada variedad en charolas con masa constante por triplicado. Las muestras se colocaron en una
estufa con temperatura de 60 °C por 12 h (Figura 4-2a). Se calculó el porcentaje de humedad en las
muestras con la siguiente ecuación:
4.4.2 Determinación de proteína total
Se determinó por la técnica de micro-Kjeldalh para lo cual se pesaron 0.040 g de harina seca y
desengrasada en papel libre de cenizas (papel arroz) por triplicado. Simultáneamente se corrió un
blanco el cual solo contenía el papel arroz. La muestra fue colocada en un matraz tipo Kjeldalh con la

Materiales y Métodos
40
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
adición de 2 mL de H2SO4 concentrado y una mezcla de catalizadores (1.9 g de K2SO4 y 40 mg de CuO),
y digerida hasta la desaparición de color en la muestra (Figura 4-2d). Posteriormente, la muestra
digerida fue destilada en un equipo automático K-350 (BUCHI, Suiza) con la adición de NaOH al 32 %
(p/v) y el destilado recibido en 5 mL de H3BO3 al 2 % (p/v) con dos gotas del reactivo de Wesslow
(preparado con dos partes de solución etanólica al 60 % (v/v) de rojo de metilo al 0.2 % (p/v) y una
parte de solución acuosa de azul de metileno al 0.2 % (p/v)). Finalmente el destilado atrapado en el
ácido bórico se tituló con una solución valorada de HCl 0.1 N y la adición de dos gotas de fenolftaleína
al 1 % (p/v). El contenido de proteína se calculó utilizando las ecuaciones siguientes:
4.4.3 Determinación del extracto etéreo
Se pesaron 3 g de harina de frijol sin humedad dentro de un cartucho de celulosa de 33 x 80
mm (Whatman, USA) y éste se colocó dentro de un equipo Soxhlet (Pyrex, México). Las
determinaciones se realizaron por triplicado. La extracción se llevó cabo con éter etílico en
recirculación por 4 h (Figura 4-2c). Al finalizar el tiempo la muestra se retiró del equipo, se secó y se
conservó en recipientes herméticos dentro de un desecador para posteriores análisis. El residuo en el
matraz balón se llevó a peso constante y se pesó para determinar la cantidad de grasa en la muestra
con la siguiente ecuación:
4.4.4 Determinación de fibra cruda
Se pesó 1 g de muestra libre de grasa a la cual se le realizó una digestión ácida con H2SO4 al
1.25 % (v/v) en ebullición por 30 min, seguida de una digestión alcalina con NaOH al 3.52 % (v/v) bajo
las mismas condiciones para llevar a cabo una digestión completa. La muestra fue filtrada con papel
filtro libre de cenizas (Whatman No. 40) en un embudo Buchner adaptado a una bomba de vacío;
durante el proceso de filtrado se realizaron varios lavados a la muestra: con agua caliente, H2SO4 1.25
%, agua a temperatura ambiente, alcohol y éter (Figura 4-2e). Las determinaciones se realizaron por

Materiales y Métodos
41
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
triplicado. El material retenido en el papel filtro se llevó a peso constante. Después de registrar la
masa de la muestra seca, ésta se pre-calcinó para ser calcinada a 600 °C por 4 h en una mufla modelo
FB1315M (Barnstead, USA). La fibra cruda se calculó con la siguiente ecuación:
4.4.5 Determinación de cenizas totales
El contenido de cenizas totales se determinó por el método de calcinación en mufla para lo cual
se pesó 1 g de harina sin humedad en crisoles a peso constante por triplicado. Se llevó a cabo una
pre-calcinación de las muestras con un mechero Fisher para evitar el desprendimiento de gases y
pérdida de muestra dentro la mufla FB1315M (Barnstead, USA). Posteriormente, las muestras se
calcinaron a 600 °C por 4 h (Figura 4-2b). El porcentaje de cenizas se determinó con la siguiente
ecuación:
4.4.6 Carbohidratos totales
El contenido de carbohidratos fue estimado como la diferencia del 100 % menos las
determinaciones de: proteína, lípidos, fibra cruda y cenizas; tomando en cuenta el contenido de
humedad y grasa en la muestra.
4.5 Proceso de desengrasado de las harinas de frijol
El proceso de desengrasado de la harina consistió en suspenderla en una proporción 1:5 (p/v)
con hexano dentro de un matraz Erlenmeyer con baño de hielo y mantener la mezcla en agitación por
8 h (Chung y col., 2007). Posteriormente, la harina desengrasada fue separada por medio de un tamiz
de pruebas físicas malla No. 80 (0.18 mm, Montimex, México) con el fin de obtener una muestra
homogénea. Las harinas desengrasadas fueron conservadas en bolsas de cierre hermético a
temperatura ambiente hasta su posterior utilización.

Materiales y Métodos
42
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.6 Obtención de los concentrados proteicos de frijol
La extracción proteica se llevó a cabo por precipitación isoeléctrica de acuerdo con Carrasco-
Castilla y col. (2012). La extracción se realizó al suspender harina desengrasada al 10 % (p/v) en agua
destilada a 4 °C, se ajustó el pH de la suspensión en 8.0 mediante la adición de NaOH 1.0 N y se
mantuvo en agitación por al menos 30 min. En seguida, las muestras se centrifugaron a 12,400 x g en
una centrífuga J2-MC (Beckman Coulter, USA) por 30 min a 4 °C. Se tomó el sobrenadante para
ajustarlo a pH 4.5 con adición de HCl 1.0 N con el fin de lograr la precipitación de las proteínas y se
mantuvo en agitación por al menos 30 min. Las muestras se centrifugaron bajo las mismas
condiciones anteriores. El concentrado proteico acumulado en el fondo del recipiente fue congelado
para ser sometido a un proceso de liofilización por un periodo de 24 a 48 h en un equipo 6 Plus
(Labconco, USA) a 0.030 mBar y -77 °C en el colector. Las muestras liofilizadas fueron conservadas en
tubos falcon a temperatura ambiente y dentro de un desecador para ser caracterizadas e hidrolizadas
posteriormente.
4.7 Caracterización de los concentrados proteicos de frijol
4.7.1 Contenido total de proteína en los concentrados de frijol
Se determinó el porcentaje de proteína total con base en la cantidad de nitrógeno contenida en
los concentrados proteicos de las tres variedades de frijol por el método de micro-Kjeldahl (método
954.01 AOAC, 1997), descrito en 4.4.2. Se utilizaron 0.010 g de cada concentrado de proteína
liofilizado para este procedimiento y se llevó a cabo por triplicado.
4.7.2 Determinación de proteína soluble en los concentrados proteicos de frijol
Se llevó a cabo mediante el método de Bradford (1976), el cual se basa en la unión del
colorante azul de Comassie G-250 a las proteínas presentes en una muestra. El colorante en solución
ácida existe en dos formas una azul y otra naranja, las proteínas se unen a la forma azul para formar
un complejo proteína-colorante con un coeficiente de extinción mayor que el colorante libre. El
reactivo fue adquirido preparado (Sigma-Aldrich, USA) y las muestras fueron leídas a 595 nm en un
espectrofotómetro Lambda XLS (Perkin Elmer, USA). También, se adaptó esta metodología a
microplacas de 96 pozos con la adición de 200 µL del reactivo y 10 µL de la muestra, en este caso la

Materiales y Métodos
43
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
lectura fue a 620 nm. La curva tipo se elaboró utilizando albúmina de suero bovino (BSA) en
concentración de 0 - 2 mg/mL para ambos casos.
4.7.3 Perfil proteico de los concentrados de proteína de frijol
Se realizó una electroforesis en gel de poliacrilamida/bisacrilamida con dodecilsulfato de sodio
(SDS-PAGE por sus siglas en inglés) para la observación del perfil proteico de las proteínas contenidas
en los concentrados de frijol. En una cámara Mini PROTEAN 3 cell (BIO-RAD Hercules, EUA) y a una
concentración del 15 % (p/v), se preparó el gel siguiendo el protocolo de Laemmli (1970). La
determinación se realizó en condiciones reductoras (en presencia de β–mercaptoetanol) y no
reductoras. Los concentrados de proteína fueron mezclados en proporción 1:2 (v/v) con una solución
búfer que contenía 200 g/L SDS, 25 mL/L glicerol, 10 mL/L β-mercaptoetanol, 0.20 g/L azul de
bromofenol y 10 mL de agua desionizada. El gel consistió en 2.1 g/L de SDS, 200 g/L de acrilamida
disuelta en el búfer separador (pH 8.8) y 50 g/L de acrilamida disuelta en búfer concentrador (pH 6.8).
El grosor del gel fue de 1.5 mm, con 6 cm en el separador y 2 cm en el concentrador. El gel se corrió a
una intensidad de corriente de 100 V por 90 min, las bandas proteicas fueron teñidas por inmersión
en una solución azul de Comassie R-250, 400 mL de metanol, 70 mL de ácido acético y 530 mL de
agua desionizada. El marcador de peso molecular utilizado fue RPN 2107 (fosforilasa 97 kDa,
albúmina 66 kDa, ovoalbúmina 45 kDa, anhidrasa carbónica 30 kDa, inhibidor de tripsina 20.1 kDa, α–
lactoglobulina 14.4 kDa, ECL, Amersham Biosciences).
Figura 4-3. Equipo de (a) isoelectroenfoque y (b) electroforesis en gel de poliacrilamida dodecil sulfato de sodio de las proteínas de frijol.
a) b)

Materiales y Métodos
44
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.7.4 Electroforesis bidimensional de los concentrados de proteína de frijol
Mediante la técnica de electroforesis bidimensional en gel de poliacrilamida/bisacrilamida
con dodecilsulfato de sodio (SDS-PAGE 2D) el perfil proteico de las proteínas contenidas en los
concentrados de frijol puede ser observado mediante la separación de proteínas en un equipo de
isoelectroenfoque por su punto isoeléctrico (pI) y por su masa molecular en un equipo estándar para
electroforesis (Aguilar y col., 2009). Para llevar a cabo el corrimiento de las proteínas en primera
dimensión o isoelectroenfoque las proteínas (100 µg) fueron solubilizadas en búfer de rehidratación
(urea 7 M, tiourea 2 M, CHAPS 4 %, azul de bromofenol 0.1 %, DTT 40 mM), y se aplicaron en tiras IPG
de 7 cm de largo con intervalo de pH 3-10 (ReadyStrip, BIO-RAD). Las tiras se cubrieron con aceite
mineral y se enfocaron mediante un equipo (GE, USA) por 2.5 h en un programa a pasos de 500 a
4000 V, 50 µA y 20 °C. En seguida, las tiras fueron acondicionadas al permanecer inmersas en búfer
de equilibrio 1 (urea 6 M, tris-HCl pH 8.8 5 mM, SDS 2 %, glicerol 20 %, DTT 2 %) y en búfer de
equilibrio 2 (urea 6 M, tris-HCl pH 8.8 5 mM, SDS 2 %, glicerol 20 %, yodoacetamida 2.5 %) por al
menos 30 min en cada solución. Finalmente, las tiras se colocaron en la parte superior de un gel SDS-
PAGE al 12 % y se corrieron de acuerdo con lo descrito en 4.7.3 (Figura 4-3). El marcador de peso
molecular utilizado fue SM0671 (Fermentas, USA).
4.8 Hidrólisis enzimática de las proteínas de frijol
Figura 4-4. Montaje experimental utilizado durante el proceso de hidrólisis enzimática.

Materiales y Métodos
45
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
La hidrólisis enzimática de las proteínas de frijol se realizó con el método descrito por Torruco-
Uco y col. (2009). Los concentrados proteicos (liofilizados) al 4 % (p/v) fueron suspendidos en agua
dentro de un reactor de vidrio con doble pared a una temperatura controlada de 55°C, agitación
constante por 120 min y pH ajustado en 8.0 con adición de NaOH 0.1 N mediante un equipo pH-STAT
Titrino 702 (Metrohm, Suiza) (Figura 4-4). La hidrólisis fue realizada usando la enzima Alcalasa® (EC
3.4.21.62 endoproteinasa de Bacillus licheniformis, 2.4 AU/g) de Novozymes® (Sigma-Aldrich, USA) en
proporción 0.3 AU/g (enzima/sustrato), el tiempo de reacción, las concentraciones de sustrato y
enzima fueron establecidos de acuerdo con trabajos anteriores (Ahumada-González, 2007; Torruco-
Uco y col., 2009; Boye y col., 2010) y pruebas preliminares a 1, 2 y 3 h de reacción. El grado de
hidrólisis alcanzado durante la reacción se determinó por medio de la ecuación de Adler-Nissen
(1979).
Dónde: B es el consumo (mL) de NaOH 0.1 N, NB es la normalidad del NaOH, α es el promedio
del grado de disociación de los grupos α–NH (1.105 para pH 8.0 y 55 °C, Adler-Nissen, 1986), MP es la
masa (g) de la proteína (N x fN) y htot el número total de enlaces peptídicos en el sustrato proteínico,
se utilizó el valor de 8.0 meq/g prot reportado por Arcila (2011) para frijol bayo.
4.9 Fraccionamiento de los hidrolizados proteicos por ultrafiltración
Los hidrolizados fueron fraccionados en un equipo de ultrafiltración Amicon LabscaleTM system
(Millipore, Alemania) con presión de nitrógeno a través de membranas con cortes de peso molecular
de 10 kDa (NMWL:10000), 3 kDa (NMWL:3000) y 1 kDa (NMWL:1000) hasta concentrar
aproximadamente un 80 % del volumen inicial, para finalmente obtener cuatro fracciones peptídicas,
lo que resulta similar a lo reportado por Palma-Rodríguez (2008). Después de este proceso se
obtuvieron cuatro muestras de pesos moleculares: fracción menor de 1 kDa (f<1), fracción de 1-3 kDa
(f1-3), fracción de 3-10 kDa (f3-10) y la fracción mayor de 10 kDa (f>10) de cada una de las variedades
de frijol ensayadas. A cada muestra se le determinó el contenido de proteína soluble por el método
de Bradford (1976) descrito en 4.7.2. Las muestras fueron congeladas y liofilizadas para ser utilizadas
en la determinación de la actividad antibacteriana, o sólo congeladas en el caso de la determinación
de la actividad antioxidante y la actividad inhibidora de la enzima convertidora de la angiotensina I.

Materiales y Métodos
46
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.10 Caracterización del fraccionamiento por ultrafiltración
La caracterización de las fracciones separadas por ultrafiltración se realizó con una
electroforesis en gel de poliacrilamida/bisacrilamida con dodecilsulfato de sodio al 18 %, en
condiciones reductoras para la observación del perfil de las proteínas en cada una de las fracciones
obtenidas por ultrafiltración para lo cual se colocaron 5 µg de proteína en cada pozo y las condiciones
fueron las mismas que se describieron en la sección 4.7.3.
4.11 Determinación de la actividad antibacteriana de los hidrolizados totales y fracciones
peptídicas de frijol
4.11.1 Prueba de susceptibilidad microbiana
Las cepas utilizadas para la determinación de actividad antibacteriana fueron mencionadas en
4.1. De las cepas indicadoras utilizadas para esta prueba, cinco se caracterizan por ser Gram (+): B.
subtilis, B. cereus, S. aureus cepa 1, S. aureus cepa 2, L. monocytogenes; y las otras por ser Gram (-): Y.
enterocolítica, E. coli, K. pneumoniae, K. rhinoscleromatis, S. typhi, S. dysenteriae y E. agglomerans.
Las cepas indicadoras fueron reactivadas en caldo Müeller Hinton (Fluka, Suiza) y posteriormente
crecidas en agar Müeller Hinton II (Fluka, Suiza).
Se realizó la prueba de susceptibilidad microbiana por la técnica de difusión en agar según el
protocolo de Bauer y col. (1966). Las muestras analizadas fueron: las harinas de frijol desengrasadas,
los hidrolizados totales obtenidos a partir de las harinas desengrasadas, los concentrados proteicos
de frijol sin hidrolizar, los hidrolizados totales (HT) obtenidos a partir de los concentrados proteicos
de frijol y las cuatro fracciones peptídicas (f<1, f1-3, f3-10 y f>10) obtenidas a partir del proceso de
hidrólisis y ultrafiltración. Los ensayos se realizaron a diferentes concentraciones (5, 10 y 20 µg en el
sensidisco). La metodología consistió en ajustar las cepas indicadoras en solución salina al 0.9 % (p/v)
a una turbidez 0.5 de la escala de McFarland (1.5 x 108 UFC/mL) y con ayuda de un isopo estéril
realizar la inoculación de la superficie del agar cubriendo toda la superficie de la caja Petri. De manera
simultánea se adicionó la muestra en discos de papel filtro (Whatman No. 1) de 6 mm de diámetro y a
los cuales se les dejó secar en condiciones de esterilidad. Finalmente se colocaron los sensidiscos
sobre la superficie de agar Müeller-Hinton previamente inoculado y se las cajas Petri se incubaron a
35 ± 2 °C por 24 h. Pasado este tiempo se midieron los halos formados en la superficie del agar
(Figura 4-5a).

Materiales y Métodos
47
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Se usaron dos controles positivos que fueron cloranfenicol (30 µg) y ampicilina (10 µg), así
como agua como control negativo (Taroco y col., 2006). El control positivo se interpretó de acuerdo
con lo reportado por Konneman y col. (2004).
Figura 4-5. Determinación de la actividad antibacteriana de los hidrolizados totales y fracciones peptídicas.
Dónde: a) prueba de susceptibilidad y b) determinación de la CMI.
4.11.2 Determinación de la concentración mínima inhibitoria
Para esta prueba sólo se utilizaron los microorganismos que presentaron halos de inhibición en
la prueba de sensibilidad (4.11.1). La concentración mínima inhibitoria (CMI) se define como la
concentración más baja de un compuesto con características antibacterianas que inhibe el
crecimiento visible de un microorganismo después de su incubación y fue determinada por la técnica
de dilución en microplaca (Langfield y col., 2004).
La prueba consistió en igualar la turbidez de cada microorganismo al estándar 0.5 de la escala
de Mc Farland y realizar una dilución 10-2 en caldo Müeller-Hinton. Posteriormente en placas de 96
pozos se adicionaron por triplicado 50 µL de la solución bacteriana (concentración final de 1.5 x 104
UFC/mL) con 50 µL de las diferentes concentraciones de proteína de las muestras (hidrolizados
totales o fracciones peptídicas de frijol en 100, 200, 300, 400 y 500 µg/mL); éstas cajas se incubaron a
35 ± 2 °C por 24 h. Se incubó también un control positivo que consistió en 100 µL de solución
bacteriana y un control negativo que consistió en solución bacteriana con cloranfenicol (30 µg).
Después del periodo de incubación se agregó cloruro de tetrazolio como indicador de
crecimiento microbiano, el colorante vira de incoloro a violeta en presencia de células vivas; se
adicionaron 40 µL en cada pozo de una solución al 0.1 % (p/v) disuelto en una solución 2:3 de K2HPO4
a) b)

Materiales y Métodos
48
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
0.1 M y NaH2PO4 0.1 M. La CMI se determinó a partir de la concentración en la cual no se observó
coloración después de 30 min de haber sido adicionado el reactivo (Figura 4-5b).
4.12 Determinación de la actividad antioxidante de los hidrolizados totales y fracciones
peptídicas de frijol
4.12.1 Actividad antioxidante como captura del radical libre DPPH+
Esta técnica determina la capacidad de una muestra para capturar radicales libres 2,2-difenil-1-
picril-hidracilo (DPPH+) a una longitud de onda de 517 nm, mediante la donación de electrones (Figura
4-6); siendo dicha actividad antioxidante indicada por el cambio de coloración del rosa al amarillo del
radical en solución metanólica. El método fue descrito por Von-Gadow y col. (1997) y modificado por
Kim y col. (2002).
Figura 4-6. Estructura química (a) del radical libre difenilpicrilhidrazilo y (b) del compuesto estable difenilpicrilhidrazina (Molyneux, 2004).
Se preparó una solución metanólica 0.1 mM de DPPH+ de la cual se tomaron 2.9 mL para cada
ensayo, a esos se agregaron 0.1 mL de la muestra problema o del reactivo antioxidante de referencia.
Los ensayos se realizaron por triplicado. Las muestras a analizar fueron los hidrolizados totales de
frijol y las fracciones peptídicas de frijol obtenidas por ultrafiltración, y los antioxidantes de referencia
fueron: ácido-6-hidroxi-2,5,7,8-tetramethilcromano-2-carboxílico (Trolox 0 - 1.2 mM), ácido ascórbico
(AASC 0 - 1.2 mM) y ácido gálico (AGAL 0 - 0.75 mM) utilizados para calcular la actividad antioxidante
eequivalente. La absorbancia (Abs) se leyó a los 30 y 60 min de reacción a una longitud de onda de
517 nm en un espectrofotómetro Lambda XLS (Perkin Elmer, USA) y se usó metanol como blanco. La
actividad antioxidante (% AA) en función de la captura del radical libre DPPH por las muestras se
determinó mediante la ecuación:
a) b)

Materiales y Métodos
49
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.12.2 Actividad antioxidante en función de la decoloración del radical libre ABTS+
Esta técnica se basa en la generación del radical catión ABTS+ (cromóforo azul-verde) mediante
la oxidación del ácido 2,2′-azino-bis(3-etilbenzotiazolin-6-sulfónico) (ABTS) con K2S2O8 (Figura 4-7), y
determina la capacidad de una muestra de decolorar dichos radicales libres generados por ésta sal
diamonical (Ré y col., 1999).
Figura 4-7. Reacción de formación del radical libre ABTS+.
El radical ABTS+ se generó al preparar una solución de ABTS 7 mM con K2S2O8 2.45 mM, a la
cual en condiciones de oscuridad y temperatura ambiente se le permitió el reposo por un periodo de
18 h antes de ser utilizado, su coloración es azul/verde intenso y permanece estable por 48 h. El
radical ABTS+ fue diluido con etanol hasta obtener una absorbancia de 0.7 ± 0.1 a una longitud de
onda de 734 nm. Del reactivo ajustado se tomaron 2 mL para cada ensayo, a esos se agregaron 20 µL
de la muestra problema o del reactivo antioxidante de referencia. Las muestras fueron los
hidrolizados totales de frijol y las fracciones peptídicas de frijol obtenidas por ultrafiltración, y los
antioxidantes de referencia fueron: Trolox (0 – 2.5 mM), AASC (0 – 2.5 mM) y AGAL (0 - 1 mM)
utilizados para calcular la actividad antioxidante equivalente. La absorbancia (Abs) se leyó a los 1 y 6
min de reacción a una longitud de onda de 734 nm en un espectrofotómetro Lambda XLS (Perkin
Elmer, USA) y se usó etanol como blanco. Los ensayos se realizaron por triplicado. La actividad
antioxidante (% AA) en función de la decoloración del radical libre por las muestras se determinó
mediante la ecuación:

Materiales y Métodos
50
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.12.3 Actividad antioxidante en función de la decoloración del radical libre DMPD+
Esta técnica permite determinar la capacidad de una muestra para decolorar los radicales libres
generados por el diclorohidrato de N,N-dimetil-p-fenilendiamina (DMPD), y se basa en la generación
del radical DMPD+ (rosa-rojo) mediante la oxidación con FeCl3 (Fogliano y col., 1999) (Figura 4-8).
Figura 4-8. Reacción de decoloración del radical libre DMPD+ (Chagas y col., 2009).
El radical DMPD+ fue generado al preparar un 1 mL de solución acuosa de DMPD 100 mM, que
posteriormente fue adicionada en solución búfer de acetatos 0.1 M (ácido acético/acetato de sodio,
pH 5.25) con 200 µL de FeCl3 0.05 M. El radical DMPD+ fue diluido con búfer de acetatos hasta
obtener una absorbancia de 0.9 ± 0.1 a una longitud de onda de 506 nm, la solución se mantuvo en
condiciones de oscuridad. Del reactivo ajustado se mezclaron 2 mL para cada ensayo con 100 µL de la
muestra problema o del reactivo antioxidante de referencia. Las determinaciones fueron realizadas
por triplicado. Las muestras a analizar fueron los hidrolizados totales de frijol y las fracciones
peptídicas de frijol obtenidas por ultrafiltración. Los antioxidantes de referencia fueron: Trolox (0 – 1
mM), AASC (0 – 0.5 mM) y AGAL (0 – 2 mM) utilizados para calcular la actividad antioxidante
equivalente. La absorbancia (Abs) se leyó a los 10 min de reacción a una longitud de onda de 506 nm
en un espectrofotómetro Lambda XLS (Perkin Elmer, USA) y se usó búfer de acetatos como blanco. La
actividad antioxidante (% AA) en función de la decoloración del radical libre por las muestras se
determinó mediante la ecuación:
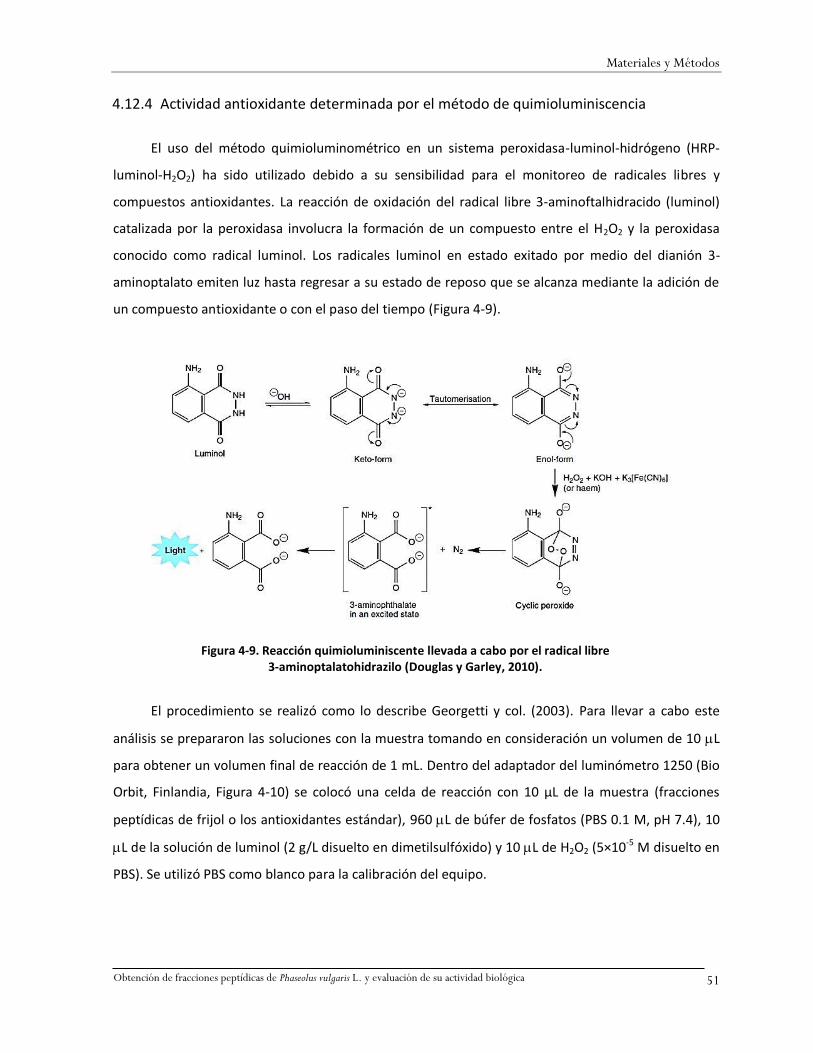
Materiales y Métodos
51
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.12.4 Actividad antioxidante determinada por el método de quimioluminiscencia
El uso del método quimioluminométrico en un sistema peroxidasa-luminol-hidrógeno (HRP-
luminol-H2O2) ha sido utilizado debido a su sensibilidad para el monitoreo de radicales libres y
compuestos antioxidantes. La reacción de oxidación del radical libre 3-aminoftalhidracido (luminol)
catalizada por la peroxidasa involucra la formación de un compuesto entre el H2O2 y la peroxidasa
conocido como radical luminol. Los radicales luminol en estado exitado por medio del dianión 3-
aminoptalato emiten luz hasta regresar a su estado de reposo que se alcanza mediante la adición de
un compuesto antioxidante o con el paso del tiempo (Figura 4-9).
Figura 4-9. Reacción quimioluminiscente llevada a cabo por el radical libre 3-aminoptalatohidrazilo (Douglas y Garley, 2010).
El procedimiento se realizó como lo describe Georgetti y col. (2003). Para llevar a cabo este
análisis se prepararon las soluciones con la muestra tomando en consideración un volumen de 10 L
para obtener un volumen final de reacción de 1 mL. Dentro del adaptador del luminómetro 1250 (Bio
Orbit, Finlandia, Figura 4-10) se colocó una celda de reacción con 10 µL de la muestra (fracciones
peptídicas de frijol o los antioxidantes estándar), 960 L de búfer de fosfatos (PBS 0.1 M, pH 7.4), 10
L de la solución de luminol (2 g/L disuelto en dimetilsulfóxido) y 10 L de H2O2 (5×10-5 M disuelto en
PBS). Se utilizó PBS como blanco para la calibración del equipo.

