Embed Size (px)
Citation preview

Sedimentary Geology 234 (2011) 42–55
Contents lists available at ScienceDirect
Sedimentary Geology
j ourna l homepage: www.e lsev ie r.com/ locate /sedgeo
Ichnological analysis of the Bidart and Sopelana Cretaceous/Paleogene(K/Pg) boundary sections (Basque Basin, W Pyrenees): Refiningeco-sedimentary environment
F.J. Rodríguez-Tovar a,⁎, A. Uchman b, X. Orue-Etxebarria c, E. Apellaniz c, Juan I. Baceta c
a Departamento de Estratigrafía y Paleontología, Facultad de Ciencias, Universidad de Granada, 18002 Granada, Spainb Jagiellonian University, Institute of Geological Sciences, Oleandry Str. 2a, PL-30-063 Kraków, Polandc Departamento de Estratigrafía y Paleontología, Facultad de Ciencia y Tecnología, Universidad del País Vasco, E-48080 Bilbao, Spain
⁎ Corresponding author. Tel.: +34 958 242724; fax:E-mail addresses: [email protected] (F.J. Rodríguez-Tov
(A. Uchman), [email protected] (X. [email protected] (E. Apellaniz), juanignacio.ba
0037-0738/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.sedgeo.2010.11.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 12 August 2010Received in revised form 23 October 2010Accepted 22 November 2010Available online 1 December 2010
Editor: M.R. Bennett
Keywords:Trace fossilsCretaceous/Paleogene boundaryZoophycos ichnofaciesEco-sedimentary conditionsBidartSopelana
Ichnological analysis of two Cretaceous–Paleogene boundary sections at Bidart (SE France) and Sopelana (NSpain) has been conducted in order to refining eco-sedimentary environment, and to amake comparison withprevious interpretations based on microfossils. In both sections, trace fossil assemblage is low diverse,consisting of Chondrites, Planolites, Thalassinoides, Trichichnus, Zoophycos, and ?Phycosiphon, ascribed to theZoophycos ichnofacies, however, with distinct differences. In the Bidart section, early Danian dark-filled tracefossil assemblage is more abundant in large Thalassinoides, Zoophycos and larger Chondrites, and less abundantin Trichichnus and small Chondrites in comparison to the Sopelana section. Sopelana is thus interpreted as amore offshore, deeper section than Bidart although both were located in the upper bathyal zone of the basin.Oxygenation below the sediment–water interface was enough to favour a generalized total bioturbation ofsediments. The presence of chemichnia and trace fossils which tracemakers sequestered sediments from thesea floor in deep burrows (Zoophycos) suggests at least periodical food deficiency. The distribution andcontent of trace fossils across the K/Pg boundary do not change significantly, revealing minor incidence of theK/Pg boundary event on the macrobenthic environment. The K/Pg boundary transition, including theboundary layer, is nearly totally bioturbated; hence a redistribution of microbiotic and abiotic components bytracemakers is possible.
+34 958 248528.ar), [email protected]),[email protected] (J.I. Baceta).
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The Cretaceous–Paleogene (K/Pg) boundary event is one of themost spectacular phenomenon in the Phanerozoic history of theEarth causing a global major extinction. In pelagic environments it isrecorded in the so-called boundary layer, which is a thin, usually 5–10 cm thick, dark and calcium carbonate depleted claystone ormarlstone. Commonly, the base of the boundary layer is defined by amillimeter-thick rusty layer which contains the iridium and othergeochemical anomalies related with the extraterrestrial bolideimpact at the northern Yucatan peninsula (Schulte et al., 2010;Smit, 1999) which define the formal position of the K/Pg boundary(Molina et al., 2006). Publications showing the presence of tracefossils at the K/Pg boundary interval are scarce (see Rodríguez-Tovarand Uchman, 2008 for a review), especially those including adetailed ichnological characterization (e.g., Ekdale and Bromley,
1984; Ekdale and Stinnesbeck, 1998). However, in the last yearsichnological analyses conducted in several K/Pg sections, reveal thatthe boundary layer is bioturbated, containing diverse trace fossils,which help to refine several aspects in the interpretation of thepalaeoenvironmental conditions that prevailed during sedimenta-tion of the K/Pg boundary event interval (Rodríguez-Tovar, 2005;Rodríguez-Tovar and Uchman, 2004a,b, 2006, 2008; Rodríguez-Tovar et al., 2002, 2004, 2006).
In the Basque Basin (western Pyrenees) several outcrops of deepmarine (hemi)pelagic facies has been studied, revealing the welldeveloped, expanded and relatively complete K–Pg boundaryinterval (see Apellaniz et al., 1997; Pujalte et al., 1998). Thesefavourable features determined the numerous studies conducted inthe uppermost Maastrichtian–lowermost Danian sediments of thisarea, involving sequence stratigraphic, mineralogical, isotopic,geochemical, magnetostratigraphical and biostratigraphical analy-ses (see Alegret, 2007 for a recent review). In this exhaustiveresearch, however, detailed ichnological analysis has been onlyrecently initiated (Rodríguez-Tovar et al., 2010). In this paper wepresent the ichnological analysis of two of the best-preserved K/Pgboundary sections, Bidart, in SW France, and Sopelana, in N Spain(Figs. 1 and 2). Their ichnological features have been investigated in

Fig. 1. A) Paleogeographic map of the Pyrenean Basin, with location of Bidart, Sopelana and Zumaia sections. B) Schematic lithologic logs of the Bidart and Sopelana sections,including sequence stratigraphic distribution (bottom). Note: The Zumaia section is included for comparison.
43F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
order to show some biogenic and sedimentary phenomena throughthe K/Pg boundary interval, and to define the similarities anddifferences between these two sections. The latter aspect is of specialinterest, because in deep basins that presumably record homoge-neous hemipelagic sediment supply, ichnological features maybecome a powerful tool to recognize subtle lateral changes in thesedimentation regime and the palaeoenvironmental conditions atthe sea floor.
2. Location and setting
The late Cretaceous and Paleogene in the Pyrenees are representedby a wide variety of sedimentary rocks, including continental alluvialclastics, shallow marine carbonates and deep-water hemipelagitesand turbidites accumulated in a broad E–W elongated embaymentopened into the Bay of Biscay and the North Atlantic (Baceta, 1996;Baceta et al., 2004; Plaziat, 1981; Pujalte et al., 1998). Basinal

Fig. 2. Lithological columns of the Sopelana B and Bidart sections with sample locations and range of trace fossils.
44 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
hemipelagites and turbidites were deposited in the central part of theembayment, the so-called Basque Basin (Pujalte et al., 1998), atdepths ranging from 1000 to1500 m (Fig. 1).
The late Maastrichtian–earliest Ypresian was a phase or relativetectonic quiescence, during which turbidite deposition was reducedand the Basque Basin became dominated by regular alternations ofhemipelagic limestones and marls, usually grouped in beddingcouplets and bundles as representative of precession and short-eccentricity climatic cycles (Dinarès-Turell et al., 2003, 2007).
The best known K/Pg boundary sections being of the Basque Basinare located in coastal cliffs between the north of Bilbao and the southof Baiona. Up to twelve of these K/Pg sections can be identified, theZumaia section being the reference for all of them (see Apellaniz et al.,1997 for a synthesis). Bidart and Sopelana, the two K/Pg sectionsselected for ichnological analysis, are 100 km apart (see Fig. 1 forcorrelation with the Zumaia section according the most significantcorrelation levels a to e). The Bidart section is located in south-western France, in a long beach few kilometres to the north of the

45F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
Bidart town (43º27′N and 1º35′E), within an upper Cretaceous–Eocene succession that dips 15–35º to the north. The Sopelana sectionis located north of Bilbao, in northern limb of the Cretaceous–Paleogene Byscay Sinclinorium (Fig. 1, 43º23′N and 2º59′W). Theupper Cretaceous–Paleocene basinal succession of the Sopelanasection is affected by reverse and strike-slip faults, which causestratigraphic repetitions and the existence of three different K/Pgboundary outcrops. Two of these sections have been analysed:Sopelana A (Sopelana Beach) and Sopelana B (above the cliff)sections, which in previous papers were labelled, respectively, asSopelana III (Apellaniz, 1998) and Sopelana I (Orue-Etxebarria, 1983,1984, 1985).
3. Lithostratigraphy and biostratigraphy of the K/Pg boundary inthe Basque Basin
3.1. Bidart and Sopelana sections
A profuse research has been conducted in the last decades in theBidart section (see Gallala et al., 2009 for a review). Recently, the K/Pgboundary transition at Bidart section was studied, focusing on thetrace fossil assemblage (Rodríguez-Tovar et al., 2010; Figs. 2 and 3).The upper 4 m of the Maastrichtian contain grey and pinkish marlsand marly limestones. The rusty layer marking the K/Pg boundary iswell preserved and about 2 mm thick. Above it, the first Daniandeposits occur in an about 15 cm thick dark clay layer, overlain bynearly 23 cm thick reddish to greenish marls and marly limestones.Above them, a 93 cm thick interval of partly parallel laminatedwhite to reddish limestones is present, followed by a 1–1.5 m thickpebbly mudstone bed made up of roundish clasts of hemipelagiclimestones embedded in a marl-lime matrix. This clastic deposit isinterpreted as reflecting an episode of erosion and redeposition bysubmarine debris flows. The upper part of the studied section consists
Fig. 3. Outcrop of the K/Pg boundary layer at Bidart. A) General view showing top of the MaStars pointed by arrows indicate debris flow calciruditic beds; note their convex up upper bDanian (indicated by stars); compare to A.
of white and reddish bioturbated limestones with thin (2–3 cm)marlstone interbeds.
