Embed Size (px)
Citation preview

643GEODIVERSITAS • 2004 • 26 (4) © Publications Scientifiques du Muséum national d’Histoire naturelle, Paris. www.geodiversitas.com
The pes of Pyramiodontherium bergi(Moreno & Mercerat, 1891) (Mammalia, Xenarthra,Phyllophaga): the most complete pes of a Tertiary Megatheriinae
Diego BRANDONICentro de Investigación Científica y de Transferencia Tecnológica a la Producción (CICYTTP),
Materi y España, 3105 Diamante, Entre Ríos (Argentina)and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
Alfredo A. CARLINIDepartamento Científico de Paleontología de Vertebrados,
Museo de La Plata, Paseo del Bosque s/n°, 1900 La Plata (Argentina)and Cátedra de Anatomía Comparada, Facultad de Ciencias Naturales y Museo (UNLP),
calle 122 y 60, 1900 La Plata (Argentina)and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
François PUJOSInstitut français d’Études andines, Casilla 18-1217, av. Arequipa 4595, Lima 18 (Peru)
and Département Histoire de la Terre, Muséum national d’Histoire naturelle, UMR 5143 du CNRS,case postale 38, 8 rue Buffon, F-75231 Paris cedex (France)
Gustavo J. SCILLATO-YANÉDepartamento Científico de Paleontología de Vertebrados,
Museo de La Plata, Paseo del Bosque s/n°, 1900 La Plata (Argentina)and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
Brandoni D., Carlini A. A., Pujos F. & Scillato-Yané G. J. 2004. — The pes of Pyra-miodontherium bergi (Moreno & Mercerat, 1891) (Mammalia, Xenarthra, Phyllophaga): themost complete pes of a Tertiary Megatheriinae. Geodiversitas 26 (4) : 643-659.
ABSTRACTThe aim of this paper is to enlarge the description of the only knowncomplete pes of a Tertiary megatherine and make comparisons with those ofother members of the subfamily. The complete pes of the type specimen ofPyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66) was foundat the informally named “Araucanense” s.l. levels of Bajo de Andalhuala,Catamarca Province (late Miocene-Pliocene), Argentina. The elements of thepes show the general characters of other megatherines. However, the astra-galar depression is not as deep as in Megatherium americanum Cuvier, 1796;the calcaneum is more slender than Eremotherium laurillardi (Lund, 1842);the navicular has two facets to articulate with the ectocuneiform; there are

Brandoni D. et al.
644 GEODIVERSITAS • 2004 • 26 (4)
seven separate tarsal elements (calcaneum, astragalus, navicular, ento-cuneiform, mesocuneiform, ectocuneiform, and cuboid) while in othermegatherines, usually the mesocuneiform and the entocuneiform are fusedinto a single element termed the mesoentocuneiform complex; the third digitis formed only by the proximal phalanx fused with the second phalanx, anddigit IV, unlike Quaternary megatherines, has three phalanges. The presenceof separated entocuneiform and mesocuneiform and three phalanges in thedigit IV, are interpreted as plesiomorphic condition of the characters presentin the Plio-Pleistocene genera. The plesiomorphic character states observedhere could be useful to solve the megatherine polytomy proposed by De Iuliis(1996), and indicate more precisely the way the states of the characters werechanged.
RÉSUMÉLe pied de Pyramiodontherium bergi (Moreno & Mercerat, 1891) (Mammalia,Xenarthra, Phyllophaga) : le pied le plus complet d’un Megatheriinae tertiaire.Ce travail se propose de compléter la description du seul pied complet demégathère tertiaire connu et ainsi de pouvoir réaliser des comparaisons avecceux des autres membres de la sous-famille. Le pied complet de l’holotype dePyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66) provientdu niveau « Araucanense » s.l. de Bajo de Andalhuala, Province de Catamarca(Miocène supérieur-Pliocène), Argentine. Les éléments du pied présentent lastructure habituellement rencontrée chez les mégathères. Cependant, ladépression astragalienne est moins profonde que chez Megatherium america-num Cuvier, 1796 et le calcanéum est moins gracile que chez Eremotheriumlaurillardi (Lund, 1842). Le naviculaire présente deux facettes articulairespour l’ectocunéiforme. Le pied est constitué de sept éléments (calcanéum,astragale, naviculaire, entocunéiforme, mésocunéiforme, ectocunéiforme etcuboïde) alors que chez les autres mégathères dont le pied est connu, méso-cunéiforme et entocunéiforme sont fusionnés en un seul élément, dénommécomplexe méso-entocunéiforme. Le troisième doigt n’est constitué que dedeux phalanges, la phalange proximale résultant de la fusion de deux pha-langes. Contrairement à tous les mégathères quaternaires, le doigt IV deP. bergi possède trois phalanges. La présence d’un entocunéiforme et d’unmésocunéiforme séparés ainsi que de trois phalanges pour le doigt IV chezle genre plio-pléistocène est considérée ici comme étant la condition primi-tive. Les états de caractères plésiomorphes présents ici pourraient aider àrésoudre la polytomie des mégathères proposée par De Iuliis (1996) ainsi qu’àappréhender leurs différents changements.
KEY WORDSMammalia, Xenarthra,
Megatheriinae,Pyramiodontherium bergi,
Tertiary, pes,
morphology.
MOTS CLÉSMammalia, Xenarthra,
Megatheriinae, Pyramiodontherium bergi,
Tertiaire, pied,
morphologie.
INTRODUCTION
Quaternary Megatheriinae are among the bestknown of all extinct South American xenar-thrans. Paradoxically, the most ancient Mega-theriinae from the middle Miocene and Pliocenehave remained largely unknown because of the
relative scarceness of the material. Relying onlyon published accounts, there is strong uncertain-ty concerning the taxonomic validity, anatomicalcharacterization, and phylogenetic relationshipsof the described taxa.The oldest records of Megatheriinae date back asfar as the middle Miocene, and include the gen-

era of the “Friasian” interval, represented by threesuccessive faunas: Colloncuran, Laventan andMayoan (see Scillato-Yané & Carlini 1998, for adiscussion of the use and extension of the term“Friasian” to include the three faunas includedhere). These genera are: 1) MegathericulusAmeghino, 1904 (including M. patagonicusAmeghino, 1904 and M. primaevus Cabrera,1939, both from the Colloncuran [middleMiocene] of Patagonia); 2) a Megatheriinaeindet. from the Laventan (middle Miocene) ofColombia described by Hirschfeld (1985: 47-49)and probably closely related to Eomegatheriumnanum (Burmeister, 1892) (= Promegatherium forHirschfeld 1985); and 3) EomegatheriumKraglievich, 1926 (including both E. andinumKraglievich, 1930 and E. cabrerai Kraglievich,1930 from the Mayoan [middle Miocene] ofPatagonia). Despite Hirschfeld’s (1985) claimthat Megathericulus primaevus, a taxon based onan astragalus found in sediments attributed to theSantacrucian (lower Miocene) (Cabrera 1939), isthe oldest record of the subfamily Megatheriinae,we now know that these fossil-bearing sedimentsare somewhat younger and of Colloncuran age(Bondesio et al. 1980). We see then that old mis-takes in the systematic identification andchronology of the oldest Megatheriinae persist tothe present. Likewise, the general panorama oflate Miocene and Pliocene megathere evolutionhas not been fully clarified, despite numerouspapers on the subject such as the contributions ofCabrera (1928) and De Iuliis (1996). Elementsof the megatherine pes have been described byseveral authors, mainly Owen (1860), Roth(1911), Cabrera (1929), Paula Couto (1978),Cartelle (1992), De Iuliis (1996), and Pujos &Salas (2004). Except for the contributions ofRoth (1911) and De Iuliis (1996), the remainingpapers concern mainly Quaternary species ofMegatherium and Eremotherium. Both, Roth(1911) and De Iuliis (1996) described brieflysome bones of the pes of Pyramiodontheriumbergi (Moreno & Mercerat, 1891), and paid spe-cial attention to the astragalus and calcaneum ofthis “Araucanian” or late Miocene-Pliocenespecies.
