Embed Size (px)
Citation preview

UNIDAD 2
Enzimología vegetal
Dra. María Inés Isla
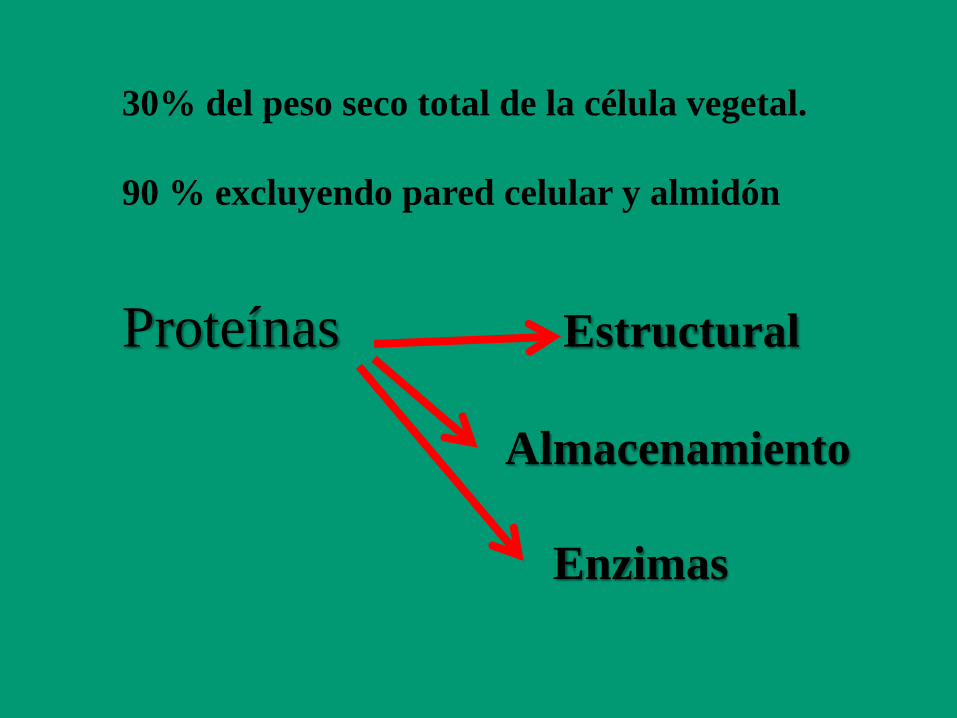
30% del peso seco total de la célula vegetal.
90 % excluyendo pared celular y almidón
Proteínas Estructural
Almacenamiento
Enzimas

A. Propiedades generales de las enzimas
B. Clasificación de las Enzimas
C. Principales problemas en el manejo de las Enzimas
D. Principios fundamentales de su acción catalítica
E. Introducción a la cinética enzimática
F. Enzimas reguladores
Enzimas

A) Propiedades generales de las enzimas
1. Son los catalizadores de las reacciones químicas en
los sistemas biológicos
2. Aceleran muchísimo la velocidad de las reacciones
(106 – 1014 veces).
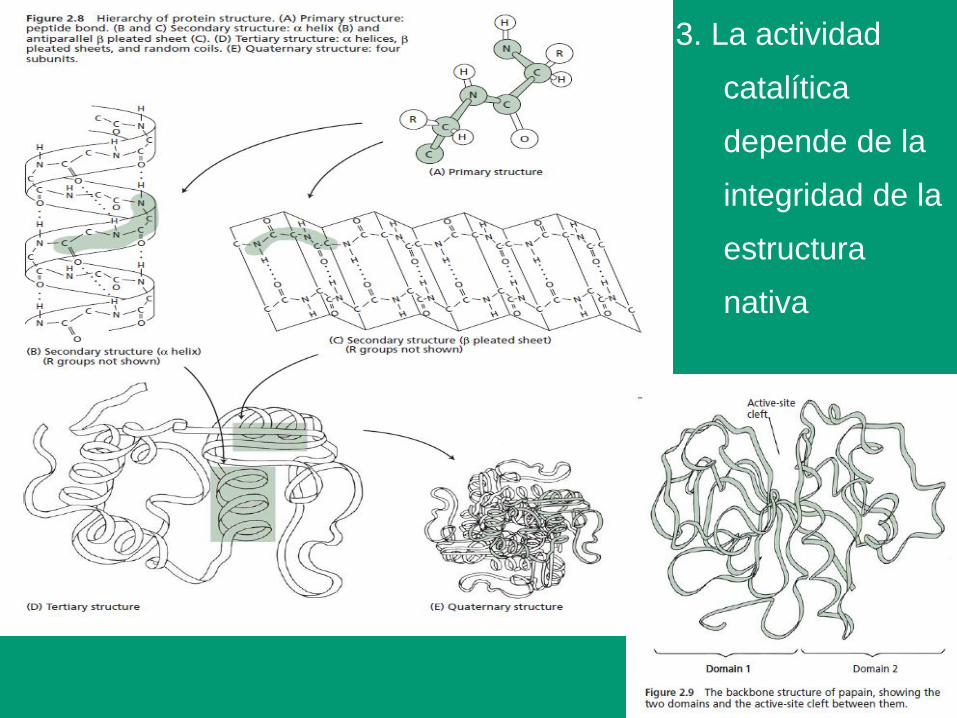
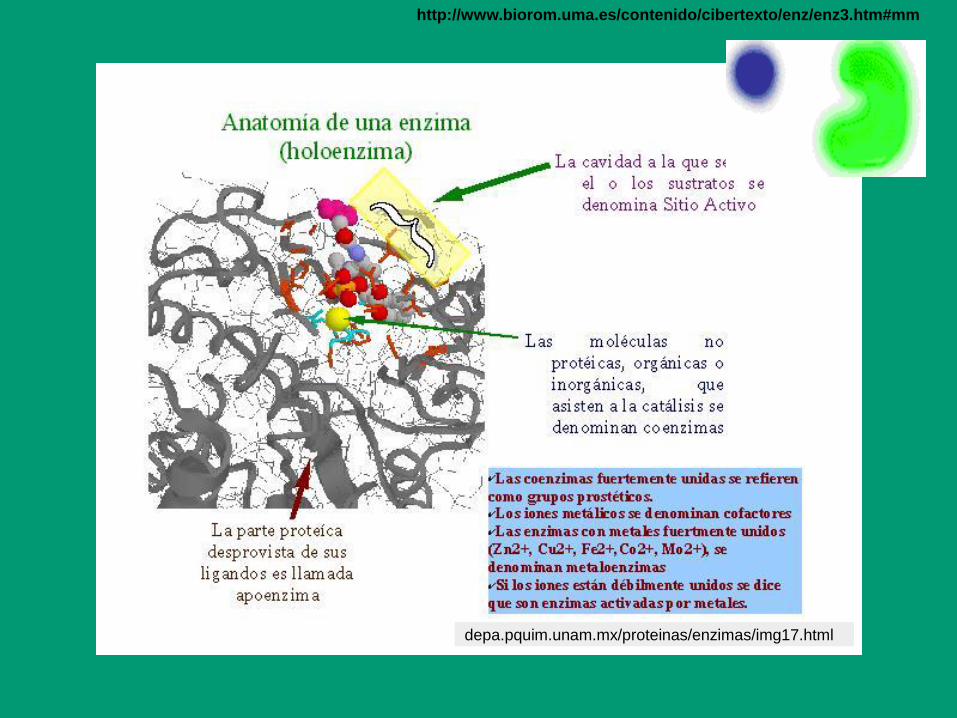
3. La actividad
catalítica
depende de la
integridad de la
estructura
nativa
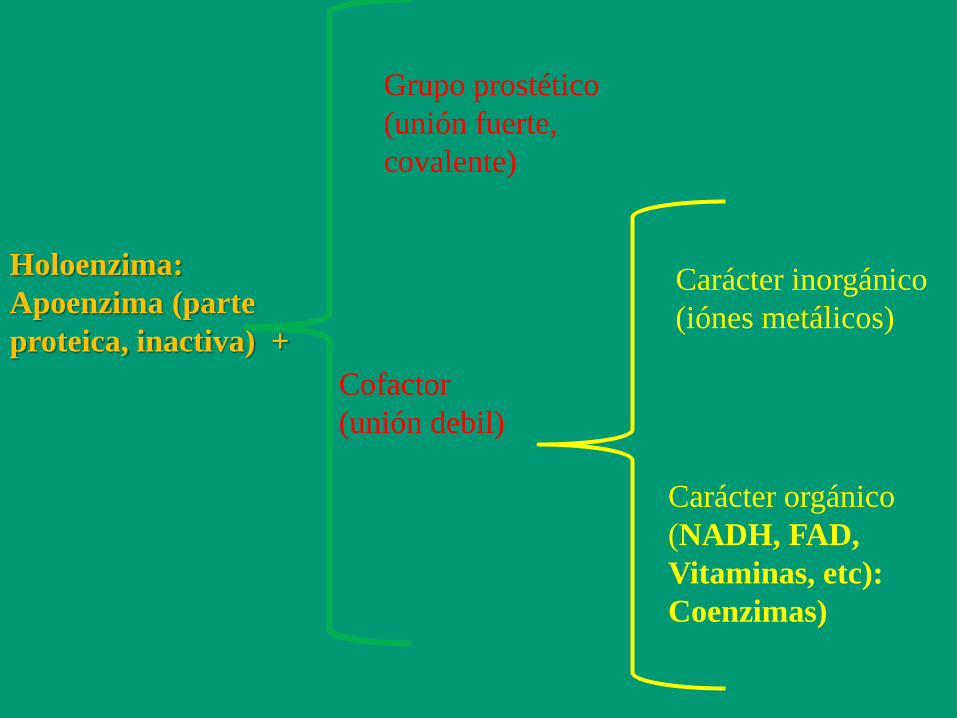
Holoenzima:
Apoenzima (parte
proteica, inactiva) +
Grupo prostético
(unión fuerte,
covalente)
Cofactor
(unión debil)
Carácter inorgánico
(iónes metálicos)
Carácter orgánico
(NADH, FAD,
Vitaminas, etc):
Coenzimas)
depa.pquim.unam.mx/proteinas/enzimas/img17.html
http://www.biorom.uma.es/contenido/cibertexto/enz/enz3.htm#mm
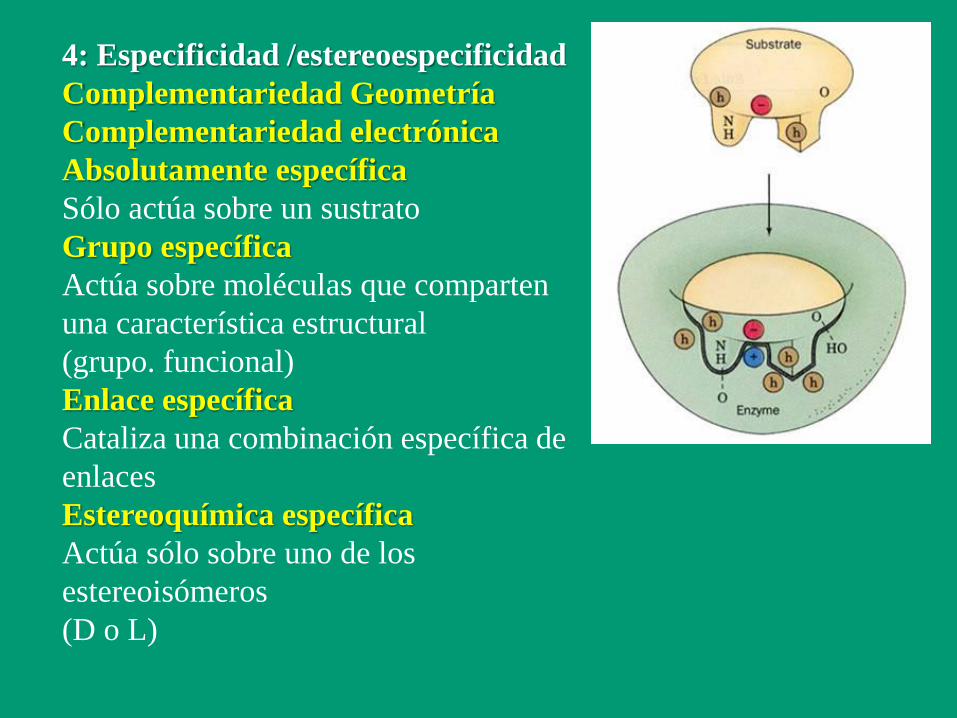
4: Especificidad /estereoespecificidad
Complementariedad Geometría
Complementariedad electrónica
Absolutamente específica
Sólo actúa sobre un sustrato
Grupo específica
Actúa sobre moléculas que comparten
una característica estructural
(grupo. funcional)
Enlace específica
Cataliza una combinación específica de
enlaces
Estereoquímica específica
Actúa sólo sobre uno de los
estereoisómeros
(D o L)

5. La ACTIVIDAD de las E depende no solo del
mantenimiento de su estructura nativa sino del pH,
temperatura
6. Control regulatorio:
Efecto sobre la actividad catalítica por un efector
(activador o inhibidor) hasta el control de la expresión
y el turnover de proteínas

A partir de 1982, se sabe que no todas las
enzimas son proteínas; existen moléculas de
RNA con actividad catalítica, las llamadas
ribozimas

Maduración del DNA
• Transcrito primario mRNA
ribozimas
1, 2, 3
1. Splicing ( corte y
empalme) eliminación de
intrones. Intrón de
autocorte y empalme
2. Introducción del
casquete en 5´
3. Introducción de una
cadena rica en adenina
(PoliA) en el extremo 3´.

A. Propiedades generales de las enzimas
B. Clasificación de las Enzimas
C. Principales problemas en el manejo de las Enzimas
D. Principios fundamentales de su acción catalítica
E. Introducción a la cinética enzimática
F. Enzimas reguladores
Enzimas

Las enzimas se clasifican según la reacción
catalizada: • Nomenclatura:
- Número clasificatorio de 4 dígitos (E.C.)
- Nombre sistemático
- Nombre trivial
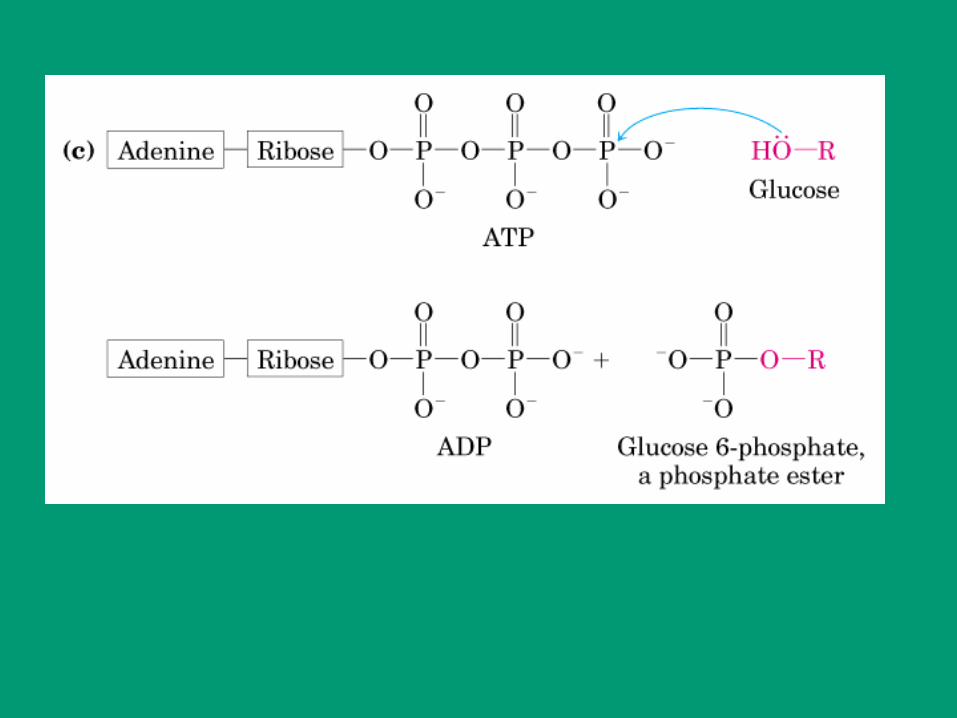
ATP + D-Glucosa ADP + D-Glucosa-fosfato
Número clasificatorio: E.C. 2.7.1.1.
2. Clase: Transferasa
7. Subclase: Fosfotransferasa
1. Fosfotransferasas con OH como aceptor
1. D-glucosa como aceptor del fosfato
Nombre sistemático: ATP:glucosa fosfotransferasa
Nombre trivial: hexoquinasa
Nomenclatura: se adiciona sufijo “asa” al nombre del sustrato o de la reacción que cataliza
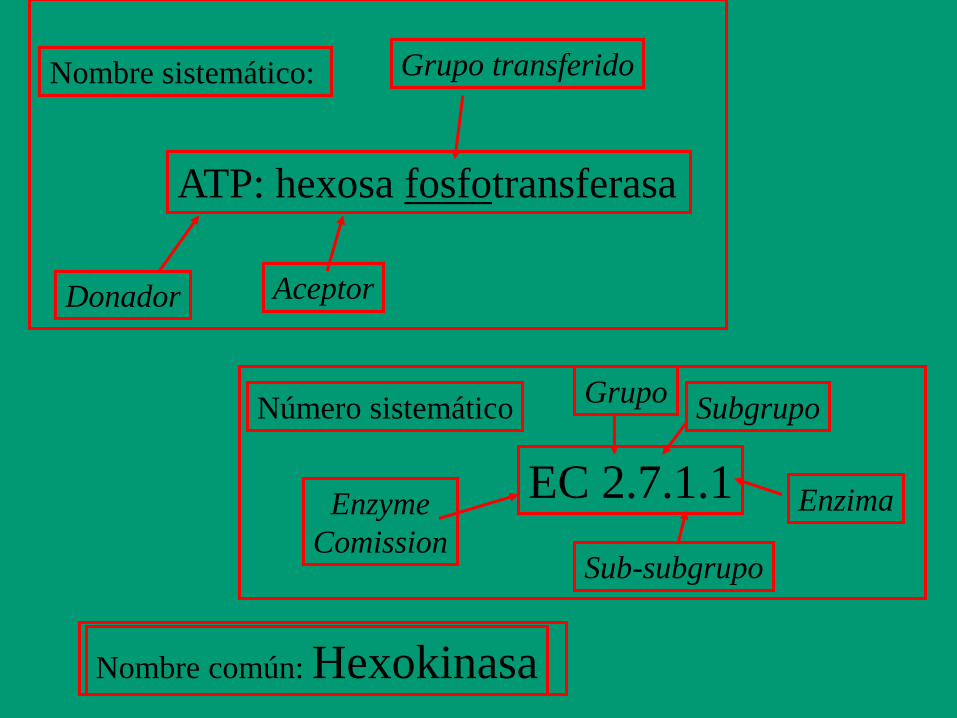
ATP: hexosa fosfotransferasa
Nombre sistemático:
Donador Aceptor
Grupo transferido
EC 2.7.1.1
Número sistemático
Enzyme
Comission
Grupo Subgrupo
Sub-subgrupo
Enzima
Nombre común: Hexokinasa

1. Oxidorreductasas
2. Transferasas
3. Hidrolasas
4. Liasas
5. Isomerasas
6. Ligasas
Clasificación de enzimas:
Grupos
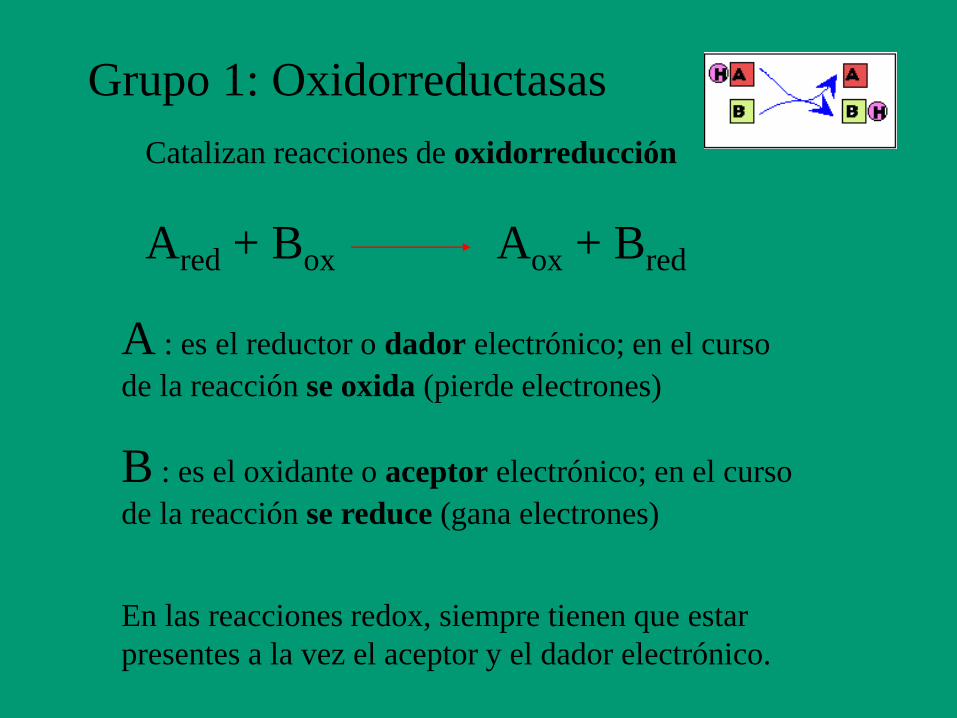
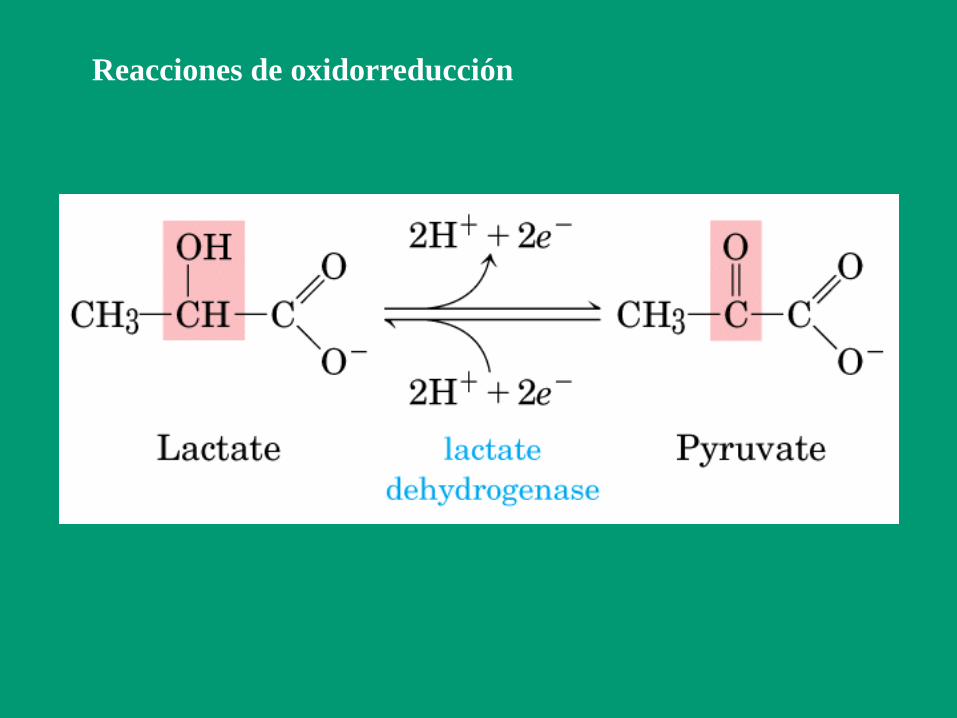
Grupo 1: Oxidorreductasas
Catalizan reacciones de oxidorreducción
En las reacciones redox, siempre tienen que estar
presentes a la vez el aceptor y el dador electrónico.
Ared + Box Aox + Bred
A : es el reductor o dador electrónico; en el curso
de la reacción se oxida (pierde electrones)
B : es el oxidante o aceptor electrónico; en el curso
de la reacción se reduce (gana electrones)

Dador: Aceptor oxidorreductasa
Glucosa : O2 oxidorreductasa
Dador Aceptor Nombre común:
Glucosa oxidasa
Los subgrupos se forman según la naturaleza del dador:
1.1.-.- Sobre grupos alcohol
1.2.-.- Sobre grupos aldehido
1.3.-.- Sobre grupos -CH-CH-
etc.
EC 1.1.3.4

Nomenclatura alternativa en oxidorreductasas:
1. Deshidrogenasas
2. Oxidasas
3. Peroxidasas
4. Oxigenasas
5. Hidroxilasas
6. Reductasas
Reacciones de oxidorreducción
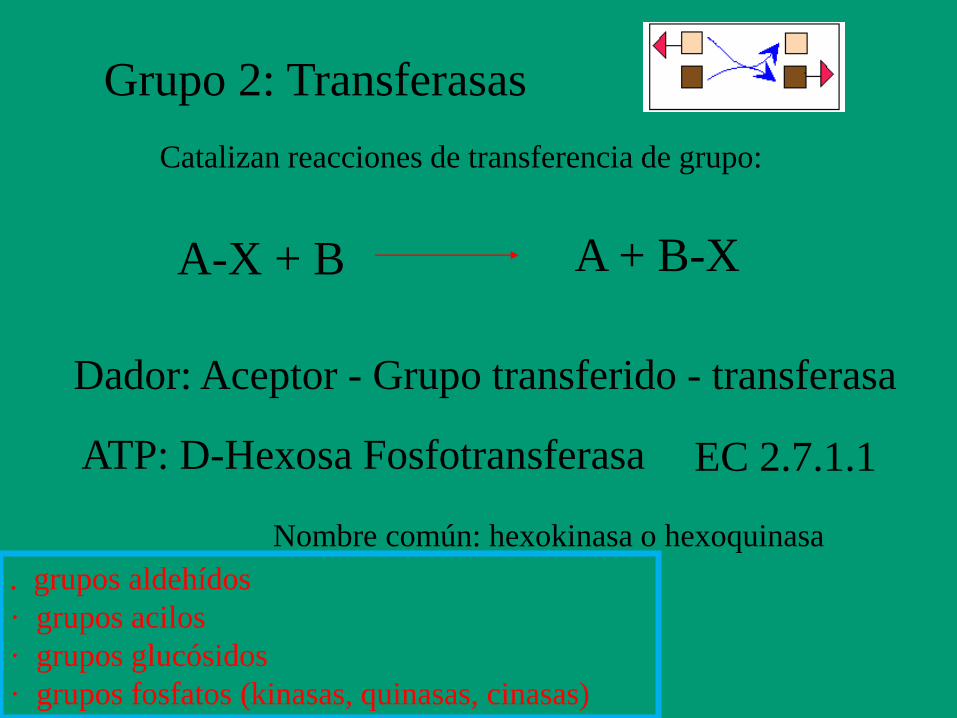
Grupo 2: Transferasas
Catalizan reacciones de transferencia de grupo:
A-X + B A + B-X
Dador: Aceptor - Grupo transferido - transferasa
ATP: D-Hexosa Fosfotransferasa EC 2.7.1.1
Nombre común: hexokinasa o hexoquinasa
. grupos aldehídos
· grupos acilos
· grupos glucósidos
· grupos fosfatos (kinasas, quinasas, cinasas)

