Embed Size (px)
Citation preview

El Colegio de la Frontera Sur
Edad y crecimiento del pulpo rojo Octopus maya(Voss y Solís, 1966) por medio de la lectura de incrementos
en el estilete.
TESISpresentada como requisito parcial para optar al grado de
Maestría en Ciencias en Recursos Naturales y Desarrollo Rural
por
Natalia Celia Albarrán Mélzer
2010

I
AGRADECIMIENTOS
Agradezco al Colegio de la Frontera Sur (Unidad Campeche) por la oportunidad
de crecer en el ámbito de la investigación permitiéndome realizar este trabajo en
sus instalaciones.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el apoyo otorgado
a través de la beca para la realización de esta investigación.
Al Dr. Unai Marcaida Aburto por su apoyo y guía durante la realización de esta
tesis.
Agradezco a mi comité revisor: Dr. Alberto de Jesús Navarrete, Dr. Manuel
Mendoza Carranza, Dr. Juan Carlos Pérez Jiménez, M. en C. Iván Méndez
Loeza por sus observaciones y sugerencias a este documento que enriquecieron
la tesis.
Mi agradecimiento al M. en C. Iván Méndez Loeza por sus recomendaciones,
explicaciones y ayuda en el conteo de las lecturas de los incrementos en el
estilete.
Con amor a mi familia por estar ahí siempre cuando los necesite, por creer en mi
y animarme a seguir adelante.
Al M. en C. Carlos Cesar Guzmán Aguirre por su incondicional apoyo, cariño y
paciencia.
A mis amigos por brindarme su amistad y hacerme reír en los mejores y peores
momentos.

II
CONTENIDO Página
LISTA DE FIGURAS................................................................................. IVLISTA DE TABLAS................................................................................... VRESUMEN................................................................................................ VI1. INTRODUCCIÓN.................................................................................. 1.1. Antecedentes................................................................................ 1.1.1. Biología de Octopus maya.................................................. 1.1.2. Ubicación taxonómica......................................................... 1.1.3. Pesquería de la especie...................................................... 1.1.4. Estudios de edad y crecimiento en pulpos.......................... 1.1.5. Estudios de edad y crecimiento en estructuras duras......... 1.1.6. Justificación.........................................................................
177
1112131518
2. OBJETIVOS......................................................................................... 2.1. Objetivo general........................................................................... 2.2. Objetivos particulares...................................................................
202020
3. HIPÓTESIS………………………………………………………………… 204. AREA DE ESTUDIO............................................................................. 215. MATERIALES Y MÉTODOS................................................................ 5.1. Trabajo de campo......................................................................... 5.2. Trabajo de laboratorio................................................................... 5.2.1. Preparación y montaje de las secciones del estilete.......... 5.2.2. Lectura de los incrementos................................................ 5.3. Análisis de datos.......................................................................... 5.3.1. Estructura de talla y peso................................................... 5.3.2. Relación entre dimensiones del estilete y el pulpo rojo….. 5.3.3. Estimación de la edad........................................................ 5.3.4. Precisión de las lecturas.................................................... 5.3.5. Patrón del crecimiento........................................................
2424262628292929303032
6. RESULTADOS..................................................................................... 6.1. Estructura de talla y peso.............................................................. 6.2. Estilete........................................................................................... 6.2.1. Relación entre dimensiones del estilete y el pulpo rojo...... 6.3. Estimación de la edad................................................................... 6.3.1. Microestructura del estilete y lectura de incrementos......... 6.4. Precisión de lecturas..................................................................... 6.5. Estimación del crecimiento............................................................ 6.5.1. Relación longitud de manto y peso con la edad.................. 6.5.2. Tasas de crecimiento.......................................................... 6.5.3. Fechas de nacimiento.........................................................
3535373739394042424648

III
7. DISCUSIÓN.......................................................................................... 7.1. Estilete........................................................................................... 7.1.1. Preservación y preparación de los estiletes........................ 7.1.2. Relación entre dimensiones del estilete y el pulpo rojo...... 7.2. Estimación de la edad................................................................... 7.2.1. Microestructura del estilete y lectura de incrementos......... 7.2.2 Deposición periódica de los incrementos............................ 7.3. Estimación del crecimiento............................................................ 7.3.1. Relación longitud de manto y peso con la edad.................. 7.3.2. Tasas de crecimiento.......................................................... 7.3.3. Fechas de nacimiento.........................................................
4949495255555658586368
8. CONCLUSIONES............................................................................... 709. LITERATURA CITADA....................................................................... 7210. ANEXO............................................................................................... 84

IV
LISTA DE FIGURAS Página
Figura 1. Pulpo rojo, Octopus maya (Voss y Solís, 1966). 8
Figura 2. Ubicación del Banco de Campeche en el Golfo de México.
Tomado de Jiménez (2005).
22
Figura 3. Localización de las zonas de muestreo. 24
Figura 4. Ubicación del estilete en la zona corporal interna de O.
maya. Modificado de Bizikov (2004).
25
Figura 5. Morfometría del estilete. Modificado de Bizikov (2004). 26
Figura 6. Técnica utilizada para el corte en el estilete. Modificado de
Bizikov (1996).
27
Figura 7. Estructura de tallas y pesos en machos y hembras. 36
Figura 8. Relaciones longitud de manto y peso total con longitud
total y diámetro máxima del estilete.
38
Figura 9. Microestructura del estilete en corte transversal, indicando
marca de nacimiento e incrementos de primer y segundo
orden.
39
Figura 10. Curva de tendencia de los conteos entre lectores. 41
Figura 11. Relaciones entre edad, longitud de manto y peso total en
hembras y machos.
46
Figura 12. Tasas de crecimiento diario e instantáneas en longitud de
manto y peso total en hembras y machos.
47
Figura 13. Distribución de las frecuencias de fechas de nacimiento
semanal del pulpo rojo en la temporada de 2007.
48

V
LISTA DE TABLAS Página
Tabla 1. Número de organismos seleccionados para la preparación
del estilete por localidad y sexo.
35
Tabla 2. Parámetros de la relación longitud de manto y peso total
entre el diámetro máximo y longitud total del estilete (mm)
para hembras y machos.
37
Tabla 3. Porcentaje de acuerdo entre lectores. 40
Tabla 4. Resultados de las pruebas de precisión y reproducibilidad
de conteos por lector.
42
Tabla 5. Parámetros entre longitud de manto (mm) y la edad en
hembras y machos.
43
Tabla 6. Parámetros entre peso total (g) y la edad en hembras y
machos.
44

VI
RESUMEN
Se estimó la edad y se describió el crecimiento del pulpo rojo, Octopus maya,
por medio de la lectura de los incrementos de crecimiento depositados en los
estiletes de 100 organismos seleccionados de cinco puertos de desembarque
pesqueros en el litoral de Campeche, México, durante 2007. La técnica a mano
alzada fue utilizada para el corte de los incrementos; los estiletes fueron
preservados en formol al 4% e incluidos en un portaobjetos con gelatina de
glicerol. La relación encontrada entre la longitud total (LTE) y el diámetro
máximo del estilete (DME) con la longitud de manto de los organismos (r2= 0.79
en LTE y r2= 0.80 en DME) es directamente proporcional al cuerpo somático del
pulpo. Esto podría sugerir que los estiletes del pulpo rojo son buenos
indicadores del crecimiento. Se identificaron cuatro zonas de incrementos de
crecimiento en el estilete y dos tipos de patrones de lectura. La relación entre la
longitud del manto y la edad para ambos sexos fue mayor (r2= 0.80), el modelo
logístico fue el que mejor se ajustó a los parámetros de crecimiento. Los machos
maduran antes que las hembras, iniciando a los 89 - 144 días y de 99 - 157 días
en hembras. La tasa de crecimiento diario en talla y peso total para hembras fue
de 0.59 mm/día, 18.7 g/día y en machos de 0.57 mm/día, 17.9 g/día.
Palabras clave: Octopus maya, estiletes, incrementos, edad, crecimiento.

1
1. INTRODUCCION
Los cefalópodos son organismos que presentan características muy particulares
que los distinguen de los demás moluscos. Las más notables son la ausencia de
una concha externa, a excepción de Nautilus, y la modificación del pie en brazos
y tentáculos. Son de crecimiento rápido, con gran variabilidad individual y vida
corta. Por consiguiente poseen una tasa alta de renovación, lo cual hace que se
encuentren mejor adaptados a soportar una fuerte presión de pesca, en
comparación con especies de peces con menor tasa de crecimiento (Caddy y
Rodhouse, 1998; Caddy, 1999).
La clase Cephalopoda se divide en tres subclases: Ammonoidea, Nautiloidea y
Coleoidea. En esta última se encuentra la familia Octopodidae, la cual
comprende a pulpos incirrados de hábitos principalmente bentónicos que se
caracterizan por tener ocho brazos circumorales que están provistos de
ventosas dispuestas en una o dos líneas. El género Octopus es el que sobresale
de esta familia por su importancia pesquera y su gran variedad de especies, se
han descrito alrededor de 100 especies, además han sido la base para el
conocimiento general de los cefalópodos (Boyle y Rodhouse, 2005).
Los octópodos son un recurso importante en las pesquerías de muchas regiones
costeras del mundo, ya que representan hasta el 10% de la producción mundial
(FAO 2001). En México, la pesquería de octópodos se encuentra sobre todo

2
establecida en las costas del Golfo de México y Mar Caribe (Nepita y Defeo,
2001). Ocupa el doceavo lugar nacional en cuanto a captura y el tercer lugar por
su valor comercial, superado sólo por la pesquería del camarón y el atún (INP-
SAGARPA, 2005). Se ha reportado que el 90% de la producción nacional se
encuentra localizada en los estados de Campeche y Yucatán (Nepita y Defeo,
2001; CONAPESCA-ITESM, 2004; INP-SAGARPA, 2005), basándose en la
explotación de dos especies, el pulpo rojo Octopus maya (Voss y Solís, 1966) y
el pulpo patón Octopus vulgaris (Cuvier, 1797) (Nepita y Defeo, 2001).
El pulpo rojo O. maya es una especie endémica que habita en la plataforma
continental de la península de Yucatán, se distribuye desde Ciudad del Carmen,
Campeche hasta Isla Mujeres, Quintana Roo (Solís, 1994, 1997). Constituye el
principal recurso pesquero en términos económicos y el segundo en volúmenes
de captura en Campeche y Yucatán, representando una de las pesquerías con
mayor potencial mundial (Nepita y Defeo, 2001; INP-SAGARPA, 2005). A partir
de 1996 se han realizado capturas importantes de las cuales cerca del 80% se
han exportado a los mercados europeos (Italia y España) y asiáticos (Corea y
Japón; INP-SAGARPA, 2005).
La información generada a partir de estudios de edad y crecimiento es crítica
para el desarrollo de modelos reales en pesquerías, siendo herramientas
esenciales para un manejo efectivo de las especies explotadas y la evaluación
del impacto sobre el ecosistema (Márquez y Ré, 2006). El proceso de

3
crecimiento en cefalópodos es uno de los más rápidos en el grupo de los
moluscos, alcanzado grandes tallas en poco tiempo (Callow, 1987). El
crecimiento es un aspecto biológico complejo, que en octópodos no ha sido
cuantificado hasta el momento con suficiente certeza, debido a su gran
plasticidad corporal.
Los cefalópodos presentan en general dos fases de crecimiento, la primera
exponencial y relativamente larga, y una segunda logarítmica, durante la cual la
tasa de crecimiento disminuye paulatinamente. La duración de ambas fases
dependerá de la especie y las condiciones del medio (Forsythe y Van Heukelem,
1987). Sin embargo, Jackson y Choat (1992) sugieren que para los cefalópodos
el crecimiento puede ser lineal o exponencial, pero siempre uniforme a lo largo
de todo el ciclo de vida del animal.
La mayoría de los estudios de crecimiento en cefalópodos se basan en métodos
indirectos como el análisis de la distribución de frecuencias de tallas, por medio
de las cuales se construye una curva de crecimiento conectando los valores
modales o de la media para los diferentes intervalos de tiempo (Arkhipkin, 1991).
Sin embargo la aplicación del análisis modal para el estudio de edad y
crecimiento en cefalópodos ha sido ampliamente criticada (Arkhipkin, 1991;
Caddy 1991; Hatfield y Rodhouse, 1994; Jackson, 1994; Yatsu et al., 1997).

4
Entre los inconvenientes más importantes están: etapa reproductiva extendida a
la mayor parte del año, poblaciones constituidas por individuos de distintas
generaciones en la misma área reproductiva o reemplazamiento gradual de
generaciones en las áreas de alimentación, especies altamente migratorias y
crecimiento individual muy variable (Arkhipkin, 1991; Caddy, 1991; Jackson,
1994; Jackson y Choat, 1992).
Es por ello que se ha recomendado que en la evaluación de poblaciones de
especies tropicales, se cuente con el mayor número de datos de frecuencia de
tallas, combinado con un número pequeño de lecturas de edad, basada en
incrementos diarios de estructuras duras corporales. Esta combinación permite
calibrar los resultados obtenidos de las tallas y explicar de una manera más
sencilla los conceptos teóricos del crecimiento a partir de datos de edad y de
talla, que solo sobre la base de la talla (Sparre y Venema, 1995).
El uso de técnicas directas, en base a la observación de incrementos en
diferentes estructuras duras corporales en cefalópodos como estatolito, pluma o
gladio y mandíbula, han permitido avanzar en el estudio de su crecimiento y
edad (Arkhipkin, 1991). El estatolito y la pluma son estructuras corporales más
ampliamente utilizadas en las especies del Superorden Decápoda (calamares y
sepias, entre otros) y las mandíbulas en el Orden Octópoda. Recientemente se
han realizado trabajos preliminares con otro tipo de estructuras internas
llamados estiletes presentes en pulpos (Bizikov, 2004) que han sido validados

5
para la estimación de la edad por Doubleday, et al. (2006) proveyendo
resultados similares en la fiabilidad y la precisión de estatolitos de calamar,
otolitos y vértebras de peces teleósteos.
La diversidad de los modelos matemáticos utilizados para describir el
crecimiento de cefalópodos se debe, al menos en parte, a la incapacidad de un
modelo único para tener en cuenta la amplia variabilidad en el crecimiento a
nivel individual. Numerosos factores bióticos y abióticos son capaces de
modificar drásticamente el crecimiento de los cefalópodos (Forsythe y Van
Heukelem, 1987). Como resultado de esta plasticidad del crecimiento, se
pueden utilizar diferentes modelos para describir el crecimiento de una sola
especie de pulpo. Las tallas de los individuos y las edades estimadas a partir de
estructuras duras son a menudo combinadas para describir el crecimiento
(Weatherley y Gill, 1987).
La tasa de crecimiento es un componente crítico de la estrategia de vida en
todas las especies. La tasa de crecimiento puede definir la longitud o edad de
madurez, la longitud y edad máxima y el potencial de reproducción (Musick,
1999). El crecimiento puede definirse en términos cuantitativos de varias
maneras, pero las más utilizadas son los modelos matemáticos de von
Bertalanffy, Gompertz y logístico. El modelo de von Bertalanffy es el más
aplicado ya que ha sido muy eficiente para describir el crecimiento de la mayoría
de los peces (Musick, 1999).

6
En México, a pesar de la importancia económica que representan los pulpos,
incluyendo a Octopus maya, la poca información biológica sobre datos de edad y
crecimiento no han sido contundentes para esclarecer aspectos como
longevidad, tasas de crecimiento, edad de madurez, dinámica de poblaciones y
(Voss y Solís, 1966) debido a que la mayoría de los estudios se han enfocado a
su utilización comercial.

7
1.1. ANTECEDENTES
1.1.1. Biología de Octopus maya
Los primeros trabajos sobre biología del pulpo en el Golfo de México se inician
con De la Peña (1957), Solís (1962) y Fuentes et al., (1965) cuando se
consideraba que la especie reportada era Octopus vulgaris. Posteriormente se le
define como especie nueva Octopus maya (Voss y Solís, 1966). Una descripción
del desarrollo de las investigaciones de esta especie se encuentra en Solís
(1987). Los estudios más relevantes sobre esta especie, se han hecho por Solís
(1962, 1967) quien analizó biometrías para definir periodos de veda.
El pulpo rojo Octopus maya es un organismo bentónico endémico de la
plataforma continental de la Península de Yucatán (Voss y Solís, 1966; Pérez-
Lozada et al., 2002), donde constituye una sola población. Presenta una
distribución que va desde los 20 km al sur de Punta Zacatal, Campeche, por el
Norte y Oeste de la Península de Yucatán, hasta Puerto Morelos, Quintana Roo
(Zambrano, 1992). Recientemente se reporta que su distribución se amplía
desde Ciudad del Carmen, Campeche, hasta Isla Mujeres, Quintana Roo (Solís,
1994, 1997).
El rasgo más distintivo de O. maya es la presencia de manchas oscuras u ocelos
debajo de los ojos (Fig. 1) y en la base del segundo y tercer brazo, haciéndose
menos aparente en los adultos (Voss y Solís, 1966). Llegan a medir casi un
metro de largo total y 3.5 g en promedio (Rosas et al., 2006).

8
Alcanzan grandes tallas en comparación con otras especies de Octópodos.
Llegan a medir hasta 250 mm de longitud de manto (LM) (Solís, 1967) y 1,300
mm de longitud total (FAO, 1989). Poseen un cuello corto y una cabeza pequeña
con ojos discretos. Los brazos son largos, corpulentos y reducidos hacia las
puntas. Poseen ventosas que se encuentran acomodadas en dos hileras que
son moderadamente grandes y más cortas en los machos (Voss y Solís, 1966).
El color de la especie es café oscuro, tornándose a café rojizo cuando se
estresa; sin embargo, su coloración varía dependiendo del medio circundante ya
que tienden a mimetizarse. El manto es café-grisáceo claro con manchas
oscuras en la papila contráctil (Voss y Solís, 1966; Zambrano, 1992).
Figura 1. Octopus maya (INP-SAGARPA, 2005).
Se estima que el apareamiento tiene lugar a partir de septiembre y se continúa
en octubre. Durante este periodo es frecuente encontrar ejemplares hembras
perfectamente maduras. En noviembre se observan huevos o puestas recién
depositadas e incluso hembras incubando. En diciembre se aprecian dos

9
regiones conspicuas en cada huevo: el embrión y el saco vitelino. En enero es
frecuente encontrar huevos recién eclosionados. En febrero esta situación se
hace mas patente, dándose por terminado en periodo normal reproductivo de la
especie (Solís, 1967). El desarrollo embrionario varía de 30 hasta 65 días
aproximadamente (Solís, 1967). Sin embargo, Van Heukelem (1977) indica un
desarrollo embrionario de 45 días a temperaturas de 25°C.
Fuentes et al., (1965) indicaron que a mediados de septiembre se inicia la
reproducción, culminando en marzo del siguiente año, entre septiembre y
noviembre las hembras alcanzan su máxima madurez sexual, cercanas a los 10
meses de edad (Solís y Chávez, 1986). Los datos de capturas y estudios de
fecundidad (Solís, 1967; Morales y Solís, 1987), describen un desove estacional
en el Banco de Campeche, con un pico en noviembre y diciembre. Morales y
Solís (1987), encontraron que en noviembre se produce el máximo desove y
ovoposición.
En el medio natural, la hembra fija los huevecillos a modo de racimos en el techo
de una cavidad o cueva, al mes del apareamiento (Voss y Solís, 1966).
Posteriormente realiza cuidados parentales que consisten en la defensa de los
huevos ante la depredación y la limpieza de estos por medio de su sifón o
brazos para protegerlos contra la invasión de hongos y bacterias (Zambrano,
1992). Durante la etapa de cuidados parentales la hembra no se alimenta
muriendo poco tiempo después de la eclosión de las crías. Los huevos son

10
grandes (17 mm de largo y 4.5 mm de ancho), piriformes, de color blanco
lechoso y su número por puesta fluctúa entre 1,500 y 2,000 (Solís, 1967).
Octopus maya no presenta paralarva. Las crías al eclosionar poseen todas las
características anatómicas y conductuales propias de un adulto, adoptan una
vida bentónica inmediatamente (INP-SAGARPA, 2005). Al nacer pesan
aproximadamente 0.1 g (Hanlon y Forsythe, 1985), nadan con facilidad,
presentan cambios de coloración e incluso expulsan tinta cuando son
estresados. Sus brazos son suficientemente hábiles para reptar y atrapar
alimentos y es normal que adopten una vida bentónica de inmediato (Solís y
Chávez, 1985).
El sexo se determina entre los tres y cinco meses de edad (25 a 450 g), cuando
empiezan a desarrollarse las gónadas (Rosas et al., 2006). La especie alcanza
su madurez aproximadamente a los 257 días después de su nacimiento
pesando alrededor de 3,262 g (Wood y O’Dor, 2000). El ciclo de vida dura en
condiciones de laboratorio entre 8 y 12 meses (Hanlon y Forsythe, 1985; Van
Heukelem, 1983).
Octopus maya es un organismo carnívoro y se le considera un consumidor
secundario, se alimenta básicamente de invertebrados, principalmente
crustáceos (cangrejos ermitaños) y algunos caracoles como Strombus gigas y S.
costatus sin embargo, se han encontrado escamas de peces en análisis

11
realizados a su contenido estomacal (Voss y Solís, 1966). Sus madrigueras
comúnmente están bordeadas por fragmentos de exoesqueletos de crustáceos y
restos de moluscos bivalvos. También ocupan conchas de grandes
gasterópodos (S. gigas, S. costatus, Pleuroploca gigantea) y diversos objetos
sumergidos, como envases de hojalata, plástico y neumáticos (Solís y Chávez,
1985).
1.1.2. Ubicación taxonómica de la especie de Octopus maya, de acuerdo
con Sweeney y Roper (1998)
Reino: Animalia
Phyllum: Mollusca
Clase: Cephalopoda Cuvier, 1979
Subclase: Coleoidea Bather, 1888
Superorden: Octopodiformes Young et al., 1998
Orden: Octopoda Leach, 1818
Suborden: Incirrata Grimpe, 1916
Familia: Octopodidae Orbigny, 1840
Subfamilia: Octopodinae Grimpe, 1921
Género: Octopus Cuvier, 1797
Especie: Octopus maya Voss y Solís, 1966

12
1.1.3. Pesquería de Octopus maya
La pesca de Octopus maya inició en Campeche en 1949 a causa de una
disminución de la pesca de otras especies. Se extendió al estado de Yucatán en
1970, registrando una producción de 304 t, (SAGARPA, 2005). A partir de 1979
Yucatán domina la producción de pulpo, en torno a las 5,000-7,000 t anuales, y
a partir de 1990 supera las 12,000 t. Las capturas en Campeche oscilaron
alrededor de las 2,500 t anuales (Solís, 1996, 1997; SEMARNAP, 1999), aunque
en los últimos tres años se superaron los 6,000 t equiparándose a las de
Yucatán.
En el período 1998-2006 se han registrado capturas a nivel nacional entre
16,000 a 24,000 t (Anuario Estadístico de Pesca 2004, Subdelegaciones de
Pesca en Campeche y Yucatán). Esta actividad genera un total de 15,000
empleos directos y una derrama económica anual de aproximadamente de 360
millones de pesos (SEMARNAP, 1999). Como medida de manejo la
administración de O. maya se regula basándose en la NOM-008-PESC-1993
(D.O.F. 21/12/93), establece una talla mínima de captura de 110 mm de longitud
de manto y se prohibe el empleo de ganchos, fisgas y arpones (D.O.F.
18/10/93). Según la NOM-009 PESC-1993 y D.O.F. 04/03/94, se establece un
periodo de veda del 16 de diciembre al 31 de julio de cada año en las aguas de
Campeche, Yucatán y Quintana Roo (Diario Oficial de la Federación, 2006).