The aim of this paper is to extend the descriptionof the only known complete pes of a Tertiarymegatherine, that of P. bergi, to include elementsthat have been less well studied, and to makemore detailed and extensive comparisons withthe same elements in other members of thefamily. The complete pes of the holotype ofP. bergi (MLP 2-66) is housed in the collection ofDepartamento Científico de Paleontología deVertebrados, Facultad de Ciencias Naturales yMuseo, Universidad Nacional de La Plata,Argentina. It was recovered from informallynamed “Araucanense” s.l. levels at Bajo deAndalhuala in Catamarca Province, Argentina, ofpresumed late Miocene-Pliocene age (see Carliniet al. 2002). According to Moreno & Mercerat(1891) the remains were collected by Methfesseland staff of the Museo de La Plata during archae-ological and paleontological explorations inCatamarca Province at the end of the 19th century.
ABBREVIATIONSFMNH Field Museum of Natural History,
Chicago;MACN Museo Argentino de Ciencias Naturales
“Bernardino Rivadavia”, Buenos Aires;MLP Museo de La Plata, La Plata;MNHN Muséum national d’Histoire naturelle,
Paris;MUSM Museo de Historia Natural de la
Universidad Mayor de San Marcos, Lima;ROM Royal Ontario Museum, Toronto;UNA Universidad de la Ingenieria, Lima.
SYSTEMATICS
Order XENARTHRA Cope, 1889Suborder PHYLLOPHAGA Owen, 1842Superfamily MEGATHERIOIDEA Gray, 1821
Family MEGATHERIIDAE Gray, 1821Subfamily MEGATHERIINAE Gray, 1821
Genus Megatherium Cuvier, 1796
Pyramiodontherium bergi(Moreno & Mercerat, 1891)
Megatherium burmeisteri Moreno & Mercerat, 1891:229.
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
645GEODIVERSITAS • 2004 • 26 (4)

Megatherium bergi Moreno & Mercerat, 1891: 231.
Pyramiodontherium dubium Rovereto, 1914: 89.
Pyramiodontherium bergi – Cabrera 1928: 344.
MATERIAL EXAMINED. — MLP 2-66 (holotype). Inaddition to the remains described and illustrated inRoth (1911), De Iuliis (1996), and Carlini et al.(2002), the following well preserved material is heredescribed: left and right astragali, left calcaneum, rightnavicular, left and right cuboids, left and right ecto-cuneiforms, left and right mesocuneiforms, left andright entocuneiforms, right metatarsal III, leftmetatarsal IV, right metatarsal V, phalanx 1+2 of leftand right digit III, left and right ungual phalanx, andphalanx 1 and 2 of right digit IV. Comparison speci-mens listed in Table 1.
COMPARATIVE DESCRIPTION
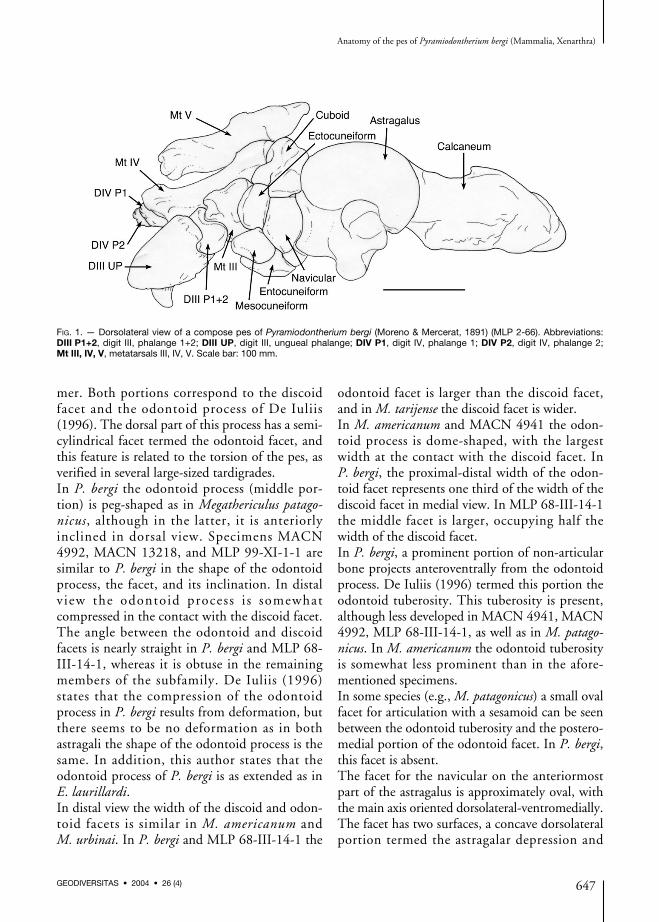
The foot bones of P. bergi (Fig. 1) are describedin anatomical position during locomotion. As inthe other megatherines, the pes has undergonetorsion, resulting in a deviation of its axis withrespect to the horizontal. Consequently, pedalelements are inclined along a dorsolateral-ventro-medial major axis.
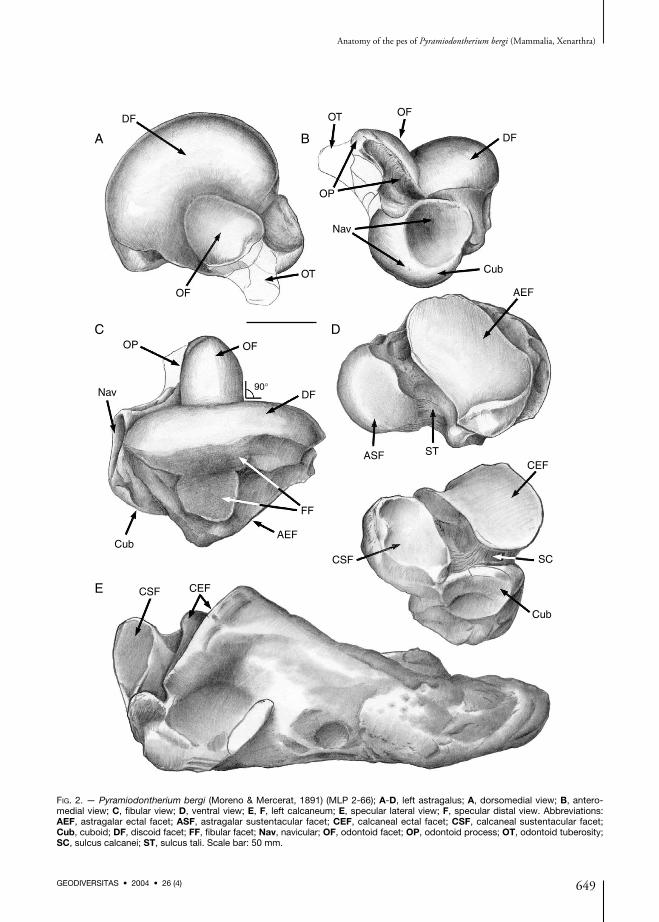
AstragalusThe astragalus of P. bergi (Fig. 2A-D), partiallydescribed by Roth (1911), has the typical shapefor the subfamily Megatheriinae. It is massive,
with a well developed central odontoid process indorsomedial view, and a navicular facet in anteriorview.As in other megatherines, the fibular facet inP. bergi is divided into two portions, an antero-posteriorly elongate dorsal division that joins thediscoid facet, and a ventral portion with a rathercircular surface that is slightly extended ventrally.This latter division has well defined marginsanteriorly. A similar morphology is also seen inMLP 68-III-14-1, but in this specimen the ven-tral portion is oval with the long axis perpendi-cular to the anteroposterior axis of the fibularfacet, and the dorsal portion is reduced. InMACN 4992 the ventral portion of the fibularfacet is similar in shape to that of P. bergi,although in some specimens of M. americanum(MLP 2-29), the ventral portion of the facet issemilunar in shape and in general moredepressed. On the basis of his observations onE. laurillardi, De Iuliis (1996) states that theshape of the fibular facet may have intraspecificvariation. In addition, he outlines the welldefined margins and a bilobate facet in M. ameri-canum (MNHN P1871).The troclea tali is composed of two portions: alateral, wide, discoid portion, and another semi-cylindrical, positioned in the middle of the for-
Brandoni D. et al.