The Bidart section can be considered, together with the classicalZumaia section (Fig. 1), as one of the most complete land-based K/Pgboundary sections exposed in the eastern margin of the AtlanticOcean (Apellaniz et al., 1997; Haslett, 1994; Seyve, 1990), as recentlycorroborated by magnetostratigraphic, calcareous nannofossils andplanktic foraminifera analyses (Galbrun and Gardin, 2004; Gallala etal., 2009). These works reveal a continuous sedimentation across theK/Pg boundary transition, without any biostratigraphically significanthiatus. The planktic foraminifera biozonation (Gallala et al., 2009),is as follow: a) at the Upper Maastrichtian, the Abathomphalusmayaroensis Zone is differentiated, b) the Guembelitria cretacea Zonethat marks the base of the Lower Danian, around 15 cm thick in theBidart section, and subdivided into the Hedbergella holmdelensis andParvularugoglobigerina longiapertura subzones, and c) the P. eugubinaZone, around 100 cm thick in the Bidart section, that is subdividedinto the P. sabina and Eoglobigerina simplicissima subzones.
The K/Pg boundary transition at the Sopelana section (A and B) hasbeen comparatively less studied than that at Bidart. Descriptions ofthe Sopelana A section can be found in Lamolda et al. (1983) andOrue-Etxebarria (1985). Especially interesting is the complex iridiumdistribution, with two iridium spikes, the first coeval with the finalCretaceous planktonic foraminiferal extinction, and the second incoincidence with the nannoplankton crisis (Rocchia et al., 1988).Detailed analysis of the Sopelana section (A and B) corroboratessimilar facies composition as in the Bidart section, but differentthicknesses (Figs. 2 and 4). The upper 7 m of the Maastrichtianconsists of grey and pinkish marls of changing endurance. Above, the4–5 cm thick dark clay K/Pg boundary layer from the base of theDanian, containing the 2 mm thick rusty layer at the base, is followedby 22–25 cm of green-grey marls showing primary lamination in theupper part. Overlapping it, a 90 cm thick interval of white and reddish
astrichtian marlstones, the boundary layer and the Danian limestones and marlstones.edding planes. B) Detail of A. C) Boundary layer. D) Debris flow calciruditic beds in the

Fig. 4. The Sopelana section and its trace fossils in the totally bioturbated Maastrichtian marlstones. A, B) The K–Pg boundary section; the boundary layer marked by the dashed line;a fault in A marked by solid line. C) Light-filled Chondrites isp. (Ch), vertical section, sample So-103. D) Dark-filled Chondrites isp., sub-horizontal parting surface, sample So-85e. E)Chondrites isp. (Ch), dark-filled Thalassinoides (Thd), light-filled Thalassinoides (Thi) and Planolites isp. (Pl), vertical section, sample So-105. F) Thalassinoides isp. (Th) and Chondritesisp. (Ch), horizontal parting surface, sample So-85c. G) Thalassinoides isp. (Th), Chondrites isp. (Ch), Planolites isp. (Pl) and Trichichnus isp. (Tr), vertical section, sample So-85 h. H)Zoophycos isp., sample So-91 polished along the spreite. I) ?Zoophycos isp. (?Zo) and ?Phycosiphon isp. (Phy), sub-horizontal section, sample So-91.
46 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
limestones which includes a level with some evidence of soft-sediment synsedimentary deformation, in the form of irregularfolding and fracturation with differentiation of isolated limestoneclasts. Biostratigraphic analysis based on planktonic foraminifera andcalcareous nannoplankton revealed that the first occurrence of theTertiary planktonic foraminifera Globigerina (E.) eugubina is a fewcentimetres (8–10 cm) above the extinction of Upper Cretaceous
planktonic foraminifera (Lamolda et al., 1983). Thus, the lowermostpart of the Paleogene corresponding to the Guembelitria cretacea Zonewas not identified (Lamolda et al., 1983; Mary et al., 1991). This zonecould be represented in the 2–3 cm just above the contact with theMaastrichtian, but the relative scarcity and poor preservation ofplanktonic foraminifera above the rusty layer avoid a conclusiveinterpretation (Canudo, 1994). The incompleteness of the Sopelana A

Fig. 5. Ichnofabrics and trace fossils of the pre-event Maastrichtian marlstones at Bidart.A) Thalassinoides isp. (Th) cross cut by Chondrites isp. (Ch), Planolites isp. (Pl), all in thetotally bioturbated background; polished slab; sample Bd2. B) Zoophycos isp. (Zo) in thetotally bioturbated background. Polished slab, sample Bd11. C) Trichichnus isp. (Tr) on arough surface, sample Bd8.
47F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
section was also revealed based on the graphic correlation analysis;the 2 cm separating the red layer from the FAD of P. eugubina implythe absence of the Zone P0 estimated in 50 000 yr in duration(MacLeod and Keller, 1991; MacLeod et al., 1997). The K/Pg boundaryinterval in the Sopelana B section situated 60 m from the formerabove the seacliffs, is more complete and significantly betterpreserved. According to Apellaniz et al. (1997) and Apellaniz (1998)the boundary layer marking the boundary is 7 cm thick and contains aplanktic foraminifer assemblage characteristic of the Guembelitriacretacea Zone. The overlying marl-marlstones that are 28 cm thick arealso well preserved. Their lower 22–23 cm corresponds to theParvularugoglobigerina longiapertura Zone.
4. Trace fossil analysis
Only occasionally trace fossils or bioturbational structures havebeen mentioned in the K/Pg boundary section at Bidart (Gorostidi andLamolda, 1995; Haslett, 1994; Smit and Ten Kate, 1982), while in theSopelana section, Lamolda et al. (1983) refer to “a distinct burrowedsurface” at the top of the purple marls corresponding to the latestMaastrichtian. Recently, Rodríguez-Tovar et al. (2010), present apreliminary ichnological analysis of the K/Pg boundary transition atthe Bidart section, focusing on the effects of different tracemakers onthe redistribution of calcareous nannofossils throughout the K/Pgboundary. The section shows a relatively abundant and conspicuousdark-coloured trace fossil assemblage with Chondrites, Planolites,Thalassinoides, Trichichnus and Zoophycos, and less abundant light-coloured ichnofossils including Planolites and Thalassinoides. Calcar-eous nannofossils are re-distributed in fillings of some ichnotaxa inthe Bidart section, with danian calcareous nannofossil assemblagespresent in Maastrichtian samples, evidenced by abundant Paleogenecalcareous nannofossils just below the K/Pg boundary (Rodríguez-Tovar et al., 2010).
The sections were analyzed layer by layer. Selected samples werecollected continuously at the K/Pg boundary transition, and moresparsely in the rest of the succession, and then cut and polished in thelaboratory. The cut and polished surfaces were wetted in order toobtain better contrast for photographs.
4.1. Synopsis of trace fossils
The trace fossil assemblage identified in the K/Pg sections of Bidartand Sopelana is low diverse, with 5 common ichnogenera (Chondrites,Planolites, Thalassinoides, Trichichnus and Zoophycos), together with ?Phycosiphon only registered in the Sopelana section.
Chondrites isp. (Figs. 4C–D, F–G, 5A, 6A, D–G, 8B–C) is observed (1)on parting surfaces as branched burrow system composed ofcompressed straight or curved equidimensional cylindrical, unwalledelements (Figs. 4D, 6A, D), or in (2) cross section as patches of circularto elliptical spots and short bars, some of them branched (Figs. 4C, E–G); they are cross sections of a branched tubular burrow system. Thebranches are under acute angle. The filling is darker (Fig. 4D–G), rarelylighter than the host rock (Fig. 4C). Larger forms (Fig. 4C–G), 1.2–2 mm in diameter, and smaller forms, 0.3–0.9 mm diameter (Fig. 4E),can be distinguished. At least some of the larger forms belong toChondrites targionii (Brongniart, 1828) and at least some of thesmaller forms belong to Chondrites intricatus (Brongniart, 1823) butlimited observations along the tunnels preclude accurate determina-tions. The smaller form can be found in the fillings of someThalassinoides or Planolites, and the larger forms in Thalassinoides.
Chondrites von Sternberg, 1833 is a feeding system of unknowntracemakers (e.g., Osgood, 1970) which, according to Kotake (1991a),are surface ingestors, packing their faecal pellets inside burrows.According to Seilacher (1990) and Fu (1991), the tracemaker may beable to live at the aerobic–anoxic interface as a chemosymbioticorganism.
?Phycosiphon isp. (Fig. 4I) is a patch of sub-horizontal small lobeswith a marginal tunnel about 0.7 mmwide. The curvature and patchyoccurrence suggests Phycosiphon incertum Fischer-Ooster, 1858, butfragmentary preservation only in one specimen precludemore certaindetermination. Phycosiphon is common in fine-grained marinesediments and is produced by deposit feeders (Wetzel and Bromley,1994). Its geometry was lately studied by Naruse and Nifuku (2008).