13
1.1.4. Estudios de Edad y crecimiento en pulpos
Los estudios sobre edad y crecimiento de Octopus maya se han realizado
empleando métodos indirectos como el análisis modal de tallas. Solís y Chávez
(1986) indican que presenta una longevidad cercana a los 12 meses y de
crecimiento muy rápido (parámetros de Bertalanffy: L = 229.5 mm, K = 0.425
mm/día). Arreguín et al. (2000) realizan estimaciones de tasas de crecimiento
por medio del modelo de von Bertalanffy de las flotas artesanales para la
temporada de 1994 (1 de agosto a 15 de diciembre). Los parámetros de
crecimiento fueron L∞ = 252 mm de longitud de manto y una K = 0.467 mm/día.
Nepita y Defeo (2001) realizaron estimaciones en 19,251 individuos de O. maya
analizando la distribución de frecuencia de longitudes de un stock para estimar
los parámetros de crecimiento, por medio de varios métodos. Concluyen que el
método ELEFAN empleando el jackknife conjuntamente presentaron los mejores
valores de L∞ que varió entre 213 y 285 mm y K = 0.44 mm/día
En condiciones de cultivo Van Heukelem (1977,1983) determina que el ciclo de
vida dura entre 9 y 10 meses a temperaturas de 25 y 30°C con una tasa de 4.1%
(g/día) del crecimiento relativo al peso corporal. Hanlon y Forsythe (1985) citan
que esta especie presenta un crecimiento rápido, debido a sus elevadas tasas
de ingestión y de conversión de alimento, ubicadas entre 30 y 60%. Domínguez
et al. (2007) mencionan que la tasa de crecimiento en O. maya fue 3,3 ± 0,2 y -
0,0 ± 0,3% de peso corporal (g/día).

14
Hasta la fecha, se han aplicado cuatro enfoques principales con métodos
directos para estimar la edad en pulpos con diferentes grados de éxito: (1)
análisis de la microestructura en estructuras duras, que investiga la formación de
los anillos de crecimiento (Raya y Hernández, 1998; Hernández y Castro, 2001;
Oosthuizen 2003), (2) estudios de crecimiento de laboratorio que investiga el
efecto de la temperatura, las dietas y tasas metabólicas (Forsythe, 1984;
Forsythe y Hanlon, 1988; Segawa y Nomoto, 2002; Iglesias et al., 2004; Miliou et
al., 2005), (3) estudios de marcado de pulpos que determinan su crecimiento
durante un período específico de tiempo (Domain et al., 2000), y (4)
cuantificación histológica de lipofuscina en el tejido nervioso, que es proporcional
a la edad fisiológica y la cual se utiliza como indicador de la edad cronológica
(Sobrino y Real, 2003).
Todos estos cuatro métodos tienen limitaciones intrínsecas: (1) el análisis de
incrementos en algunas estructuras duras como picos es afectado por el
desgaste de esa estructura debido a la alimentación, resultando en estimaciones
inexactas (Hernández et al., 2001), (2) los resultados obtenidos en el laboratorio,
no garantizan que sean transferibles a las poblaciones silvestres (Joll, 1977;
Pecl y Moltschaniwskyj, 1999), (3) factores como la mortalidad causada por las
marcas, la reducción del crecimiento debido al estrés, el desconocimiento de la
edad antes del marcado, se deben considerar antes de la realización de un
estudio de recuperación de marcas (Semmens et al., 2004), y (4) la
cuantificación histológica de lipofuscina aún no se ha validado en animales de

15
edad conocida (Semmens, et al., 2004) .
1.1.5. Estudios de edad y crecimiento en estructuras duras en pulpos
El método de determinación de la edad que utiliza las estructuras duras similares
a los encontrados en los peces teleósteos (otolitos y vértebras), que no depende
de las características externas tales como los picos, evitan la incertidumbre de
los estudios de laboratorio y no es tan limitada en su alcance como los estudios
de marcado (Leporati et al., 2008).
Tres tipos de estructuras duras están presentes en los pulpos: las mandíbulas o
picos, los estatolitos (similares a los otolitos), que son pequeñas estructuras
pares calcáreas asociadas con el epitelio sensorial, que se encuentra en la
cabeza (Lombarte et al., 2006) y los estiletes, que son una pareja de estructuras
alargadas, también conocidos como concha residual o vestigial, que se
encuentra en la musculatura del manto cerca de la base de las branquias
(Bizikov 2004, Doubleday, et al., 2006).
Young (1960) observó la existencia de anillos concéntricos de estatolitos en
Octopus vulgaris. Sin embargo los estatolitos de octópodos son en su mayoría
hialinos y, a diferencia de los de calamares, no es posible observar incrementos
en toda la estructura.

16
Lombarte et al. (2006) menciona que el uso de los estatolitos, no ha dado
resultados para el pulpo debido a la ausencia de incrementos y la morfología
que estos representan.
Gonçalves (1993) examinó la presencia de incrementos en la concha vestigial
(estiletes) de Octopus vulgaris. Sousa y Fernández (2002) realizaron
observaciones en líneas de crecimiento de los estiletes de Octopus vulgaris para
determinar la edad, pero no encontraron diferencias significativas entre el
número de incrementos y la longitud del manto.
Hernández et al. (2001) analizaron 275 picos de O. vulgaris en individuos
cultivados. Indicaron que los incrementos en la zona superior de las paredes
laterales de los picos se depositan diariamente iniciando un día después de la
eclosión. Estimaron edades de entre 51 y 91 días. Sin embargo mencionan la
dificultad de las lecturas debido a la erosión de la estructura subestimando el
conteo de algunos incrementos.
Oosthuizen y Smale (2003) examinaron 320 picos de O. vugaris en condiciones
de laboratorio y campo. Concluyeron que los incrementos se depositan cada día,
presentando longevidades de entre 57 y 352 días. El ciclo de vida es de 10 a 13
meses.

17
Doubleday et al. (2006) observaron estiletes de Octopus pallidus cultivados (de
edad conocida) en Tasmania, Australia y determinaron que la periodicidad en los
incrementos del estilete se presentan diariamente. Este estudio valida con éxito
los incrementos diarios en estiletes y demuestra así un método para determinar
la edad en el pulpo. Sin embargo, fue un estudio preliminar y recomiendan
realizar más trabajos sobre estas estructuras. En particular se necesita refinar el
tipo de preservativo de estiletes y la preparación y montaje de los cortes a
observar.
Leporati et al. (2008) trabajando con la microestructura de los estiletes en pulpos
silvestres y cultivados de Octopus pallidus, menciona que no existen diferencias
en la observación de los incrementos en pulpos cultivados y silvestres,
estimando una edad máxima de 532 días, lo que sugiere una vida típica de 12
meses después de la eclosión.
Hermosilla et al. (2010) analizaron 25 estiletes de Octopus vulgaris en
condiciones controladas en el laboratorio y determinan que la tasa media de
formación de un incremento es de 1.02 por día, lo que sugiere una periodicidad
diaria de los incrementos por día.

18
1.1.6. Justificación
Sin un método preciso, fácil y confiable en cuanto a la determinación de la edad,
la estimación de tasa de crecimiento del pulpo, la productividad y la estructura
de la población, se basan entonces en suposiciones derivadas de las
evaluaciones morfológicas (como talla y peso) y de los datos de capturas
(Jackson et al., 1997, Campana 2001). Un método de aplicación común que
utiliza la longitud del manto como la medida del crecimiento, son las estructuras
de tallas que se basan en el supuesto de que la edad está estrechamente
relacionada con el tamaño (Challier et al., 2002), sin embargo, el crecimiento de
cefalópodos en general es muy variable (Forsythe y Van Heukelem, 1987;
Semmens et al., 2004), lo que hace esta suposición incongruente. Además dado
a que la esperanza de vida de las especies de pulpo en general es corta (<2
años) (Boyle y Rodhouse, 2005) y a que este tipo de técnicas indirectas pueden
producir tasas de crecimiento y longevidad inexactas.
Por ello es necesario un estudio integral en cuanto a recursos pesqueros con
importancia biológica, comercial y social. El reciente incremento de capturas de
Octopus maya en Campeche, y la subsiguiente competencia por el recurso,
ameritan su investigación en este Estado. Los volúmenes de captura se han
triplicado durante la última década, aún después de haberse reconocido el
riesgo de sobreexplotación por Solís (1996).

19
Se desconoce la biología de los juveniles y en general la biología del recurso en
temporada de veda. Los estudios sobre crecimiento, longevidad y reclutamiento
con objeto de manejo del pulpo rojo se basan en el análisis modal de tallas
(Solís, 1986) aunque su uso en cefalópodos ha sido ampliamente criticado y son
preferibles los métodos de estimación directas (Semmens et al., 2004), como la
lectura de incrementos periódicos en estructuras duras.
Los estudios de crecimiento de pulpo usando incrementos diarios en estructuras
duras como picos no han sido concluyentes, debido al desgaste de los
incrementos que presenta esta estructura, al momento de alimentarse el
organismo (Hernández et al., 2001). No obstante la reciente validez en la lectura
diaria de los incrementos en estiletes en Octopus pallidus (Doubleday et al.,
2006) es de gran relevancia, debido a que es una estructura interna que no
presenta deterioro externo y podrán ser utilizadas en estudios futuros, con el fin
de estimar el crecimiento de pulpos silvestres.
La importancia de la información obtenida en este trabajo será esencial para
conocer la estructura de edad de la población de las pesquería de pulpo, para
complementar sesgos en muestreos indirectos, como la utilización de las
frecuencias en tallas (Caddy, 1991). Este trabajo tiene como objetivo aportar la
información biológica necesaria sobre la edad y crecimiento de Octopus maya
para contribuir a un mayor conocimiento de este importante recurso pesquero.

20
2. OBJETIVOS
2.1. Objetivo general
· Evaluar la estructura del estilete en el pulpo Octopus maya para su
uso en estudios de edad y crecimiento.
2.2. Objetivos particulares
· Determinar si los estiletes se desarrollan proporcionalmente al
crecimiento somático de O. maya.
· Aplicar una metodología reproducible y efectiva para examinar los
incrementos en los estiletes de O. maya.
· Estimar la edad del pulpo mediante el análisis de los incrementos en
los estiletes, partiendo del argumento: un incremento es igual a un día.
· Estimar los parámetros de crecimiento mediante la aplicación de los
modelos de regresión que mejor describan este proceso.
3. HIPÓTESIS
Existe una relación entre el número de los incrementos de los estiletes del pulpo
Octopus maya y las dimensiones (longitud de manto y peso) del animal.

21
4. ÁREA DE ESTUDIO
El Banco de Campeche esta representado por la plataforma continental
aproximadamente entre los 18° y 24° de latitud Norte y los 87° y 93° de longitud
Oeste (Fig. 2). Este sistema es amplio y su plataforma se extiende
significativamente alcanzando 150 km, se modifica conforme se incrementa su
profundidad, pero en general la superficie es convexa, configurándose en un
área de aproximadamente 90 000 km2 desde la línea de playa hasta los 200 m
de profundidad (Antoine y Gilmore, 1970).
La plataforma continental presenta un gradiente de profundidad de 0.3 a 1 m por
km con sedimentos formados por proporciones variables de fango, arena y
conchas. Se caracteriza por la abundancia de bancos y arrecifes coralinos
(García, 1980). De acuerdo al tipo de fondo el Banco puede ser dividido en dos
regiones: la primera se extiende desde el estrecho de Yucatán hasta el norte de
Cayo Triángulo, se caracteriza por el predominio de fondos coralinos y arenosos,
encontrándose el fango en zonas más profundas; la segunda se extiende hacia
el suroeste y predominan los fondos fangosos (García y Gómez, 1974).
El clima es cálido subhúmedo con lluvias en verano y temperatura máxima de
36°C y una mínima de 17°C. La precipitación pluvial alcanza valores de entre
1200 y 200 mm anuales y el periodo de mayor precipitación es de junio a
noviembre. Los vientos dominantes presentan dirección noreste y sureste

22
(Botello, 1978). Existen tres épocas climáticas marcadas: a) el período de secas
que abarca de febrero a mayo, b) período de lluvias de junio a octubre y c)
período de nortes que comprende de octubre a febrero. La temperatura
promedio mensual del aire varía entre los 24.4°C en febrero a 28.0° C en agosto.
La dinámica de las aguas muestra de forma general que el movimiento de las
corrientes es de Este a Oeste en la zona norte y de Norte a Sur en la zona
suroeste, siguiendo la configuración del Banco (García, 1980).
Figura 2. Ubicación del Banco de Campeche en el Golfo de México (Tomado de Jiménez, 2005).
24
20
18
22
93° 91° 89 87
100° 90° 80°
30°
20°
10°

23
En 1968 se hace referencia a dos tipos de circulación o tendencias dinámicas
sobre el Banco, causadas por cambios en la intensidad de la Corriente de
Yucatán que vierte aguas de menor temperatura sobre el Banco (Bessonov y
González, 1967). El alcance de esta masa de agua de menor temperatura esta
directamente relacionada con la fluctuación del borde oeste de la Corriente de
Yucatán, la cual durante el cambio de posición de este a oeste lleva al Banco, en
forma de pulso, una parte de la masa de agua fría de alta mar (Bulanienkov y
García, 1973). De esta manera, la intensidad del afloramiento dependerá del
grado de proximidad del borde izquierdo de dicha corriente con el talud, este
afloramiento favorece la producción primaria por cuanto vierte sobre la
plataforma los nutrientes provenientes del talud continental (Bulanienkov y
García, 1973).
Piñeiro y Giménez (2001) señalan que la temperatura de las aguas superficiales
del Banco está muy relacionada con la radiación solar, con el período del año y
posee una estrecha relación con la temperatura del aire, por lo que en la época
de verano e inicio de otoño (julio - septiembre) se observan los valores más
elevados de temperatura, variando entre 28°C y 29.5°C, siendo agosto, el mes
más cálido. Los valores mínimos se observan en invierno y principios de
primavera (diciembre - marzo) con cifras entre 25°C y 23.5°C. En las aguas del
fondo, la temperatura muestra un patrón diferente. Los valores mínimos
corresponden con la primavera - verano, y el mes de julio es el de menor valor
con 20.5°C.

24
5. MATERIALES Y MÉTODOS
5.1. Trabajo de campo
Las muestras fueron recolectadas en cinco cooperativas de producción de pulpo,
resultado de la pesca artesanal en los principales puertos de desembarque
pesqueros: Sabancuy, Champotón, Seybaplaya, Campeche e Isla arena (Fig. 3)
durante los meses de agosto a diciembre del 2007.
Figura 3. Localización de las zonas de muestreo
Estas cooperativas fueron elegidas debido a que se dedican casi exclusivamente
a la pesca de pulpo en temporada, tomando como criterios la accesibilidad,
logística, operatividad y volumen de acopio. De 300 organismos desembarcados
por localidad se midió la longitud dorsal del manto (LM) distancia en cm, medida
desde el ápice posterior del mismo hasta una línea interocular imaginaria, con el
ejemplar extendido sobre una superficie plana, mediante un vernier (precisión

25
0.01 mm). El peso total (kg) con una balanza digital OHAUS de 6000 g de
capacidad (precisión 1 g). El sexo y el grado de madurez sexual se
determinaron mediante la observación del desarrollo del aparato reproductor
(gónadas y glándulas accesorias). Se consideraron cuatro estadios de madurez
sexual por sexo: inmaduros, madurando, maduros y desovados.
De 100 individuos se extrajeron los estiletes de la zona dorsal del manto, cerca
de la inserción de las branquias (Fig. 4) con ayuda de unas pinzas y se
colocaron en bolsas de plástico debidamente etiquetadas, conservándolas en
formol al 4%.
Figura 4. Localización del estilete en la zona corporal interna de O. maya
(Modificado de Bizikov, 2004).
Estilete
Corazón
Sifón
Músculo retractordel sifón
Branquia

26
5.2. Trabajo de laboratorio
5.2.1. Preparación y montaje de las secciones del estilete
Los estiletes fueron lavados en agua dulce para quitar el formol, con el objetivo
de evitar la deshidratación. Para realizar el corte se seleccionó la región
posterior del estilete (Fig. 5) debido a que es una zona que presenta mejor
visibilidad de incrementos y menos daño al momento de extraer la estructura. La
zona central del estilete es la primera en formarse por lo tanto contiene todos los
incrementos.
Figura 5. Morfometría del estilete (Modificado de Bizikov, 2004).
Figura 5. Morfometría del estilete (Modificado de Bizikov, 2004).
Posteriormente se midió la longitud de cada estilete de la zona anterior hasta la
zona posterior y el diámetro máximo, usando un vernier digital (precisión de 0.01
mm) con el objetivo de determinar, si es proporcional al crecimiento del
organismo. Se pesó el estilete con una balanza digital OHAUS de 400 g de
capacidad (precisión de 0.01 g).
Se realizaron cortes transversales del estilete con un grosor de 0.1-0.2 mm. Para
el hacer el corte se utilizaron dos pequeños bloques de poliestireno, con
Zona anterior
Codo
Zona posterior
Región de análisisde incrementos

27
dimensiones correspondientes al tamaño del estilete de 1.0 cm × 1.5 cm × 5.0
cm. Posteriormente se colocó la estructura dentro de los dos bloques, de tal
manera que su eje longitudinal estuviera alineado para evitar rupturas. Los
cortes se realizaron con una navaja de afeitar marca Croma con medidas de 4.3
cm de largo y 2.2 cm de ancho, efectuando movimientos oblicuos lentamente
sobre la superficie de los bloques hasta llegar al estilete (Bizikov, 1996) (Fig. 6).
Figura 6. Técnica utilizada para el corte en el estilete (Modificado de Bizikov, 1996).
Después de cortar varias muestras (aproximadamente 30 secciones) se
recogieron las secciones con un pincel y se colocaron rápidamente en una caja
de petri en agua dulce para evitar la deshidratación.
El medio de montaje que se utilizó para muestras permanentes, fue la gelatina
de glicerol (Bizikov, 1996). Y se preparó disolviendo las siguientes sustancias a
37 °C con ayuda de una parrilla eléctrica: 100 ml de agua destilada, 100 g de
glicerina, 17 g de gelatina pura y 1 g o 1.0 ml de fenol (fungicida para preservar
Navaja
Estilete
Bloques de poliestireno

28
la preparación). Cuando se presentó una apariencia pastosa o gomosa,
rápidamente se retiró del calor y se dejó secar durante una hora. Posteriormente
se conservó en un frasco de vidrio, lejos de la luz solar directa y se mantuvo a
temperatura ambiente.
Las secciones del estilete se montaron en un portaobjetos de vidrio, utilizando
una gota de glicerol para montajes temporales. En montaje permanente se
aplicó un pedazo de gelatina de glicerol en un portaobjetos con ayuda de unas
pinzas de relojero y se calentó con un encendedor, esperando hasta que la
gelatina se convierta en una gota o se funda evitando que hierva.
Posteriormente se introdujo una sección transversal del estilete en la gota y se
colocó un cubreobjetos en la muestra evitando burbujas de aire.
5.2.2. Lectura de los incrementos
Los cortes de los estiletes se observaron, por medio de un microscopio ocular
marca Zeizz a 400 x aumentos. Los incrementos en los estiletes se contaron con
un contador de mano marca Upgreen. Se contaron los incrementos de primer
orden identificados como una estructura bipartita, compuesta por un incremento
oscuro y un incremento claro con un patrón regular empaquetados dentro de los
incrementos de segundo orden, el cual presentaban amplias bandas con límites
difusos e irregulares (Bizikov, 2004; Doubleday et al., 2006; Márquez y Ré,
2009). El primer incremento se definió como el primer anillo visible después del

29
núcleo embrionario y fue considerado como la marca de nacimiento. El último
anillo fue definido como la última circunferencia regular visible en la sección
(Doubleday et al., 2006; Leporati et al., 2008 y Márquez y Ré, 2009) (Fig. 6). El
número de incrementos presentes en cada sección del estilete representan la
edad en días, esto es, “un incremento igual a un día” (Doubleday et al., 2006).
5.3. Análisis de datos
5.3.1. Estructura de talla y peso
La estructura de tallas se obtuvo agrupando los datos de longitud de manto (LM)
en intervalos de 5 mm y el peso total (PT) con intervalos de 100 g, construyendo
histogramas de frecuencia ordenados por sexo. La estructura de talla y peso
para hembras y machos fueron comparadas a través de una prueba no
paramétrica (Kolmogorov-Smirnov) con un nivel de confianza del 95%, con la
finalidad de probar si la distribución de hembras fue de diferente a la de los
machos.
5.3.2. Relación entre dimensiones del estilete y dimensiones del pulpo rojo
Para establecer la relación entre el diámetro máximo y longitud total del estilete
(DME y LTE) con la longitud de manto y peso (LM y PT) del pulpo Octopus
maya, se realizó una regresión lineal entre estas variables para hembras y
machos. Con el objetivo de determinar si la estructura crece proporcionalmente
a la longitud y peso del organismo. Los coeficientes de la regresión se

30
obtuvieron a través del método de mínimos cuadrados.
5.3.3. Estimación de la edad
La edad se estimó por medio de la lectura y conteo de los incrementos de
crecimiento observados por dos lectores de manera independiente, con objeto
de comparar los resultados para determinar la reproducibilidad del
procedimiento. El máximo error entre las lecturas fue del 10% y se usó el
promedio entre ambas para la estimación de la edad. Las lecturas fueron
efectuadas en tres ocasiones para reducir el sesgo en los conteos y se
realizaron a partir de la marca de nacimiento identificada como la primera
circunferencia después del núcleo.
5.3.4. Precisión de las lecturas
Como una medida de precisión y reproducibilidad de las lecturas, se realizó el
método de Porcentaje de Acuerdo (Campana, 2001) mediante la siguiente
fórmula: PA = (LI / TL)*100
Donde:
LI = Número de lecturas iguales
TL= Total de lecturas realizadas
Para comprobar la precisión de cada lector, se calculó el Porcentaje de Error
Promedio (APE) (Beamish y Fournier, 1981), el coeficiente variación (CV) y el