Grupo 3: Hidrolasas
Catalizan reacciones de hidrólisis
A-B + H2O A-OH + H-B
No se suelen utilizar nombres sistemáticos en las
hidrolasas. Muchas de ellas conservan el nombre
primitivo: Tripsina, Pepsina, Papaína, etc.
Transforman polímeros en
monómeros.
Actúan sobre:
· enlace Ester
· enlace glucosídico
· enlace peptídico
· enlace C-N
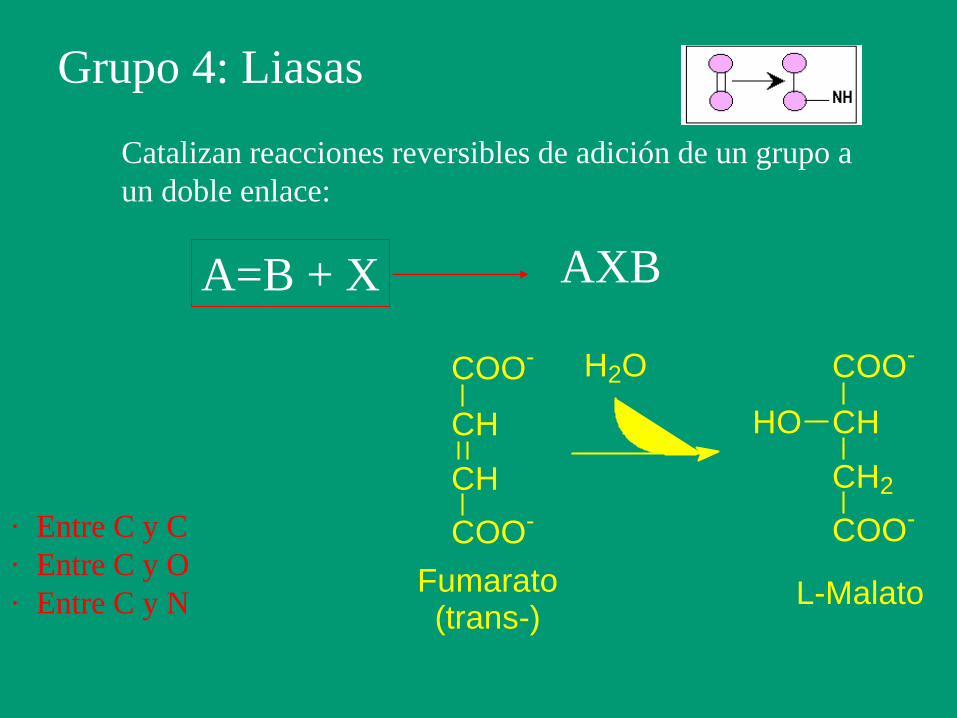
Grupo 4: Liasas
Catalizan reacciones reversibles de adición de un grupo a
un doble enlace:
A=B + X AXB
COO-
CH
CH
COO-
H2O COO-
CH
CH2
COO-
HO
Fumarato(trans-)
L-Malato
· Entre C y C
· Entre C y O
· Entre C y N
Grupo 5: Isomerasas
Catalizan reacciones de isomerización
Algunas reacciones isomerásicas:
- Racemasas
- Oxidorreductasas intramoleculares
- Mutasas o transferasas intramoleculares
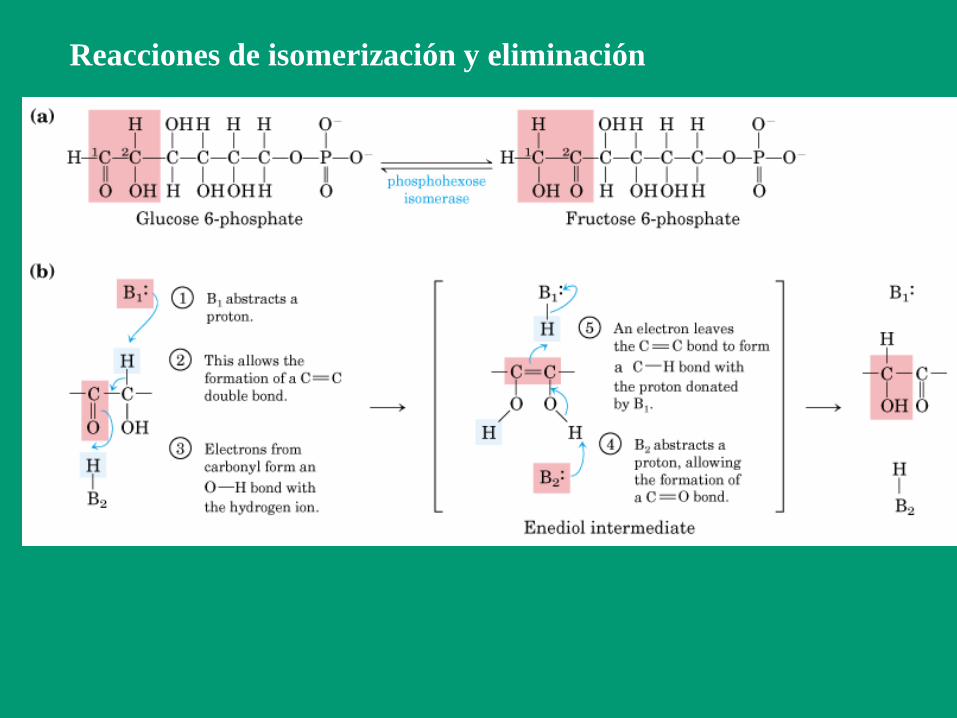
Reacciones de isomerización y eliminación

Grupo 6: Ligasas
Catalizan la unión de dos grupos químicos a expensas
de la hidrólisis de un enlace de alta energía.
A + B + ATP A-B + ADP + Pi
O bien
C + D + ATP C-D + AMP + PPi Entre C y O
· Entre C y S
· Entre C y N
· Entre C y C

A. Propiedades generales de las enzimas
B. Clasificación de las Enzimas
C. Principales problemas en el manejo de las Enzimas
(Extracción y purificación de Enzimas)
D. Principios fundamentales de su acción catalítica
E. Introducción a la cinética enzimática
F. Enzimas reguladores
Enzimas

EXISTEN DISTINTAS TÉCNICAS
QUE, COMBINADAS CONVENIENTEMENTE,
RESULTAN EN LA PURIFICACIÓN
DE UNAS DETERMINADAS PROTEÍNAS
Definir objetivos
• de pureza, actividad y cantidad requerida del producto final
• desarrollar un método
• Definir las propiedades de la proteína deseada
• simplificar las técnicas de selección y optimización
• Desarrollar ensayos analíticos
• para una rápida detección de la actividad/rendimiento de
nuestra proteína
• Minimizar el manejo de la muestra en cada paso y reducir el número de
pasos en el proceso de purificación
• evitar procedimientos largos que conllevan el riesgo de pérdida
de actividad/rendimiento
• Minimizar el uso de aditivos
• pueden interferir en ciertos ensayos y requeriría un paso extra
de purificación para eliminarlos
Eliminar contaminantes perjudiciales cuanto antes
• por ejemplo, proteasas, compuestos fenólicos, etc
Estabilizantes o conservantes de la actividad enzimática

ESTRATEGIA GENERAL DE PURIFICACIÓN
SE DEBE PROCURAR SEGUIR UN PROTOCOLO
LO MÁS SIMPLE Y SENCILLO POSIBLE

1. Obtención del extracto crudo
2. Fraccionamiento del extracto crudo por precipitación con sulfato de
amonio, polímeros (PEG), ácido poliacrílico, solventes, cambios de pH,
centrifugación, y diálisis
3. Cromatografía en columna
4. Electroforesis en gel:
•Electroforesis en poliacrilamida-SDS
• isoelectroenfoque
• Electroforesis bidimensional
PURIFICACIÓN Y ANÁLISIS DE PROTEÍNAS
Ejemplo
AFINIDAD (GST)
(columna)
GEL FILTRACIÓN
(columna)
INTERCAMBIO IÓNICO
(columna)
HIDROFÓBICA
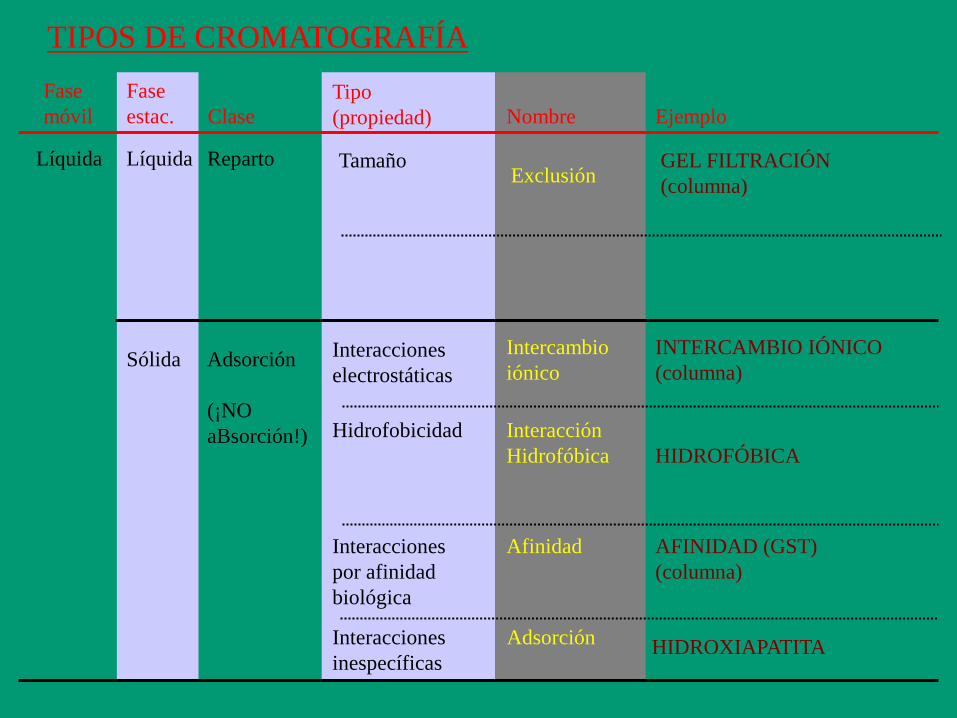
TIPOS DE CROMATOGRAFÍA
Interacciones
por afinidad
biológica
Fase
móvil
Fase
estac.
Líquida
Sólida
Tamaño
Adsorción
(¡NO
aBsorción!)
Líquida
Interacciones
electrostáticas
Nombre
Afinidad
Exclusión
Intercambio
iónico
Interacción
Hidrofóbica
Reparto
Hidrofobicidad
Interacciones
inespecíficas
Clase
Tipo
(propiedad)
Adsorción HIDROXIAPATITA
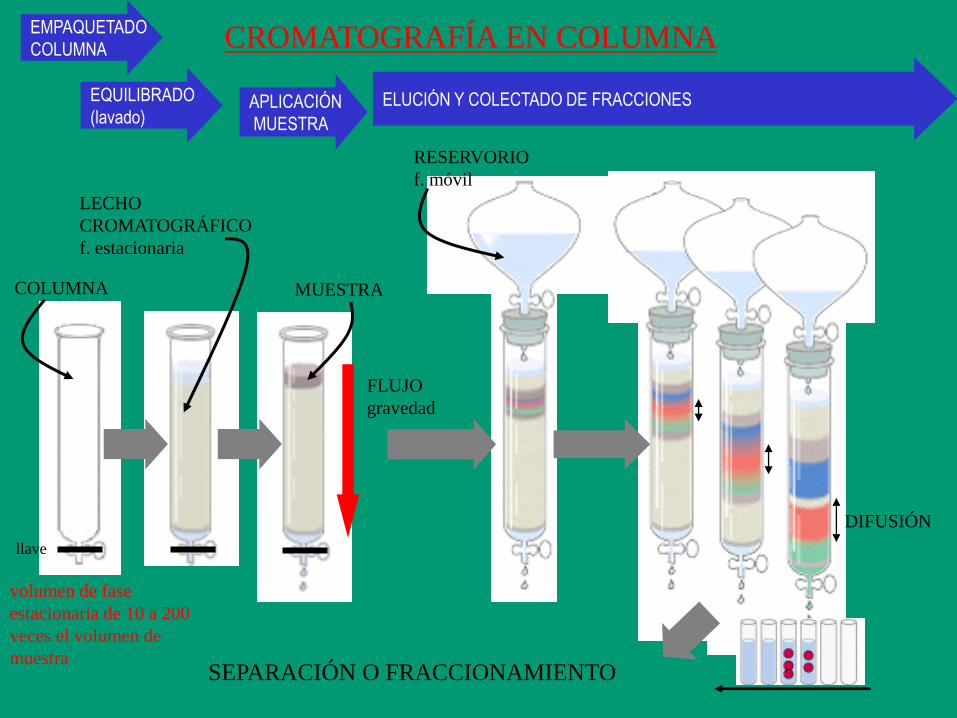
CROMATOGRAFÍA EN COLUMNA
LECHO
CROMATOGRÁFICO
f. estacionaria
EMPAQUETADO
COLUMNA
COLUMNA
llave
MUESTRA
APLICACIÓN
MUESTRA
RESERVORIO
f. móvil
ELUCIÓN Y COLECTADO DE FRACCIONES
FLUJO
gravedad
DIFUSIÓN
SEPARACIÓN O FRACCIONAMIENTO
volumen de fase
estacionaria de 10 a 200
veces el volumen de
muestra
EQUILIBRADO
(lavado)
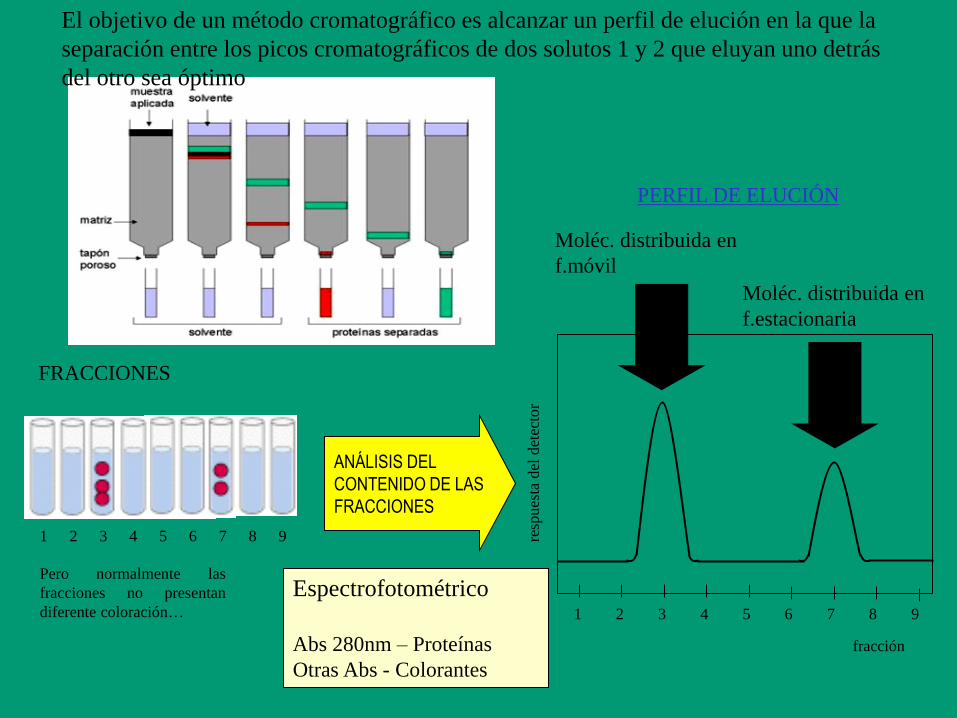
Moléc. distribuida en
f.móvil
Moléc. distribuida en
f.estacionaria
PERFIL DE ELUCIÓN
ANÁLISIS DEL
CONTENIDO DE LAS
FRACCIONES
1 2 4 5 6 7 8 9 3
resp
ues
ta d
el d
etec
tor
fracción
Espectrofotométrico
Abs 280nm – Proteínas
Otras Abs - Colorantes
FRACCIONES
1 2 4 5 6 7 8 9 3
Pero normalmente las
fracciones no presentan
diferente coloración…
El objetivo de un método cromatográfico es alcanzar un perfil de elución en la que la
separación entre los picos cromatográficos de dos solutos 1 y 2 que eluyan uno detrás
del otro sea óptimo

PROPIEDAD: Tamaño (y forma)
CROMATOGRAFÍA DE FILTRACIÓN POR GEL, DE
EXCLUSION POR TAMAÑO, DE TAMIZADO
MOLECULAR
MICROPARTÍCULAS
FASE MÓVIL
SOPORTE Resina de micropartículas esféricas porosas formadas
por polímeros hidrofílicos (dextrano, agarosa, poliacrilamida o mezcla
de ellos). Diámetro de poro determinado.
Tamaños de poro intervalo o rango de fraccionamiento.
P.e. Sephadex G25: 1000 – 5000 Da
Sephadex G200: 5000 – 250000 Da
F. MÓVIL (LÍQUIDA) solvente acuoso que atraviesa la columna
por los espacios intersticiales (entre las micropartículas).
F. ESTACIONARIA (LÍQUIDA) solvente acuoso (el mismo
que el de la fase móvil) que se encuentra dentro de micropartículas.
F. ESTACIONARIA
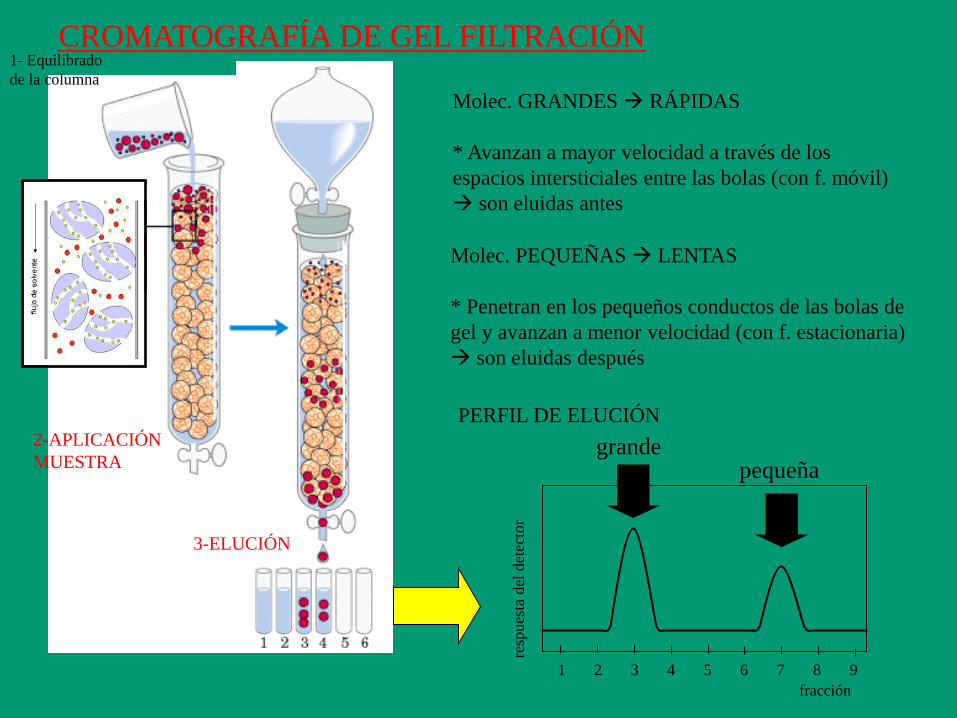
1 2 4 5 6 7 8 9 3
resp
ues
ta d
el d
etec
tor
fracción
PERFIL DE ELUCIÓN
CROMATOGRAFÍA DE GEL FILTRACIÓN
Molec. GRANDES RÁPIDAS
* Avanzan a mayor velocidad a través de los
espacios intersticiales entre las bolas (con f. móvil)
son eluidas antes
grande
Molec. PEQUEÑAS LENTAS
* Penetran en los pequeños conductos de las bolas de
gel y avanzan a menor velocidad (con f. estacionaria)
son eluidas después
pequeña
2-APLICACIÓN
MUESTRA
3-ELUCIÓN
1- Equilibrado
de la columna
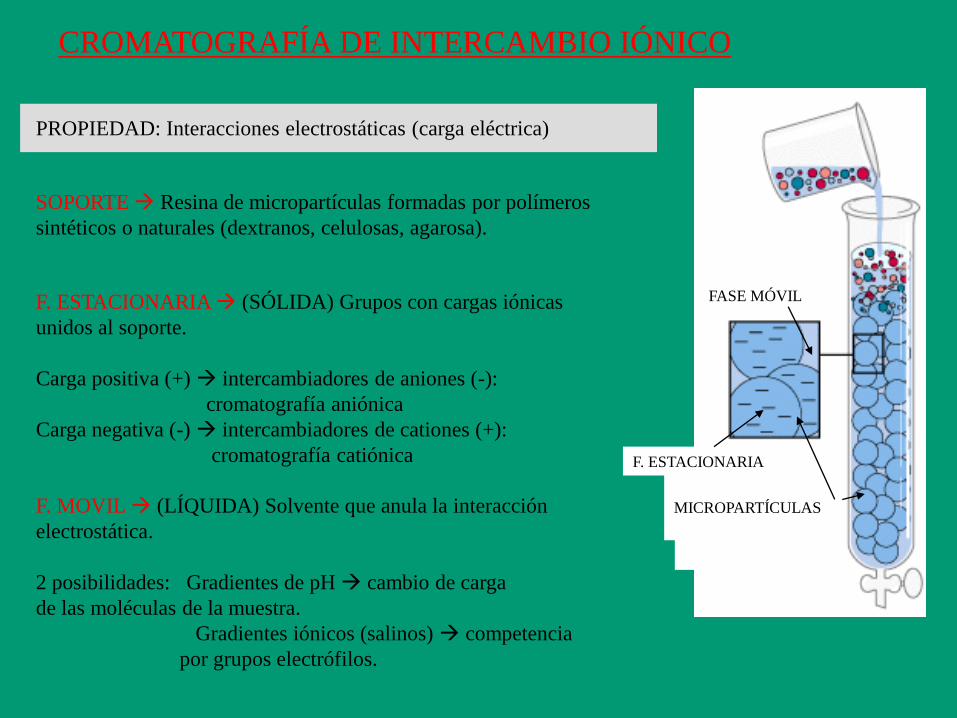
PROPIEDAD: Interacciones electrostáticas (carga eléctrica)
CROMATOGRAFÍA DE INTERCAMBIO IÓNICO
MICROPARTÍCULAS
FASE MÓVIL
SOPORTE Resina de micropartículas formadas por polímeros
sintéticos o naturales (dextranos, celulosas, agarosa).
F. MOVIL (LÍQUIDA) Solvente que anula la interacción
electrostática.
2 posibilidades: Gradientes de pH cambio de carga
de las moléculas de la muestra.
Gradientes iónicos (salinos) competencia
por grupos electrófilos.
F. ESTACIONARIA (SÓLIDA) Grupos con cargas iónicas
unidos al soporte.
Carga positiva (+) intercambiadores de aniones (-):
cromatografía aniónica
Carga negativa (-) intercambiadores de cationes (+):
cromatografía catiónica F. ESTACIONARIA
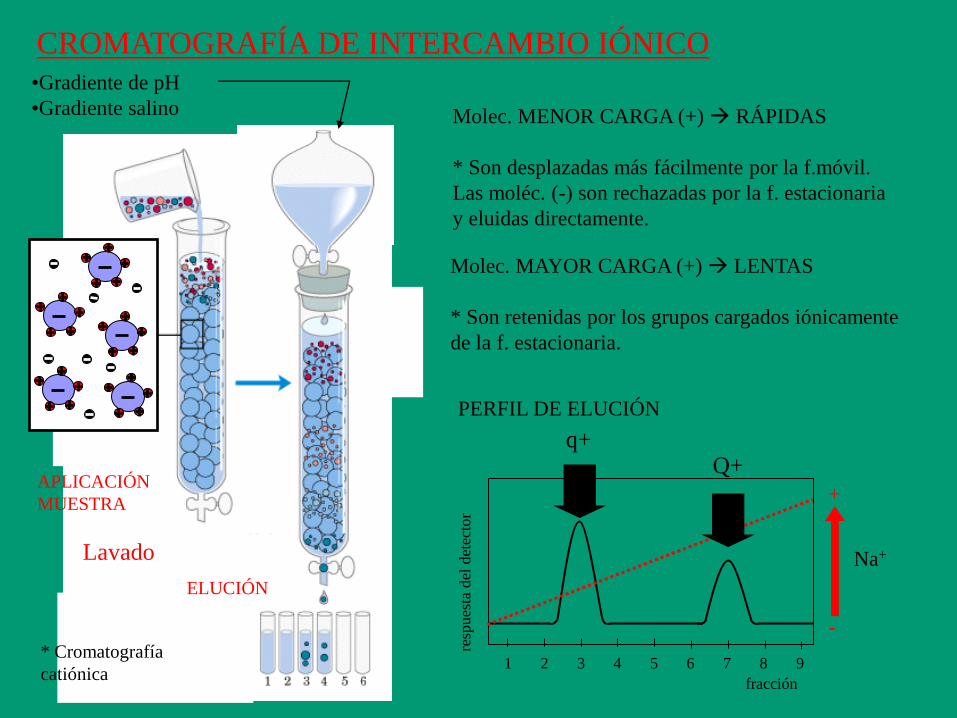
CROMATOGRAFÍA DE INTERCAMBIO IÓNICO
Molec. MENOR CARGA (+) RÁPIDAS
* Son desplazadas más fácilmente por la f.móvil.
Las moléc. (-) son rechazadas por la f. estacionaria
y eluidas directamente.
Molec. MAYOR CARGA (+) LENTAS
* Son retenidas por los grupos cargados iónicamente
de la f. estacionaria.
•Gradiente de pH
•Gradiente salino
1 2 4 5 6 7 8 9 3
resp
ues
ta d
el d
etec
tor
fracción
PERFIL DE ELUCIÓN
+
-
Na+
q+ Q+
APLICACIÓN
MUESTRA
ELUCIÓN
* Cromatografía
catiónica
Lavado