Planolites isp. (Figs. 4E, G, 5A, 6D–E, H, 8B–D) occurs as horizontalto oblique, straight to slightly winding, simple flattened cylinders,2.5–4 mm, mostly 3 mm wide, without lining. They are filled withdark grey rarely light grey fine-grained material. Some fillings arepenetrates with the small form of Chondrites. Planolites is a faciescrossing form. It has been discussed by Pemberton and Frey (1982)and Keighley and Pickerill (1995) and interpreted as a pascichnion,probably produced by a number of different organisms.
Thalassinoides isp. (Figs. 4C–G, 5A, 6A–B, E, G–H, 7A–B, F, 8B–C)occurs as straight or slightly winding, horizontal to oblique flattened,unlined cylinders, showing Y-shaped, rarely T-shaped branches andrare, vertical to oblique shafts. The cylinders are swollen at thebranching points, which are mostly 55–110 mm apart. The cylindersare filled with homogenous, or rarely meniscate, mostly darkersediment. The menisci are composed of thicker (up to 7 mm) darkportions and narrower (up to 3 mm) light portions. Rarely, granulatedfilling composed of light and dark chips of fine-grained sediment, lessthat 1 mm in diameter, can be present. Some fillings are reworkedwith the larger or smaller Chondrites (Figs. 4F, 5A). Prevailing smallerforms are 4–9 mm in diameter, with swellings up to 15 mm wide.Some forms, especially in the Danian (Fig. 8C), are larger, 15–30 mm
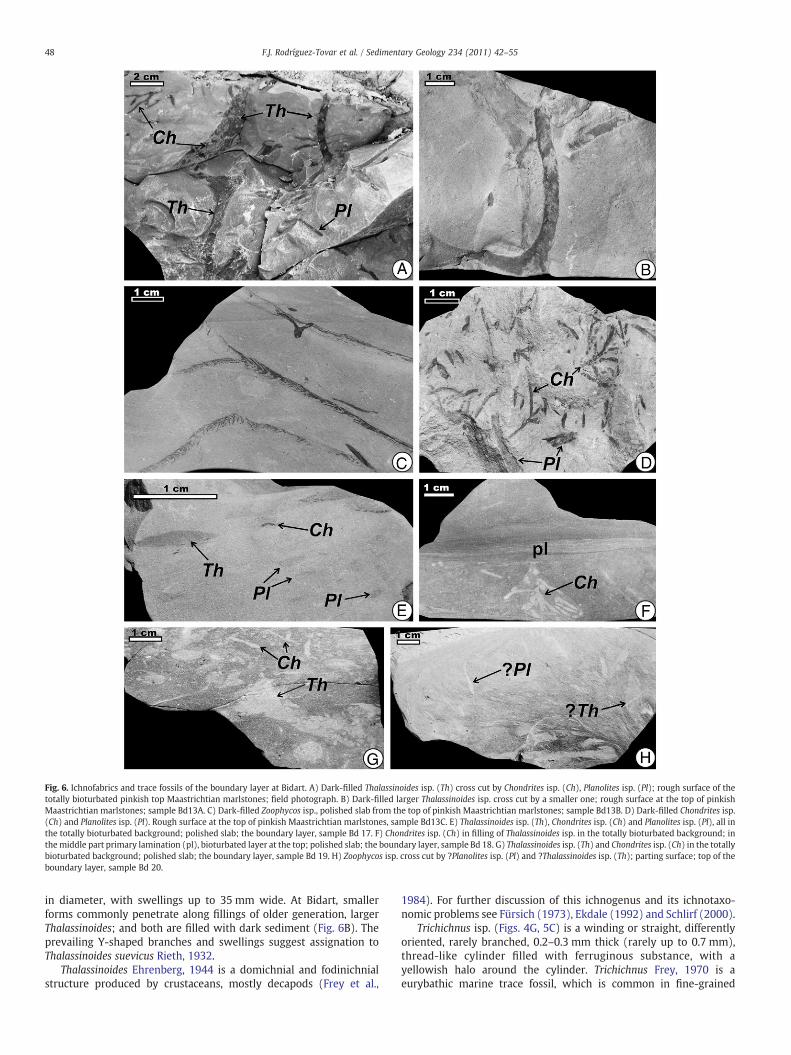
Fig. 6. Ichnofabrics and trace fossils of the boundary layer at Bidart. A) Dark-filled Thalassinoides isp. (Th) cross cut by Chondrites isp. (Ch), Planolites isp. (Pl); rough surface of thetotally bioturbated pinkish top Maastrichtian marlstones; field photograph. B) Dark-filled larger Thalassinoides isp. cross cut by a smaller one; rough surface at the top of pinkishMaastrichtian marlstones; sample Bd13A. C) Dark-filled Zoophycos isp., polished slab from the top of pinkish Maastrichtian marlstones; sample Bd13B. D) Dark-filled Chondrites isp.(Ch) and Planolites isp. (Pl). Rough surface at the top of pinkish Maastrichtian marlstones, sample Bd13C. E) Thalassinoides isp. (Th), Chondrites isp. (Ch) and Planolites isp. (Pl), all inthe totally bioturbated background; polished slab; the boundary layer, sample Bd 17. F) Chondrites isp. (Ch) in filling of Thalassinoides isp. in the totally bioturbated background; inthemiddle part primary lamination (pl), bioturbated layer at the top; polished slab; the boundary layer, sample Bd 18. G) Thalassinoides isp. (Th) and Chondrites isp. (Ch) in the totallybioturbated background; polished slab; the boundary layer, sample Bd 19. H) Zoophycos isp. cross cut by ?Planolites isp. (Pl) and ?Thalassinoides isp. (Th); parting surface; top of theboundary layer, sample Bd 20.
48 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
in diameter, with swellings up to 35 mm wide. At Bidart, smallerforms commonly penetrate along fillings of older generation, largerThalassinoides; and both are filled with dark sediment (Fig. 6B). Theprevailing Y-shaped branches and swellings suggest assignation toThalassinoides suevicus Rieth, 1932.
Thalassinoides Ehrenberg, 1944 is a domichnial and fodinichnialstructure produced by crustaceans, mostly decapods (Frey et al.,
1984). For further discussion of this ichnogenus and its ichnotaxo-nomic problems see Fürsich (1973), Ekdale (1992) and Schlirf (2000).
Trichichnus isp. (Figs. 4G, 5C) is a winding or straight, differentlyoriented, rarely branched, 0.2–0.3 mm thick (rarely up to 0.7 mm),thread-like cylinder filled with ferruginous substance, with ayellowish halo around the cylinder. Trichichnus Frey, 1970 is aeurybathic marine trace fossil, which is common in fine-grained
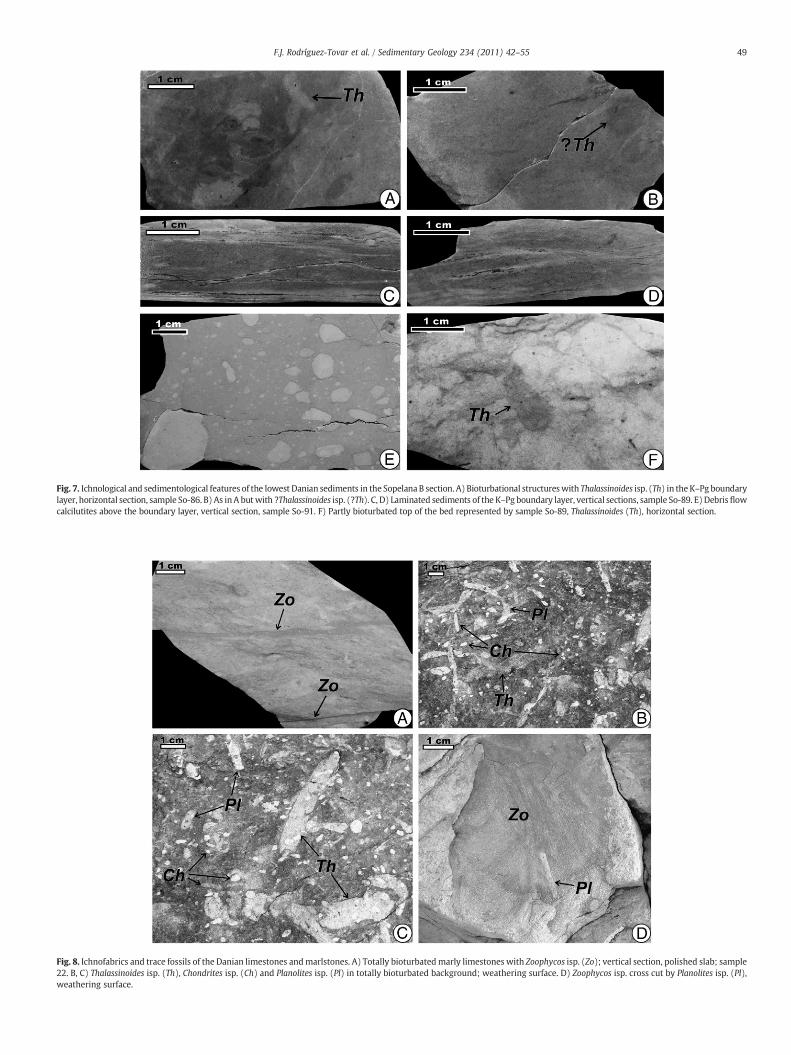
Fig. 7. Ichnological and sedimentological features of the lowest Danian sediments in the Sopelana B section. A) Bioturbational structureswith Thalassinoides isp. (Th) in theK–Pg boundarylayer, horizontal section, sample So-86. B) As inA butwith ?Thalassinoides isp. (?Th). C, D) Laminated sediments of the K–Pg boundary layer, vertical sections, sample So-89. E) Debrisflowcalcilutites above the boundary layer, vertical section, sample So-91. F) Partly bioturbated top of the bed represented by sample So-89, Thalassinoides (Th), horizontal section.