Materiales y Métodos
52
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
La reacción de oxidación se inició al adicionar 10 L de la solución enzimática de peroxidasa
(HRP 0.2 IU/mL disuelta en agua), dando seguimiento a la misma en el programa computacional
WinDaq (DataQ Instruments Inc., USA) durante 10 min para obtener las curvas de actividad
luminiscente. Los antioxidantes de referencia utilizados como control positivo fueron: Trolox (0 – 12
mM), AASC (0 – 12 mM) y AGAL (0 – 7.5 mM).
Figura 4-10. Montaje experimental del equipo de quimioluminiscencia.
Los resultados fueron expresados como porcentaje de inhibición en función del control
negativo calculados mediante la siguiente ecuación:
4.13 Determinación de la actividad inhibidora de la enzima convertidora de la
angiotensina
La determinación de la actividad inhibidora de la enzima convertidora de la angiotensina (EC
3.4.15.1), se trata de un ensayo enzimático, y se llevó a cabo siguiendo el método descrito por
Cushman y Cheung (1971). El sustrato consistió en 110 μL de una solución de hipuril-L-histidil-L-
leucina (HHL) 10 mM disuelto en búfer de fosfatos (PBS 0.1 mM, pH 8.3) con NaCl 0.3 M. Al sustrato
se añadieron 25 μL de una solución que contenía 26 mU/mL de la enzima convertidora de
angiotensina (ECA), adicionalmente se adicionaron 15 μL de la fracción de péptidos de pesos
moleculares entre 1 y 3 kDa (concentración final 20 µg/mL). Las muestras se incubaron a 37 °C por 80

Materiales y Métodos
53
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
min para posteriormente inactivar la enzima por la adición de 110 μL de ácido clorhídrico 1 N. El ácido
hipúrico fue extraído mediante agitación después de la adición de 1 mL de acetato de etilo. La
muestra fue centrifugada a 3000 × g por 10 min, se colocaron en un nuevo tubo 750 µL de la fase
orgánica y se procedió a la eliminación del acetato de etilo por evaporación. El residuo de ácido
hipúrico se disolvió en 1 mL de agua destilada y finalmente se midió la absorbancia de la muestra a
una longitud de onda de 228 nm en un espectrofotómetro Lambda XLS (Perkin Elmer, USA). La
actividad inhibitoria de la enzima convertidora de la angiotensina (I-ECA) se determinó por triplicado.
El control positivo contenía al sustrato en ausencia de enzima mientras que el control negativo
contenía al sustrato y la enzima en ausencia de inhibidores. El porciento de I-ECA se determinó
mediante la siguiente ecuación:
Además, se realizó un estudio in vivo utilizando machos de ratas Wistar espontáneamente
hipertensas (RWH) donadas por la Facultad de Estudios Superiores de Iztacala de la UNAM. Todos los
procedimientos con los animales se llevaron a cabo de acuerdo con lo estipulado por SAGARPA
(NOM-062-ZOO-1999, Mexico). Inicialmente las ratas Wistar hipertensas tenían las siguientes
características: masa = 205 ± 11.8 g, frecuencia cardiaca = 468.7 ± 27.3 lpm, presión sanguínea
sistólica (PS) = 188.7 ± 8.71 mmHg, y presión sanguínea diastólica (PD) = 148.41 ± 10.06. Las ratas se
mantuvieron a 24 °C con ciclos de luz y oscuridad de 12 h y libre acceso a alimento (Rodent
Laboratory Chow 5001 Reg SAGARPA A-0207-171, México) y agua.
Para la determinación de la actividad hipotensiva de los péptidos de frijol, se utilizó la fracción
f3-10 del FAH. Y las RWH se dividieron en tres grupos: el grupo control negativo (agua destilada 2.5
mL/kg), el grupo administrado con el péptido (FAH f3-10 en 4 mg/kg), y el grupo control positivo
(Captopril® 10 mg/kg). La presión sanguínea de los animales se midió por triplicado en la cola de las
RWH con un equipo automático LE 5007 Letica1 (PanLab, España), a las 0, 1, 2, 4, y 6 h después de
haber administrado interperitonealmente la dosis correspondiente a cada uno. Los datos fueron
tratados estadísticamente y presentados como la diferencia en la presión sanguínea (PS) (Li y col.,
2006a).

Materiales y Métodos
54
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
4.14 Purificación de las fracciones peptídicas con actividad antioxidante
Las fracciones entre 1 - 3 kDa de las tres variedades de frijol (FNP, FAH y FPS) fueron separadas
a una longitud de onda de 214 nm en una columna C-18 (small pore, Grace Vydac, USA) adaptada a
un cromatógrafo de líquidos de alta eficacia HPLC LC-20AT (Shimadzu, Japón), de las que se corrieron
100 µL de la muestra (1 mg/mL) en un programa en gradiente de acetonitrilo/TFA 0.1% (ácido
trifluoroacético). Se colectaron las muestras cada 5 min y se colocaron en un sistema SpeedVac para
la eliminación de solvente y concentración de la muestra (Srihongthong y col., 2012). A cada grupo se
le determinó su capacidad antioxidante como apagamiento del radical luminol, según el método
descrito en 4.12.4 (Georgetti y col., 2003). Los grupos que resultaron con actividad fueron inyectados
al cromatógrafo nuevamente y los picos fueron colectados individualmente. Se concentró la muestra
y se realizó el análisis de capacidad antioxidante a cada uno de los picos. El pico que resultó con
mayor actividad antioxidante fue analizado por espectrometría de masas.
4.15 Determinación de la masa molecular del péptido con actividad antioxidante
La determinación de la masa molecular del péptido con mayor actividad antioxidante obtenido
según el protocolo descrito en la sección 4.14 se llevó a cabo en un espectrómetro de masas MALDI-
TOF MS siguiendo método descrito por Alemán y col. (2011). Por duplicado se colocó 1 µL de muestra
sobre una placa de MALDI a la cual se le permitió secar a temperatura ambiente. En seguida, se
agregaron 0.4 µL de la matriz (3 mg/mL de ácido α-ciano-4-hidrixi-transcinámico en 50 % (v/v) de
acetonitrilo) y se le permitió secar antes de introducir la placa en un equipo 4800 Proteomics Analyzer
MALDI-TOF/TOF (Applied Biosystems, USA) donde la masa fue determinada operando con un
reflector en modo positivo a un voltaje de 20,000 V.
4.16 Determinación de la secuencia de aminoácidos por bioinformática del péptido con
actividad antioxidante
La determinación de la secuencia de aminoácidos del péptido aislado por cromatografía de
líquidos de alta eficacia y que presentó la máxima actividad antioxidante por el método de
quimioluminiscencia, se realizó mediante el sistema ExPASy (Bioinformatics Resource Portal, Suiza)
con la herramienta Find Peptide. El ingreso de los datos fue con base en la secuencia de aminoácidos
de la faseolina (UniProt: P02853) que es la proteína que se encuentra mayoritariamente en las
semillas de frijol, además se realizó el mismo procedimiento con la proteína lectina (UniProt:

Materiales y Métodos
55
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Q8RVH1) que es la proteína que le secunda a la faseolina en la composición proteíca del frijol; y las
masas moleculares obtenidas en el experimento anterior (4.15).
4.17 Análisis estadístico de datos
Se realizaron análisis de varianza (ANOVA por sus siglas en inglés) de una vía y análisis de
varianza dos vías para los procedimientos descritos en esta sección (según la necesidad), y de acuerdo
con los diseños experimentales de Montgomery (2006), posteriormente se llevó a cabo la
comparación de medias por Duncan a un nivel de significancia de α = 5 % (Duncan, p ≤ 0.05).
Además, para el estudio in vivo se realizó el ANOVA de una vía, seguido de un estudio de t-
Student a un nivel de significancia de α = 1 y 0.1 % (t-Student, p ≤ 0.01 o p ≤ 0.001).
Ambos análisis se llevaron a cabo mediante el paquete estadístico SAS system 9.0 (SAS Institute
Inc., USA) y los gráficos fueron realizados con el programa GraphPad Prism 4.0 (GraphPad Software
Inc., USA).

Resultados y discusión
56
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5. Resultados y Discusión
5.1 Evaluación y caracterización física de las semillas de frijol
Las semillas de frijol fueron evaluadas según lo descrito en 4.2 (Cuadro 5-1). De acuerdo con
López-Herrera (2002) si la masa de 100 semillas es menor de 25 g la especie se considera de grano
pequeño, entre 25 y 40 g de grano mediano y si supera los 40 g de grano grande. Por lo que las
semillas de frijol de la variedad negro plus (FNP) fueron clasificadas como especies de grano pequeño,
las de frijol pinto Saltillo (FPS) como especies de grano mediano y las de frijol azufrado higuera (FAH)
como especies de grano grande (Figura 5-1).
Cuadro 5-1. Características físicas de las semillas de las tres variedades de frijol
Característica Variedades de frijol (Phaseolus vulgaris L.)
Negro plus Azufrado higuera Pinto Saltillo
Masa (g) 0.2197 ± 0.0298 c 0.4619 ± 0.0716 a 0.2858 ± 0.0414 b
Largo (cm) 1.01 ± 0.07 c 1.29 ± 0.08 a 1.18 ± 0.07 b
Alto (cm) 0.70 ± 0.05 b 0.76 ± 0.05 a 0.69 ± 0.03 b
Ancho (cm) 0.44 ± 0.04 c 0.64 ± 0.04 a 0.47 ± 0.04 b
Esfericidad (cm3) 0.6706 ± 0.0392 a 0.6634 ± 0.0259 a 0.6156 ± 0.0215 b
En el cuadro se presentan los resultados (promedio ± desviación estándar) de la medición de cincuenta semillas. a, b, c
Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p<0.05)
Figura 5-1. Semilla de frijol de las tres variedades estudiadas en este trabajo. Dónde: a) frijol negro plus, b) frijol azufrado higuera, y c) frijol pinto Saltillo.
a)
b)
c)
Resultados y discusión
57
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Además, se observó de manera general que la variedad FAH es estadísticamente mayor en sus
dimensiones y masa que las variedades FNP y FPS (Duncan, p ≤ 0.05). Ya que los parámetros de
esfericidad determinados fueron cercanos a 0.6, y las semillas de las leguminosas pueden tener muy
variadas formas (lenticular, ovoide, redondeada, acorazonada, reniforme, arqueada, prismática,
globosa, etc); entonces se trata de semillas ovides ligeramente redondeadas; esta forma es aparente
la más común en las legumbres (White y González, 1990). Shahin y col. (2006) realizaron estas
mismas determinaciones con semillas de soya y reportaron esta misma forma en las semillas, incluso
las comparan con la forma de un riñon.
5.2 Obtención de las harinas de frijol
Se procesaron 1 kg de semillas de frijol de cada variedad. Durante la obtención de harina de
frijol (P. vulgaris L.) se realizó una primera molienda y un primer tamizado por malla No. 40 (0.42
mm), en éste procesos las tres variedades de frijol presentaron rendimientos alrededor de: FNP =
50.98 %, FAH = 47.53 % y FPS = 66.13 %. Durante el proceso de eliminación de grasas con hexano y el
segundo tamizado por malla No. 80 (0.18 mm), el rendimiento de las harinas disminuyó aún más
(Cuadro 5-2). Por lo que, los rendimientos finales fueron entre 31 y 38 %; ésta harina desengrasada
de tamaño igual o menor de 0.18 mm fue almacenada en bolsas de cierre hermético para su posterior
caracterización y uso en la preparación de los concentrados proteicos de frijol (FNP, FAH y FPS); y
demás experimentos de este trabajo.
Cuadro 5-2. Masas y rendimientos de las harinas de frijol después de la molienda y tamizado.
Variedad de frijol Masa inicial
(g)
Masa de harina malla No. 40
(g)
Masa de harina desengrasada
malla No. 80 (g)
Rendimiento
(%)
Negro plus 1020 520 320 31.37
Azufrado higuera 1200 570.4 390.2 32.52
Pinto Saltillo 1100 727.4 420.4 38.22
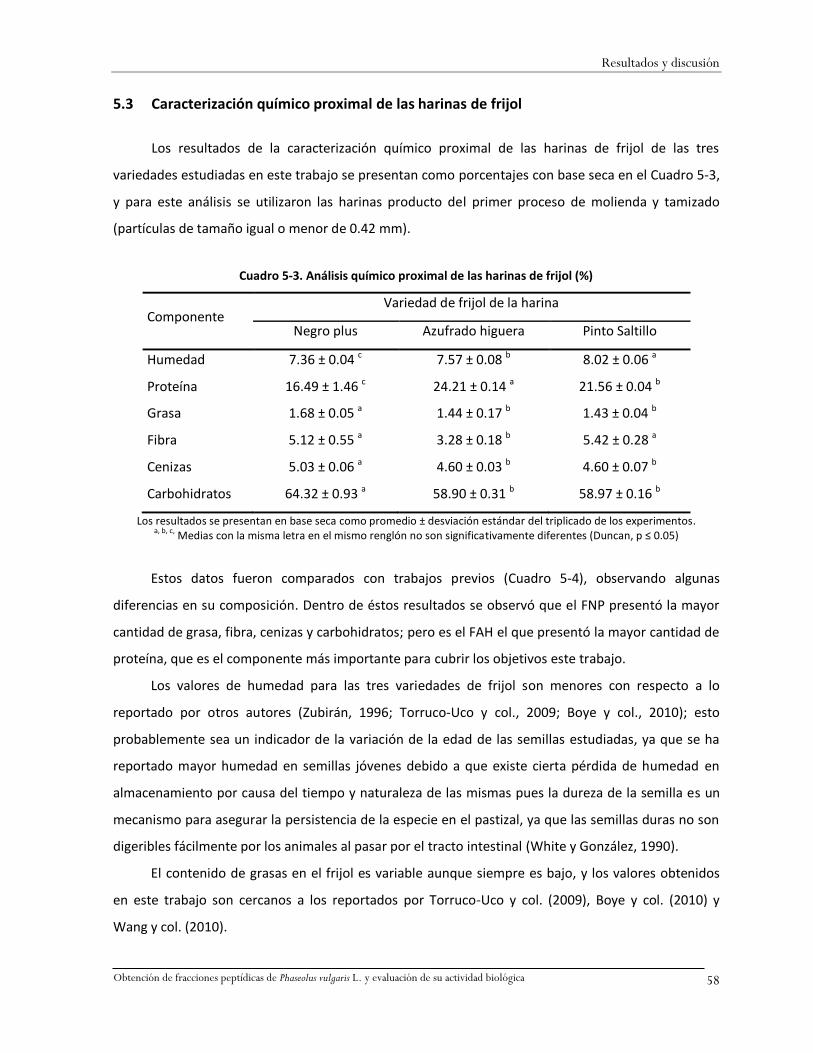
Resultados y discusión
58
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.3 Caracterización químico proximal de las harinas de frijol
Los resultados de la caracterización químico proximal de las harinas de frijol de las tres
variedades estudiadas en este trabajo se presentan como porcentajes con base seca en el Cuadro 5-3,
y para este análisis se utilizaron las harinas producto del primer proceso de molienda y tamizado
(partículas de tamaño igual o menor de 0.42 mm).
Cuadro 5-3. Análisis químico proximal de las harinas de frijol (%)
Componente Variedad de frijol de la harina
Negro plus Azufrado higuera Pinto Saltillo
Humedad 7.36 ± 0.04 c 7.57 ± 0.08 b 8.02 ± 0.06 a
Proteína 16.49 ± 1.46 c 24.21 ± 0.14 a 21.56 ± 0.04 b
Grasa 1.68 ± 0.05 a 1.44 ± 0.17 b 1.43 ± 0.04 b
Fibra 5.12 ± 0.55 a 3.28 ± 0.18 b 5.42 ± 0.28 a
Cenizas 5.03 ± 0.06 a 4.60 ± 0.03 b 4.60 ± 0.07 b
Carbohidratos 64.32 ± 0.93 a 58.90 ± 0.31 b 58.97 ± 0.16 b
Los resultados se presentan en base seca como promedio ± desviación estándar del triplicado de los experimentos. a, b, c,
Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p ≤ 0.05)
Estos datos fueron comparados con trabajos previos (Cuadro 5-4), observando algunas
diferencias en su composición. Dentro de éstos resultados se observó que el FNP presentó la mayor
cantidad de grasa, fibra, cenizas y carbohidratos; pero es el FAH el que presentó la mayor cantidad de
proteína, que es el componente más importante para cubrir los objetivos este trabajo.
Los valores de humedad para las tres variedades de frijol son menores con respecto a lo
reportado por otros autores (Zubirán, 1996; Torruco-Uco y col., 2009; Boye y col., 2010); esto
probablemente sea un indicador de la variación de la edad de las semillas estudiadas, ya que se ha
reportado mayor humedad en semillas jóvenes debido a que existe cierta pérdida de humedad en
almacenamiento por causa del tiempo y naturaleza de las mismas pues la dureza de la semilla es un
mecanismo para asegurar la persistencia de la especie en el pastizal, ya que las semillas duras no son
digeribles fácilmente por los animales al pasar por el tracto intestinal (White y González, 1990).
El contenido de grasas en el frijol es variable aunque siempre es bajo, y los valores obtenidos
en este trabajo son cercanos a los reportados por Torruco-Uco y col. (2009), Boye y col. (2010) y
Wang y col. (2010).
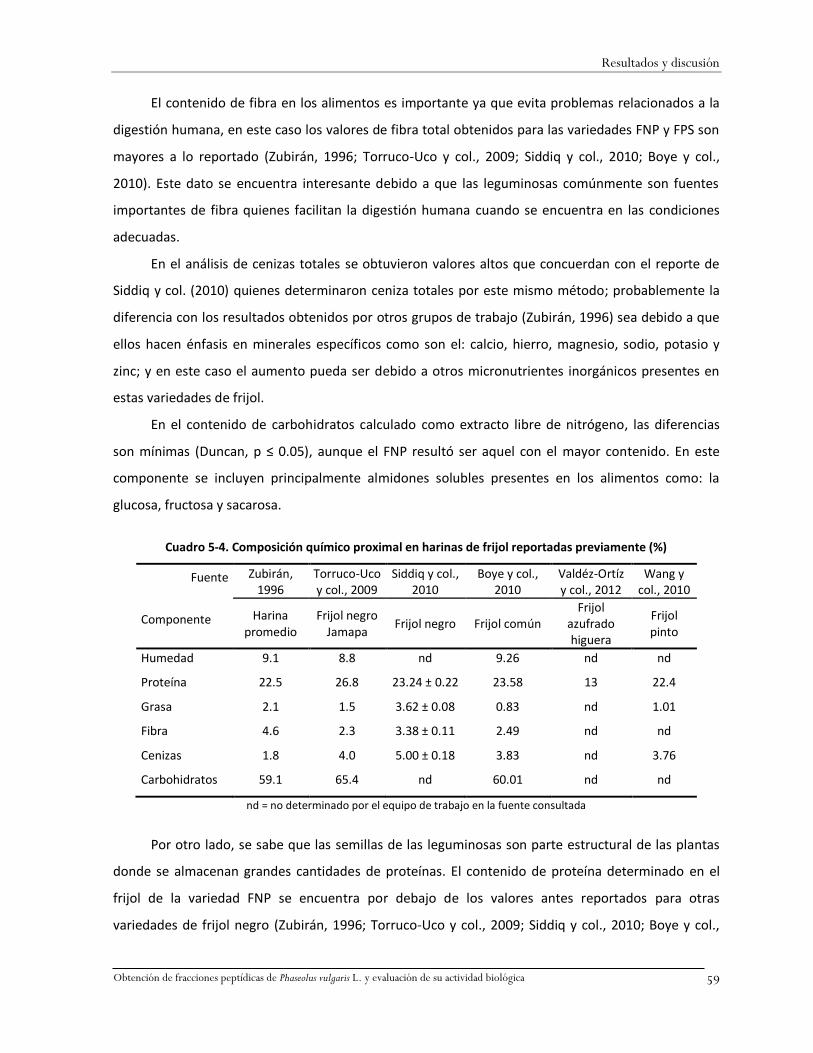
Resultados y discusión
59
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
El contenido de fibra en los alimentos es importante ya que evita problemas relacionados a la
digestión humana, en este caso los valores de fibra total obtenidos para las variedades FNP y FPS son
mayores a lo reportado (Zubirán, 1996; Torruco-Uco y col., 2009; Siddiq y col., 2010; Boye y col.,
2010). Este dato se encuentra interesante debido a que las leguminosas comúnmente son fuentes
importantes de fibra quienes facilitan la digestión humana cuando se encuentra en las condiciones
adecuadas.
En el análisis de cenizas totales se obtuvieron valores altos que concuerdan con el reporte de
Siddiq y col. (2010) quienes determinaron ceniza totales por este mismo método; probablemente la
diferencia con los resultados obtenidos por otros grupos de trabajo (Zubirán, 1996) sea debido a que
ellos hacen énfasis en minerales específicos como son el: calcio, hierro, magnesio, sodio, potasio y
zinc; y en este caso el aumento pueda ser debido a otros micronutrientes inorgánicos presentes en
estas variedades de frijol.
En el contenido de carbohidratos calculado como extracto libre de nitrógeno, las diferencias
son mínimas (Duncan, p ≤ 0.05), aunque el FNP resultó ser aquel con el mayor contenido. En este
componente se incluyen principalmente almidones solubles presentes en los alimentos como: la
glucosa, fructosa y sacarosa.
Cuadro 5-4. Composición químico proximal en harinas de frijol reportadas previamente (%)
Fuente Zubirán, 1996
Torruco-Uco y col., 2009
Siddiq y col., 2010
Boye y col., 2010
Valdéz-Ortíz y col., 2012
Wang y col., 2010
Componente Harina promedio
Frijol negro Jamapa
Frijol negro Frijol común Frijol
azufrado higuera
Frijol pinto
Humedad 9.1 8.8 nd 9.26 nd nd
Proteína 22.5 26.8 23.24 ± 0.22 23.58 13 22.4
Grasa 2.1 1.5 3.62 ± 0.08 0.83 nd 1.01
Fibra 4.6 2.3 3.38 ± 0.11 2.49 nd nd
Cenizas 1.8 4.0 5.00 ± 0.18 3.83 nd 3.76
Carbohidratos 59.1 65.4 nd 60.01 nd nd
nd = no determinado por el equipo de trabajo en la fuente consultada
Por otro lado, se sabe que las semillas de las leguminosas son parte estructural de las plantas
donde se almacenan grandes cantidades de proteínas. El contenido de proteína determinado en el
frijol de la variedad FNP se encuentra por debajo de los valores antes reportados para otras
variedades de frijol negro (Zubirán, 1996; Torruco-Uco y col., 2009; Siddiq y col., 2010; Boye y col.,

Resultados y discusión
60
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
2010), y es probablemente debido a características propias de la variedad. La variedad de frijol FAH
presentó mayor cantidad de proteína que la misma variedad reportada previamente por Valdéz-Ortíz
y col. (2012). Mientras que el contenido de proteína en la variedad FPS coincidió con lo reportado por
Wang y col. (2010). Existe diferencia significativa en el contenido de proteína de las tres variedades
de frijol estudiadas, siendo el FAH la variedad con mayor contenido de proteína y FNP la variedad con
menor contenido (Duncan, p ≤ 0.05).
5.4 Obtención del concentrado de proteínas de frijol
Los concentrados de proteína de frijol obtenidos por solubilización y precipitación isoeléctrica
fueron congelados, liofilizados y almacenados para posteriores análisis. El contenido de proteína en
los concentrados proteicos de frijol fue determinado por el método de micro-Kjeldahl y los resultados
se presentan en el Cuadro 5-5. De acuerdo con lo reportado, un concentrado proteico debe contener
un mínimo de 70 % de proteína en base seca y se extrae a partir de harina desengrasada obteniendo
un producto rico en azúcares insolubles y proteínas, y un aislado proteico supone una serie de etapas
encaminadas a eliminar o disminuir los componentes no proteicos para lograr un producto final con
un mínimo del 80 % de proteínas (Vioque y col., 2001; De Luna-Jiménez, 2007).
Cuadro 5-5. Contenido de proteína en los concentrados de frijol (%)
Variedad de frijol
Muestra Negro plus Azufrado higuera Pinto Saltillo
Harina 16.49 ± 1.45 Bc 24.21 ± 0.14 Ba 21.55 ± 0.04 Bb
Concentrado 79.49 ± 2.30 Aab 82.35 ± 0.82 Aa 78.41 ± 1.78 Ab
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. A,B,
Medias con la misma letra en la misma columna no son significativamente diferentes (Duncan, p ≤ 0.05).
a, ab, b, c, Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p ≤ 0.05).
De acuerdo con las definiciones anteriores, los resultados obtenidos lograron la concentración
de la proteína constituyente de las variedades FNP, FAH y FPS hasta niveles alrededor del 80 %. El
contenido de proteína en el concentrado proteico de FNP incrementó 4.82 veces con respecto a la
harina, el FHA 3.4 veces y el FPS 3.64 veces, esto es importante ya que refleja la capacidad de
eliminación de almidón y otros carbohidratos solubles de la harina de frijol principalmente de la
variedad FNP. Según el análisis de medias existe diferencia significativa entre harinas y concentrados
(Duncan, p ≤ 0.05); y también hay diferencia significativa entre los concentrados de proteína de las
tres variedades (Duncan, p ≤ 0.05).
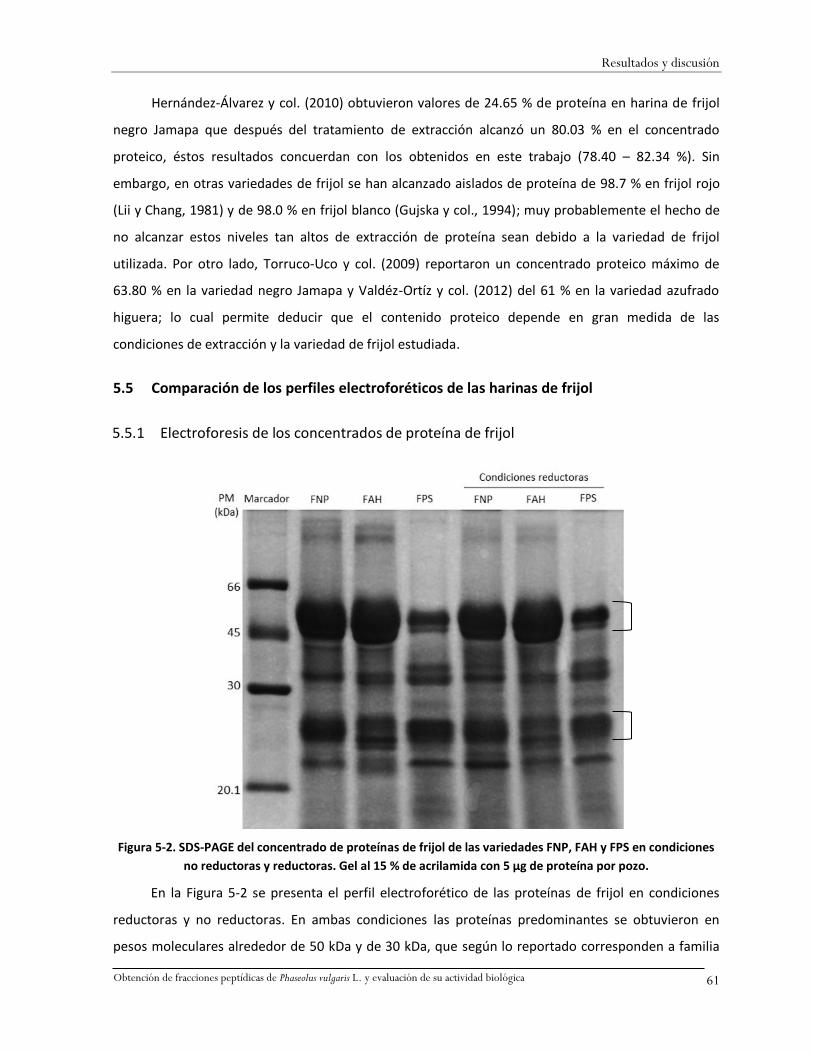
Resultados y discusión
61
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Hernández-Álvarez y col. (2010) obtuvieron valores de 24.65 % de proteína en harina de frijol
negro Jamapa que después del tratamiento de extracción alcanzó un 80.03 % en el concentrado
proteico, éstos resultados concuerdan con los obtenidos en este trabajo (78.40 – 82.34 %). Sin
embargo, en otras variedades de frijol se han alcanzado aislados de proteína de 98.7 % en frijol rojo
(Lii y Chang, 1981) y de 98.0 % en frijol blanco (Gujska y col., 1994); muy probablemente el hecho de
no alcanzar estos niveles tan altos de extracción de proteína sean debido a la variedad de frijol
utilizada. Por otro lado, Torruco-Uco y col. (2009) reportaron un concentrado proteico máximo de
63.80 % en la variedad negro Jamapa y Valdéz-Ortíz y col. (2012) del 61 % en la variedad azufrado
higuera; lo cual permite deducir que el contenido proteico depende en gran medida de las
condiciones de extracción y la variedad de frijol estudiada.
5.5 Comparación de los perfiles electroforéticos de las harinas de frijol
5.5.1 Electroforesis de los concentrados de proteína de frijol
Figura 5-2. SDS-PAGE del concentrado de proteínas de frijol de las variedades FNP, FAH y FPS en condiciones
no reductoras y reductoras. Gel al 15 % de acrilamida con 5 µg de proteína por pozo.
En la Figura 5-2 se presenta el perfil electroforético de las proteínas de frijol en condiciones
reductoras y no reductoras. En ambas condiciones las proteínas predominantes se obtuvieron en
pesos moleculares alrededor de 50 kDa y de 30 kDa, que según lo reportado corresponden a familia

Resultados y discusión
62
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
de las faseolinas y lectinas respectivamente (Camacho-Espinoza y col., 2010), éstos dos grupos
principales de proteínas se observaron en las tres variedades de frijol. Emani y Hall (2008) reportaron
que el 50 % de las proteínas contenidas en el frijol pertenecen al grupo de las faseolinas, las cuales a
su vez difieren en el contenido del aminoácido asparagina (Asn) lo que hace que se formen tres
estructuras de diferentes masas muy cercanas: α (51,000-53,000 Da), β (47,000-48,000 Da) y
(43,000-46,000 Da).
No se observa diferencia entre los patrones electroforéticos de las proteínas con y sin agente
desnaturalizante (-mercaptoetanol), lo cual coincide con lo reportado por Camacho-Espinoza y col.
(2010) quienes trabajaron con las variedades Azufrado regional 87, azufrado higuera y azufrado
noroeste. Con las pocas diferencias observadas, se podría deducir que es probable que las proteínas
de estas variedades de frijol no se encuentren unidas en gran medida por puentes disulfuro. Además,
es importante mencionar que la variedad FPS aparentemente contiene menor cantidad de proteínas
del grupo de las faseolinas que las variedades FNP y FAH, lo que indicaría menor probabilidad de
encontrar Asn en ésta variedad; aún más, se observaron dos bandas de peso molecular mayor de 30
kDa que no son visibles en las otras dos variedades. Por lo que, es probable que la variación en el
perfil electroforético de los concentrados de proteína de frijol, dependa del contenido de
aminoácidos de cada variedad, a sus condiciones de cultivo y otras características propias de la
semilla.
5.5.2 Electroforesis bidimensional de los concentrados de proteína de frijol
Mediante la técnica de electroforesis bidimensional se observó un grupo importante de
proteínas con pI cercanos a 5 y algunos puntos en pIs menores a 5 en el proceso de isoelectroenfoque
(1D), los puntos de proteínas observadas presentaron pesos moleculares mayores de 40 kDa al
correrse la segunda dimensión de la electroforesis (Figura 5-3).
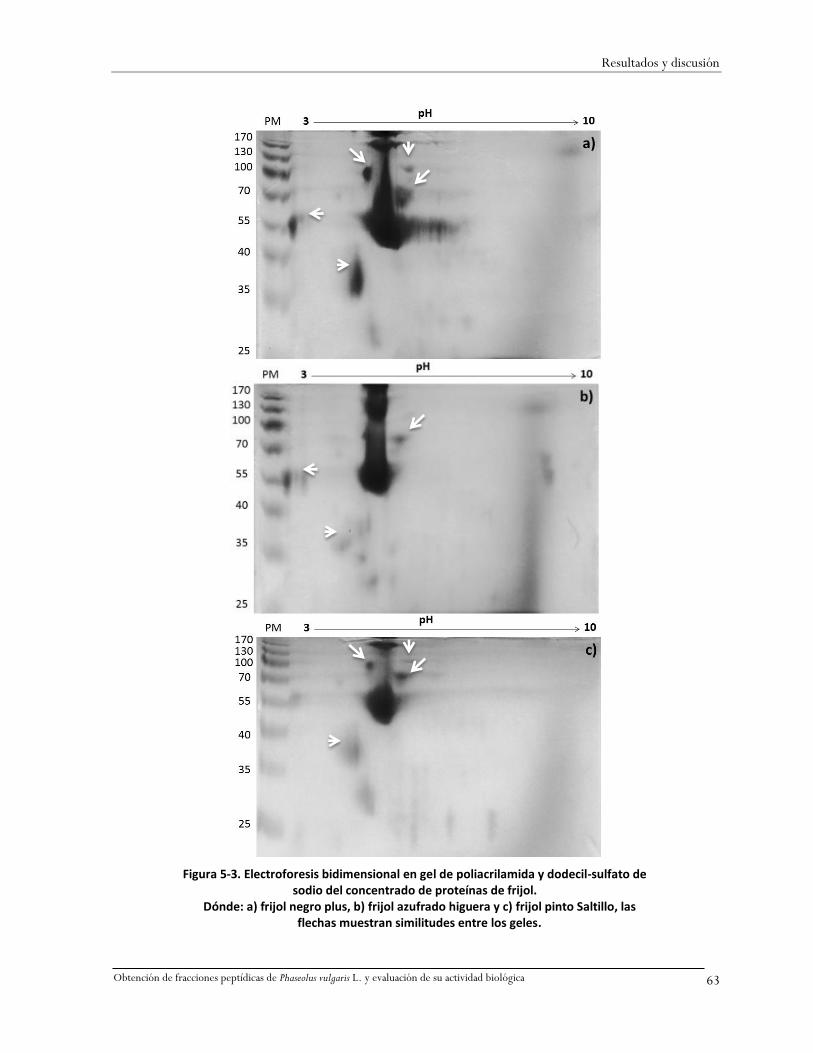
Resultados y discusión
63
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-3. Electroforesis bidimensional en gel de poliacrilamida y dodecil-sulfato de sodio del concentrado de proteínas de frijol.
Dónde: a) frijol negro plus, b) frijol azufrado higuera y c) frijol pinto Saltillo, las flechas muestran similitudes entre los geles.