CROMATOGRAFÍA DE AFINIDAD
PROPIEDAD: Interacciones específicas entre dos moléculas
P.e. hormona-receptor
proteína-metal o cofactores como ATP
antígeno-anticuerpo
F. ESTACIONARIA
MICROPARTÍCULAS
F. ESTACIONARIA (SÓLIDA) Ligandos (grupos químicos)
específicos (con afinidad por alguna de las moléculas a aislar)
unidos a la matriz.
-- Comerciales: Concavalina A une glicoproteínas
Proteína A une IgG (anticuerpos)
-- No comerciales: anclaje covalente ligando - matriz
SOPORTE Matriz de micropartículas hidrofílicas sin carga, con
rigidez, inerte y estable (agarosa, acrílica).
F. MÓVIL (LÍQUIDA) Solución que anula o disminuye la afinidad
con el ligando de la fase estacionaria.
-- Solución con el propio ligando Molécula + ligando libre
-- Solución con algo que interacciona con el ligando Molécula
FASE MÓVIL
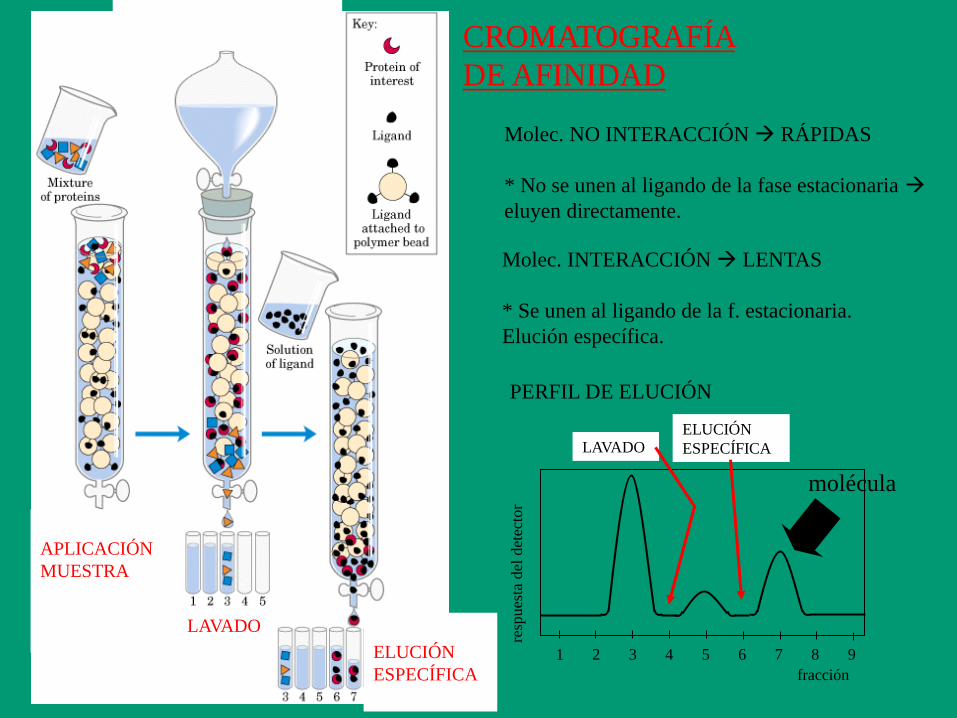
fracción
Molec. NO INTERACCIÓN RÁPIDAS
* No se unen al ligando de la fase estacionaria
eluyen directamente.
Molec. INTERACCIÓN LENTAS
* Se unen al ligando de la f. estacionaria.
Elución específica.
APLICACIÓN
MUESTRA
LAVADO
ELUCIÓN
ESPECÍFICA
1 2 4 5 6 7 8 9 3
resp
ues
ta d
el d
etec
tor
PERFIL DE ELUCIÓN
LAVADO ELUCIÓN
ESPECÍFICA
molécula
CROMATOGRAFÍA
DE AFINIDAD
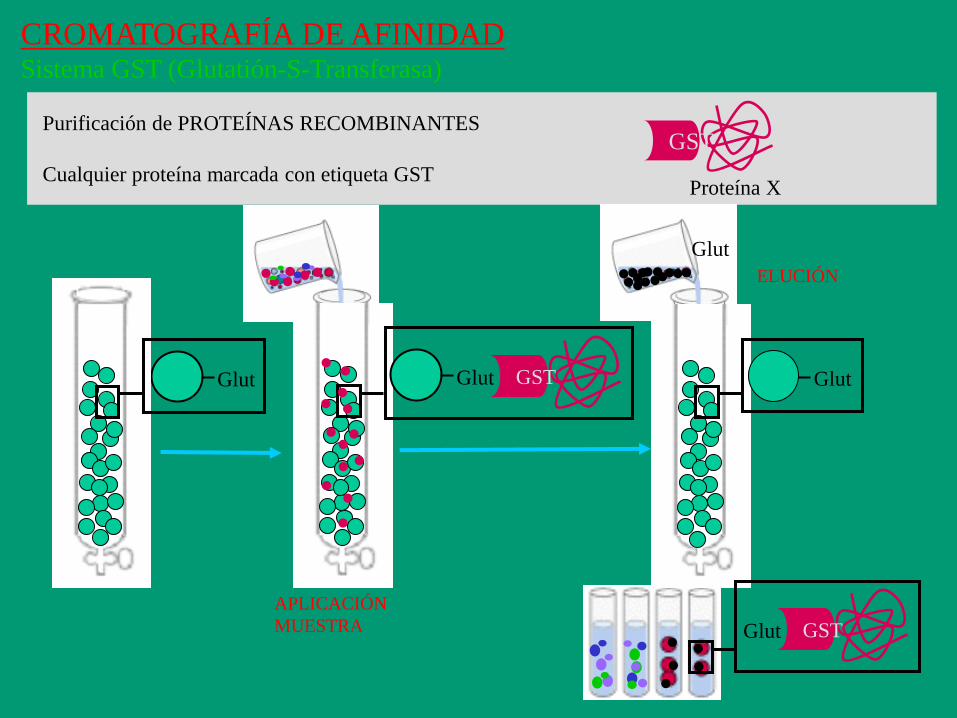
CROMATOGRAFÍA DE AFINIDAD Sistema GST (Glutatión-S-Transferasa)
GST
Proteína X
Purificación de PROTEÍNAS RECOMBINANTES
Cualquier proteína marcada con etiqueta GST
Glut GST Glut
GST Glut
Glut
APLICACIÓN
MUESTRA
ELUCIÓN
Glut
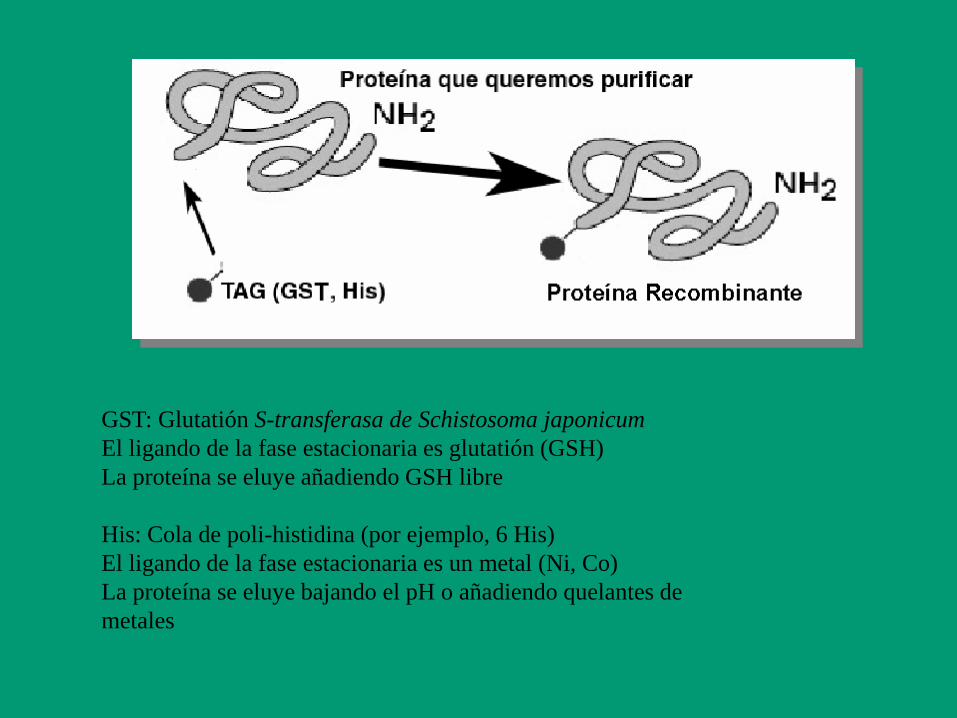
GST: Glutatión S-transferasa de Schistosoma japonicum
El ligando de la fase estacionaria es glutatión (GSH)
La proteína se eluye añadiendo GSH libre
His: Cola de poli-histidina (por ejemplo, 6 His)
El ligando de la fase estacionaria es un metal (Ni, Co)
La proteína se eluye bajando el pH o añadiendo quelantes de
metales
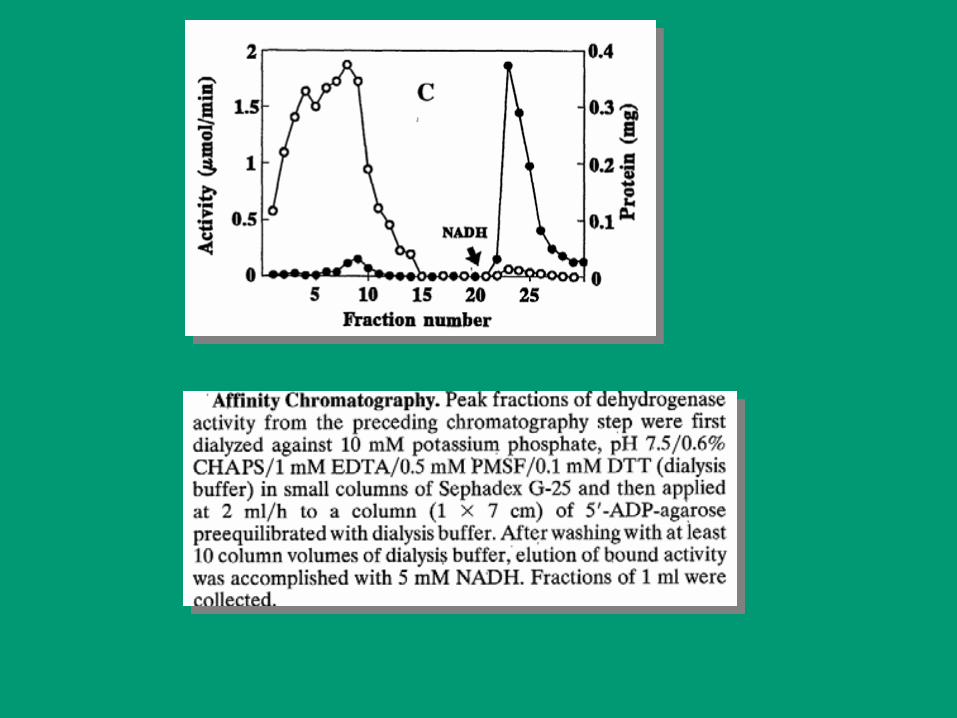


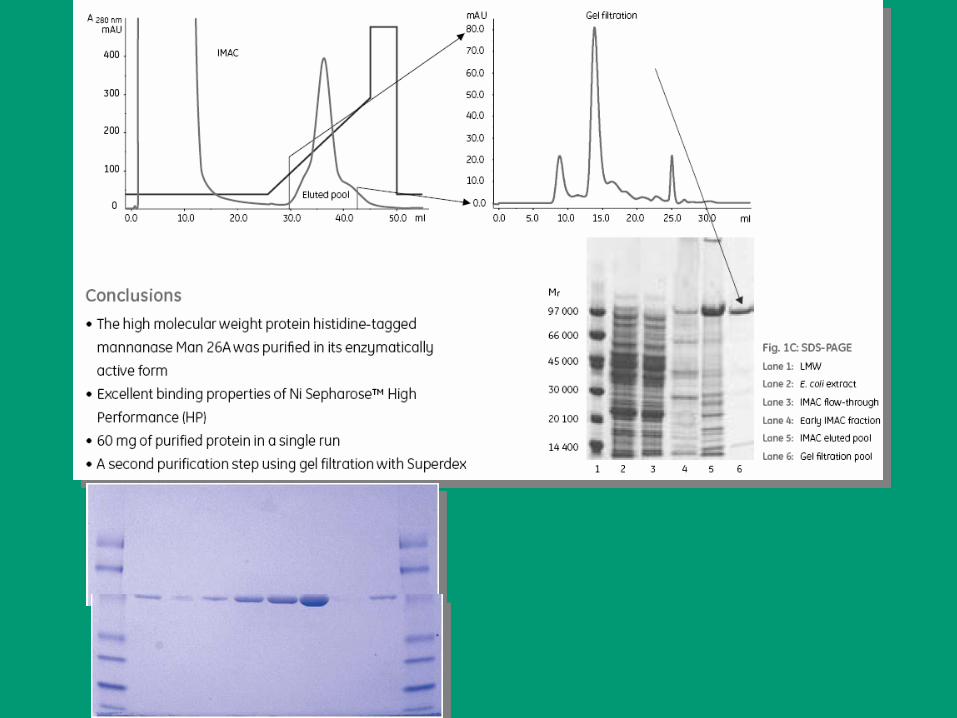
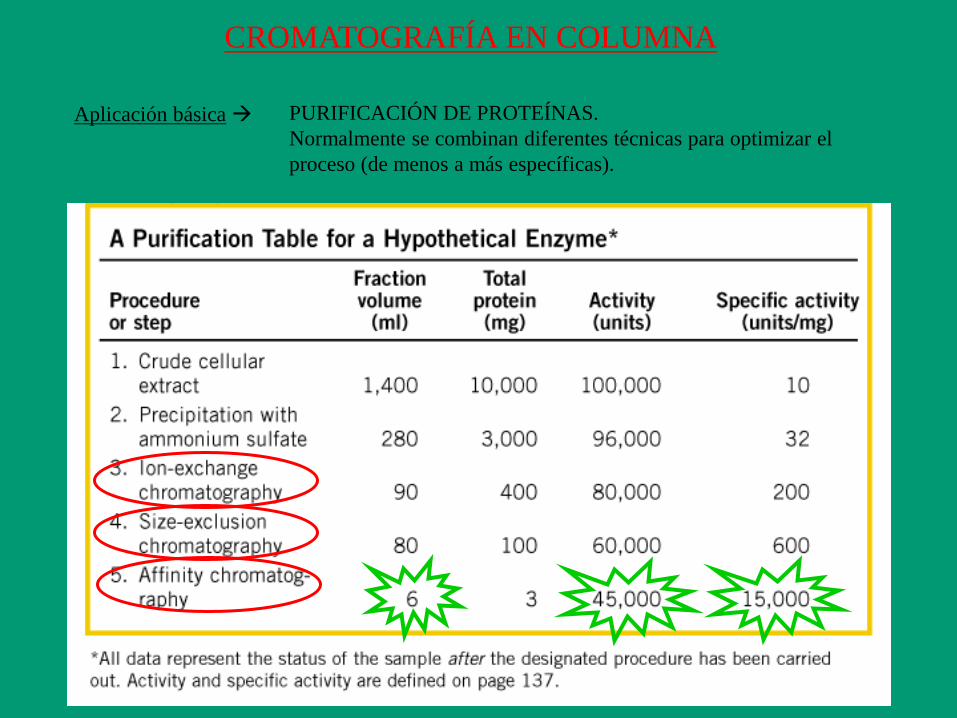
CROMATOGRAFÍA EN COLUMNA
Aplicación básica PURIFICACIÓN DE PROTEÍNAS.
Normalmente se combinan diferentes técnicas para optimizar el
proceso (de menos a más específicas).

Unidad de enzima: Se define como la cantidad de E que cataliza la
transformación de 1 µmol de sustrato por minuto bajo condiciones
definidas de trabajo (pH, T, ¨concent de S y cofactores)
Actividad específica: La actividad específica está dada por el
cociente entre el numero de unidades de enzima y los mg de proteína
del preparado.

A. Propiedades generales de las enzimas
B. Clasificación de las Enzimas
C. Principales problemas en el manejo de las Enzimas
D. Principios fundamentales de su acción catalítica
E. Introducción a la cinética enzimática
F. Enzimas reguladores
Enzimas

Principios fundamentales de la acción
catalítica de las enzimas
¿ cómo funcionan las enzimas ?
• En condiciones fisiológicas las reacciones sin catalizar
tienden a ser lentas (estabilidad de biomoléculas)
• Muchas reacciones bioquímicas suponen situaciones
poco probables
• Una enzima soluciona estos problemas proporcionando
un ambiente tridimensional favorable a la reacción (sitio
activo)
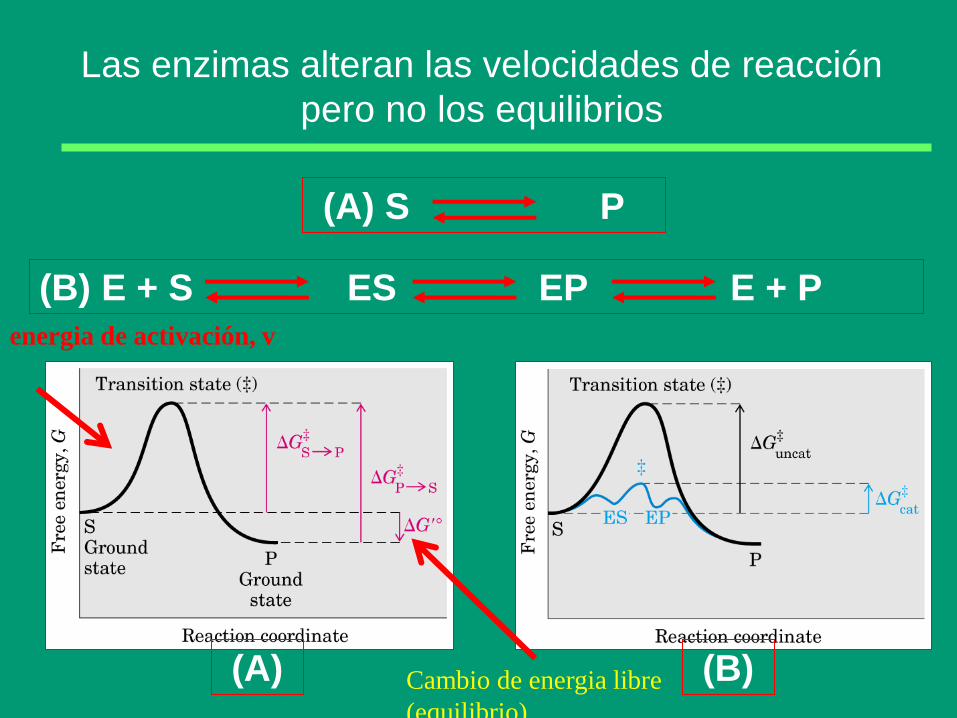
Las enzimas alteran las velocidades de reacción
pero no los equilibrios
(B) E + S ES EP E + P
(A) S P
(A) (B) Cambio de energia libre
(equilibrio)
energia de activación, v

SITIO ACTIVO:
Grupos catalíticos: (the reactive side chains of amino acids
or cofactors,which carry out the bond-breaking and bond-forming
reactions involved)
Sitio de ligamiento: attracts and positions the substrate
Interacciones no covalentes
Cambios
conformacionales
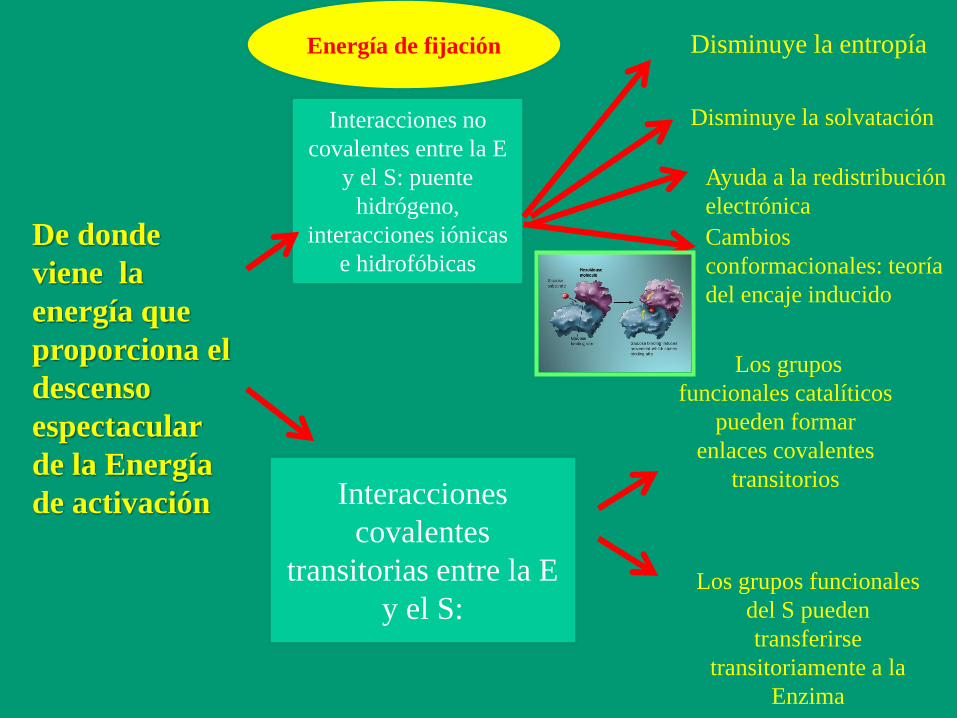
De donde
viene la
energía que
proporciona el
descenso
espectacular
de la Energía
de activación
Interacciones no
covalentes entre la E
y el S: puente
hidrógeno,
interacciones iónicas
e hidrofóbicas
Interacciones
covalentes
transitorias entre la E
y el S:
Energía de fijación Disminuye la entropía
Disminuye la solvatación
Ayuda a la redistribución
electrónica
Cambios
conformacionales: teoría
del encaje inducido
Los grupos
funcionales catalíticos
pueden formar
enlaces covalentes
transitorios
Los grupos funcionales
del S pueden
transferirse
transitoriamente a la
Enzima
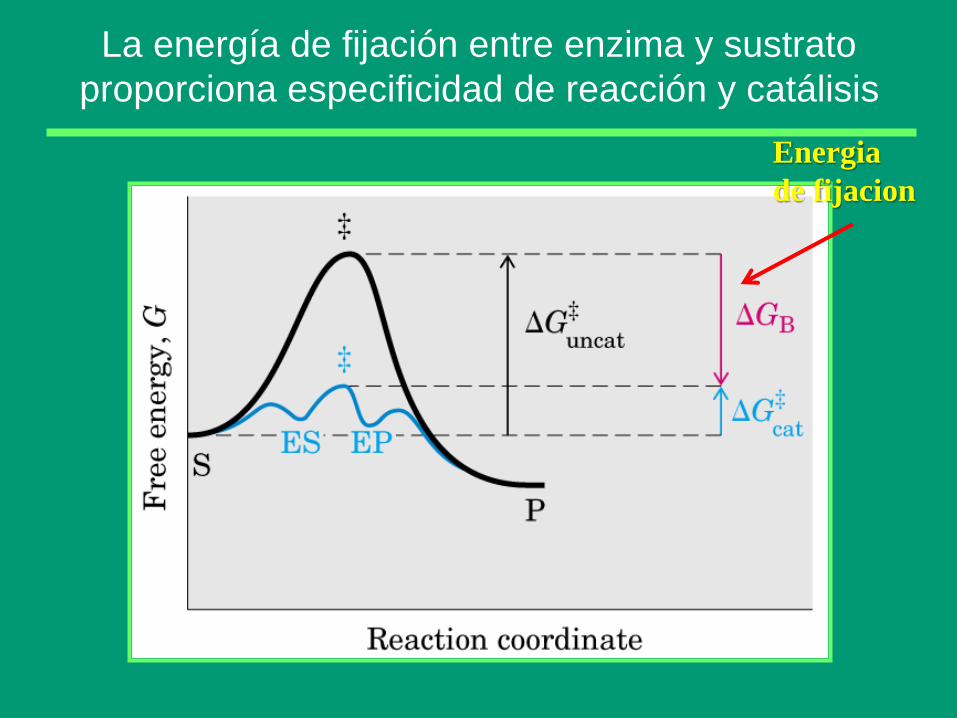
La energía de fijación entre enzima y sustrato
proporciona especificidad de reacción y catálisis
Energia
de fijacion
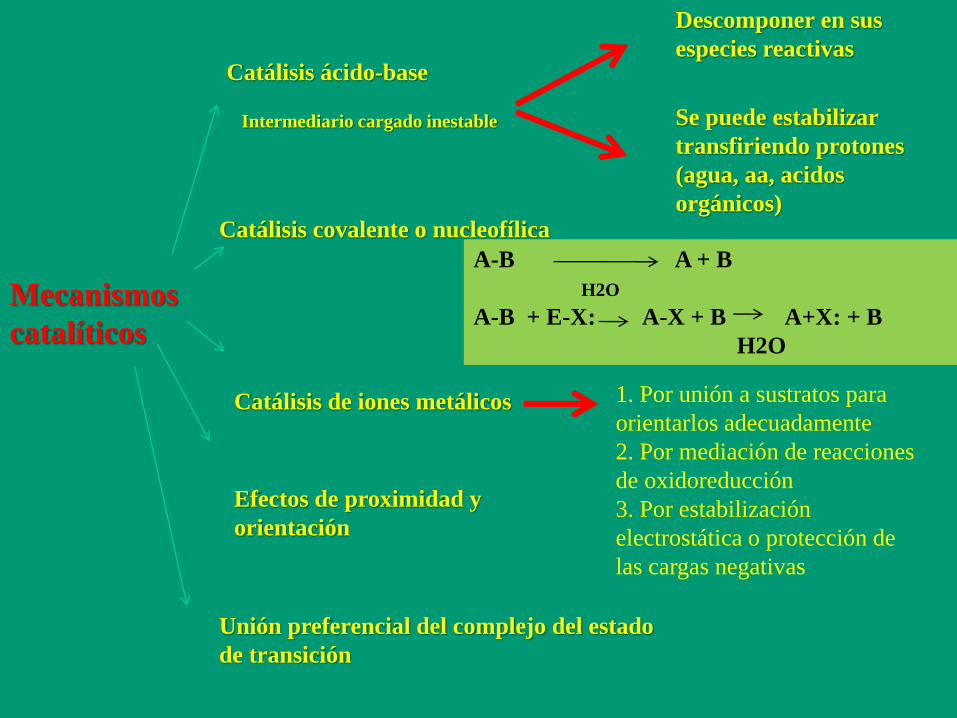
Mecanismos
catalíticos
Catálisis ácido-base
Catálisis covalente o nucleofílica
Catálisis de iones metálicos
Efectos de proximidad y
orientación
Unión preferencial del complejo del estado
de transición
Intermediario cargado inestable
Descomponer en sus
especies reactivas
Se puede estabilizar
transfiriendo protones
(agua, aa, acidos
orgánicos)
A-B A + B
H2O
A-B + E-X: A-X + B A+X: + B
H2O
1. Por unión a sustratos para
orientarlos adecuadamente
2. Por mediación de reacciones
de oxidoreducción
3. Por estabilización
electrostática o protección de
las cargas negativas