31
índice de precisión (D) (Chang, 1982) de la siguiente manera:
Donde:
R = Número de lecturas realizadas
Xij = Edad estimada para un organismo en cada una de las lecturas
Xj = Edad promedio estimada para el mismo organismo
(Campana, 2001).
Los índices APE, CV, D proporcionan una medida directa de la consistencia en
el conteo y entre más pequeño el valor de cada índice existe mayor
reproducibilidad y precisión en las lecturas.
R (│Xij-Xj│)/Xj
APEj = 100% x 1/R Σ i=1 Xj
R (Xij-Xj)2
∑ CVj = 100% x i=1 R-1 Xj
CVjDj =
√ R

32
5.3.5. Patrón del crecimiento
El patrón de crecimiento se describió a partir del ajuste de diferentes modelos a
los datos de edad-longitud de manto y edad-peso, seleccionando el modelo de
acuerdo al mayor coeficiente de determinación (r2) y menor coeficiente de
varianza (CV%) de los parámetros estimados.
El modelo seleccionado se utilizó para estimar la talla a la edad y peso a la edad
para machos y hembras, además de calcular las tasas de crecimiento. La
estimación de los parámetros de los modelos se realizó utilizando el software
STATISTICA 7.0 (STATSOFT®, 2007) aplicando el método iterativo no lineal por
medio de mínimos cuadrados. Los datos se ajustaron a los siguientes modelos:
Lineal: L = a+b*X
Potencial: L= a*Xb
Exponencial: L= a*ebt
Gompertz de crecimiento Lt= LBB0BB*exp(-exp(-k*(t-to)))
Von Bertalanffy Lt= L∞ *(1-exp(K*(t-to)))
Logística integral Lt =L∞/ (1+exp(-k*(t-to)))
Donde LBBtBB es la longitud del manto a una edad dada; t es la edad en días; LBB0BB es
la longitud de manto o peso cuando t es igual a 0; k es un parámetro
adimensional; c es la tasa especifica de crecimiento cuando t = tBBoBB; LBB∞BB longitud

33
asintótica (longitud máxima teórica); K tasa de crecimiento anual o parámetro de
curvatura; y tBBoBB es la edad a la longitud de manto o peso cero.
Para evaluar las diferencias en las curvas de crecimiento entre sexos se utilizó
una prueba propuesta por Chen et al. (1992), que básicamente analiza la suma
de cuadrados residuales (RSS) que resultan del proceso de ajuste no lineal de
los parámetros de crecimiento. La formula de la prueba (Ratkowsky, 1983) es:
RSSp - RSSs
3 (K – 1)
RSSs
(N – 3) K
F =
Donde:
N= número total de muestras
K= número de curvas comparadas
RSSs= suma de cuadrados residuales de ambos grupos (machos más hembras)
RSSp= suma de cuadrados residuales del ajuste agrupado (sin diferenciar por
sexo).
Para cada sexo se cálculo la tasa diaria de crecimiento (TDC, mm o g por día) y
la tasa instantánea de crecimiento (G x 100). Se calcularon los valores de
longitud de manto (LM) y peso en intervalos de 10 días utilizando los parámetros
de las curvas seleccionadas (Forsythe y Van Heukelem, 1987):

34
TCD = (S2 – S1)/ΔT
G = (lnS2 – lnS1)/ΔT
Donde: S1 y S2 son los tamaños calculados (LM en mm o peso en g) al inicio y
al final de cada intervalo de tiempo (ΔT=10 días).
Por medio del retrocálculo se estimaron las fechas de nacimiento de cada
individuo, sustrayendo la edad asignada mediante el conteo de incrementos en
el estilete a la fecha de captura.

35
6. RESULTADOS
6.1. Estructura de talla y peso
Durante este estudio se seleccionaron 100 individuos de distintos tamaños para
la preparación de los estiletes, de los cuales 62 correspondieron a hembras y 38
a machos (Tabla 1). En general la longitud de manto (LM) osciló entre 67 y 152
mm, con un promedio de 104.9 mm (± 20.92 DE); el peso total (PT) fluctuó de
185 a 2315 g, con un promedio de 640.87 g (± 405.82 DE).
Tabla 1. Número de organismos de O. maya seleccionados para la preparacióndel estilete. (H= hembras, M= machos).
Para las hembras la longitud de manto (LM) fluctuó entre 67 y 152 mm, con un
valor medio de 105.93 mm (±20.63 DE) y una moda en 90 mm; en machos varió
entre 69 y 151 mm, con un valor medio de 103.21 mm (±21.56 DE) con una
MesesAgosto Septiembre Octubre Noviembre Diciembre Total
PuertosH M H M H M H M H M
Campeche 5 2 2 3 2 1 - - - -
Champotón - - 8 5 3 5 11 5 2 1
Isla Arena 7 3 2 1 - - 2 1 - -
Sabancuy 6 6 - - - - 2 - 1 -
Seybaplaya 5 4 - - - - - - 4 1
15
40
16
15
14
Total 38 21 11 21 9 100
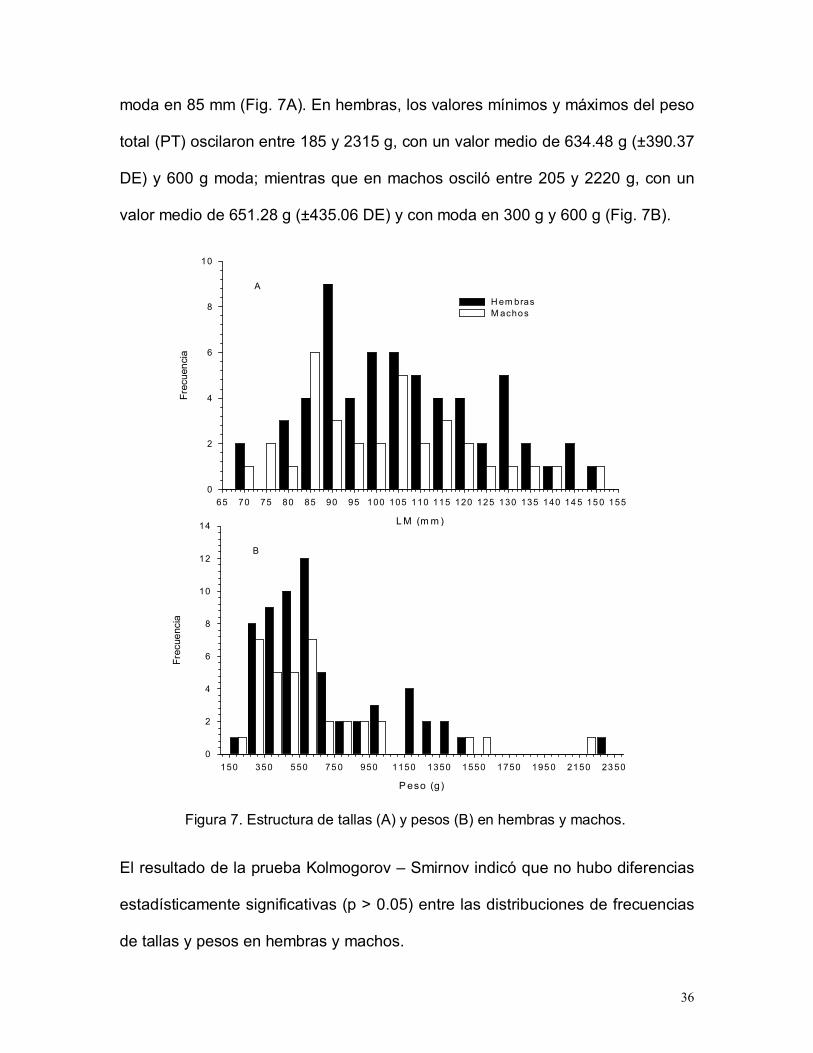
36
moda en 85 mm (Fig. 7A). En hembras, los valores mínimos y máximos del peso
total (PT) oscilaron entre 185 y 2315 g, con un valor medio de 634.48 g (±390.37
DE) y 600 g moda; mientras que en machos osciló entre 205 y 2220 g, con un
valor medio de 651.28 g (±435.06 DE) y con moda en 300 g y 600 g (Fig. 7B).
A
L M (m m )
65 70 75 80 85 90 95 100 105 110 115 120 125 130 135 140 14 5 15 0 155
Frec
uenc
ia
0
2
4
6
8
10
H em brasM achos
P eso (g)
150 350 550 75 0 950 1150 1350 1550 1750 195 0 2150 23 50
Frec
uenc
ia
0
2
4
6
8
10
12
14
B
Figura 7. Estructura de tallas (A) y pesos (B) en hembras y machos.
El resultado de la prueba Kolmogorov – Smirnov indicó que no hubo diferencias
estadísticamente significativas (p > 0.05) entre las distribuciones de frecuencias
de tallas y pesos en hembras y machos.
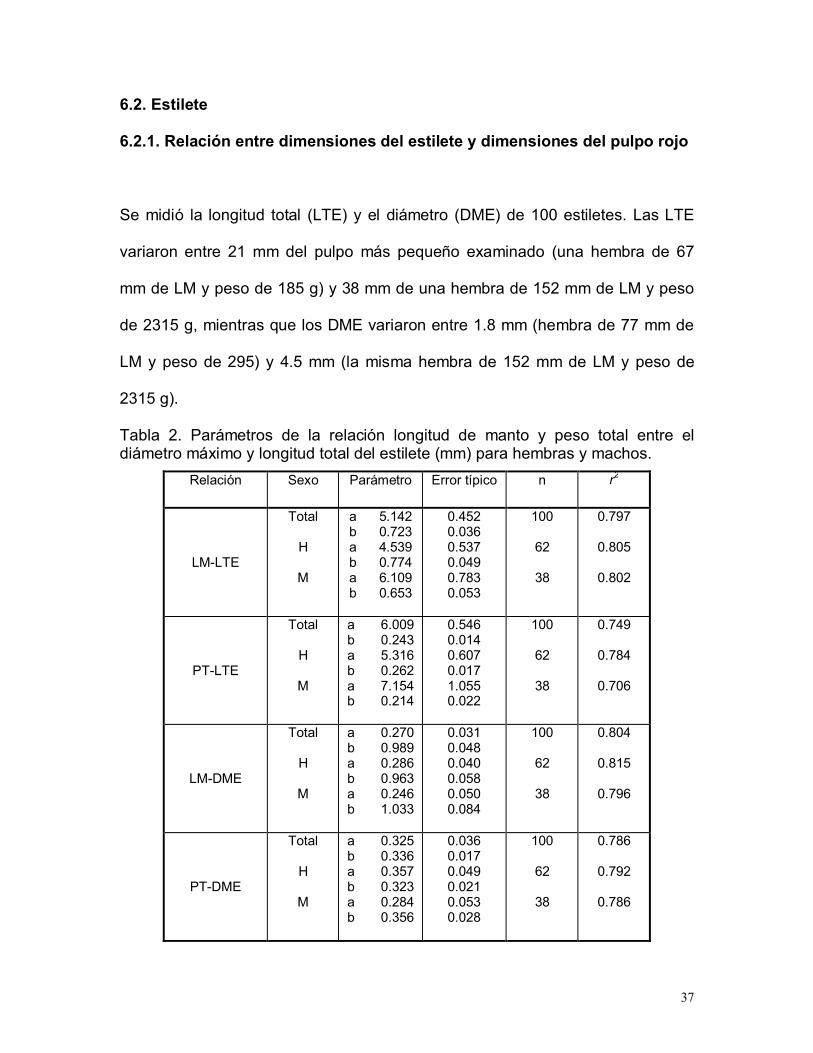
37
6.2. Estilete
6.2.1. Relación entre dimensiones del estilete y dimensiones del pulpo rojo
Se midió la longitud total (LTE) y el diámetro (DME) de 100 estiletes. Las LTE
variaron entre 21 mm del pulpo más pequeño examinado (una hembra de 67
mm de LM y peso de 185 g) y 38 mm de una hembra de 152 mm de LM y peso
de 2315 g, mientras que los DME variaron entre 1.8 mm (hembra de 77 mm de
LM y peso de 295) y 4.5 mm (la misma hembra de 152 mm de LM y peso de
2315 g).
Tabla 2. Parámetros de la relación longitud de manto y peso total entre eldiámetro máximo y longitud total del estilete (mm) para hembras y machos.
Relación Sexo Parámetro Error típico n r2
LM-LTE
Total
H
M
a 5.142b 0.723a 4.539b 0.774a 6.109b 0.653
0.4520.0360.5370.0490.7830.053
100
62
38
0.797
0.805
0.802
PT-LTE
Total
H
M
a 6.009b 0.243a 5.316b 0.262a 7.154b 0.214
0.5460.0140.6070.0171.0550.022
100
62
38
0.749
0.784
0.706
LM-DME
Total
H
M
a 0.270b 0.989a 0.286b 0.963a 0.246b 1.033
0.0310.0480.0400.0580.0500.084
100
62
38
0.804
0.815
0.796
PT-DME
Total
H
M
a 0.325b 0.336a 0.357b 0.323a 0.284b 0.356
0.0360.0170.0490.0210.0530.028
100
62
38
0.786
0.792
0.786
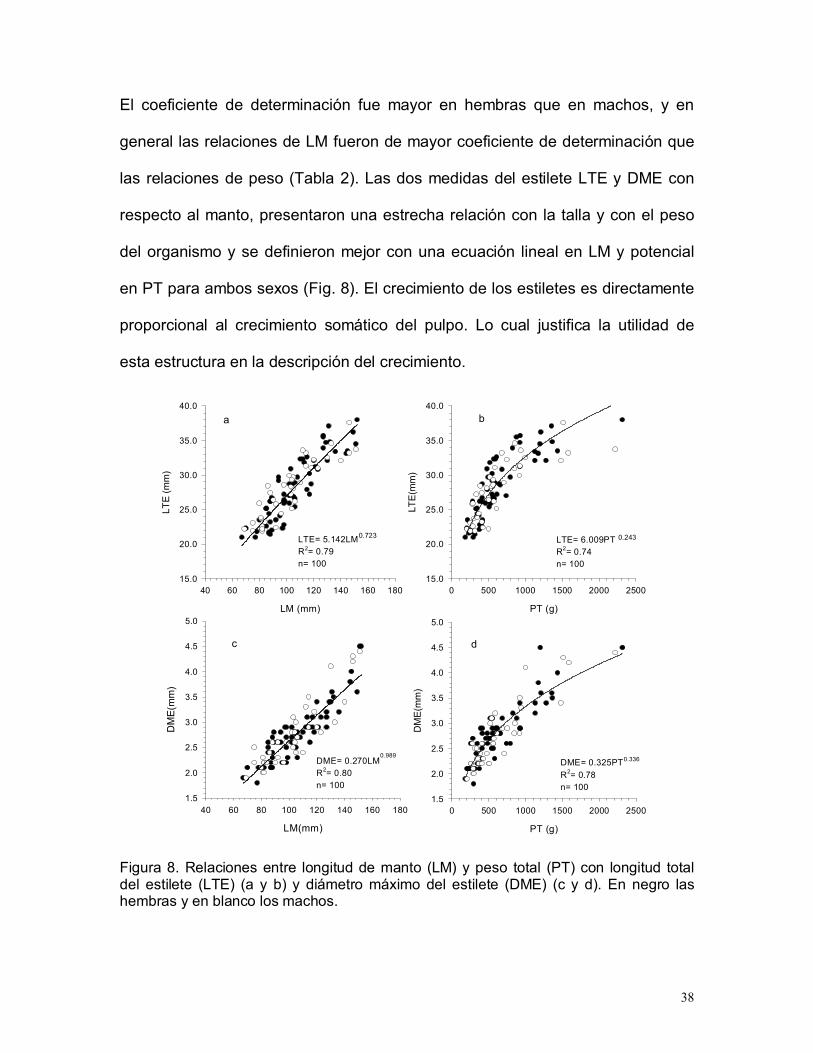
38
El coeficiente de determinación fue mayor en hembras que en machos, y en
general las relaciones de LM fueron de mayor coeficiente de determinación que
las relaciones de peso (Tabla 2). Las dos medidas del estilete LTE y DME con
respecto al manto, presentaron una estrecha relación con la talla y con el peso
del organismo y se definieron mejor con una ecuación lineal en LM y potencial
en PT para ambos sexos (Fig. 8). El crecimiento de los estiletes es directamente
proporcional al crecimiento somático del pulpo. Lo cual justifica la utilidad de
esta estructura en la descripción del crecimiento.
LM (mm)
40 60 80 100 120 140 160 180
LTE
(mm
)
15.0
20.0
25.0
30.0
35.0
40.0
PT (g)
0 500 1000 1500 2000 2500
LTE(
mm
)
15.0
20.0
25.0
30.0
35.0
40.0
PT (g)
0 500 1000 1500 2000 2500
DM
E(m
m)
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
LTE= 5.142LMR2= 0.79n= 100
0.723
a
LTE= 6.009PTR2= 0.74n= 100
b
0.243
d
DME= 0.325PTR2= 0.78n= 100
0.336
LM(mm)
40 60 80 100 120 140 160 180
DM
E(m
m)
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
DME= 0.270LMR2= 0.80n= 100
0.989
c
Figura 8. Relaciones entre longitud de manto (LM) y peso total (PT) con longitud totaldel estilete (LTE) (a y b) y diámetro máximo del estilete (DME) (c y d). En negro lashembras y en blanco los machos.
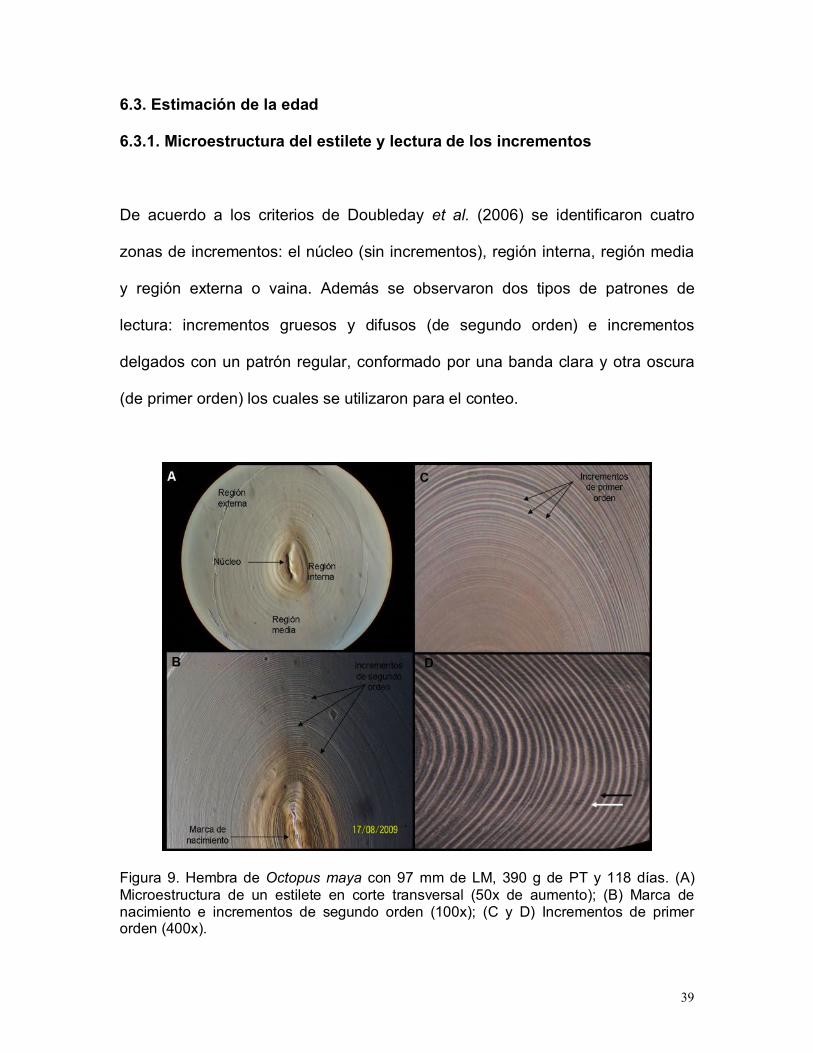
39
6.3. Estimación de la edad
6.3.1. Microestructura del estilete y lectura de los incrementos
De acuerdo a los criterios de Doubleday et al. (2006) se identificaron cuatro
zonas de incrementos: el núcleo (sin incrementos), región interna, región media
y región externa o vaina. Además se observaron dos tipos de patrones de
lectura: incrementos gruesos y difusos (de segundo orden) e incrementos
delgados con un patrón regular, conformado por una banda clara y otra oscura
(de primer orden) los cuales se utilizaron para el conteo.
Figura 9. Hembra de Octopus maya con 97 mm de LM, 390 g de PT y 118 días. (A)Microestructura de un estilete en corte transversal (50x de aumento); (B) Marca denacimiento e incrementos de segundo orden (100x); (C y D) Incrementos de primerorden (400x).