Resultados y discusión
64
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
En la electroforesis bidimensional de las muestras de FNP y FAH se observó corrimiento en los
puntos con mayor saturación de proteína de frijol en el gel; Görg y col. (2000) atribuyen este
comportamiento a la concentración de proteínas de la misma familia quienes tienen en su mayoría
las mismas características y sugieren tomar en cuenta el gradiente de inmovilización para la
optimización del ensayo. Por otra parte, Camacho-Espinoza y col. (2010) reportaron este tipo de
puntos con corrimiento como resultado de una posible contaminación con ácidos nucleicos o
azúcares que impiden la correcta separación de las proteínas, y sugieren el uso del kit 2D Clean-Up
(Amersham Biosciences, USA), y este kit fue usado en el tratamiento previo de éstas proteínas de
frijol. Estos mismos autores reportan la presencia de proteínas de FAH con PM cercano a 50 kDa, lo
que resulta similar a lo obtenido en este trabajo.
Además de este grupo mayoritario de proteínas, se observaron tres puntos similares entre el
FNP y el FAH, cuatro puntos entre el FNP y el FPS, y dos entre el FAH y el FPS. Cada uno de estos
puntos representa una proteína diferente que podría ser analizada mediante un densitómetro y un
software adecuado, tal como lo realizaron Egito y col. (2002) para la separación e identificación
proteómica de α-, β-, κ- y, γ- caseínas en cinco péptidos derivados de leche. Además, utilizando éste
tipo de herramientas Aguilar y col. (2009) también lograron la separación de proteínas de soya y las
compararon con proteínas conocidas (α-lactoalbúmina, BSA y quimotripsina).
Por otro lado, Natarajan y col. (2006) usaron la electroforesis bidimensional en tres diferentes
intervalos de pH (3 - 10, 4 - 7 y 6 - 11) para comparar la composición proteínica de semillas de soya en
cultivos nativos (Glycine soja) y comerciales (G. max), y determinaron que el intervalo de pH permite
la localización óptima de las proteínas de ésta leguminosa. Por lo que probablemente las proteínas
contenidas en los concentrados proteicos de las variedades de frijol trabajadas en esta investigación
(FNP, FAH y FPS), pudieran ser separadas con mayor eficiencia en tiras IPG (ReadyStrip, BIO-RAD) con
intervalo de pH 4-7 durante el proceso de isoelectroenfoque, ya que los puntos isoeléctricos de las
proteínas contenidas en el frijol son principalmente ácidas, incluso si se utilizaran tiras de mayor
longitud habría mayor separación. También se propone un análisis de los puntos principales según lo
propuesto por O’Farrell (1975), quién utiliza regletas estándar en la separación de proteínas
contenidas en E. coli.

Resultados y discusión
65
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.6 Hidrólisis enzimática de las proteínas de frijol
5.6.1 Hidrólisis enzimática de las proteínas contenidas en las harinas de frijol
La hidrólisis de las proteínas de frijol se llevó a cabo en harinas al 10 % (p/v) bajo los
parámetros establecidos en la metodología; el contenido de proteína en éstas muestras fue de 1.6 g,
2.4 g y 2.1 g para el FNP, FAH y FPS respectivamente. Y los resultados de la hidrólisis se presentan en
el Cuadro 5-6, donde se observa una diferencia significativa en el grado de hidrólisis según el tiempo
de reacción (Duncan, p ≤ 0.05); obteniéndose el mayor % GH en dos y tres horas; y siendo la variedad
FAH la que alcanza el mayor grado de hidrólisis. Así que es notable la diferencia entre el grado de
hidrólisis de una a dos horas de proceso; pero sin diferencia significativa en el periodo de dos a tres
horas de tratamiento. Sin embargo, los porcentajes del grado de hidrólisis pueden ser considerados
bajos si se comparan con otros trabajos realizados con concentrados de proteína (Valdéz-Ortíz y col.,
2012).
Por otro lado, se ha reportado que la enzima Alcalasa® rompe a las proteínas principalmente en
aminoácidos hidrofóbicos como: Phe, Tyr, Trp, Leu, Ile, Val y Met (Markland y Smith, 1971), por lo que
es probable que éste tipo de reacción haya sido generada durante la hidrólisis de las proteínas de
frijol, aunque aún los carbohidratos solubles interfieren de tal forma que no hay un alto % GH. La
Alcalasa® tiene una ventaja económica ante otras enzimas de grado alimenticio como la pepsina,
tripsina, quimotripsina, papaina y termolisina; todas ellas con la capacidad de ser aprovechadas para
la obtención de péptidos activos (Li y col., 2005); y otro punto favorable en ésta enzima es su
capacidad para obtener péptidos de menos de 15 residuos de aminoácidos, en los cuales se ha
demostrado actividad inhibitoria de la enzima convertidora de la angiotensina (Li y col., 2004).
Cuadro 5-6. Grado de hidrólisis alcanzado por las harinas de frijol durante el tratamiento (%)
Variedad 1 h 2 h 3 h
Negro plus 8.91 ± 0.79 Bb 21.25 ± 1.61 Aa 21.80 ± 1.43 Aa
Azufrado higuera 14.28 ± 3.09 Ab 22.04 ± 2.33 Aa 23.54 ± 0.82 Aa
Pinto Saltillo 12.94 ± 1.07 Ab 21.28 ± 2.50 Aa 22.25 ± 2.01 Aa
Los resultados se presentan en base seca como promedio ± desviación estándar del triplicado de los experimentos. a,b
. Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p<0.05).
A,B. Medias con la misma letra en la misma columna no son significativamente diferentes (Duncan, p<0.05).

Resultados y discusión
66
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.6.2 Hidrólisis enzimática de los concentrados proteicos de frijol
Se llevó a cabo el proceso de hidrólisis enzimática de los concentrados proteicos (liofilizados de
proteína de frijol) al 4 % (p/v), con contenido de proteína de 3.18 g, 3.29 g y 3.14 g para el FNP, FAH y
FPS, respectivamente. A partir de estas hidrólisis se obtuvieron los resultados presentados en el
Cuadro 5-7, donde se observa un incremento en el grado de hidrólisis con respecto a los hidrolizados
de harinas para las tres variedades de frijol (FNP, FAH y FPS). Esto indica que es posible obtener un
mayor grado de hidrólisis bajo las mismas condiciones de proceso, siempre y cuando la proteína se
encuentre más concentrada y libre de otros compuestos (almidones y carbohidratos solubles,
principalmente). En este caso los grados de hidrólisis no son mayores del 24.71 %, aunque
Hernández-Álvarez y col. (2010) reportaron un grado de hidrólisis máximo de 35.24 % para frijol
negro Jamapa en 60 min de reacción determinado como porcentaje de nitrógeno soluble en 10 % de
TCA (Kim y col., 1990). Por su parte Valdéz-Ortíz y col. (2012) determinaron un grado de hidrólisis de
60 % en el concentrado de la variedad azufrado higuera y hasta del 73 % para la variedad azufrado
noroeste en 120 min de reacción; y en otras semillas como la semilla de girasol se reportó el grado de
hidrólisis máximo de 18.8 % (Villanueva y col., 1999). Por lo que los resultados obtenidos en este
trabajo se encuentran en el intervalo obtenido en otros trabajos similares.
Cuadro 5-7. Grado de hidrólisis obtenido en 2 h de reacción con los concentrados de proteína de frijol (%)
Variedad Harina de frijol Concentrado proteico
Negro plus 21.25 ± 1.61 Aa
22.59 ± 0.44 Ba
Azufrado higuera 22.04 ± 2.33 Aa 24.72 ± 0.83 Aa
Pinto Saltillo 21.28 ± 2.50 Aa
22.49 ± 0.72 Ba
Los resultados se presentan en base seca como promedio ± desviación estándar del triplicado de los experimentos. a,b
. Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p<0.05).
A,B. Medias con la misma letra en la misma columna no son significativamente diferentes (Duncan, p<0.05).
Para comprobar que el proceso de hidrólisis separó el grupo mayoritario de proteínas de frijol
(probablemente el de las faseolinas 50 kDa y en menor cantidad el grupo de las lectinas 30 kDa), se
elaboró un gel de electroforesis con docecil-sulfato de sodio de los concentrados proteicos y los
hidrolizados totales de frijol (Figura 5-4). En ésta imagen se puede observar que el grupo mayoritario
de proteínas (mayor de 45 kDa) y otro menor (mayor de 30 kDa) presente en las tres variedades de
frijol se ha hidrolizado, dando como resultado proteínas y péptidos con pesos moleculares menores
de 30 kDa.
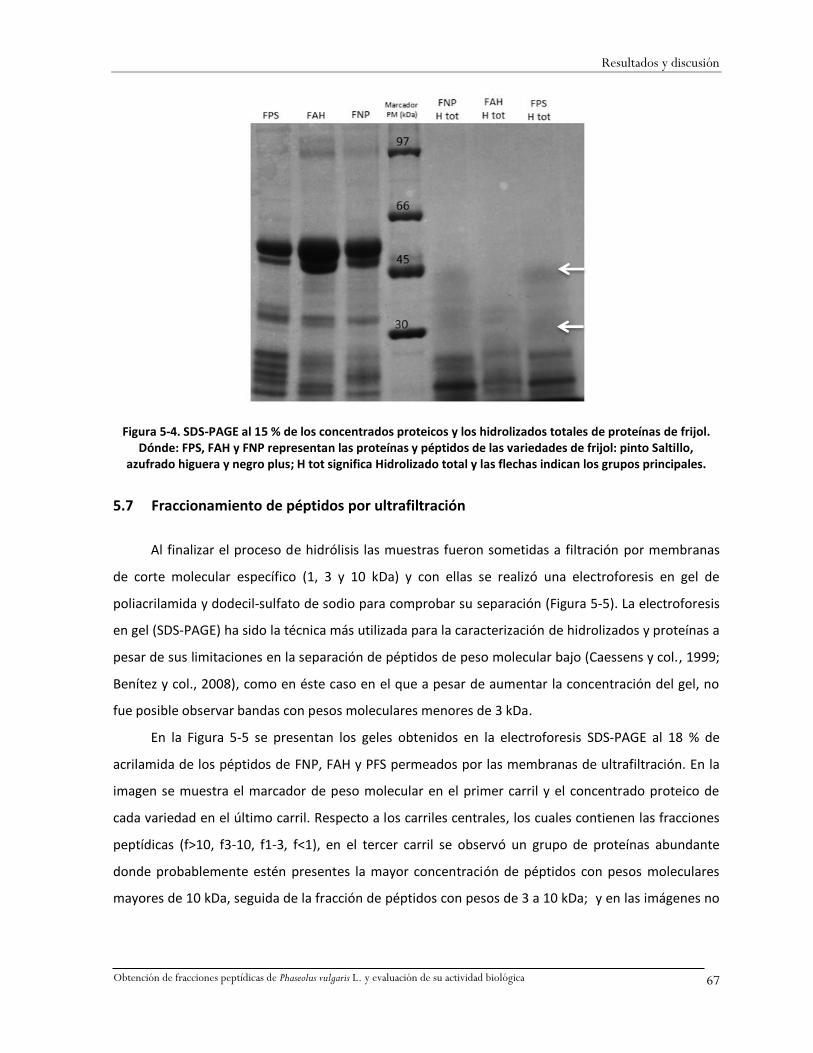
Resultados y discusión
67
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-4. SDS-PAGE al 15 % de los concentrados proteicos y los hidrolizados totales de proteínas de frijol. Dónde: FPS, FAH y FNP representan las proteínas y péptidos de las variedades de frijol: pinto Saltillo,
azufrado higuera y negro plus; H tot significa Hidrolizado total y las flechas indican los grupos principales.
5.7 Fraccionamiento de péptidos por ultrafiltración
Al finalizar el proceso de hidrólisis las muestras fueron sometidas a filtración por membranas
de corte molecular específico (1, 3 y 10 kDa) y con ellas se realizó una electroforesis en gel de
poliacrilamida y dodecil-sulfato de sodio para comprobar su separación (Figura 5-5). La electroforesis
en gel (SDS-PAGE) ha sido la técnica más utilizada para la caracterización de hidrolizados y proteínas a
pesar de sus limitaciones en la separación de péptidos de peso molecular bajo (Caessens y col., 1999;
Benítez y col., 2008), como en éste caso en el que a pesar de aumentar la concentración del gel, no
fue posible observar bandas con pesos moleculares menores de 3 kDa.
En la Figura 5-5 se presentan los geles obtenidos en la electroforesis SDS-PAGE al 18 % de
acrilamida de los péptidos de FNP, FAH y PFS permeados por las membranas de ultrafiltración. En la
imagen se muestra el marcador de peso molecular en el primer carril y el concentrado proteico de
cada variedad en el último carril. Respecto a los carriles centrales, los cuales contienen las fracciones
peptídicas (f>10, f3-10, f1-3, f<1), en el tercer carril se observó un grupo de proteínas abundante
donde probablemente estén presentes la mayor concentración de péptidos con pesos moleculares
mayores de 10 kDa, seguida de la fracción de péptidos con pesos de 3 a 10 kDa; y en las imágenes no
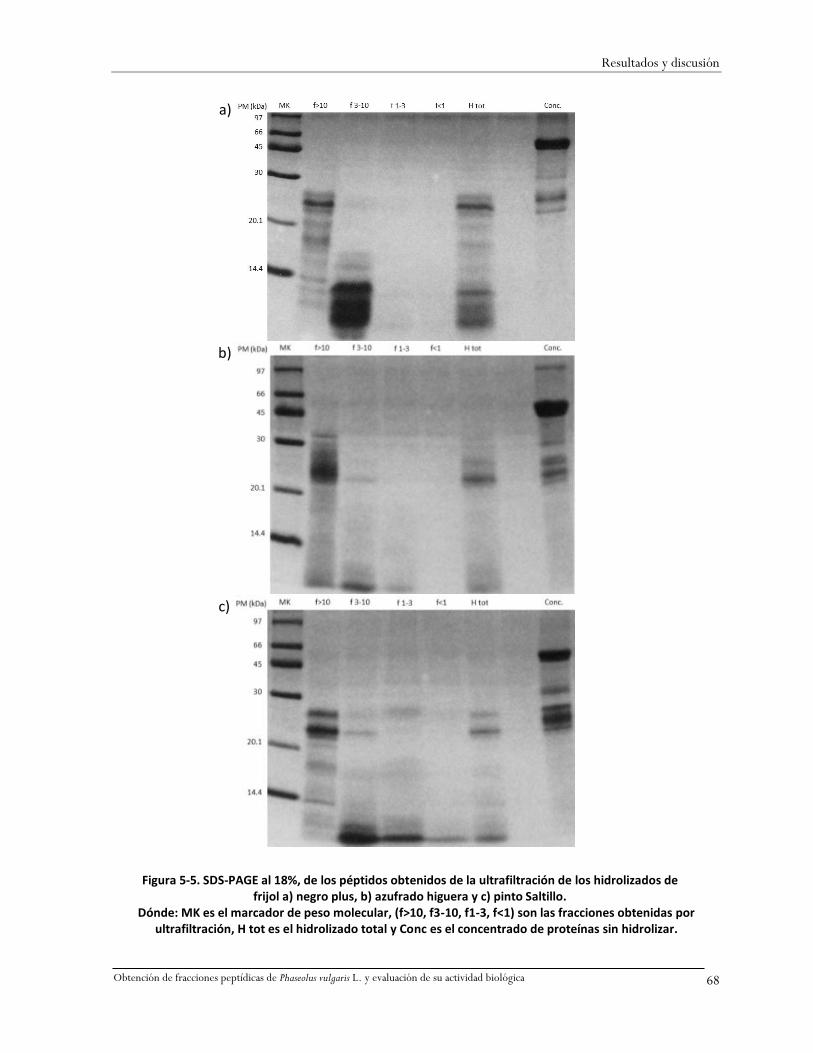
Resultados y discusión
68
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-5. SDS-PAGE al 18%, de los péptidos obtenidos de la ultrafiltración de los hidrolizados de frijol a) negro plus, b) azufrado higuera y c) pinto Saltillo.
Dónde: MK es el marcador de peso molecular, (f>10, f3-10, f1-3, f<1) son las fracciones obtenidas por ultrafiltración, H tot es el hidrolizado total y Conc es el concentrado de proteínas sin hidrolizar.
a)
b)
c)

Resultados y discusión
69
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
se observaron bandas o grupos de proteínas en las fracciones f1-3 y f<1 debido a que son pesos
moleculares muy bajos, y como ya se mencionó ésta es una de las limitaciones principales de la
electroforesis SDS-PAGE (Benítez y col., 2008); por lo cual podría utilizarse otra técnica para
corroborar la presencia de péptidos como fraccionamiento por cromatografía por exclusión de
tamaño y HPLC en fase reversa, o electroforesis capilar (Torres-Llanez y col., 2005)
Para abordar el principal objetivo de este trabajo y comprobar que cada una de éstas
fracciones peptídicas presentaba algún tipo de actividad biológica, se procedió a realizar las pruebas
in vitro de actividad antimicrobiana, actividad antioxidante y actividad inhibidora de la enzima
convertidora de la angiotensina.
5.8 Determinación de la actividad antibacteriana
5.8.1 Controles positivos - Resistencia de las cepas indicadoras a los antibióticos
Cuadro 5-8. Determinación de la resistencia de las cepas a los antibióticos de referencia (control positivo).
Diámetro de los halos de Inhibición de los controles positivos (mm)
Tinción de
Gram
Cloranfenicol (30 µg)
Ampicilina (10 µg)
Halo
Resistencia al antibiótico
Halo Resistencia al
antibiótico
B. subtilis G (+) 33.0 ± 0.4 b Susceptible
46.1 ± 0.1 a Susceptible
B. cereus G (+) 26.2 ± 1.0 a Susceptible
26.7 ± 1.3 a Intermedio
S. aureus (cepa 1) G (+) 32.6 ± 0.6 b Susceptible
46.8 ± 0.5 a Susceptible
S. aureus (cepa 2) G (+) 34.8 ± 0.4
b Susceptible
49.7 ± 0.6
a Susceptible
L. monocytogenes G (+) 33.4 ± 1.3 a Susceptible
33.3 ± 0.4 a Susceptible
Y. enterocolitica G (-) 32.5 ± 0.3 a Susceptible
17.1 ± 0.8 b Susceptible
E. coli G (-) 24.8 ± 0.7 b Susceptible
34.8 ± 0.6 a Susceptible
K. pneumoniae G (-) 34.9 ± 0.5 b Susceptible
44.9 ± 0.2 a Susceptible
K. rhinoscleromatis G (-) 35.4 ± 1.0
b Susceptible
43.6 ± 0.5
a Susceptible
S. typhi G (-) 29.6 ± 0.9 a Susceptible
11.7 ± 0.5 b Intermedio
S. dysenteriae G (-) 34.0 ± 0.3 b Susceptible
48.3 ± 0.3 a Susceptible
E. agglomerans G (-) 34.3 ± 0.3 b Susceptible
45.6 ± 0.4 a Susceptible
Los resultados se presentan como promedio del diámetro en mm ± desviación estándar del triplicado de los experimentos. a,b
. Medias con la misma letra en el mismo renglón no son significativamente diferentes (Duncan, p ≤ 0.05).
Los resultados de la determinación antibacteriana se presentan en el Cuadro 5-8 como
promedio de los diámetros de los halos de inhibición obtenidos para los controles positivos que
fueron cloranfenicol y ampicilina sobre cada una de las cepas probadas como lo indica la metodología

Resultados y discusión
70
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
en la sección 4.11, y clasificadas de acuerdo a su resistencia al antibiótico en susceptible, intermedio
o resistente como lo indica Konneman y col. (2004). De acuerdo a la clasificación estándar se observó
que todas las cepas fueron susceptibles al cloranfenicol como control positivo (antibiótico de
referencia) y casi todas fueron susceptibles a la ampicilina con excepción de B. cereus y S. typhi que
resultaron tener una resistencia intermedia, esto resalta su importancia en el momento que se desea
establecer una relación entre las características de la cepa y su resistencia al antibiótico con los
péptidos obtenidos por hidrólisis de los concentrados proteicos. Entre las cepas analizadas y los
controles positivos existen diferencias significativas (Duncan, p ≤ 0.05) respecto a los halos de
inhibición que presentaron.
De acuerdo con trabajos previos las cepas con mayor susceptibilidad a los antibióticos son las
Gram (-) debido a que su pared celular está conformada por una capa delgada de péptidoglicano
unida a una membrana exterior por lipoproteínas que fácilmente puede ser afectada por enzimas
hidrolíticas, enzimas inactivadoras de antibióticos y proteínas de transporte, como el cado de S. typhi,
aunque B. cereus es una excepción a este comportamiento ya que este patógeno está clasificado
como Gram (+) por lo que teóricamente debería resistir mejor ante los antibióticos.
5.8.2 Actividad antibacteriana en harinas de frijol hidrolizadas
Se realizaron ensayos preliminares con suspensiones de las harinas de frijol en cantidades de
10 y 20 µg de harina en el sensidisco y en ninguno de los casos se observó actividad antibacteriana.
En ninguna de las tres variedades se presentó actividad antibacteriana, ni a ninguna de las
concentraciones; además, no existen datos reportados acerca de actividad antibacteriana en harinas
de ningún tipo de leguminosas. De manera contraria, alrededor del sensidisco con el hidrolizado se
observó una aglomeración de bacterias, probablemente este comportamiento se debió a la alta
cantidad de azucares disponibles (glucosa, fructosa y sacarosa) en las harinas de frijol (alrededor del
60 % según los resultados presentados en el Cuadro 5-3).
5.8.3 Actividad antibacteriana en harinas desengrasadas e hidrolizadas de frijol
Las harinas desengrasadas de las tres variedades de frijol a una concentración del 10 % (p/v)
fueron sometidas a un proceso de hidrólisis enzimática con Alcalasa® seguido de un proceso de
liofilización. Posteriormente este liofilizado fue suspendidos en agua y colocados en concentraciones
de 5, 10 y 20 µg en sensidiscos para su análisis. En este tratamiento, no se observó actividad
antibacteriana en ninguna variedad de frijol ni en ninguna de las concentraciones utilizadas,
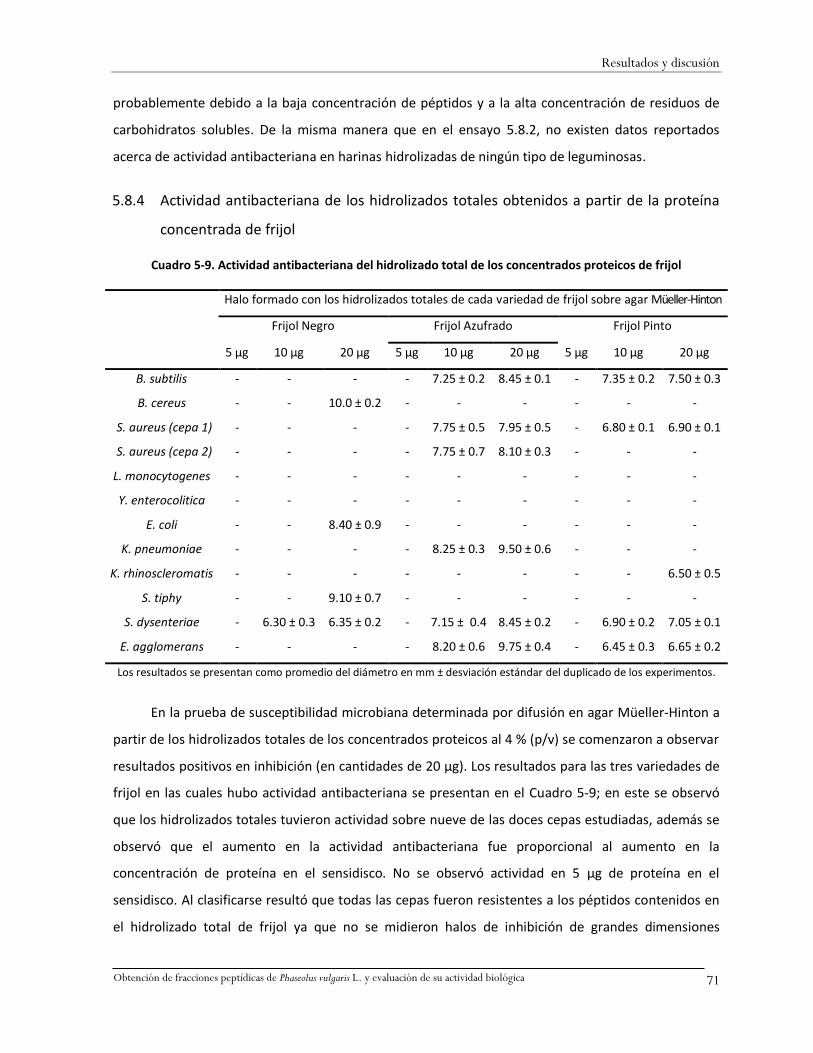
Resultados y discusión
71
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
probablemente debido a la baja concentración de péptidos y a la alta concentración de residuos de
carbohidratos solubles. De la misma manera que en el ensayo 5.8.2, no existen datos reportados
acerca de actividad antibacteriana en harinas hidrolizadas de ningún tipo de leguminosas.
5.8.4 Actividad antibacteriana de los hidrolizados totales obtenidos a partir de la proteína
concentrada de frijol
Cuadro 5-9. Actividad antibacteriana del hidrolizado total de los concentrados proteicos de frijol
Halo formado con los hidrolizados totales de cada variedad de frijol sobre agar Müeller-Hinton
Frijol Negro Frijol Azufrado Frijol Pinto
5 µg 10 µg 20 µg 5 µg 10 µg 20 µg 5 µg 10 µg 20 µg
B. subtilis - - - - 7.25 ± 0.2 8.45 ± 0.1 - 7.35 ± 0.2 7.50 ± 0.3
B. cereus - - 10.0 ± 0.2 - - - - - -
S. aureus (cepa 1) - - - - 7.75 ± 0.5 7.95 ± 0.5 - 6.80 ± 0.1 6.90 ± 0.1
S. aureus (cepa 2) - - - - 7.75 ± 0.7 8.10 ± 0.3 - - -
L. monocytogenes - - - - - - - - -
Y. enterocolitica - - - - - - - - -
E. coli - - 8.40 ± 0.9 - - - - - -
K. pneumoniae - - - - 8.25 ± 0.3 9.50 ± 0.6 - - -
K. rhinoscleromatis - - - - - - - - 6.50 ± 0.5
S. tiphy - - 9.10 ± 0.7 - - - - - -
S. dysenteriae - 6.30 ± 0.3 6.35 ± 0.2 - 7.15 ± 0.4 8.45 ± 0.2 - 6.90 ± 0.2 7.05 ± 0.1
E. agglomerans - - - - 8.20 ± 0.6 9.75 ± 0.4 - 6.45 ± 0.3 6.65 ± 0.2
Los resultados se presentan como promedio del diámetro en mm ± desviación estándar del duplicado de los experimentos.
En la prueba de susceptibilidad microbiana determinada por difusión en agar Müeller-Hinton a
partir de los hidrolizados totales de los concentrados proteicos al 4 % (p/v) se comenzaron a observar
resultados positivos en inhibición (en cantidades de 20 µg). Los resultados para las tres variedades de
frijol en las cuales hubo actividad antibacteriana se presentan en el Cuadro 5-9; en este se observó
que los hidrolizados totales tuvieron actividad sobre nueve de las doces cepas estudiadas, además se
observó que el aumento en la actividad antibacteriana fue proporcional al aumento en la
concentración de proteína en el sensidisco. No se observó actividad en 5 µg de proteína en el
sensidisco. Al clasificarse resultó que todas las cepas fueron resistentes a los péptidos contenidos en
el hidrolizado total de frijol ya que no se midieron halos de inhibición de grandes dimensiones