Fig. 8. Ichnofabrics and trace fossils of the Danian limestones andmarlstones. A) Totally bioturbatedmarly limestones with Zoophycos isp. (Zo); vertical section, polished slab; sample22. B, C) Thalassinoides isp. (Th), Chondrites isp. (Ch) and Planolites isp. (Pl) in totally bioturbated background; weathering surface. D) Zoophycos isp. cross cut by Planolites isp. (Pl),weathering surface.
49F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55

50 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
sediments. Trichichnus is regarded as the domichnial burrows ofmarine meiofaunal deposit feeders (Frey, 1970). Possibly, theproducer of Trichichnus was a chemosymbiont (Uchman, 1995), asin the case of the Chondrites producer (Fu, 1991; Seilacher, 1990).McBride and Picard (1991) suggest that Trichichnus had a moreopportunistic character than Chondrites because it occurs more deeplyin very poorly oxygenated sediments. For taxonomic discussion seeUchman (1999).
Zoophycos isp. (Figs. 4H–I, 5B, 6C, 8A–D) occurs on parting surfacesas horizontal or oblique lobes and tongues, up to 40 cm wide, filledwith spreite laminae encircled by a thin marginal tunnel. In crosssection it is seen as straight or curved stacked stripes, 1.5–4 mm thick,7–28 mm apart, which can be massive or filled with spreitemanifested on the surface as menisci-like structures. They are a partof a helical burrow system.
Zoophycos s.l. is generally considered a structure produced by someyet undiscovered deposit-feeder, which is referred to sipunculids(Wetzel and Werner, 1981), polychaete annelids, arthropods (Ekdaleand Lewis, 1991), or echiuran worms (Kotake, 1992). Kotake (1989,1991b) reported that Zoophycos is produced by surface ingestors oforganic detritus. However, the precise ethological interpretation ofZoophycos remains still controversial. Bromley and Hanken (2003)suggested that the upper helical part of a large Pliocene Zoophycosfrom Rhodes, Greece, is a deposit-feeding structure, and lateral lobesdeveloping from its lower part are sulphide wells for chemosymbioticbacteria. The same interpretation refers to a similar but smallerZoophycos from the Miocene of Austria, which displays very steeplobes in its lowermost part (Pervesler and Uchman, 2004).
4.2. Ichnological features
4.2.1. Abundance of trace fossils, and size of ThalassinoidesThe trace fossil abundance (number of specimens and visual
density estimations), and size of Thalassinoides, varies significantlybetween the Bidart and Sopelana sections (Sopelana A and B sectionsconsidered together). Thalassinoides is distinctly more abundant andlarger at Bidart (mean diameter of 35 specimens=10.1 mm, maximalwidth 30 mm) than at Sopelana (mean diameter of 28 speci-mens=6.4 mm, maximal width 14 mm). The larger Chondrites isaround two times more abundant in the K/Pg boundary at Bidart thanat Sopelana. Reverse tendency is clear for the smaller Chondrites andvery distinctly (by more or less four times) for Trichichnus. Zoophycosis comparatively more abundant at Bidart than at Sopelana. ?Phycosiphon has been only recognized at Sopelana.
4.2.2. Feeding behaviourThe trace fossil assemblage reveals different feeding behaviour.
They belong to domichnia/fodinichnia (Thalassinoides), chemichnia(Chondrites, Trichichnus), pascichnia (Planolites) and fodinichnia(Zoophycos, ?Phycosiphon). All of them indicate that the tracemakersobtained food from sediment.
4.2.3. Depth of penetrationThe dark filling of some trace fossils located in the uppermost
Maastrichtian sediments clearly derives from the dark sediments ofthe K/Pg boundary layer (black-filled trace fossil ranges in Fig. 9).Detailed microfacies analysis at Sopelana, especially in section B,reveals the presence of numerous dark-filled trace fossils comingdown from the dark-boundary layer into the light upper Maastrich-tian sediments. Dark-filled Chondrites and Zoophycos are present fromseveral centimetres above to below the K/Pg boundary. Zoophycos andChondrites are the deeper trace fossils. In the Bidart section, largedark-coloured Zoophycos isp. is the deepest trace, in some casespenetrating up to −45 cm (45 cm below the K/Pg boundary layer) inthe marlstones underlying the K/Pg boundary surface. In the SopelanaB section it occurs at depths of −22 cm below the boundary.
Chondrites penetrates up to −25 cm at Bidart, −40 cm at SopelanaB and −22 cm at Sopelana A. Thalassinoides was found up to −32 cmat Bidart and −40 cm at Sopelana B, and −20 cm at Sopelana A.Planolites is still present at −40 cm at Bidart, −30 cm at Sopelana B;dark-filled Planolites was not found at Sopelana A.
The values are minimal because the burrows can be filled fromdifferent levels above the boundary rusty layer, corresponding to38 cm thick (15 cm thick dark clay layer + 23 cm thick reddish togreenishmarls) at Bidart, and at least 22 cm thick at Sopelana A and atleast 25 cm thick at Sopelana B of green-grey marls. Data from theSopelana A section are not fully representative because of limitednumber of samples obtained by digging due to outcrop constraints.These data should be rather treated as complementary to theSopelana B section.
Trichichnus is filled with pyritized sediment and ?Phycosiphonwithlight material. Therefore, depth of penetration of these ichnotaxacannot be measured in respect to the boundary layer. However, it isknown that Phycosiphon belongs to shallow and moderate deep tiersand Trichichnus is a deep-tier burrow (e.g., Uchman, 1999).
4.2.4. Trace fossil distribution through the K/Pg boundary intervalThe boundary layer at Bidart contains the rusty layer at the base,
just above 1–2 cm thick, grey, thin, totally bioturbated marlymudstone (Fig. 9). The rusty layer is only partly bioturbated. Above,the boundary layer is totally bioturbated, except for two laminatedintervals, which are crossed only by some trace fossils (Fig. 6F). AtSopelana (Fig. 4A–B), the boundary layer is disturbed by tectonicsliding, therefore its primary internal structure is partly obliterated. Inthe Sopelana A section, the boundary layer is totally bioturbated, withtwo rusty laminae in the lower part (Fig. 9). In the Sopelana B section,the boundary layer is totally bioturbated (Fig. 7A–B) except for itsuppermost laminated few centimetres (Fig. 7C–D). The trace fossilscontent of the boundary layer is almost the same as the dark-filledtrace fossil assemblage.
In the lowest 2 m of Danian above the boundary layer at Bidart,trace fossils are poorly recorded (Fig. 9); partly by diageneticobliteration in the lower part and by the presence of the debris flowcalcirudite beds (Fig. 3D) in the upper part. The top of the firstcalcirudite layer is bioturbated and contains ?Thalassinoides. The bedsof the first metres are at least partly bioturbated, but no discrete tracefossils was recognized. Above, well preserved Chondrites, Planolites,Thalassinoides and Zoophycos, are well visible on bioturbatedbackground (Fig. 8A–D).
At Sopelana, the first metre of sediments above the boundary layeris totally bioturbated (Fig. 9) except for the debris flow deposits(Fig. 7E–F). In the bioturbated parts, trace fossils are poorly visible,probably due to poor colour contrast and diagenetic obliteration.
5. Discussion
5.1. Trace fossil assemblage, abundance of trace fossils, size ofThalassinoides and depositional environment
The trace fossil assemblage is low diverse. All the ichnotaxaidentified are rather common in fine-grained, low-energy open-marine sediments of the Mesozoic and Cenozoic. Their low diversityand the presence of Zoophycos suggest the Zoophycos ichnofacies,which typically occurs down the offshore but beyond the areas ofturbiditic sedimentation (e.g., Frey and Seilacher, 1980). The totalbioturbation of the fine-grained calcareous sediments that character-ized the two K/Pg boundary sections studied, and the general lack ofstorm or turbiditic beds agrees with this general interpretation. Thus,ichnological data point to upper bathyal zone, with no significantdifferences between the depositional settings corresponding to bothsections. This interpretation accords partly with previous data basedon microfossil assemblages. The dominance of calcareous taxa in the

Fig. 9. The K/Pg boundary sections at Bidart and Sopelana showing the boundary layer and trace fossil ranges.