40
La marca de nacimiento se consideró como el primer incremento visible (uno
claro y otro oscuro), después del núcleo embrionario que suele ser amorfo y
opaco (Fig. 9). A partir de este incremento, los siguientes incrementos fueron
enumerados consecutivamente y considerados para conformar la estimación de
la edad.
6.4. Precisión de las lecturas
El porcentaje de acuerdo (PA) se evaluó entre dos lectores, encontrándose un
50% de lecturas iguales, así como un 94% de acuerdo con una diferencia de ±10
anillos de crecimiento y el 100% con ±20 anillos (Tabla 3).
Tabla 3. Porcentaje de acuerdo entre lectores.
Diferencia de anillos Frecuencia Anillos asignados PA
-30 0
-20 0
-10 7
-0 50 ±10 94
10 37 ±20 100
20 6
30 0
41
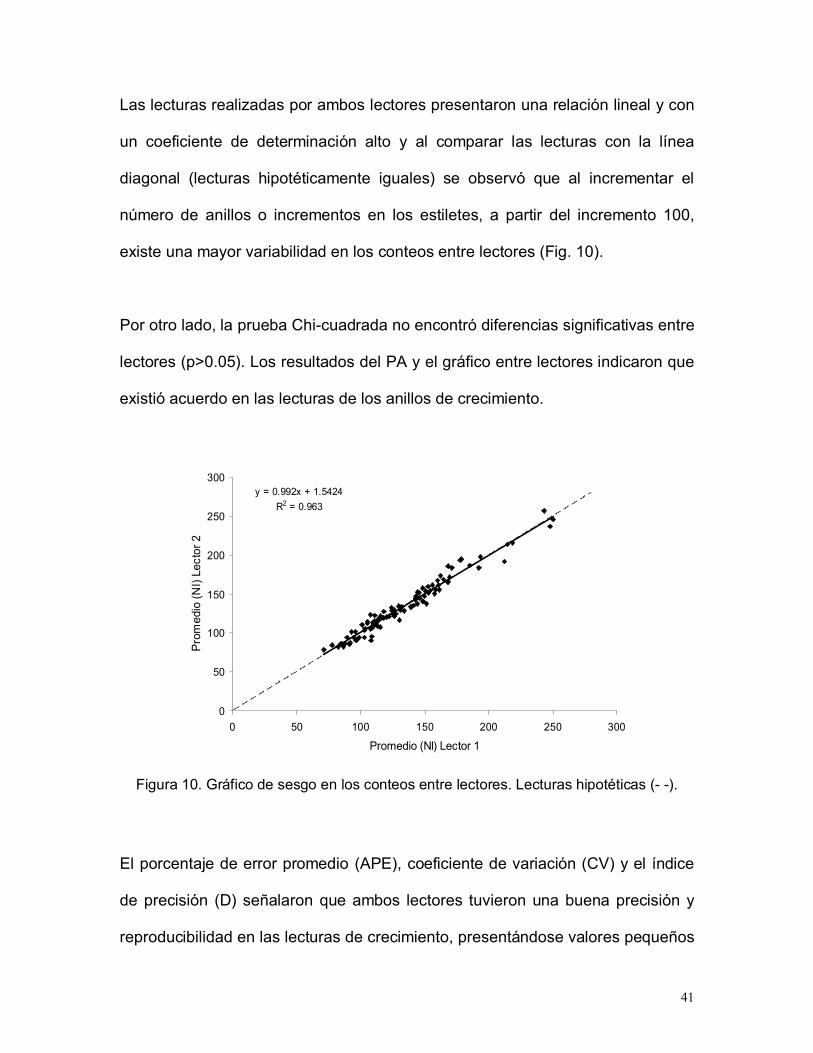
Las lecturas realizadas por ambos lectores presentaron una relación lineal y con
un coeficiente de determinación alto y al comparar las lecturas con la línea
diagonal (lecturas hipotéticamente iguales) se observó que al incrementar el
número de anillos o incrementos en los estiletes, a partir del incremento 100,
existe una mayor variabilidad en los conteos entre lectores (Fig. 10).
Por otro lado, la prueba Chi-cuadrada no encontró diferencias significativas entre
lectores (p>0.05). Los resultados del PA y el gráfico entre lectores indicaron que
existió acuerdo en las lecturas de los anillos de crecimiento.
y = 0.992x + 1.5424R2 = 0.963
0
50
100
150
200
250
300
0 50 100 150 200 250 300
Promedio (NI) Lector 1
Prom
edio
(NI)
Lect
or 2
Figura 10. Gráfico de sesgo en los conteos entre lectores. Lecturas hipotéticas (- -).
El porcentaje de error promedio (APE), coeficiente de variación (CV) y el índice
de precisión (D) señalaron que ambos lectores tuvieron una buena precisión y
reproducibilidad en las lecturas de crecimiento, presentándose valores pequeños

42
en los tres parámetros APE, CV y D (Tabla 6). De igual forma, se puede
observar que el lector 2 tuvo mayor precisión, que el lector 1.
Tabla 6. Resultados de las pruebas de precisión y reproducibilidad en losconteos de incrementos por lector.
6.5. Estimación del crecimiento
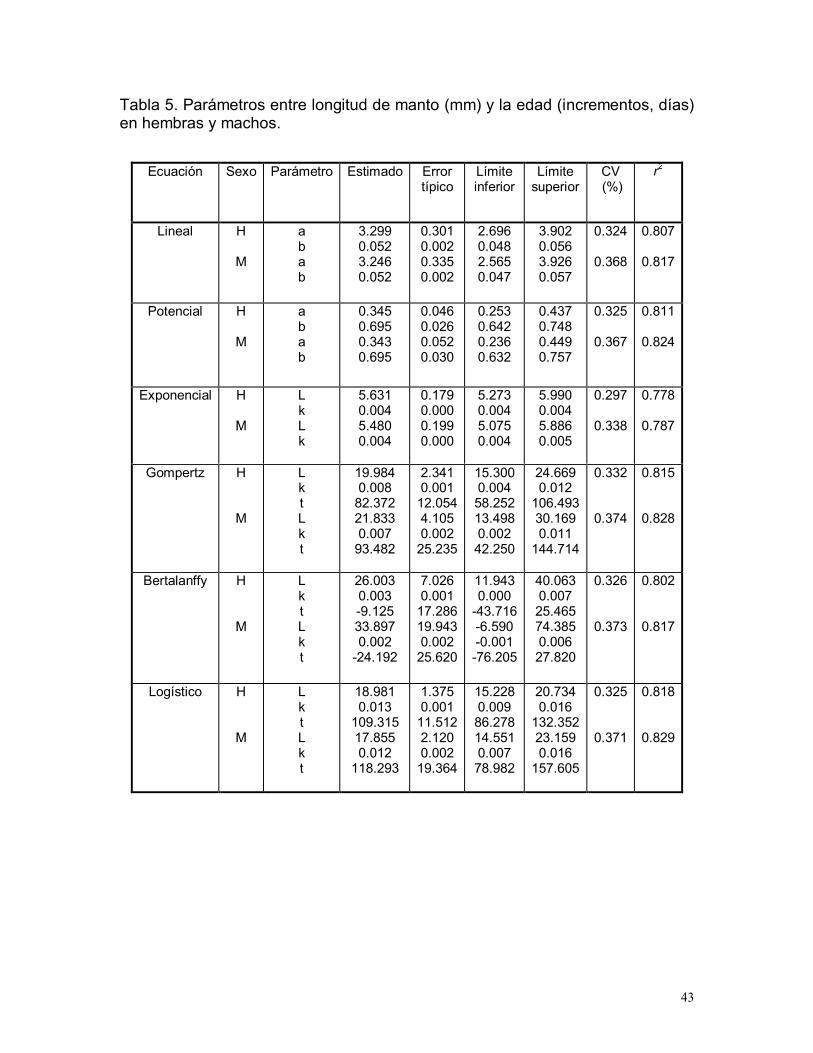
6.5.1. Relación longitud de manto-edad y peso total-edad
Los modelos de crecimiento de longitud de manto (LM) y peso total (PT) que
mejor se ajustaron con los datos de edad, fueron los modelos de Gompertz y
logístico demostrando los mayores coeficientes de determinación. El modelo
logístico presentó el mayor coeficiente de determinación (r2= 0.82) el menor
intervalo de confianza, por lo que se eligió este modelo para describir el
crecimiento del pulpo rojo (Tabla 5 y 6). Las curvas teóricas de crecimiento del
modelo logístico, obtenidas a partir de los incrementos en estiletes se presentan
en la figura 11.
Lector No. Muestrasobservadas
No.Observaciones
por muestra
APE CV D
1
2
100
100
3
3
2.79
2.68
0.02
0.02
0.01
0.01
43
Tabla 5. Parámetros entre longitud de manto (mm) y la edad (incrementos, días)en hembras y machos.
Ecuación Sexo Parámetro Estimado Errortípico
Límiteinferior
Límitesuperior
CV (%)
r2
Lineal H
M
abab
3.2990.0523.2460.052
0.3010.0020.3350.002
2.6960.0482.5650.047
3.9020.0563.9260.057
0.324
0.368
0.807
0.817
Potencial H
M
abab
0.3450.6950.3430.695
0.0460.0260.0520.030
0.2530.6420.2360.632
0.4370.7480.4490.757
0.325
0.367
0.811
0.824
Exponencial H
M
LkLk
5.6310.0045.4800.004
0.1790.0000.1990.000
5.2730.0045.0750.004
5.9900.0045.8860.005
0.297
0.338
0.778
0.787
Gompertz H
M
LktLkt
19.9840.008
82.37221.8330.007
93.482
2.3410.001
12.0544.1050.002
25.235
15.3000.00458.25213.4980.00242.250
24.6690.012
106.49330.1690.011
144.714
0.332
0.374
0.815
0.828
Bertalanffy H
M
LktLkt
26.0030.003-9.12533.8970.002
-24.192
7.0260.001
17.28619.9430.002
25.620
11.9430.000
-43.716-6.590-0.001-76.205
40.0630.00725.46574.3850.00627.820
0.326
0.373
0.802
0.817
Logístico H
M
LktLkt
18.9810.013
109.31517.8550.012
118.293
1.3750.001
11.5122.1200.002
19.364
15.2280.00986.27814.5510.00778.982
20.7340.016
132.35223.1590.016
157.605
0.325
0.371
0.818
0.829
44
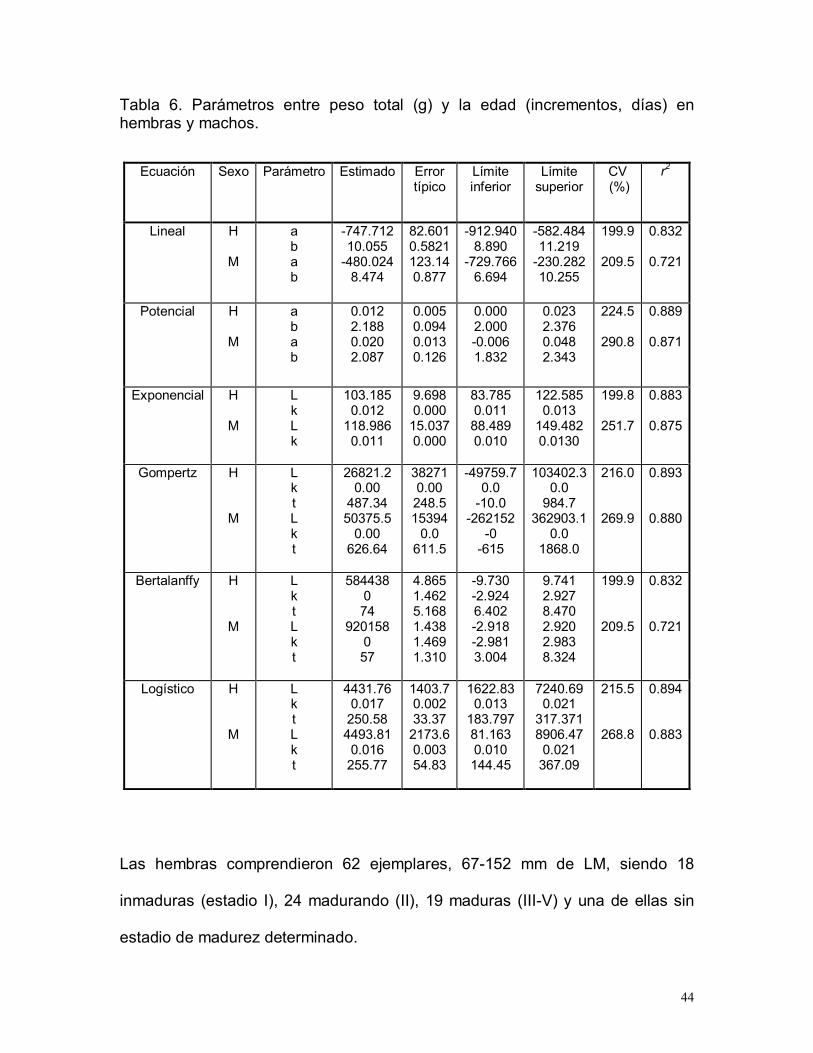
Tabla 6. Parámetros entre peso total (g) y la edad (incrementos, días) enhembras y machos.
Las hembras comprendieron 62 ejemplares, 67-152 mm de LM, siendo 18
inmaduras (estadio I), 24 madurando (II), 19 maduras (III-V) y una de ellas sin
estadio de madurez determinado.
Ecuación Sexo Parámetro Estimado Errortípico
Límiteinferior
Límitesuperior
CV (%)
r2
Lineal H
M
abab
-747.71210.055
-480.0248.474
82.6010.5821123.140.877
-912.9408.890
-729.7666.694
-582.48411.219
-230.28210.255
199.9
209.5
0.832
0.721
Potencial H
M
abab
0.0122.1880.0202.087
0.0050.0940.0130.126
0.0002.000-0.0061.832
0.0232.3760.0482.343
224.5
290.8
0.889
0.871
Exponencial H
M
LkLk
103.1850.012
118.9860.011
9.6980.000
15.0370.000
83.7850.01188.4890.010
122.5850.013
149.4820.0130
199.8
251.7
0.883
0.875
Gompertz H
M
LktLkt
26821.20.00
487.3450375.5
0.00626.64
382710.00248.515394
0.0611.5
-49759.70.0
-10.0-262152
-0-615
103402.30.0
984.7362903.1
0.01868.0
216.0
269.9
0.893
0.880
Bertalanffy H
M
LktLkt
584438074
920158057
4.8651.4625.1681.4381.4691.310
-9.730-2.9246.402-2.918-2.9813.004
9.7412.9278.4702.9202.9838.324
199.9
209.5
0.832
0.721
Logístico H
M
LktLkt
4431.760.017
250.584493.810.016
255.77
1403.70.00233.37
2173.60.00354.83
1622.830.013
183.79781.1630.010144.45
7240.690.021
317.3718906.470.021
367.09
215.5
268.8
0.894
0.883

45
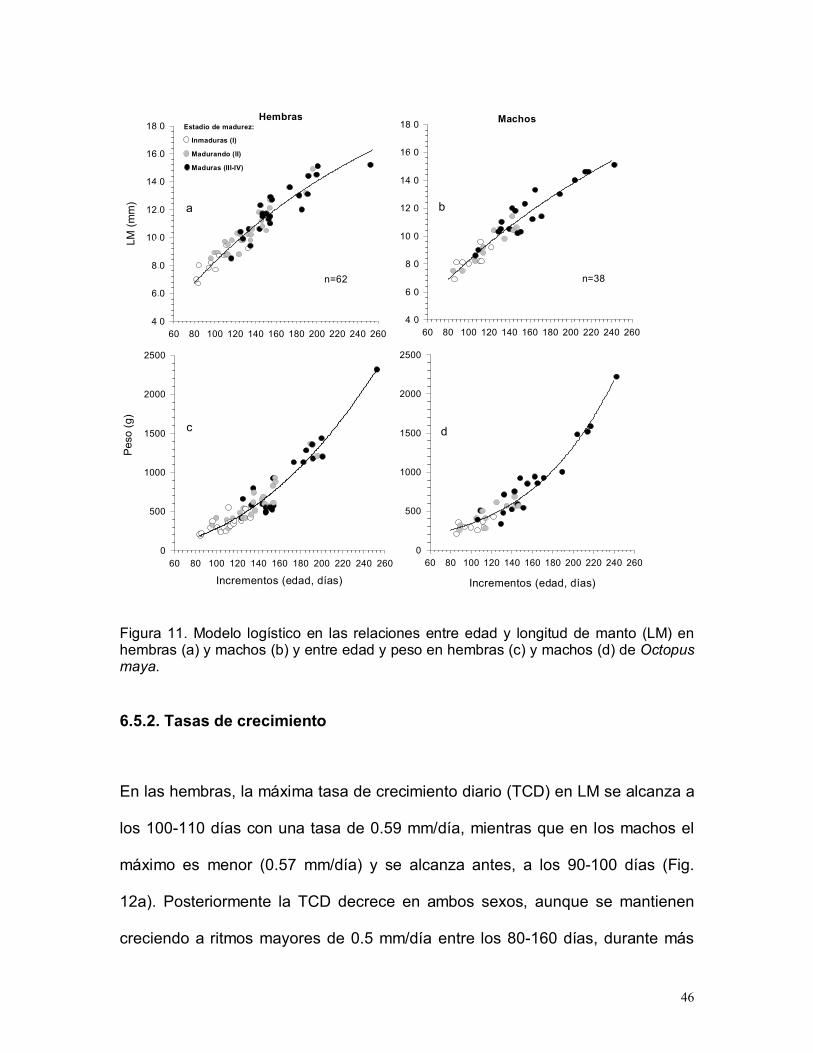
Los 38 machos midieron 69-151 mm de LM, 8 eran inmaduros, 11 estaban
madurando y 19 eran maduros.
La hembra más joven fue un ejemplar inmaduro de 80 días con 78 mm LM y 290
g, con 99 días se presentó un ejemplar madurando con 85 mm LM y 330 g y la
más longeva estaba completamente madura y tenia 253 días, 152 mm LM y
2315 g. El ejemplar de menor talla de este estudio, una hembra inmadura de 67
mm LM y 185 g, tenía 84 días. El macho inmaduro más joven tenia 86 días, 69
mm LM y 205 g, otro macho que estaba madurando presentó 89 días, 75 mm
LM y 255 g y el más longevo era un ejemplar maduro de 243 días, 151 mm LM y
2220 g (10 días mas joven que la hembra mas longeva) (Fig. 11).
Tomando en cuenta la talla de madurez reportada y los parámetros de
crecimiento estimados en el presente trabajo, las hembras inician la maduración
entre los 99 y 157 días y estarán completamente maduras entre los 125 – 253
días. Por su parte, los machos comienzan a madurar a los 89 -144 días y están
completamente maduros a los 108 -243 días de vida.
46
Hembras
A
Machos
B
60 80 100 120 140 160 180 200 220 240 260
LM (m
m)
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
60 80 100 120 140 160 180 200 220 240 2604.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 2600
500
1000
1500
2000
2500
a b
Incrementos (edad, días)60 80 100 120 140 160 180 200 220 240 260
Pes
o (g
)
0
500
1000
1500
2000
2500
c d
Estadio de madurez:
Inmaduras (I)
Madurando (II)
Maduras (III-IV)
n=62 n=38
Figura 11. Modelo logístico en las relaciones entre edad y longitud de manto (LM) enhembras (a) y machos (b) y entre edad y peso en hembras (c) y machos (d) de Octopusmaya.
6.5.2. Tasas de crecimiento
En las hembras, la máxima tasa de crecimiento diario (TCD) en LM se alcanza a
los 100-110 días con una tasa de 0.59 mm/día, mientras que en los machos el
máximo es menor (0.57 mm/día) y se alcanza antes, a los 90-100 días (Fig.
12a). Posteriormente la TCD decrece en ambos sexos, aunque se mantienen
creciendo a ritmos mayores de 0.5 mm/día entre los 80-160 días, durante más
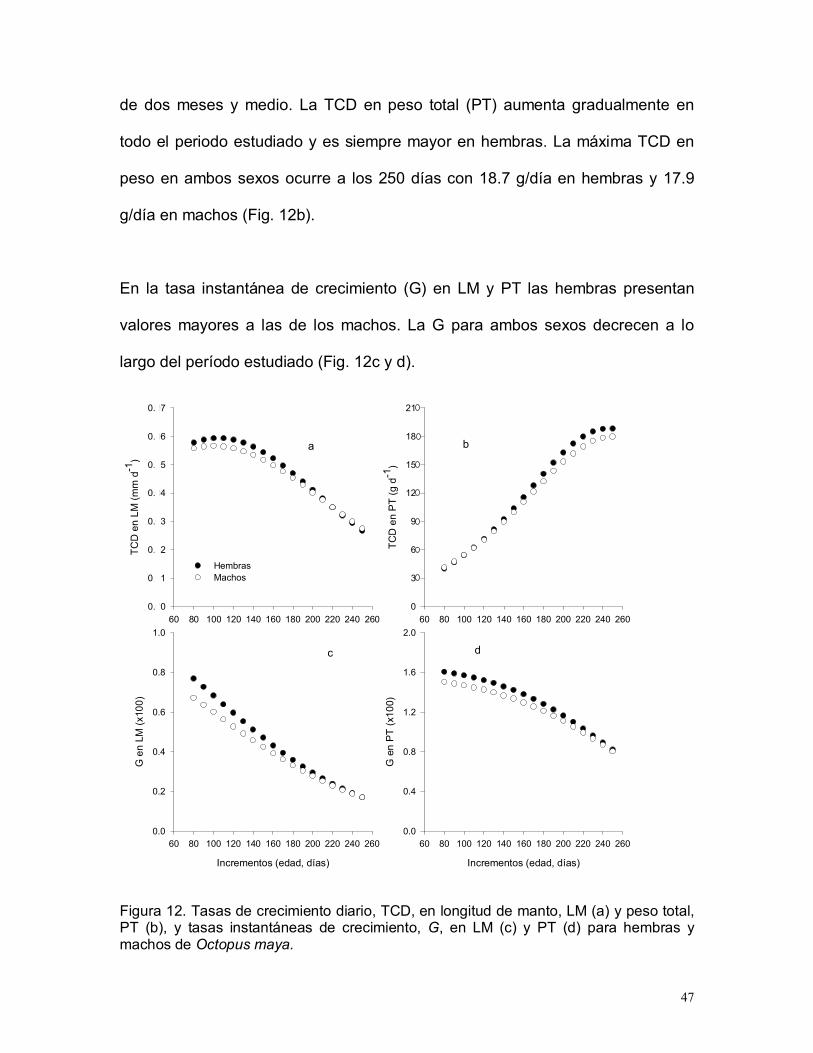
47
de dos meses y medio. La TCD en peso total (PT) aumenta gradualmente en
todo el periodo estudiado y es siempre mayor en hembras. La máxima TCD en
peso en ambos sexos ocurre a los 250 días con 18.7 g/día en hembras y 17.9
g/día en machos (Fig. 12b).
En la tasa instantánea de crecimiento (G) en LM y PT las hembras presentan
valores mayores a las de los machos. La G para ambos sexos decrecen a lo
largo del período estudiado (Fig. 12c y d).
a
60 80 100 120 140 160 180 200 220 240 260
TCD
en
LM (m
m d
-1)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
HembrasMachos
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 260
G e
n LM
(x10
0)
0.0
0.2
0.4
0.6
0.8
1.060 80 100 120 140 160 180 200 220 240 260
TCD
en
PT
(g d
-1)
0
3
6
9
12
15
18
21
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 260
G e
n PT
(x10
0)
0.0
0.4
0.8
1.2
1.6
2.0
b
c d
Figura 12. Tasas de crecimiento diario, TCD, en longitud de manto, LM (a) y peso total,PT (b), y tasas instantáneas de crecimiento, G, en LM (c) y PT (d) para hembras ymachos de Octopus maya.
48
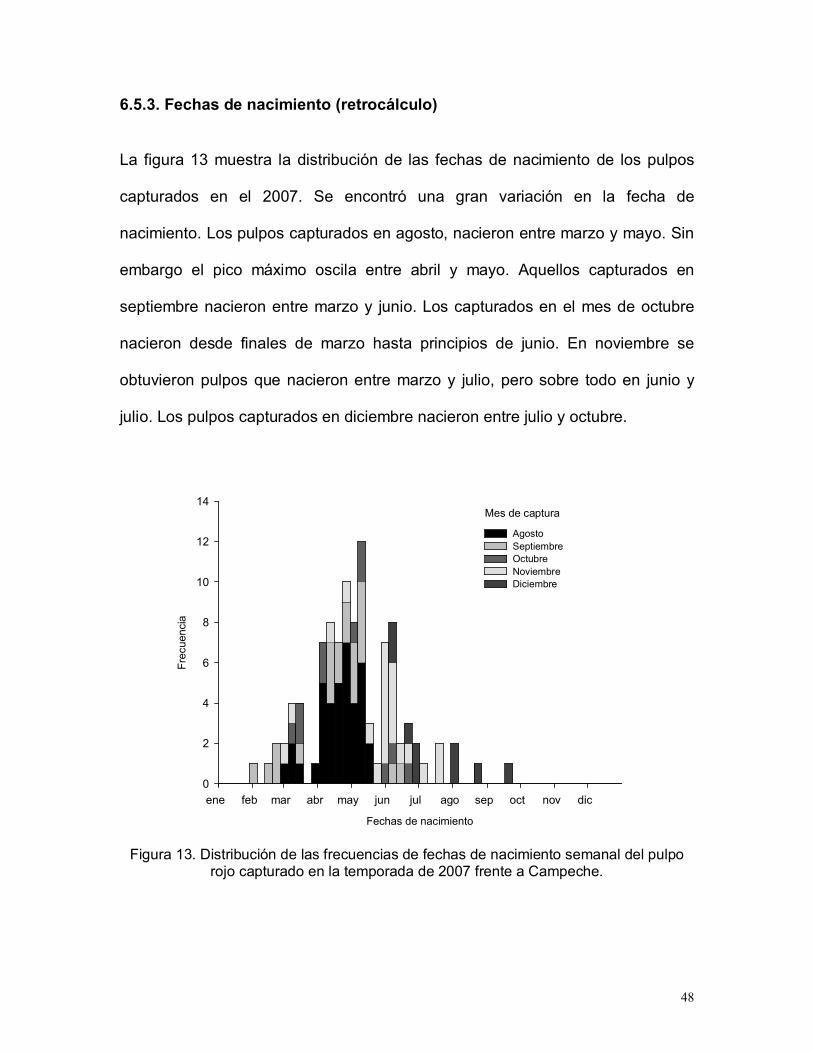
6.5.3. Fechas de nacimiento (retrocálculo)
La figura 13 muestra la distribución de las fechas de nacimiento de los pulpos
capturados en el 2007. Se encontró una gran variación en la fecha de
nacimiento. Los pulpos capturados en agosto, nacieron entre marzo y mayo. Sin
embargo el pico máximo oscila entre abril y mayo. Aquellos capturados en
septiembre nacieron entre marzo y junio. Los capturados en el mes de octubre
nacieron desde finales de marzo hasta principios de junio. En noviembre se
obtuvieron pulpos que nacieron entre marzo y julio, pero sobre todo en junio y
julio. Los pulpos capturados en diciembre nacieron entre julio y octubre.
Fechas de nacimiento
ene feb mar abr may jun jul ago sep oct nov dic
Frec
uenc
ia
0
2
4
6
8
10
12
14
AgostoSeptiembreOctubreNoviembreDiciembre
Mes de captura
Figura 13. Distribución de las frecuencias de fechas de nacimiento semanal del pulporojo capturado en la temporada de 2007 frente a Campeche.