646 GEODIVERSITAS • 2004 • 26 (4)
TABLE 1. — Specimens used for comparison.
Specimen No. Taxon Locality and age
MLP 91-IX-7-18 Megathericulus patagonicus Santa Cruz Province, Argentina. Middle MioceneMACN 4941 Megatherium? antiquum Entre Ríos Province, Argentina. Late MioceneMACN 4983 Megatherium? antiquum Entre Ríos Province, Argentina. Late MioceneMACN 4992 Eomegatherium nanum Entre Ríos Province, Argentina. Late MioceneMLP 99-XI-1-1 Pliomegatherium Entre Ríos Province, Argentina. Late MioceneMACN 13218 Pliomegatherium Entre Ríos Province, Argentina. Late MioceneMLP 68-III-14-1 Plesiomegatherium sp. La Rioja Province, Argentina. PlioceneMLP 2-29 Megatherium americanum Buenos Aires Province, Argentina. PleistoceneMNHN PAM 295 Megatherium americanum Buenos Aires Province, Argentina. PleistoceneROM 21928 Eremotherium laurillardi La Carolina, Ecuador and Daytona Beach, Florida, USAROM 21932ROM 22003ROM 22013ROM 22014ROM 30768FMNH P14216 Megatherium tarijense Tarija Valley, near Padcaya, Bolivia. PleistoceneMNHN TAR 269 Megatherium tarijense Tarija Valley, near Padcaya, Bolivia. PleistoceneMUSM 15 Megatherium urbinai Sacaco, Arequipa Department, Peru. PleistoceneUNA V2642 Megatherium urbinai Tres Ventanas Cave, Peru. Pleistocene
mer. Both portions correspond to the discoidfacet and the odontoid process of De Iuliis(1996). The dorsal part of this process has a semi-cylindrical facet termed the odontoid facet, andthis feature is related to the torsion of the pes, asverified in several large-sized tardigrades.In P. bergi the odontoid process (middle por-tion) is peg-shaped as in Megathericulus patago-nicus, although in the latter, it is anteriorlyinclined in dorsal view. Specimens MACN4992, MACN 13218, and MLP 99-XI-1-1 aresimilar to P. bergi in the shape of the odontoidprocess, the facet, and its inclination. In distalv iew the odontoid process i s somewhatcompressed in the contact with the discoid facet.The angle between the odontoid and discoidfacets is nearly straight in P. bergi and MLP 68-III-14-1, whereas it is obtuse in the remainingmembers of the subfamily. De Iuliis (1996)states that the compression of the odontoidprocess in P. bergi results from deformation, butthere seems to be no deformation as in bothastragali the shape of the odontoid process is thesame. In addition, this author states that theodontoid process of P. bergi is as extended as inE. laurillardi.In distal view the width of the discoid and odon-toid facets is similar in M. americanum andM. urbinai. In P. bergi and MLP 68-III-14-1 the
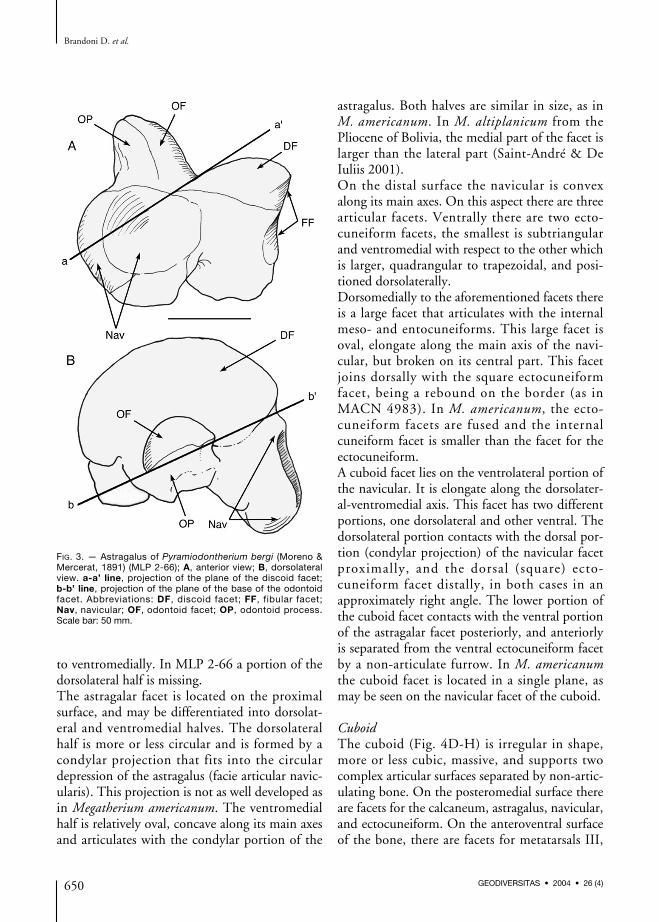
odontoid facet is larger than the discoid facet,and in M. tarijense the discoid facet is wider.In M. americanum and MACN 4941 the odon-toid process is dome-shaped, with the largestwidth at the contact with the discoid facet. InP. bergi, the proximal-distal width of the odon-toid facet represents one third of the width of thediscoid facet in medial view. In MLP 68-III-14-1the middle facet is larger, occupying half thewidth of the discoid facet.In P. bergi, a prominent portion of non-articularbone projects anteroventrally from the odontoidprocess. De Iuliis (1996) termed this portion theodontoid tuberosity. This tuberosity is present,although less developed in MACN 4941, MACN4992, MLP 68-III-14-1, as well as in M. patago-nicus. In M. americanum the odontoid tuberosityis somewhat less prominent than in the afore-mentioned specimens.In some species (e.g., M. patagonicus) a small ovalfacet for articulation with a sesamoid can be seenbetween the odontoid tuberosity and the postero-medial portion of the odontoid facet. In P. bergi,this facet is absent.The facet for the navicular on the anteriormostpart of the astragalus is approximately oval, withthe main axis oriented dorsolateral-ventromedially.The facet has two surfaces, a concave dorsolateralportion termed the astragalar depression and
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
647GEODIVERSITAS • 2004 • 26 (4)
FIG. 1. — Dorsolateral view of a compose pes of Pyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66). Abbreviations:DIII P1+2, digit III, phalange 1+2; DIII UP, digit III, ungueal phalange; DIV P1, digit IV, phalange 1; DIV P2, digit IV, phalange 2;Mt III, IV, V, metatarsals III, IV, V. Scale bar: 100 mm.

another ventromedial convex portion. The astra-galar depression is deeper in M. americanum thanin P. bergi, but similar to P. bergi in MACN4941, MACN 4992, MACN 13218, MLP 99-XI-1-1, and MLP 68-III-14-1.In P. bergi, the dorsomedial half of the navicularfacet is positioned dorsally with respect to theplane of the discoid facet (Fig. 3A). This situationis similar in M. patagonicus (MLP 68-III-14-1,MACN 4992, MACN 13218, MLP 99-XI-1-1,and MACN 4941). By contrast, in M. ameri-canum the dorsomedial part of the navicular facetis at the same level with discoid facet. At theother extreme, in E. laurillardi and M. urbinaithe dorsal margin projects by one third over thediscoid facet. The navicular facet in M. tarijensedisplays an intermediate position betweenM. americanum and M. urbinai.In dorsomedial view the odontoid facet is semi-cylindrical in section and the projection of thebase exceeds the navicular facet in P. bergi(Fig. 3B) as in all other Tertiary astragali. InM. americanum this plane cuts the navicular facetwhereas in MLP 68-III-14-1 the plane scarcelycuts the facet.The cuboid facet appears on the external side ofthe astragalus where it is slightly differentiatedfrom the convex portion of the navicular facet.The cuboid facet is convex and subtriangular topyriform in shape.There are two facets for articulation with the cal-caneum; the smaller sustentacular facet and theectal facet. These two facets are separated by adeep and wide non-articular bony canal, the sul-cus tali. A similar arrangement can be verified inmost megatherines except one specimen ofM. americanum (Kraglievich 1926), one speci-men of E. laurillardi (ROM 22006 in De Iuliis1996 and Pujos 2001 pers. obs.), one specimenof M. urbinai (UNA V2642 in Pujos & Salas2004) in which these facets are fused, and inM. tarijense (MNHN TAR 269 Pujos 2002 pers.obs.) where they are partially fused. In MLP 68-III-14-1, the bony canal is deep but narrow dueto the large size of the ectal facet.The sustentacular facet is continuous with thecuboid facet but is separated from the ventral
portion of the navicular facet. It is almost planaror flat and subtriangular in shape. The ectal facetis oval in shape, with the main axis anterolateralto posteromedial, parallel to the sulcus tali. Theectal facet is concave along the main axis andconvex along the other axis. In dorsolateral viewthe distance between the ectal and the discoidfacets is greater than in M. americanum, whereasin MLP 68-III-14-1, MACN 4941, MACN4992, MACN 13218, MLP 99-XI-1-1, althoughseparated, the distance is shorter.