* Estos compuestos pueden formarse por donación de protones desde
restos aminoacídicos ácidos del centro activo de la enzima, tales como:
- grupos carboxílicos (-COOH) Glu, Asp
- grupos imidazólicos protonados His
- grupos fenólicos (Ph-OH) Tyr
- grupos guanidinios protonados Arg
- grupos aminos protonados (-NH3+) Lys
Estos grupos pueden convertirse potencialmente en donadores de
protones en una catálisis ácida.
- grupos carboxilato (-COO-) Glu, Asp
- grupo imidazol His
- grupo guanidino Arg
- Grupo amino (-NH2) Lys
Por otra parte:
Pueden servir de aceptores de protones en una catálisis básica.
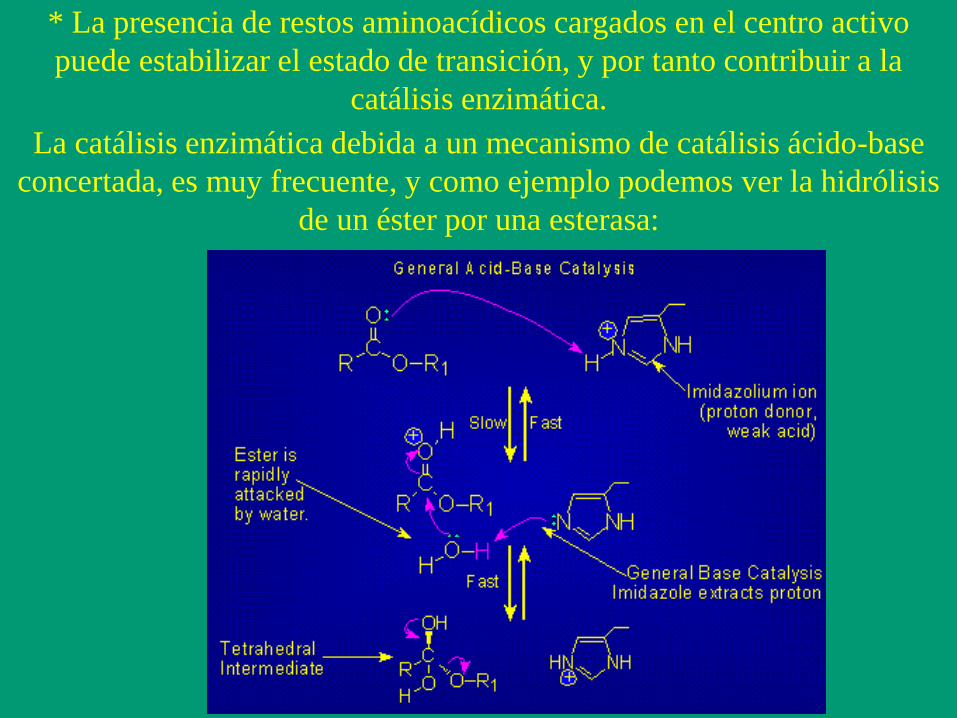
* La presencia de restos aminoacídicos cargados en el centro activo
puede estabilizar el estado de transición, y por tanto contribuir a la
catálisis enzimática.
La catálisis enzimática debida a un mecanismo de catálisis ácido-base
concertada, es muy frecuente, y como ejemplo podemos ver la hidrólisis
de un éster por una esterasa:
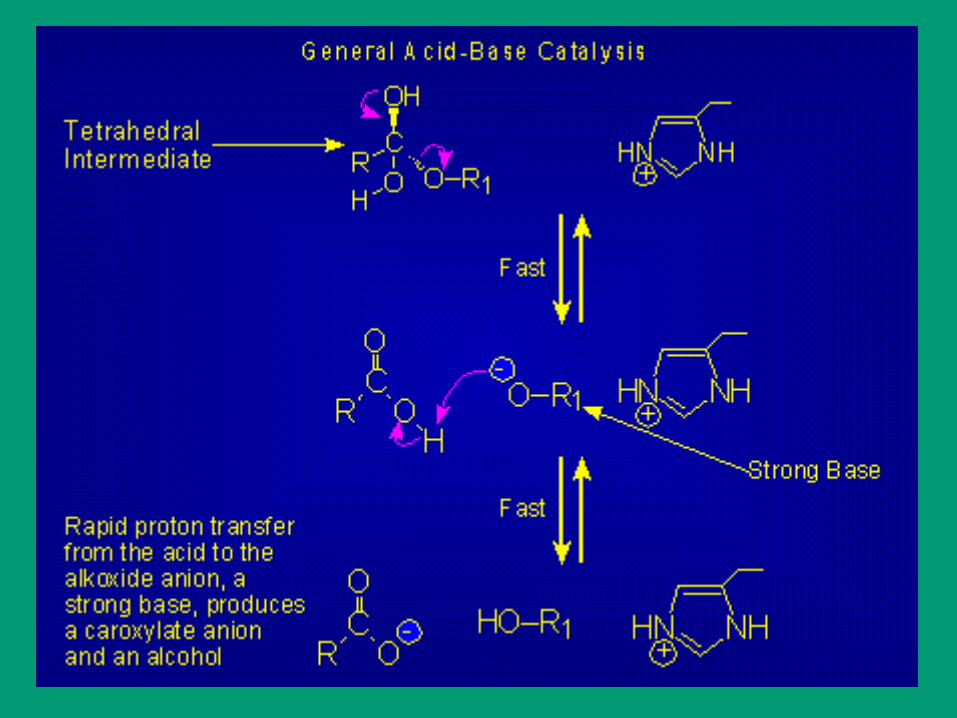
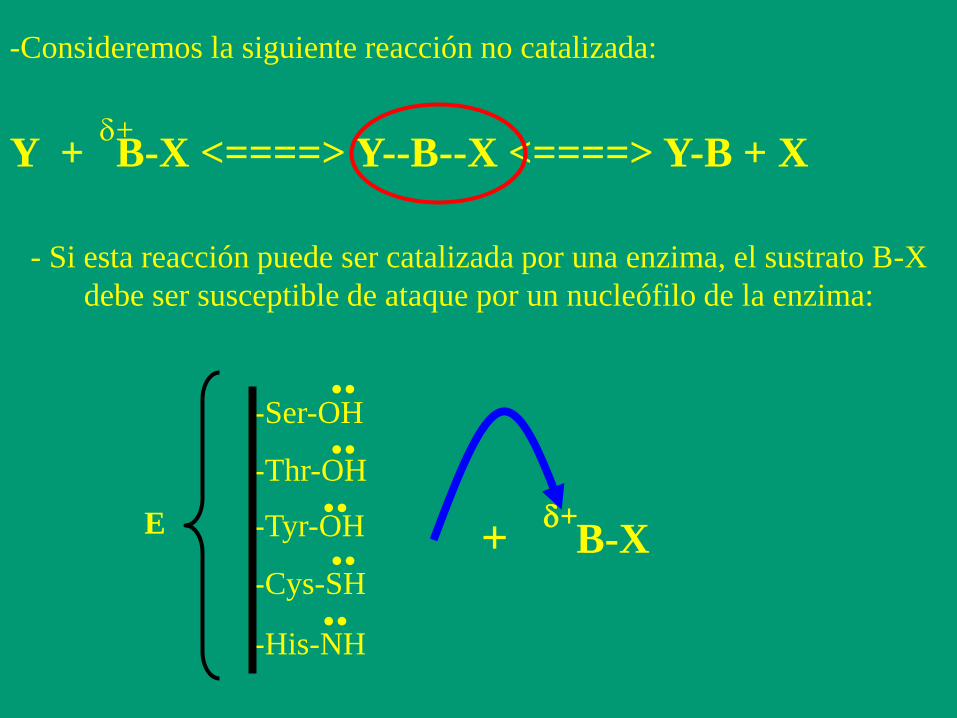
-Consideremos la siguiente reacción no catalizada:
Y + B-X <====> Y--B--X <====> Y-B + X d+
- Si esta reacción puede ser catalizada por una enzima, el sustrato B-X
debe ser susceptible de ataque por un nucleófilo de la enzima:
..
-His-NH
-Ser-OH
-Tyr-OH
-Thr-OH
-Cys-SH
E ..
..
.. ..
B-X d+
+

Para que haya un aumento de la velocidad de reacción, la enzima debe
ser un buen nucleófilo, y también un buen grupo saliente.
Así pues, en presencia de la enzima:
E-N: + B-X <==> X + E-N-B + Y <==> Y-B + E-N: d+
La estabilidad del compuesto intermedio E-N-B nos dice que N: , es
……..
- mejor nucleófilo que Y
- mejor grupo saliente que X
- Existe un elevado número de reacciones enzimáticas que transcurren a
través de catálisis covalente
E-N: (grupo nucleofílico)
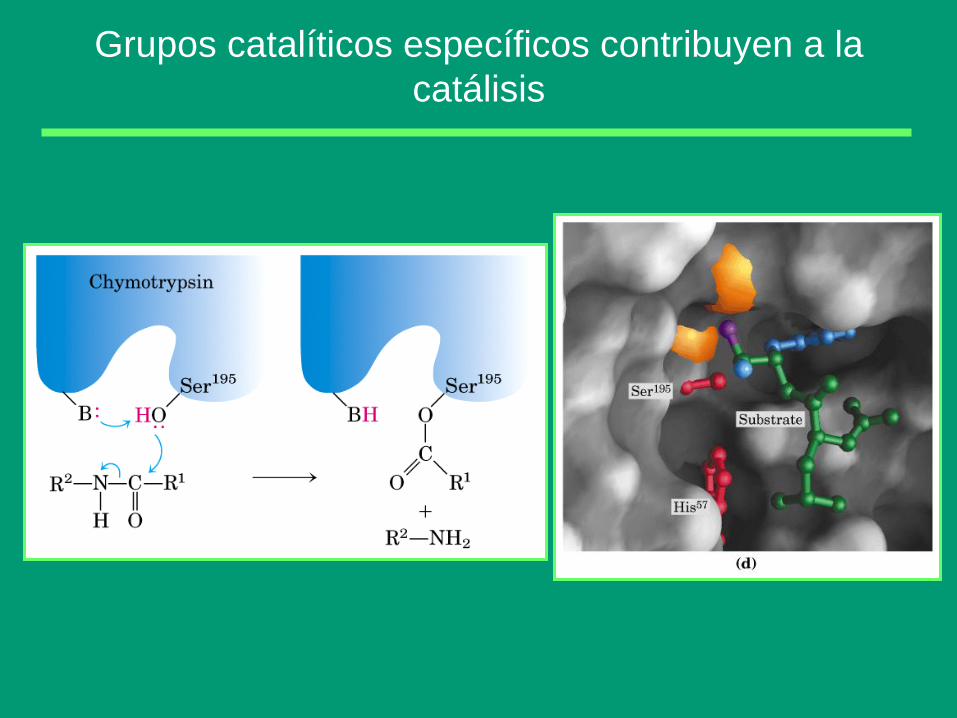
Grupos catalíticos específicos contribuyen a la
catálisis

A. Propiedades generales de las enzimas
B. Clasificación de las Enzimas
C. Principales problemas en el manejo de las Enzimas
D. Principios fundamentales de su acción catalítica
E. Introducción a la cinética enzimática
F. Enzimas reguladores
Enzimas

Cinética del estado estacionario (1913)
Postularon que la enzima se combina en primer lugar
con el sustrato, de forma reversible
El complejo se descompone en una reacción más
lenta, dando lugar al producto y enzima libre
Leonor Michaelis Maud Menten
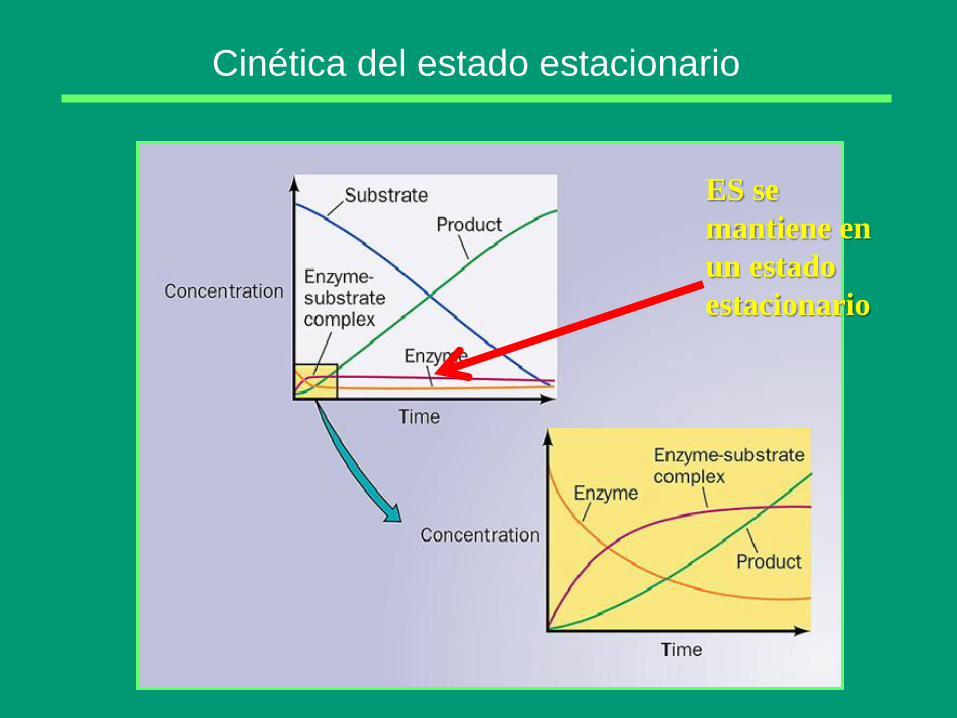
Cinética del estado estacionario
ES se
mantiene en
un estado
estacionario

Cinética del estado estacionario
d[ES] = k1 [E][S] Formación de ES dt
- d[ES] = k-1 [ES] + k2[ES] Descomposición de ES
dt
E + S ↔ ES → E + P k1
k-1
k2

ET = EL + ES
EL = ET - ES
Formación de ES: k1[EL][S]
Descomposición de ES: k-1 [ES] + k2 [ES]
k1[EL][S] = k-1 [ES] + k2 [ES]
k1[EL][S] = (k-1 + k2 )[ES]
k-1+ k2 = [EL][S]
k1 [ES]
E + S ↔ ES → E + P k1
k-1
k2

k-1+ k2 = [ET - ES][S]
k1 [ES] KM =
Constante de Michaelis:

Relación entre concentración de sustrato y
velocidad de reacción enzimática
La velocidad inicial de la reacción siempre
corresponde a la ecuación:
vo = k2 [ES]
Cuando toda la enzima se encuentra formando
complejo ES:
vmax = k2 [ET]

Relación entre concentración de sustrato y
velocidad de reacción enzimática
k1[EL][S] = k-1 [ES] + k2 [ES]
k1[ET - ES][S] = k-1 [ES] + k2 [ES]
k1[ET][S] – k1[ES][S] = k-1 [ES] + k2 [ES]
k1[ET] [S] = k-1 [ES] + k2 [ES] + k1[ES][S]
k1[ET] [S] = [ES] (k-1 + k2 + k1[S])
Dividimos por k1: [ET] [S] = [ES] ((k-1 + k2 ) + [S])
k1

[ET] [S] = [ES] (KM +[S])
(KM +[S])
[ET] [S] [ES] =
como vo = k2 [ES]
vo = k2 [ET] [S]
KM + [S]
vo = vmax [S]
KM + [S] Ecuación de
Michaelis-Menten
vmax = k2 [ET]
vmax
[ET]
K2 =

Variables o factores que influyen en la velocidad de
una reacción enzimática
1. Concentración de sustrato
2. Concentración de enzima
3. pH
4. Temperatura
5. Efectores (Activadores e Inhibidores)
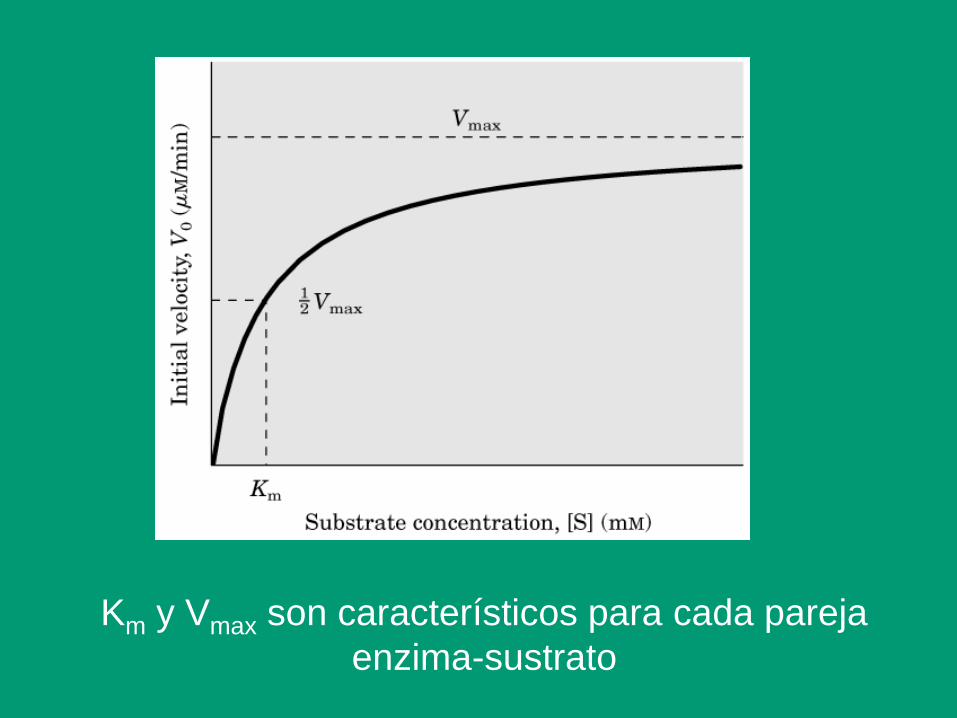
Km y Vmax son característicos para cada pareja
enzima-sustrato

Significado de la constante Km
1. Constante de equilibrio de disociación del complejo ES
2. Medida inversa de la afinidad de la enzima por el sustrato
3. Mide la función de fijación
4. Concentración de sustrato para la que la velocidad se hace
igual a la mitad de la máxima (V0.5)
5. Se define para un complejo enzima-sustrato
6. Se mide en unidades de concentración
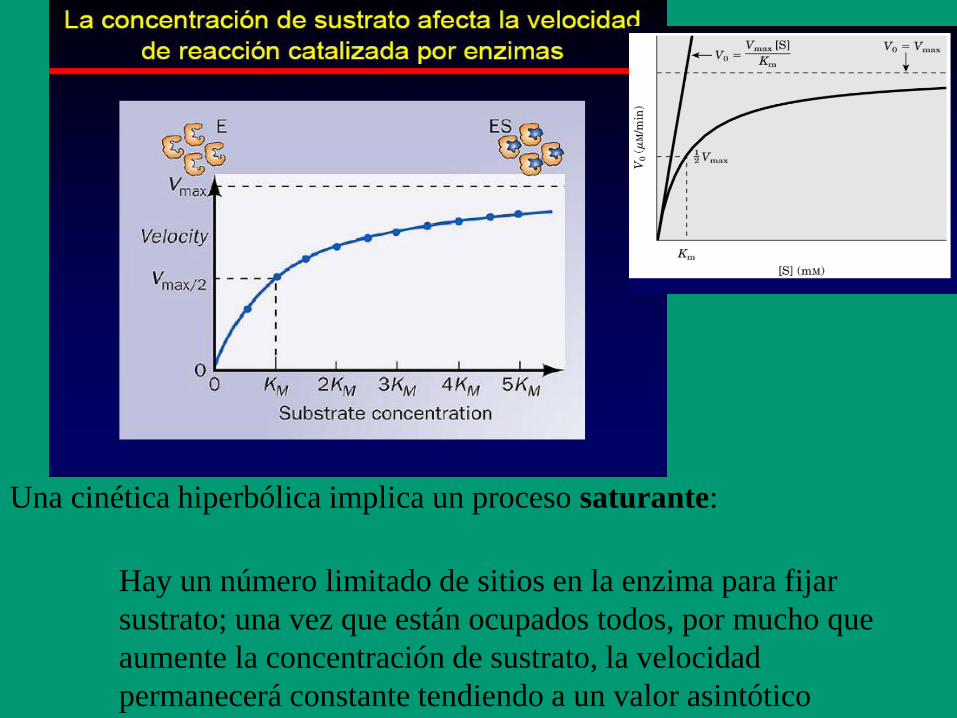
Una cinética hiperbólica implica un proceso saturante:
Hay un número limitado de sitios en la enzima para fijar
sustrato; una vez que están ocupados todos, por mucho que
aumente la concentración de sustrato, la velocidad
permanecerá constante tendiendo a un valor asintótico
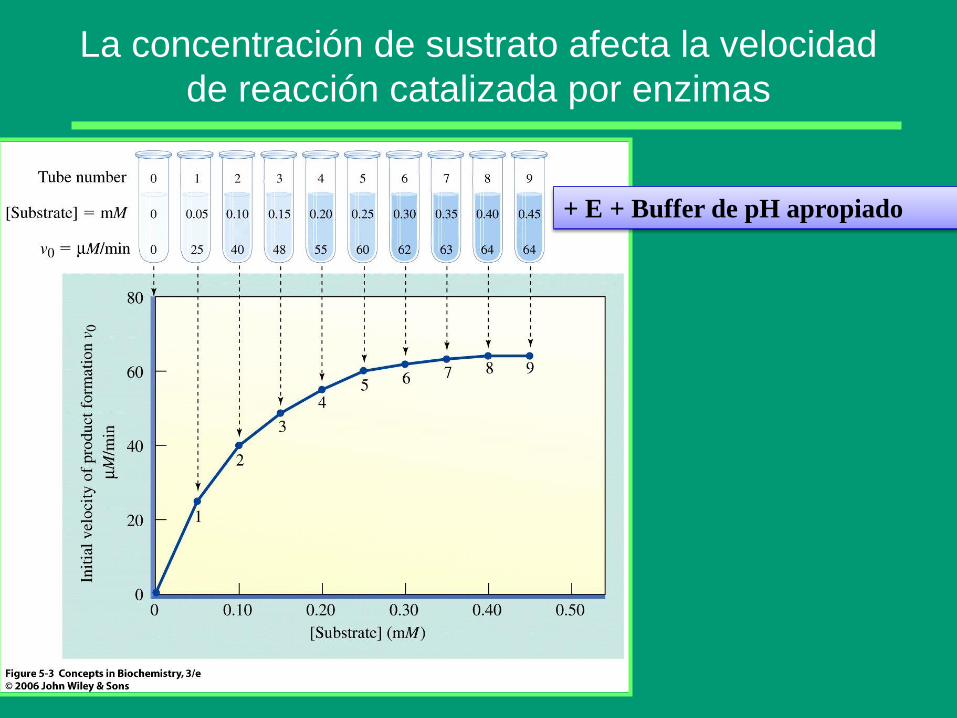
La concentración de sustrato afecta la velocidad
de reacción catalizada por enzimas
+ E + Buffer de pH apropiado
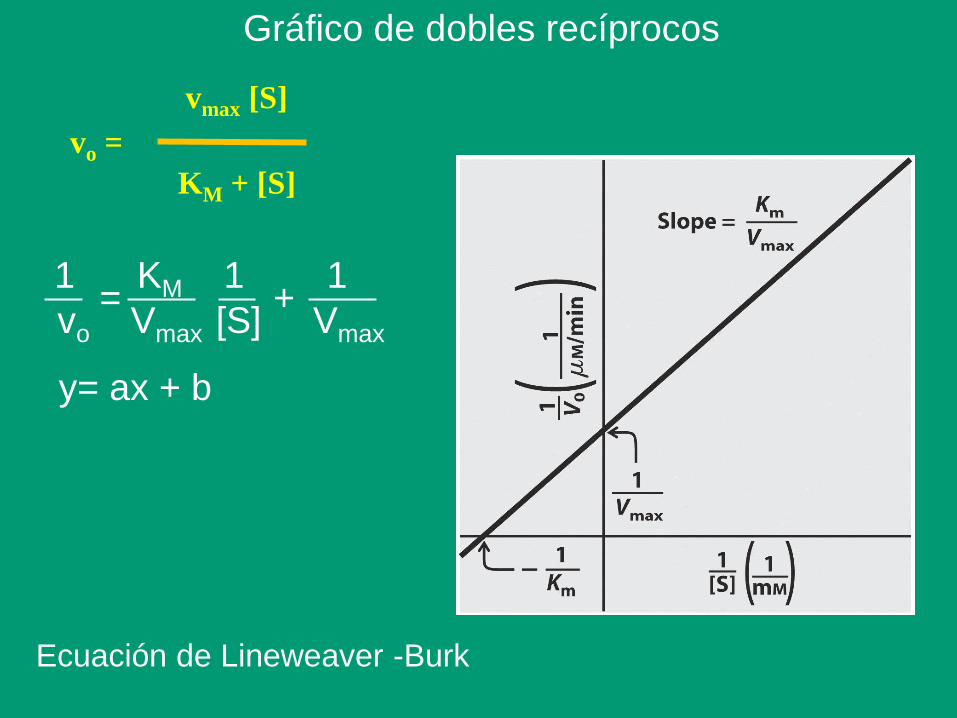
Gráfico de dobles recíprocos
1 KM 1 1
vo Vmax [S] Vmax
= +
Ecuación de Lineweaver -Burk
y= ax + b
vo =
vmax [S]
KM + [S]