49
7. DISCUSIÓN
7.1. Estilete
7.1.1. Preservación y preparación de los estiletes
Los métodos utilizados en estudios de edad y crecimiento en estructuras duras
para los octópodos presentan ventajas y desventajas desde su preparación
hasta su preservación (Doubleday et al., 2006).
Los estiletes se preservaron en formol al 4%. Esta preservación presenta la
desventaja ya que desprende una ligera capa externa o película en la estructura
(también observado por Leporati et al., 2008). Sin embargo, no afecta en la
observación de los incrementos ya que no hay anillos en esa película,
conclusión obtenida también por Cabido (1984). Sousa y Fernándes (2002)
trabajando con estiletes de O. vulgaris mencionaron que la preservación en
formol es mejor y que nunca debe utilizarse en concentraciones mayores al 4%
para mantener la estructura interna en buenas condiciones.
No se recomienda el uso de alcohol ya que causa deshidratación y proporciona
el aspecto de grietas similares a lo que sucede con el uso de muestras
congeladas. Esa es la razón principal para no utilizar los métodos clásicos
histológicos, así como la inclusión en parafina y resinas sintéticas (Cabido,
1984). Sin embargo, Leporati et al. (2008) no observaron diferencias
significativas entre la preservación en formol al 4% y etanol al 70 % en cuanto a

50
la estimación de la edad en los estiletes de O. pallidus en condiciones de
laboratorio, por lo que la utilización de los dos preservativos es aplicable. Por
otro lado Doubleday et al. (2006) proponen que el etanol al 70% en estiletes de
O. pallidus es el mejor preservativo ya que conserva mejor las propiedades de la
estructura.
En este trabajo los resultados obtenidos por medio de la aplicación de la técnica
a mano alzada o manual de Bizikov (1996) fueron satisfactorios. La ventaja que
presenta esta técnica es que la estructura no se somete a cambios bruscos de
temperatura debido a lijados o pulidos que pudieran dañarla de cara a la
observación de los incrementos. El montaje con gelatina de glicerol además
aparentemente preserva intacta la estructura pudiendo almacenarse por cierto
tiempo (dos años hasta la fecha) sin recurrir solamente a reservas de
fotografías. Asimismo es un método simple, fácil y económico.
Doubleday et al. (2006) y Leporati et al. (2008) prepararon los estiletes
empleando la técnica de lijado y pulido. Encontraron que la visualización de los
incrementos en la región interior de la estructura es limitada e insuficiente,
debido a la opacidad o pigmentación de los incrementos después de someterla a
altas temperaturas debido a la inclusión de cemento termoplástico. Pasados los
cinco minutos la estructura tiende a ser ilegible porque oscurece, ya que
presentan una desintegración microestructural después de la preparación. Es

51
por ello que esta técnica requiere de fotografiar secciones del corte antes de los
cinco minutos. El conteo de incrementos se realiza sobre imágenes.
Las desventajas que se observó en el uso de la técnica a mano alzada en este
trabajo fue lo siguiente: 1) El cortador debe entrenarse constantemente con el
método, para que el corte sea uniforme y permita observar los incrementos
legibles durante el conteo. 2) La lectura de los incrementos cercana al núcleo fue
difícil de contar, por lo tanto los conteos se realizaban en base a la mejor
visibilidad que presentaba el diámetro de la estructura, siguiendo no una misma
dirección de lectura. 4) No aplicó a organismos pequeños con estiletes de 0.9 a
4 mm de longitud, debido a que el estilete era muy pequeño y fue imposible
orientar horizontalmente durante el proceso del corte, además la estructura es
más blanda y susceptible a la ruptura, asimismo de no soportar la presión
ejercida durante el corte.
Sin embargo Barrat y Alcock (2010) realizaron una nueva técnica de
preservación permanente con resina de baja y alta viscosidad, el cual
mencionan cortes de 1 a 2 mm de longitud en estiletes de las especies Octopus
vulgaris y Eledone cirrosa. También indican que su método de corte en los
estiletes les permitió identificar incrementos en el núcleo, a diferencia de la
técnica utilizada por Doubleday et al. (2006). La desventaja que mencionan
estos autores, es que la resina de alta viscosidad no penetra en su totalidad a la

52
estructura, obstaculizando la lectura y recomiendan utlizar resina de baja
viscosidad ya que acelera la polimerización en la estructura.
Es probable que las distinciones de los incrementos desde el núcleo hasta la
periferia en los cortes del estilete, esten asociados a la técnica utilizada. Por lo
tanto se sugiere afinar la técnica de mano alzada, para posteriores trabajos en
lectura de incrementos de crecimiento en Octopus maya.
7.1.2. Relación entre dimensiones del estilete y dimensiones del pulpo
Los estudios de edad y crecimiento en estructuras duras de cefalópodos se han
venido utilizando en los últimos años como indicadores del crecimiento
somático (Young, 1960; Arkhipkin, 1993). Parece posible encontrar una
solución parcial en la relación existente entre el tamaño del estatolito, el número
de incrementos y el tamaño del cefalópodo (LM). Se ha observado que existe
una buena relación entre el número de incrementos y la longitud del estatolito
en muchas especies de calamares (Morris y Aldrich, 1985; Rodhouse y Hatfield,
1990; Villanueva, 1992; Rocha, 1994; Bettencourt, 1994; Collins et al., 1995;
Bettencourt et al., 1996; González et al., 1996). Por otra parte, existe una
relación entre el estatolito y el peso o talla del animal (Rocha, 2003).
Sin embargo los estudios de crecimiento en las especies de pulpos han sido
poco numerosos. El uso de picos para determinar edad no ha demostrado

53
buenos resultados debido al desgaste que presenta la estructura por usarse en
alimentación (Leporati et al., 2008). El primero en hallar incrementos periódicos
en los estatolitos de pulpos fue Young (1960), indicando que podrían servir para
estudios sobre la edad (Arkhipkin, 1991). Los intentos realizados para contar
incrementos en los estatolitos han sido infructuosos, debido a que en éstos el
proceso de mineralización y la orientación de los cristales que les componen son
totalmente diferentes al de otras especies más pelágicas (Rocha, 2003).
Lombarte et al. (2006) menciona que el uso de los estatolitos no ha dado
resultados para el pulpo debido a la ausencia de anillos de crecimiento.
Es por ello que la reciente validez en el uso de estructuras internas como los
estiletes y que no se desgasten como los picos, evitan la incertidumbre de los
estudios de laboratorio y no es tan limitada en su alcance como los estudios de
marcado (Leporati et al., 2008).
Para que una estructura pueda ser considerada como un buen indicador en la
estimación de la edad debe cumplir al menos tres condiciones: que no haya
reabsorción, que los patrones de las bandas de crecimiento sean producto de
diferencias en el crecimiento y por último que exista un crecimiento proporcional
entre la estructura y la longitud corporal del organismo (Cailliet et al., 1983).
Considerando la proporcionalidad hacia una tendencia rectilínea, así como el
alto valor de coeficiente de determinación (r2) de 0.80 y 0.79 entre el diámetro

54
máximo y longitud total del estilete con la longitud de manto del organismo
obtenidos en el presente trabajo, se asume que los estiletes representan
estructuras que reflejan adecuadamente el crecimiento de los organismos con el
paso del tiempo.
Doubleday et al. (2006) mencionan en estudios con O. pallidus en condiciones
de laboratorio, que las dimensiones de los estiletes están estrechamente
relacionados con el peso corporal del organismo. Además sugieren que la
estructura del estilete pone en manifiesto, que sus incrementos se depositan de
manera regular y sobre todo crecen en proporción con el pulpo.
Sin embargo Leporati et al. (2008) concluyen que la longitud del estilete no
presenta concordancia con el peso corporal de O. pallidus (r2 = 0.066 - 0.120
para ambos sexos) ni en relación al diámetro máximo del estilete (r2 = 0.026 -
0.068). En este caso en nuestro trabajo la variable en peso con respecto a la
longitud total del estilete, no presentó un alto coeficiente de determinación (r2 =
0.74), mientras que entre el diámetro máximo del estilete del organismo la r2 fue
de 0.78.
Es probable que el peso en los estiletes, haya presentado una disminución del
peso real debido a la manipulación y el estado de las estructuras al momento de
la preservación.

55
7.2. Estimación de la edad
7.2.1. Microestructura del estilete y Lectura de los incrementos
En este estudio se pudo observar la presencia de cuatro regiones en la zona
interior de la microestructura del estilete y dos tipos de patrones de lectura, los
incrementos de primer orden y segundo orden. Resultados similares reportan
Doubleday et al. (2006) en 52 estiletes de O. pallidus cultivados especificando
cuatro regiones distintas en la microestructura, así como la identificación del
primer incremento de nacimiento (first post-hatch increment) de un conjunto
compuesto por un incremento oscuro y un incremento de luz. Estos autores
indican que la microestructura del estilete en organismos silvestres es muy
similar en apariencia a la de los organismos cultivados.
Otro trabajo que apoya nuestros resultados son los de Sousa y Fernándes
(2002) con 80 estiletes de distintos tamaños de O. vulgaris en la costa
portuguesa, indicando cuatro zonas o etapas de vida del organismo, el cual
presentan varios incrementos concéntricos y algunas de estos con incrementos
muy marcados, que podrían corresponder a distintas fases de crecimiento.
La precisión se define como la reproducibilidad que existe al realizar el conteo
en la misma estructura varias veces sin importar si el valor obtenido es
verdadero o no. No existe un valor predeterminado para estudios de edad,

56
debido a que la precisión está fuertemente influenciada por la especie, la
naturaleza de la estructura y no solamente por el lector (Campana, 2001).
Aunque en los cefalópodos comúnmente se ha establecido que cuando los
incrementos son ilegibles y confusos o los conteos entre lectores difieren, la
precisión de las lecturas no debe de sobrepasar al 10% de la media (Jackson y
Lu, 1994; Pecl, 2004), lo cual concuerda para el resultado obtenido en este
estudio, el cual presentó un 94% de acuerdo entre lectores, con una diferencia
de ±10 incrementos (10%) De igual forma Doubleday et al. (2006) mencionaron
que para la mayoría de las secciones utilizadas en O. pallidus había un alto nivel
de precisión entre los conteos y replicas, con desviaciones muy por debajo del
10%.
7.2.2. Deposición periódica de los incrementos
Existe una relación lineal positiva entre el número de incrementos con la edad
conocida en Octópodos, presentando un alto grado de significancia (r2=0.97)
(Leporati et al., 2008). Estos resultados sostienen la hipótesis de que los
incrementos se producen a diario.
La periodicidad diaria en la formación de los incrementos en estiletes parece ser
un proceso endógeno (Leporati et al., 2008). Doubleday et al. (2006) explican
que los estiletes en O. pallidus criados en cautiverio en el laboratorio no
mostraron influencia de la temperatura o patrones de luz en la formación de los

57
incrementos diariamente de la estructura y había microestructuras similares a la
de los individuos silvestres. Además sugieren que la periodicidad en los
incrementos del estilete está impulsado por un ritmo endógeno circadiano en
lugar de factores extrínsecos, que han sido propuestos en la formación de
incrementos en los otolitos en teleósteos (Campana y Neilson, 1985).
En otros cefalópodos, los incrementos en los estatolitos Loligo vulgaris reynaudii
fueron depositados diariamente en la naturaleza, rechazando así la posibilidad
de periodicidad artificial inducida (Lipinski et al., 1998). Además la temperatura y
las condiciones de luz probablemente tengan un efecto insignificante en la
formación en estatolitos de Sepia officinalis (Bettencourt y Guerra, 2001)
indicando una influencia importante en la formación de los incrementos de tipo
endógeno.
Sin embargo mientras no se sigan planteando más experimentos para evaluar el
efecto de la luz y la temperatura en condiciones de laboratorio, es poco lo que
podemos concluir de lo que hasta ahora se ha sugerido. Justamente la discusión
es especulativa, y Barrat y Allcock (2010) se inclinan a que, en el caso de O.
vulgaris y E. cirrhosa, la formación de los incrementos podría deberse a factores
endógenos asociados a la temperatura y luz, señalan que estos factores
ambientales pueden generar configuraciones específicas en el estilete ya que se
debe precisamente porque éstos regulan procesos fisiológicos y reproductivos
importantes, sobretodo en pulpos costeros como O. maya.

58
7.3. Estimación del crecimiento
7.3.1. Relación longitud de manto-edad y peso total-edad
Mangold y Boletzky (1973) y Hixon (1980) sugieren que la talla y el peso no son
tan buenos indicadores de la edad en los cefalópodos de poblaciones naturales,
ya que el crecimiento está influenciado por factores ambientales y la
disponibilidad de alimento, por lo que la determinación de la edad se tiene que
realizar mediante experimentos de cultivo o lectura de incrementos.
En este trabajo la utilidad del estilete fue satisfactoria, se detectó una correlación
significativa entre el crecimiento somático y el número de incrementos del
estilete. La relación en peso y edad presentó un alto coeficiente de
determinación para ambos sexos. En este sentido se coincide con Doubleday et
al. (2006) en las relaciones entre la edad o número de incrementos con las
dimensiones de las variables en talla y peso se presenta un crecimiento
proporcional con el cuerpo del pulpo. Este autor lo confirma trabajando con O.
pallidus en cautiverio logrando un crecimiento de hasta ocho meses de edad y
mencionan la estrecha relación entre el numero de incrementos, la edad
conocida y el peso del pulpo.
En contraste nuestros resultados difieren con los encontrados por Leporati et al.
(2008) en O. pallidus en organismos en cautiverio, el cual indican que no hubo
correlación entre la edad o número de incrementos con el peso del organismo.

59
Además mencionan que la edad máxima para O. pallidus se estimó en 589 días
para machos y 475 días para hembras, sugiriendo una vida típica de 12 meses a
18 meses aproximadamente.
Estos distintos resultados, probablemente correspondan a la correlación
existente entre la talla (o el peso) y el número de incrementos que es
relativamente baja y/o distinta en numerosas especies y probablemente se
deban a variaciones individuales y estacionales en las tasas de crecimiento
(Rocha y Guerra, 1999). También, pueden estar causadas por cambios en las
tasas de deposición de los incrementos. O'Dor y Wells (1978) mencionan en O.
vulgaris que las hembras disminuyen de peso debido a que no se alimentan
durante la incubación y mueren poco después de la eclosión de los huevos. A
diferencia de los machos que presentan un comportamiento más transitorio y no
presentan el cuidado parental. Leporati et al. (2008) mencionan que la falta de
correlación con la edad y el tamaño del organismo indica un nivel de plasticidad
en el crecimiento y que esta influenciado por factores medioambientales.
Las dimensiones lineales son complicadas de medir de manera consistente en
los animales de cuerpo suave, especialmente cuando hay diferencias en los
métodos de manipulación y procesamiento del material. La congelación y la
fijación o la forma redondeada o globular del pulpo y la relativa falta de
demarcación entre el manto y la cabeza dificultan la medición precisa del animal
(Pierce et al., 1994). El peso también puede estar abierto a sesgos debido al

60
agua retenida en la cavidad del manto, especialmente en pulpos que se pesan
vivos (Boyle y Rodhouse, 2005).
El crecimiento de cualquier organismo puede ser definido como el incremento en
talla (o peso) en un intervalo de tiempo. Aunque es sencillo de medir, es un
proceso complejo que intervienen aspectos fisiológicos y conductuales, que
incluye la búsqueda de alimento, captura de la presa, ingestión digestión
absorción, asimilación, gasto metabólico y excreción (Brett, 1979).
Mangold y Boletzky (1973) indican que la talla (y probablemente el peso) puede
no ser un indicador realista de la edad de los cefalópodos procedentes de
poblaciones silvestres debido a que el crecimiento puede variar
considerablemente dependiendo de factores tales como el alimento y la
temperatura, así como de demostrar una gran variabilidad morfológica con la
latitud como consecuencia de la influencia del ambiente en su desarrollo y
probablemente en su longevidad (Van Heukelem, 1979; Mangold-Wirz, 1963).
Es por ello que se han ajustado diversos modelos de crecimiento como lineales,
asintóticos, exponenciales, cíclicos, sigmoidales y exponencial seguido de
logarítmico (Forsythe y Van Heukelem, 1987 y Jackson, 1994). No hay que
olvidar que el cumplimiento de los modelos supone la aceptación de una serie
de hipótesis que se alejan de la realidad. Además, se emplean modelos que en

61
muchos casos (si no en todos), dan resultados diferentes, debiéndose
homogeneizar los datos obtenidos de los diferentes modelos.
El uso de modelos asintóticos, principalmente el de von Bertalanffy, para
describir el crecimiento de cefalópodos se ha criticado ampliamente por la
inexistencia de evidencias de que realmente haya un estancamiento o límite del
crecimiento a una talla máxima o asintótica en estos organismos. Forsythe y Van
Heukelem (1987) plantean la existencia de dos fases de crecimiento. La primera
fase, entre la eclosión y el reclutamiento, donde las ecuaciones que mejor se
ajustan son del tipo exponencial, y la segunda de tipo logarítmica que ocurre en
juveniles y adultos. Durante esta fase la tasa de crecimiento declina, pero
asegura un crecimiento continuo sin límite superior. Mangold-Wirz (1963)
sugieren que el crecimiento no es asintótico sino que viene limitado por la
maduración sexual y la puesta.
Los resultados de Jackson y Choat (1992) parecen indicar que, al menos en los
cefalópodos tropicales, el crecimiento puede ser lineal o exponencial, pero
siempre uniforme a lo largo de todo el ciclo de vida del animal (es decir, el
crecimiento no constituiría de dos fases diferenciadas). Cortez et al. (1999)
señala sobre O. mimus basado en datos de cultivo y de campo, presenta un
crecimiento exponencial en la primera etapa de vida seguido de un crecimiento
logarítmico o potencial. Itami et al. (1963) mencionan una fase exponencial en el
crecimiento en peso de O. vulgaris desde los primeros días de vida bentónica,

62
seguida de una fase logarítmica iniciando el declive en el ritmo de crecimiento,
posiblemente como consecuencia del inicio de la madurez sexual.
En este estudio las ecuaciones (lineal, potencial y exponencial) no rindieron los
mejores coeficientes de determinación, ni de variación (Tablas 5 y 6). Esto
probablemente se deba a que al momento del cambio de una fase exponencial a
un crecimiento logarítmico, esta fase no se encuentre vinculado a factores
externos como la temperatura o alimentación, sino por el comienzo de la
madurez sexual del individuo, en donde el crecimiento podría cesar o se vuelve
errático y el peso corporal aumenta. El modelo que representa la segunda fase
de crecimiento suele incluir varios métodos matemáticos, debido a la escasa
representación de tallas menores de la colecta en el campo. Y utilizando un
modelo de crecimiento único para todo el ciclo de vida del organismo sería poco
adecuado y representaría sesgos en base a la naturaleza del animal.
Después de probar varios modelos, la estimación del crecimiento en O. maya
determinó que se ajusta mejor a un modelo logístico. Es necesario comentar que
la aplicación del modelo incluyó individuos que oscilaron en talla entre 67 y 152
mm de LM y entre 185 y 2315 g de peso, por lo que nuestros valores solo
representan una pequeña fracción de sub-adultos y adultos de la especie.
Guerra y Sánchez (1998) consideran que los modelos de Gompertz y logísticos
son expresiones que discurren en el crecimiento como una función sigmoidal

63
con un valor superior, en ocasiones otro inferior y con un punto de inflexión. El
modelo de von Bertalanffy aportó valores fuera del contexto de nuestros datos
en estudio, lo que reafirma su poca utilidad a efectos de estimar el crecimiento
en el pulpo. El uso de modelos asintóticos para describir el crecimiento de
cefalópodos debe ser tomado con mucha cautela ya que no existen evidencias
de que realmente exista un estancamiento en el crecimiento a una talla máxima,
por el contrario el crecimiento continúa y la talla se incrementa aunque este
incremento sea mínimo (Forsythe y Van Heukelem, 1987; Moltschaniwskyj,
2004).
En nuestro trabajo, al observar la curva de la función logística en O. maya de
talla ó peso respecto al número de incrementos en hembras y machos, estos
últimos maduran a los 89 días y las hembras a los 99 días de edad,
manteniendo un incremento de crecimiento alto, empezando a disminuir
gradualmente en talla y aumentando en peso.
7.3.2. Tasas de crecimiento
Los cefalópodos son uno de los grupos que alcanzan mayores tasas de
crecimiento (hasta un 20% de su peso al día), pero con una esperanza de vida
relativamente corta. La mayoría de las especies vive solamente un año. Tanto el
crecimiento como la esperanza de vida parecen estar en directa relación con las
variaciones en las condiciones ambientales (Rocha, 2003).