CalcaneumThe calcaneum of P. bergi (Fig. 2E, F) is shapedas in other megatherines, although somewhatmore gracile than in M. americanum and E. lau-rillardi and both more compressed and elongatedposteriorly behind the articular end. The calca-neum has three articular facets on its anteriorend; two articulate with the astragalus and onewith the cuboid. The largest facet articulates withthe ectal facet of the astragalus. This facet is alsothe most dorsal and is oval in shape with themain axis oriented anterolateral to posteromedi-ally, parallel to the sulcus calcanei (placed ven-trally to the facet). The facet is convex along themain axis, medially concave along the secondaryaxis, and flat laterally. A second facet articulateswith the sustentacular facet of the astragalus. Thisfacet is relatively quadrangular to pentagonal inoutline, planar, and is separated from the formerby the sulcus calcanei. The third or cuboid facetis smaller, positioned lateroventrally with respectto the sustentacular facet, and semicircular tooval in shape. In P. bergi, this facet and the sus-tentacular facet are not in contact. The sulcus cal-canei, separating the ectal and sustentacularfacets, runs transversally and of uniform widththroughout its full extent. Finally, it should benoted that the calcaneum of MLP 68-III-14-1 ismore robust and shorter than that of the type ofP. bergi.
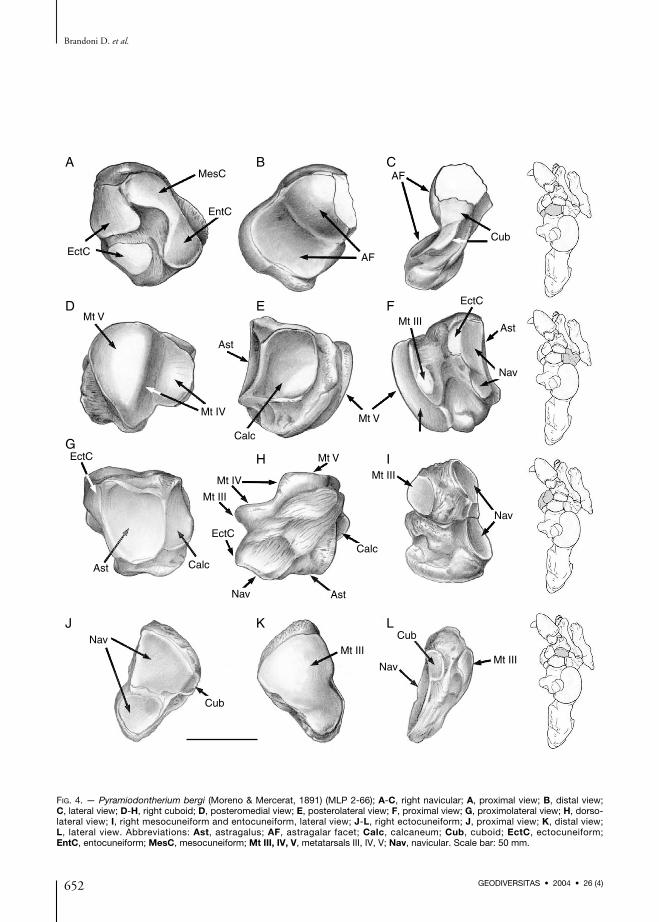
NavicularThe navicular (Fig. 4A-C) is oval to subrectan-gular in shape (distal view), antero-posteriorlycompressed with the main axis oriented dorsolateral
Brandoni D. et al.
648 GEODIVERSITAS • 2004 • 26 (4)
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
649GEODIVERSITAS • 2004 • 26 (4)
DF
DF
OT OF
OP
Nav
CubOT
OF
OP
Nav
OF
DF
ASF
FF
AEFCub
90°
ST
AEF
CEF
CSF SC
Cub
CSF CEF
A B
C
E
D
FIG. 2. — Pyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66); A-D, left astragalus; A, dorsomedial view; B, antero-medial view; C, fibular view; D, ventral view; E, F, left calcaneum; E, specular lateral view; F, specular distal view. Abbreviations:AEF, astragalar ectal facet; ASF, astragalar sustentacular facet; CEF, calcaneal ectal facet; CSF, calcaneal sustentacular facet;Cub, cuboid; DF, discoid facet; FF, fibular facet; Nav, navicular; OF, odontoid facet; OP, odontoid process; OT, odontoid tuberosity;SC, sulcus calcanei; ST, sulcus tali. Scale bar: 50 mm.
to ventromedially. In MLP 2-66 a portion of thedorsolateral half is missing.The astragalar facet is located on the proximalsurface, and may be differentiated into dorsolat-eral and ventromedial halves. The dorsolateralhalf is more or less circular and is formed by acondylar projection that fits into the circulardepression of the astragalus (facie articular navic-ularis). This projection is not as well developed asin Megatherium americanum. The ventromedialhalf is relatively oval, concave along its main axesand articulates with the condylar portion of the
astragalus. Both halves are similar in size, as inM. americanum. In M. altiplanicum from thePliocene of Bolivia, the medial part of the facet islarger than the lateral part (Saint-André & DeIuliis 2001).On the distal surface the navicular is convexalong its main axes. On this aspect there are threearticular facets. Ventrally there are two ecto-cuneiform facets, the smallest is subtriangularand ventromedial with respect to the other whichis larger, quadrangular to trapezoidal, and posi-tioned dorsolaterally.Dorsomedially to the aforementioned facets thereis a large facet that articulates with the internalmeso- and entocuneiforms. This large facet isoval, elongate along the main axis of the navi-cular, but broken on its central part. This facetjoins dorsally with the square ectocuneiformfacet, being a rebound on the border (as inMACN 4983). In M. americanum, the ecto-cuneiform facets are fused and the internalcuneiform facet is smaller than the facet for theectocuneiform.A cuboid facet lies on the ventrolateral portion ofthe navicular. It is elongate along the dorsolater-al-ventromedial axis. This facet has two differentportions, one dorsolateral and other ventral. Thedorsolateral portion contacts with the dorsal por-tion (condylar projection) of the navicular facetproximally, and the dorsal (square) ecto-cuneiform facet distally, in both cases in anapproximately right angle. The lower portion ofthe cuboid facet contacts with the ventral portionof the astragalar facet posteriorly, and anteriorlyis separated from the ventral ectocuneiform facetby a non-articulate furrow. In M. americanumthe cuboid facet is located in a single plane, asmay be seen on the navicular facet of the cuboid.
CuboidThe cuboid (Fig. 4D-H) is irregular in shape,more or less cubic, massive, and supports twocomplex articular surfaces separated by non-artic-ulating bone. On the posteromedial surface thereare facets for the calcaneum, astragalus, navicular,and ectocuneiform. On the anteroventral surfaceof the bone, there are facets for metatarsals III,
Brandoni D. et al.
650 GEODIVERSITAS • 2004 • 26 (4)
FIG. 3. — Astragalus of Pyramiodontherium bergi (Moreno &Mercerat, 1891) (MLP 2-66); A, anterior view; B, dorsolateralview. a-a’ line, projection of the plane of the discoid facet;b-b’ line, projection of the plane of the base of the odontoidfacet. Abbreviations: DF, discoid facet; FF, fibular facet;Nav, navicular; OF, odontoid facet; OP, odontoid process.Scale bar: 50 mm.