Muchas enzimas catalizan reacciones con dos
o más sustratos
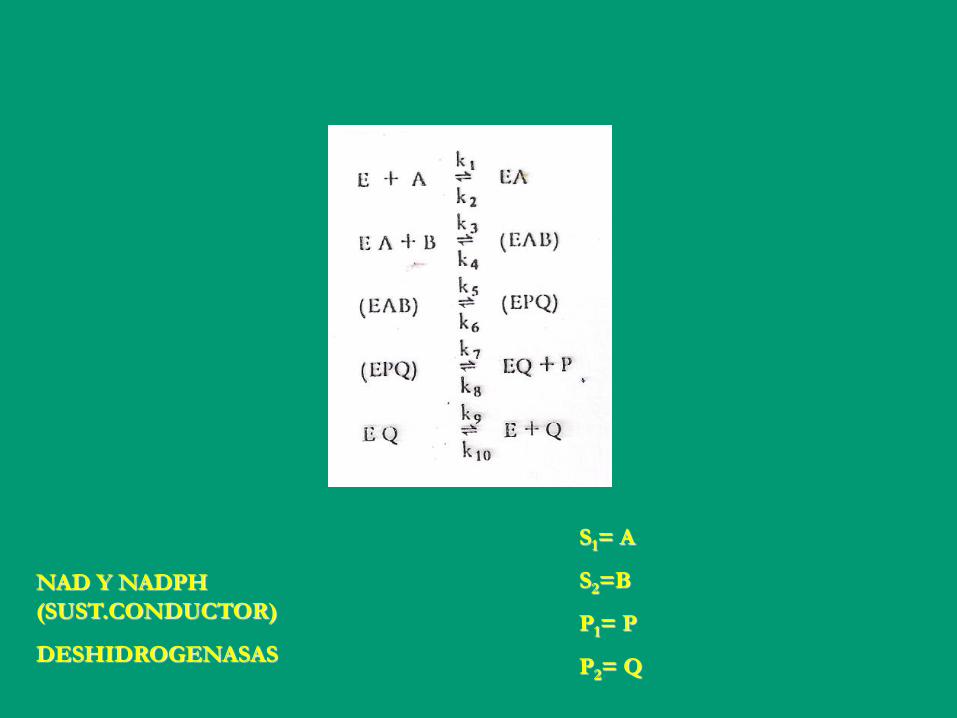
S1= A
S2=B
P1= P
P2= Q
NAD Y NADPH
(SUST.CONDUCTOR)
DESHIDROGENASAS
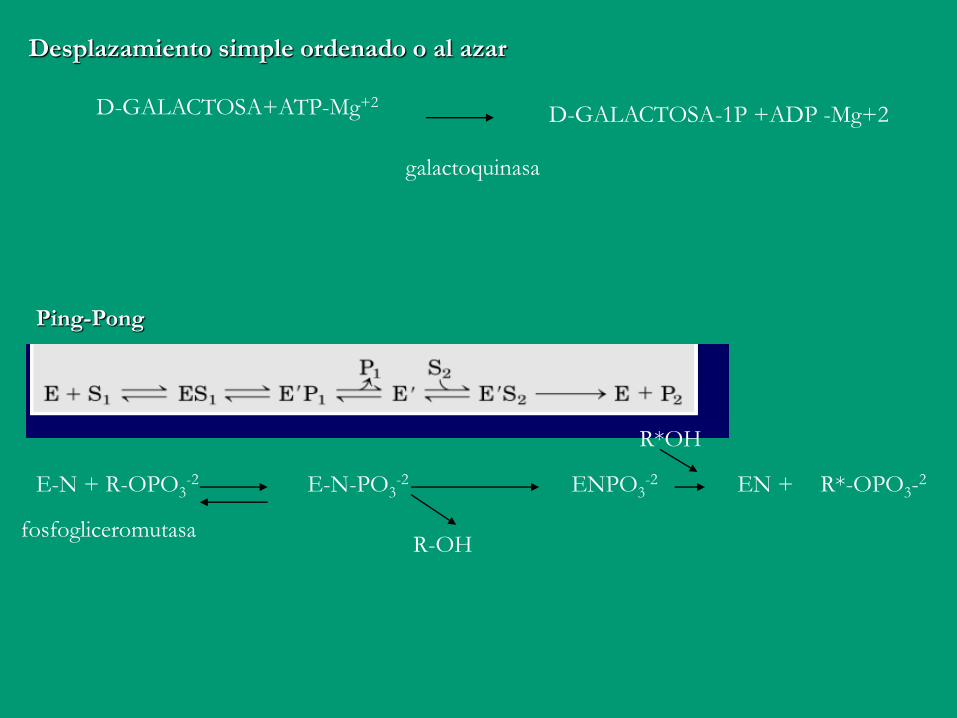
D-GALACTOSA+ATP-Mg+2 D-GALACTOSA-1P +ADP -Mg+2
galactoquinasa
Desplazamiento simple ordenado o al azar
Ping-Pong
E
E-N + R-OPO3-2 E-N-PO3
-2
R-OH
ENPO3-2 EN + R*-OPO3-
2
R*OH
fosfogliceromutasa
COOH
CHH2N
R1
+
COOH
C O
R2
COOH
CHH2N
R2
COOH
C O
R1
+
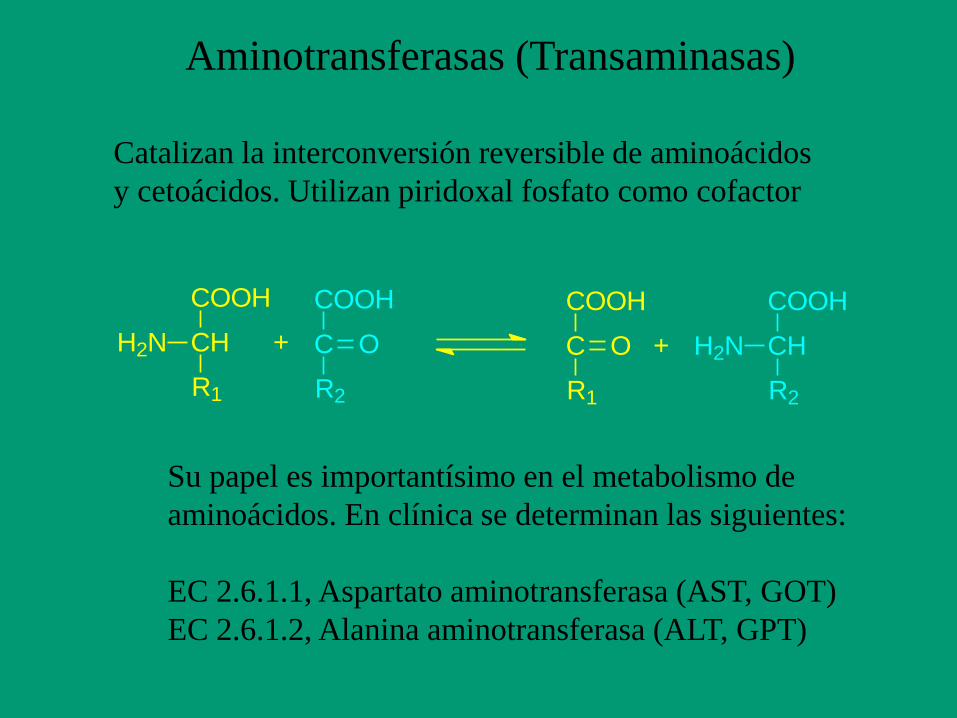
Aminotransferasas (Transaminasas)
Catalizan la interconversión reversible de aminoácidos
y cetoácidos. Utilizan piridoxal fosfato como cofactor
Su papel es importantísimo en el metabolismo de
aminoácidos. En clínica se determinan las siguientes:
EC 2.6.1.1, Aspartato aminotransferasa (AST, GOT)
EC 2.6.1.2, Alanina aminotransferasa (ALT, GPT)
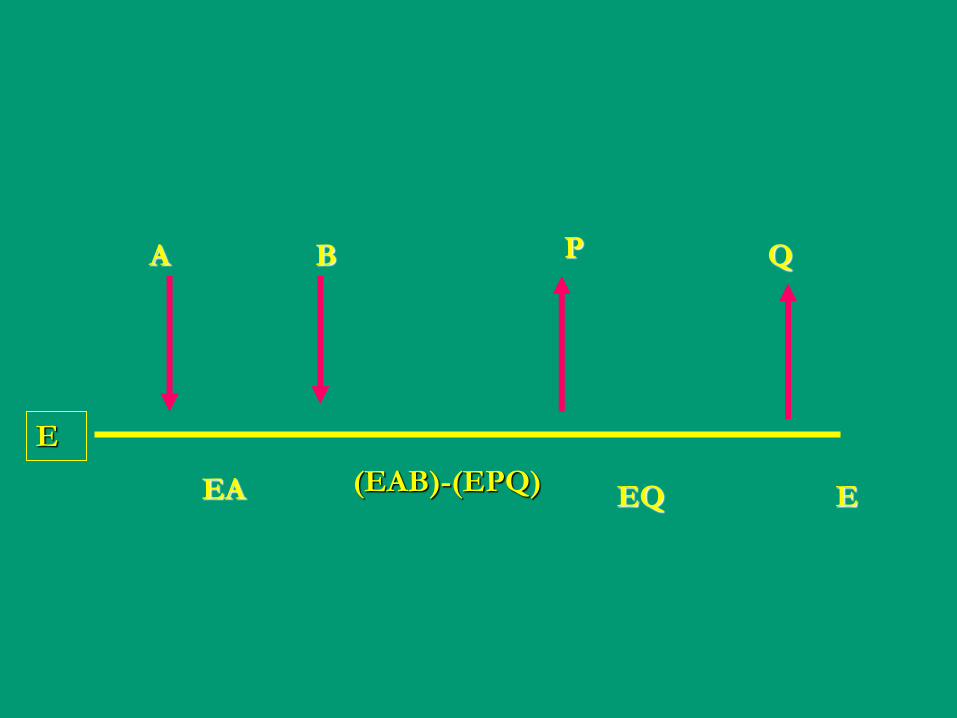
E
A B P Q
EA (EAB)-(EPQ) EQ E
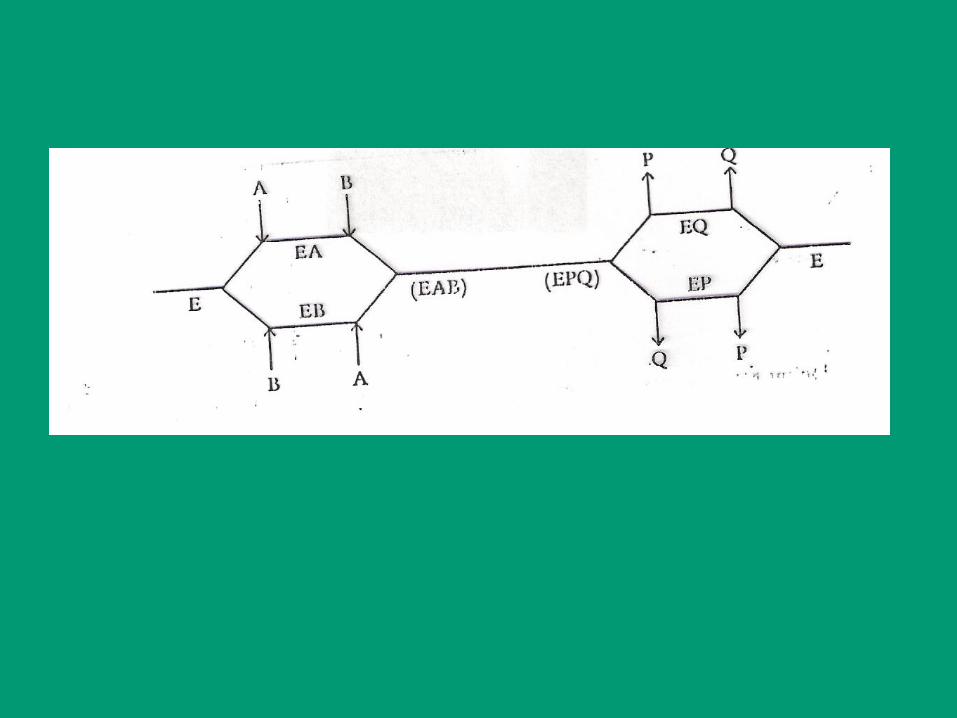
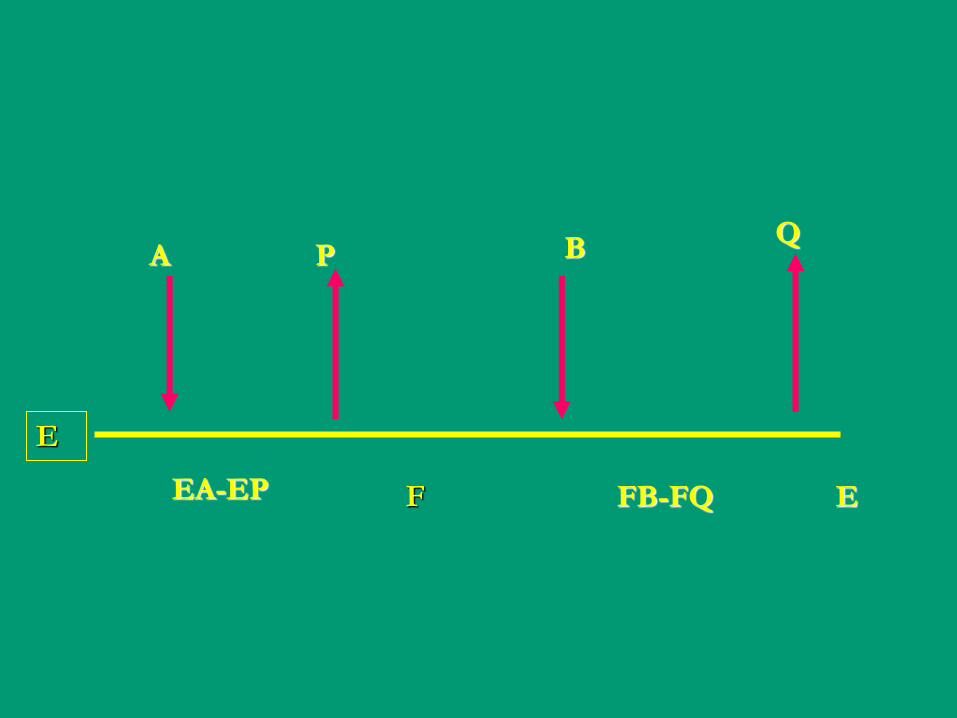
E
A P B Q
EA-EP F FB-FQ E
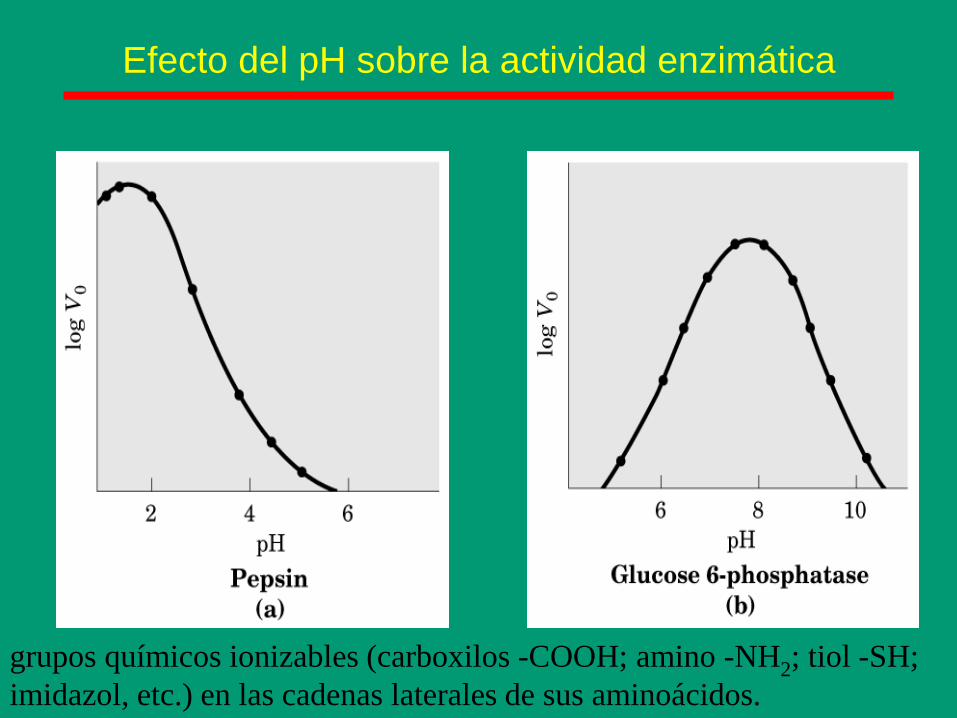
Efecto del pH sobre la actividad enzimática
grupos químicos ionizables (carboxilos -COOH; amino -NH2; tiol -SH;
imidazol, etc.) en las cadenas laterales de sus aminoácidos.

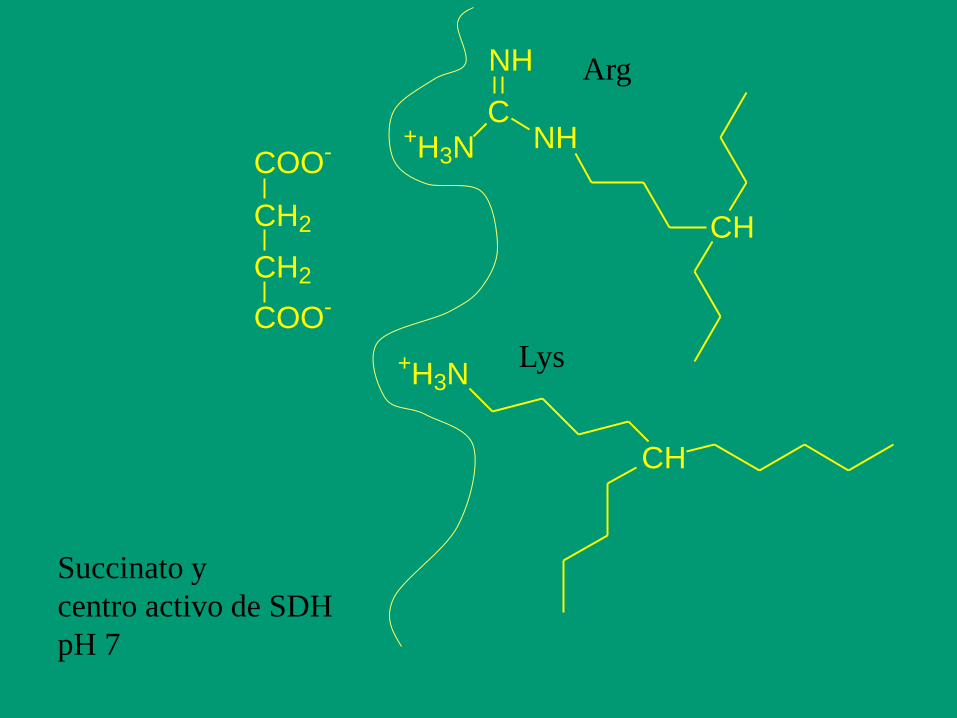
Succinato y
centro activo de SDH
pH 7
COO-
CH2
CH2
COO-
+H3NC
NH
NH
CH
+H3N
CH
Arg
Lys
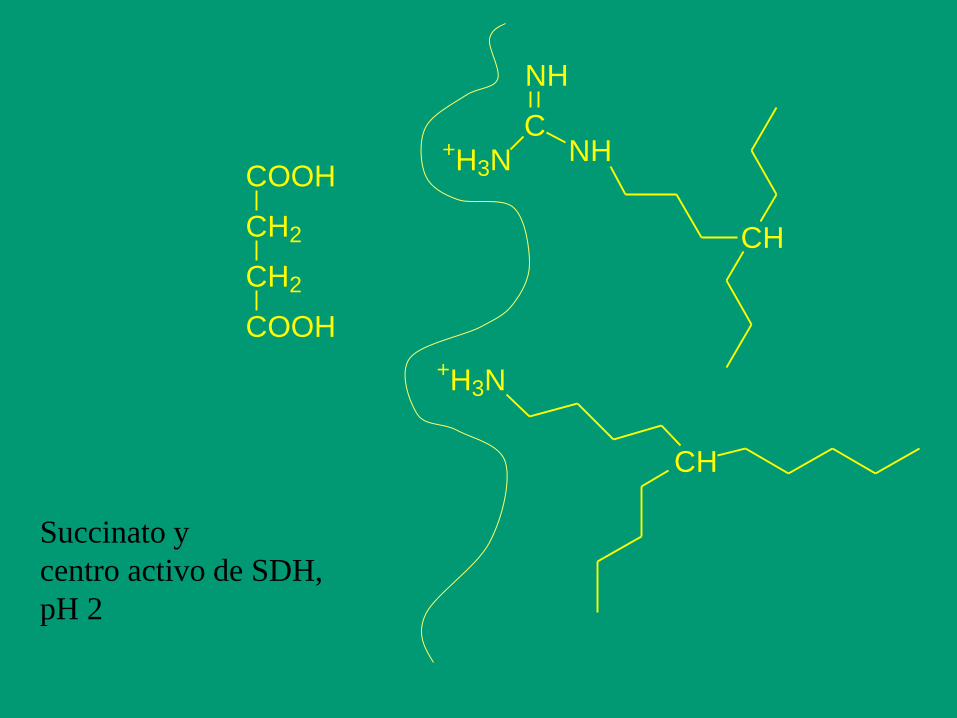
Succinato y
centro activo de SDH,
pH 2
COOH
CH2
CH2
COOH
+H3NC
NH
NH
CH
+H3N
CH
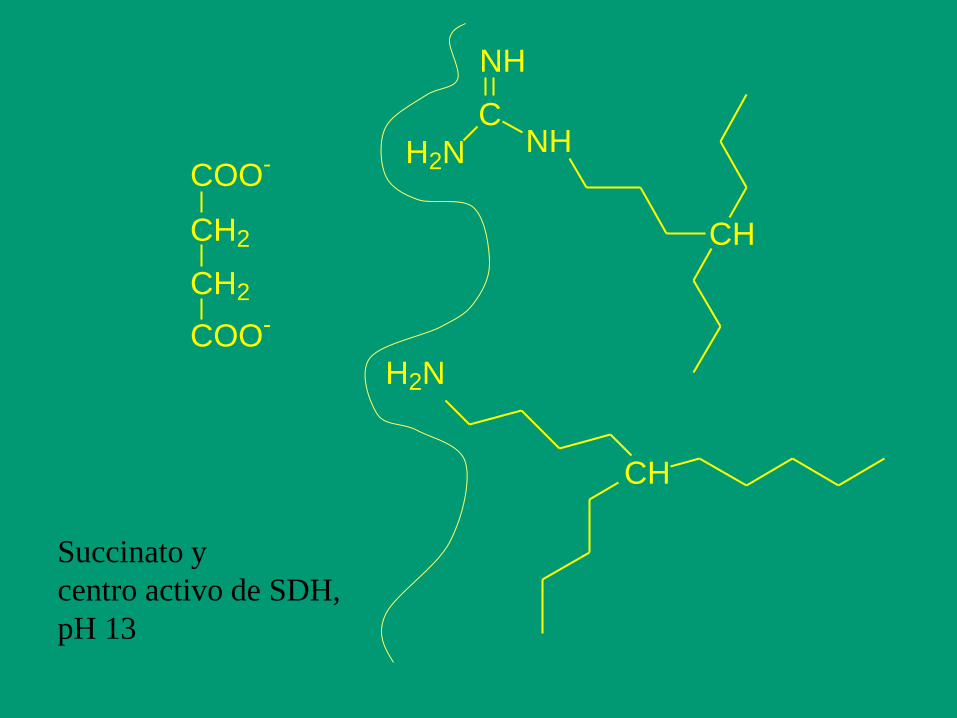
COO-
CH2
CH2
COO-
H2N
C
NH
NH
CH
H2N
CH
Succinato y
centro activo de SDH,
pH 13




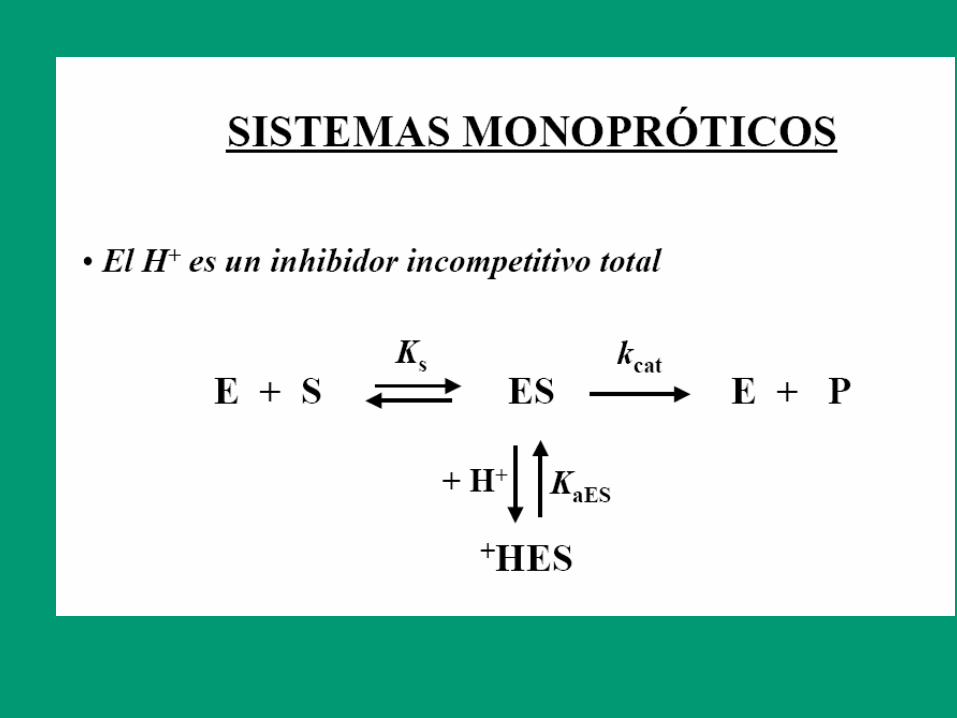

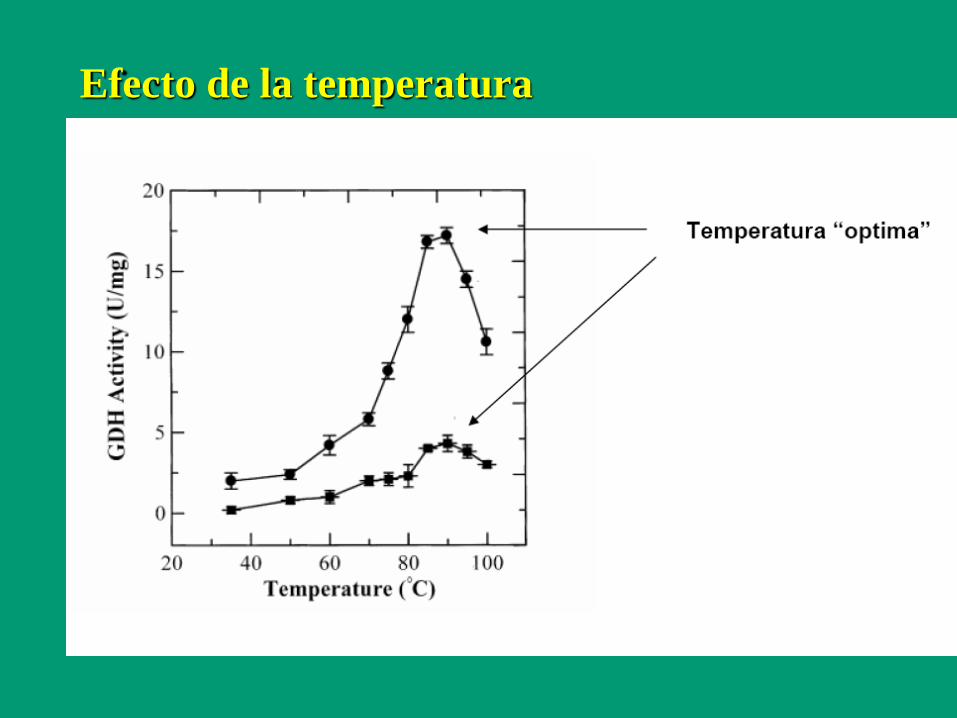
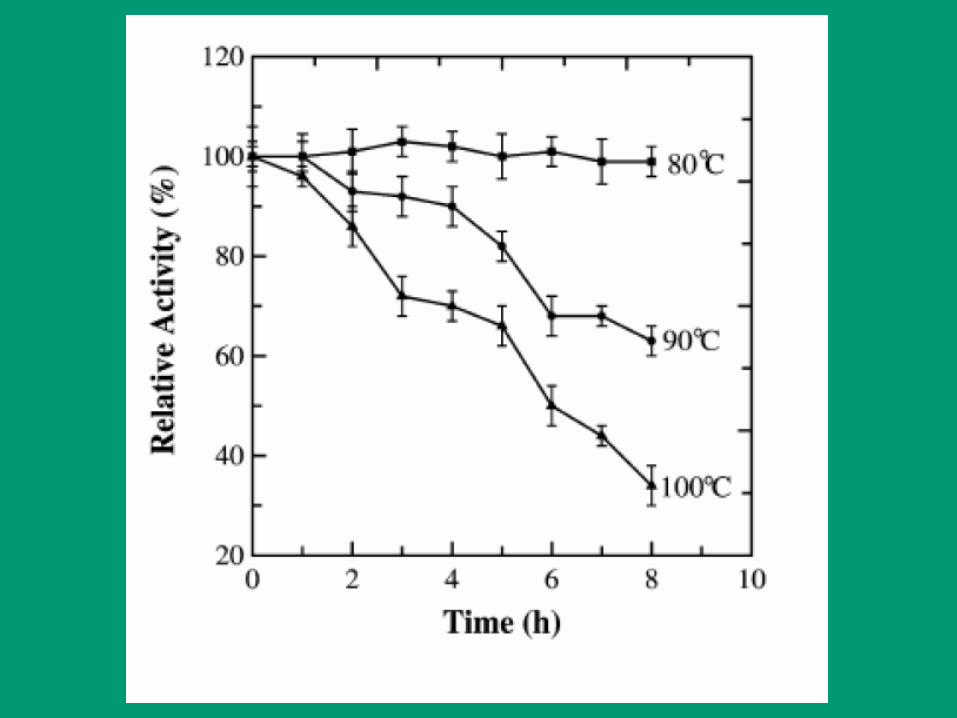
Efecto de la temperatura
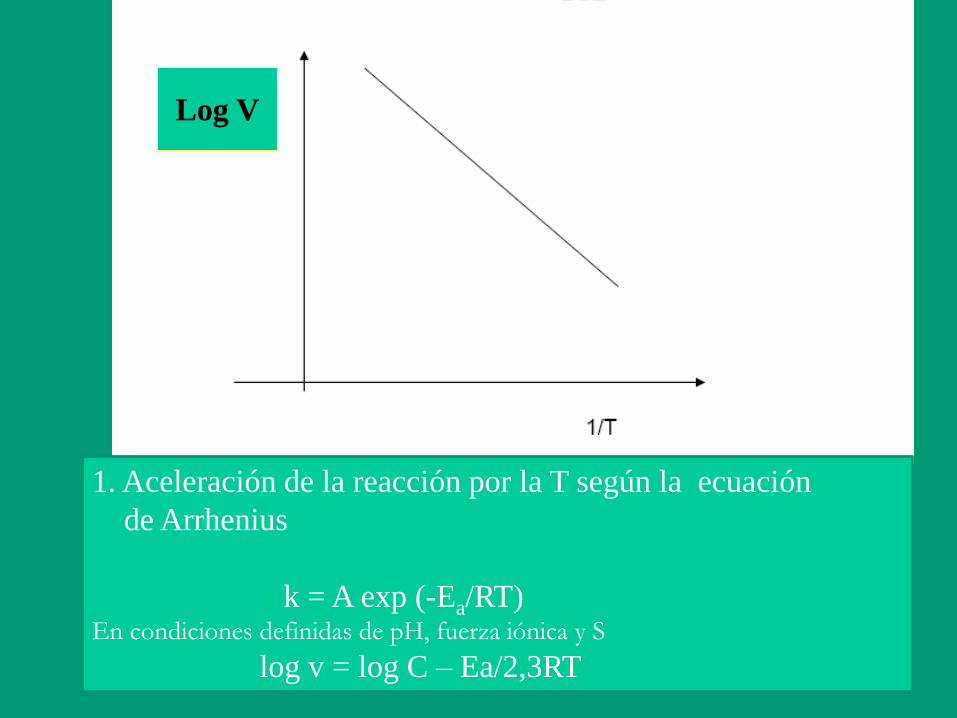
Log V
1. Aceleración de la reacción por la T según la ecuación
de Arrhenius
k = A exp (-Ea/RT) En condiciones definidas de pH, fuerza iónica y S
log v = log C – Ea/2,3RT

Inhibidor:
Efector que hace disminuir la actividad enzimática, a través de
interacciones con el centro activo u otros centros específicos
(alostéricos).
Esta definición excluye todos aquellos agentes que inactivan a
la enzima a través de desnaturalización de la molécula enzimática
De esta forma, habrá dos tipos de inhibidores:
I. Isostéricos: ejercen su acción sobre el centro activo
II. Alostéricos: ejercen su acción sobre otra parte de la
molécula, produciendo un cambio conformacional con
repercusión negativa en la actividad enzimática.

