Embed Size (px)
Citation preview

1
EFECTO DE LA FUENTE DE CARBONO Y LA ADICIÓN DE SALES
DE CALCIO Y/O MAGNESIO EN LA PRODUCCIÒN DE
NARINGINASA POR MEDIO DE Aspergillus niger RESUMEN El presente trabajo aporta información del efecto de la naringina, melaza o
ramnosa, como fuentes de carbono, y las sales de Ca2+ y/o Mg2+ en la producción
de naringinasa por medio de Aspegillus niger ATCC1015, el cual es un
microorganismo productor de ácido cítrico y no produce toxinas.
Primero se evaluó por cromatografía en capa fina la capacidad productora de
naringinasa por la cepa empleada, lo cual permitió conocer que el microorganismo
empleado tuviera presuntivamente la capacidad de producir naringinasa. Para
conocer el efecto de las diferentes fuentes de carbono de interés, se empleó un
caldo nutritivo base, adicionado con naringina, ramnosa o melaza, y/o sales de
calcio y magnesio. Las fermentaciones se llevaron a cabo bajo las mismas
condiciones de temperatura y agitación. Cada 24 horas durante los siete días de la
fermentación se cuantificó el peso del micelio, la producción de proteína y la
actividad enzimática de la naringinasa producida, además se determinó mediante
diferentes modelos gráficos las contantes de Vmáx y Km para la naringinasa que
mostró mayor actividad enzimática. Las constates de Vmáx y Km de la naringinasa
obtenida fueron comparadas contra las de la naringinasa comercial de Penicillium
decumbens, la cual también se determinó en este trabajo. A partir de las
investigaciones realizadas en la presente investigación se encontró que la melaza
y la adición de carbonato de calcio favorecieron significativamente el crecimiento
del microorganismo, pero no la producción de proteína y la actividad enzimática.
También se encontró que para favorecer la producción proteica se debe emplear
como fuente de carbono a la naringina y se debe adicionar la mezcla de las sales
de calcio y magnesio. Por otra parte, la fuente de carbono que permitió obtener
una mayor actividad enzimática fue la ramnosa sin adición de sales. Finalmente,

2
se encontró que la naringinasa producida bajo las condiciones de este trabajo
presenta la misma actividad específica que la naringinasa comercial, lo cual la
convierte en una enzima que puede competir con la enzima que se encuentra en
el mercado.

3
INTRODUCCIÓN
La implementación de enzimas, con aplicación en la industria de los alimentos, ha
aumentado en años recientes. Un ejemplo de esto son las glucanasas, utilizadas
en la panificación, la extracción y clarificación de jugos (Villena y Gutiérrez, 2003)
o las enzimas quimosina y pepsina, que forman el cuajo para la fabricación de
quesos, solo por mencionar algunas (Montes y Magaña, 2002; Manzanares y col,
1997).
La aplicación de enzimas en los alimentos presenta una serie de ventajas de
índole económica o tecnológica. La gran especificidad de acción que tienen las
enzimas hace que no se produzcan reacciones laterales imprevistas. Además, las
enzimas pueden inactivarse fácilmente cuando se considere que ya han realizado
su misión, quedando entonces asimiladas el resto de las proteínas presentes en el
alimento (Montes y Magaña, 2002).
La naringinasa es una enzima que está siendo utilizada en la industria de los
alimentos con una aplicación importante en el proceso de desamargor en los jugos
de uva y otras frutas cítricas. La naringinasa es un complejo enzimático que
presenta actividad α-L-ramnosidasa (EC.3.2.1.40) y β-D- glucosidasa
(EC.3.2.1.21) capaz de hidrolizar a la naringina, el cual es el mayor y principal
componente del amargor en jugos de uva y varias frutas cítricas (Thomas y col,
1958; Griffith, 1969; Dunlap y col, 1962; Ono y col, 1978). La naringina es
abundante en frutas inmaduras y su concentración decrece en frutas maduras. Su
presencia ha sido la mayor limitación en la aceptación comercial de los jugos de
frutas. La hidrólisis de la naringina produce naringenina la cual es insípida, lo cual
ayuda a la mejor aceptación de los jugos por la disminución de su amargor (Yusof
y col, 1990).
La información sobre la α-L-ramnosidasa de la naringinasa es limitada y su
producción ha sido asociada con plantas (Hall, 1938; Thomas y col, 1958, Ting,

4
1958; Kaji y Ichimi, 1973), gastrópodos marinos (Kurosawa y col, 1973), y
microorganismos (Manzanares y col, 199, Young y col, 1989; Shanmugam y
Yadav, 1995). Por razones de viabilidad, solo los procesos basados en
naringinasas microbianas son viables (Munish y Uttam, 2000). Tal es el caso de
Aspergillus niger quien es considerado como el mejor productor de naringinasa
extracelular (Kishi, 1955) y Aspergillus terreus quien es bien conocido como un
hongo productor eficiente de enzimas extracelulares de interés para la industria de
alimentos (Gallego y col, 1996).
Existe poca información disponible en cuanto a la producción de la naringinasa y
mucha información está guardada como secreto industrial o está patentada (Ito y
Takiguchi, 1970; Fukumoto y Okada, 1973; Wulbrandt y col, 1995). Algunos
reportes científicos ofrecen una información muy ambigua acerca de las
condiciones de producción de la naringinasa. Se ha reportado que la producción
de naringinasa mediante hongos filamentosos esta favorecida por los iones
metálicos como Ca2+ y Mg2+. Sin embargo, se ha encontrado que dichos iones no
son indispensables (Munish y col, 2005; Gallego y col, 2001). También se ha
reportado que la enzima es reprimida por la presencia de glucosa (Bram y
Salomons, 1965) lactato, citrato, sacarosa, fructosa y almidón (Munish y Uttam,
2000). Por otra parte se ha encontrado que la producción de la enzima disminuye
abajo de pH 4 y es estimulada en presencia de los sustratos naringina, ramnosa,
melaza (Munish y col, 2005; Gallego y col, 2001) y licor de maíz (Munish y Uttam,
2000).
La naringinasa de origen microbiano que existen actualmente en el mercado
presenta una eficacia muy deficiente de la hidrólisis de la naringina, por lo que es
necesario llevar a cabo estudios para conocer las condiciones de producción y
encontrar una enzima naringinasa que presente una mejor capacidad de hidrólisis.
Por ello en este estudio se utilizarán diferentes fuentes de carbono y iones
metálicos para conocer su efecto en la producción de la enzima naringinasa y en

5
su capacidad de hidrólisis. Asimismo, se llevará a cabo una comparación de la
actividad de la naringinasa producida por Aspergillus niger (ATCC 1015), la cual
es una cepa principalmente productora de ácido cítrico, y la naringinasa comercial
pura (producida por Penicillum decumbens) para conocer si la enzima producida
por A. niger presenta mayor capacidad de hidrólisis que la enzima comercial.
6
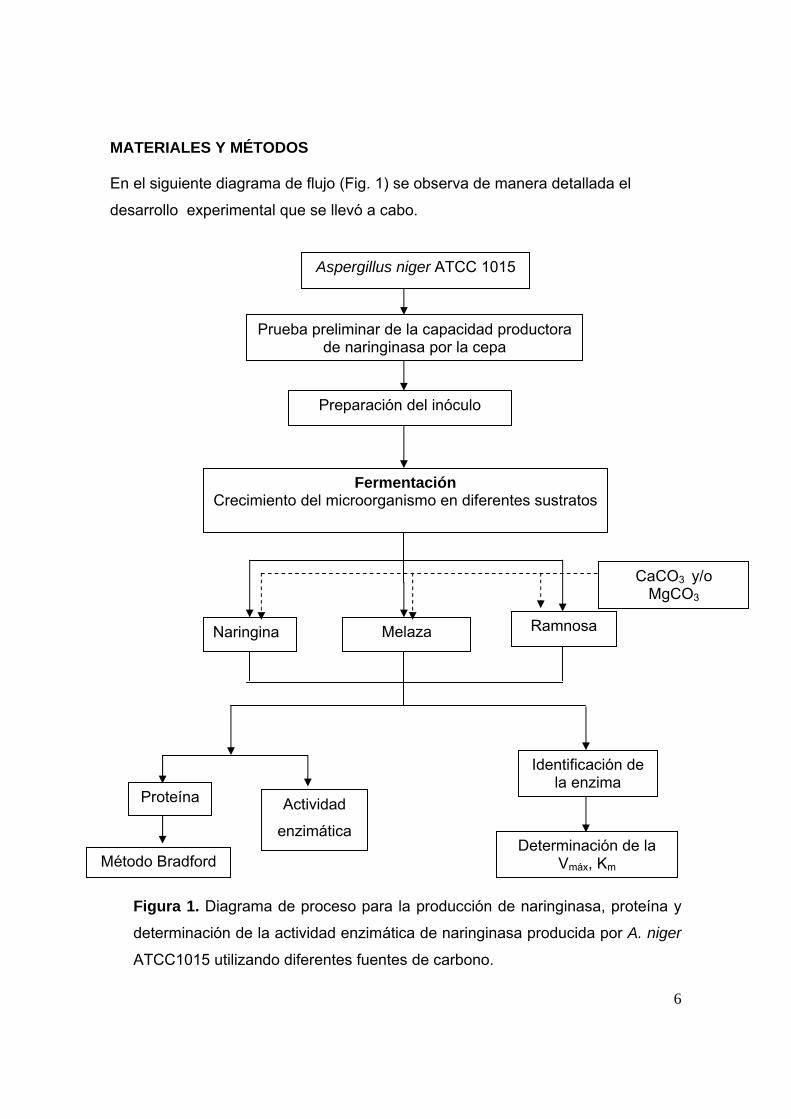
MATERIALES Y MÉTODOS En el siguiente diagrama de flujo (Fig. 1) se observa de manera detallada el
desarrollo experimental que se llevó a cabo.
Figura 1. Diagrama de proceso para la producción de naringinasa, proteína y
determinación de la actividad enzimática de naringinasa producida por A. niger
ATCC1015 utilizando diferentes fuentes de carbono.
Preparación del inóculo
Fermentación Crecimiento del microorganismo en diferentes sustratos
Ramnosa Naringina Melaza
Identificación de la enzima
Proteína
Método Bradford Determinación de la
Vmáx, Km
Prueba preliminar de la capacidad productora de naringinasa por la cepa
CaCO3 y/o MgCO3
Actividad
enzimática
Aspergillus niger ATCC 1015

7
Materias primas.
1. Naringinasa de Penicillium decumbens (Sigma)
2. Naringina (Sigma)
3. Ramnosa (BDDIFCO)
4. Melaza (Ingenio San Miguelito, Córdoba, Ver.)
5. Glucosa (Sigma)
6. Medio de cultivo Agar Saboraud (BDDIFCO)
7. Extracto de malta (BDDIFCO)
• Microorganismo
Aspergillus niger ATCC 1015 obtenido de Colección Nacional de Cepas
Microbianas y Cultivos Celulares del Centro de Investigación y de Estudios
Avanzados (CINVESTAV) y mantenidas a 5ºC en agar papa dextrosa. Este
microorganismo esta reportado como productor de ácido cítrico y antígenos
(CDBB: 177).
Material de laboratorio y reactivos. Ácido ortofosfórico (Baker), acido 3,5-dns (Baker), agujas de jeringa (plastipack),
agua destilada-desionizada, agar maltosa de sabouraud (BDDIFCO), albúmina
sérica bobina (bsa), azul de comassie-g (Baker), cajas petri, cloroformo.
Equipo. Balanza analítica (Ohaus, Explorer® Po), baño metabólico (Shaker, G25), baño
maría (BM), centrifuga ALC4239R, espectrofotómetro visible-UV (Termo
Spectronic, Genesys 20), fotocolorímetro (Klett- Summerson), horno (American),
autoclave, parrilla de calentamiento (Termolyne, HP-A1915B), vortex (Daigger,
Vortex Genie 2).

8
Métodos. Evaluación preliminar de la capacidad productora de naringinasa de la cepa
Aspergillus niger ATCC 1015 por medio de cromatografía en capa fina.
Para conocer si la cepa empleada tenía la capacidad de producir naringinasa, se
utilizaron muestras de jugo de dos lotes de naranjas verdes, un lote sin inocular y
otro inoculado por picadura con Aspergillus niger ATCC 1015, con la finalidad de
evaluar la presencia de naringina en el jugo y la hidrolisis de esta por medio de la
enzima producida por el microorganismo inoculado.
Las muestras de jugo de cada lote fueron tomadas cada 24h y en el caso del lote
de naranjas inoculadas las muestras fueron tomadas del punto de inoculación
debido a que el microorganismo no alcanza a colonizar toda la naranja en 24h. El
lote sin inocular sirvió como control para determinar que, la hidrólisis de la
naringina no fuera efecto de la maduración de la naranja.
Para dar seguimiento a la presencia o ausencia de naringina se utilizó la técnica
de cromatografía en capa fina, para lo cual se cortaron placas cromatográficas de
5 x 2 cm de largo y ancho respectivamente, a las que, se les aplicó una gota de la
muestra problema (naringina pura o jugo de naranja) a una distancia de 0.5 cm del
inicio de la placa cromatográfica (punto de aplicación). Cada placa cromatográfica
con la muestra previamente aplicada, se introdujo en una cámara de vidrio que
contenía 2 mL de disolvente aproximadamente. La muestra aplicada en la placa
ascendió por capilaridad utilizando diferentes disolventes.
Después de que la muestra ascendió por capilaridad y el disolvente recorrió una
distancia de hasta 0.5 cm antes del final de la placa cromatográfica, se sacó la
placa de la cámara de vidrio y se dejó evaporar al ambiente el disolvente que ésta
había absorbido.