IV, and V. The dorsal surface of the cuboid isrugose, relatively flat, and devoid of articularfacets. The ventral portion is more irregular, withsome plantar projections, and a posterolaterallyprojecting crest that reaches the dorsal surface.A deep dorsoventral furrow occupies the medialside of the cuboid and separates the articularareas.The calcaneal facet is posterolateral, square, withthe dorsomedial half concave, and the ventrolat-eral convex. The astragalar facet is posteromedial,and contacts the calcaneal facet forming a straightangle between them. It is approximately ovalwith the main axis dorsolateral to ventromedialand concave in both main axes.The navicular facet is placed medially in front ofthe astragalar facet. The latter is subdivided intotwo different planes corresponding to thosedeveloped for the lateral facet of the navicular.The navicular facet is dorsoventrally elongate andnarrow; the posterior margin contacts the astra-galar facet and the anterior margin contacts theectocuneiform facet.The small and semicircular ectocuneiform facetcontacts the navicular facet posteriorly and themedial furrow anteriorly. This facet is absent inM. americanum, occasionally present in E. lauril-lardi (Cartelle 1992) and always present inM. tarijense (Pujos & Salas 2004).The facet for metatarsal IV has two portions,each plane almost perpendicular to the other.The posterior portion is elongate along the dor-solateral-ventromedial axis, and the more anteriorand dorsal plane is oval in shape, and placed onan anterior laminar projection of the cuboid. InM. americanum the angle between these two por-tions is wider.The metatarsal V facet is next to the most elon-gate portion of the Mt IV facet, in a straightangle. It is almost triangular with a ventral apex,convex in both axis.On the anteromedial end of the cuboid, support-ed medially by the laminar projection, there is asmall oval facet for metatarsal III that is extendeddorsoventrally. This facet is present in M. ameri-canum, but M. tarijense has no articulationbetween the cuboid and metatarsal III.
MesocuneiformThe mesocuneiform (Fig. 4I) is trapezoidal inmedial view. It has three articular facets, twolateral facets, one proximal for the navicular,another distal for metatarsal III, and one ventralfacet (on a slightly inclined plane) for the ento-cuneiform. The navicular facet is oval but has adownward projection that is continuous with theascending lateral proximal entocuneiform facet.Both these facets form a continuous, dorsoventrally-oriented functional surface. The Mt III facet iscircular and articulates with the medial proximalfacet of metatarsal III. The two lateral meso-cuneiform facets, equal in size, are separated by afragment of non-articulating bone, also similar insize. The entocuneiform facet contacts the navic-ular facet and is separated from the Mt III facet.It is the largest of the articular facets and is ovalto triangular in shape.
EntocuneiformThe entocuneiform (Fig. 4I) is rectangular inmedial view, larger than the mesocuneiform, andhas two articular facets: one lateral and the otherdorsal, which articulate with the navicular andthe mesocuneiform, respectively, and are incontact proximally. The navicular facet is ovaland is continuous upward with the descendingportion of the mesocuneiform lateral proximalfacet. The mesocuneiform facet is placed locatedalmost horizontally and perpendicular to the lat-eral facet. It is larger and matches in shape withthe mesocuneiform ventral facet. On the internallateral surface there is a fragment of non-articu-lating bone that bears a semilunar depression offlat bottom that divides the bone in two parts,one principal and other reminiscent of a plantartuberosity.In P. bergi there is no fusion of the meso- andentocuneiform, termed the mesoentocuneiformcomplex by De Iuliis (1996). In megatherines(e.g., Megatherium sp., Eremotherium sp.) thathave this complex, it is triangular (or pentagonal)with the apex anteriorly directed. The proximalsurface has an elongate navicular facet, and thedistal surface displays a small oval Mt III facetdorsolaterally.
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
651GEODIVERSITAS • 2004 • 26 (4)
Brandoni D. et al.
652 GEODIVERSITAS • 2004 • 26 (4)
EctC
MesC
EntC
AF
AF
Cub
Mt V
Mt IV
Ast
Calc
Mt V
Mt III
EctC
Ast
Nav
EctC
Ast Calc
EctC
Mt III
Mt IV
Mt V
Calc
AstNav
Mt III
Nav
Nav
Nav
Cub
Mt IIICub
Mt III
A B C
D E F
GH I
J K L
FIG. 4. — Pyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66); A-C, right navicular; A, proximal view; B, distal view;C, lateral view; D-H, right cuboid; D, posteromedial view; E, posterolateral view; F, proximal view; G, proximolateral view; H, dorso-lateral view; I, right mesocuneiform and entocuneiform, lateral view; J-L, right ectocuneiform; J, proximal view; K, distal view;L, lateral view. Abbreviations: Ast, astragalus; AF, astragalar facet; Calc, calcaneum; Cub, cuboid; EctC, ectocuneiform;EntC, entocuneiform; MesC, mesocuneiform; Mt III, IV, V, metatarsals III, IV, V; Nav, navicular. Scale bar: 50 mm.

EctocuneiformThe ectocuneiform (Fig. 4J-L) is triangular withits base dorsolateral, apex ventromedial, andelongate in its main axis. It is anteroposteriorlycompressed, convex anteriorly, and diminishes inthickness dorsolaterally-ventromedially. It ismore gracile, elongate and tapered than in othergenera (i.e. Megathericulus and Megatherium).The proximal and distal surfaces of the ecto-cuneiform are almost entirely articular, with twomain areas (proximal and distal) that follow thetriangular shape of the ectocuneiform, and areseparated by non-articular bone. Proximally, twonavicular articular facets are developed: one lat-erodorsal facet that is subtrapezoidal to square,and another ventromedial facet that is subtrian-gular. Both facets fit in shape and size with theircounterparts developed on the distal surface ofthe navicular. Also like those of the navicular,they are separated by a transverse fringe of non-articular bone. At the same level, the laterodorsalfacet is continuous ventrally with a subrectangu-lar facet and their articular planes meet in a120° angle. The subrectangular facet articulateswith the small semicircular cuboid facet, whichis absent in M. americanum and M. urbinai, andpresent in M. tarijense. Most of the distal surfaceof the ectocuneiform is occupied by a facet forMt III that is approximately triangular andconvex, although it has a small furrow near theventromedial apex. In other genera (i.e.Megatherium), the distal facet is completely con-vex. In M. urbinai, the ectocuneiform articulateswith metatarsals III and IV (Pujos & Salas2004). There is no articulation between theectocuneiform and the internal cuneiforms inP. bergi. In M. urbinai (MUSM 15) and M. tari-jense (FMNH P14216), the ectocuneiform artic-ulates with the mesoentocuneiform complex anda similar articulation is also present in somespecimens of M. americanum (e.g., MNHNPAN 295).