Resultados y discusión
72
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
(Konneman y col., 2004). El hidrolizado total del FNP formó el halo de inhibición más grande (10 mm),
sin embargo, no es un valor comparable con los controles positivos ya que existe entre ellos
diferencia significativa (Duncan, p ≤ 0.05). La variedad FAH presentó actividad sobre seis de las doce
cepas analizadas, mientras que el FPS sobre cinco y FNP sobre cuatro. En esta prueba preliminar S.
dysenteriae resultó ser la única cepa con susceptibilidad a los hidrolizados de las tres variedades, lo
cual resulta interesante ya que ésta enterobacteria es importante desde el punto de vista de
seguridad alimentaria, pues es causante de diversas enfermedades debido a que produce una
exotoxina termolábil que afecta el intestino (diarrea e inhibición de la absorción de azúcar y amino
ácidos) y el sistema nervioso central (neurotoxina que puede contribuir a la gravedad extrema y
naturaleza mortal de las infecciones) (O’Brien y col., 1982).
5.8.5 Actividad antibacteriana de las fracciones peptídicas de los hidrolizados de los
concentrados proteicos de frijol
Los resultados de la determinación de actividad antibacteriana determinada como
susceptibilidad de las diferentes fracciones obtenidas por ultrafiltración de los hidrolizados totales de
los concentrados proteicos de frijol se presentan en el Cuadro 5-10 para el FAH. En éstos resultados,
se observó que las fracciones peptídicas de FAH presentaron la mayor actividad antibacteriana en la
fracción de péptidos con pesos moleculares de 3 a 10 kDa con halos de inhibición de hasta 10.25 mm
(en K. pneumoniae). Las cepas sobre las cuales se observó susceptibilidad debido a la presencia de
péptidos de la fracción peptídica de 3 a 10 kDa, coinciden con las del Cuadro 5-9, en donde fue
evidente que el FAH es potencialmente un antibacteriano ante ciertas cepas indicadoras; tres de ellas
Gram(-) y dos Gram(+), además la actividad de éstas se vio incrementada al aumentar la
concentración de proteína en el sensidisco. Los mismos análisis se realizaron para las otras dos
variedades de frijol (FPS y FNP). En FPS se observó un halo de 7.0 ± 0.7 mm sobre K. pneumoniae en la
fracción de 3 a 10 kDa y fue la única cepa que presentó susceptibilidad ante los péptidos de esta
variedad de frijol. Mientras que en la variedad FNP no se observó actividad alguna. Tampoco se
observaron halos de inhibición con las fracciones peptídicas de tamaño superior de 10 kDa y menor
de 3 kDa, lo que hace suponer que la amplia gama de péptidos de distintos pesos moleculares
contenidos en los HT de frijol permiten ejercer su actividad antibacteriana sobre las cepas indicadoras
estudiadas en este trabajo.
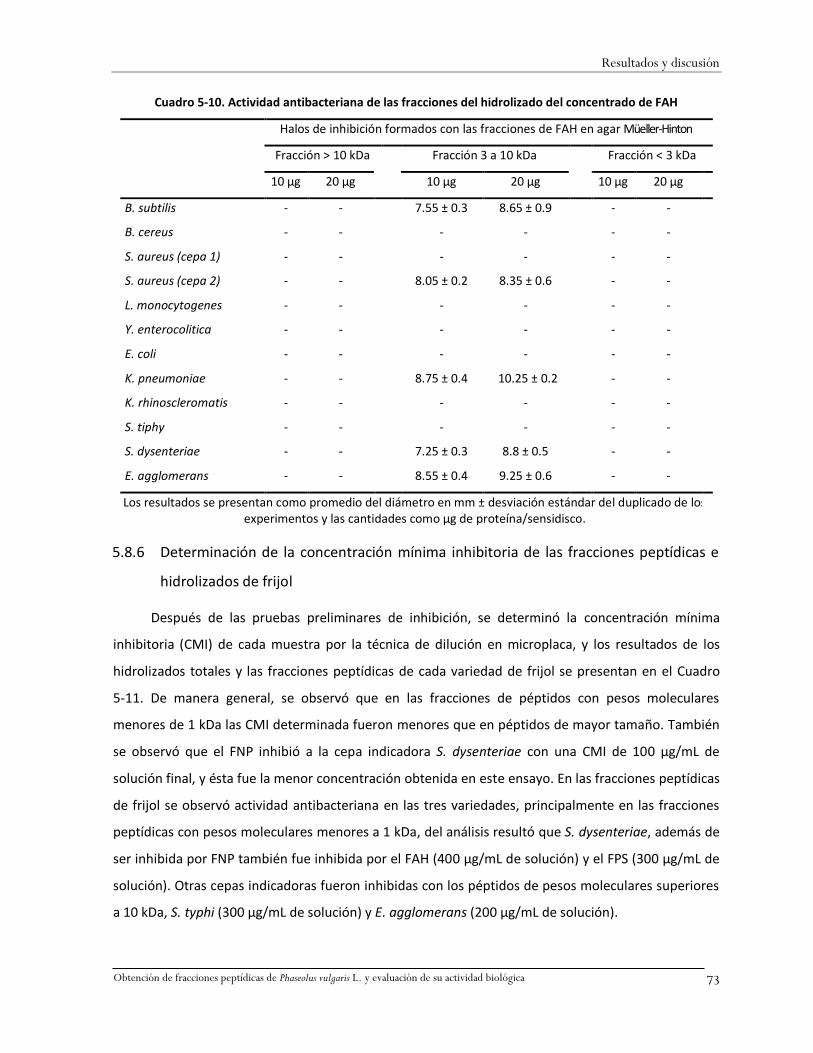
Resultados y discusión
73
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Cuadro 5-10. Actividad antibacteriana de las fracciones del hidrolizado del concentrado de FAH
Halos de inhibición formados con las fracciones de FAH en agar Müeller-Hinton
Fracción > 10 kDa Fracción 3 a 10 kDa Fracción < 3 kDa
10 µg 20 µg
10 µg 20 µg
10 µg 20 µg
B. subtilis - -
7.55 ± 0.3 8.65 ± 0.9
- -
B. cereus - -
- -
- -
S. aureus (cepa 1) - -
- -
- -
S. aureus (cepa 2) - -
8.05 ± 0.2 8.35 ± 0.6
- -
L. monocytogenes - -
- -
- -
Y. enterocolitica - -
- -
- -
E. coli - -
- -
- -
K. pneumoniae - -
8.75 ± 0.4 10.25 ± 0.2
- -
K. rhinoscleromatis - -
- -
- -
S. tiphy - -
- -
- -
S. dysenteriae - -
7.25 ± 0.3 8.8 ± 0.5
- -
E. agglomerans - -
8.55 ± 0.4 9.25 ± 0.6
- -
Los resultados se presentan como promedio del diámetro en mm ± desviación estándar del duplicado de los experimentos y las cantidades como µg de proteína/sensidisco.
5.8.6 Determinación de la concentración mínima inhibitoria de las fracciones peptídicas e
hidrolizados de frijol
Después de las pruebas preliminares de inhibición, se determinó la concentración mínima
inhibitoria (CMI) de cada muestra por la técnica de dilución en microplaca, y los resultados de los
hidrolizados totales y las fracciones peptídicas de cada variedad de frijol se presentan en el Cuadro
5-11. De manera general, se observó que en las fracciones de péptidos con pesos moleculares
menores de 1 kDa las CMI determinada fueron menores que en péptidos de mayor tamaño. También
se observó que el FNP inhibió a la cepa indicadora S. dysenteriae con una CMI de 100 µg/mL de
solución final, y ésta fue la menor concentración obtenida en este ensayo. En las fracciones peptídicas
de frijol se observó actividad antibacteriana en las tres variedades, principalmente en las fracciones
peptídicas con pesos moleculares menores a 1 kDa, del análisis resultó que S. dysenteriae, además de
ser inhibida por FNP también fue inhibida por el FAH (400 µg/mL de solución) y el FPS (300 µg/mL de
solución). Otras cepas indicadoras fueron inhibidas con los péptidos de pesos moleculares superiores
a 10 kDa, S. typhi (300 µg/mL de solución) y E. agglomerans (200 µg/mL de solución).
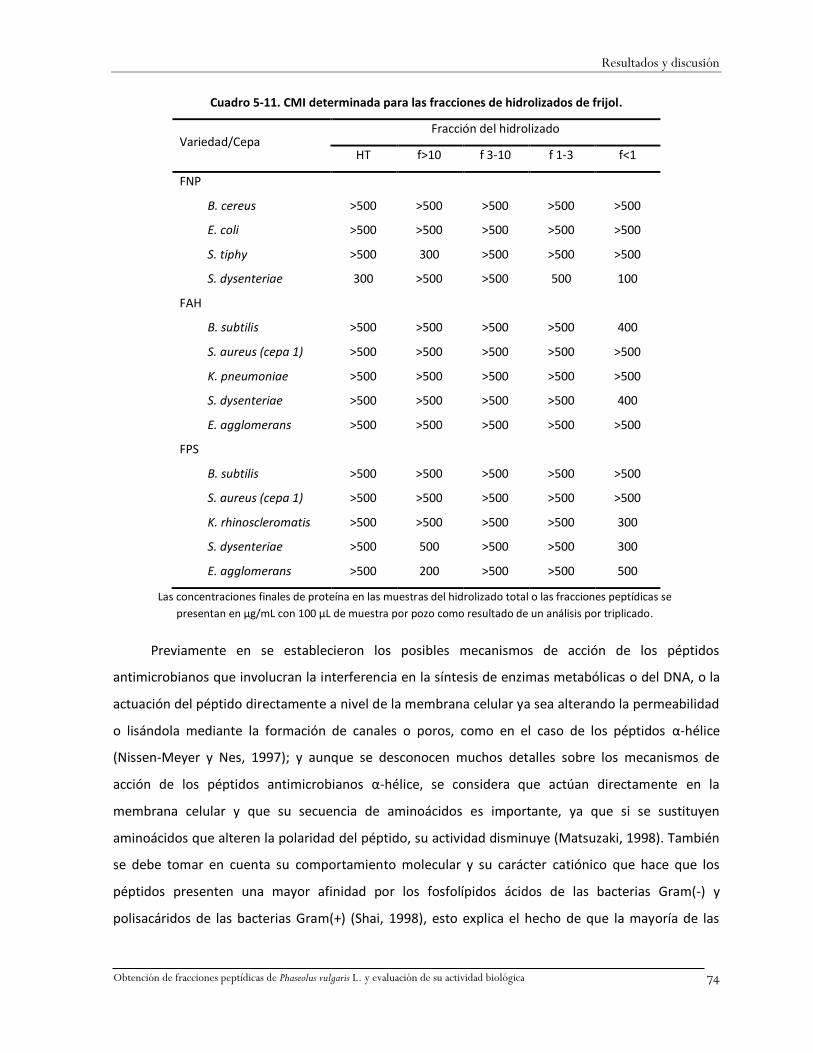
Resultados y discusión
74
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Cuadro 5-11. CMI determinada para las fracciones de hidrolizados de frijol.
Variedad/Cepa Fracción del hidrolizado
HT f>10 f 3-10 f 1-3 f<1
FNP
B. cereus >500 >500 >500 >500 >500
E. coli >500 >500 >500 >500 >500
S. tiphy >500 300 >500 >500 >500
S. dysenteriae 300 >500 >500 500 100
FAH
B. subtilis >500 >500 >500 >500 400
S. aureus (cepa 1) >500 >500 >500 >500 >500
K. pneumoniae >500 >500 >500 >500 >500
S. dysenteriae >500 >500 >500 >500 400
E. agglomerans >500 >500 >500 >500 >500
FPS
B. subtilis >500 >500 >500 >500 >500
S. aureus (cepa 1) >500 >500 >500 >500 >500
K. rhinoscleromatis >500 >500 >500 >500 300
S. dysenteriae >500 500 >500 >500 300
E. agglomerans >500 200 >500 >500 500
Las concentraciones finales de proteína en las muestras del hidrolizado total o las fracciones peptídicas se
presentan en µg/mL con 100 µL de muestra por pozo como resultado de un análisis por triplicado.
Previamente en se establecieron los posibles mecanismos de acción de los péptidos
antimicrobianos que involucran la interferencia en la síntesis de enzimas metabólicas o del DNA, o la
actuación del péptido directamente a nivel de la membrana celular ya sea alterando la permeabilidad
o lisándola mediante la formación de canales o poros, como en el caso de los péptidos α-hélice
(Nissen-Meyer y Nes, 1997); y aunque se desconocen muchos detalles sobre los mecanismos de
acción de los péptidos antimicrobianos α-hélice, se considera que actúan directamente en la
membrana celular y que su secuencia de aminoácidos es importante, ya que si se sustituyen
aminoácidos que alteren la polaridad del péptido, su actividad disminuye (Matsuzaki, 1998). También
se debe tomar en cuenta su comportamiento molecular y su carácter catiónico que hace que los
péptidos presenten una mayor afinidad por los fosfolípidos ácidos de las bacterias Gram(-) y
polisacáridos de las bacterias Gram(+) (Shai, 1998), esto explica el hecho de que la mayoría de las

Resultados y discusión
75
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
bacterias inhibidas por los péptidos de frijol sean del tipo Gram(-). Por lo que es de importancia
identificar los aminoácidos presentes en la cadena α-hélice de los péptidos analizados en este trabajo.
Por otro lado, la actividad antibacteriana de hidrolizados de origen vegetal ha sido
escasamente reportada, sin embargo, dicha actividad se ha presentado en extractos de plantas o sus
semillas. Amarowicz y col. (2008) reportaron que en constituyentes de frijol (fenoles y taninos)
obtenidos por extracción acetónica la CMI fue de 125 a 250 µg/mL, aún que no se trata del mismo
tipo de muestra sin embargo éste puede ser un patrón de comparación y muestra que los
hidrolizados de frijol y sus fracciones peptídicas inhiben a los microorganismos patógenos en
concentraciones similares. Por su parte, Wong y Ng (2005) extrajeron de P. lunatus la lunatusina, y
ésta proteína de aproximadamente 7 kDa de peso molecular presentó además de actividad
antibacteriana sobre Bacillus megaterium, B. subtilis, Proteus vulgaris y Mycobacterium phlei;
también actividad antifúngica sobre Fusarium oxysporum, Mycosphaerella arachidicola y Botrytis
cinerea.
Estos antecedentes demuestran que los péptidos de frijol tienen actividad antibacteriana sobre
bacterias principalmente Gram(-) y en menor grado Gram(+), y de acuerdo con lo reportado podrían
actuar como agentes antifúngicos sobre algunas cepas indicadoras.
5.9 Determinación de la actividad antioxidante
Los métodos de determinación de la actividad antioxidante son diversos debido a que cada uno
de ellos presenta sus beneficios y limitaciones. En este caso los métodos utilizados fueron
espectrofotométricos y tienen la limitación de no ser selectivos ya que no determinan el grupo de
compuestos que actúan (como: polifenoles totales, flavonoides totales, antocianinas totales,
catequinas totales, epi-catequina, epigalocatequingalato); sin embargo, son prácticos y se pueden
estandarizar al ser comparados con antioxidantes de referencia conocidos (Fernández-Pachón y col.,
2006).
5.9.1 Método de captación del radical libre DPPH+
En el Cuadro 5-12 se presentan los resultados de la actividad antioxidante correspondiente al
concentrado proteico, hidrolizado total y fracciones peptídicas de las tres variedades de frijol en
porcentaje de captación del radical libre DPPH.

Resultados y discusión
76
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Se observa que existe diferencia significativa entre las muestras leídas a 30 min y 60 min lo cual
muestra un evidente incremento en la captación del radical libre DPPH+ al incrementar el tiempo de
reacción (Duncan, p ≤ 0.05). Aunque no existen reportes con este tipo de muestras para realizar las
comparaciones pertinentes, hay en la literatura trabajos con proteínas vegetales similares como el
trabajo reportado por Rocha-Guzmán y col. (2007) quienes determinaron la actividad antioxidante de
5 mg/L de extracto metanólico 50 % (v/v) de harina de frijol en periodos de tiempo de 10 min por 1 h
y reportaron entre el 20 y 30 % de inhibición del radical libre DPPH+, y demostraron que el tiempo es
importante en la reacción del DPPH+ con la muestra. En el Cuadro 5-12 también se observó que el
efecto de inhibición depende de la variedad de frijol, siendo FAH y FNP las variedades con mayor
efecto antioxidante mientras el FPS presentó la menor actividad antioxidante.
Cuadro 5-12. Actividad antioxidante de péptidos de frijol determinada por captación de DPPH+ (%)
Tiempo % AA presentada por cada fracción peptídica de proteínas de frijol
Frijol (min) Concentrado H. Total f>10 kDa 3-10 kDa 1-3 kDa f<1 kDa
FNP 30 20.26 ± 1.82Da2 23.92 ± 2.60 Ca2 20.79 ± 2.38 Da2 29.89 ± 3.90 Ba2 36.35 ± 3.33 Aa2 24.27 ± 0.46 Ba2
60 21.16 ± 1.83 Da1 26.15 ± 1.46 Ca1 27.68 ± 1.30 Da1 34.46 ± 4.65 Ba1 40.19 ± 3.83 Aa1 26.09 ± 0.97 Ba1
FAH 30 20.78 ± 0.64 Da2 23.16 ± 0.50 Ca2 14.10 ± 2.36 Da2 34.51 ± 0.70 Ba2 35.24 ± 1.21 Aa2 33.79 ± 0.58 Ba2
60 21.59 ± 0.86 Da1 24.16 ± 0.70 Ca1 18.02 ± 1.92 Da1 35.50 ± 0.63 Ba1 36.91 ± 0.94 Aa1 35.31 ± 0.71 Ba1
FPS 30 10.13 ± 0.38 Db2 15.45 ± 3.01 Cb2 14.86 ± 4.46 Db2 25.58 ± 7.19 Bb2 28.61 ± 1.92 Ab2 34.02 ± 1.68 Bb2
60 12.70 ± 0.74 Db1 17.59 ± 3.94 Cb1 20.31 ± 3.18 Db1 30.18 ± 7.97 Bb1 32.30 ± 1.80 Ab1 37.71 ± 1.63 Bb1
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. A,B,C,D
. Medias en la misma línea con la misma letra no son significativamente diferentes (Duncan, p ≤ 0.05). a,b
. Medias en la misma columna con la misma letra no son significativamente diferentes (Duncan, p ≤ 0.05). 1,2
. Medias en la misma columna y variedad con el mismo número no son significativamente diferentes (Duncan, p ≤ 0.05).
Independientemente de la variedad de frijol analizada, la fracción que contiene el grupo de
péptidos con pesos moleculares de 1-3 kDa fue la que presentó mayor actividad antioxidante con
valores de 40.19, 36.91 y 32.30 % en FNP, FAH y FPS, respectivamente. Sin embargo, los controles
positivos inhiben hasta en 91.84 ± 1.5 (1.2 mM de Trolox), 90.50 ± 0.42 (0.75 mM de ácido gálico) y
89.56 ± 0.37 % (1.2 mM de ácido ascórbico).
En este tipo de análisis, se ha reportado que algunos compuestos fenólicos y taninos extraídos
de semillas de leguminosas poseen actividad antibacteriana (Valdés y col., 2011); sin embargo, han
sido pocos los trabajos publicados en hidrolizados de origen vegetal y sus fracciones peptídicas.
Aunque no se trata del mismo radical libre, el grupo de trabajo de Torruco-Uco y col. (2009)
determinaron que el hidrolizado total de frijol negro Jamapa hidrolizado con Alcalasa durante 60 min
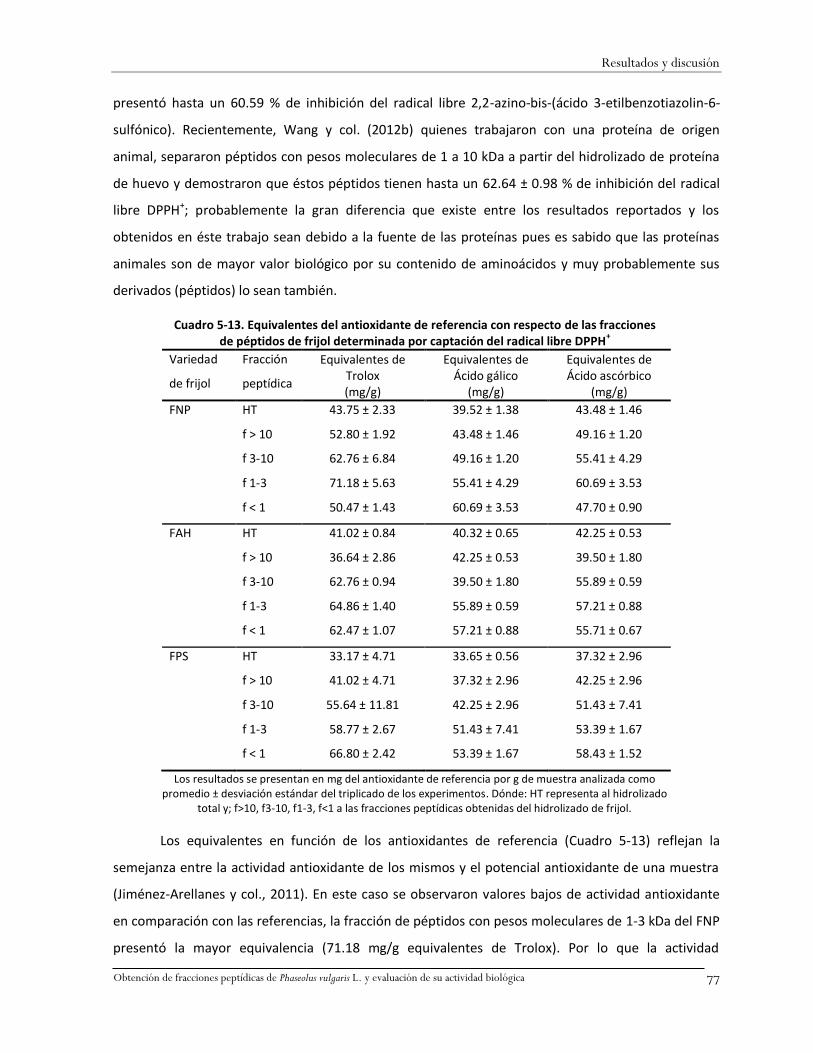
Resultados y discusión
77
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
presentó hasta un 60.59 % de inhibición del radical libre 2,2-azino-bis-(ácido 3-etilbenzotiazolin-6-
sulfónico). Recientemente, Wang y col. (2012b) quienes trabajaron con una proteína de origen
animal, separaron péptidos con pesos moleculares de 1 a 10 kDa a partir del hidrolizado de proteína
de huevo y demostraron que éstos péptidos tienen hasta un 62.64 ± 0.98 % de inhibición del radical
libre DPPH+; probablemente la gran diferencia que existe entre los resultados reportados y los
obtenidos en éste trabajo sean debido a la fuente de las proteínas pues es sabido que las proteínas
animales son de mayor valor biológico por su contenido de aminoácidos y muy probablemente sus
derivados (péptidos) lo sean también.
Cuadro 5-13. Equivalentes del antioxidante de referencia con respecto de las fracciones de péptidos de frijol determinada por captación del radical libre DPPH
+
Variedad
de frijol
Fracción
peptídica
Equivalentes de Trolox (mg/g)
Equivalentes de Ácido gálico
(mg/g)
Equivalentes de Ácido ascórbico
(mg/g)
FNP HT 43.75 ± 2.33 39.52 ± 1.38 43.48 ± 1.46
f > 10 52.80 ± 1.92 43.48 ± 1.46 49.16 ± 1.20
f 3-10 62.76 ± 6.84 49.16 ± 1.20 55.41 ± 4.29
f 1-3 71.18 ± 5.63 55.41 ± 4.29 60.69 ± 3.53
f < 1 50.47 ± 1.43 60.69 ± 3.53 47.70 ± 0.90
FAH HT 41.02 ± 0.84 40.32 ± 0.65 42.25 ± 0.53
f > 10 36.64 ± 2.86 42.25 ± 0.53 39.50 ± 1.80
f 3-10 62.76 ± 0.94 39.50 ± 1.80 55.89 ± 0.59
f 1-3 64.86 ± 1.40 55.89 ± 0.59 57.21 ± 0.88
f < 1 62.47 ± 1.07 57.21 ± 0.88 55.71 ± 0.67
FPS HT 33.17 ± 4.71 33.65 ± 0.56 37.32 ± 2.96
f > 10 41.02 ± 4.71 37.32 ± 2.96 42.25 ± 2.96
f 3-10 55.64 ± 11.81 42.25 ± 2.96 51.43 ± 7.41
f 1-3 58.77 ± 2.67 51.43 ± 7.41 53.39 ± 1.67
f < 1 66.80 ± 2.42 53.39 ± 1.67 58.43 ± 1.52
Los resultados se presentan en mg del antioxidante de referencia por g de muestra analizada como promedio ± desviación estándar del triplicado de los experimentos. Dónde: HT representa al hidrolizado
total y; f>10, f3-10, f1-3, f<1 a las fracciones peptídicas obtenidas del hidrolizado de frijol.
Los equivalentes en función de los antioxidantes de referencia (Cuadro 5-13) reflejan la
semejanza entre la actividad antioxidante de los mismos y el potencial antioxidante de una muestra
(Jiménez-Arellanes y col., 2011). En este caso se observaron valores bajos de actividad antioxidante
en comparación con las referencias, la fracción de péptidos con pesos moleculares de 1-3 kDa del FNP
presentó la mayor equivalencia (71.18 mg/g equivalentes de Trolox). Por lo que la actividad

Resultados y discusión
78
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
antioxidante de los péptidos de frijol analizados en este trabajo, no puede competir con las
referencias desde el punto de vista de bioactividad, sin embargo, puede ser considerado como una
alternativa ante los antioxidantes de origen químico.
5.9.2 Método de captación del radical libre ABTS+
En el Cuadro 5-14 se presentan los resultados de la actividad antioxidante correspondiente a la
técnica de decoloración del radical libre ABTS. Los porcentajes de actividad antioxidante más altos
fueron obtenidos cuando la fracción peptídica fue de peso molecular inferior a 1 kDa (Duncan, p ≤
0.05). En la fracción peptídica de 1–3 kDa se observó que la actividad antioxidante fue superior al 50
%, mientras que en la fracción de peso molecular menor de 1 kDa la actividad fue superior al 80 %. De
las tres variedades el FNP demostró tener el mayor potencial antioxidante (Duncan, p ≤ 0.05), ya que
desde el hidrolizado total se observó casi un 50 % de actividad antioxidante y llegó hasta el 99 % en la
fracción de menor tamaño. En lo que respecta a los antioxidantes de referencia, éstos llegaron hasta
el 97.25 ± 2.93 (2.5 mM de Trolox), 97.85 ± 2.87 (1 mM de ácido gálico) y 91.01 ± 3.23 (3.5 mM de
ácido ascórbico).
Cuadro 5-14. Actividad antioxidante de péptidos de frijol determinada por decoloración del ABTS+ (%)
Fracción (kDa) FNP FAH FPS
HT 48.38 ± 1.44 Ad 38.04 ± 0.32 Bd 19.90 ± 2.24 Cd
f > 10 54.48 ± 2.44 Ad 26.27 ± 3.59 Bd 30.75 ± 6.59 Cd
f 3-10 61.82 ± 4.19 Ac 55.31 ± 1.19 Bc 35.41 ± 3.92 Cc
f 1-3 78.62 ± 5.21 Ab 69.48 ± 2.61 Bb 51.62 ± 2.46 Cb
f < 1 99.17 ± 0.91 Aa 87.63 ± 0.79 Ba 82.69 ± 2.05 Ca
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. A,B,C
Letras iguales en el mismo renglón no representan diferencia significativa (Duncan, p ≤ 0.05). a,b,c,d
Letras iguales en la misma columna no representan diferencia significativa (Duncan, p ≤ 0.05)
Recientemente, Ruiz-Ruiz y col. (2013a) reportaron una actividad antioxidante de 6.09 a 28.2 %
en hidrolizados totales y fracciones de péptidos de frijol negro Jamapa (f>10, 5–10, 3–5, 1–3 y f<1
kDa) obtenidos con una combinación de Alcalasa-Flavourzima o pepsina-pancreatina; siendo sus
valores inferiores a los resultados obtenidos en éste trabajo. En otro trabajo similar determinaron
que el hidrolizado total de frijol negro Jamapa hidrolizado con Alcalasa durante 60 min presentó hasta
un 60.59 % de inhibición de éste radical libre (Torruco-Uco y col., 2009). Por su parte, Dávalos y col.
(2004) reportaron las secuencias de péptidos obtenidos de la hidrólisis de proteínas de huevo TTYY,
TPIL y TPI, los cuales presentaron actividad antioxidante (hasta de 3.8 mM equivalentes de Trolox por
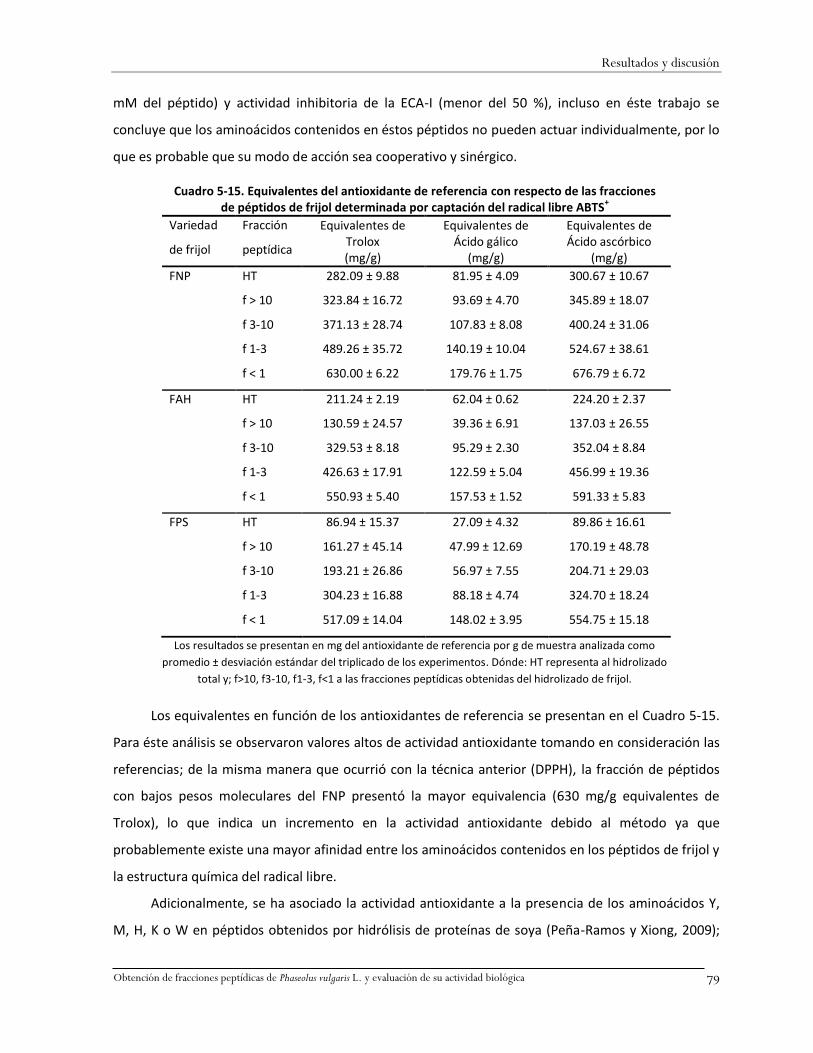
Resultados y discusión
79
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
mM del péptido) y actividad inhibitoria de la ECA-I (menor del 50 %), incluso en éste trabajo se
concluye que los aminoácidos contenidos en éstos péptidos no pueden actuar individualmente, por lo
que es probable que su modo de acción sea cooperativo y sinérgico.
Cuadro 5-15. Equivalentes del antioxidante de referencia con respecto de las fracciones de péptidos de frijol determinada por captación del radical libre ABTS+
Variedad
de frijol
Fracción
peptídica
Equivalentes de Trolox (mg/g)
Equivalentes de Ácido gálico
(mg/g)
Equivalentes de Ácido ascórbico
(mg/g)
FNP HT 282.09 ± 9.88 81.95 ± 4.09 300.67 ± 10.67
f > 10 323.84 ± 16.72 93.69 ± 4.70 345.89 ± 18.07
f 3-10 371.13 ± 28.74 107.83 ± 8.08 400.24 ± 31.06
f 1-3 489.26 ± 35.72 140.19 ± 10.04 524.67 ± 38.61
f < 1 630.00 ± 6.22 179.76 ± 1.75 676.79 ± 6.72
FAH HT 211.24 ± 2.19 62.04 ± 0.62 224.20 ± 2.37
f > 10 130.59 ± 24.57 39.36 ± 6.91 137.03 ± 26.55
f 3-10 329.53 ± 8.18 95.29 ± 2.30 352.04 ± 8.84
f 1-3 426.63 ± 17.91 122.59 ± 5.04 456.99 ± 19.36
f < 1 550.93 ± 5.40 157.53 ± 1.52 591.33 ± 5.83
FPS HT 86.94 ± 15.37 27.09 ± 4.32 89.86 ± 16.61
f > 10 161.27 ± 45.14 47.99 ± 12.69 170.19 ± 48.78
f 3-10 193.21 ± 26.86 56.97 ± 7.55 204.71 ± 29.03
f 1-3 304.23 ± 16.88 88.18 ± 4.74 324.70 ± 18.24
f < 1 517.09 ± 14.04 148.02 ± 3.95 554.75 ± 15.18
Los resultados se presentan en mg del antioxidante de referencia por g de muestra analizada como
promedio ± desviación estándar del triplicado de los experimentos. Dónde: HT representa al hidrolizado
total y; f>10, f3-10, f1-3, f<1 a las fracciones peptídicas obtenidas del hidrolizado de frijol.
Los equivalentes en función de los antioxidantes de referencia se presentan en el Cuadro 5-15.
Para éste análisis se observaron valores altos de actividad antioxidante tomando en consideración las
referencias; de la misma manera que ocurrió con la técnica anterior (DPPH), la fracción de péptidos
con bajos pesos moleculares del FNP presentó la mayor equivalencia (630 mg/g equivalentes de
Trolox), lo que indica un incremento en la actividad antioxidante debido al método ya que
probablemente existe una mayor afinidad entre los aminoácidos contenidos en los péptidos de frijol y
la estructura química del radical libre.
Adicionalmente, se ha asociado la actividad antioxidante a la presencia de los aminoácidos Y,
M, H, K o W en péptidos obtenidos por hidrólisis de proteínas de soya (Peña-Ramos y Xiong, 2009);