9
Para observar la distancia recorrida por las muestras a lo largo de la placa, se
utilizó una lámpara de luz ultravioleta de longitud corta (253 nm) y una de longitud
larga (350 nm), las manchas observadas fueron marcadas con lápiz.
Posteriormente se le aplicó con ayuda de un algodón y unas pinzas sulfato sérico
amoniacal que sirvió como revelador cuando se sometió la placa con el sulfato
sérico amoniacal a calentamiento mediante de una parrilla por 1 min
aproximadamente. El experimento se realizó por triplicado.
Determinación del mejor disolvente que permite la ascendencia por capilaridad de
la naringina pura y la naringina presente en el jugo de naranja en cromatografía en
capa fina.
En primer lugar se empleó una muestra de naringina pura (N) que se empleó
como patrón de comparación en una concentración de 50 mg/1000ml, para poder
hacer los experimentos que permitieran conocer si la cepa empleada tenía
capacidad de producir naringinasa. Por ello se evaluaron las mezclas de
disolventes presentes el Cuadro 1.
La elección del disolvente que permitió la mejor elución de la naringina en
cromatografía en capa fina se realizó en base a los Rf (factor de elución de un
compuesto en una placa cromatográfica) calculados para cada placa
cromatográfica, es decir para cada muestra de naringina eluída utilizando
diferentes disolventes.
El Rf expresa la posición de un compuesto sobre la placa cromatográfica como
una fracción decimal; el máximo Rf es 1 y a mayor Rf mejor factor de elución. El Rf
fue calculado dividiendo la distancia que recorrió la muestra en la placa
cromatográfica entre la distancia que recorrió el disolvente.

10
Cuadro 1. Disolventes utilizados para determinar el factor de elución de una
muestra de naringina pura en cromatografía en capa fina.
Disolvente / Concentración %
hexano / 100
acetato / 100
hexano: acetato de etilo / 80:20
acetato de etilo: hexano / 80:20
metanol: acetato de etilo / 80:20
metanol / 100.
acetona / 100
metanol: acetona / 95:5
metanol: acetona / 90:10
metanol: acetona / 80:20
Además se utilizaron diferentes disolvente o mezcla de disolventes para conocer
la elución de la naringina presente en el jugo de naranja (n) en una placa
cromatográfica las cuales se muestran en el Cuadro 2.
Cuadro 2. Disolventes y proporción utilizados para determinar el factor de elución
de la naringina presente en el jugo de naranja.
Disolvente / Concentración %
metanol / 100
acetona / 100
metanol: acetona / 95:5
metanol: acetona / 90:10
metanol: acetona / 80:20

11
Se buscó el mejor sistema de elución tanto para la naringina pura (N) como para la
naringina presente en el jugo de naranja (n). Para ello se evaluaron las mezclas de
disolventes presentes en el Cuadro 3.
Cuadro 3. Disolventes y proporción utilizados para determinar el factor de elución
de la naringina pura y de la naringina presente en el jugo de naranja en una misma
placa cromatográfica.
Recuperación de la cepa de A. niger a partir de un cultivo en tubo inclinado.
La cepa Aspergillus niger ATCC 1015 (obtenida del cepario del Cinvestav) se
adquirió en tubos de ensaye en agar papa dextrosa, por lo que fue resembrada
para obtener una cantidad suficiente para que pudiera emplearse en las
fermentaciones a realizar (Díaz y col, 1998).
Preparación del inóculo de A. niger por medio de turbidimetria.
Para la obtención del inóculo a emplear en cada fermentación se preparó para
cada ensayo y en el momento del inicio de la fermentación una solución que
contenía 900 x 106 esporas/ mL por medio de turbidimetria utilizando una escala
de MacFarland previamente preparada como se muestra en el Cuadro 4 (Díaz y
col, 1998).
Disolvente / Concentración %
metanol / 100
metanol: acetona / 80:20
12
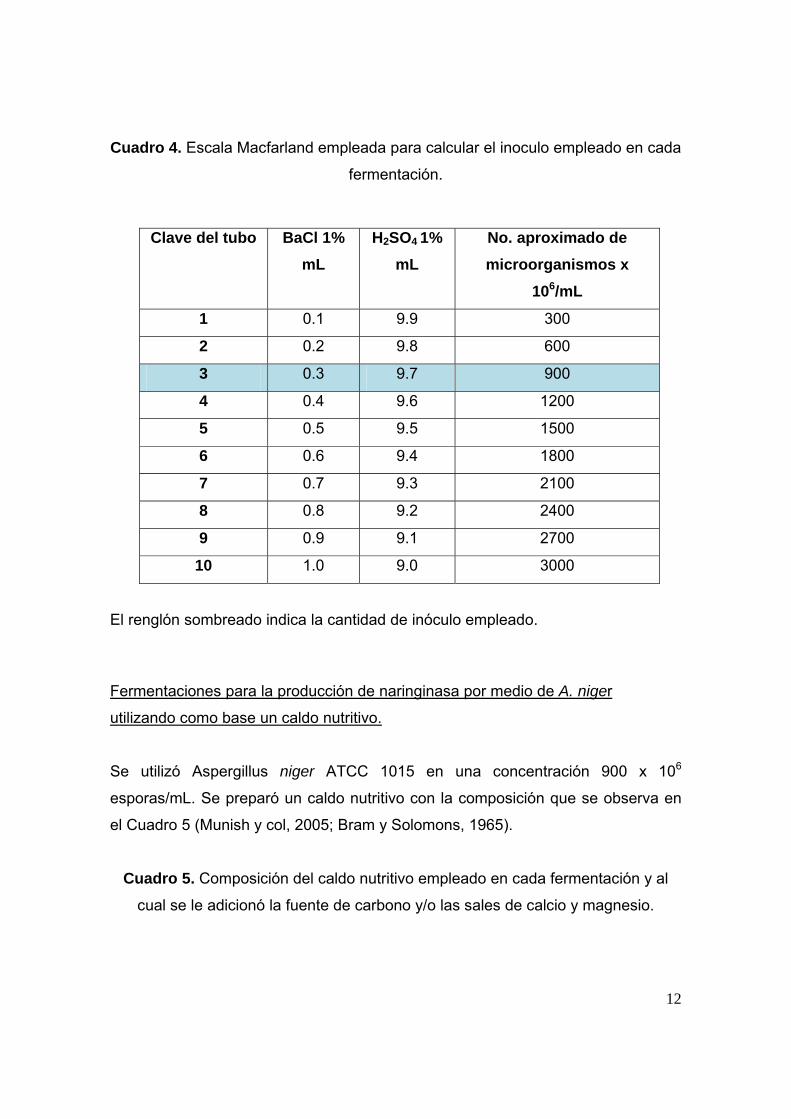
Cuadro 4. Escala Macfarland empleada para calcular el inoculo empleado en cada
fermentación.
El renglón sombreado indica la cantidad de inóculo empleado.
Fermentaciones para la producción de naringinasa por medio de A. niger
utilizando como base un caldo nutritivo.
Se utilizó Aspergillus niger ATCC 1015 en una concentración 900 x 106
esporas/mL. Se preparó un caldo nutritivo con la composición que se observa en
el Cuadro 5 (Munish y col, 2005; Bram y Solomons, 1965).
Cuadro 5. Composición del caldo nutritivo empleado en cada fermentación y al
cual se le adicionó la fuente de carbono y/o las sales de calcio y magnesio.
Clave del tubo BaCl 1% mL
H2SO4 1% mL
No. aproximado de microorganismos x
106/mL
1 0.1 9.9 300
2 0.2 9.8 600
3 0.3 9.7 900
4 0.4 9.6 1200
5 0.5 9.5 1500
6 0.6 9.4 1800
7 0.7 9.3 2100
8 0.8 9.2 2400
9 0.9 9.1 2700
10 1.0 9.0 3000
13
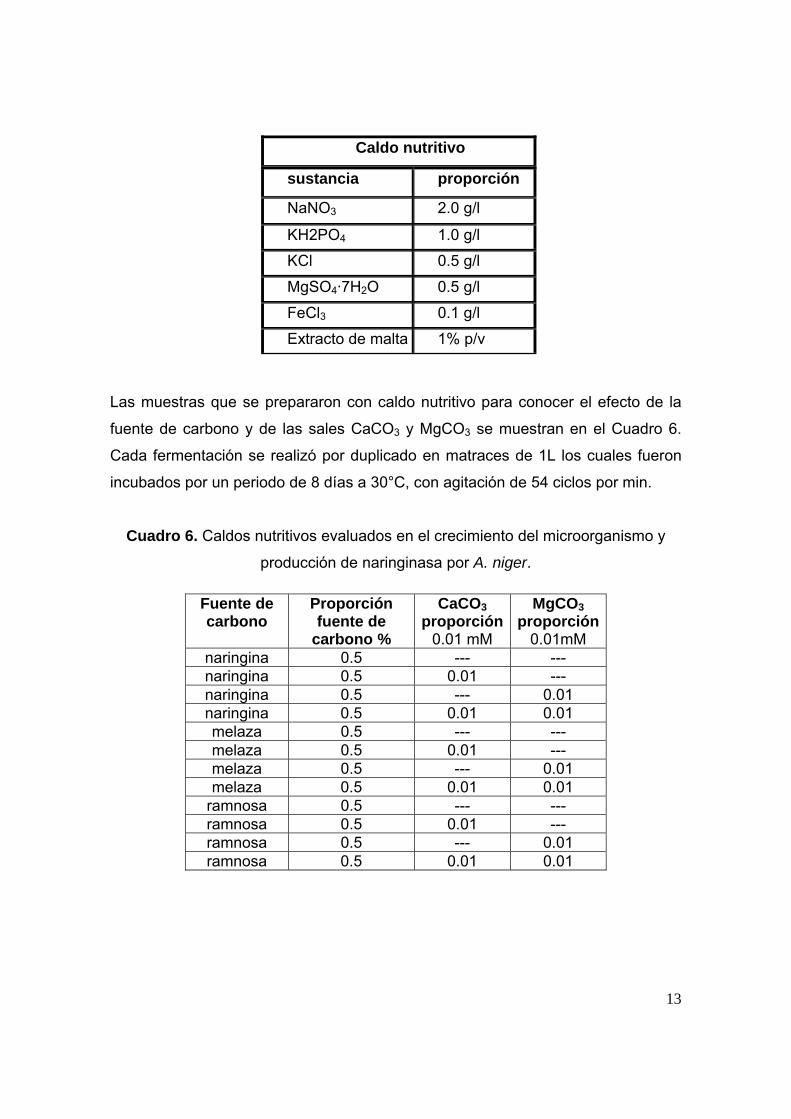
Las muestras que se prepararon con caldo nutritivo para conocer el efecto de la
fuente de carbono y de las sales CaCO3 y MgCO3 se muestran en el Cuadro 6.
Cada fermentación se realizó por duplicado en matraces de 1L los cuales fueron
incubados por un periodo de 8 días a 30°C, con agitación de 54 ciclos por min.
Cuadro 6. Caldos nutritivos evaluados en el crecimiento del microorganismo y
producción de naringinasa por A. niger.
Caldo nutritivo
sustancia proporción
NaNO3 2.0 g/l
KH2PO4 1.0 g/l
KCl 0.5 g/l
MgSO4·7H2O 0.5 g/l
FeCl3 0.1 g/l
Extracto de malta 1% p/v
Fuente de carbono
Proporción fuente de
carbono %
CaCO3 proporción
0.01 mM
MgCO3 proporción
0.01mM naringina 0.5 --- --- naringina 0.5 0.01 --- naringina 0.5 --- 0.01 naringina 0.5 0.01 0.01 melaza 0.5 --- --- melaza 0.5 0.01 --- melaza 0.5 --- 0.01 melaza 0.5 0.01 0.01
ramnosa 0.5 --- --- ramnosa 0.5 0.01 --- ramnosa 0.5 --- 0.01 ramnosa 0.5 0.01 0.01

14
Crecimiento del microorganismo utilizando diferentes fuentes de carbono.
Se tomó una alícuota de 20 mL cada día que duró la fermentación, esta fue filtrada
en papel filtro de poro de 1mm y se secó a 80ºC por 10 min aproximadamente, así
se cuantificó el peso del micelio por diferencia (Bucio Villalobos y col, 2007).
MÉTODOS ANALÍTICOS. Determinación de proteína en el medio de cultivo.
Para la determinación de la proteína en el medio de cultivo se utilizó el método de
Bradford, el cual es un método rápido y sensible para la cuantificación de micro
cantidades de proteína utilizando el principio de formación de color por unión a la
proteína (Bradford, 1976; Kirk y col, 2004).
El método de Bradford involucra la unión no covalente de la proteína al reactivo
Azul brillante de Coomasie G-250 ya que este interacciona con los grupos básicos
y aromáticos de las proteínas. Este colorante absorbe a una longitud de onda de
365 sin embargo la unión de la proteína al colorante causa un cambio en el
máximo de absorción del colorante de 365 a 595nm absorbancia a la que se
realiza la determinación (Bradford, 1976).
- Preparación del reactivo de Bradford.
Se disuelven 100 mg de azul de Comassie en 50 mL de etanol al 96 %, después
se añaden 100 mL de ácido fosfórico 85 %.Se diluye con 1000 mL de H2O
destilada y se deja reposar 24 h en oscuridad, se filtra dos veces con papel filtro y
se conserva en una botella oscura.
-Patrón de albúmina sérica bobina (ASB).
Se realiza una curva de ASB de acuerdo con el Cuadro 7. Agitando
perfectamente y después de incubar por 10 minutos a temperatura ambiente.
Cada ensayo se lee a 595 nm usando como blanco el tubo 6 de la serie y se
determinan por triplicado. Esta curva solo es estable por 30 min y se deberá agitar

15
perfectamente antes de leer. Para determinar el contenido de la proteína en
cuestión se realiza el mismo procedimiento sustituyendo la ASB por la proteína
problema e interpolando los resultados obtenidos para la proteína problema en la
curva estándar. Durante la determinación se desarrolla una coloración azulada
que se incrementa conforme se incrementa la concentración de proteínas.
Cuadro 7. Curva tipo de albumina sérica bovina, para la determinación de
proteína en el medio de cultivo por el método de Bradford.
Tubo Solución Proteína (ASB, 250 μg/mL)
Agua destilada (mL)
Reactivo de Bradford (mL)
1 0.1 2.9 3 2 0.2 2.8 3 3 0.3 2.7 3 4 0.4 2.6 3 5 0.5 2.5 3 6 0.0 3 3
La curva tipo de albumina sérica bovina, para la determinación de proteína en el
medio de cultivo por el método de Bradford.se reporta en el Anexo número 1 del
presente trabajo y presentó una ecuación de la recta de Y=5.78x + 0.1408;
R2=0.9579.
Determinación de la actividad enzimática de la naringinasa comercial de
Penicillium decumbens.
Para llevar a cabo la determinación de la naringinasa producida por el
microorganismo empleado fue importante conocer el comportamiento de esta in
vitro por tanto se realizaron las siguientes determinaciones utilizando naringinasa
de Penicillium decumbens (Sigma).
Estudio del efecto de la concentración de naringinasa de Penicillium decumbens
en la hidrólisis de naringina.