Los inhibidores isostéricos pueden ser de dos tipos:
1. Inhibidor reversible: establece un equilibrio con la enzima libre,
con el complejo enzima-sustrato o con ambos:
E + I EI
2. Inhibidor irreversible: modifica químicamente a la enzima:
E + I E’
ES + I ESI

Inhibición reversible
(a) El inhibidor se fija al centro activo de la enzima libre,
impidiendo la fijación del sustrato: Inhibición Competitiva
(b) El inhibidor se fija a la enzima independientemente de que lo
haga o no el sustrato; el inhibidor, por tanto, no impide la
fijación del sustrato a la enzima, pero sí impide la acción
catalítica: Inhibición No Competitiva
(c) El inhibidor se fija únicamente al complejo enzima-sustrato
una vez formado, impidiendo la acción catalítica; este tipo
se conoce como Inhibición Anticompetitiva
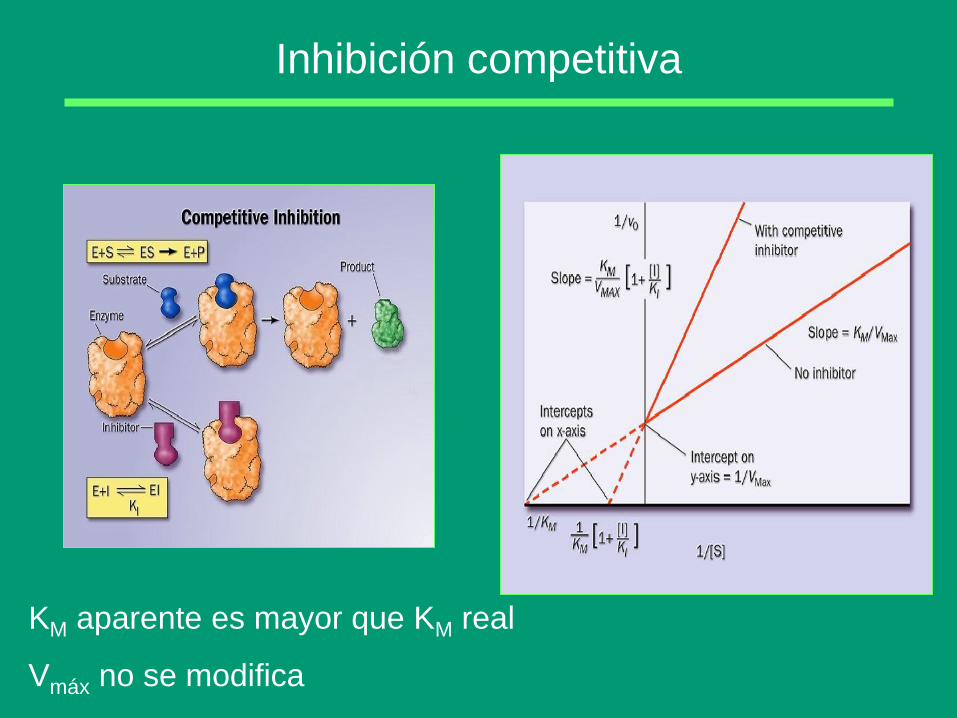
Inhibición competitiva
KM aparente es mayor que KM real
Vmáx no se modifica

E ES
EI
I
S
E + P
Características:
- Las fijaciones de sustrato e inhibidor son mutuamente exclusivas
- A muy altas concentraciones de sustrato desaparece la inhibición
- Por lo general, el inhibidor competitivo es un análogo químico del
sustrato.
- El inhibidor es tan específico como el sustrato
Se define una constante de
equilibrio de disociación del
inhibidor: Ki =
[E] [I]
[EI]
Inhibición Competitiva

Por tanto, en la inhibición competitiva,
1. El efecto cinético del inhibidor es el aumento aparente de la
Km, que aparece multiplicada por el factor (1 + I/Ki)
2. La Vmax no aparece modificada; para concentraciones muy
altas del sustrato, v = Vmax, igual que en ausencia de inhibidor
3. Cuanto más pequeño sea el valor de Ki mayor será la potencia
del inhibidor competitivo.
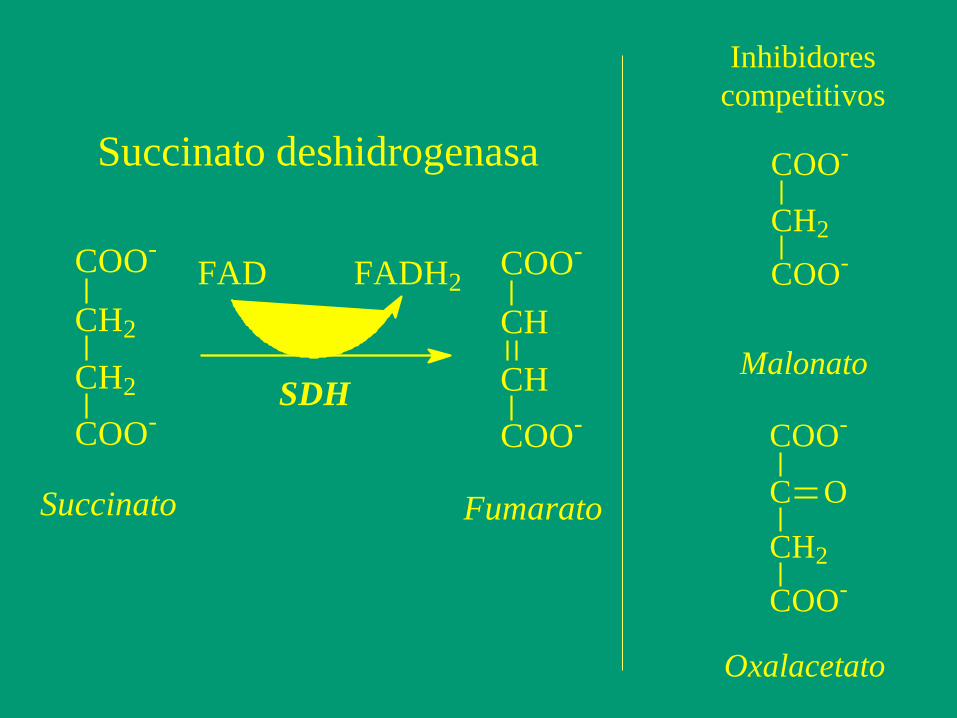
COO-
CH2
CH2
COO-
FAD FADH2COO-
CH
CH
COO-
Succinato Fumarato
SDH
Succinato deshidrogenasa COO-
CH2
COO-
Malonato
COO-
C O
CH2
COO-
Oxalacetato
Inhibidores
competitivos

Un paso más allá en el desarrollo de inhibidores potentes
es el concepto de Análogo de Estado de Transición (AET)
- El inhibidor no es estrictamente análogo del sustrato,
sino del Estado de Transición de la reacción.
- La afinidad de las enzimas por los AET es enorme, del
orden nM o pM, con lo que la fijación es tan fuerte que
puede considerarse irreversible
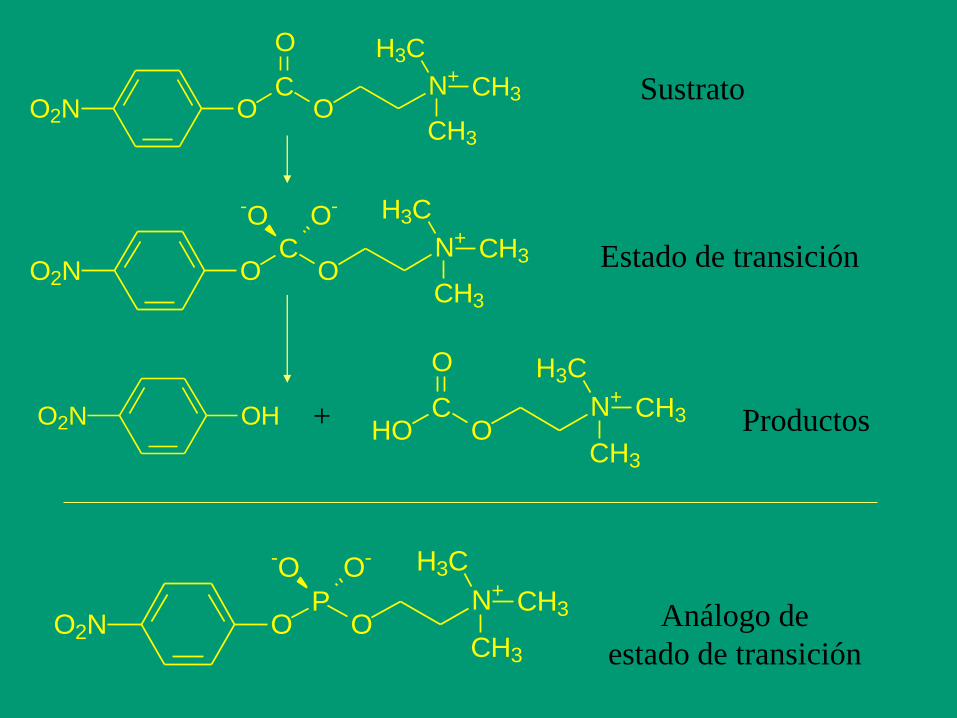
O2N OC
O
O
N+
H3C
CH3
CH3
O2N OP
ON+
H3C
CH3
CH3
-O O-
Sustrato
O2N OC
ON+
H3C
CH3
CH3
-O O-
Estado de transición
O2N OH + O
N+
H3C
CH3
CH3
C
O
HO Productos
Análogo de
estado de transición

Metanol formaldehido
alcohol deshidrogenasa hepática
Etanol acetaldehido
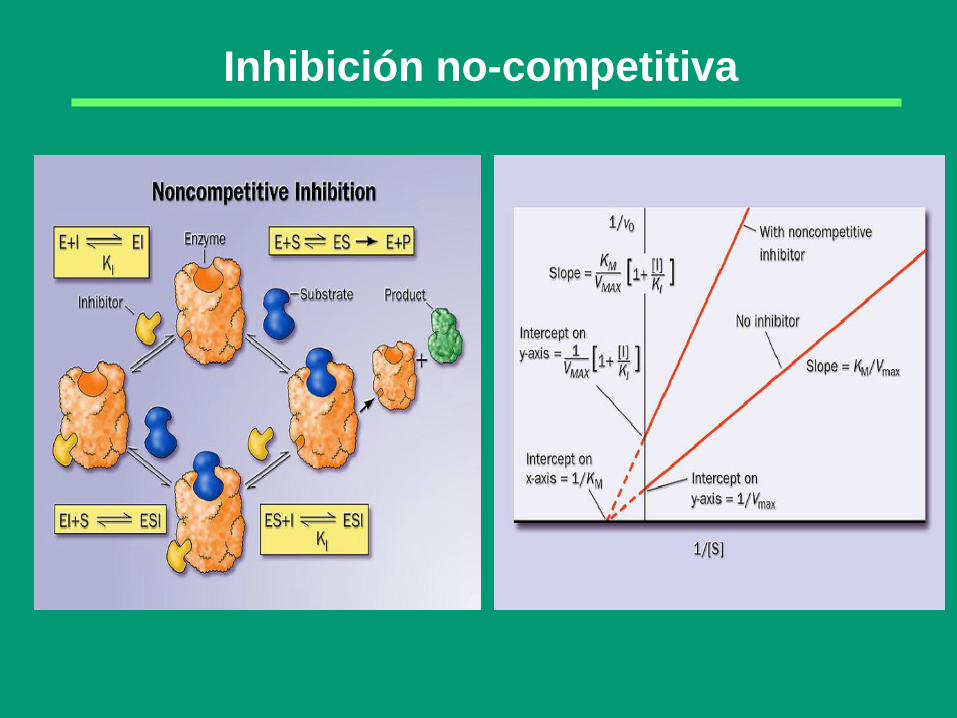
Inhibición no-competitiva

E ES
EI
I
S
E + P
I
ESI
S Inhibición
No Competitiva
El inhibidor se fija indistintamente a la enzima libre E
y al complejo enzima-substrato ES; ni el complejo EI
ni el complejo ESI son productivos
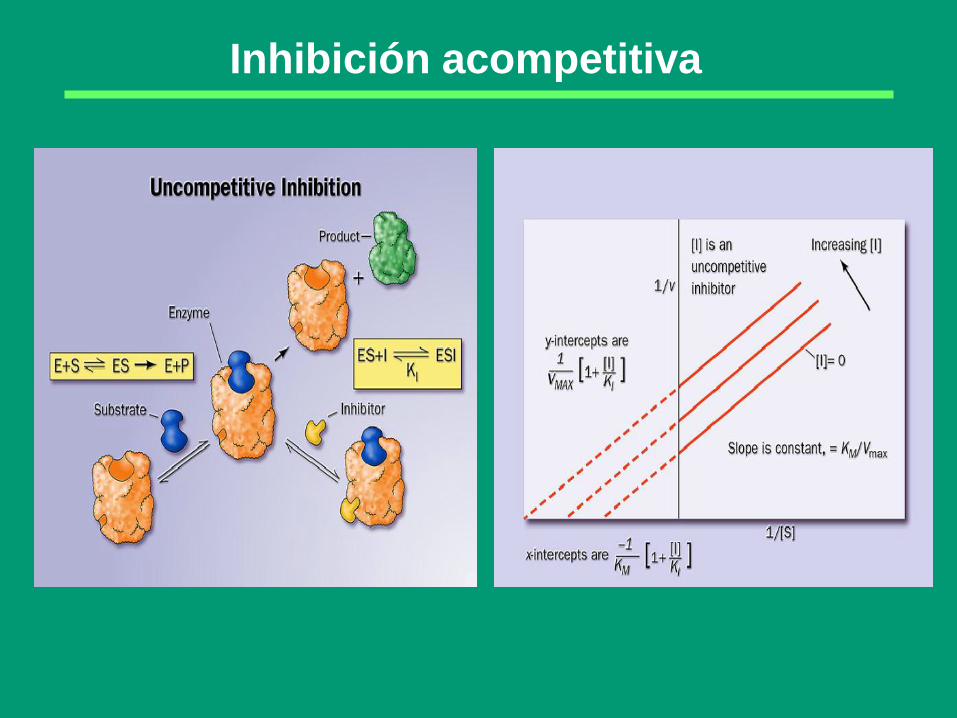
Inhibición acompetitiva

E ES
S
E + P
I
ESI
Inhibición
Anticompetitiva
El inhibidor sólo puede fijarse reversiblemente al complejo ES;
el complejo ESI no es productivo
Ks K3
KI

Inhibición Irreversible
- Los inhibidores irreversibles reaccionan con un grupo
químico de la enzima, modificándola covalentemente
- Su acción no se describe por una constante de equilibrio Ki,
sino por una constante de velocidad ki:
E + I E’
- A diferencia de la inhibición reversible, el efecto de los
inhibidores irreversibles depende del tiempo de actuación
del inhibidor.
- Los inhibidores irreversibles son, por lo general, altamente
tóxicos.

Algunos tipos de inhibidores irreversibles
1. Reactivos de grupos -SH
2. Organofosfóricos
3. Ligandos de metales
4. Metales pesados
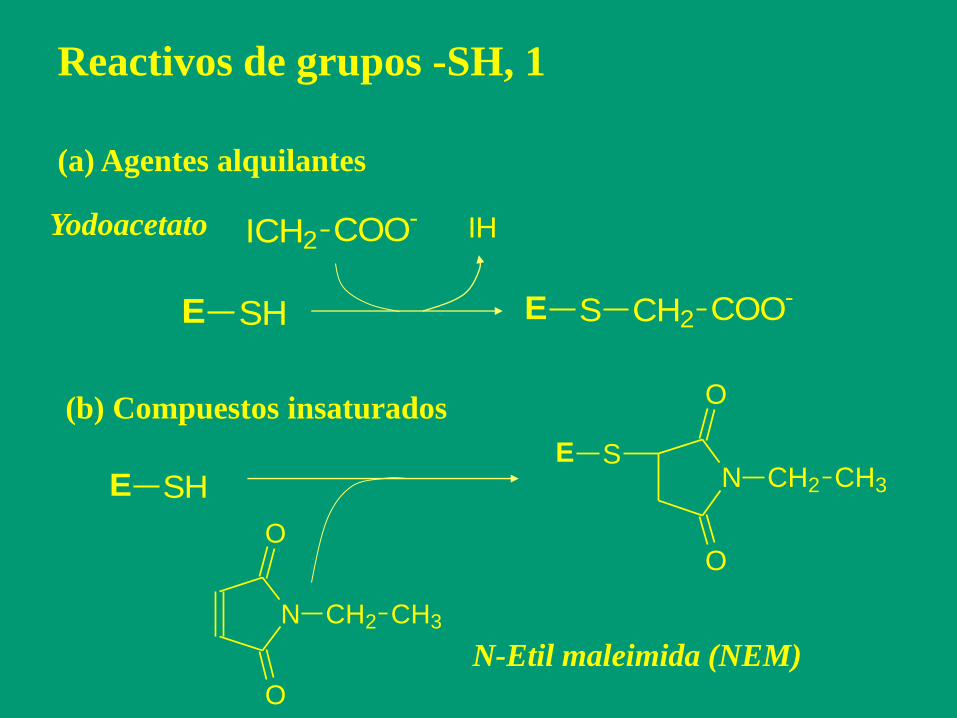
Reactivos de grupos -SH, 1
(a) Agentes alquilantes
E SH E S CH2 COO-
ICH2 COO- IHYodoacetato
(b) Compuestos insaturados
N CH2 CH3
O
O
E SHE S
N CH2 CH3
O
O
N-Etil maleimida (NEM)
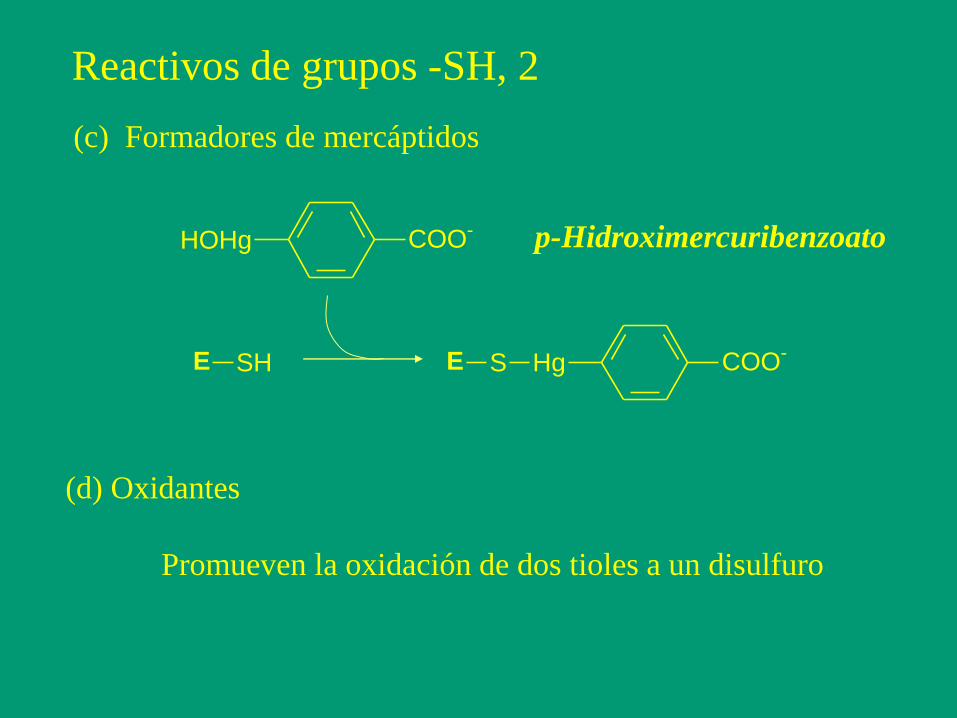
Reactivos de grupos -SH, 2
(c) Formadores de mercáptidos
HOHg COO-
E SH E S Hg COO-
p-Hidroximercuribenzoato
(d) Oxidantes
Promueven la oxidación de dos tioles a un disulfuro
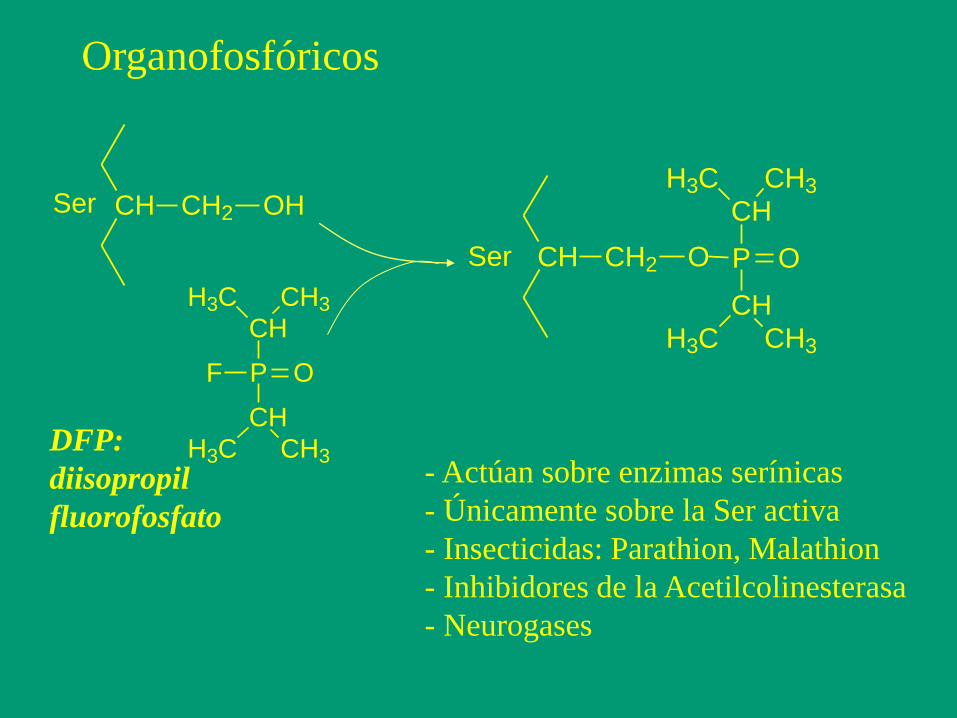
Organofosfóricos
CH CH2 OHSer
PF O
CH
CH
H3C CH3
CH3H3C
CH CH2 O P O
CH
CH
H3C CH3
CH3H3C
Ser
DFP:
diisopropil
fluorofosfato
- Actúan sobre enzimas serínicas
- Únicamente sobre la Ser activa
- Insecticidas: Parathion, Malathion
- Inhibidores de la Acetilcolinesterasa
- Neurogases

A. Propiedades generales de las enzimas
B. Principios fundamentales de su acción catalítica
C. Introducción a la cinética enzimática
D. Enzimas reguladoras
Enzimas