Resultados y discusión
80
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
con este conocimiento se puede suponer que algunos de estos aminoácidos se encuentran presentes
en las fracciones ensayadas ya que la soya pertenece a la misma familia que el frijol. En otros trabajos
han reportado que los aminoácidos básicos poseen la capacidad de unirse a iones metálicos, y los
aminoácidos aromáticos y sulfurados poseen la capacidad de donar protones para estabilizar los
radicales libres (Arcan y Yemenicioğlu, 2007); lo que permite suponer que en los péptidos de frijol se
encuentren presentes estos tipos de aminoácidos. La actividad antioxidante de los HT de FNP de este
trabajo es similar a la obtenida por Torruco-Uco y col. (2009) con los HT de frijol negro Jamapa,
mientras que las variedades FAH y FPS tuvieron menores porcentajes de actividad antioxidante; lo
que permite deducir que las semillas de color negro poseen mayor potencial antioxidante que las
variedades de color café o amarillo, y posiblemente se deba a su contenido de polifenoles (Rocha-
Guzmán y col., 2007), además de la acción de los aminoácidos contenidos en la muestra. Además,
Ruiz-Ruiz y col. (2013a) reportaron que las fracciones peptídicas de frijol de bajo peso molecular
fueron responsables de al menos otra actividad biológica, como la hipertensiva.
5.9.3 Método de captación del radical libre DMPD+
En el Cuadro 5-16 se presentan los resultados de la actividad antioxidante correspondiente a
la técnica de decoloración del radical libre DMPD.
Cuadro 5-16. Actividad antioxidante de péptidos de frijol determinada por decoloración del DMPD+ (%)
Fracción (kDa) FNP FAH FPS
HT 7.78 ± 0.37 Bd
31.24 ± 1.53 Ad
11.97 ± 2.24 Bd
f >10 17.03 ± 1.74 Bd 23.32 ± 0.77 Ad 16.34 ± 6.59 Bd
f 3-10 44.22 ± 3.25 Bc 47.57 ± 3.92 Ac 27.54 ± 3.92 Bc
f 1-3 67.89 ± 2.96 Bb 66.04 ± 1.59 Ab 66.25 ± 2.05 Bb
f < 1 76.16 ± 1.23 Ba 67.09 ± 3.62 Aa 89.08 ± 2.46 Ba
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. A,B
Letras iguales en el mismo renglón no representan diferencia significativa (Duncan, p ≤ 0.05). a,b,c,d
Letras iguales en la misma columna no representan diferencia significativa (Duncan, p ≤ 0.05)
En esta determinación el incremento en el porcentaje de la actividad antioxidante en
presencia de péptidos de bajo peso molecular fue menor a la obtenida por el método de ABTS, sin
embargo, es probable que sea la técnica más adecuada para determinar la actividad antioxidante de
los hidrolizados de frijol debido a que el disolvente es una solución acuosa. Los porcentajes de
actividad antioxidante más altos fueron obtenidos cuando la fracción peptídica fue peso molecular
inferior a 1 kDa (Duncan, p ≤ 0.05), lo cual concuerda con los resultados obtenidos con el método de
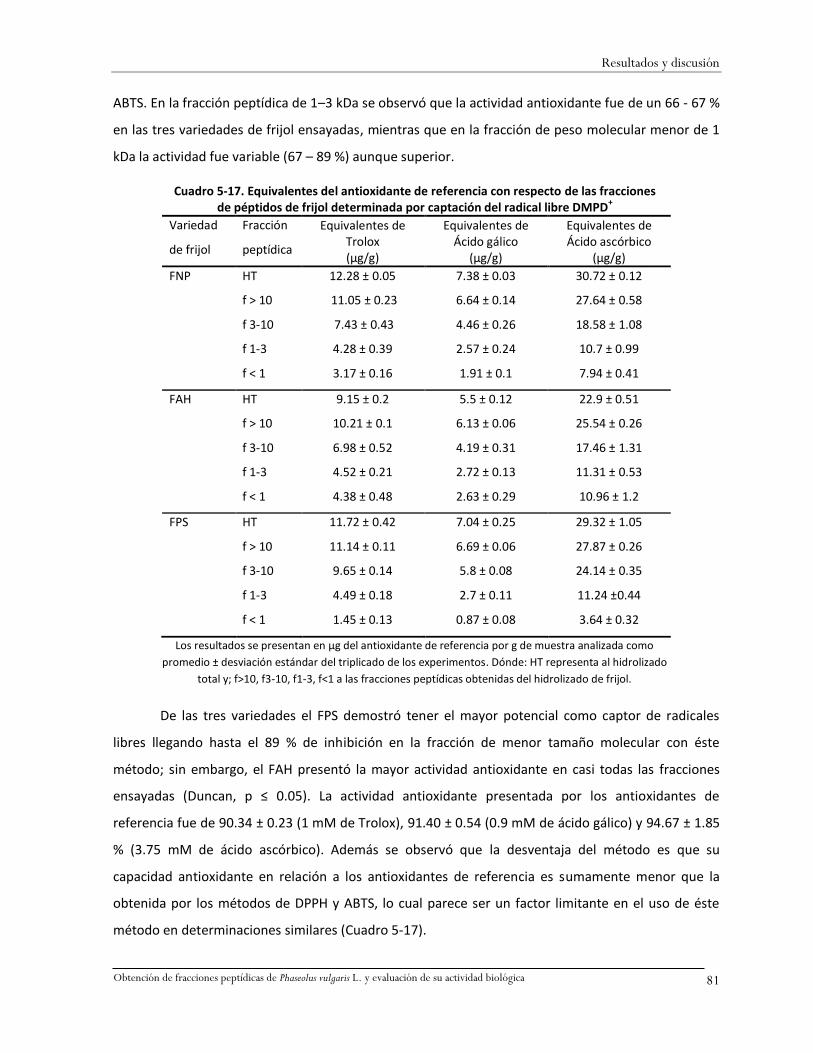
Resultados y discusión
81
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
ABTS. En la fracción peptídica de 1–3 kDa se observó que la actividad antioxidante fue de un 66 - 67 %
en las tres variedades de frijol ensayadas, mientras que en la fracción de peso molecular menor de 1
kDa la actividad fue variable (67 – 89 %) aunque superior.
Cuadro 5-17. Equivalentes del antioxidante de referencia con respecto de las fracciones de péptidos de frijol determinada por captación del radical libre DMPD+
Variedad
de frijol
Fracción
peptídica
Equivalentes de Trolox (µg/g)
Equivalentes de Ácido gálico
(µg/g)
Equivalentes de Ácido ascórbico
(µg/g)
FNP HT 12.28 ± 0.05 7.38 ± 0.03 30.72 ± 0.12
f > 10 11.05 ± 0.23 6.64 ± 0.14 27.64 ± 0.58
f 3-10 7.43 ± 0.43 4.46 ± 0.26 18.58 ± 1.08
f 1-3 4.28 ± 0.39 2.57 ± 0.24 10.7 ± 0.99
f < 1 3.17 ± 0.16 1.91 ± 0.1 7.94 ± 0.41
FAH HT 9.15 ± 0.2 5.5 ± 0.12 22.9 ± 0.51
f > 10 10.21 ± 0.1 6.13 ± 0.06 25.54 ± 0.26
f 3-10 6.98 ± 0.52 4.19 ± 0.31 17.46 ± 1.31
f 1-3 4.52 ± 0.21 2.72 ± 0.13 11.31 ± 0.53
f < 1 4.38 ± 0.48 2.63 ± 0.29 10.96 ± 1.2
FPS HT 11.72 ± 0.42 7.04 ± 0.25 29.32 ± 1.05
f > 10 11.14 ± 0.11 6.69 ± 0.06 27.87 ± 0.26
f 3-10 9.65 ± 0.14 5.8 ± 0.08 24.14 ± 0.35
f 1-3 4.49 ± 0.18 2.7 ± 0.11 11.24 ±0.44
f < 1 1.45 ± 0.13 0.87 ± 0.08 3.64 ± 0.32
Los resultados se presentan en µg del antioxidante de referencia por g de muestra analizada como
promedio ± desviación estándar del triplicado de los experimentos. Dónde: HT representa al hidrolizado
total y; f>10, f3-10, f1-3, f<1 a las fracciones peptídicas obtenidas del hidrolizado de frijol.
De las tres variedades el FPS demostró tener el mayor potencial como captor de radicales
libres llegando hasta el 89 % de inhibición en la fracción de menor tamaño molecular con éste
método; sin embargo, el FAH presentó la mayor actividad antioxidante en casi todas las fracciones
ensayadas (Duncan, p ≤ 0.05). La actividad antioxidante presentada por los antioxidantes de
referencia fue de 90.34 ± 0.23 (1 mM de Trolox), 91.40 ± 0.54 (0.9 mM de ácido gálico) y 94.67 ± 1.85
% (3.75 mM de ácido ascórbico). Además se observó que la desventaja del método es que su
capacidad antioxidante en relación a los antioxidantes de referencia es sumamente menor que la
obtenida por los métodos de DPPH y ABTS, lo cual parece ser un factor limitante en el uso de éste
método en determinaciones similares (Cuadro 5-17).

Resultados y discusión
82
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Desde el punto de vista de aplicación, el tiempo de reacción y lectura necesario para realizar
las medidas de DPPH (30 y 60 min) en comparación con el método ABTS (1 y 6 min), supone una
desventaja en adición también al elevado costo del radical libre DPPH, sin embargo, los resultados
obtenidos con el método DMPD (10 min) podrían ser más útiles ya que el sistema en el cual se lleva a
cabo la reacción involucra un medio acuoso en la prueba de actividad antioxidante, esto permitiría
deducir la actividad antioxidante de los péptidos de frijol adicionados en alimentos para consumo
humano (Kuskosky y col., 2005).
5.9.4 Método de quimioluminiscencia
Otro método utilizado para determinar la actividad antioxidante de los hidrolizados y sus
fracciones peptídicas fue el método de quimioluminiscencia (o de inhibición de los radicales libres
luminol) y cuyos resultados se presentan en el Cuadro 5-19.
Cuadro 5-18. Actividad quimioluminiscente de los controles positivos y blancos de control.
Antioxidante
de referencia % inhibición Blanco de
control % inhibición
Trolox 70.19 ± 2.71 Metanol 10.86 ± 5.20
Á. ascórbico 99.20 ± 0.06 Acetonitrilo 10.81 ± 1.63
Á. gálico 55.81 ± 2.76 PBS 0.0 ± 0.44
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos.
Para la estandarización del método se determinó la actividad antioxidante de tres
antioxidantes conocidos, utilizados como referencia (12 mM Trolox, 10 mM de ácido ascórbico y 12
mM ácido gálico) bajo los estándares de la técnica descrita en (4.12.4); además, se determinó la
posible actividad antioxidante de los disolventes utilizados en el tratamiento de los péptidos
(metanol, acetonitrilo y búfer de fosfatos) a fin de determinar alguna interferencia en la
interpretación de los resultados siguientes (Cuadro 5-18). Y se observó que la variación debido a los
disolventes utilizados fue de hasta un 10 %. Además, se observó que en este método el ácido
ascórbico fue quien inhibió hasta el 99 % del radical luminol, a diferencia de los otros métodos donde
el Trolox mostró mayor afinidad por el atrapamiento de los radicales libres.
En la determinación de la actividad antioxidante por el método de quimioluminiscencia se
observó que el potencial efecto antioxidante de las muestras depende de la variedad de frijol
empleada, siendo FAH la variedad con mayor actividad seguida de FNP y finalmente FPS (Cuadro

Resultados y discusión
83
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5-19). Además se observó que todas las fracciones son significativamente diferentes entre sí (Duncan,
p ≤ 0.05), siendo la fracción 1-3 kDa la de mayor actividad antioxidante seguida por la f<1 kDa lo que
indica que péptidos de bajo peso molecular tienen mayor actividad antioxidante que aquellos con
largas cadenas de péptidos lo que concuerda con Wang y col. (2012b). Las fracciones peptídicas
presentaron valores de actividad antioxidante en el intervalo de 36.07 - 94.02 %. Las fracciones con
mayor actividad antioxidante (determinada como inhibición del radical libre luminol) fueron las de
pesos moleculares de 1-3 kDa con 92.68 %, 94.02 % y 93.85 % en FNP, FAH y FPS, respectivamente;
seguidas por la fracción de péptidos de pesos moleculares menores de 1 kDa. Por su parte, el FAH
presentó actividad antioxidante significativamente mayor que las variedades FNP y FPS, hasta del
91.49 %.
Cuadro 5-19. Actividad antioxidante determinada por quimioluminiscencia de péptidos de frijol (%)
% AA presentada por cada fracción de frijol
Variedad Concentrado HT f>10 kDa 3-10 kDa 1-3 kDa f<1 kDa
FNP 36.07 ± 1.77 Fb 77.49 ± 1.43 Db 83.72 ± 2.48 Eb 87.33 ± 0.73 Cb 92.68 ± 3.07 Ab 76.32 ± 0.91 Bb
FAH 72.84 ± 2.42 Fa 74.14 ± 2.51 Da 47.06 ± 3.11 Ea 85.57 ± 1.07 Ca 94.02 ± 1.09 Aa 91.49 ± 1.15 Ba
FPS 42.77 ± 2.91 Fc 74.21 ± 1.58 Dc 41.97 ± 1.92 Ec 74.84 ± 2.42 Cc 93.85 ± 0.83 Ac 90.60 ± 1.32 Bc
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. A,B,C,D,E,F
. Medias en la misma línea con la misma letra no son significativamente diferentes (Duncan, p ≤ 0.05) a,b,c
. Medias en la misma columna con la misma letra no son significativamente diferentes (Duncan, p ≤ 0.05)
Al realizar la comparación de la actividad antioxidante de los hidrolizados totales y sus
fracciones peptídicas por el método del DPPH+ y por el método de quimioluminiscencia se
presentaron tendencias similares, sin embargo, con el método de luminiscencia se presenta un
incremento en la actividad antioxidante que puede estar asociado con la naturaleza de los radicales
(hidrofóbicos o hidrófilos), con los compuestos antioxidantes usados como referencia y con la
sensibilidad del método. Se ha reportado a la luminiscencia como un método de determinación de la
actividad antioxidante que ofrece ventajas al requerir muestras pequeñas y presentar alta
sensibilidad en el monitoreo de la reacción (Jakubowski y col., 1998) lo que permitió su aplicación
para determinar actividad antioxidante de péptidos de frijol usando cantidades pequeñas de dichos
péptidos.
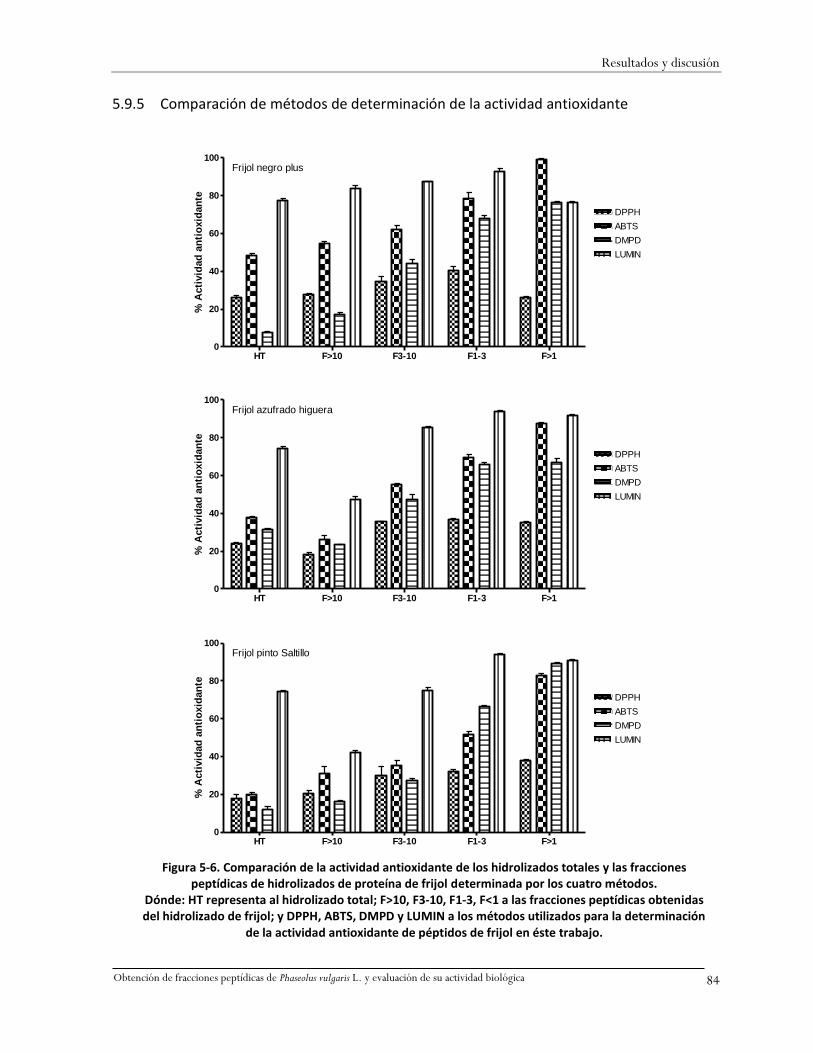
Resultados y discusión
84
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.9.5 Comparación de métodos de determinación de la actividad antioxidante
Figura 5-6. Comparación de la actividad antioxidante de los hidrolizados totales y las fracciones peptídicas de hidrolizados de proteína de frijol determinada por los cuatro métodos.
Dónde: HT representa al hidrolizado total; F>10, F3-10, F1-3, F<1 a las fracciones peptídicas obtenidas del hidrolizado de frijol; y DPPH, ABTS, DMPD y LUMIN a los métodos utilizados para la determinación
de la actividad antioxidante de péptidos de frijol en éste trabajo.
HT F>10 F3-10 F1-3 F>10
20
40
60
80
100
DPPH
ABTS
DMPD
LUMIN
Frijol negro plus
% A
cti
vid
ad
an
tio
xid
an
te
HT F>10 F3-10 F1-3 F>10
20
40
60
80
100
DPPH
ABTS
DMPD
LUMIN
Frijol azufrado higuera
% A
cti
vid
ad
an
tio
xid
an
te
HT F>10 F3-10 F1-3 F>10
20
40
60
80
100
DPPH
ABTS
DMPD
LUMIN
Frijol pinto Saltillo
% A
cti
vid
ad
an
tio
xid
an
te

Resultados y discusión
85
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
En la Figura 5-6 se observa un comportamiento similar entre las diferentes técnicas de
determinación de actividad antioxidante. Sin embargo, es notorio el incremento en la actividad
antioxidante cuando esta se determinó por los métodos de quimioluminiscencia y apagamiento del
radical ABTS, presentando una mayor sensibilidad ante los radicales libres. Los métodos menos
sensibles fueron DPPH y DMPD.
En general, las fracciones con pesos moleculares de 1-3 y f<1 kDa presentaron la mayor
actividad antioxidante, éstos datos concuerdan con lo reportado por Carrasco-Castilla y col. (2012)
quienes encontraron el máximo efecto de reducción del radical ABTS en hidrolizados de frijol negro
Jamapa con pesos moleculares menores o iguales a 700 Da. Estos datos concuerdan con lo reportado
por Gallegos-Tintoré y col. (2013) quienes mencionan que los péptidos más reactivos son de pesos
moleculares entre 500 a 1800 Da, y además contienen aminoácidos hidrofóbicos.
5.10 Determinación de la actividad inhibidora de la enzima convertidora de la
angiotensina
En el Cuadro 5-20 se presentan los resultados de la actividad inhibidora de la enzima
convertidora de la angiotensina I (% ECA-I) de la fracción que contiene péptidos de pesos moleculares
entre 1 y 3 kDa de las variedades de frijol estudiadas a una concentración final de 20 µg/mL. Se
observó que la actividad mínima obtenida fue del 88 % en frijol negro plus y máxima del 95 % en las
otras dos variedades de frijol (azufrado higuera y pinto Saltillo). Estos porcentajes de inhibición in
vitro sugieren el potencial antihipertensivo de éstos péptidos de proteínas hidrolizadas de frijol.
Cuadro 5-20. Inhibición de la ECA-I por los péptidos de frijol de 1-3 kDa (%)
Variedad de frijol Inhibición de la ECA-I
Negro plus 88.73 ± 1.35 b
Azufrado higuera 95.63 ± 2.24 a
Pinto Saltillo 95.54 ± 1.98 a
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos. a,b,c
Medias en la misma columna con la misma letra no son significativamente diferentes (Duncan, p ≤ 0.05).
Con éstas mismas variedades de frijol, se analizaron los hidrolizados totales y la fracción de
péptidos con pesos moleculares de 3-10 kDa bajo las mismas condiciones y concentración (Figura
5-7). Se observó que la actividad inhibitoria de la enzima convertidora de la angiotensina se
incrementó conforme la fracción peptídica contenía péptidos de menor tamaño molecular. Se
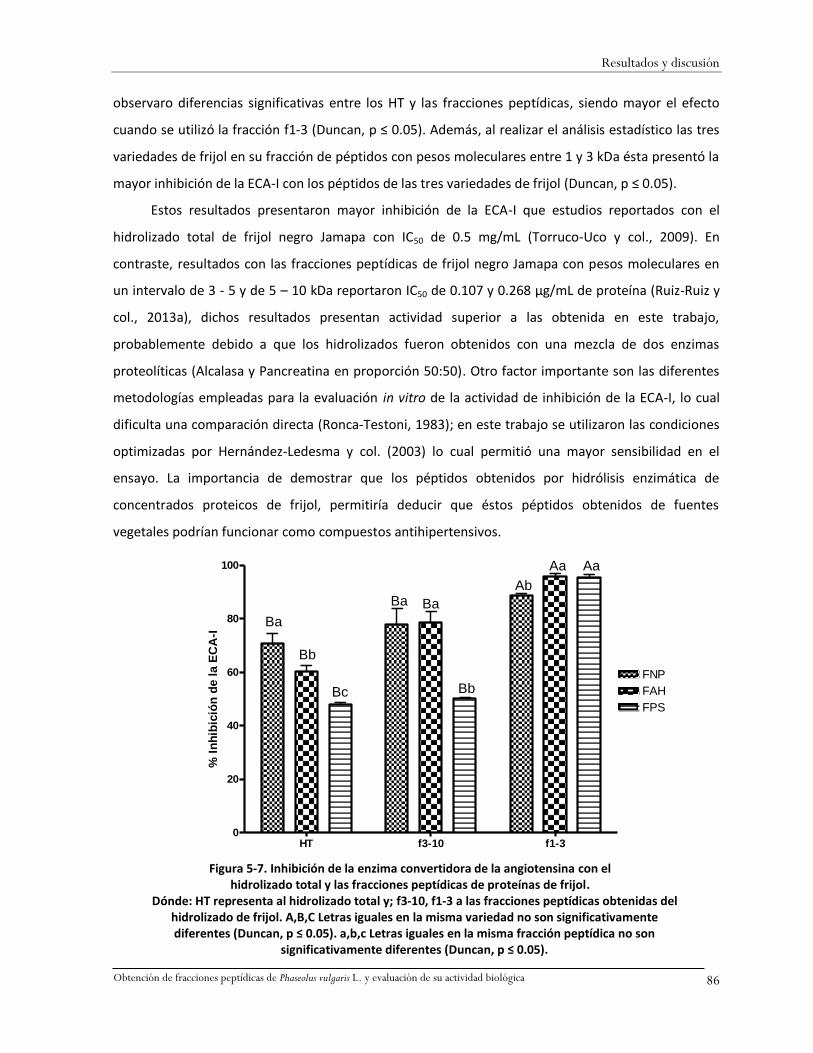
Resultados y discusión
86
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
observaro diferencias significativas entre los HT y las fracciones peptídicas, siendo mayor el efecto
cuando se utilizó la fracción f1-3 (Duncan, p ≤ 0.05). Además, al realizar el análisis estadístico las tres
variedades de frijol en su fracción de péptidos con pesos moleculares entre 1 y 3 kDa ésta presentó la
mayor inhibición de la ECA-I con los péptidos de las tres variedades de frijol (Duncan, p ≤ 0.05).
Estos resultados presentaron mayor inhibición de la ECA-I que estudios reportados con el
hidrolizado total de frijol negro Jamapa con IC50 de 0.5 mg/mL (Torruco-Uco y col., 2009). En
contraste, resultados con las fracciones peptídicas de frijol negro Jamapa con pesos moleculares en
un intervalo de 3 - 5 y de 5 – 10 kDa reportaron IC50 de 0.107 y 0.268 µg/mL de proteína (Ruiz-Ruiz y
col., 2013a), dichos resultados presentan actividad superior a las obtenida en este trabajo,
probablemente debido a que los hidrolizados fueron obtenidos con una mezcla de dos enzimas
proteolíticas (Alcalasa y Pancreatina en proporción 50:50). Otro factor importante son las diferentes
metodologías empleadas para la evaluación in vitro de la actividad de inhibición de la ECA-I, lo cual
dificulta una comparación directa (Ronca-Testoni, 1983); en este trabajo se utilizaron las condiciones
optimizadas por Hernández-Ledesma y col. (2003) lo cual permitió una mayor sensibilidad en el
ensayo. La importancia de demostrar que los péptidos obtenidos por hidrólisis enzimática de
concentrados proteicos de frijol, permitiría deducir que éstos péptidos obtenidos de fuentes
vegetales podrían funcionar como compuestos antihipertensivos.
Figura 5-7. Inhibición de la enzima convertidora de la angiotensina con el hidrolizado total y las fracciones peptídicas de proteínas de frijol.
Dónde: HT representa al hidrolizado total y; f3-10, f1-3 a las fracciones peptídicas obtenidas del hidrolizado de frijol. A,B,C Letras iguales en la misma variedad no son significativamente diferentes (Duncan, p ≤ 0.05). a,b,c Letras iguales en la misma fracción peptídica no son
significativamente diferentes (Duncan, p ≤ 0.05).
HT f3-10 f1-30
20
40
60
80
100
FNP
FAH
FPS
Ab
Aa Aa
Ba
Ba
Bb
Bc
Ba
Bb
% I
nh
ibic
ión
de l
a E
CA
-I