16
Se preparó una solución de naringina 0.005 mM en regulador de citratos 0.05 M
pH 4.5 de la cual siempre se adicionó 1.9 mL a cada tubo de ensayo, además de
diferentes soluciones de naringinasa de concentración final de 10, 30, 50, 75, 100,
125 y 150 μg/mL; las cuales fueron elaboradas a partir de una solución
concentrada de 1500 μg/ mL, siendo siempre la alícuota que se adicionó al
sustrato de 0.1 mL. Así el volumen total de cada tubo de ensayo fue de 2 mL.
Cada ensayo se incubo durante 15 minutos, tiempo en el cual se llevaría a cabo la
hidrólisis de naringina por la naringinasa y se obtendría glucosa como producto
final y esta es directamente proporcional a la cantidad de naringenina producida
por dicha hidrólisis. Terminado el tiempo de reacción, la reacción se detiene con la
adición de 4 mL de DNS y se calienta durante 5 minutos a ebullición para revelar
cada tubo, los cuales una vez fríos fueron leídos a 540nm con filtro verde. (Ting,
1958; Olson y col, 1979; Olson y Gray, 1981; Chaplin y Kennedy, 1986). Los
resultados de este estudio se reporta en el Anexo 2 apartado 2a.1 del presente
trabajo.
Estudio del efecto del tiempo en la velocidad de hidrólisis de la naringina por la
naringinasa de Penicillium decumbens.
Del estudio anterior se seleccionó una concentración de naringinasa a la cual se
observara claramente la hidrólisis de la naringina la cual fue 50 μg/ mL, dicha
concentración se mantuvo constante durante todo el experimento donde se
evaluaron diferentes tiempos de hidrolisis los cuales fueron 5,10, 20, 30, 40, 50, 60
minutos adicionando a cada tubo de ensayo 1.9 mL de naringina 0.005M y 0.1 mL
de naringinasa. Terminado el tiempo de reacción, la reacción se detuvo con la
adición de 4 mL de DNS y se calentó durante 5 minutos para revelar cada tubo,
los cuales una vez fríos fueron leídos en el fotocolorímetro con filtro verde. (Ting,
1958; Olson y col, 1979; Olson y Gray, 1981; Chaplin y Kennedy, 1986). Los
resultados de este estudio se reporta en el Anexo 2, apartado 2a.2 del presente
trabajo.

17
-Curva tipo de naringina.
Para el montaje de la curva tipo de naringina una vez conocidos tanto la
concentración de naringinasa a emplear (50 μg/ mL) y el tiempo de hidrólisis más
adecuado para observar tal efecto (10 min), se preparó una solución de
naringinasa en concentración de 50 μg/ mL en regulador de citratos 0.005 mM pH
4.5 de la cual se adición 0.5mL, la concentración de naringina empleada fue de
0.005M y se emplearon los siguientes volúmenes: 0.05, 0.1, 0.2, 0.4, 0.6, 0.8, 1,
1.2, 1.4, 1.6 y 1.8 mL para alcanzar un volumen final de cada ensayo de 2.5 mL. El
tiempo de incubación fue de 10 minutos, posteriormente la reacción fue detenida
agregando 4 mL de DNS y los tubos fueron puestos a ebullición durante 5
minutos, una vez fríos se llevo a cabo la lectura a 450nm con filtro verde. (Ting,
1958; Olson y col, 1979; Olson y Gray, 1981; Chaplin y Kennedy, 1986). Los
resultados de este estudio se reporta en el Anexo 3, Figura 4A del presente
trabajo.
-Determinación de las constantes de Vmáx y Km para la naringinasa de
P. decumbens por los métodos gráficos de Michaelis-Menten, Lineweaber-Burk,
Eadie-Hofsteen, Agustinson y Wilkinson.
Con base en base los resultados obtenidos de la curva de naringinasa y haciendo
una curva patrón de glucosa se calcularon las concentraciones reales en μg/ ml
que representa cada volumen empleado en la curva tipo de naringina 0.005M.
Dichos resultados fueron empleados para trazar las curvas de Michaelis- Mentes
(Anexo 5, Figura 6A), Lineweaber-Burk (Anexo 5, Figura 7A), Eadie-Hofsteen
(Anexo 5, Figura 8A) y Agustinson (Anexo 5, Figura 9A), y se determino la Vmáx y
Km para cada uno, a demás se determinaron dichas constantes por el método
estadístico de Wilkinson (1960). La Vmáx esta expresada en U/mg proteína donde
una U = cantidad de enzima para liberar 1μmol de glucosa/ min.

18
Estudio de la actividad enzimática de la naringinasa producida por A. niger en
muestras de la fermentación de cada formulación.
Para llevar a cabo la determinación de la actividad enzimática de la naringinasa
producida por el microorganismo empleado. Se tomó 0.5 mL de cada muestra de
la fermentación de cada formulación, se le adicionó 1.5 mL de una solución de
naringina (Sigma) 5.26 mM en regulador de citratos 0.05M pH 6, como sustrato, y
se incubó a 35°C por 24 h, la reacción fue detenida adicionando 4 mL de ácido
3,5-dinitrosalicilico y con ebullición en baño María durante 5 minutos,
posteriormente se leyó a 450 nm en un fotocolorímetro con filtro verde. Todos los
ensayos se realizaron por triplicado. Como blanco se utilizó 1.5 mL de una
solución de naringina 5.26mM. Para el tratamiento de los resultados se utilizó una
curva tipo de glucosa con una ecuación de la curva de Y=0.6456x -2.2889;
R2=0.987, reportada en el Anexo 4, Figura 5A del presente trabajo.
Estudio de la actividad enzimática de la naringinasa producida por A. niger del
caldo nutritivo que presentó mayor actividad enzimática en el estudio anterior.
A partir de los resultados obtenidos en el estudio anterior se seleccionó el caldo
nutritivo seleccionado que presentó mayor actividad enzimática.
El caldo nutritivo elegido, fue tratado para concentrar la proteína y separarla del
medio de cultivo por lo que se le adicionó 65 g de sulfato de amonio por cada 100
mL de medio de cultivo para precipitar la proteína en baño de hielo logrando una
saturación de 99% de acuerdo con Dawson (1968). La solución fue centrifugada a
17100 rpm (107442.46875277 radianes /min) 4°C por 30 min; el pellet fue re-
suspendido en 15 mL de regulador de citratos 0.05M pH 6 y se le determinó el
contenido proteico por medio del método de Bradford.
Para la determinación de la actividad enzimática del concentrado proteico obtenido
se emplearon 1.5 mL de una solución de naringina de diferente molaridad (0.02,
0.008, 0.00526, 0.0026, 0.0013, 0.001, 0.0006 M) en regulador de citratos 0.05 M
pH 6 a la que se le adicionaron 0.5 mL del concentrado proteico y se incubaron a

19
35°C por 24 h en baño María, una vez transcurrido el tiempo de incubación la
reacción fue detenida adicionando 4 mL de ácido 3,5-dinitrosalicilico y con
ebullición en baño María durante 5 minutos, posteriormente la lectura se realizó en
un fotocolorímetro con filtro verde. Todos los ensayos se realizaron por triplicado.
Para el tratamiento de los resultados se utilizó una curva tipo de glucosa con una
ecuación de la curva de Y=0.6456x -2.2889; R2=0.987, reportada en el Anexo 4,
Figura 5A del presente trabajo. Los resultados obtenidos permitieron calcular los
gráficos de Michaelis-Menten, Lineweaer-Burk, Eadie-Hoffsten y Agustinson. A
demás se determinó la Vmáx y Km para cada uno. El valor de Vmáx y Km para
Wilkinson se calculó de acuerdo al modelo estadístico de Wilkinson (1960).
Análisis estadístico
Los resultados de peso del micelio, proteína y actividad enzimática fueron
analizados estadísticamente por medio del programa MINITAB13, a través de un
análisis de varianza (ANOVA) con un p ≤ 0.05 utilizando un diseño factorial
general completo de 3x4x5 (fuente de carbono, sales y días) con tres repeticiones.

20
RESULTADOS Y DISCUSIÓN
Evaluación de la capacidad productora de naringinasa de la cepa Aspergillus niger
ATCC 1015 por medio de cromatografía en capa fina.
Se encontró que la naringina pura (N) y la naringina presente en la naranja (n),
ascendieron por capilaridad en metanol 100% alcanzando un Rf= 0.86, pero su
ascendencia por capilaridad mejoró con metanol: acetona en 80:20 presentando
un Rf= 1 el cual fue calculado con respecto a la muestra de referencia lo que nos
sugiere que el mejor sistema para permitir la ascendencia por capilaridad en
cromatografía en capa fina de las muestras de naringina pura y naringina de la
naranja es el de metanol: acetona en 80:20. Por tanto para la siguiente etapa
experimental se trabajo con este sistema de disolventes.
Evaluación de la degradación de la naringina por la naringinasa producida por
Aspergillus niger ATCC 1015 una vez inoculado en naranjas.
Pequeñas muestras de jugo de las naranjas utilizadas en el experimento (n) se
compararon en una placa cromatográfica contra una muestra de naringina pura
(N) tanto de las naranjas inoculadas por picadura con Aspergillus niger ATCC
1015 como las no inoculadas (controles). Después de revelar las placas se
confirmó la presencia de la naringina en el lote de naranjas sin inocular y de igual
manera se confirmo la presencia de la naringina en el lote de naranjas acabadas
de inocular.
A las 48 y 72 horas se observó la presencia de naringina en las naranjas no
inoculadas junto con la presencia de otros compuestos, lo que podría sugerir el
inicio de la degradación de la naringina presente en la naranja probablemente por
efecto de la maduración. A 120 hrs los controles mostraron la presencia de
naringina junto con la presencia de otros compuestos que solo absorven en
longitud de onda larga en el UV. En las muestras de las naranjas inyectadas con

21
el hongo no se observa la presencia de naringina probablemente debido a una
hidrólisis inducida por el microorganismo inoculado.
preparación del inóculo de A. niger a partir de un cultivo en tubo inclinado.
La cepa Aspergillus niger ATCC 1015 (obtenida del cepario del CINVESTAV; fue
resembrada en agar Sabouraud el cual fue preparado, esterilizado y vaciado
previamente en cajas petri, las cuales fueron resembradas por picadura. Dichas
cajas se mantuvieron a temperatura ambiente y cubiertas con papel aluminio por
una semana para obtener un óptimo crecimiento del microorganismo. El hongo
comenzó a crecer desde el primer día y alcanzo su óptimo a los 7 días de haberse
sembrado. Este procedimiento se realizó mensualmente a manera de conservar la
cepa. El caldo nutritivo, como fue descrito en materiales y métodos, fue adicionado
con diferentes fuentes de carbono (0.5% p/v) y con sales de calcio y magnesio al 1
M y 0.1mM. Sin embargo, en estas muestras no se observó el crecimiento del
hongo (peso del micelio).
Crecimiento del microorganismo utilizando diferentes fuentes de carbono.
Se observó crecimiento del microorganismo cuando se empleó como fuente de
carbono naringina, ramnosa o melaza (0.5% p/v) en el caldo nutritivo. Cuando se
adición las sales de calcio y/o magnesio en concentración de 0.01mM se observó
crecimiento del microorganismo al contrario de cuando se utilizó una
concentración de 1 y 0.1mM. Por ello se empleó en todos los caldos nutritivos que
se trabajaron en este estudio una concentración de 0.01 mM para las sales.
Es importante resaltar que cuando no se adicionó sal alguna al medio de cultivo,
no se encontró diferencia significativa para las tres diferentes fuentes de carbono
empleadas. Lo cual concuerda con Bramn y Solomons (1965) que sugieren que no
es necesaria la adición de factores de crecimiento para Aspergillus niger NRRL
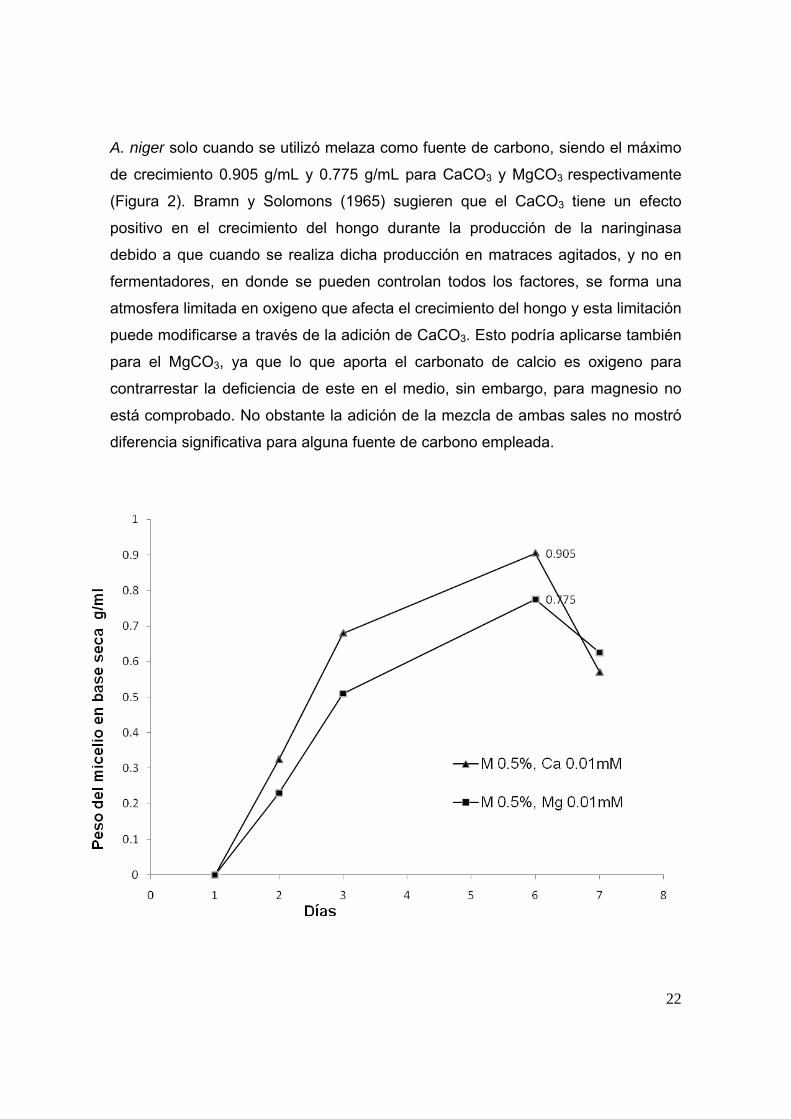
72-4 en la producción de naringinasa. Sin embargo, la presencia de CaCO3 y
MgCO3 individualmente favoreció significativamente (p≤0.05) el crecimiento de
22
A. niger solo cuando se utilizó melaza como fuente de carbono, siendo el máximo
de crecimiento 0.905 g/mL y 0.775 g/mL para CaCO3 y MgCO3 respectivamente
(Figura 2). Bramn y Solomons (1965) sugieren que el CaCO3 tiene un efecto
positivo en el crecimiento del hongo durante la producción de la naringinasa
debido a que cuando se realiza dicha producción en matraces agitados, y no en
fermentadores, en donde se pueden controlan todos los factores, se forma una
atmosfera limitada en oxigeno que afecta el crecimiento del hongo y esta limitación
puede modificarse a través de la adición de CaCO3. Esto podría aplicarse también
para el MgCO3, ya que lo que aporta el carbonato de calcio es oxigeno para
contrarrestar la deficiencia de este en el medio, sin embargo, para magnesio no
está comprobado. No obstante la adición de la mezcla de ambas sales no mostró
diferencia significativa para alguna fuente de carbono empleada.