51F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
Upper Maastrichtian and lower Danian benthic foraminiferal assem-blages from the Bidart section are interpreted as indicating upper–middle bathyal depths between 500 and 700 m (Alegret et al., 2004).Kuhnt and Kaminski (1993) assigned middle bathyal water depth tothe Maastrichtian–Paleocene of the Zumaia section, comparable withthe Sopelana section, based on agglutinated foraminiferal assem-blages, with absolute values at about 1.000 m depth suggested byHerm (1965).
According to sequence stratigraphic analyses (Fig. 1), the K/Pgboundary transition is included in a transgressive system tract (TST)belonging to a 3rd order depositional sequence (Apellaniz et al., 1997;Baceta, 1996; Pujalte et al., 1998). The top of this sequence is related toan erosive event, resulting from a sea-level drop, reflected by brecciaswell developed at the Bidart section. This interpretation accords withthat from Peybernés et al. (1996), interpreting that the breccias weredeposited under conditions of low sea level, and the underlyinglimestones under high sea-level conditions. Other possibility is thatthey could be interpreted as a slump triggered by local tectonicsduring the early Danian (Alegret et al., 2004). For Seyve (1984), theBidart section can be interpreted as located close to a palaeoslope.Alegret et al. (2004) interpreted that the dominance of calcareousbenthic foraminifera through the Bidart section may be indicative of asmall supply of terrigenous sediment at Bidart as compared to thecoeval Sopelana section (Kuhnt and Kaminski, 1993), where agglu-tinated foraminifera are dominant.
The higher abundance of Thalassinoides and its larger size, higherabundance of Zoophycos and larger Chondrites, lower abundance ofTrichichnus and small Chondrites at Bidart than at Sopelana can berelated to palaeogeographic position and the distance of the basinmargin. The Sopelana section occupied a more basinal position thanthe Bidart section during the Maastrichtian and Paleocene. This canresult in a slightly greater depth, as confirmed by the foraminiferaldata, and lesser food supply at Sopelana than at Bidart. Environmentalstress related to decrease in food can result in decrease in abundanceof larger trace fossils (Thalassinoides, larger Chondrites, and Zoophycos)
and thus a dominance of smaller forms (small Chondrites, andTrichichnus). This is in accordance to the decrease of both macro-benthic biomass (Rowe, 1983) and number of macrofaunal organismswith increasing depth, especially rapid in the first 2 km (Rowe, 1983;Rowe et al., 1982), and the decrease of biomass with distance to theshore (if not disturbed by upwellings and other currents) (Gage andTyler, 1991, and references therein). Also the size of the animalsdecreases generally with increasing depth (e.g., Gage and Tyler, 1991;Thiel, 1975). Thus, the variability within the Zoophycos ichnofacies canbe a proxy of food availability related in simple cases to the distancefrom the shore and water depth. Smaller size of ichnotaxa anddecrease in abundance of larger trace fossils can point to more distantand deeper settings.
5.2. Feeding behaviour, nutrients availability and oxygenation ofsediments
The presence of chemichnia and trace fossils which tracemakerssequestered sediments from the sea floor in deep burrows (Zoophy-cos) suggest at least periodical food deficiency pressing animals tomore sophisticated searching for food.
Variations between oligotrophic and mesotrophic conditions havebeen alluded to interpret marked quantitative changes in benthiccommunity structure through the K/Pg boundary transition in bothsections (Alegret et al., 2004; Kuhnt and Kaminski, 1993). In theSopelana sections, major change in the quantitative composition ofdeep-sea agglutinated foraminiferal community structure is related,probably, to a dramatic drop of productivity in surface waters, nearlyinstantaneously with the terminal Cretaceous event, that determinedthe collapse of the food web for benthic communities (Kuhnt andKaminski, 1993). At the lower part of the boundary clay, just after theK/Pg boundary, they record the predominance of low-oxygen tolerantand infaunal agglutinated foraminiferal assemblages, in relation tooligotrophic surface conditions. Coarsely agglutinated forms arecommon, indicating the increase in detrital supply to the deep sea,

52 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
which Kuhnt and Kaminski (1993) interpret in terms of a short-livedregressive event. Although not present in Sopelana, cm-thicksiliciclastic or mixed carbonate-siliciclastic turbidites occur overlyingor within the K/Pg clay in several basinal sections in the area betweenSopelana and Bidart (e.g., Zumaia, Urrutxua, Herrera or Monte Urko;Baceta, 1996; Apellaniz, 1998). Thus, the relative abundance ofcoarsely agglutinated foraminifers could be explained rather by thepresence of discrete levels rich in resedimented sand detrital grains.
In the Bidart section, benthic assemblage shows a similar pattern(Alegret et al., 2004). Upper Maastrichtian benthic foraminiferalassemblages are diverse and heterogeneous, with infaunal andepifaunal forms, revealing mesotrophic conditions. At the K/Pgboundary, a drastic decrease in diversity and heterogeneity isrecorded, which is coeval with a major diminution in the percentageof infaunal taxa. This drastic change is related to a significant decreasein the food supply from surface waters to the sea floor, oligotrophicconditions, due to the collapse of the food web accompanying themass extinction of primary producers (during very limited foodsupply to the sea floor, food particles were consumed at the sedimentsurface by epifaunal species, whereas the percentage of infaunalspecies decreased due to the lack of organic matter in the underlyingsediment). In the early Danian, the low diversity of the assemblages,together with the low percentage of infaunal forms, suggests thatprimary productivity had not completely recovered for more than200 kyr after the K/Pg boundary event. However, even if benthicforaminifera indicate oligotrophic conditions just after the K/Pgboundary, blooms of dinoflagellates have been documented in thedark clay layer (Peybernés et al., 1996), which would have providedlarge amounts of food that reached the sea floor. For Alegret et al.(2004), primary productivity during the early Danian was dominatedby blooms of non-calcareous primary producers; such a change inphytoplankton composition induced a stressful environment for thebenthic fauna. These authors indicated that it was not only the drasticdecrease in primary productivity, but also the changes in phytoplank-ton type (from calcareous to organic-walled phytoplankton) just afterthe K/Pg boundary that caused the low diversity, high-dominancebenthic foraminiferal assemblages from the lower part of the dark claylayer (Alegret et al., 2004). It seems, however, that the changes did notinfluenced tracemakers.
Total bioturbation of sediments (except for some interval of theboundary layer and the debris flow sediments in the Danian) suggestgenerally good oxygenation below sediment–water interface. Datafrom benthic microfossils do not reveal a significant incidence of lowoxygenation inmicrobenthic environment (Alegret et al., 2004; Kuhntand Kaminski, 1993); only, as previously indicated, just after the K/Pgboundary at the Sopelana section, low-oxygen tolerant and infaunalagglutinated foraminiferal assemblages are dominant, but mainlyrelated to oligotrophic conditions (Kuhnt and Kaminski, 1993).
5.3. Depth of penetration and rate of sedimentation
The penetration depth of the dark-filled trace fossils can be treatedas frozen tiering pattern of burrows (Bromley, 1996). At Bidart thedeepest trace fossil is Zoophycos, similarly to the Caravaca section inthe Betic Cordillera (Rodríguez-Tovar and Uchman, 2006). Zoophycostracemaker probably preferred the partly stiff or partly firm substrateof the Maastrichtian marls, which were dewatered during the slowlyaccumulating Danian dark-boundary layer. The increased cohesionmay have prevented the Chondrites tracemaker to get deeperpenetration. Commonly, Chondrites, is the deepest tier burrow in theCretaceous marls (Ekdale and Bromley, 1991). Such a situation is metat Sopelana, however Thalassinoides penetrates almost to the samedepth. Zoophycos is distinctly rarer here than at Bidart, what suggestsit was approaching to its environmental limit. In such a situation, theZoophycos tracemaker could penetrate to smaller depth than in itsoptimal environment. A comparatively higher cohesion for the
Maastrichtian sediments at the Bidart section could be related witha higher rate of sedimentation; a 40 mm/kyr sedimentation rate forthe Maastrichtian is calculated for the Bidart section (Nelson et al.,1991) against 25 mm/ky for the Sopelana section (Mary et al., 1991).
5.4. The Cretaceous–Paleogene (K–Pg) event incidence
The trace fossil assemblage can be considered with respect to theK–Pg event. The content of pre-event latest Maastrichtian ichnoas-semblage, characterized by light-filled trace fossils is nearly the sameas the lowest Danian ichnoassemblage characterized by dark fillingderiving from the boundary layer (Figs. 4D–I, 6A–D). Light-filledZoophycos has not been observed in the uppermost 7 m of Maas-trichtian at Sopelana, but it is present below. Trichichnus was notdetected at above the boundary layer at Bidart and at Sopelana. Thus,according to trace fossil assemblages, the K/Pg boundary event revealsa minor incidence onmacrobenthic environment. Previous trace fossilanalyses in other K/Pg boundary sections have revealed that changesin environmental parameters at the seafloor during the deposition ofthe K/Pg boundary sediments did not significantly affect colonizationbymacrobenthic tracemakers, showing a relatively rapid colonization,specially by organisms with a high independence with respect tosubstrate features (i.e., Chondrites and Zoophycos tracemakers;Rodríguez-Tovar and Uchman, 2004a, 2006, 2008; Rodríguez-Tovar,2005; Rodríguez-Tovar and Martín-Peinado, 2009). This compara-tively minor incidence in sections the here studied, accords with thatinterpreted for the benthic environment based on microfossilsdata. Benthic foraminifera assemblage reveals similar responsethrough the K/Pg boundary transition in the Sopelana (Kuhnt andKaminski, 1993) and Bidart (Alegret et al., 2004) sections. As isgenerally recognized at the K/Pg boundary, benthic foraminifers didnot experienced significant extinction (only three species disappearsin Bidart; Alegret et al., 2004) in any of both sections, comparing withthe well known mass extinction of planktic organisms which isinterpreted as revealing a weak benthopelagic coupling at that time(Alegret et al., 2004 and references therein). However, markedquantitative changes in their community structure (percentage ofparticular morphogroups, diversity, etc.), are evident and mainlyrelated to changes in productivity at the K/Pg boundary (see above;Kuhnt and Kaminski, 1993; Alegret et al., 2004). In the Sopelanasection, benthic foraminiferal populations show a slow recovery thattook at least 50 ky following the K/Pg boundary event (Kuhnt andKaminski, 1993).