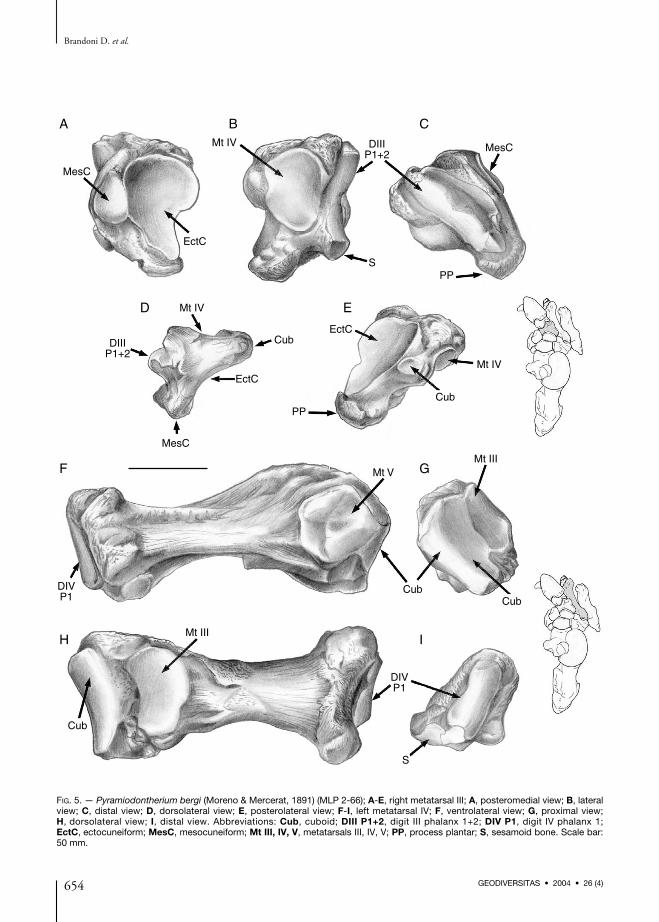
Metatarsal IIIIn P. bergi, the metatarsal III (Fig. 5A-E) isanteroposteriorly very compressed compared tometatarsals IV and V. The posterior portion is
strongly compressed transversally, while in othermegatheres (i.e. E. laurillardi), this portion isshorter (Paula Couto 1978).The ectocuneiform facet is posteromedial,almost triangular with ventral apex, and concavealong both axes. A small mesocuneiform facet,oval in shape, occupies the proximomedial sideof the metatarsal III. This facet contacts posteri-orly with the ectocuneiform facet in straightangle.The facet for metatarsal IV is lateral, oval inshape, and extended dorsoventrally. The surfaceis undulate, markedly concave anteriorly, andconvex posteriorly. The articular surface is nearlyparallel to the ectocuneiform facet. A small acces-sory Mt IV facet appears on the dorsolateral sur-face of the metatarsal III.Between the facets for ectocuneiform and Mt IV,there is a posterolateral projection that supportsan oval cuboid facet.On the distal surface a crest articulates withphalanx 1+2 of digit III. It is slightly concavedorsoventrally, it narrows ventrally becomingalso more acute, and is inclined along a dorsolat-eral to ventromedial axis. We conclude that thiscrest is almost entirely articular because in themiddle of its dorsoventral extent (on the deepestpart of the concavity) there is a small non-articulating area that fits with the correspondingphalanx 1+2 of digit III. An elongate internal sur-face continues beyond the crest to complete thearticulation. In M. americanum the crest is inter-rupted and continues medially as an independentsmall oval facet. The ventromedial end of the keelhas an articular facet probably for a sesamoid. Onthe most ventrolateral portion of metatarsal IIIthere is a plantar prominence.
Metatarsal IVMetatarsal IV (Fig. 5F-I) is long and not ascompressed as metatarsal III. The diaphysis istriangular in section with lateral apex, whereasin M. americanum the section is subtriangu-lar to oval. The proximal portion is laterallycompressed and dorsoventrally expanded.Three articular facets appear on the proximalepiphysis, an anteromedial facet for Mt III, a
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
653GEODIVERSITAS • 2004 • 26 (4)
Brandoni D. et al.
654 GEODIVERSITAS • 2004 • 26 (4)
MesC
EctC
Mt IV DIIIP1+2
DIIIP1+2
MesC
SPP
Mt IV
Cub
EctC
MesC
EctC
PP
Mt IV
Cub
DIVP1
DIVP1
Mt V
Cub
Cub
Cub
Mt III
Mt III
S
A
F
H I
G
B C
D E
FIG. 5. — Pyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66); A-E, right metatarsal III; A, posteromedial view; B, lateralview; C, distal view; D, dorsolateral view; E, posterolateral view; F-I, left metatarsal IV; F, ventrolateral view; G, proximal view;H, dorsolateral view; I, distal view. Abbreviations: Cub, cuboid; DIII P1+2, digit III phalanx 1+2; DIV P1, digit IV phalanx 1;EctC, ectocuneiform; MesC, mesocuneiform; Mt III, IV, V, metatarsals III, IV, V; PP, process plantar; S, sesamoid bone. Scale bar:50 mm.

Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
655GEODIVERSITAS • 2004 • 26 (4)
posteromedial facet for the cuboid, and a lateralfacet for Mt V. The facet for Mt III is oval,dorsoventrally extended, anteroposteriorlyconvex at the anterior part, and concave in bothdirections posteriorly. This facet is separated pos-teriorly from the cuboid facet. The cuboid facet isrectangular, dorsoventrally extended, and itsanterior two-thirds is dorsoventrally concave andoccupies the medial side. The posterior third isproximal in position.The metatarsal V facet is located on the externalside of the proximal epiphysis. This facet is pen-tagonal, with a central depression and a smallextension that is continuous with the posteriorportion of the cuboid facet.The distal epiphysis is triangular with laterodor-sal apex and tripod-like base. The facet for theproximal phalanx of digit IV is crested, as that formetatarsal III, but less acute in section, and nar-rowing ventrally. Ventromesially there is a small,and convex subcircular facet for a sesamoid. Thisfacet was first mentioned by Cabrera (1929)when describing the pes of Megatherium. Thissame extension is present in E. laurillardi, but nosesamoid bone can be seen (De Iuliis 1996).Occasionally there occurs ankylosis betweenMt IV and Mt III in E. laurillardi (Paula Couto1978; Cartelle 1992).
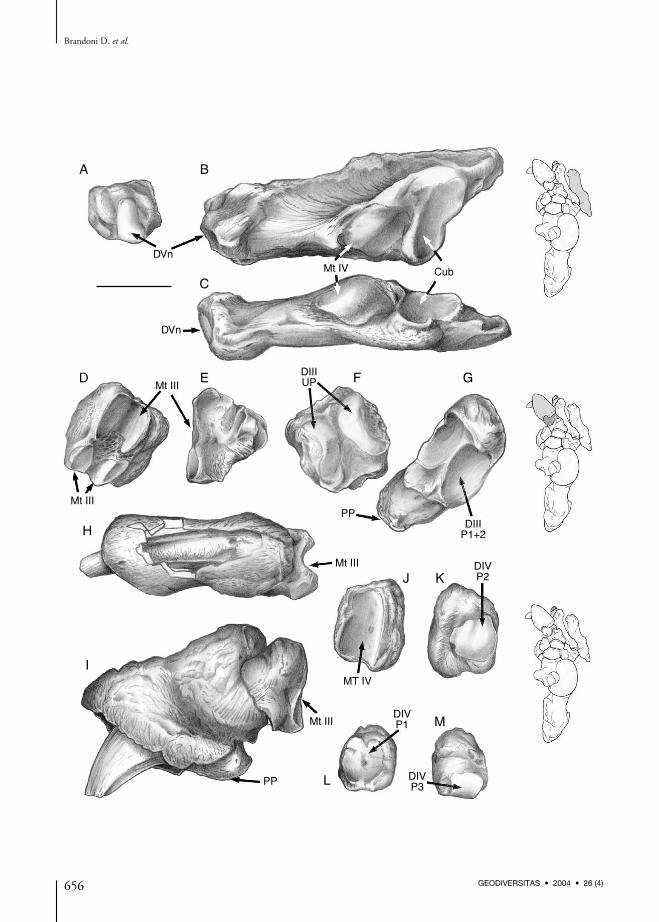
Metatarsal VMetatarsal V (Fig. 6A-C) is slightly longer thanmetatarsal IV but depressed dorsoplantarly withthe lateral margin expanded and dorsoventrallycompressed (especial ly the proximal twothirds). The distal portion is almost isodiamet-ric. The medial side has two contiguous articu-lar facets that are inclined dorsomedially. Theanteriormost facet for Mt IV is pentagonal, rela-tively flat, and continuous posteriorly with thecuboid facet. The cuboid facet is concave andtriangular with ventral base and anterior apex.The posterior end of metatarsal V is prominentand point-shaped in P. bergi, while in M. ameri-canum it is blunted. The distal surface has asmall oval, convex facet elongated along thedorsolateral-ventromedial axis for a nodular ves-tigial phalanx.