23
Figura 2. Crecimiento del micelio de A. niger ATCC1015 en melaza con adición de
CacO3 (▲) y melaza con adición de MgCO3 (■).
Determinación de proteína en el medio de cultivo
La determinación de la proteína durante el crecimiento del microorganismo (no
toda la proteína producida es enzima naringinasa) se realizó por triplicado para
todas las muestras tomadas cada 24 horas durante el transcurso de la
fermentación y se utilizó como blanco la fuente de proteína inicial (1% p/v).
Para la producción de proteína se encontró que existe diferencia significativa
(p≤0.05) dependiendo de la fuente de carbono que se utilice, siendo la naringina la
fuente de carbono que favorece más la producción proteica, lo cual se observa en
la Figura 3, Curva A.
También se encontró diferencia significativa en la producción de proteína
dependiendo de la presencia o ausencia de sales de Ca2+ y/o Mg2+. En la Figura 3,
Curva B se observa que la adición de las sales favorece positivamente la
producción de proteína, teniendo un mayor efecto positivo el MgCO3 que el
CaCO3, sin embargo, se observa que el efecto positivo se incrementa cuando se
adiciona la mezcla de sales de Ca2+ y Mg2+ y cuando se utiliza naringina como
fuente de carbono (Figura 34, Curva A). Probablemente el efecto de la mezcla de
las sales es mayor porque al adicionarlas juntas suman su efecto actuando de
manera sinergica sobre la producción proteica.
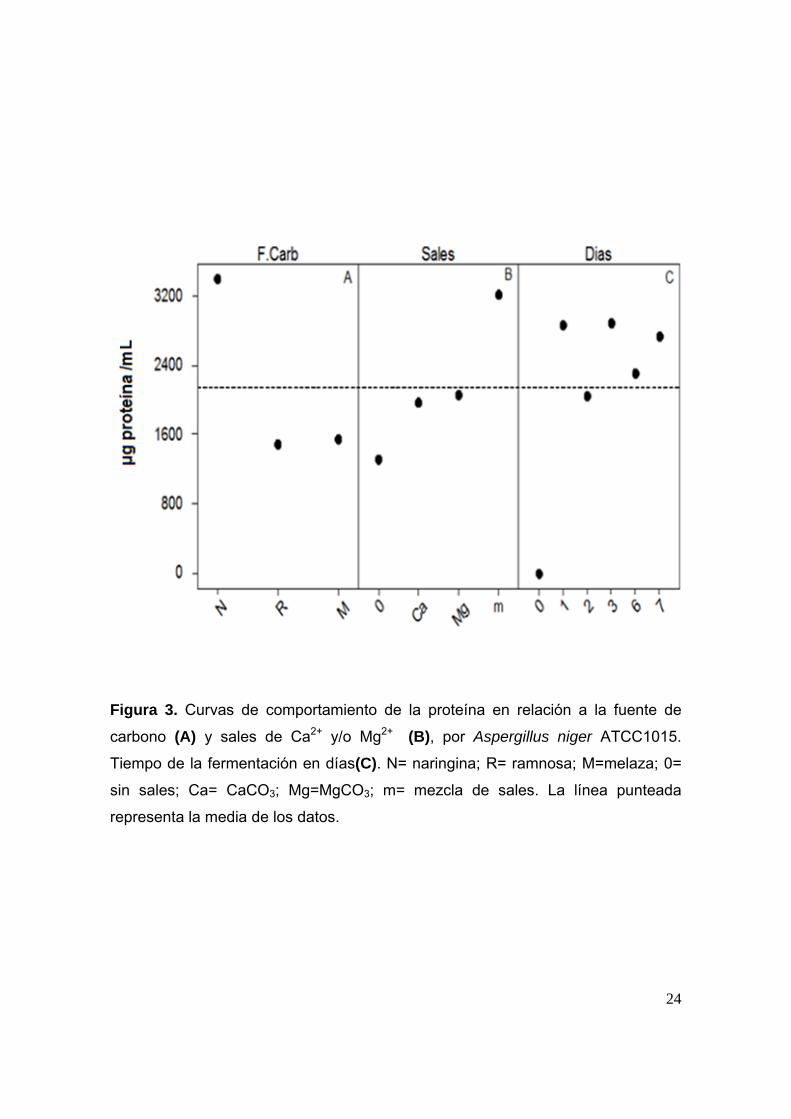
24
Figura 3. Curvas de comportamiento de la proteína en relación a la fuente de
carbono (A) y sales de Ca2+ y/o Mg2+ (B), por Aspergillus niger ATCC1015.
Tiempo de la fermentación en días(C). N= naringina; R= ramnosa; M=melaza; 0=
sin sales; Ca= CaCO3; Mg=MgCO3; m= mezcla de sales. La línea punteada
representa la media de los datos.

25
Figura 4. Curvas de producción de proteína mostrando las relaciones: fuente de
carbono-sales (A) y fuente de carbono-días (B), para naringina, melaza y ramnosa
y (C) la relación sales-días para naringina. N= naringina; R= ramnosa; M=melaza;
0= sin sales; Ca= CaCO3; Mg= MgCO3; m= mezcla de sales.
Al analizar individualmente las diferentes formulaciones en función de la fuente de
carbono utilizada se encontró para el caso de naringina que existe diferencia
significativa (p≤0.05) en ausencia y presencia de las sales de Ca2+ y/o Mg2+.
Siendo el máximo de producción de proteína (Cuadro 9) para naringina sin adición
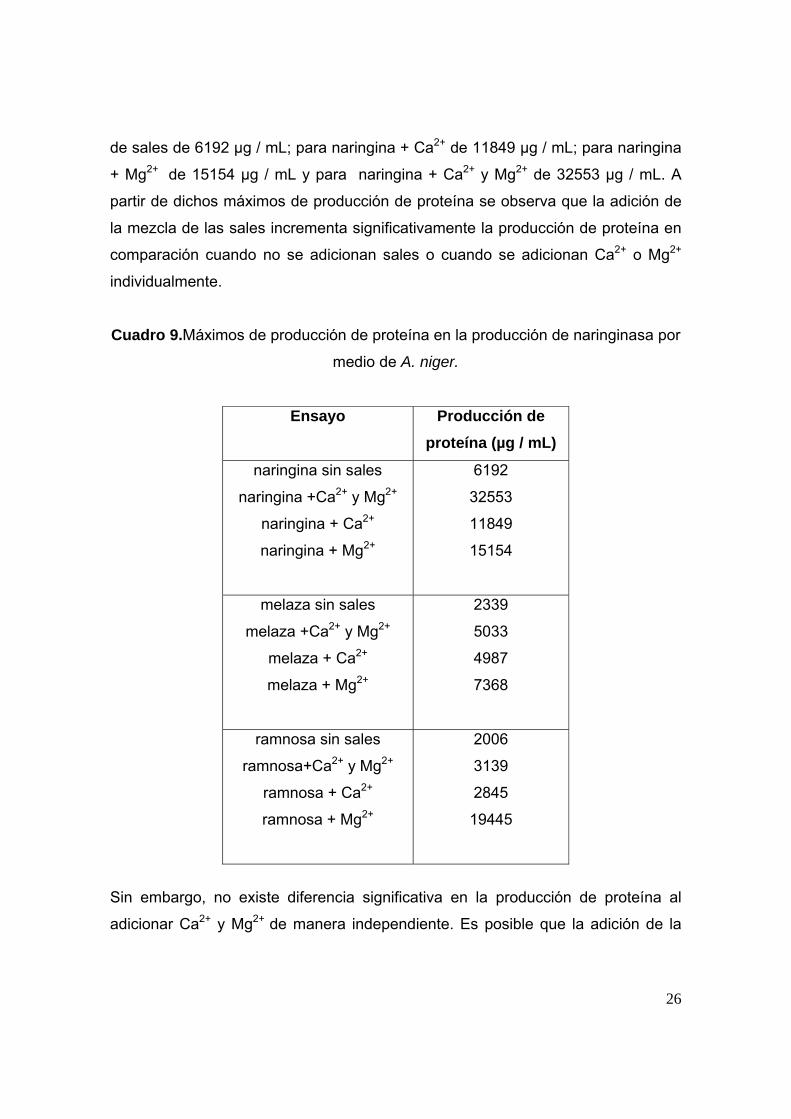
26
de sales de 6192 µg / mL; para naringina + Ca2+ de 11849 µg / mL; para naringina
+ Mg2+ de 15154 µg / mL y para naringina + Ca2+ y Mg2+ de 32553 µg / mL. A
partir de dichos máximos de producción de proteína se observa que la adición de
la mezcla de las sales incrementa significativamente la producción de proteína en
comparación cuando no se adicionan sales o cuando se adicionan Ca2+ o Mg2+
individualmente.
Cuadro 9.Máximos de producción de proteína en la producción de naringinasa por
medio de A. niger.
Sin embargo, no existe diferencia significativa en la producción de proteína al
adicionar Ca2+ y Mg2+ de manera independiente. Es posible que la adición de la
Ensayo
Producción de proteína (µg / mL)
naringina sin sales
naringina +Ca2+ y Mg2+
naringina + Ca2+
naringina + Mg2+
6192
32553
11849
15154
melaza sin sales
melaza +Ca2+ y Mg2+
melaza + Ca2+
melaza + Mg2+
2339
5033
4987
7368
ramnosa sin sales
ramnosa+Ca2+ y Mg2+
ramnosa + Ca2+
ramnosa + Mg2+
2006
3139
2845
19445