The planktic foraminiferal extinction pattern in the Bidart sectionsuggest a sudden catastrophic extinction at the K/Pg boundary, at thebase of the 2 mm thick rusty layer, affecting, at least 53 species out ofthe 72 species registered at the uppermost Maastrichtian (Gallalaet al., 2009).
5.5. Bioturbational redistribution of sediment components through theK/Pg boundary event
One especially important aspect to be taken into consideration is thepossible bioturbational disturbance of the K/Pg boundary, a fact whichis usually underestimated. Detailed ichnological analysis in theCaravaca section (SE Spain), revealed that trace fossils range contin-uously from the lowermost Danian to the uppermost Maastrichtiansediments, with the rusty layer being cross cut by Zoophycos andChondrites, and also penetrated laterally by Chondrites (Rodríguez-Tovar and Uchman, 2008). Thus, the authors claimed on the possibleredistribution of the components related to the K/Pg boundary impactdue to the tracemakers activity. Recently, the effects of differenttracemakers on the redistribution of calcareous nannofossils through-out the K/Pg boundary at the Bidart section have been revealed(Rodríguez-Tovar et al., 2010). Bloom of the Paleogene calcareousnannofossil appearances just below the K/Pg boundary are related to

53F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
infiltration into dark trace fossil infillings proceeding from the earliestPaleogene. In the Sopelana sections a complex iridium distribution isrecognized, with two iridium spikes registered in the dark-boundarylayer (the first coeval with the final Cretaceous planktonic foraminiferalextinction, and the second in coincidence with the nannoplanktoncrisis; plate B in Rocchia et al., 1988). As previously indicated, distri-bution of dark-trace fossil in the Sopelana sections (Fig. 9) reveals thatthe boundary layer is totally bioturbated, with a continuous record ofChondrites from the entire dark-boundary layer, except for itsuppermost laminated few centimetres, to the uppermost Maastrichtiansediments. Moreover, Planolites and Thalassinoides range from thelower part of the dark-boundary layer to the uppermost Maastrichtiansediments, except for Zoophycos registered only in the dark-boundarylayer. Thus the interval of the dark-boundary layer recording the twoiridium spikes is affected by an important significant bioturbation.Therefore, a redistribution by tracemakers activity affecting iridiumconcentration in the dark-boundary layer is possible in the Sopelanasections, but amore detailed analysis, out of the focus of this research, isnecessary for confirmation, even more taking into account thatrecently, the relatively mobile behaviour of iridium within thesediments in depth has been demonstrated (Martín-Peinado andRodríguez-Tovar, 2010; Miller et al., 2010; Racki et al., in press).
5.6. Continuity at the K/Pg boundary transition
As previously commented, the Bidart section can be considered asone of the most complete K/Pg boundary sections exposed in theeastern margin of the Atlantic Ocean (Apellaniz et al., 1997; Haslett,1994; Seyve, 1990). Recent magnetostratigraphic, calcareous nanno-fossil and planktic foraminifera analyses reveal a continuous sedi-mentation across the K/Pg boundary transition, without anybiostratigraphically significant hiatus (Galbrun and Gardin, 2004;Gallala et al., 2009). However at the Sopelana section, the lowermostpart of the Paleogene (Guembelitria cretacea Zone) was not identified(Lamolda et al., 1983; Mary et al., 1991), and graphic correlationanalysis indicated the incompleteness of the Sopelana section withthe absence of the Zone P0 estimated in 50,000 yr in duration(MacLeod and Keller, 1991; MacLeod et al., 1997). The distribution oftrace fossils in the Sopelana section does not reveal any evidentdisruption of ichnological features, but a continuous and similarrecord from the dark-boundary layer to the upper Maastrichtiansediments. Macrobenthic colonization of the dark-boundary layercould be conducted 50,000 yr after the K/Pg boundary, according tothe estimation of the duration of the absent Zone P0 (MacLeod andKeller, 1991; MacLeod et al., 1997). However, during this timechanges in substrate features (i.e., substrate consistency) around theK/Pg boundary must be envisaged, determining variations in theichnological features which were no registered. Thus, the absence ofsediments corresponding to an estimated duration of 50,000 yr is notconfirmed by ichnological analysis.
6. Concluding remarks
Ichnological analysis at the K/Pg boundary transition in the Bidart(SE France) and Sopelana (N Spain) sections reveals a low diversetrace fossil assemblage, with five common ichnogenera (Chondrites,Planolites, Thalassinoides, Trichichnus and Zoophycos), together with ?Phycosiphon registered only in the Sopelana section. This assemblagesuggests the Zoophycos ichnofacies, which typically occurs down theoffshore but beyond the areas of turbiditic sedimentation.
Minor differences in the earliest Danian ichnoassemblage (dark-filled trace fossils) between sections are related to palaeogeographicposition within the upper bathyal zone and to the distance to thebasinmargin. The higher abundance of large Thalassinoides, Zoophycosand larger Chondrites, and lower abundance of Trichichnus and smallChondrites, at Bidart than at Sopelana, is in agreement with a more
distal position of Sopelana relative to Bidart, with a slightly higherdepth and smaller food supply.
Feeding strategies showing the presence of chemichnia and tracefossils which tracemakers sequestered sediments from the sea floor indeep burrows (Zoophycos) denotes some intervals of periodical fooddeficiency pressing animals to more sophisticated searching for food.
A generalized total bioturbation of sediments point to goodoxygenation below sediment–water interface.
The similarity between the pre-event (light-filled trace fossils) andthe post-event (dark-filled trace fossils) trace fossil assemblagereveals a minor incidence of the K/Pg boundary event on the macro-benthic environment. The near continuous record of dark-filled tracefossils through the K/Pg boundary interval, from the upper part of thedark-boundary layer, especially in the Sopelana section, to theuppermost Maastrichtian sediments prevent on a possible redistri-bution of the biotic and abiotic components.
Acknowledgements
The field research was partially supported by the Basque CountryUniversity and by the Zaragoza University. Additional support for A.U.was given by the Jagiellonian University (DS funds). This research wassupported by projects CGL2008-03007/CLI, CGL2008-00009/BTE, andRNM-3715, and the research group RNM-178.
References
Alegret, L., 2007. Recovery of the deep-sea floor after the Cretaceous–Paleogeneboundary event: the benthic foraminiferal record in the Basque–Cantabrian basinand in South-eastern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology255, 181–194.
Alegret, L., Kaminski, M.A., Molina, E., 2004. Paleoenvironmental recovery after theCretaceous/Paleogene boundary crisis: evidence from the marine Bidart section(SW France). Palaios 19, 574–586.
Apellaniz, E., 1998. Los foraminíferos planctónicos en el tránsito Cretácico-Terciario.Ph.D. Thesis, Universidad del País Vasco, España.
Apellaniz, E., Baceta, J.I., Bernaola-Bilbao, G., Nuñez-Betelu, K., Orue-Etxebarria, X.,Payros, A., Pujalte, V., Robin, E., Rocchia, R., 1997. Analysis of uppermostCretaceous-lowermost Tertiary hemipelagic successions in the Basque Country(western Pyrenees): evidence for a sudden extinction of more than half plankticforaminifer species at the K/T boundary. Bulletin de la Societé Géologique de France168, 783–793.
Baceta, J.I., 1996. El Maastrichtiense superior, Paleoceno e Ilerdiense inferior de laRegión Vasco-Cantábrica: secuencias, cronoestratigrafía y paleoceanografía. Ph.D.Thesis, Universidad de Zaragoza, España.
Baceta, J.I., Pujalte, V., Serra-Kiel, J., Robador, A., Orue-Etxebarria, X., 2004. La CordilleraPirenaica: Maastrichtiense superior, Paleoceno e Ilerdiense inferior. In: Vera, J.A.(Ed.), Geología de España: SGE-IGME, pp. 308–313.
Bromley, R.G., 1996. Trace Fossils. Biology, Taphonomy and Applications, SecondEdition. Chapman & Hall, London.
Bromley, R.G., Hanken, N.-M., 2003. Structure and function of large, lobed Zoophycos,Pliocene of Rhodes, Greece. Palaeogeography, Palaeoclimatology, Palaeoecology192, 79–100.
Brongniart, A.T., 1823. Observations sur les Fucoïdes. Société d'Historie Naturelle deParis, Mémoire 1, 301–320.