Digit IIIDigit III is the only posterior clawed digit inMegatheriinae. It is composed of a singleproximal bone (fused phalanxes 1+2 accord-ing to Cartel le 1992) (Fig. 6D-F), and astrong ungual phalanx that is claw-shaped(Fig. 6G-I).The proximal surface of the phalanx 1+2 bears awide facet for metatarsal III. This facet is canal-shaped and elongated along the dorsolateral-ventromedial axis. Non-articular bonesubdivides it into two parts. Distally it bears atrocleated surface that is transversely elongateand bears an incomplete non-articular bonyarea. The phalanx is obliquely oriented andconsequently the ungual phalanx is deviatedventromedially.The ungual phalanx articulates with the proxi-mal phalanx (1+2) by a deeply depressed surfacedivided into two oval parts. This surface is ori-ented obliquely to the sagittal plane, very welldeveloped, relatively short and robust, and high-er than wide (especially the distal portion).Proximally it bears a crown-like expansion thatwould have covered the base of a large cornealclaw. The central bone is also short and high, tri-angular in section, and projects beyond the bonethat sheathed it on the proximal two-thirds ofthe phalanx.
Digit IVUnlike other megatherines that have only twophalanxes, digit IV of P. bergi has three phalanx-es. Phalanx 1 (Fig. 6J, K) is anteroposteriorlycompressed and elongate along the dorsoventralaxis. The proximal surface for the Mt IV facet iswell developed and almost entirely articular. Thisfacet is elongate and strongly concave transversely.The distal surface bears a dorsoventrally convexbilobate facet. The second phalanx articulates onthis facet. Phalanx 2 (Fig. 6L, M) is smaller andmore anteroposteriorly compressed than phalanx 1.The proximal facet is bilobate, somewhat cir-cular, and concave on both directions. A smallalmost circular facet on the distal surface musthave articulated with a third, probably vestigialphalanx.
Brandoni D. et al.
656 GEODIVERSITAS • 2004 • 26 (4)
DVn
DVn
Mt IV Cub
Mt III
Mt III
Mt III
Mt III
DIIIUP
DIIIP1+2
DIVP2
PP
MT IV
PP
DIVP1
DIVP3
A
D E F G
B
C
H
I
J K
M
L

DISCUSSION
The pes of P. bergi displays typical characters ofthe subfamily, i.e. astragalus and calcaneumstrongly developed, three digits (III to V) ofwhich the third is the only one clawed. Two ofthe characters (number of phalanxes and tarsalbones morphology) are intermediate between thecondition observed in primitive megatherioids(i.e. Hapalops) and that found in Quaternaryspecies of Megatheriinae (e.g., M. americanumand E. laurillardi).The number of tarsal elements of the megather-ine pes has been discussed and debated since thefirst half of the 19th century. Cuvier (1834-1836), Owen (1859), and Burmeister (1865),thought that Megatherium had only six elements,but did not explain the reason for the reduction,whether by loss or fusion of elements (see discus-sion in Cabrera 1929). Cabrera (1929) thoughtthere were seven tarsal elements in Megatherium,as the mesocuneiform and ectocuneiform appearseparated in some specimens (MLP 2-29, MLP2-30, and MLP 27-VII-1-1 assigned toMegatherium). In addition, he stated that thefusion of the internal distal tarsal elements into asingle bone occurred with advancing age. On theother hand, Cabrera reported that one falciformextratarsal bone, or sesamoid, articulated with theentocuneiform in these same specimens. Cabrerafurther speculated that the falciform element sub-stituted for the absence of a distal phalanx ondigit I and provided a supporting point duringbipedalism. Cartelle (1992) stated that the falci-form is part of the meso-entocuneiform complexin E. laurillardi. In turn, De Iuliis (1996) pro-posed that “there is some evidence that the falci-form as identified by Cabrera (1929) mayoccasionally fuse with the mesocuneiform and
entocuneiform into a single element”. In generalthe ventromedial surface of the complex, repre-sented by the portion corresponding to the ento-cuneiform, ends in a blunt projection. However,in M. tarijense FMNH P14216 this complex isventromedially extended by an elongate taperedprocess that may represent the fusion of the falci-form (De Iuliis 1996).Primitive Megatherioidea (e.g., Hapalops) hadseven separate tarsal elements (calcaneum, astra-galus, navicular, entocuneiform, mesocuneiform,ectocuneiform, and cuboid) (Scott 1903-1905).This situation, similar to that of P. bergi, is prob-ably a primitive condition seen in other mam-mals, and the fusion of different bones into acomplex is the derived and most common condi-tion among Quaternary megatheres.In the pes of P. bergi there is no falciform arti-culating with the entocuneiform, nor is there anyfacet on the bone that suggests the presence ofthis bone. The absence of a falciform elementthat would act as supporting point for the pes (asbelieved by Cabrera 1929) suggests that P. bergimay not have reached a degree of bipedalismattributed to other megatherines (Casamiquela1974; Aramayo & Manera de Bianco 1987).However, some specimens of M. americanumlack a falciform, and it is difficult to imagineconspecifics with less proficient bipedalism.As in all other megatherines, P. bergi has onlythree digits (III to V) on the pes. It shows no ves-tige of more internal digits nor articular facets onthe corresponding tarsal elements. Some authors(see below) consider that in Megatherium vestigesof digits I and II may have fused to different ele-ments. Cabrera (1929) reported vestiges of Mt II,Scott (1913) observed that the pes had only threefunctional digits and that the first and second arereduced to vestiges, and Cartelle (1992) believed
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
657GEODIVERSITAS • 2004 • 26 (4)
FIG. 6. — Pyramiodontherium bergi (Moreno & Mercerat, 1891) (MLP 2-66); A-C, right metatarsal V; A, distal view; B, dorsomedialview; C, dorsal view; D-F, left digit III 1+2 phalanx; D, proximal view; E, medial view; F, distal view; G, right digit III ungueal phalanx,proximal view; H, I, right digit III ungueal and 1+2 phalanxes; H, dorsolateral view; I, dorsomedial view; J, K, right digit IV phalanx 1;J, proximal view; K, distal view; L, M, right digit IV phalanx 2; L, proximal view; M, distal view. Abbreviations: Cub, cuboid; DIIIP1+2, digit III, phalanx 1+2; DIII UP, digit III, ungueal phalanx; DIV P1, digit IV, phalanx 1; DIV P2, digit IV, phalanx 2; DIV P3, digit IV,phalanx 3; DVn, digit V, nodular; Mt III, IV, metatarsal III, IV; PP, process plantar. Scale bar: 50 mm.
▲

that remnants of Mt II may be fused to the meso-cuneiform to form part of a mesoentocuneiformcomplex.The third digit has only two phalanxes. The proxi-mal phalanx results from the fusion of two pha-lanxes and it is the most primitive conditionknown for megatherines. Unlike all Quaternarymegatherines, digit IV of P. bergi has three pha-lanxes. This situation, unique among species of thesubfamily, is the primitive mammalian condition(and it could be present in other Tertiary mega-therines). On the navicular of P. bergi and onespecimen from Paraná assigned to “Megatheriumantiquum” Ameghino, 1885 (MACN 4983), thereare two articular facets for the ectocuneiform.However, these two facets are fused in a single facetin the naviculars of Quaternary species.Most of Quaternary megatherines species are wellknown, and particularly those of Megatherium,both in their morphological and phylogeneticalaspects (see Cartelle 1992; De Iuliis 1994, 1996;Saint-André & De Iuliis 2001; Pujos & Salas2004). Saint-André & De Iuliis (2001) indicatethat M. americanum and M. altiplanicum are sis-ter species and the most specialized species ofMegatherium. Pujos & Salas (2004) postulatethat the small sized Megatherium species,M. urbinai, is more similar to (M. medinae,M. elenense, and M. tarijense) than (M. ameri-canum + M. altiplanicum). Among the Tertiaryspecies, the relationships are more difficult toestablish, because the scarceness and fragmentarycondition of the remains (especially post cranialmaterial). The genus Pyramiodontherium is repre-sented only by two species P. bergi and P. brevi-rostrum Carlini, Brandoni, Scillato-Yané &Pujos, 2002 (Carlini et al. 2002), both from the“Araucanense” (late Miocene-Pliocene) ofCatamarca Province; this genus is also present inthe “Conglomerado osífero” ItuzaingóFormation (late Miocene), Entre Ríos Province,Argentina (Carlini et al. 2001; Brandoni et al.2001). The remains assigned to Pyramio-dontherium found in the “Conglomerado osífero”could be more closely related to those assigned to“M. antiquum” than with the medium sizedspecies of Pliomegatherium Kraglievich, 1930.