27
mezcla de las sales tenga un efecto sinergista positivo en la producción de
proteína cuando se utiliza naringina como fuente de carbono.
Cuando se utilizó melaza como fuente de carbono se encontró diferencia
significativa en la producción de proteína en ausencia y presencia de las sales de
Ca2+ y/o Mg2+. Siendo el máximo de producción de proteína (Cuadro 9) para
melaza sin adición de sales de 2339 µg /mL; para melaza + Ca2+ de 4987 µg / mL;
para melaza + Mg2+ de 7368 µg / mL y para melaza + Ca2+ y Mg2+ de 5033 µg /
mL. Es importante mencionar que, a pesar de que la producción de proteína al
adicionar Mg2+ al caldo nutritivo es mayor, esta no es significativamente diferente
en relación a la proteína producida cuando no se hizo adición de sales o cuando
se adicionó solo Ca2+, sin embargo, sí es significativamente mayor en relación a la
producida cuando se adicionó la mezcla de sales.
Finalmente cuando se utilizó ramnosa como fuente de carbono también se
encontró diferencia significativa en ausencia y presencia de las sales de Ca2+ y/o
Mg2+ en la producción de proteína (Cuadro 9). Siendo el máximo de producción de
proteína para ramnosa sin adición de sales de 2006 µg /mL; para ramnosa + Ca2+
de 3139 µg / mL; para ramnosa + Mg2+ de 2845 µg / mL y para ramnosa + Ca2+ y
Mg2+ de 19445 µg / mL. A partir de los máximos obtenidos se observa que la
adición de la mezcla de sales de Ca2+ y Mg2+ al caldo nutritivo permite una mayor
producción de proteína con respecto a cuando no se adicionaron dichas sales, es
decir, la adición de la mezcla de sales tiene un efecto positivo en la producción de
proteína. Por otra parte la adición de Ca2+ al caldo nutritivo favoreció de igual
manera la producción de proteína en relación a cuando no se adicionó, pero esta
producción no fue significativamente diferente a la obtenida cuando se adicionó
Mg2+ o cuando se adicionó la mezcla de Ca2+ y Mg2+. Cabe señalar que la adición
de Mg2+ no tuvo un efecto significativo en la producción de proteína en relación a
los otros caldos nutritivos.
Al hacer una comparación entre las tres fuentes de carbono (naringina, melaza y
ramnosa) bajo las mismas condiciones, se encontró que al no adicionar sales
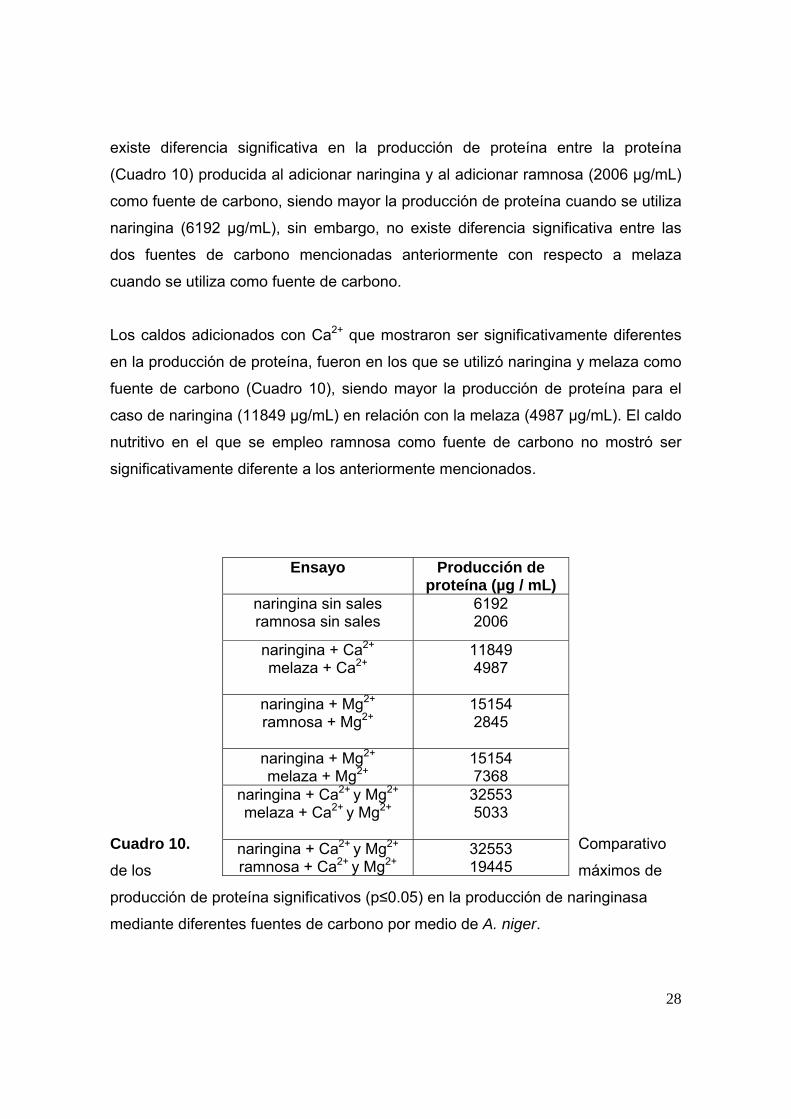
28
existe diferencia significativa en la producción de proteína entre la proteína
(Cuadro 10) producida al adicionar naringina y al adicionar ramnosa (2006 µg/mL)
como fuente de carbono, siendo mayor la producción de proteína cuando se utiliza
naringina (6192 µg/mL), sin embargo, no existe diferencia significativa entre las
dos fuentes de carbono mencionadas anteriormente con respecto a melaza
cuando se utiliza como fuente de carbono.
Los caldos adicionados con Ca2+ que mostraron ser significativamente diferentes
en la producción de proteína, fueron en los que se utilizó naringina y melaza como
fuente de carbono (Cuadro 10), siendo mayor la producción de proteína para el
caso de naringina (11849 µg/mL) en relación con la melaza (4987 µg/mL). El caldo
nutritivo en el que se empleo ramnosa como fuente de carbono no mostró ser
significativamente diferente a los anteriormente mencionados.
Cuadro 10. Comparativo
de los máximos de
producción de proteína significativos (p≤0.05) en la producción de naringinasa
mediante diferentes fuentes de carbono por medio de A. niger.
Ensayo
Producción de proteína (µg / mL)
naringina sin sales ramnosa sin sales
6192 2006
naringina + Ca2+ melaza + Ca2+
11849 4987
naringina + Mg2+ ramnosa + Mg2+
15154 2845
naringina + Mg2+ melaza + Mg2+
15154 7368
naringina + Ca2+ y Mg2+ melaza + Ca2+ y Mg2+
32553 5033
naringina + Ca2+ y Mg2+ ramnosa + Ca2+ y Mg2+
32553 19445

29
En el caso de los caldos a los que se les adiciono Mg2+ se encontró diferencia
significativa en la producción de proteína (Cuadro 10), entre el que se utilizo como
fuente de carbono la naringina (15154 µg/mL) y en el que se utilizo ramnosa (2845
µg/mL), también se encontró diferencia significativa en la producción de proteína
entre naringina y melaza (7368 µg/mL). Sin embargo, la producción de proteína
entre melaza y ramnosa no fue significativamente diferente.
En el caso de los caldos adicionados con la mezcla de las sales de Ca2+ y Mg2+,
también se encontró diferencia significativa en la producción de proteína (Cuadro
10), entre naringina (32553 µg/mL) y melaza (5033 µg/mL) utilizada como fuente
de carbono, de igual manera entre naringina y ramnosa (19445 µg/mL).
Por otra parte el tiempo (días) mostró tener efecto en la producción de proteína
dependiendo de la fuente de carbono que se empleo, lo cual se observa en la
Figura 4B donde claramente se aprecia que los ensayos realizados con naringina
como fuente de carbono presentan mayor producción de proteína ya que a pesar
de que presenta variaciones a lo largo de la fermentación, siempre se mantiene
por arriba de la producción de proteína producida con melaza y con ramnosa. Por
otra parte la Figura 4C, muestra la producción de proteína a lo largo del tiempo de
fermentación utilizando naringina como fuente de carbono en relación a la
ausencia y/o presencia de las sales de Ca2+ y Mg2+ donde se observa que es con
la mezcla donde se ve favorecida significativamente la producción de proteína a
pesar de la variación de esta a lo largo de los días debido al metabolismo del
hongo.
Estudio de la actividad enzimática de la naringinasa de A. niger en muestras de
cada fermentación.
La determinación de la actividad enzimática se realizó durante el crecimiento del
hongo en el caldo nutritivo en diferentes condiciones por triplicado a lo largo de 7

30
días de fermentación. Se utilizó como blanco una solución de naringina 5.26M pH
6 en buffer de citratos.
En la Figura 2A se observa el efecto de las tres diferentes fuentes de carbono en
la actividad enzimática. A través del análisis estadístico se encontró que existe
diferencia significativa entre ellas (p≤0.05) siendo la ramnosa la fuente de carbono
que favorece más la producción de naringinasa. Por otra parte en la figura 5B se
observa que la adición de sales no influye significativamente en la actividad
enzimática de esta enzima.
Figura 5. Curvas de Actividad enzimática expresada en microgramos de glucosa
producida después de 24hrs de hidrolisis, en relación a las diferentes fuentes de
carbono 0.5%, sales de Ca2+ y/o Mg2+ 0.01mM y días. La línea punteada
representa la media de los datos. N= naringina; R= ramnosa M=melaza; 0= sin
sales; m= mezcla de sales.
La producción enzimática obtenida en el presente trabajo puede explicarse a
través de la teoría económica de los microorganismos (Koch, 1985) que establece

31
que la producción de una enzima inducida, solo es favorecida cuando la
producción de esta permite al microorganismo obtener más nutrientes para su
crecimiento y multiplicación. Además este efecto puede incrementar si no existen
sustratos fácilmente asimilables, los microorganismos no regularan fuertemente la
producción enzimática de las enzimas constitutivas, o los costos enzimáticos son
suficientemente bajos para permitir una continua producción, y de acuerdo con
Stevens y col, (2005), las enzimas extracelulares como en este caso la
naringinasa son el principal medio por el cual los microorganismos degradan
compuestos orgánicos complejos en moléculas pequeñas que pueden ser
asimilables.
Por otra parte en la Figura 5C se observa que la producción de naringinasa varía
de acuerdo con el metabolismo del hongo durante los días que duró la
fermentación, siendo esta mayor el primer día después del inicio de la
fermentación y claramente se observa una disminución de esta los últimos días de
la fermentación, quizá debido a una represión por catabolito (glucosa) (Munish y
col 2005, Bram y Solomons, 1965).También el comportamiento ascendente y
descendente de la actividad enzimática a lo largo del tiempo de fermentación
puede explicarse a través de lo citado por Chróst (1991) que establece que una
vez que la concentración de productos incrementa lo suficiente, la síntesis
enzimática se ve reprimida y la producción regresa a un nivel basal.
Comparando la actividad enzimática que se observa a través de los días (Fig. 5C)
con la variación de la proteína presente en el medio de cultivo con respecto al
tiempo (Fig. 3C) se observa que la cantidad la actividad enzimática no es
proporcional a la producción de proteína ya que la actividad enzimática mayor se
observa en el primer día después del inicio de la fermentación (Figura 6B)
después de lo cual desciende y es hasta el sexto día cuando vuelve a subir pero
no alcanza el máximo alcanzado el primer día, lo que es contrario para la
producción de proteína donde se observa (Figura 3B) que esta aumenta y
disminuye repetitivamente, es decir no muestra la misma tendencia que en el caso

32
de la actividad enzimática . De acuerdo con Koch, 1985; Pelletier y Sygush, 1990;
Chróst, 1991; Sinsabaugh y Moorhead, 1994, debido a la producción de nitrógeno
y energía, los microorganismos solo deben producir enzimas a expensas del
crecimiento y metabolismo, es decir; cuando la disponibilidad de nutrientes es
escasa, la producción de enzimas aumenta, ya que de esta manera los
microorganismos pueden movilizar los nutrimentos de las fuentes complejas
(Harder y Dijkhuizen, 1983).
En este trabajo no se cuantificó la cantidad de enzima producida sino la actividad
enzimática de la enzima producida, en este sentido, la actividad enzimática de la
naringinasa producida no, fue proporcional a la producción de proteína.
Por otra parte también se encontró que las relaciones de Fuente de carbono-Sales
(Figura 6A), Fuente de Carbono- Días (Figura 39B), Sales-Días (Figura 6C), tienen
un efecto significativo en la actividad enzimática determinada.

33
Figura 6. Curvas de actividad enzimática mostrando las relaciones fuente de
carbono-sales, fuente de carbono-días y sales-días. N= naringina; R= ramnosa
M=melaza; 0= sin sales; m= mezcla de sales. La línea punteada representa la
media de los datos.
Al analizar los diferentes caldos nutritivos en los que se empleó naringina como
fuente de carbono se encontró que no existe diferencia significativa entre ellos en
cuanto a la actividad enzimática determinada (Cuadro 11), sin embargo, el máximo
de actividad enzimática encontrado para naringina sin sales fue de 2605 μg de
glucosa/mL; para naringina + Ca2+ fue de 5536 μg de glucosa / mL; para naringina
+ Mg2+ fue de 3122 μg de glucosa/mL y para naringina + Ca2+ y Mg2+ fue de 3122
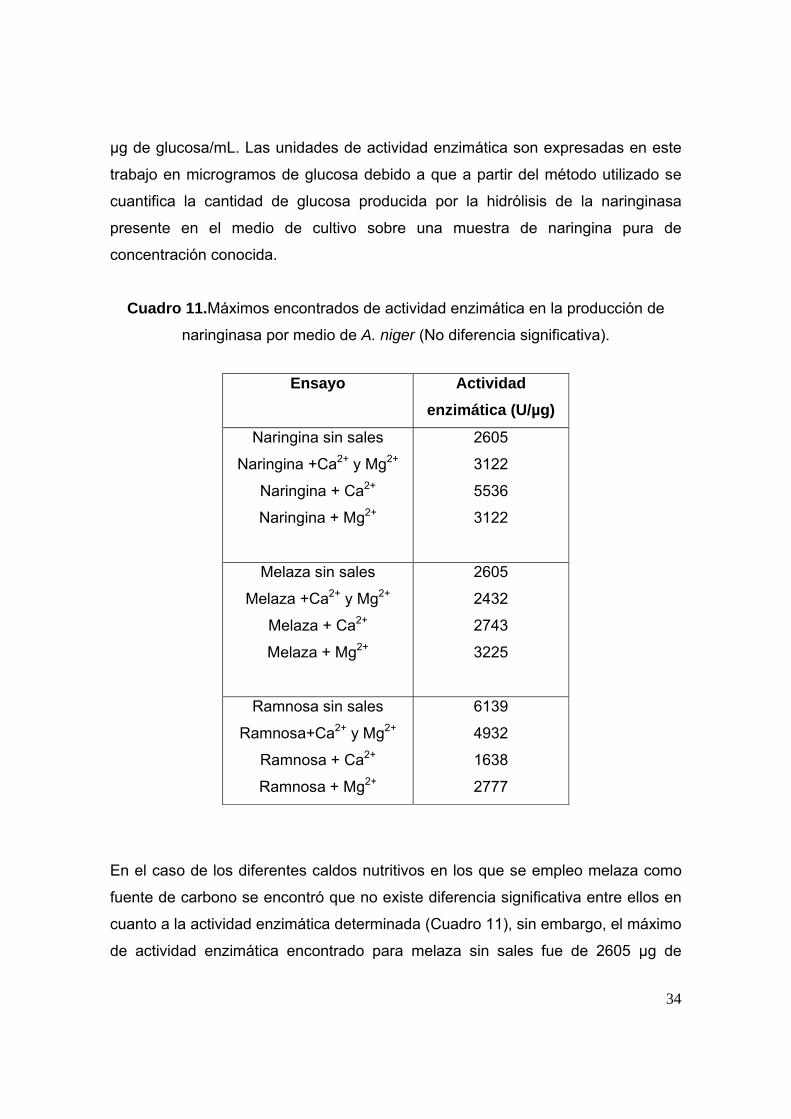
34
μg de glucosa/mL. Las unidades de actividad enzimática son expresadas en este
trabajo en microgramos de glucosa debido a que a partir del método utilizado se
cuantifica la cantidad de glucosa producida por la hidrólisis de la naringinasa
presente en el medio de cultivo sobre una muestra de naringina pura de
concentración conocida.
Cuadro 11.Máximos encontrados de actividad enzimática en la producción de
naringinasa por medio de A. niger (No diferencia significativa).
En el caso de los diferentes caldos nutritivos en los que se empleo melaza como
fuente de carbono se encontró que no existe diferencia significativa entre ellos en
cuanto a la actividad enzimática determinada (Cuadro 11), sin embargo, el máximo
de actividad enzimática encontrado para melaza sin sales fue de 2605 μg de
Ensayo
Actividad enzimática (U/µg)
Naringina sin sales
Naringina +Ca2+ y Mg2+
Naringina + Ca2+
Naringina + Mg2+
2605
3122
5536
3122
Melaza sin sales
Melaza +Ca2+ y Mg2+
Melaza + Ca2+
Melaza + Mg2+
2605
2432
2743
3225
Ramnosa sin sales
Ramnosa+Ca2+ y Mg2+
Ramnosa + Ca2+
Ramnosa + Mg2+
6139
4932
1638
2777