Brongniart, A.T. 1828. Histoire des végétaux fossiles ou recherches botaniques etgéologiques sur les végétaux renfermés dans les diverses couches du globe, 1. G.Dufour and E. d'Ocagne, Paris.
Canudo, J.J., 1994. Bioestratigrafía y evolución de los foraminíferos planctónicos enel tránsito Cretácico-Terciario en España. In: Molina, E. (Ed.), Evolución yregistro fossil: Cuadernos Interdisciplinares de la Universidad de Zaragoza, vol. 5,pp. 141–165.
Dinarès-Turell, J., Baceta, J.I., Pujalte, V., Orue-Etxebarria, X., Bernaola, G., Lorito, S.,2003. Untangling the Paleocene climatic rhythm: and astronomical calibrated EarlyPaleocene magnetostratigraphy and biostratigraphy at Zumaia (Basque basin,northern Spain). Earth and Planetary Science Letters 216, 483–500.
Dinarès-Turell, J., Baceta, J.I., Bernaola, G., Orue-Etxebarria, X., Pujalte, V., 2007. Closingthe Mid-Palaeocene gap: toward a complete astronomically tuned PalaeoceneEpoch and Selandian and Thanetian GSSPs at Zumaia (Basque Basin, W Pyrenees).Earth and Planetary Science Letters 262, 450–467.
Ehrenberg, K., 1944. Ergänzende Bemerkungen zu den seinerzeit aus dem Miozän vonBurgschleinitz beschrieben Gangkernen und Bauten dekapoder Krebse. Paläonto-logische Zeitschrift 23, 345–359.
Ekdale, A.A., 1992. Muckraking and mudslinging: the joys of deposit-feeding. In:Maples, C.G., West, R.R. (Eds.), Trace Fossils: The Paleontological Society ShortCourses in Paleontology, Vol. 5, pp. 145–171.

54 F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
Ekdale, A.A., Bromley, R.G., 1984. Sedimentology and ichnology of the Cretaceous–Tertiary boundary in Denmark: implications for the causes of the terminalCretaceous extinction. Journal of Sedimentary Petrology 54, 681–703.
Ekdale, A.A., Bromley, R.G., 1991. Analysis of composite ichnofabrics: an example inuppermost Cretaceous chalk of Denmark. Palaios 6, 232–249.
Ekdale, A.A., Lewis, D.W., 1991. The New Zealand Zoophycos revisited. Ichnos 1,183–194.
Ekdale, A.A., Stinnesbeck, W., 1998. Trace fossils in Cretaceous–Tertiary (KT) boundarybeds in northeastern Mexico: implications for sedimentation during the KTboundary event. Palaios 13, 593–602.
Fischer-Ooster, C., 1858. Die fossilen Fucoiden der Schweizer-Alpen, nebst Erörterun-gen über deren geologisches Alter. Huber, Bern.
Frey, R.W., 1970. Trace fossils of Fort Hays Limestone Member of Niobrara Chalk (UpperCretaceous), West-Central Kansas. The University Kansas Paleontological Con-tributions 53, 1–41.
Frey, R.W., Seilacher, A., 1980. Uniformity in marine invertebrate ichnology. Lethaia 23,183–207.
Frey, R.W., Curran, A.H., Pemberton, G.S., 1984. Tracemaking activities of crabs and theirenvironmental significance: the ichnogenus Psilonichnus. Journal of Paleontology58, 511–528.
Fu, S., 1991. Funktion, Verhalten und Einteilung fucoider und lophoctenoiderLebensspuren. Courier Forschungsinstitut Senckenberg 135, 1–79.
Fürsich, F.T., 1973. A revision of the trace fossils Spongeliomorpha, Ophiomorpha andThalassinoides. Neues Jahrbuch für Geologie und Paläontologie Monatschefte 12,719–735.
Gage, J.D., Tyler, P.A., 1991. Deep-Sea Biology: A Natural History of Organisms at theDeep-Sea Floor. Cambridge Univ. Press, Cambridge.
Galbrun, B., Gardin, S., 2004. New chronostratigraphy of the Cretaceous–Paleogeneboundary interval at Bidart (France). Earth and Planetary Science Letters 224, 19–32.
Gallala, N., Zaghbib-Turki, D., Arenillas, I., Arz, J.A., Molina, E., 2009. Catastrophic massextinction and assemblage evolution in planktic foraminifera across the Creta-ceous/Paleogene (K/Pg) boundary at Bidart (SW France). Marine Micropaleontol-ogy 72, 196–209.
Gorostidi, A., Lamolda, M.A., 1995. La nanoflora calcárea y el tránsito KT de la sección deBidart (SW de Francia). Revista Española de Paleontología extra, pp. 153–168.
Haslett, S.K., 1994. Planktonic foraminiferal biostratigraphy and paleoceanography ofthe Cretaceous–Tertiary boundary section at Bidart, South-west France. CretaceousResearch 15, 179–192.
Herm, D., 1965. Mikropalaeontologische-stratigraphische Untersuchungen in Kreide-flysch zwischen Deva und Zumaya (Prov. Guipúzcua, Nordspanien). Zeitschrift derDeutschen Geologischen Gesellschaft 15, 277–348.
Keighley, D.G., Pickerill, R.K., 1995. The ichnotaxa Palaeophycus and Planolites: historicalperspectives and recommendations. Ichnos 3, 301–309.
Kotake, N., 1989. Paleoecology of the Zoophycos producers. Lethaia 22, 327–341.Kotake, N., 1991a. Packing process for filling material in Chondrites. Ichnos 1, 277–285.Kotake, N., 1991b. Non-selective surface deposit feeding by the Zoophycos producers.
Lethaia 24, 379–385.Kotake, N., 1992. Deep-sea echiurans: possible producers of Zoophycos. Lethaia 25,
311–316.Kuhnt, W., Kaminski, M.A., 1993. Changes in the community structure of deep water
agglutinated foraminifers across the K/T boundary in the Basque Basin (NorthernSpain). Revista Española de Micropaleontología 25, 57–92.
Lamolda, M.A., Orue-Etxebarria, X., Proto-Decima, F., 1983. The Cretaceous–Tertiaryboundary in Sopelana (Biscay, Basque country). Zitteliana 10, 663–670.
MacLeod, N.M., Keller, G., 1991. How complete are Cretaceous/Tertiary boundarysections? A chronostratigraphic estimate based on graphic correlation. GeologicalSociety of America Bulletin 103, 1439–1457.
MacLeod, N., Rawson, P.F., Forey, P.L., Banner, F.T., Boudagher-Fadel, M.K., Bown, P.R.,Burnett, J.A., Chambers, P., Culver, S., Evans, S.E., Jeffery, C., Kaminski, M.A., Lord, A.R., Milner, A.C., Milner, A.R., Morris, N., Owen, E., Rosen, B.R., Smith, A.B., Taylor, P.D.,Urquhart, E., Young, J.R., 1997. The Cretaceous–Tertiary biotic transition. Journal ofthe Geological Society of London 154, 265–292.
Martín-Peinado, F.J., Rodríguez-Tovar, F.J., 2010. Mobility of iridium in terrestrialenvironments: implications for the interpretation of impact-related mass-extinc-tions. Geochimica et Cosmochimica Acta 74, 4531–4542.
Mary, C., Moreau, M.G., Orue-Etxebarria, X., Apellaniz, E., Courtillot, V., 1991.Biostratigraphy and magnetostratigraphy of the Cretaceous/Tertiary Sopelanasection (Basque country). Earth and Planetary Science Letters 106, 133–150.
McBride, E.F., Picard, D.M., 1991. Facies implications of Trichichnus and Chondrites inturbidites and hemipelagites, Marnoso-arenacea Formation (Miocene), NorthernApennines, Italy. Palaios 6, 281–290.
Miller, K.G., Sherrell, R.M., Browning, J.V., Paul Field, M., Gallagher, W., Olsson, R.K.,Sugarman, P.J., Tourto, S., Wahyudi, H., 2010. Relationship between mass extinctionand iridium across the Cretaceous–Paleogene boundary in New Jersey. Geology 38,867–870.
Molina, E., Alegret, L., Arenillas, I., Arz, J.A., Gallala, N., Hardenbol, J., Von Salis, K., Steurbaut,E., Vandenbeghe, N., Zaghbib-Turki, D., 2006. The global boundary stratotype sectionandpoint for thebaseof theDanianStage (Paleocene, Paleogene, "Tertiary", Cenozoic)at El Kef, Tunisia: original definition and revision. Episodes 29, 263–273.
Naruse, H., Nifuku, K., 2008. Three-dimensionalmorphology of the ichnofossils Phycosiphonincertum and its implication for paleoslope inclination. Palaios 23, 270–279.
Nelson, B.K., MaCleod, G.K., Ward, P.D., 1991. Rapid change in strontium isotopiccomposition of sea water before the Cretaceous/Tertiary boundary. Nature 351,644–647.
Orue-Etxebarria, X., 1983. Los Foraminíferos planctónicos del Paleógeno del Sinclinoriode Bizkaia (Corte de Sopelana—Punta de la Galea). Kobie 13, 175–249.