Resultados y discusión
87
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Al haber obtenido los mayores porcentajes de actividad inhibitoria de la ECA-I con fracciones
de bajo peso molecular provenientes de FAH, y también de actividad antioxidante y actividad
antibacteriana; se puede suponer que ésta variedad de frijol, específicamente sus péptidos de menor
tamaño, podría presentar otro tipo de actividad biológica ya que otros autores han reportado que un
péptido puede actuar de varias maneras en un mismo sistema (Srihongthong y col., 2012).
Cuadro 5-21. Efecto en la presión sanguínea (PS) de la administración de la F3-10 del FAH sobre RWH.
Tiempo Péptido FAH f3-10 Captopril® Control
(h) (4 mg/kg) (10 mg/kg) (agua 2.5 mL/kg)
1 -15.55 ± 12.30 - 28.44 ± 14.94 -12.86 ± 8.78
2 - 27.13 ± 11.17 * - 29.67 ± 18.89 ** - 15.70 ± 12.78
4 - 23.55 ± 12.44 * - 40.44 ± 1.07 ** - 5.11 ± 9.55
6 - 12.05 ± 13.50 - 41.11 ± 22.44 - 6.78 ± 10.25
Los valores son significativamente diferentes de la presión sanguínea basal a las 0 h, (t-Student, p≤0.01, *) y (t-Student, p≤0.001, **), n=3. Los valores de PS a las 0 h fueron 188.6 ± 13.6 para la fracción peptídica y
201.22 ± 5.6 para el Captopril.
Se observó que a una dosis de 4 mg/kg de animal (RWH) la presión sanguínea disminuyó
después de 2 h de haber sido administrado el péptido (FAH f3-10) hasta -27.13 ± 11.17 mmHg
(Cuadro 5-21). Los resultados indicaron que la fracción peptídica disminuyó la presión sanguínea con
diferencia significativa respecto al control (t-Student, p ≤ 0.01). Además, se observó que la
disminución de la PS fue similar a la que produce el control positivo a las 2 h (-29.67 ± 18.89 mmHg).
Aunque las proteínas de origen vegetal han sido escasamente estudiadas, la actividad
antihipertensiva en péptidos derivados de proteínas de leche y otras fuentes de origen animal han
sido ampliamente estudiadas y demostradas (Torres-Llanez y col., 2005). Algunos péptidos derivados
de la hidrólisis enzimática con Alcalasa de diferentes fuentes de origen vegetal como la soya, el frijol
de mung, arroz y chícharo demostraron decrementos en la actividad antihipertensiva se 31.8 mmHg
(600 mg/kg), 25.6 mmHg (600 mg/kg), 7 mmHg (100 mg/kg), y 44.4 mmHg (50 mg/kg),
respectivamente (Li y col., 2006a; Li y col., 2004; Wu y Ding, 2001; Vermeirssen y col., 2005). Éstos
resultados se encuentran en el intervalo de valores obtenidos en este trabajo. Aunque es cierto que
se requieren más experimentos para confirmar el potencial hipotensivo de los péptidos de frijiol, este
trabajo podría ser una aproximación de la demostración in vivo de dicha actividad biológica.
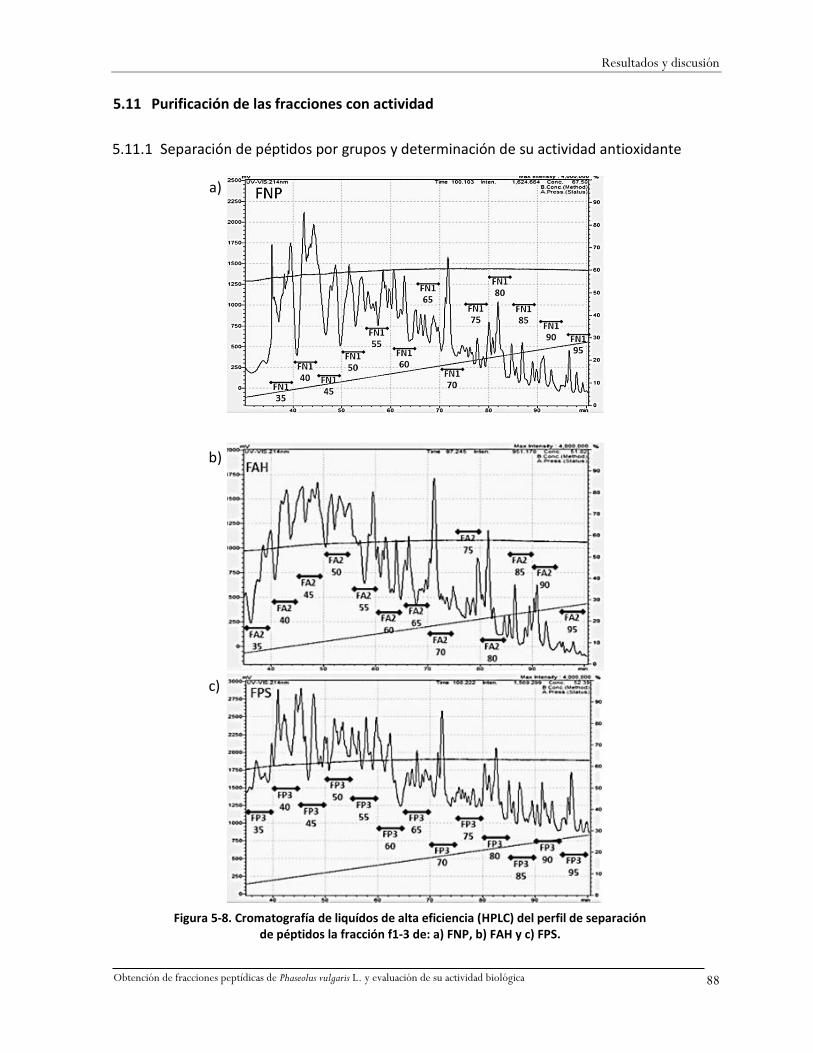
Resultados y discusión
88
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.11 Purificación de las fracciones con actividad
5.11.1 Separación de péptidos por grupos y determinación de su actividad antioxidante
Figura 5-8. Cromatografía de liquídos de alta eficiencia (HPLC) del perfil de separación de péptidos la fracción f1-3 de: a) FNP, b) FAH y c) FPS.
a)
b)
c)
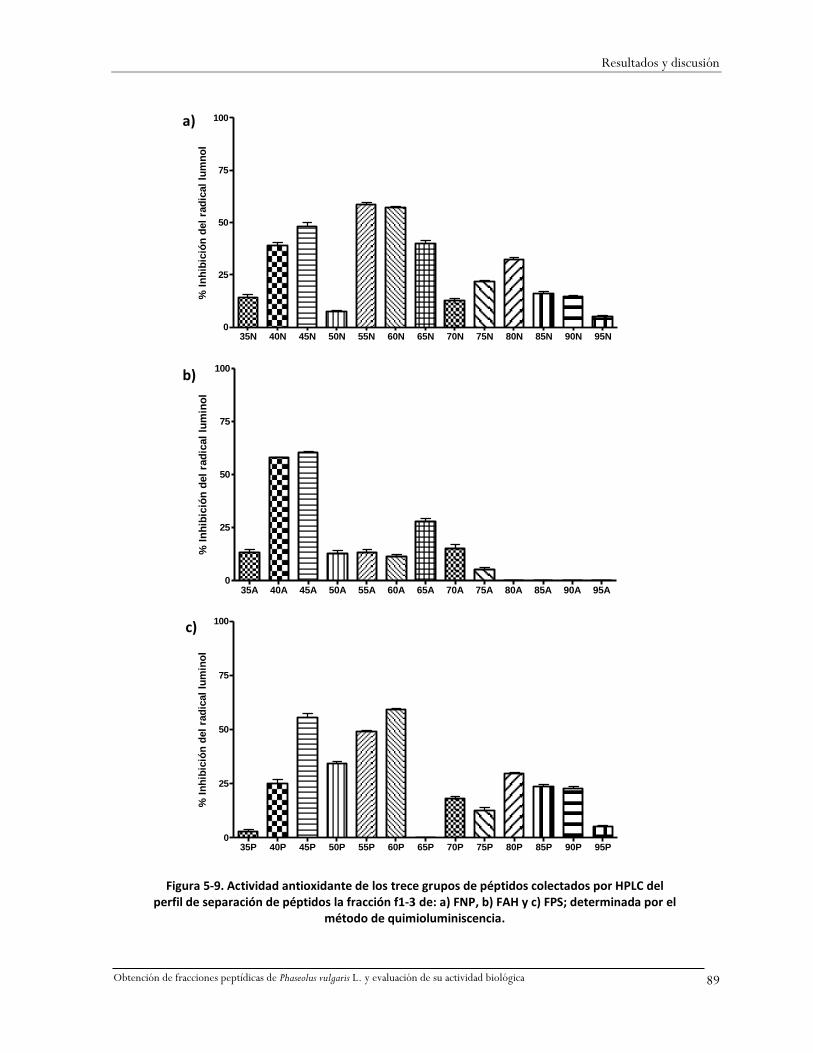
Resultados y discusión
89
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-9. Actividad antioxidante de los trece grupos de péptidos colectados por HPLC del perfil de separación de péptidos la fracción f1-3 de: a) FNP, b) FAH y c) FPS; determinada por el
método de quimioluminiscencia.
35N 40N 45N 50N 55N 60N 65N 70N 75N 80N 85N 90N 95N0
25
50
75
100
% I
nh
ibic
ión
del
rad
ical
lum
no
l
35A 40A 45A 50A 55A 60A 65A 70A 75A 80A 85A 90A 95A0
25
50
75
100
% I
nh
ibic
ión
del
rad
ical
lum
ino
l
35P 40P 45P 50P 55P 60P 65P 70P 75P 80P 85P 90P 95P0
25
50
75
100
% I
nh
ibic
ión
del
rad
ical
lum
ino
l
a)
b)
c)

Resultados y discusión
90
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Los cromatogramas de los perfiles de los péptidos y los grupos obtenidos como resultado de la
separación por HPLC de los péptidos provenientes de frijol se presentan en la Figura 5-8. Al ser
colectadas las muestras en periodos de tiempo de 5 min, se obtuvieron 13 grupos de péptidos de
cada variedad que fueron posteriormente analizados por el método de quimioluminiscencia ya que
este método requiere la menor cantidad de muestra, ofrece alta sensibilidad y proporciona datos
reproducibles, esto para determinar en qué grupos posiblemente se encontraba el péptido o los
péptidos con mayor actividad antioxidante.
La actividad antioxidante para cada grupo de péptidos de las tres variedades de frijol fue
determinada según el método descrito en 4.12.2 y los resultados se presentan en la Figura 5-9. El
criterio para considerar el grupo como activo fue que presentara al menos el 50 % de inhibición del
radical luminol. En la variedad FNP se determinaron dos grupos importantes el 55N y 60N con
actividad antioxidante superior al 50 %. En la variedad FAH se determinaron dos grupos con actividad
importe, el 40A y 45A y en la variedad FPS el 45P y 60P. Estos grupos importantes fueron
concentrados y sometidos a cromatografía de líquidos de alta eficiencia (HPLC) por segunda vez.
5.11.2 Separación de péptidos individualmente y determinación de su actividad antioxidante
La separación individual de péptidos se realizó bajo la misma gradiente de concentración de
acetonitrilo que las corridas anteriores, a cada pico se le asignó una letra y un número para poderse
identificar en pruebas posteriores. En la Figura 5-10 se presenta uno de los cromatogramas obtenidos
para la variedad de frijol azufrado higuera y las dos fracciones con actividad de inhibición grupal
superior al 80 % (40A y 45A). Se observaron para esta variedad 12 péptidos, para el FNP seis péptidos
y para la variedad FPS cinco.
En el Cuadro 5-22 se presentan los resultados de la actividad antioxidante de cada pico
colectado individualmente para las tres variedades de frijol. La variedad FAH presentó actividad en los
picos 4A y 10A con porcentajes de inhibición superiores a 80 %, en la variedad FNP se determinó un
76 % de inhibición en el pico 2P, mientras que en la variedad FNP se determinaron los picos 2N y 5N
con actividad del 75%. Se colectaron los dos péptidos importantes de la variedad azufrado higuera
para las pruebas posteriores.
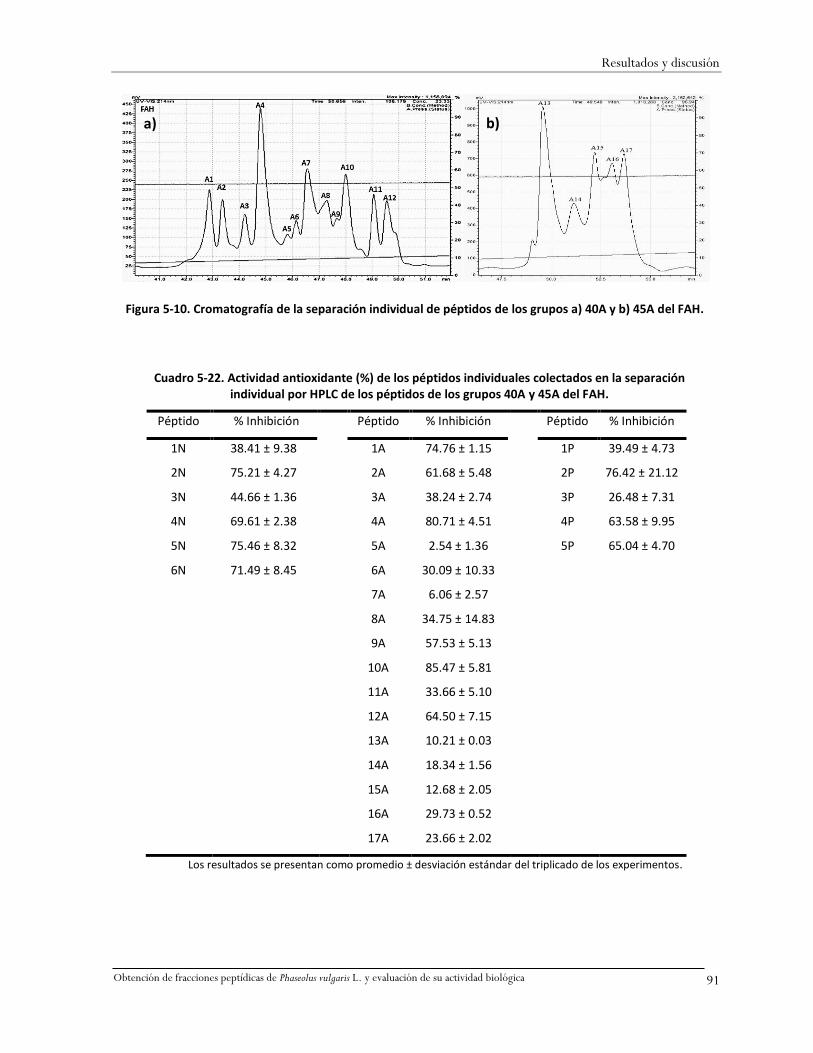
Resultados y discusión
91
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-10. Cromatografía de la separación individual de péptidos de los grupos a) 40A y b) 45A del FAH.
Cuadro 5-22. Actividad antioxidante (%) de los péptidos individuales colectados en la separación individual por HPLC de los péptidos de los grupos 40A y 45A del FAH.
Péptido % Inhibición
Péptido % Inhibición Péptido % Inhibición
1N 38.41 ± 9.38 1A 74.76 ± 1.15 1P 39.49 ± 4.73
2N 75.21 ± 4.27 2A 61.68 ± 5.48 2P 76.42 ± 21.12
3N 44.66 ± 1.36 3A 38.24 ± 2.74 3P 26.48 ± 7.31
4N 69.61 ± 2.38 4A 80.71 ± 4.51 4P 63.58 ± 9.95
5N 75.46 ± 8.32 5A 2.54 ± 1.36 5P 65.04 ± 4.70
6N 71.49 ± 8.45 6A 30.09 ± 10.33
7A 6.06 ± 2.57
8A 34.75 ± 14.83
9A 57.53 ± 5.13
10A 85.47 ± 5.81
11A 33.66 ± 5.10
12A 64.50 ± 7.15
13A 10.21 ± 0.03
14A 18.34 ± 1.56
15A 12.68 ± 2.05
16A 29.73 ± 0.52
17A 23.66 ± 2.02
Los resultados se presentan como promedio ± desviación estándar del triplicado de los experimentos.
a) b)
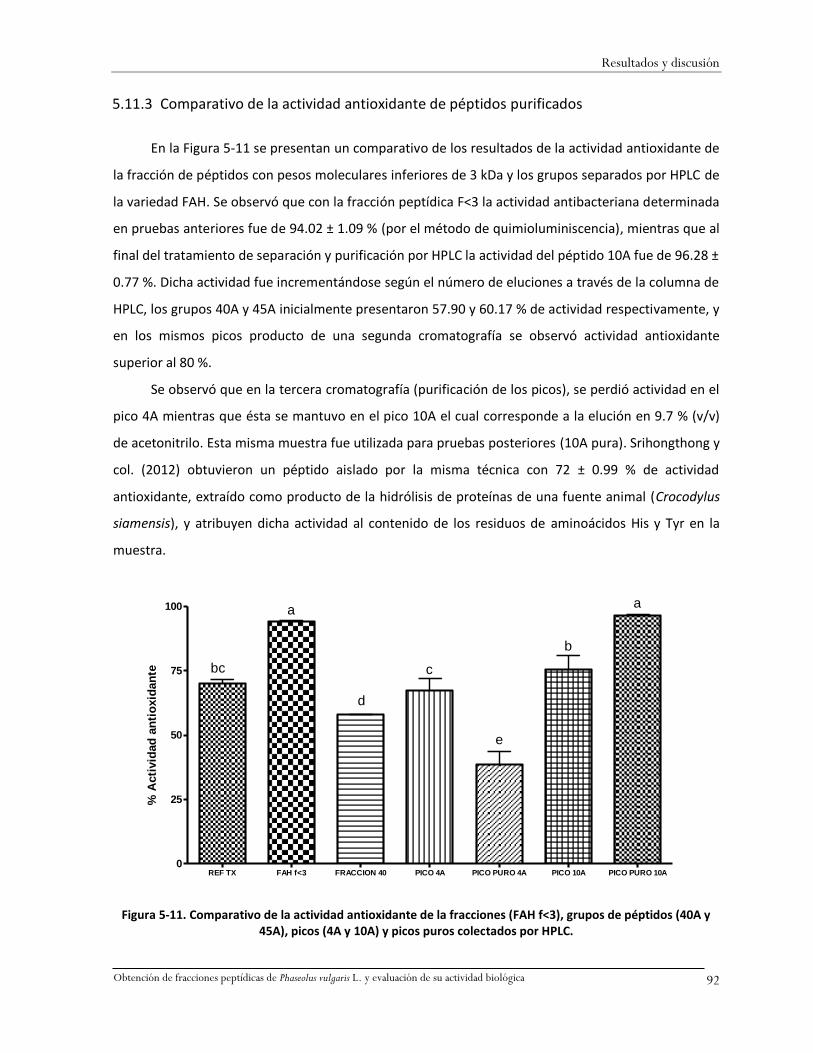
Resultados y discusión
92
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.11.3 Comparativo de la actividad antioxidante de péptidos purificados
En la Figura 5-11 se presentan un comparativo de los resultados de la actividad antioxidante de
la fracción de péptidos con pesos moleculares inferiores de 3 kDa y los grupos separados por HPLC de
la variedad FAH. Se observó que con la fracción peptídica F<3 la actividad antibacteriana determinada
en pruebas anteriores fue de 94.02 ± 1.09 % (por el método de quimioluminiscencia), mientras que al
final del tratamiento de separación y purificación por HPLC la actividad del péptido 10A fue de 96.28 ±
0.77 %. Dicha actividad fue incrementándose según el número de eluciones a través de la columna de
HPLC, los grupos 40A y 45A inicialmente presentaron 57.90 y 60.17 % de actividad respectivamente, y
en los mismos picos producto de una segunda cromatografía se observó actividad antioxidante
superior al 80 %.
Se observó que en la tercera cromatografía (purificación de los picos), se perdió actividad en el
pico 4A mientras que ésta se mantuvo en el pico 10A el cual corresponde a la elución en 9.7 % (v/v)
de acetonitrilo. Esta misma muestra fue utilizada para pruebas posteriores (10A pura). Srihongthong y
col. (2012) obtuvieron un péptido aislado por la misma técnica con 72 ± 0.99 % de actividad
antioxidante, extraído como producto de la hidrólisis de proteínas de una fuente animal (Crocodylus
siamensis), y atribuyen dicha actividad al contenido de los residuos de aminoácidos His y Tyr en la
muestra.
Figura 5-11. Comparativo de la actividad antioxidante de la fracciones (FAH f<3), grupos de péptidos (40A y 45A), picos (4A y 10A) y picos puros colectados por HPLC.
REF TX FAH f<3 FRACCION 40 PICO 4A PICO PURO 4A PICO 10A PICO PURO 10A0
25
50
75
100 aa
b
bc c
d
e
% A
cti
vid
ad
an
tio
xid
an
te
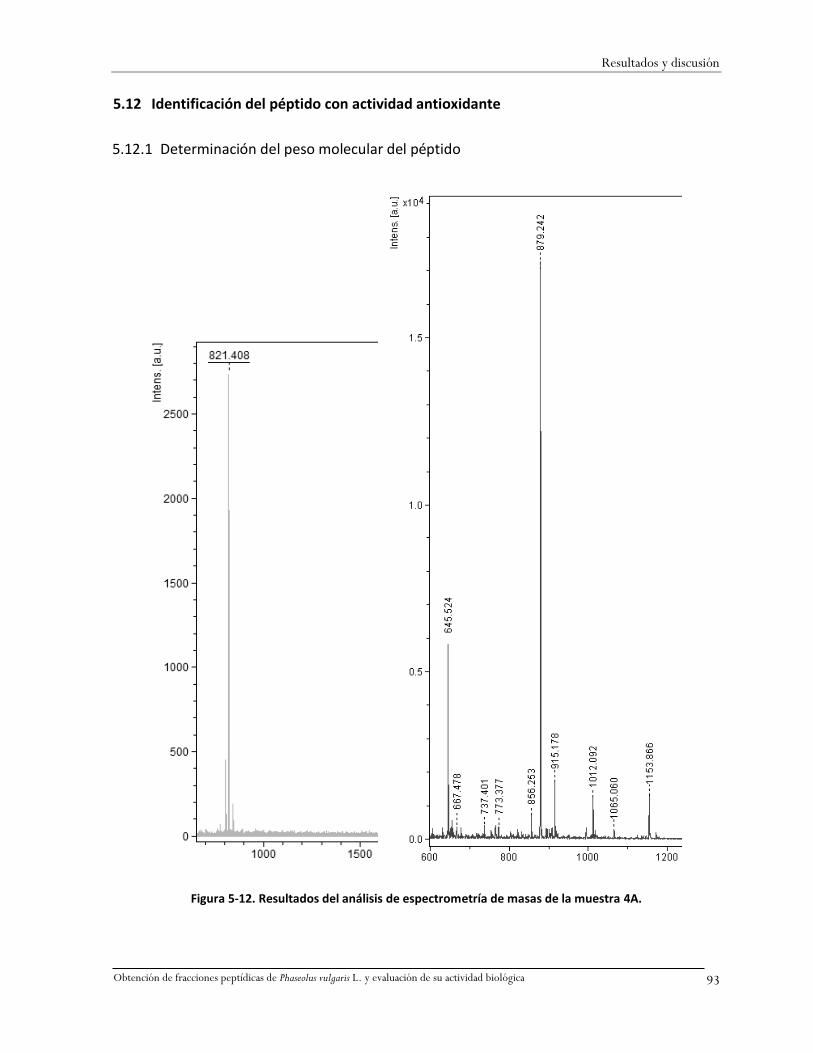
Resultados y discusión
93
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
5.12 Identificación del péptido con actividad antioxidante
5.12.1 Determinación del peso molecular del péptido
Figura 5-12. Resultados del análisis de espectrometría de masas de la muestra 4A.
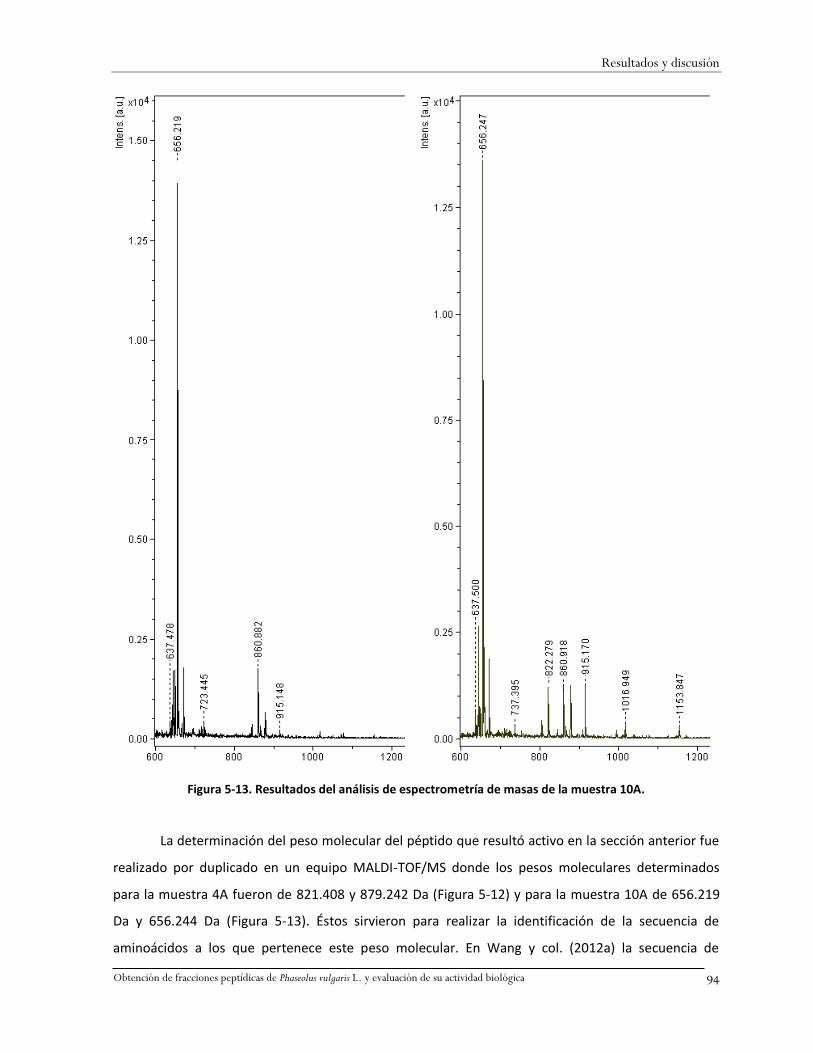
Resultados y discusión
94
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Figura 5-13. Resultados del análisis de espectrometría de masas de la muestra 10A.
La determinación del peso molecular del péptido que resultó activo en la sección anterior fue
realizado por duplicado en un equipo MALDI-TOF/MS donde los pesos moleculares determinados
para la muestra 4A fueron de 821.408 y 879.242 Da (Figura 5-12) y para la muestra 10A de 656.219
Da y 656.244 Da (Figura 5-13). Éstos sirvieron para realizar la identificación de la secuencia de
aminoácidos a los que pertenece este peso molecular. En Wang y col. (2012a) la secuencia de
Resultados y discusión
95
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
aminoácidos de dos péptidos provenientes de una fuente de origen animal (Sphyrna lewini) fueron
identificadas como Trp – Asp – Arg (PM 475.50 Da) y Pro – Tyr – Phe – Asn – Lys (PM 667.80 Da); éste
último peso molecular es cercano a los obtenidos en este trabajo aunque en éste caso provienen de
una fuente vegetal (frijol común).
5.12.2 Identificación molecular del péptido
Para la identificación del péptido activo, los pesos moleculares 821.408, 879.242, 656.219 y
656.244 Da fueron ingresados en el sistema ExPASy (Bioinformatics Resource Portal, Suiza) con la
herramienta Find Peptide y basado en la secuencia de aminoácidos de la faseolina (UniProt: P02853,
Cuadro 5-23) y de la lectina (UniProt: Q8RVH1, Cuadro 5-24) que son las proteínas más abundantes
en las semillas de frijol. El sistema virtual ExPASy presentó coincidencias entre la secuencia de
aminoácidos de ambas proteínas y los pesos moleculares obtenidos anteriormente por
espectrometría de masas.
Cuadro 5-23. Análisis de aminoácidos realizado en el sistema de predicción virtual ExPASY (UniProt: P02853).
Pico Masa ingresada
por el usuario
Masa generada
por ExPASY
Masa
(Daltons) Péptido Posición
4A 821.408 821.390 -0.017 (L) QRFDQQ (S) 61-66
821.426 0.018 (P) VNNPQIH (E) 162-168
821.452 0.043 (E) AHVELVGP (K) 295-302
821.477 0.068 (I) LFLASLSA (S) 14-21
879.242 879.497 0.255 (D) VFVIPAAY (P) 323-330
10A 656.219 656.263 0.044 (V) NEGEAH (V) 291-296
656.365 0.146 (G) TIFYL (V) 140-144
656.434 0.215 (I) VILVVN (E) 286-291
656.470 0.251 (S) KAIVIL (V) 283-288
656.247 656.263 0.016 (V) NEGEAH (V) 291-296
656.365 0.118 (G) TIFYL (V) 140-144
656.434 0.187 (I) VILVVN (E) 286-291
656.470 0.223 (S) KAIVIL (V) 283-288
La secuencia de aminoácidos de la faseolina cuenta con 421 residuos de aminoácidos, y no
hay cisteína. En ésta se encontraron cinco posibles secuencias para el péptido aislado del pico 4A y
cuatro secuencias para el péptido 10A las cuales se obtuvieron por duplicado. En dicha secuencia de
aminoácidos (Cuadro 5-23) se identificaron desde un pentapéptido hasta un octapéptido (Figura
5-14).
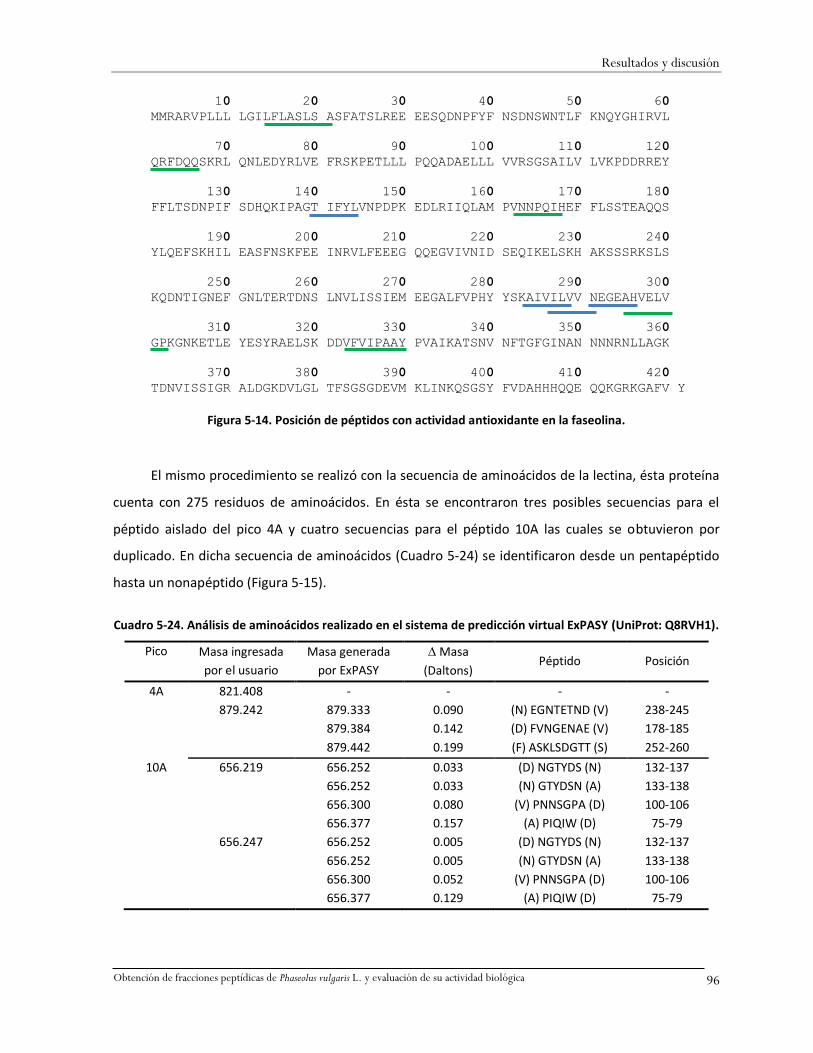
Resultados y discusión
96
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
10 20 30 40 50 60
MMRARVPLLL LGILFLASLS ASFATSLREE EESQDNPFYF NSDNSWNTLF KNQYGHIRVL
70 80 90 100 110 120
QRFDQQSKRL QNLEDYRLVE FRSKPETLLL PQQADAELLL VVRSGSAILV LVKPDDRREY
130 140 150 160 170 180
FFLTSDNPIF SDHQKIPAGT IFYLVNPDPK EDLRIIQLAM PVNNPQIHEF FLSSTEAQQS
190 200 210 220 230 240
YLQEFSKHIL EASFNSKFEE INRVLFEEEG QQEGVIVNID SEQIKELSKH AKSSSRKSLS
250 260 270 280 290 300
KQDNTIGNEF GNLTERTDNS LNVLISSIEM EEGALFVPHY YSKAIVILVV NEGEAHVELV
310 320 330 340 350 360
GPKGNKETLE YESYRAELSK DDVFVIPAAY PVAIKATSNV NFTGFGINAN NNNRNLLAGK
370 380 390 400 410 420
TDNVISSIGR ALDGKDVLGL TFSGSGDEVM KLINKQSGSY FVDAHHHQQE QQKGRKGAFV Y
Figura 5-14. Posición de péptidos con actividad antioxidante en la faseolina.
El mismo procedimiento se realizó con la secuencia de aminoácidos de la lectina, ésta proteína
cuenta con 275 residuos de aminoácidos. En ésta se encontraron tres posibles secuencias para el
péptido aislado del pico 4A y cuatro secuencias para el péptido 10A las cuales se obtuvieron por
duplicado. En dicha secuencia de aminoácidos (Cuadro 5-24) se identificaron desde un pentapéptido
hasta un nonapéptido (Figura 5-15).
Cuadro 5-24. Análisis de aminoácidos realizado en el sistema de predicción virtual ExPASY (UniProt: Q8RVH1).
Pico Masa ingresada
por el usuario
Masa generada
por ExPASY
Masa
(Daltons) Péptido Posición
4A 821.408 - - - -
879.242 879.333 0.090 (N) EGNTETND (V) 238-245
879.384 0.142 (D) FVNGENAE (V) 178-185
879.442 0.199 (F) ASKLSDGTT (S) 252-260
10A 656.219 656.252 0.033 (D) NGTYDS (N) 132-137
656.252 0.033 (N) GTYDSN (A) 133-138
656.300 0.080 (V) PNNSGPA (D) 100-106
656.377 0.157 (A) PIQIW (D) 75-79
656.247 656.252 0.005 (D) NGTYDS (N) 132-137
656.252 0.005 (N) GTYDSN (A) 133-138
656.300 0.052 (V) PNNSGPA (D) 100-106
656.377 0.129 (A) PIQIW (D) 75-79