64
Las tasas de crecimiento en los cefalópodos dependerán principalmente de su
elevada tasa de ingestión de alimentos y en la eficiencia en su alta conversión
en el crecimiento somático (Wells y Clarke, 1996). Semmens et al. (2004)
mencionan que estas tasas en los pulpos están fuertemente influenciadas por la
temperatura y cantidad de alimento. Sin embargo Hoyle (2002) sugiere que el
crecimiento general del pulpo no se encuentra fuertemente influenciado por la
temperatura estacional sino con la reproducción.
O. maya es un ejemplo de cefalópodo de crecimiento rápido y en el que las
hembras mueren después del desove y cuidado parental. La tasa de crecimiento
diaria e instantánea en peso y talla (LM) estimada con base en los parámetros
del modelo logístico se encuentran dentro de los intervalos reportados para
octópodos y cefalópodos en general entre 3 y 10% peso por día (Amaratunga,
1983; Lee, 1994; Lee et al., 1998; Domingues et al., 2002).
En este trabajo la tasa de crecimiento en O. maya presentó el 3.2% en peso
(g/día), valores similares son reportadas por Van Heukelem (1976, 1983) el cual
menciona una tasa de crecimiento de 4.1% del peso corporal (g/día) y una tasa
de conversión del 40% debido a las altas tasas de alimentación. Domínguez et
al. (2007) reporta una tasa de 3,3 ± 0,2 y -0,0 ± 0,3% de peso corporal (g/día),
con alimento artificial a base de cangrejos congelados.

65
En otros octópodos las tasas de crecimiento promedio en peso de O. vulgaris
cultivados, se han realizado por Itami et al. (1963) el cual estimó una tasa diaria
de crecimiento relativo en peso de 10.6% en pulpos juveniles (g/día) y por
Mangold y Boletzky (1973) que obtuvieron una tasa entre 4.9% y 6% (g/día)
igualmente con pulpos juveniles. Cortez et al. (1999) determinaron que la tasa
instantánea de crecimiento en peso G de O. mimus con peso total entre 40 y 500
g, fue de 5.5% en promedio (g/día).
La tasa diaria de crecimiento de O. maya obtenida por medio del modelo
logístico (0.59 en hembras y 0.57 para machos mm/día) estimada en este
estudio es mayor a lo reportado por Solís y Chávez (1986) para O. maya en los
estados de Yucatán y Campeche (0.425 mm/día). Por otro lado nuestros
resultados también difieren a lo obtenido por Arreguín et al. (2000) para O. maya
en el Banco de Campeche, Golfo de México (0.467 mm/día) y con Nepita y
Defeo (2001) estimando una tasa diaria de 0.44 mm/día. Las diferencias entre
estos resultados, probablemente se deba al tamaño de la muestra, a variaciones
ambientales o al método utilizado, ya que estos tres autores realizaron las
estimaciones en base a datos con frecuencias de tallas en las capturas y en este
estudio fue realizado en base a un método directo como la lectura de
incrementos en estructuras duras.
Solís et al. (1997) menciona que las fuentes de variación de las estimaciones
obtenidas en el crecimiento de O. maya, pueden tener origen en la variabilidad

66
natural y aquella asociada a los métodos de estimación. Arreguín (1994) sugiere
que los cambios en las estimaciones del crecimiento, depende de la inclusión de
tallas pequeñas en los muestreos y de la presencia de patrones estacionales
como variaciones de temperatura.
Las tasas de crecimiento en pulpos son mayores que las de sepia o el calamar
ya que aquellos, tienden a gastar menos energía debido al camuflaje o el estar
escondidos en las madrigueras (Domínguez et al., 2007). Probablemente la
alimentación, la madurez sexual y la temperatura pueden haber jugado un papel
importante en el crecimiento de O. maya que se desarrolla a temperaturas que
oscilan entre 25 y 26°C presentando una alta tasa de ingestión y conversión de
alimentos de 30 y 60% (Domínguez et al., 1999).
Las diferencias entre los valores de las tasas de crecimiento diario entre sexos,
sugieren la presencia de dimorfismo sexual ya que las hembras de O. maya
crecen más que los machos, sin embargo los machos maduran ligeramente
antes que las hembras.
Avila et al. (2009) indicaron mediante exámenes histológicos que los machos de
esta especie maduran antes que las hembras, aproximadamente entre los tres y
cuatro meses después de la eclosión. Lo que sugiere que se mantienen en
condición reproductiva durante un tiempo prolongado y en consecuencia, el
potencial de transferir sus gametos más de una vez durante su vida. A diferencia

67
de las hembras que presentan solo un período de desove y por lo general
coincide en el momento en que han alcanzado su tamaño máximo. Esto sugiere
que los machos tienen una etapa muy corta de sub-adulto.
Las tasas de crecimiento decaen debido a la madurez sexual de la especie, la
progenie de una hembra que incuba en primavera crecerá mas rápido que una
que incuba en invierno, debido a la mayor temperatura del agua. Además las
especies que maduran a tallas menores crecen más rápido, quizás para evitar a
los depredadores y sobrevivir, ya que el periodo de reproducción es más corto
entre su nacimiento y madurez (Wood y O’Dor, 2000).
En los cefalópodos, las altas temperaturas aceleran el proceso de madurez,
mientras que las bajas lo retrasan (Mangold, 1987), es entonces que las
diferencias entre la tasa de crecimiento entre especies del Orden Teuthoidea,
otros octópodos, O. vulgaris, O. mimus, y O. maya probablemente se deba a los
distintos ambientes pelágico y bentónico, latitudinal y probablemente se asocien
a ambientes oceanográficos distintos. Mientras que Loligo forbesi vive en
temperaturas templadas o frías, O. maya vive en un ambiente sensiblemente
más cálido, entre 20º y 28º C (Fuentes et al., 1965).
En general los cefalópodos no presentan un modelo de crecimiento único, ni
puede éste expresarse en función de un único tipo de curva de crecimiento. El
crecimiento desde el reclutamiento hasta el final de la vida de estas especies

68
puede describirse mediante varios tipos de funciones matemáticas (lineal,
potencial, logarítmica, sigmoidal). La gran variabilidad de las tasas de
crecimiento en una especie determinada, puede deberse a varias causas
posibles como la existencia de tasas de crecimiento estacional diferente o las
discrepancias en las tasas de crecimiento entre individuos pertenecientes a
diferentes poblaciones en áreas geográficas con distintas condiciones
ambientales. De esta manera, la alta variabilidad inherente a las tasas de
crecimiento (individual) permite ajustar varios tipos de curva con tan altos y
semejantes coeficientes de correlación que la elección sobre cuál usar sea
cuestión de preferencia, más que de criterios estrictamente estadísticos (Rocha,
2003).
7.3.3. Fechas de nacimiento (retrocálculo)
Los cefalópodos en general, excluyendo al género Nautilus, son animales de
crecimiento rápido, cuya forma de reproducirse es sexual, se reproducen
generalmente hacia el término de sus vidas sin regeneración gonadal y mueren
(Mangold et al., 1993).
En este trabajo por medio de retrocálculo en las fechas de nacimiento a través
del análisis de estiletes, permiten sugerir que O. maya es una especie con
desoves a lo largo del año o desovante anual y no abarca más de 12 meses de

69
vida (Solís y Chávez, 1985), con un pico mayor al desove entre los meses de
abril y mayo.
Solís (1967) indica que la temporada de desove de O. maya ocurre entre
noviembre a diciembre. La incubación dura cerca de dos meses y en la vida
silvestre de 30 hasta 65 días aproximadamente. Mientras que Van Heukelem
(1977) menciona que el periodo de incubación en organismos cultivados dura 45
días a 25 ºC. Esto quiere decir que la eclosión estaría oscilando a inicios de
febrero coincidiendo con nuestros resultados, sin embargo el pico con mayor
abundancia se presenta en los siguientes meses por lo que probablemente las
hembras es este estudio empezaron a desovar a finales de diciembre e inicios
de enero. Esto implicaría que la distribución de frecuencias de fechas de
nacimiento se deba a diferentes cohortes, variaciones según los cambios
ambientales (temperatura, fotoperiodo, alimentación).
En O. vulgaris el desarrollo embrionario tiene una duración que oscila entre 25 y
125 días, con puestas entre los meses de marzo y agosto en aguas someras y
una eclosión masiva de larvas en el verano. Sin embargo cuando la puesta que
se realiza mayor profundidad, la eclosión de los huevos puede retrasarse hasta
el otoño y el invierno, debido a la baja temperatura del agua (Mangold y
Boletzky, 1973). Estos autores sugieren que puede presentarse distribuciones
de frecuencias distintas pertenecientes a una misma cohorte, ya que pueden
presentar variabilidad, destacando la duración del desarrollo embrionario, las

70
características genéticas propias de cada animal y la influencia de las
condiciones ambientales.
8. CONCLUSIONES
Se indica que la relación del diámetro máximo en el estilete con la longitud de
manto de los organismos, es directamente proporcional al cuerpo somático del
pulpo. Esto podría sugerir que los estiletes del pulpo rojo son buenos
indicadores del crecimiento.
Se identificaron cuatro zonas de incrementos de crecimiento en el estilete y dos
tipos de patrones de lectura conformada por incrementos de segundo orden
(gruesos y difusos) e incrementos de primer orden (delgados y con un patrón
regular).
El modelo logístico fue el que mejor se ajustó a los parámetros de crecimiento
en longitud de manto y peso para O. maya. Las hembras inician la maduración
entre los 99 y 157 días y están completamente maduras entre los 125 – 253
días. Por su parte, los machos comienzan a madurar a los 89 -144 días y están
completamente maduros a los 108 -243 días de vida.
Se encontró que la tasa de crecimiento diario (TCD) en longitud de manto para
hembras es de 0.59 mm/día, mientras que en los machos es menor de 0.57

71
mm/día. En peso para ambos sexos la TCD ocurre a los 250 días con 18.7 g/día
en hembras y 17.9 g/día en machos. Las hembras presentan valores mayores
que los machos en la tasa instantánea de crecimiento (G) en talla y peso.
En base al retrocálculo en la fechas de nacimiento de los pulpos capturados en
los meses de agosto a diciembre de 2007, se encontró una gran variación en la
fecha de nacimiento.

72
9. LITERATURA CITADA
Amaratunga, T. 1983. The role of cephalopods in the marine ecosystem. FAO
Fish. Tech. Pap. 231: 379-415.
Antoine, J.W. y Gilmore, J.C. 1970. Geology of the Gulf of México. Ocean
Industry 5: 34-38.
Arreguín, S.F., Solís, M.J., Sánchez, J.A., Valero, E. y González, M.E. 1994. Age
and growth study of the Octopus maya of the Yucatán continental shelf.
44 th Ann. Sess. Gulf and Caribb. Fish. Inst.
Arreguín, S.F. 2000. Octopus red grouper interaction in the exploited ecosystem
of the Northern continental shelf of Yucatán, México. Ecol. Model. 129:
119-129.
Arkhipkin, A.I. 1991. Methods for cephalopod age and growth studies with
emphasis on statolith ageing techniques. In P. Jereb, S. Ragonese y S. v.
Boletzky, eds. Squid age determination using statoliths, Pp. 11-17.
Proceedings of the international workshop, 9-14 October 1989, Istituto di
Tecnologia della Pesca e del Pescato, N.T.R.-I.T.P.P. Publ. Especial
No.1,Mazara del Vallo, Sicilia, Italia. 127 p.
Arkhipkin, A. 1993. Age, growth, stock structure and migratory rate of pre-
spawning short-finned Illex argentinus based on statolith ageing
investigations. Fish. Res. 16(4): 313-338.
Avila, P.O., Colin, F.R. y Rosas, C. 2009. Gonad Development During the Early
Life of Octopus maya (Mollusca: Cephalopoda). Biol. Bull. 216: 94-102.
Barrat, L.M. y Allcock, A.L. 2010. Ageing octopods from stylets: development of a
technique for permanent preparations. ICES J. Mar. Sci. 67: 1452-1457.
Bettencourt, V. 1994. Estudo da estrutura da população de Loligo vulgaris
(Lamarck, 1799) da costa sul de Portugal através da leitura dos anéis de
crescimento nas estrutural duras, os estatólitos. Tese de Mestrado.
Universidade do Algarve, Portugal. 68 p.

73
Bettencourt, V., Coelho, L., Andrade, J.P. y Guerra, A. 1996. Age and growth of
the squid Loligo vulgaris off the south coast of Portugal, using statolith
analysis. J. Moll. Stud. 62: 359–366.
Bettencourt, V. y Guerra, A. 2001. Age studies based on daily growth increments
in statoliths and growth lamellae in cuttlebone of cultured Sepia officinalis.
Mar. Biol. 139: 327–334.
Beamish, R.J. y Fournier, D.A. 1981. A method for comparing the precision of a
set of age determinations. Can. J. Fish. Aquat. Sci. 38: 982 – 983.
Benssonov, N.M. y González, O. 1967. Principales particularidades de la
formación de la productividad de las aguas del Banco de Campeche. III
Congreso de Oceanografía, México, Inst. Nacional de la Pesca, CIP.
Archivo CIP. 18 p.
Bizikov, V.A. 1996. Atlas of Morphology and Anatomy of the Gladius of Squids.
VNIRO Publishing, Moscu, Rusia. 248 p.
Bizikov V.A. 2004. The shell in Vampyropoda (Cephalopoda): morphology,
functional role and evolution. Ruthenica. 3:1–88.
Boyle, P. y Rodhouse P. 2005. Cephalopods: Ecology and Fisheries. Blackwell
Publishing. Oxford. 452 p.
Botello, A. 1978. Variación de los parámetros hidrológicos en las épocas de
sequía y lluvia (May. Y Nov. De 1974) en la Laguna de Términos
Campeche, México. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón.
México. 5: 159-178.
Brett, J.R. 1979. Environmental factors and growth. In: Fish. Physiol. Vol: 8. 599-
675 p. Hoar, W.S., Randall, D.J. y Brett, J.R. (Eds). Nueva York:
Academic Press.
Bulanienkov, S.K. y García, C. 1973: Influencia de los procesos atmosféricos del
Banco de Campeche. Informe de investigación No.2. IV Reunión de
Balance de trabajo. Centro Inv. Pesq. Inst. Nac. Pesca. 28p
Cabido, M. 1984. Aspectos da biologia de Eledone cirrhosa (Lamark, 1798),
Eledone moschata (Lamark, 1798) e Octopus vulgaris (Cuvier, 1797)

74
(Mollusca: Cephalopoda) na costa portuguesa. PhD thesis, Faculdade de
Ciencias de Lisboa. 94 p.
Caddy, J.F. 1991. Daily rings on squid statoliths: an opportunity to test standard
population models? In P. Jereb, S. Ragonese y S. v. Boletzky, eds. Squid
age determination using statoliths, pp. 53-66. Proceedings of the
international workshop, 9-14 October 1989, Istituto di Tecnologia della
Pesca e del Pescato, N.T.R.-I.T.P.P. Publ. Especial No. 1, Mazara del
Vallo, Sicilia, Italia. 127 p.
Caddy, J.F. y Rodhouse, P.G. 1998. Cephalopod and groundfish landings:
evidence for ecological change in global fisheries? Rev. Fish Biol. Fish. 8:
431–444.
Caddy, J.F. 1999. Fisheries management in the twenty-first century: will new
paradigms apply? Rev. Fish Biol. Fish. 9: 1–43.
Chang, W.Y. 1982. A statistical method for evaluating the reproducibility of age
determination Can. J. Fish. Aquat. Sci. 39: 1208 – 1210.
Cailliet, G.M., Martin L.K., Kusher D., Wolf P. y Welden B.A. 1983. Techniques
for enhancing vertebral bands in age estimation of California
elasmobranchs, 157-165. En: Prince, E.D. y L.M. Pulos (Eds) Proceedings
of the international workshop on age determination of oceanic pelagic
fishes: tunas, billfishes, and sharks. U.S. Dep. Commer., NOAA Tech.
Rep. NMFS 8.
Callow, P. 1987. Fact and theory an overview. En: Cephalopod Life Cycles. Vol
II. Comparative Reviews. Boyle, P.R. (ed.) Academic Press. Londres: 351-
365.
Campana, S.E. y Neilson, J.D. 1985. Microstructure of fish otoliths. Can. J. Fish.
Aquat. Sci. 42: 1014–1032.
Campana, S.E. 2001. Accuracy, precision and quality control in age
determination, including a review of the use and abuse of age validation
methods. J. Fish Biol. 59: 197–242.

75
Challier, L., Royer, J., y Robin, J.P. 2002. Variability in age-at-recruitment and
early growth in English Channel Sepia officinalis described with statolith
analysis. Aquat. Living. Resour. 15: 303-311.
Chen, Y., Jackson, A. y Harvey, H. 1992. A comparison of von Bertalanffy and
polynomial functions in modelling fish growth data. Can. J. Fish. Aquat.
Sci., 49: 1228-1235.
Collins, M.A., Burnell, G.M. y Rodhouse, P.G. 1995. Age and growth of the squid
Loligo forbesi (Cephalopoda: Loliginidae) in Irish waters. J. Mar. Biol.
Assoc. United Kingdom. 75: 605-620.
CONAPESCA e ITESM. 2004. Características generales, aspectos
océanologicos y geográficos del pulpo. SAGARPA. 175 p.
Cortez, T.A., González F. y Guerra A. 1999. Growth of cultured Octopus mimus
(Cephalopoda, Octopidae). Fish. Res. 40: 81-89.
De la Peña, M.E. 1957. Estudios anatómicos a cerca del pulpo (Octopus vulgaris
Lam) Tesis profesional UNAM Facultad de Ciencias México.
Domain F., Jouffre, D. y Caverivière, A. 2000. Growth of Octopus vulgaris from
tagging in Senegalese waters. J. Mar. Biol. Assoc UK 80: 699–705.
Domingues, P.M. 1999. Development of alternative diets for the mass culture of
the European cuttlefish Sepia officinalis. Ph.D. thesis, University of the
Algarve, Portugal, 95 p.
Domingues, P.M., Sykes, T. y Andrade, J.P. 2002. The effects of temperature in
the life cycle of two consecutive generations of the cuttlefish Sepia
officinalis (Linnaeus, 1758), cultured in the Algarve (South Portugal).
Aquacult. Int. 10: 207-220.
Domingues, P., López, N., Muñoz, J.A., Maldonado, T., Gaxiola, G. y Rosas, C.
2007. Effects of an artificial diet on growth and survival of the Yucatan
octopus, Octopus maya. Aquacult. Int. 13: 1–9.
Doubleday Z., Semmens J.M., Pecl G.T., y Jackson G. 2006. Assessing the
validity of stylets as ageing tools in Octopus pallidus. J. Exp. Mar. Biol.
Ecol. 338: 35–42.

76
FAO, 1989. Species catalogue. Vol 3 Cephalopods of world. An annotated and
illustrated catalogue of species of interest to fisheries. FAO. Fish. Synop.,
(125) Vol. 3:277 p.
FAO, 2001. Yearbook of Fishery Statistics. Capture Production. Vo. 88/1. Roma,
FAO. 752 p.
Forsythe, J.W. 1984. Octopus joubini (Mollusca: Cephalopoda): a detailed study
of growth through the full life cycle in a closed seawater system. J. Zool.
202: 393–417.
Forsythe, J.W. y Hanlon, R.T. 1988. Effect of temperature on laboratory growth,
reproduction and life span of Octopus bimaculoides. Mar Biol 98: 369–379
Forsythe, J.W. y Van Heukelem, W.F. 1987. Growth. In: Boyle, P.R. (Ed.),
Cephalopod Life Cycles. Vol. 11. Academic Press, London, p: 135-156.
Fuentes, D.M., Solís, R. y de la Garza, J.M. 1965. Algunos aspectos de la
reproducción del pulpo Octopus vulgaris Lamarck de la Sonda de
Campeche. Secretaria de Industria y Comercio. Dirección General de
Industrias conexas. García, C. 1980. Caracterización general del Banco
de Campeche. Rev. Cub. Inv. Pesq. 5(2).
García, C., y Gómez, J. 1974. Carta preliminar de fondos del Banco de
Campeche INP/CIP, Cuba, Res. Invest. p.37.
García, C. 1980. Caracterización general del Banco de Campeche. Rev. Cub.
Inv. Pesq. 5(2).
Guerra, S.A. y Sánchez, L.J. 1998. Fundamentos de Explotación de Recursos
Vivos Marinos. Editorial Acriba. España. 249 p.
Gonçalves, J. 1993. Octopus vulgaris Cuvier, 1797 (polvo-comum). Sinopse da
biologia e exploraçao. Dept. Oceanografia e Pesca da Univ. dos Açores.
470 p.
González, A.F., Castro B.G. y Guerra A., 1996. Age and growth of the short-
finned squid Illex coindetii in Galician waters (NW Spain) based on
statolith analysis. ICES, J. Mar. Sci. 53: 802-810.
Hanlon, R.T. y Forsythe, J.W. 1985. Advances in the laboratory culture of
octopuses for biomedical research. Lab. Anim. Sci. 33-40.