35
glucosa / mL; para melaza + Ca2+ fue de 2743 μg de glucosa / mL; para melaza +
Mg2+ fue de 3225 μg de glucosa / mL y para melaza + Ca2+ y Mg2+ fue de 2432 μg
de glucosa/mL.
Finalmente, para el caso de los diferentes caldos nutritivos en los que se empleo
ramnosa como fuente de carbono tampoco se encontró diferencia significativa
entre ellos en cuanto a la actividad enzimática determinada (Cuadro 11), sin
embargo, el máximo de actividad enzimática encontrado para ramnosa sin sales
fue de 6139 μg de glucosa / mL; para ramnosa + Ca2+ fue de 1638 μg de glucosa /
mL; para ramnosa + Mg2+ fue de 2777 μg de glucosa / mL y para ramnosa + Ca2+
y Mg2+ fue de 4932 μg de glucosa / mL.
Por otra parte, al hacer una comparación entre las tres fuentes de carbono
(naringina, melaza y ramnosa) bajo las mismas condiciones, se encontró que al no
adicionar sales sí existe diferencia significativa entre en la actividad enzimática
determinada en el caldo nutritivo adicionado con naringina y el adicionado con
ramnosa, encontrándose un máximo de actividad enzimática para naringina de
2605 µg de glucosa /mL y de 6139 µg de glucosa /mL para ramnosa (Cuadro 12).
Cuadro 12. Comparativo de los máximos de actividad enzimática significativos
(p≤0.05) de la naringinasa producida utilizando diferentes fuentes de carbono por
medio de A. niger.
Ensayo
Actividad enzimática (U/µg)
Naringina sin sales
Ramnosa sin sales
2605
6139
Melaza sin sales
Ramnosa sin sales
2605
6139
Naringina + Ca2+
Melaza + Ca2+
5536
2743

36
También se encontró diferencia significativa entre la actividad enzimática
determinada en el caldo nutritivo adicionado con melaza y el adicionado con
ramnosa, encontrándose un máximo de actividad enzimática para melaza de 2605
µg de glucosa /mL, lo que claramente significa que el caldo de cultivo en el que se
emplea ramnosa como fuente de carbono permite obtener una mayor actividad
enzimática comparada con la de las otras dos fuentes de carbono (Cuadro 12).
Cabe señalar que no se encontró diferencia significativa entre la actividad
enzimática de la naringinasa determinada en el caldo nutritivo adicionado con
Naringina y el adicionado con melaza.
Los caldos adicionados con Ca2+ que mostraron ser significativamente diferentes
en la actividad enzimática fueron en los que se utilizó naringina y melaza como
fuente de carbono (Cuadro 12), siendo mayor la producción de proteína para el
caso de Naringina (5536 µg de glucosa /mL) en relación con la melaza para la que
se encontró una actividad enzimática de 2743 µg de glucosa /mL. El caldo de
nutritivo en el que se empleo Ramnosa como fuente de carbono no mostró ser
significativamente diferente a los anteriormente mencionados.
En el caso de los caldos a los que se les adiciono Mg2+ no se encontró diferencia
significativa en la actividad enzimática, entre el alguna de las fuentes de carbono
empleadas en este trabajo. En los caldos adicionados con la mezcla de las sales
de Ca2+ y Mg2+, tampoco se encontró diferencia significativa en la actividad
enzimática determinada entre los caldos de cultivo adicionados con las diferentes
fuentes de carbono.
De acuerdo a los resultados anteriores se seleccionó el caldo nutritivo que
presentó mayor actividad enzimática. De tal manera que la elección se realizó ente
la actividad enzimática para los caldos nutritivos en los que no se adicionó sales y
cuando se adicionó Ca2+, para las fermentaciones llevadas a cabo sin adición de
sales, el caldo nutritivo adicionado con ramnosa fue quién mostro mayor actividad
enzimática (6139 µg de glucosa /mL) y para las llevadas a cabo con adición de

37
Ca2+ la actividad enzimática mayor encontrada fue cuando se utilizó Naringina
(5536 µg de glucosa /mL) como fuente de carbono. Por lo que la fracción de
ramnosa sin sales fue la elegida para realizar la siguiente parte del experimento.
Estudio de la actividad enzimática del caldo nutritivo seleccionado.
La fracción seleccionada para la concentración de la proteína y la determinación
de las constantes fue la de la ramnosa sin adición de sales del primer día después
del inicio de la fermentación. El uso de la ramnosa como fuente de carbono
permite obtener una naringinasa con mayor actividad comparada con la producida
utilizando como fuente de carbono naringina o melaza. Quizá este
comportamiento pude explicarse por lo sugerido por Munish y colaboradores en el
2005, quienes señalan que los medios utilizados para producir naringinasa por A.
niger, adicionados con ramnosa, producen elevadas cantidades de enzima quizá
ya que estos medios tienen un bajo contenido de carbohidratos. Sin embargo,
Elinbaum y col. 2002 sugieren que en el caso de una fermentación sólida la
naringina es mejor inductor que la ramnosa para la producción de la naringinasa
utilizando A. terreus, además también sugieren que una fermentación solida
provee mejores resultados para la producción de naringinasa que una
fermentación liquida.
Haciendo una comparación de la producción proteica a lo largo del tiempo de
fermentación en relación a la actividad enzimática se observa que no son
proporcionales y por tanto una mayor producción proteica no conlleva a una mayor
actividad enzimática aunque la naturaleza de la enzima sea proteica, ya que una
actividad enzimática elevada puede estar dada tanto por haber alto contenido de
proteína en el medio de cultivo o por que la enzima es altamente activa. También
es importante resaltar que la actividad enzimática puede ser reprimida debido a
una inhibición por glucosa (producto de la hidrólisis de la naringina por la
naringinasa) ya que de acuerdo con lo reportado por Munish y colaboradores en el

38
2005; Gallego y col en el 2001; Bram y Solomons en 1965 y Clarke y Brammer en
1964, la glucosa reprime la producción de naringinasa.
Como se mencionó con anterioridad, el caldo nutritivo donde se encontró mayor
actividad enzimática fue el adicionado con ramnosa como fuentes de carbono y sin
adición de sales durante las primeras 24 horas de la fermentación. Sin embargo,
debido a que se contaba con una pequeña cantidad de muestra se decidió juntar
todas las muestras tomadas cada día durante el periodo de la fermentación y se
procedió a una concentración de proteína. El contenido de proteína determinado
después del proceso de concentración al que se sometió la muestra fue de 66851
μg/mL de proteína.
La determinación de la actividad enzimática de las fracciones de ramnosa (del
conjunto de muestras obtenidas cada día) sin adición de sales se probó sobre la
hidrólisis de naringina y se obtuvo una actividad enzimática de 0.010U/mg
(U=1µmol de glucosa liberada / min) la cual se reporta en el Cuadro 13. Tal vez
este valor tan pequeño encontrado se debe a que al juntar todas las fracciones
disminuyó la actividad enzimática inicial la cual fue de 0.947 U / min.
La actividad encontrada para la naringinasa presente en el concentrado de
proteína es mayor comparada con la informada por Bram y Solomons 1965 para la
naringinasa de A. niger NRRL 72-4 (Cuadro 13, No 5) y es menor comparada con
la informada por diferentes autores, los valores se presentes en el Cuadro 13, sin
embargo, es importante resaltar que la mayoría de ellos determinaron la actividad
enzimática utilizando sustratos inespecíficos como el p-nitrofenol o el dietilen glicol
alcalino, que esta actividad fue determinada utilizando diferentes condicione, y que
la producción de la enzima se realizó utilizando diferentes condiciones de
fermentación como en el caso de Gülten y col, 2006 (Cuadro13, No 2).
Por otra parte tal vez la producción de naringinasa por el hongo en estudio podría
mejorarse si se hace una adición de la fuente de carbono en diferentes etapas de

39
la fermentación de acuerdo con lo que sugiere Bram y Solomons (1965), sin
embargo, como ellos mismos establecen la adición continua de la fuente de
carbono a lo largo de la fermentación es un método más difícil y que requiere de
mayor control y de un fermentador.
Cuadro 13. Actividad enzimática de la naringinasa producida por diferentes
microorganismos y determinada mediantes diferentes sustratos.
Sustrato Actividad
narignina (1) 0.010 U/mg
naringina (2) 469 U/mg
p-nitrofenol (3) 91 U/mg
p-nitrofenol (4) 1.7 U/mL
dietilen glicol alcalino (5) 90-94 U/mL
dietilen glicol alcalino (6) 3.99x 10-4 IU/mL
p-nitrofenol (7) 0.059 U
(1) Naringinasa de A. niger ATCC1015 producida en el presente trabajo. Medio de
cultivo adicionado con ramnosa como fuente de carbono. Regulador de citratos
0.05M, pH=6, 35°C, [naringina]=0.00526mM. U=1µmol de glucosa liberada / min.
(2) Naringinasa de P. decumbens, Gülten y col, 2006. Fermentación solida.
Regulador de acetato de amonio 0.1M, pH= 4, 40°C, U= 1 µmol de glucosa
liberado/ min.
(3) α-L-ramnosidasa de A. terreus, Gallego y col, 2001. Medio de cultivo
adicionado con ramnosa. Rregulador de succinato 50 mM, pH= 5.5, 50°C. U=
cantidad de enzima necesaria para liberar 1μmol p-nitrofenol/min.
(4) α-L-ramnosidasa de A. terreus, Elinbaum y col, 2002. Medio de cultivo
adicionado con carbono de bagazo de caña de azúcar. Regulador de ácido.
40
succínico /succinato de sodio 50mM, pH=5.5, 30°C. U=cantidad de enzima
necesaria para liberar 1μmol p-nitrofenol/min.
(5) Naringinasas de A. niger NRRL 72-4, Bram y Solomons 1965. Regulador de
citratos sin molaridad especifica reportada, pH=4, 45°C. U=cantidad de enzima
necesaria para hidrolizar 1μmol naringina/min. Método de Davis 1947.
(6) Naringinasa de A. niger MTCC 1344, Munish y col en el 2005. Medio de cultivo
adicionado con ramnosa. Regulador de acetato de sodio 0.1M, pH= 4, 30°C.
IU=cantidad de enzima necesaria para liberar 1μmol naringina/min. Método de
Davis 1947.
(7) α-ramnosidasa de la naringinasa de A. niger. Manzanares y col, 1997. Medio
de cultivo adicionado con naringina. Regulador MacIIvaine, pH=4.5, 30°C.
U= μmol de ramnosa/min.
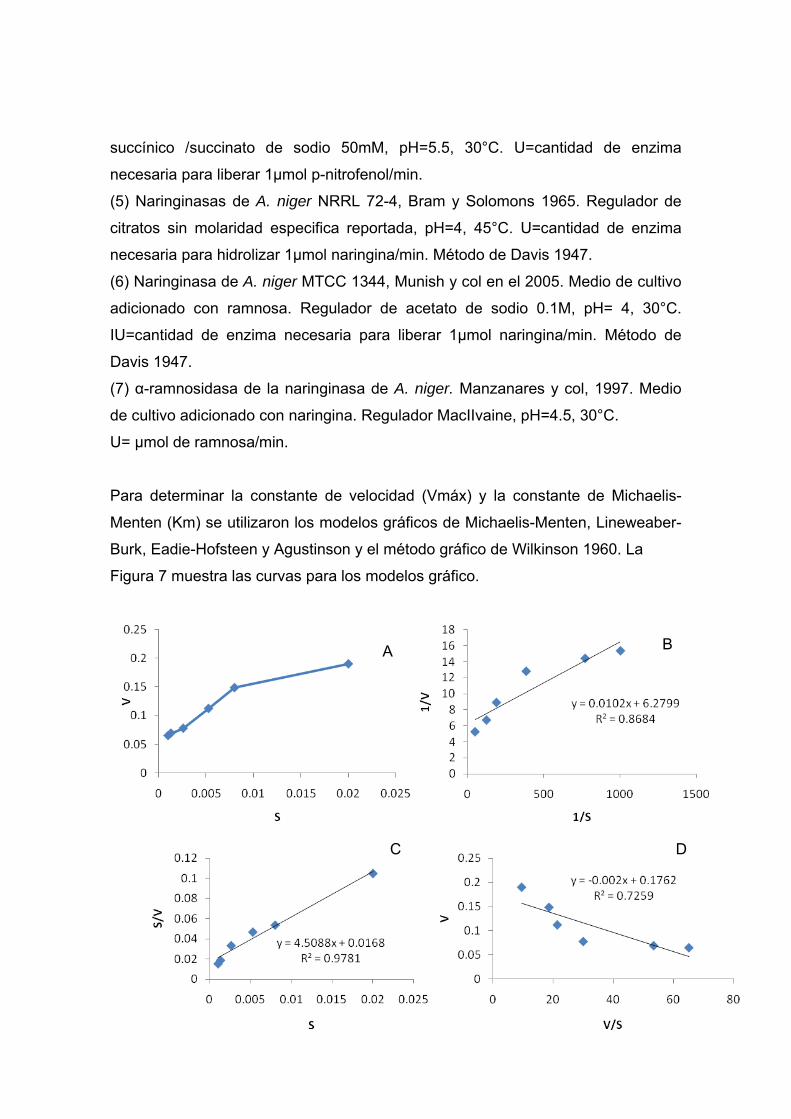
Para determinar la constante de velocidad (Vmáx) y la constante de Michaelis-
Menten (Km) se utilizaron los modelos gráficos de Michaelis-Menten, Lineweaber-
Burk, Eadie-Hofsteen y Agustinson y el método gráfico de Wilkinson 1960. La
Figura 7 muestra las curvas para los modelos gráfico.
A B
C D