Resultados y discusión
97
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
10 20 30 40 50 60
MASSNLLSLA LFLVLLTYAN SASETSFSFQ RFNETNLILQ RDATVSSKGQ LRLTNVNDNG
70 80 90 100 110 120
EPTLSSLGRA FYSAPIQIWD NTTGAVASFA TSFTFNIDVP NNSGPADGLA FVLVPVGSQP
130 140 150 160 170 180
KTNAGLLGLF DNGTYDSNAH TVAVEFDTCI NLGWDPKQRH IGIDVNSIKS IKTTPWDFVN
190 200 210 220 230 240
GENAEVLITY NSSTKLLVTS LVYPSQKTSF IISDRVELES VLPEWVSVGF SATSGINEGN
250 260 270
TETNDVLSWS FASKLSDGTT SEGLNLANSL LNQIL
Figura 5-15. Posición de péptidos con actividad antioxidante en la lectina.
Aunque la predicción del sistema virtual determinó las posibles secuencias, y sabiendo que la
enzima utilizada durante el proceso de hidrólisis no es específica, entonces se tiene la misma
posibilidad de encontrar a cada una de ellas. Positivamente, la posibilidad de tener Leu, Ile o Val en la
secuencia de aminoácidos del péptido incrementa el potencial del péptido como antioxidante o
antihipertensivo, según los trabajos previamente reportados (Martínez-Maqueda y col., 2012;
Yamada y col., 2008).
Hay trabajos donde se ha reportado la secuencia de péptidos conformados por seis
aminoácidos con actividad biológica, la novokinina (Arg-Pro-Leu-Lys-Pro-Trp) obtenida sintéticamente
basada en la secuencia de la ovokinina (Arg-Ala-Asp-His-Pro-Phe) que han sido usados como péptidos
con actividad biológica antihipertensiva (Yamada y col., 2008). Por otra parte, Alemán y col. (2011)
identificaron una secuencia más grande (Gly-Pro-Leu-Gly-Leu-Leu-Gly-Phe-Leu-Gly-Pro-Leu-Gly-Leu-
Ser) con peso molecular de 1410.63 Da, algo mayor que la reportada en este trabajo, sin embargo
dentro del intervalo de pesos moleculares con actividad biológica, de hecho ésta secuencia presentó
tanto actividad antioxidante como antihipertensiva; y ambas actividades fueron atribuidas a residuos
de Leu e Hyp contenidas en el péptido.
La identificación del péptido aislado con actividad antioxidante in vitro comprobada podría
permitir su aplicación como sustituto de compuestos de origen químico sobre células, incluso su uso
podría extenderse debido a que es posible que éste péptido también tenga actividad antioxidante e
inhibidora de la enzima convertidora de la angiotensina. Además, es importante el conocimiento de
saber que los hidrolizados de frijol, un alimento de consumo cotidiano, puede proporcionar algunos
beneficios secundarios a su solo consumo. Y de manera contrastante, la identificación de aminoácidos

Resultados y discusión
98
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
podría permitir la síntesis química del péptido para generar nuevas alternativas para el control de
enfermedades o mejoramiento en la salud del ser humano.
Por lo que este trabajo contribuye al conocimiento del efecto biológico de los péptidos
obtenidos vía hidrólisis enzimática de proteínas de frijol común (Phaseolus vulgaris L.) de tres
variedades. Se demostró que los péptidos además de presentar tres actividades biológicas in vitro
(antioxidante, antimicrobiana e inhibidora de la enzima convertidora de la angiotensina), también
tuvieron actividad biológica antihipertensiva in vivo.
Cabe destacar que aún son necesarios más experimentos de evaluación de la actividad
biológica de péptidos de proteínas de frijol. In vitro existe la posibilidad de encontrar una amplia
gama de actividades con potencial biológico, e in vivo se deben hacer pruebas con diferentes
fracciones y concentraciones del péptido, y vía de administración oral. Inclusive, la caracterización de
la secuencia de aminoácidos de péptidos responsables de alguna actividad biológica permitiría el
desarrollo de alternativas seguras para enfermedades frecuentes en la población.

Conclusiones
99
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
6. Conclusiones
Los hidrolizados de proteínas y fracciones peptídicas de frijol presentaron actividad biológica in
vitro (antioxidante, antimicrobiana e inhibidora de la enzima convertidora de la angiotensina I) e in
vivo (antihipertensiva).
Los hidrolizados totales de proteínas de frijol presentaron actividad antibacteriana sobre cepas
indicadoras. Principalmente los péptidos de la F<1 kDa, en los cuales se observó inhibición de Shigella
dysenteriae con concentración mínima inhibitoria de 10 a 40 µg de péptidos, siendo la variedad
azufrado higuera la que presentó el mayor efecto de inhibición del crecimiento bacteriano.
Los hidrolizados totales y las fracciones peptídicas de frijol presentaron actividad antioxidante,
y se observó la tendencia a incrementar ésta actividad biológica al disminuir la masa molecular de los
péptidos. La actividad antioxidante máxima obtenida por los métodos espectrofotométricos fue de:
DPPH+ = 40.19 ± 3.83 % (FNP, f1-3), ABTS+ = 99.17 ± 0.91 % (FNP, f<1) y DMPD+ = 89.08 ± 2.46 % (FPS,
f<1); y por el método de quimioluminiscencia de 94.02 ± 1.09 % (FAH, f1-3).
Los hidrolizados totales y las fracciones peptídicas de frijol presentaron actividad inhibidora de
la enzima convertidora de la angiotensina. El mayor porcentaje de actividad fue del 95 % con la
fracción f1-3 kDa de las variedades azufrado higuera y pinto Saltillo. Y se comprobó la actividad
antihipertensiva de los péptidos de FAH (f3-10) in vivo.
Se determinó la masa de dos péptidos con actividad antioxidante (656.219 y 879.242 Da), y por
bioinformática se determinaron múltiples coincidencias en la secuencia de las proteínas faseolina (9
secuencias) y lectina (7 secuencias).

Perspectivas
100
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
7. Perspectivas
Los hidrolizados de proteínas de frijol y sus fracciones peptídicas demostraron tener la capacidad
de actuar como antimicrobianos, antioxidantes, e inhibidores de la enzima convertidora de la
angiotensina; por lo tanto, se sugiere ampliar el estudio orientándolo a otras actividades biológicas in
vitro como la antifúngica o la antiinflamatoria.
Respecto a la actividad antihipertensiva aún son necesarios más experimentos de evaluación in
vivo con las diferentes fracciones peptídicas, variando las concentraciones de los péptidos, y
aministrando la dosis vía oral.
Se debe establecer un esquema de purificación de péptidos o proteínas donde se combinen
varios métodos, de tal manera que al complementarse van dejando atrás fracciones que no interesan
y se va enriqueciendo la fracción pura (como cromatografía de intercambio iónico, cromatografía de
exclusión y al final cromatografía de líquidos de alta resolución garantizan la pureza de la muestra).

Referencias
101
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
8. Referencias
Acuña-Goycolea P. A. 2001. Estudio de la prefactibilidad técnico-económico para la obtención de hidrolizado de
proteína vegetal a partir de Lupino. Tesis de licenciatura. Universidad de la Frontera. Chile.
Adler-Nissen J. 1979. Determination of the degree of hydrolysis of food protein hydrolysates by
trinitrobenzenesulfonic acid. Journal of Agricultural Food Chemistry. 27(6): 1256-1262.
Adler-Nissen J. 1986. Enzymatic hydrolysis of food proteins. New York: Elsevier Applied Science Publication.
Estados Unidos.
Aguilar O., Rito-Palomares M., y Glatz C. E. 2009. Caracterización tridimensional de proteínas de soya mediante
electroforesis de dos dimensiones y partición en fases acuosas. Revista Mexicana de Ingeniería Química.
8(1): 57-65.
Ahumada-González R. E. 2007. Producción de un aislado de proteína vegetal: determinación de parámetros de
proceso y caracterización del producto. Tesis de licenciatura. Universidad de la Frontera. Chile.
Aimutis W. R. 2004. Bioactive properties of milk proteins with particular focus on anticariogenesis. The Journal
of Nutrition. 134(4): 989–995.
Alemán A., Giménez B., Pérez-Santin E., Gómez-Guillén M. C., y Montero P. 2011. Contribution of Leu and Hyp
residues to antioxidant and ACE-inhibitory activities of peptide sequences isolated from squid gelatin
hydrolysate. Food Chemistry. 125: 334–341.
Alvarado-Carrasco C. y Guerra M. 2010. Lactosuero como fuente de péptidos bioactivos. Anales Venezolanos de
Nutrición. 23(1): 42-49.
Amarowicz R., Dykes G. A., Pegg R. B. 2008. Antibacterial activity of tannin constituents from Phaseolus vulgaris,
Fagoypyrum esculentum, Corylus avellana and Juglans nigra. Fitoterapia. 79: 217-219.
Arcan I. y Yemenicioğlu A. 2007. Antioxidant activity of protein extracts from heat-treated or thermally
processed chickpeas and white beans. Food Chemistry. 103(2): 301-312.
Arcila N. 2011. Aplicación de un proceso enzimático para la obtención de un hidrolizado proteico a partir del
frijol bayo (Vigna sinensis). Revista Electrónica Encuentro Transdisciplinar. 1(1):10-34.
Asociación Oficial de Químicos Analistas (AOAC). 1997. En: William Horwitz (Ed.), Official methods of analysis.
17a edición. Washington, D. C. Estados Unidos.
Barrón-Contreras J. L. 2006. Variedades de frijol de temporal para la zona media de San Luis Potosí. Instituto
Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP).
Bauer A. W., Kirby W. M. M., Sherris J. C., y Turck M. 1966. Antibiotic susceptibility testing by a standardized
single disk method. Amer I C/in Pathol. 45: 493-496.
Beermann C., Euler M., Herzberg J., y Stahl B. 2009. Anti-oxidative capacity of enzymatically released peptides
from soybean protein isolate. European Food and Research Technology. 229: 637-644.
Belén-Camacho D. R., Moreno-Álvarez M. J., García D., Medina C. y Sidorovas A. 2007. Caracterización de un
hidrolizado proteico enzimático obtenido del pez Caribe colorado (Pygocentruscariba, HUMBOLDT,
1821). Interciencia. 32(3): 188-194.

Referencias
102
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Benítez R., Ibarz A. y Pagan J. 2008. Hidrolizados de proteína: procesos y aplicaciones. Acta Bioquímica Clínica
Latinoamericana. 42(2): 227-236.
Bing-Zhang S., Wang Z., y Ying-Xu S. 2009. Purification and characterization of a radical scavenging peptide from
rapeseed protein hydrolysates. J Am Oil Chem Soc. 86: 959-966.
Borja-Bravo M. y García-Salazar J. A. 2008. Políticas para disminuir las importaciones de frijol (Phaseolus vulgaris
L.) en México: un análisis por tipo de variedad. Agrociencia. 42: 949-958.
Bradford M. M. 1976. A rapid and sensitive method for the quantization of microgram quantities of protein
utilizing the principle of dye binding. Anal Biochem. 72: 248-254.
Boye J., Zare F. y Pletch A. 2010. Pulse proteins: Processing, characterization, functional properties and
applications in food and feed. Food Research International. 43: 414-431.
Cachofeiro V., Guan H., y Nasjletti A. 1991. Mecanismos de acción de los inhibidores de la ECA: nuevas
perspertivas. Hipertension. 10(1): 27-31.
Caessens P. W. J. R., Daamn W. F., Gruppen H., Visser S., y Voragen A. G. J. 1999. -Lactoglobulin hydrolysis. II.
Peptide identification, SH/SS exchange, and functional properties of hydrolysate fractions formed by the
action of plasmin. J Agric Food Chem. 47: 2980-2990.
Camacho-Espinoza M. K., Peinado-Guevara L. I., López-Valenzuela J. A., Valdéz-Ortíz A., Salinas-Pérez R. A.,
Moreno-Herrera C. G. y Medina-Godoy S. 2010. Caracterización proteómica de granos de frijol azufrado
(Phaseolus vulgaris) cultivados en el Estado de Sinaloa. Ra Ximhai. 6(1): 23-36.
Cardador-Martínez A., Loarca-Piña G. y Oomah B. D. 2002. Antioxidant activity in common beans (Phaseolus
vulgaris L.). Journal of Agricultural and Food Chemistry. 50: 6975 – 6980.
Carrasco-Castilla J., Hernández-Álvarez A. J., Jiménez-Martínez C., Jacinto-Hernández C., Alaiz M., Girón-Calle J.,
Vioque J. y Dávila-Ortiz G. 2012. Antioxidant and metal chelating activities of peptide fractions from
phaseolin and bean protein hydrolysates. Food Chemistry. 135: 1789–1795.
Centro de análisis estadístico de datos. GraphPad Prism versión 4.0. Demo disponible en: http://www.
graphpad.com/scientific-software/prism/#1
Chagas M. P., Santos J. C. C., Santos E. B. G. N., Oliveira T. D., y Korn M. 2009. Exploiting iminoquinone free
radical production for thiol based drugs determination in pharmaceutical formulations. J Braz Chem Soc.
20(9): 1646-1652.
Chan Y. S., Wong J. H., Fang E. F., Pan W. L., y Ng T. B. 2012. An antifungal peptide from Phaseolus vulgaris cv.
brown kidney bean. Acta Biochim Biophys Sin. 44 (4): 307-315.
Hsieh C. C., Hernández-Ledesma B., Jeong H. J., Park J. H., Lumen B. O. 2010. Complementary roles in cancer
prevention: Protease inhibitor makes the cancer preventive peptide lunasin bioavailable. PLoS ONE 5(1):
e8890. doi:10.1371/journal.pone.0008890.
Chung, H. J., Liu, Q., Donner, E., Hoover, R., Warkentin, T. D., y Vandenberg, B. 2007. Composition, molecular
structure, thermal properties, pasting characteristics and in vitro digestibility of starches from newly
released Canadian cultivars of pea, lentil and chickpea. Cereal Chemistry, 85(4): 471-479.
Chung, H. J., Liu, Q., Pauls K. P., Fan M. Z. y. Yada R. 2008. In vitro starch digestibility, expected glycemic index
and some physicochemical properties of starch and flour from common bean (Phaseolus vulgaris)
varieties grown in Canada. Food Research International. 41: 869-875.

Referencias
103
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Comisión Veracruzana de Comercialización Agropecuaria (COVECA). 2012. Monografía del frijol. Gobierno del
Estado de Veracruz. Disponible en: http://portal.veracruz.gob.mx/pls/portal/docs/PAGE/COVECAINICIO
/IMAGENES/ARCHIVOSPDF/ARCHIVOSDIFUSION/MONOGRAFIA%20FRIJOL%202011.PDF
Cushman D. W., y Cheung H. S. 1971. Spectrophotometric assay and properties of the angiotensin-converting
enzyme of rabbit lung. Biochem Pharm. 20: 1637-1648.
Dávalos E., Miguel M., Bartolomé B., y López-Fadiño R. 2004. Antioxidant activity of peptides derived from egg
white proteins by enzymatic hydrolysis. Journal of Food Proteins. 67(9): 1939-1944.
De Luna-Jiménez A. 2007. Composición y procesamiento de la soya para consumo humano. Investigación y
ciencia de la Universidad Autónoma de Aguascalientes. 15(37): 35-44.
Delgado-Sánchez P., Saucedo-Ruiz M., Guzmán-Maldonado S. H., Villordo-Pineda E., González-Chavira M.,
Fraire-Velázquez S., Acosta-Gallegos J. A. y Mora-Avilés A. 2006. An organoleptic plant regeneration
system for common bean (Phaseolus vulgaris L.). Plant Science. 170: 822-827.
Douglas P., y Garley M. 2010. Chemistry and light. Science in School. 14: 63-68.
Drago-Serrano M. E., López-López M. y Sainz-Espuñes T. R. 2006. Componentes bioactivos de alimentos
funcionales de origen vegetal. Revista mexicana de ciencias farmacéuticas. 37(4): 58-68.
Dziuba M. y Darewicz M. 2007. Food proteins as precursors of bioactive peptides: Classification into families.
Food Science and Technology International. 13(6): 393-404.
Egito A. S., Miclo L., López C., Adam A., Girardet J. M., Gaillard J. L. 2002. Separation and characterization of
Mares’ milk αs1-, β-, κ-caseins, γ-casein-Like, and proteose peptone component 5-like peptides. Journal
of Dairy Science. 85(4): 697-706.
Emani C., y Hall T. C. 2008. Phaseolin: Structure and evolution. The Open Evolution Journal. 2:66-74.
Fernández-Pachón M. S., Villaño D., Troncoso A. M., García-Parrilla M. C. 2006. Revisión de los métodos de
evaluación de la actividad antioxidante in vitro del vino y valoración de sus efectos in vivo. Archivos
latinoamericanos de nutrición. 56(2): Disponible en línea en: http://www.alanrevista.org/ediciones/
2006-2/actividad_antioxidante_vino.asp.
Figueroa-Hernández C. Y. 2007. Utilización del sistema proteolítico de Lactococcus lactis para la generación de
péptidos potencialmente activos. Tesis de maestría. UAM-I. México.
Fogliano V., Verde V., Randazzo G., y Ritieni A. 1999. Method for measuring antioxidant activity and its
application to monitoring the antioxidant capacity of wines. J Agric Food Chem. 47(3): 1035-1040.
Fraile M. E., García-Suárez M. D., Martínez-Bernal A. y Slomianski R. 2007. Nutritivas y apetecibles: conozca de
leguminosas comestibles. Parte I. Hojas, vainas y semillas. ContactoS. 66: 27-35.
Fraga-Silva R. A., Sorg B. S, Wankhede M., Deugd C., Jun J. Y., Baker M. B., Li Y., Castellano R. K., Katovich M. J.,
Raizada M. K. y Ferreira A. J. 2010. ACE2 activation promotes antithrombotic activity. Molecular
Medicine. 16(5-6): 210–215.
Fritz M., Vecchi B., Rinaldi G., y Añón M. C. 2011. Amaranth seed protein hydrolysates have in vivo and in vitro
antihypertensive activity. Food Chemistry. 126: 878-884.
Gallegos-Tintoré S., Chel-Guerrero L., Corzo-Ríos L. J., y Martínez-Ayala A. L. 2013. Péptidos con actividad
antioxidante de proteínas vegetales. En: Bioactividad de péptidos derivados de proteínas alimentarias.
Páginas 111-121. Barcelona, España.

Referencias
104
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Georgetti S. R., Casagrande R., Di Mambro V. M. Azzolini A. E. C. S., y Fonseca M. J. V. 2003. Evaluation of the
antioxidant activity of different flavonoids by the chemiluminescence Method. AAPS Pharm Sci. 5(2):
Article 20. Disponible en línea en: http://www.pharmsci.org.
Gómez-Ruiz J. A., López-Expósito I., Pihlanto A., Ramos M., Recio I. 2008. Antioxidant activity of ovine casein
hydrolysates: identification of active peptides by HPLC–MS/MS. European Food Research Technology.
227:1061–1067.
González-García V. M., Fernández-Machín L. M., Ruibal-León A. J., y Durán-Torres G. 2002. Inhibidores de la
enzima conversora de angiotensina. Revisión de estudios multicéntricos. Revista Cubana de Medicina.
41(5). Disponible en línea en: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0034-7523200200
0500007
Görg A., Obermaier C., Boguth G., Harder A., Scheibe B., Wildgruber R., Weiss W. 2000. The current state of
two-dimensional electrophoresis with immobilized pH gradients. Electroporesis. 21: 1037-1053.
Guadix A., Guadix E. M., Páez-Dueñas M. P., González-Tello P. y Camacho F. 2000. Procesos tecnológicos y
métodos de control en la hidrólisis de proteínas. Ars Pharmaceutica. 41(1): 78-89.
Gujska E., Reinhard D., y Khan K. 1994. Physicochemical properties of field pea, pinto and navy bean starches. J
Food Sci. 59(3): 634-636.
Haque E., y Chand R. 2008. Antihypertensive and antimicrobial bioactive peptides from milk proteins. European
Food Research and Technology. 227: 7-15.
Hernández-Álvarez A., Carrasco-Castilla J., Jiménez-Martínez C., y Dávila-Ortiz G. 2010. Production of Phaseolus
vulgaris L. seeds stressed protein hydrolysates with antihypertensive activity. International Conference
of Food Innovation. Universidad Politécnica de Valencia.
Hernández-Ledesma B., Dávalos A., Bartolomé B., y Amigo L. 2005. Preparation of antioxidant enzymatic
hydrolysates from r-lactalbumin and a-lactoglobulin. Identification of active peptides hy HPLC-MS/MS.
Journal of Agricultural and Food Chemistry. 53: 588-593.
Hernández-Ledesma B., Martín-Álvarez P. J., y Pueyo E. 2003. Assessment of the spectrophotometric method
for determination of angiotensin-converting-enzyme activity: Influence of the inhibition type. Journal of
Agricultural and Food Chemistry. 51(15): 4175-4179.
Huang H. W. 2000. Action of antimicrobial peptides: two-state model. Biochemistry 39: 1-6.
Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 2013. SAGARPA. México.
Disponible en: http://www.inifap.gob.mx/SitePages/default.aspx.
Jiménez-Arellanes M. A., Román-Cortés N. R., y García I. 2011. Antioxidant and antimicrobial activities of hexane
extracts and pure compounds from Aristolochia taliscana rhizome. Revista Mexicana de Ciencias
Farmacéuticas. 42(3): 35-41.
Jakubowski W., Ertel D., Bilinski T., Kedziora J., y Bartosz G. 1998. Luminil luminescence induced by oxidants in
antioxidant deficient yeast Saccharomyces cerevisiae. Biochemistry and molecular biology international.
45(1): 191-203.
Katayama K., Tomatsu M., Kawahara S., Yamauchi K., Fuchu H., Kodama Y., Kawamura Y., y Muguruma M. 2004.
Inhibitory profile of nonapeptide derived from porcine troponin C against angiotensin I-converting
enzyme. Journal of Agricultural Food Chemistry. 52: 771-775.

Referencias
105
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Keidar S., Kaplan M., y Gamliel-Lazarovich A. 2007. ACE2 of the heart: From angiotensin I to angiotensin (1–7).
Cardiovascular Research. 73: 463-469.
Kim D. O., Lee K. W., Lee H. J., y Lee C. Y. 2002. Vitamin C equivalent antioxidant capacity (VCEAC) of phenolic
phytochemicals. Journal of Agricultural and Food Chemistry. 50: 3713-3717.
Kim S. Y., Park P. S., y Ree K. C. 1990. Functional properties of enzyme modified soy protein isolate. J Agric Food
Chem. 38: 651-656.
Konneman E. W., Allen S. D., Janda W. M., Schreckenberger P. C., y Winn W. C. 2004. Diagnóstico
Microbiológico. 5a edición. Editorial médica panamericana. Argentina. Páginas: 1286-1287.
Knorr D. 1998. Technology aspects related to microorganisms in functional foods. Trends in Food Science and
Technology. 9: 295-306.
Kuskosky E. M., Asuero A. G., Troncoso A. M., Mancini-Filho J., Fett R. 2005 Aplicación de diversos métodos
químicos para determinar actividad antioxidante en pulpa de frutos. Ciênc Tecnol Aliment, Campinas.
25(4): 726-732.
Laemmli K. U. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4.
Nature. 227(5259): 680-685.
Langfield R. D., Scarano F. J., Heitzman M. E., Kondo M., Hammond G. B., y Neto C. C. 2004. Use of a modified
microplate bioassay method to investigate antibacterial activity in the Peruvian medicinal plant
Peperomia galioides. Journal of Ethnopharmacology. 94: 279–281.
Li G. H., Le G. W., Liu H., y Shi Y. H. 2005. Mung-bean protein hydrolysates obtained with Alcalase exhibit
angiotensin I-converting enzyme inhibitory activity. Food Science Technology International. 11(4): 281-
287.
Li G. H., Le G. W., Shi Y. H., Shrestha S. 2004. Angiotensin I converting enzyme inhibitory peptides derived from
food proteins and their physiological and pharmacological effects. Nutrition Research. 27: 469-486.
Li G. H., Qu M. R., Wan J. Z., y You J. M. 2007. Antihypertensive effect of rice protein hydrolysate with in vitro
angiotensin I-converting enzyme inhibitory activity in spontaneously hypertensive rats. Asia Pac J Clin
Nutr. 16(1): 275-280.
Li G. H., Shi Y. H., Liu H., Le G. W. 2006a. Antihypertensive effect of Alcalase® generated mung bean protein
hydrolysates in spontaneously hypertensive rats. Eur Food Res Technol. 222:733-736.
Li G. H., Wan J. Z., Le G. W., y Shi Y. H. 2006b. Novel angiotensin I-converting enzyme inhibitory peptides
isolated from Alcalase hydrolysate of mung bean protein. Journal of Peptide Science. 12: 509-514.
Lii C. Y., y Chang S. M. 1981. Characterization of red bean (Phaseolus vulgaris var. Aurea). Starch and noodle
quality. J. Food Science. 46(1): 78-81.
Liu J. R., Chen M. J., y Lin C. W. 2005. Antimutagenic and antioxidant properties of milk-kefir and soymilk-kefir.
Journal of Agricultural and Food Chemistry. 53: 2467-2474.
López-Expósito I., Quirós A., Amigo L. y Recio I. 2007. Casein hydrolysates as a source of antimicrobial,
antioxidant and antihypertensive peptides. Lait. 87:241-249.
López-Herrera A. 2002. Manual de prácticas de laboratorio de producción y tecnología de semillas. Universidad
Autónoma de Chapingo. México.