In summary, the pes of P. bergi is more special-ized with respect to that of Megathericulus patago-nicus, and its morphology fits better with thatassumed to be present in Upper Tertiary mega-therines. The plesiomorphic states of charactersobserved here may help to resolve the polytomyproposed by De Iuliis (1996) and offer a moreprecise knowledge of the way characters stateswere transformed during the evolution of thegroup.
AcknowledgementsWe thank Drs. R. Pascual, M. Reguero andS. Bargo (MLP), Drs. J. Bonaparte andA. Kramarz (MACN), who facilited the studyof the specimens housed in their institutions,Mr. J. González for the draws, and Dr. C. Des-champs for the translation. Thanks are due toDr. Gregory McDonald (Geologic ResourcesDivision, National Park Service, Denver,Colorado, USA) and Dr. Claude Guérin(Université Claude Bernard, Lyon, France) foruseful review to improve the manuscript. Thiswork was partially supported by PEI-CONICETNo. 107-98 and PICT-Redes 02-00074 toA.A.C.
REFERENCES
ARAMAYO S. A. & MANERA DE BIANCO T. 1987. —Hallazgo de una icnofauna continental (PleistocenoTardío) en la localidad de Pehuén-Có (partido deCoronel Rosales) provincia de Buenos Aires,Argentina. Parte Y: Edentata, Litopterna, Pro-boscidea. Parte II: Carnívora, Artiodactyla y Aves.IV Congreso Latinoamericano de Paleontología SantaCruz de la Sierra, Bolivia, Actas 1: 532-547.
BONDESIO P., RABASSA J., PASCUAL R., VUCETICH M.G. & SCILLATO-YANÉ G. J. 1980. — La FormaciónCollón-Curá de Pilcaniyeu Viejo y sus alrededores(Río Negro, República Argentina). Su antigüedad ylas condiciones ambientales según su distribución,su litogénesis y sus vertebrados. Actas II CongresoArgentino de Paleontología y Bioestatigrafía (BuenosAires, 1978) 3: 85-99.
BRANDONI D., CARLINI A. A. & SCILLATO-YANÉ G. J.2001. — Una nueva especie de Pyramiodontherium(Xenarthra, Megatheriinae) en el “Mesopo-tamiense” (Mioceno tardío-Plioceno temprano) de
Brandoni D. et al.
658 GEODIVERSITAS • 2004 • 26 (4)

Entre Ríos, Argentina. Jornadas Argentinas dePaleontología Vertebrados, Esquel 2001.Ameghiniana 38 (4) supl.: 4R-5R.
BURMEISTER H. G. 1865. — Fauna Argentina:primera parte, Mamíferos fósiles. Anales del MuseoPúblico de Buenos Aires 1: 87-300.
CABRERA A. 1928. — Sobre algunos megaterios plio-cenos. Revista del Museo de La Plata 31: 339-352.
CABRERA A. 1929. — La estructura de la mano y delpie en el Megaterio. Anales de la Sociedad CientíficaArgentina 107: 425-443.
CABRERA A. 1939. — Un nuevo representanteSantacrusense de la Familia Megatheriidae. Notasdel Museo de La Plata, Paleontología 4: 475-483.
CARLINI A. A., SCILLATO-YANÉ G. J., NORIEGA J. I. &ACEÑOLAZA F. 2001. — Perezosos terrestres(Xenarthra, Tardigrada) del “Mesopotamiense”(Fm. Ituzaingó, Mioceno tardío-Plioceno) de laProvincia de Entre Ríos, Argentina. StudiaGeologica Salmanticensia Salamanca 36: 13-27.
CARLINI A. A., BRANDONI D., SCILLATO-YANÉ G. J. &PUJOS F. 2002. — Una nueva especie de megaterino(Xenarthra, Megatheriidae) del Mioceno Tardío-Plioceno de Catamarca, Argentina. Ameghiniana 39(3): 367-377.
CARTELLE C. 1992. — Edentata e megamamíferos her-bívoros extintos da Toca dos Ossos (Ourolândia, Bahia,Brasil). Tese de Doutorado, Universidade Federal deMinas Gerais, Belo Horizonte, Brazil, 301 p.
CASAMIQUELA R. M. 1974. — El bipedismo de losMegatherioideos. Estudios de pisadas fósiles en laFormación Río Negro típica. Ameghiniana 11:249-282.
CUVIER G. 1834-1836. — Recherches sur les ossementsfossiles, où l’on rétablit les caractères de plusieurs ani-maux dont les révolutions du globe ont détruit lesespèces. Vol. 2. 4th ed. E. d’Ocagne, Paris, 500 p.
DE IULIIS G. 1994. — Relationships of the Mega-theriinae, Nothrotheriinae, and Planopsinae: someskeletal, characteristics and their importance forphylogeny. Journal of Vertebrate Paleontology 14 (4):577-591.
DE IULIIS G. 1996. — A Systematic Review of theMegatheriinae (Mammalia: Xenarthra: Mega-theriidae). Unpublished Ph.D. Dissertation,University of Toronto, Canada, 781 p.
HIRSCHFELD S. E. 1985. — Ground sloths from theFriasian La Venta Fauna, with additions to the Pre-
Friasian Coyaima Fauna of Colombia, SouthAmerica. University of California Publications inGeological Sciences 128: 1-91.
KRAGLIEVICH L. 1926. — Notas sobre gravígrados deSud América. Anales del Museo Nacional de HistoriaNatural de Buenos Aires 34: 21-36.
MORENO F. P. & MERCERAT A. 1891. — Exploraciónarqueológica de la Provincia de Catamarca:Paleontología. Revista del Museo de La Plata 1:222-236.
OWEN R. 1859. — On the Megatherium (Mega-therium Americanum Cuvier and Blumenbach) V,bones of the posterior extremities. PhilosophicalTransactions of the Royal Society of London 149 (2):809-829.
OWEN R. 1860. — Memoir on the Megatherium(Megatherium Americanum Cuvier), or GiantGround-Sloth of America. Taylor & Francis,London, 84 p.
PAULA COUTO C. DE 1978. — Mamíferos fósseis doPleistoceno do Espírito Santo. Anales de laAcademia Brasileira de Ciencias 50 (3): 365-379.
PUJOS F. & SALAS R. 2004. — A new species ofMegatherium (Mammalia: Xenarthra: Mega-theriidae) from the Pleistocene of Sacaco and TresVentanas, Peru. Palaeontology 47 (3): 579-604.
ROTH S. 1911. — Un nuevo género de la familiaMegatheridae. Revista del Museo de La Plata 18:7-21.
ROVERETO C. 1914. — Los estratos araucanos y susfósiles. Anales del Museo de Historia natural 25:1-247.
SAINT-ANDRÉ P.-A. & DE IULIIS G. 2001. — Thesmallest and most ancient representative of thegenus Megatherium Cuvier, 1796 (Xenarthra,Tardigrada, Megatheriidae), from the Pliocene ofthe Bolivian Altiplano. Geodiversitas 23 (4):625-645.
SCILLATO-YANÉ J. G. & CARLINI A. A. 1998. —Nuevos Xenarthra del Friasense (Mioceno medio)de Argentina. Studia Geologica Salmanticensia 34:43-67.
SCOTT W. B. 1903-1905. — Mammalia of the SantaCruz Beds. Part 1: Edentata. Reports of the PrincetonUniversity to Patagonia 5: 1-364.
SCOTT W. B. 1913. — A History of Land Mammals inthe Western Hemisphere. Macmillan, New York,693 p.
Submitted on 22 October 2003;accepted on 16 April 2004.
Anatomy of the pes of Pyramiodontherium bergi (Mammalia, Xenarthra)
659GEODIVERSITAS • 2004 • 26 (4)





