77
Hatfield, E.M. y Rodhouse P.G. 1994. Migration as a source of bias in the
measurement of cephalopod growth. Antarctic Science, 6(2): 179-184.
Hermosilla, C.A., Rocha, F., Fiorito, G., González, A.F. y Guerra, A. 2010. Age
validation in common octopus Octopus vulgaris using stylet increment
analysis. ICES J. Mar. Sci. 67: 1458-1463.
Hernández, L.J., Hernández, C.J y Hernández, G.V. 2001. Age determined from
daily deposition of concentric rings on common octopus (Octopus vulgaris)
beaks. Fish. Bull. 99: 679–684.
Hernández, F.A., Solís, R.M., Espinoza, M.J., Aguilar, R.M. y Gil, F.R. 2001.
Pulpo. In: Sustentabilidad y Pesca Responsable en México. Evaluación y
Manejo. INP /SAGARPA. 1111 p
Hixon, R.F. 1980. Growth, reproductive biology, distribution and abundance of
three species of loliginid squid (Myopsida, Cephalopoda) in the northwest
Gulf of México. Ph.D. thesis, University of Miami. Coral Gables, Florida.
233 p.
Hoyle, K. 2002. Growth and life history of Octopus pallidus: effects of seasonal
temperature change. Honours Thesis. School of Zoology, University of
Tasmania. Hobart.
Iglesias J., Otero J.J, Moxica., C., Fuentes, L y Sánchez, F.J. 2004. The
completed life cycle of the octopus (Octopus vulgaris, Cuvier) under
culture conditions: paralarval rearing using Artemia and zoeae, and first
data on juvenile growth up to 8 months of age. Aquac Int 12: 481–487.
INP-SAGARPA. 2005. Breviario de la pesquería del pulpo. Instituto Nacional de
la Pesca. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y
Alimentación Gobierno Federal. México.
Itami, K., Izawa, Y., Maeda, S. y Nakai, K. 1963. Notes on the laboratory culture
of the Octopus larvae. Bull. Jap. Soc. Sci. Fish. 29: 514-520.
Jackson, G.D. y Choat, J.H. 1992. Growth in tropical cephalopods: An analysis
based on statolith microestructure. Can. J. Fish. Aquat. Sci. 49: 218-228.
Jackson, G.D. 1994. Applications and future potential on statolith increments
analysis in squids and sepioids. Can. J. Fish. Aquat. Sci., 51: 2612-2625.

78
Jackson, G.D. y Lu. C.C. 1994. Statolith microstructure of seven species of
Antarctic squid captured in Prydz Bay, Antarctica. In Rodhouse.P.G.,
Piatkowski.U. and Lu.C.C. (eds), Southern Ocean Cephalopods: Life
Cycles and Populations. Antarct. ScL, 6: 195-200.
Jackson, G.D., Forsythe, J.W., Hixon, R.F y Hanlon, R.T. 1997. Age, growth, and
maturation of Lolliguncula brevis (Cephalopoda: Loliginidae) in the
northwestern Gulf of Mexico with a comparison of length-frequency versus
statolith age analysis. Can. J. Fish. Aquat. Sci. 54: 2907-2919.
Joll, L.M. 1977. Growth and food intake of Octopus tetricus (Mollusca:
Cephalopoda) in aquaria. Aust. J. Mar. Fresh. Res. 28: 45–56.
Lee, P.G. 1994. Nutrition of cephalopods: fuelling the system. Mar. Fresh. Behav.
Physiol. 25: 35–51.
Leporati, S.C., Semmens., J.M y Pecl, G.T. 2008. Determining the age and
growth of wild octopus using stylet increment analysis. Mar. Ecol. Prog.
Ser. 367: 213-222.
Lipinski, M.R., Durholtz, M.D. y Underhill, L.G. 1998. Field validation of age
readings from the statoliths of chokka squid (Loligo vulgaris reynaudii
d’Orbigny, 1845) and an assessment of associated errors. ICES J. Mar.
Sci. 55: 240-257.
Lombarte, A., Rufino, M. M. y Sánches, P. 2006. Statolith identification of
Mediterranean Octopodidae, Sepiidae, Loliginidae, Ommastrephidae and
Enoploteuthidae based on warp analyses. J. Mar. Biol. Assoc. of the
United Kingdom, 86: 767-771.
Márquez, F. Ré, M.E y Ortiz, N. 2006. Evaluación de la conchilla vestigial de
Enteroctopus megalocyathus como herramienta para la determinación de
edad. Córdoba: Libro de resúmenes. 2006. Resumen. Congreso. XXII
Reunión Argentina de Ecología. Asociación Argentina de Ecología.
Márquez, F. y Ré, M. E. 2009. Morphological and chemical description of the
stylets of the red octopus, Enteroctopus megalocyathus (Mollusca:
Cephalopoda). Moll. Res. 29(1): 27–32.

79
Mangold, W.K. 1963. Biologie des cephalopods benthiques et nectonique del la
Mer Catalane. Vie Milieu, 13: 1-285.
Mangold, K. y Boletzky, S.V. 1973. New data on reproductive biology and growth
of Octopus vulgaris. Mar. Biol. 19: 7-12.
Mangold, K. 1987. Reproduction. In P.R. Boyle (ed.) Comparative Reviews. Vol.
II, 157–200. London, Academic Press.
Mangold, K.M., Young R.E. y Nixon, M. 1993. Growth versus maturation in
cephalopods. En: Okutani,T., R.K. O’Dor & T. Kubodera (eds.), Recent
advances in cephalopod fisheries biology. Tokai University press, Tokyo.
pp. 697-703.
Miliou, H., Fintikaki, M., Kountouris, T. y Verriopoulos, G. 2005. Combined effects
of temperature and body weight on growth and protein utilization of the
common Octopus, Octopus vulgaris. Aquaculture 249: 245–256.
Moltschaniwskyj, N. 2004. Understanding the process of growth in cephalopods.
Mar. Fresh. Res. 55: 379-386.
Morales, G. y Solís, R. M. 1987. Fecundidad y maduración de Octopus maya
Voss y Solís (Mollusca cephalopoda). Ix Congreso Nacional de Zoología.
Villahermosa Tabasco, México. 9p.
Morris, C.C. y Aldrich, F.A. 1985. Statolith length and increment number for age
determination of Illex illecebrosus (LeSeur, 1821) (Cephalopoda,
Omastrephidae) NAFO Sci. Council Studies, 9: 101-106.
Musick, J. A. 1999. Life in the Slow Lane. Am. Fish. Soc. Symp. 23: 2-7.
Nepita, V.M. y Defeo, O. 2001. Crecimiento del pulpo Octopus maya (Mollusca:
Cephalopoda) de la costa de Yucatán, México: un análisis de largo plazo.
Rev. Biol. Trop 49(1): 93 - 101.
O'Dor, R.K. y Wells, M.J. 1978. Reproduction versus somatic growth: hormonal
control in Octopus vulgaris. J. Exp. Biol. 77: 15- 31.
Oosthuizen, A. y Smale, M.J. 2003. Population biology of O. vulgaris on the
temperate south-eastern coast of South Africa. J. Mar. Biol. Ass. U.K. 83:
535-541.

80
Pecl, G.T y Moltschaniwskyj, N. 1999. Somatic growth processes: How are they
altered in captivity. Proc. R. Soc. Lond. Biol. Sci. 266: 1133–1139.
Pecl, G.T. 2004. The in situ relationship between season of hatching, growth and
condition in the southern calamary, Sepioteuthis australis. Mar. Fresh.
Res. 55: 429-438.
Pérez, L.M., Guerra, A. y Sanjuán, A. 2002. Allozime divergence supporting the
taxonomic separation of Octopus mimus and Octopus maya from Octopus
vulgaris (Cephalopoda: Octopus). Bull. Mar. Sci. 71: 653-664.
Pierce, G.J. y Guerra, A. 1994. Stock assessment methods used for cephalopod
fisheries. Fish. Res. 21: 255–286.
Piñeiro, R y Giménez, E. 2001. Características térmicas del Banco de
Campeche. Ciencia Pesquera 15. Julio, 2001 p. 83 -87.
Poder Ejecutivo Federal. 2006. Carta Nacional Pesquera. SAGARPA. Publicada
en el Diario Oficial de la Federación el 25 de agosto de 2006. Ciudad de
Mexico.
Ratkowsky, D. 1983. Nonlinear regression modelling. Marcel Dekker, New York,
276 p.
Raya, C. y Hernández, G.C. 1998. Growth lines within the beak microstructure of
the octopus Octopus vulgaris Cuvier, 1797. S. Afr. J. mar. Sci. 20: 135-
142.
Rocha, F. 1994. Aspectos biológicos y ecológicos de Loligo vulgaris y Loligo
forbesi (Cephalopoda, Loliginidae) en las costas de Galicia (NW de
España). Tesis doctoral. Universidad de Oviedo. 240 p.
Rocha, F. y Guerra, A. 1999. Age and growth of two sympatric squids, Loligo
vulgaris and Loligo forbesi, in Galician waters (north-west Spain). J. Mar.
Biol. Assoc. of the United Kingdom. 79: 697-707.
Rocha, F. 2003. Biología, ecología, cultivo y pesquerías de cefalópodos. Curso
de posgrado, Universidad Austral de Chile. Instituto de Investigaciones
Marinas (Consejo Superior de Investigaciones Marinas). Vigo, España.
Rodhouse, P.G. y Hatfield E.M. 1990. Dynamics of growth and maturation in the
cephalopod Illex argentinus de Castellanos, 1960 (Teuthoidea:

81
Ommastrephidae). Philosophycal Transactions of Royal Society of London
Serie B. 329: 229-241.
Rosas, C., Caamal, C., Cázares, R., Rodríguez, D., Romero, M. y Chay, D.
2006. Manual preeliminar para el cultivo del pulpo Octopus maya. Unidad
Multidisciplinaria de Docencia e Investigación, Facultad de Ciencias,
UNAM. Sisal, Yucatán. 36 p.
SAGARPA. 2005. Diario Oficial de la Federación 16 de marzo de 1994. En
Aguas de Jurisdicción Federal del Golfo de México y Mar Caribe, artículo
segundo, fracción XIX.
Segawa, S. y Nomoto, A. 2002. Laboratory growth, feeding, oxygen consumption
and ammonia excretion of Octopus ocellatus. Bull. Mar. Sci. 7: 801–813.
SEMARNAP. 1999. Anuario Estadístico de Pesca 1998. Secretaria del Medio
Ambiente, Recursos Naturales y Pesca. Gobierno Federal. México.
Semmens, J.M., Pecl, G., Villanueva, R., Jouffre, D., Sobrino, I., Wood, J.W. y
Rugby, P. 2004. Understanding octopus growth: patterns, variability and
physiology. Mar. Fresh. Res. 55: 367–377.
Sobrino, I. y Real, M. 2003. First approach to the quantification of age pigment
lipofuscin in brains from Octopus vulgaris (Mollusca: Cephalopoda). In:
Biology, recruitment, and culture. Cephalopod International Advisory
Council (CIAC) Symposium, Aberdeen (Abstract).
Solís, R.M. 1962. Contribución al estudio del pulpo (Octopus vulgaris Lamarck)
de la Sonda de Campeche. Trabs. Divulg. III (24): 1-30 p.
Solís, R.M. 1967. Aspectos biológicos del pulpo Octopus maya Voss y Solís.
Publ. Inst Nal. Inv. Biol. Pesq. 18: 190.
Solís, R.M y Chávez, E.A. 1985. Evaluación y Régimen Óptimo de Pesca del
Pulpo en la Península de Yucatán, México. Anales del Centro de Ciencias
del Mar y Limnología.
Solís, R.M y Chávez, E.A. 1986. Evaluación y régimen óptimo de pesca del
pulpo en la Península de Yucatán. Anales del Instituto de Ciencias del
Mar y Limnología UNAM. 13: 1-18.

82
Solís, R.M. 1987. El recurso pulpo del Golfo de México y Caribe. 25 Aniv. INP.
Ciclo: Los recursos pesqueros del país. SEPÉSCA. México.
Solís, R.M. 1994. La pesquería del pulpo del golfo de México y Caribe mexicano.
Err. Atlas Pesquero y Pesquerías Relevantes de México. C.D. Multimedia.
Secretaria de Pesca, INP. CENEDIC. Univ. de Colima, México.
Solís, R.M., Espinoza, J.C. y Ramírez, F. 1996. Pesca exploratoria y
experimental de pulpo con trampas japonesa en la plataforma continental
de Yucatán. Mem. II Simposium sobre Investigaciones en Biología y
Oceanografía Pesquera en México. La Paz, B. C. S.
Solís, R.M. 1997. The Octopus maya fishery of the Yucatán Peninsula. En:
Lang, Hochberg y Ambrose (eds). The Fishery and Market Potential of
Octopus in California, CMSC: 1-10.
Sousa, R.C. y Fernandes, R. 2002. Growth observations on Octopus vulgaris
Cuvier, 1797 from the Portuguese waters: growth lines in the vestigial
shell as possible tools for age determination. Bull. Mar. Sci. 71: 1099-
1103.
Sparre, P. y Venema, S.C. 1995. Introducción a la evaluación de recursos
pesqueros tropicales. Parte 1. Manual FAO. Documento Técnico de
Pesca. N°306/1. Roma. 420 p.
Sweeney, M.J y Roper, C.F.E. 1998. Classification, type localities, and type
repositories of recent cephalopoda. En: Voss GL, M Vecchione, RB Toll &
MJ Sweeney. Systematics and biogeography of cephalopods. Smith.
Cont. Zoo. 586: 561-599
Van Heukelem, W.F. 1976. Growth, bioenergetics and life span of Octopus
cyanea and Octopus maya. University of Hawaii, Hawaii, p 1–224.
Van Heukelem, W.F. 1977. Laboratory maintenance, breeding, rearing and
biomedical research potential of the Yucatan octopus (Octopus maya).
Lab. Anim. Sci. 27: 852-859.
Van Heukelem, W.F. 1979. Environmental control of reproduction and life span in
octopus, an hypothesis. In S.E. Stancyk (ed.) Reproductive Ecology of
Marine Invertebrates, 123–133. Columbia, University of Carolina Press.

83
Van Heukelem, W.F. 1983. Octopus maya In: Boyle, P.R. (Ed.), Cephalopod Life
Cycles. Vol. I. Academic Press, London, p: 311-323.
Villanueva, R. 1992. Interannual growth differences in the oceanic squid
Todarodes angolensis Adam in the Northern Benguela upwelling system,
based on statolith growth increment analysis. J. Exp. Mar. Biol. and Ecol.
159: 157-177.
Voss, G L. y Solís, M.J. 1966. Octopus maya, a new species from the Bay of
Campeche. Bull. Mar. Sci. 16: 615-625.
Weatherley, A.H. y Gill, H.S. 1987. Chapter 5:Tissues and growth. In The
Biology of Fish Growth. p. 147–175. (Academic Press: London, UK.).
Wells, M.J. y Clarke, A. 1996. Energetics, the costs of living and reproducing for
an individual cephalopod. Philosophical Transactions of the Royal Society
of London, Series B. Biol. Sci. 351: 1083–1104.
Word, J.B. y O'Dor, R.K. 2000. Do larger cephalopods live longer? Effects of
temperature and phylogeny on interspecific comparisons of age and size
at maturity. Mar. Biol. 136: 91-99.
Yatsu, A.S., Midorikawa, T., Shimada, y Uozumi, Y.1997. Age and growth of the
neon flying squid, Ommastrephes bartrami, in the North Pacific Ocean.
Fish. Res. 29: 257-270.
Young, J. 1960. The statocysts of Octopus vulgaris. Proc. R. Soc. Ser. B. 152: 3–
29.
Zambrano, G.L. 1992. Contribución al conocimiento de la mortalidad natural de
Octopus maya en la plataforma continental de la Península de Yucatán,
México. Tesis de Licenciatura, Facultad de Ciencias UNAM, México D.F.,
México.

84
10. ANEXO
2010
N Albarrán/ I Méndez-Loeza/ U Markaida
EDAD Y CRECIMIENTO DEL PULPO ROJO OCTOPUS MAYA
(VOSS Y SOLÍS, 1966) POR MEDIO DE LA LECTURA DE INCREMENTOS EN
EL ESTILETE

85
Edad y crecimiento del pulpo rojo Octopus maya
(Voss y Solís, 1966) por medio de la lectura de incrementos en el estilete
Age and growth for the red octopus, Octopus maya (Voss and Solís, 1966)
through stylet increments reading
Natalia Albarrán1, Iván Méndez-Loeza1, Unai Markaida1,*
1Departamento de Aprovechamiento y Manejo de Recursos Acuáticos, El
Colegio de la Frontera Sur (ECOSUR), Calle 10 # 264, Col. Centro, 24000
Campeche, México.
* Corresponding author E-mail: [email protected]
Resumen
Se estimó la edad y se describió el crecimiento del pulpo rojo, Octopus maya,
a través de los incrementos de crecimiento depositados en los estiletes de 100
organismos seleccionados de cinco puertos de desembarque pesqueros en el
litoral de Campeche, México, durante 2007. Los estiletes fueron preservados en
formol al 4% y se montaron con gelatina de glicerol. La relación encontrada entre
la longitud total (LTE) y el diámetro máximo del estilete (DME) con la longitud de
manto (LM) es lineal. Se identificaron cuatro zonas de incrementos de
crecimiento en el estilete y dos tipos de patrones de lectura. La relación entre el
diámetro máximo del estilete y la edad para ambos sexos fue mayor. El modelo

86
logístico fue el que mejor se ajusto a los parámetros de crecimiento. La
maduración en las hembras inicia a los 99 -157 días y en machos de 89 -144
días. Las hembras maduran entre los 125 -253 días y los machos entre 108-243
días. La tasa de crecimiento diario en talla y peso total para hembras fue (0.59
mm LM y 18.7 g por día) y en machos (0.57 mm LM y, 17.9 g por día). En base
al retrocálculo en la fechas de nacimiento de los pulpos capturados en los meses
de agosto a diciembre de 2007, se encontró una gran variación en la fecha de
nacimiento.
Palabras clave: Octopus maya, estiletes, incrementos, edad, crecimiento.
Abstract
We estimated the age and described the growth of the red octopus, Octopus
maya, through growth increments deposited in the stylets of 100 bodies selected
from five fish landing ports on the coast of Campeche, Mexico, during 2007. The
freehand technique was used for observation of increments, the stylets were
preserved in formalin at 4% and glycerol gelatin mounting apparently preserves
the structure intact. The relationship that was found between total length (LTE)
and the maximum diameter of the stylet (DME) with the size of the organisms
(LM) is rectilinear, indicating that both variables increase proportionally. We
identified four areas of growth increments in the stylet and two types of reading
patterns. The relationship between the maximum diameter of the stylet and the
age for both sexes was greater, the logistic model was the one that best fits the
parameters of growth. The maturation in females starts at 99 -157 days and in
males at 89-144 days, mature between 125 -253 days in females and 108-243

87
days in males. The daily growth rate in size and total weight for females was
(0.59 mm / day, 18.7 g / day) and for males (0.57 mm / day, 17.9 g / day). Based
on back-in dates of birth of the octopus caught in the months August to
December 2007, there was a large variation in birth date.
Keywords: Octopus maya, stylets, increments, age, growth.

88
Introducción
El pulpo rojo Octopus maya (Voss y Solís 1966) es una especie endémica de la
península de Yucatán que se distribuye desde Ciudad del Carmen, Campeche
hasta Islas Mujeres, Quintana Roo (Solís, 1994, 1997). Constituye el principal
recurso pesquero en términos económicos y el segundo en volúmenes de
captura en los estados de Campeche y Yucatán, representando una de las
mayores pesquerías de pulpo en el mundo (Nepita y Defeo, 2001; INP-
SAGARPA, 2005).
El crecimiento es un aspecto biológico complejo, que en octópodos no ha sido
cuantificado hasta el momento con suficiente certeza, debido a la falta de
estructuras duras donde analizar incrementos periódicos. Se requiere
información sobre las características del ciclo de vida para la interpretación de
gran parte de su ecología y para la evaluación de la pesca (Boyle y Rodhouse,
2005). La mayoría de los estudios de edad y crecimiento en cefalópodos se
basan en métodos indirectos como el análisis de la distribución de frecuencias
de tallas que han sido ampliamente criticados (Arkhipkin, 1991; Hatfield y
Rodhouse, 1994). El uso de técnicas directas, en base al descubrimiento de
incrementos de crecimiento en diferentes estructuras duras corporales en
cefalópodos como estatolito, pluma o gladio y picos, ha permitido avanzar en el
estudio de su edad y crecimiento (Arkhipkin, 1991). Recientemente se han
realizado trabajos preliminares con el vestigio de la concha interna,de los pulpos
llamados estiletes (Bizikov, 2004) que han sido validados para la estimación de
edad en Octopus pallidus (Doubleday, et al., 2006).