Regulación alostérica (union no covalente y reversible de un efector)
Procesos a nivel celular; regulación de ajuste fino
de la actividad enzimática, a través de efectos de
retroalimentación (negativa o positiva)
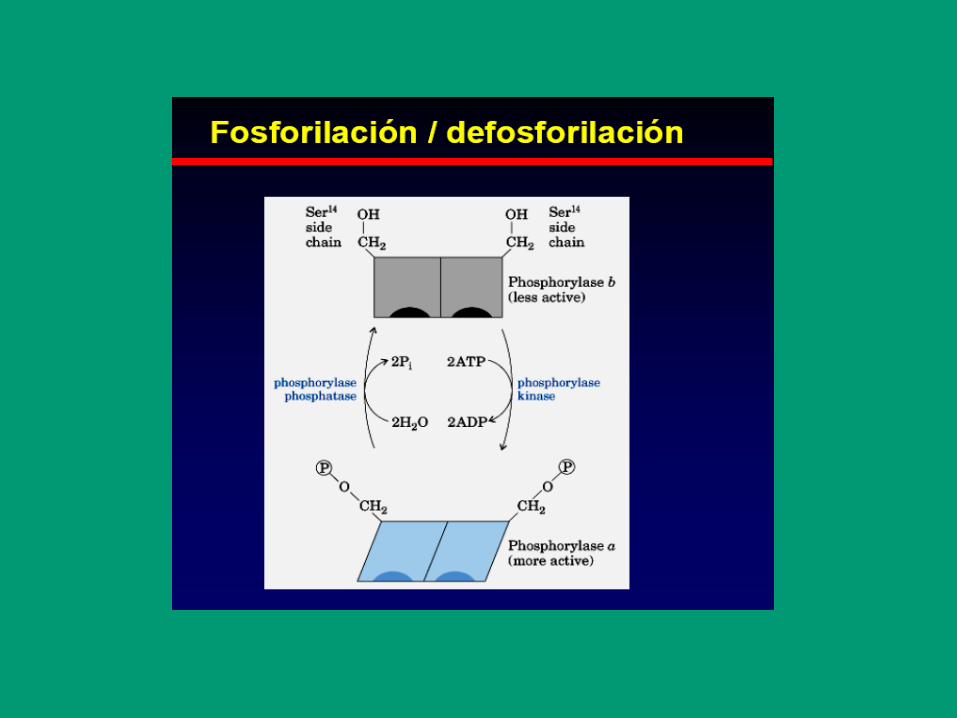
Regulación por modificación covalente
Procesos a nivel supracelular (orgánico); regulación
a gran escala de actividades enzimáticas, a través de
modificación covalente de enzimas, provocadas por
señales (transducción de señales)
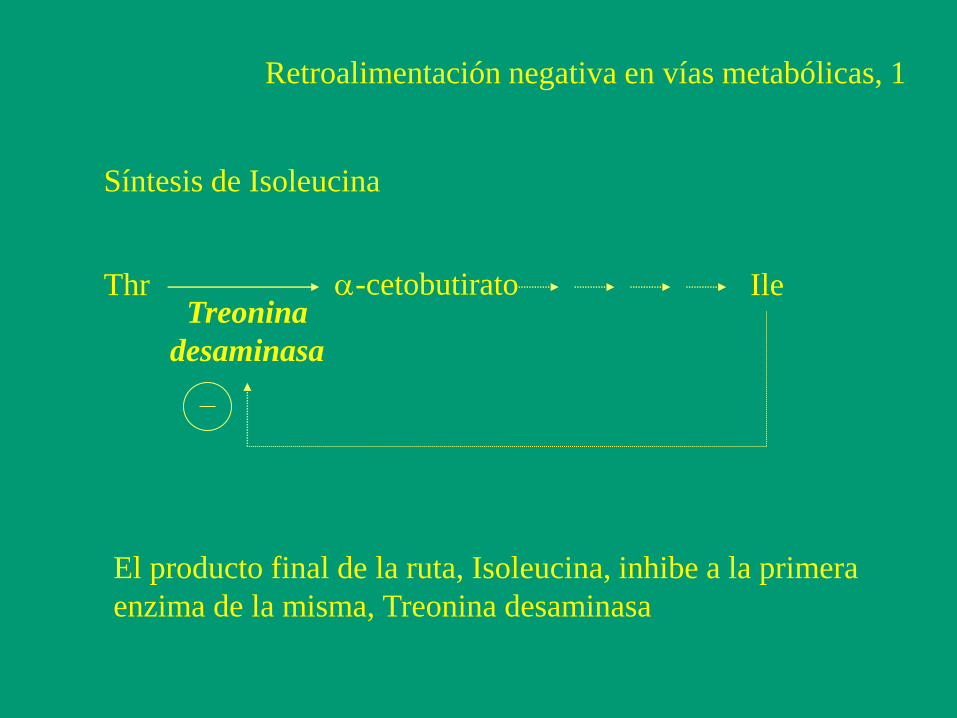
Retroalimentación negativa en vías metabólicas, 1
Thr a-cetobutirato Ile Treonina
desaminasa
Síntesis de Isoleucina
El producto final de la ruta, Isoleucina, inhibe a la primera
enzima de la misma, Treonina desaminasa
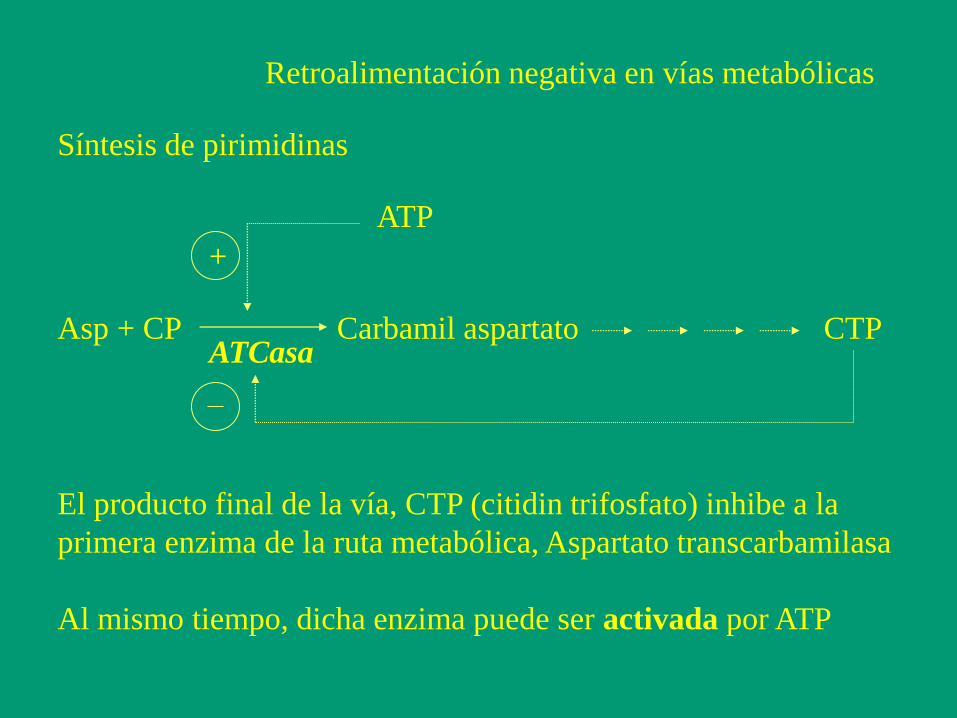
Retroalimentación negativa en vías metabólicas
Asp + CP Carbamil aspartato CTP
Síntesis de pirimidinas
ATCasa
El producto final de la vía, CTP (citidin trifosfato) inhibe a la
primera enzima de la ruta metabólica, Aspartato transcarbamilasa
Al mismo tiempo, dicha enzima puede ser activada por ATP
ATP
+


Rutas metabólicas controladas por retroalimentación
Glicolisis Fosfofructokinasa ATP
Neoglucogénesis FBP fosfatasa AMP
Bios. Ács. Grasos AcetilCoA carboxilasa AcilCoA
Bios. Colesterol HMGCoA reductasa Colesterol
Bios. Purinas PRPP sintetasa AMP,GMP,IMP
Ruta Enzima Inhibidor
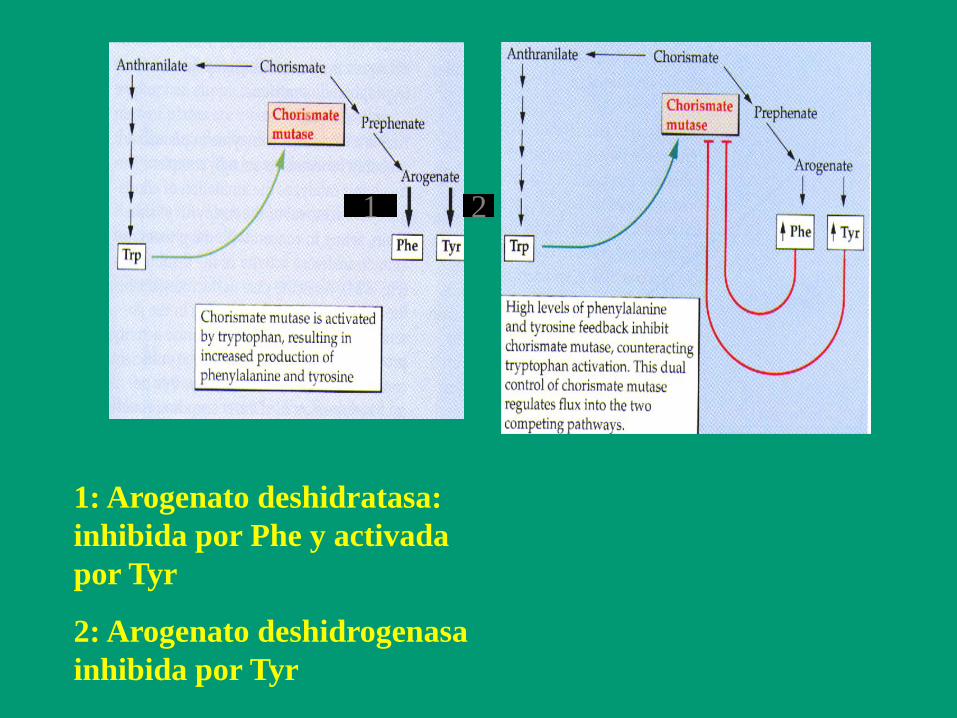
1
1: Arogenato deshidratasa:
inhibida por Phe y activada
por Tyr
2: Arogenato deshidrogenasa
inhibida por Tyr
2

En el estudio de las enzimas controladas por realimentación
negativa, se comprobaron una serie de regularidades en las mismas:
1. Primer paso de una ruta metabólica o punto de ramificación
2. Enzimas de naturaleza compleja: subunidades; la enzima puede
desensibilizarse a sus inhibidores por diversos métodos.
3. Los inhibidores se comportan como competitivos (elevan el valor
de la Km aparente, pero sin embargo no son análogos estructurales
del sustrato
Estas últimas características lleva al concepto de alosterismo:
Una acción sobre la actividad enzimática que se desarrolla
fuera del Centro Activo (Jacob y Monod, 1960)
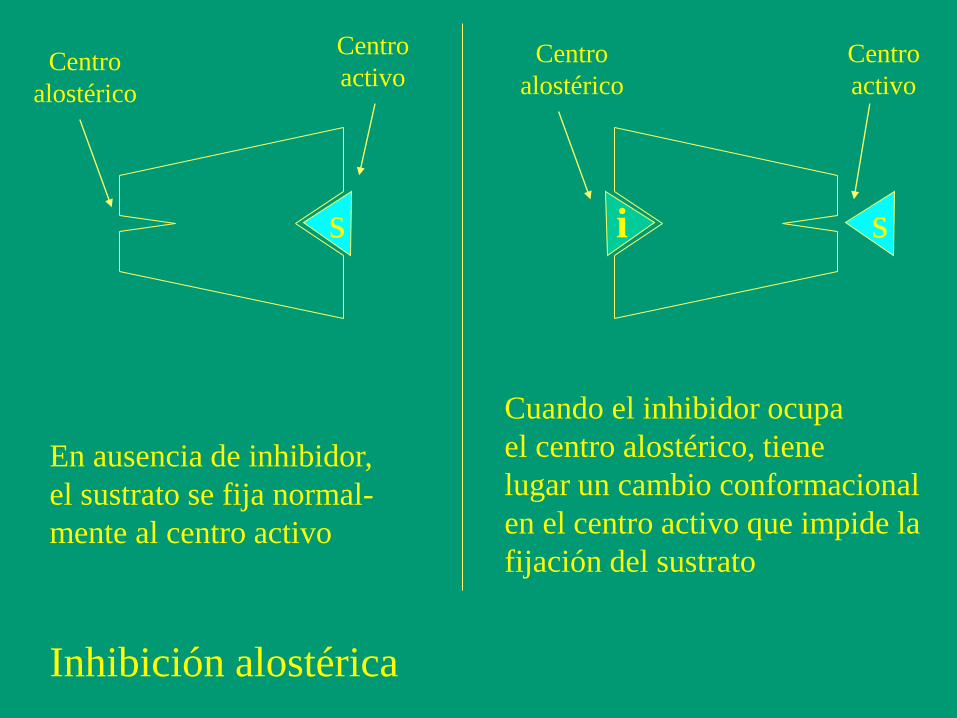
s s i
Centro
activo Centro
alostérico
Centro
alostérico
Centro
activo
En ausencia de inhibidor,
el sustrato se fija normal-
mente al centro activo
Cuando el inhibidor ocupa
el centro alostérico, tiene
lugar un cambio conformacional
en el centro activo que impide la
fijación del sustrato
Inhibición alostérica
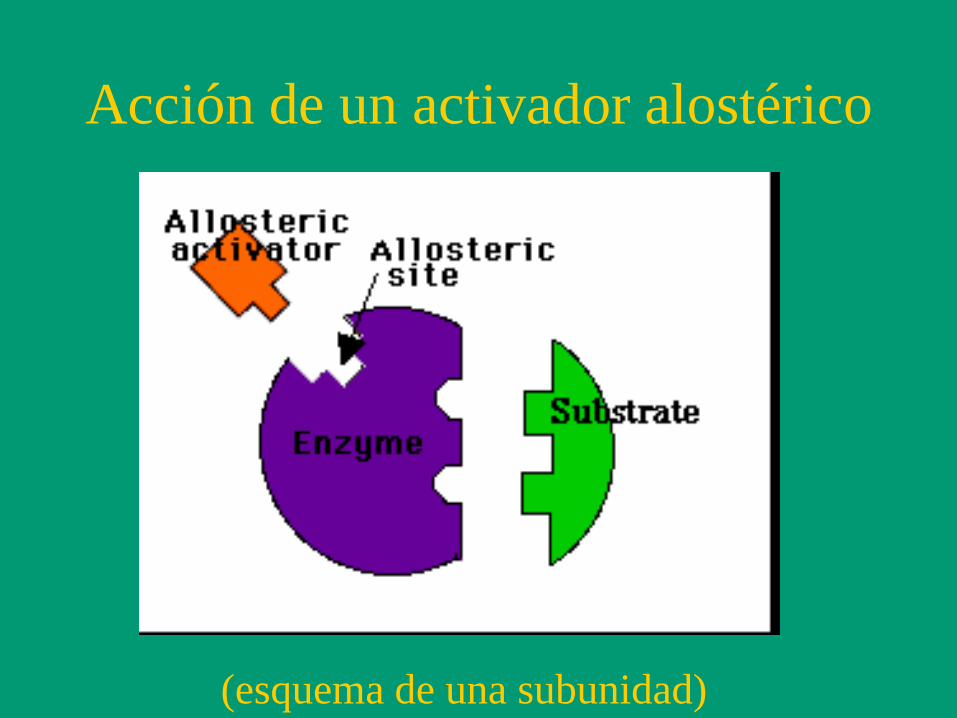
Acción de un activador alostérico
(esquema de una subunidad)
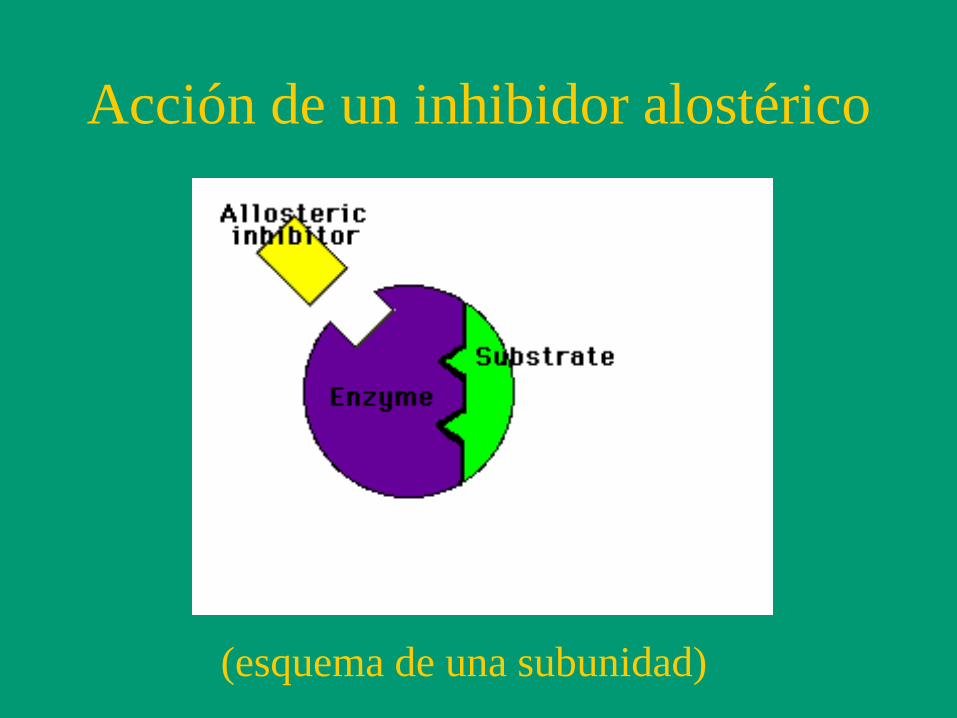
Acción de un inhibidor alostérico
(esquema de una subunidad)
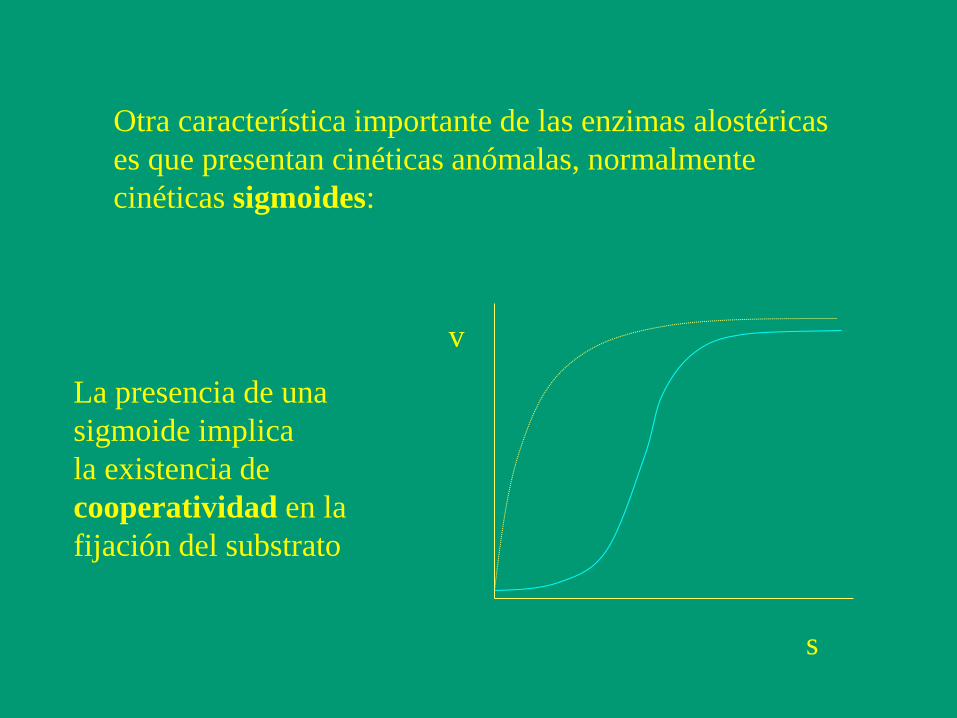
Otra característica importante de las enzimas alostéricas
es que presentan cinéticas anómalas, normalmente
cinéticas sigmoides:
v
s
La presencia de una
sigmoide implica
la existencia de
cooperatividad en la
fijación del substrato
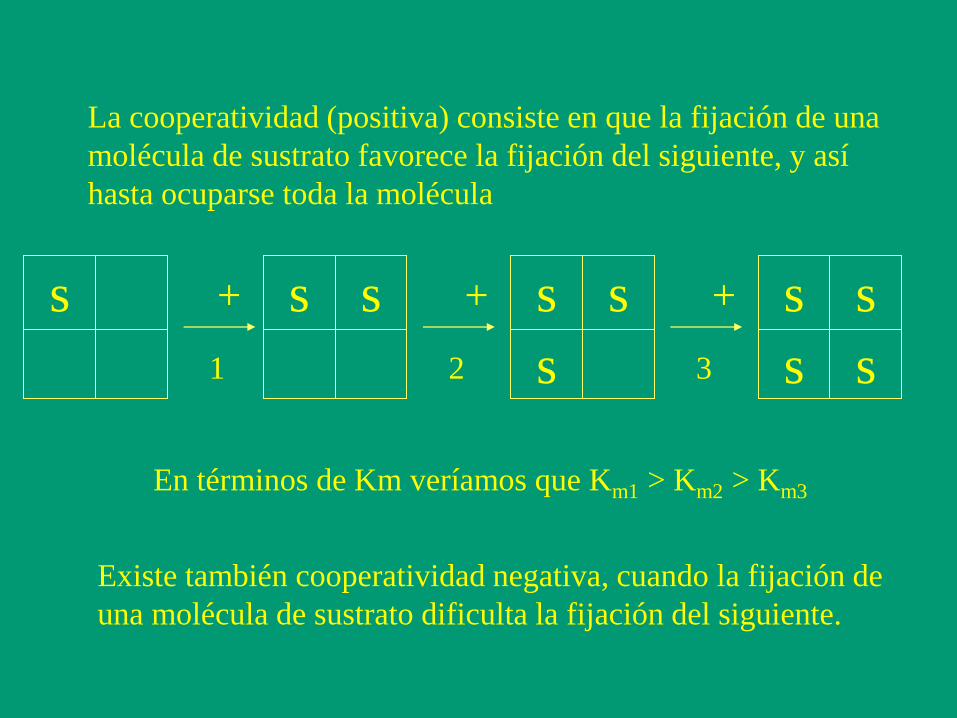
La cooperatividad (positiva) consiste en que la fijación de una
molécula de sustrato favorece la fijación del siguiente, y así
hasta ocuparse toda la molécula
s s s s s s
s
s
s s
+ + +
1 2 3
En términos de Km veríamos que Km1 > Km2 > Km3
Existe también cooperatividad negativa, cuando la fijación de
una molécula de sustrato dificulta la fijación del siguiente.

El comportamiento cooperativo implica:
1. Que hay más de un sitio de fijación de substrato
por molécula de enzima (estr.cuaternaria)
2. Que hay interacciones entre los sitios de fijación
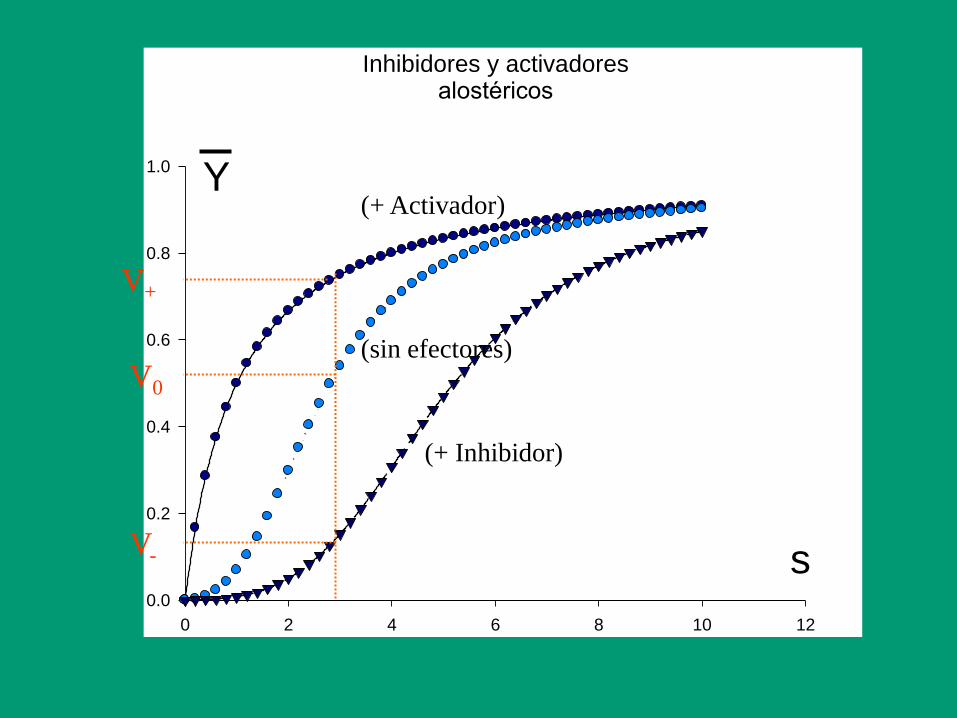
Inhibidores y activadoresalostéricos
s
0 2 4 6 8 10 12
Y
0.0
0.2
0.4
0.6
0.8
1.0
(+ Inhibidor)
(+ Activador)
(sin efectores)
V+
V0
V-

Enzimas reguladoras
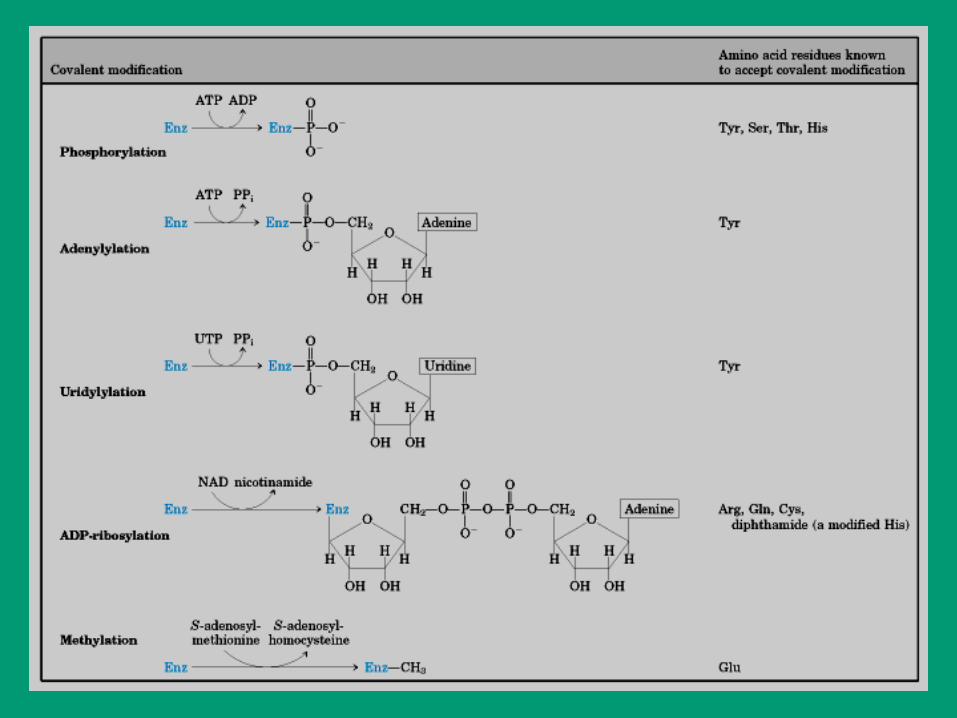
2. Modulación covalente reversible
• Fosforilación
• Adenililación
• Uridililación
• ADP-ribosilación
• Metilación
• Carbamilación