41
Figura 7. (A)Curva de Michaelis –Menten para Naringinasa donde Y=
6.683x+0.068; R2=0.908; (B) Curva de Lineweaber-Burk para Naringinasa; (C) Curva de Agustinson para Naringinasa; (D) Curva de Eadie-Hofsteen para la
naringinasa de A. niger ATCC1015.
Los valores correspondientes a Vmáx y Km para la naringinasa de A. niger a partir
de cada modelo gráfico y el calculado por el método estadístico de Wilkinson se
muestran en el cuadro 15.
Cuadro 15. Parámetros de Vmax y Km para las curvas de Michaelis-Menten,
Lineweaber-Burk, Eadi-Hoffsten, Agustinson y Wilkinson para la naringinasa de A.
niger ATCC1015.
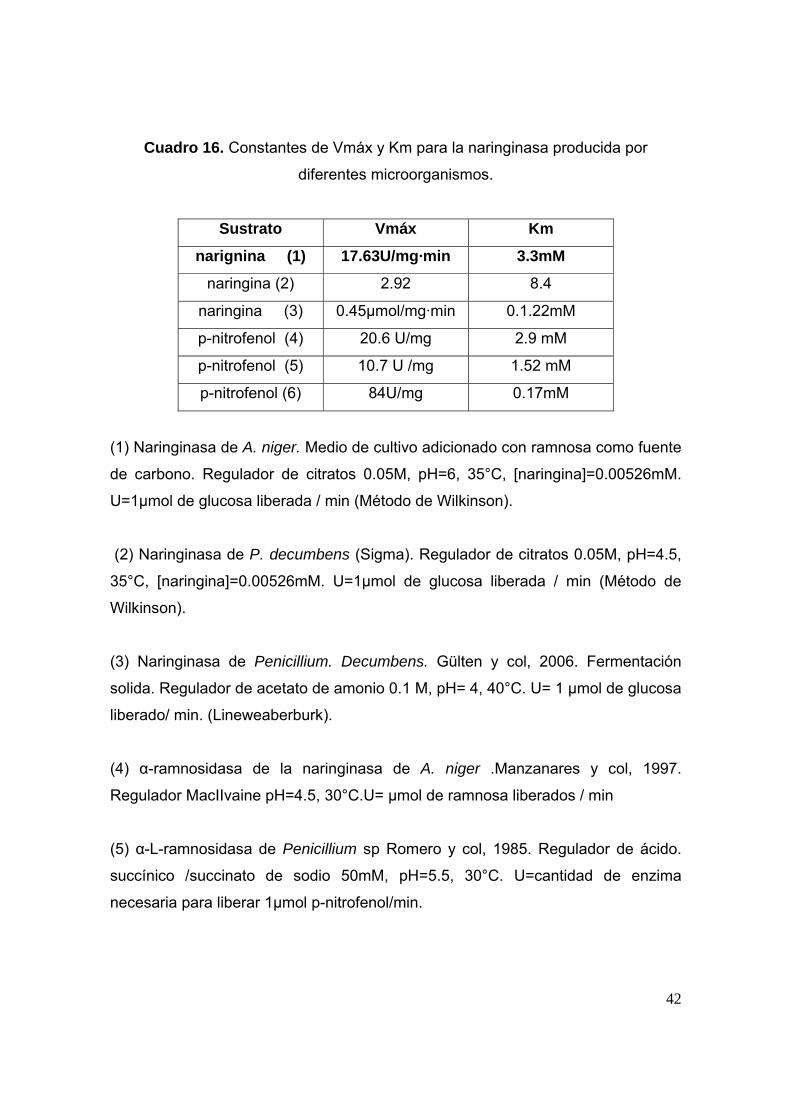
En el Cuadro 16 se presentan los valores para la Vmáx y Km determinadas para
A. niger y P. decumbens, así como los valores reportados por diferentes autores
para la naringinasa.
Curva Vmax ( U/mg min)
Km (mM)
Michaelis-Menten 0.0158 0.0954
Lineweaverburk 13.24 1.6
Eadie-Hoffsten 18.46 7.21
Agustinson 14.64 2.1
Wilkinson* 17.63 3.3
42
Cuadro 16. Constantes de Vmáx y Km para la naringinasa producida por
diferentes microorganismos.
Sustrato Vmáx Km
narignina (1) 17.63U/mg·min 3.3mM
naringina (2) 2.92 8.4
naringina (3) 0.45µmol/mg·min 0.1.22mM
p-nitrofenol (4) 20.6 U/mg 2.9 mM
p-nitrofenol (5) 10.7 U /mg 1.52 mM
p-nitrofenol (6) 84U/mg 0.17mM
(1) Naringinasa de A. niger. Medio de cultivo adicionado con ramnosa como fuente
de carbono. Regulador de citratos 0.05M, pH=6, 35°C, [naringina]=0.00526mM.
U=1µmol de glucosa liberada / min (Método de Wilkinson).
(2) Naringinasa de P. decumbens (Sigma). Regulador de citratos 0.05M, pH=4.5,
35°C, [naringina]=0.00526mM. U=1µmol de glucosa liberada / min (Método de
Wilkinson).
(3) Naringinasa de Penicillium. Decumbens. Gülten y col, 2006. Fermentación
solida. Regulador de acetato de amonio 0.1 M, pH= 4, 40°C. U= 1 µmol de glucosa
liberado/ min. (Lineweaberburk).
(4) α-ramnosidasa de la naringinasa de A. niger .Manzanares y col, 1997.
Regulador MacIIvaine pH=4.5, 30°C.U= μmol de ramnosa liberados / min
(5) α-L-ramnosidasa de Penicillium sp Romero y col, 1985. Regulador de ácido.
succínico /succinato de sodio 50mM, pH=5.5, 30°C. U=cantidad de enzima
necesaria para liberar 1μmol p-nitrofenol/min.

43
(6) α-L-ramnosidasa de A. terreus, Gallego y col, 2001. Medio de cultivo
adicionado con ramnosa. Rregulador de succinato 50 mM, pH= 5.5, 50°C. U=
cantidad de enzima necesaria para liberar 1μmol p-nitrofenol/min. (Michaelis)
En este trabajo se encontró que los valores de Vmáx y Km para la naringinasa de
A. niger ATCC1015 (Cuadro16, No.1) coinciden con los encontrados para la
naringinasa de P. decumbens (Cuadro16, No.2), lo que significa que a pesar de
haber una aparente producción proteica baja para la naringinasa producida por A.
niger, las constantes resultaron ser del mismo orden que las encontradas para la
naringinaa de P. decmbens por lo cual los resultados obtenidos son
comparativamente buenos.
El valor de Vmáx reportados por Gallego y colaboradores en el 2001 (Cuadro 16,
No 6) para la α-L-ramnosidasa de A. terreus es menor comparada con el obtenido
en el presente trabajo (Cuadro 16, No. 1) sin embargo, la enzima obtenida
presenta una menor afinidad por el sustrato naringina ya que obtuvo una Km
mayor (Cuadro 16, No.1) quizá porque esta enzima no solo presenta la actividad
de α-L-ramnosidasa sino también la de β-glucosidasa.
Por otra parte el valor informado para la α-L-ramnosidasa de A. niger por
Manzanares y colaboradores en 1997 y el informado por Romero y colaboradores
en 1985 es menores que el encontrado en el presente trabajo mediante el método
de Wilkinson ya que este engloba de manera estadística a todos los demás.
El valor de Vmáx para la α-L-ramnosidasa de A. niger informado por Manzanares y
col. 1997 (Cuadro 16, No.4) es mayor a lo encontrado en el presente trabajo lo
que significa que la actividad enzimática de la enzima encontrada es menor
teniendo ambas actividades (α-L-ramnosidasa y β-D-glucosidasa).
Gülten y col. en el 2006 determinaron las constantes de actividad enzimática de la
naringinasa de P. decumbens a través del método de Linewaver-Burk,

44
encontrando una Km menor a la encontrada, lo que significa que dicha naringinasa
tiene más afinidad por el sustrato que la que tiene la naringinasa producida
(Cuadro 64, No.3).
Sin embargo todos los autores anteriormente mencionados determinaron la las
constantes de Vmáx y Km utilizando un sustrtos inespecíficos como el p-nitrofenol,
obteniendo los valores presentes en el Cuadro 16.
CONCLUSIONES
El presente trabajo nos permitió encontrar una cepa capaz de producir una enzima
con las características encontrada en una enzima comercial, por lo que usando
fuentes de carbono económicas se puede encontrar un nicho que a nivel comercial
puede ser más rentable.
La prueba presuntiva en cromatografía en capa fina mostró presuntivamente que
la cepa de Aspergillus niger ATCC 1015 es capaza de producir naringinasa.
Las diferentes fuentes de carbono y la concentración en la que se emplearon
produjeron naringinasa a partir de A. niger ATCC 1015 como respuesta por parte
del microorganismo a la presencia de compuestos orgánicos que pueden ser
asimilables a través de estas enzimas.
Cuando se adicionaron las sales de calcio y magnesio se encontró que un factor
determinante para el crecimiento del microorganismo es la concentración en la
que éstas se adicionan.
La fuente de carbono que favorece más el crecimiento del microorganismo es la
melaza y este crecimiento incrementa cuando se adiciona carbonato de calcio.
La mayor producción proteica se obtuvo cuando se empleó naringina como fuente
de carbono y ésta fue favorecida positivamente al adicionar la mezcla de sales.

45
La mayor actividad enzimática encontrada fue la de la enzima producida utilizando
el caldo nutritivo adicionado con ramnosa como fuente de carbono sin adición de
sales.
Por lo tanto las condiciones que favorecieron el crecimiento del hongo y la
producción de la proteína no fueron las ideales para favorecer la mayor actividad
enzimática de la naringinasa producida. De acuerdo a lo encontrado las
condiciones más adversas utilizadas en este estudio para el crecimiento del hongo
favorecieron la producción de enzima con mayor capacidad de hidrólisis de la
naringina. Lo que significa que no existe relación alguna entre el crecimiento del
hongo y la actividad enzimática de la enzima producida. A demás tampoco existe
relación alguna entre la producción proteica con respecto una mayor actividad
enzimática ni entre el crecimiento del microorganismo con respecto a la
producción proteica. Sin embargo en este trabajo si existió una relación entre la
actividad enzimática y la adquisición de nutrientes por parte del microorganismo.
Debido a que la naturaleza de A. niger es producir acido cítrico, la tendencia
natural del medio de cultivo tiende a ser acida, quizás controlando estas
condiciones puede mejorarse la producción enzimática y la actividad de esta y
esto puede lograrse haciendo un escalamiento de dicha producción a
fermentadores controlados ya que con una aeración controlada y vigorosa puede
inhibirse la producción de acido cítrico por este hongo y favorezca la producción
de naringina. Además el proceso podría ser optimizado ya que existen
investigaciones que suguieren que la adición de fuentes de carbono en diferentes
etapas podría proveer mejores resultados en cuanto a la actividad enzimática de la
naringinasa que se produce utilizando A. niger.

46
BIBLIOGRAFÍA
1) Abarca Ma. L. 2000. Taxonomía e identificación de especies implicadas en
la aspergillosis nosocomial. Rev Iberoam Micol., 17:S79-S84.
2) Asmar F., F. Eiland and N. E. Nielson. 1994. Effect of extracellular enzyme
activities on solubilization rate of soil organic nitrogen. Biology and Fertility of Soils., 17:32-38.
3) Atkinson M. R., J.F. Jackson and R. K. Morton. 1961.Nicotinamide
Mononucleotide Adenylyltransferase of Pig-Liver Nuclei. Biochem. J. 80:318-323.
4) Bradford. 1976. A rapid and sensitive method for the quantization o protein
utilizing the principle of protein-dye binding. Anal. Biochem., 72:248-254.
5) Bram B. and G.L. Solomons. 1965. Production of the enzyme naringinasa
by Aspergillus niger. Appl Microbiol., 13:842-845.
6) Bourbouze R., F. Perchero and J.E. Courtois. 1976. α-L-rhamnosidasa de
Fagopyrum esculentum. Purification et vuelques propiétés. Eur. J. Biochem., 63:331-337.
7) Bucio Villalobos C.M., H. A. Luna Olvera, O.A. Martínez Jaime y D. G.
Guzmán de Peña. Efecto del extracto de estigmas de maíz sobre
Aspergillus spp. Acta universitaria, enero-abril, vol. 17, número 001.
Universidad de Guanajuato. Guanajuato, México. 2001. pp 59-62.
8) Caldini C., F. Bonomi, P.G. Pifferi, G. Lanzarini and Galante YM. 1994.
Kinetic and immobilization studies on fungal glycosidases for aroma
enhancement in wine. Enzym Microb Technol., 16:286-291.