Referencias
106
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Lutz M. y Edel-León A. editores. 2009. Aspectos nutricionales y saludables de los productos de panificación.
Universidad de Valparaíso- Editorial. Chile.
Ma Y., Xiong Y. L., Zhai J., Zhu H., Dziubla T. 2010. Fractionation and evaluation of radical scavenging peptides
from in vitro digest of Buckwheat protein. Food Chem. 118: 582-588.
Makri E. A., y Doxastakis G. I. 2006. Emulsifying and foaming properties of Phaseolus vulgaris and P. coccineus
proteins. Food Chemistry. 98: 558-568.
Markland F. S., y Smith E. L. 1971. Subtilisins: primary structure, chemical and physical properties. En: The
Enzyme. New York: Academic Press. Páginas 561-608. Estados Unidos.
Martínez-Agustín O. y Martínez de Victoria-Muñoz E. 2006. Proteínas y péptidos en nutrición enteral. Nutrición
hospitalaria. 21(2): 1-14.
Martínez-Maqueda D., Miralles B., Recio I., Hernández-Ledesma B. 2012. Antihypertensive peptides from food
proteins: a review. Food Funct. 3(4): 350-361. Antihypertensive peptides from food proteins: a review.
Marxen K., Vanselow K. H., Lippemeier S., Hintze R., Ruser A. y Hansen V. F. 2007. Determination of DPPH
radical oxidation caused by methanolic extracts of some microalgal species by linear regression analysis
of spectrophotometric measurements. Sensors. 7: 2080-2095.
Matsuzaki K. 1998. Magainins as paradigm for the mode of action of pore forming polypeptides. Biochim
Biophys Acta. 1376: 391-400.
Medina-Godoy S., Ambriz-Pérez D. L., Fuentes-Gutiérrez C. I., Germán-Báez L. J., Gutiérrez-Dorado R. Reyes-
Moreno C., y Valdéz-Ortíz A. 2012. Angiotensin-converting enzyme inhibitory and antioxidative activities
and functional characterization of protein hydrolysates of hard-to-cook chickpeas. J Sci Food Agric. 92:
1974–1981.
Megías C., Pedroche J., Yust M. M., Alaiz M., Girón-Calle J., Millán F. y Vioque J. 2009. Purification of angiotensin
converting enzyme inhibitory peptides from sunflower protein hydrolysates by reverse-phase
chromatography following affinity purification. Food Science and Technology. 42: 228–232.
Meisel H. y Bockelmann W. 1999. Bioactive peptides encrypted in milk proteins: proteolytic activation and
tropho-functional properties. Antonie van Leeweenhoek. 76: 207-215.
Mohamad H., Abas F., Permana D., Lajis N. H., Ali A. M., Sukari M. A., Hin T. Y. Y., Kikizaki H., y Nakatani N. 2004.
DPPH free radical scavenger components from the fruits of Alpinia rafflesiana Wall. ex. Bak.
(Zingiberaceae). Z. Naturforsch. 59c: 811-815.
Möller N. P., Scholz-Ahrens K. E., Roos N. y Schrezenmeir J. 2008. Bioactive peptides and proteins from foods:
indication for health effects. European Journal of Nutrition. 47: 171-182.
Molyneux P. 2004. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant
activity. J Sci Technol. 26:211-219.
Montaño-Pérez K. y Vargas-Albores F. 2002. Péptidos antimicrobianos: un mecanismo de defensa ancestral con
mucho futuro. Interciencia. 27(1): 21-27.
Montgomery D. C. 2006. Diseño y análisis de experimentos. Segunda edición. Editorial Limusa Wiley. México. pp
686.

Referencias
107
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Nakahara T., Sano A., Yamaguchi H., Sugimoto K., Chikata H., Kinoshita E., y Uchida R. 2010. Antihypertensive
effect of peptide-enriched soy sauce-like seasoning and identification of its angiotensin I-converting
enzyme inhibitory substances. J Agric Food Chem. 58: 821-827.
Natarajan S. S., Xu C., Bae H., Caperna T. J., y Garrett W. M. 2006. Characterization of storage proteins in wild
(Glycine soja) and cultivated (Glycine max) soybean seeds using proteomic analysis. J Agric Food Chem.
54: 3114-3120.
Nissen-Meyer J., Nes I. F. 1997. Ribosomally synthesized antimicrobial peptides: Their function, structure,
biogenesis and mechanism of action. Arch Microbiol. 167: 67-77.
Norton I., Fryer P. y Moore S. 2006. Product/ Process Integration in food manufacture: Engineering Sustained
Health. American Institute of Chemical Engineers. 52(2): 1632-1640.
O’Brien A. D., LaVeck G. D., Thompson M. R., y Formal S. B. 1982. Production of Shigella dysenteriae type 1-like
cytotoxin by Escherichia coli. J Infect Dis. 146(6): 763-769.
O’Farrell P. H. 1975. High resolution two-dimensional electrophoresis of proteins. The Journal of Biological
Chemistry. 250: 4007-4021.
Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). 2013. Disponible en:
http://www.fao.org/home/es/.
Padilla F. C., Guédez T., Alfaro M. J., Regnault M. y Rincón-C A. M. 2010. Fraccionamiento y caracterización de
las proteínas solubles de la harina de nuez de Barinas (Caryodendrono rinocense K.). Rev Inst Nac Hig
“Rafael Rangel”. 41(1): 38-42.
Palencia Mendoza Y. 1999. Sustancias Bioactivas en Alimentos. Universidad de Zaragoza, España. Consultado en
octubre 2012. Disponible en: www.unizar.es/med_naturista/bioactivos%20en%20alimentos.pdf
Palma-Rodríguez H. M. 2008. Efecto de la hemisfericina refinada en la hidrólisis de proteínas de suero lácteo.
IPN-CEPROBI. Tesis de maestría. México.
Peña-Ramos E. A., y Xiong Y. L. 2009. Antioxidant activity of soy protein hydrolysates in a liposomal system.
Journal of Food Science. 67: 2952-2956.
Portal de recursos de bioinformática (ExPASy) del Instituto Suizo de Bioinformática (SIB). 2014. Disponible en
línea en: http://web.expasy.org/findpept/.
Ré R., Pellegrini R., Proteggente A., Pannala A., Yang M., y Rice-Evans C. 1999. Antioxidant activity applying an
improved ABTS radical cation decolorization assay. Free Radical Biology & Medicine 26(9/10): 1231-1237.
Rocha-Guzmán N. E., Herzog A., González-Laredo R. F., Ibarra-Pérez F. J., Zambrano-Galván G. y Gallegos-Infante
J. A. 2007. Antioxidant and antimutagenic activity of phenolic compounds in three different color groups
of common bean cultivars (Phaseolus vulgaris). Food Chemistry. 103:521-527.
Ronca-Testoni S. 1983. Direct spectrophotometric assay for angiotensin-converting enzyme in serum. Clinical
chemistry. 29(6): 1093-1096.
Rui X., Boye J. I., Simpson B. K., y Prasher S. O. 2013. Purification and characterization of angiotensin I-
converting enzyme inhibitory peptides of small red bean (Phaseolus vulgaris) hydrolysates. J Funct
Foods. 5: 1116-1124.

Referencias
108
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Ruiz-Ruiz J., Dávila-Ortiz G., Chel-.Guerrero L., y Betancour-Ancona D. 2013a. Angiotensin I-converting enzyme
inhibitory and antioxidant peptide fractions from hard-to-cook bean enzymatic hydrolysates. Journal of
Food Biochemistry 37: 26-35.
Ruiz-Ruiz J. C., Segura-Campos M., Betancour-Ancona D., y Chel-Guerrero L. 2013b. Capítulo 1. Proteínas y
péptidos biológicamente activos con potencial nutracéutico. En: Bioactividad de péptidos en proteínas
alimentarias. Páginas 11-28. México.
Salinas-Moreno Y., Rojas-Herrera L., Sosa-Montes E. y Pérez-Herrera P. 2005. Composición de antocianinas en
variedades de frijol negro (Phaseolus vulgaris L.) cultivadas en México. Agrociencia. 39: 385-394.
Samaranayaka A. G. P. & Li-Chan E. 2011. Food-derived peptidic antioxidants: A review of their production,
assessment, and potential applications. Journal of Functional Foods. 3(4): 229-254.
Sánchez R. G., Manriquez N., Martínez M., y López I. 2001. El frijol en México: competitividad y oportunidades
de desarrollo. Boletín Informativo FIRA Banco. México. 13(33): 1-74.
Sánchez-Contreras P. M. E. 2007. Obtención de un concentrado proteico a partir de torta de raps (Brassican
apus). Tesis de licenciatura. Universidad de la Frontera. Chile.
Sarmadi B. H., & Ismail A. 2010. Antioxidative peptides from food proteins: A review. Peptides. 31(10): 1949-
1956.
Shai Y. 1998. Mode of action of antibacterial peptides. En: Molecular mechanisms of immune responses in
insects. Editorial Chapman & Hall. Páginas 111-131. Londres, Reino Unido.
Shahin M. A., Symons S. J., Poysa V. W. 2006. Determining soya bean seed size uniformity with image analysis.
Biosystems Engineering. 94(2): 191-198.
Servicio de Información y Estadística Agroalimentaria y Pesquera (SIAP). 2013. Secretaría de Agricultura,
Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). México. Disponible en:
www.siap.gob.mx/wp-content/uploads/2013/EXCEL/SiembrasCosechas/IO/Frijol.xls
Siddiq M., Ravi R., Harte J. B., y Dolan K. D. 2010. Physical and functional characteristics of selected dry bean
(Phaseolus vulgaris L.) flours. LWT-Food Science and Technology. 43: 232-237.
Silveira R. M. B., Moreno M. S., y Molina B. B. 2003. Alimentos funcionales y nutrición óptima. ¿cerca o lejos?.
Revista Española de Salud Pública. 77: 317-331.
Sistema de análisis estadístico para Windows (Statistical analysis sysmem SAS). Versión 9.0. Disponible en:
http://www.sas.com/en_us/software/analytics/stat.html
Sistema de Información de Organismos Vivos Modificados (SIOVM). 2011. Frijol negro. Phaseolus vulgaris. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). México. Disponible en: http://www.conabio.gob.mx/conocimiento/bioseguridad/pdf/21059_sg7.pdf
Software para adquisición de datos multitareas para Windows. WinDaq versión Lite 2014. Disponible en:
http://www.dataq.com/products/software/acquisition.htm.
Srihongthong S., Pakdeesuwan A., Daduang S., Araki T., Dhiravisit A., y Thammasirirak S. 2012. Complete amino
acid sequence of globin chains and biological activity of fragmented crocodile hemoglobin (Crocodylus
siamensis). Protein Journal. 31(6): 466-76.
Sreerama Y. N., Sashikala V. B., y Pratape V. M. 2009. Expansion properties and ultrastructure of legumes: Effect
of chemical and enzyme pre-treatments. LWT-Food Science and Technology. 42: 44-49.

Referencias
109
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Suárez-López M. M., Kizlansky A. y López L. B. 2006. Evaluación de la calidad de las proteínas en los alimentos
calculando el escore de aminoácidos corregido por digestibilidad. Nutrición Hospitalaria. 21(1): 47-51.
Taroco R., Seija V. y Vignoli R. 2006. Métodos de estudio de la sensibilidad antibiótica. Capítulo 36. Páginas: 663-
671. Disponible en: http://www.higiene.edu.uy/cefa/2008/BacteCEFA36.pdf.
Tavares T. G., Monteiro K. M., Possenti A., Pintado M. E., Carvalho J. E. y Malcata F. X. 2011. Antiulcerogenic
activity of peptide concentrates obtained from hydrolysis of whey proteins by proteases from Cynara
cardunculus. International Dairy Journal. 21(12): 934–939.
Tessera V., Guida F., Juretic D., y Tossi A. 2012. Identification of antimicrobial peptides from teleosts and
anurans in expressed sequence tag databases using conserved signal sequences. FEBS Journal. 279: 724-
736.
Torres-Llanez M. J., Vallejo-Cordoba B. y González-Córdova A. F. 2005. Péptidos bioactivos derivados de las
proteínas de la leche. Revista electrónica ALAN. 55(2): 111-117.
Torruco-Uco J., Chel-Guerrero L., Martínez-Ayala A., Dávila-Ortíz G. y Betancour-Ancona D. 2009. Angiotensin-I
converting enzime inhibitory and antioxidant activities of protein hydrolysates from Phaseolus lunatus
and Phaseolus vulgaris sedes. LWT-Food Science and Technology. 42: 1597-1604.
Valdés S., Medeiros-Coelho C. M, Michelluti D. J. y García-Tramonte V. L. C. 2011. Association of genotype and
preparation methods on the antioxidant activity, and antinutrients in common beans (Phaseolus vulgaris
L.). LWT-Food Science and Technology. 44: 2104-2111.
Valdéz-Ortíz A., Fuentes-Gutiérrez C. I., Germán-Báez L. J., Gutiérrez-Dorado R., y Medina-Godoy S. 2012.
Protein hydrolysates obtained from Azufrado (sulphur yellow) beans (Phaseolus vulgaris): Nutritional,
ACE-inhibitory and antioxidative characterization. LWT-Food Science and Technology. 46: 91–96.
Vásquez-Cardeño A., Cala-Molina M., Miranda I., Tafurt-García G., Martínez-Morales J. y Stashenko E. E. 2007.
Actividad antioxidante y contenido total de fenoles de los extractos etanólicos de Salvia aratocensis,
Salvia sochensis, Bidens reptons y Montanoa ovalifolia. Scientia et Technica. 33: 205-207.
Vaucher R. A., Teixeira M. L., Brandelli A. 2010. Investigation of the cytotoxicity of antimicrobial peptide P40 on
eukaryotic cells. Current Microbiology. 60(1): 1-5.
Vermeirssen V., Augustins P., Morel N., Camp J. V., Opsomer A., Verstraete W. 2005. In vitro intestinal transport
and antihypertensive activity of ACE inhibitory pea and whey digests. Int J Food Sci Nutr. 56:415-430.
Villanueva A., Vioque J., Sánchez-Vioque R., Clemente A., Bautista J. y Millán F. 1999. Production of an
extensive sunflower protein hydrolysate by sequential hydrolysis with endo- and exo- proteases. Grasas
y aceites. 50(6): 472-476.
Vioque J., Sánchez-Vioque R., Clemente A., Pedroche J., Yust M. M., y Millán F. 2000. Péptidos bioactivos en
proteínas de reserva. Grasas y aceites. 51: 361-365.
Vioque J., Sánchez-Vioque R., Pedroche J., Del Mar-Yust M. y Millán F. 2001. Obtención y aplicaciones de
concentrados y aislados proteicos. Grasas y aceites. 52(2): 127-131.
Von-Gadow A., Joubert E., y Hansmann C. F. 1997. Comparision of the antioxidant activity of aspalathin with
that of the other plant phenols of rooibos tea (Aspalathus linearis), alpha-tocopherol, BHT, and BHA.
Journal of agricultural and food chemistry. 45: 632-638.
Vrese M. y Schrezenmeir J. 2008. Probiotics, prebiotics and synbiotics. Adv Biochem Engin/biotechnol. 111:1-66.

Referencias
110
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Wang B., Li Z. R., Chi C. F., Zhang Q. H. y Luo, H. Y. 2012a. Preparation and evaluation of antioxidant peptides
from ethanol-soluble proteins hydrolysate of Sphyrna lewini muscle. Peptides. 36(2): 240-250.
Wang J., Wang K., Lin S., Zhao P., Jones G., Trang H., Liu J. y Ye H. 2012b. Improvement of antioxidant activity of
peptides with molecular weights ranging from 1 to 10 kDa by PEF technology. International Journal of
Biological Macromolecules. 51: 244–249.
Wang N., Hatcher D. W., Tyler R. T., Toews R., y Gawalko E. J. 2010. Effect of cooking on the composition of
beans (Phaseolus vulgaris L.) and chickpeas (Cicer arietinum L.). Food Research International. 43: 589-
594.
Wang W., y González de Mejía E. 2005. A new frontier in soy bioactive peptides that may prevent age-related
chronic diseases. Compr Rev Food Sci F. 4: 63-76.
White J. W., y González A. 1990. Characterization of the negative association between seed yield and seed size
among genotypes of common bean. Field Crops Research. 23(3-4): 159-175.
Wong J. H. y Ng T. B. 2005. Lunatusin, a trypsin-stable antimicrobial peptide from lima beans (Phaseolus lunatus
L.). Peptides. 26: 2086-2092.
Wu J., Ding X. 2001. Hypotensive and physiological effect of angiotensin converting enzyme inhibitory peptides
derived from soy protein on spontaneously hypertensive rats. J Agric Food Chem. 49: 501-506.
Xiong J., Fang W., Fang W., Bai L., Huo J., Kong Y. y Yunman L. 2010. Anticoagulant and antithrombotic activity of
a new peptide pENW (pGlu-Asn-Trp). Journal of Pharmacy and Pharmacology. 61(1): 89–94.
Yamada Y., Nishizawa K., Yokoo M., Zhao H., Onishi K., Teraishi M., Utsumi S., Ishimoto M. y Yoshikawa M. 2008.
Anti-hypertensive activity of genetically modified soybean seeds accumulating novokinin. Peptides. 29:
331-337.
Yust M. M., Millán-Linares M. C., Alcaide-Hidalgo J. M., Millán F. y Pedroche J. 2012. Hypocholesterolaemic and
antioxidant activities of chickpea (Cicer arietinum L.) protein hydrolysates. Journal of Science and Food
Agricultural. 92: 1994–2001.
Zhang J., Zhang H., Wang L., Gou X., Wang X., y Yao H. 2010. Isolation and identification of antioxidative
peptides from rice endosperm protein enzymatic hydrolysate by consecutive chromatography. Food
Chem. 119: 226-234.
Zhang T., Li Y., Miao M., y Jiang B. 2011. Purification and characterization of a new antioxidant peptide from
chickpea (Cicer arietium L.) protein hydrolysates. Food Chem. 128: 28-33.
Zubirán S. 1996. Valor nutritivo de los alimentos de mayor consumo en México. Limusa-Noriega. México.

Anexos
111
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
9. Anexos
9.1 Productos derivados de ésta Tesis
Carteles en Congresos u otras reuniones científicas
Ariza-Ortega TJ, Salas CE, Capettiti L, Soares-Lemoes V, Oliver-Salvador MC, Yáñez-Fernández J. 2012.
Determinación quimioluminométrica de la actividad antioxidante en péptidos de frijol
azufrado higuera (Phaseolus vulgaris L.). VII Congreso Internacional de Ingeniería Bioquímica.
Ariza-Ortega TJ, Ramírez-Sotelo MG, Valencia-Del Toro G, Oliver-Salvador MC, Yáñez-Fernández J.
2012. Evaluation of the antioxidant activity of Phaseolus vulgaris L. peptides by DPPH. 5th
International Congress of Food Science and Food Biotechnology in developing countries.
Ariza-Ortega TJ, Berber-Aceves GE, Ramírez-Sotelo MG, Yáñez-Fernández J. 2013. Phytochemical
screening of methanolic extracts from Jamapa and Zacatecas black bean (Phaseolus vulgaris
L.). XV National Congress of Biotechnology and Bioengineering.
Ariza-Ortega TJ, Berber-Aceves GE, Ramírez-Sotelo MG, Yáñez-Fernández J. 2013. Antioxidant activity
of methanolic extracts from Jamapa and Zacatecas black bean (Phaseolus vulgaris L.). XV
National Congress of Biotechnology and Bioengineering.
Ariza-Ortega TJ, Oliver-Salvador MC, Ramírez-Sotelo MG, Yáñez-Fernández J. 2014. Reducción del
radical libre DMPD como indicador de la actividad antioxidante de hidrolizados de proteínas
aisladas de frijol (Phaseolus vulgaris L.). VIII Congreso Internacional de Ingeniería Bioquímica.
Exposiciones orales en Congresos u otras reuniones científicas
Ariza-Ortega TJ, Ramírez-Sotelo MG, Valencia-Del Toro G, Oliver-Salvador MC, Yañez-Fernández J.
2012. Evaluación de la actividad antimicrobiana determinada por difusión en agar de péptidos
de frijol azufrado higuera (Phaseolus vulgaris L.). VII Congreso Internacional de Ingeniería
Bioquímica.
Ariza-Ortega TJ, Salas-Bravo CE, Capettiti L, Soares-Lemoes V, Oliver-Salvador MC, Yáñez-Fernández J.
2012. Antioxidant activity of Phaseolus vulgaris L. peptides by the chemiluminescence
method. 5th International Congress of Food Science and Food Biotechnology in developing
countries.

Anexos
112
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
Artículos
Ariza-Ortega TJ, Berber-Aceves GE, Ramírez-Sotelo MG, Yáñez-Fernández J. 2013. Methanolic
extracts antioxidant and antimicrobial activities from five varieties of common beans (Phaseolus
vulgaris L.). International Journal of Engineering Research & Technology. 2(11): 3196-3201.
Ariza-Ortega TJ, Zenón-Briones EY, Castrejón-Flores JL, Yáñez-Fernández J, Gómez-Gómez YM, Oliver-
Salvador MC. 2014. Angiotensin I-converting enzyme inhibitory, antimicrobial and antioxidant
effect of bioactive peptides obtained from different varieties of common beans (Phaseolus
vulgaris L.) with in vivo antihypertensive activity in spontaneously hypertensive rats. European
Food Research and Technology. DOI: 10.1007/s00217-014-2271-3. En línea.
Ariza-Ortega TJ, Oliver-Salvador MC, Ramírez-Sotelo MG, Valencia-Del Toro G, Salas-Bravo CE,
Capettini L, Yáñez-Fernández J. Obtención e identificación de péptidos con potencial actividad
antioxidante de concentrados proteicos de tres variedades de frijol (Phaseolus vulgaris L.).
Advances in Bioresearch. En revisión.
Ariza-Ortega TJ, Berber-Aceves GE, Yáñez-Fernández J. Propiedades tecnofuncionales de los
concentrados proteicos de cinco variedades de frijol (Phaseolus vulgaris L.). En proceso.
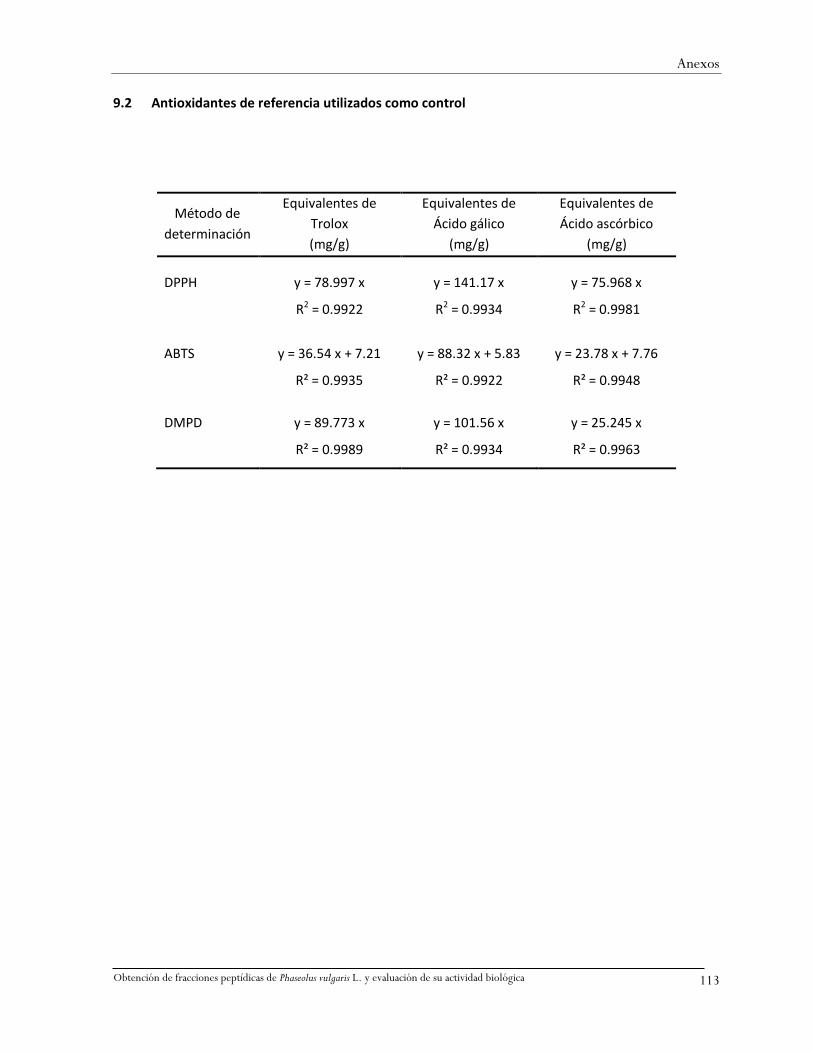
Anexos
113
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
9.2 Antioxidantes de referencia utilizados como control
Método de
determinación
Equivalentes de
Trolox
(mg/g)
Equivalentes de
Ácido gálico
(mg/g)
Equivalentes de
Ácido ascórbico
(mg/g)
DPPH y = 78.997 x y = 141.17 x y = 75.968 x
R2 = 0.9922 R2 = 0.9934 R2 = 0.9981
ABTS y = 36.54 x + 7.21 y = 88.32 x + 5.83 y = 23.78 x + 7.76
R² = 0.9935 R² = 0.9922 R² = 0.9948
DMPD y = 89.773 x y = 101.56 x y = 25.245 x
R² = 0.9989 R² = 0.9934 R² = 0.9963
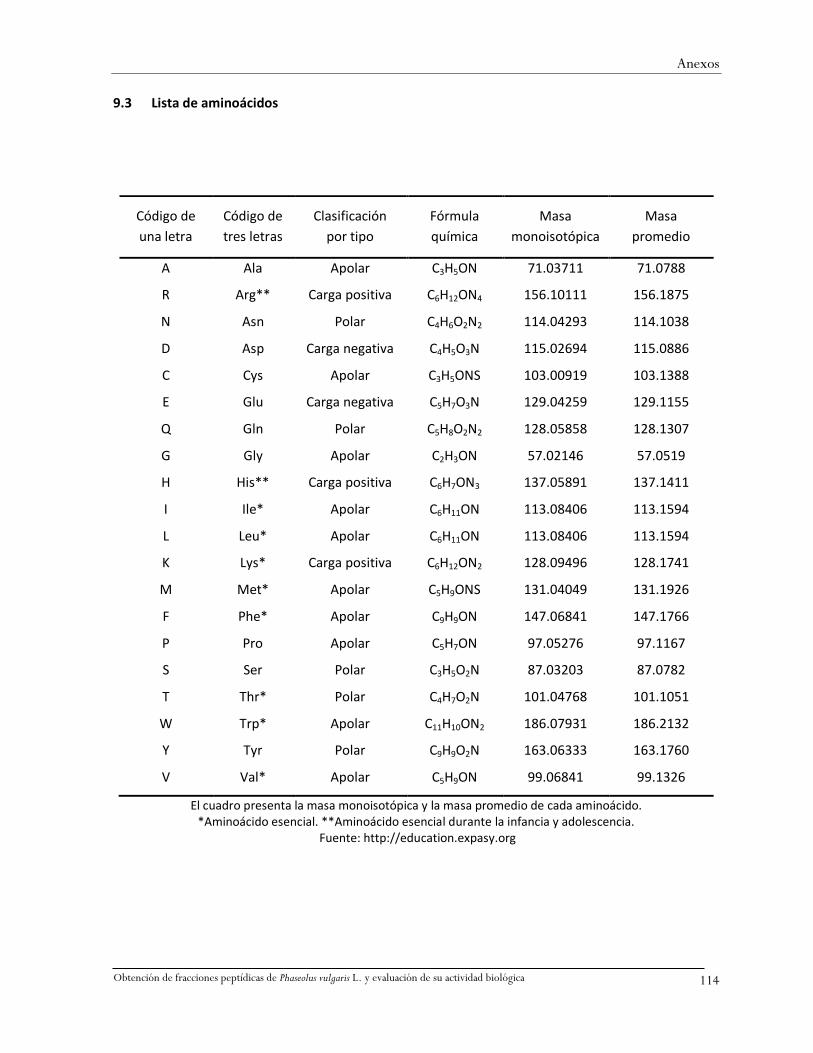
Anexos
114
Obtención de fracciones peptídicas de Phaseolus vulgaris L. y evaluación de su actividad biológica
9.3 Lista de aminoácidos
Código de
una letra
Código de
tres letras
Clasificación
por tipo
Fórmula
química
Masa
monoisotópica
Masa
promedio
A Ala Apolar C3H5ON 71.03711 71.0788
R Arg** Carga positiva C6H12ON4 156.10111 156.1875
N Asn Polar C4H6O2N2 114.04293 114.1038
D Asp Carga negativa C4H5O3N 115.02694 115.0886
C Cys Apolar C3H5ONS 103.00919 103.1388
E Glu Carga negativa C5H7O3N 129.04259 129.1155
Q Gln Polar C5H8O2N2 128.05858 128.1307
G Gly Apolar C2H3ON 57.02146 57.0519
H His** Carga positiva C6H7ON3 137.05891 137.1411
I Ile* Apolar C6H11ON 113.08406 113.1594
L Leu* Apolar C6H11ON 113.08406 113.1594
K Lys* Carga positiva C6H12ON2 128.09496 128.1741
M Met* Apolar C5H9ONS 131.04049 131.1926
F Phe* Apolar C9H9ON 147.06841 147.1766
P Pro Apolar C5H7ON 97.05276 97.1167
S Ser Polar C3H5O2N 87.03203 87.0782
T Thr* Polar C4H7O2N 101.04768 101.1051
W Trp* Apolar C11H10ON2 186.07931 186.2132
Y Tyr Polar C9H9O2N 163.06333 163.1760
V Val* Apolar C5H9ON 99.06841 99.1326
El cuadro presenta la masa monoisotópica y la masa promedio de cada aminoácido. *Aminoácido esencial. **Aminoácido esencial durante la infancia y adolescencia.
Fuente: http://education.expasy.org