Suele haber fenómenos de modificación covalente de
enzimas en la respuesta celular a señales químicas:
1. Neurotransmisores
2. Hormonas
3. Factores de crecimiento
4. Estímulos morfogenéticos y de diferenciación
5. Muerte celular programada (apoptosis)
6. Estímulos antigénicos
7. Luz y otros agentes físico-químicos
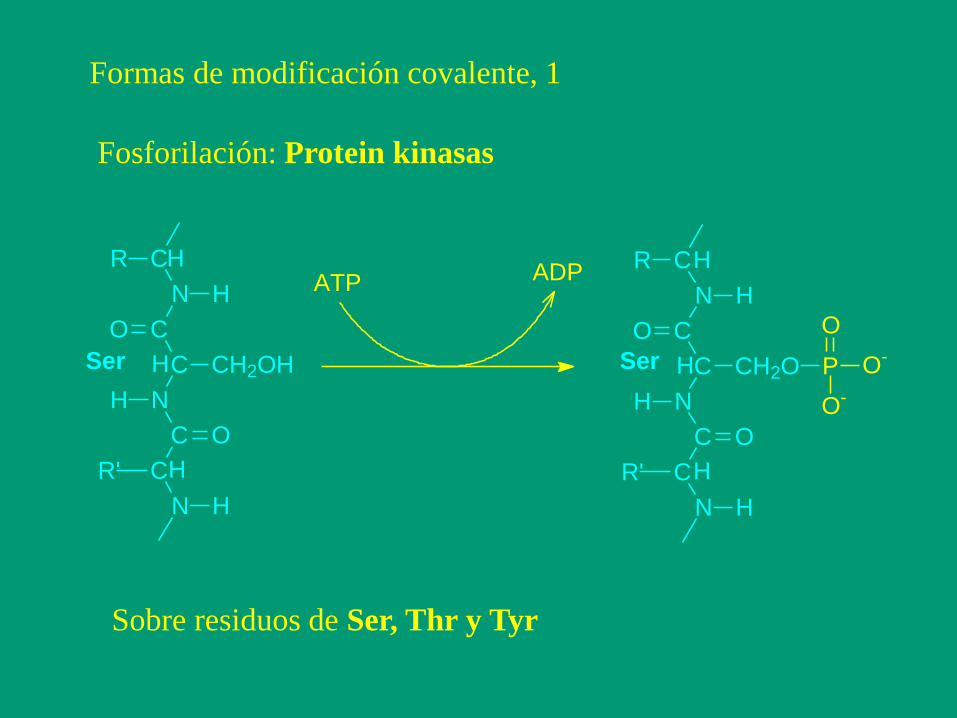
Formas de modificación covalente, 1
Fosforilación: Protein kinasas
Sobre residuos de Ser, Thr y Tyr
Ser
ATP ADP
Ser
C
N
C
C
N
C
C
N
CH2OH
O
H
R
H
O
R'
H
H
H
H
C
N
C
C
N
C
C
N
CH2O
O
H
R
H
O
R'
H
P O-
O
O-
H
H
H
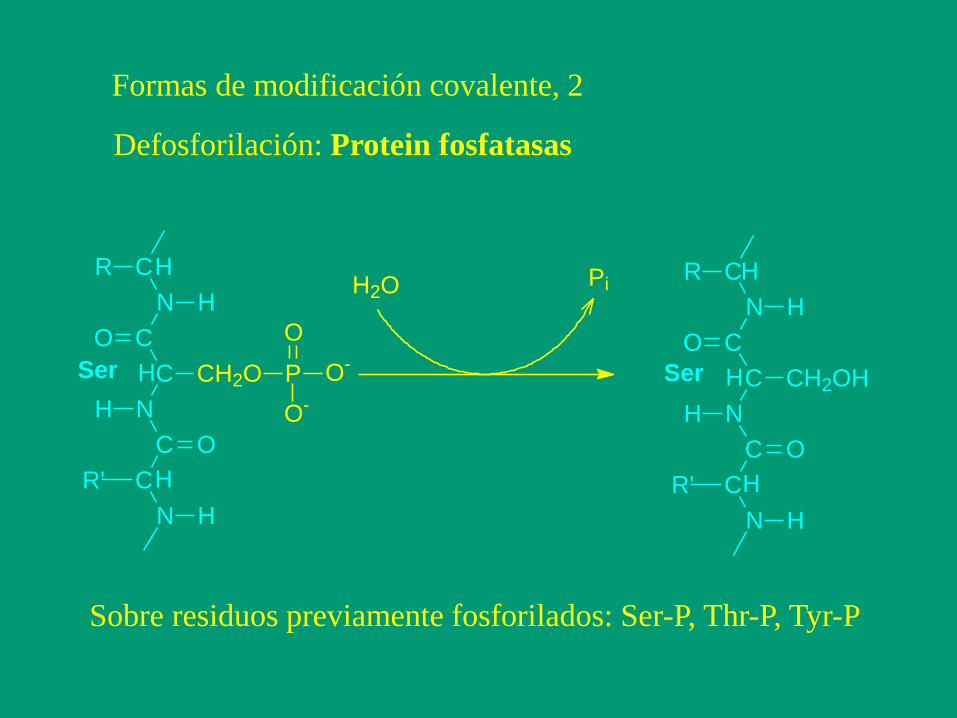
Formas de modificación covalente, 2
Defosforilación: Protein fosfatasas
Sobre residuos previamente fosforilados: Ser-P, Thr-P, Tyr-P
Ser Ser
C
N
C
C
N
C
C
N
CH2OH
O
H
R
H
O
R'
H
H
H
H
C
N
C
C
N
C
C
N
CH2O
O
H
R
H
O
R'
H
P O-
O
O-
H
H
H
H2O Pi
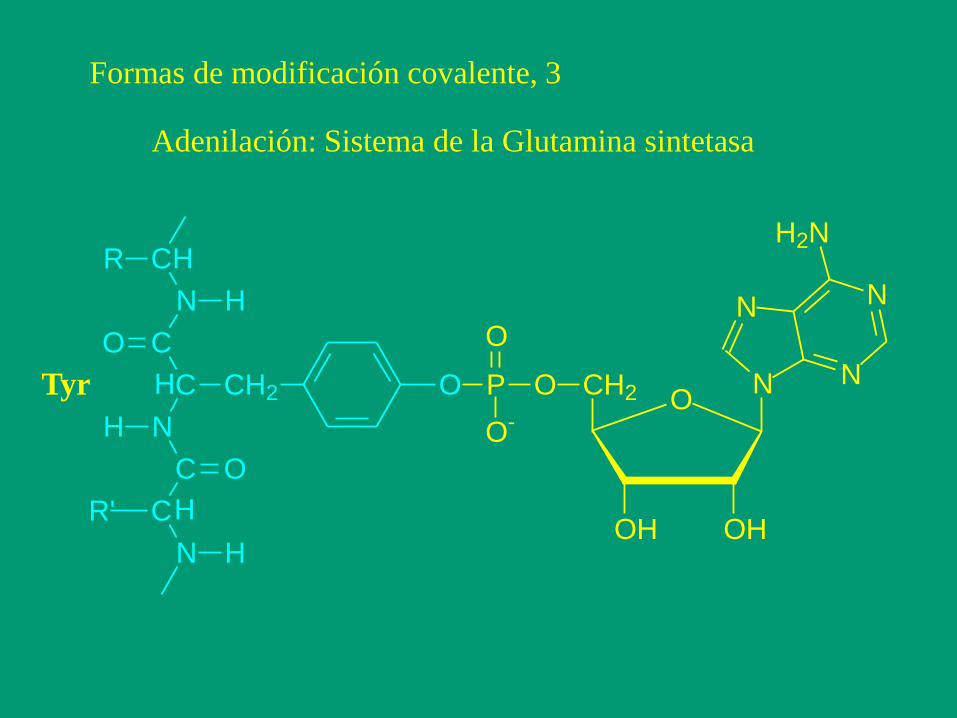
Formas de modificación covalente, 3
Adenilación: Sistema de la Glutamina sintetasa
Tyr O
OH OH
N
N N
N
H2NC
N
C
C
N
C
C
N
O
H
R
H
O
R'
H
CH2 O P
O
O-
O CH2
H
H
H
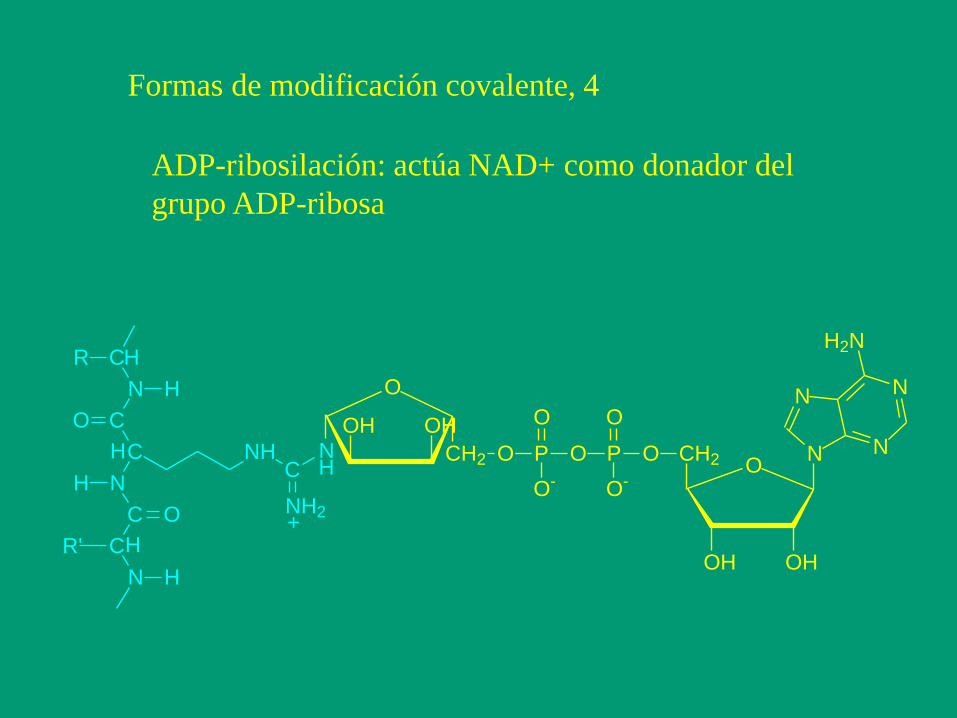
Formas de modificación covalente, 4
ADP-ribosilación: actúa NAD+ como donador del
grupo ADP-ribosa
H
H
H
O
OH OH
N
N N
N
H2N
+
C
N
C
C
N
C
C
N
O
H
R
H
O
R'
H
NHC
NH2
N
O
OH OH
CH2 O P O P O CH2
O O
O-O-H
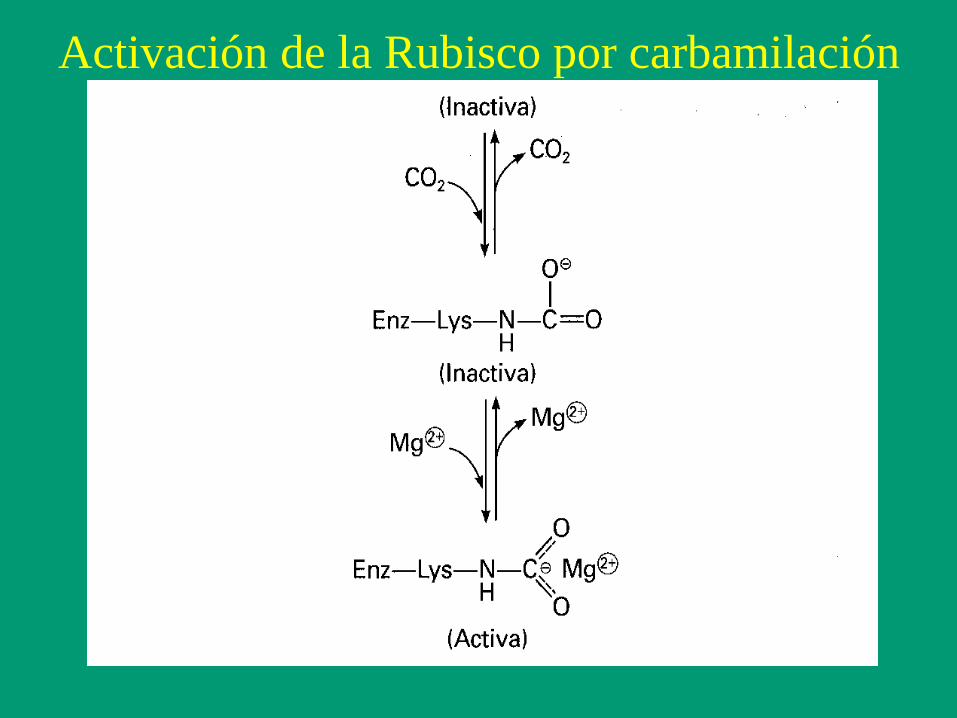
Activación de la Rubisco por carbamilación
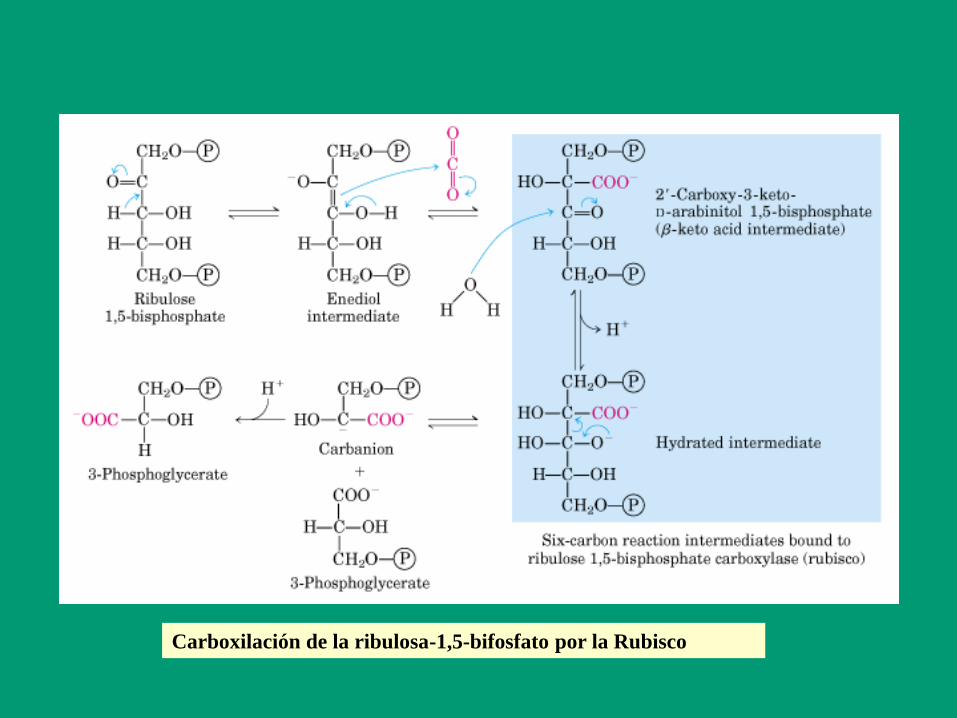
Carboxilación de la ribulosa-1,5-bifosfato por la Rubisco
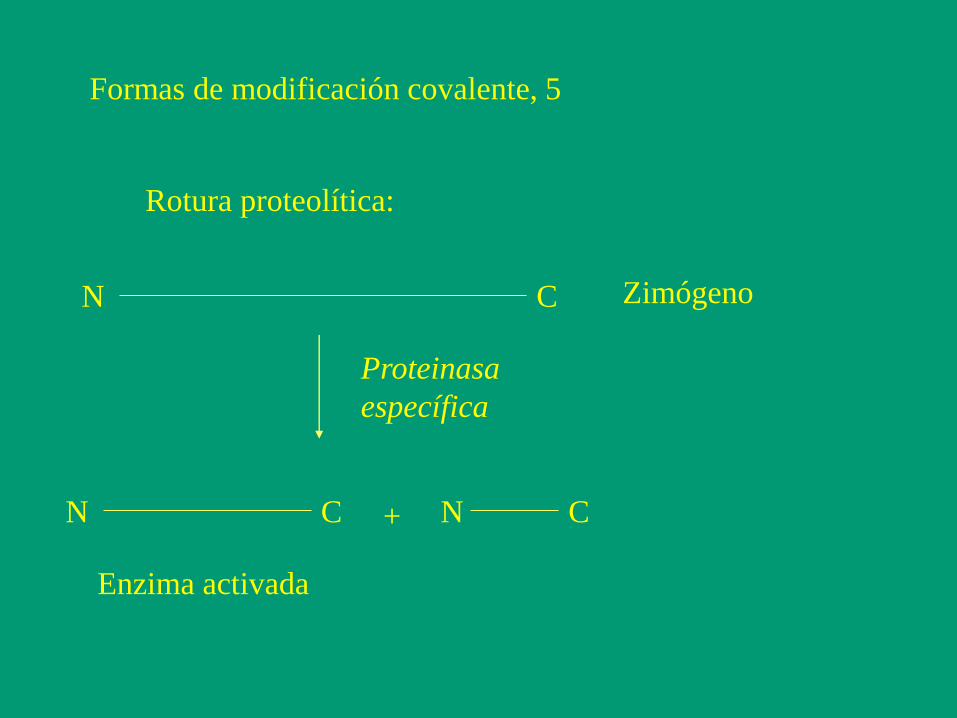
Formas de modificación covalente, 5
Rotura proteolítica:
Proteinasa
específica
+
N C
N N C C
Zimógeno
Enzima activada
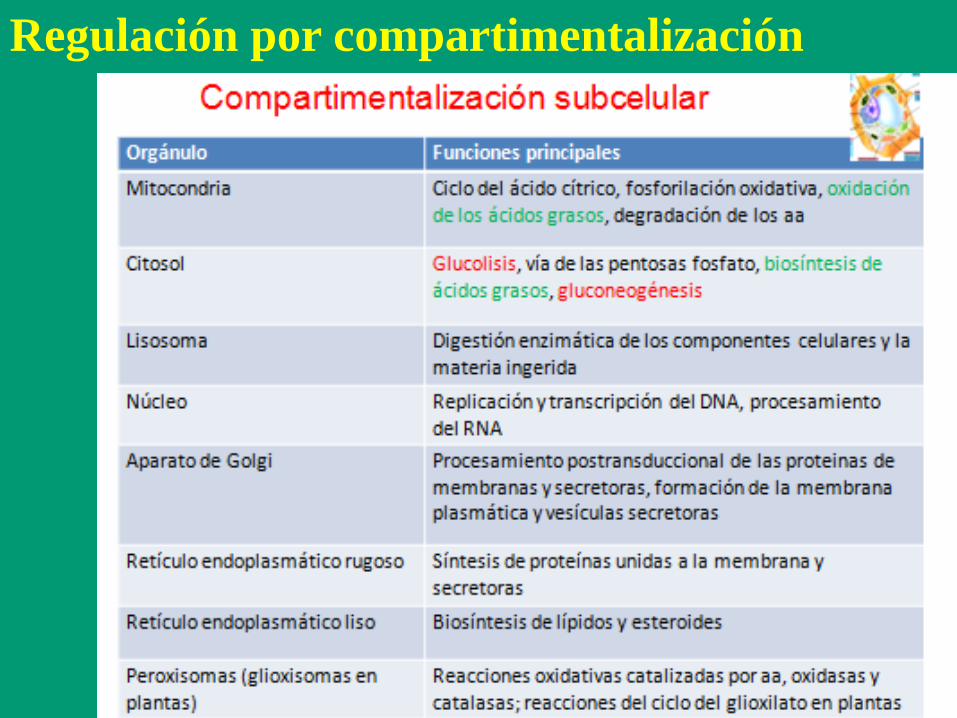
Regulación por compartimentalización


Taninos condensados
Gimnospermas
Monocotiledón
eas
Musgos ,
hepáticas y
helechos
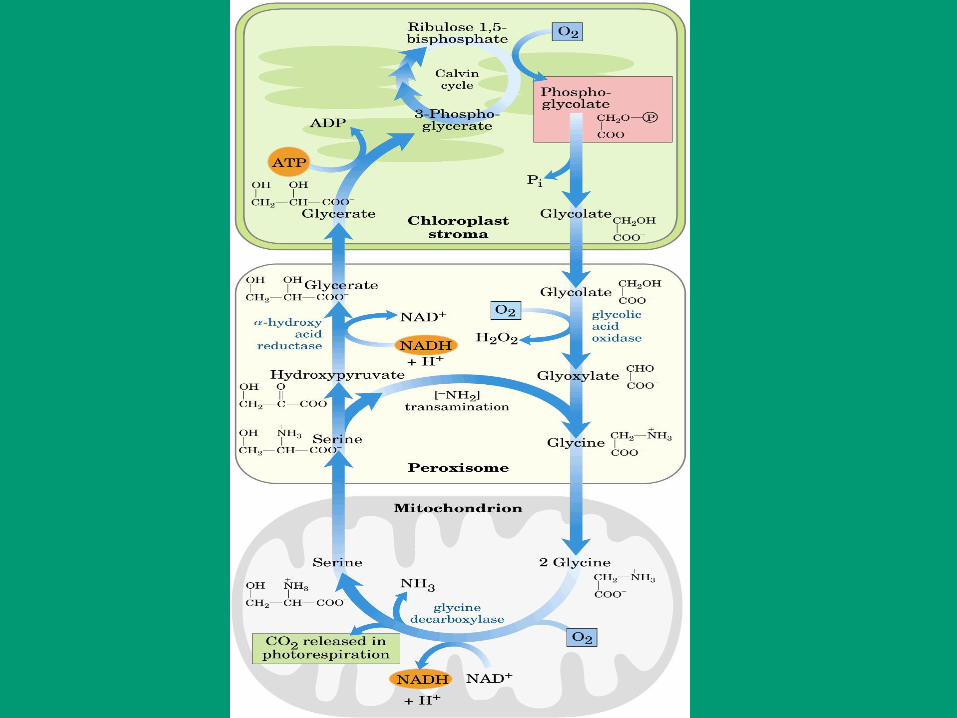
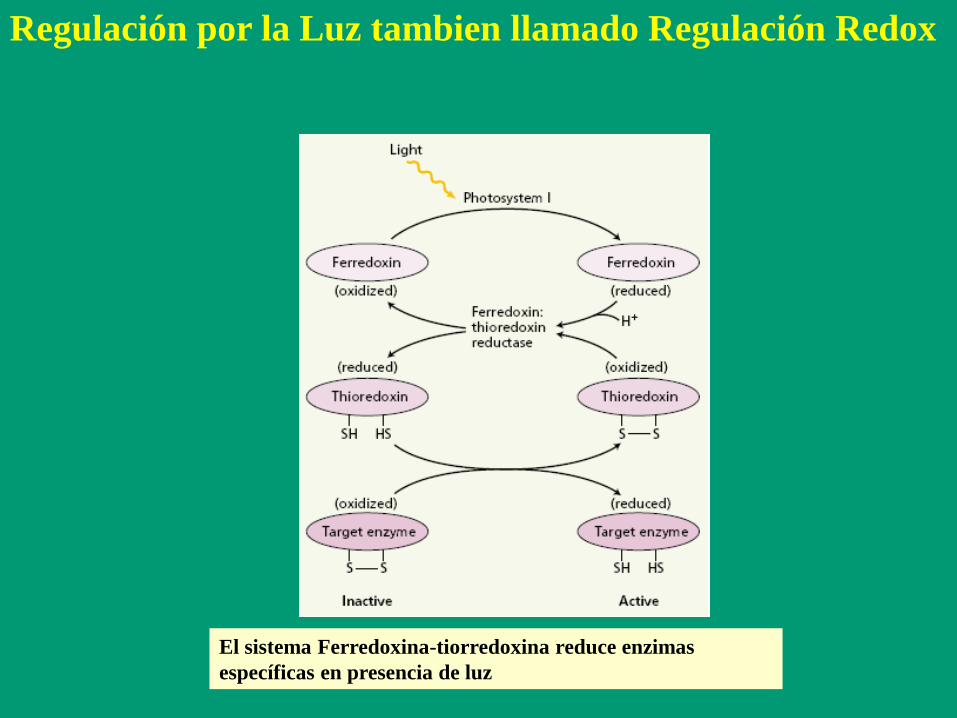
El sistema Ferredoxina-tiorredoxina reduce enzimas
específicas en presencia de luz
Regulación por la Luz tambien llamado Regulación Redox
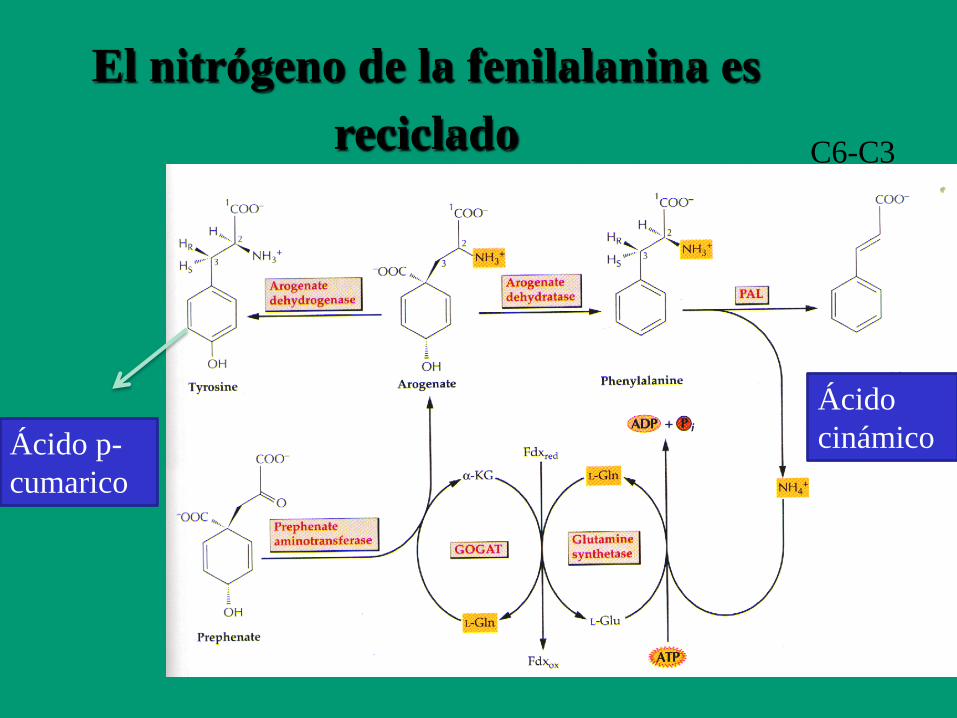
El nitrógeno de la fenilalanina es
reciclado C6-C3
Ácido p-
cumarico
Ácido
cinámico

ENZIMAS REGULADAS POR LUZ
NADP: Gliceraldehído 3-fosfato deshidrogenasa
PAL: Fenilalanina amonio liasa
Fructosa 1,6-bifosfatasa
Ribulosa 5-fosfato quinasa
Sedoheptulosa 1,7-bifosfatasa
Rubisco













![Simulación molecular de la elución de asfaltenos ... · del peso molecular [18], que ha pasado de 2000 g/mol en las últimas décadas, hasta definirse un intervalo de 300 a 1400](https://img.pdfslide.es/doc/110x75/5e50a21559d06b0e374905ce/simulacin-molecular-de-la-elucin-de-asfaltenos-del-peso-molecular-18.jpg)










![Metabolismo de Azufre [Modo de compatibilidad]ecaths1.s3.amazonaws.com/fitoquimicafbqf/437262345...Triptamina Protoalcaloides (fenilalquilaminas) Noradrenalina Mescalina Efedrina y](https://img.pdfslide.es/doc/110x75/5b03d36b7f8b9aba168c6b41/metabolismo-de-azufre-modo-de-compatibilidadecaths1s3-protoalcaloides-fenilalquilaminas.jpg)