47
9) Chandler B.V. and K.J. Nicol. 1975. Some relationships of naringina: their
importance in orange juice bitterness. CCCSIRO Food Res Quart., 35:79-88.
10) Chaplin M.F. and J.F. Kennedy. Carbohydrate Analysis: A Practical
Approach. IRL Press. Oxford, England. 1986. pp 1-36.
11) Chisti Y. Solid substrate fermentation, enzyme production, food enrichment
In: Flickinger MC, Drew SW, editors. Encyclopedia of bioprocess
Technology: Fermentation, Biocatalysis, and Bioseparation, vol. 5. New York: Wiley. 1999. pp 2446-62.
12) Chróst, R.J. Environmental control of the synthesis and activity of aquatic
microbial ectoenzymes, In: Chróst, R.J, (Ed), Microbial Enzymes in Aquatic
Environments. Springer-Verlag. New York. 1991.pp 29-59.
13) Clarke, P. M. and W. J. Brammer. 1964. Regulation of bacterial enzyme
synthesis by induction and repression. Nature., 203:1153-1155.
14) Coghlan A. 1997. Squeezing bitterness out of grapefruit. Nature Biotechnol., 15:202.
15) Currier N. J. 1917. The citric acid fermentation of Aspergillus niger. J. Biol. Chem., 31:15.
16) Daniela L., Rj. Linhardt, B.A. Brryan, F. Mayerl and M. Pickenhagen. 1990.
Methods for producing rhamnosa. US Patent 4,933, 281.
17) Davis WB. 1947. Determination of Flavonones in citrus fruits. Anal Chem., 19:476.

48
18) Dawson M.C. Data for Biochemical Research. 2nd Edition. Oxford. 1968. p 616.
19) Díaz R., C. Gamazo e I. López-Goñi. Manual práctico de Microbiología. 1ª
Reimpresión. MASSON, S. A. Ed. España.1998.
20) Dixon M. and E.C. Webb. Enzymes. Green and Co. Ltd. London: Longmans. 1958.
21) Dunlap W.J., R. E. Hagen and S.H. Winder. 1962. Preparation and
properties of rhamnosidase and glucosidase fractions from fungal flavoniod
glycosidase fractions from a fungal flavonoid glycosidase preparation,
“Naringinase C-100”. J. Food Sci., 27:597-601.
22) Elinbaum S., H. Ferreyra., G. Ellenrieder and C. Cuevas. 2002. Production
of Aspergillus terreus α-L-rhamnosidase by solid state fermentation. Letters in Applied Microbiology., 34:67-71.
23) Ellenrieder G., S. Blanco and M. Daz. 1998. Hydrolysis of supersaturated
naringin solutions by free and immobilized naringinase. Biotechnology Techniques., 12:63-65.
24) Fernandez M. Alternativas para los residuos alimentarios. Consuma
seguridad (el diario de la seguridad alimentaria). Fundación Eroski. 2004.
www.consumaseguro.com
25) Fukumoto J. and S. Okada. Naringinase production by fermenmtation.
Japanese Patent 7, 306, 554, 1973.
26) Gallego M.V., F. Piñaga., D. Ramón and S. Valés. 1996. Production and
characterization of an Aspergillus terreus α-L-rhamnosidase of enological
interes. Z. Lebensm. Unters. Forsch., 203:522-527.

49
27) Gallego M.V., F. Piñaga., D. Ramón and S. Vallés. 2001. Purification and
characterization of an α-L-rhamnosidasa from Aspergillus terreus of interest
in winemaking. Food Chemistry and Toxicology., 66:2:204-209.
28) Grassin C and P. Fauquembergue. 1996. Fruit juices. In: Godfrey T, West
S, editors. Industrial Enzymology. New York: Macmillan Press. 2:225-64.
29) Griffith F.P. Process for reactivating polyamides resin used in debittering
citrus juice. US Patent 3,463,763.1969.
30) Gunata YZ., C.L. Bayonove., R.L. Baumes and R.E. Cordonnier. 1985. The
aroma of grapes. 1. Extraction and determination of free and glycosidicaly
bound fractions of some grape aroma components. J. Chromatography., 331:83-90.
31) Gülten Sekeroglu., Sibel Fadiloglu., Fahrettin Gögüs. 2006. Immobilization
and characterization of naringinasa for the hydrolysis of naringina. Eur Food Res Technol., 224:55-60.
32) Habelt K., and F. Pittner. 1983. A rapid method for the determination of
naringina, pruning, and naringina applied to the assay of naringinase. Anal Biochem., 134:393-7.
33) Habelt K and F. Pittner. 1983. A rapid method for the determination of
naringin, prunin and naringenin applied to the assay of naringinase. Anal Biochem., 134:393-397.
34) Hall D.H. 1938. A new enzyme of the glycosidase type. Chem Ind., 57:473.
35) Harder W and L. Dijkhuizen. 1983. Physiological responses to nutrient
limitation. Annual Review of Microbiology., 37:1-23.

50
36) Hofstee B. H. J. 1952. On the Evaluation of the Constants Vmáx and Km in
Enzyme Reactions. Science., 116:329-331.
37) Horuichi S., H. Yamamoto., T. Asaloshi and T. Tanaka. 1985. High
pressure liquid chromatographic determination of naringinasa activity. J. Jpn Soc Food Sci Technol., 32(8):582-5.
38) Ito T. and Y. Takiguchi. Naringinase production by Cochibolus miyabeanus.
Japanese Patent 7,014,875.1970.
39) Jonson R.L. and B.V. Chandler. 1988. Adsorptive removal of bitter
principles and titrable acid from citrus juice. Food Technol., 45:130-7.
40) Kaji A. and T. Ichimi. 1973. A L- rhamnosidase activity in culture filtrate of
Corticium rolfsii. Enzymic activity at low pH. Agr Biol Chem., 37:431-432.
41) Kimball D.A. 1987. Debittering of citrus juices using supercritical carbon
dioxide. J Food Sci., 52:481-2.
42) Kimball D.A. Citrus Processing: Quality Control Technology. New York: Van Nostrand Reinhold, 1991.
43) Kirk R.S., R. Sawyer y H. Egan. Composición y análisis de los alimentos de
pearson. CECSA. México 2004.
44) Kishi K. 1955. Production of naringinasa from Aspergillus niger. Kagaku to kogyo. Chemistry and Industry, Japan., 29:140.
45) Klonowska A., C. Gaudin., A.M. Fournel., M. Asson., J. le Petit., M. Giorgi
and T. Tron. 2002. Characterization of a low redox potential laccase from

51
the basidiomycete C30. European Journal of Biochemistry., 269:6119-6125.
46) Koch A. L. 1985. The macroeconomics of bacterial growth, in: Fletchjer M
and G.D. Floodgate (Eds). Bacteria in their Natural Environments.
Academic Press, London. pp 1-42.
47) Koroljova Skorobogatko O.V., E.V. Stepanova., V.P. Gavrilova., O.V.
Morozova., N.V. Lubimova., A.N. Dzchafarova., A.I. Jaropolov and A.
Makower. 1998. Purification and characterization of the constitutive form of
laccase from the basidiomycete Coriolus hirsutus and effect of inducers on
laccase synthesis. Biotechnology and Applied Biochemistry., 28:47-54.
48) Kurosawa Y., K. Ikeda and F. Egami.1973. α-L-rhamnosidasa from the liver
of Turbo cornutus and Aspergillus niger. J Biochem., 73:31-37.
49) Lineweaver, H and D. Burk, 1934. The determination of enzyme
dissociation constants. J. Amer. Chem. Soc., 56:658-666.
50) Lehninger Albert. Bioquímica. Las bases moleculares de la estructura y
función celular. Ediciones Omega, S. A. Barcelona. 1990. pp 990-991.
51) Madigan M.T., J.M. Martinko and J. Parker. BROCK Biología de los
microorganismos. Pearson Educación, Madrid. 2004. pp 207,215.
52) Magindag .Amylase, polygalacturonase and naringinase co-inmobilization.
German Patent EP. 298:954.1989.
53) Manzanares P., L. de Graaff and J. Viseer. 1997. Purification and
characterization of an α-L-rhamnosidasa from Aspergillus niger. FEMS Microbiol Lett., 157:279-283.

52
54) Michon F. 1987. Structure of the complex group-specific polysaccharide of
group B Streptococcus. Biochemistry., 26:476-478.
55) Montes Mª del C., y I. Magaña. 2002. Enzimas con aplicación industrial.
Avances y perspectivas., 21:279-282.
56) Munish P., S.S. Marwaha and R.M. Kothari. 1996. Biochemical basis of
bitterness in citrus fruit juices and biotech approaches for debittering. Crit
Rev Biotechnol., 16:145-55.
57) Munish P. and C. Uttam. 2000. Production, purification, and
characterization of the debittering enzyme naringinase. Biotechnology Advances., 18:207-217.
58) Munish P., B. Anirban and U.C. Banerjee. 2005 Optimization of process
parameters for the production of naringinasa by Aspergillus niger MTCC
1344. Process biochemistry., 40:195-201.
59) Mutter M., G. Beidman., H.A. Schols and A.G.J. Voragen. 1994.
Rhamnogalacturonan α-L-Rahamnopyranohydrolase. A novel enzyme
specific for the terminal nonreducing rhamnosyl unit in rhamnogalacturonan
regions of pectin. Plant Physiol., 106:241-250.
60) Olson A.C., M.G. Gray., G. Dante. 1979. Naringina bitterness of grapefruit
juice debittered with naringinase immobilized in a hollow fiber. Food Sci., 44:1358-1361.
61) Olson A.C., M.G. Gray. 1981. Hydrolysis of high levels of naringin in
grapefruit juice using a hollow fiber naringinasa reactor. J. Agr. Food Chem., 29:1301-1304.

53
62) Ono M., T. Tosa and I. Chibata. 1978. Preparation and properties of
immobilized naringinase using tannin-aminohexyl cellulose. Agric. Biol. Chem., 42:1847-1853.
63) Pelletier A. and J. Sygush. 1990. Purification and characterization of three
chitosanase activities from Bacillus megaterium P1. Applied and Environmental Microbiology., 56:844-848.
64) Pritcher D.E. Extraction of the bitter principle from navel orange juice. US Patent 2,816,033.1957.
65) Puri A. Preparation and properties of citrus juices, concentrates and dried
powders which are reduced in bitterness. US Patent 4,439,458. 1984.
66) Roitner M., T. Schalkhammer and F. Pittner. 1984a Preparation of pruning
with the help of immobilized naringinase pretreated with alkaline buffer.
Applied Biochemistry and Biotechnology., 9:483-488.
67) Roitner M., T. Schalkhammer and F. Pittner. 1984b. Characterization of
naringinasa from aspergillus niger. Monatsh Chemme., 114:1255-67.
68) Romero C., A. Manjón., J. Bastida and J.L. Iborra. 1985. A method for
assaying the rahmnosidase activity of naringinase. Anal biochem., 149:566-571.
69) Sankyo. Preparation of antibiotic chloropolysporin-C. Japanese Patent 63, 146, 797, 1988.

54
70) Shackle V. J., C. Freeman and B. Reinolds. 2000. Carbon supply and the
regulation of enzyme activity in constructed wetlands. Soil Biology & Biochemistry., 32:1935-1940.
71) Shanmugan V. and K.D.S. Yadav. 1995. Extracellular production of alpha-
rhamnosidase by Rhizopus nigricans. Ind I Exp Biol., 33:705-7.
72) Shaw P.E. and C.W Wilson. 1983. Debitterin citrus juices with β-
cyclodextrin polymer. J Food Sci., 48:646-7.
73) Sinsabaugh R. L. and D. L Moorhead. 1994. Resource allocation to
extracellular enzyme production: a model for nitrogen and phosphorus
control of litter decomposition. Soil Biology & Biochemistry., 26:1305-1311.
74) Steven D. A. and P. M. Vitousek. 2005. Response of extracellular enzymes
o simple and complex nutrient inputs. Soil Biology & Biochemistry., 37:937-944.
75) Tereda Y., T. Kometani., T. Nishimura., H. Takii and S. Okada. 1995.
Prevention of hesperidin crystal formation in canned mandarin orange syrup
and clarified orange huice by hesperidin glycodises. Food Science and Tecnology Int., 1(1):29-33.
76) Ting S.V. 1958. Enzymatic hydrolysis of naringin in grapefruit. J Agric Food Che., 6:54-9.
77) Thomas D.W., C.V. Smythe and M.D. Labee.1958. Enzymatic hydrolysis of
naringin the bitter principle of grapefruit. Food Res., 23:591-598.

55
78) Vila Real H., J.A.J. Alfada., A. R. Calado and M. Ribeiro. 2006. High
pressure–Temperature effects on enzymatic activity: Naringin
bioconversion. Food Chem., 102:565-5790.
79) Villena K. y M. Gutiérrez. 2003. Biopelículas de Aspergillus niger para la
producción de celulasas: algunos aspectos estructurales y fisiológicos. Rev. peru biol., v.10 n.1.
80) Voet D. and J. Voet. Bioquímica. Editorial Panamericana. México D. F. 2006. 3a Ed. Capítulo 14.
81) Wagner C.J., C.W. Wilson and P.E. Shaw. 1991. Reduction of grapefruit
bitter components in a fluidized β-cyclodextrin polymer bed. J Food Sci., 56:31-4.
82) Wilkinson G.D. 1960. Statical Estimation in Enzyme Kinetics, J. Biochem,. 80:324-332.
83) Williams P.J., C.R. Strauss., B. Wilson and R.A. Massy–Westropp. 1982.
Novel monoterpene disaccharide glycosides of Vitis vinifera grapes and
wines. Phytochem., 21:2013-2020.
84) Wulbrandt D., C. Gian and J. Meiwes. 1995. Alpha-L-rhamnosidases for
obtaining rhamnosa, a process for its preparation and use. European Patent 599159.
85) Young N.M., R.A.Z. Jhonston and J.C. Richards. 1989.Purification of the
α-L-rhamnosidasa from Penicillum decumbens and characterization of two
glycopeptides components. Cargohydr Res., 191:53-62.

56
86) Yusof S., H.M. Gazali and G.S. King. 1990. Naringin content in local citrus
fruit. Food Chem., 37:113-121.





