Orue-Etxebarria, X., 1984. Los foraminíferos planctónicos del Paleógeno del Sinclinoriode Bizkaia (Corte de Sopelana–Punta de la Galea). Kobie 14, 351–429.
Orue-Etxebarria, X., 1985. Descripción de Globigerina hillebralldti n. sp. en ellimiteCretácico/Terciario de 1a sección de Sopelana (Pais Vasco). Evolución de losprimeros foraminíferos planctónicos al comienzo del Terciario. Newsletters onStratrigraphy 15, 71–80.
Osgood, R.G., 1970. Trace fossils of the Cincinnati Area. Palaeontographica Americana 6,193–235.
Pemberton, G.S., Frey, R.W., 1982. Trace fossil nomenclature and the Planolites–Palaeophycus dilemma. Journal of Paleontology 56, 843–881.
Pervesler, P., Uchman, A., 2004. Ichnofossils from the type area of the Grund Formation(Miocene, Lower Badenian) in northern Lower Austria (Molasse Basin). GeologicaCarpathica 55, 103–110.
Peybernés, B., Fondecave-Wallez, M.J., Eichène, P., Bost, J., Sibe, B., Marais, M., Quilès, G.,1996. La Limite Crétacé–Paléocène: Phénomènes Biologiques, Evénements Géolo-giques d'après les Sites de la Côte Basque. Centre Régional de DocumentationPédagogique d'Aquitaine, Bordeaux.
Plaziat, J.C., 1981. Late Cretaceous to late Eocene paleogeographic evolution ofsouthwest Europe. Palaeogeography, Palaeoclimatology, Palaeoecology 36,263–320.
Pujalte, A., Baceta, J.I., Orue-Etxebarria, X., Payros, A., 1998. The Paleocene of the BasqueCountry, W Pyrenees, Spain: facies and sequence development in a deep-waterstarved basin. In: de Graciansky, P.C., Hardenbol, J., Jacquin, T., Vail, P.R. (Eds.),Mesozoic and Cenozoic Sequence Stratigraphy of European Basins: SEPM, Conceptsin Sedimentology and Paleontology, vol. 60, pp. 311–325.
Racki, G., Machalski, M., Koeberl, C., Harasimiuk, M., in press. The weathering-modifiedrecord of a new Cretaceous-Palaeogene site at Lechówka near Chelm, SE Poland,and its palaeobiologic implications. Acta Paleontologica Polonica 55, (in press),doi:10.4202/app.2010.0062.
Rieth, A., 1932. Neue Funde spongeliomorpher Fucoiden aus dem Jura Schwabens.Geologische und Paläontogische Abhandlungen, N.F. 19, 257–294.
Rocchia, R., Boclet, D., Bonté, P., Buffetaut, E., Orue-Etxebarria, X., Jaeger, J.-J., Jéhanno,C., 1988. Structure de l'anomalie en iridium à la limite Crétacé-Tertiarie du site deSopelana (Pays Basque Espagnol). Comptes Rendus de l´Académie des Sciences deParis 307, 1217–1223.
Rodríguez-Tovar, F.J., 2005. Fe-oxide spherules infilling Thalassinoides burrows at theCretaceous–Paleogene (K–P) boundary: evidence of a near-contemporaneousmacrobenthic colonization during the K–P event. Geology 33, 585–588.
Rodríguez-Tovar, F.J., Martín-Peinado, F.J., 2009. The environmental disaster ofAznalcóllar (southern Spain) as an approach to the Cretaceous–Palaeogene massextinction event. Geobiology 7, 533–543.
Rodríguez-Tovar, F.J., Uchman, A., 2004a. Trace fossils after the K–T boundary eventfrom the Agost section, SE Spain. Geological Magazine 141, 429–440.
Rodríguez-Tovar, F.J., Uchman, A., 2004b. Ichnotaxonomic analysis of the Cretaceous/Palaeogene boundary interval in the Agost section, south-east Spain. CretaceousResearch 25, 635–647.
Rodríguez-Tovar, F.J., Uchman, A., 2006. Ichnological analysis of the Cretaceous–Palaeogene boundary interval at the Caravaca section, SE Spain. Palaeogeography,Palaeoclimatology, Palaeoecology 242, 313–325.
Rodríguez-Tovar, F.J., Uchman, A., 2008. Bioturbational disturbance of the Cretaceous–Palaeogene (K–Pg) boundary layer: implications for the interpretation of the K–Pgboundary impact event. Geobios 41, 661–667.
Rodríguez-Tovar, F.J., Martínez-Ruiz, F., Bernasconi, S.M., 2002. Carbon isotopecomposition of bioturbation infills as indication of the macrobenthic-colonisationtiming across the Cretaceous–Tertiary boundary (Agost section, SE Spain).Geochimica et Cosmochimica Acta 66, A644.
Rodríguez-Tovar, F.J., Martínez-Ruiz, F., Bernasconi, S.M., 2004. Carbon isotope evidencefor the timing of the Cretaceous–Palaeogene macrobenthic colonisation at theAgost section (southeast Spain). Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 203, 65–72.
Rodríguez-Tovar, F.J., Martínez-Ruiz, F., Bernasconi, S.M., 2006. Use of high-resolutionichnological and stable isotope data for assessing completeness of a K–P boundarysection, Agost, Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 237,137–146.
Rodríguez-Tovar, F.J., Uchman, A., Molina, E., Monechi, S., 2010. Bioturbational re-distribution of Danian calcareous nannofossils in the uppermost Maastrichtianacross the K–Pg boundary at Bidart, SW France. Geobios 43, 569–579.
Rowe, 1983. Biomass and production of deep-sea biomass. In: Rowe, G.T. (Ed.), Deep-sea Biology. Wiley-Interscience, New York, pp. 97–121.
Rowe, G.T., Polloni, P.T., Haedrich, R.L., 1982. The deep-sea macrobenthos on theContinentalmarginof thenorthwestAtlanticOcean.Deep-SeaResearch 29A, 257–278.
Schlirf, M., 2000. Upper Jurassic trace fossils from the Boulonnais (northern France).Geologica et Palaeontologica 34, 145–213.
Schulte, P., Alegret, L., Arenillas, I., Arz, J.A., Barton, P.J., Bown, P.R., Bralower, T.J.,Christeson, G.L., Claeys, P., Cockell, C.S., Collins, G.S., Deutsch, A., Goldin, T.J., Goto,K., Grajales-Nishimura, J.M., Grieve, R.A.F., Gulick, S.P.S., Johnson, K.R., Kiessling,W., Koeberl, C., Kring, D.A., MacLeod, K.G., Matsui, T., Melosh, J., Montanari, A.,Morgan, J.V., Neal, C.R., Nichols, D.J., Norris, R.D., Pierazzo, E., Ravizza, G.,Rebolledo-Vieyra, M., Reimold, W.U., Robin, E., Salge, T., Speijer, R.P., Sweet, A.R.,Urrutia-Fucugauchi, J., Vajda, V., Whalen, M.T., Willumsen, P.S., 2010. TheChicxulub asteroid impact and mass extinction at the Cretaceous–PaleogeneBoundary. Science 327, 1214–1218.
Seilacher, A., 1990. Aberration in bivalve evolution related to photo- and chemosym-biosis. Historical Biology 3, 289–311.
Seyve, C., 1984. Le passage Crétacé–Tertiaire à Pont Labau. Bulletin des Centres deRecherches Exploration-Production Elf-Aquitaine 8, 385–423.

55F.J. Rodríguez-Tovar et al. / Sedimentary Geology 234 (2011) 42–55
Seyve, C., 1990. Nannofossil biostratigraphy of the Cretaceous–Tertiary boundary in theFrench Barque Country. Bulletin Centre Recherches Exploration-Production Elf-Aquitaine 14, 553–572.
Smit, J., 1999. The global stratigraphy of the Cretaceous–Tertiary boundary impactejecta. Annual review of Earth and Planetary Sciences 27, 75–113.
Smit, J., Ten Kate, W.G.H.Z., 1982. Trace-element patterns at the Cretaceous–Tertiaryboundary: consequences of a large impact. Cretaceous Research 3, 307–332.
Sternberg, G.K., 1833. Versuch einer geognostisch-botanischen Darstellung der Florader Vorwelt. IV Heft. C. E. Brenck, Brenck, Regensburg.
Thiel, H., 1975. The size structure of the deep-sea benthos. Internationale Revue derGesamten Hydrobiologie 60, 575–606.
Uchman, A., 1995. Taxonomy and palaeoecology of flysch trace fossils: the Marnoso-arenacea formation and associated facies (Miocene, Northern Apennines, Italy).Beringeria 15, 3–115.
Uchman, A., 1999. Ichnology of the Rhenodanubian flysch (Lower Cretaceous-Eocene)in Austria and Germany. Beringeria 25, 65–171.
Wetzel, A., Bromley, R., 1994. Phycosiphon incertum revisited: Anconichnus horizontalisis its junior subjective synonym. Journal of Paleontology 68, 1396–1402.
Wetzel, A., Werner, F., 1981. Morphology and ecological significance of Zoophycos indeep-sea sediments off NW Africa. Palaeogeography, Palaeoclimatology, Palaeoe-cology 32, 185–212.





