89
En México, a pesar de la importancia económica que representa la pesquería del
pulpo Octopus maya, la poca información biológica sobre datos de edad y
crecimiento no han sido contundentes para esclarecer aspectos como
longevidad, fecundidad, dinámica de poblaciones y migraciones (Voss y Solís,
1966) debido a que la mayoría de los estudios se han enfocado a su utilización
comercial. En el presente estudio se describe las estimaciones de la edad y
crecimiento realizadas por lectura directa de estiletes en el pulpo rojo O. maya
durante la temporada de 2007 en el litoral de Campeche.
Materiales y métodos
Se muestrearon los principales puertos de desembarque pesqueros del litoral
campechano, Sabancuy, Champotón, Seybaplaya, Campeche e Isla arena
durante la temporada de pesca de pulpo del 2007 (agosto a diciembre) (fig. 1)
(Tabla 1). De 100 organismos por mes y por localidad se midieron la longitud
dorsal del manto (LM) en mm, el peso total (kg) y el sexo y se les extrajeron los
estiletes para preservarlos en formol al 4% En laboratorio se midió la longitud y
se peso cada estilete. Se obtuvieron cortes transversales con un grosor de 0.1-
0.2 mm trabajando manualmente con una navaja de afeitar (fig. 2) (Bizikov,
1996). Los cortes se realizaron en la región posterior del estilete cerca del codo
debido a que es una zona que presenta mejor visibilidad de incrementos y
menos daño al momento de extraer la estructura. La edad se estimó por medio
del conteo de los incrementos de crecimiento observados por dos lectores de
manera independiente. El patrón de crecimiento se describió a partir del ajuste

90
de diferentes modelos a los datos de edad-longitud de manto y edad-peso,
seleccionando el modelo de acuerdo al mayor coeficiente de determinación (r2) y
menor coeficiente de varianza (CV%) de los parámetros estimados. Para cada
sexo se estimaron tasas de crecimiento diaria (TCD, mm o g por día) TCD = (S2
– S1)/ΔT y tasas instantánea de crecimiento (G x 100) G = (lnS2 – lnS1)/ΔT
calculando los valores de longitud de manto (LM) y peso en intervalos de 10 días
utilizando los parámetros de las curvas seleccionadas (Forsythe y Van
Heukelem, 1987). Por medio del retrocálculo se estimaron las fechas de
nacimiento de cada individuo, sustrayendo la edad asignada mediante el conteo
de incrementos en el estilete a la fecha de captura.
(INSERTAR FIGURA 1)
(INSERTAR TABLA 1)
(INSERTAR FIGURA 2)
Resultados
Microestructura del estilete
Se identificaron cuatro zonas de incrementos de crecimiento: el núcleo (sin
incrementos), región interna, región media y región externa (fig. 3). Se
observaron dos grupos de incrementos de crecimiento: incrementos delgados
(de primer orden) e incrementos gruesos (de segundo orden) (Doubleday et al.,
2006; Márquez y Ré, 2006). La marca de nacimiento se identificó como el primer
incremento visible, después del núcleo embrionario que suele ser amorfo.
(INSERTAR FIGURA 3)

91
Estimación del crecimiento
El modelo logístico fue el que mejor se ajustó a los parámetros de crecimiento
en longitud de manto y peso para O. maya. Las hembras inician la maduración
entre los 99 y 157 días y están completamente maduras entre los 125 – 253
días. Por su parte, los machos comienzan a madurar a los 89 -144 días y están
completamente maduros a los 108 -243 días de vida (fig. 4).
(INSERTAR FIGURA 4)
Tasas de crecimiento
En hembras, la máxima tasa de crecimiento diario (TCD) en LM se alcanza a los
100-110 días con una tasa de 0.59 mm/día, mientras que en los machos el
máximo es menor (0.57 mm/día) y se alcanza antes, a los 90-100 días (fig. 5).
Las tasas instantáneas de crecimiento (G) en LM y PT de las hembras son
siempre mayores a la de los machos.
(INSERTAR FIGURA 5)
Fechas de nacimiento
Los pulpos capturados en agosto, nacieron entre marzo y mayo con un pico en
abril y mayo. Aquellos capturados en septiembre nacieron entre marzo y junio.
Los capturados en el mes de octubre nacieron desde finales de marzo hasta
principios de junio. En noviembre se muestrearon pulpos que nacieron entre
marzo y julio, pero sobre todo en junio y julio, y los de diciembre nacieron entre
julio y octubre (fig. 6).
(INSERTAR FIGURA 6)

92
Discusión
Los resultados obtenidos por medio de la preparación de estiletes con la técnica
manual de Bizikov (1996) fueron satisfactorios. La ventaja que presenta esta
técnica es que la estructura no se somete a cambios bruscos de temperatura
debido a montajes, lijados o pulidos que pudieran dañar los estiletes dificultando
la observación de sus incrementos. En el presente trabajo, se asume que los
estiletes representan estructuras que reflejan adecuadamente el crecimiento de
los organismos con el paso del tiempo. La presencia de cuatro regiones en la
zona interior de la microestructura del estilete y dos tipos de patrones de lectura
son similares a los que reportan Doubleday et al. (2006). Se detectó una
correlación significativa entre el crecimiento somático y el número de
incrementos del estilete.
La estimación del crecimiento en O. maya determinó que se ajusta mejor a un
modelo logístico. Es necesario comentar que la aplicación del modelo incluyó
individuos que oscilaron en talla entre 67 y 152 mm de LM y entre 185 y 2315 g
de peso, por lo que nuestra muestra solo representa una pequeña fracción de
sub-adultos y adultos de la especie. Las hembras inician la maduración entre los
99 y 157 días y están completamente maduras entre los 125 – 253 días. Por su
parte, los machos comienzan a madurar a los 89 -144 días y están
completamente maduros a los 108 -243 días de vida. Esto sugiere que los
machos maduran ligeramente antes que las hembras y se mantienen en
condición reproductiva durante un tiempo prolongado y en consecuencia, el
potencial de transferir sus gametos más de una vez durante su vida (Ávila et al.,

93
2009). A diferencia de las hembras que presentan solo un período de desove y
por lo general coincide en el momento en que han alcanzado su tamaño
máximo. Esto podría indicar que los machos tienen una etapa muy corta de sub-
adulto.
La tasa de crecimiento diaria e instantánea en peso y longitud de manto
estimada con base en los parámetros del modelo logístico se encuentran dentro
de los intervalos reportados para octópodos y cefalópodos en general de entre 3
y 10% peso por día (Amaratunga, 1983; Lee, 1994; Lee et al., 1998; Domingues
et al., 2002).
La tasa diaria de crecimiento obtenida por medio del modelo logístico (0.59 en
hembras y 0.57 para machos mm/día) estimada en este estudio es mayor a lo
reportado por Solís y Chávez (1986) para O. maya en los estados de Yucatán y
Campeche (0.425 mm/día). Por otro lado nuestros resultados también difieren a
lo obtenido por Arreguín et al. (2000) para O. maya en el Banco de Campeche,
Golfo de México (0.467 mm/día) y con Nepita y Defeo (2001) estimando una
tasa diaria de 0.44 mm/día. Las diferencias entre estos resultados,
probablemente se deba al tamaño de la muestra, a variaciones ambientales o al
método utilizado, ya que estos tres autores realizaron las estimaciones en base
a datos con frecuencias de tallas en las capturas y en este estudio fue realizado
en base a un método directo como la lectura de incrementos en estructuras
duras. La tasa de crecimiento de O. maya fue 3.2% en peso corporal (g/día) y
un rango en longitud de manto de 0.57 y 0.59 mm/día-1, además de una
longevidad de 253 días en hembras y 243 días en machos, valores similares son

94
reportadas en condiciones de cultivo por Van Heukelem (1977, 1983) con una
tasa de crecimiento de 4.1% del peso corporal (g/día) y una longevidad media de
295 días para los machos y 305 para las hembras. Domínguez et al. (2007)
mencionan una tasa de 3,3 ± 0,2 y -0,0 ± 0,3% de peso corporal (g/día).
La distribución de las fechas de nacimiento a partir del retrocálculo permite
sugerir que O. maya es una especie que se reproduce en gran parte del año,
con un pico de desove entre los meses de abril y mayo. Solís (1967) reporta
desoves de esta especie en diciembre, 1964, y enero y febrero, 1965. Morales y
Solís (1987) revelan desoves estacionales, con un pico en noviembre. Por otro
lado Arreguín (1994), sugiere la existencia de dos picos de reproducción en el
año como resultado de dos temporadas de desove o una temporada larga con
dos periodos de mayor intensidad, uno en otoño y el otro en primavera. La
eclosión estaría oscilando a inicios de febrero coincidiendo con nuestros
resultados, sin embargo el pico con mayor abundancia se presenta en los
siguientes meses por lo que probablemente las hembras es este estudio
empezaron a desovar a finales de diciembre e inicios de enero. Esto implicaría
que la distribución de frecuencias de fechas de nacimiento se deba a diferentes
cohortes, variaciones en los cambios ambientales (temperatura, fotoperiodo,
alimentación).

95
Agradecimientos
El presente estudio fue realizado dentro del proyecto “Aspectos de la biología
pesquera, el cultivo y la repoblación del pulpo Octopus maya en el litoral de
Campeche. Finalmente se agradece a FOMIX-2007 por el apoyo económico
ofrecido a este trabajo, el cual se enmarca dentro de la tesis de maestría del
primer autor.
Referencias
Amaratunga, T. 1983. The role of cephalopods in the marine ecosystem. FAO
Fish. Tech. Pap. 231: 379-415.
Arreguín, S.F., Solís, M.J., Sánchez, J.A., Valero, E. y González, M.E. 1994. Age
and growth study of the Octopus maya of the Yucatán continental shelf.
44 th Ann. Sess. Gulf and Caribb. Fish. Inst.
Arreguín, S.F. 2000. Octopus red grouper interaction in the exploited ecosystem
of the Northern continental shelf of Yucatán, México. Ecol. Model. 129:
119-129.
Arkhipkin, A.I. 1991. Methods for cephalopod age and growth studies with
emphasis on statolith ageing techniques. In P. Jereb, S. Ragonese y S. v.
Boletzky, eds. Squid age determination using statoliths, Pp. 11-17.
Proceedings of the international workshop, 9-14 October 1989, Istituto di
Tecnologia della Pesca e del Pescato, N.T.R.-I.T.P.P. Publ. Especial
No.1,Mazara del Vallo, Sicilia, Italia. 127 p.
Avila, P.O., Colin, F.R. y Rosas, C. 2009. Gonad Development During the Early
Life of Octopus maya (Mollusca: Cephalopoda). Biol. Bull. 216: 94-102.
Bizikov, V.A. 1996. Atlas of Morphology and Anatomy of the Gladius of Squids.
VNIRO Publishing, Moscu, Rusia. 248 p.
Bizikov V.A. 2004. The shell in Vampyropoda (Cephalopoda): morphology,
functional role and evolution. Ruthenica. 3:1–88.

96
Boyle, P. y Rodhouse P. 2005. Cephalopods: Ecology and Fisheries. Blackwell
Publishing. Oxford. 452 p.
Domingues, P.M., Sykes, T. y Andrade, J.P. 2002. The effects of temperature in
the life cycle of two consecutive generations of the cuttlefish Sepia
officinalis (Linnaeus, 1758), cultured in the Algarve (South Portugal).
Aquacult. Int. 10: 207-220.
Domingues, P., López, N., Muñoz, J.A., Maldonado, T., Gaxiola, G. y Rosas, C.
2007. Effects of an artificial diet on growth and survival of the Yucatan
octopus, Octopus maya. Aquacult. Int. 13: 1–9.
Doubleday Z., Semmens J.M., Pecl G.T., y Jackson G. 2006. Assessing the
validity of stylets as ageing tools in Octopus pallidus. J. Exp. Mar. Biol.
Ecol. 338: 35–42.
Forsythe, J.W. y Van Heukelem, W.F. 1987. Growth. In: Boyle, P.R. (Ed.),
Cephalopod Life Cycles. Vol. 11. Academic Press, London, p: 135-156.
Hatfield, E.M. y Rodhouse P.G. 1994. Migration as a source of bias in the
measurement of cephalopod growth. Antarctic Science, 6(2): 179-184.
INP-SAGARPA. 2005. Breviario de la pesquería del pulpo. Instituto Nacional de
la Pesca. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y
Alimentación Gobierno Federal. México.
Lee, P.G. 1994. Nutrition of cephalopods: fuelling the system. Mar. Fresh. Behav.
Physiol. 25: 35–51.
Márquez, F. Ré, M.E y Ortiz, N. 2006. Evaluación de la conchilla vestigial de
Enteroctopus megalocyathus como herramienta para la determinación de
edad. Córdoba: Libro de resúmenes. 2006. Resumen. Congreso. XXII
Reunión Argentina de Ecología. Asociación Argentina de Ecología.
Nepita, V.M. y Defeo, O. 2001. Crecimiento del pulpo Octopus maya (Mollusca:
Cephalopoda) de la costa de Yucatán, México: un análisis de largo plazo.
Rev. Biol. Trop 49(1): 93 - 101.
Solís, R.M y Chávez, E.A. 1986. Evaluación y régimen óptimo de pesca del
pulpo en la Península de Yucatán. Anales del Instituto de Ciencias del
Mar y Limnología UNAM. 13: 1-18.

97
Solís, R.M. 1994. La pesquería del pulpo del golfo de México y Caribe mexicano.
Err. Atlas Pesquero y Pesquerías Relevantes de México. C.D. Multimedia.
Secretaria de Pesca, INP. CENEDIC. Univ. de Colima, México.
Solís, R.M. 1967. Aspectos biológicos del pulpo Octopus maya Voss y Solís.
Publ. Inst Nal. Inv. Biol. Pesq. 18: 190.
Solís, R.M. 1987. El recurso pulpo del Golfo de México y Caribe. 25 Aniv. INP.
Ciclo: Los recursos pesqueros del país. SEPÉSCA. México.
Solís, R.M. 1997. The Octopus maya fishery of the Yucatán Peninsula. En:
Lang, Hochberg y Ambrose (eds). The Fishery and Market Potential of
Octopus in California, CMSC: 1-10.
Van Heukelem, W.F. 1977. Laboratory maintenance, breeding, rearing and
biomedical research potential of the Yucatan octopus (Octopus maya).
Lab. Anim. Sci. 27: 852-859.
Van Heukelem, W.F. 1983. Octopus maya In: Boyle, P.R. (Ed.), Cephalopod Life
Cycles. Vol. I. Academic Press, London, p: 311-323.
Voss, G L. y Solís, M.J. 1966. Octopus maya, a new species from the Bay of
Campeche. Bull. Mar. Sci. 16: 615-625.
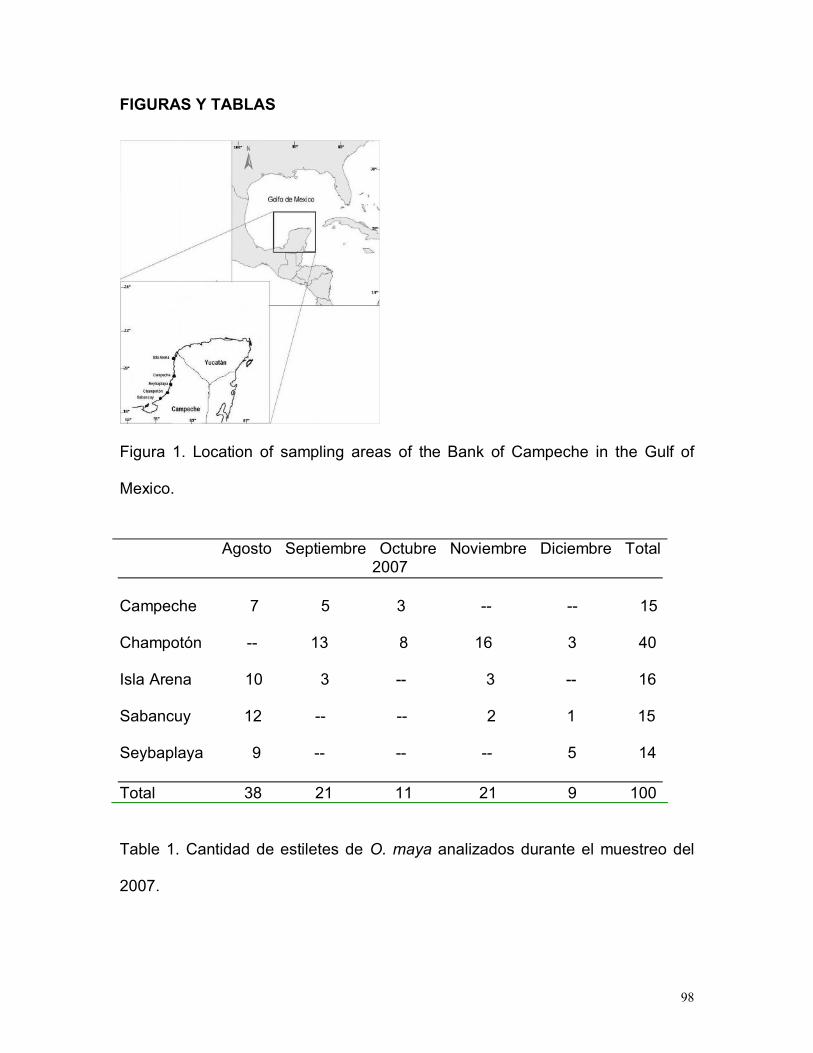
98
FIGURAS Y TABLAS
Figura 1. Location of sampling areas of the Bank of Campeche in the Gulf of
Mexico.
Table 1. Cantidad de estiletes de O. maya analizados durante el muestreo del
2007.
Agosto Septiembre Octubre Noviembre Diciembre Total2007
Campeche 7 5 3 -- -- 15
Champotón -- 13 8 16 3 40
Isla Arena 10 3 -- 3 -- 16
Sabancuy 12 -- -- 2 1 15
Seybaplaya 9 -- -- -- 5 14
Total 38 21 11 21 9 100

99
Figura 2. Technique used for cutting the stylus (Modified from Bizikov, 1996).
Figure 3. Female of Octopus maya with 97 mm ML, 390 g TW and 118 days. (A)
Microstructure of a stylet in cross section (50x), (B) Brand of birth and second
order increments (100x), (C and D) First order increments (400x).
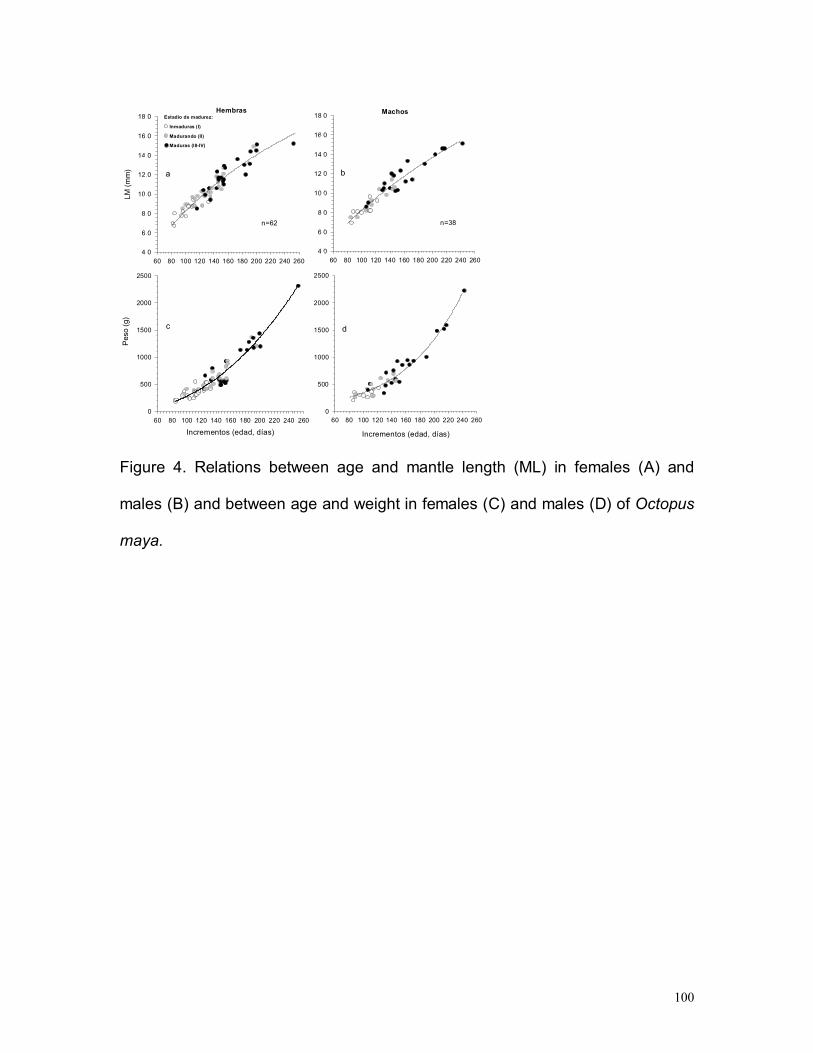
100
Hembras
A
Machos
B
60 80 100 120 140 160 180 200 220 240 260
LM (m
m)
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
60 80 100 120 140 160 180 200 220 240 2604.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 2600
500
1000
1500
2000
2500
a b
Incrementos (edad, días)60 80 100 120 140 160 180 200 220 240 260
Pes
o (g
)
0
500
1000
1500
2000
2500
c d
Estadio de madurez:
Inmaduras (I)
Madurando (II)
Maduras (III-IV)
n=62 n=38
Figure 4. Relations between age and mantle length (ML) in females (A) and
males (B) and between age and weight in females (C) and males (D) of Octopus
maya.
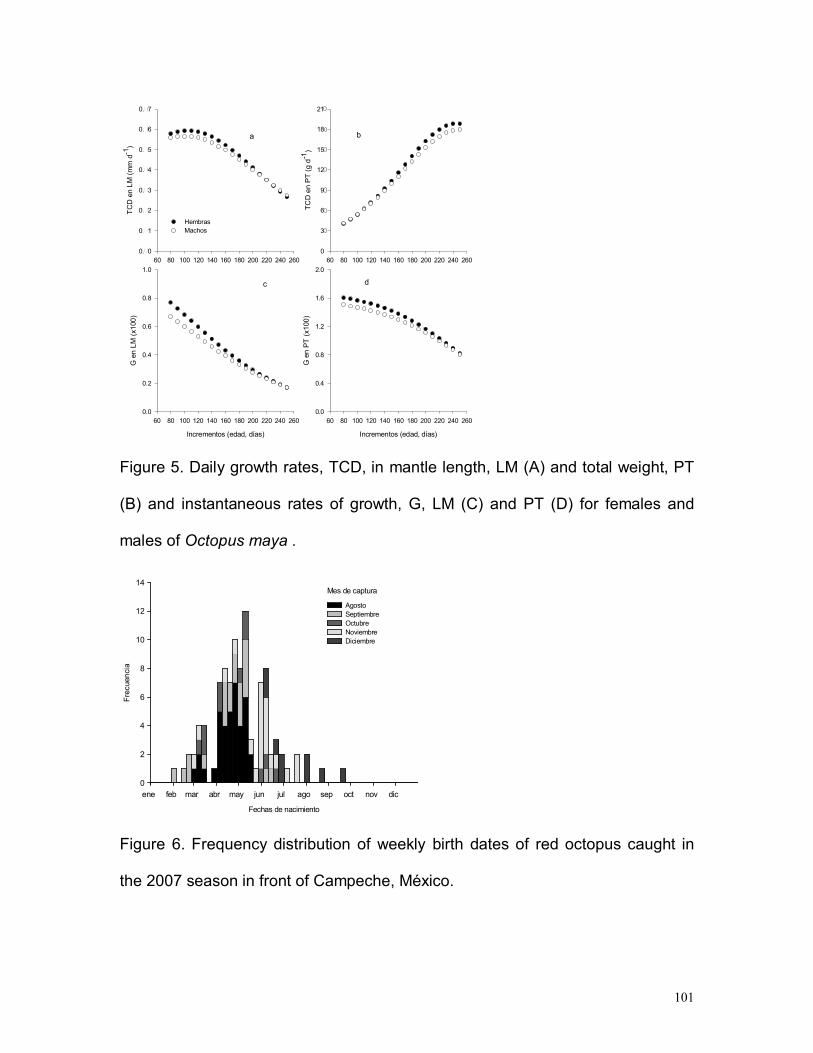
101
a
60 80 100 120 140 160 180 200 220 240 260
TCD
en
LM (m
m d
-1)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
HembrasMachos
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 260
G e
n LM
(x10
0)
0.0
0.2
0.4
0.6
0.8
1.060 80 100 120 140 160 180 200 220 240 260
TCD
en
PT (g
d-1
)
0
3
6
9
12
15
18
21
Incrementos (edad, días)
60 80 100 120 140 160 180 200 220 240 260
G e
n PT
(x10
0)
0.0
0.4
0.8
1.2
1.6
2.0
b
c d
Figure 5. Daily growth rates, TCD, in mantle length, LM (A) and total weight, PT
(B) and instantaneous rates of growth, G, LM (C) and PT (D) for females and
males of Octopus maya .
Fechas de nacimiento
ene feb mar abr may jun jul ago sep oct nov dic
Frec
uenc
ia
0
2
4
6
8
10
12
14
AgostoSeptiembreOctubreNoviembreDiciembre
Mes de captura
Figure 6. Frequency distribution of weekly birth dates of red octopus caught in
the 2007 season in front of Campeche, México.





























